
Aqueous extract of black tea ( Camellia sinensis) prevents ethanol + cholecystokinin-induced...
10
Aqueous extract of black tea (Camellia sinensis ) prevents ethanol + cholecystokinin-induced pancreatitis in a rat model Dolan Das, Sandip Mukherjee, Asankur S. Das, Maitrayee Mukherjee, Chandan Mitra * Department of Physiology, Presidency College, Calcutta, India Received 17 June 2005; accepted 7 September 2005 Abstract Black Tea Extract (BTE), a phytocompound has been attributed with a plethora of health-promoting actions. We have previously demonstrated that BTE inhibits chronic hepatitis in a rat model induced with high-fat and ethanol (EtOH). This study reports that BTE prevents altered pancreatic acinar cell functions, oxidative stress, inflammatory changes and DNA damage in the EtOH + cholecystokinin (CCK)-induced model of pancreatitis. The EtOH + CCK model rats were administered with BTE, and were examined the activity of pancreatic digestive enzymes (amylase and lipase), proinflammatory cytokines (IL-6 and TNF-a), oxidative and antioxidative enzymes (nitric oxide, NO; malondialdehyde, MDA; superoxide dismutase, SOD; catalase, CAT), antioxidant level (glutathione, GSH), histopathological changes and the integrity of genomic DNA. Results show that because of chronic EtOH treatment, serum level of amylase and lipase (two biomarkers for pancreatitis) and pancreatic levels of MDA and NO (two biomarkers of oxidative stress) increased significantly, which could be effectively blunted by BTE. BTE could normalize EtOH+CCK-induced suppressed activities of SOD and CAT, and GSH content of pancreatic tissue. Also, histopathological and inflammatory changes during EtOH + CCK-induced pancreatitis could be blunted by BTE. Furthermore, BTE could effectively reduce EtOH + CCK-induced increase in DNA fragmentation and damage. These findings suggest that BTE prevents pancreatitis caused by chronic EtOH + CCK toxicity presumably by enhancing antioxidant, anti-inflammatory and antiapoptotic activity in rats. D 2005 Elsevier Inc. All rights reserved. Keywords: Ethanol+CCK-induced pancreatitis; Oxidative stress; Proinflammatory cytokines; DNA fragmentation; Apoptosis; Black Tea Extract (BTE) Introduction Alcohol abuse is the main cause of pancreatitis in about 80% of cases (Aleynik et al., 1999). In developed countries, 60–70% of patients with chronic pancreatitis have a long history (6–12 years) of heavy consumption of alcohol (190– 220 ml/day) before the onset of clinically apparent disease (Hunger et al., 1997). Although the mechanisms by which ethanol leads to pancreatic damage remain unknown, there is growing evidence that ethanol exerts direct toxic effect on the pancreatic acinar cells (Rydzewska et al., 2001). Ethanol can change systemic and pancreatic lipid metabolism with accu- mulation of lipid droplets within the acinar cells, alter membrane fluidity and integrity. Ethanol even sensitizes the acinar cells to cholecystokinin (CCK)-stimulated intracellular zymogen proteolysis. In presence of combined-stimulation of CCK and alcohol, it is easier to induce acute pancreatitis (Wu, 2000). Mounting evidence has accumulated that oxygen radicals and other reactive oxygen species (ROS) may play a pivotal role in the pathogenesis of acute and chronic pancreatitis (Tsai et al., 1998; Norton et al., 1998; Casini et al., 2000; Andican et al., 2005). Evidence further suggests that chronic alcoholic pancreatitis may develop as a result of repeated necro-inflammatory episodes of acute pancreatitis (Rydzewska et al., 2001). It has been reported that the combination of an ethanol (15%) containing diet and infusion of a low dose of CCK- 8 causes pancreatitis in experimental animal (rat) with increased levels of amylase and lipase, infiltration of neutro- phils and macrophages, and the number of apoptotic acinar cells (Pandol et al., 1999, 2003). All these observations have led to the concept that the development of alcoholic pancre- atitis requires cofactors (Jerrells et al., 2003). In our present study we selected an animal model of alcoholic pancreatitis 0024-3205/$ - see front matter D 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.lfs.2005.09.020 * Corresponding author. 14/17A Golf Club Road, Calcutta—700 033, India. Tel.: +91 33 2413 1383/2417 8645 (R), +91 33 2219 8149 (O). E-mail address: chandan _ [email protected] (C. Mitra). Life Sciences 78 (2006) 2194 – 2203 www.elsevier.com/locate/lifescie
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Aqueous extract of black tea ( Camellia sinensis) prevents ethanol + cholecystokinin-induced...

sevier.com/locate/lifescie
Life Sciences 78 (200
Aqueous extract of black tea (Camellia sinensis) prevents
ethanol+cholecystokinin-induced pancreatitis in a rat model
Dolan Das, Sandip Mukherjee, Asankur S. Das, Maitrayee Mukherjee, Chandan Mitra *
Department of Physiology, Presidency College, Calcutta, India
Received 17 June 2005; accepted 7 September 2005
Abstract
Black Tea Extract (BTE), a phytocompound has been attributed with a plethora of health-promoting actions. We have previously demonstrated
that BTE inhibits chronic hepatitis in a rat model induced with high-fat and ethanol (EtOH). This study reports that BTE prevents altered
pancreatic acinar cell functions, oxidative stress, inflammatory changes and DNA damage in the EtOH+cholecystokinin (CCK)-induced model of
pancreatitis. The EtOH+CCK model rats were administered with BTE, and were examined the activity of pancreatic digestive enzymes (amylase
and lipase), proinflammatory cytokines (IL-6 and TNF-a), oxidative and antioxidative enzymes (nitric oxide, NO; malondialdehyde, MDA;
superoxide dismutase, SOD; catalase, CAT), antioxidant level (glutathione, GSH), histopathological changes and the integrity of genomic DNA.
Results show that because of chronic EtOH treatment, serum level of amylase and lipase (two biomarkers for pancreatitis) and pancreatic levels of
MDA and NO (two biomarkers of oxidative stress) increased significantly, which could be effectively blunted by BTE. BTE could normalize
EtOH+CCK-induced suppressed activities of SOD and CAT, and GSH content of pancreatic tissue. Also, histopathological and inflammatory
changes during EtOH+CCK-induced pancreatitis could be blunted by BTE. Furthermore, BTE could effectively reduce EtOH+CCK-induced
increase in DNA fragmentation and damage. These findings suggest that BTE prevents pancreatitis caused by chronic EtOH+CCK toxicity
presumably by enhancing antioxidant, anti-inflammatory and antiapoptotic activity in rats.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Ethanol+CCK-induced pancreatitis; Oxidative stress; Proinflammatory cytokines; DNA fragmentation; Apoptosis; Black Tea Extract (BTE)
Introduction
Alcohol abuse is the main cause of pancreatitis in about
80% of cases (Aleynik et al., 1999). In developed countries,
60–70% of patients with chronic pancreatitis have a long
history (6–12 years) of heavy consumption of alcohol (190–
220 ml/day) before the onset of clinically apparent disease
(Hunger et al., 1997). Although the mechanisms by which
ethanol leads to pancreatic damage remain unknown, there is
growing evidence that ethanol exerts direct toxic effect on the
pancreatic acinar cells (Rydzewska et al., 2001). Ethanol can
change systemic and pancreatic lipid metabolism with accu-
mulation of lipid droplets within the acinar cells, alter
membrane fluidity and integrity. Ethanol even sensitizes the
acinar cells to cholecystokinin (CCK)-stimulated intracellular
0024-3205/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.lfs.2005.09.020
* Corresponding author. 14/17A Golf Club Road, Calcutta—700 033, India.
Tel.: +91 33 2413 1383/2417 8645 (R), +91 33 2219 8149 (O).
E-mail address: [email protected] (C. Mitra).
zymogen proteolysis. In presence of combined-stimulation of
CCK and alcohol, it is easier to induce acute pancreatitis (Wu,
2000). Mounting evidence has accumulated that oxygen
radicals and other reactive oxygen species (ROS) may play a
pivotal role in the pathogenesis of acute and chronic
pancreatitis (Tsai et al., 1998; Norton et al., 1998; Casini et
al., 2000; Andican et al., 2005). Evidence further suggests that
chronic alcoholic pancreatitis may develop as a result of
repeated necro-inflammatory episodes of acute pancreatitis
(Rydzewska et al., 2001).
It has been reported that the combination of an ethanol
(15%) containing diet and infusion of a low dose of CCK-
8 causes pancreatitis in experimental animal (rat) with
increased levels of amylase and lipase, infiltration of neutro-
phils and macrophages, and the number of apoptotic acinar
cells (Pandol et al., 1999, 2003). All these observations have
led to the concept that the development of alcoholic pancre-
atitis requires cofactors (Jerrells et al., 2003). In our present
study we selected an animal model of alcoholic pancreatitis
6) 2194 – 2203
www.el

D. Das et al. / Life Sciences 78 (2006) 2194–2203 2195
induced by a combination of chronic ethanol and low-dose of
CCK in rat.
During the last decade, numerous in vitro and in vivo
studies have suggested that tea has numerous potentially
beneficial medicinal properties (Chen et al., 2002). The
chemicals that make tea a potential protector of health are
called polyphenols and these polyphenolic compounds are
collectively known as the tea flavonoids. Tea flavonoids
possess antioxidant properties in vitro and have been proposed
as a protective dietary component, reducing the risk of various
cancers and other diseases (Langley-Evans, 2000; Chen et al.,
2002). Further, it has been reported that antioxidants in tea are
more potent than those found in many fruits and vegetables and
neutralize highly reactive molecules, called free radicals, which
damage cells (Luczaj and Skrzydlewska, 2004). It has also
been reported that consumption of black tea resulted in a
significant increase in plasma antioxidant activity (Leenan et
al., 2000; Rizvi et al., 2005). Earlier it was observed that green
tea catechins have a protective effect on the pathogenesis of
cerulein-induced pancreatitis (Takabayashi and Harada, 1997).
The aim of the present study was to examine the efficacy of
black tea extract (BTE) in preventing chronic EtOH+CCK-
induced pancreatitis in a rat model.
Materials and methods
Plant material
The black tea extract was prepared from CTC (Curl, Tear
and Crush) BOP (Broken Orange Pickoe) grade black clonal
tea. Preparation of aqueous extract of black tea was done by
following the method as described elsewhere (Wei et al., 1999).
Briefly, 1.25 g of tea leaves were added to 25 ml of boiling
water and was steeped for 15 min. The infusion was cooled to
room temperature and then filtered. The tea leaves were
extracted a second time with 25 ml of boiling water and
filtered, and the two filtrates were combined to obtain a 2.5%
aqueous-tea extract. The resulting clear solution was similar to
tea brews consumed by human. This black tea extract (BTE)
was fed to animals by gavage technique at a dose of 1 ml/100 g
body weight/day at a temperature of 37 -C (Wei et al., 1999).
Animal model and study protocol
Male albino rats, weighing 145–165 g, were used in the
experiment. They were housed in a temperature controlled
room (25T2 -C) with 12-h light–dark cycles for at least 1
week. After the stabilization period, rats were randomly
divided into four groups: Control (Gr. A), CCK (Gr. B),
EtOH+CCK (Gr. C) and EtOH+CCK+BTE (Gr. D). All rats
were pair-fed with a normal laboratory diet containing 7% of
total energy as fat, 18% as protein and 75% as carbohydrates,
part of which, in the ethanol formula, are isocalorically
replaced by ethanol (to the extent of 36% of total energy)
(Lieber and Decarli, 1994) and free access to drinking water.
Animals of EtOH+CCK and EtOH+CCK+BTE groups were
pair-fed with 15% (v/v) ethanol (Luma and Vorne, 1976)
(volume that contributed isocalorically 36% of total energy)
once daily for 30 days by gavage technique. Animals of
EtOH+CCK+BTE were fed with 2.5% (w/v) BTE at a single
dose of 1 ml/100 g body weight/day for that same period (30
days) by gavage technique (Wei et al., 1999).
All ethical care and treatment of animals were undertaken by
following the guidelines of the Animal Care and Ethics
Committee of Presidency College, Calcutta. After the treatment
period (30 days) was over, rats were fasted and anesthetized by
intramuscular injection of urethane (500 mg/kg body weight)
(Miura et al., 1997). A polyethylene cannula (PE 50) was placed
in the right jugular vein for CCK infusion. Animals of CCK,
EtOH+CCK and EtOH+CCK+BTE groups were infused with
CCK in physiological saline solution at a dose of 50 ng/kg body
weight/h (Pandol et al., 1999) for 1 h. Animals of control group
were infused with physiological saline solution alone. A
midline celiotomy was performed and, the common pancreat-
ic-bile duct was cannulated extraduodenally with a PE 10
catheter through which pancreatic secretions were collected.
The bile duct was ligated on the liver side of the pancreas to
prevent the flow of bile into the common pancreatic-bile duct.
Core temperature was maintained by using a heating pad and
overhead lamp. The animals were allowed to stabilize for 30
min before the onset of infusion. After surgery, the abdomen
was covered with a gauze soaked in liquid Vaseline to avoid
tissue dryness. Animals were killed at the end of the experiment
with an overdose of anesthesia (Saluja et al., 1997). Blood
samples were taken from the heart before the animals were
killed in order to measure TNF-a and IL-6.
Preparation of tissue extracts
The abdomen was opened, pancreas was quickly removed
and placed in a beaker containing ice-cold Tris–HCl buffer
(pH 7.4). It was minced into small pieces on ice and
homogenized immediately in a glass homogenizing tube
equipped with a Teflon pestle. The homogenate was processed
according to the method of Koyama et al. (1983) for the
estimation of NO, MDA and SOD. For CAT estimation, the
tissue was homogenized in ice-cold isotonic phosphate buffer
(pH 7.4) and processed according to the method of Cohen et al.
(1970). For GSH estimation, pancreatic tissue was homoge-
nized according to the method of Ellman (1959) in ice-cold
phosphate buffer (pH 8).
Measurement of amylase and lipase
Biochemical assay of amylase and lipase were performed
from collected pancreatic secretions by using kit methods.
Amylase and lipase kits were obtained from E. Merck (India)
Ltd. and Dyasis (Germany), respectively.
Measurement of tumor necrosis factor-a (TNF-a) and inter-
leukin-6 (IL-6)
Serum TNF-a level was estimated by using the CYTELISA
rat TNF-a kit obtained from CYTIMMUNE Sciences Inc.,

D. Das et al. / Life Sciences 78 (2006) 2194–22032196
Maryland, USA. All samples were assayed in duplicate. The
intra assay variation was 6.7%. To avoid inter assay variation
all samples were run at one time. Optical density of each well
was determined by using a microplate reader (Thermo
Labsystems, Finland).
Serum IL-6 was estimated by using the ELISA Quantikine
rat IL-6 immunoassay kit obtained from R & D Systems Inc.,
Minneapolis, MN, USA. All samples were assayed in
duplicate. The intra assay variation was 5.5%. To avoid inter
assay variation all samples were run at one time. Optical
density of each well was determined by using a microplate
reader (Thermo Labsystems, Finland).
Measurement of NO production
NO production was estimated by Griess reaction (Giusep-
pina et al., 1999) which was expressed in the form of nitrite
accumulation. In brief, 100 Al of tissue extract was mixed with
100 Al of Griess reagent [equal volumes of 1% (w/v)
sulphanilamide in 5% (v/v) phosphoric acid and 0.1% (w/v)
napthylethylenediamine hydrochloride (NEDH)] and incubated
at room temperature for 10 min, and then the absorbance at 550
nm was measured in a UV-double beam spectrophotometer
(Shimadzu 160A). The amount of nitrite in the sample (AMunit) was calculated from a sodium nitrite standard curve. The
results were expressed as AM nitrite/mg of protein.
Measurement of lipid peroxidation
The quantitative measurement of lipid peroxidation was
performed following the thiobarbituric acid (TBA) test. The
amount of MDA formed was quantitated by reaction with TBA
and used as an index of lipid peroxidation. In brief, 2 ml of
sample was added with 1 ml of 20% trichloroacetic acid and 2
ml of 0.67% (w/w) TBA and heated in a lightly stoppered tube
for 10 min in a boiling water bath. The tubes were cooled and
centrifuged and the absorbance was taken at 530 nm. The
results were expressed as nM MDA/mg of protein using molar
extinction co-efficient of the chromophore (1.56�105 cm2/
mM) (Wills, 1987).
Measurement of antioxidant enzymes: SOD and CAT
SOD was assayed according to the method of Misra and
Fridovich (1972) by comparing the adrenochrome formation in
control, CCK, EtOH+CCK, and EtOH+CCK+BTE-supple-
mented groups of animals. In brief, 0.5 ml of 1.8 mM
epinephrine (freshly prepared), 0.5 ml of 0.6 mM EDTA and
0.5 ml of 0.3 M sodium carbonate (pH 10.2) was added with
the sample which contained 100 Ag of protein determined by
trial and error. The reaction was initiated by addition of
epinephrine and the increase in absorbance at 480 nm was
observed.
CAT was assayed by the method described by Cohen et al.
(1970). The enzyme-catalyzed decomposition of H2O2 was
measured. In brief, 0.5 ml aliquot of cold CAT sample and a
blank consisting of 0.5 ml distilled water was taken in test
tubes and the enzymatic reactions was initiated by adding 5 ml
of cold 6 mM H2O2 and mixed thoroughly. After exactly 3 min
the reaction was stopped by rapidly adding 1 ml 6 N H2SO4
and mixed thoroughly. Then 7 ml of 0.01 N KMnO4 reagent
was added, mixed thoroughly and the reading was taken at 480
nm within 30–60 s.
Measurement of GSH
GSH was estimated in the pancreas samples according to the
method of Ellman (1959) and the results were expressed as AMGSH/mg of protein. In brief, 20 Al DTNB (5,5V-dithiobis 2-
nitrobenzoic acid) was added to 3 ml of reagent mixture
(containing 3 ml sample, 2 ml buffer and 5 ml water) in a
cuvette. The colour developed rapidly and the absorbance was
read at 412 nm after 2 min in a UV-double beam spectropho-
tometer (Shimadzu 160A).
Measurement of protein
Protein in the homogenate of pancreas was estimated by the
method of Lowry et al. (1951) using BSA as standard.
Quantification of fragmented DNA by diphenylamine (DPA)
Fresh pancreas samples were collected in phosphate buffer
(pH 8) and DNA damage was quantified in each sample
individually according to the method described earlier by
Kurita-Ochiai et al. (1997). To measure pancreatic DNA
fragmentation by spectrophotometry, a portion of the pancreas
was homogenized in chilled phosphate buffer and the
homogenates were centrifuged at 400 �g for 10 min and
the supernatant was discarded. Pellet was then resuspended in
400 Al of hypotonic lysis buffer (0.2% Triton X-100, 10 mM
Tris, 1 mM EDTA, pH 8.0) and centrifuged for 15 min at
13,800 �g to separate intact chromatin in the pellet from
fragmented/damaged DNA in the supernatant. Pellets were
resuspended in 400 Al hypotonic lysis buffer and then 400
Al 0.5 N perchloric acid was mixed with both pellets and
supernatants. Then, 800 Al diphenylamine (DPA) reagent
[0.088 M DPA, 98% (v/v) glacial acetic acid, 1.5% (v/v) conc.
H2SO4 and 0.5% of 1.6% acetaldehyde solution] was added
and the samples were kept at 4 -C for 48 h. Absorbance was
measured at 575 nm using a UV-double beam spectropho-
tometer (Shimadzu 160A). DNA fragmentation in samples
[(frag. DNA in sup.) / (frag. DNA in sup.+ intact DNA in
pellet)] were expressed as percentage of total DNA appearing
in the supernatant fraction.
Light microscopy and histopathological evaluation
Histological studies of the pancreas were conducted by
methods as described earlier (Bancroft et al., 1996). Briefly, the
pancreas from rats of all groups were carefully isolated,
trimmed of fat, and fixed in 10% buffered formalin. The
formalin-fixed pancreas were then dehydrated with ethanol and
embedded in paraffin. Sections of 4–5 Am were cut and stained

0
500
1000
1500
2000
2500
3000
3500
4000
4500
Pan
crea
tic
Am
ylas
e (U
/L)
#
Co
ntr
ol
CC
K
CC
K+E
tOH
CC
K+E
tOH
+BT
E
*
**
**
0
500
1000
1500
2000
2500
3000
Pan
crea
tic
Lip
ase
(U/L
) #
Co
ntr
ol
CC
K
CC
K+E
tOH
CC
K+E
tOH
+BT
E
*
**
**
A B
Fig. 1. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g body weight/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30 days)+CCK
(50 ng/kg body weight/h for 1 h)-induced pancreatic amylase and lipase activity of rat. Error bars represent meansTSEM (n =6). # Denotes significance based on
Kruskal–Wallis nonparametric ANOVA test at p <0.01; ** denotes significance based on Mann–Whitney U multiple comparison test at p <0.01; * denotes not
significant ( p >0.05).
D. Das et al. / Life Sciences 78 (2006) 2194–2203 2197
with hematoxylin and eosin. All slides were evaluated in a
blinded manner by one of the authors. Pathological changes in
the pancreas were scored as described by Tsukamoto et al.
(1988) as follows: steatosis (the percentage of acinar cells
containing fat droplets): <25%=1+, <50%=2+, <75%=3+,
75%�4+; inflammation and necrosis: 1 focus per low-power
view=1+; >1 focus=2+.
Statistical analysis
Values are expressed as meanTSEM. Statistical significance
was tested by the Mann–Whitney U multiple comparison test
for comparison of data between groups, and the Kruskal–
Wallis nonparametric ANOVA test for the comparison of
sequential data within a group. SPSS 10 software was used for
0
10
20
30
40
50
60
70
80
Ser
um
TN
F-a
lph
a (p
g/m
l) #
Co
ntr
ol
CC
K
EtO
H+C
CK
EtO
H+C
CK
+BT
E
**
***
Ser
um
IL-6
(p
g/m
l) #
A B
Fig. 2. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g body weight/d
(50 ng/kg body weight/h for 1 h)-induced serum cytokine levels (TNF-a and IL-6) o
Kruskal–Wallis nonparametric ANOVA test at p <0.01; ** denotes significance b
significant ( p >0.05).
statistical analysis. Differences were considered significant if
p <0.05.
Results
Effects of administration of chronic EtOH+CCK on pancreatic
amylase and lipase: protective effects of BTE
Fig. 1A and B depict the effect of administration of
EtOH+CCK on pancreatic amylase and lipase activity. Fig.
1A shows that, compared to control, there was no significant
change in the amylase activity level after infusion of CCK alone.
However, in the EtOH+CCK-infused group, this pancreatic
amylase level was found significantly ( p <0.01) higher com-
pared to both control and CCK groups. BTE supplementation
0
50
100
150
200
250
300
350
Co
ntr
ol
EtO
H+C
CK
EtO
H+C
CK
+BT
E
CC
K
*
**
**
ay for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30 days)+CCK
f rat. Error bars represent meansTSEM (n =6). # Denotes significance based on
ased on Mann–Whitney U multiple comparison test ( p <0.01); * denotes not

0
0.5
1
1.5
2
2.5
3
nM
MD
A /m
g o
f p
rote
in #
*
**
**
Co
ntr
ol
CC
K
EtO
H+C
CK
EtO
H+C
CK
+BT
E
Fig. 4. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g/body
weight/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30
days)+CCK (50 ng/kg body weight/h for 1 h)-induced MDA production in rat
pancreatic tissue. Error bars represent meansTSEM (n =6). # Denotes
significance based on Kruskal–Wallis test ( p <0.01); ** denotes significance
based on Mann–Whitney U multiple comparison test ( p <0.01); * denotes not
significant ( p >0.05).
D. Das et al. / Life Sciences 78 (2006) 2194–22032198
was found effective in reducing such EtOH+CCK-induced
increase level of amylase activity ( p <0.01) near to control level.
Similar to amylase, lipase activity was found increased in
EtOH+CCK-infused group, compared to both control and CCK
groups. Like amylase, BTE supplementation in an identical way
reduced EtOH+CCK-induced increased lipase activity near to
control level ( p <0.01) (Fig. 1B).
Effects of administration of chronic EtOH+CCK on TNF-aand IL-6: protective effects of BTE
To ascertain that inflammatory responses occur due to
EtOH+CCK administration and BTE supplementation may
recover this, the serum concentration of pro-inflammatory
cytokines TNF-a and IL-6 were assayed. Both the cytokines,
TNF-a and IL-6 levels in serum (pg/ml) were significantly
higher ( p <0.01) in EtOH+CCK group, compared to CCK and
control groups of animals. Such increased levels of TNF-a and
IL-6 were significantly restored ( p <0.01) after BTE supple-
mentation (Fig. 2A and B).
Effects of administration of chronic EtOH+CCK on NO and
MDA production: protective effects of BTE
Figs. 3 and 4 show the effect of administration of
EtOH+CCK on nitrite accumulation, an indicator of NO
synthesis, and MDA production, an index of lipid peroxidation,
in pancreatic tissue of different groups of rats. Results show
that compared to control, there was no significant change in
NO and MDA production in the CCK group. However, in the
EtOH+CCK-infused group, both NO and MDA production
were found significantly ( p <0.01) higher compared to both
control and CCK groups. BTE supplementation was found
effective in reducing such EtOH+CCK-induced increase level
of NO and MDA production ( p<0.01).
0
1
2
3
4
5
6
7
µM N
itri
te /m
g o
f p
rote
in #
#
Co
ntr
ol
CC
K
EtO
H+C
CK
EtO
H+C
CK
+BT
E
*
**
**
Fig. 3. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g/body
weight/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30
days)+CCK (50 ng/kg body weight/h for 1 h)-induced NO production in rat
pancreatic tissue. Error bars represent meansTSEM (n =6). ## Denotes
significance based on Kruskal–Wallis test ( p <0.001); ** denotes significance
based on Mann–Whitney U multiple comparison test ( p <0.01); * denotes not
significant ( p >0.05).
Effects of administration of chronic EtOH+CCK on SOD and
CAT: protective effects of BTE
Figs. 5 and 6 represent the result of administration of
CCK+EtOH on antioxidant enzymes SOD and CAT in
pancreatic tissue of different groups of rats. Results indicate
that, compared to control, rats infused with CCK showed no
significant ( p >0.05) change in both SOD and CAT activity.
However, EtOH+CCK group of rats showed marked increase
in the activity of both SOD and CAT ( p <0.01) compared to
control and CCK. When BTE was supplemented, this increased
0
1
2
3
4
5
6
IU S
OD
/ m
g o
f p
rote
in #
#
Co
ntr
ol
CC
K
EtO
H+C
CK
EtO
H+C
CK
+BT
E
**
**
*
Fig. 5. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g/body
weight/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30
days)+CCK (50 ng/kg body weight/h for 1 h)-induced SOD activity in ra
pancreatic tissue. Error bars represent meansTSEM (n =6). ## Denotes
significance based on Kruskal–Wallis test ( p <0.001); ** denotes significance
based on Mann–Whitney U multiple comparison test ( p <0.01); * denotes no
significant ( p >0.05).
t
t

0
0.5
1
1.5
2
2.5
3
1st
ord
er r
eact
ion
rat
e co
nst
ant
of
CA
T /
mg
of
pro
tein
##
Co
ntr
ol
CC
K
EtO
H+C
CK
EtO
H+C
CK
+BT
E
**
**
*
Fig. 6. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g/body
weight/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30
days)+CCK (50 ng/kg body weight/h for 1 h)-induced CAT activity in rat
pancreatic tissue. Error bars represent meansTSEM (n =6). ## Denotes
significance based on Kruskal–Wallis test ( p <0.001); ** denotes significance
based on Mann–Whitney U multiple comparison test ( p <0.01); * denotes not
significant ( p >0.05).
D. Das et al. / Life Sciences 78 (2006) 2194–2203 2199
production of SOD and CAT levels were found to be reduced
significantly ( p <0.01).
Effects of administration of chronic EtOH+CCK on GSH:
protective effects of BTE
Fig. 7 depicts the result of administration of EtOH+CCK on
GSH content of pancreas of different groups of rats. Figure
shows that there is no significant change in GSH content in the
0
20
40
60
80
100
µM G
SH
/mg
of
pro
tein
##
Co
ntr
ol
CC
K
EtO
H+C
CK
+BT
E
EtO
H+C
CK
*
**
**
Fig. 7. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g/body
weight/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30
days)+CCK (50 ng/kg body weight/h for 1 h)-induced GSH content in whole
pancreas of rat. Error bars represent meansTSEM (n =6). ## Denotes
significance based on Kruskal–Wallis test ( p <0.001); ** denotes significance
based on Mann–Whitney U multiple comparison test ( p <0.01); * denotes not
significant ( p >0.05).
CCK group, compared to control. In EtOH+CCK group, GSH
level was found significantly ( p <0.01) lower as compared to
control and CCK. On BTE supplementation, this lower level of
GSH was recovered significantly ( p <0.01).
Effects of administration of chronic EtOH+CCK on genomic
DNA damage: protective effects of BTE
The effect of EtOH+CCK treatment on the integrity of
genomic DNA is presented in Fig. 8. Compared to physiolog-
ical saline infused control group (44.8%), there was no
significant ( p >0.05) change in pancreatic genomic DNA
fragmentation in CCK group (46.5%). However, infusion of
EtOH+CCK caused an increase in genomic DNA fragmenta-
tion (70.2%) which was significantly ( p <0.01) higher com-
pared to control and CCK. BTE-supplementation was found
effective in blunting such increased fragmentation to as low as
45% ( p <0.01).
Histopathological observation
No histological alterations were detected in the pancreas
of control (Fig. 9A) and CCK infused groups of rats.
However, the pancreas of rats of EtOH+CCK showed
extensive acinar vacuolization, steatosis, inflammatory
changes and also changes in normal histoarchitecture of the
pancreatic acini (Fig. 9B). All these histopathological
changes, viz., steatosis, acinar vacuolization and inflamma-
tion were markedly reduced after BTE supplementation, and
histoarchitecture of pancreatic acini was returned near to
normal (Fig. 9C). Total pathology scores of pancreas from
different groups of rats are summarized in Fig. 10. Results
show that, compared to control, different pathological
changes, viz., inflammation, necrosis and steatosis were
significantly aggravated in EtOH+CCK-administered groups.
0
10
20
30
40
50
60
70
80
% D
NA
Fra
gm
enta
tio
n #
Co
ntr
ol
CC
K
EtO
H+C
CK
EtO
H+C
CK
+BT
E
*
**
**
Fig. 8. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g/body
weight/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30
days)+CCK (50 ng/kg body weight/h for 1 h)-induced genomic DNA
fragmentation in whole pancreas of rat. Error bars represent meansTSEM
(n =6). # Denotes significance based on Kruskal–Wallis test ( p <0.01); **
denotes significance based on Mann–Whitney U multiple comparison test
( p <0.01); * denotes not significant ( p >0.05).
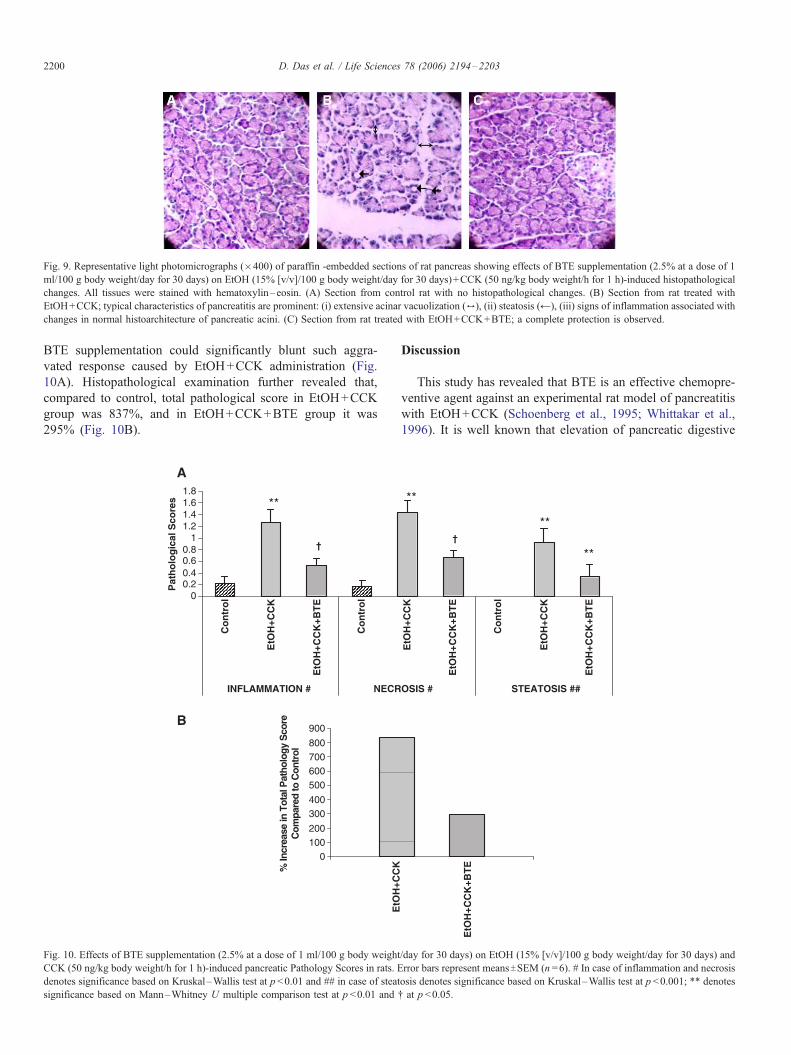
Fig. 9. Representative light photomicrographs (�400) of paraffin -embedded sections of rat pancreas showing effects of BTE supplementation (2.5% at a dose of 1
ml/100 g body weight/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30 days)+CCK (50 ng/kg body weight/h for 1 h)-induced histopathological
changes. All tissues were stained with hematoxylin–eosin. (A) Section from control rat with no histopathological changes. (B) Section from rat treated with
EtOH+CCK; typical characteristics of pancreatitis are prominent: (i) extensive acinar vacuolization (6), (ii) steatosis (@), (iii) signs of inflammation associated with
changes in normal histoarchitecture of pancreatic acini. (C) Section from rat treated with EtOH+CCK+BTE; a complete protection is observed.
D. Das et al. / Life Sciences 78 (2006) 2194–22032200
BTE supplementation could significantly blunt such aggra-
vated response caused by EtOH+CCK administration (Fig.
10A). Histopathological examination further revealed that,
compared to control, total pathological score in EtOH+CCK
group was 837%, and in EtOH+CCK+BTE group it was
295% (Fig. 10B).
00.20.40.60.8
11.21.41.61.8
Co
ntr
ol
EtO
H+C
CK
EtO
H+C
CK
+BT
E
Co
ntr
ol
INFLAMMATION # NECR
Pat
ho
log
ical
Sco
res **
†
0100200
300400
500600700800
900
% In
crea
se in
Tot
al P
atho
logy
Sco
reC
ompa
red
to C
ontr
ol
A
B
Fig. 10. Effects of BTE supplementation (2.5% at a dose of 1 ml/100 g body weight
CCK (50 ng/kg body weight/h for 1 h)-induced pancreatic Pathology Scores in rats. E
denotes significance based on Kruskal–Wallis test at p <0.01 and ## in case of steat
significance based on Mann–Whitney U multiple comparison test at p <0.01 and
Discussion
This study has revealed that BTE is an effective chemopre-
ventive agent against an experimental rat model of pancreatitis
with EtOH+CCK (Schoenberg et al., 1995; Whittakar et al.,
1996). It is well known that elevation of pancreatic digestive
EtO
H+C
CK
EtO
H+C
CK
+BT
E
Co
ntr
ol
EtO
H+C
CK
EtO
H+C
CK
+BT
E
OSIS # STEATOSIS ##
**
†
**
**
EtO
H+C
CK
EtO
H+C
CK
+BT
E
/day for 30 days) on EtOH (15% [v/v]/100 g body weight/day for 30 days) and
rror bars represent meansTSEM (n =6). # In case of inflammation and necrosis
osis denotes significance based on Kruskal–Wallis test at p <0.001; ** denotes
. at p <0.05.

D. Das et al. / Life Sciences 78 (2006) 2194–2203 2201
enzymes lipase and amylase in the blood or pancreatic juice
suggests pancreatitis (Kono et al., 2001; Pfefferkorn et al.,
2002). Significant elevation in the activities of both of these
enzymes in our studies by chronic EtOH+CCK confirmed that
pancreatitis was developed in our experimental rat model. BTE
supplementation could significantly reduce such elevated
activities of these enzymes suggesting that BTE possibly has
a certain degree of protective influence against EtOH+CCK-
induced pancreatitis and altered pancreatic acinar cell func-
tions. Earlier, we observed similar chemopreventive action of
BTE against EtOH-induced chronic hepatitis in a rat model
(Das et al., 2004). Such regulatory influence of BTE against
pancreatitis-induced altered enzymatic function are in quite
agreement with plethora of medicinal effects attributed to BTE
earlier (Krishnamoorthy, 1991; Matsumoto et al., 1991; Chen
et al., 2002).
The role of various cytokines, such as IL-1, IL-6, and TNF-
a in the development of local inflammatory changes and
release of pro-inflammatory cytokines (Grewal et al., 1994),
and also in the development of acute and chronic pancreatitis
(Rongione et al., 1997; Gukovsky et al., 1998; Kuldip et al.,
2003) are well documented. In the present study, combined
EtOH+CCK administration significantly raised both serum IL-
6 and TNF-a levels together with increased activities of serum
amylase and lipase, and thus confirmed that local inflammatory
changes and release of pro-inflammatory cytokines did occur in
our experimental model which possibly caused pancreatitis and
changes in acinar cell functions. All these experimentally
induced pathological changes were blunted significantly by
BTE supplementation, indicating that BTE possibly has an
ameliorative role against the deleterious effects on the pancreas
caused by combined EtOH+CCK administration.
There is growing evidence that ethanol exerts direct toxic
effects on pancreatic acinar cells. Our results from the
pathologic findings showed that chronic EtOH+CCK caused
significant histological changes in the exocrine pancreas as
evidenced by the appearance of steatosis, extensive acinar
vacuolization, inflammatory changes and also changes in
normal histoarchitecture of the pancreatic acini. The appear-
ance of fat droplets in the acinar cells has been used as an
indicative criterion for pancreatitis development in rats (Kono
et al., 2001). Because the exocrine pancreas of rats after BTE
supplementation appeared to be normal in contrast to that of
rats on combined EtOH+CCK, the results indicated that
supplementing rats with BTE may have significant recovery
effects on the histological changes of pancreas.
It was reported earlier that reactive oxygen species (ROS)
generated in pancreatic acinar cells play an important role in
chronic ethanol-induced pancreatitis (Norton et al., 1998;
Szuster-Ciesielska et al., 2001; Wittel et al., 2003;). Further,
ethanol-induced augmented ROS production has been impli-
cated with lipid peroxidation (Takabayashi et al., 1995; Kono et
al., 2001) which induces either apoptosis or necrosis depending
on its extent (Andican et al., 2005). An earlier report also
suggest that ethanol sensitizes the pancreas in inflammatory
responses, cell death and fibrosis caused by CCK (Pandol et al.,
2003). We observed identical results after chronic (30 days)
ethanol ingestion. MDA formation, the index of lipid perox-
idation, was significantly increased after ethanol treatment and
BTE supplementation was potentially effective in reducing
lipid peroxidation, suggesting that BTE might have antioxidant
principles to produce such response.
NO is a cytotoxic agent involved as mediator in inflamma-
tory disorders (Kalf et al., 2000), and because of its
cytotoxicity, overproduction is deleterious to cells (Krippeit-
Drews et al., 1995). It has also been reported that NO can
induce oxidative stress (Lynn et al., 1998; Andican et al.,
2005). In our studies, EtOH+CCK administration was found to
cause a significant increase in NO production which, we
speculate, might be responsible for pancreatic injury and
inflammatory changes. BTE supplementation could reduce this
enhanced NO production, even lower than the control value.
Earlier, it was reported that black tea scavenges NO and
peroxynitrite, and also inhibits the excessive production of NO
by the inducible form of NO synthase (iNOS) (Paquay et al.,
2000). Therefore, it seems logical to infer that BTE, because of
its antioxidant principles, might be capable of protecting the
pancreas from EtOH+CCK-induced injury and inflammatory
changes.
Our study further revealed that chronic (30 days) EtOH
exposure decreased the activities of the ROS scavenging
enzymes, SOD and CAT. This is in line with an assumption
suggested earlier by Sandhir and Gill (1999) that a decrease in
the activity of antioxidant enzymes SOD, glutathione perox-
idase (GPx) and glutathione following ethanol exposure may
be due to the damaging effects of free radicals or alternatively
could be due to a direct effect of acetaldehyde, formed from
oxidation of ethanol, on these enzymes. BTE supplementation,
in our studies, could restore the activity of both of these
antioxidant enzymes and possibly could reduce generation of
free radicals and pancreatic damage. This was found to be
similar with that observed earlier that tea flavonoids have anti-
oxidant properties (Langley-Evans, 2000; Chen et al., 2002)
and is a powerful chemopreventive agent against oxidative
damage caused by free radicals (Sarkar and Bhaduri, 2001).
Thus, results of these studies together with those of earlier
findings, suggest that BTE possibly because of its rich content
of antioxidative active principles has an ability to protect the
pancreas from EtOH+CCK-induced damage by its direct
antioxidative effect.
GSH is a naturally occurring antioxidant important in the
antioxidant defence of the body. Increased lipid peroxidation
and depletion of intracellular GSH in pancreatic tissue in
alcohol-induced acute pancreatitis have been reported (Andican
et al., 2005). Recently it has been reported that tea polyphenols
function as antioxidants through the induction of antioxidant
enzymes, such as glutathione S-transferases and superoxide
dismutases (Frei and Higdon, 2003). This observation is well in
line with our results that BTE supplementation could effec-
tively restore EtOH+CCK-induced reduction in GSH content
of pancreas.
Earlier, it has been reported that DNA fragmentation and
damage signify lipid peroxidation and increased NO produc-
tion which might lead to cell death (Heller et al., 1995; Lynn et

D. Das et al. / Life Sciences 78 (2006) 2194–22032202
al., 1998; Ray et al., 2001). Measurement of DNA fragmen-
tation also has been used for quantification of apoptotic events
in cells (Suenobu et al., 1999). Indeed, our quantitative study
has revealed that EtOH+CCK-induced increased fragmenta-
tion and damage of DNA are correlated with increased lipid
peroxidation and NO production. BTE supplemented reduction
in DNA fragmentation and damage along with the reduction of
lipid peroxidation and NO production therefore suggest that
BTE has potentiality to sustain integrity of genomic DNA by
reducing oxidative stress factors. It is well documented that
green tea is an efficient radical scavenger, activates enzymes
important for detoxification and prevent in vivo DNA damage
and lipid peroxidation (Cheng et al., 1991). Our results indeed
support such notion because BTE supplementation could
effectively reduce EtOH+CCK-induced increase in DNA
fragmentation and damage, as well as NO and MDA formation.
In summary, it may be proposed that BTE prevents
EtOH+CCK-induced alterations in pancreatic acinar cell
functions, oxidative stress, inflammatory changes, DNA
damage and apoptotic changes. Since this model of
EtOH+CCK-induced pancreatic damage in rat simulates many
of the characteristic features of human alcoholic pancreatitis,
we suggest that natural antioxidants and scavenging principles
present in BTE might be equally effective in reducing human
pancreatic damage. Thus, BTE may have some obvious
therapeutic implications against chronic EtOH-induced pancre-
atitis which are to be explored.
Acknowledgements
This work was financially sponsored by the National Tea
Research Foundation (N.T.R.F.), Calcutta, India. The authors
thank Dr. J. R. Vedasiromoni, Drug Development Division,
Indian Institute of Chemical Biology, Calcutta, India for the
generous gift of black tea samples.
References
Aleynik, S.I., Leo, M.A., Aleynik, M.K., Lieber, C.S., 1999. Alcohol-induced
pancreatic oxidative stress: protection by phospholipid repletion. Free
Radical Biology Medicine 26 (5/6), 609–619.
Andican, G., Gelisgen, R., Unal, E., Tortum, O.B., Dervisoglu, S., Karahasa-
noglu, T., Burcak, G., 2005. Oxidative stress and nitric oxide in rats with
alcohol-induced acute pancreatitis. World Journal of Gastroenterology 11
(15), 2340–2345.
Bancroft, J.D., Stevens, A., Turner, D.R., 1996. Theory and Practice of
Histological Techniques. Churchill Livingstone, New York, p. 104.
Casini, A., Galli, A., Pignalosa, P., Frulloni, L., Grappone, C., Milani, S.,
Pederzoli, P., Cavallini, G., Surrenti, C., 2000. Collagen type I synthesized
by pancreatic periacinar stellate cells (PSC) co-localizes with lipid
peroxidation-derived aldehydes in chronic alcoholic pancreatitis. Journal
of Pathology 192 (1), 81–89.
Chen, L., Yang, X., Jiao, H., Zhao, B., 2002. Tea catechins protect against lead-
induced cytotoxicity, lipid peroxidation, and membrane fluidity in HepG2
cells. Toxicological Sciences 69, 149–156.
Cheng, S., Lin, P., Ding, L., Hu, X., Oguni, I., Hara, Y., 1991. Inhibition of
green tea extract on mutagenicity and carcinogenicity. Proceedings of the
International Symposium on Tea Science, Japan. , pp. 195–199.
Cohen, G., Dembiec, D., Marcus, J., 1970. Measurement of catalase activity in
tissue extract. Analytical Biochemistry 34, 30–37.
Das, D., Mukherjee, S., Mukherjee, M., Das, A.S., Mitra, C., 2004. Aqueous
extract of black tea (Camellia sinensis) prevents chronic ethanol toxicity.
Current Science 88 (6), 952–961.
Ellman, G.L., 1959. Tissue sulfhydryl groups. Archives of Biochemistry 82,
70–77.
Frei, B., Higdon, J.V., 2003. Antioxidant activity of tea polyphenols in vivo:
evidence from animal studies. Journal of Nutrition 133 (10), 3275S–3284S.
Giuseppina, M.R., Rosario, M., Oreste, G., Maria, P., Raffaele, D.C., 1999.
Prolactin induction of nitric oxide synthase in rat C6 glioma cells. Journal
of Neurochemistry 73 (6), 2272–2277.
Grewal, H.P., Kotb, M., el.Din, A.M., Ohman, M., Salem, A., Gaber, L.,
Gaber, A.O., 1994. Induction of tumor necrosis factor in severe acute
pancreatitis and its subsequent reduction after hepatic passage. Surgery
115 (2), 213.
Gukovsky, I., Gukovskaya, A.S., Blinman, T.A., Zaninovic, V., Pandol, S.J.,
1998. Early NF kappa B activation is associated with hormone-induced
pancreatitis. American Journal of Physiology 275, G1402–G1414.
Heller, B., Wang, Z.Q., Wagner, E.F., Radons, J., Burkle, A., Fehsel, K.,
Burkart, V., Kolb, H., 1995. Inactivation of the poly (ADP-robose)
polymerase gene affects oxygen radical and nitric oxide toxicity in islet
cells. Journal of Biological Chemistry 270 (19), 11176–11180.
Hunger, R.E., Christoph, M., Z’graggen, K., Friess, H., Buchler, M.W., 1997.
Cytotoxic cells are activated in cellular infiltrates of alcoholic chronic
pancreatitis. Gastroenterology 112, 1656–1663.
Jerrells, T.R., Chapman, N., Clemens, D.L., 2003. Animal model of alcoholic
pancreatitis: role of viral infections. Pancreas 27 (4), 301–304.
Kalf, J.C., Schraut, W.H., Billiar, J.R., Simmons, R.L., Bauer, A.J., 2000. Role
of inducible nitric oxide synthase in postoperative intestinal smooth muscle
dysfunction in rodents. Gastroenterology 118 (2), 316–327.
Kono, H., Nakagami, M., Rusyn, I., Connor, H.D., Stefanovic, B., Brenner,
D.A., Mason, R.P., Arteel, G.E., Thurman, R.G., 2001. Development of an
animal model of chronic alcohol-induced pancreatitis in the rat. American
Journal of Physiology—Gastrointestinal and Liver Physiology 280 (6),
G1178–G1186.
Koyama, I., Tsugikazu, K., Yoshikatsu, S., Munetsugu, K., 1983. A possible
mechanism for the changes in hepatic and intestinal alkaline phosphatase
activities in bile duct ligated rats and guinea pigs. Biochemical Biophysical
Acta 760, 169–174.
Krippeit-Drews, P., Kroncke, K.D., Welker, S., Zempel, G., Roenfeldt, M.,
Ammon, H.P.T., Lang, F., Drews, G., 1995. The effects of Nitric Oxide on
the membrane potential and ionic currents of mouse pancreatic B cells.
Endocrinology 136 (12), 5363–5369.
Krishnamoorthy, K.K., 1991. The nutritional and therapeutic value of
tea. Proceedings of the International Symposium on Tea Science, Japan. ,
pp. 6–11.
Kuldip, S., Narang, A.P., Singh, R.P., Goyal, S.C., 2003. Evaluation of the
prognostic role of inflammatory markers: tumor necrosis factor (TNF-a)
and interleukin-6 (IL-6)—in patients with acute pancreatitis. Indian Journal
of Surgery 65, 480–482.
Kurita-Ochiai, T., Fukushima, K., Ochiai, K., 1997. Butyric acid-induced
apoptosis of murine thymocytes, splenic T cells, and human Jurkat T cells.
Infection Immunity 65 (1), 35–41.
Langley-Evans, S.C., 2000. Antioxidant potential of green and black tea
determined using the ferric reducing power (FRAP) assay. International
Journal of Food Science and Nutrition 51 (3), 181–188.
Leenan, R., Roodenburg, A.J., Tijburg, L.B., Wiseman, S.A., 2000. A single
dose of tea with or without milk increases plasma antioxidant activity in
humans. European Journal of Clinical Nutrition 54 (1), 87–92.
Lieber, C.S., Decarli, L.M., 1994. Animals models of chronic ethanol toxicity.
Methods in Enzymology 233, 585–594.
Lowry, O.H., Rosenbrouhg, N.J., Farr, A.L., Randall, R.J., 1951. Protein
measurement with Folin phenol reaction. Journal of Biological Chemistry
193, 265–271.
Luczaj, W., Skrzydlewska, E., 2004. Antioxidant properties of black tea in
alcohol intoxication. Food Chemical Toxicology 42 (12), 2045–2051.
Luma, P., Vorne, M., 1976. Changes in the hepatic microsomal enzyme activity
during long-term ethanol feeding in rat. Acta Pharmacology and Toxicology
38 (3), 260–266.

D. Das et al. / Life Sciences 78 (2006) 2194–2203 2203
Lynn, S., Shiung, J.N., Gurr, J.R., Jan, K.Y., 1998. Arsenite stimulates poly
(ADP-ribosylation) by generation of nitric oxide. Free Radical Biology
Medicine 24 (3), 442–449.
Matsumoto, N., Tono-Oka, F., Ishigaki, A., Okushio, K., Hara, Y., 1991. The
fate of (�)-epigallocatechin gallate (EGCg) in the digestive tract of rats.
Proceedings of the International Symposium on Tea Science, Japan,
pp. 253–257.
Misra, H.P., Fridovich, I., 1972. The superoxide anion in the antioxidation of
epinephrine: a simple assay for superoxide dismutase. Journal of Biological
Chemistry 247, 3170–3175.
Miura, Y., Murayama, H., Tsuzuki, S., Sugimoto, E., Torii, K., Fushiki, T.,
1997. Long-term consumption of an amino acid diet reduces the pancreatic
enzyme secretion response to a trypsin inhibitor in rats. Journal of Nutrition
127 (7), 1377–1381.
Norton, I.D., Apte, M.V., Haber, P.S., McCaughan, G.W., Pirola, R.C., Wilson,
J.S., 1998. Cytochrome P450E1 is present in rat pancreas and is induced by
chronic ethanol administration. Gut 42 (3), 426–430.
Pandol, S.J., Periskic, S., Gukovsky, I., Zaninovic, V., Jung, Y., Zong, Y.,
Solomon, T.E., Gukovskaya, S., Hidekazu Tsukamoto, H., 1999. Ethanol
diet increases the sensitivity of rats to pancreatitis induced by cholecysto-
kinin octapeptide. Gastroenterology 117, 706–716.
Pandol, S.J., Gukovsky, I., Satoh, A., Lugea, A., Gukovskaya, A.S., 2003.
Animal and in vitro models of alcoholic pancreatitis: role of cholecysto-
kinin. Pancreas 27 (4), 297–300.
Paquay, J.B., Haenen, G.R., Stender, G., Wiseman, S.A., Tijburg, L.B., Bast,
A., 2000. Protection against nitric oxide toxicity by tea. Journal Agricultural
and Food Chemistry 48 (11), 5768–5772.
Pfefferkorn, M.D., Fitzgerald, J.F., Croffie, J.M., Gupta, S.K., Caffrey, H.M.,
2002. Direct measurement of pancreatic enzymes: a comparison of
secretagogues. Digestive Diseases and Science 47 (10), 2211–2216.
Ray, S.D., Balasubramanian, G., Bagchi, D., Reddy, C.S., 2001. Ca2+-
calmodulin antagonist chlorpromazine and poly (ADP-robose) polymerase
modulators 4-aminobenzamide and nicotinamide influence hepatic expres-
sion of BCL-XL and P53 and protect against acetaminophen-induced
programmed and unprogrammed cell death in mice. Free Radical Biology
Medicine 31 (3), 277–291.
Rizvi, S.I., Zaid, A.M., Anis, R., Mishra, N., 2005. Protective role of tea
catechins against oxidation-induced damage of type 2 diabetic erythrocytes.
Clinical and Experimental Pharmacology and Physiology 32 (1–2), 70.
Rongione, A.J., Kusske, A.M., Reber, H., Ashley, S.W., Mc-Fadden, D.W.,
1997. Interleukin-10 reduces circulating levels of serum cytokines in experi-
mental pancreatitis. Journal of Gastrointestinal Surgery 1 (2), 159–166.
Rydzewska, G., Jurkowska, G., Dzieciol, J., Faszczewska, A., Wroblewski, E.,
Gabryelewicz, A., 2001. Does chronic ethanol administration have influence
on pancreatic regulation in the course of caerulein induced acute pancreatitis
in rats. Journal of Physiology and Pharmacology 52 (4), 835–849.
Saluja, A.K., Lu, L., Yamaguchi, Y., Hofbauer, B., Runzi, M., Dawra, R.,
Bhatia, M., Steer, M.L., 1997. A cholecystokinin-releasing factor mediates
ethanol-induced stimulation of rat pancreatic secretion. Journal of Clinical
Investigation 99 (3), 506–512.
Sandhir, R., Gill, K.D., 1999. Hepatoprotective effects of Liv-52 on ethanol
induced liver damage in rats. Indian Journal of Experimental Biology 37,
762–766.
Sarkar, A., Bhaduri, A., 2001. Black tea is a powerful chemopreventor of
reactive oxygen and nitrogen species: comparison with its individual
catechin constituents and green tea. Biochemical Biophysics Research
Communication 284 (1), 173–178.
Schoenberg, M.H., Buchler, M., Pietrzyk, C., Uhl, W., Birk, D., Eisele, S.,
Marzinzig, M., Beger, H.G., 1995. Lipid peroxidation and glutathione
metabolism in chronic pancreatitis. Pancreas 10, 36–43.
Suenobu, N., Shichiri, M., Iwashina, M., Marumo, F., Hirata, Y., 1999.
Natriuretic peptides and nitric oxide induce endothelial apoptosis via
cGMP-dependent mechanism. Arteriosclerosis, Thrombosis, and Vascular
Biology 19, 140–146.
Szuster-Ciesielska, A., Daniluk, J., Kandfer-Szerszen, M., 2001. Alcohol-
related cirrhosis with pancreatitis: the role of oxidative stress in the
progression of the disease. Archivum Immunologiae et Therapiae Experi-
mentalis 49 (2), 139–146.
Takabayashi, F., Harada, N., 1997. Effects of green tea catechins (Poly-
phenon 100) on cerulein-induced acute pancreatitis in rats. Pancreas 14
(3), 276–279.
Takabayashi, F., Harada, N., Hara, Y., 1995. The effects of green tea catechins
(Polyphenon) on DL-ethionine-induced acute pancreatitis. Pancreas 11 (2),
127–131.
Tsai, K., Wang, S.S., Chen, T.S., Kong, C.W., Chang, F.Y., Lee, S.D.,
Lu, F.J., 1998. Oxidative stress: an important phenomenon with
pathogenetic significance in the progression of acute pancreatitis. Gut
42, 850–855.
Tsukamoto, H., Towner, S.J., Yu, G.S.M., French, S.W., 1988. Potentiation of
ethanol-induced pancreatic injury by dietary fat. American Journal of
Pathology 131, 246–257.
Wei, H., Zhang, X., Zhao, J.F, Wang, Z.Y, Bickers, D., Lebwohl, M., 1999.
Scavenging of hydrogen peroxide and inhibition of ultraviolet light-induced
oxidative DNA damage by aqueous extracts from green and black teas. Free
Radical Biology Medicine 26 (11/12), 1427–1435.
Whittakar, P., Hines, F., Robl, M.G., Dunkel, V.C., 1996. Histopathological
evaluation of liver, pancreas, spleen and heart from iron-overloaded
Sprague–Dawley rats. Toxicologic Pathology 24, 558–563.
Wills, E.D., 1987. Evaluation of lipid peroxidation in lipids and biological
membranes. In: Snell, K., Mullock, B. (Eds.), Biochemical Toxicology —
A Practical Approach. IRL Press, Oxford, p. 138.
Wittel, U.A., Bachem, M., Siech, M., 2003. Oxygen radical production
precedes alcohol-induced acute pancreatitis in rats. Pancreas 26 (4),
E74–E80.
Wu, X.N., 2000. Current concept of pathogenesis of severe acute pancreatitis.
World Journal of Gastroenterology 6 (1), 32–36.




















