
Treading LighTLy in BaxTer STaTe Park - Appalachian Trail ...
Upload
georgiasouthernCategory
view
2download
0
ARCHAEOFAUNAL REMAINS FROM THE RAVENSFORD SITE: A REGIONAL AND ETHNOHISTORICAL PERSPECTIVE ON ANIMAL USE IN THE
APPALACHIAN SUMMIT
J. Matthew Compton
DO NOT CITE IN ANY CONTEXT WITHOUT PERMISSION OF AUTHOR
J. Matthew Compton, Southeastern Zooarchaeological Research, LLC, 3216 Arrowwood Dr., Raleigh, NC 27604-2406 ([email protected]) Paper presented at the 71st Annual Meeting of the Southeastern Archaeological Conference, Greenville, SC, November 12-15, 2014

2
Abstract. The Ravensford site is a large multi-component site located in the Appalachian
Summit Region of North Carolina. Excavations produced a large well-preserved archaeofaunal
assemblage dating primarily to the Late Pisgah, Early Qualla, and Late Qualla phases. Analysis
indicates a pattern of animal use consistent with contemporaneous sites and ethnohistorical
accounts of Cherokee subsistence practices. Deer and bear were the most significant animals
utilized, but a wide variety of terrestrial and aquatic fauna were harvested. Notably, a large
number of toad remains are present. Toads are conclusively identified as a foodstuff based on a
recently discovered historic account consistent with the archaeological record.

3
(TITLE SLIDE) The Ravensford site (31SW78/136) is a large multi-component site
located in the Appalachian Summit Region of North Carolina. Excavations conducted from 2004
to 2008 produced a large well-preserved archaeofaunal assemblage dating primarily to the Late
Pisgah, Early Qualla, and Late Qualla phases. The occupation of Ravensford spans a period
which saw increasing interaction with European settlers during the seventeenth and eighteenth
centuries. However, from the late-seventeenth century through the first decade of the eighteenth
century the Cherokee remained relatively isolated from European contact (Hatley 1995:5, 32-33;
Marcoux 2012:9). The remaining decades of the eighteenth century saw increasing interaction
with Europeans that resulted in major economic and social change for the Cherokee. Among the
economic changes related to animal use were the Cherokee’s increasing participation in the
deerskin trade and the gradual adoption of Eurasian domesticates (Goodwin 1977:95; Marcoux
2012:9, 14-15; Riggs 1999; Rodning 2010).
It was hoped that the Ravensford assemblage might provide some of the earliest evidence
for these shifts in animal use associated with increasing interaction with European colonists.
Instead, analysis indicates a relatively stable animal use pattern from the Late Pisgah to Late
Qualla phases consistent with contemporaneous sites and ethnohistorical accounts of early-
eighteenth century Cherokee subsistence.
Perhaps the most curious aspect about the Ravensford faunal assemblage is the
abundance of amphibian remains, particularly those of toads. Though generally not a common
pattern observed among southeastern archaeological sites, the same pattern of amphibian
abundance has been observed at other late-period sites of the Appalachian Summit region
(Runquist 1979; VanDerwarker and Detwiler 2000; Vavrasek 2012). In this paper I discuss

4
previous researchers’ interpretations of high-toad abundance in faunal assemblages of the region
and ultimately conclude that toads were indeed a food resource.
RAVENSFORD
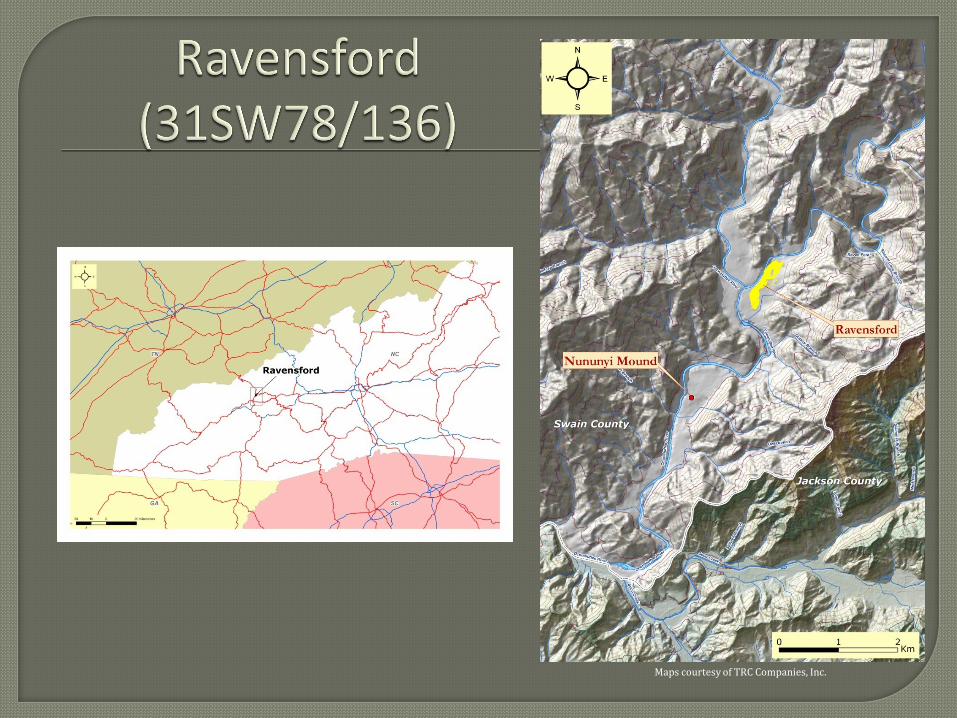
(MAP SLIDE) The Ravensford site is situated in Swain County, North Carolina along the
Raven Fork near its confluence with the main branch of the Oconaluftee River. The site is
located on the Qualla Boundary just north of the town of Cherokee. The land was acquired by the
Eastern Band of Cherokee Indians from the Great Smoky Mountains National Park in a land
exchange in order to construct a K-12 educational complex. The archaeological excavations
conducted at Ravensford are the direct result of this land exchange and were completely funded
by the Eastern Band of Cherokee. Data recovery excavations at Ravensford were conducted from
2004 to 2008 by TRC Companies, Inc. under the direction of Tasha Benyshek and Paul Webb
(Benyshek and Webb 2008, 2009; Webb and Benyshek 2008).
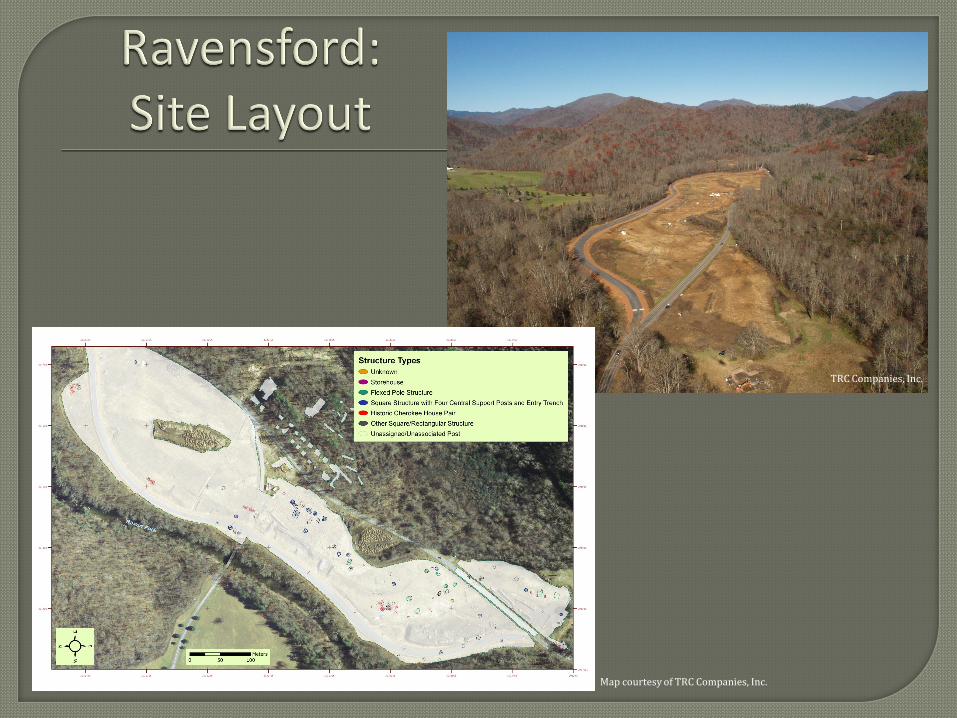
(SITE LAYOUT) Ravensford is a multi-component site covering over 18 ha of alluvial
terrace and neighboring slopes. Aboriginal occupations date from the Early Archaic period to
Historic Cherokee occupations as late as the early-eighteenth century. The site was also the
location of a Euro-American lumber town from 1918 to 1934 (Benyshek and Webb 2008, 2009;
Webb and Benyshek 2008).
(EXCAVATIONS) In order to reveal cultural features over a large portion of the terrace,
an area of approximately 13 ha was stripped of plow zone using a backhoe. Mechanical stripping
allowed for the identification of 114 aboriginal structures, hundreds of pit features, and 98 likely
burials that were left unexcavated (Benyshek and Webb 2008, 2009; Webb and Benyshek 2008).

5
(COMPONENTS) The most significant occupations at Ravensford date to the late
prehistoric and Historic Cherokee periods. Major late-period components include Early Pisgah
(A.D. 1150-1250), Early Qualla (early- to mid-fifteenth century), and Late Qualla (late-
seventeenth to early-eighteenth centuries) phase occupations (Benyshek and Webb 2008, 2009;
Webb and Benyshek 2008). Each of these major occupations includes multiple structures and
features from which much of the animal remains studied were recovered.
(ANALYTICAL UNIT 1) For this study I present Ravensford faunal remains as two
temporally assigned analytical units. The first combines materials from the minor Late Pisgah
phase occupation with the major Early Qualla phase occupation. (ANALYTICAL UNIT 2) The
second analytical unit combines materials from the minor occupation of the Middle Qualla phase
with the major occupation of the Late Qualla phase.
ZOOARCHAEOLOGICAL METHODS
(METHODS) Animal remains were recovered from Ravensford using a variety of
recovery strategies. Individual specimens were collected by hand during mechanical stripping
operations as well as during piece-plotting activities. However, the vast majority of faunal
remains were recovered from water screening (1/16 in, 1.6 mm) and flotation.
The Ravensford faunal materials were identified following the quality assurance standard
developed by Driver (1992; see also Driver 2011a, 2011b) that was recently “condensed and
modified” by Wolverton (2013:283). All identifications were made by me using my personal
comparative skeletal collection, the comparative collection of the Georgia Museum of Natural
History Zooarchaeology Laboratory, the comparative collection of the Appalachian State

6
Department of Anthropology Zooarchaeology Laboratory, and comparative materials housed at
the North Carolina Museum of Natural Sciences.
RAVENSFORD ANIMAL USE
(LATE PISGAH/EARLY QUALLA FAUNAL) In order to assess the relative abundance
of different groups of animals identified, taxa are summarized into faunal categories based on
major taxonomic groups. These faunal categories are compared in terms of Number of Identified
Specimens (NISP), Minimum Number of Individuals (MNI), specimen weight, and biomass
(Quitmyer 1985; Reitz and Quitmyer 1988; Reitz et al. 1987; Reitz and Wing 2008:68, 237-239).
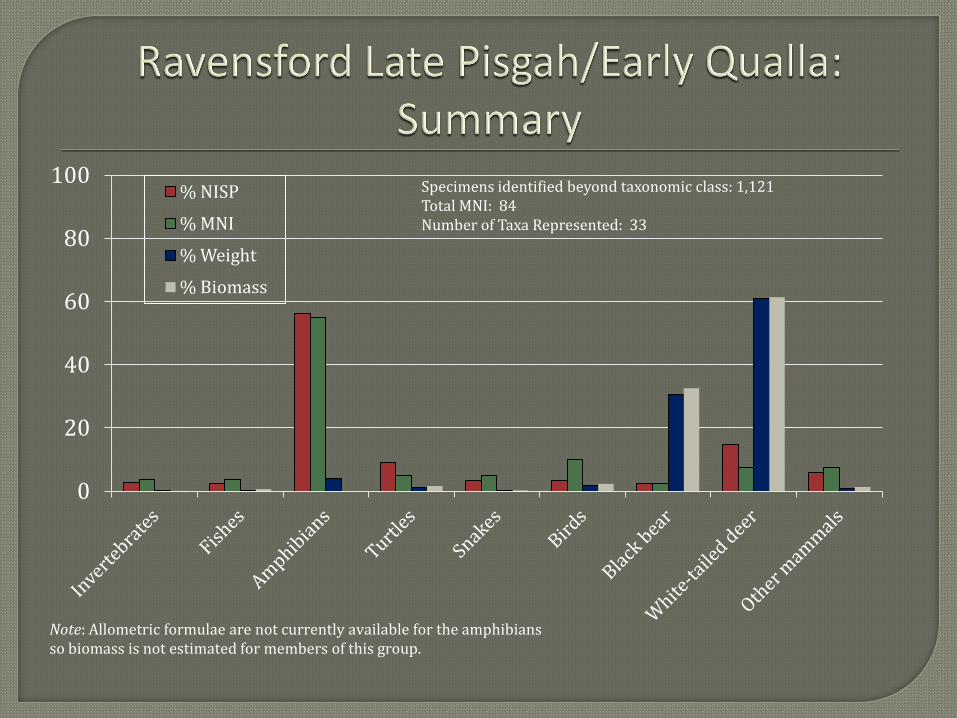
The first faunal analytical unit represents a combination of materials recovered from Late
Pisgah and Early Qualla contexts. The majority of the remains, however, date to the Early Qualla
phase. The collection includes 1,121 specimens identified beyond taxonomic class (i.e., excludes
Mammalia, Vertebrata, etc. counts). Thirty-three taxa are present representing a minimum of 84
individuals.
The Late Pisgah/Early Qualla collection includes a variety of terrestrial and aquatic taxa.
Not surprisingly, the most abundant species in terms of weight, biomass and therefore meat
contribution was the white-tailed deer (Odocoileus virginianus). Black bear (Ursus americanus)
is the only category other than the white-tailed deer that contributes a significant amount of
biomass. When these two mammal species are combined they contribute 94 percent of the total
biomass, suggesting a heavy reliance on large mammals for food. But the presence of an
additional 31 smaller animal species representing 90 percent of the estimated number of
individuals points to a more diverse resource base than the dominance of large mammals in the
biomass calculations indicates.

7
Excluding amphibians for the moment, the most abundant taxa other than black bear and
white-tailed deer are fish of the sucker family (Catostomidae), box turtle (Terrapene carolina),
colubrid snakes (Colubridae), passenger pigeon (Ectopistes migratorius), wild turkey (Meleagris
gallopavo), and squirrels (Sciurus spp.). These results, along with the abundance of white-tailed
deer and black bear, compare favorably to other zooarchaeological collections recovered from
Native American sites in the Appalachian Summit region (Kimball et al. 2004; Runquist 1979;
VanDerwarker and Detwiler 2000; Vavrasek 2012).
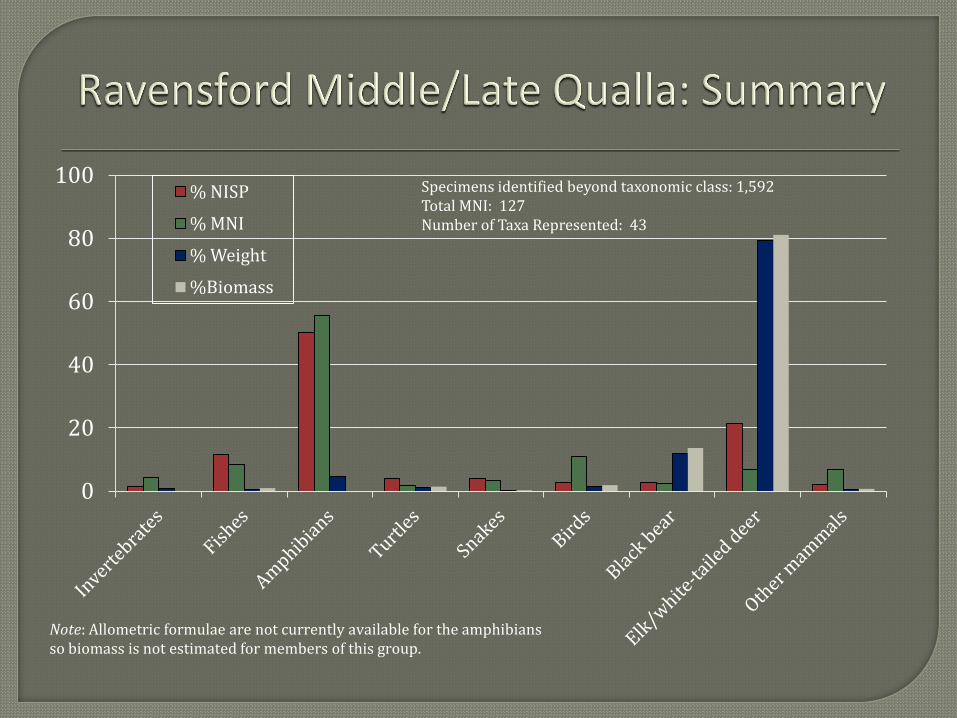
(MIDDLE/LATE QUALLA FAUNAL) The second faunal analytical unit represents a
combination of materials recovered from Middle and Late Qualla phase contexts. But again the
majority of the remains date to the latter of these two phases. The Middle/Late Qualla collection
is somewhat larger than the Late Pisgah/Early Qualla collection including 1,592 specimens
identified beyond taxonomic class. Forty-three taxa are present representing a minimum of 127
individuals.
As you can see the Middle/Late Qualla collection is very similar to the Late Pisgah/Early
Qualla collection in terms of the representation of the different groups of animals. The primary
difference between the two collections is the greater representation of the Elk/White-tailed deer
category in the Middle/Late Qualla collection. Elk (Cervus elaphus) is grouped with white-tailed
deer but contributes a minimal amount of biomass to the category since it is represented by only
two specimens. The higher representation of white-tailed deer in terms of biomass is countered
by a lower representation of black bear. However, the two categories still contribute nearly 95
percent of the total biomass indicating a continued reliance on large mammals for the majority of
animal products consumed. Smaller animals present in the collection are also similar to those
observed for the Late Pisgah/Early Qualla collection with fish of the sucker family, box turtle,

8
colubrid snakes, members of the ducks, geese, and swans family (Anatidae), wild turkey, rabbit
(Sylvilagus spp.), turkey, and squirrel the most abundant animals represented other than
amphibians.
While it was hoped there might be some indication of changes in the animal use
strategies of the Late Qualla inhabitants of Ravensford associated with increasing interaction
with Europeans, there seems to be great continuity over much of the duration of the Ravensford
occupation with little change in animal use practices. It is tempting to suggest that the higher
abundance of deer in the Middle/Late Qualla collection may be related to participation in the
deerskin trade, but this is far from certain without additional lines of evidence. Further, the low
representation of European trade goods in the Ravensford artifact assemblage seems to indicate
only limited or indirect contact with European colonists. Although peach pits have been
recovered from Ravensford, indicating the adoption of some Eurasian domesticates, no
domesticated animals other than the native domestic dog (Canis lupus familiaris) are present in
the Middle/Late Qualla collection.
AMPHIBIANS AT RAVENSFORD
Perhaps the most striking similarity and unusual characteristic of the two faunal
collections is the abundance of amphibians, which dominate the collections in terms of NISP and
MNI. (AMPHIBIANS DATA) When all faunal materials assigned to the Late Pisgah and Qualla
phases are combined the number of amphibian individuals is estimated at 133 with at least four
taxa represented. The four taxa identified include probable American toad (Anaxyrus cf.
americanus), American bullfrog (Lithobates catesbeianus), hellbender (Cryptobranchus
alleganiensis), and lungless salamanders (Plethodontidae). In some cases, specimens could not

9
conclusively be identified to these lower-level taxonomic categories and were therefore assigned
to the broader categories of frogs and toads (Anura), North American toads (Anaxyrus spp.),
American water frogs (Lithobates spp.), and salamanders (Caudata).
Of the 1,964 amphibian specimens identified only 65 were identified as salamander
remains. Frogs and toads are far more represented at Ravensford than salamanders with 1,872
identified specimens representing a minimum of 127 individuals. American water frogs are
represented by 37 specimens with only three specimens positively identified to the species
American bullfrog.
Far exceeding the number of American water frogs in the collection are North American
toads with 1,275 specimens representing a minimum of 116 individuals. Of these, 52 specimens
representing 19 individuals were tentatively identified as probable American toad. The ability of
American toads and Fowler’s toads to hybridize limits the ability to absolutely assign specimens
to one taxon or the other. Nevertheless, close examination of specimens, particularly the femur
and tibiofibula, points to the American toad as the primary species represented.
The high abundance of toads in the collection suggests they may have been a source of
food. Although biomass estimates are not currently available for frogs and toads, the
considerable number of individuals represented does suggest frogs and toads could have supplied
a substantial amount of meat.
At least two ethnohistoric accounts suggest some familiarity with eating frogs and/or
toads by the Cherokee even if the association was not necessarily a favorable one. In Schwarze
(1923:55) book on Moravian missions among the Southeastern Indians he states that the Lower
Cherokee referred to the Upper Cherokee as “frog-eaters…because they were said to not be
averse to eating frogs”. This was apparently meant as a derogatory nickname but how it relates to

10
actual frog consumption is not known. In another example Hudson (1976:411) notes that
Cherokee stickball players were not to eat frogs before games so there bones did not break like
those of frogs during competition.
While these ethnohistoric accounts provide some evidence of Cherokee familiarity with
eating frogs and/or toads it does not address one significant factor about toads that make them an
unlikely food source: they are poisonous. (PHARMACOLOGICAL AGENT) North American
toads possess an enlarged pair of granular glands atop the neck called parotoid glands. These
glands produce a variety of chemical compounds that act to guard against bacterial and fungal
infections as well as to ward off predators. The primary chemical compounds produced by the
parotoid glands include biogenic amines, alkaloids, peptides, and the steroids bufogenines and
bufotoxins (Clarke 1997). Combined, these secretions can produce significant effects on human
vascular, muscular, and neurological systems including inflammation of the mouth and throat,
nausea, vomiting, irregular heartbeat, seizures, hallucinations, and even death (Clarke 1997; Weil
and Davis 1994).
The abundance of toad remains among some Mesoamerican archaeological sites has been
attributed by some researchers to the use of toad venom for its hallucinogenic properties by the
Olmec and Maya (Weil and Davis 1993). A similar interpretation has been made for toad-
bearing sites of the Appalachian Summit region by Jeanette Runquist (1979).
(SITES WITH TOADS) As part of her dissertation research, Runquist (1979) analyzed
faunal remains from the Warren Wilson (31BN29) and Coweeta Creek (31MA34) sites. The
Warren Wilson site is a medium size Pisgah phase (A.D. 1200-1400) village located along the
Swannanoa River in Buncombe County, North Carolina. Coweeta Creek is a seventeenth-century
Cherokee village located in Macon County, North Carolina near the junction of Coweeta Creek

11
and the Little Tennessee River. One striking characteristic of these two faunal assemblages is
their similarity in terms of the amphibian representation to that of the Ravensford assemblage. At
Warren Wilson, frogs and toads account for 18 percent of the specimens identified (NISP=1,508)
and 43 (MNI=145) percent of the estimated number of individuals. Amphibian representation at
Coweeta Creek is even higher with frogs and toads accounting for 59 percent of the specimens
identified (NISP=9,058) and 74 percent of the estimated number of individuals (MNI=621). Frog
and toad taxa identified at the two sites include North American toads, American water frogs,
American bullfrog, and eastern spadefoot (Scaphiopus holbrookii). Only the eastern spadefoot is
not present in the Ravensford assemblage. Like at Ravensford, North American toads are the
dominant amphibian group represented at Warren Wilson and Coweeta Creek.
Although Runquist considers the possibilities that the high abundance of toads at the
Warren Wilson and Coweeta Creek sites were either the result of natural entrapment or food use,
she ultimately attributes their abundance to their use as medicinal or hallucinogenic agents by the
inhabitants of the two villages (Runquist 1979:261).
A different interpretation of these two sites’ faunal assemblages is offered by Tom Whyte
(1988) based on his experimental study of pit entrapment among small animals. The experiment
consisted of 15 pits dug near Knoxville, Tennessee that were observed for over a year. The study
recorded an extraordinary variety and number of animals that became entrapped in the pits over
the course of the experiment (Whyte 1988, 1991). Based on the information gained from the
experimental study, Whyte proposes the large number of toad remains found at the Warren
Wilson and Coweeta Creek sites is the result of natural entrapment.
A fourth site containing abundant frog and toad remains is the Smokemont (31SW393)
site in Swain County, North Carolina (Vavrasek 2012). Smokemont is located very close to

12
Ravensford, only a few kilometers to the north along the Oconaluftee River. Smokemont is
interpreted as an early Late Qualla phase Cherokee farmstead dating to the first half of the
eighteenth century. The Smokemont faunal assemblage is dominated by frog and toad remains
with 70 percent of specimens and 80 percent of the individuals represented. In her study of the
Smokemont faunal assemblage Jessica Vavrasek (2012) concludes that the high abundance of
frog and toad remains is not the result of natural entrapment due to the higher frequency of limb
elements represented. She suggests toads were possibly consumed in a stew, but cannot rule out
some type of “ritual-based” use due to the toxic nature of toad skin (Vavrasek 2012:132).
TESTING THE RAVENSFORD AMPHIBIAN REMAINS
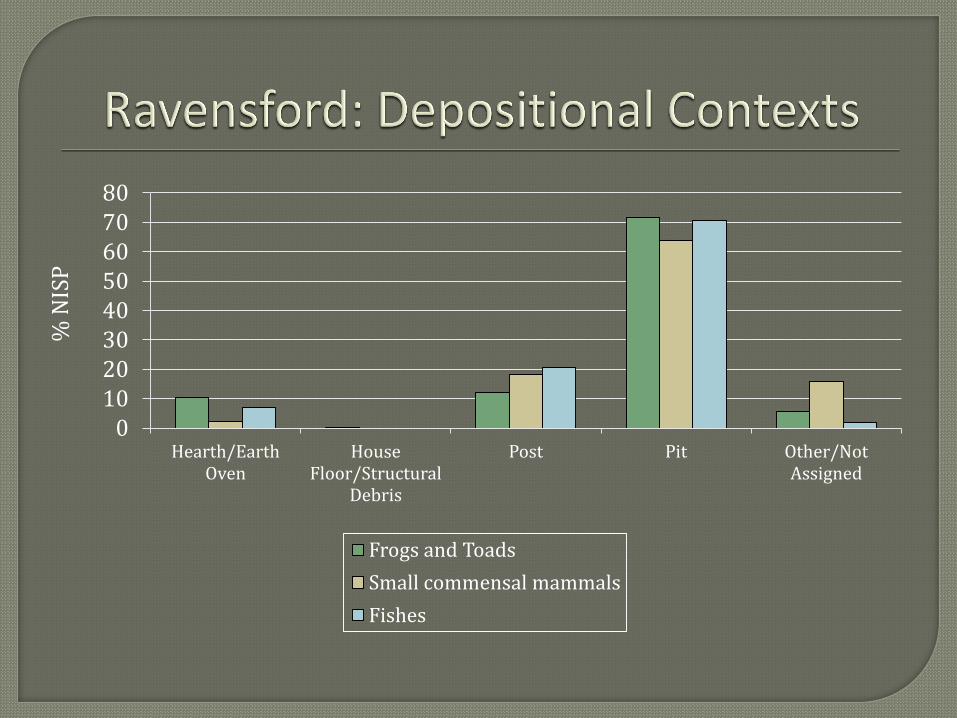
(DEPOSITIONAL CONTEXTS) To investigate natural entrapment as a source of the
large number of toads present in the assemblage an analysis of depositional contexts was
conducted. Frog and toad remains were quite ubiquitous at Ravensford, occurring in 29 percent
of the proveniences studied. This ubiquity is comparable to the white-tailed deer, found to occur
in 30 percent of proveniences studied. Analysis of depositional contexts in which frogs and toads
occur reveals that frog and toad remains were most often recovered from pit contexts. This
pattern is what would be expected if the frog and toad remains represent naturally entrapped
individuals. But fish, clearly a food item and not the result of natural entrapment, were also most
frequently recovered from pit features. This correlation probably has to do with the volume of
materials recovered from pit features compared to other depositional contexts rather than to the
mode of deposition. From this analysis is does not appear that toads are more frequently
associated with features likely to cause entrapment, any more so than known food items.

13
(MODIFICATIONS) Animals utilized by humans for food or other uses tend to exhibit
signs of being processed including butchery marks and differential representation of body parts
(Stahle 1996; Thomas 1971). So in order to determine if the Ravensford frogs and toads
represent cultural debris or natural deposition, a series of analyses were performed to examine
bone modification and skeletal representation.
The first investigates modifications of frog and toad skeletal remains. Unfortunately, no
butchery marks have been observed among frog or toad remains at Ravensford. Direct evidence
of the consumption of frogs or toads is provided by the identification of four digested specimens,
but it cannot be determined if these specimens were consumed by humans or some other animal
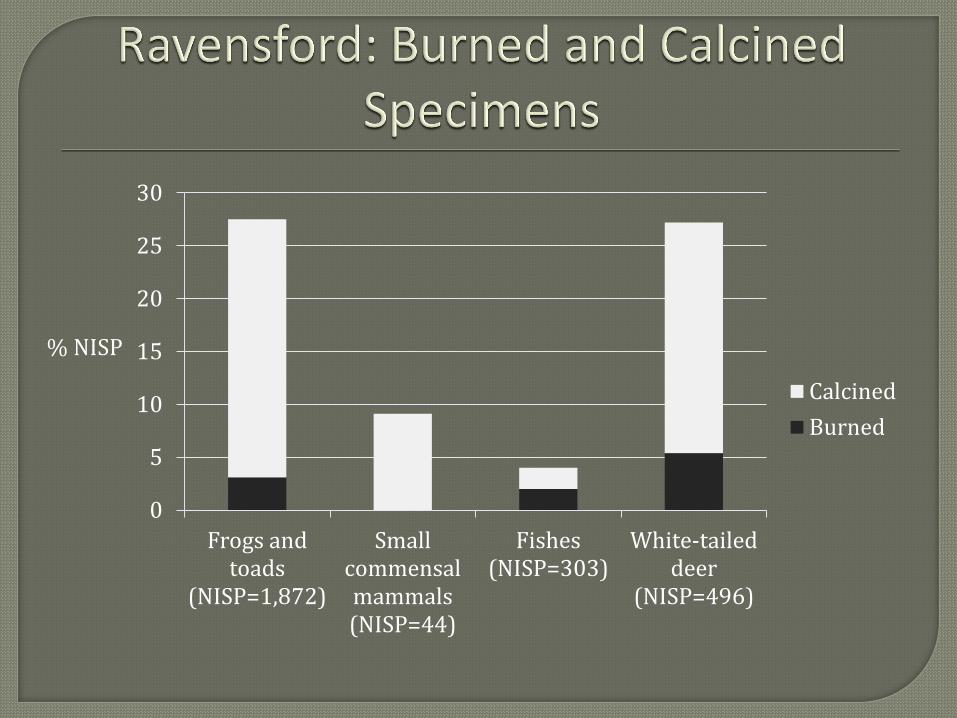
such as dogs. The most frequently observed modifications among frog and toad remains are
burning and calcination. Twenty-eight percent of the frog and toad specimens are either burned
or calcined. While this might seem to suggest the cooking of frogs or toads, the amount of heat
required to burn and/or calcine skeletal bone would render the flesh of the animal inedible. More
likely, burning and calcination among frog and toad remains represents burning of garbage by
the inhabitants of Ravensford. Whether or not these burned and calcined specimens represent
intentional burning of food remains or secondary burning of naturally interred skeletal remains is
not as easily discerned. The presence of some calcined small mammal remains, assumed to
represent commensal taxa, seems to support the later burning of naturally deposited skeletons of
commensal species.
The difference in the frequency of burning and calcination among known food animals,
fish and white-tailed deer, is not completely understood, but may represent differential
preservation between the more fragile bones of fish and the more robust bones of the white-tailed
deer. Presumably, frogs and toads would exhibit a pattern more similar to fish in this regard but

14
this discrepancy cannot be immediately resolved and may require investigation of a larger
sample of fish remains.
(ELEMENT DISTRIBUTION) Differential representation of skeletal elements is often
cited as evidence of human utilization of animals (Thomas 1971). This attribution is most often
stated in regard to large-bodied animals where transport and distribution of large portions of
meat play a role in producing differentially represented skeletal element profiles. Smaller
animals would not be subject to the same transport and economic redistribution factors.
Ethnohistoric accounts often describe small animals as being eaten in their entirety or boiled
whole to produce a broth or stew. The consumption of entire animals would produce little if any
archaeological evidence other than digested bone (Whyte 1988:12). If small animals were boiled
whole to produce a broth or stew the remaining uneaten hard parts would likely show little
evidence of modification and represent a nearly complete skeleton, similar to what would be
expected for animals deposited naturally (Whyte 1988:12).
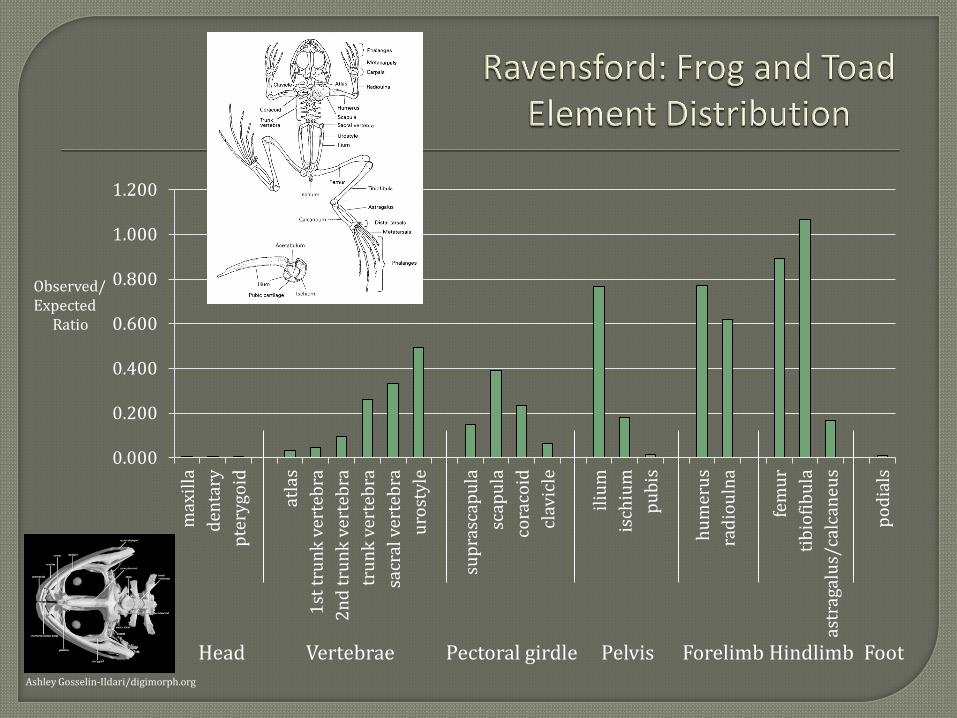
Toads, however, seem unlikely to have been eaten or cooked in their entirety. The
toxicity of toad venom contained in the animal’s parotoid glands would have required some
processing to make the animals edible. Frog and toad elements represented from Ravensford
indicate most portions of the body were represented except for one notable exception: the head.
Of the 1,872 frog and toad specimens identified, only four are skull elements. A similar pattern
was observed by Runquist (1979:53, Table 10) at the Warren Wilson and Coweeta Creek sites
and Vavrasek (2012) at the Smokemont site. It is possible the low representation of skull
elements at the three sites is the result of differential preservation of cranial elements. But the
proportion of head elements represented by salamanders seems to argue against this. Of the 65
salamander remains identified, 28 percent (NISP=18) are skull elements.

15
It is possible the lack of head elements is the result of the removal of toad’s poisonous
parotoid glands so they could be consumed. During my initial investigations of the Ravensford
site I came up with two speculative ways in which this may have been accomplished: The first
simply involves cutting off the head and neck, including the parotoid glands, with a knife. The
second involves grabbing the head with the thumb below the chin and a flexed indexed finger on
top of the head and then pulling the head off and over the back while at the same time removing
the skin and parotoid glands.
(RECIPE BOOK) And then I found what I consider to be the final word on what and how
toads were being used by the Cherokee in the Appalachian Summit Region. From a small book
on Cherokee cooking traditions produced by the Museum of the Cherokee Indian in 1951, I
present this description of toad preparation from Aggie Ross Lossiah “Catch toads, twist of their
heads, pull off the skin while all the time holding the animals under running water lest the meat
become very bitter. Parboil, then cook as any other meat (Ulmer and Beck 1951:51)” Aggie Ross
Lossiah was born in 1880 and was the granddaughter of John Ross, Principle Chief of the
Cherokee Nation at the time of Removal in the 1830s. She provided this description of how to
prepare toads and many other traditional Cherokee foods to the authors Mary Ulmer and Samuel
Beck from her home near Cherokee, North Carolina in 1949 (Ulmer and Beck 1951:42).
I could not have asked for a better description of the preparation technique. It addresses
how the parotoid glands were removed without tainting the meat and why the skeletal elements
of the head are not present in the archaeological assemblage. In all likelihood, the toads were
prepared by decapitation in the adjacent Ravens Fork where the skull bones likely remained.

16
CONCLUSION
(ACKNOWLEDGMENTS) In conclusion, the Ravensford faunal assemblage exhibits
patterns of animal use similar to those found at other late-period sites of the Appalachian Summit
region. The white-tailed deer and black bear were the greatest contributors of meat to the diet of
the inhabitants of Ravensford, but fish, amphibians, turtles, snakes, birds, and other smaller
mammals also played a significant role in the subsistence strategy. Animal use from the Late
Pisgah phase to the early part of the Late Qualla phase was relatively stable with very little shift
in animal use that can be attributed to increasing European contact.
Perhaps the most significant finding of the current research is the conclusion that the
abundance of toads observed among numerous sites in the Appalachian Summit Region is the
result of the use of toads as a source of food rather than use as a pharmacological agent or due to
natural entrapment. Beyond identifying toads as a source of food, the greater significance of this
finding is that it documents a culinary tradition passed down from generation to generation for
over 500 years until it was finally put into print in 1951. Further, the finding illustrates the utility
of conducting element distribution analysis among small animals to better understand
archaeofaunal assemblages.

17
ACKNOWLEDGMENTS
I would like to thank Paul Webb and Tasha Benyshek of TRC Companies, Inc. for the
opportunity to study the Ravensford faunal materials. Funding was provided by TRC Companies,
Inc. through a contract with the Eastern Band of Cherokee Indians to perform data recovery
excavations of the Ravensford site. I would also like to thank Jeffrey C. Beane, Arthur E. Bogan,
Becky Desjardins, Lisa J. Gatens, John A. Gerwin, Benjamin M. Hess, Gabriela M. Hogue,
Brian J. O’Shea, Wayne C. Starnes, and Bryan L. Stuart for providing access to comparative
materials housed at the North Carolina State Museum of Natural Sciences. Access to the
comparative collection of the Georgia Museum of Natural History Zooarchaeology Laboratory
was graciously provided by Elizabeth J. Reitz. Thomas R. Whyte of the Appalachian State
Department of Anthropology Zooarchaeology Laboratory also generously allowed use of
comparative materials.

18
REFERENCES CITED
Benyshek, Tasha and Paul Webb 2008 Mississippian and Historic Cherokee Structure Types and Settlement Plans at Ravensford. Paper presented at the 65th annual meeting of the Southeastern Archaeological Conference, Charlotte, NC. 2009 The Ravensford and Macon County Airport Sites. Paper presented at the symposium North Carolina Appalachian Summit Archaeology: New Visions of Ancient Times. Appalachian State University, Boone, North Carolina. Clarke, B. T. 1997 The Natural History of Amphibian Skin Secretions, Their Normal Functioning and Potential Medical Applications. Biological Review 72:365-379. Driver, Jonathan C. 1992 Identification, Classification and Zooarchaeology. Circaea 9(1):35-47. 2011a Identification, Classification and Zooarchaeology (featured reprint and invited comments). Ethnobiology Letters 2:19-39. 2011b Twenty Years after Identification, Classification and Zooarchaeology. Ethnobiology Letters 2:36-39. Goodwin, Gary C. 1977 Cherokees in Transition: A Study of Changing Culture and Environment Prior to 1775. Research Paper No. 181. University of Chicago Department of Anthropology, Chicago. Hatley, Tom 1993 The Dividing Paths: Cherokees and the South Carolinians Through the Era of Revolution. Oxford University Press, New York. Hudson, Charles 1976 The Southeastern Indians. University of Tennessee Press, Knoxville. Kimball, Larry R., M. Scott Shumate, Thomas R. Whyte, and Gary D. Crites 2004 Hopewellian Subsistence and Ceremonialism at Biltmore Mound, Biltmore Estate, North Carolina. Appalachian State University Laboratories of Archaeological Science. Submitted to the Committee for Research and Exploration, National Geographic Society, Washington, D.C. Copies available from Appalachian State University Laboratories of Archaeological Science, Boone, NC. Marcoux, Jon 2012 The Cherokees of Tuckaleechee Cove. Memoirs, No. 52. Museum of Anthropology, University of Michigan, Ann Arbor.

19
Quitmyer, Irvy R. 1985 Zooarchaeological Methods for the Analysis of Shell Middens at Kings Bay. In Aboriginal Subsistence and Settlement Archaeology of the Kings Bay Locality, Volume 2: Zooarchaeology, edited by William H. Adams, pp. 33-48. Reports of Investigations 2. University of Florida, Department of Anthropology, Gainesville. Reitz, Elizabeth J. and Irvy R. Quitmyer 1988 Faunal Remains from two Coastal Georgia Swift Creek Sites. Southeastern Archaeology 7(2):95-108. Reitz, Elizabeth J., Irvy R. Quitmyer, H. Stephen Hale, Sylvia J. Scudder, and Elizabeth S. Wing 1987 Application of Allometry to Zooarchaeology. American Antiquity 52(2):304-317. Reitz, Elizabeth J., and Elizabeth S. Wing 2008 Zooarchaeology, 2nd edition. Cambridge University Press, Cambridge. Riggs, Brett H. 1999 Removal Period Cherokee Households in Southwestern North Carolina: Material Perspectives on Ethnicity and Cultural Differentiation. Ph.D. dissertation, Department of Anthropology, University of Tennessee, Knoxville. University Microfilms International, Ann Arbor. Rodning, Christopher B. 2010 European Trade Goods at Cherokee Settlements in Southwestern North Carolina. North Carolina Archaeology 39:1-84. Runquist, Jeannette 1979 Analysis of the Flora and Faunal Remains from Proto-historic North Carolina Cherokee Indian Sites. Ph.D. dissertation, Department of Zoology, North Carolina State University, Raleigh. University Microfilms International, Ann Arbor. Schwarze, Edmund 1923 The History of the Moravian Missions Among Southern Indian Tribes of the United States. Transactions of the Moravian Historical Society, Special Series, Vol. 1. Times Publishing, Bethlehem, Pennsylvania. Stahle, Peter W. 1996 The Recovery and Interpretation of Microvertebrate Bone Assemblages from Archaeological Contexts. Journal of Archaeological Method and Theory 3(1):31-75. Thomas, David H. 1971 On Distinguishing Natural from Cultural Bone in Archaeological Sites. American Antiquity 36:366-371.

20
Ulmer, Mary and Samuel E. Beck (editors) 1951 Cherokee Cooklore: Preparing Cherokee Foods. Mary and Goingback Chiltoskey, Stephens Press, Cherokee, North Carolina. VanDerwarker, Amber M. and Kandace R. Detwiler 2000 Plant and Animal Subsistence at the Coweeta Creek Site (31MA34), Macon County, North Carolina. North Carolina Archaeology 49:59-77. Vavrasek, Jessica L. 2012 Faunal Analysis. In Archaeological Investigations at Site 31SW393, Smokemont, Swain County, North Carolina, by Michael G. Angst, pp. 121-132. Archaeological Research Laboratory, University of Tennessee, Department of Anthropology, Knoxville. Submitted to Erik Kreusch, Great Smoky Mountains National Park, Gatlinburg, TN. Webb, Paul and Tasha Benyshek 2008 Historic Cherokee Homesteads at the Ravensford Site. Paper presented at The Social Archaeology of Southeastern Frontiers Conference, University of South Carolina, Columbia. Weil, Andrew T. and Wade Davis 1994 Bufo alvarius: A Potent Hallucinogen of Animal Origin. Journal of Ethnopharmacology 41:1-8. Whyte, Thomas R. 1988 An Experimental Study of Small Animal Remains in Archaeological Pit Features. Ph.D. dissertation, Department of Anthropology, University of Tennessee, Knoxville. 1991 Small-Animal Remains in Archaeological Pit Features. In Beamers, Bobwhites, and Blue- points: Tributes to the Career of Paul W. Parmalee, edited by James R. Purdue, Walter E. Klippel, and Bonnie W. Styles, pp. 163-176. Scientific Papers Vol. 23. Illinois State Museum, Springfield. Wolverton, Steve 2013 Data Quality in Zooarchaeological Faunal Identification. Journal of Archaeological Method and Theory 20(3):381-396.

Paper presented at the 71st Annual Meeting of the Southeastern Archaeological Conference, Greenville, SC, November 12-15, 2014
Matthew Compton Southeastern Zooarchaeological Research
photo: M. Compton
Maps courtesy of TRC Companies, Inc.
TRC Companies, Inc.
Map courtesy of TRC Companies, Inc.

TRC Companies, Inc.
TRC Companies, Inc.
map courtesy of TRC Companies, Inc.
TRC Companies, Inc.
M. Compton

Major Occupations: Early Pisgah phase (A.D. 1150-
1250) Early Qualla phase(early to
mid 15th century Late Qualla phase (late 17th to
early 18th century)
Minor Occupations: Archaic-Early, Middle, & Late
(beginning 8000 B.C.) Early and Middle Woodland Late Pisgah phase (14th & 15th
centuries Middle Qualla phase (A.D.
1500-1650)
Historic Euro-American: Ravensford Lumber Town
(A.D. 1918-1934)

Major Occupations: Early Pisgah phase (A.D. 1150-
1250) Early Qualla phase(early to
mid 15th century Late Qualla phase (late 17th to
early 18th century)
Minor Occupations: Archaic-Early, Middle, & Late
(beginning 8000 B.C.) Early and Middle Woodland Late Pisgah phase (14th & 15th
centuries) Middle Qualla phase (A.D.
1500-1650)
Historic Euro-American: Ravensford Lumber Town
(A.D. 1918-1934)

Major Occupations: Early Pisgah phase (A.D. 1150-
1250) Early Qualla phase(early to
mid 15th century Late Qualla phase (late 17th to
early 18th century)
Minor Occupations: Archaic-Early, Middle, & Late
(beginning 8000 B.C.) Early and Middle Woodland Late Pisgah phase (14th & 15th
centuries Middle Qualla phase (A.D.
1500-1650)
Historic Euro-American: Ravensford Lumber Town
(A.D. 1918-1934)

Comparative collections consulted: Georgia Museum of Natural History,
Zooarchaeology Laboratory Appalachian State University,
Department of Anthropology, Zooarchaeology Laboratory
North Carolina Museum of Natural Sciences, Mollusk, Fish, Reptile and Amphibian, Bird, and Mammal collections
Author’s personal comparative collection
waterscreening at Ravensford M. Compton
M. Compton
Identification procedures: follow the quality assurance standard
developed by Driver (1992) as “condensed and modified” by Wolverton (2013:283)
0
20
40
60
80
100 % NISP
% MNI
% Weight
% Biomass
Note: Allometric formulae are not currently available for the amphibians so biomass is not estimated for members of this group.
Specimens identified beyond taxonomic class: 1,121 Total MNI: 84 Number of Taxa Represented: 33
0
20
40
60
80
100 % NISP
% MNI
% Weight
%Biomass
Note: Allometric formulae are not currently available for the amphibians so biomass is not estimated for members of this group.
Specimens identified beyond taxonomic class: 1,592 Total MNI: 127 Number of Taxa Represented: 43

Taxon Common Name NISP MNI Amphibia Indeterminate amphibian 27 Anura Frogs and toads 557 127 Anaxyrus spp. (formerly Bufo spp.) North American toads 1,223 (116) Anaxyrus cf. americanus (formerly Bufo cf. americanus) Probable American toad 52 (19) Lithobates spp. (formerly Rana spp.) American water frogs 37 (10) Lithobates catesbeianus (formerly Rana catesbeiana) American bullfrog 3 (1) Caudata Salamanders 1 Cryptobranchus alleganiensis Hellbender 58 5 Plethodontidae Lungless salamanders 6 1 Total 1,964 133
American Toad American bullfrog Hellbender Ravensford frog and toad remains during analysis
M. Compton M. Compton C. Compton

Parotoid Glands Guard against bacterial and fungal infection Predator defense Pharmacological Agent Can cause significant effects on human body Toad remains among some Olmec and Maya
sites attributed to use of toad venom as a hallucinogen
Charles H. Warren/life.nbii.gov
American toad illustrating parotoid glands
Toad licking in American popular culture

Warren Wilson (31BN29) Pisgah phase (A.D. 1200-1400),
medium size village
Coweeta Creek (31MA34) 17th-century Cherokee village Smokemont (31SW393) early Late Qualla Cherokee
farmstead (ca. A.D. 1715-1750)
Frogs and Toads NISP % NISP MNI % MNI
Warren Wilson1 1,508 18% 145 43% Coweeta Creek1 9,058 59% 621 74% Smokemont2 2,645 70% 113 80%
1Adapted from Runquist 1979:37, Table 8; 2Adapted from Vavrasek 2012:122, Table 22
Smokemont
0 10 20 30 40 50 60 70 80
Hearth/Earth Oven
House Floor/Structural
Debris
Post Pit Other/Not Assigned
Frogs and Toads Small commensal mammals Fishes
% N
ISP
0
5
10
15
20
25
30
Frogs and toads
(NISP=1,872)
Small commensal mammals (NISP=44)
Fishes (NISP=303)
White-tailed deer
(NISP=496)
Calcined Burned
% NISP
0.000
0.200
0.400
0.600
0.800
1.000
1.200 m
axill
a de
ntar
y pt
eryg
oid
atla
s 1s
t tru
nk v
erte
bra
2nd
trun
k ve
rteb
ra
trun
k ve
rteb
ra
sacr
al v
erte
bra
uros
tyle
supr
asca
pula
sc
apul
a co
raco
id
clav
icle
ilium
is
chiu
m
pubi
s
hum
erus
ra
diou
lna
fem
ur
tibio
fibul
a as
trag
alus
/cal
cane
us
podi
als
Observed/ Expected
Ratio
Head Vertebrae Pectoral girdle Pelvis Forelimb Hindlimb Foot Ashley Gosselin-Ildari/digimorph.org

Aggie Ross Lossiah ca. 1949

Funding provided by TRC Companies, Inc. through a contract with the Eastern Band of Cherokee Indians
Tasha Benyshek and Paul Webb of TRC Companies, Inc. for the opportunity to study the Ravensford faunal materials
Elizabeth Reitz of the University of Georgia, Georgia Museum of Natural History, Zooarchaeology Laboratory for providing access to comparative materials
Tom Whyte of Appalachian State University, Department of Anthropology, Zooarchaeology Laboratory for providing access to comparative materials
Jeffrey Beane, Arthur Bogan, Becky Desjardins, Lisa Gatens, John Gerwin, Benjamin Hess, Gabriela Hogue, Brian O’Shea, Wayne Starnes, and Bryan Stuart of the North Carolina Museum of Natural Sciences for providing access to comparative materials
Copyright © 2022 FDOKUMEN







![Inscribing Interaction: Middle Woodland Monumentality in the Appalachian Summit, 100 B.C. - A.D. 400 [Table of Contents]](https://static.fdokumen.com/doc/165x107/631838244de2f6960b03f7c9/inscribing-interaction-middle-woodland-monumentality-in-the-appalachian-summit.jpg)












