
A new blunt-snouted dyrosaurid, Anthracosuchus balrogus gen. et sp. nov. (Crocodylomorpha,...
23
A new blunt-snouted dyrosaurid, Anthracosuchus balrogus gen. et sp. nov. (Crocodylomorpha, Mesoeucrocodylia), from the Palaeocene of Colombia Alexander K. Hastings a,b *, Jonathan I. Bloch a and Carlos A. Jaramillo c a Florida Museum of Natural History, University of Florida, Gainesville, FL, USA; b Geiseltalmuseum, Zentralmagazin Naturwissenschaftlicher Sammlungen, Martin Luther Universita ¨t Halle-Wittenberg, Halle (Saale), Germany; c Smithsonian Tropical Research Institute, Balboa-Ancon, Panama (Received 23 March 2014; accepted 24 April 2014) A new exceptionally brevirostrine dyrosaurid is described from the middle Palaeocene (58 – 60 million years ago) Cerrejo ´n Formation, northeastern Colombia, based on four partial skulls and associated postcrania. This taxon is unique among dyrosaurids not only in skull shape, but also in having orbital tuberosities, and osteoderms that are dorsoventrally thick and unpitted, a trait otherwise unknown in Crocodylomorpha. Results from a cladistic analysis of Dyrosauridae suggest that the new taxon, together with Cretaceous – Palaeocene Chenanisuchus lateroculi from Africa and Cerrejonisuchus improcerus also from the Cerrejo ´n Formation, are the most basal members of the family. Results from a biogeographic analysis indicate at least three independent dispersals of dyrosaurids from Africa to the New World occurred in the Late Cretaceous or early Palaeocene. Widely set orbits in the new taxon indicate a deviation from surface-based predation, characteristic of other dyrosaurids, to sub-surface predation, as in modern Gavialis. Tooth impressions found on turtle shells recovered from the same locality match well with teeth of the new taxon indicating possible predation. http://www.zoobank.org/urn:lsid:zoobank.org:pub:AB2B24A5-27CC-4D3F-B580-F11F17851CE6 Keywords: Dyrosauridae; Crocodylomorpha; Palaeocene; Colombia; Cerrejo ´n; biogeography Introduction Crocodylomorpha has long been considered a ‘living fossil’ group (McGregor 2005). This largely stems from the fact that they have displayed conservative morphology in maintain- ing at least some members within the same body plan as extant crocodylians since shortly after the group arose (Ross 1989). However, numerous instances of incredibly varied morphology have evolved time and again within the crocodylomorph lineage. Forms have varied from slender terrestrial ancestors (e.g. Sphenosuchus; Walker 1990), to heavily armoured forms (Armadillosuchus; Marinho and Carvalho 2009) to fully marine with paddled feet and short powerful jaws skulls (Dakosaurus; Gasparini et al. 2006). Most dyrosaurids are thought to have been relatively limited in ecological diversity, characterised as large near-shore marine longirostrine piscivores with some variation associated with juveniles possibly inhabiting freshwater environments (Jouve, Bardet, et al. 2008). Freshwater deposits (Jaramillo et al. 2007; Figure 1) of the middle Palaeocene-aged (58–60 million years ago) Cerrejo ´n Formation in northeastern Colombia have yielded not only plant fossils documenting the earliest evidence of a neotropical rainforest (Wing et al. 2009), but also vertebrate fossils including side-necked turtles (Cadena et al. 2010; Cadena, Bloch, et al. 2012; Cadena, Ksepka, et al. 2012), a giant boid snake (Head et al. 2009a, 2009b) and a diversity of dyrosaurids that expand our understanding of the ecological diversity of the group, with a previously published dwarf species (Cerrejonisuchus improcerus; Hastings et al. 2010) residing in freshwater habitats in adulthood, and a large- bodied longirostrine taxon (Acherontisuchus guajiraensis; Hastings et al. 2011) also in freshwater sediments deposited in a large river system that drained into what is now the Caribbean Sea. Dyrosauridae originated and diversified during the late Cretaceous, somehow survived the K – P extinction event that wiped out other large-bodied marine reptiles (Benson et al. 2010) and ultimately became more diverse in the early Cenozoic (Jouve, Bardet, et al. 2008). The ability of dyrosaurids to inhabit both marine and fresh water may have contributed to their preferential survival (Jouve, Bardet, et al. 2008). Isotopic values for carbon and oxygen may provide empirical evidence for a freshwater habitat, supporting this hypothesis (Wheatley 2010). Described here is a new genus and species of Dyrosauridae with a very different skull shape relative to other dyrosaurids, likely reflecting a unique ecology associated with the invasion of the freshwater ecosystem in tropical South America. The first skull of this new dyrosaurid (referred specimen UF/IGM 69) was discovered in 2005, but the skull was missing its anterior end. Preservation of the posterior external nares indicated that the individual likely q 2014 Taylor & Francis *Corresponding author. Email: [email protected] Historical Biology , 2014 http://dx.doi.org/10.1080/08912963.2014.918968 Downloaded by [University of Florida] at 08:16 26 May 2014
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of A new blunt-snouted dyrosaurid, Anthracosuchus balrogus gen. et sp. nov. (Crocodylomorpha,...

A new blunt-snouted dyrosaurid, Anthracosuchus balrogus gen. et sp. nov. (Crocodylomorpha,Mesoeucrocodylia), from the Palaeocene of Colombia
Alexander K. Hastingsa,b*, Jonathan I. Blocha and Carlos A. Jaramilloc
aFlorida Museum of Natural History, University of Florida, Gainesville, FL, USA; bGeiseltalmuseum, ZentralmagazinNaturwissenschaftlicher Sammlungen, Martin Luther Universitat Halle-Wittenberg, Halle (Saale), Germany; cSmithsonian TropicalResearch Institute, Balboa-Ancon, Panama
(Received 23 March 2014; accepted 24 April 2014)
A new exceptionally brevirostrine dyrosaurid is described from the middle Palaeocene (58–60 million years ago) CerrejonFormation, northeastern Colombia, based on four partial skulls and associated postcrania. This taxon is unique amongdyrosaurids not only in skull shape, but also in having orbital tuberosities, and osteoderms that are dorsoventrally thick andunpitted, a trait otherwise unknown in Crocodylomorpha. Results from a cladistic analysis of Dyrosauridae suggest that thenew taxon, together with Cretaceous–Palaeocene Chenanisuchus lateroculi from Africa and Cerrejonisuchus improcerusalso from the Cerrejon Formation, are the most basal members of the family. Results from a biogeographic analysis indicateat least three independent dispersals of dyrosaurids from Africa to the New World occurred in the Late Cretaceous or earlyPalaeocene. Widely set orbits in the new taxon indicate a deviation from surface-based predation, characteristic of otherdyrosaurids, to sub-surface predation, as in modern Gavialis. Tooth impressions found on turtle shells recovered from thesame locality match well with teeth of the new taxon indicating possible predation.
http://www.zoobank.org/urn:lsid:zoobank.org:pub:AB2B24A5-27CC-4D3F-B580-F11F17851CE6
Keywords: Dyrosauridae; Crocodylomorpha; Palaeocene; Colombia; Cerrejon; biogeography
Introduction
Crocodylomorpha has long been considered a ‘living fossil’
group (McGregor 2005).This largely stems from the fact that
they have displayed conservative morphology in maintain-
ing at least some members within the same body plan as
extant crocodylians since shortly after the group arose (Ross
1989). However, numerous instances of incredibly varied
morphology have evolved time and again within the
crocodylomorph lineage. Forms have varied from slender
terrestrial ancestors (e.g. Sphenosuchus; Walker 1990), to
heavily armoured forms (Armadillosuchus; Marinho and
Carvalho 2009) to fully marine with paddled feet and short
powerful jaws skulls (Dakosaurus; Gasparini et al. 2006).
Most dyrosaurids are thought to have been relatively limited
in ecological diversity, characterised as large near-shore
marine longirostrine piscivores with some variation
associated with juveniles possibly inhabiting freshwater
environments (Jouve, Bardet, et al. 2008). Freshwater
deposits (Jaramillo et al. 2007; Figure 1) of the middle
Palaeocene-aged (58–60 million years ago) Cerrejon
Formation in northeastern Colombia have yielded not only
plant fossils documenting the earliest evidence of a
neotropical rainforest (Wing et al. 2009), but also vertebrate
fossils including side-necked turtles (Cadena et al. 2010;
Cadena, Bloch, et al. 2012; Cadena, Ksepka, et al. 2012), a
giant boid snake (Head et al. 2009a, 2009b) and a diversity of
dyrosaurids that expand our understanding of the ecological
diversity of the group, with a previously published dwarf
species (Cerrejonisuchus improcerus; Hastings et al. 2010)
residing in freshwater habitats in adulthood, and a large-
bodied longirostrine taxon (Acherontisuchus guajiraensis;
Hastings et al. 2011) also in freshwater sediments deposited
in a large river system that drained into what is now the
Caribbean Sea.
Dyrosauridae originated and diversified during the late
Cretaceous, somehow survived the K–P extinction event
that wiped out other large-bodied marine reptiles (Benson
et al. 2010) and ultimately becamemore diverse in the early
Cenozoic (Jouve, Bardet, et al. 2008). The ability of
dyrosaurids to inhabit both marine and fresh water may
have contributed to their preferential survival (Jouve,
Bardet, et al. 2008). Isotopic values for carbon and oxygen
may provide empirical evidence for a freshwater habitat,
supporting this hypothesis (Wheatley 2010). Described
here is a new genus and species of Dyrosauridae with a very
different skull shape relative to other dyrosaurids, likely
reflecting a unique ecology associated with the invasion of
the freshwater ecosystem in tropical South America.
The first skull of this new dyrosaurid (referred
specimen UF/IGM 69) was discovered in 2005, but the
skull was missing its anterior end. Preservation of the
posterior external nares indicated that the individual likely
q 2014 Taylor & Francis
*Corresponding author. Email: [email protected]
Historical Biology, 2014
http://dx.doi.org/10.1080/08912963.2014.918968
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

had a short snout. A second skull with the anterior snout
preserved was discovered in early 2007, and indeed this
skull exhibited a very short snout while maintaining other
diagnostic characters of Dyrosauridae. Two more skulls
were discovered later in 2007 bringing the minimum
number of individuals discovered thus far to 4.
All specimens of the new taxon are deposited at the
Museo Geologico Jose Royo y Gomez, Colombian
Geological Survey, Bogota, Colombia. Each specimen
has been assigned a unique number using the abbreviation
UF/IGM. Casts of these specimens are also deposited at
the collections of the Florida Museum of Natural History
(UF).
Institutional abbreviations
IGM, Colombian Geological Survey, Bogota, Colombia;
NJSM, New Jersey State Museum, Trenton, NJ, USA; UF,
University of Florida, Florida Museum of Natural History,
Vertebrate Palaeontology Collection, Gainesville, FL,
USA; USGS SAP, US Geological Survey – Saudi Arabian
collection.
Terminology and anatomical abbreviations
Teeth and alveoli will be referred to by number with 1
being the most anterior. Premaxillary teeth and alveoli will
have the initial ‘pm’, maxillary teeth and alveoli will use
‘m’, and dentary teeth and alveoli will use ‘d’. For
example, the first tooth or alveolus of the premaxilla will
be referred to as ‘pm1’ and the second tooth or alveolus of
the maxilla will be referred to as ‘m2’. These follow the
abbreviations of Hastings et al. (2010).
Systematic palaeontology
Crocodylomorpha Walker, 1970
Crocodyliformes Hay, 1930
Mesoeucrocodylia Whetstone and Whybrow, 1983
Dyrosauridae de Stefano, 1903
Anthracosuchus balrogus gen. et sp. nov.
Etymology. Anthraco-, meaning coal, referring to the coal
mine in which the type locality is located; -suchus,
meaning crocodile. The specific term, balrogus, is named
from a literary beast discovered within a deep mine,
originally written by Tolkien (1954).
Diagnosis. Possesses the following apomorphies: (1) a
snout that is 44–53% of overall skull length; (2) rugose
orbital tuberosities anterior to medial margin of orbits; (3)
osteoderms that are dorsoventrally thick, unpitted and not
imbricated; (4) premaxillae that are wider than long and
(5) external nares that are wider than long.
Further differs from all other dyrosaurids in the
following unique combination of characters: (1) posterior
parietal margin straight; (2) interfenestral bar wide,
contributes to.10% of the skull table width, as measured
at anteroposterior midpoint of supratemporal fenestrae; (3)
8–10 maxillary teeth, of which 5–7 are antorbital; (4)
strong constriction at the premaxilla–maxilla suture; (5)
nasal–prefrontal and nasal–lacrimal sutures subequal in
Figure 1. (Colour online) Locality map and stratigraphiccolumn for locality where all known Anthracosuchus balrogusgen. et sp. nov. fossils are found. (A) Northern South Americawith star indicating Cerrejon fossil locality in northeasternColombia. (B) Stratigraphic column of Cerrejon Formation (fromJaramillo et al. 2007). Fossil locality of Anthracosuchus isunderclay below Coal Seam 90, indicated with arrow above.
2 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

length; (6) orbits laterally placed and (7) teeth that are
robust, round and weakly striated.
Holotype. UF/IGM 67, nearly complete skull from
premaxilla to occipital condyle (missing medial portion of
pterygoids and ventral braincase), nine ribs, seven
vertebrae and unidentifiable bone.
Paratype. UF/IGM 68, skull from partial premaxilla to
occipital condyle (missing pterygoids and ventral brain-
case), right articular, five osteoderms, five vertebrae, eight
ribs, distal pubis, distal ischium, haemal arch, three
isolated teeth, proximal phalanx, sacral rib and unidentifi-
able bone.
Referred specimens. UF/IGM 69, skull from peri-
orbital snout to posterior parietal (missing all but a
fragment of the pterygoids and the ventral braincase). UF/
IGM 70, skull from premaxilla to quadratic condyles
(missing pterygoids and ventral braincase).
Type locality and horizon. The holotype and all
known specimens of Anthracosuchus balrogus were
recovered from a single stratigraphic layer, broadly
exposed within the Cerrejon coal mine, Guajira Depart-
ment, northeastern Colombia. The fossils were discovered
in the underlying clay layer below Coal Seam 90
(Figure 1). The geologic setting is of fine-grained, grey
clay and mud with fragmented lignite interspersed
throughout (Jaramillo et al. 2007). The geologic setting
is indicative of a large fluvial floodplain, likely supporting
abundant large plant life. The age of the layer is middle-
late Palaeocene, 58–60 million years old (Jaramillo et al.
2007).
Comparative description
General. The rostrum is very stout and short in all
specimens of Anthracosuchus, representing between 44%
and 53% of the total skull length (Table 1). The rostrum of
Anthracosuchus is much shorter than that of any other
known dyrosaurid (Table 1). Cerrejonisuchus and
Chenanisuchus have much more longirostrine skulls
(Jouve, Bouya, et al. 2005; Hastings et al. 2010). The
anterior rostrum of Anthracosuchus is much wider than
that of the other short-snouted dyrosaurids, even in the
case of UF/IGM 68 that has a snout only 2.5% shorter than
Cerrejonisuchus (Hastings et al. 2010). The premaxilla
and external nares of Anthracosuchus are both wider than
long (Figures 2 and 3), a very atypical character for
Dyrosauridae. Even the relatively short-snouted forms of
Cerrejonisuchus and Chenanisuchus have anteroposter-
iorly elongate premaxillae and external nares (Jouve,
Bouya, et al. 2005; Hastings et al. 2010).
The overall shape of the skull is more square than
triangular, which is most apparent in the holotype
(Figure 2) and the paratype (Figure 3). The holotype
skull has experienced dorsoventral compression, but the
overall shape is similar in the uncompressed paratype,
indicating that compression did not significantly alter
these features. The skull table is flat and broad, comprising
most of the posterior half of the skull in dorsal view.
Again, this general feature does not appear to have been
significantly altered by compression due to its presence in
the uncompressed paratype (Figure 3). The wide, large and
flat skull table of Anthracosuchus (Figure 2) is similar to
that of Phosphatosaurus (Buffetaut 1978a), Sokotosuchus
Table 1. Snout proportions of all members of Dyrosauridae with material complete enough for skull length estimation.
DL (cm) PreoL (cm) R (PreoL/DL) £ 100 TBL (m)
Anthracosuchus balrogus (UF/IGM 67) 62 27.5 44.35 4.58Anthracosuchus balrogus (UF/IGM 68) 68 35.5 52.21 5.05Cerrejonisuchus improcerus (UF/IGM 31)a 31.4 17.2 54.78 1.72–2.22Cerrejonisuchus improcerus (UF/IGM 29)a 25.8 15.3 59.30 1.22–1.79Chenanisuchus lateroculib 57.6 36.5 63.37 3.57–4.24Acherontisuchus guajiraensis (UF/IGM 35)c 77.5–86.3 43.2/52.1 55.74–60.37 5.04–6.46Acherontisuchus guajiraensis (UF/IGM 34)c 72.4–78.7 41.9/50.8 57.87–64.52 4.66–5.87Congosaurus bequaerti d 63 41 65.08 3.97–4.66Guarinisuchus munizi e 47.1 30.8 65.41 2.79–3.43H. rogersiib 42.9 28.2 65.73 2.48–3.11Sokotosuchus ianwilsonib 60.1 39.7 66.06 3.75–4.44P. gavialoidesb 107 72 67.29 7.22–8.05Arambourgisuchus khouribgaensisf 100 71.5 71.50 6.71–7.51Dyrosaurus phosphaticusb 104 75 72.12 7.00–7.82Dyrosaurus maghribensisg 89–97 – 73.00 5.89–7.28R. keiniensisb 73.1 53.4 73.05 4.72–5.44Atlantosuchus coupatezih 104.5 83 79.43 7.04–7.86
Notes: DL, dorsal skull length; PreoL, preorbital skull length; R, ratio of preorbital skull length to dorsal skull length; TBL, estimated total body lengthusing method by Sereno et al. (2001). Citations are marked by superscript alphabets: a, Hastings et al. (2010); b, Jouve, Bouya, et al. (2005); c, Hastingset al. (2011); d, an estimation from Jouve and Schwarz (2004); e, an estimation from Figure 2 in Barbosa et al. (2008); f, Jouve, Iarochene, et al. (2005);g, Jouve et al. (2006); h, Jouve, Bouya, et al. (2008).
Historical Biology 3
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

(Buffetaut 1979), Chenanisuchus (Jouve, Bouya, et al.
2005) and Cerrejonisuchus as opposed to the elongate and
narrow skull table present in Hyposaurinae. The posterior
margin of the skull table in dorsal view is transversally
straight in Anthracosuchus (Figure 2), but highly indented
anteriorly in Rhabdognathus keiniensis (Jouve 2007),
Hyposaurus rogersii (Troxell 1925) and Atlantosuchus
coupatezi (Jouve, Bouya, et al. 2008).
Each of the skulls of Anthracosuchus possesses wide
and rugose tuberosities immediately anterior to the orbits.
These orbital tuberosities are most pronounced in
dorsolateral view in UF/IGM 69 (Figure 4) and appear to
be variable as to whether they are fully on the prefrontal,
lacrimal or split between the two (Figures 2 and 3). The
orbital tuberosities are dorsoventrally short and roughly
conform to the contour of the skull (Figure 4). The rugose
tuberosities near the anteromedial margins of the orbits,
visible in dorsal view (Figure 4), are unusual for
crocodyliforms. The only other known instance of
preorbital tuberosities within Crocodylomorpha is that of
Caryonosuchus pricei from the Late Cretaceous of Brazil,
but these are more akin to small horns (Kellner et al. 2011).
Alligatorids have highly rugose anteromedial orbital
margins, roughly where the palpebral attaches, but this is
not accompanied by surrounding unpitted surfaces as in
Anthracosuchus. These may have been keratinised, but
likely did not support large structures.
Cranial openings. The external nares are wider than
long in dorsal view and completely surrounded by the
premaxillae with lateral margins that are ventral to both
the anterior and posterior margins (Figures 2 and 3). The
external nares of all other dyrosaurids are anteroposter-
iorly elongate in dorsal view. No portion of the incisive
foramen is preserved on any specimen. The orbits of
Anthracosuchus are widely placed from the midline, in a
lateral position (Figures 2–4) like only one other
dyrosaurid, Chenanisuchus lateroculi from the Palaeocene
of Morocco (Jouve, Bouya, et al. 2005). All other
dyrosaurids have much more medially positioned orbits.
The supratemporal fenestrae of Anthracosuchus are very
large, much larger than the orbits and anteroposteriorly
elongate (Figures 2–4). The supratemporal fenestrae are
generally wider in dorsal view (Figures 2 and 4) than in
hyposaurine dyrosaurids, which have much more elongate
fenestrae. The infratemporal fenestrae of Anthracosuchus
are laterally placed, with minimal dorsal exposure and are
bound anteroventrally by the jugals and posterodorsally by
the quadratojugals (Figures 2–4). The infratemporal
Figure 2. (Colour online) Holotype (UF/IGM 67) of Anthracosuchus balrogus gen. et sp. nov. from Cerrejon locality in northeasternColombia, middle-late Palaeocene: (A, B) in dorsal view, (C, D) inventral view and (C) insert displays an in situ tooth in lingual and anteriorviews; scale bar: 1 cm. bf, bone fragment; bo, basioccipital; bsp, basisphenoid; en, external nares; eo, exoccipital; ep, ectopterygoid; f,frontal; j, jugal; l, lacrimal; max,maxilla; m1, 4, 5 and 8,maxillary teeth/alveoli; n, nasal; or t, orbital tuberosity; ost, osteoderm (displaced);o t, occipital tuberosities; p, parietal; pal, palatine; pm2–4, premaxillary teeth/alveoli; pmx, premaxilla; po, postorbital; prf, prefrontal; pt,pterygoid; q, quadrate; qj, quadratojugal; sq, squamosal; stf, supratemporal fenestra; vert, vertebra (displaced). Scale bar: 10 cm.
4 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

fenestrae are large in hyposaurine dyrosaurids, but more
transversally compressed and laterally placed in Anthra-
cosuchus, which is apparent in the uncompressed UF/IGM
68 (visible in dorsal view, Figure 3). The suborbital
fenestrae of Anthracosuchus are bordered by the palatines
and maxillae and are anteriorly concave in ventral view
(Figures 2–4).
Premaxilla. The premaxillae are wider than long and
completely surround the external nares, excluding the
nasal from contact with the external nares (Figures 2 and
3). All dyrosaurids except Anthracosuchus possess
premaxillae that are longer than wide. Anthracosuchus
instead has wide and stout premaxillae and a constriction
at the premaxilla–maxilla suture that is very pronounced
in both dorsal and ventral views (Figures 2 and 3), as in
Phosphatosaurus (Buffetaut 1978a), and very different
from the gentle curvature seen in hyposaurines such as
Dyrosaurus (Jouve 2005). The premaxilla of Anthraco-
suchus possesses four alveoli, although they are not well
preserved in any specimen (see ventral views of Figures 2
and 3). The third premaxillary tooth is by far the largest,
and the fourth premaxillary tooth appears to be severely
reduced. There is a strong indentation of the lateral margin
at the premaxilla–maxilla suture, noticeable in dorsal and
ventral views (Figures 2 and 3).
Maxilla. The maxilla makes broad contact with the
premaxilla, nasal and lacrimal in Anthracosuchus along its
dorsal snout (Figures 2–4). The maxilla broadly contacts
the palatine on the palatal surface in ventral view
(Figures 2–4). The maxilla of Anthracosuchus comprises
a similar proportion of the snout to other dyrosaurids, even
if its overall shape is very different. Eight maxillary alveoli
are preserved in ventral view of UF/IGM 67 and 68
(Figures 2 and 3). Aventral portion of the posterior maxilla
may be missing in all of the specimens, so a complete
count is not certain (Figures 2–4). The first and third
maxillary alveoli appear to be the largest (Table 2).
The fifth to seventh alveoli are smaller, but the eighth
alveolus appears to be more robust. Even the posteromost
alveoli appear uncompressed and retain the rounded shape
of the anterior alveoli. The tooth count is smaller than
most dyrosaurids. The tooth count of typical dyrosaurids
such as Dyrosaurus maghribensis is 19–21 (Jouve et al.
2006), as opposed to 8 in Anthracosuchus and 11 in
Cerrejonisuchus (Hastings et al. 2010). Most dyrosaurids
have approximately homodont teeth, but the maxilla of
Figure 3. (Colour online) Paratype (UF/IGM 68) of Anthracosuchus balrogus gen. et sp. nov. from Cerrejon locality in northeasternColombia, middle-late Palaeocene: (A, B) skull in dorsal view, (C) associated tooth in lingual view, (D) associated tooth in posterior viewand (E, F) skull in ventral view. bo, basioccipital; en, external nares; eo, exoccipital; eo a, articulation for exoccipital; ep, ectopterygoid; f,frontal; j, jugal; l, lacrimal; max, maxilla; m1–8, maxillary teeth/alveoli; n, nasal; orb, orbit; or t, orbital tuberosity; ot, occipitaltuberosities; p, parietal; pal, palatine; pm2–4, premaxillary teeth/alveoli; pmx, premaxilla; po, postorbital; prf, prefrontal; pt, pterygoid;q, quadrate; qj, quadratojugal; qc, quadratic condyle; sq, squamosal; stf, supratemporal fenestra. Scale bar for A–B and E–F equals10 cm. Scale bar for C–D equals 1 cm.
Historical Biology 5
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

Anthracosuchus, Cerrejonisuchus and Phosphatosaurus
(Buffetaut 1978a; Hastings et al. 2010) has notably
enlarged third maxillary teeth. In Anthracosuchus, the
maxillary margin is modestly sinuous (nonlinear) in dorsal
view (Figures 2 and 3). The maxilla of Anthracosuchus is
somewhat intermediate in terms of degree to which the
lateral margin undulates, or is ‘festooned’, in dorsal view
(Figures 2 and 3). In hyposaurines, the margin is linear, but
in phosphatosaurines, it is much more sinuous (Buffetaut
1979). Anthracosuchus instead has only a slight expansion
posterior to the premaxilla–maxilla suture.
Nasal. The nasal is variably fused in Anthracosuchus
in dorsal view. The nasals are unfused in UF/IGM 67
and 68 (Figures 2 and 3), but fully fused in UF/IGM 69
(Figure 4). The nasal is fused in most dyrosaurids,
including Dyrosaurus (Jouve 2005), Congosaurus (Jouve
and Schwarz 2004) and adult specimens of Cerrejoni-
suchus. However, Atlantosuchus coupatezi (Jouve,
Bouya, et al. 2008), Rhabdognathus aslerensis (Jouve
2007) and Sokotosuchus ianwilsoni (Buffetaut 1979) all
have unfused nasals. The largest two specimens of
Anthracosuchus have unfused nasals (UF/IGM 67 and
68; Figures 2 and 3), while the smaller UF/IGM 69 has
fully fused nasals (Figure 4), visible in dorsal view. The
bone is broad and medially placed in Anthracosuchus,
expanding from a narrow tip within the premaxilla and
extending posteriorly into a rather straight margin with
the frontal. The nasal does not reach the external nares
in dorsal view.
Lacrimal. The lacrimal forms much of the anterior
margin of the orbit in dorsal view and is anteroposteriorly
thin (Figures 2–4). The lacrimal–prefrontal contact
appears to be greater than the lacrimal–nasal contact.
The length of contact is similar between the prefrontal–
nasal and the lacrimal–nasal sutures. The prefrontal–
nasal suture is longer than the lacrimal–nasal suture in
Figure 4. (Colour online) Referred skull (UF/IGM 69) of Anthracosuchus balrogus gen. et sp. nov. fromCerrejon locality in northeasternColombia, middle-late Palaeocene: (A, B) skull in dorsal view, (C,D) skull in ventral view and (E, F) oblique view of right orbital tuberosity.f, frontal; j, jugal; l, lacrimal; max, maxilla; m2, 5 and 6, maxillary teeth/alveoli; n, nasal; orb, orbit; or t, orbital tuberosity; p, parietal;pal, palatine; pm2 and 3, premaxillary teeth/alveoli; pmx, premaxilla; po, postorbital; prf, prefrontal; sq, squamosal. Scale bar: 10 cm.
6 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

Dyrosaurus (Jouve et al. 2006), but the reverse is true of
Congosaurus (Jouve and Schwarz 2004). Anthracosuchus
appears to have an intermediate condition with roughly
equal proportions of contact to the nasal, in dorsal view
(Figures 2 and 3).
Prefrontal. The prefrontal contributes largely to the
medial margin of the orbit in dorsal view as well as to the
overall interorbital width in Anthracosuchus (Figures 2–
4). The prefrontal–frontal contact on the dorsal surface is
extensive and penetrates broadly into the frontal, rather
than being slender and reduced. The prefrontal–frontal
contact of Anthracosuchus in dorsal view is much longer
and extensive than the reduced state of most dyrosaurids
such as Congosaurus (Jouve and Schwarz 2004) and
Dyrosaurus (Jouve 2005), and most similar to Chenani-
suchus lateroculi (Jouve, Bouya, et al. 2005). Anthraco-
suchus is also most similar to Chenanisuchus lateroculi in
prefrontal contribution to interorbital width, which is
broad in both taxa.
Jugal. The jugal makes up the lateral margin of
the orbit, visible in dorsal view (Figures 2 and 3). The
postorbital bar is missing or obscured in each of the
specimens. The jugal extends posteriorly to its contact
with the quadratojugal in the posterolateral corner of the
infratemporal fenestra, visible in dorsal view (Figures 2
and 3). The jugal of Anthracosuchus is thicker and more
robust than that of all hyposaurine dyrosaurids (Troxell
1925; Jouve and Schwarz 2004; Jouve 2005, 2007; Jouve,
Iarochene, et al. 2005; Jouve et al. 2006; Jouve, Bouya,
et al. 2008; Barbosa et al. 2008).
Table 2. Measurements of alveoli for Anthracosuchus balrogus gen. et sp. nov.
Left Right
SpecimenAlveolarposition
Anteroposteriorlength
Medeolateralwidth
Alveolarposition
Anteroposteriorlength
Medeolateralwidth
UF/IGM 67 pm1 – – pm1 – –pm2 17.98 18.27 pm2 17.52 18.65pm3 25.92 27.01 pm3 27.58 25.04pm4 – – pm4 – –m1 29.47 32.55 m1 – –m2 – – m2 – –m3 – – m3 – –m4 16.70 26.92 m4 – –m5 – – m5 11.71 11.69m6 – – m6 – –m7 – – m7 – –m8 18.43 – m8 15.41 14.63
UF/IGM 68 pm1 – – pm1 – –pm2 – – pm2 – –pm3 14.49 22.83 pm3 – –pm4 – – pm4 – –m1 – – m1 20.21 19.26m2 – – m2 – –m3 26.19 21.76 m3 – –m4 14.04 12.61 m4 12.88 12.52m5 16.43 15.38 m5 – –m6 – – m6 21.49 20.56m7 – – m7 – –m8 – – m8 – –
UF/IGM 69 pm1 – – pm1 – –pm2 – 13.34 pm2 – –pm3 16.95 25.41 pm3 15.71 17.75pm4 – – pm4 – –m1 – – m1 – –m2 – – m2 – –m3 – – m3 – –m4 – – m4 – –m5 – – m5 – –m6 – – m6 – –m7 – – m7 – –m8 – – m8 – –
Notes: Alveolar walls that were either not preserved at all or not preserved well enough for measurement are denoted with a ‘–’. All measurements are inmillimetres. Abbreviations for the alveoli follow those of Hastings et al. (2010).
Historical Biology 7
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

Frontal. The frontal is roughly cruciform in dorsal
view, with an anterior projection penetrated by the paired
nasals in UF/IGM 67, slightly penetrated in UF/IGM 68
and with a fairly linear anterior connection to the nasals in
UF/IGM 69 (Figures 2–4). The frontal in most dyrosaurids
comes to a point anteriorly, bifurcating the nasals, which
can be found in Dyrosaurus maghribensis (Jouve et al.
2006) and Congosaurus bequaerti (Jouve and Schwarz
2004). However, Anthracosuchus has nasals that instead
slightly bifurcate the frontal, in dorsal view, at least in the
holotype (Figures 2). Alternatively, in Cerrejonisuchus
improcerus, the nasal terminates along a relatively straight
suture with the frontal, as in UF/IGM 69 (Figure 4). The
frontal contributes to the posteromedial margin of the orbit
and has a broad contact with the prefrontal. The left orbit
of UF/IGM 67 has been slightly modified by compression,
making the prefrontal appear to exclude the frontal from
the orbit (Figure 2), but the better preserved right orbit
shows the same relationship as the other two skulls, and
seems to more accurately reflect the character in
Anthracosuchus. The frontal is slightly ornamented and
forms a broad bar on the lateral side where it joins with the
postorbital on the posterior margin of the orbit. The frontal
contributes to about one-quarter of the overall dorsal
length of the interfenestral bar in UF/IGM 67 (Figure 2)
and closer to one-half in UF/IGM 69 (Figure 4).
Palatine. The palatine extends anteriorly on the palate
in ventral view to the level of the m3 in the holotype (UF/
IGM 67; Figure 2) but only to the m6 in UF/IGM 68
(Figure 3). The suture of UF/IGM 69 appears to reach as
far forward as m2, but the suture is difficult to identify in
this specimen (Figure 4). The palatine forms the wide bar
extending posteriorly from the palate towards the brain-
case and also forms the medial margin of the suborbital
fenestra in ventral view (Figures 2 and 3). The maxilla–
palatine suture occurs at the anteromedial corner of the
suborbital fenestra, with the palatine contributing very
minimally to the anterior margin. The palatine of
Dyrosaurus (Jouve et al. 2006) and Rhabdognathus
(Jouve 2007) meets the maxilla slightly more laterally
than in Anthracosuchus, coming to the anterior point of the
suborbital fenestra. No part of the choanal opening is
preserved in any of the described skulls.
Parietal. The parietal forms the largest contribution to
the skull table in dorsal view, which is ornamented with a
shallow divot on the triangular section between the
supratemporal fenestrae, posterior to the interfenestral bar
(Figures 2 and 3). The parietal contributes between half
and three-quarters to the dorsal length of the interfenestral
bar. The interfenestral bar is square in cross section, with a
slight overhang. The interfenestral bar of dyrosaurids is
typically T shaped and thin, as inDyrosaurus phosphaticus
(Jouve 2005) and Arambourgisuchus khouribgaensis
(Jouve, Iarochene, et al. 2005) or forms a thin sagittal
crest as in R. aslerensis (Jouve 2007). In Anthracosuchus,
Cerrejonisuchus improcerus and Chenanisuchus latero-
culi, the interfenestral bar is much more square shaped and
robust, but still with a bit of an overhanging lip onto the
supratemporal fenestra (Jouve, Bouya, et al. 2005). The
straight posterior parietal margin of Anthracosuchus in
dorsal view (Figures 2–4) is most similar to that of other
dyrosaurids such as Dyrosaurus (Jouve et al. 2006) and
Phosphatosaurus (Buffetaut 1978a), and differs from the
anteriorly indented margins of R. keiniensis (Jouve 2007)
and H. rogersii (Troxell 1925).
Postorbital. The postorbital forms the anterolateral
portion of the supratemporal fenestrae and does not appear
to contact the parietal on the dorsal surface in any of the
specimens (Figures 2–4). The anterolateral process is well
developed in UF/IGM 68 (Figure 3) and UF/IGM 69
(Figure 4), and modest in UF/IGM 67 (Figure 2).
Dyrosaurids typically have well-developed anterolateral
postorbital processes, as in Dyrosaurus phosphaticus
(Jouve 2005) but are reduced in Phosphatosaurus
gavialoides (Buffetaut 1978a) and practically non-existent
in Chenanisuchus lateroculi (Jouve, Bouya, et al. 2005).
The ventral portion of the postorbital is not preserved well
enough in any Anthracosuchus specimen to determine
whether it contacted the infratemporal fenestra anteriorly.
Squamosal. The squamosal forms the posterolateral
portion of the supratemporal fenestrae in dorsal view
(Figures 2–4). The squamosal and postorbital appear to
contribute roughly equally to the lateral bridges of the
skull table. The posterodorsal prong of the squamosal is
most pronounced in UF/IGM 68 and extends posteriorly to
the level of the posterior end of the occipital tuberosities
(Figure 3). However, the other two specimens have much
more reduced posterodorsal squamosal projections
(Figures 2 and 4). The degree of posterior projection of
the squamosals in Anthracosuchus (Figures 2–4) is most
similar to those of Dyrosaurus (Jouve et al. 2006), and not
nearly as pronounced and elongate as R. aslerensis
(Brochu et al. 2002; Jouve 2007).
Pterygoid. The pterygoid is flattened onto the brain-
case of UF/IGM 67, but clearly shows in ventral view a
well-developed wing extending laterally from the midline
(Figure 2). The pterygoidian wing of UF/IGM 67 is broad
and flat (Figure 2), and its relative thickness may be similar
to that of Dyrosaurus maghribensis (Jouve et al. 2006), but
preservation makes it difficult to discern. A fragment of
this wing is also preserved on the ventral surface of UF/
IGM 68 (Figure 3). The suture with the ectopterygoid
forms a distinct, posteriorly directed angle (Figure 2).
Ectopterygoid. The ectopterygoid is firmly sutured to
the pterygoid on the ventral surface and connects this bone
to the maxilla and jugal (Figure 2). The bone forms the
posterolateral portion of the suborbital fenestra in ventral
view. The ectopterygoid of UF/IGM 69 (Figure 4) does not
appear twisted as in Dyrosaurus maghribensis (Jouve et al.
2006) but is instead thick and linear.
8 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

Exoccipital. The exoccipital has wide lateral expansion
and constitutes much of the posterior surface of the
braincase. Due to oblique deformation of this part of the
skull, it is visible on the dorsal surface of UF/IGM 68
(Figure 3) and to a lesser degree on the ventral surface of
UF/IGM 67 (Figure 2). The exoccipital has a large
contribution of roughly one-third from each side to the
occipital condyle, which can be observed in dorsal view in
UF/IGM 68 (Figure 3). The occipital condyle of UF/IGM
67 is largely obscured in dorsal view and difficult to
discern exoccipital contribution (Figure 2). The exoccipi-
tals of all dyrosaurids contribute largely to the occipital
condyle (Jouve et al. 2006) and that of Anthracosuchus is
no exception. Well-developed, wide and flat occipital
tuberosities extend from the occipital surface, just ventral
to the skull table (Figures 2–4). The occipital tuberosities
are developed to varying degrees within Dyrosauridae.
Anthracosuchus is similar to Chenanisuchus and Sokoto-
suchus in having wide and flat tuberosities (Figure 2), but
differs from Rhabdognathus and Hyposaurus that have
elongate and narrow tuberosities (Jouve, Bouya, et al.
2005).
Basioccipital. The basioccipital forms most of the
wide occipital condyle, as seen in both dorsal and ventral
views (Figures 2–4). It has an oblong, smooth and rounded
surface with a strong ventral lip where the surface recurves
dorsally, best seen in ventral view. The basioccipital of
Anthracosuchus differs from that of Cerrejonisuchus in
being less arched dorsally and lacking the wide flat
anteroventrally directed tuberosity on the ventral surface
(Figure 2). No portion of the basioccipital tubera is
preserved in any specimen.
Basisphenoid. A small portion of the basisphenoid is
preserved in UF/IGM 67, but lacks any part of the
eustachian foramen in ventral view (Figure 2). It is situated
between the pterygoid and the exoccipitals and makes at
least some contact with the basioccipital. The basi-
sphenoid is not well preserved in Anthracosuchus, but does
appear to be mediolaterally narrow in ventral view
(Figure 2) as that of Dyrosaurus maghribensis (Jouve et al.
2006).
Quadratojugal. The quadratojugal reaches the postero-
most portion of the skull in dorsal and ventral views and
contributes at least one-quarter to the craniomandibular
joint (Figures 2 and 3), a similar capacity as Dyrosaurus
maghribensis (Jouve et al. 2006). This bone forms the
posteromedial margin of the infratemporal fenestra, in
dorsal view, and extends anteriorly to the postorbital,
ventral to the skull table (Figures 2 and 3).
Quadrate. The quadrate forms approximately three-
quarters of the craniomandibular condyle as well as much
of the braincase, seen in both dorsal and ventral views of
UF/IGM 67 and 68 (Figures 2 and 3). The quadrate
extends dorsally to at least the squamosal, but possibly the
postorbital in dorsal view (Figures 2 and 3). Quadratic
crests on the ventral surface are understated in
Anthracosuchus, if present at all (Figure 2), much like
that of Cerrejonisuchus improcerus, but differing from the
prominent crest in Arambourgisuchus khouribgaensis
(Jouve, Iarochene, et al. 2005), R. aslerensis (Brochu
et al. 2002) and R. keiniensis (Jouve 2007).
Dentition. The in situ teeth of UF/IGM 67 are wide,
blunt and at least one tooth has a well-preserved carina that
defines the labial and lingual surfaces (Figure 2). The
associated, yet unattached teeth preserved with UF/IGM
68 also have striations and well-developed carinae
(Figure 3). All associated teeth are blunt and low crowned.
The teeth vary in size, but all are roughly circular with
minimal labiolingual compression towards the posterior
alveoli (Figures 2 and 3). The dentition of Anthracosuchus
(Figures 2 and 3) is most similar to that described for
Phosphatosaurus (Buffetaut 1978a) in being round and
blunt, and differs from the elongate recurved teeth typical
of Hyposaurus (Denton et al. 1997), Dyrosaurus (Jouve
et al. 2006) and the anterior dentition of Acherontisuchus.
The teeth of Anthracosuchus are more low crowned and
not spade shaped as in the posterior dentition of
Cerrejonisuchus.
Articular. An isolated articular (but no other portion of
the mandible) was recovered with UF/IGM 68 (Figure 5).
The articular surface where it would fuse with the
surangular is preserved well, but much of the retroarticular
process is missing. Although the surangular is missing, due
to the location of articular–surangular suture marks along
the lateral margin of the articular, the surangular likely did
contribute to the articular surface. The lateral shelf of the
retroarticular process is well developed in dorsal view and
the glenoid fossa is smooth and deeply curved. The lateral
shelf of the retroarticular process forms an L shape in
dyrosaurids (Jouve, Bouya, et al. 2005) and is well
represented in UF/IGM 68 (Figure 5). The anteromedial
wing described for Dyrosaurus maghribensis (Jouve et al.
2006) and Congosaurus bequaerti (Jouve and Schwarz
2004) is also present in Anthracosuchus, although much
more rounded and robust (Figure 5).
Dorsal vertebrae. A total of seven dorsal vertebrae
were preserved with UF/IGM 67 (Figure 6). These appear
to be more or less from a continuous series from dorsal 4 to
dorsal 9, as well as a more posterior vertebra from around
the position of dorsal 16. Even the anteromost of these
vertebrae have fully fused neurocentral sutures, indicating
maturity (sensu Brochu 1996). The neurocentral sutures of
Anthracosuchus were likely straight prior to compression,
as in Hyposaurinae (Schwarz et al. 2006).
The hypapophysis of the dorsal 4 is robust, short and
the vertebra lacks a parapophysis, indicating a transition to
the rib cage (Figure 6). Dorsals 6–9 show clear attachment
points for ribs on the transverse processes, indicating
bifurcated attachment surfaces. The dorsal 4 vertebra of
Anthracosuchus bears a hypapophysis that is roughly half
Historical Biology 9
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

as high as the vertebral body, as described for
Hyposaurinae (Schwarz et al. 2006). Dorsals 6–9 may
have possessed the ventral keel on the centrum described
for Congosaurus (Schwarz et al. 2006), but preservation
makes this difficult to discern. The transverse processes of
Anthracosuchus dorsals 4 and 6–9 have faintly apparent
parapophyses and diapophyses, making this taxon more
similar to Dyrosaurus, Hyposaurus and Congosaurus than
to cf. Rhabdognathus, which has divergent parapophyses
and diapophyses until dorsal 5 (Schwarz et al. 2006). The
dorsal 16 vertebra shows no articular surface for ribs and
likely represents the lumbar region of the vertebral
column, posterior to the rib cage (Figure 6).
These vertebrae are all wider, larger and more robust
(Figure 6) than any described thus far for Dyrosauridae
(Schwarz et al. 2006). Vertebral series are best known for
Dyrosaurus, but have also been described for Rhabdog-
nathus (Langston 1995) and Congosaurus (Jouve and
Schwarz 2004). The neural spines are more mediolaterally
thick and anteroposteriorly wide than those of other
dyrosaurids, but not nearly as expanded as those of
Sarcosuchus (Sereno et al. 2001). Compression has made
it difficult to tell whether the transverse processes widen
posteriorly as in Hyposaurinae, but they do appear to
lengthen laterally (Schwarz et al. 2006). From what can be
determined from incomplete specimens, the neural spines
do appear to be of similar length, as in Hyposaurinae
(Schwarz et al. 2006).
Caudal vertebrae. Three caudal vertebrae were found
in association with UF/IGM 68 (Figure 6) and represent
positions between Caudal 28 and 32. One of these
specimens has the entire neural spine preserved, which is
very long and anteroposteriorly narrow in lateral view
(Figure 6). The distal tip of the neural spine does not
appear swollen or enlarged relative to the rest of the shaft.
The caudal vertebrae from this section of the tail typically
have a swelling at the distal tip in Dyrosaurus and
Congosaurus, while UF/IGM 68 does not (Figure 6). The
anterior and posterior centra of the caudal vertebrae are
quadrangular in cross section as well as their articular
facets (Figure 6) and appear very similar to those described
for Dyrosaurus and Congosaurus (Schwarz et al. 2006).
Cervical ribs. Two cervical ribs were discovered with
UF/IGM 68 (Figure 7) and represent positions around
articulation with cervicals 4–6. Although compressed, the
widely divergent tuberculum and capitulum, best seen in
medial view (Figure 7), indicate a relatively vertical
position of the cervical ribs in this portion of the neck. The
lateral surface is smooth, with an anterior surface that
likely imbricated with the adjacent anterior cervical rib.
The internal surface of the cervical ribs forms a concave
trough in Anthracosuchus (Figure 6) and in all hyposaurine
dyrosaurids (Schwarz et al. 2006).
Dorsal ribs.Most ribs associated with UF/IGM 67 and
68 are poorly preserved and only represent partial shafts
Figure 5. (Colour online) Associated articular bone of UF/IGM68: (A, B) in medial view, (C, D) in lateral view and (E, F) indorsal view. as sa, articular–surangular surface of articulation;gf, glenoid fossa; lsrap, lateral shelf of retroarticular process; rap,retroarticular process. Scale bar equals 10 cm.
10 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

with little to no discernible morphology. Two large dorsal
ribs were well preserved with UF/IGM 67 though and are
shown in Figure 7. These are large and robust, much like
the dorsal vertebrae, with thick dorsoventral shafts. These
have clearly distinct tubercula and capitula, indicating
positions around dorsals 7 and 8. The dorsal ribs of
Anthracosuchus (Figure 7) appear more thick and robust
than those figured for either Dyrosaurus or Congosaurus
(Schwarz et al. 2006). Anthracosuchus has dorsoventrally
tall ribs, as in all other described Dyrosauridae, and
both Congosaurus and Anthracosuchus have thickened
distal tips.
Haemal arch. The single-preserved haemal arch
associated with a skull (UF/IGM 68) is from the mid-
caudal section, with a likely position between the 11th and
15th caudal vertebrae (Figure 7). The vertebral attachment
points are split between anterior (13.6mm long) and
posterior facets (9.1mm long), indicating its position in
the tail section. The bone is 73.7mm long, is constricted in
lateral view ventral to the haemal canal (10.5mm wide)
and flares ventrally forming a spatulate shape (max ventral
width of 16.7mm). The haemal arch does not appear to
articulate with any of the three preserved caudal vertebrae.
The left and right articular surfaces do not appear as fully
fused in Anthracosuchus (Figure 7) as in Congosaurus
bequaerti (Jouve and Schwarz 2004) and seem most
similar to the unfused condition described for Rhabdog-
nathus sp. (Langston 1995).
Sacral rib. The second right sacral rib is preserved with
UF/IGM 68 (Figure 8). The iliac surface flares poster-
olaterally and appears to receive an anteroposteriorly
oriented ridge towards the posterior end of the pelvis. UF/
IGM 68 exhibits the roughly hourglass shape in ventral
view (Figure 8) typical of Dyrosauridae (Schwarz et al.
2006). As in Rhabdognathus sp. (USGS SAP 37-CR-1),
the medial surface articulates with the sacral vertebra, but
does not participate in the centrum (Langston 1995).
Ischium. The left distal ischium is preserved with UF/
IGM 68 (Figure 8). The anterior and posterior margins are
well preserved and do not indicate flaring in either
direction, but instead a relatively narrow termination. The
bone is also very dorsoventrally straight, with little to no
curvature laterally. A modest, laterally deflected crest
extends from the anterior distal edge, which is slightly
Figure 6. (Colour online) Vertebrae associated with skulls of Anthracosuchus balrogus gen. et sp. nov. (A, B) Three articulated dorsalvertebrae associated with the skull of UF/IGM 67, likely positions 6 to 8. (C, D) Dorsal vertebra associated with the skull of UF/IGM 67,likely position 16. (E, F) Dorsal vertebra associated with the skull of UF/IGM 67, likely position 4. (G, H) Anterior view of caudalvertebra associated with the skull of UF/IGM 68, likely position 28. (I) Same as (G), but in lateral view. (J) Lateral view of caudal vertebraassociated with the skull of UF/IGM 68, likely position 30. cen, centrum; hyp, hypopophysis; it, isolated tooth; nc, neural canal; ncs,traces of neurocentral suture; ns, neural spine; przy, prezygapophyses; pozy, postzygapophyses; tp, transverse process. Scale bar for A–F:10 cm; scale bar for G–J: 5 cm.
Historical Biology 11
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

laterally offset from the anterior margin, but then curves
slightly medially to form the anterior edge. The well-
preserved narrow distal end of the ischium of UF/IGM 68
(Figure 8) is consistent with the preserved partial ischium
of Acherontisuchus (Hastings et al. 2011) and interpret-
ations of H. rogersii (Troxell 1925), and less consistent
with Dyrosaurus (Jouve et al. 2006). While known ischia
of Acherontisuchus and Hyposaurus are incomplete, the
distal portion of the ischium of Anthracosuchus is
complete, possessing a more narrow and pointed distal
extremity than the anteroposteriorly long ventral margin of
Dyrosaurus maghribensis (Jouve et al. 2006).
Pubis. The right distal pubis was preserved with UF/
IGM 68 (Figure 8). The entire symphyseal surface that
would have joined the other pubis is preserved and reflects
an angle between the pubes at around 408, narrower thanthat seen in H. rogersii of North America (,508; Troxell1925). However, the overall distal pubic shape is still much
more similar between Anthracosuchus and Cretaceous
Hyposaurus of New Jersey than either is to Palaeocene–
Eocene Dyrosaurus or cf. Rhabdognathus of Africa
(Langston 1995). The pubis of Anthracosuchus is
remarkably flat, with very little arching.
Phalanx. A proximal phalanx was preserved with UF/
IGM 68 (Figure 9), likely pertaining to the pes, due to its
elongate form. The dorsal surface is tubular with a slightly
flattened lateral surface. The ventral (plantar) side has a
wide longitudinal groove along the bone’s centreline. The
articular facet is roughly ovular, with a width of 24.8mm
and height of 15.7mm. The proximal articular surface of
the phalanx of Anthracosuchus (Figure 9) appears more
ovular than does the triangular shape described for
hyposaurine dyrosaurids (Schwarz et al. 2006). The
preserved portion is 57.1mm long and lacks any part of the
distal condyle.
Osteoderms. Five osteoderms were found associated
with the skull of UF/IGM 68. The osteoderms of UF/IGM
68 are very thick and completely unpitted (Figure 10).
Both dorsal and ventral surfaces are smooth, with faint
grooves radiating from the centre, particularly evident
around the edges. The osteoderms swell to the thickest at
the middle and are not curved upward, but are either flat
ventrally or swell ventrally as well as dorsally. None of the
preserved osteoderms have any indication of an imbricat-
ing surface. These osteoderms are very different from any
other published dyrosaurid osteoderms (see Schwarz et al.
2006).
Dyrosaurids typically have thin, slightly dorsally
curved osteoderms with wide shallow pitting. Pitting is
consistent on dorsal, accessory and gastral osteoderms.
Pitting is unlikely to have been lost in Anthracosuchus due
to taphonomic weathering as more subtle features such as
cranial ornamentation were preserved with the same
specimen. Moreover, smaller Cerrejonisuchus osteoderms
from the same locality have been preserved with pitting
intact as well as a more typically dyrosaurid thin cross
section (Hastings et al. 2010). Hyposaurus rogersii is very
typical for a dyrosaurid in this respect and a representative
osteoderm is shown in Figure 10. Osteoderms may be a
little thicker in Congosaurus than other dyrosaurids (Jouve
and Schwarz 2004; Schwarz et al. 2006) but cross-
sectional diagrams of these in Schwarz-Wings et al. (2009)
still appear much thinner than those of Anthracosuchus.
Many dyrosaurid osteoderms are imbricated and have a
Figure 7. (Colour online) Ribs and haemal arch associated withskulls of Anthracosuchus balrogus gen. et sp. nov. (A) Lateralview of dorsal ribs associated with UF/IGM 67, likely positions 7and 8. (B) Medial view of cervical rib associated with UF/IGM68, likely between positions 4 and 6. (C) Same as in (B), in lateralview. (D) Posterior view of haemal arch associated with UF/IGM68, likely positions between 11 and 15. (E) Same as in (D), inlateral view. capm, capitulum; hc, haemal canal; inc ct, incisuracapitulotubercularis (capitulotubercular incision); p art f,posterior articular facet; tubm, tuberculum.
12 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

thick articular band where one osteoderm overlaps the
next-most posterior one.
The osteoderms of Anthracosuchus are not only
atypical for Dyrosauridae, but they are highly unusual for
Crocodylomorpha, or any armoured vertebrate. Osteo-
derms from all parts of the body are known in most
crocodyliform groups, but none resemble the unpitted and
thickened form seen in Anthracosuchus. Even outside of
Crocodylomorpha, osteoderms are well known in testu-
dines, but are not as square, inflated and untextured as
these (AKH). Osteoderms in sauropods are much more
conical or shaped like a thickened disc, and not rectangular
(Dodson et al. 1998). Somewhat similar forms are seen in
dorsal osteoderms of ankylosaurs (Burns 2008), but not
nearly as square as in UF/IGM 68. Osteoderms in
xenarthrans such as armadillos and glyptodonts have
surface patterns and also have very different overall shapes
(Hill 2006).
Phylogenetic analysis
Relationship to other dyrosaurids
The morphologic data generated by Hastings et al. (2010,
2011) were combined with new data for Anthracosuchus to
better understand the relationships of the Cerrejon taxa
within Dyrosauridae (Table 3). A cladistic analysis was
conducted using branch and bound searches with the
program PAUP version 4.0b10 (Swofford 2003). The data-
set was small enough that branch and bound searches were
practical, while exhaustive searches (which are more
comprehensive) were far too computationally intensive.
The cladistics analysis was rooted with Sarcosuchus
imperator, Elosuchus cherifiensis and Terminonaris
Figure 8. (Colour online) Portions of the pelvis of Anthracosuchus balrogus gen. et sp. nov., UF/IGM 68. (A) Distal left ischium inlateral view. (B) Distal left ischium in posterior view. (C) Distal right pubis in ventral view. (D) Distal right pubis in medial (articular)view. (E) Diagram showing appropriate position of ischium and pubis in ventral view with muscle attachments overlaid, outline indicatesposition in the body. (F) Second right sacral rib in ventral view. (G) Same as (F), in lateral view. ant, anterior; art pub, articular surface forattachment to other pubis bone; f sym il, facies symphysialis ilii (articular surface of the sacral rib with the ilium); gas, gastralia; is,ischium; isc, ischiocaudalis muscle; isp, ischiopubis muscle; ist, ischiotruncus muscle; pub, pubis; pub car, pubic cartilage; r ab, rectusabdominis muscle. Scale bar for A–D and E equals 10 cm; scale bar for F–G equals 5 cm.
Figure 9. (Colour online) Proximal phalanx of Anthracosuchusbalrogus gen. et sp. nov. UF/IGM 68. (A) Dorsal view, (B)plantar view and (C) proximal view. Scale bar for A–B equals5 cm; scale bar for C equals 1 cm.
Historical Biology 13
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

robusta treated as outgroup taxa. The characters were
ordered as in Hastings et al. (2010, 2011) and multistate
codings were treated as uncertain. For the initial analysis,
we included all 14 ingroup taxa, as in Hastings et al.
(2011), with the addition of the new taxon described in this
study (Table 3).
With all taxa included, the initial results are presented
in a strict consensus cladogram of 44 trees (Figure 11).
This topology is not well resolved, with a large polytomy
including all 15 dyrosaurid species, i.e. the ingroup. Some
resolution was recovered with a monophyletic Rhabdog-
nathus and Dyrosaurus, and a pairing of Sokotosuchus
ianwilsoni and P. gavialoides, but no other relationships
were retained.
The initial results revealed two wildcard taxa,
Congosaurus and Acherontisuchus. These two taxa
were recovered as wildcard taxa in the analysis of
Hastings et al. (2011) as well. An Adams consensus
cladogram placed Acherontisuchus at an unresolved
polytomy at the base of Dyrosauridae with Chenani-
suchus and a monophyletic group including the 13 other
species of Dyrosauridae. Congosaurus placed as sister to
a clade uniting a monophyletic Rhabdognathus and
Atlantosuchus þ Guarinisuchus. The reason for the
unresolved status of these two taxa is that each has
characters that are both derived and primitive and that
there are not enough coded characters to discern which
is more prevalent in the taxon. Acherontisuchus coded
primitively for characters: 1 (snout ,68% of dorsal skull
length), 72 (minimal occlusal pits) and 74 (symphysis
wider than high). This taxon also possessed the derived
state for characters: 23 (linear maxillary margin), 25
(alveolar walls level with maxillary surface) and 71
(mandibular symphysis ends posterior to anterior 3/4
alveoli). This character conflict, combined with only
18.3% of the characters coded for the taxon, resulted in
poor resolution within Dyrosauridae. Acherontisuchus is
known from fragmentary cranial remains and mandibles
(Hastings et al. 2011), and Congosaurus is known only
from the snout section of the skull and not from the
braincase (Jouve and Schwarz 2004). Despite the lack of
material, an approximation of the snout-to-skull length
ratio of Congosaurus was determined from what cranial
material was preserved (Jouve and Schwarz 2004) and
from two nearly complete mandibles of Acherontisuchus
(Hastings et al. 2011) (see values in Table 1).
Congosaurus too possessed primitive and derived
characters. The primitive states were characters: 1
Figure 10. (Colour online) Dyrosaurid osteoderms. (A, C) Dorsal osteoderm, UF/IGM 68, in dorsal view; (B, D) same specimen as (A, C)in ventral view; (E) another dorsal osteoderm, UF/IGM 68, in dorsal view; (F) same specimen as (E), in ventral view; (G) lateral osteoderm,UF/IGM68, in dorsal view; (H) same specimen as G, in ventral view; (I) dorsal osteoderm,UF/IGM68 in dorsal view; (J) same specimen as(I), in ventral view; (K) same specimen as (I), in cross-sectional view at fracture point visible in (I) and (J); (L) osteoderm of H. rogersii,NJSM 12293, in dorsal view; (M) same specimen as (L), in ventral view; (N) Same specimen as (L), in lateral view. Scale bar equals 5 cm.
14 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

(snout ,68% of dorsal skull length), 3 (thick anterior
margin of external nares) and 14 (absence of lateral
expansion of premaxilla). Congosaurus possessed more
derived characters: 4 (medially positioned orbits), 13
(posterodorsal premaxillary process proximal/anterior to
second alveolus), 22 (premaxilla nearly three times
longer than wide), 23 (linear maxillary margin), 33
(nasal ceases posterior to first maxillary tooth) and 81
(alveoli more widely spaced in posterior versus anterior
snout).
When the two wildcard taxa are removed from the
analysis, the result is a single-most parsimonious
cladogram (Figure 11). This topology shows Chenanisu-
chus lateroculi from Africa as the most primitive
dyrosaurid. The next most basal dyrosaurids are
Anthracosuchus balrogus and Cerrejonisuchus impro-
cerus, respectively. The next most basal clade includes H.
rogersii as the sister taxon to Sokotosuchus ianwilsoni and
P. gavialoides. Arambourgisuchus khouribgaensis is the
sister taxon to a clade that includes a monophyletic
Dyrosaurus, and another nested clade that includes
Atlantosuchus coupatezi þ Guarinisuchus munizi and a
monophyletic Rhabdognathus. For Figure 11, the wildcard
taxa have been given tentative positions based on the
analysis from Hastings et al. (2011), from a 50% majority
rule consensus cladogram where they were placed within
a polytomy with the clade including Atlantosuchus þGuarinisuchus and a monophyletic Rhabdognathus, a
clade also recovered in the current study. As Hyposaurus
was recovered as more primitive than the analysis of
Hastings et al. (2011), it is not presented as part of this
polytomy. Instead, the monophyletic Dyrosaurus is
retained in its position as sister to the clade including
Atlantosuchus þ Guarinisuchus and a monophyletic
Rhabdognathus. The positions of Acherontisuchus and
Congosaurus are denoted with a dashed line to indicate
that they were not part of this particular analysis, but are
included to provide an approximation of where they likely
fit within Dyrosauridae.
Three characters unambiguously support the clade
including all dyrosaurids except the shortest-snouted
genera Chenanisuchus, Anthracosuchus and Cerrejonisu-
chus (characters 7, 8 and 71; see snout proportions in
Table 1). These characters are a narrow interfenestral bar,
an unornamented interfenestral bar and a mandibular
symphysis that ends posterior to the anterior three-quarters
of the alveoli. The pairing of Hyposaurus with
Sokotosuchus and Phosphatosaurus has not been recov-
ered in previous phylogenetic analyses. In this analysis,
the only unambiguous synapomorphy supporting this
clade is character 45, a reversal to state 0, which is the
presence of deep pits on the dorsal surface of the
parietal. Character 47, a pronounced anterolateral
process of the postorbital, unites all dyrosaurids except
Chenanisuchus.Table
3.
Morphologic
characterstates
ofAnthracosuchusbalrogusgen.et
sp.nov.followingthecharactermatrixofHastingset
al.(2010).
Anthracosuchus
balrogusgen.
etsp.nov.
00
00
00
00
01
11
11
11
11
12
22
22
22
22
23
33
33
33
33
34
23
33
33
33
33
34
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
90
12
34
56
78
90
11
01
01
00
10
0?
11
??
?1
?1
10
10
0A
11
BB
00
1?
?0
00
10
BB
00
1?
?0
00
10
Anthracosuchus
balrogusgen.
etsp.nov.
44
44
44
44
45
55
55
55
55
56
66
66
66
66
67
77
77
77
77
78
88
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
12
?0
1?
20
10
??
?1
2?
??
0?
?1
??
??
??
??
??
??
??
??
?1
?0
??
Notes:Multistate
characters
aretreatedas
uncertain.Fornexusfile,seeElectronic
Supplement.A¼
1/2;B¼
0/1.
Historical Biology 15
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

Relationship of Dyrosauridae to other crocodyliforms
In many phylogenetic studies of crocodyliforms, long-
irostrine taxa group together despite instances of clear
convergence (Clark 1994). Characters largely tied to
longirostry most often yield a close phylogenetic
relationship (Jouve et al. 2006). A long-standing problem
has been the largely Jurassic-aged thalattosuchians pairing
with the Cretaceous pholidosaurids and Cretaceous–
Eocene dyrosaurids (Jouve et al. 2006; Pol and Gasparini
2009; de Andrade et al. 2011).
The most primitive dyrosaurids in the analysis above
were also the shortest snouted. We carried out an
additional phylogenetic analysis to test the hypothesis
that longirostrine thalattosuchians are much more
primitive within Crocodyliformes than other analyses
suggest and that inclusion of the new brevirostrine
dyrosaurids will result in a more disparate relationship
between Dyrosauridae and Thalattosuchia. This crocodyli-
form analysis utilised a cladistic data-set that included
representatives from all major lineages of Crocodyli-
formes (Jouve et al. 2006) and added to it the new short-
snouted Anthracosuchus balrogus and Cerrejonisuchus
improcerus. The morphological matrix of Jouve et al.
(2006) was chosen because 234 characters were coded
across a large sampling of Crocodyliformes (n ¼ 47) that
included the dyrosaurids Dyrosaurus and Chenanisuchus.
Although more recent studies have also investigated this
problem thoroughly (e.g. Pol and Gasparini 2009; de
Adrade et al. 2011), these studies included only long-
irostrine dyrosaurids, excluding the short-snouted Chena-
nisuchus.
The two Cerrejon dyrosaurids were coded and added to
the matrix (Table 4) that was then run with a heuristic
search with 10,000 repetitions and the random seed
function in PAUP version 4.0b10 (Swofford 2003). The
Figure 11. Cladograms resulting from a phylogenetic analysis of Dyrosauridae. Left cladogram is a strict consensus with all namedspecies known from more than dentary fragments. Right cladogram represents a single cladogram resulting from analysis with the twowildcard taxa removed (Acherontisuchus guajiraensis and Congosaurus bequaerti). In the right cladogram, dotted lines for the wildcardtaxa represent approximate placement based on cladistic analysis of Hastings et al. (2011). The right cladogram is placed in stratigraphicand geographic context. Dates were obtained from Gradstein et al. (2004). CI, consistency index; RI, retention index; RC, rescaledconsistency index; HI, homplasy index.
16 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

Table
4.
CladisticdataforAnthracosuchusbalrogusgen.et
sp.nov.andCerrejonisuchusimprocerususedin
theanalysisbyJouveet
al.(2006).
11
11
11
11
11
22
22
22
22
22
31
23
45
67
89
01
23
45
67
89
01
23
45
67
89
0Anthracosuchusbalrogus
10
?0
2/3
00
11
30
10
??
??
??
??
??
?1
10
11
3Cerrejonisuchusimprocerus
10
?0
20
01
13
01
10
11
??
??
??
?0
11
01
13
33
33
33
33
34
44
44
44
44
45
55
55
55
55
56
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
Anthracosuchusbalrogus
01
00
02
00
?0
00
00
12
01
10
11
??
0?
?1
??
Cerrejonisuchusimprocerus
01
00
02
00
?0
00
00
12
01
10
11
??
0?
?1
??
66
66
66
66
67
77
77
77
77
78
88
88
88
88
89
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
Anthracosuchusbalrogus
?1
10
2?
–0
1?
0?
?0
??
??
0?
??
01
01
01
00
Cerrejonisuchusimprocerus
?1
10
2?
–0
1?
01
?0
??
00
01
0?
01
01
11
00
11
11
11
11
11
11
11
11
11
11
19
99
99
99
99
00
00
00
00
00
11
11
11
11
11
21
23
45
67
89
01
23
45
67
89
01
23
45
67
89
0Anthracosuchusbalrogus
01
?1
1?
?0
?1
0?
?1
??
?1
12
2?
00
00
??
0?
Cerrejonisuchusimprocerus
01
11
10
10
01
0?
?1
0?
?1
11
20
00
00
??
??
11
11
11
11
11
11
11
11
11
11
11
11
11
11
11
22
22
22
22
23
33
33
33
33
34
44
44
44
44
45
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
Anthracosuchusbalrogus
01
1?
00
10
0?
1?
??
??
??
0?
00
11
??
?1
??
Cerrejonisuchusimprocerus
??
??
??
??
0?
11
??
?2
10
00
00
1?
??
??
?1
11
11
11
11
11
11
11
11
11
11
11
11
11
11
11
55
55
55
55
56
66
66
66
66
67
77
77
77
77
78
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
Anthracosuchusbalrogus
??
0?
1?
00
??
??
??
??
??
??
??
??
??
??
??
Cerrejonisuchusimprocerus
01
01
1?
??
??
1?
01
??
??
??
?2
??
??
??
??
11
11
11
11
11
11
11
11
11
12
22
22
22
22
22
88
88
88
88
89
99
99
99
99
90
00
00
00
00
01
12
34
56
78
90
12
34
56
78
90
12
34
56
78
90
Anthracosuchusbalrogus
?1
1?
00
0/1
00
10
00
00
00
??
1?
??
??
??
??
1Cerrejonisuchusimprocerus
?1
00
00
10
10
00
00
00
0?
11
??
??
??
??
?1
22
22
22
22
22
22
22
22
22
22
22
22
11
11
11
11
12
22
22
22
22
23
33
33
12
34
56
78
90
12
34
56
78
90
12
34
Anthracosuchusbalrogus
??
??
??
??
??
??
0?
?1
0?
??
1?
??
Cerrejonisuchusimprocerus
??
??
00
??
??
??
0?
?1
1?
??
1?
??
Note:A
totalof234characters
werecoded
andtheother
47taxausedin
thepreviousanalysisremained
unchanged
inthecurrentanalysis.Fornexusfile,seeElectronic
Supplement.
Historical Biology 17
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

result was 30 equally most parsimonious cladograms
(Figure 12), as opposed to 124 in the heuristic search of
Jouve et al. (2006). The overall results were similar to
those of Jouve et al. (2006). Thalattosuchians shared a
close relationship with dyrosaurids and pholidosaurids. All
four dyrosaurids were monophyletic, although the only
internal resolution had Cerrejonisuchus and Anthracosu-
chus as sister taxa. Despite the short-snouted dyrosaurids
being present in the analysis, the resultant topology
was very similar to other studies addressing the
thalattosuchian issue (e.g. Pol and Gasparini 2009; de
Adrade et al. 2011).
Following Jouve et al. (2006), the same data-set was
used for another heuristic search with 10,000 replicates
with the 15 characters suggested as associated with
longirostry removed (characters 5, 7, 8, 12, 15, 30, 46, 47,
68, 83, 103, 150, 161, 172 and 189). The result of this
analysis was very similar to the heuristic search of Jouve
et al. (2006), with Thalattosuchia having a more basal
position within Crocodyliformes, basal to nearly all of
Mesoeucrocodylia (Figure 12). Pholidosauridae and
Dyrosauridae were united in a node, but different from
the previous analysis in that Elosuchus had an unresolved
relationship in a polytomy including Dyrosauridae and a
clade that includes all other pholidosaurids. Jouve et al.
(2006) found Elosuchus as primitive to both Pholidosaur-
idae and Dyrosauridae. Pol and Gasparini (2009) studied
the correlation between longirostrine characters and the
thalattosuchian phylogenetic issue. They concluded from
their analyses that because these characters are not
uniformly found in longirostrine taxa, they are in fact
informative of the phylogenetic evolution of the taxa. In
the present study, the addition of short-snouted dyrosaurids
did not resolve the problem of convergence driving the
topology between thalattosuchians, dyrosaurids and
pholidosaurids. These results suggest that these convergent
characters are not limited to longirostry and are present
even in non-longirostrine taxa.
Figure 12. Two cladograms from analyses utilising a matrix with representatives across Crocodylomorpha. (A) Strict consensuscladogram resulting from analysis using all 234 characters of Jouve et al. (2006). (B) Strict consensus cladogram resulting from analysisthat omitted the 15 characters associated with longirostry. Note the very different placement of Thalattosuchia with respect toDyrosauridae in the two analyses. CI, consistency index; RI, retention index; RC, rescaled consistency index; HI, homplasy index.
18 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

Biogeography
In order to discern possible methods of dispersal within
Dyrosauridae, a biogeographic analysis was run on the
dyrosaurid matrix (Table 3) using the program S-DIVA
(Yu et al. 2010a), which utilises a tree file as generated by
PAUP (Swofford 2003). We assigned each taxon to a
region: Africa, North America and South America,
assigned as regions A, B and C, respectively (Figure 13).
The program generates a consensus cladogram from the
tree file, incorporating the biogeographic region of the taxa
included (Yu et al. 2010b). The program generates likely
regions of origin for the ancestors of the taxa included in
the analysis. The program creates a pie chart at each of
these nodes with per cent probability by region for the
ancestor. The program can be set to allow for multiple
regions to be occupied simultaneously.
Four limitations of the software should be kept in
mind. The first is that the program from which it was
derived (DIVA; Ronquist 1996) treats dispersal between
different regions as equally possible, with no preference
for a certain direction or between certain regions
(Kodandaramaiah 2010). The second is that the program
must assign a topology with no polytomies. As a result,
taxa with unresolved relationships in non-biogeographic
studies should be interpreted with caution when using S-
DIVA (Yu et al. 2010b). For this reason, both wildcard
taxa recognised in the earlier analysis were not included.
The third limitation is due to S-DIVA utilising phylogeny
to determine dispersal; taxonomically uninformative
fossils cannot be included. Thus, the present analysis and
discussion are limited to identifiable taxa known from
nearly complete fossil skulls and do not take into account
regions where dyrosaurid fossils were found that could not
be diagnosed to genus. Finally, the program cannot
incorporate temporal data, and timing of dispersal must be
determined using ages of the fossil taxa.
All past phylogenetic studies of Dyrosauridae have
recovered an African origin for the family, and the current
analysis supports this as well. The biogeographic analysis
recovered five optimised dispersal histories, with a
minimum of three dispersal events from Africa to the
New World within Dyrosauridae. The first of these
occurred with the ancestor of Anthracosuchus to South
America (Figure 14). The second dispersal occurred with
the ancestor of Guarinisuchus munizi reaching eastern
Brazil. The third independent trans-Atlantic dispersal
would have been the ancestor ofH. rogersii immigrating to
North America. Had Acherontisuchus guajiraensis been
included, it would likely have represented a fourth
dispersal, as its relationships were most often close to
Arambourgisuchus and Dyrosaurus of Africa and did not
have a direct relationship with Guarinisuchus or any other
Figure 13. Cladogram resulting from biogeographic analysis of Dyrosauridae using the program S-DIVA (Yu et al. 2010a). Solid arrowsindicate dispersal events. The dotted line indicates a possible back dispersal from South America to Africa.
Historical Biology 19
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014
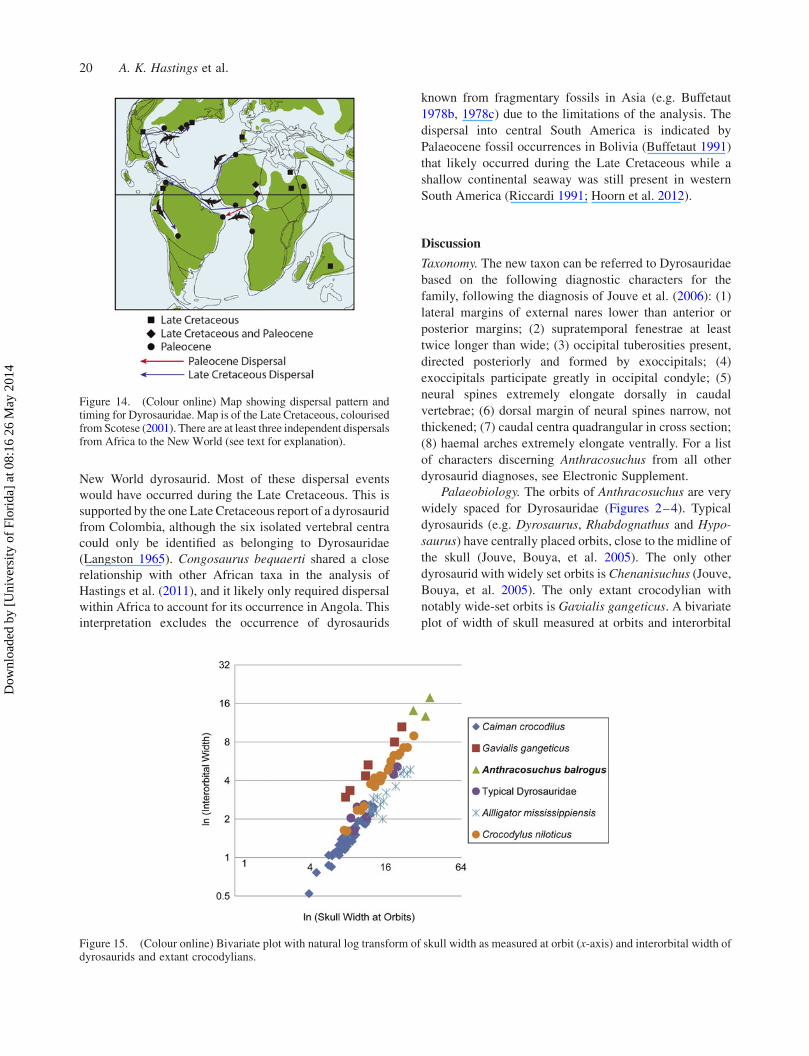
New World dyrosaurid. Most of these dispersal events
would have occurred during the Late Cretaceous. This is
supported by the one Late Cretaceous report of a dyrosaurid
from Colombia, although the six isolated vertebral centra
could only be identified as belonging to Dyrosauridae
(Langston 1965). Congosaurus bequaerti shared a close
relationship with other African taxa in the analysis of
Hastings et al. (2011), and it likely only required dispersal
within Africa to account for its occurrence in Angola. This
interpretation excludes the occurrence of dyrosaurids
known from fragmentary fossils in Asia (e.g. Buffetaut
1978b, 1978c) due to the limitations of the analysis. The
dispersal into central South America is indicated by
Palaeocene fossil occurrences in Bolivia (Buffetaut 1991)
that likely occurred during the Late Cretaceous while a
shallow continental seaway was still present in western
South America (Riccardi 1991; Hoorn et al. 2012).
Discussion
Taxonomy. The new taxon can be referred to Dyrosauridae
based on the following diagnostic characters for the
family, following the diagnosis of Jouve et al. (2006): (1)
lateral margins of external nares lower than anterior or
posterior margins; (2) supratemporal fenestrae at least
twice longer than wide; (3) occipital tuberosities present,
directed posteriorly and formed by exoccipitals; (4)
exoccipitals participate greatly in occipital condyle; (5)
neural spines extremely elongate dorsally in caudal
vertebrae; (6) dorsal margin of neural spines narrow, not
thickened; (7) caudal centra quadrangular in cross section;
(8) haemal arches extremely elongate ventrally. For a list
of characters discerning Anthracosuchus from all other
dyrosaurid diagnoses, see Electronic Supplement.
Palaeobiology. The orbits of Anthracosuchus are very
widely spaced for Dyrosauridae (Figures 2–4). Typical
dyrosaurids (e.g. Dyrosaurus, Rhabdognathus and Hypo-
saurus) have centrally placed orbits, close to the midline of
the skull (Jouve, Bouya, et al. 2005). The only other
dyrosaurid with widely set orbits is Chenanisuchus (Jouve,
Bouya, et al. 2005). The only extant crocodylian with
notably wide-set orbits is Gavialis gangeticus. A bivariate
plot of width of skull measured at orbits and interorbital
Figure 14. (Colour online) Map showing dispersal pattern andtiming for Dyrosauridae. Map is of the Late Cretaceous, colourisedfromScotese (2001). There are at least three independent dispersalsfrom Africa to the New World (see text for explanation).
Figure 15. (Colour online) Bivariate plot with natural log transform of skull width as measured at orbit (x-axis) and interorbital width ofdyrosaurids and extant crocodylians.
20 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

width (with natural log transform) revealed that Anthra-
cosuchus had greater similarity in terms of orbital spacing
with Gavialis than with typical dyrosaurids, Alligator
mississippiensis, Caiman crocodilus or Crocodylus nilo-
ticus (Figure 15). Extant Gavialis typically acquire their
prey, almost exclusively fish, below the water surface
(Thorbjarnarson 1990; Grenard 1991). The three extant
species with more narrowly placed orbits more commonly
acquire prey from a position at the surface of the water
(Grenard 1991; Thorbjarnarson 1993). More widely set
orbits may be tied to a non-surficial prey acquisition
feeding strategy and likely indicate a deviation by
Anthracosuchus and Chenanisuchus from the typical
behaviour of dyrosaurids. This subsurface predation
method may also be inferred in the highly marine
metriorhynchid crocodyliforms, which have laterally
placed orbits (Pol and Gasparini 2009).
The distal pubis is rarely recovered for New World
dyrosaurids, with the only other figured specimen from H.
rogersii from the Late Cretaceous marine deposits of New
Jersey (Troxell 1925). Both have a more elongate suture
between left and right pubes as compared to extant
crocodylians. That of Anthracosuchus (Figure 8) is
proportionally longer than that of H. rogersii. The pubic
cartilage attaches to the anterior portions of these bones in
extant crocodylians, and the symphyseal region is where
the ischiopubis and ischiotruncus muscles attach (Uriona
and Farmer 2008; Schwarz-Wings et al. 2009). A non-
enlarged distal ischium, also preserved with UF/IGM 68,
indicates highly reduced attachment surfaces for the
ischiopubis, ischiotruncus and ischiocaudalis muscles
(Figure 8). The ischiopubis muscle in particular is used
primarily in buoyancy control (Uriona and Farmer 2008).
Much like the implications for Acherontisuchus (Hastings
et al. 2011), it would appear Anthracosuchus also relied
less on these muscles for pitch control within the water.
The pubis is heavily utilised in extant crocodylian
respiration and the bone is pulled ventrally and rotated
slightly by the rectus abdominus and diaphragmaticus
muscles, bringing air into the lungs (Claessons 2004). The
pubic cartilage is what dominantly attaches these two bones
in extant crocodylians (Uriona andFarmer 2008). The longer
and more rigid suture marks in Anthracosuchus may have
reduced the flexibility of this joint, but provided a firmer
basis from which to depress the pubes and take in air. This
firmer suture may have also meant that Anthracosuchuswas
less capable of pubic rotation as in extant crocodylians
(Claessons 2004). The Eocene Dyrosaurus maghribensis of
Morocco possessed a pubis that is spatulate and much more
similar to that of extant crocodylia (Jouve et al. 2006).
The preserved ribs of Anthracosuchus indicate similar
vertical positions for muscle attachments, as indicated for
Hyposaurinae (Schwarz-Wings et al. 2009) and Acheronti-
suchus (Hastings et al. 2011). The resultant angle for axial
muscles was found to imply dyrosaurids likely swam in a
much more undulating fashion than extant crocodylians,
which are nearly entirely tail-based swimmers (MacDonald
2005; Schwarz-Wings et al. 2009). The osteoderms of
hyposaurine dyrosaurids are known to imbricate dorsally,
providing bracing for the vertebral column.
Osteoderms are not as extensive along the lateral
portion of the dyrosaurid body as in some extant
crocodylians, likely affording dyrosaurids greater flexi-
bility at the sacrifice of reduced stability and armoured
Figure 16. (Colour online) Portions of pelomedosoid turtlecarapace,UF/IGM71,with crocodyliformbitemarks. (A,C)Costalfragment in dorsal view; (B, D) same fragment in ventral view; (E,G) neural fragment in dorsal view; (F, H) peripheral in dorsal view;(I, K) m1 tooth from holotype of Anthracosuchus balrogus gen. etsp. nov., UF/IGM67, placedwithin the impact shell scar featured in(I); (J, L) peripheral indorsal view. bm, bitemark; bm?, possible bitemark; bm&p, bite mark and regrown bone plug; dm, drag marks;m1, first maxillary tooth; pr, proximal rib. Scale bar for A–H, J andL equals 5 cm; scale bar for I and K equals 1 cm.
Historical Biology 21
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

protection. This flexibility would have been necessary for
the more undulating locomotion of dyrosaurids (Schwarz-
Wings et al. 2009). The osteoderms of Anthracosuchus
likely did not imbricate at all, implying an even greater
degree of flexibility, at the sacrifice of reduced stability,
particularly for rostroterminal torsion.
Pitting in osteoderms has been associated with thermal
regulation in extant crocodyliforms, with the increased
surface area and veination allowing a greater amount of
heat transfer to take place along the animal’s broadly
exposed dorsal surface (Seidel 1979). Bones of the skull
that are unpitted are separated from the dermis by muscle,
as opposed to pitted surfaces that are very near to the
dermis (Seidel 1979). The osteoderms of Anthracosuchus
are unpitted and likely resided deeper in the dorsal
musculature than in extant crocodylians, and were likely
not utilised as much for thermoregulation. The small
rivulets and thin crevices radiating from the centre of the
dorsal surface are instead likely indicative of keratinisa-
tion, which has been proposed for similar features on the
cranium as a display feature in the extinct crocodyliform
Aegisuchus (Holliday and Gardner 2012).
Diet. The teeth of Anthracosuchus are blunt and robust,
which may be indicative of a durophagous diet with a
strong crushing bite (Massare 1987). A large turtle shell
(UF/IGM 71) has preserved several dents, scores and
scratch marks on dorsal and ventral surfaces that resemble
predation attempts by a crocodyliform (Noto et al. 2012).
There are straight puncture marks, tooth slide marks and at
least one rehealed puncture mark (Figure 16). The largest
of these puncture marks fits very well with the largest
preserved tooth (m1) of UF/IGM 67 (Figure 2).
Acknowledgements
Many thanks to L. Teicher, F. Chavez, G. Hernandez, C. Montesand the rest of the crew at the Cerrejon coal mine for access to thesite and housing during field work. Thanks to Ecopetrol S.A.-ICPfor logistic support. Thanks to H. Garcia, A. Rincon, E. Cadena,F. Herrera, S. Wing and M. Ramirez for help collecting fossils atCerrejon. J. Bourque provided useful guidance for specimenpreparation and conservation. Thanks to E. Whiting for takingadditional measurements during manuscript revision. Thanks toR. Pellegrini for access to and assistance with the collections at theNJSM. Thanks to J. Head for assistance and guidance during thedevelopment of this manuscript. Lastly, thanks to the ColombianGeological Survey (SGC) in Bogota, Colombia, for access tospecimens and facilitating ongoing research of the remarkablevertebrate fossils of the Palaeocene–Eocene of Colombia.
Funding
Funding for research and fieldwork was made possible by theNational Science Foundation [grant numbers DEB-0733725 andDSGC 640179], funds from the Florida Museum of NaturalHistory, the Smithsonian Tropical Research Institute Paleobiol-ogy Fund, the Geological Society of America Graduate StudentResearch Grant, the R. Jerry Britt, Jr Paleobiology Award
(Florida Museum of Natural History), Gary S. Morgan StudentResearch Award and the Cerrejon Coal mine.
References
Barbosa JA, Kellner WA, Viana MSS. 2008. New dyrosauridcrocodylomorph and evidences for faunal turnover at the K–Ptransition in Brazil. Proc R Soc B. 275(1641):1385–1391.
Benson RBJ, Butler RJ, Lindgren J, Smith AS. 2010. Mesozoic marinetetrapod diversity: mass extinctions and temporal heterogeneity ingeological megabiases affecting vertebrates. Proc R Soc B. 277(1683):829–834.
Brochu CA. 1996. Closure of neurocentral sutures during crocodilianontogeny: implications for maturity assessment in fossil archosaurs.J Vert Paleontol. 16(1):49–62.
Brochu CA, Bouare ML, Sissoko F, Roberts EM, O’Leary MA. 2002.A dyrosaurid crocodyliform braincase from Mali. J Paleontol. 76(6):1060–1071.
Buffetaut E. 1978a. Les Dyrosauridae (Crocodylia, Mesosuchia) desphosphates de l’Eocene inferieur de Tunisie: Dyrosaurus, Rhabdog-nathus, Phosphatosaurus. Geol Mediter. 5(2):237–256.
Buffetaut E. 1978b. A dyrosaurid (Crocodilia, Mesosuchia) from theupper Eocene of Burma. Neues Jahrb Geol Palaontol Monatshefte.5:273–281.
Buffetaut E. 1978c. Crocodilian remains from the Eocene of Pakistan.Neues Jahrb Geol Palaontol Abh. 156:262–283.
Buffetaut E. 1979. Sokotosuchus ianwilsoni and the evolution of thedyrosaurid crocodiles. Niger Field Monogr. 1(6):31–41.
Buffetaut E. 1991. Fossil crocodilians from Tiupampa, (Santa LuciaFormation, Early Paleocene) Bolivia: a preliminary report. Rev TecYacim Pet Fisc Boliv. 12(3–4):541–544.
Burns ME. 2000. Taxonomic utility of ankylosaur (Dinosauria,Ornithischia) osteoderms: Glyptodontopelta mimus Ford, 2000: atest case. J Vert Paleontol. 28(4):1102–1109.
Cadena E, Bloch JI, Jaramillo C. 2010. New podocnemidid turtle(Testudines: Pleurodira) from the middle-upper Paleocene of SouthAmerica. J Vert Paleontol. 30(2):367–382.
Cadena EA, Bloch JI, Jaramillo C. 2012. New bothremydid turtle(Testudines, Pleurodira) from the Paleocene of NortheasternColombia. J Paleontol. 86(4):689–699.
Cadena EA, Ksepka DT, Jaramillo CA, Bloch JI. 2012. Newpelomedusoid turtles from the late Palaeocene Cerrejon Formationof Colombia and implications for phylogeny and body size evolution.J Syst Palaeontol. 10(2):313–331.
Claessons LPAM. 2004. Archosaurian respiration and the pelvic girdleaspiration breathing of crocodyliforms. Proc R Soc Lond B.271(1547):1461–1465.
Clark JM. 1994. Patterns of evolution in Mesozoic Crocodyliformes.In: In the shadow of the dinosaurs. New York, NY: CambridgeUniversity Press; p. 84–97.
de Andrade MB, Edmonds R, Benton MJ, Schouten R. 2011. A newBerriasian species of Goniopholis (Mesoeucrocodylia, Neosuchia)from England, and a review of the genus. Zool J Linn Soc. 163(s1):S66–S108.
de Stefano G. 1903. Nuovi rettili degli strati a fosfato della Tunisia. BollSoc Geol Ital. 22(1):51–80.
Denton RK, Dobie JL, Parris DC. 1997. The marine crocodilianHyposaurus in North America. In: Ancient marine reptiles.New York, NY: Academic Press; p. 375–397.
Dodson PD, Krause W, Forster CA, Sampson SD, Ravoavy F. 1998.Titanosaurid (Sauropoda) osteoderms from the Late Cretaceous ofMadagascar. J Vert Paleontol. 18(3):563–568.
Gasparini Z, Pol D, Spalletti LA. 2006. An unusual marine crocodyliformfrom the Jurassic–Cretaceous boundary of Patagonia. Science.311(70):70–73.
Gradstein F, Ogg J, Smith A. 2004. A geologic time scale 2004.Cambridge: Cambridge University Press.
Grenard S. 1991. Handbook of alligators and crocodiles. Malabar, FL:Krieger Publishing Company.
Hastings AK, Bloch JI, Jaramillo CA. 2011. A new longirostrinedyrosaurid (Crocodylomorpha, Mesoeucrocodylia) from the Paleo-
22 A. K. Hastings et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014

cene of north-eastern Colombia: biogeographic and behaviouralimplications for new-world Dyrosauridae. Palaeontology. 54(5):1095–1116.
Hastings AK, Bloch JI, Cadena EA, Jaramillo CA. 2010. A new smallshort-snouted dyrosaurid (Crocodylomorpha, Mesoeucrocodylia)from the Paleocene of Northeastern Colombia. J Vert Paleontol.30(1):139–162.
Hay OP. 1930. Second bibliography and catalogue of the fossil Vertebrataof North America. Vol. 2. Carnegie Inst Washington Publ.390:1–1074.
Head JJ, Bloch JI, Hastings AK, Bourque JR, Cadena EA, Herrera FA,Polly PD, Jaramillo CA. 2009a. Giant boid snake from the Paleoceneneotropics reveals hotter past equatorial temperatures. Nature.457:715–717.
Head JJ, Bloch JI, Hastings AK, Bourque JR, Cadena EA, Herrera FA,Polly PD, Jaramillo CA. 2009b. Brief communication arising (reply).Nature. 460:E4–E5.
Hill RV. 2006. Comparative anatomy and histology of xenarthranosteoderms. J Morphol. 267(12):1441–1460.
Holliday CM, Gardner NM. 2012. A new eusuchian crocodyliform withnovel cranial integument and its significance for the origin andevolution of Crocodylia. PLoS ONE. 7:e30471.
Hoorn C, Wesselingh FP, ter Steege H, Bermudez MA, Mora A, Sevink J,Sanmartın I, Sanchez-Meseguer, Anderson CL, Figueirido JP. 2012.Amazonia through time: andean uplift, climate change, landscapeevolution, and biodiversity. Science. 330(927):927–931.
Jaramillo C, Pardo-Trujillo A, Rueda M, Harrington G, Bayona G, TorresV, Mora G. 2007. Palynology of the Upper Paleocene CerrejonFormation. North Colombia Palynol. 31(1):153–189.
Jouve S. 2005. A new description of Dyrosaurus phosphaticus (Thomas,1893) (Mesoeucrocodylia: Dyrosauridae) from the Lower Eocene ofNorth Africa. Can J Earth Sci. 42(3):323–337.
Jouve S. 2007. Taxonomic revision of the dyrosaurid assemblage(Crocodyliformes: Mesoeucrocodylia) from the Paleocene of theIullemmeden Basin, West Africa. J Paleontol. 81(1):163–175.
Jouve S, Bardet N, Jalil N-E, Suberbiola X, Bouya B, AmaghzazM. 2008.The oldest African crocodylian: phylogeny, paleobiogeography, anddifferential survivorship of marine reptiles through the Cretaceous-Tertiary boundary. J Vert Paleontol. 28(2):409–421.
Jouve S, Bouya B, Amaghzaz M. 2005. A short-snouted dyrosaurid(Crocodyliformes, Mesoeucrocodylia) from the Palaeocene ofMorocco. Palaeontolgy. 48(2):359–369.
Jouve S, Bouya B, Amaghzaz M. 2008. A long-snouted dyrosaurid(Crocodyliformes, Mesoeucrocodylia) from the Paleocene ofMorocco: phylogenetic and palaeobiogeographic implications.Palaeontology. 51(2):281–294.
Jouve S, Iarochene M, Bouya B, Amaghzaz M. 2005. A newcrocodyliform dyrosaurid from the Palaeocene of Morocco and aphylogenetic analysis of Dyrosauridae. Acta Palaeontol Pol. 50(3):581–594.
Jouve S, Iarochene M, Bouya B, Amaghzaz M. 2006. A new species ofDyrosaurus (Crocodylomorpha, Dyrosauridae) from the earlyEocene of Morocco: phylogenetic implications. Zool J Linn Soc.148(4):603–656.
Jouve S, Schwarz D. 2004. Congosaurus bequaerti, a Paleocenedyrosaurid (Crocodyliformes; Mesoeucrocodylia) from Landana(Angola). Bull Inst R Sci Nat Belg Sci Terre. 74:129–146.
Kellner AWA, Campos DA, Riff D, de Andrade MB. 2011. A newcrocodylomorph (Sphagesauridae, Notosuchia) with horn-liketubercles from Brazil. Zool J Linn Soc. 163(s1):S57–S65.
Kodandaramaiah U. 2010. Use of dispersal-vicariance analysis inbiogeography – a critique. J Biogeogr. 37(1):3–11.
LangstonWL. 1965. Fossil crocodilians from Colombia and the Cenozoichistory of the Crocodilia in South America. U Calif Publ Geol Sci.52:1–168.
Langston WL. 1995. Dyrosaurs (Crocodilia, Mesosuchia) from thePaleocene Umm Himar Formation, Kingdom of Saudi Arabia. USGeol Surv Bull. 2093:F1–F36.
MacDonald S. 2005. Activation of the axial muscles during aquaticlocomotion in the estuarine crocodile (Crocodylus porosus) [honoursthesis]. Brisbane: University of Queensland.
Marinho TS, Carvalho IS. 2009. An armadillo-like sphagesauridcrocodyliform from the Late Cretaceous of Brazil. J S Am EarthSci. 27(2009):36–41.
Massare JA. 1987. Tooth morphology and prey preference of Mesozoicmarine reptiles. J Vert Paleontol. 7(2):121–137.
McGregor J. 2005. Crocodile crimes: people versus wildlife and thepolitics of postcolonial conservation on Lake Kariba. Zimbabwe.Geoforum. 36(2005):353–369.
Noto CR, Main DJ, Drumheller SK. 2012. Feeding traces andpaleobiology of a Cretaceous (Cenomanian) crocodyliform: examplefrom the Woodbine Formation of Texas. Palaios. 27(2):105–115.
Pol D, Gasparini Z. 2009. Skull Anatomy of Dakosaurus andiniensis(Thalattosuchia: Crocodylomorpha) and the phylogentic position ofThalattosuchia. J Syst Palaeontol. 7:165–197.
Riccardi AC. 1991. Jurassic and Cretaceous marine connections betweenthe Southeast Pacific and Tethys. Palaeogeogr Palaeoecol.87:155–189.
Ronquist F. 1996. DIVA, version 1.1. Computer program and manualavailable by anonymous FTP from Uppsala University, Uppsala,Sweden. Available at: ftp.uu.se or ftp.systbot.uu.se.
Ross CA. 1989. Crocodiles and alligators. New York, NY: Facts on File.Schwarz D, Frey E, Martin T. 2006. The postcranial skeleton of the
Hyposaurinae (Dyrosauridae; Crocodyliformes). Palaeontology.49(4):695–718.
Schwarz-Wings D, Frey E, Martin T. 2009. Reconstruction of the bracingsystem of the trunk and tail in hyposaurine dyrosaurids(Crocodylomorpha; Mesoeucrocodylia). J Vert Paleontol. 29(2):453–472.
Scotese CR. 2001. Digital paleogeographic map archive on CD-ROM.Arlington, TX: PALEOMAP Project.
Seidel MR. 1979. The osteoderms of the American Alligator and theirfunctional significance. Herpetologica. 35(4):375–380.
Sereno PC, Larsson HC, Sidor CA, Gado B. 2001. The giantcrocodyliform Sarcosuchus from the Cretaceous of Africa. Science.294(5546):1516–1519.
Swofford D. 2003. PAUP* 4.0b10. Sunderland, MA: Sinauer Associates.Thorbjarnarson JB. 1990. Notes on the feeding behavior of the gharial
(Gavialis gangeticus) under semi-natural conditions. J Herpetol.24(1):99–100.
Thorbjarnarson JB. 1993. Fishing behavior of spectacled caiman in theVenezuelan Llanos. Copeia. 1993(4):1166–1171.
Troxell EL. 1925. Hyposaurus, a marine crocodilian. Am J Sci Ser 5.9(54):489–514.
Tolkien JRR. 1954. The fellowship of the ring. London: Unwin Books.Uriona TJ, Farmer CG. 2008. Recruitment of the diaphragmaticus,
ischiopubis and other respiratory muscles to control pitch and roll inthe American alligator (Alligator mississippiensis). J Exp Biol.211(7):1141–1147.
Walker AD. 1970. A revision of the Jurassic reptile Hallopus victor(Marsh), with remarks on the classification of crocodiles. PhilosTrans R Soc Lond Biol Sci. 257(816):323–372.
Walker AD. 1990. A revision of Sphenosuchus acutus Haughton,a crocodylomorph reptile from the Elliot Formation (Late Triassic orEarly Jurassic) of South Africa. Philos Trans R Soc Lond Biol Sci.330(1256):1–120.
Wheatley PV. 2010. Understanding saltwater tolerance and marineresource use in the Crocodylia: a stable isotope approach[dissertation]. Santa Cruz, CA: University of California.
Whetstone KN, Whybrow PJ. 1983. A ‘cursorial’ crocodilian from theTriassic of Lesotho (Basutoland), southern Africa. Occas Pap UnivKans Mus Nat Hist. 106:1–37.
Wing S, Herrera F, Jaramillo C, Gomez-Navarro C,Wilf P, Labandeira C.2009. Late Paleocene fossils from the Cerrejon Formation,Colombia, are the earliest record of Neotropical rainforest. ProcNatl Acad Sci. 106(44):18627–18632.
Yu Y, Harris AJ, He XJ. 2010a. S-DIVA (Statistical Dispersal–Vicariance Analysis) 1.5. Accessed from http://mnh.scu.edu.cn/soft/blog/sdiva on September 2, 2011.
Yu Y, Harris AJ, He XJ. 2010b. S-DIVA (statistical dispersal-vicarianceanalysis): a tool for inferring biogeographic histories. MolPhylogenet Evol. 56(2):848–850.
Historical Biology 23
Dow
nloa
ded
by [
Uni
vers
ity o
f Fl
orid
a] a
t 08:
16 2
6 M
ay 2
014




















