
Apratylenchus vietnamensis gen. n., sp. n. and A. binhi gen. n., sp. n., sedentary Pratylenchidae...
17
Nematology, 2009, Vol. 11(4), 565-581 Apratylenchus vietnamensis gen. n., sp. n. and A. binhi gen. n., sp. n., sedentary Pratylenchidae (Nematoda: Tylenchida) from coffee in Vietnam, with proposal of Apratylenchinae subfam. n. Phap Q. TRINH 1,2,3 , Lieven WAEYENBERGE 3,4 , Chau N. NGUYEN 1 , James G. BALDWIN 4 , Gerrit KARSSEN 5,6 and Maurice MOENS 2,3,∗ 1 Institute of Ecology and Biological Resources, 18 Hoang Quoc Viet Road, Hanoi, Vietnam 2 Laboratory for Agrozoology, Ghent University, Coupure links 653, 9000 Ghent, Belgium 3 Institute for Agriculture and Fisheries Research, Burg. Van Gansberghelaan 96, 9820 Merelbeke, Belgium 4 Department of Nematology, University of California, Riverside, CA 92521, USA 5 Plant Protection Service, Box 9102, 6700 HC Wageningen, The Netherlands 6 Department of Biology, Ghent University, K.L. Ledeganckstraat 35, 9000 Ghent, Belgium Received: 10 July 2007; revised: 17 October 2008 Accepted for publication: 20 October 2008 Summary – Apratylenchinae subfam. n. is proposed within the Pratylenchidae to include Apratylenchus gen. n. and two new species, A. vietnamensis gen. n., sp. n. and A. binhi gen. n., sp. n., from coffee plantations in Vietnam. Apratylenchus gen. n. is distinguished by the presence of transverse rows of cuticular tubercles on the annuli and by sexual dimorphism in which the males are stout and cylindrical, but mature sedentary females are club-shaped. The sedentary females are broadest in the posterior third although narrowing posterior to the vulva (V 85%). The female cephalic region is low and dome-shaped with all labial sectors fused to one another and the oral disc and the pharyngeal glands overlap the intestine ventrally. Males have a weak stylet with reduced knobs and a conical tail with a blunt terminus, arcuate spicules and a non-protrusible gubernaculum. Apratylenchus vietnamensis gen. n., sp. n. is characterised by a sedentary mature female with a 368-515 μm long club-shaped body, cephalic region low, dome shaped with three annuli, body annuli bearing a single or double row of refractive tubercles, stylet 17.0-19.9 μm long, V = ca 88-92, spermatheca filled with sperm and males known, if rarely found, in the type population. Males have a weak, 15 μm long stylet with reduced knobs, 16 μm spicules, a non-protrusible gubernaculum and low bursa. It differs from A. binhi gen. n., sp. n., the only other nominal species of the new genus, by longer female body (368-515 vs 326-364 μm), longer distance from anterior end to anus (353-512 vs 310-348 μm) and longer stylet (17.0-19.9 vs 14.7-16.9 μm). In addition, the spermatheca is filled with round sperm vs empty in A. binhi gen. n., sp. n. and the cuticle has less densely spaced and less pronounced tubercles. D2/D3 sequences of the 28 rDNA gene clearly support the separation of Apratylenchus gen. n. from other Pratylenchidae and further distinguish the two new species from each other. Keywords – D2/D3, description, molecular, morphology, morphometrics, phylogeny, SEM, taxonomy. During nematode surveys conducted in 2005 in cof- fee plantations of Vietnam, we detected Pratylenchidae populations of two undescribed species infecting, in as- sociation with Pratylenchus coffeae (Zimmermann, 1898) Filipjev & Schuurmans Stekhoven, 1941, roots of both Robusta and Arabica coffee plants. The juveniles, imma- ture females and males of these populations, like many other Pratylenchidae, are migratory endoparasites char- acterised by a low cephalic region and a ventrally over- lapping pharyngeal gland lobe. However, the mature fe- ∗ Corresponding author, e-mail: [email protected] males are sedentary and club-shaped with a widened pos- terior region. All the post-embryonic life stages are char- acterised by the body annuli bearing transverse rows of tubercles. These morphological characters are incongru- ent with all known genera in the Pratylenchidae and can- not be accommodated in any of the extant subfamilies as currently defined (see Luc, 1987; Siddiqi, 2000). The classification of the family Pratylenchidae is in a state of flux. Systems proposed by Luc (1987) and Sid- diqi (2000) predate more recent insight from molecular © Koninklijke Brill NV, Leiden, 2009 DOI:10.1163/138855409X12465362560593 Also available online - www.brill.nl/nemy 565
-
Upload
universiteitgent -
Category
Documents
-
view
4 -
download
0
Transcript of Apratylenchus vietnamensis gen. n., sp. n. and A. binhi gen. n., sp. n., sedentary Pratylenchidae...

Nematology, 2009, Vol. 11(4), 565-581
Apratylenchus vietnamensis gen. n., sp. n. and A. binhi gen. n.,sp. n., sedentary Pratylenchidae (Nematoda: Tylenchida) from
coffee in Vietnam, with proposal of Apratylenchinae subfam. n.
Phap Q. TRINH 1,2,3, Lieven WAEYENBERGE 3,4, Chau N. NGUYEN 1, James G. BALDWIN 4,Gerrit KARSSEN 5,6 and Maurice MOENS 2,3,∗
1 Institute of Ecology and Biological Resources, 18 Hoang Quoc Viet Road, Hanoi, Vietnam2 Laboratory for Agrozoology, Ghent University, Coupure links 653, 9000 Ghent, Belgium
3 Institute for Agriculture and Fisheries Research, Burg. Van Gansberghelaan 96, 9820 Merelbeke, Belgium4 Department of Nematology, University of California, Riverside, CA 92521, USA
5 Plant Protection Service, Box 9102, 6700 HC Wageningen, The Netherlands6 Department of Biology, Ghent University, K.L. Ledeganckstraat 35, 9000 Ghent, Belgium
Received: 10 July 2007; revised: 17 October 2008Accepted for publication: 20 October 2008
Summary – Apratylenchinae subfam. n. is proposed within the Pratylenchidae to include Apratylenchus gen. n. and two new species,A. vietnamensis gen. n., sp. n. and A. binhi gen. n., sp. n., from coffee plantations in Vietnam. Apratylenchus gen. n. is distinguishedby the presence of transverse rows of cuticular tubercles on the annuli and by sexual dimorphism in which the males are stout andcylindrical, but mature sedentary females are club-shaped. The sedentary females are broadest in the posterior third although narrowingposterior to the vulva (V � 85%). The female cephalic region is low and dome-shaped with all labial sectors fused to one another andthe oral disc and the pharyngeal glands overlap the intestine ventrally. Males have a weak stylet with reduced knobs and a conical tailwith a blunt terminus, arcuate spicules and a non-protrusible gubernaculum. Apratylenchus vietnamensis gen. n., sp. n. is characterisedby a sedentary mature female with a 368-515 μm long club-shaped body, cephalic region low, dome shaped with three annuli, bodyannuli bearing a single or double row of refractive tubercles, stylet 17.0-19.9 μm long, V = ca 88-92, spermatheca filled with spermand males known, if rarely found, in the type population. Males have a weak, 15 μm long stylet with reduced knobs, 16 μm spicules,a non-protrusible gubernaculum and low bursa. It differs from A. binhi gen. n., sp. n., the only other nominal species of the new genus,by longer female body (368-515 vs 326-364 μm), longer distance from anterior end to anus (353-512 vs 310-348 μm) and longerstylet (17.0-19.9 vs 14.7-16.9 μm). In addition, the spermatheca is filled with round sperm vs empty in A. binhi gen. n., sp. n. and thecuticle has less densely spaced and less pronounced tubercles. D2/D3 sequences of the 28 rDNA gene clearly support the separation ofApratylenchus gen. n. from other Pratylenchidae and further distinguish the two new species from each other.
Keywords – D2/D3, description, molecular, morphology, morphometrics, phylogeny, SEM, taxonomy.
During nematode surveys conducted in 2005 in cof-fee plantations of Vietnam, we detected Pratylenchidaepopulations of two undescribed species infecting, in as-sociation with Pratylenchus coffeae (Zimmermann, 1898)Filipjev & Schuurmans Stekhoven, 1941, roots of bothRobusta and Arabica coffee plants. The juveniles, imma-ture females and males of these populations, like manyother Pratylenchidae, are migratory endoparasites char-acterised by a low cephalic region and a ventrally over-lapping pharyngeal gland lobe. However, the mature fe-
∗ Corresponding author, e-mail: [email protected]
males are sedentary and club-shaped with a widened pos-terior region. All the post-embryonic life stages are char-acterised by the body annuli bearing transverse rows oftubercles. These morphological characters are incongru-ent with all known genera in the Pratylenchidae and can-not be accommodated in any of the extant subfamilies ascurrently defined (see Luc, 1987; Siddiqi, 2000).
The classification of the family Pratylenchidae is in astate of flux. Systems proposed by Luc (1987) and Sid-diqi (2000) predate more recent insight from molecular
© Koninklijke Brill NV, Leiden, 2009 DOI:10.1163/138855409X12465362560593Also available online - www.brill.nl/nemy 565

P.Q. Trinh et al.
data suggesting that the group is paraphyletic (Al-Bannaet al., 1997; Subbotin et al., 2006; Bert et al., 2008), butadditional data are needed to substantiate further thesephylogenetic patterns so that they can then be reflected, ifwarranted, in a revised classification. In this paper, for thesake of clarity and as a conservative approach during thistransition period, we use taxonomic categories and termi-nology consistent with Pratylenchidae as proposed by Sid-diqi (2000). Based on morphology and molecular data, in-cluding comparisons with extant taxa, Apratylenchus gen.n. is best placed in a new subfamily within the Praty-lenchidae. Accordingly, Apratylenchinae subfam. n. isproposed to hold the new genus and the two new species:A. vietnamensis gen. n., sp. n. and A. binhi gen. n., sp. n.
Materials and methods
Of the 375 sites sampled during the survey, 11 wereinfested with specimens of the new genus. These sampleswere taken in the outskirts of Buon Ma Thuot City (DakLak province), Chu Se (Gia Lai province) and Khe Sanh(Quang Tri province), Vietnam.
Nematodes were extracted from soil by Cobb’s (1918)sieving method combined with filtering (Nguyen &Nguyen, 1993). In addition, nematodes were extractedfrom root samples by a combination of maceration andflotation-centrifugation (Coolen & D’Herde, 1972). Insome cases, obese sedentary females were gently dis-lodged from the root with the aid of dissecting tools un-der a stereomicroscope. After examination, the remain-ing root pieces were macerated and subjected to flotation-centrifugation, resulting in collection of immature andobese females. Attempts to initiate living cultures of thesenematodes on carrot disk (O’Bannon & Taylor, 1968)were not successful. Nematodes were killed in hot wa-ter (70◦C), fixed in TAF and mounted in anhydrous glyc-erin using the slow method of Hooper and Evans (1993).All morphometric data were collected and measured usingan Olympus BX51 microscope and photographed with anOlympus U-TV 0.5xC-3 digital camera.
For scanning electron microscopy (SEM), specimenspreserved in anhydrous glycerin were transferred to adrop of 4% formalin. Ultrasonic treatment for ca 10min was used to remove any particles adhering to thespecimens. The nematodes were then passed through anaqueous ethanol gradient of 25% (overnight), 50, 75, 95%(3 h each), and 100% (overnight) at 25◦C. Dehydrationwas followed by critical point drying with liquid CO2.Specimens were then mounted on stubs and coated with
gold-palladium (25 nm) for observation and photographyusing a Jeol LSM-840 at 15 kV.
Molecular analysis
For each of the nematode species, DNA was extracted(Waeyenberge et al., 2000) from female individuals pre-viously identified by morphology. The crude DNA extractwas kept at −70◦C until use. Five μl of the DNA ex-tract was used for amplification of the D2/D3 extensionregion of the 28 rDNA gene (De Ley et al., 1999). Afterelectrophoresis in 1% TAE-buffered agarose gel (1 h, 100V), the PCR product was visualised under UV light, ex-cised from the gel and purified using the Wizard SV Geland PCR Clean-Up System (Promega Benelux, Leiden,The Netherlands). Purified PCR products were cloned us-ing pGEM-T vectors (Protocols and Application Guide,3rd edn, Promega, Madison, WI, USA). Five clones werepicked by blue/white selection and further grown in 1ml liquid LB-medium (LB-broth, Sigma, Bornem, Bel-gium). One μl of a cultured colony was used for PCRwith SP6 and T7 vector primers (Promega Benelux). PCRconditions were the same as for the D2/D3 primers. Af-ter electrophoresis it was possible to determine if the in-sert was present. If so, the corresponding PCR productswere purified (Wizard SV Gel and Clean-Up System)and the DNA concentration was determined (NanodropND-1000, Isogen Life Sciences, Sint-Pieters-Leeuw, Bel-gium). Twenty ng were used for sequencing both DNAstrands using the vector primers according to the manu-facturer’s instructions (Protocol BigDye terminator v3.1Cycle Sequencing Kit, ABI Prism 377 DNA Sequencer,Applied Biosystems, Foster City, CA, USA). Finally, thesequences were edited using the software packages Chro-mas 2.00 (Technelysium, Helensvale, QLD, Australia)and BioEdit 7.0.4.1 (Hall, 1999). The resulting sequenceswere deposited in GenBank under the accession numbersEF645137 (A. binhi gen. n., sp. n.) and EF645138 (A. viet-namensis gen. n., sp. n.).
To investigate the phylogenetic position and relation-ship of the new species, sequences from both specieswere aligned with sequences of species of additional ge-nera of Pratylenchidae and two Paratylenchus spp., tothe extent that those sequences were available in theNational Center for Biotechnology Information (NCBI,http://www.ncbi.nlm.nih.gov/) or the ILVO-Nematologydatabase. To obtain the alignment, Clustal X 1.64 (Thomp-son et al., 1997) was used (pairwise alignment parame-ters: gap opening set on 10, gap extension set on 0.1; mul-
566 Nematology

A new genus and two new species of sedentary pratylenchid from Vietnam
tiple alignment parameters: gap opening set on 10, gap ex-tension set on 0.2). The alignment was subsequently putinto PAUP*4.0b10 (Swofford, 1998) to construct a Max-imum Parsimony (MP) tree. The MP was calculated bya heuristic search with default settings changed to 1000replicates and ‘adding of sequences’ randomly. Subse-quently, bootstrapping was done with 100 replicates tocalculate the robustness of the trees.
Family Pratylenchidae Thorne, 1949= Nacobbidae Chitwood in Chitwood & Chitwood,
1950= Radopholidae Allen & Sher, 1967
DIAGNOSIS (EMENDED FROM SIDDIQI, 2000)
Hoplolaimoidea. Vermiform (except mature femalesof Nacobbus Thorne & Allen, 1944 and, to a lesserdegree, Achlysiella Hunt, Bridge & Machon, 1989 andApratylenchus gen. n.). Cuticle prominently annulated.Lateral fields each with four to six incisures, very rarelyareolated posterior to pharynx. Deirids absent (exceptin Pratylenchoides Winslow, 1958). Phasmids pore-like,on tail located far posterior to anal level, extending intocaudal alae as pseudoribs, except in Hirschmanniella Luc& Goodey, 1964.
Female
Amphidial apertures pore-like, indistinct. Cephalic re-gion low, anteriorly flattened to broadly rounded, annu-lated; framework strongly sclerotised; labial disc incon-spicuous, en face lip pattern highly variable, includingwithin genera. Stylet strong, length not exceeding threecephalic region diam. (except in Hirschmanniella); conusabout as long as posterior part; knobs large, rounded,usually closely applied to shaft. Orifice of dorsal glandclose (usually 2-3 μm) to stylet base. Procorpus slen-der. Postcorpus strongly muscular, with prominent re-fractive thickenings. Isthmus short. Pharyngo-intestinaljunction indistinct, with a refringent valvula. Pharyngealglands extending over intestine; subventral glands asym-metrical, extending beyond dorsal gland; three gland nu-clei usually lying in tandem. Vulva a transverse slit, po-sitioned submedian to more posterior but not subtermi-nal; lips not modified; lateral membranes absent. Vaginadirected inward. Didelphic, amphidelphic or pseudo-mono-prodelphic. Ovaries outstretched; posterior ovarydegenerate in pseudo-monodelphic forms. Spermathecalarge, rounded, with small round or rod-like sperm when
impregnated. Tail conoid, sub-cylindrical to elongate-conoid, ca twice or more anal body diam. long, with roundto pointed tip that may bear a mucro (Hirschmanniella),generally with inconspicuous hyaline terminal portion.
Male
Stylet and pharynx similar to female or reduced as inRadopholinae Allen & Sher, 1967 and Apratylenchinaesubfam. n. Tail elongate-conoid, bursa terminal or subter-minal. Testis single, outstretched, spermatocytes in one ortwo rows. Spicules similar, cephalated, arcuate, pointedwith subterminal opening on dorsal or ventral side. Gu-bernaculum simple, fixed, or complex with telamon or tit-illae, protrusible. Hypoptygma present or absent.
Juveniles
Resembling female in anterior region and tail.Obligate migratory endoparasites of roots, some with
sedentary mature female (Nacobbus, Achlysiella, Apraty-lenchus gen. n.).
TYPE SUBFAMILY
Pratylenchinae Thorne, 1949
OTHER SUBFAMILIES
Apratylenchinae subfam. n.Hirschmanniellinae Fotedar & Handoo, 1978Nacobbinae Chitwood in Chitwood & Chitwood, 1950Radopholinae Allen & Sher, 1967
Apratylenchinae subfam. n.
DIAGNOSIS
Pratylenchidae. Marked sexual dimorphism in bodyshape. Juveniles, young females and males stout. Maturefemale sedentary, body club-shaped, swollen in posteriorthird but abruptly narrowing posterior to vulva. Cuticlesurface pattern of annuli comprising transverse rows oftubercles. Deirids absent. Phasmids located posterior toanal level. Cephalic region low, continuous, annulated,framework with moderately strong sclerotisation in bothsexes. Lip pattern with all labial sectors fused with one an-other and with oral disc. Amphidial apertures oval. Styletwell developed, basal knobs rounded to ellipsoid, some-times sloping slightly posteriorly. Orifice of dorsal glandclose to stylet base. Median bulb strongly muscular. Pha-ryngeal glands elongate, overlapping intestine ventrally.
Vol. 11(4), 2009 567

P.Q. Trinh et al.
Lateral field at mid-body marked by four indistinct in-cisures intersected by rows of tubercles. Vulva a trans-verse slit, located far posterior (V = ca 90), reproductivetract monodelphic, prodelphic. Post-uterine sac present.Tail conical, bluntly pointed or truncate with smooth orannulated terminus. Immature female motile, with bodystout but not swollen, otherwise similar to mature females.Male body cylindrical. Tubercles on annuli comprisingless pronounced than in female. Stylet slender, poorlydeveloped, stylet knobs weak. Pharynx well developed.Testis monorchic, outstretched. Spicules ventrally curved,gubernaculum small, fixed, wedge-shaped. Caudal alaenarrow, extending posteriorly almost to tail terminus. Tailbroadly conical with rounded terminus. Juveniles slender,with anterior and posterior body similar to immature fe-males.
TYPE AND ONLY GENUS
Apratylenchus gen. n.
RELATIONSHIPS
Apratylenchinae subfam. n. is unique in the Praty-lenchidae by virtue of the presence of cuticular tubercleson the annuli. Apratylenchinae subfam. n. is similar toPratylenchinae and Hirschmanniellinae with respect to thepharyngeal glands overlapping the intestine mostly ven-
trally and ventrolaterally (Table 1), but differs in showingmarked sexual dimorphism with club-shaped females andmales having a reduced stylet. Apratylenchinae subfam. n.shares with Nacobbinae and Radopholinae some aspectsof sexual dimorphism, but differs by the presence of ven-trally overlapping pharyngeal glands.
Molecular phylogenetics further establishes Apraty-lenchinae subfam. n. as sharing a clade distinct from anydefining Radopholinae or Nacobbinae.
Apratylenchus* gen. n.
DIAGNOSIS
Mature female
Sedentary. Body moderately small (<600 μm), club-shaped, broadening in posterior third but abruptly nar-rowing posterior to vulva. Annuli ornamented by trans-verse single or double rows of rounded tubercles arrangedlike kernels on an ear of corn. Deirids absent. Cepha-lic region low, dome-shaped, framework with moderatelystrong sclerotisation in both sexes. Cephalids not seen. Enface lip pattern with all labial sectors fused with one an-other and with oral disc. Amphidial apertures oval. Stylet
* Specific epithet derived from Greek and consisting of the α
prefix and Pratylenchus.
Table 1. Characters differentiating genera of Pratylenchidae.
Subfamily Genera Deirids Mature Male Pharyngeal glands Female Lateral Caudal alaefemale secondary Abutting Over- branches field Terminal Sub-shape sexual lapping* genital incisures terminal
dimorphism
Radopholinae Pratylenchoides + vermiform ++ + D 2 6 (4) +Apratylenchoides − vermiform + D 1 4 +Radopholus − vermiform + + + D 2 4 + (+)Achlysiella − sausage + + + D 2 4 +Radopholoides − vermiform + + + D 1 4 +Hoplotylus − vermiform + + + D 1 4 +Zygradus − vermiform + D 2 4 +
Pratylenchinae Zygotylenchus − vermiform − V 2 4 +Pratylenchus − vermiform − V 1 6 (4) +
Hirschmanniel- Hirschmanniella − vermiform − V 2 4 +linae
Nacobbinae Nacobbus − saccate − D 1 4 +Apratylenchinae Apratylenchus − club + V 1 4 +
subfam. n. gen. n.
* V = ventral; D = dorsal.
568 Nematology

A new genus and two new species of sedentary pratylenchid from Vietnam
robust with large ellipsoidal or rounded knobs, sometimesslightly sloping posteriorly. Pharyngeal glands overlap-ping intestine ventrally. Dorsal gland nucleus located pos-terior to pharyngo-intestinal valve. Lateral field markedby four indistinct incisures, areolated by transverse rowsof tubercles. Vulva a transverse slit, located far posterior(V = ca 90). Vulval lips smooth, not tuberculate. Gen-ital tract monodelphic, prodelphic. Post-uterine branchvery short, pouch-like. Phasmids located anterior to pos-terior third of tail. Tail conical with terminus bluntlypointed or truncate and with smooth, annulated or in-dented terminus.
Immature female
Mobile, body stout but not swollen or club-shaped;morphologically similar to mature female.
Male
Mobile, cylindrical, not swollen. Annuli marked withless pronounced tubercles than female. Stylet slender,poorly developed, with weak basal knobs. Pharynx welldeveloped but with metacorpus and pharyngeal glandssmaller than in female. Spicules curved ventrally withasymmetrical, oval, head. Gubernaculum simple, wedge-shaped, non-protrusible. Caudal alae narrow, extendingposteriorly to point near tail terminus. Tail broadly conicalwith rounded terminus. Phasmids located posterior tocloaca, slightly extending into bursa as a pseudorib.
Juveniles
Slender and similar in shape, digestive tract and cuticu-lar pattern to immature female.
TYPE SPECIES
A. vietnamensis gen. n., sp. n.
OTHER SPECIES
A. binhi gen. n., sp. n.
RELATIONSHIPS
Apratylenchus gen. n. is unique because of the presenceof cuticular tubercles on the body annules. It shares somecharacters with Pratylenchus Filipjev, 1936 and Zygoty-lenchus Siddiqi, 1963, including the pharyngeal glandsoverlapping the intestine ventrally (Table 1). The lip pat-tern of Apratylenchus gen. n., including a high degree of
fusion of labial sectors to one another and the labial disc,resembles the patterns of some species of Hirschman-niella and Pratylenchus (Baldwin & Bell, unpubl.; Sher &Bell, 1975; Corbett & Clark, 1983; Duncan et al., 1999;Subbotin et al., 2008). However, Apratylenchus gen. n.can be distinguished from Pratylenchus by the markedsexual dimorphism, including the swollen body of themature sedentary female and the poorly developed styletin the male. Apratylenchus gen. n. differs from Zygoty-lenchus by monodelphy vs didelphy.
The swollen body and sedentary endoparasitism ofmature female Apratylenchus gen. n. resembles that ofAchlysiella. However, it differs from the latter by mon-odelphy vs didelphy and ventral vs dorsal overlap of thepharyngeal glands. The male pharynx of Achlysiella isalso more reduced than in Apratylenchus gen. n.
Within the Pratylenchidae, sexual dimorphism ispresent in Radopholus Thorne, 1949, Hoplotylus s’-Jacob,1960, Pratylenchoides, Apratylenchoides Sher, 1973, Zy-gradus Siddiqi, 1991 and Nacobbus Thorne & Allen,1944. Apratylenchinae subfam. n. differs from all of thesegenera by the ventral vs dorsal overlap of the pharyngealglands.
The ventral overlap of the pharyngeal glands in Apraty-lenchus gen. n. is shared with Hirschmanniella fromwhich it otherwise differs by monodelphy vs didelphy, theswollen body of sedentary mature females vs vermiform,phasmids located in a slightly more anterior position andthe cuticular pattern of the annuli comprising rows of tu-bercles vs not.
In its tuberculate cuticle, swollen body and sexual di-morphism, the new genus shares several superficial char-acteristics with representatives of the Tylenchulidae Skar-bilovich, 1947 (Criconematoidea Taylor, 1936) such asCacopaurus Thorne, 1943 (viz. C. pestis Thorne, 1943)and Paratylenchus Micoletzky, 1922 (viz. P. colina Huang& Raski, 1986 and P. punctata Huang & Raski, 1986).However, Apratylenchus gen. n. is clearly distinguishedby a typical pratylenchid head shape, ventral overlap ofthe elongate pharyngeal glands, heavily sclerotised cepha-lic region and relatively much shorter stylet. Furthermore,the cuticular tubercles in Cacopaurus and Paratylenchusare less prominent and sparser than in Apratylenchus gen.n. (Bello & Belart, 1975; Huang & Raski, 1986).
Molecular phylogenies from this and previous studiesfurther establish that Paratylenchus and other Criconema-toidea are a monophyletic group distinct from any Praty-lenchidae (Subbotin et al., 2006).
Vol. 11(4), 2009 569

P.Q. Trinh et al.
Apratylenchus vietnamensis* gen. n., sp. n.(Figs 1-3)
MEASUREMENTS
See Table 2.
DESCRIPTION
Mature female
Sedentary. Body relatively short (<520 μm), club-shaped, becoming broader in posterior third but abruptlynarrowing posterior to vulva. Cuticle surface with an-nuli comprising transverse single or double rows of tu-bercles. With SEM, tubercles appearing rounded withsmooth margins, arranged like kernels on an ear of corn;with light microscopy appearing as rows of punctations.Cephalic region low, dome-shaped, framework with mod-erately strong sclerotisation in both sexes. Cephalids notseen. En face lip pattern with all labial sectors fusedwith one another and with oral disc; additional lip annulisometimes discontinuous. Amphidial apertures oval. Lat-eral field tuberculate with four indistinct incisures at mid-body, occasionally with five incisures in vulval region.Stylet robust with large ellipsoidal or rounded knobs,sometimes slightly sloping posteriorly. Dorsal pharyngealgland opening situated less than 5 μm posterior to stylet.Hemizonid less than one annulus anterior to excretorypore and two body annuli long. Hemizonion not seen.Pharyngeal glands elongate, overlapping intestine ven-trally. Dorsal gland nucleus located posterior to pharyngo-intestinal valve. Nuclei of subventral glands in mid- andposterior third of pharyngeal lobe. Lateral field ca 3.5-6μm wide at mid-body, marked by four indistinct incisures,areolated by transverse rows of tubercles. Lateral field ex-tending from metacorpus level to posterior to vulva. Vulvaa transverse slit, located far posterior (V = ca 90). Vul-val lips smooth, not tuberculate; lateral flaps and epiptyg-mata absent. Anterior vulvar lip salient, almost coveringposterior lip. Genital tract monodelphic, prodelphic, out-stretched, ovary extending to mid-body region. Spermath-eca spherical, axial, filled with rounded sperm. Ovary withoocytes arranged in one or two rows. Post-uterine branchvery short, pouch-like. Phasmids located 6-8 annuli pos-terior to anus, distinct only with light microscopy. Cau-dalids not seen. Tail broadly conical with 14-20 annuli,
* Specific epithet derived from Vietnam, the country where thespecies was found.
either bluntly pointed with smooth terminus or with atruncate, annulated or indented, terminus.
Immature female
Motile, body stout but not swollen or club-shaped asin sedentary female; morphologically similar to maturefemale.
Male
Mobile, cylindrical, not swollen. Annuli marked withless pronounced tubercles than in female. Stylet slender,poorly developed, with weak basal knobs. Pharynx welldeveloped but with metacorpus and pharyngeal glandssmaller than in female. Lateral field at mid-body withfour indistinct incisures. Testis monorchic, outstretched.Spicules curved ventrally with asymmetrical, oval, head.Gubernaculum simple, wedge shaped, non-protrusible.Caudal alae narrow, extending posteriorly to point neartail terminus. Tail broadly conical with rounded terminus.Phasmids located posterior to cloacal aperture.
Juveniles
Slender and similar in shape, digestive tract and cuticu-lar pattern to immature females.
TYPE HOST AND LOCALITY
Isolated from roots of 13-year-old Coffea canephoraPierre (Robusta coffee) growing in clay soil at Buon MaThuot, Dak Lak, Vietnam.
OTHER HOST AND LOCALITIES
A few specimens were also found on roots of Arabicacoffee (Coffea arabica L. cv. Catimor) and in sandy clayloam soil at Khe Sanh, Quang Tri, Vietnam, and also onroots of Robusta coffee growing in sandy clay soil at ChuSe, Gia Lai, Vietnam.
TYPE MATERIAL
Holotype female and 40 female paratypes deposited inthe Nematode Collection of Institute of Ecology and Bi-ological Resources, Vietnamese Academy of Science andTechnology, 18 Hoang Quoc Viet Road, Hanoi, Vietnam(IEBR). Five female and one male paratypes depositedat the University of California Riverside Nematode Col-lection (UCRNC), Riverside, CA, USA, and five femaleparatypes in the nematode collection of the Zoology Mu-seum, Ghent University, K.L. Ledeganckstraat 35, Ghent,Belgium (UGMD 104107).
570 Nematology

A new genus and two new species of sedentary pratylenchid from Vietnam
Fig. 1. Apratylenchus vietnamensis gen. n., sp. n. A: Entire female; B: Entire male; C: En face; D: Female head region; E: Femaleanterior region; F: Male anterior region; G: Female cuticular surface structure; H: Female lateral field; I: Female posterior bodyvariation; J: Female, gonad and posterior region; K: Male, posterior region.
Vol. 11(4), 2009 571
P.Q. Trinh et al.
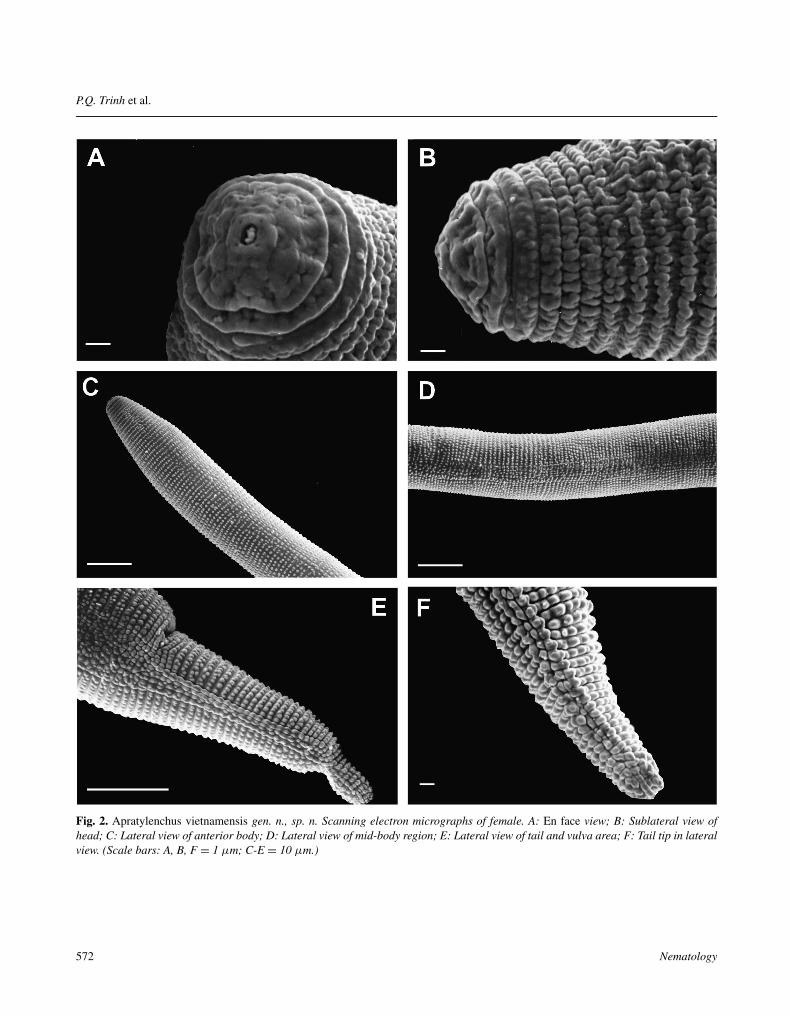
Fig. 2. Apratylenchus vietnamensis gen. n., sp. n. Scanning electron micrographs of female. A: En face view; B: Sublateral view ofhead; C: Lateral view of anterior body; D: Lateral view of mid-body region; E: Lateral view of tail and vulva area; F: Tail tip in lateralview. (Scale bars: A, B, F = 1 μm; C-E = 10 μm.)
572 Nematology
A new genus and two new species of sedentary pratylenchid from Vietnam
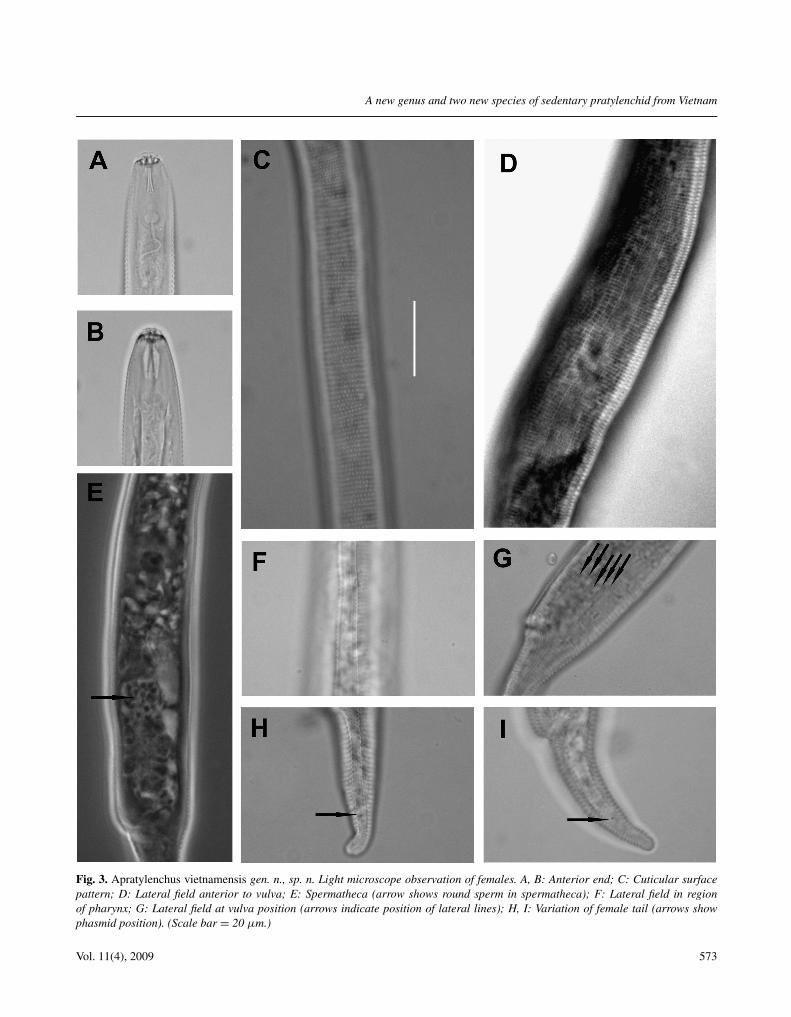
Fig. 3. Apratylenchus vietnamensis gen. n., sp. n. Light microscope observation of females. A, B: Anterior end; C: Cuticular surfacepattern; D: Lateral field anterior to vulva; E: Spermatheca (arrow shows round sperm in spermatheca); F: Lateral field in regionof pharynx; G: Lateral field at vulva position (arrows indicate position of lateral lines); H, I: Variation of female tail (arrows showphasmid position). (Scale bar = 20 μm.)
Vol. 11(4), 2009 573

P.Q. Trinh et al.
Table 2. Morphometrics of Apratylenchus vietnamensis gen. n., sp. n. and A. binhi gen. n., sp. n. All measurements in μm and in theform: mean ± s.d. (range).
Character Species
A. vietnamensis gen. n., sp. n. A. binhi gen. n., sp. n.
Female Male Female
Holotype Paratypes Paratype Holotype Paratypes
n – 40 1 – 12L 416 421 ± 35.3 392 364 349 ± 13.1
(368-515) (326-364)L′ 399.7 406 ± 35.8 368 344 332 ± 13.2
(353-512) (310-348)a 16.2 18.3 ± 1.9 23.1 18.1 17.7 ± 2.6
(14.7-23.6) (14.8-21.5)b 5.6 6.2 ± 0.6 4.6 4.9 5.6 ± 0.5
(5.3-7.4) (4.5-6.2)b′ 4.5 4.3 ± 0.4 3.5 3.9 3.8 ± 0.3
(3.5-5.2) (3.3-4.1)c 25.5 23.5 ± 2.3 16 18.9 21.8 ± 2.5
(19.1-29.5) (18.9-26.6)c′ 1.3 1.9 ± 0.3 2.1 1.9 1.9 ± 0.1
(1.3-2.7) (1.7-2.2)V 90.9 90.0 ± 0.8 – 90.2 89.9 ± 0.7
(87.7-91.6) (88.1-90.6)V′ 94.6 93.6 ± 1.5 – 95.2 94.3 ± 0.7
(89.2-95.3) (93.0-95.3)Max. body diam 25.7 23.2 ± 3.1 17 20.1 20.1 ± 2.9
(18.8-30.1) (15.5-24.1)Lip region height 2.6 2.3 ± 0.3 2.5 2.2 2.2 ± 0.3
(1.7-3.0) (1.8-3.0)Lip region diam. 8.4 6.6 ± 0.7 6 6.0 6.2 ± 0.7
(5.4-8.4) (5.3-7.5)Stylet length 18.3 18.0 ± 0.8 15 16.5 16.0 ± 0.8
(17.0-19.9) (14.7-16.9)Basal knobs width 4.5 4.0 ± 0.5 2 4.0 4.0 ± 0.2
(3.0-6.7) (3.6-4.3)Basal knobs height 3.0 2.6 ± 0.4 2 2.8 2.5 ± 0.3
(2.0-4.2) (2.0-2.9)DGO 4.6 4.1 ± 0.7 4 3.9 4.1 ± 0.9
(2.6-5.7) (2.9-5.6)Anterior to median bulb 48.1 45.5 ± 3.8 37.5 47.2 42.9 ± 3.5
(36.0-55.7) (38.3-48.9)Anterior to pharyngo-intestinal 60 62 ± 6.6 85 61 60 ± 6.9
junction (48-77) (52-74)Anterior to tip of pharyngeal 92 97 ± 6.7 112 92 93 ± 7.4
gland lobe (87-115) (85-108)Pharyngeal gland lobe 65 65 ± 9.0 44 66 65 ± 6.9
(40-82) (57.5-79.1)Anterior to excretory pore 73 71 ± 4.9 70 71 65 ± 4.1
(62-83) (58-71)
574 Nematology

A new genus and two new species of sedentary pratylenchid from Vietnam
Table 2. (Continued).
Character Species
A. vietnamensis gen. n., sp. n. A. binhi gen. n., sp. n.
Female Male Female
Holotype Paratypes Paratype Holotype Paratypes
Width of annuli at mid-body 1.1 1.2 ± 0.1 1.1 1.1 1.2 ± 0.1(1.0-1.4) (1.0-1.4)
Spermatheca diam. 12.8 12.4 ± 1.5 – 10.9 10.5 ± 2.5(10.6-16.1) (6.6-14.1)
Tail length 16.3 18.0 ± 1.9 24 19.3 16.2 ± 1.7(15-22) (13.2-19.3)
Anal body diam. 12.8 9.5 ± 1.3 14.4 10.4 8.6 ± 1.0(7.2-12.8) (6.9-10.4)
Vulval body diam. 20.5 16.4 ± 2.1 – 14.9 13.9 ± 1.5(13.2-20.8) (11.8-15.9)
Annuli on tail 15 16 ± 1.6 19 18 17 ± 1.4(14-20) (14-19)
Annuli from anus to vulva 22 19 ± 2.1 – 19 18 ± 2.1(15-24) (14-21)
Ovary length 92 141 ± 28.6 – 94 102 ± 18.8(92-202) (75-126)
Post-uterine sac length 13.3 9.2 ± 2.1 – 10.2 7.8 ± 2(6.1-13.3) (5.4-11.0)
MB 52.4 47.0 ± 2.5 33.5 51.1 46.5 ± 2.5(41.6-52.4) (43.8-51.1)
Lip diam. / lip height 3.2 2.8 ± 0.4 2.4 2.8 2.8 ± 0.3(2.2-3.8) (2.4-3.4)
Metacorpus length / diam. 1.0 1.2 ± 0.1 1.2 1.3 1.2 ± 0.2(0.8-1.5) (1.0-1.4)
Tail length / stylet length 0.9 1.0 ± 0.1 1.6 1.2 1.0 ± 0.1(0.8-1.2) (0.9-1.2)
Spicule length – – 16 – –Gubernaculum length – – 4 – –
DIAGNOSIS AND RELATIONSHIP
Apratylenchus vietnamensis gen. n., sp. n. is charac-terised by a sedentary mature female with a 368-515 μmlong club-shaped body, cephalic region low, dome-shapedwith three annuli, body annuli bearing a single or dou-ble row of refractive tubercles, stylet 17.0-19.9 μm long,vulva located at ca 88-92% of body length, spermath-eca filled with sperm and males known, if rarely found,in the type population. Males have a weak stylet 15 μmlong with reduced knobs, 16 μm long spicules, a non-protrusible gubernaculum and low bursa.
Apratylenchus vietnamensis gen. n., sp. n. differs fromA. binhi gen. n., sp. n., the only other nominal species ofthe genus, by a longer female body (368-515 vs 326-364
μm), longer distance from the anterior end to the anus(353-512 vs 310-348 μm) and a longer stylet (17.0-19.9vs 14.7-16.9 μm). In addition, in A. vietnamensis gen. n.,sp. n. the spermatheca is filled with round sperm vs emptyin A. binhi gen. n., sp. n. and the cuticle has less denselyspaced and less pronounced tubercles.
Apratylenchus binhi* gen. n., sp. n.(Figs 4-6)
MEASUREMENTS
See Table 2.
* Named after Mr Binh Thanh Phan who collected the samplesinfested with the new species.
Vol. 11(4), 2009 575
P.Q. Trinh et al.
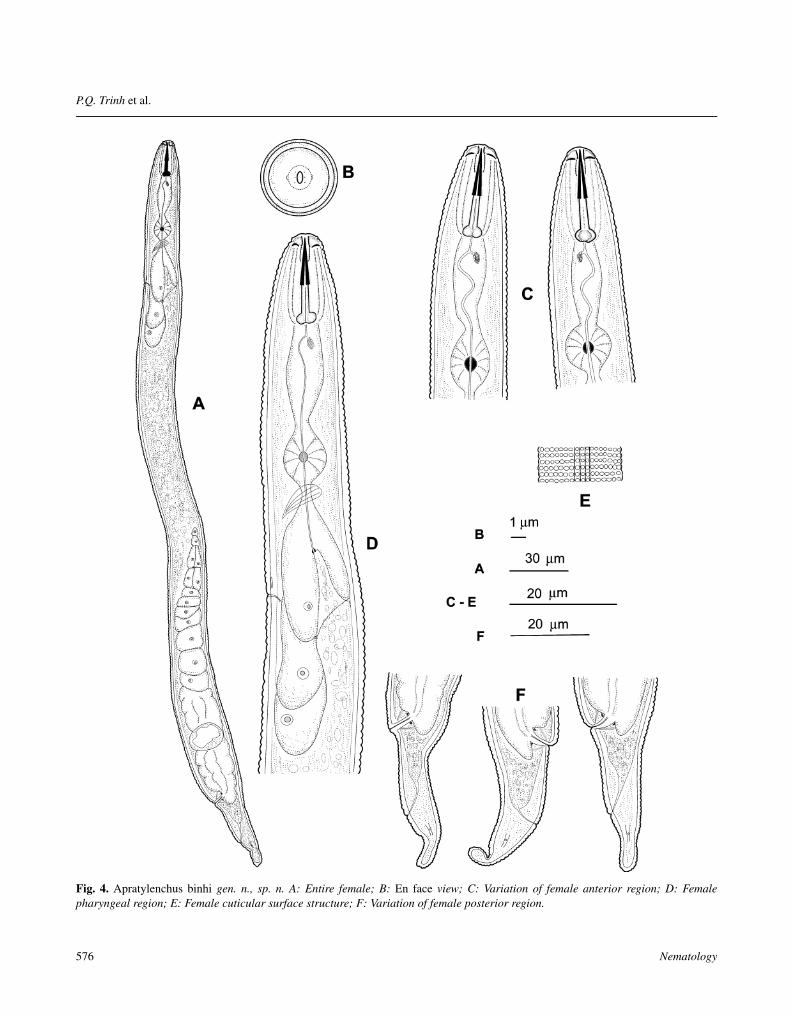
Fig. 4. Apratylenchus binhi gen. n., sp. n. A: Entire female; B: En face view; C: Variation of female anterior region; D: Femalepharyngeal region; E: Female cuticular surface structure; F: Variation of female posterior region.
576 Nematology

A new genus and two new species of sedentary pratylenchid from Vietnam
Fig. 5. Apratylenchus binhi gen. n., sp. n. Scanning electron micrographs of female. A: En face view; B: Sublateral view of head; C:Lateral view of anterior body; D: Lateral view of mid-body region; E: Lateral view of tail and vulva region; F: Tail tip in lateral view.(Scale bars: A, B, F = 1 μm; C-E = 10 μm.)
DESCRIPTION
Mature female
Sedentary, body < 370 μm long, generally straightto slightly curved ventrally, resembling a club, slightlyswollen anterior to vulva, abruptly narrower posteriorto vulva. Surface body annuli marked by single row ofdense refractive cuticular tubercles with smooth marginsarranged like kernels on an ear of corn, appearing as rowsof refractive dots with light microscopy. Cephalic regionlow, dome-shaped, with three lip annuli. Lip pattern withlabial sectors fused together and with oral disc. Anterior-most (labial) annulus slightly narrower than second, con-
vex in profile. Second lip annulus distinctly narrower thanthird; lip annuli sometimes discontinuous. Cephalids notseen. Deirids absent. Lateral field areolated, ca 3.5-5 μmwide at mid-body, with four indistinct incisures intersect-ing transverse rows of tubercles. Lateral lines convergingposterior to vulva, reducing to two at phasmid level. Am-phidial apertures ovoid. Oral orifice distinctly oval. Styletrobust, stout, conus and shaft equal in length, basal knobsrounded, ellipsoid or sloping posteriorly. Hemizonid lessthan one annulus anterior to excretory pore and two an-nuli long. Hemizonion not seen. Metacorpus roundedto slightly oval with well developed ovoid valve. Pha-ryngeal gland lobe overlapping intestine ventrally. Dor-
Vol. 11(4), 2009 577

P.Q. Trinh et al.
Fig. 6. Apratylenchus binhi gen. n., sp. n. Light microscope observation of females. A, B: Anterior end; C: Cuticular surface pattern; D:Lateral field at mid-body; E: Vulva and spermatheca (arrow shows collapsed spermatheca without sperm); F, G: Female tail variation(arrows show phasmid position); H: Lateral field in posterior body. (Scale bar = 20 μm.)
sal gland nucleus located posterior to pharyngo-intestinalvalve. Sublateral gland nuclei situated in posterior thirdof pharyngeal lobe. Excretory pore located posterior tonerve ring and pharyngo-intestinal valve. Vulva a trans-verse slit, located far posterior (V = ca 90). Genital tractmonodelphic, prodelphic, outstretched, ovary extendingto mid-body region. Oocytes typically arranged in one,rarely two, rows. Spermatheca rounded to broadly oval,with large central cavity and lacking sperm. Anterior vul-
val lip salient. Post-uterine sac very short, ca 0.5 vulvalbody diam. long. Tail broadly conical with 14-19 annuli,terminus bluntly pointed to truncate, annulated or slightlyindented. Phasmids distinct, located 4-5 annuli posteriorto anus.
Immature female
Mobile with stout, but not swollen, body. Morphologi-cally similar to mature female.
578 Nematology

A new genus and two new species of sedentary pratylenchid from Vietnam
Male
Not found.
Juveniles
Mobile with stout, but not swollen, body. Morphologi-cally similar to mature female.
TYPE HOST AND LOCALITY
Rhizosphere and roots of coffee (Coffea canephora,‘Robusta’) collected at Buon Ma Thuot, Vietnam.
TYPE MATERIAL
Holotype female and five female paratypes deposited inthe Nematode Collection of the Institute of Ecology andBiological Resources, Vietnamese Academy of Scienceand Technology, 18 Hoang Quoc Viet Road, Hanoi,Vietnam (IEBR), and five female paratypes deposited inthe nematode collection of the Zoology Museum, GhentUniversity, K.L. Ledeganckstraat 35, Ghent, Belgium(UGMD104108).
DIAGNOSIS AND RELATIONSHIP
Apratylenchus binhi gen. n., sp. n. is characterised bya sedentary mature female with a 326-364 μm long club-shaped body, cephalic region low, dome shaped with threeannuli, body annuli bearing a single row of refractive,prominent, densely packed tubercles, stylet 14.7-16.9 μmlong, vulva located at ca 88-91% of body length, emptyspermatheca and males apparently absent.
The two new species of Apratylenchus gen. n. are sim-ilar in most morphological and morphometric characters.However, A. binhi gen. n., sp. n. differs from A. vietna-mensis gen. n., sp. n., the type and only other species ofthe genus, by a shorter female body (326-364 vs 368-515 μm), shorter distance from the anterior end to theanus (310-348 vs 353-512 μm) and a shorter stylet (14.7-16.9 vs 17.0-19.9 μm). In addition, in A. binhi gen. n., sp.n. the spermatheca is empty vs filled with round sperm inA. vietnamensis gen. n., sp. n., and the annuli bear moredensely packed and more pronounced tubercles than in A.vietnamensis gen. n., sp. n.
MOLECULAR CHARACTERISATION
The D2/D3 sequences of both Apratylenchus gen.n. species are of similar length, being 776 bp in A.vietnamensis gen. n., sp. n. and 778 bp in A. binhi gen.n., sp. n. However, the composition is only 84% similar.
Comparison of the sequences with the nucleotide data-bases accessible via BLAST (NCBI) reveals the se-quences to be most similar to Pratylenchus and Hirsch-manniella species. The similarity is only 85 and 80%, withquery coverage of at least 95%, for A. binhi gen. n., sp. n.and A. vietnamensis gen. n., sp. n., respectively. This sup-ports the hypothesis that both species belong to a cladethat includes most genera of Pratylenchidae. However, onthe basis of the molecular data currently available in Gen-Bank (Pratylenchus, Hirschmanniella and Radopholus) itdoes not support Apratylenchus gen. n. as a member orsister taxon of any clade representing an extant genus.
To position the new species and genus among the othergenera and to provide a phylogenetic context and re-veal relationships with other species, the sequences ofboth species were aligned with those of species belong-ing to several genera of Pratylenchidae. Paratylenchusnanus Cobb, 1923 was used as an outgroup. The MPtrees (Fig. 7) were calculated based on 315 parsimony-informative characters. At the end of the heuristic search1 580 543 rearrangements were tried and only one mostparsimonious tree was retained. The tree showed twoclades (I and II). Clade I is composed of Radopholus andHoplotylus. Clade II contains the remaining members ofPratylenchidae including Apratylenchus gen. n. Clade IIcan further be subdivided into two sub-clades (Ia and Ib)(Fig. 7). The two new Apratylenchus n. gen. species aregrouped in sub-clade Ia as a separate group. Based onthese results, the two new species can be regarded as beingmembers of a new genus closely related to Pratylenchus,Hirschmanniella and Nacobbus.
Acknowledgements
This work received financial support from a VLIR PhDscholarship (2005-0018) for Phap Quang Trinh at GhentUniversity, Belgium. The authors thank Mrs Rita VanDriessche and Mrs Marjolein Courveur, Department ofBiology, Ghent University, for their assistance in SEMpreparation and Mr Hoa Xuan Nguyen from the WesternHighlands Agro-forestry Science Technical Institute inDak Lak province, Vietnam, for his help in sampling.
Vol. 11(4), 2009 579

P.Q. Trinh et al.
Fig. 7. Maximum Parsimony tree after heuristic search (1000 replicates) using PAUP* 4.0b10. Bootstrap values greater than 50% areindicated.
References
AL-BANNA, L., WILLIAMSON, V. & GARDNER, S.L. (1997).Phylogenetic analysis of nematodes of genus Pratylenchususing nuclear 26S rDNA. Molecular Phylogenetics and Evo-lution 7, 94-102.
ALLEN, M.W. & SHER, S.A. (1967). Taxonomic problemsconcerning the phytoparasitic nematodes. Annual Review ofPhytopathology 5, 247-264.
BAERMANN, G. (1917). Eine einfache Methode zur Auffindungvon Ankylostomum (Nematoden) Larven in Erdproben. Ge-neeskundig Tijdschrift Nederlands-Indië 57, 131-137.
BELLO, A. & BELART, C. (1975). Cacopaurus pestis Thorne,1943 (Nematoda: Criconematoidea) en España. NematologiaMediterranea 3, 167-171.
BERT, W., LELIAERT, F., VIERSTRAETE, A.R., VAN-FLETEREN, J.R. & BORGONIE, G. (2008). Molecular phy-logeny of the Tylenchina and evolution of the female gon-oduct (Nematoda: Rhabditida). Molecular Phylogenetics andEvolution 48, 728-744.
CHITWOOD, B.G. (1950). An outline classification of thenematodes. In: Chitwood, B.G. & Chitwood, M.B. (Eds).An introduction to nematology. I. Anatomy. Baltimore, MD,USA, Monumental Printing, pp. 12-27.
COBB, N.A. (1918). Estimating the nema population of the soil.Agricultural Technology Circular 1. Washington, DC, Bureauof Plant Industry, United States Department of Agriculture,48 pp.
COOLEN, W.A. & D’HERDE, C.J. (1972). A method for thequantitative extraction of nematodes from plant tissue. StateAgricultural Research Centre, Ghent, Belgium, 77 pp.
CORBETT, D.C.M. & CLARK, S.A. (1983). Surface features inthe taxonomy of Pratylenchus species. Revue de Nématologie6, 85-98.
DE LEY, P., FÉLIX, M.A., FRISSE, L.M., NADLER, S.A.,STERNBERG, P.W. & THOMAS, W.K. (1999). Molecu-lar and morphological characterisation of two reproduc-tively isolated species with mirror-image anatomy (Nema-toda: Cephalobidae). Nematology 1, 591-612.
DUNCAN, L.W., INSERRA, R.N., THOMAS, W.K., DUNN, D.,MUSTIKA, I., FRISSE, L.M., MENDES, M.L., MORRIS,
580 Nematology

A new genus and two new species of sedentary pratylenchid from Vietnam
K. & KAPLAN, D.T. (1999). Molecular and morphologicalanalysis of isolates of Pratylenchus coffeae and closelyrelated species. Nematropica 29, 61-80.
FILIPJEV, I.N. (1936). On the classification of the Tylenchinae.Proceedings of the Helminthological Society of Washington3, 80-82.
FOTEDAR, D.N. & HANDOO, Z.A. (1978). A revised schemeof classification to order Tylenchida Thorne, 1949 (Nema-toda). Journal of Science, University of Kashmir 3 (1975),55-82.
HALL, T.A. (1999). BioEdit: a user-friendly biological se-quence alignment editor and analysis program for Win-dows95/98/NT. Nucleic Acids Symposium Series 41, 95-98.
HOOPER, D.J. & EVANS, K. (1993). Extraction, identifica-tion and control of plant parasitic nematodes. In: Evans, K.,Trudgill, D.L. & Webster, J.M. (Eds). Plant parasitic nema-todes in temperate agriculture. Wallingford, UK, CABI Pub-lishing, pp. 1-59.
HUANG, C.S. & RASKI, D.J. (1986). Four new species ofGracilacus Raski, 1962 (Criconematoidea: Nemata). Revuede Nématologie 9, 347-356.
HUNT, D.J., BRIDGE, J. & MACHON, J.E. (1989). OnAchlysiella, a new genus of obese Pratylenchidae (Nematoda:Tylenchoidea). Revue de Nématologie 12, 401-407.
S’-JACOB, J.J. (1960). Hoplotylus femina n. g., n. sp. (Praty-lenchinae: Tylenchida) associated with ornamental trees. Ne-matologica 4 (1959), 317-321.
KHAN, E. & NANJAPPA, C.K. (1972). Four new species in thesuperfamily Hoplolaimoidea (Tylenchida: Nematoda) fromIndia. Bulletin of Entomological Research 11 (1970), 143-149.
LUC, M. (1987). A reappraisal of Tylenchina (Nemata). 7. Thefamily Pratylenchidae Thorne, 1949. Revue de Nématologie10, 203-218.
LUC, M. & GOODEY, J.B. (1964). Hirschmanniella nom. nov.for Hirschmannia. Nematologica 9 (1963), 471.
MICOLETZKY, H. (1922). Die freilebenden Erd-Nematoden.Archive Naturgeschelschaft Berlin Abteilung A. 87 (1921), 1-650.
NGUYEN, N.C. & NGUYEN, V.T. (1993). [A modified tech-nique for extraction of nematodes from soil and plant tissues.]Achievements applied into practices. National Center for Nat-ural Sciences and Technology, pp. 41-45.
O’BANNON, J.H. & TAYLOR, A.L. (1968). Migratory en-doparasitic nematodes reared on carrot discs. Phytopathology58, 385.
SHER, S.A. (1973). The esophageal glands of PratylenchusFilipjev and Apratylenchoides belli n. gen., n. sp. (Nematoda:Tylenchoidea). Journal of Nematology 5, 218-221.
SHER, S.A. & BELL, A.H. (1975). Scanning electron mi-croscopy of the anterior region of some species of Tylen-
choidea (Tylenchina: Nematoda). Journal of Nematology 7,69-83.
SIDDIQI, M.R. (1963). On the classification of the Pratylenchi-dae (Thorne, 1949) nov. grad. (Nematoda: Tylenchida), witha description of Zygotylenchus browni nov. gen. et nov. sp.Zeitschrift für Parasitenkunde 23, 390-396.
SIDDIQI, M.R. (1991). Tanzanius coffeae gen. n., sp. n. andZygradus rector gen. n., sp. n. (Nematoda: Tylenchina) fromAfrica. Afro-Asian Journal of Nematology 1, 101-107.
SIDDIQI, M.R. (2000). Tylenchida parasites of plants andinsects, 2nd edition. Wallingford, UK, CABI Publishing, viii+ 834 pp.
SUBBOTIN, S.A., STURHAN, D., CHIZHOV, V.N., VOVLAS,N. & BALDWIN, J.G. (2006). Phylogenetic analysis of Ty-lenchida Thorne, 1949 as inferred from D2 and D3 expansionfragments of the 28S rRNA gene sequences. Nematology 8,455-474.
SUBBOTIN, S.A., RAGSDALE, E.J., MULLENS, T., ROBERTS,P.A., MUNDO-OCAMPO, M. & BALDWIN, J.G. (2008).A phylogenetic framework for root lesion nematodes of thegenus Pratylenchus (Nematoda): evidence from 18S andD2-D3 expansion segments of 28S ribosomal RNA genesand morphological characters. Molecular Phylogenetics andEvolution 48, 491-505.
SWOFFORD, D.L. (1998). PAUP*. Phylogenetic analysis usingparsimony and other methods. Version 4. Sunderland, MA,USA, Sinauer Associates, 128 pp.
THOMPSON, J.D., GIBSON, T.J., PLEWNIAK, F., JEANMOU-GIN, F. & HIGGINS, D.G. (1997). The ClustalX Windowsinterface: flexible strategies for multiple sequence alignmentaided by quality analysis tools. Nucleic Acids Research 24,4876-4882.
THORNE, G. (1943). Cacopaurus pestis, nov. gen., nov. spec.(Nematoda: Criconematinae), a destructive parasite of thewalnut, Juglans regia Linn. Proceedings of the Helmintho-logical Society of Washington 10, 78-83.
THORNE, G. (1949). On the classification of the Tylenchida,new order (Nematoda, Phasmidia). Proceedings of theHelminthological Society of Washington 16, 37-73.
THORNE, G. & ALLEN, M.W. (1944). Nacobbus dorsalis, nov.gen., nov. spec. (Nematoda: Tylenchidae) producing gallson the roots of alfileria, Erodium cicutarium (L.) L’Hér.Proceedings of the Helminthological Society of Washington11, 27-31.
WAEYENBERGE, L., MOENS, M., PINOCHET, J. & VRAIN,T.C. (2000). Molecular characterisation of Pratylenchusspecies using rDNA restriction fragment length polymor-phisms. Nematology 2, 135-142.
WINSLOW, R.D. (1958). The taxonomic position of Anguil-lulina obtusa Goodey, 1932 and 1940. Nematologica 3, 136-139.
Vol. 11(4), 2009 581
All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately.




















