
Aproximación multivariada a la dinámica del ensamblaje algal epifítico en un sistema de planos...
13
Aproximación multivariada a la dinámica del ensamblaje algal epifítico en un sistema de planos inundables tropical Multivariate approach to dynamics of epiphytic algal assemblage in a tropical system flood plains Yimmy Montoya-Moreno * , Néstor Aguirre * Resumen Objetivo: Identificar la dinámica del ensamble algal epifítico a través del pulso de inundación durante tres ciclos hidrológicos en una planicie inundable tropical. Materiales y métodos: En diferentes fases del pulso de inundación entre los años 2006 y 2009 se realizaron nueve muestreos donde se evaluó la variación espacial y temporal de los ensambles de algas epifíticas aso- ciadas con raíces de macrófitas en 14 sitios de muestreo, 7 zonas con condiciones lénticas y 7 con condicio- nes lóticas del sistema de planos inundables de Ayapel. Se tomaron muestras de raíces que se fijaron en lugol, en las que se observó el ensamble algal epifítico, lo que permitió hacer el estudio multivariado de su dinámica. Resultados: La similaridad florística entre los ensamblajes ficoperifíticos fue baja y denota especificidad de las especies que habitan cada uno de estos compartimentos del sistema de planos inundables. En ambos sistemas se observó una ubicación mayoritaria de especies alrededor del amonio. Las pocas especies o géne- ros que se repiten entre ambos ecosistemas se ordenan con respecto a condiciones diferentes, lo que puede estar asociado con el grado de adaptación de los organismos a las condiciones ambientales. Conclusión: El pulso de inundación en su fase de expansión no alcanza a homogenizar los ensamblajes ficoperifíticos, situación que sucede también en la fase de contracción, razón por la cual cada subsistema responde de una manera particular a las características hidrológicas, respuesta mediada por su resiliencia. Se hipotetiza que al presentarse un tiempo de retención hidráulico bajo en la ciénaga de Ayapel este factor redu- ce el efecto del pulso de inundación y debido a la morfometría variada del sistema, cortocircuitos hidráulicos y aporte de tributarios con características diferenciales, estas condiciones reflejan situaciones particulares para el ensamblaje, lo que favorece la riqueza alta de especies encontradas. El ficoperifiton mostró cambios en su estructura en relación con el pulso de inundación. 1) Aguas bajas. Aumento del efecto de cada sistema (caños y ciénagas) sobre el ensamble debido a la reducción del caudal del río y caños (efecto del pulso fase de desecación). Ello determinó que las estructuras de los ensambles epifíticos fueran diferentes de un sistema a otro. 2) Aguas bajas en ascenso. Reducción del efecto de cada sistema sobre el ensamble e incremento del efecto del pulso de inundación. 3) Aguas altas. Uniformidad de las condiciones fisicoquímicas en los caños y ciénagas por la entrada de agua desde el río. Similitud de la composición específica del ficoperifiton en los diferentes ambientes. 4) Aguas altas en descenso. Reducción de la uniformidad en las condiciones fisicoquímicas del agua por la disminución del caudal procedente del río. Reducción en la similitud de la composición especí- fica de los ensambles de diferentes ambientes. Palabras clave: Epifiton; Perifiton; Ciénaga; Colombia. Abstract Objective: Identify dynamics of periphytic algal assembly through flood pulse during three hydrological cycles on a tropical flood plain. Materials and methods: In different phases of flood pulse between 2006 and 2009, nine samplings were carried out which evaluated the spatial and temporal variation of epiphytic algal assemblage associated with macrophytes roots in 14 sites of sampling, 7 zones with lentic and 7 lotic conditions of the Ayapel flood plain were taken at different stages of the flood pulse between 2006 and 2009. Macrophytes roots were taken in lugol, in which it was observed epiphytic algal assemblage, what allowed the multivariate study of their dynamic. Results: Similarity floristic between the algae assemblages was low and denoted specificity of species that inhabit each of these compartments of the system. A majority location of species around the ammonium was observed in both systems. The few species or genera that are repeated between two ecosystems are ordered with respect to different conditions, which may be associated with the degree of adaptation of organisms to environmental conditions. © Investigación, Biodiversidad y Desarrollo 2012; 31 (2): 83-95 Ciencias biológicas y medio ambiente * Grupo Geolimna, Facultad de Ingeniería, Universidad de Antioquia, Medellín, Colombia. e-mail: [email protected] [email protected] Fecha de recibido: Abril 9, 2012 Fecha de aprobación: Julio 13, 2012
Transcript of Aproximación multivariada a la dinámica del ensamblaje algal epifítico en un sistema de planos...

83
Aproximación multivariada a la dinámica del ensamblaje algal epifítico en unsistema de planos inundables tropical
Multivariate approach to dynamics of epiphytic algal assemblage in atropical system flood plains
Yimmy Montoya-Moreno*, Néstor Aguirre*
Resumen
Objetivo: Identificar la dinámica del ensamble algal epifítico a través del pulso de inundación durante tresciclos hidrológicos en una planicie inundable tropical.Materiales y métodos: En diferentes fases del pulso de inundación entre los años 2006 y 2009 se realizaronnueve muestreos donde se evaluó la variación espacial y temporal de los ensambles de algas epifíticas aso-ciadas con raíces de macrófitas en 14 sitios de muestreo, 7 zonas con condiciones lénticas y 7 con condicio-nes lóticas del sistema de planos inundables de Ayapel. Se tomaron muestras de raíces que se fijaron enlugol, en las que se observó el ensamble algal epifítico, lo que permitió hacer el estudio multivariado de sudinámica.Resultados: La similaridad florística entre los ensamblajes ficoperifíticos fue baja y denota especificidad delas especies que habitan cada uno de estos compartimentos del sistema de planos inundables. En ambossistemas se observó una ubicación mayoritaria de especies alrededor del amonio. Las pocas especies o géne-ros que se repiten entre ambos ecosistemas se ordenan con respecto a condiciones diferentes, lo que puedeestar asociado con el grado de adaptación de los organismos a las condiciones ambientales.Conclusión: El pulso de inundación en su fase de expansión no alcanza a homogenizar los ensamblajesficoperifíticos, situación que sucede también en la fase de contracción, razón por la cual cada subsistemaresponde de una manera particular a las características hidrológicas, respuesta mediada por su resiliencia. Sehipotetiza que al presentarse un tiempo de retención hidráulico bajo en la ciénaga de Ayapel este factor redu-ce el efecto del pulso de inundación y debido a la morfometría variada del sistema, cortocircuitos hidráulicos yaporte de tributarios con características diferenciales, estas condiciones reflejan situaciones particulares parael ensamblaje, lo que favorece la riqueza alta de especies encontradas. El ficoperifiton mostró cambios en suestructura en relación con el pulso de inundación. 1) Aguas bajas. Aumento del efecto de cada sistema (cañosy ciénagas) sobre el ensamble debido a la reducción del caudal del río y caños (efecto del pulso fase dedesecación). Ello determinó que las estructuras de los ensambles epifíticos fueran diferentes de un sistema aotro. 2) Aguas bajas en ascenso. Reducción del efecto de cada sistema sobre el ensamble e incremento delefecto del pulso de inundación. 3) Aguas altas. Uniformidad de las condiciones fisicoquímicas en los caños yciénagas por la entrada de agua desde el río. Similitud de la composición específica del ficoperifiton en losdiferentes ambientes. 4) Aguas altas en descenso. Reducción de la uniformidad en las condiciones fisicoquímicasdel agua por la disminución del caudal procedente del río. Reducción en la similitud de la composición especí-fica de los ensambles de diferentes ambientes.
Palabras clave: Epifiton; Perifiton; Ciénaga; Colombia.
Abstract
Objective: Identify dynamics of periphytic algal assembly through flood pulse during three hydrological cycleson a tropical flood plain.Materials and methods: In different phases of flood pulse between 2006 and 2009, nine samplings werecarried out which evaluated the spatial and temporal variation of epiphytic algal assemblage associated withmacrophytes roots in 14 sites of sampling, 7 zones with lentic and 7 lotic conditions of the Ayapel flood plainwere taken at different stages of the flood pulse between 2006 and 2009. Macrophytes roots were taken inlugol, in which it was observed epiphytic algal assemblage, what allowed the multivariate study of their dynamic.Results: Similarity floristic between the algae assemblages was low and denoted specificity of species thatinhabit each of these compartments of the system. A majority location of species around the ammonium wasobserved in both systems. The few species or genera that are repeated between two ecosystems are orderedwith respect to different conditions, which may be associated with the degree of adaptation of organisms toenvironmental conditions.
© Investigación, Biodiversidad y Desarrollo 2012; 31 (2): 83-95
Ciencias biológicas y medio ambiente
* Grupo Geolimna, Facultad de Ingeniería, Universidad de Antioquia, Medellín, Colombia.e-mail: [email protected] [email protected] de recibido: Abril 9, 2012 Fecha de aprobación: Julio 13, 2012

84
Conclusion: The flood pulse in expansion phase fails tostandardize epiphytic assemblages, situation which happensalso in contraction phase, reason by which each subsystemresponds in a particular way to the hydrological characteristics,this response its mediated by their resilience. Is hypothesizedto occur low hydraulic retention time Ayapel swamp this factorreduces the effect of flood pulse and because morphometryvaried system, hydraulic short circuits and differentialcharacteristics in effluents, these conditions reflect particularsituations for the assemblage, which favors the high richnessof species founded. Phycoperiphyton showed changes in itsstructure in relation to the flood pulse. 1) Shallow waters.Increase the effect of each system (streams and swamps) onthe assemblage due to the reduction of the flow of the riverand streams (flood pulse in desiccation phase). Thisdetermined the structures of epiphytes assemblages weredifferent from a system to another. (2) Low water rising.Reduction of the effect of each system on the assembly andincrease the effect of flood pulse. (3) High water. Uniformity inthe physico-chemical conditions in the streams and swampsby the entry of water from the river. Similarity of the specificcomposit ion of the phycoperiphyton in the dif ferentenvironments. (4) High water in descent. Reduction ofuniformity in physicochemical conditions of water flowdecreased from the river. Reduction in the similarity of thespecies composit ion of assemblages f rom dif ferentenvironments.
Keywords: Epifiton; Periphyton; Swamp; Colombia.
Introducción
Las algas epifíticas hacen parte de los organismosque sostienen la cadena alimenticia, su estudio esprioritario y en los ecosistemas poco profundos,alcanzan mayor importancia en los balances demateria y energía que fluyen a través de este com-partimiento (Goldsborough & Robinson, 1996).Estos organismos son indicadores biológicos delnivel trófico del agua, los cuales muestran varia-ción con los cambios en la calidad del agua y ladinámica del sistema (Junk et al., 1989).
En los ambientes de planos inundables existen po-cas investigaciones sobre las algas perifíticas (Ri-vera-Rondón et al., 2010; Álvarez, 2008; Martínezet al., 2008; Montoya-Moreno & Aguirre, 2008;2009; Montoya-Moreno et al., 2011; Abuhatab &Asprilla, 2006; Báez-Polo y Hernández-Jiménez,2001; Maturana et al., 2001).
A pesar de la gran abundancia de las áreas de
inundación a escala mundial, Goldsborough &Robinson (1996) consideran que las investiga-ciones ficológicas se han concentrado en otrossistemas como lagos y ríos, de modo que la in-formación sobre los ensamblajes algales en losambientes de planos de inundación, en formageneral, son bastante fragmentadas, particular-mente en las áreas tropicales y subárticas.
Los sistemas de planos inundables están someti-dos a variaciones altas del nivel de la columna deagua, como respuesta a las fluctuaciones de las pre-cipitaciones propias del ciclo hidrológico. Este con-junto de cambios debe ser internalizado por losorganismos (Montoya-Moreno & Aguirre, en pren-sa). Los humedales conectados a un río han sidocatalogados bajo la denominación de llanuras deinundación y están sujetos a fluctuaciones impor-tantes en los niveles y a cambios pronunciados desus fases terrestre y acuática (sistemas pulsantes).Estos sistemas mantienen una conexión permanentecon el río y poseen una zona litoral móvil que seconfigura como zona de transición acuático terres-tre (ATTZ) (Junk et al., 1989). El mismo autor con-sidera que el tamaño de la ATTZ, su estado tróficoy el tipo de pulso, determinan el conjunto de lascomunidades de plantas y animales, la producciónprimaria y secundaria y los ciclos de niveles en elhumedal.
Los sistemas de planos inundables en su faseextrema de expansión y contracción y sus fasesintermedias (ascenso y descenso), generan va-riaciones en las condiciones fisicoquímicas ymorfométricas (Montoya-Moreno et al., 2011),que afectan la estructura y dinámica de losensamblages algales epifíticos, por lo que el ob-jetivo de este trabajo es realizar una aproxima-ción multivariada a la dinámica de las algasepifíticas en el sistema de planos inundables deAyapel, la cual fue evaluada durante tres cicloshidrológicos consecutivos.
Investigación, Biodiversidad y Desarrollo 2012; 31 (2): 83-95
85
Materiales y métodos
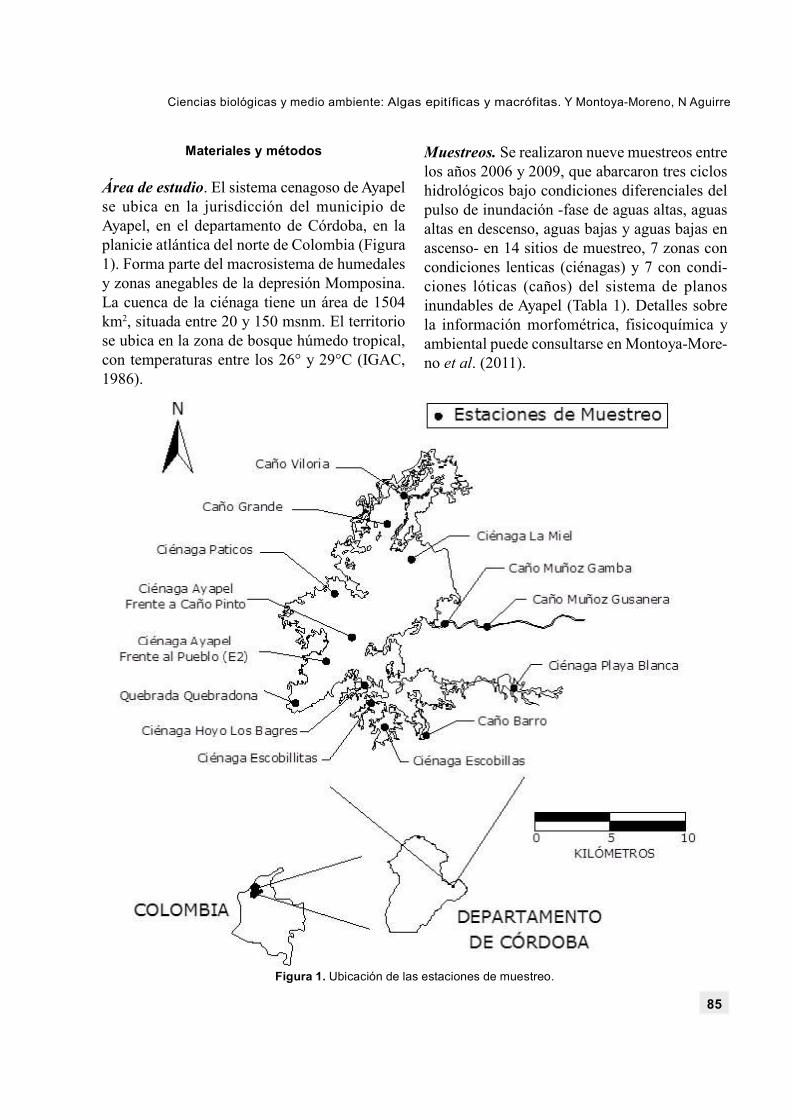
Área de estudio. El sistema cenagoso de Ayapelse ubica en la jurisdicción del municipio deAyapel, en el departamento de Córdoba, en laplanicie atlántica del norte de Colombia (Figura1). Forma parte del macrosistema de humedalesy zonas anegables de la depresión Momposina.La cuenca de la ciénaga tiene un área de 1504km2, situada entre 20 y 150 msnm. El territoriose ubica en la zona de bosque húmedo tropical,con temperaturas entre los 26° y 29°C (IGAC,1986).
Muestreos. Se realizaron nueve muestreos entrelos años 2006 y 2009, que abarcaron tres cicloshidrológicos bajo condiciones diferenciales delpulso de inundación -fase de aguas altas, aguasaltas en descenso, aguas bajas y aguas bajas enascenso- en 14 sitios de muestreo, 7 zonas concondiciones lenticas (ciénagas) y 7 con condi-ciones lóticas (caños) del sistema de planosinundables de Ayapel (Tabla 1). Detalles sobrela información morfométrica, fisicoquímica yambiental puede consultarse en Montoya-More-no et al. (2011).
Ciencias biológicas y medio ambiente: Algas epitíficas y macrófitas. Y Montoya-Moreno, N Aguirre
Figura 1. Ubicación de las estaciones de muestreo.

86
En los parches de macrofitas se determinó cuá-les eran las especies más abundantes por inspec-ción visual y del borde del parche, se tomaroncon pinzas tres muestras de raíces por cada es-pecie de planta acuática dominante. En el sitiode recolección de las raíces se midió la transpa-rencia y la temperatura del agua, el oxígeno di-suelto y su porcentaje de saturación, laconductividad eléctrica, el pH y la profundidadtotal empleándose una sonda multiparamétricaIQ. Se tomaron muestras de 3 litros de agua parala determinación en el laboratorio de la concen-tración de nutrientes, como nitrógeno amoniacal,nitratos, nitritos, fósforo soluble reactivo (FSR),alcalinidad, sulfatos, los sólidos totales disuel-tos (STD) y los sólidos suspendidos totales(SST), evaluados según APHA (2005).
Las muestras se depositaron en frascos de plás-tico de 100 ml, a los que se adicionó agua de laciénaga previamente filtrada y después se fija-ron con solución de lugol, hasta obtener una con-centración final del 4%. Las raíces recolectadasse llevaron al laboratorio para la determinaciónde las especies del ficoperifiton con base en lite-ratura especilalizada (Montoya-Moreno et al.,2011).
El conteo se realizó a partir del uso de cámaras deSegdwick-Rafter de 1 ml de capacidad, se empleó
un microscópio olimpus DHK con el objetivo de40X en 50 campos aleatorios, los cuales se deter-minaron al tener en cuenta curvas de saturaciónde especies. Para calcular la densidad delfitoperifiton se utilizó la fórmula de Ross (1979).
La composición, abundancia y diversidad de lacomunidad se evaluó mediante curvas de impor-tancia y se usaron los índices comunitarios. Paraevaluar la relación de las algas perifíticas conlas variables ambientales se utilizó la correla-ción por rangos de Spearman (rS). Para evaluarel efecto de las variables físicas y químicas so-bre la composición, abundancia y diversidad delensamblaje, se realizó un análisis de correlaciónmúltiple y un análisis de componentes principa-les. Además, se efectuó un análisis de regresiónlineal simple para evaluar el efecto de las dife-rentes variables físicas, químicas y ambientalessobre los grupos algales componentes del ensam-blaje. Se empleó un análisis de correspondenciacanónica (ACC) para identificar las tendenciasde variación entre ciénagas y caños, mediante elprograma Canoco versión 4.5.
Resultados
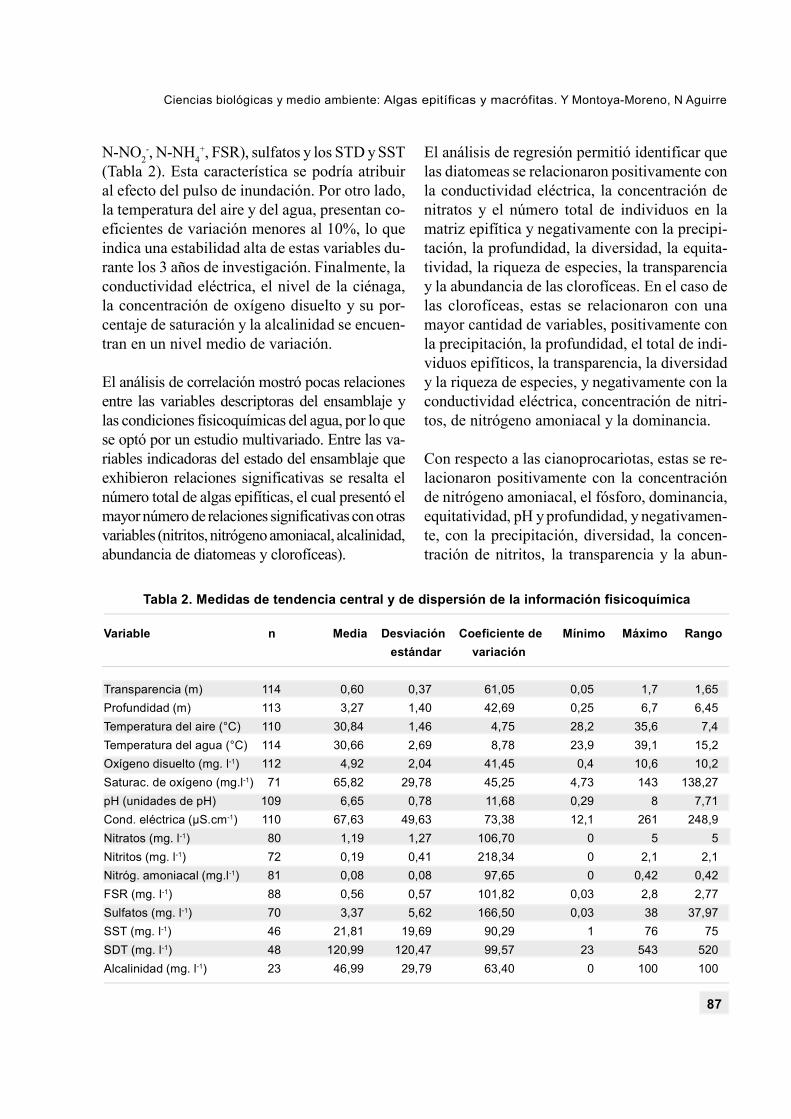
Se presentan tres tendencias, las variables queexhiben coeficientes de variación altos, talescomo la transparencia, los nutrientes (N-NO3
-,
Tabla 1. Fechas de muestreo y condición hidrológica
Muestreo Característica hidrológica
Mayo/2006 Aguas en ascensoJulio/2006 Aguas en ascensoSeptiembre/2006 Aguas altasDiciembre/2006 Aguas altas en descensoMarzo/2007 Aguas bajasAgosto2007 Aguas en ascensoEnero/2008 Aguas altas en descensoSeptiembre/2008 Aguas altasEnero/2009 Aguas altas en descenso
Investigación, Biodiversidad y Desarrollo 2012; 31 (2): 83-95
87
N-NO2-, N-NH4
+, FSR), sulfatos y los STD y SST(Tabla 2). Esta característica se podría atribuiral efecto del pulso de inundación. Por otro lado,la temperatura del aire y del agua, presentan co-eficientes de variación menores al 10%, lo queindica una estabilidad alta de estas variables du-rante los 3 años de investigación. Finalmente, laconductividad eléctrica, el nivel de la ciénaga,la concentración de oxígeno disuelto y su por-centaje de saturación y la alcalinidad se encuen-tran en un nivel medio de variación.
El análisis de correlación mostró pocas relacionesentre las variables descriptoras del ensamblaje ylas condiciones fisicoquímicas del agua, por lo quese optó por un estudio multivariado. Entre las va-riables indicadoras del estado del ensamblaje queexhibieron relaciones significativas se resalta elnúmero total de algas epifíticas, el cual presentó elmayor número de relaciones significativas con otrasvariables (nitritos, nitrógeno amoniacal, alcalinidad,abundancia de diatomeas y clorofíceas).
Tabla 2. Medidas de tendencia central y de dispersión de la información fisicoquímica
Variable n Media Desviación Coeficiente de Mínimo Máximo Rango estándar variación
Transparencia (m) 114 0,60 0,37 61,05 0,05 1,7 1,65Profundidad (m) 113 3,27 1,40 42,69 0,25 6,7 6,45Temperatura del aire (°C) 110 30,84 1,46 4,75 28,2 35,6 7,4Temperatura del agua (°C) 114 30,66 2,69 8,78 23,9 39,1 15,2Oxígeno disuelto (mg. l-1) 112 4,92 2,04 41,45 0,4 10,6 10,2Saturac. de oxígeno (mg.l-1) 71 65,82 29,78 45,25 4,73 143 138,27pH (unidades de pH) 109 6,65 0,78 11,68 0,29 8 7,71Cond. eléctrica (µS.cm-1) 110 67,63 49,63 73,38 12,1 261 248,9Nitratos (mg. l-1) 80 1,19 1,27 106,70 0 5 5Nitritos (mg. l-1) 72 0,19 0,41 218,34 0 2,1 2,1Nitróg. amoniacal (mg.l-1) 81 0,08 0,08 97,65 0 0,42 0,42FSR (mg. l-1) 88 0,56 0,57 101,82 0,03 2,8 2,77Sulfatos (mg. l-1) 70 3,37 5,62 166,50 0,03 38 37,97SST (mg. l-1) 46 21,81 19,69 90,29 1 76 75SDT (mg. l-1) 48 120,99 120,47 99,57 23 543 520Alcalinidad (mg. l-1) 23 46,99 29,79 63,40 0 100 100
El análisis de regresión permitió identificar quelas diatomeas se relacionaron positivamente conla conductividad eléctrica, la concentración denitratos y el número total de individuos en lamatriz epifítica y negativamente con la precipi-tación, la profundidad, la diversidad, la equita-tividad, la riqueza de especies, la transparenciay la abundancia de las clorofíceas. En el caso delas clorofíceas, estas se relacionaron con unamayor cantidad de variables, positivamente conla precipitación, la profundidad, el total de indi-viduos epifíticos, la transparencia, la diversidady la riqueza de especies, y negativamente con laconductividad eléctrica, concentración de nitri-tos, de nitrógeno amoniacal y la dominancia.
Con respecto a las cianoprocariotas, estas se re-lacionaron positivamente con la concentraciónde nitrógeno amoniacal, el fósforo, dominancia,equitatividad, pH y profundidad, y negativamen-te, con la precipitación, diversidad, la concen-tración de nitritos, la transparencia y la abun-
Ciencias biológicas y medio ambiente: Algas epitíficas y macrófitas. Y Montoya-Moreno, N Aguirre

88
dancia de clorofíceas. Las crisofíceas presenta-ron regresión positiva con la abundancia de lasdiatomeas y negativa con la conductividad eléc-trica, fósforo, profundidad, riqueza y la diversi-dad. Las dinofíceas presentaron regresión posi-tiva con la concentración de sulfatos en el aguay negativa con la diversidad y el total de indivi-duos. Finalmente, la euglenofíceas exhibieronregresión lineal positiva con la concentración desulfatos y negativa con la diversidad.
En el análisis de componentes principales (Figura
2), el primer eje canónico explicó el 29% de lavarianza y resultó positivamente correlacionado conla diversidad de diatomeas y la concentración denitrógeno amoniacal y negativamente con los índi-ces de diversidad, equitatividad y la abundancia declorofíceas. El segundo eje, que explicó el 18,9%de la varianza, estuvo relacionado positivamentecon la profundidad y la precipitación y negativa-mente con la concentración de nitritos, lo que indi-ca que estas variables determinan el principalgradiente al que respondió la distribución de lasespecies (Tabla 3).
Figura 2. Organización de las variables ambientales y los grupos ficoperifíticos predominantes.
Tabla 3. Correlaciones de las variablescon los ejes
Variables Eje 1 Eje 2
Profundidad -0,04 0,53NO2 -0,17 -0,47NH4 0,39 0,04Precipitación -0,25 0,40S (riqueza de especies) -0,49 -0,05D (dominancia) 0,08 -0,41H (diversidad) -0,40 -0,17Bacillariophyta 0,34 -0,12Cianophyta 0,21 0,28Chlorophyta -0,41 0,05Euglenophyta 0,14 -0,21
La ubicación de los grupos algales en el plano deordenación permite diferenciar algunas tendencias,porque las diatomeas se ubicaron cerca de las con-centraciones de nutrientes (N-NH4
+); las clorofíceasse localizaron en sentido opuesto a éstas y conmayor afinidad hacia la pluviosidad y la riqueza deespecies; las cianoprocariotas presentaron afinidadcon la profundidad de la columna de agua y con laconcentración de nitrógeno en forma amoniacal.Finalmente, las euglenofíceas se ubican hacia elcentro de la figura, en la cual registra relación conla dominancia.
Con el fin de depurar la información se realizó unACC individual para las ciénagas y los caños res-
Investigación, Biodiversidad y Desarrollo 2012; 31 (2): 83-95
89
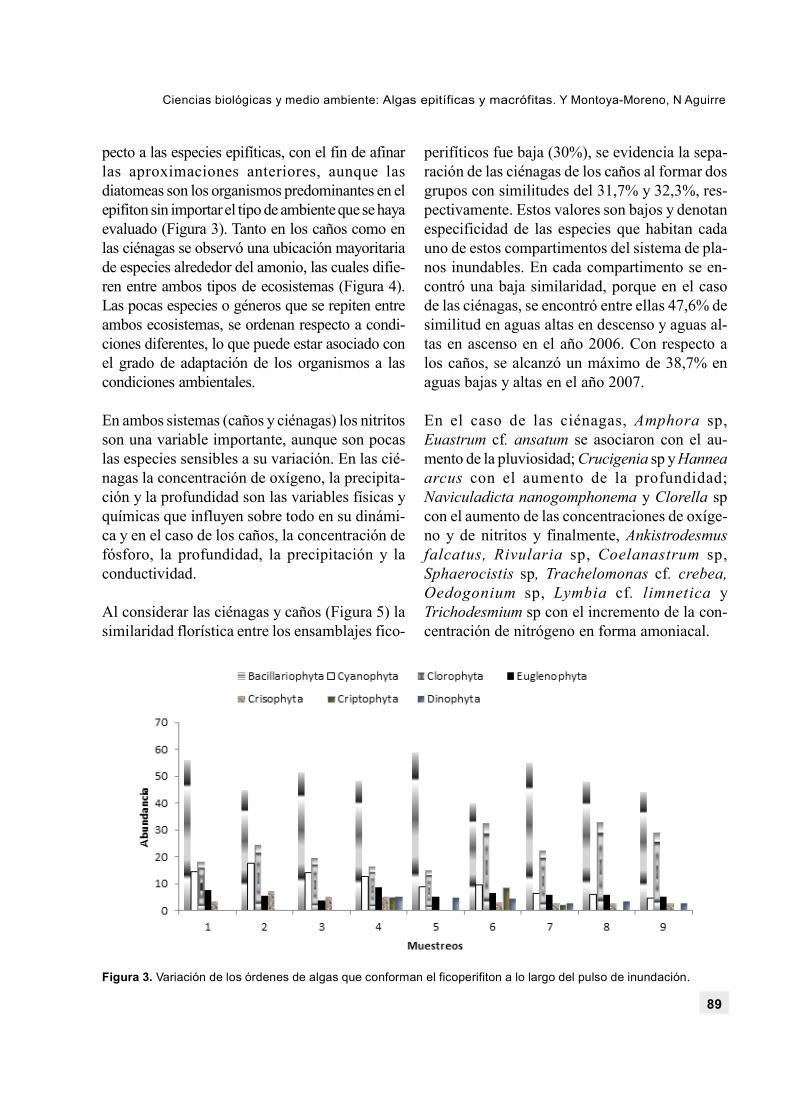
pecto a las especies epifíticas, con el fin de afinarlas aproximaciones anteriores, aunque lasdiatomeas son los organismos predominantes en elepifiton sin importar el tipo de ambiente que se hayaevaluado (Figura 3). Tanto en los caños como enlas ciénagas se observó una ubicación mayoritariade especies alrededor del amonio, las cuales difie-ren entre ambos tipos de ecosistemas (Figura 4).Las pocas especies o géneros que se repiten entreambos ecosistemas, se ordenan respecto a condi-ciones diferentes, lo que puede estar asociado conel grado de adaptación de los organismos a lascondiciones ambientales.
En ambos sistemas (caños y ciénagas) los nitritosson una variable importante, aunque son pocaslas especies sensibles a su variación. En las cié-nagas la concentración de oxígeno, la precipita-ción y la profundidad son las variables físicas yquímicas que influyen sobre todo en su dinámi-ca y en el caso de los caños, la concentración defósforo, la profundidad, la precipitación y laconductividad.
Al considerar las ciénagas y caños (Figura 5) lasimilaridad florística entre los ensamblajes fico-
perifíticos fue baja (30%), se evidencia la sepa-ración de las ciénagas de los caños al formar dosgrupos con similitudes del 31,7% y 32,3%, res-pectivamente. Estos valores son bajos y denotanespecificidad de las especies que habitan cadauno de estos compartimentos del sistema de pla-nos inundables. En cada compartimento se en-contró una baja similaridad, porque en el casode las ciénagas, se encontró entre ellas 47,6% desimilitud en aguas altas en descenso y aguas al-tas en ascenso en el año 2006. Con respecto alos caños, se alcanzó un máximo de 38,7% enaguas bajas y altas en el año 2007.
En el caso de las ciénagas, Amphora sp,Euastrum cf. ansatum se asociaron con el au-mento de la pluviosidad; Crucigenia sp y Hanneaarcus con el aumento de la profundidad;Naviculadicta nanogomphonema y Clorella spcon el aumento de las concentraciones de oxíge-no y de nitritos y finalmente, Ankistrodesmusfalcatus, Rivularia sp, Coelanastrum sp,Sphaerocistis sp, Trachelomonas cf. crebea,Oedogonium sp, Lymbia cf. limnetica yTrichodesmium sp con el incremento de la con-centración de nitrógeno en forma amoniacal.
Figura 3. Variación de los órdenes de algas que conforman el ficoperifiton a lo largo del pulso de inundación.
Ciencias biológicas y medio ambiente: Algas epitíficas y macrófitas. Y Montoya-Moreno, N Aguirre

90
Figura 4. Organización de las variables ambientales y losgrupos ficoperifíticos predominantes en las ciénagas (a) y loscaños (b).
Con respecto a los caños, Achanthes inflata yGomphonema clavatum se asociaron con el incremen-to del fósforo; Synedra montana se relacionó con elincremento de la conductividad eléctrica; Aulacoseira
granulata y Hannea arcus se relacionaroncon el aumento de los nitritos yGomphonema parvulum, Brachysiramicrocephala, Eolimna subminuscula,Nitzchia palea y Neidium aff. affine con elincremento de la concentración de nitróge-no en forma amoniacal.
Discusión
En la época de estiaje, las concentracionesde nutrientes son mayores debido a los efec-tos de la resuspensión y de la mineralizaciónde la materia orgánica, lo que favorece eldesarrollo de las diatomeas, la disminuciónde la densidad algal perifítica y la reducciónde la competencia intraespecífica y un au-mento de la competencia interespecífica. Eneste sentido, sobre el primer aspecto, lasdiatomeas en el ACP y en el ACC se ubicancerca de la concentración de nutrientes. Ri-vera-Rondón y Díaz-Quiróz (2004) encon-traron que dentro de las variables químicas,el fósforo reactivo soluble influyó sobre lacomposición de especies de diatomeas cuan-do registró concentraciones altas.
En aguas bajas varios sectores del sistemase aislan por barras de sedimento, que se-gregan el sistema en varios compartimentos,los cuales desarrollan estructuras del ensam-blaje diferentes, las cuales, en aguas altaspermiten la integración de especies y lasostenibilidad de una microdiversidad alta.Thomaz et al. (2007) consideran que enaguas bajas las comunidades biológicas decada hábitat de los planos de inundación (la-gunas, aguas detenidas, ríos) desarrollan dis-tintas trayectorias temporales debido a losefectos de las fuerzas locales (por ejemploun depredador eficiente que atrapa en lagu-nas pero no en otros ambientes). Estos sis-temas fragmentados son susceptibles al tra-bajo del viento, el cual estimula la mezcla
Investigación, Biodiversidad y Desarrollo 2012; 31 (2): 83-95

91
nocturna, lo que genera un aumento en la diná-mica del cuerpo de agua.
La ubicación de las clorofíceas en un extremoopuesto del ACP a las diatomeas podría confir-mar la sospecha de que ambos grupos algalesson indicadores de condiciones contrastastes enla ciénaga de Ayapel, aunque su abundancia seencuentra ligada entre si. Teniendo en cuenta quelas demidíaceas son el grupo predominante enlas clorofíceas, se puede considerar la investiga-ción de Rodrigues y Bicudo (2001) en la que re-lacionaron una gran riqueza de desmidíaceasperifíticas con la presencia de estanques demacrófitas, los cuales alcanzan su mayor densi-dad en el sistema de Ayapel en la fase de aguasaltas, cuando el aumento en las precipitacionesinduce la movilidad de los bancos de macrófitas.Según estos autores muchas clorofíceasplanctónicas poseen un desarrollo derivado delhábitat perifítico, por lo que en aguas bajas sefavorecería la salida de estas algas hacia el
plancton debido al incremento en la energía tur-bulenta del sistema. En este sentido, lasdiatomeas también se ven beneficiadas por losbancos de macrofitas, debido a que poseen di-versas formas de fijación al sustrato (Round,1990).
La abundancia de las diatomeas se relaciona deuna manera altamente competitiva con la abun-dancia de las clorofíceas y con la dinámica dife-rencial de cada ambiente. La disminución de larelación Si/P puede favorecer el reemplazo delas diatomeas por clorofitas y esto puede llevara un cambio en la relación N/P (Margalef, 1983).Tilman et al. (1996) demostraron que lasdiatomeas dominan si la razón N:P y la tempera-tura son bajas, las algas verdes en la medida enque la temperatura y la razón N:P presentan va-lores intermedios y las cianobacterias cuando haytemperaturas altas y bajas razones N:P. En el casodel sistema de Ayapel las temperaturas son altasdurante todo el año, no obstante las diatomeas
Figura 5. Dendrograma de afinidad entre las especies ficoperifíticas en las ciénagas y los caños a través del pulso deinundación.
HW: aguas altas, LW: aguas bajas, Des: descenso, Asc: ascenso, 06: año 2006, 07: año 2007, 08: año 2008, 09: año2009, ci: ciénaga, ca: caño.
Ciencias biológicas y medio ambiente: Algas epitíficas y macrófitas. Y Montoya-Moreno, N Aguirre

92
presentan abundancias mayores al 50% y en elperíodo de aguas bajas (mayores temperaturas),las cianofíceas alcanzan a incrementar sus abun-dancias, sin llegar a ser dominantes en elperifiton.
Las cianoprocariotas de acuerdo con la ordena-ción canónica presentaron relación con la tem-peratura del agua, la profundidad de la columnade agua y la concentración de nitrógeno en for-ma amoniacal. No obstante, esta tendencia pre-senta mayor importancia a nivel local, en dondezonas como la ciénaga de Ayapel sector La Mielen aguas bajas, presenta blooms decianoprocariotas planctónicas (Cylindrosper-mopsis raciborskii), las cuales se encuentrandentro de la matriz perifítica. La prueba de re-gresión lineal no mostró relación significativaentre estas variables y la abundancia de lascianoprocariotas.
Las euglenofíceas se ubicaron en el ACP cercadel pH. Se ha considerado que se presentan enaguas ricas en partículas y compuestos orgáni-cos (Rodrigues et al., 2008), lo que permite ex-plicar esta relación. Las euglenofíceas presenta-ron relación positiva con la concentración deoxígeno disuelto y negativa con el número totalde organismos en el ensamblaje epifítico. Peresy Senna (2000) observaron que la aparición dela clase euglenofícea estuvo relacionada con elperiodo seco en sistemas inundables brasileros.
Otro aspecto a tener en cuenta, es la heteroge-neidad de hábitats que se generan en la zona li-toral, los cuales van a ser decisivos para la di-versidad biológica. Como han enfatizadoGoldsborough & Robinson (1996) en ambientesrasos, como en las áreas inundables en general,es difícil de establecer límites de hábitat para laspoblaciones algales una vez que se presenta unagran interacción entre los compartimentos delecosistema. En este tipo de ecotono, la interac-ción epifiton/plancton merece mayor atención,
una vez que puede dar luces sobre los patronesde fluctuación de la estructura del funcionamien-to de las comunidades de algas y consecuente-mente, sobre el metabolismo y flujos de energíaen esos ecosistemas acuáticos (Rodrigues et al.,2003).
El tiempo de retención hidráulico bajo del siste-ma de planos inundables de Ayapel ejerce unalimitación a la fijación sobre los sustratos parael perifiton, por lo que las diatomeas son favore-cidas al presentar estructuras especializadas parala fijación. Además que, son rápidas y eficientescolonizadoras, capaces de ocupar sustratos en elrango de un día a varias semanas (Hoagland etal., 1986). Otro aspecto a tener en cuenta es lafisicoquímica del agua, ya que la dominancia delas diatomeas tanto en el perifiton como en elfitoplancton, esta relacionada con la baja con-centración de nutrientes y el pH ácido (Rivera-Rondón et al., 2010).
El valor bajo de similaridad para cada subsistema(ciénagas y caños) al igual que entre ellos indi-caría que el pulso en su fase de expansión noalcanza a homogenizar los ensamblajesficoperifíticos, situación que sucede también enla fase de contracción, razón por la cual cadasubsistema responde de una manera particular através del pulso, respuesta mediada por sus ca-racterísticas propias principalmente. Se podríahipotetizar que al presentarse un tiempo de re-tención hidráulico bajo en la ciénaga de Ayapeleste factor reduce el efecto del pulso de inunda-ción. Aunque debido a la morfometría variadadel sistema, corto circuitos hidráulicos y aportede tributarios con condiciones diferenciales, es-tas condiciones reflejan unas condiciones parti-culares para el ensamblaje, lo que favorece lariqueza alta de especies encontradas.
El ficoperifiton mostró cambios en su estructuraen relación con el pulso de inundación. De estaforma, se pudieron discriminar cuatro ensambles
Investigación, Biodiversidad y Desarrollo 2012; 31 (2): 83-95

93
característicos de cuatro momentos hidrológicosocurridos en el sistema (Figura 6).
1) Aguas bajas. Aumento del efecto de cada sis-tema (caños y ciénagas) sobre el ensamble debi-do a la reducción del caudal del río y caños (efec-to del pulso fase de desecación). Dinámica par-ticular de las diferentes variables físico-quími-cas en los caños y ciénagas debido al aislamien-to (o desconexión) con el río (fase de deseca-ción). Ello determinó que las estructuras de losensambles epífitos fueran fuertemente diferen-
tes de un cuerpo de agua a otro. Predominan enlas ciénagas las especies, Planothidiumlanceolatum, Encyonema jetlandicum yOscillatoria sp.1 y, en los caños, Navicula aff.cryptocephala, Stenopterobia delicatissima yEunotia naegueli.
2) Aguas bajas en ascenso. Reducción del efec-to de cada sistema sobre el ensamble e incre-mento del efecto del pulso de inundación. En lasciénagas predominaron las especies Ulnarialanceolata, Fragilaria capucina y Lyngbya cf.
Figura 6. Análisis general de las especies ficoperifíticas en las ciénagas y los caños a través de las fases del pulso deinundación.
Ciencias biológicas y medio ambiente: Algas epitíficas y macrófitas. Y Montoya-Moreno, N Aguirre

94
limnetica. En los caños fueron abundantesFragilaria capucina, Achnanthes inflata,Achnanthidium minutissimum, Gomphonemaparvulum y Nitzschia palea.
3) Aguas altas. Uniformidad de las condicionesfisicoquímicas en los caños y ciénagas por laentrada de agua desde el río. Similitud de la com-posición específica del ficoperifiton en los dife-rentes ambientes. Predominan en las ciénagasFragilaria capucina, Eunotia naegueli,Euastrum cf. ansatum, y Lyngbya cf. limneticay, en los caños, Fragilaria capucina, Plano-thidium lanceolatum y Lyngbya cf. limnetica.
4) Aguas altas en descenso. Reducción de launiformidad en las condiciones fisicoquímicasdel agua por la disminución del caudal proce-dente del río. Reducción en la similitud de lacomposición específica de los ensambles de di-ferentes ambientes. Predominaron en las ciéna-gas, las especies Synedra goulardii, Eunotianaegueli, Diadesmis confervaceae, Brachysiracf. manfredii Navicula aff. rostellataStenopterobia delicatissima, Spondylosiumrectangulare y Mougeotia sp. En los caños, fue-ron abundantes Lyngbya cf. limnetica, Fragilariacapucina y Planothidium lanceolatum.
Literatura citada
Abuhatab AY, Asprila S. 2006. Distribución y abundanciade la comunidad fitoperifitica y su relación con lasvariables físicas y químicas en diferentes cuerposde agua en los cerros Las Mojarras, Istmina-Chocó.VII Seminario Colombiano de Limnología, Ibagué.56 pp.
APHA, AWWA, WPCF. 2005. Standard methods for theexamination of water and wastewater. Washington:American Public Health Association (APHA),American Water Works Association (AWWA) andWater Environment Federation (WEF). 1325 pp.
Álvarez JP. 2008. Caracterización limnológica de loscomplejos cenagosos de Arcial-Pooor-Cintura (RíoSan Jorge) y de Bañó-Charco Pescao-PantanoBonito (Río Sinú) Córdoba, Colombia. VIIISeminario Colombiano de Limnología, Cali. 30 pp.
Báez-Polo A, Hernández-Jiménez C. 2001. Composiciónde especies y variación temporal de las comunidadesde perifiton de substratos artificiales en la ciénagade Pajaral, Caribe colombiano. Resumen extendidoen el IX Congreso Latinoamericano sobre Cienciasdel Mar. San Andrés Isla. 6 pp.
Goldsborough LG, Robinson G.G. 1996. Pattern inwetlands. In: Algal ecology, freshwater benthicecosystems. Stevenson RJ, Bothwell ML, Lowe RL,eds. San Diego: Academic Press. p.78-117.
Hoagland KD, Zlotsky A, Peterson CG. 1986. The sourceof algal colonizer on rock substrates in a freshwaterimpoundment. In: Algal biofouling. Evans LV,Hoagland KD, eds. Amsterdam: Elsevier SciencePublishing Company Inc. p. 21-39.
Instituto Geográfico Agustin Codazzi (IGAC). 1986.Estudio general de los suelos de los municipios deAyapel, Buenavista, Planeta Rica y Pueblo Rico(Departamento de Córdoba). Bogotá: IGAC. 282 pp.
Junk WJ, Bayley PB, Sparks RE. 1989. The flood pulseconcept in river floodplain systems. Can Spec PublFish. Aquat Sci. 106: 110-27.
Margalef R. 1983. Limnología. Barcelona: EdicionesOmega, SA. 951 pp.
Martínez LF, Ayarith Y, Moreno Y. 2008. Composición ydistribución de la comunidad fitoperifítica endiferentes ecosistemas lóticos de la cuenca del SanJuan, Chocó-Colombia. VIII Seminario Colombianode Limnología, Cali. 30p.
Maturana MM, Cuesta OJ, Asprilla S. 2001. Variaciónde la estructura numérica de las comunidadesficoperifíticas asociadas a Nymphoideae sp en laciénaga de Plaza Seca, corregimiento de Sanceno,Quibdó-Chocó, Colombia. Trabajo de grado debiología. Universidad Tecnológica del Chocó. 46p.
Montoya-Moreno Y, Aguirre N. 2008. Asociación de algasperifíticas en raíces de macrófitas en una ciénagatropical colombiana. Hidrobiologica. 18 (3): 189-98.
Montoya-Moreno Y, Aguirre N. 2009. Dinámica delperifiton asociado con macrofitas en la ciénaga deEscobillitas y su relación con el pulso de inundación.Revista Institucional Universidad Tecnológica delChoco 28 (2): 196-202.
Montoya-Moreno Y, Aguirre N, González EM. 2011. Elperifiton del sistema cenagoso de Ayapel, Córdoba-Colombia. Medellín: Editorial Universidad deMedellín. 76 Pp.
Peres AC, Senna PAC. 2000. Estudo quantitativo eestatístico do fitoplâncton da lagoa do Diogo emum ciclo hidrológico (1995-1996). In: EstaçãoEcológica de Jataí. Santos JE, Pires JSR, eds. São
Investigación, Biodiversidad y Desarrollo 2012; 31 (2): 83-95

95
Carlos: RiMa. 483-95.Rivera-Rondón CA, Díaz Q. 2004. Grandes taxones de
fitobentos y su relación con la hidrología, física yquímica de pequeños ríos andinos. UniversitasScientiarum. 9: 75-86.
Rivera-Rondón CA, Zapata AM, Pérez D, Morales Y,Ovalle H, Álvarez JP. 2010. Caracterizaciónlimnológica de humedales de la planicie deinundación del río Orinoco (Orinoquía, Colombia).Acta Biol Colomb. 15 (1): 1-16.
Rodrigues L, Bicudo DC. 2001. Similarity amongperiphyton algal communities in lentic-lotic gradientof the upper Paraná river floodplain, Brazil. RevBrasil Bot. 24 (3): 235-48.
Rodrigues L, Bicudo DC, Moschini-Carlos V. 2003. Opapel do perifiton em áreas alagáveis e nos diag-nósticos ambientais. In: Ecología e manejo demacrófitas aquáticas. Thomaz SM, Bini LM, eds.Maringá: Eduem. p. 211-29.
Rodrigues L, Zanon JE, Carapurnala L, Biolo S. 2008.
Perifiton en a Planície Alagável do Rio Paraná:estrutura e processo ambiental. Capítulo 4.Disponible en http://www.peld.uem.br/Relat2008/pdf/Capitulo04.pdf. Fecha de consulta: 17 de juliode 2010.
Roldán PG, Ramírez JJ. 2008. Fundamentos de limnologíaneotropical. 2a ed. Medellín: Universidad deAntioquia. 440 pp.
Round, FE, Crawford RM, Mann DG. 1990. The diatoms,biology and morphology of the genera. Cambridge:Cambridge University Press. 747 pp.
Ross J. 1979. Prácticas de ecología. Barcelona: Omega1.250 pp.
Thomaz SM, Bini LM, Bozelli RL. 2007. Floods increasesimilarity among aquatic habitats in river-floodplainsystems. Hydrobiologia. 579 (1): 1-13.
Tilman D, Wedin D, Knops J. 1996. Productivity andsustainability influenced by biodiversity in grasslandecosystems. Nature. 379: 718 20.
Ciencias biológicas y medio ambiente: Algas epitíficas y macrófitas. Y Montoya-Moreno, N Aguirre




















