
An emerging interface between life science and nanotechnology
Upload
khangminh22Category
view
0download
0
Application of nanotechnology for the
discovery of circulating proteins as novel
biomarkers of breast cancer
Antonio Castro López
Doctoral Thesis UDC 2021
Supervisors: Dr. Benigno Acea Nebril, Dra. Cristina Núñez González
Doctoral Program in Health Sciences


Application of nanotechnology for the
discovery of circulating proteins as novel
biomarkers of breast cancer
Antonio Castro López
Doctoral Thesis UDC 2021
Supervisors: Dr. Benigno Acea Nebril, Dra. Cristina Núñez González
Doctoral Program in Health Sciences




ACKNOWLEDGEMENTS


Simplemente, a la VIDA.


Esta tesis se ha realizado con el esfuerzo continuado de muchas personas que me
han ayudado y que, gracias a su apoyo y motivación, he conseguido terminarla.
Al Dr. Benigno Acea Nebril, por la co-dirección de esta tesis, por su confianza,
por su persistencia y por su capacidad de enseñarme.
A la Dra. Cristina Núñez González, por la co-dirección de la tesis, por su ayuda,
su trabajo y su apoyo.
A la Dra. Mª Carmen Cereijo Garea, por su disponibilidad, su entusiasmo y su
amistad.
A todas las pacientes, cuya colaboración han hecho posible este trabajo.


SHORT ABSTRACTS


Abstract
Breast cancer is one of the most common cancers in women and accounts for about
14% of cancer-related deaths in females around the world. Breast cancer is a
heterogeneous disease that presents a wide variety of molecular and clinical
characteristics, as well as variability in clinical progression. For the treatment choice,
patients are classified according to intrinsic biological subtypes within the breast
spectrum, using clinical-pathological criteria, i.e. the recognition of amplification and/or
overexpression of the human epidermal growth factor receptor 2 (HER2) oncogene, the
immunohistochemical classification of the estrogen receptor (ER) and the progesterone
receptor (PR), and Ki-67 labelling index. This classification allows for a more
personalized approach to medical treatments, with favorable results. However, despite
that, almost 10-15% of these patients still experience local or distant recurrences in the
first 5 years from diagnosis.
Classification of breast cancer might be markedly improved if new biomarkers
identified with the use of high-throughput “omics” approaches could support diagnosis
based on histopathological patterns. In this way, nanomaterials have been introduced into
the field of proteomics to establish a new and rapidly evolving research area termed
nanoproteomics.
It is well known that the dispersion of a nanomaterial in physiological fluid results in
the formation of a protein shell named “protein corona” (PC). PC varies depending on the
characteristics of the biological media, the physical (size, shape, curvature) and chemical
properties (composition, surface charge/chemistry, hydrophobicity/hydrophilicity) of the
nanomaterial, and the incubation time. Disease-associated biomarkers comprise less than
1% of serum proteins. In this way, through the formation of the PC, nanoparticles could
act as sorbent materials for the enrichment of low-abundance peptides/proteins presented
in serum samples before the biomarker identification by mass spectrometry (MS)
analysis. Importantly, otherwise undetectable changes in protein concentration at an early
stage of the disease (as breast cancer), after any treatment (chemotherapy,
immunotherapy) or surgery could be detected analyzing the PC composition. Thus,
characterization of the PC around NPs offers distinct advantages over sole proteomic
approaches and increases the success of identifying molecular targets.
Firstly, this thesis aims to optimize the formation of the bio-corona formed around
the surface of gold nanoparticles (AuNPs), silver nanoparticles (AgNPs), platinum

nanoparticles (PtNPs) and magnetic nanoparticles (MNPs) after their interaction with
proteins present in human serum. After that, it was developed an exhaustive qualitative
and quantitative analysis of composition of the PC through electrophoretic separation
(SDS-PAGE) in combination with liquid chromatography tandem-mass spectrometry
(LC-MS/MS). This methodology was applied for the identification of serum biomarkers
of the different breast cancer subtypes. This study shows that nanoproteomics is a
valuable tool that can facilitate comprehensive and systematic identification of the serum
proteome and the molecular classification of breast cancer.

Resumen
El cáncer de mama es uno de los tipos de cáncer más común en mujeres y supone
aproximadamente el 14% de las muertes relacionadas con el cáncer en mujeres de todo el
mundo. Se trata de una enfermedad heterogénea con una amplia variedad de
características moleculares y clínicas, así como una variabilidad en la progresión clínica.
Para la elección del tratamiento adecuado, las pacientes con cáncer de mama se clasifican
en distintos subtipos empleando criterios clínico-patológicos que se basan en los niveles
de expresión del oncogén del receptor 2 del factor de crecimiento epidérmico humano
(HER2), la clasificación inmunohistoquímica del receptor de estrógeno (RE) y el receptor
de progesterona (RP), y el índice Ki-67. La clasificación del cáncer de mama en los
distintos subtipos permite un abordaje más personalizado de los tratamientos médicos,
con resultados favorables. Sin embargo, a pesar de esto, casi el 10-15% de estos pacientes
todavía experimentan recidivas locales o distantes en los primeros 5 años tras el
diagnóstico.
Actualmente, el uso de nuevas herramientas “ómicas” permite la identificación de
nuevos biomarcadores que respalden el diagnóstico basado en patrones histopatológicos,
lo que se traducirá en una mejora en la clasificación del cáncer de mama.
Con esta finalidad se han comenzado a introducer los nanomateriales en el campo de
la proteómica, dando lugar a una nueva área de investigación denominada
nanoproteómica.
La nanoproteómica se basa en que la dispersión de un nanomaterial en un fluido
fisiológico da como resultado la formación de una capa de proteinas denominada
"corona". Esta corona proteica varía según las características del medio biológico, las
propiedades físicas (tamaño, forma, curvatura) y químicas (composición, carga
superficial/química, hidrofobicidad/hidrofilicidad) del nanomaterial y el tiempo de
incubación.
Los biomarcadores asociados a un determinada enfermedad suponen menos del 1%
de las proteínas presentes en el suero sanguíneo. Con la formación de la corona de
proteínas, los nanomateriales actuan como materiales absorbentes con los que se lleva a
cabo el enriquecimiento de péptidos/proteínas de baja abundancia presentes en las
muestras de suero sanguíneo. El análisis de estas proteínas ancladas a la superficie de los
nanomateriales mediante técnicas de espectrometría de masas permitirá la identificación
de nuevos biomarcadores asociados con una determinada enfermedad, como el cancer de

mama. Así, mediante este tipo de análisis se podrán detectar cambios en la concentración
de proteínas en una fase temprana de una enfermedad, tras cualquier tratamiento
(quimioterapia, inmunoterapia) o una intervención quirúrgica. Por lo tanto, la
caracterización de la corona de proteínas que se forma alrededor de los nanomateriales
ofrece distintas ventajas en relación con los análisis proteómicos convencionales, y es
más eficaz a la hora de llevar a cabo la identificación de nuevas dianas moleculares.
En primer lugar, esta tesis tiene como objetivo optimizar la formación de la bio-
corona formada alrededor de la superficie de nanopartículas de oro (AuNPs),
nanopartículas de plata (AgNPs), nanopartículas de platino (PtNPs) y nanopartículas
magnéticas (MNPs) tras su interacción con las proteínas presente en suero sanguíneo
humano. Posteriormente, se lleva a cabo un análisis cualitativo y cuantitativo exhaustivo
de la composición del corona proteica formada mediante la combinación de las técnicas
de separación en gel (SDS-PAGE) en combinación con la espectrometría de masas en
tándem (MS/MS) acoplada a la cromatografía de líquidos (LC-MS/MS). Tras la
optimización de esta metodología, se aplica en la identificación y cuantificación de
biomarcadores de los diferentes subtipos de cáncer de mama presentes en muestras de
suero sanguíneo.
Este estudio muestra que la nanoproteómica es una herramienta valiosa que puede
facilitar la identificación integral y sistemática del proteoma sérico y la clasificación
molecular del cáncer de mama.

Resumo
O cancro de mama é un dos cancros máis comúns nas mulleres, xa que representa
aproximadamente o 14% das mortes relacionadas co cancro en mulleres de todo o mundo.
É unha enfermidade heteroxénea cunha gran variedade de características moleculares e
clínicas, así como variabilidade na progresión clínica. Para escoller o tratamento axeitado,
as pacientes con cancro de mama clasifícanse en diferentes subtipos empregando criterios
clínicopatolóxicos baseados nos niveis de expresión do oncóxeno receptor de factor de
crecemento epidérmico humano 2 (HER2), a clasificación inmunohistoquímica do
receptor de estróxenos (RE) e o receptor de proxesterona (RP), e o índice Ki-67. A
clasificación do cancro de mama nos diferentes subtipos permite unha abordaxe máis
personalizada dos tratamentos médicos con resultados favorables. Non obstante, a pesar
diso, case o 10-15% destas pacientes aínda experimentan recorrencias locais ou distantes
nos primeiros 5 anos despois do diagnóstico.
Actualmente, o uso de novas ferramentas "ómicas" permite a identificación de
novos biomarcadores que apoien o diagnóstico baseado en patróns histopatolóxicos, o
que se traducirá nunha mellora na clasificación do cancro de mama.
Para iso, comezaron a introducirse nanomateriais no campo da proteómica, dando
lugar a unha nova área de investigación chamada nanoproteómica.
A nanoproteómica baséase no feito de que a dispersión dun nanomaterial nun
fluído fisiolóxico resulta na formación dunha capa de proteínas chamada "coroa". Esta
coroa proteica varía segundo as características do medio biolóxico, as propiedades físicas
(tamaño, forma, curvatura) e químicas (composición, carga superficial/química,
hidrofobicidade / hidrofilicidade) do nanomaterial e o tempo de incubación.
Os biomarcadores asociados a unha determinada enfermidade representan menos
do 1% das proteínas presentes no soro sanguíneo. Coa formación da coroa proteica, os
nanomateriais actúan como materiais absorbentes cos que enriquecen os
péptidos/proteínas de baixa abundancia presentes nas mostras de soro sanguíneo. A
análise destas proteínas ancoradas á superficie de nanomateriais mediante técnicas de
espectrometría de masas permitirá a identificación de novos biomarcadores asociados a
unha determinada enfermidade, como o cancro de mama. Así, a través deste tipo de
análise, pódense detectar cambios na concentración de proteínas nunha fase inicial dunha
enfermidade, despois de calquera tratamento (quimioterapia, inmunoterapia) ou

intervención cirúrxica. Polo tanto, a caracterización da coroa proteica que se forma ao
redor dos nanomateriais ofrece distintas vantaxes sobre as análises proteómicas
convencionais e é máis eficiente na identificación de novas dianas moleculares.
En primeiro lugar, esta tese ten como obxectivo optimizar a formación da bio-
coroa formada ao redor da superficie de nanopartículas de ouro (AuNPs), nanopartículas
de prata (AgNPs), nanopartículas de platino (PtNPs) e nanopartículas magnéticas (MNPs)
despois da súa interacción coas proteínas presente no soro sanguíneo humano.
Posteriormente, lévase a cabo unha análise cualitativa e cuantitativa completa da
composición da coroa proteica formada combinando técnicas de separación de xel (SDS-
PAGE) en combinación con espectrometría de masas en tándem (MS/MS) unida a
cromatografía líquida (LC-MS / MS). Despois de optimizar esta metodoloxía, aplícase na
identificación e cuantificación de biomarcadores dos diferentes subtipos de cancro de
mama presentes en mostras de soro sanguíneo.
Este estudo demostra que a nanoproteómica é unha valiosa ferramenta que pode
facilitar a identificación completa e sistemática do proteoma sérico e a clasificación
molecular do cancro de mama.

CONTENTS


Contents
Abbreviations…………………………………………………………………………...1
I. Introduction…………………………………………………………………………..7
1. Problematic: breast cancer…………………………………………………......7
2. Non-modifiable risk factors…………………………………………………....7
3. Modifiable risk factors………………………………………………………..10
4. Protective factors……………………………………………………………..11
5. Actions with sufficient evidence of benefit…………………………………...12
6. Actions without sufficient evidence of relationship………………………......14
7. Factors with sufficient evidence of no or little relationship……………….......15
8. Locoregional and systemic spread of breast cancer…………………………..15
9. Early detection of breast cancer……………………………………………....18
10. Protein biomarkers for breast cancer screening and diagnosis……………....20
11. Protein biomarkers for breast cancer prognosis……………………………..38
12. Biomarkers for response prediction and treatment monitoring of breast
cancer…………………………………………………………………………………...40
13. References…………………………………………………………………..42
II. Objectives…………………………………………………………………………..65
III. Results and Discussion…………………………………………………………....67
Results and Discussion. Chapter 1…………………………………………………....67
Abstract………………………………………………………………………....67
Keywords……………………………………………………………………….67
1. Introduction…………………………………………………………………..68
2. Experimental………………………………………………………………....70
3. Results and discussion……………………………………………………….74
4. Conclusions…………………………………………………………………..84
5. References…………………………………………………………………....85

Results and Discussion. Chapter 2…………………………………………………...93
Abstract………………………………………………………………………....93
Keywords……………………………………………………………………….94
1. Introduction…………………………………………………………………..94
2. Experimental………………………………………………………………....97
3. Results and discusión……………………………………………………….106
4. Conclusions………………………………………………………………....135
5. References…………………………………………………………………..135
Results and Discussion. Chapter 3…………………………………………………....143
Abstract………………………………………………………………………..143
Keywords……………………………………………………………………...144
1. Introduction………………………………………………………………....144
2. Experimental………………………………………………………………..146
3. Results and discusión……………………………………………………….149
4. Conclusions………………………………………………………………....163
4. References…………………………………………………………………..165
IV. Conclusions……………………………………………………………………....173
ANNEX A. Supplemental Material Chapter 1…………………………………….175
ANNEX B. Supplemental Material Chapter 2……………………………………..205
ANNEX C. Supplemental Material Chapter 3…………………………………….283
ANNEX D. Extended abstract………………………………………………………325
ANNEX E. List of publications

ABBREVIATIONS


1
AAL: aleuria aurantia lectin
ACM: antibody colocalization microarray
ACN: acetonitrile
AD: area-based breast density
AFP: α-fetoprotein
AgNPs: silver nanoparticles
AHSG: alpha 2HS-glycoprotein
ANX A3: annexin A3
APOA1: apolipoprotein A-I
APOA2: apolipoprotein A-II
APOC1: apolipoprotein C-I
APOC2: apolipoprotein CII
APOC3: apolipoprotein C-III
APOE: apolipoprotein E
APOH: apolipoprotein H
ATII: angiotensin II
AUC: area under the curve
AuNPs: gold nanoparticles
BBC: BRCA1 mutant breast cancer
BBD: benign breast disease
BCa: breast cancer
BH: BRCA1 mutant healthy
BM: bone metastasis
BTD: biotinidase
CA15-3: carbohydrate antigen 15-3
CA19-9: carbohydrate antigen 19-9
CA27.29: carbohydrate antigen 27.29
CA125: carbohydrate antigen 125
C3a desArg: C3a des-arginine anaphylatoxin
C4BPB: complement component 4 binding protein β
CDH5: cadherin-5
CEA: carcinoembryonic antigen
CFHR3: complement factor H-related 3
CGB: champedak galactose binding

2
COL10A1: collagen 10a1
COL11A1: collagen11a1
COMP: collagen oligomeric matrix protein
1CTP: pyridinoline crosslinked carboxyterminal telopeptide of type I collagen
DCIS: ductal carcinoma in situ
DE: differentially expressed
DEP: differentially expressed proteins
DL: detection limit
DM: distant metastases
dPC: digital ProteomesChip
DPV: differential pulse voltammetry
DR: dynamic range
2D-DIGE: 2-dimensional difference gel electrophoresis
2D-nanoLC-MS/MS: two-dimensional nano-liquid chromatography coupled with
tandem mass spectrometry
ECL: electrochemiluminescent
ELISA: enzyme-linked immunosorbent assay
ELLA: enzyme-linked lectin assay
ER: estrogen receptor
ESI: electrospray ionization
EVs: extracellular vesicles
FGA: fibrinogen alpha
GO: graphene oxides
HC: healthy controls
HER2: human epidermal growth factor receptor-2
HIC: hydrophobic interaction chromatography
HILIC: hydrophilic interaction chromatography
HP: human plasma
HPA: helix pomatia agglutinin
HS: human serum
HSPs: heat shock proteins
HSP90A: heat shock protein 90A
IAP: inhibitor of apoptosis
IDC: invasive ductal carcinoma

3
IEF: isoelectric focusing
IHC: immunohistochemistry
IGFBP3: insulin-like growth factor-binding protein 3
IgG Fc: immunoglobulin G crystallizable fragment
IMAC: immobilized metal affinity chromatography
ITIH4: inter-alpha trypsin inhibitor heavy chain family member H4
iTRAQ: isobaric tags for relative and absolute quantification
LA: luminal A
LAC: lectin affinity chromatography
LB: luminal B
LC: label-free quantification
LC-MS/MS: liquid chromatography tandem-mass spectrometry
LNM: lymph node metastasis
LR: local recurrence
LTA: lotus tetragonolobus agglutinin
MALDI-TOF MS: matrix-assisted laser desorption/ionization time-of-flight mass
spectrometry
MBs: magnetic beads
MIF: migration inhibitory factor
M-LAC: multi-lectin affinity chromatography
MLR: multiple logistic regression
MNPs: magnetic nanoparticles
MRM: multireaction monitoring
MWCNTs: multiwalled carbon nanotubes
m/z: mass/charge ratio
NCIs: non-cancerous individuals
NFX1: nuclear transcription factor, X box-binding protein 1
NSR: no sign of recurrence
NTNBC: non-triple-negative breast cancer
NTX: N-terminal crosslinking telopeptides of type I collagen
OPN: osteopontin
ORM-1: α-glycoprotein orosomucoid 1
OS: overall survival
PAI-1: plasminogen activator inhibitor-1

4
PAPPA: pappalysin-1
PCA: perchloric acid
PDD: primary disseminated disease
PDGF: platelet-derived growth factor
PEP: protein elution plate
pIgR: polymeric immunoglobulin receptor
PKG1: cGMP dependent protein kinase1
PR: progesterone receptor
PRM: parallel reaction monitoring
PTMs: post-translational modifications
PTHrP: parathyroid hormone-related protein
PtNPs: platinum nanoparticles
PZP: pregnancy zone protein
RALGAPA2: Ral GTPase-activating protein subunit alpha-2
RANTES/CCL5: regulated on activation normal T cell expressed and
secreted/chemokine (C-C motif) ligand 5
REC: recurrent breast cancer
ROC: receiver operating characteristic
RP: reverse phase
RPC: reversed phase chromatography
SAP: serum amyloid protein
SBA: antibody suspensión bead array
SBC: sporadic breast cancer
SDS-PAGE: sodium dodecyl sulfate polyacrylamide gel electrophoresis
SELDI-TOF MS: surface-enhanced laser desorption/ionization time-of-flight mass
spectrometry
SEREX: serological analysis of recombinant cDNA expression libraries
SISCAPA: stable isotope standards and capture by anti-peptide antibodies
SLPI: secretory leukocyte protease inhibitor
SPB: serum protein biomarker
sTfR: soluble form of transferrin receptor
TAA: tumor-associated antigens
TAAb: tumor associaced autoantibodies
TJP2: tight junction protein 2

5
TNBC: triple-negative breast cancer
TPA: tissue polypeptide antigen
TPS: tissue polypeptide specific antigen
UPLC: ultraperformance liquid chromatography
WCX: weak cation exchange
WT: wild type

6

I. INTRODUCTION


I. Introduction
______________________________________________________________________
7
1. Problematic: breast cancer
Incidence is defined as the number of new cases of a disease in a population and
at a given time. It can be expressed as the absolute number of new cases in a year or as
rates (number of new cases per-100000 people/year).
Cancer continues to be one of the main causes of death with approximately 18.1
million new cases/year in the world (SEOM, year 2018) [1]. Regarding the European
Union and breast cancer, in that year 404,900 new cases were diagnosed, which
represents 29.2 percent of all cancers in women and an incidence rate of 108.8 per-
100000. Spain has an average incidence rate in relation to its neighboring countries
(84.9 per-100000 women) (SEOM, año 2018) [1].
Breast cancer is the leading cause of cancer death in the E.U. In 2018, 98800
women died from this cause, with an age-standardized mortality rate of 21.4 per-100000
(in Spain, 15.4 per-100000) (SEOM, año 2018) [1].
The estimate of new cases of breast cancer in Spain in 2020 is 32953. It is
estimated that one in every 8 women will have breast cancer at some point in her life
[2].
Breast cancer continues to be a major problem and, at present, we do not have
effective primary prevention measures, given that the most important risk factors (age,
family history of breast cancer, sex, history of breast disease, …) are not modifiable.
It is undoubted, therefore, that our efforts should go towards secondary
prevention activities, such as a diagnosis as early as possible, ... etc. In this way, we will
be able to carry out a treatment with a much greater chance of cure, since survival is
closely linked to the stage in which the cancer is, at the time of diagnosis. (Survival in
stage I is 98%, while in stages III it drops to 25%). In 2018, 6534 women died of breast
cancer in Spain [3].
2. Non-modifiable risk factors
2.1. Age. Sex. Race and Size
The risk increases with increasing age. It is very rare, before the age of 30 (and
is usually associated with genetic alterations) [4]. In Spain, the percentage of women
with breast cancer over 65 years of age represents 0.24% of the population in that age
group, while in the range of 15 to 65 years, it represents 0.12% of that age group.

I. Introduction
______________________________________________________________________
8
population. This explains why the incidence is higher in developed countries, given that
life expectancy is higher than in underdeveloped countries.
Only 1 in 100 breast cancers occurs in men. This is explained by hormonal
differences, since exposure to sex hormones is the most determining difference.
Estrogen levels (but also progestogens and androgens) have shown a strong
association with breast cancer (more pronounced in postmenopausal women) and
especially with luminal tumors. This association is not so evident in triple negative
tumors; therefore, there are studies that report that they can even be protective against
this subtype of breast cancer.
White women are at higher risk (and within these, Hispanics have a lower risk
than Caucasians). African American women have a higher risk of "triple negative" and
"younger ages" than Caucasians of the same characteristics. And this is significant. But
lifestyle and migrant populations may see their initial risk modified in relation to race
[5].
Women who are taller than 1.75 cm are 20% more likely to develop breast
cancer than those who measure 1.60 or less (bias in relation to lifestyle and diet, in the
growth stage) [6].
2.2. Breast tissue density
Women with mammographically dense breasts have a 2- to 6-fold increased risk
of developing breast cancer compared to women with the lowest breast density.
The increase in risk is proportional to the degree of density. In a report that
groups three studies of cases (1112 pairs of cases) and controls (similar number), it was
observed that women with a density of 75% or more, compared to women with a
density lower than 10%, which ranges between 1.79 and 4.74. This increase persisted
for a minimum of 8 years and was greater in younger women [7, 8].
An increased risk of mortality has not been shown in women with dense breast
tissue.
2.3. Bone mineral density
There is a correlation between high bone mineral density (which leads to a high
hormonal load) and an increased risk of breast cancer [9]. In the Women's Health study,
it is observed that for each unit that increases the T-score, this risk increases [10].

I. Introduction
______________________________________________________________________
9
2.4. Reproductive factors
Early menarche and late menopause are higher risk factors for breast cancer; due
to increased reproductive cycles and therefore increased circulating estrogens. Even
higher, from 55 years of age and independent of the tumor phenotype [11].
2.5. Personal history of breast cancer
There is an increased risk of developing contraletral breast cancer, but it varies
depending on the age of the woman or the histological type of the tumor (in both DCIS
and ICD). LCIS is considered a risk marker for developing ipsilateral or contralateral
breast carcinoma, with a RR between 8-10 [6].
2.6. Personal history of proliferative injuries
Non-proliferative lesions (simple cysts, fibrosis, simple fibroadenoma, simple
columnar or apocrine alteration, and mild ductal hyperplasia) have not been shown to
significantly increase the risk of developing breast cancer (RR, 1.2 - 1.4).
Proliferative lesions without atypia (ductal or columnar hyperplasia, sclerosing
adenosis, papilloma and radial scar), a RR, between 1.7-2.1.
Proliferative lesions with atypia (HLA, CLIS, HDA), a clear increase in risk has
been demonstrated (RR greater than or equal to 4).
The increased risk due to flat epithelial atypia, aprocrine atypia, and secretory
atypia is unclear [12].
2.7. Heredity
Only 5-10% of women with breast cancer have inherited mutations that are the
main cause of tumors. The most common are in the BRCA1 and BRCA2 genes (55-
65%) that pose an approximate risk of developing cancer before age 80, of 70%. Other
genes also involved are: PALB2, TP53, ATM, CHEK2, PTEN or STK11.
There are populations with characteristic mutations such as the Icelandic and the
Ashkenazi Jewish community [13].

I. Introduction
______________________________________________________________________
10
3. Modifiable risk factors
3.1. Hormone therapy
Multiple population studies (Heart and estrogen/progestin replacement study,
WHY, Million Women study, etc.) have shown that long-term use of combined
estrogen-progestin HRT increases the risk of breast cancer (very low, with less use of
five years and marked with use greater than 10 years). However, the relationship of
increased risk with the use of estrogen-only HRT is controversial; since there are studies
that show a higher risk and others that show protection (it is possible that the time
elapsed between the start of E therapy and menopause, is very important) [4].
3.2. Obesity
In postmenopausal women there is a clear relationship between weight gain and
an increased risk of breast cancer. Mainly due to the fact of a greater production of
estrogens by adipose tissue (between 50 to 100%, higher in women with obesity). The
relationship between obesity, diabetes and insulin concentration and the risk of breast
cancer has been studied; but this has not been possible to define clearly [14-16].
3.3. Alcohol
Alcohol use increases the risk of breast cancer and has been shown to be
dose/dependent in many studies and is a global risk for all tumor subtypes. In a British
meta-analysis of 53 studies, comparing women with breast cancer who did not consume
alcohol, the RR was 1.32 (95% CI, 1.19-1.45; P <0.001) in women who consumed 35 to
44 g of alcohol per day and 1.46 (95% CI, 1.33-1.61; P <0.001) among those who
consumed 45 or more. The RR of breast cancer increases 10% for every 10 grams of
alcohol (one drink/day) [17].
3.4. Exposure to ionizing radiation
Although it appears that there is no increased risk of developing breast cancer, in
women exposed to ionizing radiation for diagnosis, there appears to be a certain
predisposition, especially in carriers of BRCA mutations, especially if the exposure
occurs at ages younger than 40 years.
On the other hand, exposures to radiation of a therapeutic nature (Hodking's disease,
etc.) in young women (during their breast development), if it represents an increased
risk of up to 35% of developing breast cancer around the age of 40. Higher doses of

I. Introduction
______________________________________________________________________
11
radiation and treatment between 10 and 16 years of age are associated with a higher risk
(and this risk is not reduced with follow-up time, persisting up to 25 years after
treatment). Ovarian suppression secondary to chemotherapy or targeted radiation does
seem to have a protective effect [14-27].
3.5. Nulliparity
It is a perfectly established risk factor (especially in the development of tumors
in ages over 70 years). Obesity acts synergistically for the development of cancer [28].
4. Protective factors
4.1. Pregnancy
In general, it is considered a protective factor for breast cancer. The early age in
gestation confers greater protection (there is a 12% decrease in risk for each full-term
pregnancy, in menopausal women and 3% for each one in premenopausal women). This
protective effect does not occur in women over 35 years of age, compared to nulliparous
women [28-30].
4.2. Breastfeeding
Breastfeeding is associated with a lower risk of breast cancer (this decrease in
risk is especially significant, for triple negative tumors). In a review of 47
epidemiological studies from 30 countries, with 50302 women with breast cancer and
96000 controls. It was shown that the decrease in risk was greater in women who had
children and breastfed than in those who had children and did not breastfeed. It was also
proportional to the duration of lactation. The RR decreased by 4.3% for every 12
months of lactation and by 7%, for every delivery [31].
4.3. Physical activity
Regular physical exercise can reduce the risk of breast cancer, especially in
young women who have had children. In a meta-analysis with 123574 cases, it was
observed that physical exercise decreased the risk of breast cancer, as well as the events
arising from the neoplasia [32-35].

I. Introduction
______________________________________________________________________
12
4.4. Diet
The analyzes on the impact of the Mediterranean diet, as well as the intake of
fruits and vegetables seem to show a decrease in the risk of breast cancer. In a statistical
analysis carried out on 62000 women in the Netherlands, a statistically significant
association was observed between diet and a decrease in breast cancer (especially in ER
negative tumors). Although all this is contradictory, due to the existence of other studies
(Women's Healthy Eating and living randomized trial, a branch of the WHI, ... etc) that
have not found a relationship [36].
5. Actions with sufficient evidence of benefit
5.1. Selective estrogen receptor modulators
Several trials have shown that tamoxifen reduces the recurrence rate and the
appearance of new contralateral primary breast cancers and protects bone mineral
density (BMD) in postmenopausal women. [37-42].
The Breast Cancer Prevention Trial (BCPT) randomized more than 13,000
women at high risk of breast cancer to receive treatment with tamoxifen versus placebo,
finding a 49% decrease in the incidence in the tamoxifen group, also accompanied by a
reduction in the number of fractures (as side effects, more endometrial cancers and more
thrombotic phenomena were observed) [43, 44]. An update confirmed similar results at
7 years of follow-up [45].
A meta-analysis was conducted with three other trials: one in the UK (2471
women at high risk of breast cancer due to family history) [45], another Italian (5,408
women undergoing hysterectomy, low or normal risk) [46] and the Breast Cancer
Intervetion study, with 7152 women at increased risk of breast cancer; where a 38%
reduction in the incidence of breast cancer was demonstrated (greater decrease in ER
positive tumors, up to 48%), confirming a similar incidence of adverse effects [47].
The NSABP-24 showed that in women with DCIS (who have a higher risk of
contralateral breast cancer), who had added tamoxifen to local radiotherapy, compared
to those who had not been added, a decrease was shown, statistically significant,
invasive and in situ cancers, as well as contralateral breast cancers [48].
Raloxifene is a selective estrogen receptor modulator (SERM) that acts as an
antiestrogen at the level of the breasts and endometrium, with an estrogenic effect at the
bone, coagulation and lipid levels. In the MORE study, with a sample of close to 8.000

I. Introduction
______________________________________________________________________
13
postmenopausal women with osteoporosis, treated with raloxifene, vertebral fractures
were reduced and, as a collateral benefit, the incidence of invasive breast cancer (mainly
positive for ER) was reduced, and no found an increase in the incidence of endometrial
cancer and hyperplasia [49].
In the CORE trial (MORE trial extension), in which 80% of the MORE study
participants were followed for a further 4 years, there was a 66% reduction in invasive
breast cancer and a reduction in breast cancer positive for ER was 76% [50].
Similar results were observed in the Raloxifene User for the heart (study to
evaluate coronary effects and invasive breast cancer with raloxifene) [51].
The Star study compared about 20000 women at risk to tamoxifen versus
raloxifene treatment, with a 4-year follow-up and demonstrated a similar decrease in
incidence in both groups; but fewer invasive cancers in tamoxifen. There were no
significant differences in coronary events, stroke, or fractures. Episodes of venous
thrombosis and cataracts were more frequent on tamoxifen [52, 53].
5.2. Aromatase inhibitors
These drugs interfere with the adrenal enzyme (anastrozole and letrozole inhibit
its activity and exemestane inactivates it) that allows the production of estrogens in
postmenopause. The most notable side effect is the reduction in bone mineral density
(BMD) and the increase in fractures.
Assays: Arimidex, Tamoxifen Alone or in combination (compared anastrozole
and tamoxifen as adjuvant therapy for breast cancer) [54], another 5000 women taking
tamoxifen adjuvant for five years were randomized to chance, letrozole placebo vs [55];
another controlled with placebo in which 1900 women participated who had received
adjuvant tamoxifen followed by 5 years, followed by letrozole, taking it for another five
years [56] and, another of about 4700 women with neoadjuvant tamoxifen for two years
randomized to continue tamoxifen or switch to exemestane; all demonstrated a
decreased risk of recurrence and new breast cancers in women with previous breast
cancers [57].
Both an RCT of primary prevention (comparing exemestane with placebo in
4500 women) [58], and in IBIS II (studying 3800 women with increased risk of
developing breast cancer, who were alatorized to anastrozole and placebo) [59], which
aromatase inhibitors decrease the incidence of breast cancer in patients at increased risk.

I. Introduction
______________________________________________________________________
14
5.3. Risk-reducing mastectomy (RRM)
The studies are retrospective. Bilateral RRM reduces the risk of breast cancer by
around 90%, depending on the type of surgery performed and the clinical study. There is
no clear demonstration of reduced survival in these women. But it is an option in
women carrying Brca [60].
Contralateral RRM may be an indication in patients with a previous diagnosis of
breast cancer in some circumstances (diagnosed before the age of 41, or triple negative
tumors diagnosed before the age of 50). An impact on survival of contralateral RRM in
patients at low or moderate risk of breast cancer has not been demonstrated [61].
5.4. Risk-reducing salpingo-oophorectomy (RRSO)
High-risk patients and BRCA mutation carriers are at increased risk for breast,
ovarian, tube, and primary peritoneal cancer. Since there are no reliable methods of
early detection and the poor prognosis of advanced ovarian cancer, RRSO has been
recommended, after ending the birth desire (this is associated with a decrease in the risk
of carcinoma of the ovary, tube and primary peritoneum (in carriers of the mutation) and
77% of all-cause mortality. This surgery has been associated with a reduction in the risk
of breast cancer, especially if performed in premenopausal women. In risk
assessment/benefit, the impact on reproduction, the risk of breast and ovarian cancer,
and the risks associated with premature menopause must be considered [62-65].
6. Actions without sufficient evidence of relationship
6.1. Taking hormonal contraceptives
Studies have linked a small increased risk of breast cancer in current consumers,
which decreases over time [66, 67]. Another study, in Denmark [68], also found it
among those who take them now or had recently taken them (and it increased, the
longer they were taken); but, in absolute terms, this effect was very low.
In other cases and controls, well carried out, no relationship was observed
between its consumption and the increased risk of breast cancer with respect to each
use, the duration of use or when it was used [69].
6.2. Environmental factors
In general, studies and evidence supporting a relationship between
environmental factors and specific exposures and the increase in breast cancer are often

I. Introduction
______________________________________________________________________
15
weak. Because many factors have to be considered, which leads to many difficulties in
interpreting that relationship [70-71].
7. Factors with sufficient evidence of no or little relationship
7.1. Shift work
Attempts have been made to link the nocturnal production of melatonin,
secondary to night shift work, with the development of breast cancer. There are multiple
contradictory and light-weight studies; but in 2016, the results of three prospective
studies from the United Kingdom and several additional prospective studies were
combined, with a total of 800,000 women, and it is objective that there are no data that
allow associating the incidence of breast cancer and night shift work [72].
7.2. Geographical residence
It has not been shown that geographic influence could have something to do with
the risk of breast cancer. However, a variation has been seen in the prevalence and
incidence of breast cancer in population groups that migrate (especially in the second
generation), which seems to demonstrate the incidence of lifestyle in this cancer.
The combination of the polygenic risk score (PRS) with family history and other
risk factors allows better risk stratification and the development of prediction models,
which require further studies for their validation and adaptation to other populations
[73].
8. Locoregional and systemic spread of breast cancer
(Extracted, with permission, from the doctoral thesis of Dra. Alejandra García
Novoa. https://www.researchgate.net/publication/317638293)
Between 25-30% of breast cancer patients may have a recurrence [74,75].
The metastases of any carcinoma are the spread of tumor cells to other organs.
Genetic heterogeneity makes it possible for some tumor cells to survive in other organs.
These cells, starting from the local invasion, through the blood and the lymphatics
colonize in the distance [76].
In breast cancer, tumor spread can occur by embolization or permeation through
the bloodstream, the lymphatic system, or by direct invasion through the chest wall.
Systemic spread is usually mixed: lymphovascular. Thus through the small

I. Introduction
______________________________________________________________________
16
intramammary veins a neoplastic invasion can occur. Cancer can invade the vasa-
vasorum or perivascular lymphatics, leading to intravascular cancer invasion with
consequent neoplastic embolization through the bloodstream [77].
Pathophysiological studies present indirect evidence that tumor cells in breast
cancer tend to invade lymphatic vessels first than blood vessels. However,
hematogenous spread can occur without clear identification of regional lymph node
invasion. In fact, most patients do not present with simultaneous lymphatic and bone
marrow involvement [78] and up to 40% of patients without lymph node metastases
present with bone marrow micrometastases. This supports the theory that breast cancer
spread does not occur simultaneously and that it can use different pathways.
Halsted's Mechanistic Theory. William Halsted proposed that breast cancer is a
local disease that spreads systemically in a predictable way. The disease begins in the
primary tumor in the breast, later spreading to regional lymph nodes and then
systemically to distant organs. Unfortunately, only 12% of patients treated with the
classic Halsted radical mastectomy survived 10 years, so this theory did not explain the
failure of local treatment [79].
Alternative Theory: systemic disease. In contrast to the “Halstedian” theory,
Bernard Fisher proposed a concept of systemic disease, defending the idea that tumor
cells can directly invade lymphatic or hematic capillaries and spread systemically
without passing through regional nodes [80, 81]. Therefore, the hypothesis that
metastases occur as a late "additional event" in carcinogenesis is questioned. One of the
reasons for rethinking this theory is the evidence that 10-20% of patients with metastatic
breast carcinoma at the time of surgery do not have infiltrated lymph nodes. In addition,
it has been reported that more than 30% of patients without lymph node involvement
will relapse in the next 10 years [82].
Spectrum theory. It is based on the biological heterogeneity of the tumor and its
genetic expression. Postulates that the ability to metastasize is acquired in early stages
of carcinogenesis, although it manifests much later, after mutation of other genes [64].
Other groups defend that tumor cells develop their metastatic potential as the tumor
grows and evolves clinically. Therefore, lymph node dissection is important for the
prognosis and control of the disease [82].

I. Introduction
______________________________________________________________________
17
“Seed and Soil” hypothesis. Each carcinoma has a different ability to
metastasize to each organ. In breast cancer, the bones, lung, liver, lymph nodes, chest
wall and brain are the most frequent sites of metastasis, however, cases of metastatic
invasion have been reported in almost any organ. Tumors with hormone receptors (HR)
usually metastasize initially to the bones, and those negative for HR, HER2 (human
epidermal growth factor receptor 2) positive more commonly metastasize to the viscera.
[83]. The "Seed and Soil" hypothesis could explain this fact. This hypothesis proposed
by Paget [83, 84] in 1889 explained that each cancer ("seed" or seed) has a specific
tropism for each organ ("soil" or soil).
Based on animal models, it has been shown that thousands of epithelial tumor
cells diffuse daily into the bloodstream; Most of these cells are very short-lived, some
are already apoptotic, while others are supposed to be removed by shear forces from the
bloodstream. However, in up to 30% of patients, tumor cells are able to persist in the
bloodstream after removal of the primary tumor, which can lead to late disease relapse
[84]. This theory is the subject of much modern research, which focuses on determining
the molecular environment that allows the metastatic cascade of cancer.
Plumbing theory. In contrast to the above, the "plumbing" or anatomical theory
was proposed, which defends that the ability to metastasize in certain tissues is
secondary to the anatomical relationships and the circulation that the tumor tissue
presents [77]. Thus, for example, colon cancer patients metastasize to the liver through
the portal system. Both arguments are currently defended as contributors to the tropism
of cancer cells. The dissemination of metastatic cells can occur in the early stages of the
disease, even before the tumor acquires the maximum phenotypic expression of
malignancy. This is the reason because the primary and metastatic tumors can evolve
independently with genetic diversity, acquiring different phenotypes.
Tumor Cells in the Bone Marrow. It has recently been reported that 30% to
40% of breast cancer patients may present viable tumor cells in the bloodstream after
surgery, and may even persist in the blood or bone marrow after adjuvant treatment is
completed [83, 84]. But despite the high incidence of bone marrow micrometastases in
breast cancer patients, bone marrow metastases are rare. This persistence of malignant
cells is associated with a worse prognosis, with 40% to 60% of these patients suffering a

I. Introduction
______________________________________________________________________
18
relapse [83]. Gruber et al [84] demonstrated that the persistence of tumor cells after
chemotherapy is an independent marker of residual disease and therefore reduces the
disease-free period and OS. This fact supports the "sleeping cell" hypothesis, which
proposes that the tumor cell survives in a latent state until it finds the optimal conditions
to proliferate. Disseminated tumor cells are characterized by a low expression of
proliferation markers such as Ki-67, which could explain their ability to survive as a
latent cell to antiproliferative cytotoxic treatment. In addition, these cells express few
molecules of the major histocompatibility complex class I (MHC I), which allows an
immune escape. Up to 87% of bone marrow tumor cells are HER2 positive, as opposed
to 15% to 30% of HER2 positive primary tumors [84]. Consequently, studies have been
carried out indicating specific therapy (for example, Trastuzumab) in patients with
tumor cells in the bone marrow that are positive for this marker, successfully
eliminating them. However, the clinical value of eliminating these tumor cells is
uncertain. [84]. This phenotypic difference between primary tumor cells and circulating
cells also occurs with hormone receptors; being the majority of tumor cells in bone
marrow negative for estrogen receptors and therefore resistant to TH. Consequently, for
routine clinical practice, biopsy and phenotypic identification of metastases contribute
to the choice of the appropriate specific treatment.
9. Early detection of breast cancer
Early detection of BC is important for improvement of prognosis and survival
rate. Until now, mammography has been one of the most important early diagnostic
methods for BC, but it is less effective for young women, with a sensitivity of 25-59%
[85].
On the other hand, the main cause of mortality after BC is metastatic
dissemination of the primary tumour to distant sites in the body. New markers capable
of identifying metastatic breast cancer are required to aid clinical decision making for
individual patients [86,87]. The prognostic tests in current clinical use require tumour
tissue to be obtained by biopsy or other surgical approaches.
It is desirable to minimise such invasive procedures, and new validated
serum/plasma biomarkers are urgently necessary for the early detection of BC in
asymptomatic individuals, precise prognosis and prediction of response to treatment,
and clinical detection of breast cancer metastasis [88].

I. Introduction
______________________________________________________________________
19
Whilst several serum biomarkers have been evaluated over the past three
decades, they lack the sensitivity to detect early primary BC [89] and none of these
possess sufficient accuracy in predicting recurrence [90]. Therefore, it is imperative to
find potential blood-based biomarkers in breast cancer.
Proteomics has become an attractive approach to search for novel biomarkers in
biological fluids of cancer patients using protein and peptide profiling [91]. In this way,
mass spectrometry (MS) has been used to compare proteomic patterns in cancer patients
and healthy controls [92]. The detection of early-stage cancer is based on the paradigm
that the disease develops by increasing deviations from the normal status. Thus,
potential biomarkers could be found among the specific proteins or peptides that are up
or down-regulated in serum proteomic profiling in cancer patients compared with
controls [93]. Furthermore, proteomics analysis could also complement gene analyses in
its use in the prognosis and evaluation of disease [94].
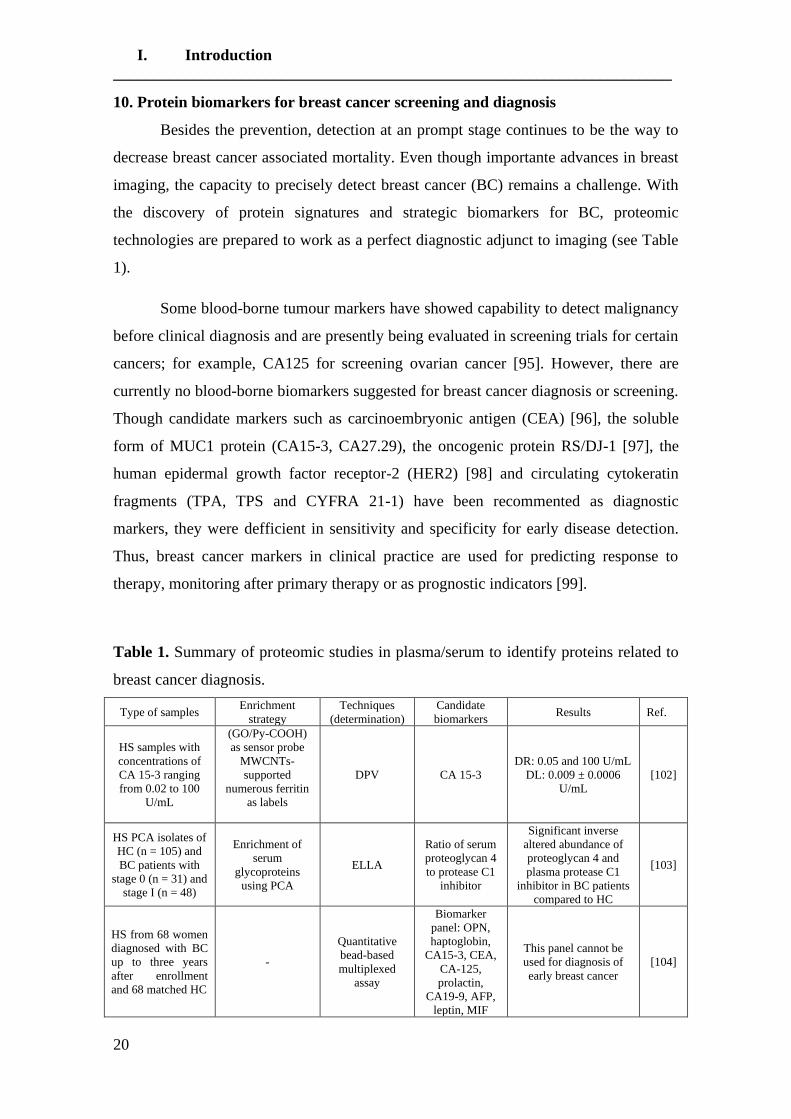
This revision summarizes studies linked to the application of proteomics in the
field of early BC detection, prognosis, and response to therapeutic treatments (see
Figure 1).
Figure 1. Classification of proteomic studies carried out in breast cancer focused on
early detection, prognosis and evaluation of response to treatment.
I. Introduction
______________________________________________________________________
20
10. Protein biomarkers for breast cancer screening and diagnosis
Besides the prevention, detection at an prompt stage continues to be the way to
decrease breast cancer associated mortality. Even though importante advances in breast
imaging, the capacity to precisely detect breast cancer (BC) remains a challenge. With
the discovery of protein signatures and strategic biomarkers for BC, proteomic
technologies are prepared to work as a perfect diagnostic adjunct to imaging (see Table
1).
Some blood-borne tumour markers have showed capability to detect malignancy
before clinical diagnosis and are presently being evaluated in screening trials for certain
cancers; for example, CA125 for screening ovarian cancer [95]. However, there are
currently no blood-borne biomarkers suggested for breast cancer diagnosis or screening.
Though candidate markers such as carcinoembryonic antigen (CEA) [96], the soluble
form of MUC1 protein (CA15-3, CA27.29), the oncogenic protein RS/DJ-1 [97], the
human epidermal growth factor receptor-2 (HER2) [98] and circulating cytokeratin
fragments (TPA, TPS and CYFRA 21-1) have been recommented as diagnostic
markers, they were defficient in sensitivity and specificity for early disease detection.
Thus, breast cancer markers in clinical practice are used for predicting response to
therapy, monitoring after primary therapy or as prognostic indicators [99].
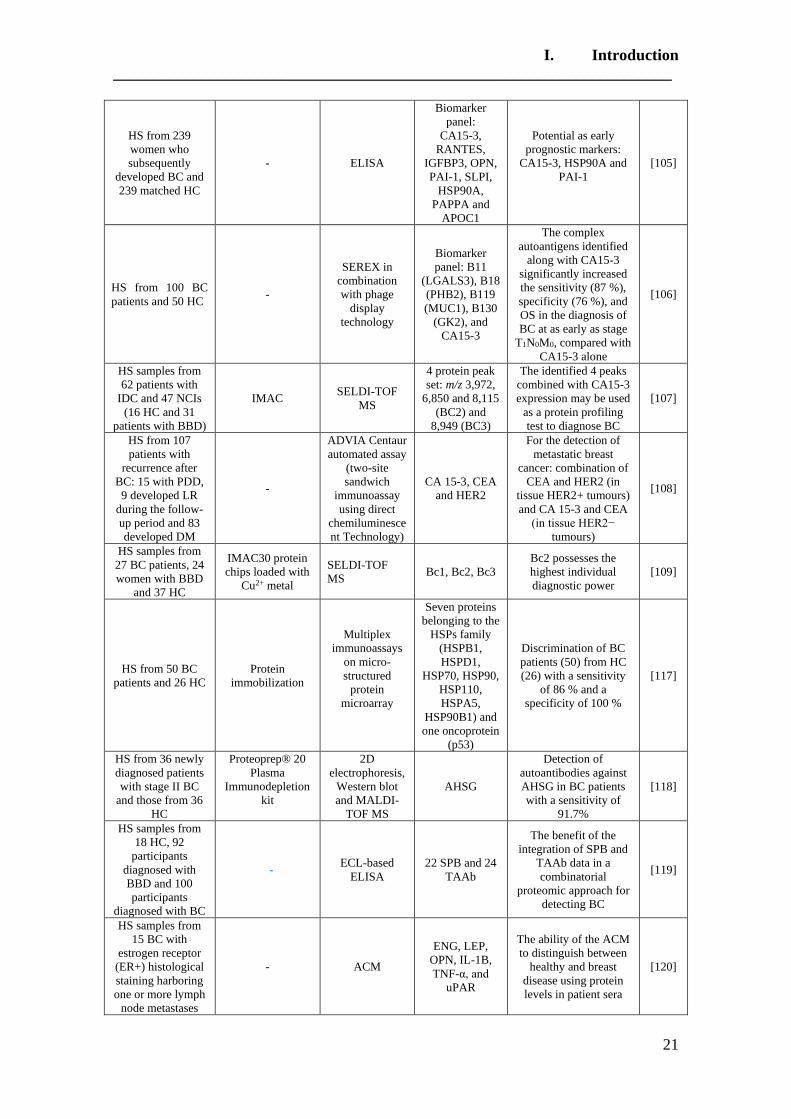
Table 1. Summary of proteomic studies in plasma/serum to identify proteins related to
breast cancer diagnosis.
Type of samples Enrichment
strategy
Techniques
(determination)
Candidate
biomarkers Results Ref.
HS samples with
concentrations of
CA 15-3 ranging
from 0.02 to 100
U/mL
(GO/Py-COOH)
as sensor probe
MWCNTs-
supported
numerous ferritin
as labels
DPV CA 15-3
DR: 0.05 and 100 U/mL
DL: 0.009 ± 0.0006
U/mL
[102]
HS PCA isolates of
HC (n = 105) and
BC patients with
stage 0 (n = 31) and
stage I (n = 48)
Enrichment of
serum
glycoproteins
using PCA
ELLA
Ratio of serum
proteoglycan 4
to protease C1
inhibitor
Significant inverse
altered abundance of
proteoglycan 4 and
plasma protease C1
inhibitor in BC patients
compared to HC
[103]
HS from 68 women
diagnosed with BC
up to three years
after enrollment
and 68 matched HC
-
Quantitative
bead-based
multiplexed
assay
Biomarker
panel: OPN,
haptoglobin,
CA15-3, CEA,
CA-125,
prolactin,
CA19-9, AFP,
leptin, MIF
This panel cannot be
used for diagnosis of
early breast cancer
[104]
I. Introduction
______________________________________________________________________
21
HS from 239
women who
subsequently
developed BC and
239 matched HC
- ELISA
Biomarker
panel:
CA15-3,
RANTES,
IGFBP3, OPN,
PAI-1, SLPI,
HSP90A,
PAPPA and
APOC1
Potential as early
prognostic markers:
CA15-3, HSP90A and
PAI-1
[105]
HS from 100 BC
patients and 50 HC -
SEREX in
combination
with phage
display
technology
Biomarker
panel: B11
(LGALS3), B18
(PHB2), B119
(MUC1), B130
(GK2), and
CA15-3
The complex
autoantigens identified
along with CA15-3
significantly increased
the sensitivity (87 %),
specificity (76 %), and
OS in the diagnosis of
BC at as early as stage
T1N0M0, compared with
CA15-3 alone
[106]
HS samples from
62 patients with
IDC and 47 NCIs
(16 HC and 31
patients with BBD)
IMAC SELDI-TOF
MS
4 protein peak
set: m/z 3,972,
6,850 and 8,115
(BC2) and
8,949 (BC3)
The identified 4 peaks
combined with CA15-3
expression may be used
as a protein profiling
test to diagnose BC
[107]
HS from 107
patients with
recurrence after
BC: 15 with PDD,
9 developed LR
during the follow-
up period and 83
developed DM
-
ADVIA Centaur
automated assay
(two-site
sandwich
immunoassay
using direct
chemiluminesce
nt Technology)
CA 15-3, CEA
and HER2
For the detection of
metastatic breast
cancer: combination of
CEA and HER2 (in
tissue HER2+ tumours)
and CA 15-3 and CEA
(in tissue HER2−
tumours)
[108]
HS samples from
27 BC patients, 24
women with BBD
and 37 HC
IMAC30 protein
chips loaded with
Cu2+ metal
SELDI-TOF
MS Bc1, Bc2, Bc3
Bc2 possesses the
highest individual
diagnostic power
[109]
HS from 50 BC
patients and 26 HC
Protein
immobilization
Multiplex
immunoassays
on micro-
structured
protein
microarray
Seven proteins
belonging to the
HSPs family
(HSPB1,
HSPD1,
HSP70, HSP90,
HSP110,
HSPA5,
HSP90B1) and
one oncoprotein
(p53)
Discrimination of BC
patients (50) from HC
(26) with a sensitivity
of 86 % and a
specificity of 100 %
[117]
HS from 36 newly
diagnosed patients
with stage II BC
and those from 36
HC
Proteoprep® 20
Plasma
Immunodepletion
kit
2D
electrophoresis,
Western blot
and MALDI-
TOF MS
AHSG
Detection of
autoantibodies against
AHSG in BC patients
with a sensitivity of
91.7%
[118]
HS samples from
18 HC, 92
participants
diagnosed with
BBD and 100
participants
diagnosed with BC
- ECL-based
ELISA
22 SPB and 24
TAAb
The benefit of the
integration of SPB and
TAAb data in a
combinatorial
proteomic approach for
detecting BC
[119]
HS samples from
15 BC with
estrogen receptor
(ER+) histological
staining harboring
one or more lymph
node metastases
- ACM
ENG, LEP,
OPN, IL-1B,
TNF-α, and
uPAR
The ability of the ACM
to distinguish between
healthy and breast
disease using protein
levels in patient sera
[120]

I. Introduction
______________________________________________________________________
22
and 11 HC
40 HP samples
from women with
BC (diagnosed with
a stage II or III or
earlier breast
cancer) and 40 HP
samples from HC
-
Identification by
LC/MS/MS and
quantification
using the
LC/MS-based
label-free
protein
quantification
software
licensed from
Eli Lilly and
Company
90 alternative
splicing
isoforms in 38
genes were
found, which
showed
statistically
significant (q <
0.05)
differences
between BC
and HC samples
The signature identified
92.5 % BC samples and
72.5 % of normal
samples
[125]
HS samples for BC
patients and HC
were pooled with
equal volumen (100
μL each)
AlbuvoidTM beas
2-D gel
separation and
subsequent PEP
Metabolic
enzymes
(hexokinases)
and proteases
Qualitative and
quantitative differences
between BC patients
and HC
[126]
Among all, carbohydrate antigen 15-3 (CA 15-3), is the most widely used serum
marker in patients with BC [100]. CA 15-3 has been used for routine breast cancer
screening, monitoring and follow-up of patients with breast cancer [7]. The median
level of CA 15-3 is 17 U/mL (range 3.9-99.5 U/mL) in patients with primary untreated
breast cancer [101].
To develop novel strategies for the ultrasensitive detection of CA 15-3, an
electrochemical nanostructured immunosensor was fabricated using non-covalent
functionalized graphene oxides (GO/Py-COOH) and multiwalled carbon nanotube
(MWCNTs)-supported numerous ferritins as labels [102]. CA 15-3 was selectively
detected as low as 0.01 ± 0.07 U/mL in human serum samples. This system showed an
excellent selectivity, and it can be regenerated for multiple uses, having a great potential
for future development of the point-of-care cancer diagnostics. On this way, perchloric
acid (PCA) was used to improve the detection of serum O-glycosylated proteins (such
as CA 27.29 and CA 15-3) using an earlier developed sandwich enzyme-linked lectin
assay (ELLA) [103]. By subjecting pre-coated champedak galactose binding (CGB)
lectin-captured glycoprotein fractions of serum PCA isolates of the stage 0 (n = 31) and
stage I (n = 48) breast cancer patients and those of controls (n = 105) to SDS-PAGE,
substantial inverse altered abundance of plasma protease C1 inhibitor and proteoglycan
4 were detected in both the early stages of breast cancer patients related to the controls.
Although it needed further validation in clinically representative populations, ratio of
serum proteoglycan 4 to protease C1 inhibitor couldy be exploited for screening of early
breast cancer.

I. Introduction
______________________________________________________________________
23
Furthermore, the potential of different biomarker panels (containing the cancer
antigen 15-3 (CA15-3)) were explored for the diagnosis of early breast cancer. It was
found that the set of ten potential breast cancer serum biomarkers and cancer antigens
(haptoglobin, osteopontin (OPN), cancer antigen 15-3 (CA15-3), cancer antigen 125
(CA-125), cancer antigen 19-9 (CA19-9), carcinoembryonic antigen (CEA), prolactin,
α-fetoprotein (AFP), leptin and migration inhibitory factor (MIF)) cannot be used to
predict early-stage breast cancer [104]. Similarly, none of these 9 candidate markers
(CA15-3 (cancer antigen 15-3), RANTES/CCL5 (regulated on activation, normal T cell
expressed and secreted/chemokine (C–C motif) ligand 5), OPN (osteopontin), PAI-1
(plasminogen activator inhibitor-1), SLPI (secretory leukocyte protease inhibitor),
HSP90A (heat shock protein 90A), IGFBP3 (insulin-like growth factor-binding protein
3), APOC1 (apolipoprotein C-I) and PAPPA (pappalysin-1) or combinations was useful
for screening breast cancer, and only links with clinico-pathological elements correlated
to prognosis were found for the candidates CA15-3, HSP90A and PAI-1 [105].
However, a panel of complex antigens consisting of B11 (LGALS3), B18
(PHB2), B119 (MUC1) and B130 (GK2) along with CA15-3 significantly increased the
sensitivity (87%), specificity (76%), and overall survival (82.7 %) in the diagnosis of
BC at as early as stage T1N0M0, compared with CA15-3 alone [106]. Even though this
panel of complex antigens required to be validated using more BC samples, it may be a
promise instrument to detect early-stage BC. Furthermore, CA15-3 was also included in
the diagnostic panel constituted of 4 protein peaks [m/z 3,972, 6,850 and 8,115 (BC2)
and 8,949 (BC3)] used to distinguish 62 BC patients with invasive ductal carcinoma
from 16 healthy controls (HCs) and 31 patients with benign breast diseases (BBDs)
[107]. Importanly, the resultant 4 peaks panel together with CA15-3 was demonstrated
to have good sensitivity and specificity for the diagnosis of BC. However, further
investigation using a larger sample size should be performed to verify these results.
The potential of (CA15-3) was also explored for the early diagnosis of
metastatic breast cancer [108]. In this fashion, the sensitivity of CA 15-3, CEA and
HER2 was investigated, and it was found that the combination of two tumour markers
enhanced the sensitivity for detection of metastatic breast cancer, and the determination
of all three tumour markers only improved the sensitivity vaguely. These authors

I. Introduction
______________________________________________________________________
24
suggested the combination of CEA and HER2 in tissue HER2+ tumours and the
combination of CA 15-3 and CEA in tissue HER2− tumours. Nevertheless, sizeable
prospective clinical randomised trials are required to explore the clinical benefits of
early detection and treatment of metastatic disease.
The efficacy of other serum biomarkers on early detection of breast cancer were
also considered. For example, after the evaluation of the efficacy of Bc1, Bc2, and Bc3
serum biomarkers on early detection of breast cancer (BC), only Bc2 was statistically
significant in comparison between the malignant disease group, control group and
benign disease group [109].
On other hand, it is well known that breast cancer is a heterogeneous disease in
which cancer cells can express a variety of aberrant proteins (tumor-associated antigens:
TAA) that are capable of eliciting an immune response (antibody production).
Interestingly, this immune response appears months or years before the clinical
diagnosis of the malignancy [110,111]. TAA and their specific antibodies may offer in
vivo amplification of an early carcinogenic signal, thus possibly allowing earlier
detection of cancer than methods used currently.
In particular, serum possesses several circulating antigens and antibodies related
with cancer progression and development [112,113]. The presence of autoantibodies in
serum against several tumor antigens, such as p53, antineural/antinuclear antigens, and
embryonic neural proteins, has been also assessed in breast cancer [114].
Cancer antigens have demonstrated incredible importance in the clinic for
screening and as prognostic indicators [115,116]. Particularly, heat shock proteins
(HSPs), over-expressed in a extensive range of human cancers, caused the stimulation
of the immune system and accordingly in elevated concentration of anti-HSP
autoantibodies, that are associated with tumor metastasis in breast cancer patients.
Consequently, screening these autoantibodies could be of prognostic and diagnostic
values. In this way, L. Shi et al. [117] immobilized seven proteins belonging to the heat
shock protein family (HSPB1, HSPD1, HSP70, HSP90, HSPA5, HSP90B1) and one
oncoprotein, P53, in six different surface chemistries. Two surface chemistries (COOH
and chitosan) were employed to detect antitumor antigen autoantibodies in 26 healthy
donor and 50 breast cancer sera. The detection of a single autoantibody did not allow
significantly discriminating breast cancer sera from healthy sera, whereas combining

I. Introduction
______________________________________________________________________
25
seven autoantibodies (autoantibodies against HSPB1, HSPD1, HSP70, HSP90, HSPA5,
HSP90B1, and P53) increased the specificity and sensitivity of the test (with a
specificity of 100 % and a sensitivity of 86 %). In this study, they have demonstrated
that customized protein microarrays could be effective tools for the rapid screening of
thousands of biomarkers in a parallel and high-throughput fashion. The performance of
protein microarray is influenced by many parameters such as spotting buffer, surface
chemistry, and protein concentration. However, larger cohorts of breast cancer patients
and healthy donors are needed to validate its performance.
An immune proteomic approach also suggested that the presence of serum
autoantibodies against alpha 2HS-glycoprotein (AHSG) protein colud be helpful as
serum biomarkers for early-stage breast cancer minimally invasive diagnosis and
screening [118]. However, the AHSG will need to be tested and validated by multiple
independent studies utilizing an adequately sized test and a training set of sera samples
from very-early-stage breast cancer. Moreover, further verification with samples from
patients with ductal carcinomain situ and breast cancer in stages III and IV would aid in
confirming the specificity of AHSG autoantibodies in this subset of patients with breast
cancer. This research provided additional preliminary, but important, data on the
potential advantage for clinical serological screening of autoantibody measurement to
detect small tumors in early stages, because autoantibody biomarkers have also been
identified in breast cancer, the majority of these have only been reported in the late-
stage, but not in the early-stage, breast cancer.
Breast tumors were found to be related with systemic changes in levels of both
serum protein biomarkers (SPB) and tumor associated autoantibodies (TAAb). Meredith
C. Henderson et al. [119] evaluated for the first time the independent and combinatorial
contribution of SPB and TAAb expression data for identifying BC using a retrospective
cohort of prebiopsy serum samples from 18 participants with no evidence of breast
disease (ND), 92 participants diagnosed with Benign Breast Disease (BBD) and 100
participants diagnosed with BC, including DCIS. It is important to mention that when
modeling integrated data from both SPB and TAAb, the clinical sensitivity and
specificity for detection of BC improved to 81.0% and 78.8%, respectively. These data
showed the advantage of the combination of SPB and TAAb data and toughly sustained

I. Introduction
______________________________________________________________________
26
the development of other similar combinatorial proteomic approaches for detecting BC
in the future.
A novel concept for multiplexing without mixing named antibody colocalization
microarray (ACM) was introduced by M. Pla-Roca et al. [120]. This technique was
validated by profiling 32 proteins in the serum of (i) 11 controls from age-matched
patients undergoing reduction mammoplasties, (ii) 15 patients with primary breast
cancer overexpressing the estrogen receptor (ER) in the primary tumor (ER+ subtype).
It was found that six proteins (ENG, LEP, OPN, IL-1B, TNF-α, and uPAR) were
associated with the cancer grade of the patient. The candidate biomarkers that were
identified agree with the findings of previous studies which described increased
concentrations of uPAR [121], TNF-RII [122], IL-1B [123], and ENG [124]. However,
all of them need to be veried in follow-up studies with more patients and controls.
Besides, recognizing and characterizing different forms of a protein (isoforms)
are critial to the study of molecular mechanisms and early detection of complex diseases
such as breast cancer. In this way, F. Zhang et al. [125] showed that isoform-specific
peptides could differenciate normal breast from breast cancer, identifying 92.5 % cancer
samples and 72.5 % of normal samples in an independent set of 40 normal samples and
40 breast cancer samples. It showed that alternative splicing isoform makers could act
as independent markers of breast cancer.
In a study developed by D. L. Wang et al. [126], a functional proteomics
technology was used to monitor protease activities and metabolic enzymes
(hexokinases) from resolved serum proteins produced by a modified 2-D gel separation
and subsequent Protein Elution Plate, a method collectively called PEP. For the first
time, substantial differences were found between breast cancer patient serum and
normal serum in both families of enzymes implicated in the cancer development and
metastasis, giving excellent biomarker candidates for breast cancer diagnosis and drug
development.

I. Introduction
______________________________________________________________________
27
10.1. Protein profiling for diagnosis of breast cancer
Protein and peptide profiling was used to find novel biomarkers in biological
fluids (as serum and plasma) of cancer patients [8]. Among the specific proteins or
peptides that are up or down-regulated in serum proteomic profiling in cancer patients
compared with controls, potential biomarkers could be found [10]. Particularly, several
studies focused on protein profiling using the two-laser desorption/ionisation (LDI)
platforms (as matrix-assisted laser desorption/ionisation time-off light (MALDI-TOF)
MS [127] and its variant surfaceenhanced laser desorption/ionisation (SELDI-TOF MS)
were developed to search novel breast cancer biomarkers [128].
On the other hand, recently novel sample preparation techniques based on
nanomaterials have developed, and applied to the separation and enrichment of peptides
and proteins in biological samples [129]. Particularly, magnetic microspheres with the
properties of the easiness to surface modification, high dispersibility and magnetic
responsivity, were considered as a promising material for the convenient and efficient
enrichment of peptides or proteins [130, 131].
Commercial n-alkyl magnetic polymeric beads (1-10 μm diameter) have widely
been used in the enrichment of low-abundance peptides and proteins in biological
samples [132, 133]. However, the commercial magnetic beads have usually showed
poor magnetic response.
C8-functionalized magnetic nanoparticles (about 50 nm diameter) with high
dispersibility, large surface area and excellent magnetic responsibility, were
successfully applied for convenient, fast and efficient enrichment of low-abundance
peptides from tryptic protein digest and human serum, followed by a direct MALDI-
TOF-MS analysis [134]. Furthermore, weak cation exchanges magnetic beads (MB-
WCX-MBs) were used for the effective enrichment of peptides and proteins in
biological samples. Both enrichment methods were applied for the detection of breast
cancer [135, 136].
Using magnetic bead-hydrophobic interaction chromatography C8 and C18
(HIC-C8-MBs and HIC-C18-MBs), and weak cation exchange (WCX) beads for the
enrichment of proteins presented in human serum samples, 14 biomarkers were found,

I. Introduction
______________________________________________________________________
28
whose combination detects breast cancer patients from non-cancer controls with a
sensitivity of 89% and specificity of 67% [137]. Of them, five biomarkers were
comparable with previously identified proteins from published data using similar
approaches (peaks at 4283 and 3972 Da [138,139], 3972 Da [140], 6630 and 6629 Da
[141,142] and 6429 Da [143]). In addition, this biomarker panel were able to
discriminate low-risk (tumor grade G1 or tumor grade G2 with a low level of uPA and
PAI-1) and high-risk breast cancer patients (tumor grade G3 or tumor grade G2 with a
high level of uPA and PAI-1) with a high sensitivity (75%) and specificity (100%).
However, further validation of biomarkers could potentially facilitate the early
diagnosis of breast cancer as an aid to imaging diagnostics.
In a similar way, combining the data resulting from two complementary workup
procedures (WCX-MBs and reversed-phase (RP) C18 magnetic beads (MBs)) improved
the classification of breast cancer, and sensitivity and specificity increased up to 84 and
95%, respectively [144]. Although MALDI-TOF peptide and protein profiles can be
used for classification of breast cancer, larger patient sets must be analyzed for
validation and MS/MS be used to identify the discriminating proteins and peptides for
its use in breast cancer screening programs. More recently, WCX-MBs fractionation
provided predictive model for BC versus healthy controls with 79.04% sensitivity and
82.18% specificity. Furthermore, FGA 605-629, ITIH4 347-356 and APOA2 43-52
were found as potential peptide biomarkers [145].
Using WCX fractionation and mass spectrometry protein profiling, C. L.
Washam et al. [146] found that 12-48aa peptide fragment of parathyroid hormone-
related protein PTHrP(12-48) was significantly increased in the plasma of bone
metastasis (BM) patients compared with patients without BM (p<0.0001). Importantly,
the clinical measurement of PTHrP(12-48) in plasma in combination with NTx in serum
improved the detection of breast cancer BM (diagnostic specificity and accuracy
(AUC=0.99). This result could provide novel opportunities for the improved diagnosis
of bone metastasis, however, some limitations of this study are that is retrospective, the
sample size was somewhat small; and it may also suffer from selection bias. Using the
same methodology, Y. Sun et al [147] found that the candidate biomarker positioned at
m/z 6447.9 identified as apolipoprotein C-I (ApoC-I) was significantly decreased in BC
patients, and its expression intensity was weaker in the triple negative breast cancer
(TNBC) and pre-surgery group compared with the NTNBC and post-surgery group.

I. Introduction
______________________________________________________________________
29
These results suggest that ApoC-I peptides may be a potential diagnostic biomarker and
therapeutic approach for BC. However, further replicated experiments with many more
samples are necessary to verify this possible protein biomarker.
Many reviews reported the advantages of immobilized metal ion affinity
chromatography (IMAC) on the purification of peptides and proteins [46]. Using this
approach for the serum proteomic analysis of 36 healthy volunteers and 37 breast cancer
patients, three peaks at m/z 698, 720 and 1866 were idetintified and used to construct
the peptidome patterns with 91.78% accuracy [148]. Using and independent group for
the validation, it was found that the peptidome patterns could differentiate the validation
group achieving a sensitivity of 91.89% (34/37) and a specitivity of 91.67% (33/36) (>
CA 15-3, p < 0.05).
Using the IMAC enrichment approach, several serum proteins that differed in
concentration between women with asymptomatic breast cancer and matched healthy
controls were also detected [149]. Particularly, two SELDI-TOF MS peaks with m/z
3323 (doubly charged apolipoprotein C-I) and m/z 8939 (C3a des-arginine
anaphylatoxin (C3adesArg)), and with 2D-nanoLC-MS/MS, afamin, apolipoprotein E
and isoform 1 of inter-alpha trypsin and inhibitor heavy chain H4 (ITIH4) were higher
in pre-diagnostic breast cancer serum. Particularly, C3adesArg and ITIH4 have
previously been related to the presence of symptomatic and/or mammographically
detectable breast cancer. However, the currently identified proteins were high abundant
and they were unlikely to be breast cancer specific. In order to find low abundant and
probably more specific tumor markers, other techniques would be employed to give
insight into ‘the deeper/low abundant proteome’.
Similarly, performing serum fractionation by IMAC30 array, WCX (CM10)
array and strong anion exchange chromatography preceding protein profiling with
SELDI-TOF MS, eight peaks showed statistically significantly different intensities
between incident breast cancer cases and controls (p < 0.05) [150]. Seven of these peaks
were tentatively identified as heterodimer of apolipoprotein A-I and apolipoprotein AII
(m/z 45435), apolipoprotein C-II (m/z 8909), oxidized apolipoprotein C-II (m/z 8925),
apolipoprotein C-III (m/z 8746), fragment of coagulation factor XIIIa (m/z 3,959),
hemoglobin B-chain (m/z 15915), and post-translational modified hemoglobin (m/z

I. Introduction
______________________________________________________________________
30
15,346). However, similar to the previous study [66], discriminating proteins were still
high abundant.
In an attemp to combine both purification methods mentioned above, J. Yang et
al. [151] used magnetic bead-based weak cation exchange chromatography (MB-WCX)
and immobilized metal ion affinity chromatography (MB-IMAC-Cu)) to preanalyze 32
patients with early stage (stages I-II) of Invasive Ductal Carcinoma (IDC) and 30
healthy control serum samples. It was found that the serum samples purified in the MB-
WCX group provided a better proteomic pattern than MB-IMAC-Cu. However, both
accurately distinguished patients with early stage IDC from healthy individuals. It was
found that two candidate biomarkers (m/z 4209 and 4264) were upregulated in patients
with IDC by MB-WCX purification, while similar potential biomarkers (m/z: 4263 and
4208) identified by MB-IMAC-Cu purification were also overexpressed in IDC breast
cancer patients. Thus, these two candidate biomarkers will be further identified by
expanding samples from patients with IDC.
In a different approach, 10 normal control and 10 stage IV breast cancer patient
serum samples were analysed by label-free mass spectrometry using a CaptureSelectTM
Transferrin Affinity Matrix [152], identifying 21 potential candidate biomarkers. After
selecting fibronectin and fibrinogen for further analysis in a larger cohort of patient
samples along with CA15-3, it was found that these molecules were significantly altered
when comparing the controls groups to stage IV breast cancer, highlighting the
usefulness of analysing the high-abundant fraction associated proteins.
Other studies to identify serum proteome patterns specific for early stage breast
cancer [153] and invasive breast cancer (IDC) patients [154] using MALDI-TOF mass
spectrometry were also developed. In the latter case, a novel mammary biomarker,
regulator and therapeutic target, Annexin A3, was identified, however large scale
prospective studies together with long term follow-up and detailed molecular analysis
are required to elucidate the role and mechanism(s) by which ANX A3 might impact
breast pathology, diagnostics, and tumourigenesis.

I. Introduction
______________________________________________________________________
31
10.1.1. Protein profiling for diagnosis of breast cancer: post-translational
modifications
To date, most of the serum/plasma proteomic work has been conducted to
analyze total protein level abundance, with only a few studies to analyze post-
translational modifications (PTMs) [155], usually glycosylation [156, 157]. As one of
the most important mechanisms for regulating protein function, PTMs, including
phosphorylation, acetylation, ubiquitination, and methylation, have been identified and
validated as critical for signaling transduction, protein degradation, and transcriptional
regulation [158]. The known importance of PTMs in cellular signaling provided the
impetus for a large-scale survey of PTMs other than glycosylation by immunoaffinity
enrichment of PTM-containing peptides.
10.1.1.1. Glycosylation
Breast carcinomas develop from mammary epithelial cells through genetic
alterations, and interactions with the surrounding stromal tissue are essential for
malignant transformation and for progression of the disease [159]. Glycosylation of cell
surface proteins and lipids is a common post-translational event that regulates the
interaction between epithelial cells and the microenvironment by altering adhesion
properties, cell-cell interaction, and the immune system and by affecting the cells’
migration properties [160].
Several studies, on both breast and other cancer types, have shown that cancer-
related alterations in glycosylation are reflected in serum [161]. Thus, the
characterization of glycan structures is expected to broaden the scope of discovery
studies beyond the protein level, and thus improves the clinical values of existing
biomarkers. For this reason, the development and application of techniques and
methodologies for enriching or fractionating the glycoproteome has become an
emerging field [46].
Experimentally, a main advantage of targeting glycosylation is that
glycopeptides or glycoproteins can be effectively enriched over nonglycosylated
molecules. In proteomics, enrichment targeted to N-glycosylation has typically been
performed using lectin based enrichment [162, 163] or hydrazide chemistry [164].

I. Introduction
______________________________________________________________________
32
In the particular case of breast cancer, multi-lectin affinity chromatography (M-
LAC) was used to isolate the serum glycoproteome. Subsequently, proteins with altered
abundance and glycosylation that could act as biomarkers were identified:
thrombospondin-1 and 5, alpha-1B-glycoprotein, serum amyloid P-component and
tenascin-X. They will be further investigated in future studies to confirm these
glycoprotein biomarker candidates in a significant group of patients for the improved
early detection for breast cancer [165].
S. Selvaraju et al. [166] developed a fully integrated platform for
capturing/fractionating human fucome from disease-free and breast cancer sera using
two lectin columns specific to fucose, namely Aleuria aurantia lectin (AAL) and Lotus
tetragonolobus agglutinin (LTA). After the comparison of the fucosylated proteins in
both groups (disease-free and breast cancer patients), a broad panel of 35 differentially
expressed proteins (DEP) from the combined LTA and AAL captured proteins and a
narrower panel of 8 DEP that were commonly differentially expressed in both LTA and
AAL captured proteins, were obtained. As advantages, the platform allowed the
“cascading” of the serum sample from column-to-column in the liquid phase with no
sample manipulation between the various steps. This guaranteed no sample loss and no
propagation and dilution or experimental biases between the various columns when
comparing the diseased serum fucome to the disease-free fucome by LC-MS/MS.
Lectin affinity chromatography (LAC) in conjunction with 2-dimensional
difference gel electrophoresis (2D-DIGE) and liquid chromatography-tandem mass
spectrometry (LC-MS/MS) were also used to identify serum markers of metastatic
breast cancer [167]. In this case, helix pomatia agglutinin (HPA) was used to isolate
glycoproteins from pooled breast cancer serum samples due to their properties of
binding aberrant glycans associated with metastatic breast cancer. Following proteomic
identification of HPA binding glycoproteins, cadherin-5 (CDH5), pregnancy zone
protein and the polymeric immunoglobulin receptor emerged as potential markers of
metastasis. It was also observed that CDH5 discriminated patients with no sign of
recurrence from those with recurrent breast cancer with 90% specificity. CDH5 showed
to be a potential marker of metastatic breast cancer with both protein levels and HPA
binding contributing to a test that is comparable to CA15.3 in terms of specificity. As
evidenced by the CDH5 data, the glycoproteomic and validatory approach employed
here presented the capacity to identify novel markers of breast cancer metastasis.

I. Introduction
______________________________________________________________________
33
Hydrophilic interaction chromatography (HILIC) separates both neutral and
charged glycans in a single separation, which facilitates total glycan characterization
with single-column chemistry. Separation is based on the hydrophilic potential of the
glycan, which is affected by its size, charge, composition, structure, linkage, and
oligosaccharide branching [168]. HILIC glycan analysis is continuously advancing, and
ultraperformance liquid chromatography (UPLC) has recently been developed for N-
glycan separation of various samples with high efficiency [85]. Improving on HPLC,
UPLC allows for a decrease in the run times and greatly increased resolution, partially
due to sub-2 μm stationary phase technology.
In previous studies using HPLC-HILIC, K. Marino et al. [78] found significant
changes in glycosylation including sialylation, fucosylation, and branching in breast
cancer, especially indicating the presence of metastasis, spread to the lymph nodes and
correlation with tumor circulating cells. More recently, R. Saldova et al. [169] described
the greatly improved separation of human serum N-glycans on UPLC, as compared with
HPLC, where more than 140 N-glycans were assigned using this technique after
profiling serum samples from healthy controls and newly diagnosed breast cancer
patients. Particularly, they also found decreases in high-mannosylated and biantennary
corefucosylated glycans in breast cancer patients compared with controls. They found
that bisected biantennary nonfucosylated glycans were decreased in patients with
progesterone-receptorpositive tumors, and core-fucosylated biantennary bisected
monogalactosylated glycans were decreased in patients with tumor TP53 mutation. In
conclusion, this UPLC-based glycan analysis technique revealed highly significant
differences between healthy women and breast cancer patients. Furthermore, significant
associations with breast carcinoma and systemic features were also described.
Recently, breast cancer (BC) patients were distinguished from cancer-free (NC)
controls by serum immunoglobulin G (IgG) crystallizable fragment (Fc) region N-
glycosylation profiling using matrix-assisted laser desorption/ionization mass
spectrometry (MALDI-MS) [170]. These results suggested that an unknown humoral
factor or soluble mediator affects IgGs from the earliest stage of breast cancer, and also
suggested that IgG Fc region N-glycosylation could play a role in tumor biology.
Although the sample size in the present study was small, the IgG Fc region N-glycan

I. Introduction
______________________________________________________________________
34
multiple logistic regression (MLR) model developed presented the potential to be used
as a diagnostic biomarker and may provide the insights into tumor immunology.
10.1.1.2. Phosphorylation
Although the status of phosphorylation events could provide evidences regarding
disease status, few phosphoproteins have been developed as disease markers.
Furthermore, with several highly abundant proteins representing more than 95% of the
mass in blood, few phosphorylated proteins in plasma/serum can be identified with
detectable and stable concentrations [171].
Recently, using label-free quantitative phosphoproteomics, 144 phosphoproteins
significantly higher in patients diagnosed with breast cancer compared with healthy
controls were indentified in plasma extracellular vesicles (EVs) [172]. Four biomarkers
were initially validated in individual patients using paralleled reaction monitoring
(PRM) for targeted quantitation: cGMP-dependent protein kinase1 (PKG1), ral GTPase-
activating protein subunit alpha-2 (RALGAPA2), tight junction protein 2 (TJP2), and
nuclear transcription factor, X box-binding protein 1 (NFX1). These four
phosphoproteins showed significant phosphorylation up-regulation in patients with
cancer, and have been associated in several breast cancer studies [173]. Although this
study demonstrated that the development of phosphoproteins in plasma EVs as disease
biomarkers could transform cancer screening, it relied on the isolation of a good
quantity of EVs with high reproducibility and the development of phosphoproteins as
biomarkers was also strictly limited by the availability of phosphospecific antibodies.
H. Gu et al. [174] combined immunoaffinity purification and LC-MS/MS
without depletion of abundant proteins for the enrichment and quantitative analysis of
post-translational modifications (PTMs) in serum samples of patients with breast cancer
(BC). It was found that lysine acetylation (AcK) and arginine mono-methylation (Rme)
were more prevalent than other PTMs, finding several AcK and Rme sites with distinct
abundance distribution patterns. Therefore, this approach could be especially useful for
patient profiling and biomarker discovery research.

I. Introduction
______________________________________________________________________
35
10.1.2. Protein profiling studies related to breast cacer subtypes
Studies on breast cancer showed substantial tumor heterogeneity consisting of
different molecular subtypes, each with distinct clinical and biological and
individualities [175]: luminal, HER2-enriched, basal-like and normal breast-like
subtype. The criteria to classify subtypes were recently refined, in that moderate of a
strong expression of PR and Ki-67 level defined the subtypes as: luminal A-ER
positive, HER2 negative, Ki-67 low, and PR high; luminal B (HER2 negative)-ER
positive, HER2 negative, and either Ki-67 high or PR low; luminal B-like (HER2
positive)-ER positive, HER2 overexpressed or amplified, any Ki-67, and any PR; HER2
positive-HER2 over-expressed or amplified, ER and PR absent; and triple negative-ER
and PR absent and HER2 negative [176,177].
Due to its lower costs and easy implementation into standard pathology
workflow, immunohistochemistry (IHC) is the method used to define surrogate protein
biomarkers for the classification of breast cancer [178]. More recently, the molecular
classification by microarray analysis agreed well to IHC classification of different
breast carcinomas [179,180]. Subsequently, IHC and molecular classifications are
concurrently used to define the breast cancer subtypes.
In this way, proteomics could also detect additional proteins or protein profiles
to improve current breast cancer classifications. Besides, proteomics might show
biological insights and recognize protein biomarkers outlining differences in therapy
resistance, prognosis and metastatic spread within a specific subtype. In this section,
recent proteomic studies developed in relation with the molecular classification of
breast cancer will be discussed.
In a proteomic study developed by H. Nakshatri et al. [181] was found that the
plasma proteome of luminal A and HER2+ breast cancer patients did not differ
significantly from healthy individuals, however, in the luminal B subtype, eight proteins
involved in immune response (as α-glycoprotein orosomucoid 1 (ORM-1) and serum
amyloid protein (SAP)) were significantly increased, whereas 12 proteins involved in
free radical scavenging were significantly decreased. Two complement factors
(complement factor H-related 3 (CFHR3) and complement component 4 binding protein
β (C4BPB)) were identified being elevated in TNBC compared with healthy controls,

I. Introduction
______________________________________________________________________
36
and a significant number of plasma proteins (20) were downregulated compared with
healthy individuals.
An integrative clustering analysis of breast cancer subtypes from plasma
proteome samples showed that luminal A and luminal B subtypes were clustered
together, as well as the basal-like and HER2+. Furthermore, luminal A and luminal B
were closer to each other than basal-like and HER2+ to each other. The results also
showed that proteomic pathway-assisted clustering of breast cancer subtypes could offer
relationships and biological insight into the intrinsic mechanisms between the diverse
breast cancer subtypes [182].
More rencently, a multipronged quantitative proteomic approach identified 307
differentially regulated subtype specific proteins: luminal A subtype consisted of 24,
luminal B subtype 38, HER2 Enriched subtype 17 and triple negative breast cancer
subtype 10 [183]. These specific proteins were further subjected to bioinformatic tools
which revealed the involvement in platelet degranulation, fibrinolysis, lipid metabolism,
immune response, complement activation, blood coagulation, glycolysis and cancer
signaling pathways in the subtypes of the breast cancer.
SRM assays in a different cohort of samples verified and confirmed that
Biotinidase (BTD) is down-regulated in LA when compared with other subtypes.
Similarly, APOL1, AATM, TTHY, THRB, AACT, and APOD found to be up-regulated
in LB. SRM assays also confirmed that CPN2 showed increased expression in TN
whereas CD5L showed elevated expression in LB and HE subtypes.
In a work to find protein biomarkers for early detection of ER-positive breast
cancer, it was found that the pre-diagnostic level of plasma EGFR was significant
elevated in women who developed breast cancer. However, this biomarker showed
moderate specificity and sensitivity and for early diagnosis [184]. In another study from
the same group [185], an increased level of glycolysis-related proteins in plasma of
breast cancer patients compared with that in controls was detected. Nevertheless, the
cause of these glycolysis proteins as well as their function in ER-positive breast cancer
was not elucidated until the moment.

I. Introduction
______________________________________________________________________
37
Using an antibody microarray, a panel of 23 proteins was screened in human
plasma from patients with an actual diagnosis of breast cancer of different subtypes
(defined by ER and HER2 status) or benign breast disease [186]. In comparison with
women with benign breast disease, four proteins (amphiregulin, RANTES, heparin-
binding EGF, TGFa) were increased in plasma of ER-positive/HER2-negative BC
patients, three proteins (heparin-binding EGF, EGF, RANTES) in ER-negative/HER2-
positive BC patients, two proteins (RANTES and platelet-derived growth factor
(PDGF)) in ER-positive/HER2-positive BC patients, and two proteins (RANTES and
VEGF) in plasma of ER-negative/HER2-negative of BC patients.
In other antibody-based assay, C. I. Li et al. [187] analyzed plasma samples from
healthy controls and TNBC patients identifying 93 differential proteins. 29 proteins
were confirmed by the validation in an independent cohort. Using a strict criteria, five
proteins (DUSP9, EED, EFNA5, ITGB1 and PTPMT1) were found and colud be
exploited as markers for early detection of TNBC.
Lymph node status is a crucial predictor for the overall survival of invasive
breast cancer. However, lymph node involvement is only detected in about half of
HER2-positive patients. Since patients with lymph node involvement has less
favourable prognosis and higher risk of recurrence, it is important to develop plasma
protein biomarkers for distinguishing lymph node metastasis. In this way, L. Chen et al.
[188] applied label-free quantitative proteomic strategy to construct plasma proteomes
of ten patients with small size HER2-positive breast cancer (five patients with lymph
node metastasis (LNM+) versus five patients without lymph node metastasis (LNM-)).
A total of 388 proteins were identified, of which 33 proteins were differentially
expressed (DE), and 24 proteins were over-expressed in LNM+ group. Western blotting
analysis showed that RARB was greatly increased in LNM+ group and FBLN5 was
slightly elevated, indicating the results of label-free quantification is highly consistent
with western blotting. Although statistical analyses suggested that this aproach is low-
cost and high-efficiency in initial screening of plasma biomarkers, the present dataset
only provided a list of differentially expressed (DE) plasma proteins which could be
used for further screening of genuine biomarkers.

I. Introduction
______________________________________________________________________
38
11. Protein biomarkers for breast cancer prognosis
Compared to diagnostic studies, proteomics studies to discover novel markers to
improve breast cancer prognostication are rather limited.
It was found that adjuvant use of bisphosphonate diminished recurrence in breast
cancer patients through suppression of bone resorption. To determine the prognostic
impact of bone resorption markers, M. Imamura et al. [189] investigated serum levels of
the pyridinoline crosslinked carboxyterminal telopeptide of type I collagen (1CTP) and
N-terminal crosslinking telopeptides of type I collagen (NTX). Relapse-free survival of
130 patients whose 1CTP changed from low at baseline to high at 6 months
postoperatively showed RFS almost as poor as that for patients with high 1CTP
throughout. Only 1CTP could be suitable not only for categorizing patients with adverse
prognosis, but also for choosing patients who colud benefit from administration of bone
modifying agents in an adjuvant scenary.
Recently, it was developed an study to identify density-associated proteins to
improve the understanding of mammographic breast density as a risk factor for breast
cancer [190]. Particularly, it was found that ABCC11, TNFRSF10D, F11R and ERRF
were positively associated with area-based breast density (AD), and SHC1, CFLAR,
ACOX2, ITGB6, RASSF1, FANCD2 and IRX5 were negatively associated with AD.
These data provided insights into the aetiology of breast density as a prominent risk
factor for breast cancer. Furthermore, this study showed that stroma-specific and
epithelial-specific proteins could be found in blood as a consequence of tissue leakage,
which would make them key candidates for future individual risk stratification.
However, further validation and follow-up studies of the shortlisted protein candidates
in independent cohorts will be needed to infer their role in breast density and also its
progression in premenopausal and postmenopausal women.
Among the major risk factors of breast cancer, other important role is played by
familial history of BC. Germ-line mutations in BRCA1/2 genes account for most of the
hereditary ovarian and/or breast cancers. Gene expression profiling studies revealed
particular molecular signatures for BRCA1/2-related breast tumors as compared to
sporadic cases, which could facilitate diagnosis and clinical follow-up. Even though, a
clear hallmark of BRCA1/2-positive BC is still needing.

I. Introduction
______________________________________________________________________
39
Tumor-specific changes in the plasma proteome of BC patients and healthy
family members sharing the same BRCA1 gene founder mutation (5083del19) were
investigated by L. Tammè et al. [191]. The proteomic analysis revealed that gelsolin
was down-expressed in plasma samples of patients with hereditary BC, and that its
levels were associated with the BRCA1 mutation status, showung that this relevant
tumor suppressor gene could stimulate BC cell proliferation, migration and invasion,
also thorough the down-regulation of gelsolin.
Women with inherited BRCA1 mutations are more likely to develop breast
cancer (BC); however, not every carrier will progress to BC. The aim of the study
developed by J. Fan et al. [192]. was to identify and characterize circulating peptides
that correlate with BC patients carrying BRCA1 mutations. After the enrichment of
circulating peptides using a nanoporous silica thin films (NanoTraps), peptides
KNG1K438-R457 and C 3fS1304-R1320 were identified as putative peptide candidates
to differentiate BRCA1 mutant BC from sporadic BC and cancer-free BRCA1 mutant
carriers. Therefore, the expression level of both peptides were associated with cancer
status in BRCA1 carriers. This approach could offer an answer to the dilema of who is
going to get cancer. The long-term longitudinal information would also be greatly
beneficial, particularly for cancer-free BRCA1 mutation carriers who maintain their
high-risk status. We intend for this strategy to improve the early examination of cancer
in the BRCA1 carriers based on the suggestions from the blood-based test. However,
future prospective studies are required to validate these findings.
It was described that the inhibitor of apoptosis (IAP) protein Survivin and its
variants (Survivin-ΔEx3 and Survivin-2B) are differentially expressed in breast cancer
tissues. Furhermore, Survivin is released from tumor cells via small membrane-bound
vesicles called exosomes. Thus, S. Khan et al. [193] developed the analysis of exosomal
Survivin, Survivin-ΔEx3 and Survivin-2B in breast cancer patient sera in parallel with
paired breast tumor tissue. After exsosomal investigation, Survivin and Survivin-ΔEx3
were detected in all of the samples examined, however, Survivin-2B was differentially
expressed depending on the disease aggressiveness: expressed mostly in primary tumors
in early stage disease, low or no expression was found in high-grade tumors and absent
in most distant metastasis. Therefore, exosomal Survivin-2B can be further investigated
as an early diagnostic or prognostic marker in breast cancer.

I. Introduction
______________________________________________________________________
40
M. Giussani et al. [194] tested plasma samples from healthy donors and from
patients with malignant or benign breast disease by ELISA for the presence of
collagen11a1 (COL11A1), collagen oligomeric matrix protein (COMP), and collagen
10a1 (COL10A1). Importantly, the combination of COL11A1, COMP, and COL10A1
was identified as potentially informative in discriminating BC patients from those with
benign disease. The three molecules resulted expressed in the stroma of BC tissue
samples, thus circulating COL11A1, COMP, and COL10A1 could be very
advantageous in diagnostic assessment of suspicious breast nodules.
12. Biomarkers for response prediction and treatment monitoring of breast cancer
Though accurate prediction of chemosensitivity in cancer therapy would allow
personalized therapy, thus avoiding toxic side effects and the use of ineffective agents,
protein profiling studies searching for markers for response prediction and treatment
monitoring of breast cancer are scarce.
In recent years, several studies have reported the diagnostic utility of the low
molecular weight fraction of the human serum peptidome in BCa [53, 195]; however, to
the best of our knowledge, only two studies investigated the putative use of peptide
signals as biomarkers to predict tumour outcome following surgery [196, 197]. M. C.
Gast et al. [113] demonstrated a strong association between serum haptoglobin
phenotype and recurrence-free survival in a group of 63 high-risk early BCa patients. A.
Gonçalves et al. [114] investigated post-operative sera of 83 high-risk BCa patients,
identifying a 40-protein signature that correctly predicted the outcome in 83% of the
cases. Major components of this signature include haptoglobin alpha-1, C3a, transferrin
and apo-lipoproteins A-I and C-I. However, both groups were unable to subsequently
validate their results in an independent patient set.
In this way, recently F. Boccardo et al. [198] employed MALDI-TOF MS to
recognise serum peptidome profiles predictive of mortality in 331 patients who
underwent an operation for infiltrating BCa. At a median follow-up time of 25.5 years
(range 1.3 to 26.9 years), 68 of the 102 patients were deceased, and 45 of these deaths
were attributed to BCa-related causes. It was found that four signals were increased in
deceased patients compared with living patients and only one having mass/charge ratio
(m/z) 1046.49 was associated with BCa-specific mortality. This pea kwas identified as

I. Introduction
______________________________________________________________________
41
Angiotensin II (ATII) whose levels were increased in women who exhibited worse
mortality outcomes, reinforcing the evidence that this peptide potentially significantly
affects the natural history of early BCa.
After clustering analysis of protein spectra to identify protein patterns related to
BC and HV groups [199], five protein peaks (m/z 3808, m/z 6624, m/z 8916, m/z 13870,
and m/z 28268) were indentified that together classified BC and HV with a receiver
operating characteristic (ROC) area-under-the-curve value of 0.961. These proteins
were identified as a fragment of apolipoprotein H (ApoH, m/z 3808), ApoCI (m/z
6624), complement C3a (m/z 8916), transthyretin (m/z 13870), and ApoAI (m/z 28268).
Importantly, this panel significantly predicted disease-free survival (P = 0.005), with
and efficacy greater in women with estrogen receptor (ER)-negative tumors (n = 50, P =
0.003) that in ER-positive (n = 131, P = 0.161). Furthermore, in women with ER-
negative tumors, the combined biomarker could be used as an adjunct to other
pathological variables in predicting patient outcome. However, this method needs to be
confirmed in larger patient cohorts, and also to be developed in a new format (for
example, a multiplexed ELISA kit) that could facilitate its further application.
Although the soluble form of transferrin (sTfR) is frequently used to identify
iron deficiency anaemia [200], a few studies have reported its elevation in a variety of
cancers, including breast cancer [201]. In this way, Q. Xu et al. [202] developed and
validated an advanced LC-MS/MS-based targeted proteomics assay coupled with
peptide immunoaffinity enrichment (SISCAPA) for the quantification of low-level sTfR
(100 ng/ml) in breast cancer patients after the onset of chemotherapy. Using this assay,
60 pairs of serum samples pre- and post-chemotherapy and the corresponding control
samples from 60 healthy volunteers were determined and compared. The results
confirmed sTfR suppression during chemotherapy and suggested that sTfR may be a
potential indicator of transfusion requirement. However, further studies using large
sample sizes in preclinical and clinical trials will be required to confirm the value of this
assay.
Using a fully validated liquid chromatography-tandem mass spectrometric
method, I. van den Broek et al. [203] have compared absolute serum concentrations of
eight peptides derived from inter-α-trypsin inhibitor heavy chain-4 (ITIH4) (ITIH4

I. Introduction
______________________________________________________________________
42
[658-687] to [667-687] (ITIH4-30 to -21)) before and after surgical substraction of the
tumor. Intra-individual comparisons of serum obtained before and after surgery showed
significantly decreased serum levels after surgery for seven of the ITIH4-derived
peptides (p < 0.02). The obtained results particularly suggest potential for these ITIH4-
derived peptides in the follow-up of breast cancer after surgery.
13. References
[1] J. Ferlay, M. Colombet, I. Soerjomataram, T. Dyba, G. Randi, M. Bettio, A. Gavin,
O. Visser, F. Bray, Cancer incidence and mortality patterns in Europe: Estimates for 40
countries and 25 major cancers in 2018, Eur J Cancer 103 (2018) 356-387.
[2] Ministerio de Sanidad, Consumo y Bienestar Social. Instituto de Informacion
Sanitaria. Mortalidad por causa de muerte. Portal estadístico interactivo de información
epidemiológica, [cited 2019 01/12/2019].
https://pestadistico.inteligenciadegestion.mscbs.es/publicoSNS/Comun/Informe.aspx?Id
Nodo=5012.
[3] https://seom.org/info-sobre-el-cancer/cancer-de-mama?showal=1.
[4] C.A. Rodríguez, A. Gómez. Cáncer de mama. En: Cruz II. Rodríguez CA, Del Barco
E. et al. Oncología clínica 6ª Ed. Elservier España; 2019. p. 131-148.
[5] R.L. Siegel, K.D. Miller, A. Jemal, Cancer statistics 2018. CA Cancer J Clin. 68(1)
(2018) 7-30.
[6] Centers for disease control and prevention (CDC). Vital signs: racial disparities in
breast cancer severety_United States, 2005-2009. MMWR Morb Mortal Wkly Rep.
61(45) (2012) 922-926.
[7] N.F. Boyd, H. Guo, L.J. Martin, L. Sun, J. Stone, E. Fishell, R.A. Jong, G. Hislop,
A. Chiarelli, S. Minkin, M.J. Yaffe, Mammographic density and the risk and detection
of breast cancer, N Engl J Med. 356(3) (2007) 227-236.
[8] H. Razzaghi, M.A. Troester, G.L. Gierach, A.F. Olshan, B.C. Yankaskas, R.C.
Millikan, Mammographic density and breast cancer risk in White and African American
Women, Breast Cancer Res Treat. 135(2) (2012) 571-580.
[9] X. Qu, X. Zang, A. Qin, G. Liu, Z. Zhai, Y. Hao, H. Li, Z. Zhu, K. Dai, Bone
mineral density and risk of breast cancer risk in postmenopausal women, Breast Cancer
Res Treat. 138(1) (2013) 261-271.

I. Introduction
______________________________________________________________________
43
[10] Z. Chen, L. Arendell, M. Aickin, J. Cauley, C.E. Lewis, R. Chlebowski, Hip bone
density predicts breast cancer risk independently of gail score: results from the
Women´s Health initiative, Cancer 113(5) (2008) 907-915.
[11] R. Ritte, A. Lukanova, Anne Tjønneland, A. Olsen, K. Overvad, S. Mesrine, G.
Fagherazzi, L. Dossus, B. Teucher, K. Steindorf, H. Boeing, K. Aleksandrova, A.
Trichopoulou, P. Lagiou, D. Trichopoulos, D. Palli, S. Grioni, A. Mattiello, R. Tumino,
C. Sacerdote, J.R. Quirós, G. Buckland, E. Molina-Montes, M.-D. Chirlaque, E.
Ardanaz, P. Amiano, B. Bueno-de-Mesquita, F. van Duijnhoven, C.H van Gils, P.H.
Peeters, N. Wareham, K.-T. Khaw, T.J. Key, R.C. Travis, S. Krum-Hansen, I. Torhild
Gram, E. Lund, M. Sund, A. Andersson, I. Romieu, S. Rinaldi, V. McCormack, E.
Riboli, R. Kaaks, Age and menarche and risk of hormone receptor-positive and negative
breast cancer: a cohort study, Int J Cancer 132(11) (2013) 2019-2029.
[12] L.C. Hartmann, D.C. Radisky, M.H. Frost, R.J. Santen, R.A. Vierkant, L.L. Benetti,
Y. Tarabishy, K. Ghosh, D.W. Visscher, A.C. Degnim, Understanding the premalignant
potential of atypical hyperplasia through its natural history: a longitudinal cohort study,
Cancer Prev Res (Phila) 7 (2014) 211-217.
[13] Y. Feng, M. Spezia, S. Huang, C. Yuan, Z. Zeng, L. Zhang, X. Ji, W. Liu, B.
Huang, W. Luo, B. Liu, Y. Lei, S. Du, A. Vuppalapati, H.H. Luu, R.C. Haydon, T.-C.
He, G. Ren, Breast cancer developement and progression: Risk factors, cancer stem
cells, signaling pathways genomics and molecular pathogenesis, Genes Dis. 5(2) (2018)
77-106.
[14] L.M. Morimoto, E. White, Z. Chen, R.T. Chlebowski, J. Hays, L. Kuller, A.M.
Lopez, J. Manson, K.L. Margolis, P.C. Muti, M.L. Stefanick, A. McTiernan, Obesity,
body size, and risk of postmenopausal breast cancer: the Women's Health Initiative
(United States), Cancer Causes Control 13(8) (2002) 741-751, 2002.
[15] K.Y. Wolin, K. Carson, G.A. Colditz, Obesity and cancer, Oncologist 15(6) (2010)
556-565.
[16] D.A. Lawlor, G.D. Smith, S. Ebrahim, Hyperinsulinaemia and increased risk of
breast cancer: findings from the British Women's Heart and Health Study, Cancer
Causes Control 15(3) (2004) 267-275.
[17] N. Hamajima, K. Hirose, K. Tajima, et al., Alcohol, tobacco and breast cancer--
collaborative reanalysis of individual data from 53 epidemiological studies, including

I. Introduction
______________________________________________________________________
44
58,515 women with breast cancer and 95,067 women without the disease, Br J Cancer
87(11) (2002) 1234-1245.
[18] N. Andrieu, D.F. Easton, J. Chang-Claude, M.A. Rookus, R. Brohet, E. Cardis,
A.C. Antoniou, T. Wagner, J. Simard, G. Evans, S. Peock, J.-P. Fricker, C. Nogues, L.
Van't Veer, F.E. Van Leeuwen, D.E. Goldgar, Effect of chest X-rays on the risk of breast
cancer among BRCA1/2 mutation carriers in the international BRCA1/2 carrier cohort
study: a report from the EMBRACE, GENEPSO, GEO-HEBON, and IBCCS
Collaborators' Group, J Clin Oncol. 24(21) (2006) 3361-3366.
[19] S. Bhatia, L.L. Robison, O. Oberlin, M. Greenberg, G. Bunin, F. Fossati-Bellani,
A.T. Meadows, Breast cancer and other second neoplasms after childhood Hodgkin's
disease, N Engl J Med. 334(12) (1996) 745-751.
[20] S.L. Hancock, M.A. Tucker, R.T. Hoppe, Breast cancer after treatment of
Hodgkin's disease, J Natl Cancer Inst. 85(1) (1993) 25-31.
[21] L.B. Travis, D.A. Hill, G.M. Dores, M. Gospodarowicz, F.E. van Leeuwen, E.
Holowaty, B. Glimelius, M. Andersson, T. Wiklund, C.F. Lynch, M.B. Van't Veer, I.
Glimelius, H. Storm, E. Pukkala, M. Stovall, R. Curtis, J.D. Boice, E. Gilbertet, Breast
cancer following radiotherapy and chemotherapy among young women with Hodgkin
disease, JAMA 290(4) (2003) 465-475.
[22] R. Sankila, S. Garwicz, J.H. Olsen, H. Döllner, H. Hertz, A. Kreuger, F. Langmark,
M. Lanning, T. Möller, H. Tulinius, Risk of subsequent malignant neoplasms among
1,641 Hodgkin's disease patients diagnosed in childhood and adolescence: a population-
based cohort study in the five Nordic countries. Association of the Nordic Cancer
Registries and the Nordic Society of Pediatric Hematology and Oncology, J Clin Oncol.
14(5) (1996) 1442-1446.
[23] F.E. Van Leeuwen, W.J. Klokman, M. Stovall, E.C. Dahler, M.B. van't Veer, E.M.
Noordijk, M.A. Crommelin, B.M. Aleman, A. Broeks, M. Gospodarowicz, L.B. Travis,
N.S. Russell, Roles of radiation dose, chemotherapy, and hormonal factors in breast
cancer following Hodgkin's disease, J Natl Cancer Inst. 95(13) (2003) 971-980.
[24] E. Obedian, D.B. Fischer, B.G. Haffty, Second malignancies after treatment of
early-stage breast cancer: lumpectomy and radiation therapy versus mastectomy, J Clin
Oncol. 18(12) (2000) 2406-2412.
[25] B. Fisher, S. Anderson, J. Bryant, R.G. Margolese, M. Deutsch, E.R. Fisher, J.-H.
Jeong, N. Wolmark, Twenty-year follow-up of a randomized trial comparing total

I. Introduction
______________________________________________________________________
45
mastectomy, lumpectomy, and lumpectomy plus irradiation for the treatment of invasive
breast cancer, N Engl J Med. 347(16) (2002) 1233-1241.
[26] U. Veronesi, N. Cascinelli, L. Mariani, M. Greco, R. Saccozzi, A. Luini, M.
Aguilar, E. Marubini, Twenty-year follow-up of a randomized study comparing breast-
conserving surgery with radical mastectomy for early breast cancer, N Engl J Med.
347(16) (2002) 1227-1232.
[27] B. Fisher, J.H. Jeong, S. Anderson, J. Bryant, E.R. Fisher, N. Wolmark, Twenty-
five-year follow-up of a randomized trial comparing radical mastectomy, total
mastectomy, and total mastectomy followed by irradiation, N Engl J Med. 347(8)
(2002) 567-575.
[28] B.E. Henderson, M.C. Pike, R.K. Ross, et al.: Epidemiology and risk factors. In:
Bonadonna G, ed.: Breast Cancer: Diagnosis and Management. Chichester, NY: John
Wiley & Sons, 1984, pp 15-33.
[29] M.H. Gail, L.A. Brinton, D.P. Byar, D.K. Corle, S.B. Green, C. Schairer, J.J.
Mulvihill, Projecting individualized probabilities of developing breast cancer for white
females who are being examined annually, J Natl Cancer Inst. 81(24) (1989) 1879-1886.
[30] H.B. Nichols, M.J. Schoemaker, J. Cai, J. Xu, L.B. Wright, M.N. Brook, M.E.
Jones, H.-O. Adami, L. Baglietto, K.A. Bertrand, W.J. Blot, M.-C. Boutron-Ruault, M.
Dorronsoro, L. Dossus, A.H. Eliassen, G.G. Giles, I.T. Gram, S.E. Hankinson, J.
Hoffman-Bolton, R. Kaaks, T.J. Key, C.M. Kitahara, S.C. Larsson, M. Linet, M.A.
Merritt, R.L. Milne, V. Pala, J.R. Palmer, P.H. Peeters, E. Riboli, M. Sund, R.M.
Tamimi, A. Tjønneland, A. Trichopoulou, G. Ursin, L. Vatten, K. Visvanathan, E.
Weiderpass, A. Wolk, W. Zheng, C.R. Weinberg, A.J. Swerdlow, D.P. Sandler, Breast
Cancer Risk After Recent Childbirth: A Pooled Analysis of 15 Prospective Studies, Ann
Intern Med. 170(1) (2019) 22-30.
[31] Collaborative Group on Hormonal Factors in Breast Cancer, Breast cancer and
breastfeeding: collaborative reanalysis of individual data from 47 epidemiological
studies in 30 countries, including 50302 women with breast cancer and 96973 women
without the disease, Lancet 360(9328) (2002) 187-195.
[32] L. Bernstein, B.E. Henderson, R. Hanisch, J. Sullivan-Halley, R.K. Ross, Physical
exercise and reduced risk of breast cancer in young women, J Natl Cancer Inst. 86(18)
(1994) 1403-1408.

I. Introduction
______________________________________________________________________
46
[33] C.M. Friedenreich, Physical activity and cancer prevention: from observational to
intervention research, Cancer Epidemiol Biomarkers Prev. 10(4) (2001) 287-301.
[34] I. Thune, T. Brenn, E. Lund, M. Gaard, Physical activity and the risk of breast
cancer, N Engl J Med. 336(18) (1997) 1269-1275.
[35] L.L. Adams-Campbell, L. Rosenberg, R.S. Rao, J.R. Palmer, Strenuous physical
activity and breast cancer risk in African-American women, J Natl Med Assoc. 93(7-8)
(2001) 267-275.
[36] P.A. Van den Brandt, M. Schulpen, Mediterranean diet adherence and risk of
postmenopausal breast cancer: results of a cohort study and meta-analysis, Int. J Cancer
140(10) (2017) 2220-2231.
[37] S.G. Nayfield, J.E. Karp, L.G. Ford, F.A. Dorr, B.S. Kramer, Potential role of
tamoxifen in prevention of breast cancer, J Natl Cancer Inst. 83(20) (1991) 1450-1459.
[38] R.R. Love, H.S. Barden, R.B. Mazess, S. Epstein, R.J. Chappell, Effect of
tamoxifen on lumbar spine bone mineral density in postmenopausal women after 5
years. Arch Intern Med. 154(22) (1994) 2585-2588.
[39] T.J. Powles, T. Hickish, J.A. Kanis, A. Tidy, S. Ashley, Effect of tamoxifen on bone
mineral density measured by dual-energy x-ray absorptiometry in healthy
premenopausal and postmenopausal women, J Clin Oncol. 14(1) (1996) 78-84.
[40] J.P. Costantino, L.H. Kuller, D.G. Ives, B. Fisher, J. Dignam, Coronary heart
disease mortality and adjuvant tamoxifen therapy, J Natl Cancer Inst, 89(11) (1997)
776-782.
[41] C.C. McDonald, H.J. Stewart, Fatal myocardial infarction in the Scottish adjuvant
tamoxifen trial. The Scottish Breast Cancer Committee, BMJ 303(6800) (1991) 435-
437.
[42] L.E. Rutqvist, A. Mattsson, Cardiac and thromboembolic morbidity among
postmenopausal women with early-stage breast cancer in a randomized trial of adjuvant
tamoxifen. The Stockholm Breast Cancer Study Group, J Natl Cancer Inst. 85(17)
(1993) 1398-1406.
[43] C.K. Redmond, D.L. Wickerham, W. Cronin, et al, The NSABP breast cancer
prevention trial (BCPT): a progress report, Proc Annu Meet Am Soc Clin Oncol. 12(A-
78) (1993).
[44] B. Fisher, J.P. Costantino, D.L. Wickerham, C.K. Redmond, M. Kavanah, W.M.
Cronin, V. Vogel, A. Robidoux, N. Dimitrov, J. Atkins, M. Daly, S. Wieand, E. Tan-

I. Introduction
______________________________________________________________________
47
Chiu, L. Ford, N. Wolmark, Tamoxifen for prevention of breast cancer: report of the
National Surgical Adjuvant Breast and Bowel Project P-1 Study, J Natl Cancer Inst. 90
(18) (1998) 1371-1388.
[45] T. Powles, R. Eeles, S. Ashley, D. Easton, J. Chang, M. Dowsetta, A. Tidya, J.
Viggers, J. Davey, Interim analysis of the incidence of breast cancer in the Royal
Marsden Hospital tamoxifen randomised chemoprevention trial, Lancet 352(9122)
(1998) 98-101.
[46] U. Veronesi, P. Maisonneuve, A. Costa, V. Sacchini, C. Maltoni, C. Robertson, N.
Rotmensz, P. Boyle, Prevention of breast cancer with tamoxifen: preliminary findings
from the Italian randomised trial among hysterectomised women. Italian Tamoxifen
Prevention Study. Lancet 352(9122) (1998) 93-97.
[47] J. Cuzick, J.F. Forbes, I. Sestak, S. Cawthorn, H. Hamed, K. Holli, A. Howell,
Long-term results of tamoxifen prophylaxis for breast cancer--96-month follow-up of
the randomized IBIS-I trial, J Natl Cancer Inst. 99(4) (2007) 272-282.
[48] B. Fisher, J. Dignam, N, Wolmark, D.L. Wickerham, E. R. Fisher, E. Mamounas,
R. Smith, M. Begovic, N. V. Dimitrov, R.G. Margolese, C.G. Kardinal, M.T. Kavanah,
L. Fehrenbacher, R.H. Oishi, Tamoxifen in treatment of intraductal breast cancer:
National Surgical Adjuvant Breast and Bowel Project B-24 randomised controlled trial,
Lancet 353(9169) (1999) 1993-2000.
[49] W. Khovidhunkit, D.M. Shoback, Clinical effects of raloxifene hydrochloride in
women, Ann Intern Med. 130(5) (1999) 431-439.
[50] S. Martino, J.A. Cauley, E. Barrett-Connor, T.J. Powles, J. Mershon, D. Disch, R.J.
Secrest, S.R. Cummings, Continuing outcomes relevant to Evista: breast cancer
incidence in postmenopausal osteoporotic women in a randomized trial of raloxifene, J
Natl Cancer Inst 96(23) (2004) 1751-1761.
[51] D. Grady, J.A. Cauley, M.J. Geiger, M. Kornitzer, L. Mosca, P. Collins, N.K.
Wenger, J. Song, J. Mershon, E. Barrett-Connor, and the Raloxifene Use for The Heart
Trial, Reduced incidence of invasive breast cancer with raloxifene among women at
increased coronary risk, J Natl Cancer Inst. 100(12) (2008) 854-861.
[52] V.G. Vogel, J.P. Costantino, D.L. Wickerham, W.M. Cronin, R.S. Cecchini, J.N.
Atkins, T.B. Bevers, L. Fehrenbacher, E.R. Pajon Jr, J.L. Wade 3rd, A. Robidoux, R.G.
Margolese, J. James, S.M. Lippman, C.D. Runowicz, P.A. Ganz, S.E. Reis, W.
McCaskill-Stevens, L.G. Ford, V.C. Jordan, N. Wolmark, National Surgical Adjuvant

I. Introduction
______________________________________________________________________
48
Breast and Bowel Project (NSABP), Effects of tamoxifen vs raloxifene on the risk of
developing invasive breast cancer and other disease outcomes: the NSABP Study of
Tamoxifen and Raloxifene (STAR) P-2 trial, JAMA 295(23) (2006) 2727-2741.
[53] S.R. Land, D.L. Wickerham, J.P. Costantino, M.W. Ritter, V.G. Vogel, M. Lee, E.R.
Pajon, J.L. Wade 3rd, S. Dakhil, J.B. Lockhart Jr, N. Wolmark, P.A. Ganz, Patient-
reported symptoms and quality of life during treatment with tamoxifen or raloxifene for
breast cancer prevention: the NSABP Study of Tamoxifen and Raloxifene (STAR) P-2
trial, JAMA 295(23) (2006) 2742-2751.
[54] The ATAC Trialists' Group, Arimidex, tamoxifen alone or in combination:
Anastrozole alone or in combination with tamoxifen versus tamoxifen alone for
adjuvant treatment of postmenopausal women with early breast cancer: first results of
the ATAC randomised trial, Lancet 359(9324) (2002) 2131-2139.
[55] P.E. Goss, J.N. Ingle, S. Martino, et al., A randomized trial of letrozole in
postmenopausal women after five years of tamoxifen therapy for early-stage breast
cancer, N Engl J Med. 349(19) (2003) 1793-1802.
[56] P.E. Goss, J.N. Ingle, K.I. Pritchard, et al., Extending Aromatase-Inhibitor
Adjuvant Therapy to 10 Years, N Engl J Med. 375(3) (2016) 209-219.
[57] R.C. Coombes, E. Hall, L.J. Gibson, et al., A randomized trial of exemestane after
two to three years of tamoxifen therapy in postmenopausal women with primary breast
cancer, N Engl J Med. 350(11) (2004) 1081-1092.
[58] P.E. Goss, J.N. Ingle, J.E. Alés-Martínez, et al., Exemestane for breast-cancer
prevention in postmenopausal women, N Engl J Med. 364(25) (2011) 2381-2391.
[59] J. Cuzick, I. Sestak, J.F. Forbes, et al., Anastrozole for prevention of breast cancer
in high-risk postmenopausal women (IBIS-II): an international, double-blind,
randomised placebo-controlled trial, Lancet 383(9922) (2014) 1041-1048.
[60] L.C. Hartmann, D.J. Schaid, J.E. Woods, et al., Efficacy of bilateral prophylactic
mastectomy in women with a family history of breast cancer, N Engl J Med. 340(2)
(1999) 77-84.
[61] K.L. Kummerow, L. Du, D.F. Penson, et al., Nationwide trends in mastectomy for
early-stage breast cancer, JAMA Surg. 150(1) (2015) 9-16.
[62] T.R. Rebbeck, A.M. Levin, A. Eisen, et al., Breast cancer risk after bilateral
prophylactic oophorectomy in BRCA1 mutation carriers, J Natl Cancer Inst. 91(17)
(1999) 1475-1479.

I. Introduction
______________________________________________________________________
49
[63] N.D. Kauff, J.M. Satagopan, M.E. Robson, et al., Risk-reducing salpingo-
oophorectomy in women with a BRCA1 or BRCA2 mutation, N Engl J Med. 346(21)
(2002) 1609-1615.
[64] T.R. Rebbeck, H.T. Lynch, S.L. Neuhausen, et al., Prophylactic oophorectomy in
carriers of BRCA1 or BRCA2 mutations, N Engl J Med. 346(21) (2002) 1616-1622.
[65] N.D. Kauff, S.M. Domchek, T.M. Friebel, et al., Risk-reducing salpingo-
oophorectomy for the prevention of BRCA1- and BRCA2-associated breast and
gynecologic cancer: a multicenter, prospective study, J Clin Oncol. 26(8) (2008) 1331-
1337.
[66] E.E. Calle, C.W. Heath Jr., H.L. Miracle-McMahill, R.J. Coates, J.M. Liff, S.
Franceschi, R. Talamini, et al., Breast cancer and hormonal contraceptives: further
results. Collaborative Group on Hormonal Factors in Breast Cancer, Contraception
54(3) (1996) 1-106.
[67] P.A. Marchbanks, J.A. McDonald, H.G. Wilson, et al., Oral contraceptives and the
risk of breast cancer, N Engl J Med. 346(26) (2002) 2025-2032.
[68] B.L. Strom, J.A. Berlin, A.L. Weber, et al., Absence of an effect of injectable and
implantable progestin-only contraceptives on subsequent risk of breast cancer,
Contraception 69(5) (2004) 353-360.
[69] L.S. Mørch, C.W. Skovlund, P.C. Hannaford, et al., Contemporary Hormonal
Contraception and the Risk of Breast Cancer, N Engl J Med. 377(23) (2017) 2228-2239.
[70] D. Berry, Multiplicities in cancer research: ubiquitous and necessary evils, J Natl
Cancer Inst. 104(15) (2012) 1124-1132.
[71] K. Dickersin, The existence of publication bias and risk factors for its occurrence,
JAMA 263(10) (1990) 1385-1359.
[72] R.C. Travis, A. Balkwill, G.K. Fensom, et al., Night Shift Work and Breast Cancer
Incidence: Three Prospective Studies and Meta-analysis of Published Studies, J Natl
Cancer Inst. 108(12) (2016).
[73] C. Rodríguez Sanchez, R. Escala Cornejo, A. Olivares Hernández, Manual de
práctica clínica en Senología, 7 (2019) 61-64.
[74] Early Breast Cancer Trialists' Collaborative Group (EBCTCG), Effects of
chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year
survival: an overview of the randomised trials, Lancet 365(9472) (2005) 1687-1717.

I. Introduction
______________________________________________________________________
50
[75] M. Banys, N. Krawczyk, T. Fehm, The Role and Clinical Relevance of
Disseminated Tumor Cells in Breast Cancer, Cancers 6 (2014) 143-152.
[76] V.T. Jr DeVita, T.S. Lawrence, S.A. Rosenberg. Cancer: Principles and practice of
oncology. 9na edición. 2011. Philadelphia.
[77] I. González, J.M. García, Historia natural del cáncer de mama, Toko-Gin Pract.
61(5) (2002) 264-269.
[78] M. Solá, M. Margelí, E. Castellá, J.F. Julian, M. Rull, J.M. Gubern, et al.,
Prognostic value of hematogenous dissemination and biological profile of the tumor in
early breast cancer patients: A prospective observational study, BMC Cancer. 11 (2011)
252.
[79] M. Cianfrocca, L.J. Goldstein, Prognostic and Predictive Factors in Early-Stage
Breast, Oncologist 9 (2004) 606-616.
[80] S.D. Nathanson, D. Kwon, A. Kapke, S.H. Alford, D. Chitale, The Role of Lymph
Node Metastasis in the Systemic Dissemination of Breast Cancer, Indian J Surg Oncol.
1(4) (2010) 313-322.
[81] K. Kawada, M.M. Taketo, Significance and Mechanism of Lymph Node Metastasis
in Cancer progression, Cancer Res. 71 (2011) 1214-1218.
[82] I.J. Fidler, The Pathogenesis of cancer metastasis: the “seed and soil” hypothesis
revisited, Nat Rev Cancer 3(6) (2003) 453-458.
[83] O. Schmidt-Kittler, T. Ragg, A. Daskalakis, M. Granzow, A. Ahr, T.J.F.
Blankensteins, et al., From latent disseminated cells to overt metastasis: Genetic
analysis of systemic breast cancer progression, PNAS 100(13) (2003) 7737-7742.
[84] M. Solá, M. Margelí, E. Castellá, J.F. Julian, M. Rull, J.M. Gubern, et al.,
Prognostic value of hematogenous dissemination and biological profile of the tumor in
early breast cancer patients: A prospective observational study, BMC Cancer 11 (2011)
252.
[85] D. Schopper, C. de Wolf, How effective are breast cancer screening programmes
by mammography? Review of the current evidence, Eur J. Cancer 45 (2009) 1916-1923.
[86] B. Weigelt, J.L. Peterse, L.J. van‘t Veer, Breast cancer metastasis: markers and
models, Nat. Rev. Cancer 5 (2005) 591-602.
[87] M.T. Weigel, M. Dowsett, Current and emerging biomarkers in breast cancer:
prognosis and prediction, Endocr. Relat. Cancer 17 (2010) R245-R262.

I. Introduction
______________________________________________________________________
51
[88] J. Yang, J. Yang, Y. Gao, L. Zhao, L. Liu, Y. Qin, X. Wang, T. Song, C. Huang,
Identification of potential serum proteomic biomarkers for clear cell renal cell
carcinoma, PLoS One 9 (2014) e111364.
[89] M.J. Duffy, Serum tumor markers in breast cancer: are they of clinical value? Clin.
Chem. 52 (2006) 345-351.
[90] M.J. Duffy, D. Evoy, E.W. McDermott, CA 15-3: uses and limitation as a
biomarker for breast cancer, Clin. Chim. Acta 411 (2010) 1869-1874.
[91] T.H. Tsai, E. Song, R. Zhu, C.D. Poto, M. Wang, Y. Luo, R.S. Varghese, M.G.
Tadesse, D.H. Ziada, C.S. Desai, K. Shetty, Y. Mechref, H.W. Ressom, LC-MS/MS-
based serum proteomics for identification of candidate biomarkers for hepatocellular
carcinoma, Proteomics 15 (2015) 2369-2381.
[92] N.J. Fan, R. Kang, X.Y. Ge, M. Li, Y. Liu, H.M. Chen, C.F. Gao, Identification
alpha-2-HS-glycoprotein precursor and tubulin beta chain as serology diagnosis
biomarker of colorectal cancer, Diagn. Pathol. 12 (2014) 9-53.
[93] A.K. Callesen, J.S. Madsen, W. Vach, T.A. Kruse, Serum protein profiling by solid
phase extraction and mass spectrometry: a future diagnostics tool?, Proteomic 9 (2009)
1428-1441.
[94] W. Clarke, Z. Zhang, D.W. Chan, The application of clinical proteomics to cancer
and other diseases, Clin. Chem. Lab. Med. 41 (2003) 1562-1570.
[95] U. Menon, A. Ryan, J. Kalsi, A. Gentry-Maharaj, A. Dawnay, M. Habib, S.
Apostolidou, N. Singh, E. Benjamin, M. Burnell, S. Davies, A. Sharma, R. Gunu, K.
Godfrey, A. Lopes, D. Oram, J. Herod, K. Williamson, M.W. Seif, H. Jenkins, T.
Mould, R. Woolas, J.B. Murdoch, S. Dobbs, N.N. Amso, S. Leeson, D. Cruickshank, I.
Scott, L. Fallowfield, M. Widschwendter, K. Reynolds, A. McGuire, S. Campbell, M.
Parmar, S.J. Skates, I. Jacobs, Risk algorithm using serial biomarker measurements
doubles the number of screen-detected cancers compared with a single-threshold rule in
the United Kingdom Collaborative Trial of ovarian cancer screening, J. Clin. Oncol.
33(18) (2015) 2062-2071.
[96] S. Hammerström, The carcinoembryonic antigen (CEA) family: structures,
suggested functions and expression in normal and malignant tissues, Semin. Cancer
Biol. 9 (1999) 67-81.

I. Introduction
______________________________________________________________________
52
[97] F. Le Naour, D.E. Misek, M.C. Krause, L. Deneux, T.J. Giordano, S. Scholl, S.M.
Hanash, Proteomics-based identification of RS/DJ-1 as a novel circulating tumor
antigen in breast cancer. Clin. Cancer Res. 7(11) (2001) 3328-3335.
[98] R. Molina, J.M. Augé, J.M. Escudero, X. Filella, G. Zanon, J. Pahisa, B. Farrus, M.
Muñoz, M. Velasco, Evaluation of tumor markers (HER-2/neu oncoprotein, CEA, and
CA 15.3) in patients with locoregional breast cancer: prognostic value. Tumour Biol.
31(3) (2010) 171-180.
[99] L. Harris, H. Fritsche, R. Mennel, L. Norton, P. Ravdin, S. Taube, M.R.
Somerfield, D.F. Hayes, R.C. Bast Jr., American Society of Clinical Oncology update
of recommendations for the use of tumor markers in breast cancer, J. Clin. Oncol. 25
(2010) 5287-5312.
[100] J. Li, Z. Zhang, J. Rosenzweig, Y.Y. Wang, D.W. Chan, Proteomics and
bioinformatics approaches for identification of serum biomarkers to detect breast
cancer, Clin. Chem. 48 (2002) 1296‑1304.
[101] D.D. Gioia, M. Dresse, D. Mayr, D. Nagel, V. Heinemann, P. Stieber, Serum
HER2 in combination with CA 15-3 as a parameter for prognosis in patients with early
breast cancer, Clin. Chim. Acta 440 (2015) 16-22.
[102] R. Akter, B. Jeong, J.S. Choi, M.A. Rahman, Ultrasensitive Nanoimmunosensor
by coupling non-covalent functionalized graphene oxide platform and numerous ferritin
labels on carbon nanotubes, Biosens Bioelectron 80 (2016) 123-130.
[103] C.S. Lee, N.A. Taib, A. Ashrafzadeh, F. Fadzli, F. Harun, K. Rahmat, S.M.
Hoong, P.S. Abdul-Rahman, O.H. Hashim, Unmasking Heavily O-Glycosylated Serum
Proteins Using Perchloric Acid: Identification of Serum Proteoglycan 4 and Protease C1
Inhibitor as Molecular Indicators for Screening of Breast Cancer, PLoS One 11(2)
(2016) e0149551.
[104] A.W. Opstal-van Winden, W. Rodenburg, J.L. Pennings, C.T. van Oostrom, J.H.
Beijnen, P.H. Peeters, C.H. van Gils, A. de Vries, A bead-based multiplexed
immunoassay to evaluate breast cancer biomarkers for early detection in pre-diagnostic
serum, Int. J. Mol. Sci. 13(10) (2012) 13587-13604.
[105] A. Kazarian, O. Blyuss, G. Metodieva, A. Gentry-Maharaj, A. Ryan, E.M.
Kiseleva, O.M. Prytomanova, I.J. Jacobs, M. Widschwendter, U. Menon, J.F. Timms,
Testing breast cancer serum biomarkers for early detection and prognosis in pre-
diagnosis samples, Br. J. Cancer 116(4) (2017) 501-508.

I. Introduction
______________________________________________________________________
53
[106] X. Zuo, L. Chen, L. Liu, Z. Zhang, X. Zhang, Q. Yu, L. Feng, X. Zhao, T. Qin,
Identification of a panel of complex autoantigens (LGALS3, PHB2, MUC1, and GK2)
in combination with CA15-3 for the diagnosis of early-stage breast cancer, Tumour
Biol. 37(1) (2016) 1309-1317.
[107] T. Yigitbasi, G. Calibasi-Kocal, N. Buyukuslu, M.K. Atahan, H. Kupeli, S. Yigit,
E. Tarcan, Y. Baskin, An efficient biomarker panel for diagnosis of breast cancer using
surface-enhanced laser desorption ionization time-of-flight mass spectrometry, Biomed.
Rep. 8(3) (2018) 269-274.
[108] A.C. Pedersen, P.D. Sørensen, E.H. Jacobsen, J.S. Madsen, I. Brandslund,
Sensitivity of CA 15-3, CEA and serum HER2 in the early detection of recurrence of
breast cancer, Clin. Chem. Lab. Med. 51(7) (2013) 1511-1519.
[109] K. Atahan, H. Küpeli, S. Gür, T. Yiğitbaşı, Y. Baskın, S. Yiğit, M. Deniz, A.
Cökmez, E. Tarcan, The value of serum biomarkers (Bc1, Bc2, Bc3) in the diagnosis of
early breast cance, Int. J. Med. Sci. 8(2) (2011) 148-155.
[110] C. Desmetz, F. Bibeau, F. Boissiere, Proteomics based identification of HSP60 as
a tumor-associated antigen in early-stage breast cancer and ductal carcinoma in situ, J.
Proteome Res. 7 (2008) 3830-3837.
[111] B.W. Park, J.W. Oh, J.H. Kim, S.H. Park, K.S. Kim, J.H. Kim, K.S. Lee,
Preoperative CA15-3 and CEA serum levels as predictor for breast cancer outcomes,
Ann. Oncol. 19 (2008) 675-681.
[112] J.W. Chang, U.B. Kang, D.H. Kim, J.K. Yi, J.W. Lee, D.Y. Noh, C. Lee, M.H.
Yu, Identification of circulating endorepellin LG3 fragment: potential use as a
serological biomarker for breast cancer, Proteomics Clin. Appl. 2 (2008) 23-32.
[113] F. Fernández-Madrid, Autoantibodies in breast cancer sera: candidate biomarkers
and reporters of tumourigenesis, Cancer Lett. 230 (2007) 187-198.
[114] M. Volkmann, H.P. Sinn, D. Gaugel, Anti-p53 in breast cancer: concordance of
different assay procedures and association with p53 antigen expression, Oncology 63
(2002) 297-305.
[115] W. Cho, Contribution of oncoproteomics to cancer biomarker discovery, Mol.
Cancer 6(25) (2007) 1-13.
[116] V. Levenson, Biomarkers for early detection of breast cancer: what, when, and
where?, Biochim. Biophys. Acta 1770 (2007) 847-856.

I. Introduction
______________________________________________________________________
54
[117] L. Shi, T. Gehin, Y. Chevolot, E. Souteyrand, A. Mangé, J. Solassol, E.
Laurenceau, Anti-heat shock protein autoantibody profiling in breast cancer using
customized protein microarray, Anal. Bioanal. Chem. 408(5) (2016) 1497-1506.
[118] A.L. Fernández-Grijalva, A. Aguilar-Lemarroy, L.F. Jave-Suarez, A. Gutiérrez-
Ortega, P.A. Godinez-Melgoza, S.E. Herrera-Rodríguez, I. Mariscal-Ramírez, M.
Martínez-Velázquez, M.A. Gawinowicz, M.G. Martínez-Silva, J.A. Cruz-Ramos, R.
Hernández-Gutiérrez, Alpha 2HS-glycoprotein, a tumor-associated antigen (TAA)
detected in Mexican patients with early-stage breast cancer, J. Proteomics 112 (2015)
301-312.
[119] M.C. Henderson, A.B. Hollingsworth, K. Gordon, M. Silver, R. Mulpuri, E.
Letsios, D.E. Reese, Integration of Serum Protein Biomarker and Tumor Associated
Autoantibody Expression Data Increases the Ability of a Blood-Based Proteomic Assay
to Identify Breast Cancer, PLoS One 11(8) (2016) e0157692.
[120] M. Pla-Roca, R.F. Leulmi, S. Tourekhanova, S. Bergeron, V. Laforte, E. Moreau,
S.J. Gosline, N. Bertos, M. Hallett, M. Park, D. Juncker, Antibody colocalization
microarray: a scalable technology for multiplex protein analysis in complex samples,
Mol. Cell Proteomics 11(4) (2012) M111.011460.
[121] R.W. Stephens, A.N. Pedersen, H.J. Nielsen, M.J. Hamers, G. Hoyer-Hansen, E.
Ronne, E. Dybkjaer, K. Dano, N. Brünner, ELISA determination of soluble urokinase
receptor in blood from healthy donors and cancer patients, Clin. Chem. 43 (1997) 1868-
76.
[122] D. Aderka, H. Englemann, V. Hornik, Y. Skornick, Y. Levo, D. Wallach, G.
Kushtai, Increased serum levels of soluble receptors for tumor necrosis factor in cancer
patients, Cancer Res. 51 (1991) 5602-5607.
[123] B.K. Kim, J.W. Lee, P.J. Park, Y.S. Shin, W.Y. Lee, K.A. Lee, S. Ye, H. Hyun,
K.N. Kang, D. Yeo, Y. Kim, S.Y. Ohn, D.Y. Noh, C.W. Kim, The multiplex bead array
approach to identifying serum biomarkers associated with breast cancer, Breast Cancer
Res. 11 (2009) R22.
[124] M.N. Vo, M. Evans, K. Leitzel, S.M. Ali, M. Wilson, L. Demers, D.B. Evans, A.
Lipton, Elevated plasma endoglin (CD105) predicts decreased response and survival in
a metastatic breast cancer trial of hormone therapy, Breast Cancer Res. Treat. 119
(2010) 767-771.

I. Introduction
______________________________________________________________________
55
[125] F. Zhang, J.Y. Chen, A method for identifying discriminative isoform-specific
peptides for clinical proteomics application, BMC Genomics 17(7) (2016) 522.
[126] D.L. Wang, C. Xiao, G. Fu, X. Wang, L. Li, Identification of potential serum
biomarkers for breast cancer using a functional proteomics technology, Biomark. Res. 5
(2017) 11.
[127] G.L. Hortin, The MALDI-TOF mass spectrometric view of the plasma proteome
and peptidome, Clin. Chem. 52 (2006) 1223-1237.
[128] M.C. Gast, J.H. Schellens, J.H. Beijnen,Clinical proteomics in breast cancer: a
review, Breast Cancer Res. Treat. 116 (2009) 17-29.
[129] C. Núñez, M.P. Chantada-Vázquez, S.B. Bravo, S. Vázquez-Estévez, Novel
functionalized nanomaterials for the effective enrichment of proteins and peptides with
post-translational modifications, J. Proteomics 181 (2018) 170-189.
[130] Y. Li, X. Zhang, C. Deng, Functionalized magnetic nanoparticles for sample
preparation in proteomics and peptidomics analysis, Chem. Soc. Rev. 42(21) (2013)
8517-39.
[131] M. Gao, C. Deng, X. Zhang, Magnetic nanoparticles-based digestion and
enrichment methods in proteomics analysis, Expert Rev. Proteomics 8(3) (2011) 379-
390.
[132] J. Villanueva, J. Philip, D. Entenberg, C.A. Chaparro, M.K. Tanwar, E.C.
Holland, P. Tempst, Serum peptide profiling by magnetic particle-assisted, automated
sample processing and MALDI-TOF mass spectrometry, Anal. Chem. 76 (2004) 1560-
1570.
[133] M.P. Ebert, D. Niemeyer, S.O. Deininger, T. Wex, C. Knippig, J. Hoffmann, J.
Sauer, W. Albrecht, P. Malfertheiner, C. Röcken, Identification and confirmation of
increased fibrinopeptide a serum protein levels in gastric cancer sera by magnet bead
assisted MALDI-TOF mass spectrometry, J. Proteome Res. 5 (2006) 2152-2158.
[134] N. Yao, H. Chen, H. Lin, C. Deng, X. Zhang, Enrichment of peptides in serum by
C8-functionalized magnetic nanoparticles for direct matrix-assisted laser
desorption/ionization time-of-flight mass spectrometry analysis, J. Chromatogr. A 1185
(2008) 93-101.
[135] M.E. de Noo, A. Deelder, M. van der Werff, A. Ozalp, B. Mertens, R. Tollenaar,
MALDI-TOF serum protein profiling for the detection of breast cancer, Onkologie 29
(2006) 501-506.

I. Introduction
______________________________________________________________________
56
[136] S. Shin, L. Cazares, H. Schneider, S. Mitchell, C. Laronga, O.J. Semmes, R.R.
Perry, R.R. Drake, Serum biomarkers to differentiate benign and malignant
mammographic lesions, J. Am. Coll. Surg. 204(5) (2007) 1065-1071.
[137] D. Böhm, K. Keller, N. Wehrwein, A. Lebrecht, M. Schmidt, H. Kölbl, F.H. Grus,
Serum proteome profiling of primary breast cancer indicates a specific biomarker
profile, Oncol. Rep. 26(5) (2011) 1051-1056.
[138] J. Song, M. Patel, C.N. Rosenzweig, Y. Chan-Li, L.J. Sokoll, E.T. Fung, N.H.
Choi-Miura, M. Goggins, D.W. Chan, Z. Zhang, Quantification of fragments of human
serum inter-alpha-trypsin inhibitor heavy chain 4 by a surface-enhanced laser
desorption/ionization-based immunoassay, Clin. Chem. 52 (2006) 1045-1053.
[139] J. Peng, A.J. Stanley, D. Cairns, P.J. Selby, R.E. Banks, Using the protein chip
interface with quadrupole time-of-flight mass spectrometry to directly identify peaks in
SELDI profiles-initial evaluation using low molecular weight serum peaks, Proteomics
9 (2009) 492-498.
[140] J. Villanueva, D.R. Shaffer, J. Philip, C.A. Chaparro, H. Erdjument-Bromage,
A.B. Olshen, M. Fleisher, H. Lilja, E. Brogi, J. Boyd, M. Sánchez-Carbayo, E.C.
Holland, C. Cordon-Cardo, H.I. Scher, P. Tempst, Differential exoprotease activities
confer tumor-specific serum peptidome patterns, J. Clin. Invest. 116 (2006) 271-284.
[141] Y. Fan, L. Shi, Q. Liu, R. Dong, Q. Zhang, S. Yang, Y. Fan, H. Yang, P. Wu, J.
Yu, S. Zheng, F. Yang, J. Wang, Discovery and identification of potential biomarkers of
papillary thyroid carcinoma, Mol. Cancer 8 (2009) 79.
[142] D.G. Ward, N. Suggett, Y. Cheng, W. Wei, H. Johnson, L.J. Billingham, T.
Ismail, M.J. Wakelam, P.J. Johnson, A. Martin, Identification of serum biomarkers for
colon cancer by proteomic analysis, Br. J. Cancer 94 (2006) 1898-1905.
[143] S.Y. Yang, X.Y. Xiao, W.G. Zhang, L.J. Zhang, W. Zhang, B. Zhou, G. Chen,
D.C. He, Application of serum SELDI proteomic patterns in diagnosis of lung cancer,
BMC Cancer 5 (2005) 83.
[144] B. Velstra, Y.E. van der Burgt, B.J. Mertens, W.E. Mesker, A.M. Deelder, R.A.
Tollenaar, Improved classification of breast cancer peptide and protein profiles by
combining two serum workup procedures, J. Cancer Res. Clin. Oncol. 138(12) (2012)
1983-1992.

I. Introduction
______________________________________________________________________
57
[145] J. Yang, X. Xiong, S. Liu, J. Zhu, M. Luo, L. Liu, L. Zhao, Y. Qin, T. Song, C.
Huang, Identification of novel serumpeptides biomarkers for female breast cancer
patients in Western China, Proteomics 16(6) (2016) 925-934.
[146] C.L. Washam, S.D. Byrum, K. Leitzel, S.M. Ali, A.J. Tackett, D. Gaddy, S.E.
Sundermann, A. Lipton, L.J. Suva, Identification of PTHrP(12-48) as a plasma
biomarker associated with breast cancer bone metastasis, Cancer Epidemiol.
Biomarkers Prev. 22(5) (2013) 972-983.
[147] Y. Sun, J. Zhang, F. Guo, W. Zhao, Y. Zhan, C. Liu, Y. Fan, J. Wang,
Identification of Apolipoprotein C-I Peptides as a Potential Biomarker and its
Biological Roles in Breast, Med. Sci. Monit. 22 (2016) 1152-1160.
[148] N.J. Fan, C.F. Gao, G. Zhao, X.L. Wang, Q.Y. Liu, Serum peptidome patterns of
breast cancer based on magnetic bead separation and mass spectrometry analysis,
Diagn. Pathol. 7 (2012) 45.
[149] A.W. Opstal-van Winden, E.J. Krop, M.H. Kåredal, M.C. Gast, C.H. Lindh, M.C.
Jeppsson, B.A. Jönsson, D.E. Grobbee, P.H. Peeters, J.H. Beijnen, C.H. van Gils, R.C.
Vermeulen, Searching for early breast cancer biomarkers by serum protein profiling of
pre-diagnostic serum; a nested case-control study, BMC Cancer 11 (2011) 381.
[150] A.W. Opstal-van Winden, J.H. Beijnen, Loof A, W.L. van Heerde, R. Vermeulen,
H.M. Peeters Petra, C.H. van Gils, Search for breast cancer biomarkers in fractionated
serum samples by protein profiling with SELDI-TOF MS, J. Clin. Lab. Anal. 26 (2012)
1-9.
[151] J. Yang, J. Zhu, K. He, L.Y. Zhao, L.Y. Liu, T.S. Song, C. Huang, Proteomic
Profiling of Invasive Ductal Carcinoma (IDC) using Magnetic Beads-based Serum
Fractionation and MALDI-TOF MS, J. Clin. Lab. Anal. 29(4) (2015) 321-327.
[152] P. Dowling, V. Palmerini, M. Henry, P. Meleady, V. Lynch, J. Ballot, G. Gullo, J.
Crown, M. Moriarty, M. Clynes, Transferrin-bound proteins as potential biomarkers for
advanced breast cancer patients, BBA Clin 2 (2014) 24-30.
[153] M. Pietrowska, L. Marczak, J. Polanska, K. Behrendt, E. Nowicka, A.
Walaszczyk, A. Chmura, R. Deja, M. Stobiecki, A. Polanski, R. Tarnawski, P.J.
Widlak, Mass spectrometry-based serum proteome pattern analysis in molecular
diagnostics of early-stage breast cancer, J. Transl. Med. 7 (2009) 60.
[154] B. Zeidan, T.R. Jackson, S.E. Larkin, R.I. Cutress, G.R. Coulton, M. Ashton-Key,
N. Murray, G. Packham, V. Gorgoulis, S.D. Garbis, P.A. Townsend, Annexin A3 is a

I. Introduction
______________________________________________________________________
58
mammary marker and a potential neoplastic breast cell therapeutic target, Oncotarget
6(25) (2015) 21421-21427.
[155] D. Galasko, T.E. Golde, Biomarkers for Alzheimer’s disease in plasma, serum
and blood-Conceptual and practical problems, Alzheimers Res. Ther. 5 (2013) 10.
[156] F.S. Berven, R. Ahmad, K.R. Clauser, S.A. Carr, Optimizing performance of
glycopeptide capture for plasma proteomics, J. Proteome Res. 9 (2010) 1706-1715.
[157] S. Chen, C. Lu, H. Gu, A. Mehta, J. Li, P.B. Romano, D. Horn, D.C. Hooper,
C.R. Bazemore-Walker, T. Block, Aleuria aurantia lectin (AAL)-reactive
immunoglobulin G rapidly appears in sera of animals following antigen exposure, PloS
One 7 (2012) e44422.
[158] H. Huang, S. Lin, B.A. Garcia, Y. Zhao, Quantitative proteomic analysis of
histone modifications, Chemical Rev. 115 (2015) 2376-2418.
[159] M.J. Bissell, W.C. Hines, Why don’t we get more cancer? A proposed role of the
microenvironment in restraining cancer progression, Nat. Med. 17(3) (2011) 320-329.
[160] I.O. Potapenko, V.D. Haakensen, T. Luders, A. Helland, I. Bukholm, T. Sorlie,
V.N. Kristensen, O.C. Lingjaerde, A.L. Borresen-Dale, Glycan gene expression
signatures in normal and malignant breast tissue; possible role in diagnosis and
progression, Mol. Oncol. 4(2) (2010) 98-118.
[161] K. Marino, R. Saldova, B. Adamczyk, P.M. Rudd, Changes in serum N-
glycosylation profiles: functional significance and potential for diagnostics. In
Carbohydrate Chemistry: Chemical and Biological Approaches; Rauter AP, Ed.; RSC
Publishing: Cambridge, U.K., 2012; Vol. 37.
[162] J. Hirabayashi, K. Kasai, Separation technologies for glycomics, J.
Chromatography B Analyt. Technol. Biomed. Life Sci. 771 (2002) 67-87.
[163] J. Bunkenborg, B.J. Pilch, A.V. Podtelejnikov, J.R. Wisniewski, Screening for N-
glycosylated proteins by liquid chromatography mass spectrometry, Proteomics 4
(2004) 454-65.
[164] H. Zhang, E.C. Yi, X. Li, P. Mallick, K.S. Kelly-Spratt, C.D. Masselon, D.G.
Camp, R.D. Smith, C.J. Kemp, R. Aebersold, High throughput quantitative analysis of
serum proteins using glycopeptide capture and liquid chromatography mass
spectrometry, Mol. Cell Proteomics 4 (2005) 144-155.
[165] Z. Zeng, M. Hincapie, S.J. Pitteri, S. Hanash, J. Schalkwijk, J.M. Hogan, H.
Wang, W.S. Hancock, A proteomics platform combining depletion, multi-lectin affinity

I. Introduction
______________________________________________________________________
59
chromatography (M-LAC), and isoelectric focusing to study the breast cancer proteome,
Anal. Chem. 83(12) (2011) 4845-4854.
[166] S. Selvaraju, Z. El Rassi, Targeting human serum fucome by an integrated liquid-
phase multicolumn platform operating in "cascade" to facilitate comparative mass
spectrometric analysis of disease-free and breast cancer sera, Proteomics 13(0) (2013)
1701-1713.
[167] S.A. Fry, J. Sinclair, J.F. Timms, A.J. Leathem, M.V. Dwek, A targeted
glycoproteomic approach identifies cadherin-5 as a novel biomarker of metastatic breast
cancer, Cancer Lett. 328 (2013) 335-344.
[168] K. Marino, J. Bones, J.J. Kattla, P.M. Rudd, A systematic approach to protein
glycosylation analysis: a path through the maze, Nat. Chem. Biol. 6(10) (2010) 713-
723.
[169] R. Saldova, A. Asadi Shehni, V.D. Haakensen, I. Steinfeld, M. Hilliard, I. Kifer,
A. Helland, Z. Yakhini, A.L. Børresen-Dale, P.M. Rudd, Association of N-
glycosylation with breast carcinoma and systemic features using high-resolution
quantitative UPLC, J Proteome Res. 13 (2014) 2314-2327.
[170] N. Kawaguchi-Sakita, K. Kaneshiro-Nakagawa, M. Kawashima, M. Sugimoto,
M. Tokiwa, E. Suzuki, S. Kajihara, Y. Fujita, S. Iwamoto, K. Tanaka, M. Toi, Serum
immunoglobulin G Fc region N-glycosylation profiling by matrix-assisted laser
desorption/ionization mass spectrometry can distinguish breast cancer patients from
cancer-free controls, Biochem. Biophys. Res.Commun 469(4) (2016) 1140-1145.
[171] A.B. Iliuk, J.V. Arrington, W. Tao, Analytical challenges translating mass
spectrometry-based phosphoproteomics from discovery to clinical applications,
Electrophoresis 35(24) (2014) 3430-3440.
[172] I.H. Chen, L. Xue, C.C. Hsu, J.S. Paez, L. Pan, H. Andaluz, M.K. Wendt, A.B.
Iliuk, J.K. Zhu, W.A. Tao, Phosphoproteins in extracellular vesicles as candidate
markers for breast cancer, Proc. Natl. Acad. Sci. USA. 114(12) (2017) 3175-3180.
[173] T. Yi, B. Zhai, Y. Yu, Y. Kiyotsugu, T. Raschle, M. Etzkorn, H. Seo, M. Nagiec,
R.E. Luna, E.L. Reinherz, J. Blenis, S.P. Gygi, G. Wagner, Quantitative
phosphoproteomic analysis reveals system-wide signaling pathways downstream of
SDF-1/CXCR4 in breast cancer stem cells, Proc. Natl. Acad. Sci. USA 111(21) (2014)
E2182-E2190.

I. Introduction
______________________________________________________________________
60
[174] H. Gu, J.M. Ren, X. Jia, T. Levy, K. Rikova, V. Yang, K.A. Lee, M.P. Stokes,
J.C. Silva, Quantitative Profiling of Post-translational Modifications by Immunoaffinity
Enrichment and LC-MS/MS in Cancer Serum without Immunodepletion, Mol. Cell
Proteomics 15(2) (2016) 692-702.
[175] T. Sørlie, R. Tibshirani, J. Parker, T. Hastie, J.S. Marron, A. Nobel, S. Deng, H.
Johnsen, R. Pesich, S. Geisler, J. Demeter, C.M. Perou, P.E. Lønning, P.O. Brown, A.L.
Børresen-Dale, D. Botstein, Repeated observation of breast tumor subtypes in
independent gene expression data sets, Proc. Natl. Acad. Sci. USA 100 (2003) 8418-
8423.
[176] M. Yanagawa, K. Ikemot, S. Kawauchi, T. Furuya, S. Yamamoto, M. Oka, A.
Oga, Y. Nagashima, K. Sasaki, Luminal A and luminal B (HER2 negative) subtypes of
breast cancer consist of a mixture of tumors with different genotype, BMC Res. Notes 5
(2012) 376.
[177] Z. Inic, M. Zegarac, M. Inic, I. Markovic, Z. Kozomara, I. Djurisic, I. Inic,
G. Pupic, S. Jancic, Difference between luminal A and luminal B subtypes according to
Ki-67, tumor size, and progesterone receptor negativity providing prognostic
information, Clin. Med. Insights Oncol. 8 (2014) 107-111.
[178] C. Sotiriou, M.J. Piccart, Taking gene-expression profiling to the clinic: when will
molecular signatures become relevant to patient care? Nat. Rev. Cancer 7 (2007) 545-
553.
[179] L.A. Carey, C.M. Perou, C.A. Livasy, L.G. Dressler, D. Cowan, K. Conway, G.
Karaca, M.A. Troester, C.K. Tse, S. Edmiston, S.L. Deming, J. Geradts, M.C.
Cheang, T.O. Nielsen, P.G. Moorman, H.S. Earp, R.C. Millikan, Race, breast cancer
subtypes, and survival in the carolina breast cancer study, JAMA 295 (2006) 2492-
2502.
[180] K.M. O’Brien, S.R. Cole, C.K. Tse, C.M. Perou, L.A. Carey, W.D. Foulkes, L.G.
Dressler, J. Geradts, R.C. Millikan, Intrinsic breast tumor subtypes, race, and long-term
survival in the Carolina Breast Cancer Study, Clin. Cancer Res. 16 (2010) 6100-6110.
[181] H. Nakshatri, G. Qi, J. You, B. Kerry, B. Schneider, R. Zon, C. Buck, F. Regnier,
M. Wang, Intrinsic subtype-associated changes in the plasma proteome in breast cancer,
Proteomics Clin. Appl. 3(11) (2009) 1305-1313.
[182] F. Zhang, J.Y. Chen, Breast cancer subtyping from plasma proteins, BMC Med.
Genomics 6(1) (2013) S6.

I. Introduction
______________________________________________________________________
61
[183] A. Gajbhiye, R. Dabhi, K. Taunk, M.G. Jagadeeshaprasad, S. RoyChoudhury, A.
Mane, S. Bayatigeri, K. Chaudhury, M.K. Santra, S. Rapole, Multipronged quantitative
proteomics reveals serum proteome alterations in breast cancer intrinsic subtypes, J.
Proteomics 163 (2017) 1-13.
[184] S.J. Pitteri, L.M. Amon, T. Busald Buson, Y. Zhang, M.M. Johnson, A. Chin, J.
Kennedy, C.H. Wong, Q. Zhang, H. Wang, P.D. Lampe, R.L. Prentice, M.W. McIntosh,
S.M. Hanash, C.I. Li, Detection of elevated plasma levels of epidermal growth factor
receptor before breast cancer diagnosis among hormone therapy users, Cancer Res.
70(21) (2010) 8598-8606.
[185] L.M. Amon, S.J. Pitteri, C.I. Li, M. McIntosh, J.J. Ladd, M. Disis, P. Porter, C.H.
Wong, Q. Zhang, P. Lampe, R.L. Prentice, S.M. Hanash, Concordant release of
glycolysis proteins into the plasma preceding a diagnosis of ER+ breast cancer, Cancer
Res. 72(8) (2012) 1935-1942.
[186] R.M. González, D.S. Daly, R. Tan, J.R. Marks, R.C. Zangar, Plasma biomarker
profiles differ depending on breast cancer subtype but RANTES is consistently
increased, Cancer Epidemiol. Biomarkers Prev. 20(7) (2011) 1543-1551.
[187] C.I. Li, J.E. Mirus, Y. Zhang, A.B. Ramirez, J.J. Ladd, R.L. Prentice, M.W.
McIntosh, S.M. Hanash, P.D. Lampe, Discovery and preliminary confirmation of novel
early detection biomarkers for triple-negative breast cancer using preclinical plasma
samples from the women’s health initiative observational study, Breast Cancer Res.
Treat. 135(2) (2012) 611-618.
[188] L. Chen, W. Zhao, J. He, L. Li, D. Meng, D. Cai, J. Yu, D. Chen, Y. Wu, T.
Zhou, Label-free quantitative proteomic screening of candidate plasma biomarkers for
the prognosis of breast cancer with different lymph node statuses, Proteomics Clin.
Appl. 12(3) (2018) e1700117.
[189] M. Imamura, A. Nishimukai, T. Higuchi, H. Ozawa, A. Yanai, Y. Miyagawa, K.
Murase, I. Sakita, T. Hatada, Y. Takatsuka, T. Katagiri, Y. Miyoshi, High levels at
baseline of serum pyridinoline crosslinked carboxyterminal telopeptide of type I
collagen are associated with worse prognosis for breast cancer patients, Breast Cancer
Res. Treat. 154 (2015) 521-531.
[190] S. Byström, M. Eklund, M.G. Hong, C. Fredolini, M. Eriksson, K. Czene, P. Hall,
J.M. Schwenk, M. Gabrielson, Affinity proteomic profiling of plasma for proteins

I. Introduction
______________________________________________________________________
62
associated to area-based mammographic breast density, Breast Cancer Res. 20(1) (2018)
14.
[191] D. Scumaci, L. Tammè, C.V. Fiumara, G. Pappaianni, A. Concolino, E. Leone,
M.C. Faniello, B. Quaresima, E. Ricevuto, F.S. Costanzo, G. Cuda, Plasma proteomic
profiling in hereditary breast cancer reveals a BRCA1-specific signature: diagnostic and
functional implications, PLoS One 10(6) (2015) e0129762.
[192] J. Fan, M.M. Tea, C. Yang, L. Ma, Q.H. Meng, T.Y. Hu, C.F. Singer, M. Ferrari,
Profiling of cross-functional peptidases regulated circulating peptides in BRCA1 mutant
breast cancer, J Proteome Res. 15(5) (2016) 1534-1545.
[193] S. Khan, H.F. Bennit, D. Turay, M. Perez, S. Mirshahidi, Y. Yuan, N.R. Wall,
Early diagnostic value of survivin and its alternative splice variants in breast cancer,
BMC Cancer 14 (2014) 176.
[194] M. Giussani, E. Landoni, G. Merlino, F. Turdo, S. Veneroni, B. Paolini, V.
Cappelletti, R. Miceli, R. Orlandi, T. Triulzi, E. Tagliabue, Extracellular matrix proteins
as diagnostic markers of breast carcinoma, J. Cell Physiol. 233(8) (2018) 6280-6290.
[195] A.K. Callesen, W. Vach, P.E. Jørgensen, S. Cold, Q. Tan, R. Depont Christensen,
O. Mogensen, T.A. Kruse, O.N. Jensen, J.S. Madsen, Combined experimental and
statistical strategy for mass spectrometry based serum protein profiling for diagnosis of
breast cancer: a case-control study, J. Proteome Res. 7 (2008) 1419-1426.
[196] M.C. Gast, H. van Tinteren, M. Bontenbal, R.Q. van Hoesel, M.A. Nooij, S.
Rodenhuis, P.N. Span, V.C. Tjan-Heijnen, E.G. de Vries, N. Harris, J.W. Twisk, J.H.
Schellens, J.H. Beijnen, Haptoglobin phenotype is not a predictor of recurrence free
survival in high-risk primary breast cancer patients., BMC Cancer 8 (2008) 389.
[197] A. Gonçalves, B. Esterni, F. Bertucci, R. Sauvan, C. Chabannon, M. Cubizolles,
V.J. Bardou, G. Houvenaegel, J. Jacquemier, S. Granjeaud, X.Y. Meng, E.T. Fung, D.
Birnbaum, D. Maraninchi, P. Viens, J.P. Borg, Postoperative serum proteomic profiles
may predict metastatic relapse in high-risk primary breast cancer patients receiving
adjuvant chemotherapy, Oncogene 25 (2006) 981-989.
[198] F. Boccardo, A. Rubagotti, P.V. Nuzzo, F. Argellati, G. Savarino, P. Romano, G.
Damonte, M. Rocco, A. Profumo, Matrix-assisted laser desorption/ionisation (MALDI)
TOF analysis identifies serum angiotensin II concentrations as a strong predictor of all-
cause and breast cancer (BCa)-specific mortality following breast surgery, Int. J. Cancer
137(10) (2015) 2394-2402.

I. Introduction
______________________________________________________________________
63
[199] L. Chung, K. Moore, L. Phillips, F.M. Boyle, D.J. Marsh, R.C. Baxter, Novel
serum protein biomarker panel revealed by mass spectrometry and its prognostic value
in breast cancer, Breast Cancer Res. 16(3) (2014) R63.
[200] C.M. Pfeiffer, J.D. Cook, Z. Mei, M.E. Cogswell, A.C. Looker, D.A. Lacher,
Evaluation of an automated soluble transferrin receptor (sTfR) assay on the Roche
Hitachi analyzer and its comparison to two ELISA assays, Clin. Chim. Acta 382 (2007)
112-116.
[201] S. Kuvibidila, T. Gauthier, R.P. Warrier, W. Rayford, Increased levels of serum
transferrin receptor and serum transferrin receptor/log ferritin ratios in men with
prostate cancer and the implications for body-iron stores, J. Lab. Clin. Med. 144 (2004)
176-182.
[202] Q. Xu, M. Zhu, T. Yang, F. Xu, Y. Liu, Y. Chen, Quantitative assessment of
human serum transferrin receptor in breast cancer patients pre- and post-chemotherapy
using peptide immunoaffinity enrichment coupled with targeted proteomics, Clin.
Chim. Acta 448 (2015) 118-123.
[203] I. van den Broek, R.W. Sparidans, A.W. van Winden, Gast MC, E.J. van Dulken,
J.H. Schellens, J.H. Beijnen, The absolute quantification of eight inter-α-trypsin
inhibitor heavy chain 4 (ITIH4)-derived peptides in serum from breast cancer patients,
Proteomics Clin. Appl. 4(12) (2010) 931-939.

I. Introduction
______________________________________________________________________
64

II. OBJECTIVES


II. Objectives
______________________________________________________________________
65
The main objective of this Doctoral Thesis has been the discovery of serum
biomarkers (minor proteins) associated to the different breast cancer subtypes throught
the use of a novel nanoproteomic approach.
The specific objectives of the Doctoral Thesis are therefore as follows:
1. To find the optimal conditions of pH, temperature, protein/nanoparticle ratio
and incubation time for the formation of the protein corona around AuNPs,
AgNPs, FeNPs and PtNPs after their interaction with human serum.
2. To qualitatively analyze and compare the functionality of the human serum
proteins adsorbed on the surface of three different nanomaterials stabilized
with citrate: 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nm
silver nanoparticles (AgNPs) and, 2.40 ± 0.30 nm platinum nanoparticles
(PtNPs).
3. To identify novel serum biomarkers of triple negative breast cancer (TNBC)
throught the qualitative and quantitative analysis of the protein corora
formed arround 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70
nm silver nanoparticles (AgNPs) and 9.30 ± 0.67 nm magnetic nanoparticles
(MNPs).
4. To identify novel serum biomarkers of the different breast cancer subtypes
(luminal A, luminal B HER2 negative, luminal B HER2 positive, HER2
positive and triple negative breast cancer) throught the qualitative and
quantitative analysis of the protein corora formed arround 12.96 ± 0.72 nm
gold nanoparticles (AuNPs).

II. Objectives
______________________________________________________________________
66

III. RESULTS AND DISCUSSION


CHAPTER 1
Proteomic analysis of the bio-corona formed on the surface of
(Au, Ag, Pt)-nanoparticles in human serum
María del Pilar Chantada-Vázquez, Antonio Castro López, Susana B. Bravo, Sergio
Vázquez-Estévez, Benigno Acea-Nebril, Cristina Núñez
Colloids Surf B Biointerfaces 177 (2019) 141-148
DOI: 10.1016/j.colsurfb.2019.01.056


III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
67
Proteomic analysis of the bio-corona formed on the surface of
(Au, Ag, Pt)-nanoparticles in human serum
María del Pilar Chantada-Vázquez,a Antonio Castro López,b Susana B. Bravo,c
Sergio Vázquez-Estévez,d Benigno Acea-Nebril,e Cristina Núñeza
a Research Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de
Saúde (SERGAS), 27002 Lugo, Spain
b Breast Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde
(SERGAS), 27002 Lugo, Spain
d Proteomic Unit, Instituto de Investigaciones Sanitarias-IDIS, Complejo Hospitalario
Universitario de Santiago de Compostela (CHUS), 15706 Santiago de Compostela,
Spain
d Oncology Division, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de
Saúde (SERGAS), 27002 Lugo, Spain
e Department of Surgery, Breast Unit, Complexo Hospitalario Universitario A Coruña
(CHUAC), SERGAS, A Coruña, Spain
Abstract
Adsorption of biomolecules onto nanoparticles surface in biological samples led
to the formation of a bio-corona, it could modify the “identity” of nanoparticles,
contributing to the determination of their toxicity and biocompatibility.
Gel electrophoresis in combination with liquid chromatography-tandem mass
spectrometry (LC-MS/MS) was employed to qualitatively analyze and identify the
human serum proteins adsorbed on the surface of three different nanomaterials
stabilized with citrate: 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nm
silver nanoparticles (AgNPs) and, 2.40 ± 0.30 nm platinum nanoparticles (PtNPs). An
exhaustive analysis and classification of all identified proteins related with their
function were also developed.
Keywords: Gold nanoparticles (AuNPs), silver nanoparticles (AgNPs), platinum
nanoparticles (PtNPs), human serum, protein corona, prognostic biomarker.

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
68
1. Introduction
In recent years, noble metal NPs have gained great interest because their diverse
material properties hold promising potential in advancing current diagnostic and
therapeutic technologies. Such properties are fundamentally dependent on their size,
shape, and composition [1].
Particularly, gold nanoparticles (AuNPs) have a wide range of applications in
the biological and medical fields, including thermotherapy, biosensors, and molecular
imaging [2]. Silver nanoparticles (AgNPs) are known to be potent antimicrobial agents
with a broad antimicrobial spectrum and high efficacy against bacteria [3], and they
have been widely used in consumer products [4]. Platinum nanoparticles (PtNPs) have
been already proposed as efficient and selective radical scavengers for therapies of
oxidative stress diseases [5, 6]. However, their clinical potential has been slowed down
by some toxicological concerns. Nevertheless, the use of PtNPs as additives in
consumer products and cosmetics has been already approved [7]. This suggests that
PtNPs will likely register an increase shortly in a wide range of applications, including
healthcare devices, diagnostics, and cosmetics [8]. Hence, the possibility of living
organisms being exposed to Ag/Au/Pt-NPs either directly or indirectly is greatly
increasing, which has resulted in safety concerns [9].
NPs can enter the human body in different ways, among which the main
exposure routes include inhalation, oral administration, intravenous injection, and
dermal exposure [10]. Once NPs enter the body, they contact various biological
molecules such as proteins, lipid, polysaccharides, and nucleic acids [11]. Particularly,
when NPs are exposed to biological fluids (as serum/plasma), proteins and other
biomolecules are easily adsorbed onto the surface to form a protein ‘corona’ around
NPs, which reduces the surface free energy of NPs [12].
Protein corona is divided into “soft corona” and “hard corona”. While soft
corona is formed by lower affinity proteins reversibly bound to NPs (which can be
further exchangeable), hard corona contains higher affinity proteins on the NP surface
that may irreversibly bind to NPs. Protein corona is a dynamic layer in which amount of
protein and the arrangement are changeable according to the conditions of biological
and physicochemical interaction [13].
During the processes, chemical or physical adsorption takes part in the formation
of protein corona. Coordination, hydrogen bonding, van der Waals forces, electrostatic

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
69
and hydrophobic interactions, steric hindrance, etc. play important roles in driving the
binding of proteins to NPs [14, 15].
Furthermore, protein corona patterns mainly depend on the physicochemical
properties of NPs (nanomaterial, size, charge, surface functional groups, shape) and
exposing environments including immersed media components, temperature, pH,
dynamic shear stress, and interaction (or exposing) time [16-19]. Proteins with large
quantities are first bound to NP surface, and then gradually replaced by higher affinity
proteins (Vroman effect) [20].
When the protein is bound to NPs to form protein corona, proteins may
reorganize their structures to adapt to surrounding environments and NPs surface. The
secondary and/or tertiary protein structure is modified, and this event is known as
“conformational changes” [21]. Furthermore, the formation of a corona may eliminate
the physiological functions of proteins, which leads to the loss of original targeting
capabilities [22], induces various cellular responses including inflammatory responses,
increased lysosomal permeability, activated caspase-related pathways, or even apoptosis
[23, 24].
Understanding how the different properties of nanoparticles affect the
composition of protein coronas is therefore important. The serum and plasma protein
corona compositions of many AuNPs with different coatings and different sizes have
been identified [25-27]. However, the high-throughput protein analyses of the AgNPs’
coronas have been limited [26, 28-30]. To better understand the relationship between
corona composition and the nanoparticles’ properties, a recent study was developed.
This analysis showed specific binding patterns for the blood plasma-derived corona
composition of AgNPs and AuNPs with three surface coatings (polyethyleneimine
(BPEI), citrate (CIT), and polyvinylpyrrolidone (PVP)) [31]. To the best of our
knowledge, the analysis of the composition of the serum protein corona formed around
PtNPs was not carried out until the moment.
On the other hand, a prognostic factor is defined as any parameter, evaluated at
diagnosis (or surgery), which is associated with treatment outcome (local control,
disease-free interval, survival) and may predict patient outcome independent of
treatment. Furthermore, prognostic factors (clinical or biological) may be defined in any
disease stage or setting.
Blood-based biomarkers can be useful as pre-treatment prognostic markers, as
they can reflect variations in the tumor microenvironment and host immune response

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
70
and can complement biopsy-based biomarkers that evaluate tumor cells directly [32]. As
whole blood provides a dynamic representation of physiological and pathological state,
serum or plasma represents the most broadly studied biological matrix for cancer
biomarkers. Therefore, analysis of the plasma or serum proteome could be important to
achieve accurate diagnosis or prognosis.
A great number of proteomics-based studies of plasma and serum have reported
differential peptide/protein ion peaks, either as identified proteins or on the basis of
their mass/charge (m/z) values, for different cancer diagnosis or prognosis as, for
example, breast cancer [33, 34], ovarian cancer [35], head and neck cancer [36], bladder
cancer [37], lung cancer [38]; or other diseases as amyotrophic lateral sclerosis [39],
ST-segment elevation myocardial infarction [40], severe sepsis [41].
Taking into consideration all the aforementioned arguments, with our
experimental work of the serum protein corona formed around AuNPs, AgNPs and
PtNPs, we are providing basic information for toxicological and immunological risk
assessment, as well as information about the properties of different nanomaterials for
the development of novel sensors with potential medical applications.
2. Experimental
2.1. Chemicals and reagents
All reagents and solvents used were HPLC-grade or higher. Sodium citrate
tribasic dihydrate, tannic acid, chloroplatinic acid (H2PtCl6), sodium borohydride
(NaBH4), trypsin, trifluoroacetic acid, DL-Dithiothreitol(DTT), Iodoacetamide (IAA),
acrylamide/bis-acrylamide 30% solution (37.5:1), Glycerol 86-88%, Tris-base,
Coomassie Brilliant Blue R250 (CBB), sodium carbonate, and the Sigma Marker wide
range 6.5-200 KDa were purchased from Sigma-Aldrich (St. Louis, MO, USA). Sodium
dodecylsulfate (SDS) and formaldehyde were purchased from Panreac (Barcelona,
Spain). β-mercaptoethanol was purchased from Merck (Hohen-brunn,Germany) and
bromophenol-blue was purchased from Riedel-de Haen (Seelze,Germany). Hydrogen
tetrachloroaurate (III) hydrate (99.9%-Au) (49%Au) at 10%w/v was purchased from
Strem Chemicals (Newburyport, MA, USA). Ammoniumbicarbonate (AMBIC) and
formic acid were purchased from Fluka (Steinheim, Germany).

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
71
2.2. Instrumentation
Microscopic characterizations of AuNPs, AgNPs and PtNPs were performed by
transmission electron microscopy (TEM) using a Jeol JEM 1011 microscope. Samples
for TEM were prepared by pipetting a drop of the colloidal dispersion onto an ultrathin
carbon-coated copper grid and allowing the solvent to evaporate. Power Pac Basic
power supply from Bio-Rad (CA, USA) was used for sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE) protein separation. Protein
quantification was accomplished by measuring the absorbance at 280 nm with the use of
a Qubit™ 4 Quantitation Starter Kit from Thermo Fisher Scientific. Gel image
acquisition was carried out with a UVP PhotoDoc-ItTM Imaging System from Analytik
Jena.
2.3. Synthesis of inorganic nanoparticles
Gold nanoparticles (AuNPs), silver nanoparticles (AgNPs) and platinum
nanoparticles (PtNPs) were synthesized by the citrate reduction method in aqueous
solution by the method reported by R. López-Cortés et al. [42], V. Puntes et al. [43] and
W. Chen et al. [44], respectively (see Supplemental Material).
2.4. Serum samples
Venous blood sample was obtained from five disease-free individuals with the
use of VACUETTE® Serum Clot Activator Tubes (10 mL). The collected blood samples
were allowed to clot for 15 min, and then centrifuged for 5 min at 4˚C and 1800 × g.
Sera were transferred into clean plastic tubes (1 mL) and immediately frozen at -80˚C at
Research Unit, Hospital Universitario Lucus Augusti (HULA).
2.5. Depletion of multiple high abundant proteins
Serum aliquots (×3) were filtered with Miller-GP® Filter Unit (Millipore) with a
size of 0.22 μm. Each aliquot of human serum (30 µL) was depleted with dithiothreitol
(DTT) according to the protocol described by Warder el al. [45, 46]. Fresh DTT 500
mM (3.3 µL) was mixed with 30 µL of human serum and vortex briefly. Samples were
then incubated until a viscous white precipitate persisted (60 min), followed by
centrifugation at 18840 × g for 20 min. Supernatants were transferred to a clean tube
prior to protein alkylation and nanoparticles (NPs) fractionation.

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
72
2.6. NPs protein alkylation and fractionation
After protein depletion, the reduced SH-groups were alkylated with iodoacetic
acid (IAA) for 45 min at room temperature and protected from light. After protein
reduction and alkylation, 75 µL of AuNPs (10.02 ± 0.91 nm), 75 µL of AgNPs (9.73 ±
1.70 nm) and 75 µL of PtNPs (2.40 ± 0.30 nm) were added to each different serum
aliquots (×3) belonging to the five disease-free individuals (9 aliquots per individual, 3
with each nanoparticle type), followed by the addition of 40 µL of citrate/citric acid
buffer to a final pH of 5.8, as described by López-Cortés et al [42]. Then, all NPs-serum
solutions were incubated at 37 °C with shaking in a thermostatic bath during 30 min.
Pellets were harvested by centrifugation at 18840 × g (AuNPs and AgNPs) and
24610 × g (PtNPs) during 30 min. In all cases, pellets containing proteins bound to
nanoparticles were washed three times with 25 µL citrate/citric acid buffer and
harvested again by centrifugation at 18840 × g (AuNPs and AgNPs) and 24610 × g
(PtNPs) during 30 min to remove unbound proteins.
All pellets were reconstituted in 10 µL of buffer with 0.2 M Tris-HCl, 2 % w/v
SDS and 20 % v/v glycerol. This 10 µL was mixed with 4 µL of SDS-PAGE loading
buffer (10 % w/v SDS, Tris-Base 40 mM, pH 6.8, 50 % v/v glycerol, 0.1 % v/v
bromophenol blue, 10 % v/v β-mercaptoethanol) in a final volume of 20 µL. Then, all
samples were denatured by heating at 100°C for 5 min and loaded into a 10%
acrylamide/bis-acrylamide, stacking gel /12.5% acrylamide/bis-acrylamide running gel,
of 1 mm thickness, and separated at 180 V (constant voltage) for 120 min. After
electrophoresis, the gel was fixed for 30 minutes with 40% (v/v) ethanol and 10% (v/v)
acetic acid and then stained overnight with Colloidal Coomassie Blue [47]. Gels were
rinsed with distilled water and a 0.5 M sodium chloride solution until a clear
background was observed. Gel imaging was carried out with a with a UVP PhotoDoc-
ItTM Imaging System.
2.7. In-gel protein digestion
Protein bands were excised manually and transferred to 2.5-mL Lo-Bind tubes,
and then washed twice with water and with 50% (v/v) acetonitrile/ 25 mM ammonium
bicarbonate (ambic) until the blue color disappeared.

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
73
Prior to trypsin digestion, gel spots were washed with 25 mM ambic and
dehydrated with acetonitrile. Then, 30 μL of trypsin (20 ng·μL-1 in 12.5 mM ambic/2%
(v/v) acetonitrile) was added to the gel spots and incubated for 60 min at 0ºC.
After this time, gel spots were inspected, trypsin solution not absorbed into the
gel was removed, and the gels were covered with 100 μL of 12.5 mM ambic. Samples
were incubated for 12 h at 37°C. Then 50 μL of 5% (v/v) formic acid was added and the
supernatant was transferred to a new Lo-Bind tube and the peptides were further
extracted from the gel twice with 50% (v/v) acetonitrile/0.1% (v/v) trifluoroacetic acid
(TFA) (×3) and acetonitrile (ACN) (x1). Samples were dried-down and stored at -20 °C
[48].
2.8. Protein identification by mass spectrometry (LC-MS/MS) and data analysis
Digested peptides of each sample were separated using Reverse Phase
Chromatography. Gradient was developed using a micro liquid chromatography system
(Eksigent Technologies nanoLC 400, SCIEX) coupled to high-speed Triple TOF 6600
mass spectrometer (SCIEX) with a micro flow source. The analytical column used was
a silica-based reversed phase column YMC-TRIART C18 150 × 0.30 mm, 3 mm
particle size and 120 Å pore size (YMC Technologies, Teknokroma). The trap column
was a YMC-TRIART C18 (YMC Technologies, Teknokroma with a 3 mm particle size
and 120 Å pore size, switched on-line with the analytical column. The loading pump
delivered a solution of 0.1% formic acid in water at 10 µL/min. The micro-pump
provided a flow-rate of 5 µL/min and was operated under gradient elution conditions,
using 0.1% formic acid in water as mobile phase A, and 0.1% formic acid in acetonitrile
as mobile phase B. Peptides were separated using a 25 minutes gradient ranging from
2% to 90% mobile phase B (mobile phase A: 2% acetonitrile, 0.1% formic acid; mobile
phase B: 100% acetonitrile, 0.1% formic acid). Injection volume was 4 µL.
Data acquisition was carried out in a TripleTOF 6600 System (SCIEX, Foster
City, CA) using a Data dependent workflow. Source and interface conditions were as
follows: ionspray voltage floating (ISVF) 5500 V, curtain gas (CUR) 25, collision
energy (CE) 10 and ion source gas 1 (GS1) 25. Instrument was operated with Analyst
TF 1.7.1 software (SCIEX, USA). Switching criteria was set to ions greater than mass
to charge ratio (m/z) 350 and smaller than m/z 1400 with charge state of 2–5, mass
tolerance 250ppm and an abundance threshold of more than 200 counts (cps). Former

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
74
target ions were excluded for 15s. Instrument was automatically calibrated every 4
hours using as external calibrant tryptic peptides from PepcalMix (Sciex).
2.9. Data Analysis
After MS/MS analysis, data files were processed using ProteinPilotTM 5.0.1
software from Sciex which uses the algorithm ParagonTM for database search and
ProgroupTM for data grouping. Data were searched using a Human specific Uniprot
database. False discovery rate was performed using a non-lineal fitting method
displaying only those results that reported a 1% Global false discovery rate or better
[49, 50].
3. Results and discussion
3.1. Serum fraction preparation and protein corona purification
Following the synthetic methods described by R. López-Cortés [42], V. Puntes
et al. [43] and W. Chen et al. [44], AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm)
and PtNPs (2.40 ± 0.30 nm) were successfully obtained and characterized (see Fig
1_SM to Fig 3_SM).
High abundance serum proteins were first depleted using DTT and separated
from low abundance serum proteins [42, 43]. Low abundance proteins were further
processed as described in the experimental section. Afterward, AuNPs (10.02 ± 0.91
nm), AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm) were mixed with serum
aliquots (×3) from five different individuals (9 protein samples per individual: 3 treated
with AuNPs, 3 with AgNPs and 3 with PtNPs).
It is well known that some variables influence the protein-capture efficiency,
namely (i) the NPs/protein ratio, (ii) sample pH and (iii) incubation time [42].
Particularly, the pH value is an important parameter because influences the charge state
of proteins, and a maximum binding capacity between NPs and proteins takes place at
the pH near to the protein pI [51, 52]. The optimum conditions found in preliminary
experiments with AuNPs [42], were used in this case to reduce the complexity and large
dynamic range of the human serum. To this aim, the pH was adjusted to 5.8 with
citrate/citric acid buffer and three incubation times (30, 60 and 90 minutes) were tested.
After the incubation, the pellets and the supernatants were separated via centrifugation
and the respective protein content was assessed through the use of 1D gel

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
75
electrophoresis. Similar results to that obtained after 30 minutes were found with
incubation times of 60 and 90 minutes (Fig 1_SM to Fig 3_SM), suggesting that 30
minutes was enough to achieve a good separation efficiency and it was selected as the
optimum incubation time.
To investigate the influence of the amount of each type of NPs during the
separation process, three volumes of each type of nanoparticles were explored: 75 μL,
100 μL and 125 μL, to get the following protein/NPs ratios: 10.7, 8.6 and 6.5. Fig 2 and
Fig 4_SM to Fig 6_SM show the result of this set of experiments, which suggest good
separation efficiency, even for the lowest amounts of each NPs tested (75 μL).
As it is shown in Figure 2, for an incubation time of 30 minutes, differences in
the protein corona formed around the three different NPs were visible after Coomassie
staining. When the gel profiles of each fraction (AuNPs-protein corona, AgNPs-protein
corona, PtNPs-protein corona) were compared, it was easily noted that there is a
different in the intensity of the bands. However, no conclusion can be drawn unless the
proteins were identified. However, the supernatants were very similar in all cases,
independently of the type of nanoparticle employed and the quantity of each one (Fig
7_SM).
After that, gel bands corresponding to the fractions AuNPs-protein corona,
AgNPs-protein corona, PtNPs-protein corona formed after the addition of 75 μL, were
excised and submitted to the sample treatment described in the experimental section.
The resulting pools of peptides were then analyzed by mass spectrometry (LC-MS/MS)
for protein identification.
A similar number of proteins were identified from the protein corona formed
around AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm)
after their incubation with serum aliquots (×3) from five different individuals (9 protein
samples per individual: 3 treated with AuNPs, 3 with AgNPs and 3 with PtNPs) (see
Table 1_SM). Importantly, 215, 215 and 198 were commonly found in all cases in the
surface of AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30
nm) (see Table 2_SM). From them, 170 proteins were commonly detected in the protein
corona of all three different types of NPs (see Fig 1). One of these proteins was serum
albumin, the most abundant protein in the blood. However, 52 different proteins were
found on the three different NPs surface (see Fig 1): 21 different proteins on the 10.02 ±
0.91 nm AuNPs (see Table 1), 17 on the 9.73 ± 1.70 nm AgNPs (see Table 2) and 14
individual proteins on the 2.40 ± 0.30 nm PtNPs (see Table 3).
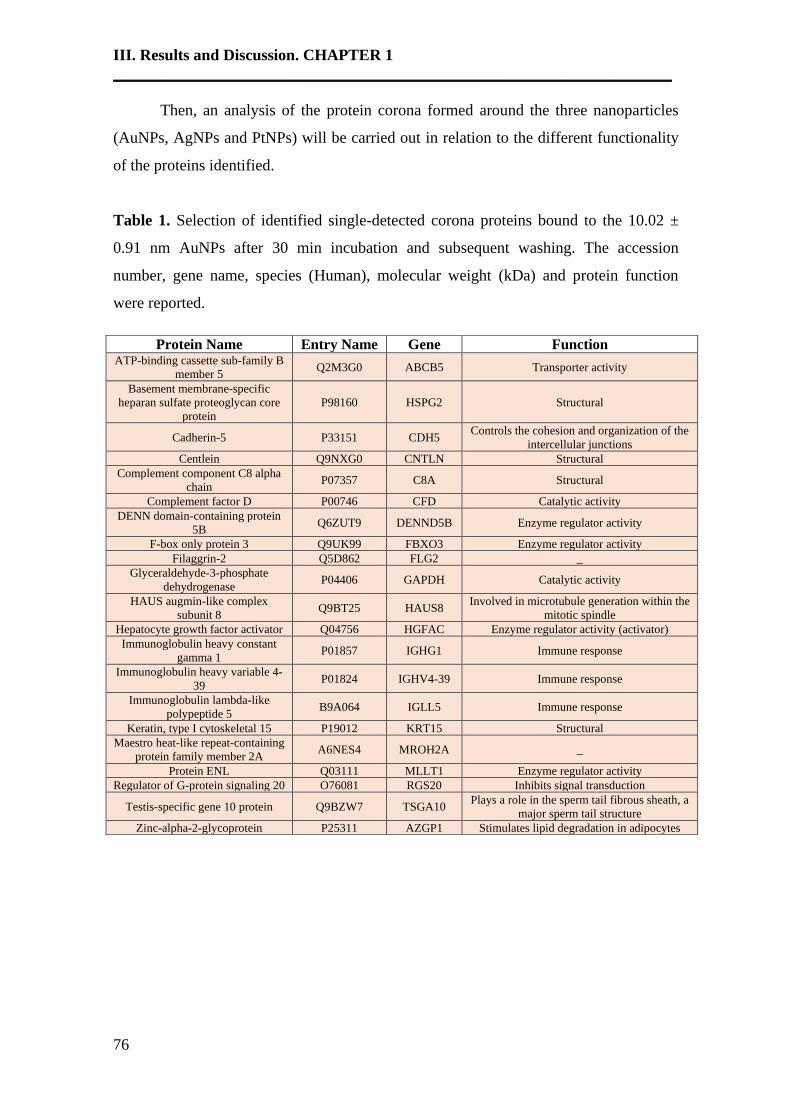
III. Results and Discussion. CHAPTER 1
______________________________________________________________________
76
Then, an analysis of the protein corona formed around the three nanoparticles
(AuNPs, AgNPs and PtNPs) will be carried out in relation to the different functionality
of the proteins identified.
Table 1. Selection of identified single-detected corona proteins bound to the 10.02 ±
0.91 nm AuNPs after 30 min incubation and subsequent washing. The accession
number, gene name, species (Human), molecular weight (kDa) and protein function
were reported.
Protein Name Entry Name Gene Function ATP-binding cassette sub-family B
member 5 Q2M3G0 ABCB5 Transporter activity
Basement membrane-specific
heparan sulfate proteoglycan core
protein
P98160 HSPG2 Structural
Cadherin-5 P33151 CDH5 Controls the cohesion and organization of the
intercellular junctions
Centlein Q9NXG0 CNTLN Structural
Complement component C8 alpha
chain P07357 C8A Structural
Complement factor D P00746 CFD Catalytic activity
DENN domain-containing protein
5B Q6ZUT9 DENND5B Enzyme regulator activity
F-box only protein 3 Q9UK99 FBXO3 Enzyme regulator activity
Filaggrin-2 Q5D862 FLG2 _
Glyceraldehyde-3-phosphate
dehydrogenase P04406 GAPDH Catalytic activity
HAUS augmin-like complex
subunit 8 Q9BT25 HAUS8
Involved in microtubule generation within the
mitotic spindle
Hepatocyte growth factor activator Q04756 HGFAC Enzyme regulator activity (activator)
Immunoglobulin heavy constant
gamma 1 P01857 IGHG1 Immune response
Immunoglobulin heavy variable 4-
39 P01824 IGHV4-39 Immune response
Immunoglobulin lambda-like
polypeptide 5 B9A064 IGLL5 Immune response
Keratin, type I cytoskeletal 15 P19012 KRT15 Structural
Maestro heat-like repeat-containing
protein family member 2A A6NES4 MROH2A _
Protein ENL Q03111 MLLT1 Enzyme regulator activity
Regulator of G-protein signaling 20 O76081 RGS20 Inhibits signal transduction
Testis-specific gene 10 protein Q9BZW7 TSGA10 Plays a role in the sperm tail fibrous sheath, a
major sperm tail structure
Zinc-alpha-2-glycoprotein P25311 AZGP1 Stimulates lipid degradation in adipocytes
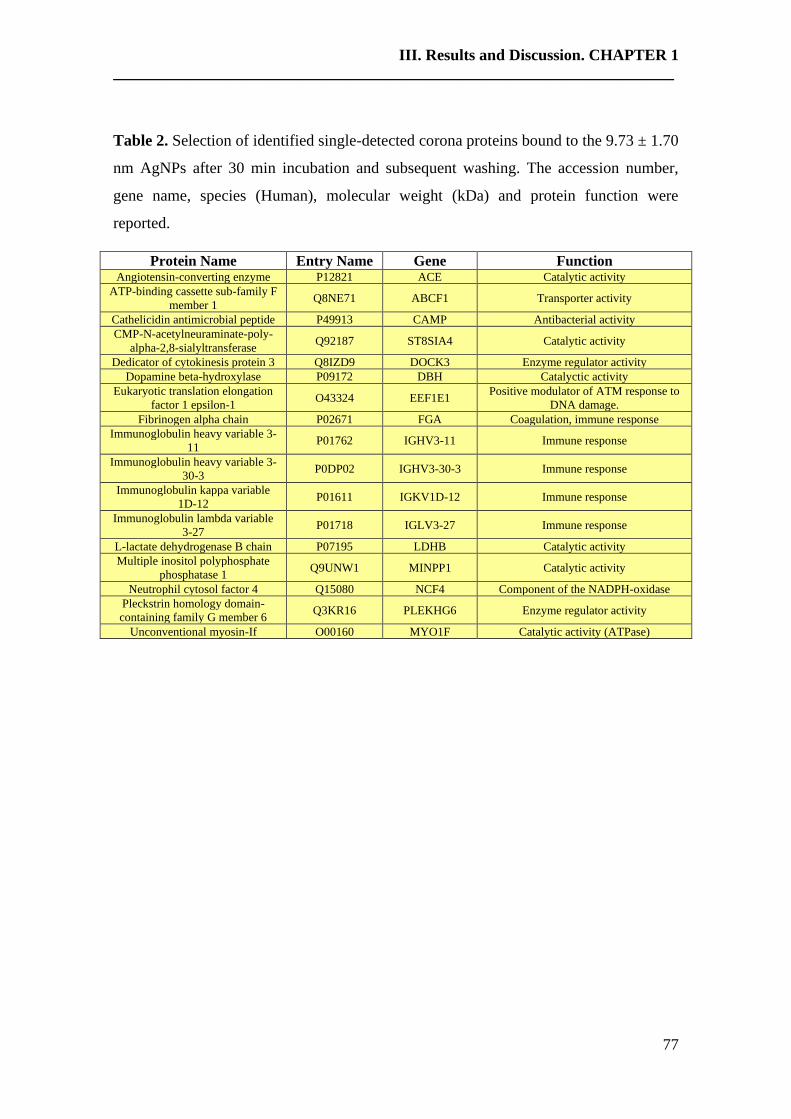
III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
77
Table 2. Selection of identified single-detected corona proteins bound to the 9.73 ± 1.70
nm AgNPs after 30 min incubation and subsequent washing. The accession number,
gene name, species (Human), molecular weight (kDa) and protein function were
reported.
Protein Name Entry Name Gene Function Angiotensin-converting enzyme P12821 ACE Catalytic activity
ATP-binding cassette sub-family F
member 1 Q8NE71 ABCF1 Transporter activity
Cathelicidin antimicrobial peptide P49913 CAMP Antibacterial activity
CMP-N-acetylneuraminate-poly-
alpha-2,8-sialyltransferase Q92187 ST8SIA4 Catalytic activity
Dedicator of cytokinesis protein 3 Q8IZD9 DOCK3 Enzyme regulator activity
Dopamine beta-hydroxylase P09172 DBH Catalyctic activity
Eukaryotic translation elongation
factor 1 epsilon-1 O43324 EEF1E1
Positive modulator of ATM response to
DNA damage.
Fibrinogen alpha chain P02671 FGA Coagulation, immune response
Immunoglobulin heavy variable 3-
11 P01762 IGHV3-11 Immune response
Immunoglobulin heavy variable 3-
30-3 P0DP02 IGHV3-30-3 Immune response
Immunoglobulin kappa variable
1D-12 P01611 IGKV1D-12 Immune response
Immunoglobulin lambda variable
3-27 P01718 IGLV3-27 Immune response
L-lactate dehydrogenase B chain P07195 LDHB Catalytic activity
Multiple inositol polyphosphate
phosphatase 1 Q9UNW1 MINPP1 Catalytic activity
Neutrophil cytosol factor 4 Q15080 NCF4 Component of the NADPH-oxidase
Pleckstrin homology domain-
containing family G member 6 Q3KR16 PLEKHG6 Enzyme regulator activity
Unconventional myosin-If O00160 MYO1F Catalytic activity (ATPase)
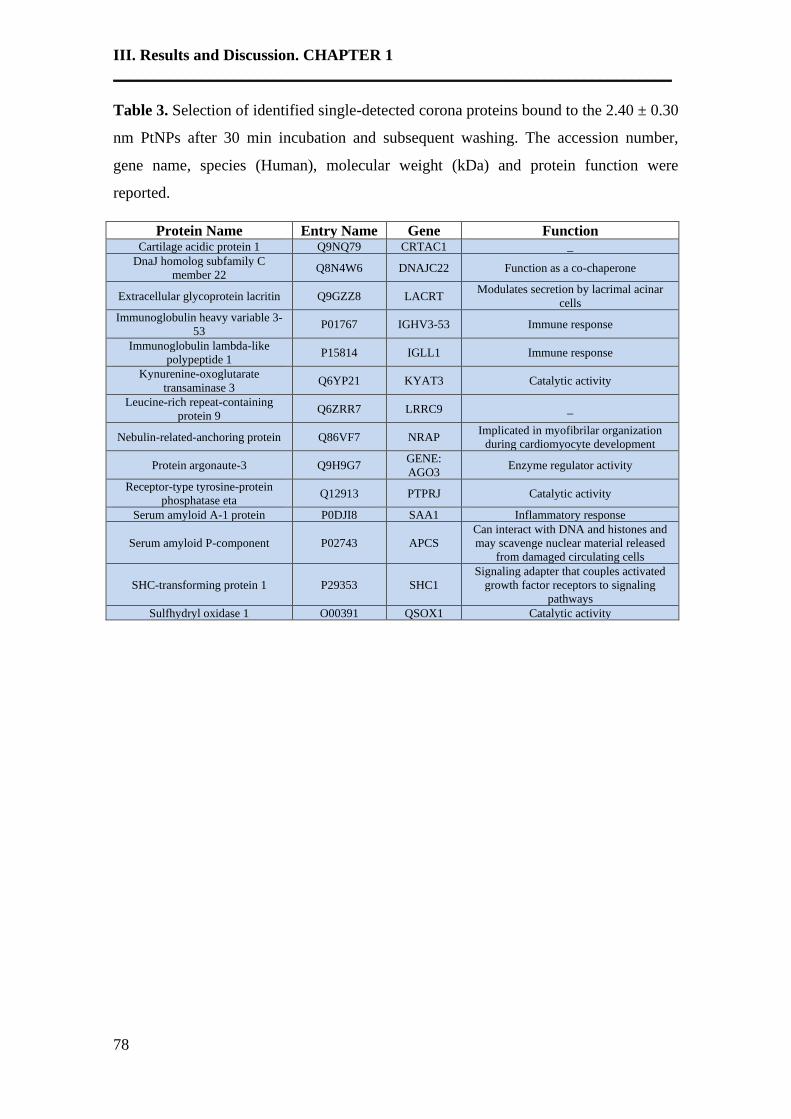
III. Results and Discussion. CHAPTER 1
______________________________________________________________________
78
Table 3. Selection of identified single-detected corona proteins bound to the 2.40 ± 0.30
nm PtNPs after 30 min incubation and subsequent washing. The accession number,
gene name, species (Human), molecular weight (kDa) and protein function were
reported.
Protein Name Entry Name Gene Function Cartilage acidic protein 1 Q9NQ79 CRTAC1 _
DnaJ homolog subfamily C
member 22 Q8N4W6 DNAJC22 Function as a co-chaperone
Extracellular glycoprotein lacritin Q9GZZ8 LACRT Modulates secretion by lacrimal acinar
cells
Immunoglobulin heavy variable 3-
53 P01767 IGHV3-53 Immune response
Immunoglobulin lambda-like
polypeptide 1 P15814 IGLL1 Immune response
Kynurenine-oxoglutarate
transaminase 3 Q6YP21 KYAT3 Catalytic activity
Leucine-rich repeat-containing
protein 9 Q6ZRR7 LRRC9 _
Nebulin-related-anchoring protein Q86VF7 NRAP Implicated in myofibrilar organization
during cardiomyocyte development
Protein argonaute-3 Q9H9G7 GENE:
AGO3 Enzyme regulator activity
Receptor-type tyrosine-protein
phosphatase eta Q12913 PTPRJ Catalytic activity
Serum amyloid A-1 protein P0DJI8 SAA1 Inflammatory response
Serum amyloid P-component P02743 APCS
Can interact with DNA and histones and
may scavenge nuclear material released
from damaged circulating cells
SHC-transforming protein 1 P29353 SHC1
Signaling adapter that couples activated
growth factor receptors to signaling
pathways
Sulfhydryl oxidase 1 O00391 QSOX1 Catalytic activity
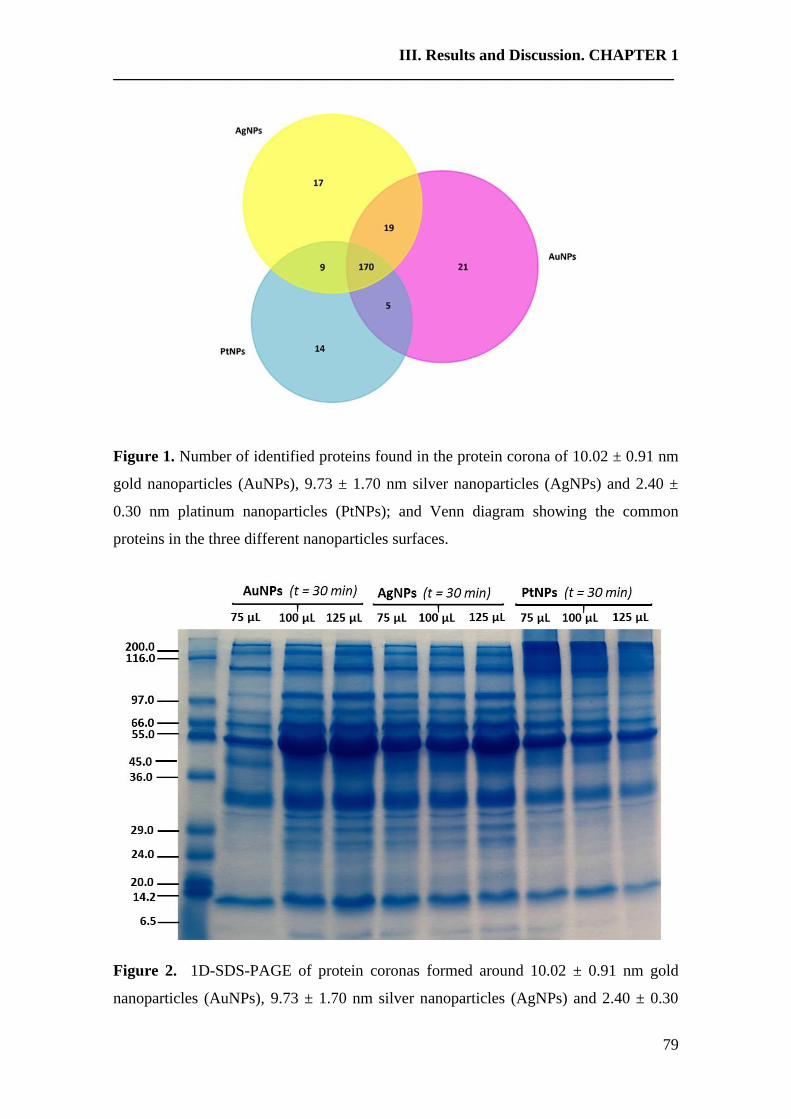
III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
79
Figure 1. Number of identified proteins found in the protein corona of 10.02 ± 0.91 nm
gold nanoparticles (AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs) and 2.40 ±
0.30 nm platinum nanoparticles (PtNPs); and Venn diagram showing the common
proteins in the three different nanoparticles surfaces.
Figure 2. 1D-SDS-PAGE of protein coronas formed around 10.02 ± 0.91 nm gold
nanoparticles (AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs) and 2.40 ± 0.30

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
80
nm platinum nanoparticles (PtNPs) in human serum (incubation time: 30 minutes;
volumes of each nanoparticles solution: 75 μL, 100 μL and 125 μL, to get the following
protein/NPs ratios: 10.7, 8.6 and 6.5, respectively). On the left, it marks the lane with
Mw protein standards with molecular weights in kDa.
3.2. Proteins implicated in the immune response
A total of 66 proteins implicated in the immune response (55 of them are
immunoglobulins) were identified from the protein corona formed around the AuNPs,
AgNPs, and PtNPs after their incubation with serum of all individuals. From them, 48
common proteins were found on the surface of the three different nanoparticles (see
Table 1_SM). Particularly, two of these common immune-related proteins were
complement C3 (187.1 kDa) and complement factor B (85.5 kDa). The according
complement system is involved in an effective strategy called opsonization for labeling
pathogens for removal by phagocytic cells from the circulation. To minimize
opsonization of AuNP, AgNPs and PtNPs in pharmaceutical applications (since the
drug-loaded NP is cleared by the organism and cannot release its drug at the desired
site) binding of the NP with complement factors should be avoided [49].
An example of a protein implicated in the immune response that was only found
on the surface of AgNPs was fibrinogen alpha chain (94.9 kDa), involved in blood
coagulation but also opsonizes surfaces of foreign bodies for immune cell recognition.
Furthermore, AuNPs nanoparticles exhibited selective adsorption towards
immunoglobulin heavy constant gamma 1 (36.1 kDa) (IGHG1), similar to that reported
previously in by zeolites in plasma [53].
3.3. Proteins with a transport function
A total of 29 proteins with a transporter function (7 of them apolipoproteins)
were identified from the protein corona formed around the AuNPs, AgNPs, and PtNPs.
From them, 25 common proteins were found on the surface of the three different
nanoparticles (see Table 2_SM). Among them, the most interesting protein regarding
toxicity and drug delivery potential is the ApoE. This transport protein is known to
mediate transcytosis across biological barriers, e.g., the blood-brain barrier [54, 55].
AuNPs and AgNPs exhibited selective adsorption towards two proteins that
belong to the ATP-binding cassette (ABC) transporter superfamily of integral
membrane proteins: ATP-binding cassette sub-family B member 5 (ABCB5) (138.6

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
81
kDa) and ATP-binding cassette sub-family F member 1 (ABCF1) (95.9 kDa),
respectively. Both proteins participate in the ATP-dependent transmembrane transport
of structurally diverse molecules ranging from sugars, small ions, and peptides to more
complex organic molecules [56].
3.4. Proteins with a structural functionality
A total of 32 proteins with a structural functionality were identified from the
protein corona formed around the AuNPs, AgNPs, and PtNPs (18 proteins were
commonly found on the surface of the three different nanoparticles) (see Table 2_SM).
AuNPs nanoparticles exhibited selective adsorption towards 7 proteins. Within
them, basement membrane-specific heparan sulfate proteoglycan core protein (also
known as perlecan) fragments showed to be biomarkers of bone stromal lysis [57], renal
dysfunction [58] and also pancreatic cancer secretome [59]; and the complement
component C8 alpha chain was also identified as a potential serum biomarker in
multiple sclerosis [60].
3.5. Proteins implicated in enzymatic processes
A total of 45 proteins with catalytic activity were identified from the protein
corona formed around the AuNPs, AgNPs, and PtNPs (25 commonly found on the
surface of the three different nanoparticles) (see Table 1_SM).
AuNPs nanoparticles exhibited selective adsorption towards two proteins:
complement factor D (27.1 kDa) and glyceraldehyde-3-phosphate dehydrogenase (36.1
kDa). While complement factor D (CFD; also known as adipsin) regulates activation of
the alternative complement pathway, which is implicated in age-related macular
degeneration (AMD) pathogenesis, glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) (a glycolytic enzyme) can interact with proteins participating in DNA repairs,
such as APE1, PARP1, HMGB1, and HMGB2 [61].
AgNPs nanoparticles exhibited a selective adsorption towards seven proteins,
within them, lactate dehydrogenase (LDH), which was an indirect marker of hypoxia,
was a potentially prognostic factor in several malignancies such as in patients with
hepatocellular carcinoma [62] and advanced pancreatic [63], both receiving sorafenib.
In the case of PtNPs nanoparticles, they exhibited selective adsorption towards
three proteins: kynurenine-oxoglutarate transaminase 3 (51.4 kDa), receptor-type
tyrosine-protein phosphatase eta (149.5 kDa) and sulfhydryl oxidase 1 (82.6 kDa).

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
82
Particularly, it was found that the expression of sulfhydryl oxidase 1 was associated
with a highly invasive phenotype and correlates with a poor prognosis in Luminal B
breast cancer [64].
A total of 39 proteins that regulates (activate or inhibit) the activity of different
enzymes were identified from the protein corona formed around the AuNPs, AgNPs,
and PtNPs. From them, 28 common proteins were found on the surface of the three
different nanoparticles.
Four (DENN domain-containing protein 5B (145.1 kDa), F-box only protein 3
(54.6 kDa), hepatocyte growth factor activator (70.7 kDa) and protein ENL (62.1 kDa)),
two (dedicator of cytokinesis protein 3 (233.1 kDa) and pleckstrin homology domain-
containing family G member 6 (88.9 kDa)) and one (protein argonaute-3 (97.4 kDa))
enzymatic regulatory proteins were found on the protein-corona of AuNPs (table 1),
AgNPs (table 2) and PtNPs (table 3), respectively.
3.6. Proteins implicated in the inflammatory response
A total of 6 proteins implicated in the inflammatory response were identified
from the protein corona formed around the AuNPs, AgNPs, and PtNPs.
Particularly, PtNPs nanoparticles exhibited selective adsorption towards one
protein: serum amyloid A-1 protein (SAA1) (13.5 kDa), a sensitive acute phase reactant
primarily produced by the liver in response to acute inflammation. It was recently
shown that SAA affects proliferation, migration, and invasion of glioblastoma cell lines,
which suggest its participation in the malignant process. Consistently, levels of SAA
have been used as a non-invasive biomarker for the prognosis of many cancers and,
particularly, serum amyloid A1 was found to be upregulated in human glioblastoma
[65].
3.7. Proteins with antibiotic activity
Dermcidin (11.3 kDa), a protein with antimicrobial activity, was identified from
the protein corona formed around the AuNPs, AgNPs, and PtNPs. Dermcidin (DCD), an
antimicrobial peptide with a broad spectrum of activity against bacteria such as
Propionibacterium acnes, is expressed constitutively in sweat in the absence of
stimulation due to injury or inflammation. It was found that reduced DCD concentration

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
83
in sweat in patients with inflammatory acne may permit proliferation of P. acnes in
pilosebaceous units, resulting in progression of inflammatory acne [66].
However, only AgNPs nanoparticles exhibited selective adsorption towards one
protein with antibacterial activity: cathelicidin antimicrobial peptide (19.3 kDa),
responsible for protecting the urinary tract against invasive bacterial infection [67].
3.8. Proteins implicated in signal transduction
There were identified three proteins implicated in the signal transduction from
the protein corona formed around the three nanosystems. While protein leucine-rich
alpha-2-glycoprotein (38.2 kDa) (implicated in the protein-protein interaction, signal
transduction and cell adhesion and development) was presented in the protein corona of
the three nanoparticles, the protein regulator of G-protein signaling 20 (31.5 kDa) (that
inhibits signal transduction) and SHC-transforming protein 1 (62.8 kDa) were only
found in the surface of AuNPs and PtNPs, respectively.
In general, in relation with the total number of proteins identified from the
protein corona surfaces (see Fig 3), the most abundant groups are constituted by the
proteins implicated in the immune response, followed by proteins with and enzymatic
function, structural, transporter, inflammatory, signal transduction and with
antibiotic/antibacterial properties.
Figure 3. Total number of identified proteins found on the surface of the three different
nanoparticles (color: black) and commonly found on the surface of AuNPs, AgNPs and
PtNPs (color: grey) with a different function.

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
84
However, we found exclusive proteins with structural function and implicated in
the signal transduction only in the protein corona of 10.02 ± 0.91 nm AuNPs (see Table
1), with antibiotic/antibacterial properties in 9.73 ± 1.70 nm AgNPs (see Table 2) and
implicated on inflammatory processes only in 2.40 ± 0.30 nm PtNPs (see Table 3).
4. Conclusions
In this study, it was shown that the interaction of AuNPs (10.02 ± 0.91 nm),
AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm) with human serum withstands the
formation of a protein corona enveloping the nanoparticle, in all cases.
The formation of this protein corona depends on the composition of the
nanoparticle (core material) and its size. In that case, it was observed that smaller NPs
(2.40 ± 0.30 nm PtNPs) have lower protein adsorption (198 proteins) than larger NPs
(10.02 ± 0.91 nm AuNPs and 9.73 ± 1.70 nm AgNPs) (215 proteins). A total of 170
proteins with different functionality were detected in the protein corona of all three
different types of NPs. However, 21, 14 and 14 different proteins were found on the
AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm),
respectively.
In general, the function of all identified proteins ranges from proteins implicated
in the immune response, followed by proteins with an enzymatic function, structural,
transporter, inflammatory, signal transduction and with antibiotic/antibacterial
properties, being the majority group the first one with 66 proteins identified implicated
in the immune response. However, if we observed the different proteins on the corona
of the three different NPs, we found exclusive proteins with structural function and
implicated in the signal transduction only in the protein corona of AuNPs (10.02 ± 0.91
nm), with antibiotic/antibacterial properties in AgNPs (9.73 ± 1.70 nm) and implicated
on inflammatory processes only in PtNPs (2.40 ± 0.30 nm).
This has implications for immune safety, biocompatibility, and information for
developing novel nanomaterials with high specificity and selectivity towards proteins
with an important biological function (prognostic and diagnostic protein biomarkers).
However, it is important to mention that the interaction of these nanoparticles with
human serum could drive to the formation of a different protein corona at a pH
dissimilar of 5.8, for example, under physiological conditions.

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
85
5. References
[1] J. Zheng, C. Zhou, M. Yu, J. Liu, Different sized luminescent gold nanoparticles,
Nanoscale 4(14) (2012) 4073-4083.
[2] Z. Miao, Z. Gao, R. Chen, X. Yu, Z. Su, G. Wei, Surface-bioengineered Gold
Nanoparticles for Biomedical Applications, Curr Med Chem. 25(16) (2018) 1920-1944.
[3] N. Duran, M. Duran, M.B. de Jesus, A.B. Seabra, W.J. Favaro, G. Nakazato, Silver
nanoparticles: A new view on mechanistic aspects on antimicrobial activity,
Nanomedicine 12(3) (2016) 789-799.
[4] K.W. Lem, A. Choudhury, A.A. Lakhani, P. Kuyate, J.R. Haw, D.S. Lee, Z. Iqbal,
C.J. Brumlik, Use of nanosilver in consumer products, Recent Pat Nanotechnol. 6
(2012) 60-72.
[5] M. Moglianetti, E. De Luca, D. Pedone, R. Marotta, T. Catelani, B. Sartori, H.
Amenitsch, S.F. Retta, P.P. Pompa, Platinum nanozymes recover cellular ROS
homeostasis in an oxidative stress-mediated disease model, Nanoscale 8 (2016) 3739-
3752.
[6] H. Hosaka, R. Haruki, K. Yamada, C. Böttcher, T. Komatsu, Hemoglobin-albumin
cluster incorporating a Pt nanoparticle: artificial O2 carrier with antioxidant activities,
PLoS One 9 (2014) e110541.
[7] S. Onizawa, K. Aoshiba, M. Kajita, Y. Miyamoto, A. Nagai, Platinum nanoparticle
antioxidants inhibit pulmonary inflammation in mice exposed to cigarette smoke, Pulm.
Pharmacol Ther. 22 (2009) 340-349.
[8] D. Pedone, M. Moglianetti, E. De Luca, G. Bardi, P.P. Pompa, Platinum
nanoparticles in nanobiomedicine, Chem Soc Rev. 46 (2017) 4951-4975.
[9] T. Zhang, L. Wang, Q. Chen, C. Chen, Cytotoxic potential of silver nanoparticles,
Yonsei Med J. 55 (2014) 283-291.
[10] B. Wang, X. He, Z.Y. Zhang, Y.L. Zhao, W.Y. Feng, Metabolism of nanomaterials
in vivo: blood circulation and organ clearance, Accounts Chem. Res. 46 (2012) 761-
769.
[11] M. Mahmoudi, I. Lynch, M.R. Ejtehadi, M.P. Monopoli, F.B. Bombelli, S.
Laurent, Protein-nanoparticle interactions: opportunities and challenges, Chem Rev. 111
(2011) 5610-5637.

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
86
[12] E. Casals, T. Pfaller, A. Duschl, G.J. Oostingh, V.F Puntes, Hardening of the
nanoparticle-protein corona in metal (Au, Ag) and oxide (Fe3O4, CoO, and CeO2)
nanoparticles, Small 7 (2011) 3479-3486.
[13] S. Milani, F. Baldelli Bombelli, A.S. Pitek, K.A. Dawson, J. Radler, Reversible
versus irreversible binding of transferrin to polystyrene nanoparticles: soft and hard
corona, ACS Nano 6 (2012) 2532-2541.
[14] L. Treuel, S. Brandholt, P. Maffre, S. Wiegele, L. Shang, G.U. Nienhaus, Impact of
protein modification on the protein corona on nanoparticles and nanoparticle-cell
interactions, ACS Nano 8 (2014) 503-513.
[15] L.M. Wang, J.Y. Li, J. Pan, X.M. Jiang, Y.L. Ji, Y.F. Li, Y. Qu, Y.L. Zhao, X.C.
Wu, Revealing the binding structure of the protein corona on gold nanorods using
synchrotron radiation-based techniques: understanding the reduced damage in cell
membranes, J Am Chem Soc. 135 (2013) 17359-17368.
[16] M.P. Monopoli, C. Aberg, A. Salvati, K.A. Dawson, Biomolecular coronas provide
the biological identity of nanosized materials, Nat Nanotechnol. 7 (2012) 779-786.
[17] M. Lundqvist, J. Stigler, G. Elia, I. Lynch, T. Cedervall, K.A. Dawson,
Nanoparticle size and surface properties determine the protein corona with possible
implications for biological impacts, PNAS 105(38) (2008)14265-14270.
[18] S. Tenzer, D. Docter, S. Rosfa, A. Wlodarski, J. Kuharev, A. Rekik, S.K. Knauer,
C. Bantz, T. Nawroth, C. Bier, J. Sirirattanapan, W. Mann, L. Treuel, R. Zellner, M.
Maskos, H. Schild, R.H. Stauber, Nanoparticle size is a critical physicochemical
determinant of the human blood plasma corona: a comprehensive quantitative proteomic
analysis, ACS Nano 5(9) (2011) 7155-7167.
[19] J. Wolfram, Y. Yang, J. Shen, A. Moten, C. Chen, H. Shen, M. Ferrari, Y. Zhao,
The nano-plasma interface: Implications of the protein corona, Colloids Surf B
Biointerfaces 124 (2014) 17-24.
[20] L. Vroman, A. Adams, G.C. Fischer, P.C. Munoz, Interaction of high molecular
weight kininogen, factor XII, and fibrinogen in plasma at interfaces, Blood 55(1) (1980)
156-159.
[21] I. Lynch, K.A. Dawson, Protein-nanoparticle interactions, Nano Today. 3(1) (2008)
40-47.
[22] A. Salvati, A.S. Pitek, M.P. Monopoli, K. Prapainop, F.B. Bombelli, D.R. Hristov,
P.M. Kelly, C. Åberg, E. Mahon, K.A. Dawson, Transferrin-functionalized

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
87
nanoparticles lose their targeting capabilities when a biomolecule corona adsorbs on the
surface, Nat Nanotechnol. 8 (2013) 137-143.
[23] C. Paviolo, J.W. Haycock, J. Yong, A. Yu, S.L. McArthur, P.R. Stoddart,
Plasmonic properties of gold nanoparticles can promote neuronal activity, Proc SPIE
8579 (2013) 85790C.
[24] J.Y. Lee, W. Park, D.K. Yi, Immunostimulatory effects of gold nanorod and silica-
coated gold nanorod on RAW 264.7 mouse macrophages, Toxicol Lett. 209 (2012) 51-
57.
[25] M. Schaffler, M. Semmler-Behnke, H. Sarioglu, S. Takenaka, A. Wenk, C. Schleh,
S.M. Hauck, B.D. Johnston, W.G. Kreyling, Serum protein identification and
quantification of the corona of 5, 15 and 80 nm gold nanoparticles, Nanotechnology 24
(2013) 265103.
[26] C.D. Walkey, J.B. Olsen, F. Song, R. Liu, H. Guo, D.W. Olsen, Y. Cohen, A.
Emili, W.C. Chan, Protein corona fingerprinting predicts the cellular interaction of gold
and silver nanoparticles, ACS Nano 8 (2014) 2439-2455.
[27] M.A. Dobrovolskaia, B.W. Neun, S. Man, X. Ye, M. Hansen, A.K. Patri, R.M.
Crist, S.E. McNeil, Protein corona composition does not accurately predict
hematocompatibility of colloidal gold nanoparticles, Nanomedicine 10 (2014) 1453-
1463.
[28] N. Duran, C.P. Silveira, M. Duran, D.S. Martínez, Silver nanoparticle protein
corona and toxicity: a mini-review, J Nanobiotechnology 13 (2015) 55.
[29] J.H. Shannahan, X. Lai, P.C. Ke, R. Podila, J.M. Brown, F.A. Witzmann, Silver
nanoparticle protein corona composition in cell culture media, PloS One 8(9) (2013)
e74001.
[30] H. Huang, W. Lai, M. Cui, L. Liang, Y. Lin, Q. Fang, Y. Liu, L. Xie, An
Evaluation of Blood Compatibility of Silver Nanoparticles, Sci Rep. 6 (2016) 25518.
[31] W. Lai, Q. Wang, L. Li, Z. Hu, J. Chen, Q. Fang, Interaction of gold and silver
nanoparticles with human plasma: Analysis of protein corona reveals specific binding
patterns, Colloids Surf B Biointerfaces 152 (2017) 317-325.
[32] L. Brøndum, J.G. Eriksen, B.S. Sørensen, L.S. Mortensen, K. Toustrup, J.
Overgaard, J. Alsner, Plasma proteins as prognostic biomarkers in radiotherapy treated
head and neck cancer patients, Clin Trans Rad Oncol. 2 (2017) 46-52.

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
88
[33] A. Kazarian, O. Blyuss, G. Metodieva, A. Gentry-Maharaj, A. Ryan, E.M.
Kiseleva, O.M. Prytomanova, I.J. Jacobs, M. Widschwendter, U. Menon, J.F. Timms,
Testing breast cancer serum biomarkers for early detection and prognosis in pre-
diagnosis samples, Br J Cancer 116 (2017) 501-508.
[34] D. Song, L. Yue, J. Zhang, S. Ma, W. Zhao, F. Guo, Y. Fan, H. Yang, Q. Liu, D.
Zhang, Z. Xia, P. Qin, J. Jia, M. Yue, J. Yu, S. Zheng, F. Yang, J. Wang, Diagnostic
and prognostic significance of serum apolipoprotein C-I in triple-negative breast cancer
based on mass spectrometry, Cancer Biol Ther. 17(6) (2016) 635-647.
[35] T. Muinao, H.P. Deka Boruah, M. Pal, Diagnostic and Prognostic Biomarkers in
ovarian cancer and the potential roles of cancer stem cells - An updated review, Exp
Cell Res. 362(1) (2018) 1-10.
[36] H.-S. Lin, F. Siddiq, H.S. Talwar, W. Chen, C. Voichita, S. Draghici, G. Jeyapalan,
M. Chatterjee, A. Fribley, G.H. Yoo, S. Sethi, H. Kim, A. Sukari, A.J. Folbe, M.A.
Tainsky, Serum prognostic biomarkers in head and neck cancer patients, Laryngoscope
124(8) (2014) 1819-1826.
[37] K. Schwamborn, N.T. Gaisa, C. Henkel, Tissue and serum proteomic profiling for
diagnostic and prognostic bladder cancer biomarkers, Expert Rev Proteomics 27(6)
(2010) 897-906.
[38] T. Targowski, K. Jahnz-Rozyk, W. Owczarek, A. Raczka, P. Janda, T. Szkoda, T.
Płusa, Telomerase activity and serum levels of p53 protein as prognostic factors of
survival in patients with advanced non-small cell lung cancer, Respir Med. 104(9)
(2010) 1356-1361.
[39] C. Lunetta, A. Lizio, E. Maestri, V.A. Sansone, G. Mora, R.G. Miller, S.H. Appel,
A. Chiò, Serum C-Reactive Protein as a Prognostic Biomarker in Amyotrophic Lateral
Sclerosis, JAMA Neurol. 74(6) (2017) 660-667.
[40] S. Baumann, A. Huseynov, J. Koepp, C. Jabbour, M. Behnes, T. Becher, M.
Renker, S. Lang, M. Borggrefe, R. Lehmann, I. Akin, Comparison of Serum Uric Acid,
Bilirubin, and C-Reactive Protein as Prognostic Biomarkers of In-Hospital MACE
Between Women and Men With ST-Segment Elevation Myocardial Infarction,
Angiology 37(3) (2016) 272-280.
[41] J. Szederjesi, E. Almasy, A. Lazar, A. Huțanu, I. Badea, A. Georgescu, An
Evaluation of Serum Procalcitonin and C-Reactive Protein Levels as Diagnostic and
Prognostic Biomarkers of Severe Sepsis, J Crit Care Med. 1(4) (2015) 147-153.

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
89
[42] R. Lopez-Cortés, E. Oliveira, C. Núñez, C. Lodeiro, María Páez de la Cadena,
Florentino Fdez-Riverola, H. López-Fernández, Miguel Reboiro-Jato, Daniel Glez-
Peña, José Luis Capelo, Hugo M. Santos, Fast human serum profiling through chemical
depletion coupled to gold-nanoparticle-assisted protein separation, Talanta 100 (2012)
239-245.
[43] N.G. Bastus, F. Merkoci, J. Piella, V. Puntes, Synthesis of Highly Monodisperse
Citrate-Stabilized Silver Nanoparticles of up to 200 nm: Kinetic Control and Catalytic
Properties, Chem. Mater. 26(9) (2014) 2836-2846.
[44] G.-W. Wu, S.-B. He, H. Peng, H.-H. Deng, A.-L. Liu, X.-H. Lin, X.-h. Xia, W.
Chen, Citrate-capped platinum nanoparticle as a smart probe for ultrasensitive mercury
sensing, Anal. Chem. 86(21) (2014) 10955-10960.
[45] S.E. Warder, L.A. Tucker, T.J. Strelitzer, E.M. McKeegan, J.L. Meuth, P.M. Jung,
A. Saraf, B. Singh, J. Lai-Zhang, G Gagne, J.C. Rogers, Reducing agent-mediated
precipitation of high-abundance plasma proteins, Anal Biochem. 387(2) (2009) 184-
193.
[46] C. Fernández, H.M. Santos, C. Ruíz-Romero, F.J. Blanco, J.L. Capelo-Martínez, A
comparison of depletion versus equalization for reducing high-abundance proteins in
human serum, Electrophoresis 32(21) (2011) 2966-2974.
[47] G. Candiano, M. Bruschi, L. Musante, L. Santucci, G.M. Ghiggeri, B. Carnemolla,
P. Orecchia, L. Zardi, P.G. Righetti, Blue silver: a very sensitive colloidal Coomassie
G-250 staining for proteome analysis, Electrophoresis 25(9) (2004) 1327-1333.
[48] E. Oliveira, J.E. Araújo, S. Gómez-Meire, C. Lodeiro, C. Perez-Melon, E. Iglesias-
Lamas, A. Otero-Glez, J.L. Capelo, H.M. Santos, Proteomics analysis of the peritoneal
dialysate effluent reveals the presence of calcium-regulation proteins and acute
inflammatory response, Clin Proteomics 11(1) (2014) 17.
[49] I.V. Shilov, S.L. Seymour, A.A. Patel, A. Loboda, W.H. Tang, S.P. Keating, C.L.
Hunter, L.M. Nuwaysir, D.A. Schaeffer, The Paragon Algorithm, a next generation
search engine that uses sequence temperature values and feature probabilities to identify
peptides from tandem mass spectra, Mol Cell Proteomics 6 (2007) 1638-1655.
[50] W.H. Tang, I.V. Shilov, S.L. Seymour, Nonlinear fitting method for determining
local false discovery rates from decoy database searches. J Proteome Res 7 (2008)
3661-3667.

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
90
[51] Z.G. Peng, K. Hidajat, M.S. Uddin, Adsorption of bovine serum albumin on
nanosized magnetic particles, J. Colloid Interface Sci. 271 (2004) 277-283.
[52] J.R. de Jesús, R. da Silva Fernandes, G de Souza Pessôa, I.M. Jr. Raimundo, M.A.
Z. Arruda, Depleting high-abundant and enriching low-abundant proteins in human
serum: An evaluation of sample preparation methods using magnetic nanoparticle,
chemical depletion and immunoaffinity techniques, Talanta 170 (2017) 199-209.
[53] M. Rahimi, E.P. Ng, K. Bakhtiari, M. Vinciguerra, H. Ali Ahmad, H. Awala, S.
Mintova, M. Daghighi, F. Bakhshandeh Rostami, M. de Vries, M.M. Motazacker, M.P.
Peppelenbosch, M. Mahmoudi, F. Rezaee, Zeolite Nanoparticles for Selective Sorption
of Plasma Proteins, Sci Rep. 5 (2015) 17259.
[54] J. Kreuter, D. Shamenkov, V. Petrov, P. Ramge, K. Cychutek, C. Koch-Brandt, R.
Alyautdin, Apolipoprotein-mediated transport of nanoparticle-bound drugs across the
blood-brain barrier, J Drug Target 10(4) (2002) 317-325.
[55] S. Wagner, A. Zensi, S.L. Wien, S.E. Tschickardt, W. Maier, T. Vogel, F. Worek,
C.U. Pietrzik, J. Kreuter, H. von Briesen, Uptake mechanism of ApoE-modified
nanoparticles on brain capillary endothelial cells as a blood-brain barrier model, PLoS
One 7(3) (2012) e32568.
[56] K.G. Chen, G. Szakács, J.P. Annereau, F. Rouzaud, X.J. Liang, J.C. Valencia, C.N.
Nagineni, J.J. Hooks, V.J. Hearing, M.M. Gottesman, Principal expression of two
mRNA isoforms (ABCB 5alpha and ABCB 5beta) of the ATP-binding cassette
transporter gene ABCB 5 in melanoma cells and melanocytes, Pigment Cell Res. 18(2)
(2005) 102-112.
[57] Patent No.: US 8.481,273 B2. Inventor: Mary C. Farach-Carson. Date of Patent:
Jul. 9, 2013.
[58] Patent No.: US 9,465,039 B2. Perlecan as a biomarker for renal dysfunction .
Inventors: Koen Kas, Schilde (BE); Griet Vanpoucke, Ingooigem (BE); Piet Moerman,
Deurle (BE). Date of Patent: Oct. 11, 2016.
[59] M. Grønborg, T.Z. Kristiansen, A. Iwahori, R. Chang, R. Reddy, N. Sato, H.
Molina, O.N. Jensen, R.H. Hruban, M.G. Goggins, A. Maitra, A. Pandey, Biomarker
discovery from pancreatic cancer secretome using a differential proteomic approach,
Mol Cell Proteomics. 5(1) (2006) 157-171.

III. Results and Discussion. CHAPTER 1 _____________________________________________________________________________
91
[60] B. Amin, A. Maurer, W. Voelter, A. Melms, H. Kalbacher, New poteintial serum
biomarkers in multiple sclerosis identified by proteomic strategies, Curr Med Chem.
21(13) (2014) 1544-1556.
[61] A.A. Kosova, S.N. Khodyreva, O.I. Lavrik, Role of Glyceraldehyde-3-Phosphate
Dehydrogenase (GAPDH) in DNA Repair, Biochemistry (Mosc) 82(6) (2017) 643-654.
[62] L. Mu-xing, Z. Hong, B. Xin-yu, L. Zhi-yu, Y. Xue-song, L. Huai, H. Zhen, H.
Yue, Z. Jian-guo, Z. Jian-jun, Z. Ye-fan, Z. Dong-bin, C. Jian-qiang, Lactate
dehydrogenase is a prognostic indicator in patients with hepatocellular carcinoma
treated by sorafenib: results from the real-life practice in HBV endemic area,
Oncotarget. 7(52) (2016) 86630-86647.
[63] L. Faloppi, M. Bianconi, R. Giampieri, A. Sobrero, R. Labianca, D. Ferrari, S.
Barni, E. Aitini, A. Zaniboni, C. Boni, F. Caprioni, S. Mosconi, S. Fanello, R. Berardi,
A. Bittoni, K. Andrikou, M. Cinquini, V. Torri, M. Scartozzi, S. Cascinu, Italian Group
for the Study of Digestive Tract Cancer (GISCAD), The value of lactate dehydrogenase
serum levels as a prognostic and predictive factor for advanced pancreatic cancer
patients receiving sorafenib Oncotarget. 6(33) (2015) 35087-35094.
[64] B.A. Katchman, I.T. Ocal, H.E. Cunliffe, Y.-H. Chang, G. Hostetter, A. Watanabe,
J. LoBello, D.F. Lake, Expression of quiescin sulfhydryl oxidase 1 is associated with a
highly invasive phenotype and correlates with a poor prognosis in Luminal B breast
cancer, Breast Cancer Res. 15(2) (2013) R28.
[65] F.H. Knebel, M. Uno, T.F. Galatro, L.P. Bellé, S.M. Oba-Shinjo, S.K.N. Marie, A.
Campa, Serum amyloid A1 is upregulated in human glioblastoma, J Neurooncol.
132(3) (2017) 383-391.
[66] T. Nakano, T. Yoshino, T. Fujimura, S. Arai, A. Mukuno, N. Sato, K. Katsuoka,
Reduced expression of dermcidin, a peptide active against propionibacterium acnes, in
sweat of patients with acne vulgaris, Acta Derm Venereol. 95(7) (2015) 783-786.
[67] M. Chromek, Z. Slamová, P. Bergman, L. Kovács, L. Podracká, I. Ehrén, T.
Hökfelt, G.H. Gudmundsson, R.L. Gallo, B. Agerberth, A. Brauner, The antimicrobial
peptide cathelicidin protects the urinary tract against invasive bacterial infection, Nat
Med. 12 (2016) 636-641.

III. Results and Discussion. CHAPTER 1
______________________________________________________________________
92

CHAPTER 2
Proteomic investigation on bio-corona of Au, Ag and Fe
nanoparticles for the discovery of triple negative breast cancer
serum protein biomarkers
María del Pilar Chantada-Vázquez, Antonio Castro López, María García Vence, Sergio
Vázquez-Estévez, Benigno Acea-Nebril, David G. Calatayud, Teresa Jardiel, Susana B.
Bravo, Cristina Núñez
Journal of Proteomics 212 (2020) 103581
DOI: 10.1016/j.jprot.2019.103581


III. Results and Discussion. CHAPTER 2
______________________________________________________________________
93
Proteomic investigation on bio-corona of Au, Ag and Fe
nanoparticles for the discovery of triple negative breast cancer serum
protein biomarkers
María del Pilar Chantada-Vázquez,a Antonio Castro Lopez,b María García Vence,c
Sergio Vázquez-Estévez,d Benigno Acea-Nebril,e David G. Calatayud,f Teresa Jardiel,f
Susana B. Bravo,c Cristina Núneza
a Research Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de
Saúde (SERGAS), 27002 Lugo, Spain
b Breast Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde
(SERGAS), 27002 Lugo, Spain
c Proteomic Unit, Instituto de Investigaciones Sanitarias-IDIS, Complejo Hospitalario
Universitario de Santiago de Compostela (CHUS), 15706 Santiago de Compostela,
Spain
d Oncology Division, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de
Saúde (SERGAS), 27002 Lugo, Spain
e Department of Surgery, Breast Unit, Complexo Hospitalario Universitario A Coruña
(CHUAC), SERGAS, A Coruña, Spain
f Department of Electroceramics, Instituto de Cerámica y Vidrio-CSIC, Kelsen 5,
Campus de Cantoblanco, 28049 Madrid, Spain
Abstract
Nowadays, there are no targeted therapeutic modalities for triple negative breast
cancer (TNBC). This disease is associated with poor prognosis and worst clinical
outcome because of the aggressive nature of the tumor, delayed diagnosis, and non-
specific symptoms in the early stages. Therefore, identification of novel specific TNBC
serum biomarkers for screening and therapeutic purposes remains an urgent clinical
requirement.
New user-friendly and cheap methods for biomarker identification are needed,
and nanotechnology offers new opportunities. When dispersed in blood, nanoparticles
(NPs) are covered by a protein shell termed “protein corona” (PC). While alterations in

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
94
protein patterns are challeging to detect by conventional blood analyses, PC acts as a
“nano-concentrator” of serum proteins with affinity for NPs’ surface. So, the
characterization of PC could allow the detection of otherwise undetectable changes in
protein concentration at an early stage of the disease or after chemotherapy or surgery.
To explore this research idea, serum samples from 8 triple negative breast cancer
(TNBC) patients and 8 patients without malignancy were allowed to interact with gold
nanoparticles (AuNPs: 10.02 ± 0.91 nm), silver nanoparticles (AgNPs: 9.73 ± 1.70 nm)
and magnetic nanoparticles (MNPs: (9.30 ± 0.67 nm). Here, in order to identify
biomarker candidates in serum of TNBC patients, these nanomaterials were combined
with electrophoretic separation (SDS-PAGE) to performed qualitative and quantitative
comparisons of the serum proteomes of TNBC patients (n = 8) and healthy controls (n =
8) by liquid chromatography tandem-mass spectrometry (LC-MS/MS) analysis. The
results were validated through a sequential window acquisition of all theoretical mass
spectra (SWATH) analysis, performed in total serum samples (patients and controls)
using this approach as a multiple reaction monitoring (MRM) analysis.
Keywords: Triple negative breast cancer (TNBC); Proteomics; Nanoparticles;
Biomarkers; SWATH-MS; Mass spectrometry (MS).
1. Introduction
Breast cancer (BC) is the most frequently diagnosed cancer and the leading
cause of cancer death in women worldwide, accounting for 23% of total new cancer
cases [1].
Mainly, triple negative breast cancer (TNBC) is a heterogeneous disease that is
characterized by a lack of estrogen receptor/progesterone receptor (ER/PgR) expression
and absence of human epidermal growth factor receptor 2 (HER2) overexpression or
amplification. This subgroup accounts for 12-15% of all types of breast cancer and
exhibits a distinct molecular profile, clinical behavior, and response to therapy [2].
Notably, triple negative tumors are usually high grade and exhibit increased
aggressiveness, poor prognosis, and worst clinical outcome [3]. Because hormonal
(tamoxifen) and HER2-directed (trastuzumab) therapies are not effective, TNBC
patients are managed with standard chemotherapy; however, a high rate of local and
systemic relapse is frequently associated with treatment. Unfortunately, no useful

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
95
biomarkers neither targeted therapeutic modalities exist for this breast cancer subtype
[4].
It is well known that proteins secreted from tumor tissues have a higher
likelihood of reaching the systemic circulation and may, therefore, serve as potential
biomarkers for early detection [5]. Serum proteomics is a valuable tool that can
facilitate comprehensive and systematic elucidation of the serum proteome under both
healthy and disease conditions as well as identification of serum protein markers used
for disease diagnosis and prognosis, particularly for identifying breast cancer-specific
markers [5].
Current proteomic technologies that promote large-scale sample screening and
facilitate the identification of proteins associated with disease and treatment are
developing rapidly [6]. Mass spectrometry (MS), a powerful proteomics tool, has
evolved to a high-throughput level, allowing rapid and accurate analysis of several
thousand proteins in a single study [7]. Several studies have addressed the possibility of
applying MS proteome analysis to diagnostics of TNBC, revealing protein patterns
specific for patients with TNBC at either early or late clinical stages [8]. The peptide
markers identified with differentiating patterns include glycolytic enzymes (as for
example MDH2, PGK1, TKT, Aldolase1), cytokeratins (CK7, 8, 9, 14, 17, 19), further
structure proteins (vimentin, fibronectin, L-plastin), for NME1-NME2, lactoferrin, and
members of the Annexin family, among others [9].
SWATH-MS is an emerging technique that combines deep proteome coverage
capabilities with quantitative consistency and accuracy [10]. Mainly, SWATH-MS
analysis offers several advantages, including high reproducibility and reliability of
quantitative information, in discovery proteomics [11]. Furthermore, SWATH-MS
methods can be interchanged to MRM approaches focused on the validated biomarkers.
Therefore, SWATH-MS is an important tool not only for the biomarker discovery but
also for the development of preliminary validation studies [12].
However, currently available proteomic tests detect only a tiny fraction of
potential biomarkers due to their deficient concentration in biofluids, in addition to the
‘swamping’ effect, caused by non-specific highly abundant molecules. The issue of
signal-to-noise exceeds the current capability of proteomic analysis and therefore limits
the diagnostic information that can be obtained [13].

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
96
To overcome these challenges, several approaches have been developed,
comprising depletion of high-abundance proteins that mask less abundant proteins [14,
15], chromatographic or electrophoretic sample prefractionation, to decrease complexity
before mass spectrometric analyses [16,17], and direct isolation of preferred proteins
[18]. Nevertheless, none of the methods can provide a standard solution to biomarker
discovery or can give a reproducible diagnostic platform for establishing biomarker
guides. In this way, one promising way taken is the use of nanoscale materials [19].
Nanotechnology-based platforms hold great promise in addressing the above
fundamental and technical issues of biomarker discovery to overcome persistent
deficiencies of conventional methods. Currently, it is well known that the surfaces of
nanoparticles (NPs) are rapidly covered by different types of biomolecules when they
contact biological media called protein corona (PC) [20].
The protein composition and content in the corona depend on several
parameters, including: i) physicochemical properties of the NPs (i.e. composition, size,
shape, curvature, surface chemistry and surface charge, hydrophobicity/hydrophilicity)
[21-23]; ii) characteristics of biological media (i.e. protein source, and temperature) [24-
26]); iii) incubation time [27].
Notably, the composition of the protein corona varies among healthy
individuals, as well as among patients with various diseases/medical conditions. Thus,
the same NPs may have different protein coronas in different individuals. These
alterations are often small and challenging to be detected by conventional blood
analyses.
On the other side, the protein corona can act as a “nano-concentrator” [28] of
those serum proteins with affinity for the NP surface. Therefore, characterization of
protein corona could allow detecting minor changes in protein concentration at the very
early stages of disease development or even after chemotherapy or surgery (i.e., when
an alteration in circulating level of proteins could be undetectable by blood tests).
Keeping in mind that each disease is characterized by different plasma/serum
proteomes, inducing the formation of different PCs on the same nanomaterial, M.
Mahmoudi, et al. introduced the novel concept of “personalized protein corona” (PPC)
[29]. More specifically, depending on the type, period and severity of the disease (which
determines the serum/plasma alterations), each patient may have a personalized protein
corona.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
97
In the present study, gold nanoparticles (AuNPs: 10.02 ± 0.91 nm), silver
nanoparticles (AgNPs: 9.73 ± 1.70 nm) and magnetic nanoparticles (MNPs: (9.30 ±
0.67 nm) were used to pre-concentrate and separate proteins from sera samples of eight
patients with TNBC as well as from eight healthy people. For protein biomarkers
identification and quantification, the proteome map changes between both groups were
detected using a proteomic approach based on electrophoretic separation (SDS-PAGE)
and mass spectrometry (nLC-MS/MS).
2. Experimental
2.1. Chemicals and reagents
All reagents and solvents used were HPLC-grade or higher. Sodium citrate
tribasic dihydrate, tannic acid, silver nitrate, ammonium hydroxide,
iron(III) chloride hexahydrate and iron(II) sulfate heptahydrate, sodium borohydride
(NaBH4), trypsin, trifluoroacetic acid, DL-Dithiothreitol (DTT), Iodoacetamide (IAA),
acrylamide/bis-acrylamide 30% solution (37.5:1), Glycerol 86-88%, Tris-base,
Coomassie Brilliant Blue R250 (CBB), sodium carbonate, and the Sigma Marker wide
range 6.5-200 kDa were purchased from Sigma-Aldrich (St. Louis, MO, USA). Sodium
dodecyl sulfate (SDS) and formaldehyde were purchased from Panreac (Barcelona,
Spain). β-mercaptoethanol was purchased from Merck (Hohen-Brunn, Germany), and
bromophenol-blue was purchased from Riedel-de Haen (Seelze, Germany). Hydrogen
tetrachloroaurate (III) hydrate (99.9%-Au) (49%Au) at 10%w/v was purchased from
Strem Chemicals (Newburyport, MA, USA). Ammonium bicarbonate (ambic) and
formic acid were purchased from Fluka (Steinheim, Germany).
2.2. Instrumentation
Microscopic characterizations of AuNPs, AgNPs, and MNPs were performed by
transmission electron microscopy (TEM) using a Jeol JEM 1011 microscope. Samples
for TEM were prepared by pipetting a drop of the colloidal dispersion onto an ultrathin
carbon-coated copper grid and allowing the solvent to evaporate. AuNPs, AgNPs and
MNPs ζ-potentials were measured at 25°C before and after protein corona formation
using a Malvern Zetasizer Nano ZS instrument. For ζ-potential measurements samples
were diluted in 1 mL milli-Q water and placed in Zetasizer disposable cuvettes. A
minimum of 3 measurements per sample were made.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
98
Power Pac Basic power supply from Bio-Rad (CA, USA) was used for sodium
dodecyl sulfate- polyacrylamide gel electrophoresis (SDS-PAGE) protein separation.
Protein quantification was accomplished by measuring the absorbance at 280 nm with
the use of a Qubit™ 4 Quantitation Starter Kit from Thermo Fisher Scientific. Gel
image acquisition was carried out with a UVP PhotoDoc-ItTM Imaging System from
Analytik Jena.
2.3. Synthesis of inorganic nanoparticles
2.3.1. Synthesis of citrate-gold nanoparticles (10.02 ± 0.91 nm)
Gold nanoparticles (AuNPs) were synthesized by the citrate reduction method in
aqueous solution [30]. Briefly, 60 ml of sodium citrate tribasic solution (0.075% w/v)
was heated to 100 °C, and then gold was added as 54 μL of 10% w/v of hydrogen
tetrachloroaurate (III) hydrate solution. The reaction mixture was kept under reflux until
a deep red color was detected. The solution of nanoparticles is chilled at room
temperature and stored at 4 ºC for a maximum of one month.
2.3.2. Synthesis of citrate-silver nanoparticles (9.73 ± 1.70 nm)
Silver nanoparticles (AgNPs) were synthesized by the citrate reduction method
in aqueous solution by the method reported by V. Puntes et al. [31]. A 100 mL volume
of an aqueous solution containing sodium citrate (SC) (5 mM) and tannic acid (TA)
(0.025 mM) was prepared and heated with a heating mantle in a three-neck round-
bottomed flask for 15 min under vigorous stirring. A condenser was used to prevent the
evaporation of the solvent. After boiling had commenced, 1 mL of AgNO3 (25 mM)
was injected into this solution. The solution became bright yellow immediately.
Resultant Ag NPs were purified by centrifugation at 18000×g to remove the excess of
TA and further redispersed in Milli-Q-water before sample characterization.
2.3.3. Synthesis of Fe3O4 magnetic nanoparticles (9.30 ± 0.67 nm)
The synthesis of magnetic nanoparticles was performed according to a
previously described procedure [32]. Briefly, 6 mL of concentrated ammonium
hydroxide and 4 mL of water saturated with N2 were mixed in a round-bottom flask
under an inert atmosphere. Subsequently, in another vessel, 1 g of FeCl3.6H2O and
0.25-0.5 g of FeSO4.7H2O were dissolved in 10 mL of water saturated with N2. After
mixing both solutions, the system was kept under constant stirring for 80 min at 80 °C.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
99
The final reaction product was washed three times with deionized water and
magnetically separated for 24 h at room temperature.
2.4. Sample resources
Venous blood samples were obtained from eight females with triple negative
breast cancer (TNBC) and eight disease-free individuals with the use of VACUETTE®
Serum Clot Activator Tubes (10 mL). The collected blood samples were allowed to clot
for 15 min and then centrifuged for 5 min at 4˚C and 1,800×g. Sera were transferred
into clean plastic tubes (1 mL) and immediately frozen at -80˚C at Research Unit,
Hospital Universitario Lucus Augusti (HULA). Clinical features of TNBC tumors,
including tumor size, histology, receptor status, clinical stage, and nodal status, are
summarized in Table 1.
Table 1. Clinical features of triple negative breast cancer tumors.
Characteristics Number
Patients 8
Age (years) < 40 1
40-70 4
> 70 3
Tumor size (cm) < 2 3
2-5 4
>5 1
Histological types Ductal invasive
carcinoma
8
Receptor status Triple negative 8
Clinical stage I 1
II 4
III 3
Nodal status N0 7
N1 1
2.5. Sample preparation
2.5.1. Optimization of protein corona formation in serum using magnetic nanoparticles
A series of serum sample aliquots were used for checking the effects of sample
pH and temperature on the high-abundance proteins depletion using DTT, MNP/protein
ratios, and pH of the medium on the washing steps.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
100
- Depletion of multiple high abundant proteins
Human serum aliquots (x8) were filtered with Miller-GP® Filter Unit
(Millipore) with a size of 0.22 μm. Four aliquots of human serum (30 µL) were depleted
with dithiothreitol (DTT) according to the protocol described by Warder el al. [33, 34].
Briefly, fresh DTT 500 mM (3.3 µL) in milli-Q water was mixed with 30 µL of human
serum and vortex quickly. Samples were then incubated at room temperature until a
viscous white precipitate persisted (60 min), followed by centrifugation at 18,840×g for
20 min. Supernatants were transferred to a clean tube before the protein alkylation and
nanoparticles (NPs) fractionation.
To evaluate the effects of sample pH and temperature on the high-abundance
proteins depletion with DTT, four aliquots of human serum (30 µL) were depleted with
dithiothreitol (DTT) following a modification of the protocol previously published by
Arruda et al. [35]. Four aliquots of 30 μL of human serum were mixed fresh DTT 500
mM (3.3 µL) in ambic (12.5 mmol L-1)] and vortexed. Samples were incubated for 60
min at 37ºC, centrifuged at 13000×g for 40 min to separate supernatants to be alkylated
and fractionated with nanoparticles (NPs).
- NPs protein alkylation and fractionation
After protein depletion, the reduced SH-groups were alkylated with iodoacetic
acid (IAA) for 45 min at room temperature and protected from light. Volumes of serum
reduced and alkylated, were diluted to a final volume of 100 μL in Tris-HCl (0.1 mol L-
1, pH 5.5), and mixed with MNPs (5 μg) to obtain the following MNP/protein ratios:
1:1, 1:2, 1:4, 1:10. Then, all NPs-serum solutions were incubated at 25 °C with shaking
(300 rpm) in a thermostatic bath for 30 min and then pellets were harvested by
centrifugation at 20,186 ×g for 30 min. To evaluate the effects of sample pH on the
stabilization of the protein corona in the washing steps, a fraction of pellets were
washed (x3) with 50 µL of Tris-HCl (0.1 mol L-1, pH 5.5) and another fraction with 50
µL of milli-Q water (x3). In both cases, pellets were harvested again by centrifugation
at 20,186×g for 30 min to remove unbound proteins.
2.5.2. Incubation of nanoparticles with serum samples
Serum aliquots (x2) belonging to the eight disease-free individuals and eight
triple negative breast cancer patients were depleted with dithiothreitol (DTT) following

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
101
the method of Warder el al. [33, 34]. After that, the reduced SH-groups were alkylated
with iodoacetic acid (IAA) at room temperature (45 min in the dark).
After protein reduction and alkylation, serum aliquots (x2) belonging to the eight
disease-free individuals and eight triple negative breast cancer patients were incubated
with AuNPs (10.02 ± 0.91 nm) and AgNPs (9.73 ± 1.70 nm) (4 aliquots per individual,
2 with each nanoparticle type) following the method described by C. Núñez et al. [36].
Briefly, 75 μL of AuNPs (10.02 ± 0.91 nm) and 75 μL of AgNPs (9.73 ± 1.70 nm) were
added to each different serum aliquots (×2) belonging to the eight disease-free
individuals and eight triple negative breast cancer patients (4 aliquots per individual, 2
with each nanoparticle type), followed by the addition of 40 μL of citrate/citric acid
buffer to a final pH of 5.8. Then, all NPs-serum solutions were incubated at 37 °C with
shaking in a thermostatic bath for 30 min. Pellets were harvested by centrifugation at
18,840×g for 30 min. In all cases, pellets containing proteins bound to nanoparticles
were washed three times with 25 μL citrate/citric acid buffer and harvested again by
centrifugation at 18,840×g for 30 min to remove unbound proteins.
In the particular case of magnetic nanoparticles, each different reduced and
alkylated serum aliquot (x2) from disease-free individuals (n = 8) and negative breast
cancer patients (n = 8) were incubated (shaking at 300 rpm, 25 °C, 30 min) with 5 µL of
MNPs (9.30 ± 0.67 nm) after the addition of 87 µL of Tris-HCl (0.1 mol L-1, pH 5.5).
After centrifuging (15000×g, 30 min), pellets were separated and washed (x3)
with 50 µL of Tris-HCl (0.1 mol L-1, pH 5.5) and centrifuged again (20,186×g, 30 min).
2.5.3. Gel electrophoresis
After that, pellets were reconstituted in 10 µL of a buffer with 0.2 M Tris-HCl, 2
% w/v SDS and 20 % v/v glycerol. This 10 µL was mixed with 4 µL of SDS-PAGE
loading buffer (10 % w/v SDS, Tris-Base 40 mM, pH 6.8, 50 % v/v glycerol, 0.1 % v/v
bromophenol blue, 10 % v/v β-mercaptoethanol) in a final volume of 20 µL. Then, all
samples were denatured by heating at 100°C for 5 min and loaded into a 10%
acrylamide/bis-acrylamide, stacking gel /12.5% acrylamide/bis-acrylamide running gel,
of 1 mm thickness, and separated at 180 V (constant voltage) for 120 min. After
electrophoresis, the gel was fixed for 30 minutes with 40% (v/v) ethanol and 10% (v/v)
acetic acid and then stained overnight with Colloidal Coomassie Blue [37]. Gels were
rinsed with distilled water and a 0.5 M sodium chloride solution until a clear

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
102
background was observed. Gel imaging was carried out with a UVP PhotoDoc-ItTM
Imaging System.
2.5.4. In-gel protein digestion
Protein bands were excised manually and transferred to 2.5-mL Lo-Bind tubes,
and then washed twice with water and with 50% (v/v) acetonitrile/ 25 mM ammonium
bicarbonate (ambic) until the blue color disappeared.
Before the trypsin digestion, gel spots were washed with 25 mM ambic and
dehydrated with acetonitrile. Then, 30 μL of trypsin (20 ng·μL-1 in 12.5 mM ambic/2%
(v/v) acetonitrile) was added to the gel spots and incubated for 60 min at 0ºC.
After this time, gel spots were inspected, trypsin solution not absorbed into the
gel was removed, and the gels were covered with 100 μL of 12.5 mM ambic. Samples
were incubated for 12 h at 37°C. Then 50 μL of 5% (v/v) formic acid was added, and
the supernatant was transferred to a new Lo-Bind tube and the peptides were further
extracted from the gel twice with 50% (v/v) acetonitrile/0.1% (v/v) trifluoroacetic acid
(TFA) (x3) and acetonitrile (ACN) (x1). Samples were dried-down and stored at -20 °C
[38].
2.6. Protein identification by mass spectrometry (LC-MS/MS) and data analysis
Digested peptides of each sample were separated using Reverse Phase
Chromatography. The gradient was developed using a micro liquid chromatography
system (Eksigent Technologies nanoLC 400, SCIEX) coupled to high-speed Triple TOF
6600 mass spectrometer (SCIEX) with a microflow source. The analytical column used
was a silica-based reversed phase column Chrom XP C18 150 × 0.30 mm, 3 mm
particle size and 120 Å pore size (Eksigen, SCIEX). The trap column was a YMC-
TRIART C18 (YMC Technologies, Teknokroma with a 3 mm particle size and 120 Å
pore size, switched on-line with the analytical column. The loading pump delivered a
solution of 0.1% formic acid in water at 10 µL/min. The micro-pump provided a flow-
rate of 5 µL/min and was operated under gradient elution conditions, using 0.1% formic
acid in water as mobile phase A, and 0.1% formic acid in acetonitrile as mobile phase
B. Peptides were separated using a 25 minutes gradient ranging from 2% to 90% mobile
phase B (mobile phase A: 2% acetonitrile, 0.1% formic acid; mobile phase B: 100%
acetonitrile, 0.1% formic acid). The injection volume was 4 µL.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
103
Data acquisition was carried out in a TripleTOF 6600 System (SCIEX, Foster
City, CA) using a Data dependent workflow. Source and interface conditions were as
follows: ion spray voltage floating (ISVF) 5500 V, curtain gas (CUR) 25, collision
energy (CE) 10 and ion source gas 1 (GS1) 25. The instrument was operated with
Analyst TF 1.7.1 software (SCIEX, USA). Switching criteria were set to ions greater
than mass to charge ratio (m/z) 350 and smaller than m/z 1400 with a charge state of 2-
5, mass tolerance 250 ppm and an abundance threshold of more than 200 counts (cps).
Former target ions were excluded for 15s. The instrument was automatically calibrated
every 4 hours using as external calibrant tryptic peptides from PepcalMix (Sciex).
After MS/MS analysis, data files were processed using ProteinPilotTM 5.0.1
software from Sciex, which uses the algorithm ParagonTM for database search and
ProgroupTM for data grouping. Data were searched using a Human-specific UniProt
database. False discovery rate was performed using a non-linear fitting method
displaying only those results that reported a 1% Global false discovery rate or better
[39, 40].
2.7. Protein quantification by SWATH (Sequential Window Acquisition of all
Theoretical Mass Spectra)
2.7.1. Creation of the spectral library
To construct the MS/MS spectral libraries, the peptide solutions were analyzed
by a shotgun data-dependent acquisition (DDA) approach by micro-LC-MS/MS. To get
a good representation of the peptides and proteins present in all samples, pooled vials of
samples from each group (control and triple negative breast cancer patients) were
prepared using equal mixtures of the original samples. 4 μL (4 mg) of each pool was
separated into a micro-LC system Ekspert nLC425 (Eksigen. Dublin. CA. USA) using a
column Chrom XP C18 150 × 0.30 mm. 3 mm particle size and 120 Å pore size
(Eksigent, Sciex) at a flow rate of 5 µL/min. Water and ACN, both containing 0.1%
formic acid, were used as solvents A and B, respectively. The gradient run consisted of
5% to 95% B for 30 min, 5 min at 90% B and finally 5 min at 5% B for column
equilibration, for a total run time of 40 min. When the peptides eluted, they were
directly injected into a hybrid quadrupole-TOF mass spectrometer Triple TOF 6600
(Sciex, Redwood City. CA. USA) operated with a data-dependent acquisition system in
positive ion mode. A Micro source (Sciex) was used for the interface between microLC

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
104
and MS, with an application of 2600 V voltage. The acquisition mode consisted of a
250 ms survey MS scan from 400 to 1250 m/z followed by an MS/MS scan from 100 to
1500 m/z (25 ms acquisition time) of the top 65 precursor ions from the survey scan, for
a total cycle time of 2.8 s. The fragmented precursors were then added to a dynamic
exclusion list for 15 s; any singly charged ions were excluded from the MS/MS
analysis.
The peptide and protein identifications were performed using Protein Pilot
software (version 5.0.1. Sciex) with a Data were searched using a Human-specific
UniProt database, specifying iodoacetamide as Cys alkylation. The false discovery rate
(FDR) was set to 1 for both peptides and proteins. The MS/MS spectra of the identified
peptides were then used to generate the spectral library for SWATH peak extraction
using the add-in for PeakView Software (version 2.2. Sciex) MS/MSALL with SWATH
Acquisition MicroApp (version 2.0. Sciex). Peptides with a confidence score above
99% (as obtained from Protein Pilot database search) were included in the spectral
library).
2.7.2. Relative quantification by SWATH acquisition
SWATH-MS (Sequential Window Acquisition of all Theoretical Mass Spectra)
acquisition was performed on a TripleTOF® 6600 LC-MS/MS system (Sciex). Samples
from control and triple negative breast cancer patients were analyzed using data-
independent acquisition (DIA) method (30 total samples). Each sample (4 μL (from a
mg/ml solution) was analyzed using the LC-MS equipment and LC gradient described
above for building the spectral library but instead using the SWATH-MS acquisition
method. The method consisted of repeating a cycle that consisted of the acquisition of
65 TOF MS/MS scans (400 to 1500 m/z, high sensitivity mode, 50 ms acquisition time)
of overlapping sequential precursor isolation windows of variable width (1 m/z overlap)
covering the 400 to 1250 m/z mass range with a previous TOF MS scan (400 to 1500
m/z. 50 ms acquisition time) for each cycle. The total cycle time was 6.3 s. For each
sample set, the width of the 100 variable windows was optimized according to the ion
density found in the DDA runs using a SWATH variable window calculator worksheet
from Sciex.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
105
2.7.3. Data analysis
The targeted data extraction of the fragment ion chromatogram traces from the
SWATH runs was performed by PeakView (version 2.2) using the SWATH Acquisition
MicroApp (version 2.0). This application processed the data using the spectral library
created from the shotgun data. Up to ten peptides per protein and seven fragments per
peptide were selected, based on signal intensity; any shared and modified peptides were
excluded from the processing. Five-minute windows and 30 ppm widths were used to
extract the ion chromatograms; SWATH quantitation was attempted for all proteins in
the ion library that were identified by ProteinPilot with an FDR below 1%.
The retention times from the peptides that were selected for each protein were
realigned in each run according to the iRT peptides spiked in each sample and eluted
along the whole-time axis. The extracted ion chromatograms were then generated for
each selected fragment ion; the peak areas for the peptides were obtained by summing
the peak areas from the corresponding fragment ions. PeakView computed an FDR and
a score for each assigned peptide according to the chromatographic and spectral
components; only peptides with an FDR below 1% were used for protein quantitation.
Protein quantitation was calculated by adding the peak areas of the corresponding
peptides.
The integrated peak areas (processed, mrkvw files from PeakView) were directly
exported to the MarkerView software (Sciex) for relative quantitative analysis. The
export will generate three files containing quantitative information about individual
ions, the summed intensity of different ions for a particular peptide, and the summed
intensity of different peptides for a specific protein. MarkerView has been used for the
analysis of SWATH-MS data reported in other proteomics studies [41-44] because of its
data-independent method of quantitation. MarkerView uses processing algorithms that
accurately find chromatographic and spectral peaks direct from the raw SWATH data.
Data alignment by MarkerView compensates for minor variations in both mass and
retention time values, ensuring that identical compounds in different samples are
accurately compared to one another.
To control for possible uneven sample loss across the different samples during
the sample preparation process, we performed a global normalization based on the total
sum of all the peak areas extracted from all the peptides and transitions across the

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
106
replicates of each sample. Unsupervised multivariate statistical analysis using principal
component analysis (PCA) was performed to compare the data across the samples. The
average MS peak area of each protein was derived from the biological replicates of the
SWATH-MS of each sample followed by Student’s t-test analysis using the
MarkerView software for comparison among the samples based on the averaged area
sums of all the transitions derived for each protein. The t-test will indicate how well
each variable distinguishes the two groups, reported as a p-value. To set of differentially
expressed proteins (p-value <0.05) with a 1.5 fold in- or decrease was selected.
Functional analysis was performed by FunRich open access software (Functional
Enrichment analysis tool) for functional enrichment and interaction network analysis
(http://funrich.org/index.html).
2.8. TNBC biomarkers validation
A SWATH-MS analysis was performed using the same conditions described in
section 2.7. In the validation phase, total serum samples from control and triple negative
breast cancer patients previously depleted with DTT were used.
To perform the biomarker validation, the SWATH library was performed using
not only serum pools from incubation with the different nanoparticles but also pools of
total serum samples. Therefore, we improve our library and perform better protein
quantification to achieve biomarker validation.
3. Results and discussion
Following the synthetic methods described by R. López-Cortés [30], V. Puntes
et al. [31] and F. Schüth et al. [32], AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm)
and MNPs (9.30 ± 0.67 nm) were successfully obtained, respectively.
The sizes and ζ-potential of AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm)
and MNPs (9.30 ± 0.67 nm) were examined before and after their incubation with two
pools of human blood serum from healthy individuals and triple negative breast cancer
patients, following the conditions described in section section 2.5.2.
TEM and ζ-potential measurements after the incubation of AuNPs (10.02 ± 0.91
nm), AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm) with serum demonstrated
that, in all cases, the size did not change significantly, and the surface charge remained

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
107
negative (see Fig 1_SM to Fig 10_SM). Upon serum incubation, the mean particle
surface charge of the AuNPs (10.02 ± 0.91 nm) increased (became less negative) from -
37.0 mV to -29.7 mV, and the same measurement for the MNPs (9.30 ± 0.67 nm)
increased from -30.5 mV to -29.3 mV. However, upon serum incubation the mean
particle surface charge of the AgNPs (9.73 ± 1.70 nm) decreased (became more
negative) from -27.4 mV to -30.0 mV. These results agree with previous studies
suggesting that negatively charged NPs do not exclusively interact with positively
charged proteins, as electrostatic interactions are not the only driving force behind NP-
corona interactions [45, 46]. Interestingly, for AuNPs (10.02 ± 0.91 nm) and MNPs
(9.30 ± 0.67 nm), where the ζ-potential was shifted toward less negative values, it could
be suggested preferential interaction with positively charged proteins. The presence of
negatively charged proteins can be explained by a sequential model of protein binding,
in which positively charged proteins initially bind the NP, followed by negatively
charged ones [45,46].
3.1. Optimization of parameters for the protein corona formation in serum using
MNPs
As mentioned above, a great number of variables could influence the efficiency
of protein adsorption on the MNPs surface [47]. For this reason, three parameters were
evaluated: (i) the effects of sample pH and temperature on the depletion of high-
abundance proteins presented in serum using DTT, (ii) MNP/protein ratios; (iii) and pH
of the medium on the washing steps. For this study, shaking and incubation temperature
were previously defined as 300 rpm and 25 ºC, respectively.
In order to evaluate the effects of sample pH and temperature on the depletion of
high-abundance proteins, four human serum samples (x2) (30 µL) were depleted with
fresh DTT 500 mM (3.3 µL) in milli-Q H2O for 60 min at room temperature (protocol
of Warder el al. [33, 34]), and four human serum samples (x2) (30 µL) were depleted
with fresh DTT 500 mM (3.3 µL) in ambic (12.5 mmol L-1) for 60 min at 37ºC
(modification of protocol described by Arruda [35]). In both cases, after the incubation
and centrifugation, supernatants were transferred to a clean tube before the protein
alkylation and nanoparticles (NPs) fractionation. Depletion with fresh DTT 500 mM in
milli-Q for 60 min at room temperature showed more reproducible results (see Figure
4_SM).

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
108
Protein concentration is another critical parameter that may affect the capacity
and kinetics of protein adsorption. To investigate the influence of the MNP/protein ratio
on the formation of the protein corona, volumes of serum reduced and alkylated (x2)
were mixed with MNPs (9.30 ± 0.67 nm), at MNP/protein ratios of 1:1, 1:2, 1:4, and
1:10 (see section 2.5.1). Maintaining the amount of adsorbent (i.e., MNPs) constant and
increasing the protein concentration, would be expected to lead to a decrease of
available adsorption sites, reducing the efficiency of protein removal [48]. As a
compromise between MNPs and protein corona formation, the 1:2 ratio (MNP/protein)
was then selected for future experiments (see Figure 5_SM).
The pH value is an essential parameter because it influences the charge state of
proteins, therefore influencing their interaction with MNPs [49]. To evaluate the effects
of sample pH on the stabilization of the protein corona in the washing steps, a fraction
of pellets was washed three times with 50 µL Tris-HCl (0.1 mol L-1, pH 5.5) and
another fraction was washed three times with milli-Q water. The first one was selected
as the preferred method, because the washes with milli-Q water promoted the
destabilization of the protein corona formed around the MNPs, due to the modifications
of the pH (data not shown).
3.2. Serum fraction preparation and protein corona purification (patients vs. controls)
Serum aliquots (x2) belonging to the eight disease-free individuals and serum
samples from eight triple negative breast cancer patients were depleted with
dithiothreitol (DTT) according to the protocol described by Warder el al. [33, 34]. After
that, the reduced SH-groups were alkylated with iodoacetic acid (IAA) for 45 min at
room temperature and protected from light.
After protein reduction and alkylation, serum aliquots (x2) belonging to the eight
disease-free individuals and eight triple negative breast cancer patients were incubated
with AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm) (6
aliquots per individual, 2 with each nanoparticle type) and further processed as
described in section 2.2.5.
Two protein fractions were thus obtained in each case, one in the supernatant
and the second one attached to the surface of each nanoparticles types (protein corona).
Then, protein fractions (AuNPs-protein corona, AgNPs-protein corona, MNPs-protein
corona) were separately loaded onto a 1D-SDS-PAGE. Proteins were separated and,
after staining, gel bands were excised and submitted to the sample incubation described

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
109
in section 2.5.4. The resulting peptides were then analyzed by mass spectrometry (LC-
MS/MS) for protein identification.
Figure 1 shows the 1D gels for the protein corona formed around the three
different NPs (AuNPs-protein corona, AgNPs-protein corona, MNPs-protein corona)
visible after Coomassie staining. As may be seen, it is quickly noted that there is a
difference in the intensity of the bands on the gel profiles for the healthy controls (from
C1 to C8) and the patients (from P1 to P8) for each type of nanoparticle. However, no
conclusion can be drawn unless the proteins are identified.
Figure 1. 1D-SDS-PAGE of protein coronas formed around 10.02 ± 0.91 nm gold
nanoparticles (AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs) and 9.30 ± 0.67
nm magnetic nanoparticles (MNPs) after their incubation with serum aliquots (x2)
belonging to the eight disease-free individuals (C1 to C8) and eight triple negative
breast cancer patients (P1 to P8). On the left, it marks the lane with Mw protein
standards.
As Figure 2 shows, 192, 161 and 142 proteins were commonly detected in the
protein corona formed around AuNPs, AgNPs, and MNPs, after their incubation with
serum samples of the eight-triple negative breast cancer patients and eight healthy
controls, respectively.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
110
Figure 2. Quantitative Venn diagrams showing the number of identified proteins found
in the protein corona of 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nm
silver nanoparticles (AgNPs) and 9.30 ± 0.67 nm magnetic nanoparticles (MNPs) after
their incubation with serum from eight triple negative breast cancer patients and eight
healthy controls.
In the case of all serum samples from healthy controls, 285, 292 and 206 were
identified on the surface of AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and
MNPs (9.30 ± 0.67 nm), respectively. From them, 149 proteins were commonly
detected in the protein corona of all three different types of NPs. However, 202 different
proteins were found on the three distinct NPs surface: 71 different proteins on the
AuNPs, 85 on the AgNPs, and 46 individual proteins on the MNPs (see Table 1_SM).
Furthermore, 231, 206 and 203 were found in the surface of AuNPs (10.02 ±
0.91 nm), AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm) after their incubation
with all serum aliquots (x2) from eight triple negative breast cancer patients (6 protein
samples per individual: 2 incubated with AuNPs, 2 with AgNPs and 2 with MNPs) (see
Table 2_SM). A total of 138 proteins were commonly found in the protein corona of all
three different types of NPs. However, 142 different proteins were found in the three
different NPs surface: 56 different proteins on the AuNPs, 33 on the AgNPs and 53
individual proteins on the MNPs.
Fractionation of the proteome using AuNPs, AgNPs, and MNPs allows for the
identification of 39 (see Table 2), 45 (see Table 3) and 61 (see Table 4) protein
biomarkers in the pellet of all patient samples, respectively (see Figure 3). Remarkably,

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
111
these proteins were not identified in the healthy control. These potential biomarkers
came from different cellular components; most of them from the nucleus and cytoplasm
(see Figure 4). Protein biomarkers also showed different functionality and are
constituted by proteins implicated in the immune response, followed by proteins with an
enzymatic function, structural, transporter, inflammatory, signal transduction, and with
antibiotic/antibacterial properties.
Importantly, the GRF-type zinc finger domain-containing protein 1 (protein
ZGRF1) was identified in the protein corona of AuNPs, AgNPs and MNPs after their
incubation with the serum samples of all triple negative breast cancer patients. Zinc-
finger proteins (ZNFs) are one of the most abundant groups of proteins and have a wide
range of molecular functions. Given the wide variety of zinc-finger domains, ZNFs can
interact with DNA, RNA, PAR (poly-ADP-ribose) and other proteins. Thus, ZNFs are
involved in the regulation of several cellular processes. ZNFs are implicated in
transcriptional regulation, ubiquitin-mediated protein degradation, signal transduction,
actin targeting, DNA repair, cell migration, and numerous other processes [50].
Notably, overexpression of similar zinc finger proteins has been shown to promote cell
growth and metastasis in laryngeal squamous cell carcinoma, glioma, non-small cell
lung cancer, gastric cancer, oral squamous cell carcinoma, gallbladder cancer, and
breast cancer [51], and also in triple negative breast cancers [52].
Matrix metalloproteinase-9 (MMP9) was identified in the protein corona of
AuNPs and AgNPs after their incubation with the serum samples of all triple negative
breast cancer patients. Members of the matrix metalloproteinase (MMP) family have
been identified as poor prognosis markers for breast cancer patients and as drivers of
many facets of the tumor phenotype in experimental models [53]. Studies of the
pathological processes involved in tumor progression and metastasis revealed matrix
metalloproteinases (MMPs) as prominent molecules engaged in shaping the tumor
microenvironment and driving cancer progression and metastasis [54-56]. Mainly,
MMP9 was investigated as a potential tumor marker in breast cancer [57]. MMP-9 is
one of 70 genes in the Rosetta poor prognosis signature for breast cancer patients [58],
the basis for the clinically implemented Mammaprint prognostic assay (Agendia Inc.,
Irvine, CA). MMP-9 was also highly expressed in node-positive tumors and the

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
112
preoperative blood serum of patients, but MMP-9 activity was appreciably inhibited in
blood serum samples collected after surgery.
Lebercilin and Immunoglobulin lambda variable 3-27 (LV327) were identified
in the protein corona of AuNPs and MNPs after their incubation with the serum samples
of all triple negative breast cancer patients. While the protein expression of lebercilin
was observed in several tissue cancers like colorectal cancer, breast cancer, prostate
cancer, lung cancer (The Human Protein Atlas,
https://www.proteinatlas.org/ENSG00000157578-LCA5L/pathology), immunoglobulin
free light chains as LV327 are biomarkers of poor prognosis in basal-like breast cancer
and are potential targets in tumor-associated inflammation [59].
LINE-1 type transposase domain-containing protein 1 (LITD1), structural
maintenance of chromosomes protein 6 (SMC6) and short coiled-coil protein (SCOC)
were identified in the protein corona of AgNPs and MNPs after their incubation with
the serum samples of all triple negative breast cancer patients.
L1TD1 is an RNA-binding protein that involved with self-renewal of
undifferentiated human embryonic stem cells and cancer cell proliferation [60].
The structural maintenance of chromosomes (SMC) proteins are essential for
successful chromosome transmission during replication and segregation of the genome
in all organisms. SMC proteins function together with other proteins in a range of
chromosomal transactions, including chromosome condensation, sister-chromatid
cohesion, recombination, DNA repair, and epigenetic silencing of gene expression.
Notably, the protein expression of SMC6 was observed in different tissues as colorectal
cancer, breast cancer, prostate cancer, lung cancer, and liver cancer (The Human Protein
Atlas, https://www.proteinatlas.org/ENSG00000163029-SMC6/pathology). In humans,
SCOC is required for autophagosome formation during amino acid starvation [61];
however, this relation with cancer is unknown until the moment.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
113
Figure 3. Quantitative Venn diagrams showing the number of identified protein
biomarkers found on the surface of the three different nanoparticles (color: black) and
commonly found on the surface of AuNPs (10.02 ± 0.91), AgNPs (9.73 ± 1.70 nm) and
MNPs (9.30 ± 0.67 nm) (color: grey).
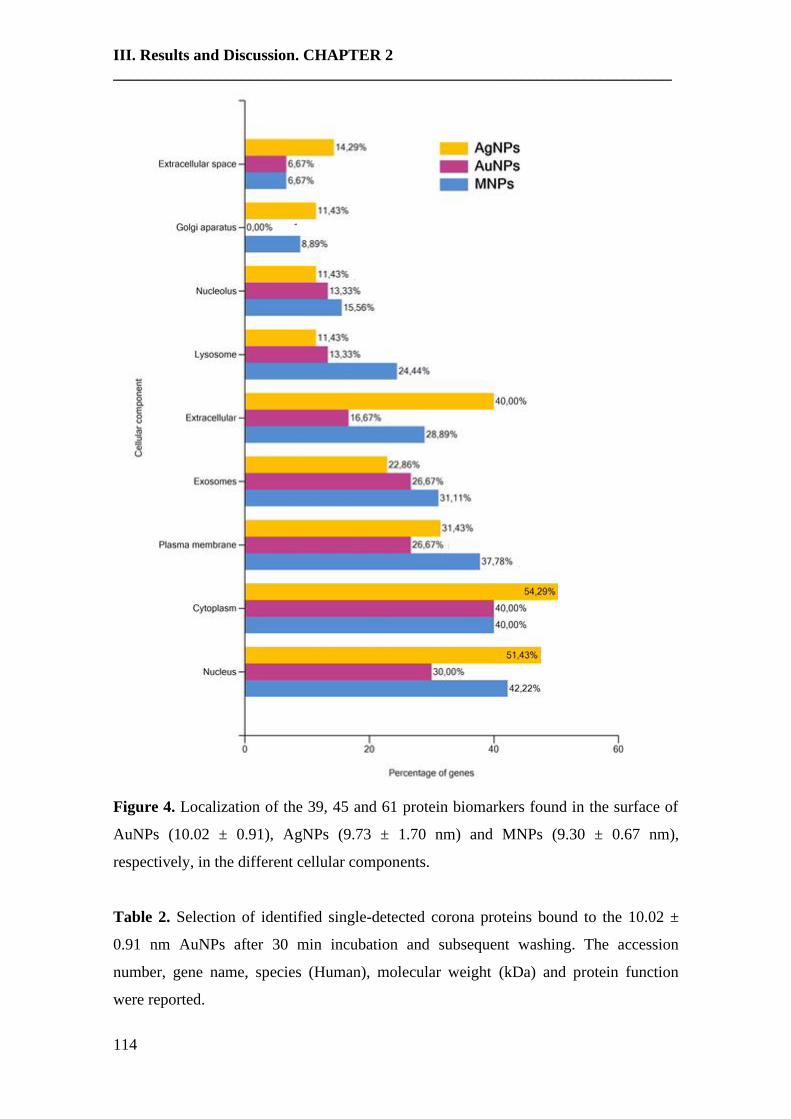
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
114
Figure 4. Localization of the 39, 45 and 61 protein biomarkers found in the surface of
AuNPs (10.02 ± 0.91), AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm),
respectively, in the different cellular components.
Table 2. Selection of identified single-detected corona proteins bound to the 10.02 ±
0.91 nm AuNPs after 30 min incubation and subsequent washing. The accession
number, gene name, species (Human), molecular weight (kDa) and protein function
were reported.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
115
Protein Name Entry Name Gene Function Actin, cytoplasmic 2 P63261 ACTG1 Cell mobility
Atypical kinase COQ8B,
mitochondrial Q96D53 COQ8B Involved in the biosynthesis of coenzyme Q
Beta-ureidopropionase Q9UBR1 UPB1 Catalytic activity
Brefeldin A-inhibited guanine
nucleotide-exchange protein 3 Q5TH69 ARFGEF3
Participates in the regulation of systemic
glucose homeostasis
85/88 kDa calcium-independent
phospholipase A2 O60733 PLA2G6 Catalytic activity
Cilia- and flagella-associated
protein 100 Q494V2 CFAP100
Plays a role in ciliary/flagellar motility by
regulating the assembly and the activity of
axonemal inner dynein arm
Coagulation factor XI P03951 F11 Catalytic activity
Contactin-associated protein-like 2 Q9UHC6 CNTNAP2
Plays a role in the formation of functional
distinct domains critical for saltatory
conduction of nerve impulses in myelinated
nerve fibers
DNA topoisomerase 1 P11387 TOP1 Catalyctic activity
Dynein assembly factor 1,
axonemal Q8NEP3 DNAAF1
Plays a role in cytoplasmic preassembly of
dynein arms
E3 ubiquitin-protein ligase SHPRH Q149N8 SHPRH Enzyme involved in DNA repair
Estrogen sulfotransferase P49888 SULT1E1
Catalytic activity. May play a role in the
regulation of estrogen receptor activity by
metabolizing free estradiol
[F-actin]-monooxygenase MICAL3 Q7RTP6 MICAL3 Catalytic activity
Glial fibrillary acidic protein P14136 GFAP Cell-specific marker during the development
of the central nervous system
Histone-lysine N-methyltransferase
2D O14686 KMT2D
Catalytic activity. Acts as a coactivator for
estrogen receptor by being recruited by
ESR1, thereby activating transcription
Immunoglobulin lambda variable
3-27 P01718 IGLV3-27 Immune response
Inositol hexakisphosphate kinase 2 Q9UHH9 IP6K2 Catalytic activity
Inositol 1,4,5-trisphosphate
receptor type 3 Q14573 ITPR3 Receptor for inositol 1,4,5-trisphosphate
Intraflagellar transport protein 74
homolog Q96LB3 IFT74 Transporter activity
Janus kinase and microtubule-
interacting protein 1 Q96N16 JAKMIP1
Plays a role in the microtubule-dependent
transport of the GABA-B receptor
Lebercilin Q86VQ0 LCA5 Transporter activity
MaFF-interacting protein Q8WZ33 MAFIP Inhibits cell growth and colony-forming
efficiency
Matrix metalloproteinase-9 P14780 MMP9 Catalytic activity
Microtubule-associated protein 2 P11137 MAP2 Stabilizes the microtubules against
depolymerization
Mucolipin-3 Q8TDD5 MCOLN3 Pays a role in the regulation of membrane
trafficking events
Multidrug resistance protein 1 P08183 ABCB1 Responsible for decreased drug accumulation
in multidrug-resistant cells
Nebulette O76041 NEBL Plays an important role in the assembly of the
Z-disk
Nucleoporin NUP188 homolog Q5SRE5 NUP188 May function as a component of the nuclear
pore complex (NPC)
PC membrane recruitment protein
2 Q8N7J2 AMER2
Negative regulator of the canonical Wnt
signaling pathway involved in
neuroectodermal patterning
Protein ELYS Q8WYP5 AHCTF1
Required for the assembly of a functional
nuclear pore complex (NPC) on the surface of
chromosomes
Protein phosphatase 1 regulatory
subunit 26 Q5T8A7 PPP1R26
Inhibits phosphatase activity of protein
phosphatase 1 (PP1) complexes. May
positively regulate cell proliferation.
Protein ZGRF1 Q86YA3 ZGRF1 Zinc ion binding that inhibits factors Xa and
XIa of the coagulation cascade
Putative transmembrane protein Q96M19 LINC00477 Product of a dubious CDS prediction. May be
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
116
encoded by LINC00477 a non-coding RNA
Serine protease 33 PRSS33 PRSS33 Catalytic activity
Serine-protein kinase ATM Q13315 ATM
Activates checkpoint signaling upon double
strand breaks (DSBs), apoptosis and
genotoxic stresses
Serum amyloid A-1 protein P0DJI8 SAA1 Inflammatory response
Sodium-dependent noradrenaline
transporter P23975 SLC6A2 Transporter activity
Suppressor of tumorigenicity 7
protein Q9NRC1 ST7 Acts as a tumor suppressor
Vasorin Q6EMK4 VASN May act as an inhibitor of TGF-beta signaling
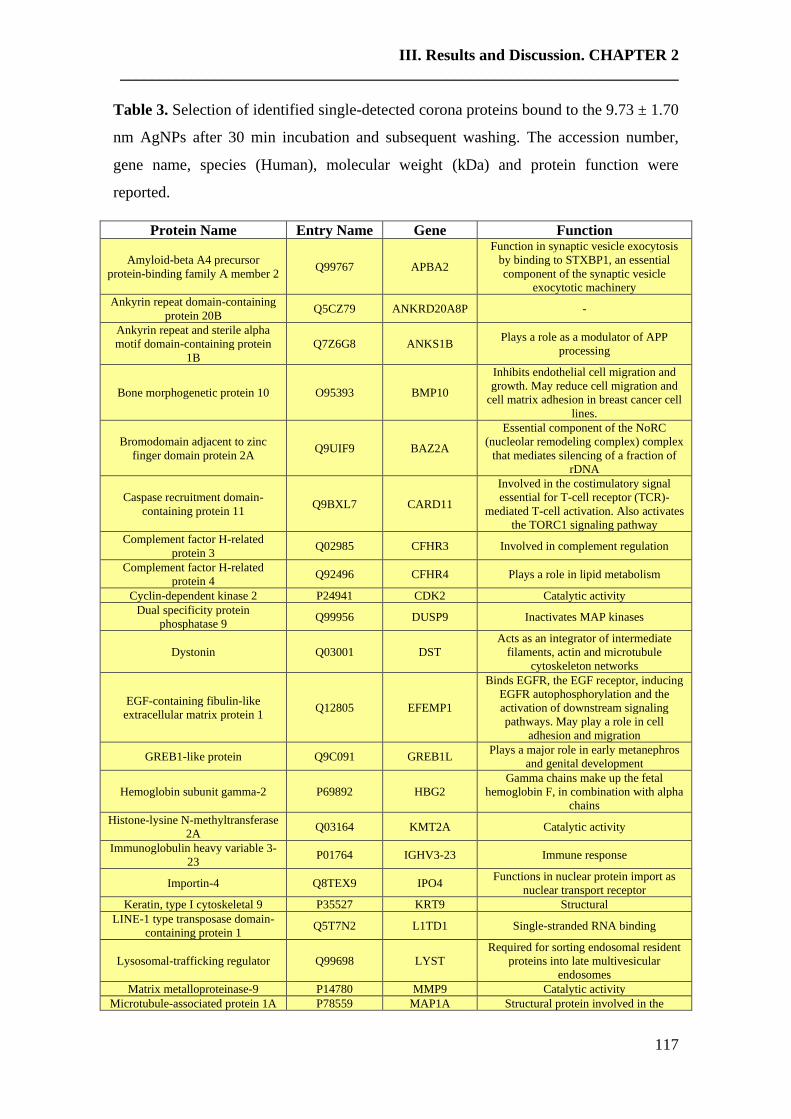
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
117
Table 3. Selection of identified single-detected corona proteins bound to the 9.73 ± 1.70
nm AgNPs after 30 min incubation and subsequent washing. The accession number,
gene name, species (Human), molecular weight (kDa) and protein function were
reported.
Protein Name Entry Name Gene Function
Amyloid-beta A4 precursor
protein-binding family A member 2 Q99767 APBA2
Function in synaptic vesicle exocytosis
by binding to STXBP1, an essential
component of the synaptic vesicle
exocytotic machinery
Ankyrin repeat domain-containing
protein 20B Q5CZ79 ANKRD20A8P -
Ankyrin repeat and sterile alpha
motif domain-containing protein
1B
Q7Z6G8 ANKS1B Plays a role as a modulator of APP
processing
Bone morphogenetic protein 10 O95393 BMP10
Inhibits endothelial cell migration and
growth. May reduce cell migration and
cell matrix adhesion in breast cancer cell
lines.
Bromodomain adjacent to zinc
finger domain protein 2A Q9UIF9 BAZ2A
Essential component of the NoRC
(nucleolar remodeling complex) complex
that mediates silencing of a fraction of
rDNA
Caspase recruitment domain-
containing protein 11 Q9BXL7 CARD11
Involved in the costimulatory signal
essential for T-cell receptor (TCR)-
mediated T-cell activation. Also activates
the TORC1 signaling pathway
Complement factor H-related
protein 3 Q02985 CFHR3 Involved in complement regulation
Complement factor H-related
protein 4 Q92496 CFHR4 Plays a role in lipid metabolism
Cyclin-dependent kinase 2 P24941 CDK2 Catalytic activity
Dual specificity protein
phosphatase 9 Q99956 DUSP9 Inactivates MAP kinases
Dystonin Q03001 DST
Acts as an integrator of intermediate
filaments, actin and microtubule
cytoskeleton networks
EGF-containing fibulin-like
extracellular matrix protein 1 Q12805 EFEMP1
Binds EGFR, the EGF receptor, inducing
EGFR autophosphorylation and the
activation of downstream signaling
pathways. May play a role in cell
adhesion and migration
GREB1-like protein Q9C091 GREB1L Plays a major role in early metanephros
and genital development
Hemoglobin subunit gamma-2 P69892 HBG2
Gamma chains make up the fetal
hemoglobin F, in combination with alpha
chains
Histone-lysine N-methyltransferase
2A Q03164 KMT2A Catalytic activity
Immunoglobulin heavy variable 3-
23 P01764 IGHV3-23 Immune response
Importin-4 Q8TEX9 IPO4 Functions in nuclear protein import as
nuclear transport receptor
Keratin, type I cytoskeletal 9 P35527 KRT9 Structural
LINE-1 type transposase domain-
containing protein 1 Q5T7N2 L1TD1 Single-stranded RNA binding
Lysosomal-trafficking regulator Q99698 LYST
Required for sorting endosomal resident
proteins into late multivesicular
endosomes
Matrix metalloproteinase-9 P14780 MMP9 Catalytic activity
Microtubule-associated protein 1A P78559 MAP1A Structural protein involved in the
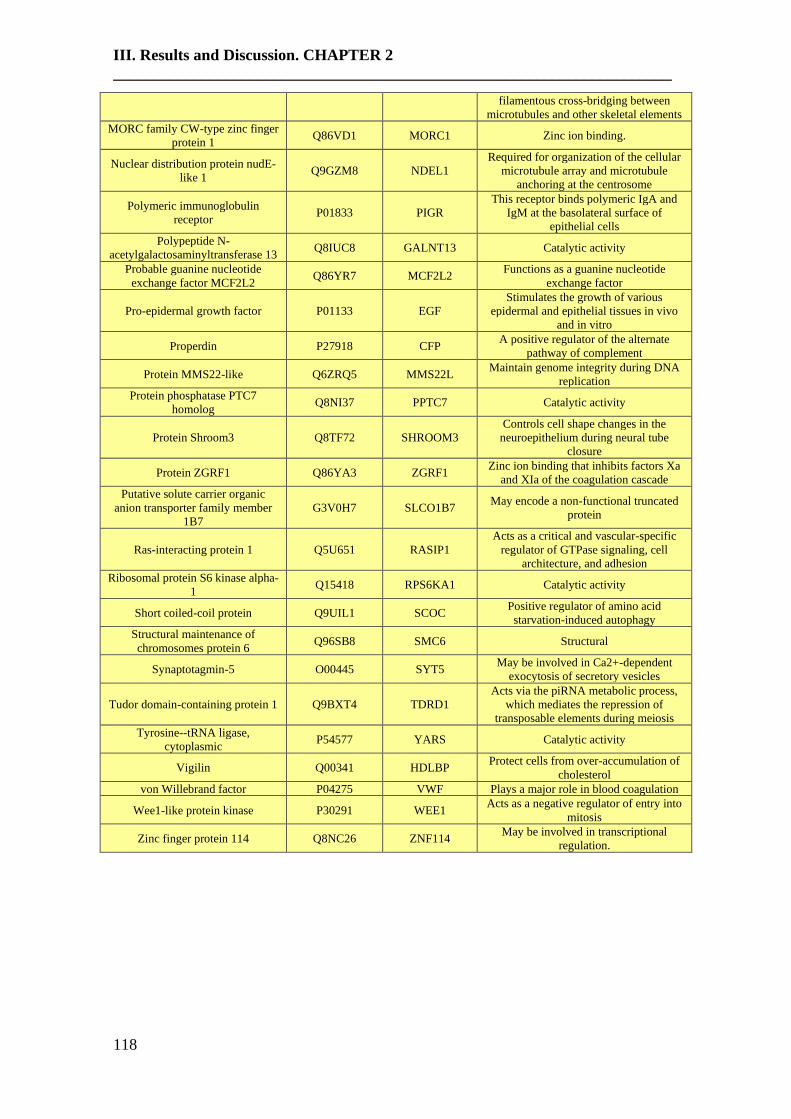
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
118
filamentous cross-bridging between
microtubules and other skeletal elements
MORC family CW-type zinc finger
protein 1 Q86VD1 MORC1 Zinc ion binding.
Nuclear distribution protein nudE-
like 1 Q9GZM8 NDEL1
Required for organization of the cellular
microtubule array and microtubule
anchoring at the centrosome
Polymeric immunoglobulin
receptor P01833 PIGR
This receptor binds polymeric IgA and
IgM at the basolateral surface of
epithelial cells
Polypeptide N-
acetylgalactosaminyltransferase 13 Q8IUC8 GALNT13 Catalytic activity
Probable guanine nucleotide
exchange factor MCF2L2 Q86YR7 MCF2L2
Functions as a guanine nucleotide
exchange factor
Pro-epidermal growth factor P01133 EGF
Stimulates the growth of various
epidermal and epithelial tissues in vivo
and in vitro
Properdin P27918 CFP A positive regulator of the alternate
pathway of complement
Protein MMS22-like Q6ZRQ5 MMS22L Maintain genome integrity during DNA
replication
Protein phosphatase PTC7
homolog Q8NI37 PPTC7 Catalytic activity
Protein Shroom3 Q8TF72 SHROOM3
Controls cell shape changes in the
neuroepithelium during neural tube
closure
Protein ZGRF1 Q86YA3 ZGRF1 Zinc ion binding that inhibits factors Xa
and XIa of the coagulation cascade
Putative solute carrier organic
anion transporter family member
1B7
G3V0H7 SLCO1B7 May encode a non-functional truncated
protein
Ras-interacting protein 1 Q5U651 RASIP1
Acts as a critical and vascular-specific
regulator of GTPase signaling, cell
architecture, and adhesion
Ribosomal protein S6 kinase alpha-
1 Q15418 RPS6KA1 Catalytic activity
Short coiled-coil protein Q9UIL1 SCOC Positive regulator of amino acid
starvation-induced autophagy
Structural maintenance of
chromosomes protein 6 Q96SB8 SMC6 Structural
Synaptotagmin-5 O00445 SYT5 May be involved in Ca2+-dependent
exocytosis of secretory vesicles
Tudor domain-containing protein 1 Q9BXT4 TDRD1
Acts via the piRNA metabolic process,
which mediates the repression of
transposable elements during meiosis
Tyrosine--tRNA ligase,
cytoplasmic P54577 YARS Catalytic activity
Vigilin Q00341 HDLBP Protect cells from over-accumulation of
cholesterol
von Willebrand factor P04275 VWF Plays a major role in blood coagulation
Wee1-like protein kinase P30291 WEE1 Acts as a negative regulator of entry into
mitosis
Zinc finger protein 114 Q8NC26 ZNF114 May be involved in transcriptional
regulation.
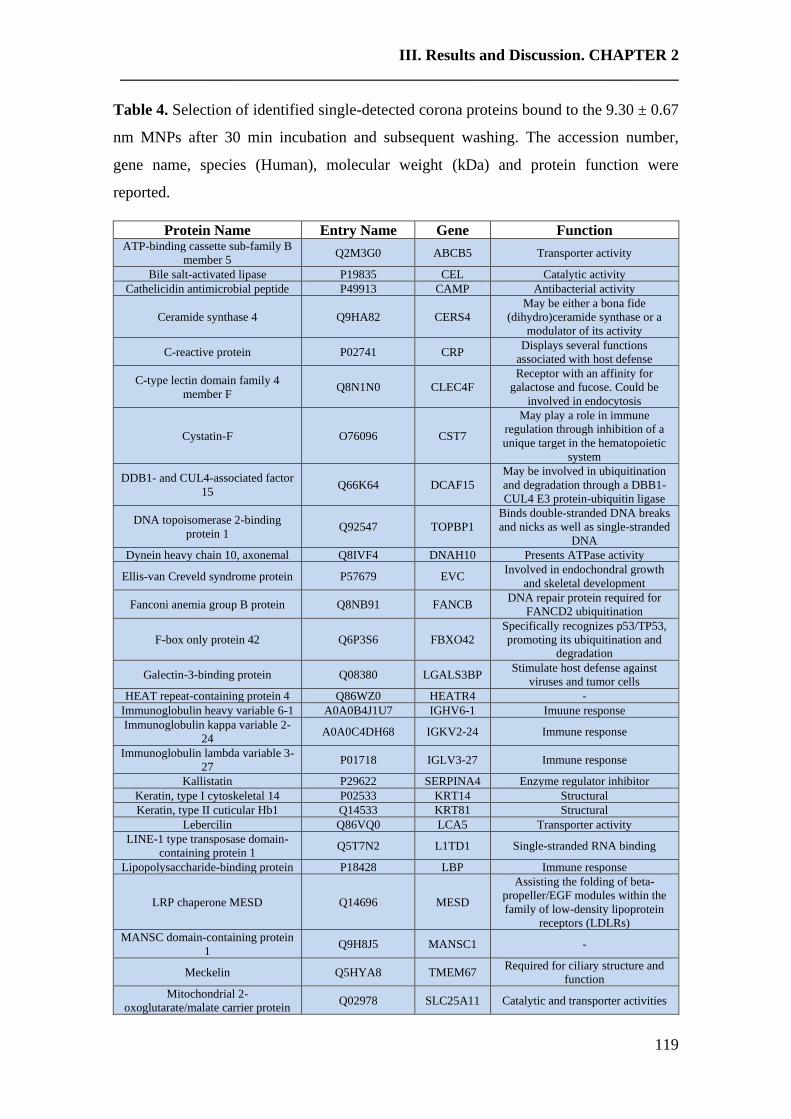
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
119
Table 4. Selection of identified single-detected corona proteins bound to the 9.30 ± 0.67
nm MNPs after 30 min incubation and subsequent washing. The accession number,
gene name, species (Human), molecular weight (kDa) and protein function were
reported.
Protein Name Entry Name Gene Function ATP-binding cassette sub-family B
member 5 Q2M3G0 ABCB5 Transporter activity
Bile salt-activated lipase P19835 CEL Catalytic activity
Cathelicidin antimicrobial peptide P49913 CAMP Antibacterial activity
Ceramide synthase 4 Q9HA82 CERS4
May be either a bona fide
(dihydro)ceramide synthase or a
modulator of its activity
C-reactive protein P02741 CRP Displays several functions
associated with host defense
C-type lectin domain family 4
member F Q8N1N0 CLEC4F
Receptor with an affinity for
galactose and fucose. Could be
involved in endocytosis
Cystatin-F O76096 CST7
May play a role in immune
regulation through inhibition of a
unique target in the hematopoietic
system
DDB1- and CUL4-associated factor
15 Q66K64 DCAF15
May be involved in ubiquitination
and degradation through a DBB1-
CUL4 E3 protein-ubiquitin ligase
DNA topoisomerase 2-binding
protein 1 Q92547 TOPBP1
Binds double-stranded DNA breaks
and nicks as well as single-stranded
DNA
Dynein heavy chain 10, axonemal Q8IVF4 DNAH10 Presents ATPase activity
Ellis-van Creveld syndrome protein P57679 EVC Involved in endochondral growth
and skeletal development
Fanconi anemia group B protein Q8NB91 FANCB DNA repair protein required for
FANCD2 ubiquitination
F-box only protein 42 Q6P3S6 FBXO42
Specifically recognizes p53/TP53,
promoting its ubiquitination and
degradation
Galectin-3-binding protein Q08380 LGALS3BP Stimulate host defense against
viruses and tumor cells
HEAT repeat-containing protein 4 Q86WZ0 HEATR4 -
Immunoglobulin heavy variable 6-1 A0A0B4J1U7 IGHV6-1 Imuune response
Immunoglobulin kappa variable 2-
24 A0A0C4DH68 IGKV2-24 Immune response
Immunoglobulin lambda variable 3-
27 P01718 IGLV3-27 Immune response
Kallistatin P29622 SERPINA4 Enzyme regulator inhibitor
Keratin, type I cytoskeletal 14 P02533 KRT14 Structural
Keratin, type II cuticular Hb1 Q14533 KRT81 Structural
Lebercilin Q86VQ0 LCA5 Transporter activity
LINE-1 type transposase domain-
containing protein 1 Q5T7N2 L1TD1 Single-stranded RNA binding
Lipopolysaccharide-binding protein P18428 LBP Immune response
LRP chaperone MESD Q14696 MESD
Assisting the folding of beta-
propeller/EGF modules within the
family of low-density lipoprotein
receptors (LDLRs)
MANSC domain-containing protein
1 Q9H8J5 MANSC1 -
Meckelin Q5HYA8 TMEM67 Required for ciliary structure and
function
Mitochondrial 2-
oxoglutarate/malate carrier protein Q02978 SLC25A11 Catalytic and transporter activities

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
120
NAD(P)H dehydrogenase [quinone]
1 P15559 NQO1 Catalytic activity
N-acetyllactosaminide beta-1,3-N-
acetylglucosaminyltransferase 2 Q9NY97 B3GNT2 Catalytic activity
Nck-associated protein 1 Q9Y2A7 NCKAP1
As component of the WAVE1
complex, required for BDNF-
NTRK2 endocytic trafficking and
signaling from early endosomes
Nesprin-1 Q8NF91 SYNE1
Forms a linking network between
organelles and the actin
cytoskeleton to maintain the
subcellular spatial organization
Nuclear receptor coactivator 6 Q14686 NCOA6 Involved in the coactivation of the
NF-kappa-B pathway
PAN2-PAN3 deadenylation
complex subunit PAN3 Q58A45 PAN3
Regulatory subunit of the poly(A)-
nuclease (PAN) deadenylation
complex
Pentatricopeptide repeat-containing
protein 1, mitochondrial O75127 PTCD1
Mitochondrial protein implicated in
negative regulation of leucine
tRNA levels, as well as negative
regulation of mitochondria-encoded
proteins and COX activity
Phosphatidylcholine translocator
ABCB4 P21439 ABCB4
Acts as a positive regulator of
biliary lipid secretion
1-phosphatidylinositol 4,5-
bisphosphate phosphodiesterase
gamma-2
P16885 PLCG2 Catalytic activity
Phospholipase D1 Q13393 PLD1 Catalytic activity
Polypeptide N-
acetylgalactosaminyltransferase 3 Q14435 GALNT3 Catalytic activity
Protein salvador homolog 1 Q9H4B6 SAV1
Regulator of STK3/MST2 and
STK4/MST1 in the Hippo signaling
pathway which plays a pivotal role
in organ size control and tumor
suppression by restricting
proliferation and promoting
apoptosis
Protein S100-A7 P31151 S100A7 -
Protein S100-A8 P05109 S100A8
Plays a prominent role in the
regulation of inflammatory
processes and immune response
Protein S100-A9 P06702 S100A9
Plays a prominent role in the
regulation of inflammatory
processes and immune response
Protein Z-dependent protease
inhibitor Q9UK55
SERPINA1
0 Enzyme regulator activity
Protein ZGRF1 Q86YA3 ZGRF1
Zinc ion binding that inhibits
factors Xa and XIa of the
coagulation cascade
Protocadherin-12 Q9NPG4 PCDH12
Cellular adhesion molecule that
plays an important role as a
regulator of cell migration,
probably via increasing cell-cell
adhesion
Roundabout homolog 4 Q8WZ75 ROBO4
Mediates the inhibition of primary
endothelial cell migration by Slit
proteins
Serum amyloid A-2 protein P0DJI9 SAA2 Inflammatory response
Short coiled-coil protein Q9UIL1 SCOC Positive regulator of amino acid
starvation-induced autophagy
Structural maintenance of
chromosomes protein 6 Q96SB8 SMC6 Structural
Supervillin O95425 SVIL Structural
Syncoilin Q9H7C4 SYNC
Plays a supportive role in the
efficient coupling of mechanical
stress between the myofibril and
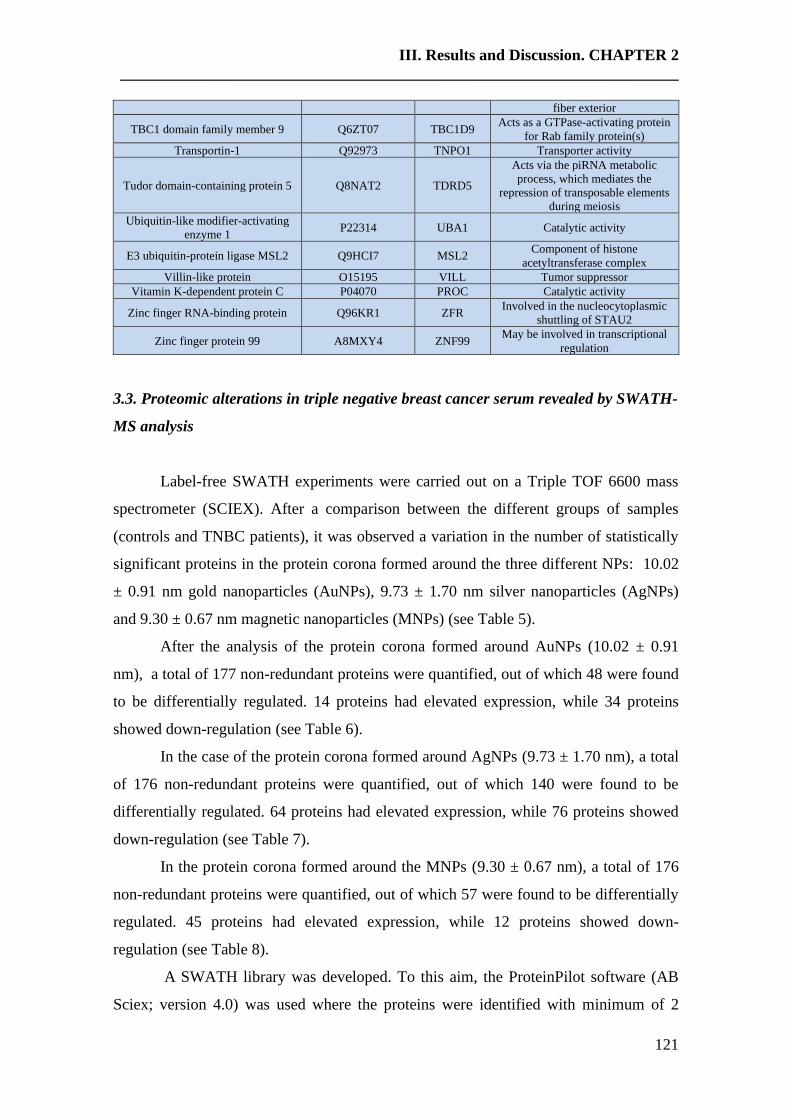
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
121
fiber exterior
TBC1 domain family member 9 Q6ZT07 TBC1D9 Acts as a GTPase-activating protein
for Rab family protein(s)
Transportin-1 Q92973 TNPO1 Transporter activity
Tudor domain-containing protein 5 Q8NAT2 TDRD5
Acts via the piRNA metabolic
process, which mediates the
repression of transposable elements
during meiosis
Ubiquitin-like modifier-activating
enzyme 1 P22314 UBA1 Catalytic activity
E3 ubiquitin-protein ligase MSL2 Q9HCI7 MSL2 Component of histone
acetyltransferase complex
Villin-like protein O15195 VILL Tumor suppressor
Vitamin K-dependent protein C P04070 PROC Catalytic activity
Zinc finger RNA-binding protein Q96KR1 ZFR Involved in the nucleocytoplasmic
shuttling of STAU2
Zinc finger protein 99 A8MXY4 ZNF99 May be involved in transcriptional
regulation
3.3. Proteomic alterations in triple negative breast cancer serum revealed by SWATH-
MS analysis
Label-free SWATH experiments were carried out on a Triple TOF 6600 mass
spectrometer (SCIEX). After a comparison between the different groups of samples
(controls and TNBC patients), it was observed a variation in the number of statistically
significant proteins in the protein corona formed around the three different NPs: 10.02
± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs)
and 9.30 ± 0.67 nm magnetic nanoparticles (MNPs) (see Table 5).
After the analysis of the protein corona formed around AuNPs (10.02 ± 0.91
nm), a total of 177 non-redundant proteins were quantified, out of which 48 were found
to be differentially regulated. 14 proteins had elevated expression, while 34 proteins
showed down-regulation (see Table 6).
In the case of the protein corona formed around AgNPs (9.73 ± 1.70 nm), a total
of 176 non-redundant proteins were quantified, out of which 140 were found to be
differentially regulated. 64 proteins had elevated expression, while 76 proteins showed
down-regulation (see Table 7).
In the protein corona formed around the MNPs (9.30 ± 0.67 nm), a total of 176
non-redundant proteins were quantified, out of which 57 were found to be differentially
regulated. 45 proteins had elevated expression, while 12 proteins showed down-
regulation (see Table 8).
A SWATH library was developed. To this aim, the ProteinPilot software (AB
Sciex; version 4.0) was used where the proteins were identified with minimum of 2
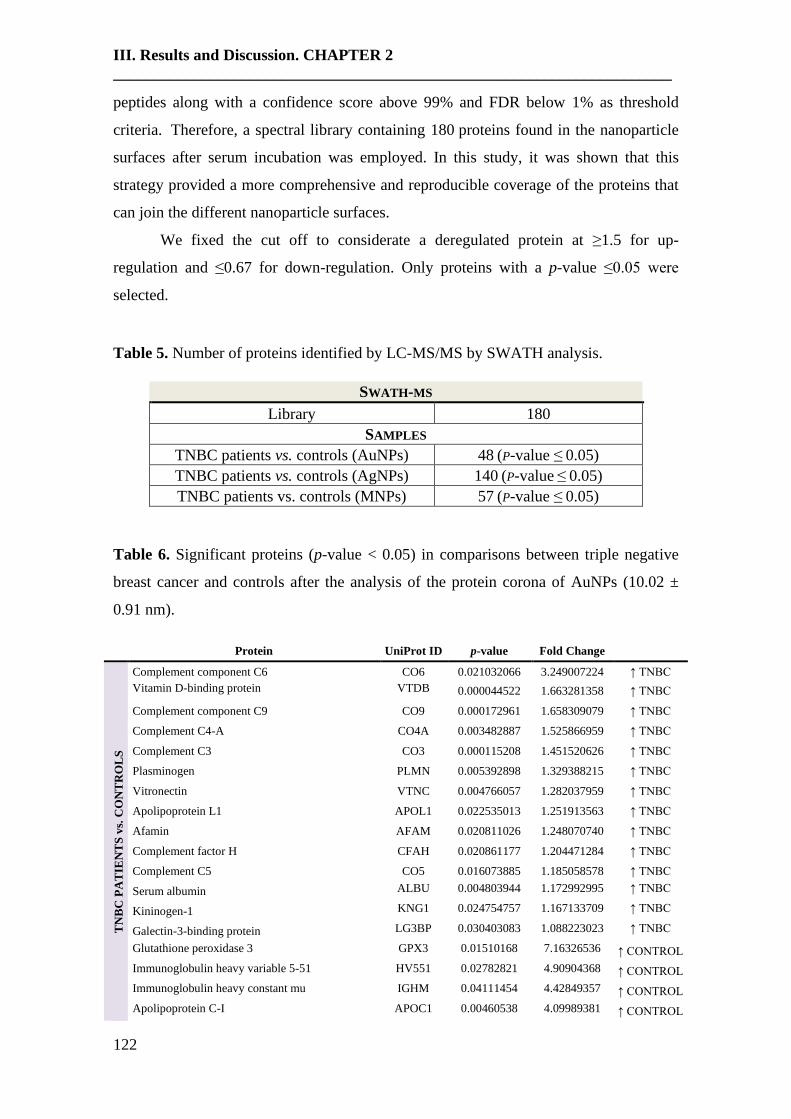
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
122
peptides along with a confidence score above 99% and FDR below 1% as threshold
criteria. Therefore, a spectral library containing 180 proteins found in the nanoparticle
surfaces after serum incubation was employed. In this study, it was shown that this
strategy provided a more comprehensive and reproducible coverage of the proteins that
can join the different nanoparticle surfaces.
We fixed the cut off to considerate a deregulated protein at ≥1.5 for up-
regulation and ≤0.67 for down-regulation. Only proteins with a p-value ≤0.05 were
selected.
Table 5. Number of proteins identified by LC-MS/MS by SWATH analysis.
SWATH-MS
Library 180
SAMPLES
TNBC patients vs. controls (AuNPs) 48 (P-value ≤ 0.05)
TNBC patients vs. controls (AgNPs) 140 (P-value ≤ 0.05)
TNBC patients vs. controls (MNPs) 57 (P-value ≤ 0.05)
Table 6. Significant proteins (p-value < 0.05) in comparisons between triple negative
breast cancer and controls after the analysis of the protein corona of AuNPs (10.02 ±
0.91 nm).
Protein UniProt ID p-value Fold Change
TN
BC
PA
TIE
NT
S v
s. C
ON
TR
OL
S
Complement component C6 CO6 0.021032066 3.249007224 ↑ TNBC
Vitamin D-binding protein VTDB 0.000044522 1.663281358 ↑ TNBC
Complement component C9 CO9 0.000172961 1.658309079 ↑ TNBC
Complement C4-A CO4A 0.003482887 1.525866959 ↑ TNBC
Complement C3 CO3 0.000115208 1.451520626 ↑ TNBC
Plasminogen PLMN 0.005392898 1.329388215 ↑ TNBC
Vitronectin VTNC 0.004766057 1.282037959 ↑ TNBC
Apolipoprotein L1 APOL1 0.022535013 1.251913563 ↑ TNBC
Afamin AFAM 0.020811026 1.248070740 ↑ TNBC
Complement factor H CFAH 0.020861177 1.204471284 ↑ TNBC
Complement C5 CO5 0.016073885 1.185058578 ↑ TNBC
Serum albumin ALBU 0.004803944 1.172992995 ↑ TNBC
Kininogen-1 KNG1 0.024754757 1.167133709 ↑ TNBC
Galectin-3-binding protein LG3BP 0.030403083 1.088223023 ↑ TNBC
Glutathione peroxidase 3 GPX3 0.01510168 7.16326536 ↑ CONTROL
Immunoglobulin heavy variable 5-51 HV551 0.02782821 4.90904368 ↑ CONTROL
Immunoglobulin heavy constant mu IGHM 0.04111454 4.42849357 ↑ CONTROL
Apolipoprotein C-I APOC1 0.00460538 4.09989381 ↑ CONTROL

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
123
Immunoglobulin lambda variable 3-9 LV39 0.0394842 3.35634858 ↑ CONTROL
Apolipoprotein D APOD 0.04050732 3.22034685 ↑ CONTROL
Immunoglobulin heavy constant alpha 1 IGHA1 0.00960489 3.19128417 ↑ CONTROL
Immunoglobulin kappa variable 2-29 KV229 0.00334596 2.48673552 ↑ CONTROL
Immunoglobulin heavy constant alpha 2 IGHA2 0.02785381 2.43044159 ↑ CONTROL
Inter-alpha-trypsin inhibitor heavy chain H4 ITIH4 0.02521756 2.41781135 ↑ CONTROL
Immunoglobulin lambda variable 4-69 LV469 0.00833282 2.37374475 ↑ CONTROL
Apolipoprotein(a) OS=Homo sapiens APOA 0.03681681 2.12362227 ↑ CONTROL
Pigment epithelium-derived factor PEDF 0.00858685 1.94114234 ↑ CONTROL
Alpha-2-macroglobulin A2MG 0.00477503 1.9190963 ↑ CONTROL
Hemoglobin subunit beta HBB 0.00500101 1.90068671 ↑ CONTROL
CD5 antigen-like CD5L 0.0397185 1.7759815 ↑ CONTROL
Serum amyloid A-4 protein SAA4 0.04115058 1.77144695 ↑ CONTROL
Tetranectin TETN 0.01611908 1.7097494 ↑ CONTROL
Apolipoprotein A-II APOA2 0.01467561 1.67488772 ↑ CONTROL
Inter-alpha-trypsin inhibitor heavy chain H2 ITIH2 0.00096553 1.61898708 ↑ CONTROL
Carboxypeptidase B2 CBPB2 0.00728968 1.46619249 ↑ CONTROL
Transthyretin TTHY 0.04987991 1.45277849 ↑ CONTROL
Complement C1r subcomponent-like protein C1RL 0.03887632 1.43784936 ↑ CONTROL
Inter-alpha-trypsin inhibitor heavy chain H1 ITIH1 0.00031251 1.41502216 ↑ CONTROL
Serum paraoxonase/arylesterase 1 PON1 0.00142352 1.40629048 ↑ CONTROL
Apolipoprotein F APOF 0.03956903 1.39106396 ↑ CONTROL
Apolipoprotein M APOM 0.00064927 1.36179665 ↑ CONTROL
Apolipoprotein A-I APOA1 0.02341567 1.34721575 ↑ CONTROL
Serotransferrin TRFE 0.0020036 1.31516423 ↑ CONTROL
Selenoprotein P SEPP1 0.00426885 1.30905254 ↑ CONTROL
Mannan-binding lectin serine protease 1 MASP1 0.03643268 1.29206286 ↑ CONTROL
Carboxypeptidase N subunit 2 CPN2 0.03700623 1.28514087 ↑ CONTROL
Protein AMBP AMBP 0.0084045 1.22749917 ↑ CONTROL
Plasma protease C1 inhibitor IC1 0.04106727 1.2095684 ↑ CONTROL
Table 7. Significant proteins (p-value < 0.05) in comparisons between triple negative
breast cancer and controls after the analysis of the protein corona of AgNPs (9.73 ± 1.70
nm).
Protein UniProt
ID p-value Fold Change
TN
BC
PA
TIE
NT
S v
s.
CO
NT
RO
LS
C-reactive protein CRP 0.00550613 4.84567669 ↑ TNBC
Histidine-rich glycoprotein HRG 3.22E-09 3.93436379 ↑ TNBC
Complement component C9 CO9 6.28E-08 3.56825414 ↑ TNBC
Complement C3 CO3 1.95E-08 3.03698879 ↑ TNBC
Complement factor B CFAB 4.13E-09 2.83840576 ↑ TNBC
Immunoglobulin kappa variable 2-24 KV224 0.00067261 2.77717871 ↑ TNBC
Ficolin-3 FCN3 7.79E-08 2.73769215 ↑ TNBC
Immunoglobulin kappa variable 1-13 KV113 0.00613408 2.71041552 ↑ TNBC
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
124
IgGFc-binding protein FCGBP 0.00010163 2.70272053 ↑ TNBC
Phosphatidylinositol-glycan-specific
phospholipase D PE=1 SV=3 PHLD 0.00305474 2.66489178 ↑ TNBC
Coagulation factor XIII B chain F13B 2.66E-07 2.60635841 ↑ TNBC
Fetuin-B FETUB 1.50E-06 2.55082102 ↑ TNBC
Apolipoprotein L1 APOL1 2.53E-10 2.52932257 ↑ TNBC
Serum albumin ALBU 4.46E-09 2.51568102 ↑ TNBC
Insulin-like growth factor-binding protein
complex acid labile subunit ALS 4.46E-09 2.49955167
↑ TNBC
Inter-alpha-trypsin inhibitor heavy chain H3 ITIH3 7.30E-07 2.38250842 ↑ TNBC
Complement factor I CFAI 1.02E-07 2.3647495 ↑ TNBC
Complement factor H CFAH 4.83E-09 2.35326116 ↑ TNBC
Vitronectin VTNC
1.40E-09 2.31946741 ↑ TNBC
C4b-binding protein alpha chain C4BPA
0.00011973 2.28632913 ↑ TNBC
Hyaluronan-binding protein 2 HABP2 3.29E-07 2.28034134 ↑ TNBC
Plasminogen PLMN 7.79E-05 2.23378207 ↑ TNBC
Keratin type II cytoskeletal 1 K2C1 0.00022394 2.20416961 ↑ TNBC
Kininogen-1 KNG1 1.18E-08 2.19276786 ↑ TNBC
Fibronectin FINC 3.36E-08 2.08858855 ↑ TNBC
Complement C5 CO5 2.42E-08 2.01278029 ↑ TNBC
Complement C1q subcomponent subunit C HUMAN 4.55E-05 2.01080564 ↑ TNBC
Apolipoprotein A-IV APOA4 4.61E-06 2.00364799 ↑ TNBC
Keratin type I cytoskeletal 10 K1C10 0.00259975 1.97893282 ↑ TNBC
Tetranectin TETN 4.94E-05 1.95250374 ↑ TNBC
Clusterin CLUS 1.82E-06 1.9173471 ↑ TNBC
Complement component C8 gamma chain CO8G 6.29E-06 1.89453705 ↑ TNBC
Serotransferrin TRFE 4.33E-05 1.8939175 ↑ TNBC
Complement C4-B CO4B 0.03028632 1.89103397 ↑ TNBC
Complement C1q subcomponent subunit B C1QB 0.00018878 1.88786101 ↑ TNBC
Plasma kallikrein KLKB1 2.85E-06 1.88182482 ↑ TNBC
Galectin-3-binding protein LG3BP 1.76E-09 1.85166575 ↑ TNBC
Alpha-2-HS-glycoprotein FETUA 1.70E-06 1.82851516 ↑ TNBC
Complement C1q subcomponent subunit A C1QA 0.00028866 1.80691958 ↑ TNBC
Prothrombin THRB 0.0030656 1.68707742 ↑ TNBC
Retinol-binding protein 4 RET4 5.98E-06 1.65111784 ↑ TNBC
Carboxypeptidase N catalytic chain CBPN 0.00562324 1.62654965 ↑ TNBC
Complement component C8 beta chain CO8B 0.00298689 1.58131021 ↑ TNBC
Sex hormone-binding globulin SHBG 0.04298875 1.5528487 ↑ TNBC
Serum amyloid P-component SAMP 0.00436056 1.55023657 ↑ TNBC
Zinc-alpha-2-glycoprotein ZA2G 0.00133379 1.54906716 ↑ TNBC
Beta-2-glycoprotein 1 APOH 0.00192644 1.53648273 ↑ TNBC
Vitamin K-dependent protein S PROS 1.47E-06 1.52146296 ↑ TNBC
Complement C4-A CO4A 0.00378789 1.50875703 ↑ TNBC
Mannan-binding lectin serine protease 1 MASP1 0.00057947 1.49286664 ↑ TNBC
Complement C2 CO2 1.04E-05 1.48927332 ↑ TNBC
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
125
Coagulation factor X FA10 0.00022691 1.47652463 ↑ TNBC
Complement C1r subcomponent C1R 0.03355753 1.46634915 ↑ TNBC
Alpha-1-acid glycoprotein 2 A1AG2 0.04347799 1.46608046 ↑ TNBC
Antithrombin-III ANT3 0.00109057 1.45858581 ↑ TNBC
Afamin AFAM 0.02956547 1.42380083 ↑ TNBC
Vitamin D-binding protein VTDB 0.00689951 1.40641874 ↑ TNBC
Hemopexin HEMO 0.00014568 1.38633651 ↑ TNBC
Carboxypeptidase N subunit 2 CPN2 0.03697862 1.36334001 ↑ TNBC
Protein AMBP AMBP 0.0013811 1.32566307 ↑ TNBC
Apolipoprotein E APOE 0.00068645 1.28506343 ↑ TNBC
Attractin ATRN 0.0072892 1.27421822 ↑ TNBC
N-acetylmuramoyl-L-alanine amidase PGRP2 0.00138778 1.23991258 ↑ TNBC
Apolipoprotein M APOM 0.01360078 1.23954254 ↑ TNBC
Immunoglobulin heavy variable 3-15 HV315 4.18E-06 75.3967905 ↑ CONTROL
Immunoglobulin heavy constant gamma 1 IGHG1 1.34E-07 42.3987588 ↑ CONTROL
Alpha-1-antichymotrypsin AACT 6.14E-10 40.3625379 ↑ CONTROL
Immunoglobulin heavy variable 3-73 HV373 6.06E-07 37.7464233 ↑ CONTROL
Immunoglobulin heavy constant gamma 3 IGHG3 3.68E-06 36.0478431 ↑ CONTROL
Ceruloplasmin CERU 2.42E-09 33.0108942 ↑ CONTROL
Immunoglobulin heavy constant gamma 2 IGHG2 1.72E-06 32.3391318 ↑ CONTROL
Alpha-1-antitrypsin A1AT 1.14E-10 28.6542019 ↑ CONTROL
Immunoglobulin heavy constant mu IGHM 7.89E-05 26.3705332 ↑ CONTROL
Corticosteroid-binding globulin CBG 1.19E-09 25.9868345 ↑ CONTROL
Immunoglobulin heavy constant gamma 4 IGHG4 0.00011456 21.9488461 ↑ CONTROL
Cholinesterase CHLE 1.48E-08 21.7678062 ↑ CONTROL
Immunoglobulin kappa variable 4-1 KV401 2.21E-07 20.676772 ↑ CONTROL
Immunoglobulin heavy variable 4-59 HV459 4.18E-08 19.791237 ↑ CONTROL
Immunoglobulin heavy variable 3-30-5 HV335 1.61E-07 18.8319962 ↑ CONTROL
Immunoglobulin lambda variable 1-47 LV147 6.19E-05 18.715722 ↑ CONTROL
Immunoglobulin lambda variable 8-61 LV861 2.74E-08 18.5700233 ↑ CONTROL
Immunoglobulin kappa variable 6-21 KV621 0.0062062 16.6110906 ↑ CONTROL
Immunoglobulin heavy variable 3-49 HV349 2.43E-08 16.3703203 ↑ CONTROL
Immunoglobulin kappa constant IGKC 2.49E-06 15.6449675 ↑ CONTROL
Immunoglobulin lambda constant 3 IGLC3 1.61E-05 15.2226153 ↑ CONTROL
MMS19 nucleotide excision repair protein
homolog
MMS19 0.01743218 15.0005868 ↑ CONTROL
Immunoglobulin kappa variable 3-20 KV320 5.98E-09 14.9565997 ↑ CONTROL
Immunoglobulin kappa variable 3-15 KV315 4.41E-09 13.7845573 ↑ CONTROL
Immunoglobulin lambda-like polypeptide 1 IGLL1 9.67E-08 11.9419924 ↑ CONTROL
Immunoglobulin lambda variable 3-9 LV39 4.22E-07 11.8083139 ↑ CONTROL
Immunoglobulin heavy variable 5-51 HV551 4.24E-07 11.4284904 ↑ CONTROL
Immunoglobulin kappa variable 3-11 KV311 5.08E-09 11.3796551 ↑ CONTROL
Immunoglobulin kappa variable 1-33 KV133 7.64E-07 11.2884931 ↑ CONTROL
Immunoglobulin heavy constant alpha 2 IGHA2 6.08E-05 10.9418774 ↑ CONTROL
Immunoglobulin lambda-like polypeptide 5 V=2 IGLL5 9.57E-05 10.7682004 ↑ CONTROL
Angiotensinogen ANGT 3.42E-11 8.97430276 ↑ CONTROL
Pigment epithelium-derived factor PEDF 8.81E-10 8.87937962 ↑ CONTROL
III. Results and Discussion. CHAPTER 2
______________________________________________________________________
126
Immunoglobulin lambda variable 7-43 LV743 3.62E-06 8.19552518 ↑ CONTROL
Hemoglobin subunit alpha HBA 0.0003338 7.79643124 ↑ CONTROL
Immunoglobulin heavy constant alpha 1 IGHA1 1.48E-07 7.66824876 ↑ CONTROL
Immunoglobulin lambda variable 3-25 LV325 1.49E-06 7.44370609 ↑ CONTROL
Immunoglobulin heavy variable 4-28 HV428 1.38E-06 7.26533223 ↑ CONTROL
Immunoglobulin kappa variable 1D-12 KVD12 1.17E-09 7.18559345 ↑ CONTROL
Heparin cofactor 2 HEP2 2.44E-07 7.13401112 ↑ CONTROL
Inter-alpha-trypsin inhibitor heavy chain H4 ITIH4 3.63E-08 6.70198308 ↑ CONTROL
Immunoglobulin lambda variable 9-49 LV949 0.00017391 6.43567288 ↑ CONTROL
Alpha-1B-glycoprotein A1BG 1.70E-09 6.00033312 ↑ CONTROL
Apolipoprotein A-I APOA1 1.65E-09 5.16689397 ↑ CONTROL
Immunoglobulin lambda variable 3-19 LV319 1.92E-08 5.05592895 ↑ CONTROL
Immunoglobulin kappa variable 2-29 KV229 2.59E-07 5.02231162 ↑ CONTROL
Immunoglobulin heavy variable 6-1 HV601 2.19E-05 4.85408469 ↑ CONTROL
Immunoglobulin heavy variable 1-46 HV146 9.86E-06 4.84652628 ↑ CONTROL
Immunoglobulin heavy variable 3-23 HV323 1.25E-05 4.75302522 ↑ CONTROL
Cholesteryl ester transfer protein CETP 1.22E-05 4.64058921 ↑ CONTROL
Apolipoprotein A-II APOA2 2.16E-08 4.62885891 ↑ CONTROL
Thyroxine-binding globulin THBG 1.80E-08 4.45994071 ↑ CONTROL
Protein Z-dependent protease inhibitor ZPI 4.44E-05 4.43951224 ↑ CONTROL
Serum amyloid A-4 protein SAA4 1.38E-07 4.39608833 ↑ CONTROL
Immunoglobulin heavy variable 1-69 HV169 0.00058138 4.33088897 ↑ CONTROL
Immunoglobulin kappa variable 1-9 KV109 3.99E-06 4.20727924 ↑ CONTROL
Gelsolin GELS 7.03E-09 4.09546999 ↑ CONTROL
CD44 antigen CD44 0.00233936 4.0837648 ↑ CONTROL
Apolipoprotein C-I APOC1 0.00038806 3.66742486 ↑ CONTROL
Alpha-2-macroglobulin A2MG 3.58E-09 3.65331919 ↑ CONTROL
Apolipoprotein D APOD 3.95E-08 3.53083248 ↑ CONTROL
Kallistatin KAIN 4.78E-10 3.48621242 ↑ CONTROL
Immunoglobulin kappa variable 1-5 KV105 8.42E-05 3.22054945 ↑ CONTROL
Leucine-rich alpha-2-glycoprotein A2GL 1.29E-08 3.00855446 ↑ CONTROL
Hemoglobin subunit beta HBB 8.64E-05 2.890312 ↑ CONTROL
Coiled-coil domain-containing protein 8 CCDC8 6.16E-07 2.88193259 ↑ CONTROL
Immunoglobulin kappa variable 2D-28 KVD28 0.00104876 2.6282284 ↑ CONTROL
Immunoglobulin kappa variable 6D-21 KVD21 0.01695916 2.61227937 ↑ CONTROL
Immunoglobulin lambda variable 4-69 LV469 8.14E-05 2.35130225 ↑ CONTROL
Biotinidase BTD 0.00622085 2.19095666 ↑ CONTROL
Plasma serine protease inhibitor IPSP 3.04E-05 2.1835559 ↑ CONTROL
Carboxypeptidase B2 CBPB2 8.55E-05 1.8115321 ↑ CONTROL
Immunoglobulin heavy variable 2-26 HV226 0.00977698 1.69947422 ↑ CONTROL
Apolipoprotein B-100 APOB 2.35E-05 1.66025805 ↑ CONTROL
Alpha-2-antiplasmin A2AP 0.00014843 1.60147275 ↑ CONTROL
Apolipoprotein C-III APOC3 0.02144075 1.39259865 ↑ CONTROL

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
127
Table 8. Significant proteins (p-value < 0.05) in comparisons between triple negative
breast cancer and controls after the analysis of the protein corona of MNPs (9.30 ± 0.67
nm).
Protein UniProt ID p-value Fold Change
TN
BC
PA
TIE
NT
S v
s. C
ON
TR
OL
C-reactive protein CRP 0.00169149 3.40849571 ↑ TNBC
Lipopolysaccharide-binding protein LBP 0.00837847 2.93851762 ↑ TNBC
Immunoglobulin kappa variable 2-24 KV224 0.00583062 2.27733934 ↑ TNBC
Serum amyloid P-component SAMP 0.00010388 2.20872021 ↑ TNBC
Keratin. type II cytoskeletal 2 epidermal K22E 0.01682722 2.10615702 ↑ TNBC
Complement C4-B CO4B 0.04180946 1.9800469 ↑ TNBC
Complement component C9 CO9 5.75E-06 1.90848624 ↑ TNBC
IgGFc-binding protein FCGBP 0.01481938 1.86546782 ↑ TNBC
Protein Z-dependent protease inhibitor ZPI 0.00252121 1.78800298 ↑ TNBC
Complement C1q subcomponent subunit B C1QB 0.00199072 1.78457812 ↑ TNBC
Phosphatidylinositol-glycan-specific
phospholipase D
PHLD 0.00435099 1.73844701 ↑ TNBC
Prothrombin THRB 0.00750791 1.72802617 ↑ TNBC
Complement C4-A CO4A 0.00510952 1.67059345 ↑ TNBC
Alpha-1-acid glycoprotein 1 A1AG1 0.02552784 1.64048407 ↑ TNBC
Apolipoprotein C-III APOC3 0.03298589 1.61371634 ↑ TNBC
Keratin. type II cytoskeletal 1 K2C1 0.01154011 1.60905471 ↑ TNBC
Sex hormone-binding globulin SHBG 0.02523265 1.59168855 ↑ TNBC
Inter-alpha-trypsin inhibitor heavy chain H3 ITIH3 0.00066933 1.55061894 ↑ TNBC
Complement C3 CO3 6.41E-06 1.53090904 ↑ TNBC
Keratin. type I cytoskeletal 10 K1C10 0.0384444 1.52567319 ↑ TNBC
C4b-binding protein alpha chain C4BPA 0.00027605 1.51193215 ↑ TNBC
Complement factor H CFAH 2.79E-05 1.46546396 ↑ TNBC
Plasma serine protease inhibitor IPSP 0.00973255 1.4623027 ↑ TNBC
Histidine-rich glycoprotein HRG 0.01164428 1.4616738 ↑ TNBC
Complement factor B CFAB 0.00061903 1.45925143 ↑ TNBC
Pregnancy zone protein PZP 0.00155229 1.44791482 ↑ TNBC
Complement C1q subcomponent subunit A C1QA 0.02763702 1.4062295 ↑ TNBC
Coagulation factor V FA5 0.01312931 1.40131004 ↑ TNBC
Complement factor I CFAI 7.52E-05 1.40080904 ↑ TNBC
Complement C2 CO2 0.00061386 1.33762187 ↑ TNBC
Hemopexin HEMO 3.66E-05 1.29733733 ↑ TNBC
Coagulation factor XIII B chain F13B 0.01256843 1.28491873 ↑ TNBC
Complement C1q subcomponent subunit C C1QC 0.03780247 1.26491165 ↑ TNBC
Kininogen-1 KNG1 0.00429848 1.26291248 ↑ TNBC
Beta-2-glycoprotein 1 APOH 0.04746608 1.24963502 ↑ TNBC
Apolipoprotein L1 APOL1 0.00237944 1.23613552 ↑ TNBC
Antithrombin-III ANT3 0.01869897 1.22888805 ↑ TNBC
Protein AMBP AMBP 0.01434176 1.22857819 ↑ TNBC

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
128
Vitronectin VTNC 0.01666796 1.21690616 ↑ TNBC
Inter-alpha-trypsin inhibitor heavy chain H1 ITIH1 0.01847152 1.20880937 ↑ TNBC
Insulin-like growth factor-binding protein
complex acid labile subunit
ALS 0.03555036 1.20445695 ↑ TNBC
Hyaluronan-binding protein 2 HABP2 0.01352977 1.18411135 ↑ TNBC
Clusterin CLUS 0.04346683 1.1756767 ↑ TNBC
Alpha-2-HS-glycoprotein FETUA 0.02891723 1.16730605 ↑ TNBC
Galectin-3-binding protein LG3BP 0.00292722 1.116735 ↑ TNBC
Apolipoprotein C-I APOC1 0.02222358 2.50213212 ↑ CONTROL
Immunoglobulin heavy constant mu IGHM 0.00022527 2.032717896 ↑ CONTROL
Immunoglobulin lambda variable 8-61 LV861 0.00063688 2.023228572 ↑ CONTROL
Immunoglobulin lambda variable 3-19 LV319 0.0082843 1.73294072 ↑ CONTROL
CD5 antigen-like CD5L 0.02066785 1.731935965 ↑ CONTROL
Immunoglobulin lambda variable 3-9 LV39 0.02497423 1.576244295 ↑ CONTROL
Apolipoprotein F APOF 0.02131359 1.556435198 ↑ CONTROL
Immunoglobulin kappa variable 6-21 KV621 0.00608326 1.473240834 ↑ CONTROL
Immunoglobulin heavy variable 3-23 HV323 0.02960462 1.38352045 ↑ CONTROL
Immunoglobulin heavy variable 4-28 HV428 0.02327515 1.304707447 ↑ CONTROL
Plasma kallikrein KLKB1 0.04204343 1.23839695 ↑ CONTROL
Apolipoprotein M APOM 0.01063485 1.232467766 ↑ CONTROL
In the analysis of the protein corona formed around the three nanoparticles
(AuNPs, AgNPs, and MNPs), eight common proteins showed to be statistically
significant and appeared quantitatively increased (up-regulated) in triple negative breast
cancer patients versus controls (healthy people) (Figure 5). These proteins are
complement component C9 (CO9), complement C4-A (CO4A), complement C3 (CO3),
vitronectin (VTNC), apolipoprotein L1 (APOL1), complement factor H (CFAH),
kininogen-1 (KNG1), galectin-3-binding protein (LG3BP). However, three common
proteins appeared quantitatively decreased (down-regulated) in triple negative breast
cancer patients versus controls (healthy people) (Figure 6). These proteins are
immunoglobulin heavy constant mu (IGHM), immunoglobulin lambda variable 3-9
(LV39) and apolipoprotein C-I (APOC1).

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
129
Figure 5. Quantitative Venn diagrams showing the number of up-regulated proteins
found in the protein corona of 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70
nm silver nanoparticles (AgNPs) and 9.30 ± 0.67 nm magnetic nanoparticles (MNPs)
after their incubation with serum from eight triple negative breast cancer patients and
eight healthy controls.
Figure 6. Quantitative Venn diagrams showing the number of down-regulated proteins
found in the protein corona of 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70
nm silver nanoparticles (AgNPs) and 9.30 ± 0.67 nm magnetic nanoparticles (MNPs)
after their incubation with serum from eight triple negative breast cancer patients and
eight healthy controls.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
130
After the analysis of the protein corona formed around AuNPs (10.02 ± 0.91
nm), AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm), the principal components
analysis (PCA) clearly revealed that the samples of the triple negative breast cancer
patients and healthy people were separated in the PC1 axis, which explains 47.7, 80.9
and 79.6 % of the variance between the samples, respectively (Figure 13_SM to Figure
15_SM).
In all cases (AuNPs, AgNPs, MNPs), the separation between the groups of
samples is visible between the group of healthy people and the group of triple negative
breast cancer.
3.4. TNBC biomarker validation
Mass spectrometry-based validation assays were performed in a different cohort
of total serum patients samples (n=8) and controls (n=8) which were run in triplicate
using a TripleTOF® 6600 LC-MS/MS system (Sciex). A correct protein validation was
performed. To this aim, the library was improved after the addition of DDA data
acquired from total serum pools (controls and patient), obtaining a total of 205
identified proteins. After comparing the results obtained by this analysis, with previous
analysis performed on serum samples after the incubation with the different
nanoparticles, fascinating results were observed.
Graphically, these variations can be observed through charts such as the volcano
plot. Volcano plots (see Figure 7) of the global quantification of proteins between
healthy and triple negative breast cancer patients with A: AuNPs (10.02 ± 0.91 nm), B:
AgNPs (9.73 ± 1.70 nm) and C: MNPs (9.30 ± 0.67 nm) were generated by plotting the
log 2-fold changes for the identified proteins against their corresponding adjusted p-
value.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
131
-8 -6 -4 -2 0 2 4 6 8
0
5
1 0
1 5
p a t ie n ts v s h e a lth y
lo g 2 ( fo ld c h a n g e )
-Lo
g 1
0 (
P v
alu
e)
P = 0 .0 5
C F AH
KN G 1VT N C
APO L 1
C O 3
L G 3 BP
AL S
C O 5
IG H M
L V3 9
APO C 1
Figure 7. Volcano plots of the SWATH analysis of proteins between healthy and triple
negative breast cancer patients with AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm)
and MNPs (9.30 ± 0.67 nm) . X-axis shows log(2)-fold change and Y-axis the statistical
significance through -log(10)-pvalue. The dot lines represent the cut off (p value ≤0.05).
Protein biomarkers are those that were common and statistical significative in all the
nanoparticles.
Validation assays verified and confirmed that transthyretin (TTHY) was
statistically significant and appeared quantitatively decreased (down-regulated) in triple
negative breast cancer patients when compared with controls (healthy people). This
result was observed after the analysis of the protein corona formed around AuNPs
(10.02 ± 0.91 nm). A similar observation was recently reported, where it was found that
transthyretin (TTHY) was predominantly (68.75%) down-regulated (n = 33/48) in the
sera of breast cancer patients [62].
Furthermore, validation assays verified and confirmed that complement
component C8 gamma chain (CO8G), ficolin-3 (FCN3), retinol-binding protein 4
(RET4), fibronectin (FINC), fetuin-B (FETUB) and apolipoprotein A-IV (APOA4),
were up-regulated in triple negative breast cancer when compared with controls.
However, apolipoprotein C-III (APOC3), immunoglobulin kappa variable 2D-28
(KVD28), immunoglobulin kappa variable 1-5 (KV105), immunoglobulin kappa
variable 4-1 (KV401), immunoglobulin kappa variable 1D-12 (KVD12),

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
132
immunoglobulin heavy variable 1-46 (HV146), immunoglobulin heavy variable 3-30-5
(HV335), immunoglobulin heavy constant gamma 2 (IGHG2), immunoglobulin heavy
constant gamma 3 (IGHG3), and immunoglobulin heavy constant gamma 4 (IGHG4)
were down-regulated in triple negative breast cancer when compared with controls.
These results were observed after the analysis of the protein corona formed around
AgNPs (9.73 ± 1.70 nm).
Recently, similar studies found that elevated serum levels of retinol binding
protein 4 (RBP4) [63] and fibronectin (FINC) [64] were associated with breast cancer
risk and they could be useful markers for predicting poor prognosis in breast cancer
patients.
In the particular case of apolipoprotein A-IV (APOA4), comparative proteomic
profiling of immunodepleted plasma of healthy and of BC individuals revealed that this
protein was also up-regulated in the plasma of the BC individuals. Furthermore, this
protein was found to be involved in the pathogenesis of cancer and played an important
role in the regulation of metastasis [65].
Validation assays verified and confirmed that pregnancy zone protein (PZP),
coagulation factor V (FA5), protein Z-dependent protease inhibitor (ZPI), alpha-1-acid
glycoprotein 1 (A1AG1) and inter-alpha-trypsin inhibitor heavy chain H1 (ITIH1), were
statistically significant and appeared quantitatively up-regulated in triple negative breast
cancer patients when compared with controls (healthy people). This result was observed
after the analysis of the protein corona formed around MNPs (9.30 ± 0.67 nm).
Mainly, the pregnancy zone protein (PZP) was found to be implicated the
pathogenesis of breast cancer [66].
It is well known that cancer is associated with hypercoagulability, and
circumstantial evidence suggests that tumor-expressed coagulation factors actively
support cancer pathogenesis and progression. Notably, single nucleotide polymorphisms
(SNPs) in F5, encoding coagulation factor V (FA5), have been found associated with
breast cancer. In 2018, M. Tinholt et al. [67] found that FA5 expression was higher in
breast tumors compared to normal tissue. Importantly, FA5 expression was significantly
increased in patients with tumors of aggressive nature (hormone receptor negative-,
triple negative-, HER2 enriched-, and basal-like tumors). These authors also suggested
that tumor-expressed FA5 could be a possible marker of aggressive breast cancer, and it
could emerge as a promising tumor suppressor candidate.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
133
Finally, protein Z-dependent protease inhibitor (ZPI) were identified by
immunochemistry in breast cancer cells, whereas they were absent from normal breast
tissue [68], and alpha-1-acid glycoprotein (AGP) was also found to be a potential
biomarker for breast cancer in 'at risk' individuals, particularly, TNBC patients [69].
3.5. Discussion
In this study, we provided by the first time the application of a combined
proteomic approach as DDA and SWATH-MS to develop the characterization and
quantification of triple negative breast cancer proteins after bio-corona nanoparticle
protein pre-concentration.
Gold nanoparticles (AuNPs: 10.02 ± 0.91 nm), silver nanoparticles (AgNPs:
9.73 ± 1.70 nm) and magnetic nanoparticles (MNPs: (9.30 ± 0.67 nm) were assessed in
biomarker discovery as a tool for the pre-concentration and separation of proteins from
complex proteomes. To this end, sera from eight healthy individuals were compared
with sera from eight patients diagnosed with triple negative breast cancer. The
application of these nanomaterials, combined with mass spectrometry, has allowed the
identification of seven potential biomarkers for the diagnostic and control of TNBC
progression: GRF-type zinc finger domain-containing protein 1 (protein ZGRF1),
Matrix metalloproteinase-9 (MMP9), Lebercilin and Immunoglobulin lambda variable
3-27 (LV327) and LINE-1 type transposase domain-containing protein 1 (LITD1),
structural maintenance of chromosomes protein 6 (SMC6) and short coiled-coil protein
(SCOC).
After performing over these samples, a SWATH analysis to quantify the protein
changes, the separation between the group of healthy people and the group of triple
negative breast cancer patients was observed. Moreover, a lot of deregulated proteins
among both groups were observed. However, these proteins are not among the altered
proteins find by the qualitative DDA assay. The proteomic methods used in this study
are complementary and allow improving characterization studies. The fact that they are
complementary and not necessarily identify the same proteins is due to the search
methods DDA (qualitative), and IDA (SWATH-quantitative) analysis is different.
So, in the analysis of the protein corona formed around the three nanoparticles
(AuNPs, AgNPs, and MNPs), eight common proteins showed to be statistically
significant and appeared quantitatively increased (up-regulated) in triple negative breast

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
134
cancer patients versus controls (healthy people). These proteins are complement
component C9 (CO9), complement C4-A (CO4A), complement C3 (CO3), vitronectin
(VTNC), apolipoprotein L1 (APOL1), complement factor H (CFAH), kininogen-1
(KNG1), galectin-3-binding protein (LG3BP). And three common proteins appeared
quantitatively decreased (down-regulated) in triple negative breast cancer patients
versus controls (healthy people). These proteins are immunoglobulin heavy constant mu
(IGHM), immunoglobulin lambda variable 3-9 (LV39), and apolipoprotein C-I
(APOC1).
Moreover, a lot of deregulated proteins not common to all samples incubated
with the different nanoparticles were found. Therefore, protein corona formed around
AuNPs in breast cancer patients showed 14 proteins up-regulated and 34 proteins down-
regulated, in comparison with the protein corona formed around AuNPs in healthy
people. In the case of the protein corona formed around AgNPs, 64 proteins were found
to be up-regulated, and 76 proteins down-regulated. Finally, in the protein corona
formed around the MNPs, 45 proteins had elevated expression, while 12 proteins
showed down-regulation. From this point of view, AgNPs seem to be more suitable for
a clinical translational purpose, because the analysis of the protein corona formed
around these systems allows better differentiation between both groups of study: healthy
and diseased individuals.
All these proteins can be considered potential triple negative breast cancer
biomarker candidates; however, these proteins were found when serum samples were
concentrated using a bio-corona nanoparticle. Thus, it was thought that the best
validation could be finding these proteins in total serum samples. To this aim, it was
performed a new SWATH-MS analysis improving the library, and it allowed us to find
several validated proteins.
When it was compared total serum SWATH with concentrated proteins in
AgNPs, we found again complement component C8 (CO8G), ficolin-3 (FCN3), retinol-
binding protein 4 (RET4), fibronectin (FINC), fetuin-B (FETUB) and apolipoprotein
A-IV (APOA4), up-regulated; and immunoglobulin heavy constant gamma 4 (IGHG4),
immunoglobulin kappa variable 2D-28 (KVD28), immunoglobulin kappa variable 1-5
(KV105), apolipoprotein C-III (APOC3), immunoglobulin heavy variable 1-46
(HV146), immunoglobulin heavy constant gamma 2 (IGHG2), immunoglobulin heavy
constant gamma 3 (IGHG3), immunoglobulin kappa variable 4-1 (KV401),

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
135
immunoglobulin kappa variable 1D-12 (KVD12) and immunoglobulin heavy variable
3-30-5 (HV335), down-regulated. It is consistent with data from previous studies, and
these proteins can be considered validated. In the comparison between total serum
SWATH and MNPs only found 5 up-regulated proteins pregnancy zone protein (PZP),
coagulation factor V (FA5), protein Z-dependent protease inhibitor (ZPI), alpha-1-acid
glycoprotein 1 (A1AG1) and inter-alpha-trypsin inhibitor heavy chain H1 (ITIH1). And
finally, in the comparison between total serum SWATH and AuNPs, only one down-
regulated protein was validated: transthyretin (TTHY).
4. Conclusions
This study shows that serum proteomics is a valuable tool that can facilitate
comprehensive and systematic identification of the serum proteome under both healthy
and disease conditions. Thus, serum proteomics could be used for disease diagnosis and
prognosis. In our case, we found several breast cancer-specific markers that can be used
in the diagnosis. However, because we found a lot of differentiated proteins, it is
necessary complementary assays to reduce the number of protein biomarkers.
5. References
[1] A. Jemal, F. Bray, M.M. Center, J. Ferlay, E. Ward, D. Forman, Global cancer
statistics, CA Cancer J. Clin. 61(2) (2011) 69-90.
[2] M. Arnedos, C. Bihan, S. Delaloge, F. Andre, Triple-negative breast cancer: are we
making headway at least?, Ther Adv Med Oncol. 4(4) (2012) 195-210.
[3] F.C. Geyer, F. Pareja, B. Weigelt, E. Rakha, I.O. Ellis, S.J. Schnitt, J.S. Reis-Filho,
The spectrum of triple-negative breast disease, high- and low-grade lesions, Am J
Pathol. 187(10) (2017) 2139-2151.
[4] S. Cleator, W. Heller, R.C. Coombes, Triple-negative breast cancer: therapeutic
options, Lancet Oncol. 8(3) (2007) 235-244.
[5] C. Núñez, Blood-based protein biomarkers in breast cancer, Clin Chim Acta 490
(2019) 113-127.
[6] Y. He, A. Mohamedali, C. Huang, M.S. Baker, E.C. Nice, Oncoproteomics: Current
status and future opportunities, Clin Chim Acta 495 (2019) 611-624.
[7] M. Wilhelm, J. Schlegl, H. Hahne, A. Moghaddas Gholami, M. Lieberenz, M.M.
Savitski, E. Ziegler, L. Butzmann, S. Gessulat, H. Marx, T. Mathieson, S. Lemeer, K.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
136
Schnatbaum, U. Reimer, H. Wenschuh, M. Mollenhauer, J. Slotta-Huspenina, J.-H.
Boese, M. Bantscheff, A. Gerstmair, F. Faerber, B. Kuster, Mass spectrometry-based
draft of the human proteome, Nature 509 (2014) 582-587.
[8] S. Miah, C.A.S. Banks, M.K. Adams, L. Florens, K.E. Lukong, M.P. Washburn,
Advancement of mass spectrometry-based proteomics technologies to explore triple
negative breast cancer, Mol Biosyst. 13(1) (2016) 42-55.
[9] D.M. Schulz, C. Böllner, G. Thomas, M. Atkinson, I. Esposito, H. Höfler, M.
Aubele, Identification of differentially expressed proteins in triple-negative breast
carcinomas using DIGE and mass spectrometry, J Proteome Res. 8(7) (2009) 3430-
3438.
[10] C. Ludwig, L. Gillet, G. Rosenberger, S. Amon, B.C. Collins, R. Aebersold, Data‐
independent acquisition‐based SWATH‐MS for quantitative proteomics: a tutorial, Mol
Syst Biol. 14(8) (2018) e8126.
[11] J.G. Meyer, B. Schilling, Clinical applications of quantitative proteomics using
targeted and untargeted data-independent acquisition techniques, Expert Rev
Proteomics. 14(5) (2017) 419-429.
[12] M. Narasimhan, S. Kannan, A. Chawade, A. Bhattacharjee, R. Govekar, Clinical
biomarker discovery by SWATH-MS based label-free quantitative proteomics: impact
of criteria for identification of differentiators and data normalization method, J Transl
Med. 17(1) (2019) 184.
[13] N. Rifai, M.A. Gillette, S.A. Carr, Protein biomarker discovery and validation: the
long and uncertain path to clinical utility, Nat. Biotechnol. 24(8) (2006) 971-983.
[14] R. Millioni, S. Tolin, L. Puricelli, S. Sbrignadello, G.P. Fadini, P. Tessari, G.
Arrigoni, High abundance proteins depletion vs low abundance proteins enrichment:
comparison of methods to reduce the plasma proteome complexity, PLoS One. 6(5)
(2011) e19603.
[15] E. Bellei, S. Bergamini, E. Monari, L.I. Fantoni, A. Cuoghi, T. Ozben, A. Tomasi,
High-abundance proteins depletion for serum proteomic analysis: concomitant removal
of non-targeted proteins, Amino Acids 40(1) (2011) 145-156.
[16] S. Chutipongtanate, S. Chatchen, J. Svasti, Plasma prefractionation methods for
proteomic analysis and perspectives in clinical applications, Proteomics Clin
Appl. (2017) 11(7-8).

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
137
[17] C.P. Albuquerque, M.B. Smolka, S.H. Payne, V. Bafna, J. Eng, H. Zhou, A
multidimensional chromatography technology for in-depth phosphoproteome analysis,
Mol. Cell Proteomics 7(7) (2008) 1389-1396.
[18] D.-G. Liu, L. Sun, Direct isolation of specific RNA-interacting proteins using a
novel affinity médium, Nucleic Acids Res. 33(15) (2005) e132.
[19] P.D. Rakowska, M.G. Ryadnov, Nano-enabled biomarker discovery and detection,
Biomark Med. 5(3) (2011) 387-96.
[20] N. Phogat, M. Kohl, I. Uddin, A. Jahan, Chapter 11 - Interaction of nanoparticles
with biomolecules, protein, enzymes, and its applications, Precision Medicine, Tools
and Quantitative Approaches, Academic Press, Editors: Deigner, Hans-Peter and Kohl,
Matthias (2018) 253-276.
[21] S. Tenzer, D. Docter, S. Rosfa, A. Wlodarski, J. Kuharev, A. Rekik, S.K. Knauer,
C. Bantz, T. Nawroth, C. Bier, J. Sirirattanapan, W. Mann, L. Treuel, R. Zellner, M.
Maskos, H. Schild, R.H. Stauber, Nanoparticle size is a critical physicochemical
determinant of the human blood plasma corona: a comprehensive quantitative proteomic
analysis, ACS Nano 5 (2011) 7155-7167.
[22] M. Lundqvist, J. Stigler, G. Elia, I. Lynch, T. Cedervall, K.A. Dawson,
Nanoparticle size and surface properties determine the protein corona with possible
implications for biological impacts, Proc. Natl. Acad. Sci. U.S.A. 105 (2008) 14265-
14270.
[23] M.P. Monopoli, D. Walczyk, A. Campbell, G. Elia, I. Lynch, F.B. Bombelli, K.A.
Dawson, Physical-chemical aspects of protein corona: relevance to in vitro and in vivo
biological impacts of nanoparticles, J. Am. Chem. Soc. 133 (2011) 2525-2534.
[24] S. Laurent, C. Burtea, C. Thirifays, F. Rezaee, M. Mahmoudi, Significance of cell
“observer” and protein source in nanobiosciences, J. Colloid Interface Sci. 392 (2013)
431-445.
[25] D. Pozzi, G. Caracciolo, L. Digiacomo, V. Colapicchioni, S. Palchetti, A. L.
Capriotti, C. Cavaliere, R. Zenezini Chiozzi, A. Puglisi, A. Laganà, The biomolecular
corona of nanoparticles in circulating biological media, Nanoscale 7 (2015) 13958-
13966.
[26] M. Mahmoudi, A.M. Abdelmonem, S. Behzadi, J.H. Clement, S. Dutz, M.R.
Ejtehadi, R. Hartmann, K. Kantner, U. Linne, P. Maffre, S. Metzler, M.K. Moghadam,
C. Pfeiffer, M. Rezaei, P. Ruiz-Lozano, V. Serpooshan, M.A. Shokrgozar, G.U.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
138
Nienhaus, W.J. Parak, Temperature: the “ignored” factor at the NanoBio interface, ACS
Nano 7 (2013) 6555-6562.
[27] S. Tenzer, D. Docter, J. Kuharev, A. Musyanovych, V. Fetz, R. Hecht, F. Schlenk,
D. Fischer, K. Kiouptsi, C. Reinhardt, K. Landfester, H. Schild, M. Maskos, S.K.
Knauer, R.H. Stauber, Rapid formation of plasma protein corona critically affects
nanoparticle pathophysiology, Nat. Nanotechnol. 8 (2013) 772-781.
[28] T. Zheng, N. Pierre-Pierre, X. Yan, Q. Huo, A.J. Almodovar, F. Valerio, I. Rivera-
Ramirez, E. Griffith, D.D. Decker, S. Chen, Gold nanoparticle-enabled blood test for
early stage cancer detection and risk assessment, ACS Appl. Mater. Interfaces 7(12)
(2015) 6819-6827.
[29] C. Corbo, R. Molinaro, M. Tabatabaei, O.C. Farokhzad, M. Mahmoudi,
Personalized protein corona on nanoparticles and its clinical implications, Biomater Sci.
5(3) (2017) 378-387.
[30] R. Lopez-Cortés, E. Oliveira, C. Núñez, C. Lodeiro, M. Páez de la Cadena, F.
Fdez-Riverola, H. López-Fernández, M. Reboiro-Jato, D. Glez-Peña, J.L. Capelo, H.M.
Santos, Fast human serum profiling through chemical depletion coupled to gold-
nanoparticle-assisted protein separation, Talanta 100 (2012) 239-245.
[31] N.G. Bastus, F. Merkoci, J. Piella, V. Puntes, Synthesis of highly monodisperse
citrate-stabilized silver nanoparticles of up to 200 nm: kinetic control and catalytic
properties, Chem. Mater. 26(9) (2014) 2836-2846.
[32] A.H. Lu, E.L. Salabas, F. Schüth, Magnetic nanoparticles: Synthesis, protection,
functionalization, and application, Angew. Chemie-Int. Ed. 46 (2007) 1222-1244.
[33] S.E. Warder, L.A. Tucker, T.J. Strelitzer, E.M. McKeegan, J.L. Meuth, P.M. Jung,
A. Saraf, B. Singh, J. Lai-Zhang, G Gagne, J.C. Rogers, Reducing agent-mediated
precipitation of high-abundance plasma proteins, Anal. Biochem. 387(2) (2009) 184-
193.
[34] C. Fernández, H.M. Santos, C. Ruíz-Romero, F.J. Blanco, J.L. Capelo-Martínez, A
comparison of depletion versus equalization for reducing high‐abundance proteins in
human serum, Electrophoresis 32(21) (2011) 2966-2974.
[35] J.R. de Jesus, R. da Silva Fernandes, G. de Souza Pessôa, I.M. Jr. Raimundo,
M.A.Z. Arruda, Depleting high-abundant and enriching low-abundant proteins in
human serum: An evaluation of sample preparation methods using magnetic

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
139
nanoparticle, chemical depletion and immunoaffinity techniques, Talanta 170 (2017)
199-209.
[36] M.P. Chantada-Vázquez, A. Castro López, S.B. Bravo, S. Vázquez-Estévez, B.
Acea-Nebril, C. Núñez, Proteomic analysis of the bio-corona formed on the surface of
(Au, Ag, Pt)-nanoparticles in human serum, Colloids Surf B Biointerfaces 177 (2019)
141-148.
[37] G. Candiano, M. Bruschi, L. Musante, L. Santucci, G.M. Ghiggeri, B. Carnemolla,
P. Orecchia, L. Zardi, P.G. Righetti, Blue silver: a very sensitive colloidal Coomassie
G-250 staining for proteome analysis, Electrophoresis 25(9) (2004) 1327-1333.
[38] E. Oliveira, J.E. Araújo, S. Gómez-Meire, C. Lodeiro, C. Perez-Melon, E. Iglesias-
Lamas, A. Otero-Glez, J.L. Capelo, H.M. Santos, Proteomics analysis of the peritoneal
dialysate effluent reveals the presence of calcium-regulation proteins and acute
inflammatory response, Clin. Proteomics 11(1) (2014) 17.
[39] I.V. Shilov, S.L. Seymour, A.A. Patel, A. Loboda, W.H. Tang, S.P. Keating, C.L.
Hunter, L.M. Nuwaysir, D.A. Schaeffer, The paragon algorithm, a next generation
search engine that uses sequence temperature values and feature probabilities to identify
peptides from tandem mass spectra, Mol. Cell Proteomics 6 (2007) 1638-1655.
[40] W.H. Tang, I.V. Shilov, S.L. Seymour, Nonlinear fitting method for determining
local false discovery rates from decoy database searches, J. Proteome Res. 7 (2008)
3661-3667.
[41] I. Ortea, I. Ruiz-Sánchez, R. Cañete, J. Caballero-Villarraso, M.D. Cañete,
Identification of candidate serum biomarkers of childhood-onset growth hormone
deficiency using SWATH-MS and feature selection, J. Proteomics 175 (2018) 105-113.
[42] J.G. Meyer, B. Schilling, Clinical applications of quantitative proteomics using
targeted and untargeted data-independent acquisition techniques, Expert Rev.
Proteomics 14(5) (2017) 419-429.
[43] Y. Luo, T.S. Mok, X. Lin, W. Zhang, Y. Cui, J. Guo, X. Chen, T. Zhang, T. Wang,
SWATH-based proteomics identified carbonic anhydrase 2 as a potential diagnosis
biomarker for nasopharyngeal carcinoma, Sci. Rep. 7 (2017) 41191.
[44] H.T. Tan, M.C.M. Chung, Label‐free quantitative phosphoproteomics reveals
regulation of vasodilator‐stimulated phosphoprotein upon stathmin‐1 silencing in a pair
of isogenic colorectal cancer cell lines, Proteomics 18(8) (2018) e1700242.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
140
[45] M.A. Dobrovolskaia, A.K. Patri, J. Zheng, J.D. Clogston, N. Ayub, P. Aggarwal,
B.W. Neun, J.B. Hall, S.E. McNeil, Interaction of colloidal gold nanoparticles with
human blood: effects on particle size and analysis of plasma protein binding profiles,
Nanomedicine: NBM 5 (2009) 106-117.
[46] R. García-Álvarez, M. Hadjidemetriou, A. Sánchez-Iglesias, L.M. Liz-Marzán, K.
Kostarelos, In vivo formation of protein corona on gold nanoparticles. The effect of size
and shape, Nanoscale 10(3) (2018) 1256-1264.
[47] S.T. Yang, Y. Liu, Y.W. Wang, A. Cao, Biosafety and bioapplication of
nanomaterials by designing protein-nanoparticle interactions, Small. 9 (2013) 1635-
1653.
[48] J. Chen, Y. Wang, X. Ding, Y. Huang, K. Xu, Magnetic solid-phase extraction of
proteins based on hydroxy functional ionic liquid-modified magnetic nanoparticles,
Anal. Methods. 6 (2014) 8358-8367.
[49] L. Treuel, D. Docter, M. Maskos, R.H. Stauber, R.H. Stauber, Protein corona -
from molecular adsorption to physiological complexity, Beilstein J. Nanotechnol. 6
(2015) 857-873.
[50] M. Cassandri, A. Smirnov, F. Novelli, C. Pitolli, M. Agostini, M. Malewicz, G.
Melino, G. Raschellà, Zinc-finger proteins in health and disease, Cell Death Discov. 3
(2017) 17071.
[51] J. Jen, Y.-C. Wang, Zinc finger proteins in cancer progression, J. Biomed. Sci. 23
(2016) 53.
[52] B.I. Bassey-Archibong, J.M. Kwiecien, S.B. Milosavljevic, R.M. Hallett, L.G.
Rayner, M.J. Erb, C.J. Crawford-Brown, K.B. Stephenson, P.A. Bédard, J.A. Hassell,
J.M. Daniel, Kaiso depletion attenuates transforming growth factor-β signaling and
metastatic activity of triple-negative breast cancer cells, Oncogenesis 5 (2016) e208.
[53] E.S. Radisky, D.C. Radisky, Matrix metalloproteinases as breast cancer drivers and
therapeutic targets, Front. Biosci. (Landmark Ed) 20 (2015) 1144-1163.
[54] K. Kessenbrock, V. Plaks, Z. Werb, Matrix metalloproteinases: regulators of the
tumor microenvironment, Cell. 141(1) (2010) 52-67.
[55] E.S. Radisky, D.C. Radisky, Stromal induction of breast cancer: inflammation and
invasion, Rev. Endocr. Metab. Disord. 8(3) (2007) 279-287.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
141
[56] E.S. Radisky, D.C. Radisky, Matrix metalloproteinase-induced epithelial-
mesenchymal transition in breast cancer, J Mammary Gland Biol Neoplasia. 15(2)
(2010) 201-212.
[57] D.P. Nanda, H. Sil, S. Moulik, J. Biswas, S.S. Mandal, A. Chatterjee, Matrix
metalloproteinase-9 as a potential tumor marker in breast cancer, J. Environ. Pathol.
Toxicol. Oncol. 32(2) (2013) 115-129.
[58] L.J. van 't Veer, H. Dai, M.J. van de Vijver, Y.D. He, A.A. Hart, M. Mao, H.L.
Peterse, K. van der Kooy, M.J. Marton, A.T. Witteveen, G.J. Schreiber, R.M.
Kerkhoven, C. Roberts, P.S. Linsley, R. Bernards, S.H. Friend, Gene expression
profiling predicts clinical outcome of breast cancer, Nature 415(6871) (2002) 530-536.
[59] T.G. Kormelink, D.G. Powe, S.A. Kuijpers, A. Abudukelimu, M.H.A.M. Fens,
E.H.E. Pieters, W.W. Kassing-van der Ven, H.O. Habashy, I.O. Ellis, B.R. Blokhuis, M.
Thio, W.E. Hennink, G. Storm, F.A. Redegeld, R.M. Schiffelers, Immunoglobulin free
light chains are biomarkers of poor prognosis in basal-like breast cancer and are
potential targets in tumor-associated inflammation, Oncotarget 5(10) (2014) 3159-3167.
[60] S.L. Anwar, W. Wulaningsih, U. Lehmann, Transposable elements in human
cancer: causes and consequences of deregulation, Int J Mol Sci. 18(5) (2017) pii: E974.
[61] C. Behrens, B. Binotti, C. Schmidt, C.V. Robinson, J.J. Chua, K. Kühnel, Crystal
structure of the human short coiled coil protein and insights into SCOC-FEZ1 complex
formation, PLoS One. 8(10) (2013) e76355.
[62] F.U. Nasim, S. Ejaz, M. Ashraf, A.R. Asif, M. Oellerich, G. Ahmad, G.A. Malik,
Attiq-Ur-Rehman, Potential biomarkers in the sera of breast cancer patients from
bahawalpur, Pakistan, Biomark Cancer. 4 (2012) 19-34.
[63] C. Jiao, L. Cui, A. Ma, N. Li, H. Si, Elevated serum levels of retinol-binding
protein 4 are associated with breast cancer risk: a case-control study, PLoS One. 11(12)
(2016) e0167498.
[64] Y.K. Bae, A. Kim, M.K. Kim, J.E. Choi, S.H. Kang, S.J. Lee, Fibronectin
expression in carcinoma cells correlates with tumor aggressiveness and poor clinical
outcome in patients with invasive breast cancer, Hum Pathol. 44 (2013) 2028-2037.
[65] D. Arora, Z. Mahmood, J. George, A.K. Yadav, S. Kumar, U.S. Singh, Y. Shukla,
Plasma protein profiling of breast cancer patients of North Indian population: A
potential approach to early detection, J Proteomics Bioinform 6 (2013) 5.

III. Results and Discussion. CHAPTER 2
______________________________________________________________________
142
[66] R. Kumar, E. Kuligina, A. Sokolenko, I. Bizin, A.K. Varma, S.K. Hasan,
Identification of pregnancy zone protein in the pathogenesis of breast cancer, December
2018, DOI: 10.13140/RG.2.2.15317.06887.
[67] M. Tinholt, Ø. Garred, E. Borgen, E. Beraki, M. Sletten, K. Kleivi Sahlberg, P.M.
Sandset, N. Iversen, Coagulation factor V is expressed in tumors and predicts favorable
outcome in aggressive breast cancer, Thrombosis Research 164 (2018) S183-S184.
[68] E. Sierko, M.Z. Wojtukiewicz, K. Ostrowska-Cichocka, L. Zimnoch, Protein Z-
dependent protease inhibitor (ZPI) is present in loco in human breast cancer tissue,
Thromb Haemost. 104(1) (2010) 183-185.
[69] K.D. Smith, J. Behan, G. Matthews-Smith, A.M. Magliocco, Alpha-1-Acid
Glycoprotein (AGP) as a potential biomarker for breast cancer, (2012) DOI:
10.5772/48177.

CHAPTER 3
Protein Corona Gold Nanoparticles Fingerprinting Reveals a
Profile of Blood Coagulation Proteins in the Serum of HER2-
Overexpressing Breast Cancer Patients
María del Pilar Chantada-Vázquez, Antonio Castro López, María García-Vence, Benigno
Acea-Nebril, Susana B. Bravo, Cristina Núñez
International Journal of Molecular Sciences 21 (2020) 8449
DOI: 10.3390/ijms21228449


III. Results and Discussion. CHAPTER 3
143
Protein corona gold nanoparticles fingerprinting reveals a profile of blood
coagulation proteins in the serum of HER2-overexpressing breast cancer
patients
María del Pilar Chantada-Vázquez,1,2 Antonio Castro López,3 María García-Vence,2 Benigno
Acea-Nebril,4 Susana B. Bravo,2 Cristina Núñez1
1 Research Unit, Lucus Augusti University Hospital (HULA), Servizo Galego de Saúde
(SERGAS), 27002 Lugo, Spain;
2 Proteomic Unit, Health Research Institute of Santiago de Compostela (IDIS), University
Clinical Hospital of Santiago de Compostela (CHUS), 15706 Santiago de Compostela, Spain;
3 Breast Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde
(SERGAS), 27002 Lugo, Spain;
4 Department of Surgery, Breast Unit, Complexo Hospitalario Universitario A Coruña
(CHUAC), Servizo Galego de Saúde (SERGAS), 15006 A Coruña, Spain;
Abstract
Breast cancer (BC) is a molecularly heterogeneous disease that encompasses five
major molecular subtypes (luminal A (LA), luminal B HER2 negative (LB-), luminal B
HER2 positive (LB+), HER2 positive (HER2+), the triple negative breast cancer (TNBC)).
BC treatment mainly depends on the identification of the specific subtype. Despite the correct
identification, therapies could fail in some patients. Thus, further insights into the genetic and
molecular status of the different BC subtypes could be very useful to improve the response
of BC patients to the range of available therapies. In this way, we use gold nanoparticles
(AuNPs, 12.96 ± 0.72 nm) as a scavenging tool in combination with Sequential Window
Acquisition of All Theoretical Mass Spectra (SWATH-MS) to quantitative analyse the serum
proteome alterations in the different breast cancer intrinsic subtypes. The differentially
regulated proteins specific of each subtype were further analysed with the bioinformatic tools
STRING and PANTHER to identify the major molecular function, biological processes,
cellular origin, protein class and biological pathways altered due to the heterogeneity in

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
144
proteome of the different BC subtypes. Importantly, a profile of blood coagulation proteins
was identified in the serum of HER2-overexpressing BC patients.
Keywords: Protein corona (PC); gold nanoparticles (AuNPs); breast cancer (BC);
fingerprinting; SWATH-MS; HER2+.
1. Introduction
Breast cancer (BC) is a heterogeneous disease that presents a wide variety of molecular
and clinical characteristics, as well as variability in clinical progression [1]. For the treatment
choice, patients are classified according to intrinsic biological subtypes within the BC
spectrum, using clinical-pathological criteria, i.e. the recognition of amplification and/or
overexpression of the human epidermal growth factor receptor 2 (HER2) oncogene, the
immunohistochemical classification of the estrogen receptor (ER) and the progesterone
receptor (PR), and Ki-67 labelling index [ 2 ]. This classification allows for a more
personalized approach to medical treatments, with favourable results. However, in spite of
that, almost 10-15% of these patients still experience local or distant recurrences in the first
5 years from diagnosis [3]. Particularly, HER2-positive BC, defined by the overexpression
of HER2 protein, represents 15-20% of BC cases [4 , 5 ] and, is correlated with poor
prognosis, high rates of recurrence and short survival [6].
Classification of BC might be markedly improved if new biomarkers identified with the
use of high-throughput “omics” approaches could support diagnosis based on
histopathological patterns [7-9]. Nanomaterials have been introduced into the field of
proteomics to establish a new and rapidly evolving research area termed nanoproteomics
[10].
It is well known that the dispersion of a nanomaterial in physiological fluid results in the
formation of a protein shell named “protein corona” (PC). PC varies depending on the
characteristics of the biological media, the physical (size, shape, curvature) and chemical
properties (composition, surface charge/chemistry, hydrophobicity/hydrophilicity) of the
nanomaterial, and the incubation time [11]. Disease-associated biomarkers comprise less
than 1% of serum proteins. In this way, through the formation of the PC, nanoparticles could
act as sorbent materials for the enrichment of low-abundance peptides/proteins presented in
serum samples before the biomarker identification by mass spectrometry (MS) analysis [12-

III. Results and Discussion. CHAPTER 3
145
14]. Importantly, otherwise undetectable changes in protein concentration at an early stage of
the disease (as breast cancer), after any treatment (chemotherapy, immunotherapy) or surgery
could be detected analyzing the PC composition [15]. Thus, characterization of the PC around
NPs offers distinct advantages over sole proteomic approaches and increases the success of
identifying molecular targets [16].
Particularly, AuNPs present some properties to be used as suitable sorbent
nanomaterials: high surface-area-to-volume ratio, colloidal stability, and the capability to
conjugate with biomolecules [17]. Here, the interaction of AuNPs (12.96 ± 0.72 nm) with the
sera of disease-free women (healthy controls, HC) (n = 42) and BC patients (n = 42) allowed
the pre-concentration of the low-abundance proteins thorough the PC formation. Then, an
exhaustive quantitative analysis of the PCs by SWATH-MS was carried out to identify novel
molecular targets associated to the different BC intrinsic subtypes (see Figure 1).
Figure 1. Simplified representation of the experimental procedure. AuNPs (12.96 ± 0.72
nm) were incubated ex vivo with human serum samples obtained from HC (n = 42) and
BC patients (n = 42). Ex vivo corona-coated AuNPs were recovered and purified from
unbound proteins by centrifugation. The formed PCs were quantitatively characterized
and analysed by SWATH-MS and compared between the groups.

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
146
2. Experimental
2.1. Chemicals and reagents
Acrylamide/bis-acrylamide 30% solution (37.5:1), β-mercaptoethanol (molecular
biology grade), Coomassie Brilliant Blue R250 (CBB), DL-dithiothreitol
(HSCH2CH(OH)CH(OH)CH2SH, 99%) (DTT), glycerol (HOCH2CH(OH)CH2OH, 86-
88%), iodoacetamide (IAA, ICH2CONH2, 99%), sodium citrate tribasic dehydrate
(HOC(COONa)(CH2COONa)2·2H2O, 99%), sodium carbonate (Na2CO3, 99%), tris-base
(NH2C(CH2OH)3), trifluoroacetic acid (CF3COOH, 99%), trypsin from bovine pancreas, and
the Sigma Marker wide range 6.5-200 KDa were purchased from Sigma-Aldrich.
Formaldehyde for molecular biology (36.5-38% in H2O) and sodium dodecylsulfate (SDS,
CH3(CH2)11SO4Na) were purchased from Panreac. Bromophenol-blue was purchased from
Riedel-de Haen. Hydrogen tetrachloroaurate (III) hydrate (HAuCl4·xH2O) (99.9%-Au) (49%
Au) at 10% w/v was purchased from Strem Chemicals. Ammoniumbicarbonate (AMBIC,
NH4HCO3, 99.5%) and formic acid (HCOOH, 95%) were purchased from Fluka.
2.2. Biological samples
Blood samples were collected from n = 42 newly diagnosed BC patients with the five
different breast cancer subtypes: n = 11 patients with the luminal A subtype (ER positive,
HER2 negative, Ki-67 low, and PR high), n = 10 patients with the luminal B-HER2 negative
subtype (ER positive, HER2 negative, and either Ki-67 high or PR low), n = 7 patients with
the luminal B-HER2 positive subtype (ER positive, HER2 overexpressed or amplified, any
Ki-67, and any PR), n = 6 patients with the HER2 positive subtype (HER2 over-expressed or
amplified, ER and PR absent), and n = 8 patients with the triple negative subtype (ER and
PR absent and HER2 negative); for ER: estrogen receptor, PR: progesterone receptor, HER2:
human epidermal growth factor receptor 2 (see Table 1_SM) [2].
Blood samples were also collected from n = 42 age-matched and gender-matched healthy
women (controls). In all cases, venous blood samples were collected in VACUETTE® Serum
Clot Activator Tubes (10 mL).
The experiment was conducted in conformity with the declaration of Helsinki and
approved by the Clinical Research Ethics Committees (CEIC) of Galicia (Spain) with

III. Results and Discussion. CHAPTER 3
147
approval number 2017-021. All participants from Lucus Augusti University Hospital (Spain)
gave written informed consent prior to their participation.
2.3. Synthesis of citrate-gold nanoparticles (AuNPs, 12.96 ± 0.72 nm)
Colloidal AuNPs with a size of 12.96 ± 0.72 nm were prepared by chemical reduction
method as per the protocol developed previously [12-14]. In short, sodium citrate tribasic
solution (0.075% w/v) was dissolved in 60 mL warm distilled water under constant magnetic
stirring. To this, 54 μL of 10% w/v of hydrogen tetrachloroaurate (III) hydrate solution was
then added drop-wise and the reaction was heated to 100 °C under constant magnetic stirring.
Reaction was allowed to proceed further the color of the solution changes from yellow to
deep red indicating the reduction of Au3+ to Au0, which spontaneously aggregates to form
colloidal AuNPs. This colloidal dispersion of AuNPs was cooled to room temperature and
preserved at 4 °C for further analysis.
2.4. Incubation of AuNPs with human serum samples: ex vivo protein corona formation
Firstly, collected blood samples were allowed to clot for 15 min. After that, samples
were centrifuged for 5 min at 4˚C and 1,800×g. Resultant serum were transferred to sterile
cryovials, frozen and stored at -80 °C until further use at Research Unit, Lucus Augusti
University Hospital (HULA). The formation of the ex vivo PC was achieved following the
steps showed in Figure 2 [12-14]. After that, unbound serum proteins from the surface of
AuNPs were removed by centrifugation at 18,840×g for 30 min.
2.4. Characterization of colloidal AuNPs
The morphology of the AuNPs was investigated by transmission electron microscopy
(TEM) with a JEM 1011, JEOL instrument. The size and ζ-potentials of colloidal AuNPs
were measured (3 determinations per sample) with a Malvern Zetasizer Nano ZS instrument
at 25°C.
Protein quantification and protein separation by SDS-PAGE were carried out with the
use of a Qubit™ 4 Quantitation Starter Kit (Thermo Fisher Scientific) and a PowerPacTM
Basic Power Supply (Bio-Rad), respectively.

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
148
2.5. Separation and digestion of the proteins presented in the corona-coated AuNPs
Corona proteins associated with AuNPs were separated by sodium dodecyl sulfate-
polyacrylamide gel electrophoresis (SDS-PAGE) and digested following the scheme
represented in Figure 2.
The digestion was stopped with the addition of 50 μL of 5% (v/v) formic acid. After that,
the extraction of the peptides from the gel was carried out with a solution of 50% (v/v)
acetonitrile/0.1% (v/v) trifluoroacetic acid (TFA) (x3) and acetonitrile (ACN) (x1). Samples
were dried-down and stored at -20 °C until their further use [18].
Figure 2. Flowchart depicting the separation and digestion of the corona proteins
associated with AuNPs.
2.6. Protein quantification by SWATH-MS
SWATH/MS experiments were carried out following the instrumental parameters
described elsewhere [13]. Briefly, two biological replicates of LA, LB-, LB+, HER+, TNBC
and HC samples were used to get extensive quantitative data by label-free SWATH-MS
analysis. Peptides of all samples were analysed with a micro-LC system Ekspert nLC425
(Eksigen, Dublin, CA. USA) couplet to a hybrid quadrupole-TOF mass spectrometer Triple
TOF 6600 (Sciex, Redwood City. CA. USA). One of the first steps is the construction of the
MS/MS spectral libraries. For that purpose, peptide solutions were analyzed by a shotgun

III. Results and Discussion. CHAPTER 3
149
data-dependent acquisition (DDA) approach by micro-LC-MS/MS. For spectral alignment
and peak extraction was employed the Peakview software (version 2.2; AB Sciex) using the
SWATH Acquisition MicroApp (version 2.0). Parameters used were: number of fragments
= 7, number of peptides = 10, peptide confidence = 95%, XIC width = 30 ppm, XIC extraction
window = 5 min. Exportation of the SWATH file to the MarkerView software (version 1.3.1;
AB Sciex) allowed the quantitative analysis of ions, peptides and proteins in the different
samples. As output result, the summed intensity of ions for the peptide, summed intensity of
the peptides for protein and Area under Curve (AUC) of the ions were provided. The test set
(LA, LB-, LB+, HER+, TNBC) was compared with the control (HC) dataset to generate fold
change ratios. For protein quantitation, only peptides with a False Discovery Rate (FDR)
below 1% were considered. Average MS peak area of each protein derived from the analysis
of the biological replicates and Student’s t-test analysis among samples was developed. t-test
indicates the capacity of each variable to distinguish between two groups, and it was reported
as a p-value. The criteria to select differentially expressed proteins was a p-value <0.05 with
a 1.5-fold in- or decrease.
2.7. Protein functional interaction network analysis and protein ontology classification
The informatic tool STRING v.10.0 database (http://string-db.org) was the used to
analyse the the functional interaction networks of the proteins, integrating direct (physical)
and indirect protein-protein interactions (PPI) [19].
Protein ontology classification was performed with the PANTHER classification system
(http:// www.pantherdb.org/). The differentially expressed proteins in the different breast
cancer subtypes were grouped according to their major molecular function, biological
processes, cellular origin, protein class and biological pathways.
3. Results and discussion
3.1. Incubation of AuNPs (12.96 ± 0.72 nm) with human serum samples: ex vivo protein
corona formation and characterization
Human serum samples from n = 42 healthy controls (HC) and n = 42 breast cancer (BC)
patients (n = 42) have been recruited, handled and analysed in the same way as it is further
detailed in Figure 3. The group of BC patients was divided into the following biological

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
150
subtypes: n = 11 patients with the luminal A (LA) subtype, n = 10 patients with the luminal
B HER2 negative (LB-) subtype, n = 7 patients with the luminal B HER2 positive (LB+)
subtype, n = 6 patients with the HER2 positive (HER2+) subtype, and n = 8 patients with the
triple negative breast cancer (TNBC) subtype. Patient clinical characteristics are summarized
in Table 1_SM.
AuNPs with a size of 12.96 ± 0.72 nm were prepared by a chemical reduction method
[12-14]. As Figure 3 shows, proteins presented in serum samples (x2) were chemically
reduced with dithiothreitol (DTT) and alkylated with iodoacetamide (IAA) before their ex
vivo incubation with AuNPs (12.96 ± 0.72 nm) to get the formation of the PCs [12-14].
Figure 3. Flowchart depicting serum samples pretreatment and protein corona formation.
After the ex vivo incubation of AuNPs with human serum samples of HC (n = 42) and
BC patients (n = 42), the resultant protein corona-coated AuNPs were centrifugated and
structurally characterized by dynamic light scattering (DLS) and negative stain transmission
electron microscopy (TEM). DLS measurements showed that the interaction of serum

III. Results and Discussion. CHAPTER 3
151
proteins with the surface of AuNPs resulted in an increase of the size of the AuNPs, from
12.96 ± 0.72 nm to 17.33 ± 1.55 nm (HC) and 17.13 ± 1.53 nm (BC) (see Table 2_SM).
Probably, the preferential interaction of positively charged proteins with the AuNPs surface
promoted the increase of the mean particle surface charge from -38.3 mV (bare AuNPs) to -
30.5 mV (HC) and -30.3 mV (BC) [20, 21]. TEM imaging revealed a well-dispersed
nanoparticles population corroborating the PC formation around AuNPs.
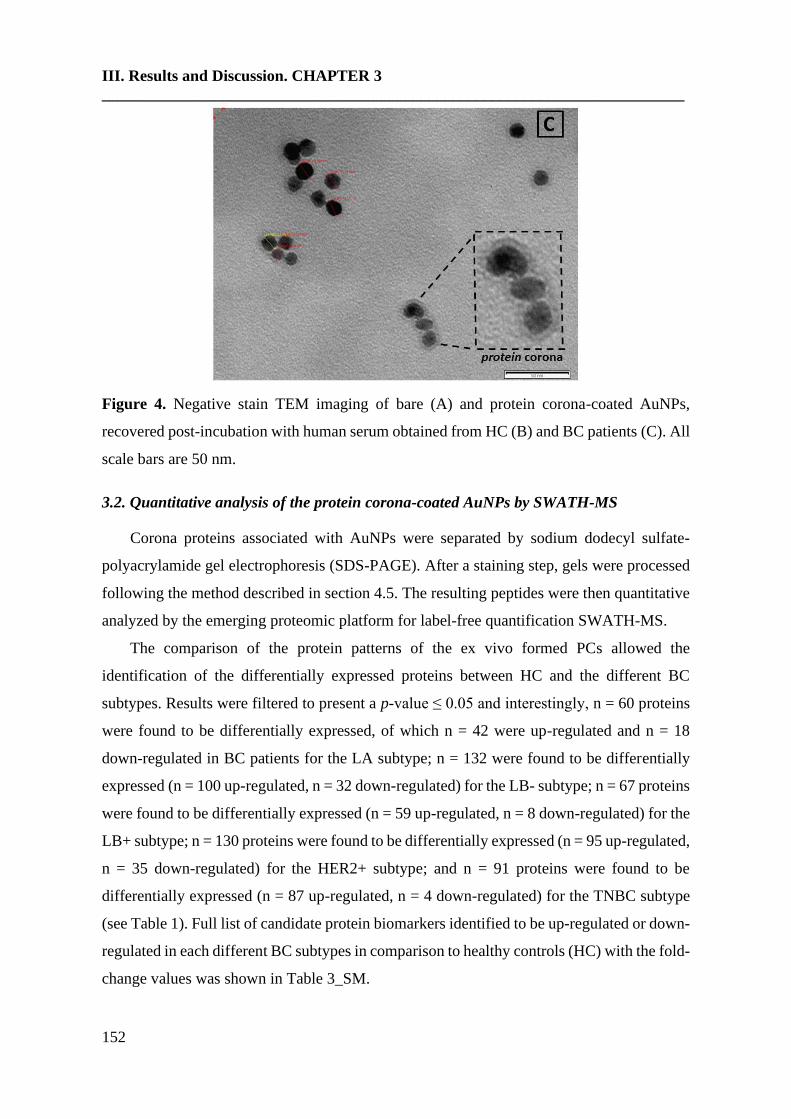
III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
152
Figure 4. Negative stain TEM imaging of bare (A) and protein corona-coated AuNPs,
recovered post-incubation with human serum obtained from HC (B) and BC patients (C). All
scale bars are 50 nm.
3.2. Quantitative analysis of the protein corona-coated AuNPs by SWATH-MS
Corona proteins associated with AuNPs were separated by sodium dodecyl sulfate-
polyacrylamide gel electrophoresis (SDS-PAGE). After a staining step, gels were processed
following the method described in section 4.5. The resulting peptides were then quantitative
analyzed by the emerging proteomic platform for label-free quantification SWATH-MS.
The comparison of the protein patterns of the ex vivo formed PCs allowed the
identification of the differentially expressed proteins between HC and the different BC
subtypes. Results were filtered to present a p-value ≤ 0.05 and interestingly, n = 60 proteins
were found to be differentially expressed, of which n = 42 were up-regulated and n = 18
down-regulated in BC patients for the LA subtype; n = 132 were found to be differentially
expressed (n = 100 up-regulated, n = 32 down-regulated) for the LB- subtype; n = 67 proteins
were found to be differentially expressed (n = 59 up-regulated, n = 8 down-regulated) for the
LB+ subtype; n = 130 proteins were found to be differentially expressed (n = 95 up-regulated,
n = 35 down-regulated) for the HER2+ subtype; and n = 91 proteins were found to be
differentially expressed (n = 87 up-regulated, n = 4 down-regulated) for the TNBC subtype
(see Table 1). Full list of candidate protein biomarkers identified to be up-regulated or down-
regulated in each different BC subtypes in comparison to healthy controls (HC) with the fold-
change values was shown in Table 3_SM.

III. Results and Discussion. CHAPTER 3
153
Figure 5. Venn diagram showing the number of shared and specific (or unique)
deregulated proteins identified in the PCs formed after the interaction of AuNPs (12.96
± 0.72 nm) with serum samples of the different BC subtypes (LA, LB-, LB+, HER2+,
TNBC).
Table 1. Number of differentially expressed proteins (up-regulated and down-regulated)
(p-value ≤ 0.05) found in the protein patterns of the ex vivo formed coronas after the
analysis by SWATH-MS for the different breast cancer subtypes (LA LB-, LB+, HER2+,
TNBC) in comparison with healthy controls (HC) samples. The number of differentially
expressed proteins (up-regulated and down-regulated) specific to each of the five
subtypes of BC found in the ex vivo formed coronas were also indicated.
SWATH-MS analysis
Comparison Protein number (p-value ≤ 0.05)
Total Up-regulated Down-regulated Specific Up-regulated Down-regulated
Controls vs LA 60 42 18 8 4 4
Controls vs LB- 132 100 32 27 25 2
Controls vs LB+ 67 59 8 2 2 0
Controls vs HER2+ 130 95 35 28 23 5
Controls vs TNBC 91 87 4 10 9 1
Venn diagram of statistically significant (up- and down-) regulated proteins showed that
seven proteins were found to be commonly altered in all BC subtypes (see Figure 5). These
common proteins were apolipoprotein C-III (APOC3), c-reactive protein (CRP), hemoglobin
subunit beta (HBB), immunoglobulin heavy variable 3-49 (IGHV3-49), serum amyloid A-4
protein (SAA4), serum amyloid P-component (APCS) and serotransferrin (TR) (Table 4_SM
and Table 5_SM).
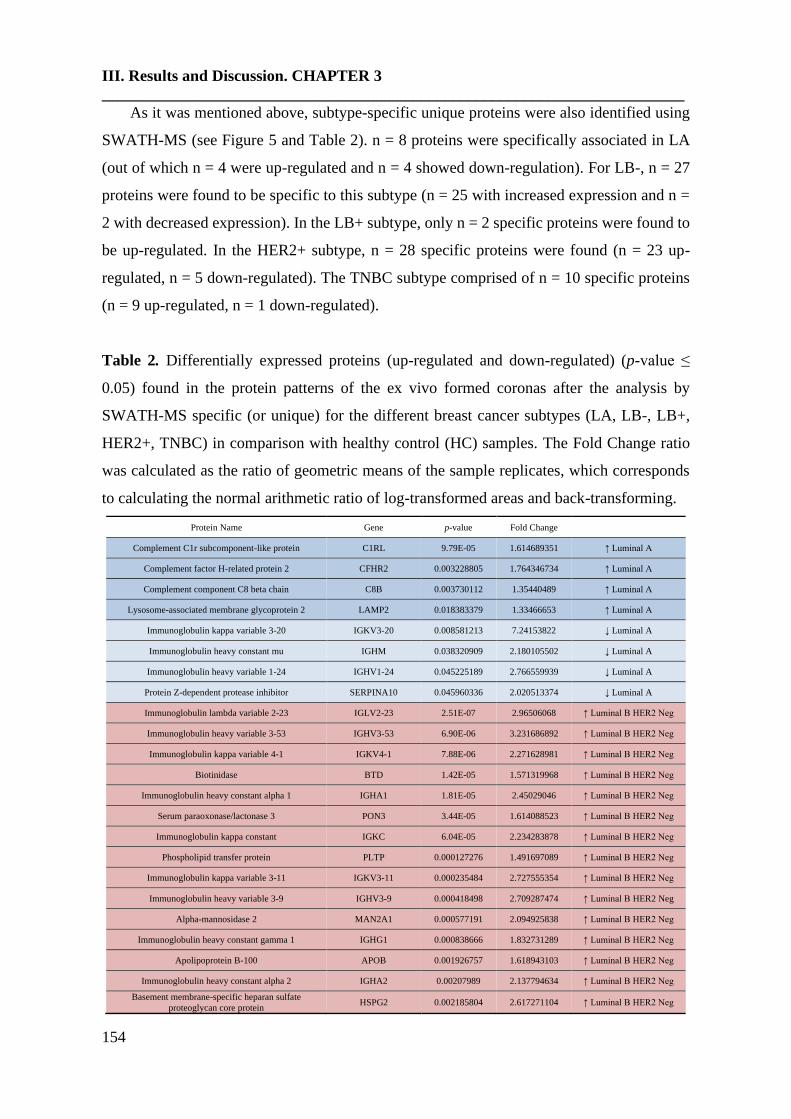
III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
154
As it was mentioned above, subtype-specific unique proteins were also identified using
SWATH-MS (see Figure 5 and Table 2). n = 8 proteins were specifically associated in LA
(out of which n = 4 were up-regulated and n = 4 showed down-regulation). For LB-, n = 27
proteins were found to be specific to this subtype (n = 25 with increased expression and n =
2 with decreased expression). In the LB+ subtype, only n = 2 specific proteins were found to
be up-regulated. In the HER2+ subtype, n = 28 specific proteins were found (n = 23 up-
regulated, n = 5 down-regulated). The TNBC subtype comprised of n = 10 specific proteins
(n = 9 up-regulated, n = 1 down-regulated).
Table 2. Differentially expressed proteins (up-regulated and down-regulated) (p-value ≤
0.05) found in the protein patterns of the ex vivo formed coronas after the analysis by
SWATH-MS specific (or unique) for the different breast cancer subtypes (LA, LB-, LB+,
HER2+, TNBC) in comparison with healthy control (HC) samples. The Fold Change ratio
was calculated as the ratio of geometric means of the sample replicates, which corresponds
to calculating the normal arithmetic ratio of log-transformed areas and back-transforming.
Protein Name Gene p-value Fold Change
Complement C1r subcomponent-like protein C1RL 9.79E-05 1.614689351 ↑ Luminal A
Complement factor H-related protein 2 CFHR2 0.003228805 1.764346734 ↑ Luminal A
Complement component C8 beta chain C8B 0.003730112 1.35440489 ↑ Luminal A
Lysosome-associated membrane glycoprotein 2 LAMP2 0.018383379 1.33466653 ↑ Luminal A
Immunoglobulin kappa variable 3-20 IGKV3-20 0.008581213 7.24153822 ↓ Luminal A
Immunoglobulin heavy constant mu IGHM 0.038320909 2.180105502 ↓ Luminal A
Immunoglobulin heavy variable 1-24 IGHV1-24 0.045225189 2.766559939 ↓ Luminal A
Protein Z-dependent protease inhibitor SERPINA10 0.045960336 2.020513374 ↓ Luminal A
Immunoglobulin lambda variable 2-23 IGLV2-23 2.51E-07 2.96506068 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 3-53 IGHV3-53 6.90E-06 3.231686892 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 4-1 IGKV4-1 7.88E-06 2.271628981 ↑ Luminal B HER2 Neg
Biotinidase BTD 1.42E-05 1.571319968 ↑ Luminal B HER2 Neg
Immunoglobulin heavy constant alpha 1 IGHA1 1.81E-05 2.45029046 ↑ Luminal B HER2 Neg
Serum paraoxonase/lactonase 3 PON3 3.44E-05 1.614088523 ↑ Luminal B HER2 Neg
Immunoglobulin kappa constant IGKC 6.04E-05 2.234283878 ↑ Luminal B HER2 Neg
Phospholipid transfer protein PLTP 0.000127276 1.491697089 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 3-11 IGKV3-11 0.000235484 2.727555354 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 3-9 IGHV3-9 0.000418498 2.709287474 ↑ Luminal B HER2 Neg
Alpha-mannosidase 2 MAN2A1 0.000577191 2.094925838 ↑ Luminal B HER2 Neg
Immunoglobulin heavy constant gamma 1 IGHG1 0.000838666 1.832731289 ↑ Luminal B HER2 Neg
Apolipoprotein B-100 APOB 0.001926757 1.618943103 ↑ Luminal B HER2 Neg
Immunoglobulin heavy constant alpha 2 IGHA2 0.00207989 2.137794634 ↑ Luminal B HER2 Neg
Basement membrane-specific heparan sulfate
proteoglycan core protein HSPG2 0.002185804 2.617271104 ↑ Luminal B HER2 Neg

III. Results and Discussion. CHAPTER 3
155
Pregnancy zone protein PZP 0.002663618 4.343426266 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 1-69D IGHV1-69D 0.00307313 2.762019962 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 3-74 IGHV3-74 0.005843078 1.63300774 ↑ Luminal B HER2 Neg
Immunoglobulin lambda-like polypeptide 5 IGLL5 0.006474796 1.674164832 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 3D-20 IGKV3D-20 0.007387257 1.722873358 ↑ Luminal B HER2 Neg
L-lactate dehydrogenase B chain LDHB 0.011088233 1.464826821 ↑ Luminal B HER2 Neg
Platelet glycoprotein Ib alpha chain GP1BA 0.014355443 1.435497779 ↑ Luminal B HER2 Neg
Apolipoprotein D APOD 0.02689663 1.565894604 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 3-21 IGLV3-21 0.030887039 1.531598796 ↑ Luminal B HER2 Neg
Mediator of RNA polymerase II transcription subunit
23 MED23 0.034189832 1.582094144 ↑ Luminal B HER2 Neg
Platelet factor 4 variant PF4V1 0.013149247 8.695958011 ↓ Luminal B HER2 Neg
Complement C1s subcomponent C1S 0.040782815 1.400513823 ↓ Luminal B HER2 Neg
Plastin-2 LCP1 0.002018957 4.501473239 ↑ Luminal B HER2 Pos
Immunoglobulin heavy variable 6-1 IGHV6-1 0.020191442 1.539338405 ↑ Luminal B HER2 Pos
Complement C5 C5 2.95E-15 2.104078338 ↑ HER2 Pos
Adiponectin ADIPOQ 8.28E-09 9.579128591 ↑ HER2 Pos
Immunoglobulin heavy variable 3-73 IGHV3-73 1.67E-06 15.87006684 ↑ HER2 Pos
Coagulation factor XII F12 2.50E-06 4.48323521 ↑ HER2 Pos
Plasma kallikrein KLKB1 2.05E-05 2.839283512 ↑ HER2 Pos
Immunoglobulin heavy variable 3-23 IGHV3-23 8.83E-05 2.952483084 ↑ HER2 Pos
Immunoglobulin lambda variable 1-51 IGLV1-51 0.000382645 2.419267666 ↑ HER2 Pos
Immunoglobulin heavy variable 3-64 IGHV3-64 0.000401774 2.217759109 ↑ HER2 Pos
Selenoprotein P SELENOP 0.000539614 3.796988329 ↑ HER2 Pos
Immunoglobulin kappa variable 1D-12 IGKV1D-12 0.00188481 4.471370936 ↑ HER2 Pos
Immunoglobulin lambda variable 5-45 IGLV5-45 0.00772078 2.858034161 ↑ HER2 Pos
Immunoglobulin lambda variable 6-57 IGLV6-57 0.009251968 5.252644969 ↑ HER2 Pos
Keratin type I cytoskeletal 10 KRT10 0.012361191 1.599012598 ↑ HER2 Pos
Immunoglobulin kappa variable 1-27 IGKV1-27 0.014636279 3.663302525 ↑ HER2 Pos
Immunoglobulin kappa variable 1-5 IGKV1-5 0.015089737 3.097611266 ↑ HER2 Pos
EGF-containing fibulin-like extracellular matrix
protein 1 EFEMP1 0.015909032 1.962284999 ↑ HER2 Pos
Immunoglobulin kappa variable 2-24 IGKV2-24 0.019946847 3.792528291 ↑ HER2 Pos
Immunoglobulin heavy constant gamma 2 IGHG2 0.020013155 1.605775681 ↑ HER2 Pos
Adipocyte plasma membrane-associated protein APMAP 0.021204299 23.89017843 ↑ HER2 Pos
Immunoglobulin kappa variable 1D-16 IGKV1D-16 0.023927557 15.36602613 ↑ HER2 Pos
Coagulation factor V F5 0.025887503 3.739380413 ↑ HER2 Pos
Cysteine-rich secretory protein 3 CRISP3 0.034870563 3.338300324 ↑ HER2 Pos
Immunoglobulin heavy variable 3-33 IGHV3-33 0.038394512 8.344444793 ↑ HER2 Pos
N-acetylmuramoyl-L-alanine amidase PGLYRP2 0.000321687 1.799194526 ↓ HER2 Pos
Alpha-1-antitrypsin SERPINA1 0.001111283 4.959761437 ↓ HER2 Pos
Trypsin-1 PRSS1 0.002454431 4.217786063 ↓ HER2 Pos
Apolipoprotein F APOF 0.005336626 7.55893037 ↓ HER2 Pos
Antithrombin-III SERPINC1 0.018612975 1.308597079 ↓ HER2 Pos
Apolipoprotein E APOE 0.005108321 1.297484301 ↑ Triple Negative
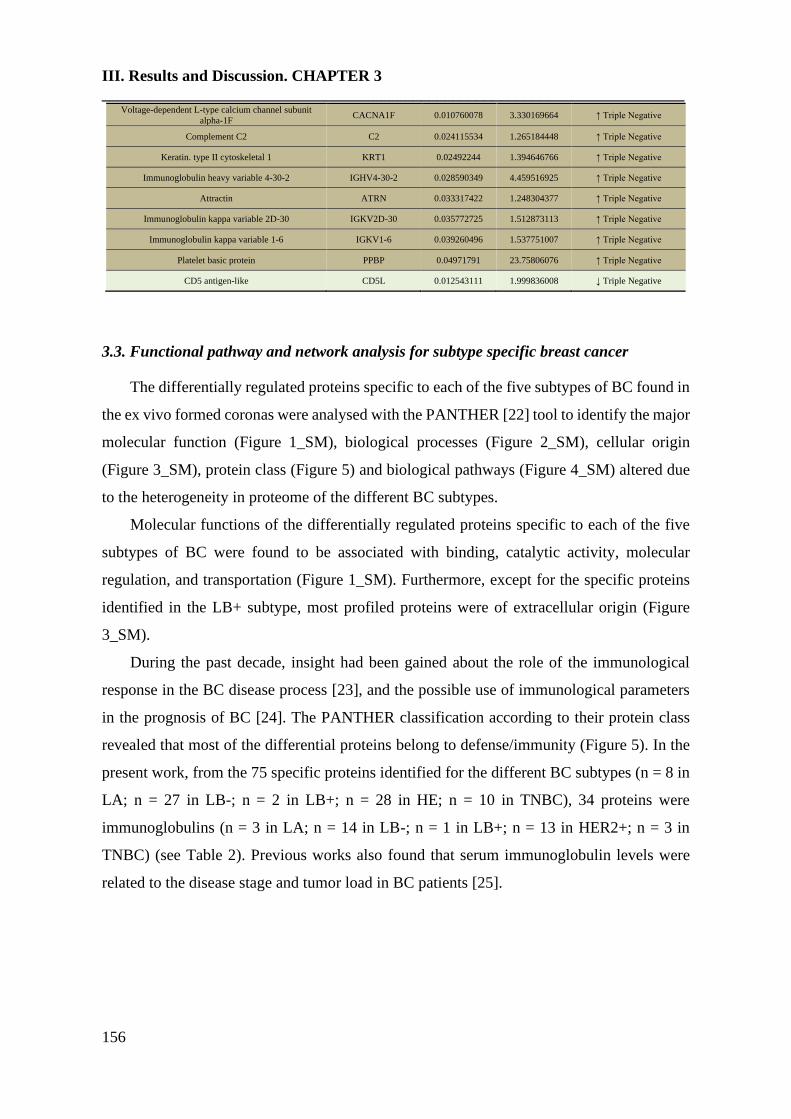
III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
156
Voltage-dependent L-type calcium channel subunit
alpha-1F CACNA1F 0.010760078 3.330169664 ↑ Triple Negative
Complement C2 C2 0.024115534 1.265184448 ↑ Triple Negative
Keratin. type II cytoskeletal 1 KRT1 0.02492244 1.394646766 ↑ Triple Negative
Immunoglobulin heavy variable 4-30-2 IGHV4-30-2 0.028590349 4.459516925 ↑ Triple Negative
Attractin ATRN 0.033317422 1.248304377 ↑ Triple Negative
Immunoglobulin kappa variable 2D-30 IGKV2D-30 0.035772725 1.512873113 ↑ Triple Negative
Immunoglobulin kappa variable 1-6 IGKV1-6 0.039260496 1.537751007 ↑ Triple Negative
Platelet basic protein PPBP 0.04971791 23.75806076 ↑ Triple Negative
CD5 antigen-like CD5L 0.012543111 1.999836008 ↓ Triple Negative
3.3. Functional pathway and network analysis for subtype specific breast cancer
The differentially regulated proteins specific to each of the five subtypes of BC found in
the ex vivo formed coronas were analysed with the PANTHER [22] tool to identify the major
molecular function (Figure 1_SM), biological processes (Figure 2_SM), cellular origin
(Figure 3_SM), protein class (Figure 5) and biological pathways (Figure 4_SM) altered due
to the heterogeneity in proteome of the different BC subtypes.
Molecular functions of the differentially regulated proteins specific to each of the five
subtypes of BC were found to be associated with binding, catalytic activity, molecular
regulation, and transportation (Figure 1_SM). Furthermore, except for the specific proteins
identified in the LB+ subtype, most profiled proteins were of extracellular origin (Figure
3_SM).
During the past decade, insight had been gained about the role of the immunological
response in the BC disease process [23], and the possible use of immunological parameters
in the prognosis of BC [24]. The PANTHER classification according to their protein class
revealed that most of the differential proteins belong to defense/immunity (Figure 5). In the
present work, from the 75 specific proteins identified for the different BC subtypes (n = 8 in
LA; n = 27 in LB-; n = 2 in LB+; n = 28 in HE; n = 10 in TNBC), 34 proteins were
immunoglobulins (n = 3 in LA; n = 14 in LB-; n = 1 in LB+; n = 13 in HER2+; n = 3 in
TNBC) (see Table 2). Previous works also found that serum immunoglobulin levels were
related to the disease stage and tumor load in BC patients [25].

III. Results and Discussion. CHAPTER 3
157
Figure 5. Classification according to the protein class of the differentially regulated
proteins specific to each of the five subtypes of BC found in the ex vivo formed coronas
analyzed with the PANTHER database.
Immune cell activation was also shown to be an altered pathway in BC which was
enriched only for the LA subtype in the present study (Figure 4_SM). Immunoglobulin heavy
constant mu (IGHM) was down-regulated for B cell activation in LA, indicating that
antibody-mediated immune response was implicated in this subtype. Probably, the tumor
alters the immune system mechanism to suppress the B cell activation promoting this down-
regulation.
Complement activation is an important factor of innate immunity and a defense system
against infecting pathogens. Furthermore, complement activation also participates in the
adaptive immune response. Particularly in BC, complement activation contributes to cancer
progression [26]. In the present work, a total of 3 complement system components implicated
in the innate immune response were identified in the PC for the different BC subtypes:
complement component C8 beta chain (C8B) for the LA, complement C5 (C5) for the
HER2+, complement C2 (C2) for the TNBC (see Table 2). Previous works also found
deregulation of some of these complement system components in the sera of BC patients [27,
28].
Other deregulated proteins which may play a role in the innate immune system were
complement C1r subcomponent-like protein (C1RL) and complement factor H-related
protein 2 (CFHR2) (both up-regulated) in the LA subtype; complement C1s subcomponent

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
158
(C1S) (down-regulated) in the LB- subtype; plastin-2 (LCP1) (up-regulated) in the LB+
subtype; cysteine-rich secretory protein 3 (CRISP3) (up-regulated) and N-acetylmuramoyl-
L-alanine amidase (PGLYRP2) (down-regulated) in the HER2+ subtype; and keratin, type II
cytoskeletal 1 (KRT1) (up-regulated) in the TNBC subtype (see Table 2). Particularly, LCP1
plays a role in the activation of T-cells and its exosomal release by breast cancer cells was
found to facilitate metastatic bone osteolysis [29].
After the analysis of the protein corona, HER2+ subtype was enriched with coagulation
factor V (F5), coagulation factor XII (F12) and plasma kallikrein (KLKB1) (up-regulated),
and alpha-1-antitrypsin (SERPINA1), trypsin-1 (PRSS1) and antithrombin-III (SERPINC1)
(down-regulated) for blood coagulation pathway (Figure 4_SM, Tables 2 and 3).
Platelets, as small cell fragments, are not only important coagulation-related factors, also
play a vital role in tumor progression [30]. Particularly, platelet factor 4 (PF4) or CXCL4, a
member of CXC chemokine family, acts as an angiogenesis inhibitor which may contribute
to prevent tumor metastasis [31]. In the present work, it was observed that the inflammation
mediated by chemokine and cytokine signaling pathway was enriched specifically for LB-
subtype with the platelet factor 4 variant (PF4V1) being down-regulated (Figure 4_SM).
Other protein of the family of platelets is the platelet basic protein (PPBP) that was up-
regulated in the TNBC subtype (see Table 2).
The gonadotropin-releasing hormone receptor pathway was found to be enriched with
adiponectin (ADIPOQ) (up-regulated) and voltage-dependent L-type calcium channel
subunit alpha-1F (CACNA1F) (up-regulated) in the HER2+ and TNBC subtypes,
respectively (Figure 4_SM).
In the present work, a group of proteins implicated in the combination and transportation
of lipids, apoliporoteins, were also found to be deregulated in the LB- subtype
(apolipoprotein B-100 (APOB), apolipoprotein D (APOD) and phospholipid transfer protein
(PLTP); up-regulated), the HER2+ subtype (apolipoprotein F (APOF); down-regulated) and
TNBC subtype (apolipoprotein E (APOE); up-regulated) (see Table 2).
Other family of potential molecular targets that were found to be deregulated in the
present study are some glycoproteins as: lysosome-associated membrane glycoprotein 2
(LAMP2) (up-regulated) in the LA subtype; platelet glycoprotein Ib alpha chain (GP1BA)
and basement membrane-specific heparan sulfate proteoglycan core protein (HSPG2) (up-
regulated) in LB- subtype; EGF-containing fibulin-like extracellular matrix protein 1

III. Results and Discussion. CHAPTER 3
159
(EFEMP1) and selenoprotein P (SELENOP) (up-regulated) in the HER2+ subtype; and CD5
antigen-like (CD5L) (down-regulated) in the TNBC subtype (see Table 2).
While some proteins with an enzymatic functionality as biotinidase (BTD), serum
paraoxonase/lactonase 3 (PON3), L-lactate dehydrogenase B chain (LDHB) and alpha-
mannosidase 2 (MAN2A1) were found to be the up-regulated in LB- subtype, protein Z-
dependent protease inhibitor (SERPINA10) were down-regulated in the LA subtype (see
Table 2).
Table 3. Candidate deregulated blood coagulation biomarkers in HER2-overexpressing BC
patients found after the proteomic analysis of the ex vivo corona-coated AuNPs. On the
bottom, a cluster of blood coagulation proteins found in the protein-protein interaction
network map of the genes encoded differentially regulated proteins for the HER2-
overexpressing BC patients found after the proteomic analysis of the ex vivo corona-coated
AuNPs.
Protein Name Gene p-value Fold Change Control vs. HER2 Positive
Coagulation factor XII F12 2.50E-06 4.48323521 ↑ HER2 Pos
Plasma kallikrein KLKB1 2.05E-05 2.839283512 ↑ HER2 Pos
Coagulation factor V F5 0.025887503 3.739380413 ↑ HER2 Pos
Alpha-1-antitrypsin SERPINA1 0.001111283 4.959761437 ↓ HER2 Pos
Trypsin-1 PRSS1 0.002454431 4.217786063 ↓ HER2 Pos
Antithrombin-III SERPINC1 0.018612975 1.308597079 ↓ HER2 Pos
3.4. Discussion
Nowadays, the different breast cancer intrinsic subtypes (LA, LB-, LB+, HER2+,
TNBC) guide the therapy selection [2]. In some patients, therapies could fail for different
reasons as cancer recurrence, therapy resistance, and/or metastasis. Thus, therapy response
in breast cancer patients could be improved with the study of the molecular alterations at the
subtype level.
The application of different omics approaches to deep insight the different BC subtypes
allowed the refinement of the complexity of tumor heterogeneity. In this way, a variety of

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
160
quantitative proteomic studies were developed to identify potential signatures for breast
cancer using clinical samples such as saliva, the ductal lavage fluids, nipple aspirate fluids
(NAFs), urine and tissue [32]. Importantly, blood may be a suitable sample source for
studying the proteomic deregulation in the different BC subtypes with minimally invasive
collection procedures. Thus, the analysis of blood-based markers at the subtype level in
biological fluids such as plasma and serum allowed to find different protein markers related
with the tumor microenvironment and the subtype-specific changes. Following this research
line, the proteomic alterations in blood plasma [7] and blood serum [8] of BC subtypes were
explored.
Potential biomarkers are presented in very low concentrations in blood (less than 1% of
blood proteins). Thus, the isolation and enrichment of low-abundance peptides/proteins from
complex mixtures is a mandatory step in the proteomic biomarkers pipeline and nanoparticles
represent an ideal alternative [33].
In the present work, an exhaustive quantitative analysis of the PCs formed around AuNPs
after their incubation in serum samples was developed by SWATH-MS to identify novel
molecular targets associated to the different BC intrinsic subtypes (see Figure 1).
Seven proteins were found to be commonly altered in all BC subtypes, namely APOC3,
CRP, HBB, IGHV3-49, SAA4, APCS and TR. CRP and SAA4 are acute-phase proteins
(APPs), a class of proteins whose serum concentrations increase or decrease in response to
inflammation. Particularly, a significant association of state of inflammation with stage of
BC was previously described [34]. A recent study found that elevated serum levels of CRP
were associated considerably with a high risk of BC and poor outcome, including metastasis
and recurrence [35]. In addition, the concentrations of SAA4 increased gradually with tumor
progression and the severity of BC stages [36]. Thus, CRP and SAA4 may be good candidate
markers for the staging and prognosis of BC. The expression of HBB and TR, members of
the globin family, was also found to be associated with BC cells aggressiveness and poor
prognosis, indicating to HBB and TR as novel biomarker for BC progression [37, 38].
On the other hand, several studies point out that blood coagulation proteins develop an
important role in tumor progression [39]. These works discussed the impact of the activation
of the blood clotting cascade on primary tumor growth [40], on tumor metastasis and cancer-
associated thrombosis [41]; and antitumor therapies that target blood-coagulation-associated
proteins [42].

III. Results and Discussion. CHAPTER 3
161
In the case of BC, different reports support blood coagulation proteins as an important
patient factor that facilitates the metastatic potential [43]. Particularly, metastatic patients
exhibited significantly higher D-dimer values when compared with early breast cancer
patients [44]. Furthermore, high plasma fibrinogen was found to be correlated with poor
response to trastuzumab treatment in HER2 positive BC patients [45] and circulating levels
of factor VIII (FVIII) were significantly associated with axillary lymph node involvement,
number of metastatic nodes, and HER2 status [ 46 ]. These studies support that the
measurement of some coagulation-related biomarkers could provide additional data for the
evaluation of HER2 positive BC patients' prognosis and could be novel molecular targets.
The present quantitative proteomic analysis revealed a profile of blood coagulation
proteins for the HER2+ subtype, namely F5, F12 and KLKB1 (up-regulated), and
SERPINA1, PRSS1 and SERPINC1) (down-regulated). While F5 is expressed in tumors and
indicates favorable outcome in aggressive BC [47], F12 is involved in the pathogenesis of
thrombosis through the induction and amplification of thrombin generation [48].
KLKB1 (up), SERPINA1 (down) and SERPINC1 (down) are serine proteases.
Particularly, SERPINA1 and SERPINC1 are serine proteases inhibitors (serpins) which
belong to the protease inhibitor family. Members of the kallikrein family, as KLKB, were
also found to be deregulated during malignant transformation [ 49 ]. Nevertheless, the
variations in expression (downregulation/up-regulation), activation and secretion are not
substantially to consider them as suitable biomarkers for follow-up disease progression.
SERPINA1 is synthesized and released by tumor cells and plays major roles in physiologic
and pathologic processes such as angiogenesis, tumor invasion and metastasis [50]. It was
found that that high expression of SERPINA1 could be predictive for a better clinical
outcome of ER+ and ER+/HER2+ patients. Thus, SERPINA1 was found to be a direct ER
target gene and a predictor of survival in BC patients [51]. SERPINC1, an antithrombin,
develops an important role as an inhibitor of the coagulation cascade. Furthermore,
SERPINC1 also functions as an anti-angiogenic, anti-inflammatory, anti-viral and anti-
apoptotic protein. The mechanism by which antithrombin controls invasion, tumor migration,
and angiogenesis is by inhibition of enteropeptidase. This inhibition showed to be a double
anti-tumor effect through producing an anti-angiogenic molecule and inhibiting a protease
implicated in metastasis [52].
In the present work, the gonadotropin-releasing hormone receptor pathway was found to
be enriched with adiponectin (ADIPOQ) in the HER2+ subtype. Although different studies

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
162
reported controversial findings in the association between ADIPOQ and BC, a recent meta-
analysis suggests that low serum adiponectin concentration may be associated with an
increased BC risk in premenopausal and postmenopausal women [53]. A negative correlation
has been also demonstrated between ADIPOQ levels and tumor size and grade.
Interestingly, the correlation between ADIPOQ and BC seems to be more prominent in
oestrogen-negative and progesterone-negative BC [54]. Therefore, it seems there may be a
set group of BC patients that are most susceptible to the effects of ADIPOQ and would benefit
most from a potential treatment. ADIPOQ may serve as a biomarker of BC risk and help to
identify subjects at high risk for BC development.
Numerous research articles have accumulated solid evidence that lipoproteins are closely
related to various types of tumorigenesis, as BC [55]. Apolipoproteins in the blood transfer
lipids to cancer cells to provide energy for cancer cell proliferation and invasion.
Apolipoproteins also function as important factors in cellular signal transduction. In the
present work, different apoliporoteins were found to be deregulated in the different BC
subtypes. Particularly, APOB, APOD and APOE in serum were found to function as a risk
factor for BC, being APOD and APOB involved in BC metastasis [56]. Particularly, a recent
study found that apolipoprotein B is a risk factor for development of intraocular metastasis
(IOM) in patients with BC [57].
Within the group of glycoproteins found to be deregulated in the present study, it was
found that LAMP2 overexpression in breast tumors promotes cancer cell survival via
chaperone-mediated autophagy (CMA) [58]. Thus, inhibiting CMA activity in breast tumor
cells (with a chemotherapeutic drug, for example) can be exploited as a potential therapeutic
application in the treatment of BC. HSPG2, also known as perlecan, is a heavily glycosylated
protein component of the extra-cellular matrix (ECM) that plays essential roles in tumor
vascularization, that is closely related to tumor growth and metastasis [59]. Although HSPG2
expression in BC has not been examined in detail, a recent study investigated the expression
of HSPG2 in human TNBC and the ability of anti-HSPG2 antibodies to specifically target
and inhibit tumor growth in a mouse xenograft model [60], showing that HSPG2 is a
promising therapeutic target in TNBC. EFEMP1, also knon as fibulin 3, may have a potential
cancer-promoting function in BC [ 61 ]. EFEMP1 expression decreases during BC
progression, with low EFEMP1 levels correlating with a poorer prognosis. Functionally, high
EFEMP1 levels inhibited TGF-β-induced EMT, migration, invasion, and endothelial
permeability, while loss of EFEMP1 expression/function promoted these TGF-β-mediated

III. Results and Discussion. CHAPTER 3
163
effects. Further, restoring EFEMP1 expression in breast cancer cells inhibited TGF-β
signaling, breast cancer cell EMT, invasion and metastasis in vivo. Although the role of
CD5L in the oncogenesis of BC is not fully understood, a recent work found that CD5L is
upregulated in hepatocellular carcinoma and promotes liver cancer cell proliferation and
antiapoptotic responses [62].
The enzyme LDHB, that was found to be up-regutaled in the LB- subtype, may help
identify breast cancers most likely to respond to neoadjuvant chemotherapy as well as those
with the highest risk of relapse that may benefit from additional adjuvant therapy [63].
All these novel molecular targets found in the serum of BC patients could detect a
missing invasion, they could be performed in ambulatory settings, they could be repeatedly
checked, and they could be applicable for BC diagnosis, the assessment of prognosis and
selection of treatment.
Importantly, further insights exploring the deregulated blood coagulation proteins as
potential effective prognosis biomarkers and targets for novel therapeutic approaches could
have a great impact in the management of HER2-overexpressing BC patients.
4. Conclusions
The quantitative comparison of the ex vivo PCs formed upon incubation of AuNPs with
serum samples obtained from BC patients revealed 75 deregulated subtype-specific unique
proteins (8, 27, 2, 28 and 10 proteins specifically associated to the LA, LB-, LB+, HER2+
and TNBC subtypes). The analysis of the ex vivo PCs formed onto AuNPs reveals a profile
of blood coagulation proteins in the serum of HER2-overexpressing BC patients, that are
implicated in breast tumor progression, including cellular transformation, proliferation,
tumor cell survival, and angiogenesis. Of all BC patients, HER2+ patients have a worse
outcome. Further insights exploring these blood coagulation proteins as potential effective
prognosis biomarkers and targets for novel therapeutic approaches could have a great impact
on the management of HER2-overexpressing BC patients.

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
164
5. References
[1] Y. Feng, M. Spezia, S. Huang, C. Yuan, Z. Zeng, L. Zhang, X. Ji, W. Liu, B. Huang,
W. Luo, B. Liu, Y. Lei, S. Du, A. Vuppalapati, H.H. Luu, R.C. Haydon, T. He, G. Ren,
Breast cancer development and progression: Risk factors, cancer stem cells, signaling
pathways, genomics, and molecular pathogenesis, Genes Dis. 5 (2018) 77-106.
[2] Z. Inic, M. Zegarac, M. Inic, I. Markovic, Z. Kozomara, I. Djurisic, I. Inic, G. Pupic,
S. Jancic, Difference between Luminal A and Luminal B Subtypes According to Ki-67,
Tumor Size, and Progesterone Receptor Negativity Providing Prognostic Information,
Clin Med Insights Oncol. 8 (2014) 107-111.
[3] M. Colleoni, Z. Sun, K.N. Price, P. Karlsson, J.F. Forbes, B. Thürlimann, L. Gianni,
M. Castiglione, R.D. Gelber, A.S. Coates, A. Goldhirsch, Annual Hazard Rates of
Recurrence for Breast Cancer During 24 Years of Follow-Up: Results From the
International Breast Cancer Study Group Trials I to V, J Clin Oncol. 34 (2016) 927-935.
[4] S. Loibl, L. Gianni, HER2-positive breast cancer, Lancet 389 (2017) 2415-2429.
[5] H. Asif, S. Sultana, S. Ahmed, N. Akhtar, M. Tariq, HER-2 Positive Breast Cancer -
a Mini-Review, Asian Pac J Cancer Prev. 17 (2016) 1609-1615.
[6] A.C. Wolff, M.E.H. Hammond, K.H. Allison, B.E. Harvey, P.B. Mangu, J.M.S.
Bartlett, M. Bilous, I.O. Ellis, P. Fitzgibbons, W. Hanna, R.B. Jenkins, M.F. Press, P.A.
Spears, G.H. Vance, G. Viale, L.M. McShane, M. Dowsett, Human epidermal growth
factor receptor 2 testing in breast cancer: American society of clinical oncology/college
of american pathologists clinical practice guideline focused update, J Clin Oncol 36
(2018) 2105-2122.
[7] S. Suman, T. Basak, P. Gupta, S. Mishra, V. Kumar, S. Sengupta, Y. Shukla,
Quantitative Proteomics revealed novel proteins associated with molecular subtypes of
breast cancer, J Proteomics 418 (2016) 183-193.

III. Results and Discussion. CHAPTER 3
165
[8] A. Gajbhiye, R. Dabhi, K. Taunk, M.G. Jagadeeshaprasad, S. RoyChoudhury, A.
Mane, S. Bayatigeri, K. Chaudhury, M.K. Santra, S. Rapole, Multipronged
quantitative proteomics reveals serum proteome alterations in breast cancer intrinsic
subtypes, J. Proteomics 163 (2017) 1-13.
[9] P. Bouchal, O.T. Schubert, J. Faktor, L. Capkova, H. Imrichova, K. Zoufalova, V.
Paralova, R. Hrstka, Y. Liu, H.A. Ebhardt, E. Budinska, R. Nenutil, R. Aebersold, Breast
Cancer Classification Based on Proteotypes Obtained by SWATH Mass Spectrometry,
Cell Rep. 28 (2019) 832-843.
[10] L. Jia, Y. Lu, J. Shao, X.J. Liang, Y. Xu, Nanoproteomics: a new sprout from
emerging links between nanotechnology and proteomics, Trends Biotechnol. 31 (2013)
99-107.
[ 11 ] D. Pozzi, G. Caracciolo, L. Digiacomo, V. Colapicchioni, S. Palchetti, A.L.
Capriotti, C. Cavaliere, R. Zenezini Chiozzi, A. Puglisi, A. Laganà, The biomolecular
corona of nanoparticles in circulating biological media, Nanoscale 7 (2015) 13958-
13966.
[12] M.P. Chantada-Vázquez, A. Castro López, S.B. Bravo, S. Vázquez-Estévez, B.
Acea-Nebril, C. Núñez, Proteomic analysis of the bio-corona formed on the surface of
(Au, Ag, Pt)-nanoparticles in human serum, Colloids Surf B Biointerfaces 177 (2019)
141-148.
[13] M.P. Chantada-Vázquez, A. Castro López, M. García Vence, S. Vázquez-Estévez,
B. Acea-Nebril, D.G. Calatayud, T. Jardiel, S.B. Bravo, C. Núñez, Proteomic
investigation on bio-corona of Au, Ag and Fe nanoparticles for the discovery of triple
negative breast cancer serum protein biomarkers, J. Proteomics 212 (2020) 103581.
[14] M.P. Chantada-Vázquez, M. García-Vence, S. Vázquez-Estévez, S.B. Bravo, C.
Núñez, Identification of a Profile of Neutrophil-Derived Granule Proteins in the Surface
of Gold Nanoparticles after Their Interaction with Human Breast Cancer Sera,
Nanomaterials (Basel) 10(6) (2020) 1223.

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
166
[15] D. Docter, D. Westmeier, M. Markiewicz, S. Stolte, S.K. Knauer, R.H. Stauber, The
nanoparticle biomolecule corona: lessons learned – challenge accepted?. Chem. Soc.
Rev. 44 (2015) 6094-6121.
[16] Z.W. Lai, Y. Yan, F. Caruso, E.C. Nice, Emerging techniques in proteomics for
probing nano-bio interactions, ACS Nano 6 (2012) 10438-10448.
[17] X. Bai, Y. Wang, Z. Song, Y. Feng, Y. Chen, D. Zhang, L. Feng, The Basic
Properties of Gold Nanoparticles and their Applications in Tumor Diagnosis and
Treatment, Int J Mol Sci 21(7) (2020) 2480.
[18] E. Oliveira, J.E. Araújo, S. Gómez-Meire, C. Lodeiro, C. Pérez-Melón, E. Iglesias-
Lamas, A. Otero-Glez, J.L. Capelo, H.M. Santos, Proteomics analysis of the peritoneal
dialysate effluent reveals the presence of calcium-regulation proteins and acute
inflammatory response, Clin Proteomics 11 (2014) 17.
[19] D. Szklarczyk, A. Franceschini, S. Wyder, K. Forslund, D. Heller, J. Huerta-Cepas,
M. Simonovic, A. Roth, A. Santos, K.P. Tsafou, M. Kuhn, P. Bork, L.J. Jensen, C. von
Mering, STRING v10: protein-protein interaction networks, integrated over the tree of
life, Nucleic Acids Res. 43(Database issue) (2015) D447-D452.
[20] M.A. Dobrovolskaia, A.K. Patri, J. Zheng, J.D. Clogston, N. Ayub, P. Aggarwal,
B.W. Neun, J.B. Hall, S.E. McNeil, Interaction of colloidal gold nanoparticles with
human blood: effects on particle size and analysis of plasma protein binding profiles,
Nanomedicine 5 (2009) 106-117.
[21] R. García-Álvarez, M. Hadjidemetriou, A. Sánchez-Iglesias, L.M. Liz-Marzán, K.
Kostarelos, In vivo formation of protein corona on gold nanoparticles. The effect of size
and shape, Nanoscale 10(3) (2018) 1256-1264.
[22] H. Mi, A. Muruganujan, P.D. Thomas, PANTHER in 2013: modeling the evolution
of gene function, and other gene attributes, in the context of phylogenetic trees, Nucleic
Acids Res. 41 (2013) D377-D386.

III. Results and Discussion. CHAPTER 3
167
[23] L.M. Coussens, W.W. Raymond, G. Bergers, Inflammatory mast cells up-regulate
angiogenesis during squamous epithelial carcinogenesis, Genes Dev. 13 (1999) 1382-
1397.
[24] B. Singh, J.A. Berry, A. Shoher, A. Lucci, COX-2 induces IL-11 production in
human breast cancer cells, J Surg Res. 131 (2006) 267-275.
[25] R.P. Singh, V.P. Singh, K.N. Udupa, E-Rosette forming lymphocytes and serum
immunoglobulins in breast cancer patients, Mater Med Pol. 23 (1991) 179-181.
[26] Y. Lu, X. Hu, C5a stimulates the proliferation of breast cancer cells via Akt-
dependent RGC-32 gene activation, Oncol Rep. 32 (2014) 2817-2823.
[27] F. Nasim, S. Ejaz, M. Ashraf, A.R. Asif, M. Oellerich, G. Ahmad, G.A. Malik, Attiq-
Ur-Rehman, Potential Biomarkers in the Sera of Breast Cancer Patients from
Bahawalpur, Pakistan, Biomark Cancer 4 (2012) 19-34.
[28] A. Liu, H. Qu, C. Yu, J. Liu, A. Jiao, P. Sun, Serum biomarkers for lymph node
metastasis in patients with triple-negative breast cancer by proteomics, Int J Clin Exp
Pathol. 10 (2017) 3377-3384.
[29] K. Tiedemann, G. Sadvakassova, N. Mikolajewicz, M. Juhas, Z. Sabirova, S.
Tabariès, J. Gettemans, P.M. Siegel, S.V. Komarova, Exosomal Release of L-Plastin by
Breast Cancer Cells Facilitates Metastatic Bone Osteolysis, Transl Oncol. 12(3) (2019)
462-474.
[30] S. Ishikawa, T. Miyashita, M. Inokuchi, H. Hayashi, K. Oyama, H. Tajima, H.
Takamura, I. Ninomiya, A.K. Ahmed, J.W. Harman, S. Fushida, T. Ohta, Platelets
surrounding primary tumor cells are related to chemoresistance, Oncol Rep. 36(2) (2016)
787.
[ 31 ] A. Goodarzi, F. Yari, M. Mohammadipour, M.R. Deyhim, H.T. Naghadeh,
Capability of Platelet Factor 4 to Induce Apoptosis in the Cancerous Cell Lines in Vitro,
IJML 5(3) (2018) 195-207.

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
168
[32] N. Pendharkar, A. Gajbhiye, K. Taunk, S. RoyChoudhury, S. Dhali, S. Seal, A.
Mane, S. Abhang, M.K. Santra, K. Chaudhury, S. Rapole, Quantitative tissue proteomic
investigation of invasive ductal carcinoma of breast with luminal B HER2 positive and
HER2 enriched subtypes towards potential diagnostic and therapeutic biomarkers, J
Proteome 132 (2016) 112-130.
[33] L. Zhang, H. Lu, P. Yang, Recent developments of nanoparticle-based enrichment
methods for mass spectrometric analysis in proteomic, Sci China Chem. 53 (2010) 695-
703.
[34] S.B. Asegaonkar, U.V. Takalkar, P. Kodlikeri, A. Pagdhune, V. Bonduliya, A.P.
Thorat, Serum high sensitivity C-reactive protein in breast cancer patients, Int J Res Med
Sci. 2 (2014) 1408-1411.
[35] R.P. Kaur, Rubal, R.P.S. Banipal, R. Vashistha, M. Dhiman, A. Munshi, Association
of elevated levels of C-reactive protein with breast cancer, breast cancer subtypes and
poor outcome, Curr Probl Cancer 43 (2019) 123-129.
[36] G. Zhang, X. Sun, H. Lv, X. Yang, X. Kang, Serum amyloid A: A new potential
serum marker correlated with the stage of breast cancer, Oncol Lett. 3 (2012) 940-944.
[37] M. Ponzetti, M. Capulli, A. Angelucci, L. Ventura, S.D. Monache, C. Mercurio, A.
Calgani, P. Sanita, A. Teti, N. Rucci, Non-conventional role of haemoglobin beta in
breast malignancy, Br J Cancer 117 (2017) 994-1006.
[38] B. Vandewalle, L. Hornez, F. Revillion, J. Lefebvre, Secretion of transferrin by
human breast cancer cells, Biochem Biophys Res Commun 163(1) (1989) 149-154.
[39] L.G. Lima, R.Q. Monteiro, Activation of blood coagulation in cancer: implications
for tumour progression, Biosci Rep. 33 (2013) e00064.
[40] A. Metelli, B.X. Wu, B. Riesenberg, S. Guglietta, J.D. Huck, C. Mills, A. Li, S.
Rachidi, C. Krieg, M.P. Rubinstein, D.T. Gewirth, S. Sun, M.B. Lilly, A.H. Wahlquist,
D.P. Carbone, Y. Yang, B. Liu, Z. Li, Thrombin contributes to cancer immune evasion
via proteolysis of platelet-bound GARP to activate LTGF-β, Sci Transl Med 12 (2020)
eaay4860.

III. Results and Discussion. CHAPTER 3
169
[41] Y. Hisada, N. Mackman, Cancer-associated pathways and biomarkers of venous
thrombosis, Blood 130 (2017) 1499-1506.
[42] W. Ruf, A.S. Rothmeier, C. Graf, Targeting clotting proteins in cancer therapy -
progress and challenges, Thromb Res. 140 (2016) S1-S7.
[43] I. Lal, K. Dittus, C.E. Holmes, Platelets, coagulation and fibrinolysis in breast cancer
progression, Breast Cancer Res. 15 (2013) 207.
[44] F. Tas, L. Kilic, D. Duranyildiz, Coagulation tests show significant differences in
patients with breast cancer, Tumor Biol. 35 (2014) 5985-5992.
[45] Y.-L. Liu, Q. Lu, J.-W. Liang, Y. Xia, W. Zhang, B.-Q. Hu, F.-F. Shang, Y.-R. Ji, J.
Wang, Q. Wang, B. Liang, High Plasma Fibrinogen is Correlated With Poor Response to
Trastuzumab Treatment in HER2 Positive Breast Cancer, Medicine (Baltimore) 94(5)
(2015) e481.
[46] E. Yigit, G. Gönüllü, I. Yücel, M. Turgut, D. Erdem, B. Cakar, Relation between
hemostatic parameters and prognostic/predictive factors in breast cancer, Eur J Intern
Med. 19 (2008) 602-607.
[47] M. Tinholt, Ø. Garred, E. Borgen, E. Beraki, M. Sletten, K. Kleivi Sahlberg, P.M.
Sandset, N. Iversen, Coagulation factor V is expressed in tumors and predicts favorable
outcome in aggressive breast cancer, Thromb Res. 164 (2011) S183.
[48] N.J. Mutch, Emerging roles for factor XII in vivo, J Thromb Haemost. 9 (2011)
1355-1358.
[ 49 ] T. Kryza, M.L. Silva, D. Loessner, N. Heuzé-Vourc’h, J.A. Clements, The
kallikrein-related peptidase family: dysregulation and functions during cancer
progression, Biochimie 122 (2016) 283-299.
[50] I. Zelvyte, H.O. Sjögren, S. Janciauskiene, Effects of native and cleaved forms of
alpha1-antitrypsin on ME 1477 tumor cell functional activity, Cancer Detect Prev. 26
(2002) 256-265.

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
170
[51] H.J. Chan, H. Li, Z. Liu, Y.C. Yuan, J. Mortimer, S. Chen, SERPINA1 is a direct
estrogen receptor target gene and a predictor of survival in breast cancer patients,
Oncotarget 6 (2015) 25815-25827.
[52] G. Luengo-Gil, M.I. Calvo, E. Martín-Villar, S. Águila, N. Bohdan, A.I. Antón, S.
Espín, F.A. de la Peña, V. Vicente, J. Corral, M. Quintanilla, I. Martínez-Martínez,
Antithrombin controls tumor migration, invasion and angiogenesis by inhibition of
enteropeptidase, Sci Rep. 6 (2016) 27544.
[53] L. Gu, C. Cao, J. Fu, Q. Li, D.-H. Li, M.-Y. Chen, Serum adiponectin in breast
cancer. A meta-analysis, Medicine (Baltimore) 97(29) (2018) e11433.
[54] S.W. Oh, C.-Y. Park, E.S. Lee, Y.S. Yoon, E.S. Lee, S.S. Park, Y. Kim, N.J. Sung,
Y.H. Yun, K.S. Lee, H.S. Kang, Y. Kwon, J. Ro, Adipokines, insulin resistance,
metabolic syndrome, and breast cancer recurrence: a cohort study, Breast Cancer Res. 13
(2011) R34.
[55] Y. Zhou, G. Luo, Apolipoproteins, as the carrier proteins for lipids, are involved in
the development of breast cancer, Clin Transl Oncol. 22(11) (2020) 1952-1962.
[56] G.H. Cibeira, J. Giacomazzi, E. Aguiar, S. Schneider, B. Ettrich, C.I. De Souza, S.
Camey, M. Caleffi, B. Weber, P. Ashton-Prolla, E.H. Moriguchi, Apolipoprotein E
genetic polymorphism, serum lipoprotein levels and breast cancer risk: A case-control
study, Mol Clin Oncol. 2(6) (2014) 1009-1015.
[57] J.-X. Liu, Q. Yuan, Y.-L. Min, Y. He, Q.-H. Xu, B. Li, W.-Q. Shi, Q. Lin, Q.-H. Li,
P.-W. Zhu, Y. Shao, Apolipoprotein A1 and B as risk factors for development of
intraocular metastasis in patients with breast cancer, Cancer Manag Res. 11 (2019) 2881-
2888.
[58] T. Saha, LAMP2A overexpression in breast tumors promotes cancer cell survival
via chaperone-mediated autophagy, Autophagy 8(11) (2012) 1643-1656.
[ 59 ] J.M. Whitelock, L.D. Graham, J. Melrose, A.D. Murdoch, R.V. Iozzo, P.A.
Underwood, Human perlecan immunopurified from different endothelial cell sources has
different adhesive properties for vascular cells, Matrix Biol. 18 (1999) 163-178.

III. Results and Discussion. CHAPTER 3
171
[60] S. Kalscheuer, V. Khanna, H. Kim, S. Li, D. Sachdev, A. DeCarlo, D. Yang, J.
Panyam, Discovery of HSPG2 (Perlecan) as a Therapeutic Target in Triple Negative
Breast Cancer, Sci Rep. 9 (2019) 12492.
[61] B. Davidson, H.T. Stavnes, A. Holth, X. Chen, Y. Yang, I.-M. Shih, T.-L. Wang,
Gene expression signatures differentiate ovarian/peritoneal serous carcinoma from breast
carcinoma in effusions, J Cell Mol Med. 15(3) (2011) 535-544.
[62] G. Aran, L. Sanjurjo, C. Bárcena, M. Simon-Coma, E. Téllez, M. Vázquez-Vitali,
M. Garrido, L. Guerra, E. Díaz, I. Ojanguren, F. Elortza, R. Planas, M. Sala, C. Armengol,
M.-R. Sarrias, CD5L is upregulated in hepatocellular carcinoma and promotes liver
cancer cell proliferation and antiapoptotic responses by binding to HSPA5 (GRP78),
FASEB J. 32(7) (2018) 3878-3891.
[63] J.B. Dennison, J.R. Molina, S. Mitra, A.M. González-Angulo, J.M. Balko, M.G.
Kuba, M.E. Sanders, J.A. Pinto, H.L. Gómez, C.L. Arteaga, R.E. Brown, G.B. Mills,
Lactate Dehydrogenase B: A Metabolic Marker of Response to Neoadjuvant
Chemotherapy in Breast Cancer, Clin Cancer Res. 19(13) (2013) 3703-3713.

III. Results and Discussion. CHAPTER 3
_________________________________________________________________________
172

IV. CONCLUSIONS


IV. Conclusions
______________________________________________________________________
173
1.- The interaction of AuNPs (10.02 ± 0.91 nm and 12.96 ± 0.72 nm), AgNPs (9.73 ±
1.70 nm), PtNPs (2.40 ± 0.30 nm) and MNPs (9.30 ± 0.67 nm) with human serum
withstands the formation of a protein corona enveloping the nanoparticle, in all cases.
2.- The formation of this protein corona depends on the composition of the nanoparticle
(core material) and its size (showing that smaller NPs as PtNPs have lower protein
adsorption than larger NPs as AuNPs), the NPs/protein ratio, the sample pH and the
incubation time.
3.- After adjusting the pH value to 5.8 with citrate/citric acid buffer, the incubation time
to 30 min, and the protein/NPs ratio to 10.7, a total of 215, 215 and 198 proteins were
identified in the surface of AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and PtNPs
(2.40 ± 0.30 nm), respectively. From them, 170 proteins were commonly detected in the
protein corona of all three different types of NPs. 52 unique proteins were found on the
three different NPs surface: 21 unique proteins on the 10.02 ± 0.91 nm AuNPs, 17 on the
9.73 ± 1.70 nm AgNPs and 14 individual proteins on the 2.40 ± 0.30 nm PtNPs.
4.- The function of all identified proteins ranges from proteins implicated in the immune
response, followed by proteins with an enzymatic function, structural, transporter,
inflammatory, signal transduction and with antibiotic/antibacterial properties, being the
majority group the first one with 66 proteins identified implicated in the immune
response.
5.- The unique proteins identified on the corona of the three different NPs were proteins
with structural function and implicated in the signal transduction in the case of AuNPs
(10.02 ± 0.91 nm), proteins with antibiotic/antibacterial properties in the surface of
AgNPs (9.73 ± 1.70 nm) and proteins implicated on inflammatory processes in PtNPs
(2.40 ± 0.30 nm).
6.- The optimal conditions found for the formation of the protein corona around MNPs
(9.30 ± 0.67 nm) were the depletion with fresh DTT 500 mM in milli-Q for 60 min at
room temperature, a 1:2 ratio (MNP/protein) and a pH value of 5.5 in the incubation and
the final washing step.

IV. Conclusions
______________________________________________________________________
174
7.- The application of AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) MNPs (9.30 ±
0.67 nm), combined with mass spectrometry, has allowed the identification of seven
potential biomarkers for the diagnostic and control of TNBC progression: GRF-type zinc
finger domain-containing protein 1 (protein ZGRF1), matrix metalloproteinase-9
(MMP9), lebercilin and immunoglobulin lambda variable 3-27 (LV327) and LINE-1 type
transposase domain-containing protein 1 (LITD1), structural maintenance of
chromosomes protein 6 (SMC6) and short coiled-coil protein (SCOC).
8.- AuNPs (12.96 ± 0.72 nm) was an optimal scavenging tool in combination with
Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS) to
quantitatively analyze the serum proteome alterations in the different breast cancer
molecular subtypes (luminal A (LA), luminal B HER2 negative (LB-), luminal B HER2
positive (LB+), HER2 positive (HER2+) and triple negative breast cancer (TNBC)).
9.- The quantitative comparison of the ex vivo PCs formed upon incubation of AuNPs
with serum samples obtained from BC patients revealed 75 deregulated subtype-specific
unique proteins (8, 27, 2, 28 and 10 proteins specifically associated to the LA, LB-, LB+,
HER2+ and TNBC subtypes, respectively).
10.- The analysis of the ex vivo PCs formed onto AuNPs revealed a profile of blood
coagulation proteins in the serum of HER2-overexpressing BC patients that are
implicated in breast tumor progression, including cellular transformation, proliferation,
tumor cell survival and angiogenesis.

ANNEX A
Supplemental Material Chapter 1


175
Figure 1_SM. TEM image of AuNPs@citrate in aqueous phase and the characterization data.
AuNPs@citrate (nm) 1 7.94 21 10.35
2 8.37 22 10.38
3 8.38 23 10.41
4 8.60 24 10.44
5 8.78 25 10.56
6 8.78 26 10.65
7 8.87 27 10.65
8 9.17 28 10.68
9 9.19 29 10.80
10 9.41 30 10.81
11 9.49 31 10.83
12 9.49 32 10.85
13 9.55 33 10.86
14 9.62 34 10.86
15 9.82 35 10.88
16 9.84 36 10.95
17 9.89 37 10.96
18 9.89 38 10.99
19 9.99 39 11.48
20 10.03 40 11.50
Count 40
Mean 10.02
Minimum 7.94
Maximum 11.50
Standar Deviation 0.91
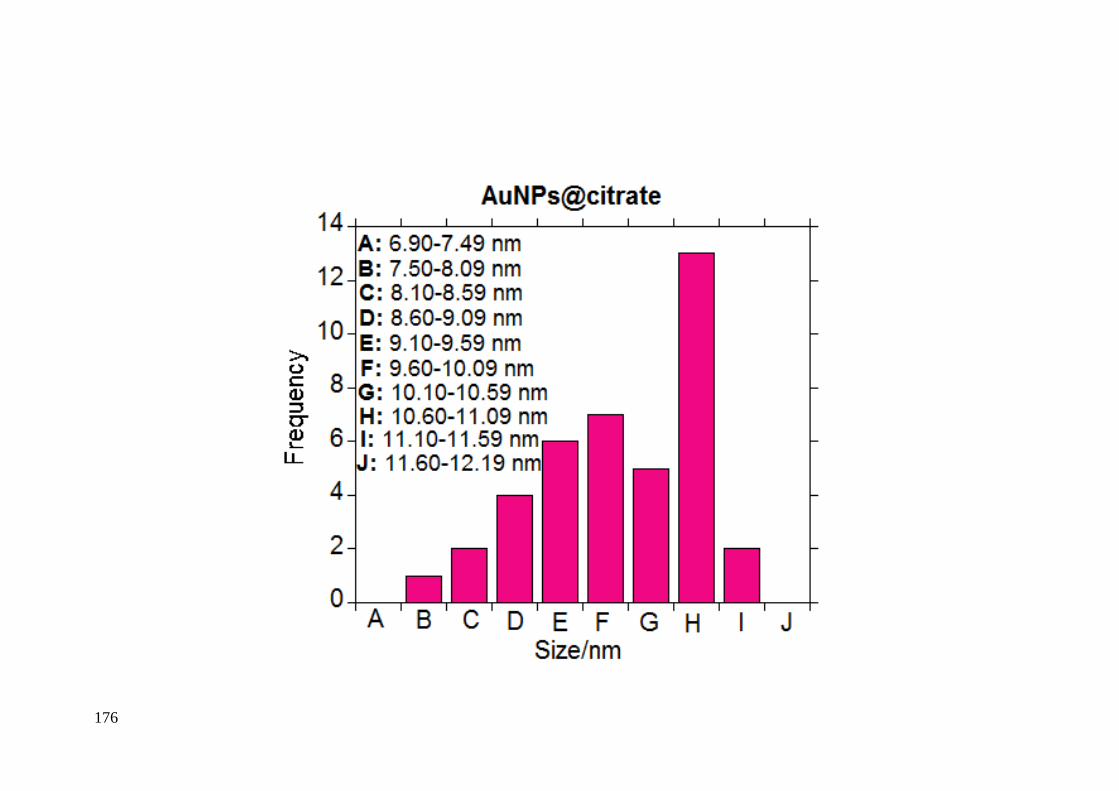
176

177

178
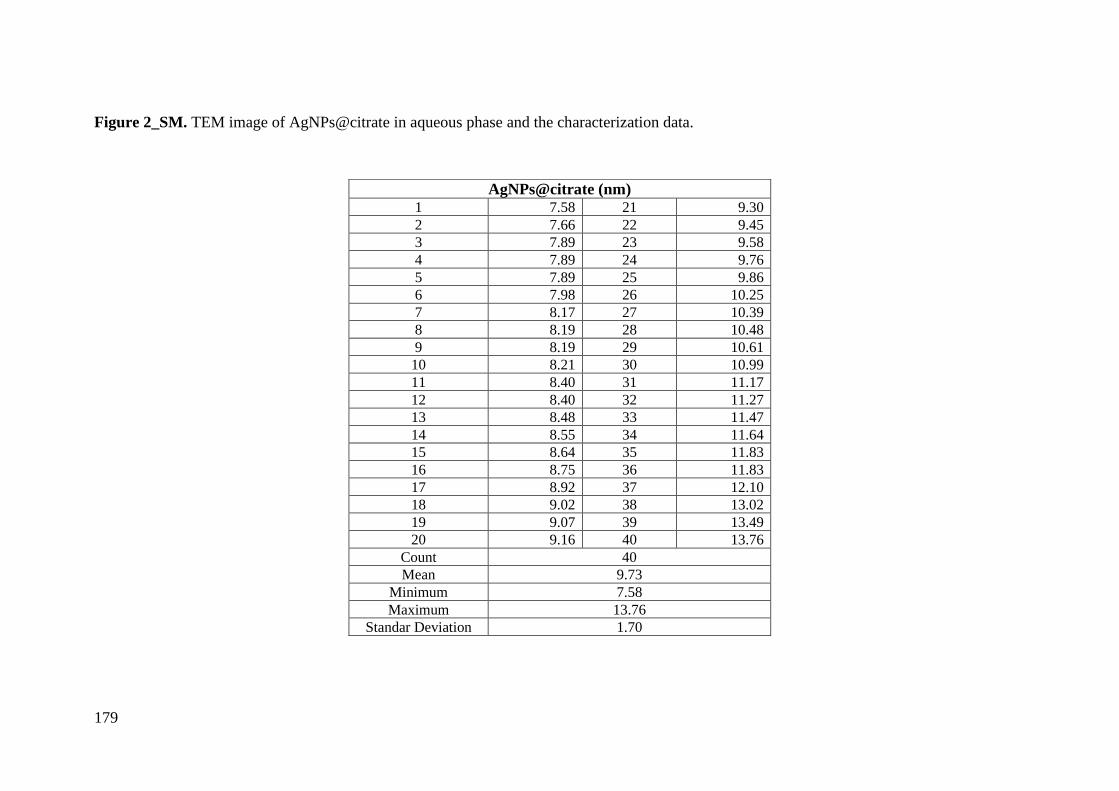
179
Figure 2_SM. TEM image of AgNPs@citrate in aqueous phase and the characterization data.
AgNPs@citrate (nm) 1 7.58 21 9.30
2 7.66 22 9.45
3 7.89 23 9.58
4 7.89 24 9.76
5 7.89 25 9.86
6 7.98 26 10.25
7 8.17 27 10.39
8 8.19 28 10.48
9 8.19 29 10.61
10 8.21 30 10.99
11 8.40 31 11.17
12 8.40 32 11.27
13 8.48 33 11.47
14 8.55 34 11.64
15 8.64 35 11.83
16 8.75 36 11.83
17 8.92 37 12.10
18 9.02 38 13.02
19 9.07 39 13.49
20 9.16 40 13.76
Count 40
Mean 9.73
Minimum 7.58
Maximum 13.76
Standar Deviation 1.70
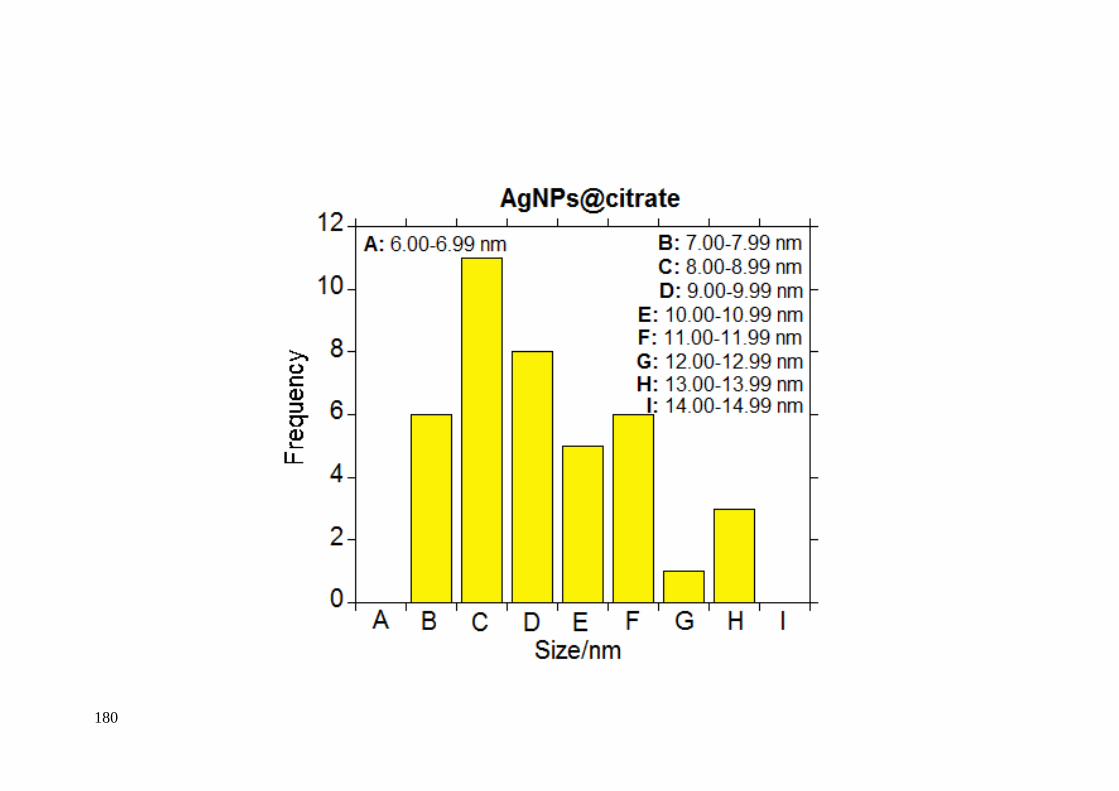
180

181

182
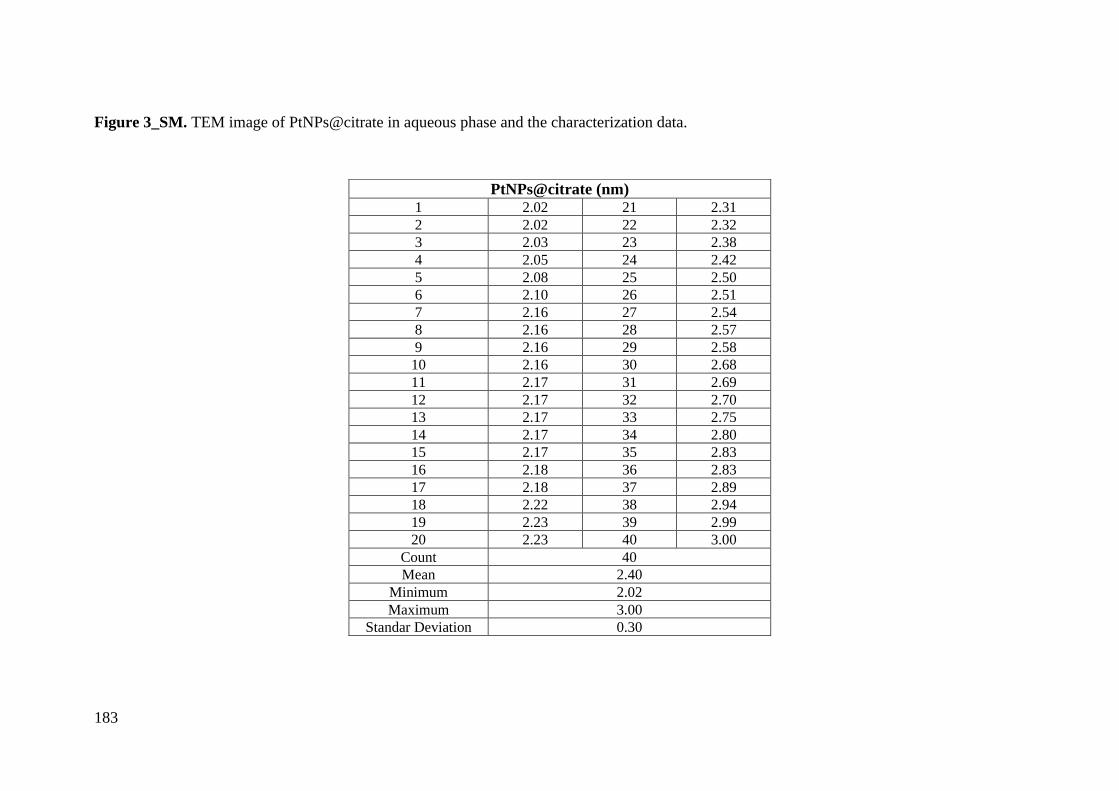
183
Figure 3_SM. TEM image of PtNPs@citrate in aqueous phase and the characterization data.
PtNPs@citrate (nm) 1 2.02 21 2.31
2 2.02 22 2.32
3 2.03 23 2.38
4 2.05 24 2.42
5 2.08 25 2.50
6 2.10 26 2.51
7 2.16 27 2.54
8 2.16 28 2.57
9 2.16 29 2.58
10 2.16 30 2.68
11 2.17 31 2.69
12 2.17 32 2.70
13 2.17 33 2.75
14 2.17 34 2.80
15 2.17 35 2.83
16 2.18 36 2.83
17 2.18 37 2.89
18 2.22 38 2.94
19 2.23 39 2.99
20 2.23 40 3.00
Count 40
Mean 2.40
Minimum 2.02
Maximum 3.00
Standar Deviation 0.30
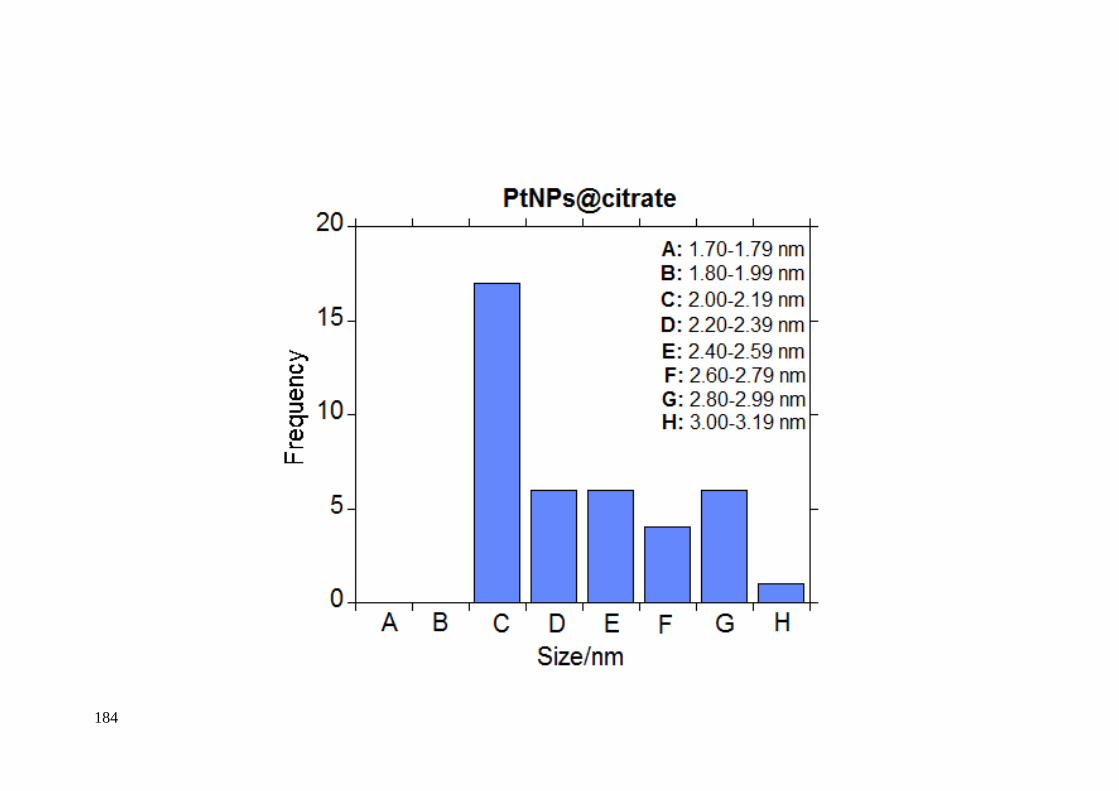
184

185

186
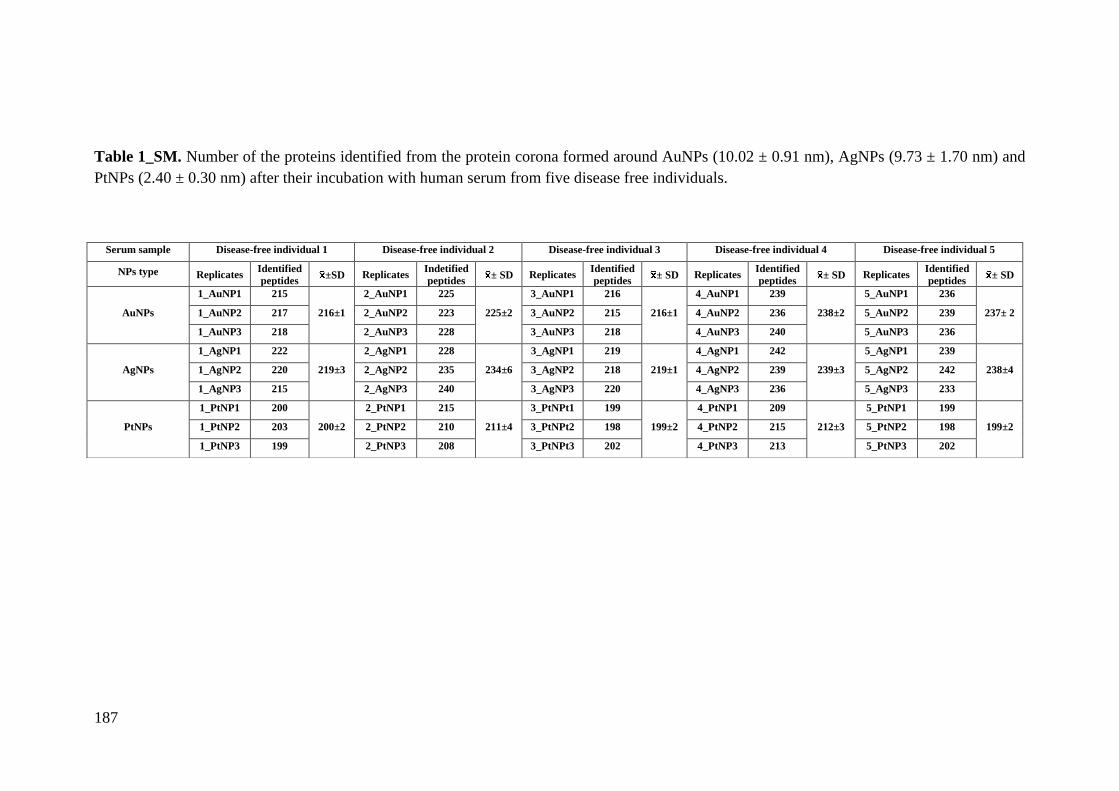
187
Table 1_SM. Number of the proteins identified from the protein corona formed around AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and
PtNPs (2.40 ± 0.30 nm) after their incubation with human serum from five disease free individuals.
Serum sample Disease-free individual 1 Disease-free individual 2 Disease-free individual 3 Disease-free individual 4 Disease-free individual 5
NPs type Replicates Identified
peptides ±SD Replicates
Indetified
peptides ± SD Replicates
Identified
peptides ± SD Replicates
Identified
peptides ± SD Replicates
Identified
peptides ± SD
AuNPs
1_AuNP1 215
216±1
2_AuNP1 225
225±2
3_AuNP1 216
216±1
4_AuNP1 239
238±2
5_AuNP1 236
237± 2 1_AuNP2 217 2_AuNP2 223 3_AuNP2 215 4_AuNP2 236 5_AuNP2 239
1_AuNP3 218 2_AuNP3 228 3_AuNP3 218 4_AuNP3 240 5_AuNP3 236
AgNPs
1_AgNP1 222
219±3
2_AgNP1 228
234±6
3_AgNP1 219
219±1
4_AgNP1 242
239±3
5_AgNP1 239
238±4 1_AgNP2 220 2_AgNP2 235 3_AgNP2 218 4_AgNP2 239 5_AgNP2 242
1_AgNP3 215 2_AgNP3 240 3_AgNP3 220 4_AgNP3 236 5_AgNP3 233
PtNPs
1_PtNP1 200
200±2
2_PtNP1 215
211±4
3_PtNPt1 199
199±2
4_PtNP1 209
212±3
5_PtNP1 199
199±2 1_PtNP2 203 2_PtNP2 210 3_PtNPt2 198 4_PtNP2 215 5_PtNP2 198
1_PtNP3 199 2_PtNP3 208 3_PtNPt3 202 4_PtNP3 213 5_PtNP3 202

188
Table 2_SM. Analysis of the commonly protein corona formed around AuNPs (10.02 ± 0.91 nm). AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ±
0.30 nm) after their incubation with serum aliquots (x3) from five different individuals (9 protein samples per individual: 3 treated with AuNPs. 3
with AgNPs and 3 with PtNPs). The accession number. gene name. species (Human). molecular weight and protein function were reported. Grey
color: 170 proteins commonly found in the protein corona of AuNPs (10.02 ± 0.91 nm). AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm).
Pink color: 21 identified corona proteins exclusively bound to the AuNPs (10.02 ± 0.91 nm). Yellow color: 17 identified corona proteins
exclusively bound to the AgNPs (9.73 ± 1.70 nm). Blue: 14 identified corona proteins exclusively bound to the PtNPs (2.40 ± 0.30 nm). Orange
color: 19 identified corona proteins commonly bound to the AuNPs (10.02 ± 0.91 nm) and AgNPs (9.73 ± 1.70 nm). Violet color: 5 identified
corona proteins commonly bound to the AuNPs (10.02 ± 0.91 nm) and PtNPs (2.40 ± 0.30 nm). Green color: 9 identified corona proteins
commonly bound to the AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm).
Protein Name UniProt Name Entry Name Gene AuNPs AgNPs PtNPs Mass (kDa) Function Actin. cytoplasmic 2 ACTG_HUMAN P63261 ACTG1 X X X 41.7 Cell mobility
Adipocyte plasma membrane-
associated protein APMAP_HUMAN Q9HDC9 APMAP X X X
46.5
Catalytic activity
Afamin AFAM_HUMAN P43652 AFM X X X 69.1 Transporter activity
Alpha-1-acid glycoprotein 1 A1AG1_HUMAN P02763 ORM1 X X X 23.5 Transporter activity
Alpha-1-acid glycoprotein 2 A1AG2_HUMAN P19652 ORM2 X X X 23.6 Transporter activity
Alpha-1-antichymotrypsin AACT_HUMAN P01011 SERPINA3 X X X 47.6 Protease ihibitor
Alpha-1-antitrypsin A1AT_HUMAN P01009 SERPINA1 X X X 46.7 Protease ihibitor
Alpha-1B-glycoprotein A1BG_HUMAN P04217 A1BG X X X 54.3 Neutrophil and platelet degranulation
Alpha-2-antiplasmin A2AP_HUMAN P08697 SERPINF2 X X X 54.6 Protease inhibitor
Alpha-2-HS-glycoprotein FETUA_HUMAN P02765 AHSG X X X 39.3
Promotes endocitosis. brain
development and the formation of
bone tissue
Alpha-2-macroglobulin A2MG_HUMAN P01023 A2M X X X 163.3 Immune response
Angiotensin-converting enzyme ACE_HUMAN P12821 ACE X 149.7 Catalytic activity
Angiotensinogen ANGT_HUMAN P01019 AGT X X X 53.1 Regulator of blood pressure. body
fluid and electrolyte homeostasis
Antithrombin-III ANT3_HUMAN P01008 SERPINC1 X X X 52.6 Protease inhibitor
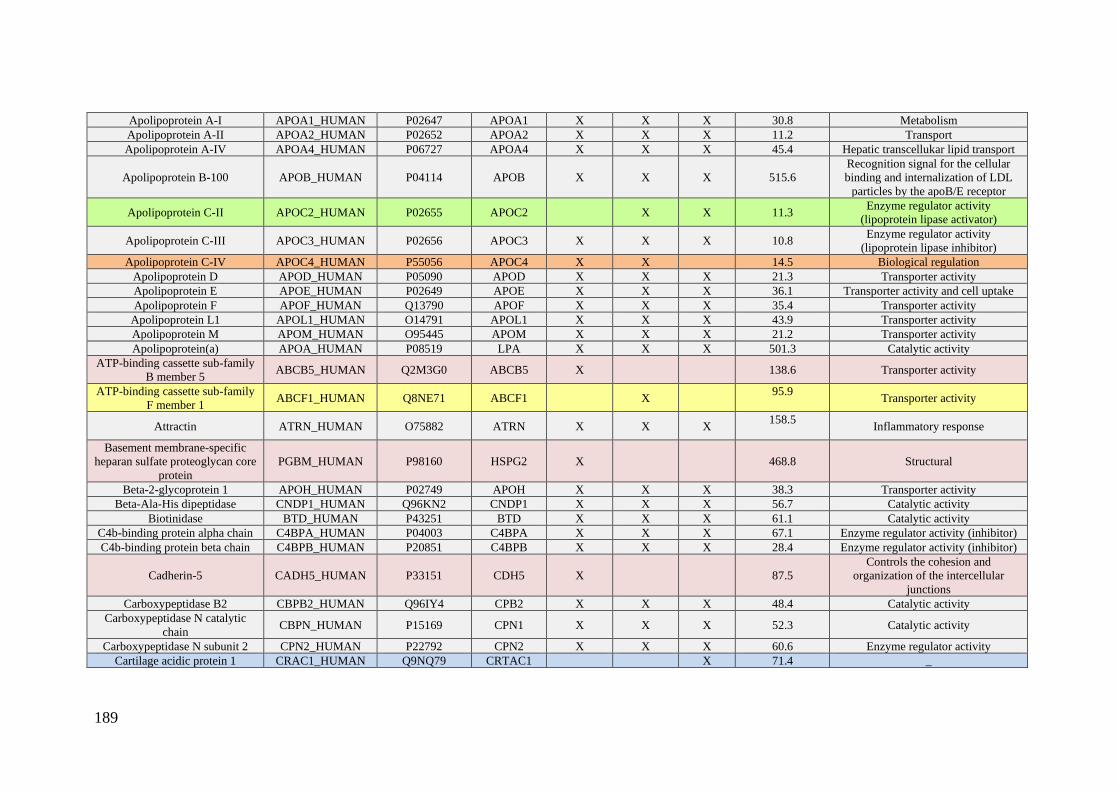
189
Apolipoprotein A-I APOA1_HUMAN P02647 APOA1 X X X 30.8 Metabolism
Apolipoprotein A-II APOA2_HUMAN P02652 APOA2 X X X 11.2 Transport
Apolipoprotein A-IV APOA4_HUMAN P06727 APOA4 X X X 45.4 Hepatic transcellukar lipid transport
Apolipoprotein B-100 APOB_HUMAN P04114 APOB X X X 515.6
Recognition signal for the cellular
binding and internalization of LDL
particles by the apoB/E receptor
Apolipoprotein C-II APOC2_HUMAN P02655 APOC2 X X 11.3 Enzyme regulator activity
(lipoprotein lipase activator)
Apolipoprotein C-III APOC3_HUMAN P02656 APOC3 X X X 10.8 Enzyme regulator activity
(lipoprotein lipase inhibitor)
Apolipoprotein C-IV APOC4_HUMAN P55056 APOC4 X X 14.5 Biological regulation
Apolipoprotein D APOD_HUMAN P05090 APOD X X X 21.3 Transporter activity
Apolipoprotein E APOE_HUMAN P02649 APOE X X X 36.1 Transporter activity and cell uptake
Apolipoprotein F APOF_HUMAN Q13790 APOF X X X 35.4 Transporter activity
Apolipoprotein L1 APOL1_HUMAN O14791 APOL1 X X X 43.9 Transporter activity
Apolipoprotein M APOM_HUMAN O95445 APOM X X X 21.2 Transporter activity
Apolipoprotein(a) APOA_HUMAN P08519 LPA X X X 501.3 Catalytic activity
ATP-binding cassette sub-family
B member 5 ABCB5_HUMAN Q2M3G0 ABCB5 X 138.6 Transporter activity
ATP-binding cassette sub-family
F member 1 ABCF1_HUMAN Q8NE71 ABCF1 X
95.9
Transporter activity
Attractin ATRN_HUMAN O75882 ATRN X X X 158.5
Inflammatory response
Basement membrane-specific
heparan sulfate proteoglycan core
protein
PGBM_HUMAN P98160 HSPG2 X 468.8 Structural
Beta-2-glycoprotein 1 APOH_HUMAN P02749 APOH X X X 38.3 Transporter activity
Beta-Ala-His dipeptidase CNDP1_HUMAN Q96KN2 CNDP1 X X X 56.7 Catalytic activity
Biotinidase BTD_HUMAN P43251 BTD X X X 61.1 Catalytic activity
C4b-binding protein alpha chain C4BPA_HUMAN P04003 C4BPA X X X 67.1 Enzyme regulator activity (inhibitor)
C4b-binding protein beta chain C4BPB_HUMAN P20851 C4BPB X X X 28.4 Enzyme regulator activity (inhibitor)
Cadherin-5 CADH5_HUMAN P33151 CDH5 X 87.5
Controls the cohesion and
organization of the intercellular
junctions
Carboxypeptidase B2 CBPB2_HUMAN Q96IY4 CPB2 X X X 48.4 Catalytic activity
Carboxypeptidase N catalytic
chain CBPN_HUMAN P15169 CPN1 X X X 52.3 Catalytic activity
Carboxypeptidase N subunit 2 CPN2_HUMAN P22792 CPN2 X X X 60.6 Enzyme regulator activity
Cartilage acidic protein 1 CRAC1_HUMAN Q9NQ79 CRTAC1 X 71.4 _
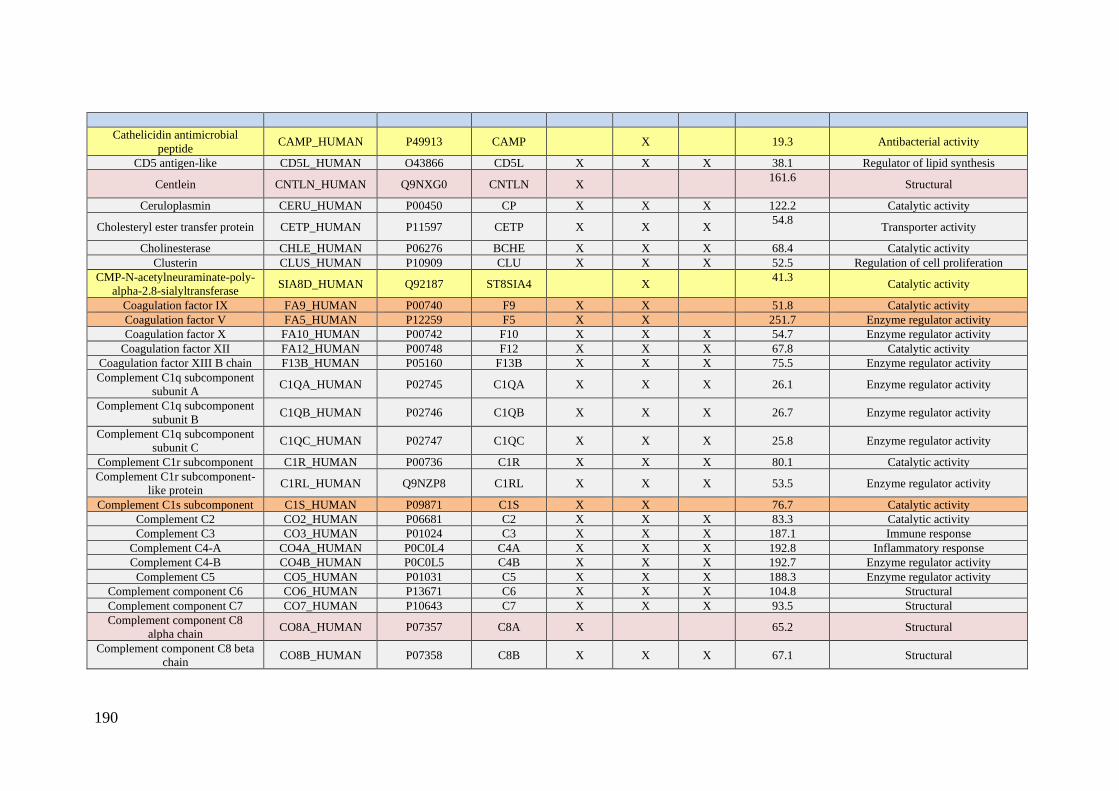
190
Cathelicidin antimicrobial
peptide CAMP_HUMAN P49913 CAMP X 19.3 Antibacterial activity
CD5 antigen-like CD5L_HUMAN O43866 CD5L X X X 38.1 Regulator of lipid synthesis
Centlein CNTLN_HUMAN Q9NXG0 CNTLN X 161.6
Structural
Ceruloplasmin CERU_HUMAN P00450 CP X X X 122.2 Catalytic activity
Cholesteryl ester transfer protein CETP_HUMAN P11597 CETP X X X 54.8
Transporter activity
Cholinesterase CHLE_HUMAN P06276 BCHE X X X 68.4 Catalytic activity
Clusterin CLUS_HUMAN P10909 CLU X X X 52.5 Regulation of cell proliferation
CMP-N-acetylneuraminate-poly-
alpha-2.8-sialyltransferase SIA8D_HUMAN Q92187 ST8SIA4 X
41.3
Catalytic activity
Coagulation factor IX FA9_HUMAN P00740 F9 X X 51.8 Catalytic activity
Coagulation factor V FA5_HUMAN P12259 F5 X X 251.7 Enzyme regulator activity
Coagulation factor X FA10_HUMAN P00742 F10 X X X 54.7 Enzyme regulator activity
Coagulation factor XII FA12_HUMAN P00748 F12 X X X 67.8 Catalytic activity
Coagulation factor XIII B chain F13B_HUMAN P05160 F13B X X X 75.5 Enzyme regulator activity
Complement C1q subcomponent
subunit A C1QA_HUMAN P02745 C1QA X X X 26.1 Enzyme regulator activity
Complement C1q subcomponent
subunit B C1QB_HUMAN P02746 C1QB X X X 26.7 Enzyme regulator activity
Complement C1q subcomponent
subunit C C1QC_HUMAN P02747 C1QC X X X 25.8 Enzyme regulator activity
Complement C1r subcomponent C1R_HUMAN P00736 C1R X X X 80.1 Catalytic activity
Complement C1r subcomponent-
like protein C1RL_HUMAN Q9NZP8 C1RL X X X 53.5 Enzyme regulator activity
Complement C1s subcomponent C1S_HUMAN P09871 C1S X X 76.7 Catalytic activity
Complement C2 CO2_HUMAN P06681 C2 X X X 83.3 Catalytic activity
Complement C3 CO3_HUMAN P01024 C3 X X X 187.1 Immune response
Complement C4-A CO4A_HUMAN P0C0L4 C4A X X X 192.8 Inflammatory response
Complement C4-B CO4B_HUMAN P0C0L5 C4B X X X 192.7 Enzyme regulator activity
Complement C5 CO5_HUMAN P01031 C5 X X X 188.3 Enzyme regulator activity
Complement component C6 CO6_HUMAN P13671 C6 X X X 104.8 Structural
Complement component C7 CO7_HUMAN P10643 C7 X X X 93.5 Structural
Complement component C8
alpha chain CO8A_HUMAN P07357 C8A X 65.2 Structural
Complement component C8 beta
chain CO8B_HUMAN P07358 C8B X X X 67.1 Structural

191
Complement component C8
gamma chain CO8G_HUMAN P07360 C8G X X X 22.3 Structural
Complement component C9 CO9_HUMAN P02748 C9 X X 63.2 Structural
Complement factor B CFAB_HUMAN P00751 CFB X X X 85.5 Immune response
Complement factor D CFAD_HUMAN P00746 CFD X 27.1 Catalytic activity
Complement factor H CFAH_HUMAN P08603 CFH X X X 139.1 Enzyme regulator activity
Complement factor H-related
protein 1 FHR1_HUMAN Q03591 CFHR1 X X X 37.6 Plays a role in lipid metabolism
Complement factor H-related
protein 2 FHR2_HUMAN P36980 CFHR2 X X X
30.6
Plays a role in lipid metabolism
Complement factor I CFAI_HUMAN P05156 CFI X X X 65.7 Catalytic activity
Corticosteroid-binding globulin CBG_HUMAN P08185 SERPINA6 X X X 45.1 Transporter activity
Dedicator of cytokinesis protein
3 DOCK3_HUMAN Q8IZD9 DOCK3 X 233.1 Enzyme regulator activity
DENN domain-containing
protein 5B DEN5B_HUMAN Q6ZUT9 DENND5B X 145.1 Enzyme regulator activity
Dermcidin DCD_HUMAN P81605 DCD X X X 11.3
Antimicrobial activity and proteolytic
activity
DnaJ homolog subfamily C
member 22 DJC22_HUMAN Q8N4W6 DNAJC22 X 38.1 Function as a co-chaperone
Dopamine beta-hydroxylase DOPO_HUMAN P09172 DBH X 69.1 Catalyctic activity
Endophilin-A3 SH3G3_HUMAN Q99963 SH3GL3 X X 39.3 Implicated in endocytosis.
Eukaryotic translation elongation
factor 1 epsilon-1 MCA3_HUMAN O43324 EEF1E1 X 19.8
Positive modulator of ATM response
to DNA damage.
Extracellular glycoprotein lacritin LACRT_HUMAN Q9GZZ8 LACRT X 14.2 Modulates secretion by lacrimal
acinar cells
Extracellular matrix protein 1 ECM1_HUMAN Q16610 ECM1 X X X 60.7 Enzyme regulator inhibitor
F-box only protein 3 FBX3_HUMAN Q9UK99 FBXO3 X 54.561 Enzyme regulator activity
Fetuin-B FETUB_HUMAN FETUB FETUB X X X 42.1 Catalytic activity
Fibrinogen alpha chain FIBA_HUMAN P02671 FGA X 94.9 Coagulation. immune response
Fibronectin FINC_HUMAN P02751 FN1 X X X 262.6 Growth. migration and differenciation
Fibulin-1 FBLN1_HUMAN P23142 FBLN1 X X 77.2
Structural
Ficolin-2 FCN2_HUMAN Q15485 FCN2 X X X 34.0 Immunity response
Ficolin-3 FCN3_HUMAN O75636 FCN3 X X X 32.9 Immunity response
Filaggrin-2 FILA2_HUMAN Q5D862 FLG2 X 248.1 _
Galectin-3-binding protein LG3BP_HUMAN Q08380 LGALS3BP X X X 65.3 Stimulate host defense against viruses
and tumor cells
Gelsolin GELS_HUMAN P06396 GSN X X X 85.7 Actin regulation (assembly)
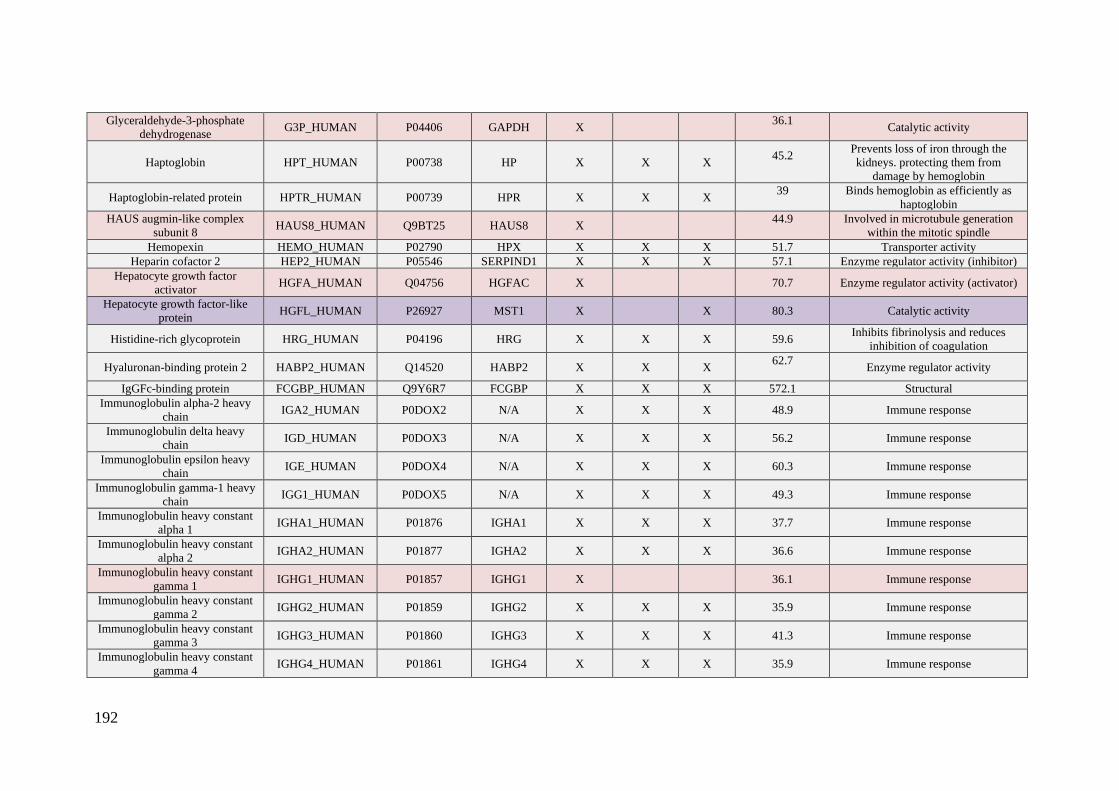
192
Glyceraldehyde-3-phosphate
dehydrogenase G3P_HUMAN P04406 GAPDH X
36.1
Catalytic activity
Haptoglobin HPT_HUMAN P00738 HP X X X 45.2
Prevents loss of iron through the
kidneys. protecting them from
damage by hemoglobin
Haptoglobin-related protein HPTR_HUMAN P00739 HPR X X X 39
Binds hemoglobin as efficiently as
haptoglobin
HAUS augmin-like complex
subunit 8 HAUS8_HUMAN Q9BT25 HAUS8 X
44.9
Involved in microtubule generation
within the mitotic spindle
Hemopexin HEMO_HUMAN P02790 HPX X X X 51.7 Transporter activity
Heparin cofactor 2 HEP2_HUMAN P05546 SERPIND1 X X X 57.1 Enzyme regulator activity (inhibitor)
Hepatocyte growth factor
activator HGFA_HUMAN Q04756 HGFAC X 70.7 Enzyme regulator activity (activator)
Hepatocyte growth factor-like
protein HGFL_HUMAN P26927 MST1 X X 80.3 Catalytic activity
Histidine-rich glycoprotein HRG_HUMAN P04196 HRG X X X 59.6 Inhibits fibrinolysis and reduces
inhibition of coagulation
Hyaluronan-binding protein 2 HABP2_HUMAN Q14520 HABP2 X X X 62.7
Enzyme regulator activity
IgGFc-binding protein FCGBP_HUMAN Q9Y6R7 FCGBP X X X 572.1 Structural
Immunoglobulin alpha-2 heavy
chain IGA2_HUMAN P0DOX2 N/A X X X 48.9 Immune response
Immunoglobulin delta heavy
chain IGD_HUMAN P0DOX3 N/A X X X 56.2 Immune response
Immunoglobulin epsilon heavy
chain IGE_HUMAN P0DOX4 N/A X X X 60.3 Immune response
Immunoglobulin gamma-1 heavy
chain IGG1_HUMAN P0DOX5 N/A X X X 49.3 Immune response
Immunoglobulin heavy constant
alpha 1 IGHA1_HUMAN P01876 IGHA1 X X X 37.7 Immune response
Immunoglobulin heavy constant
alpha 2 IGHA2_HUMAN P01877 IGHA2 X X X 36.6 Immune response
Immunoglobulin heavy constant
gamma 1 IGHG1_HUMAN P01857 IGHG1 X 36.1 Immune response
Immunoglobulin heavy constant
gamma 2 IGHG2_HUMAN P01859 IGHG2 X X X 35.9 Immune response
Immunoglobulin heavy constant
gamma 3 IGHG3_HUMAN P01860 IGHG3 X X X 41.3 Immune response
Immunoglobulin heavy constant
gamma 4 IGHG4_HUMAN P01861 IGHG4 X X X 35.9 Immune response
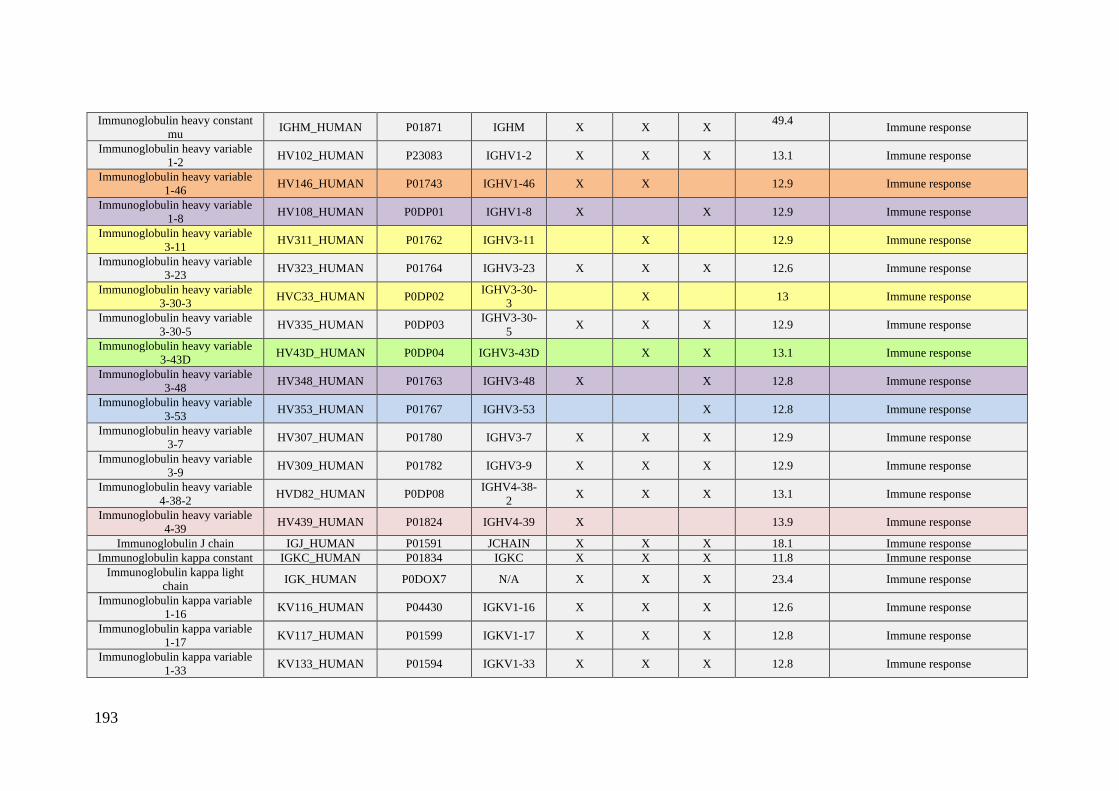
193
Immunoglobulin heavy constant
mu IGHM_HUMAN P01871 IGHM X X X
49.4
Immune response
Immunoglobulin heavy variable
1-2 HV102_HUMAN P23083 IGHV1-2 X X X 13.1 Immune response
Immunoglobulin heavy variable
1-46 HV146_HUMAN P01743 IGHV1-46 X X 12.9 Immune response
Immunoglobulin heavy variable
1-8 HV108_HUMAN P0DP01 IGHV1-8 X X 12.9 Immune response
Immunoglobulin heavy variable
3-11 HV311_HUMAN P01762 IGHV3-11 X 12.9 Immune response
Immunoglobulin heavy variable
3-23 HV323_HUMAN P01764 IGHV3-23 X X X 12.6 Immune response
Immunoglobulin heavy variable
3-30-3 HVC33_HUMAN P0DP02
IGHV3-30-
3 X 13 Immune response
Immunoglobulin heavy variable
3-30-5 HV335_HUMAN P0DP03
IGHV3-30-
5 X X X 12.9 Immune response
Immunoglobulin heavy variable
3-43D HV43D_HUMAN P0DP04 IGHV3-43D X X 13.1 Immune response
Immunoglobulin heavy variable
3-48 HV348_HUMAN P01763 IGHV3-48 X X 12.8 Immune response
Immunoglobulin heavy variable
3-53 HV353_HUMAN P01767 IGHV3-53 X 12.8 Immune response
Immunoglobulin heavy variable
3-7 HV307_HUMAN P01780 IGHV3-7 X X X 12.9 Immune response
Immunoglobulin heavy variable
3-9 HV309_HUMAN P01782 IGHV3-9 X X X 12.9 Immune response
Immunoglobulin heavy variable
4-38-2 HVD82_HUMAN P0DP08
IGHV4-38-
2 X X X 13.1 Immune response
Immunoglobulin heavy variable
4-39 HV439_HUMAN P01824 IGHV4-39 X 13.9 Immune response
Immunoglobulin J chain IGJ_HUMAN P01591 JCHAIN X X X 18.1 Immune response
Immunoglobulin kappa constant IGKC_HUMAN P01834 IGKC X X X 11.8 Immune response
Immunoglobulin kappa light
chain IGK_HUMAN P0DOX7 N/A X X X 23.4 Immune response
Immunoglobulin kappa variable
1-16 KV116_HUMAN P04430 IGKV1-16 X X X 12.6 Immune response
Immunoglobulin kappa variable
1-17 KV117_HUMAN P01599 IGKV1-17 X X X 12.8 Immune response
Immunoglobulin kappa variable
1-33 KV133_HUMAN P01594 IGKV1-33 X X X 12.8 Immune response

194
Immunoglobulin kappa variable
1D-12 KVD12_HUMAN P01611 IGKV1D-12 X 12.6 Immune response
Immunoglobulin kappa variable
1D-16 KVD16_HUMAN P01601 IGKV1D-16 X X 12.7 Immune response
Immunoglobulin kappa variable
1D-39 KVD39_HUMAN P04432 IGKV1D-39 X X X 12.7 Immune response
Immunoglobulin kappa variable
2-30 KV230_HUMAN P06310 IGKV2-30 X X X 13.2 Immune response
Immunoglobulin kappa variable
2D-28 KVD28_HUMAN P01615 IGKV2D-28 X X X 12.9 Immune response
Immunoglobulin kappa variable
3-11 KV311_HUMAN P04433 IGKV3-11 X X X 12.6 Immune response
Immunoglobulin kappa variable
3-15 KV315_HUMAN P01624 IGKV3-15 X X X 12.5 Immune response
Immunoglobulin kappa variable
3-20 KV320_HUMAN P01619 IGKV3-20 X X X 12.6 Immune response
Immunoglobulin kappa variable
4-1 KV401_HUMAN P06312 IGKV4-1 X X X 13.4 Immune response
Immunoglobulin lambda constant
2 IGLC2_HUMAN P0DOY2 IGLC2 X X X 11.3 Immune response
Immunoglobulin lambda constant
3 IGLC3_HUMAN P0DOY3 IGLC3 X X X 11.3 Immune response
Immunoglobulin lambda variable
1-47 LV147_HUMAN P01700 IGLV1-47 X X X 12.3 Immune response
Immunoglobulin lambda variable
1-51 LV151_HUMAN P01701 IGLV1-51 X X X 12.2 Immune response
Immunoglobulin lambda variable
2-11 LV211_HUMAN P01706 IGLV2-11 X X 12.6 Immune response
Immunoglobulin lambda variable
2-23 LV223_HUMAN P01705 IGLV2-23 X X 11.9 Immune response
Immunoglobulin lambda variable
3-19 LV319_HUMAN P01714 IGLV3-19 X X X 12.1 Immune response
Immunoglobulin lambda variable
3-21 LV321_HUMAN P80748 IGLV3-21 X X X 12.4 Immune response
Immunoglobulin lambda variable
3-25 LV325_HUMAN P01717 IGLV3-25 X X X 12.1 Immune response
Immunoglobulin lambda variable
3-27 LV327_HUMAN P01718 IGLV3-27 X 12.2 Immune response
Immunoglobulin lambda variable
6-57 LV657_HUMAN P01721 IGLV6-57 X X X 12.6 Immune response
195
Immunoglobulin lambda-1 light
chain IGL1_HUMAN P0DOX8 N/A X X X 22.8 Immune response
Immunoglobulin lambda-like
polypeptide 1 IGLL1_HUMAN P15814 IGLL1 X
22.9
Immune response
Immunoglobulin lambda-like
polypeptide 5 IGLL5_HUMAN B9A064 IGLL5 X
23.1
Immune response
Immunoglobulin mu heavy chain IGM_HUMAN P0DOX6 N/A X X X 63.5 Immune response
Insulin-like growth factor-
binding protein complex acid
labile subunit
ALS_HUMAN P35858 IGFALS X X X 66.1
Binds insulin-like growth factors.
increasing their half-life and their
vascular localization
Inter-alpha-trypsin inhibitor
heavy chain H1 ITIH1_HUMAN P19827 ITIH1 X X X
101.4
Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H2 ITIH2_HUMAN P19823 ITIH2 X X X 106.5 Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H3 ITIH3_HUMAN Q06033 ITIH3 X X X
99.9
Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H4 ITIH4_HUMAN Q14624 ITIH4 X X X
103.4
Inflammatory response
Interleukin-1 receptor accessory
protein IL1AP_HUMAN Q9NPH3 IL1RAP X X X
65.4
Inflammatory response
Kallistatin KAIN_HUMAN P29622 SERPINA4 X X X 48.5 Enzyme regulator inhibitor
Keratin. type I cytoskeletal 10 K1C10_HUMAN P13645 KRT10 X X X 58.8 Structural
Keratin. type I cytoskeletal 14 K1C14_HUMAN P02533 KRT14 X X X 51.6 Structural
Keratin. type I cytoskeletal 15 K1C15_HUMAN P19012 KRT15 X 49.2 Structural
Keratin. type I cytoskeletal 16 K1C16_HUMAN P08779 KRT16 X X X 51.3 Structural
Keratin. type I cytoskeletal 9 K1C9_HUMAN P35527 KRT9 X X X 62.1 Structural
Keratin. type II cytoskeletal 1 K2C1_HUMAN P04264 KRT1 X X X 66.1 Enzyme regulator activity
Keratin. type II cytoskeletal 2
epidermal K22E_HUMAN P35908 KRT2 X X X 65.4 Structural
Keratin. type II cytoskeletal 5 K2C5_HUMAN P13647 KRT5 X X X 62.4 Structural
Keratin. type II cytoskeletal 6A K2C6A_HUMAN P02538 KRT6A X X 60.1 Enzyme regulator activity (inhibitor)
Keratin. type II cytoskeletal 6C K2C6C_HUMAN P48668 KRT6C X X 60.1 Structural
Kininogen-1 KNG1_HUMAN P01042 KNG1 X X X 71.9 Enzyme regulator activity
Kynurenine-oxoglutarate
transaminase 3 KAT3_HUMAN Q6YP21 KYAT3 X
51.4
Catalytic activity
Leucine-rich alpha-2-
glycoprotein A2GL_HUMAN P02750 LRG1 X X X 38.2
Protein-protein interaction. signal
transduction. and cell adhesion and
development
Leucine-rich repeat-containing LRRC9_HUMAN Q6ZRR7 LRRC9 X 166.9 _
196
protein 9
Lipopolysaccharide-binding
protein LBP_HUMAN P18428 LBP X X X 53.4 Immune response.
L-lactate dehydrogenase A-like
6B LDH6B_HUMAN Q9BYZ2 LDHAL6B X X 41.9 Catalytic activity
L-lactate dehydrogenase B chain LDHB_HUMAN P07195 LDHB X 36.6 Catalytic activity
L-selectin LYAM1_HUMAN P14151 SELL X X 42.2
Mediates in lymphocyte-endothelial
cell interactions
Lumican LUM_HUMAN P51884 LUM X X X 38.4 Collagen binding. extracellular matrix
structural constituent
Maestro heat-like repeat-
containing protein family
member 2A
MRO2A_HUMAN A6NES4 MROH2A X 189.6 _
Mannan-binding lectin serine
protease 1 MASP1_HUMAN P48740 MASP1 X X X
79.2
Enzyme regulator activity
Mannan-binding lectin serine
protease 2 MASP2_HUMAN O00187 MASP2 X X
75.7
Catalytic activity
Mannosyl-oligosaccharide 1.2-
alpha-mannosidase IA MA1A1_HUMAN P33908 MAN1A1 X X
72.9
Catalytic activity
Monocyte differentiation antigen
CD14 CD14_HUMAN P08571 CD14 X X X 40.1 Immune response
Multiple inositol polyphosphate
phosphatase 1 MINP1_HUMAN Q9UNW1 MINPP1 X
55.1
Catalytic activity
N-acetylmuramoyl-L-alanine
amidase PGRP2_HUMAN Q96PD5 PGLYRP2 X X X
62.2
Catalytic activity
Nebulin-related-anchoring
protein NRAP_HUMAN Q86VF7 NRAP X
197.1
Implicated in myofibrilar
organization during cardiomyocyte
development
Neutrophil cytosol factor 4 NCF4_HUMAN Q15080 NCF4 X 39.1
Component of the NADPH-oxidase
Phosphatidylcholine-sterol
acyltransferase LCAT_HUMAN P04180 LCAT X X X 49.1 Catalytic activity
Phosphatidylinositol-glycan-
specific phospholipase D PHLD_HUMAN P80108 GPLD1 X X X
92.3
Catalytic activity
Phospholipid transfer protein PLTP_HUMAN P55058 PLTP X X X 54.8
Transporter activity
Pigment epithelium-derived
factor PEDF_HUMAN P36955 SERPINF1 X X X 46.3
Induces extensive neuronal
differentiation in retinoblastoma cells
Plasma kallikrein KLKB1_HUMAN P03952 KLKB1 X X X 71.4 Catalytic activity
Plasma protease C1 inhibitor IC1_HUMAN P05155 SERPING1 X X X 55.2 Protease.
197
Plasma serine protease inhibitor IPSP_HUMAN P05154 SERPINA5 X X 45.7 Enzyme regulator activuty (inhibitor)
Plasminogen PLMN_HUMAN P00747 PLG X X X 90.6 Catalytic activity
Platelet glycoprotein Ib alpha
chain GP1BA_HUMAN P07359 GP1BA X X 71.1
Participates in the formation of
platelet plugs
Platelet glycoprotein V GPV_HUMAN P40197 GP5 X X 61 Immune response
Pleckstrin homology domain-
containing family G member 6 PKHG6_HUMAN Q3KR16 PLEKHG6 X 88.9 Enzyme regulator activity
Polymeric immunoglobulin
receptor PIGR_HUMAN P01833 PIGR X X X 83.3
This receptor binds polymeric IgA
and IgM at the basolateral surface of
epithelial cells
Pregnancy zone protein PZP_HUMAN P20742 PZP X X X 163.8 Enzyme regulator activity
Prenylcysteine oxidase 1 PCYOX_HUMAN Q9UHG3 PCYOX1 X X X 56.6 Clatytic activity
Protein AMBP AMBP_HUMAN P02760 AMBP X X X 38.9 Enzyme regulator activity (inhibitor)
Protein argonaute-3 AGO3_HUMAN Q9H9G7 GENE:
AGO3 X 97.4 Enzyme regulator activity
Protein ENL ENL_HUMAN Q03111 MLLT1 X 62.1 Enzyme regulator activity
Protein Shroom3 SHRM3_HUMAN Q8TF72 SHROOM3 X X X 216.8
Controls cell shape changes in the
neuroepithelium during neural tube
closure
Protein Z-dependent protease
inhibitor ZPI_HUMAN Q9UK55
SERPINA1
0 X X X 50.7 Enzyme regulator activity
Proteoglycan 4 PRG4_HUMAN Q92954 PRG4 X X 151.1 Plays a role in boundary lubrication
within articulating joints
Prothrombin THRB_HUMAN P00734 F2 X X X 70.1 Catalytic activity
Pulmonary surfactant-associated
protein B PSPB_HUMAN P07988 SFTPB X X X 42.1
Promotes alveolar stability by
lowering the surface tension at the
air-liquid interface in the peripheral
air spaces
Receptor-type tyrosine-protein
phosphatase eta PTPRJ_HUMAN Q12913 PTPRJ X 145.9 Catalytic activity
Regulator of G-protein signaling
20 RGS20_HUMAN O76081 RGS20 X
31.5
Inhibits signal transduction
Retinol-binding protein 4 RET4_HUMAN P02753 RBP4 X X X 23.1 Transporter activity
Secreted phosphoprotein 24 SPP24_HUMAN Q13103 SPP2 X X 24.1
Bind cytokines of the TGF-β
superfamily and also activate
intracellular signaling pathways
Secretogranin-2 SCG2_HUMAN P13521 SCG2 X X 70.9 Involved in the packaging or sorting
of peptide hormones and
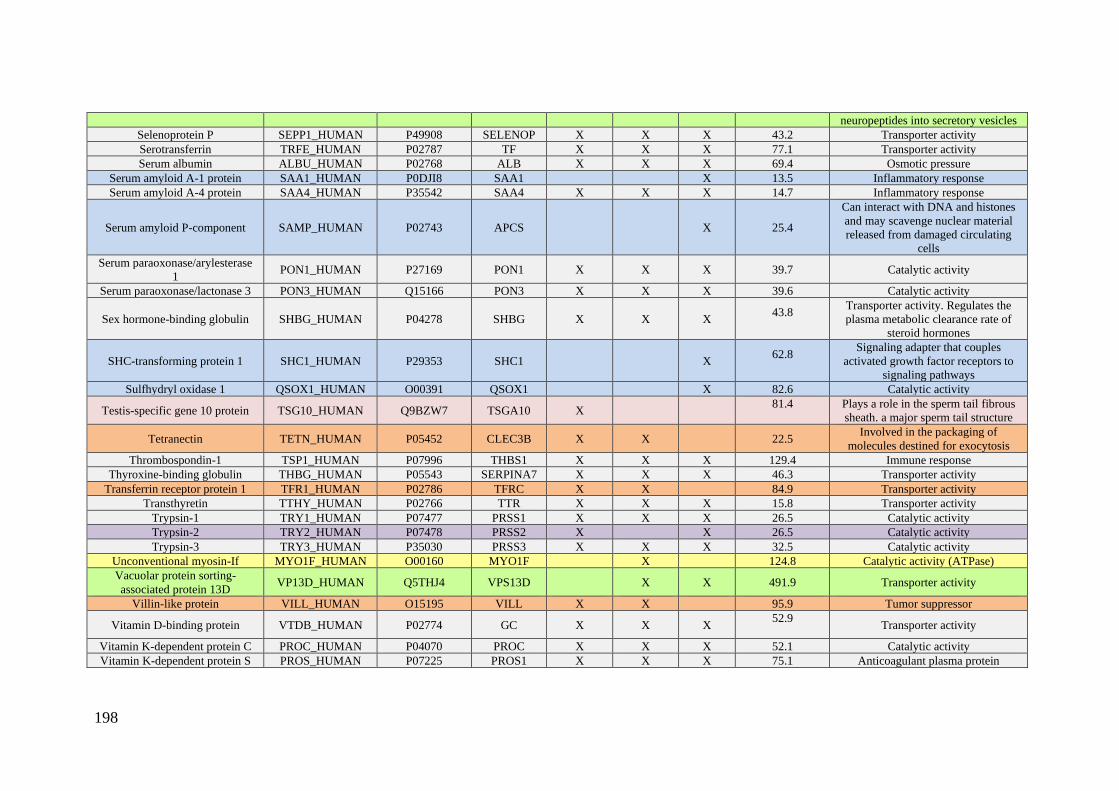
198
neuropeptides into secretory vesicles
Selenoprotein P SEPP1_HUMAN P49908 SELENOP X X X 43.2 Transporter activity
Serotransferrin TRFE_HUMAN P02787 TF X X X 77.1 Transporter activity
Serum albumin ALBU_HUMAN P02768 ALB X X X 69.4 Osmotic pressure
Serum amyloid A-1 protein SAA1_HUMAN P0DJI8 SAA1 X 13.5 Inflammatory response
Serum amyloid A-4 protein SAA4_HUMAN P35542 SAA4 X X X 14.7 Inflammatory response
Serum amyloid P-component SAMP_HUMAN P02743 APCS X 25.4
Can interact with DNA and histones
and may scavenge nuclear material
released from damaged circulating
cells
Serum paraoxonase/arylesterase
1 PON1_HUMAN P27169 PON1 X X X 39.7 Catalytic activity
Serum paraoxonase/lactonase 3 PON3_HUMAN Q15166 PON3 X X X 39.6 Catalytic activity
Sex hormone-binding globulin SHBG_HUMAN P04278 SHBG X X X 43.8
Transporter activity. Regulates the
plasma metabolic clearance rate of
steroid hormones
SHC-transforming protein 1 SHC1_HUMAN P29353 SHC1 X 62.8
Signaling adapter that couples
activated growth factor receptors to
signaling pathways
Sulfhydryl oxidase 1 QSOX1_HUMAN O00391 QSOX1 X 82.6 Catalytic activity
Testis-specific gene 10 protein TSG10_HUMAN Q9BZW7 TSGA10 X 81.4
Plays a role in the sperm tail fibrous
sheath. a major sperm tail structure
Tetranectin TETN_HUMAN P05452 CLEC3B X X 22.5 Involved in the packaging of
molecules destined for exocytosis
Thrombospondin-1 TSP1_HUMAN P07996 THBS1 X X X 129.4 Immune response
Thyroxine-binding globulin THBG_HUMAN P05543 SERPINA7 X X X 46.3 Transporter activity
Transferrin receptor protein 1 TFR1_HUMAN P02786 TFRC X X 84.9 Transporter activity
Transthyretin TTHY_HUMAN P02766 TTR X X X 15.8 Transporter activity
Trypsin-1 TRY1_HUMAN P07477 PRSS1 X X X 26.5 Catalytic activity
Trypsin-2 TRY2_HUMAN P07478 PRSS2 X X 26.5 Catalytic activity
Trypsin-3 TRY3_HUMAN P35030 PRSS3 X X X 32.5 Catalytic activity
Unconventional myosin-If MYO1F_HUMAN O00160 MYO1F X 124.8 Catalytic activity (ATPase)
Vacuolar protein sorting-
associated protein 13D VP13D_HUMAN Q5THJ4 VPS13D X X 491.9 Transporter activity
Villin-like protein VILL_HUMAN O15195 VILL X X 95.9 Tumor suppressor
Vitamin D-binding protein VTDB_HUMAN P02774 GC X X X 52.9
Transporter activity
Vitamin K-dependent protein C PROC_HUMAN P04070 PROC X X X 52.1 Catalytic activity
Vitamin K-dependent protein S PROS_HUMAN P07225 PROS1 X X X 75.1 Anticoagulant plasma protein
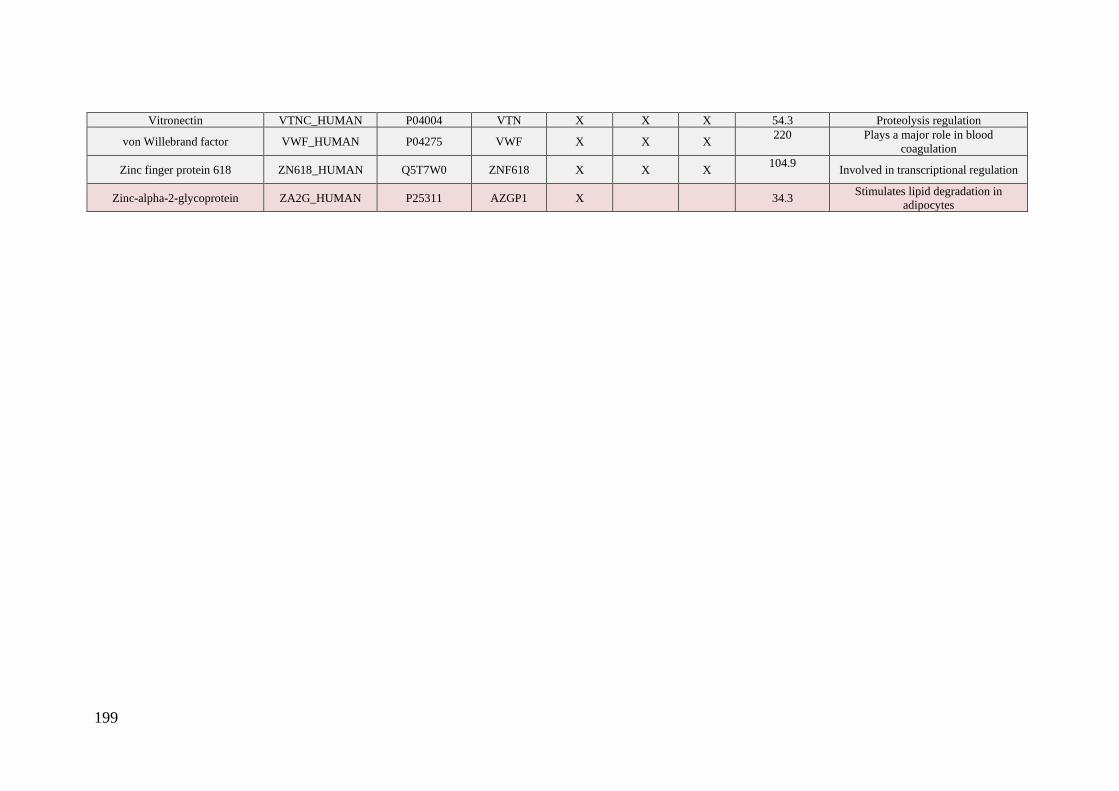
199
Vitronectin VTNC_HUMAN P04004 VTN X X X 54.3 Proteolysis regulation
von Willebrand factor VWF_HUMAN P04275 VWF X X X 220
Plays a major role in blood
coagulation
Zinc finger protein 618 ZN618_HUMAN Q5T7W0 ZNF618 X X X 104.9
Involved in transcriptional regulation
Zinc-alpha-2-glycoprotein ZA2G_HUMAN P25311 AZGP1 X 34.3 Stimulates lipid degradation in
adipocytes
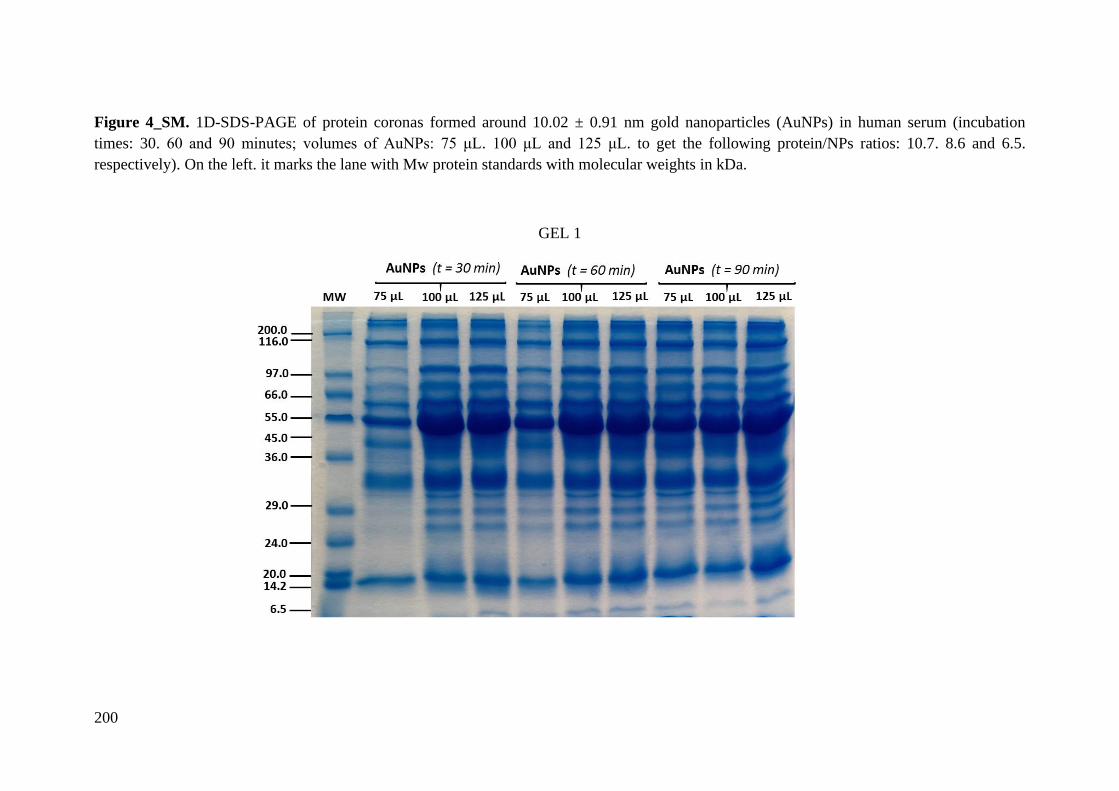
200
Figure 4_SM. 1D-SDS-PAGE of protein coronas formed around 10.02 ± 0.91 nm gold nanoparticles (AuNPs) in human serum (incubation
times: 30. 60 and 90 minutes; volumes of AuNPs: 75 μL. 100 μL and 125 μL. to get the following protein/NPs ratios: 10.7. 8.6 and 6.5.
respectively). On the left. it marks the lane with Mw protein standards with molecular weights in kDa.
GEL 1

201
Figure 5_SM. 1D-SDS-PAGE of protein coronas formed around 9.73 ± 1.70 nm silver nanoparticles (AgNPs) in human serum (incubation
times: 30. 60 and 90 minutes; volumes of AgNPs: 75 μL. 100 μL and 125 μL. to get the following protein/NPs ratios: 10.7. 8.6 and 6.5.
respectively). On the left. it marks the lane with Mw protein standards with molecular weights in kDa.
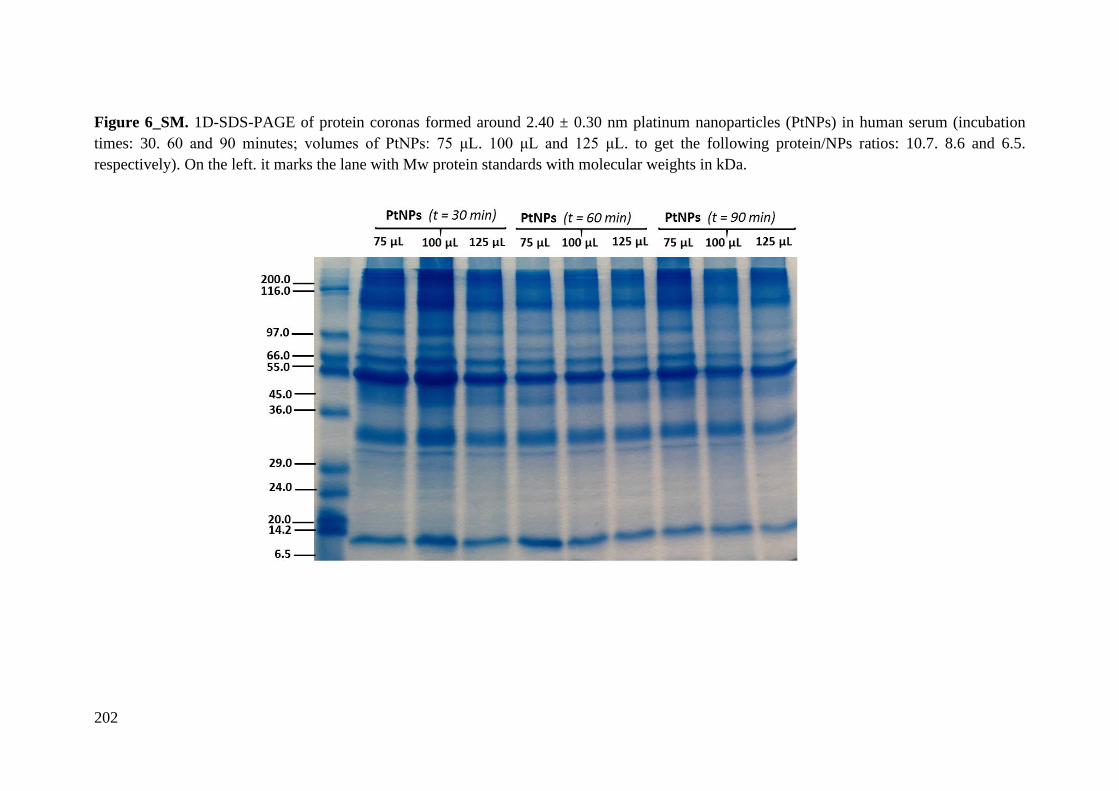
202
Figure 6_SM. 1D-SDS-PAGE of protein coronas formed around 2.40 ± 0.30 nm platinum nanoparticles (PtNPs) in human serum (incubation
times: 30. 60 and 90 minutes; volumes of PtNPs: 75 μL. 100 μL and 125 μL. to get the following protein/NPs ratios: 10.7. 8.6 and 6.5.
respectively). On the left. it marks the lane with Mw protein standards with molecular weights in kDa.

203
Figure 7_SM. 1D-SDS-PAGE of fractions of human serum (supernatants) after the separation of the pellet containing the protein coronas
formed around 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs) and 2.40 ± 0.30 nm platinum
nanoparticles (PtNPs) in human serum (incubation time: 30 minutes; volumes of each nanoparticles solution: 75 μL. 100 μL and 125 μL. to get
the following protein/NPs ratios: 10.7. 8.6 and 6.5. respectively). On the left. it marks the lane with Mw protein standards with molecular
weights in kDa.

204

ANNEX B
Supplemental Material Chapter 2

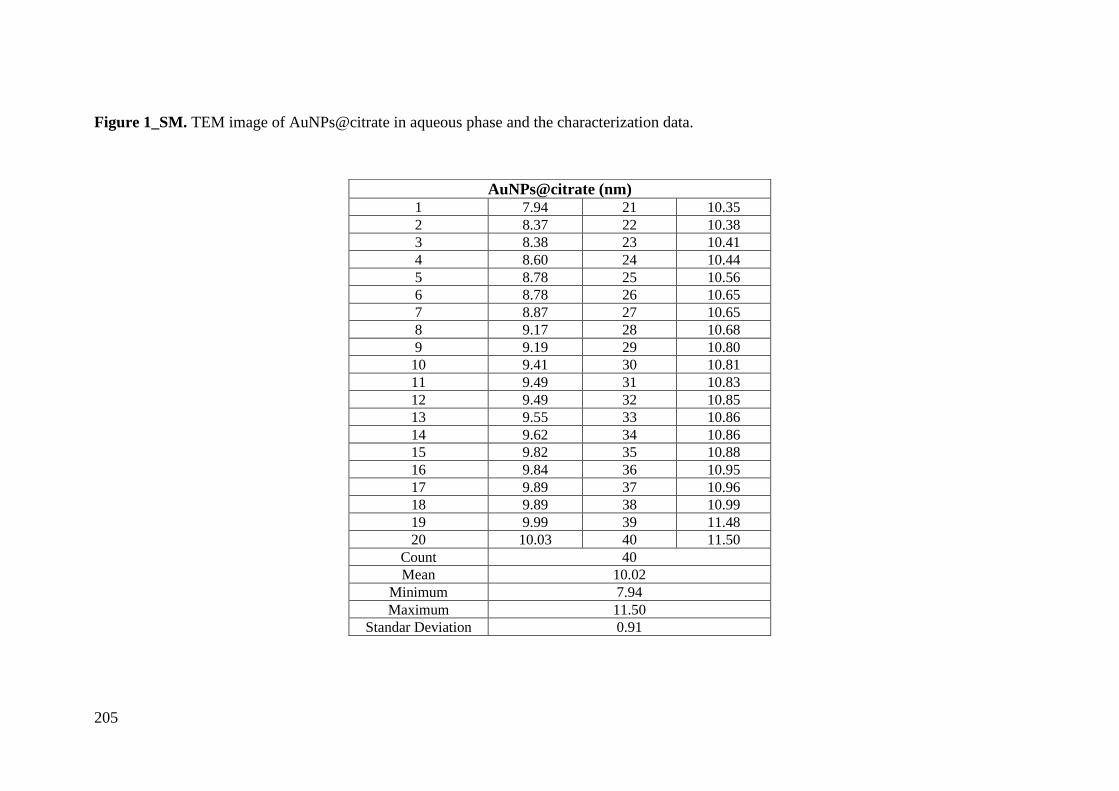
205
Figure 1_SM. TEM image of AuNPs@citrate in aqueous phase and the characterization data.
AuNPs@citrate (nm) 1 7.94 21 10.35
2 8.37 22 10.38
3 8.38 23 10.41
4 8.60 24 10.44
5 8.78 25 10.56
6 8.78 26 10.65
7 8.87 27 10.65
8 9.17 28 10.68
9 9.19 29 10.80
10 9.41 30 10.81
11 9.49 31 10.83
12 9.49 32 10.85
13 9.55 33 10.86
14 9.62 34 10.86
15 9.82 35 10.88
16 9.84 36 10.95
17 9.89 37 10.96
18 9.89 38 10.99
19 9.99 39 11.48
20 10.03 40 11.50
Count 40
Mean 10.02
Minimum 7.94
Maximum 11.50
Standar Deviation 0.91
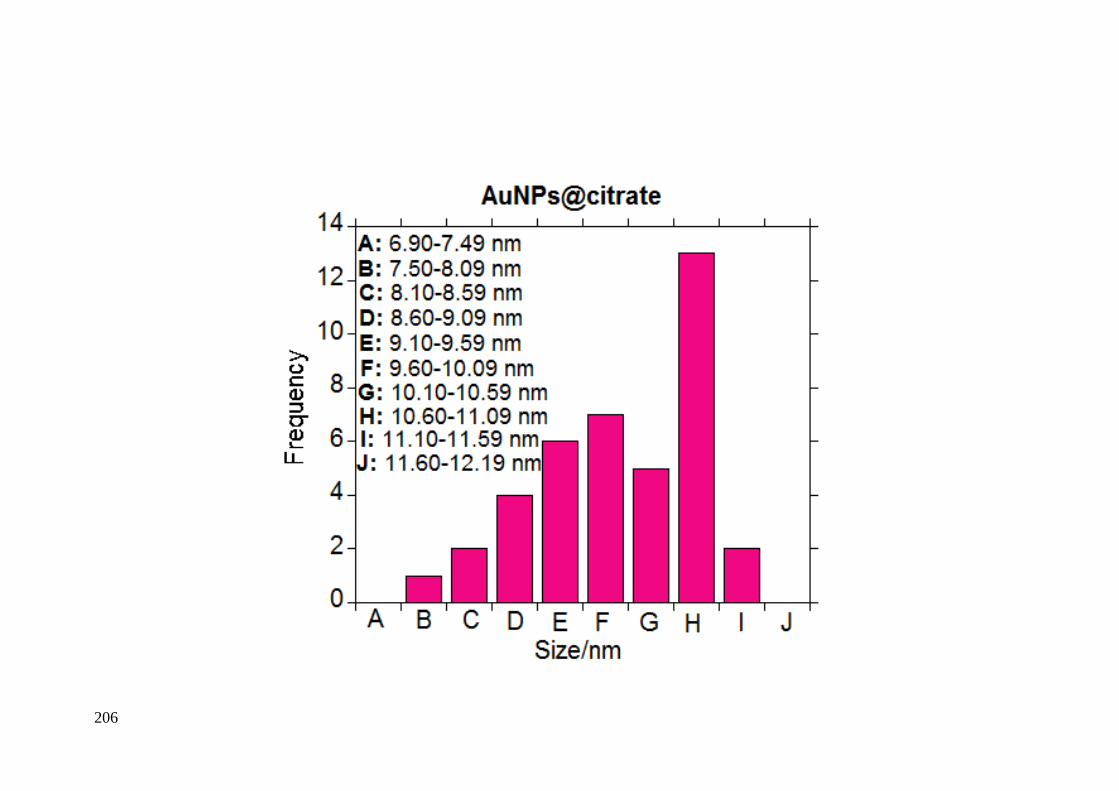
206

207

208
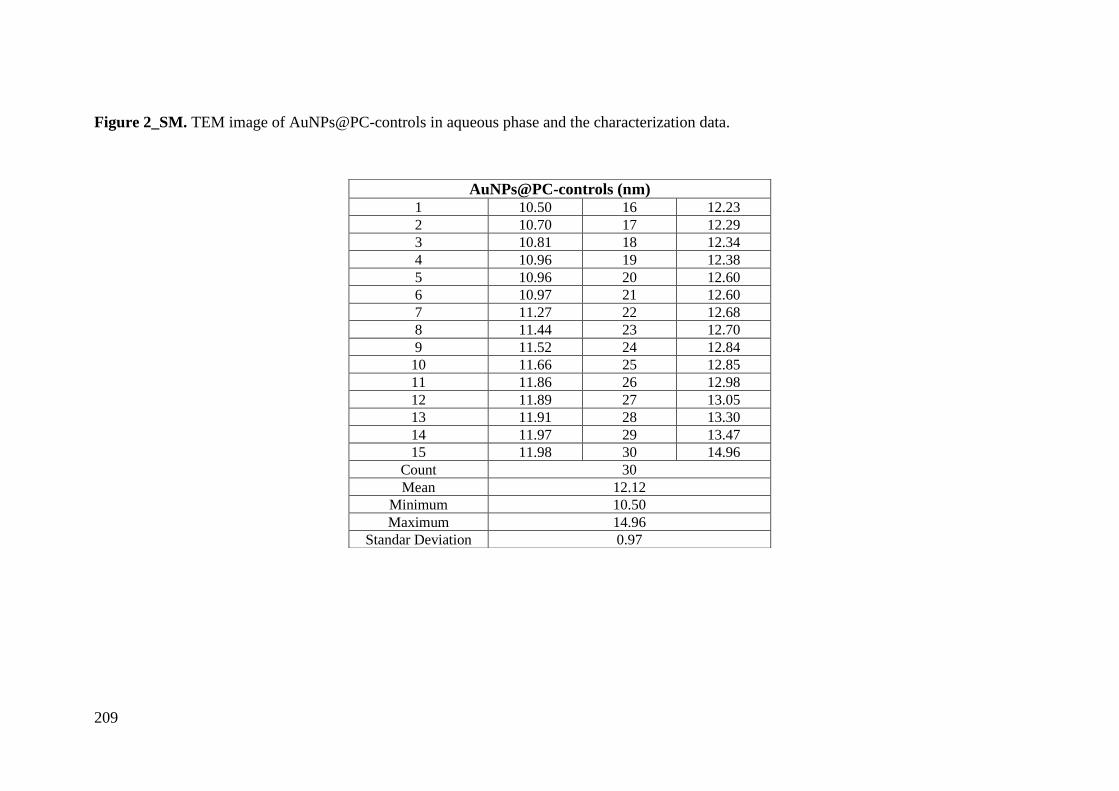
209
Figure 2_SM. TEM image of AuNPs@PC-controls in aqueous phase and the characterization data.
AuNPs@PC-controls (nm) 1 10.50 16 12.23
2 10.70 17 12.29
3 10.81 18 12.34
4 10.96 19 12.38
5 10.96 20 12.60
6 10.97 21 12.60
7 11.27 22 12.68
8 11.44 23 12.70
9 11.52 24 12.84
10 11.66 25 12.85
11 11.86 26 12.98
12 11.89 27 13.05
13 11.91 28 13.30
14 11.97 29 13.47
15 11.98 30 14.96
Count 30
Mean 12.12
Minimum 10.50
Maximum 14.96
Standar Deviation 0.97
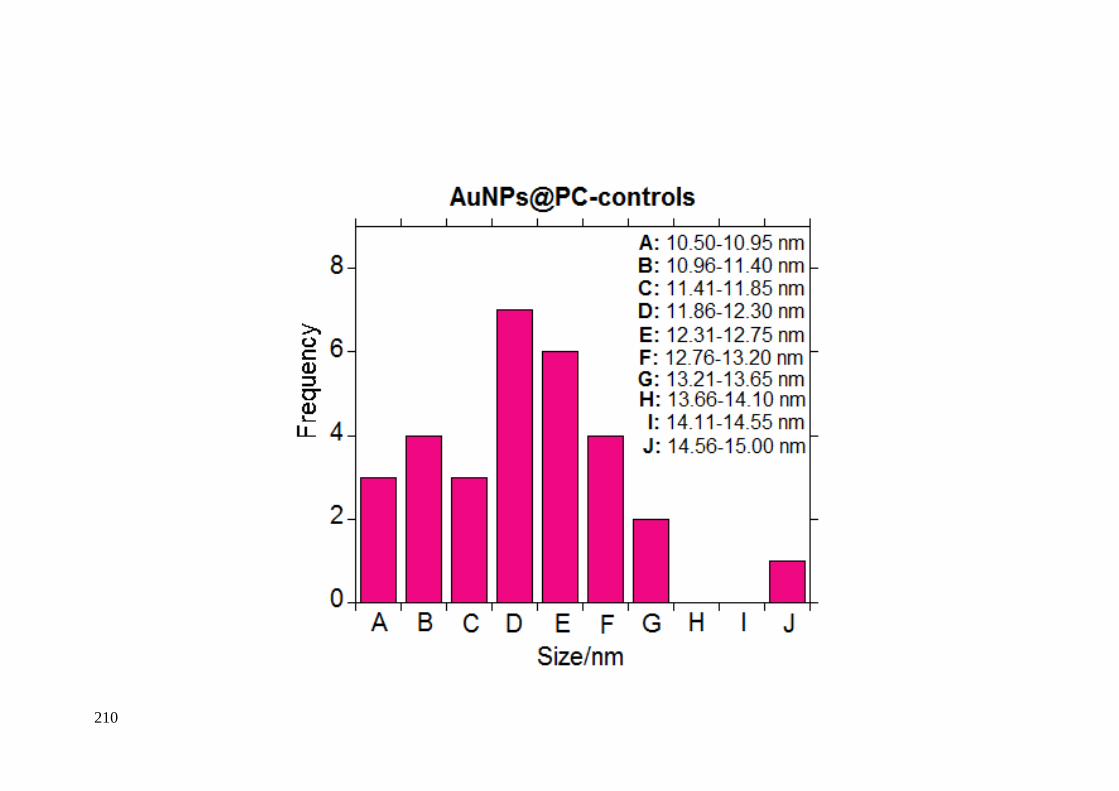
210

211
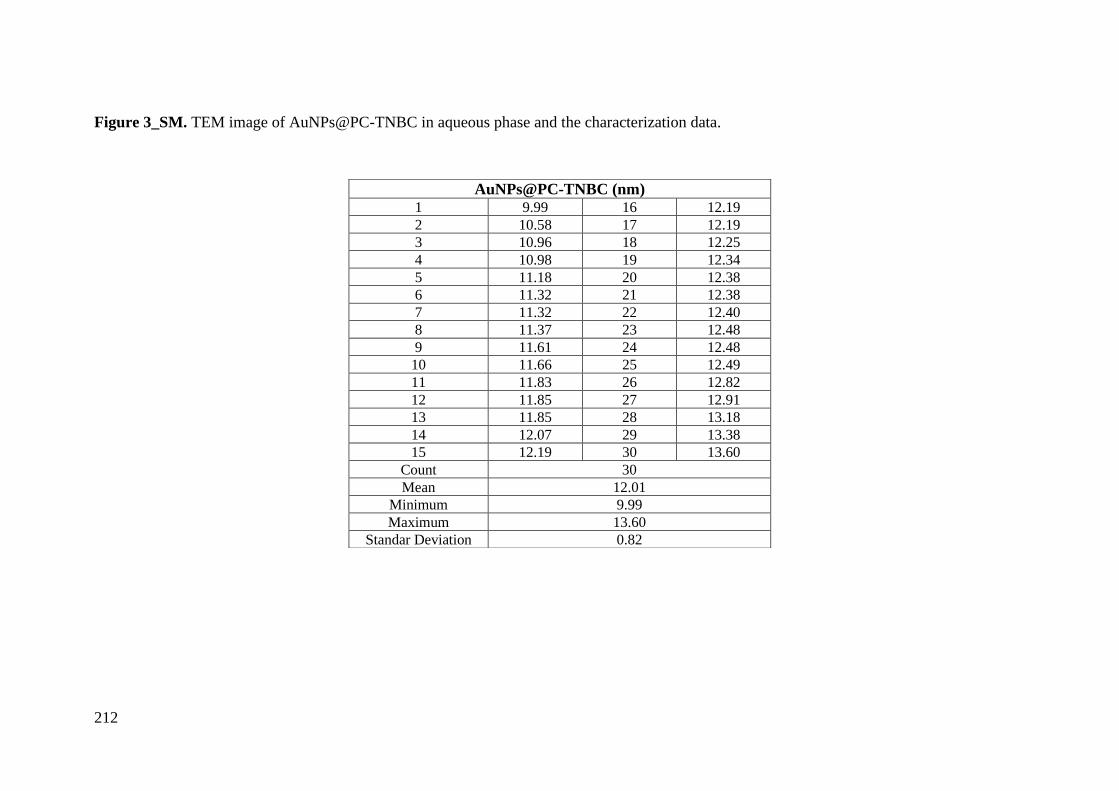
212
Figure 3_SM. TEM image of AuNPs@PC-TNBC in aqueous phase and the characterization data.
AuNPs@PC-TNBC (nm) 1 9.99 16 12.19
2 10.58 17 12.19
3 10.96 18 12.25
4 10.98 19 12.34
5 11.18 20 12.38
6 11.32 21 12.38
7 11.32 22 12.40
8 11.37 23 12.48
9 11.61 24 12.48
10 11.66 25 12.49
11 11.83 26 12.82
12 11.85 27 12.91
13 11.85 28 13.18
14 12.07 29 13.38
15 12.19 30 13.60
Count 30
Mean 12.01
Minimum 9.99
Maximum 13.60
Standar Deviation 0.82
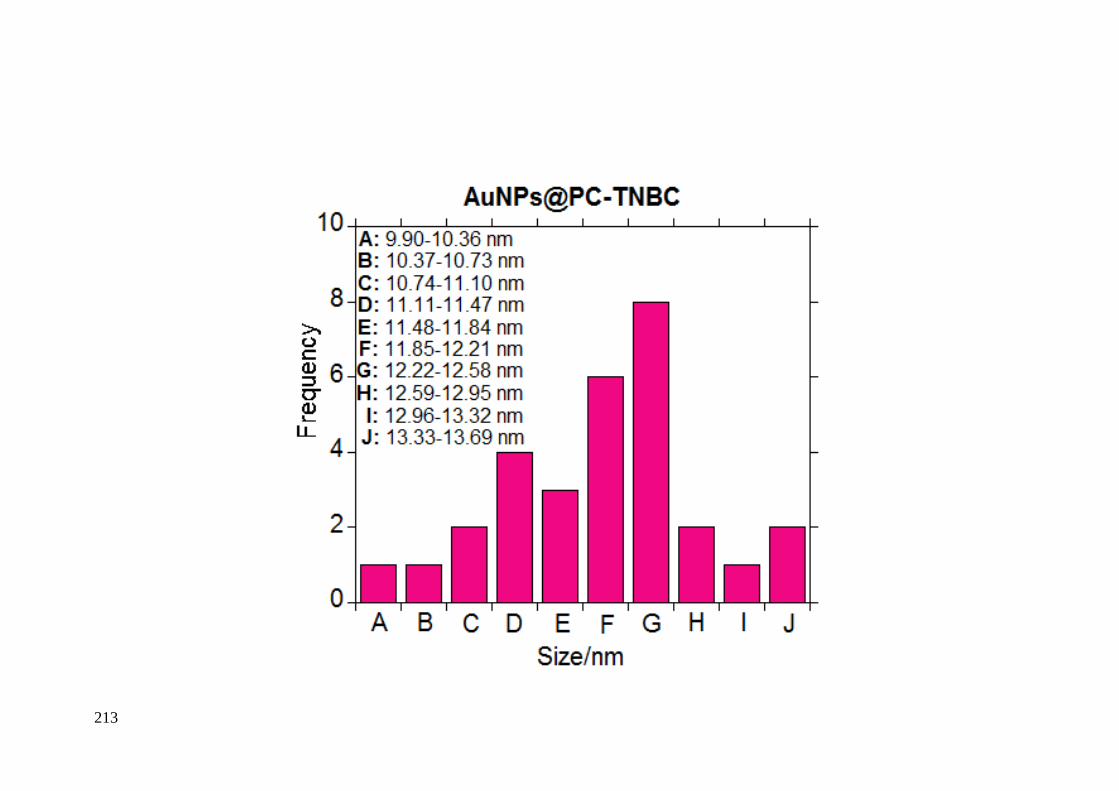
213

214

215
Figure 4_SM. TEM image of AgNPs@citrate in aqueous phase and the characterization data.
AgNPs@citrate (nm) 1 7.58 21 9.30
2 7.66 22 9.45
3 7.89 23 9.58
4 7.89 24 9.76
5 7.89 25 9.86
6 7.98 26 10.25
7 8.17 27 10.39
8 8.19 28 10.48
9 8.19 29 10.61
10 8.21 30 10.99
11 8.40 31 11.17
12 8.40 32 11.27
13 8.48 33 11.47
14 8.55 34 11.64
15 8.64 35 11.83
16 8.75 36 11.83
17 8.92 37 12.10
18 9.02 38 13.02
19 9.07 39 13.49
20 9.16 40 13.76
Count 40
Mean 9.73
Minimum 7.58
Maximum 13.76
Standar Deviation 1.70

216

217

218
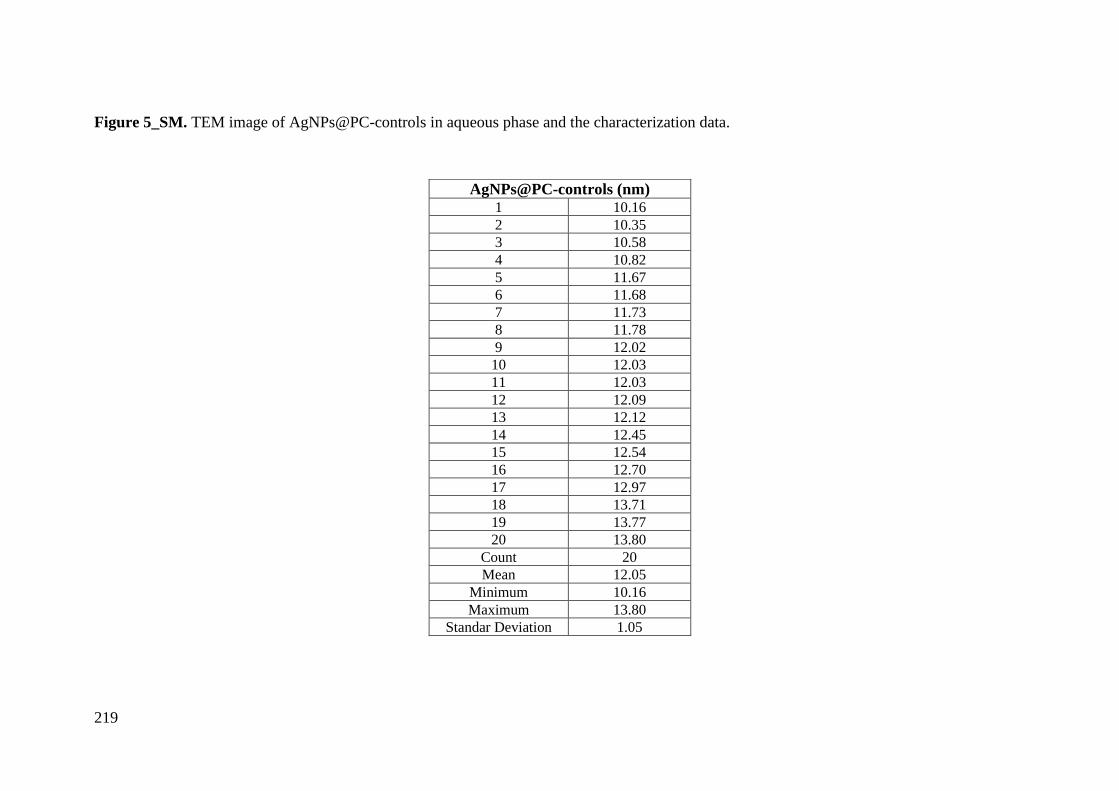
219
Figure 5_SM. TEM image of AgNPs@PC-controls in aqueous phase and the characterization data.
AgNPs@PC-controls (nm) 1 10.16
2 10.35
3 10.58
4 10.82
5 11.67
6 11.68
7 11.73
8 11.78
9 12.02
10 12.03
11 12.03
12 12.09
13 12.12
14 12.45
15 12.54
16 12.70
17 12.97
18 13.71
19 13.77
20 13.80
Count 20
Mean 12.05
Minimum 10.16
Maximum 13.80
Standar Deviation 1.05
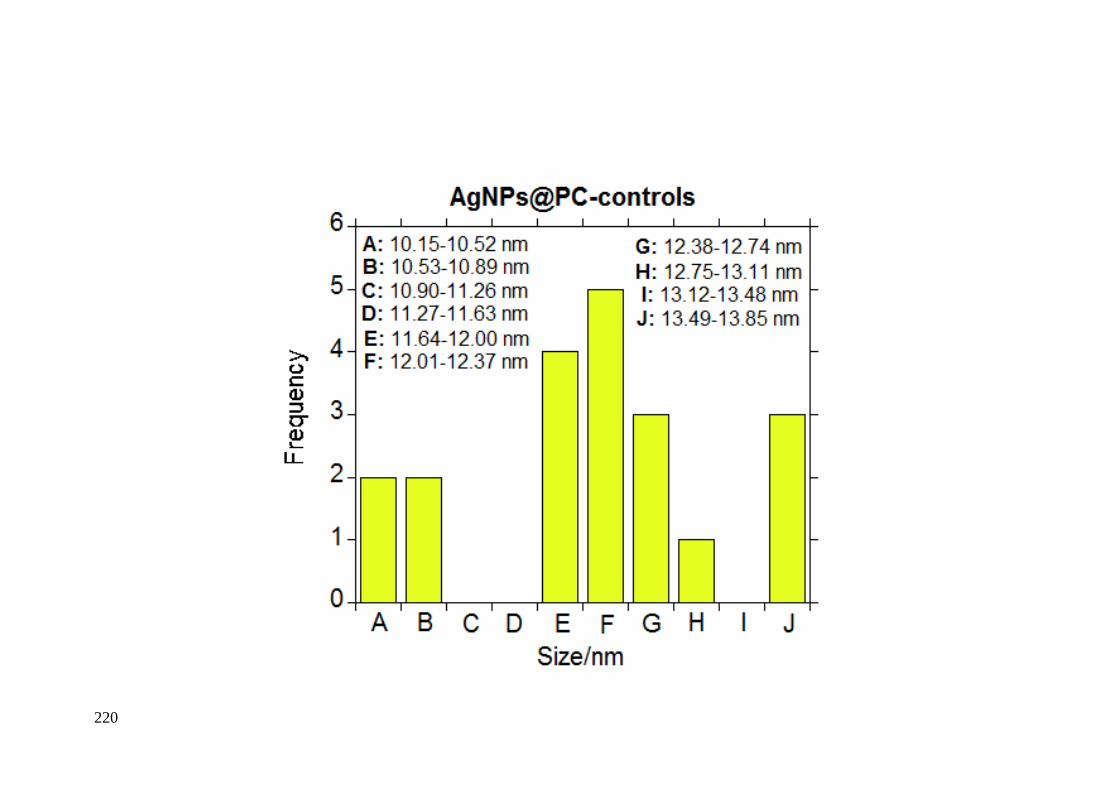
220
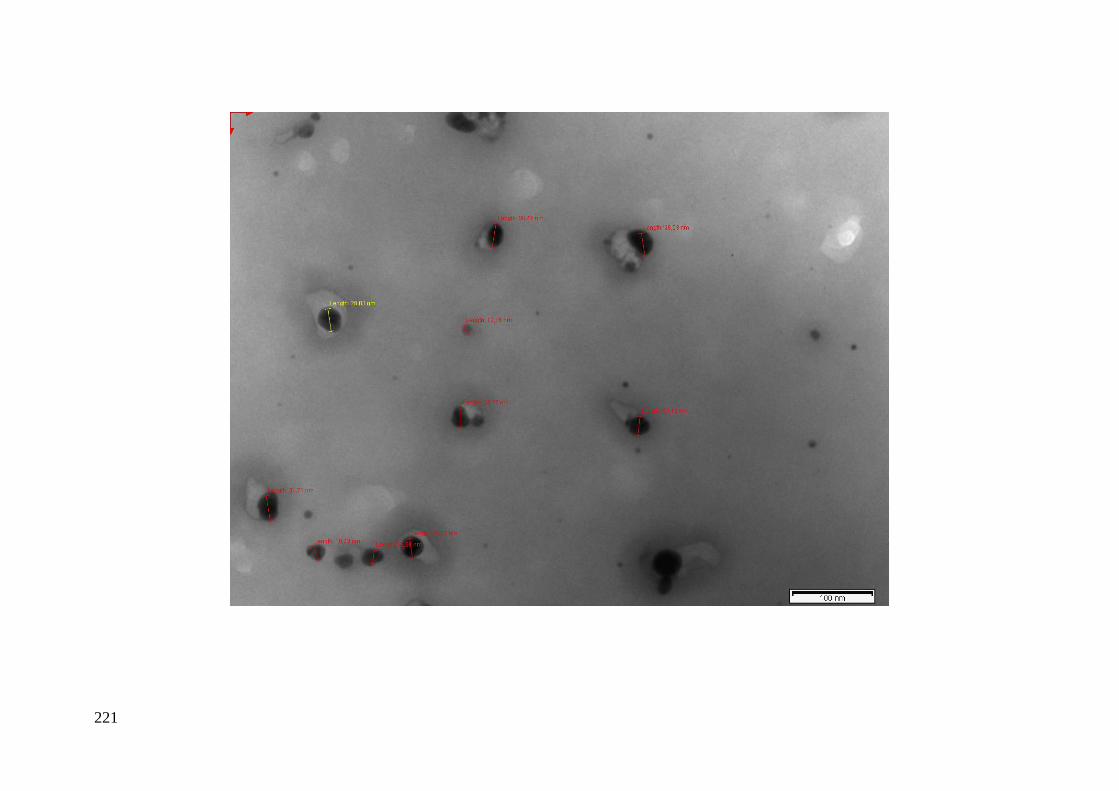
221
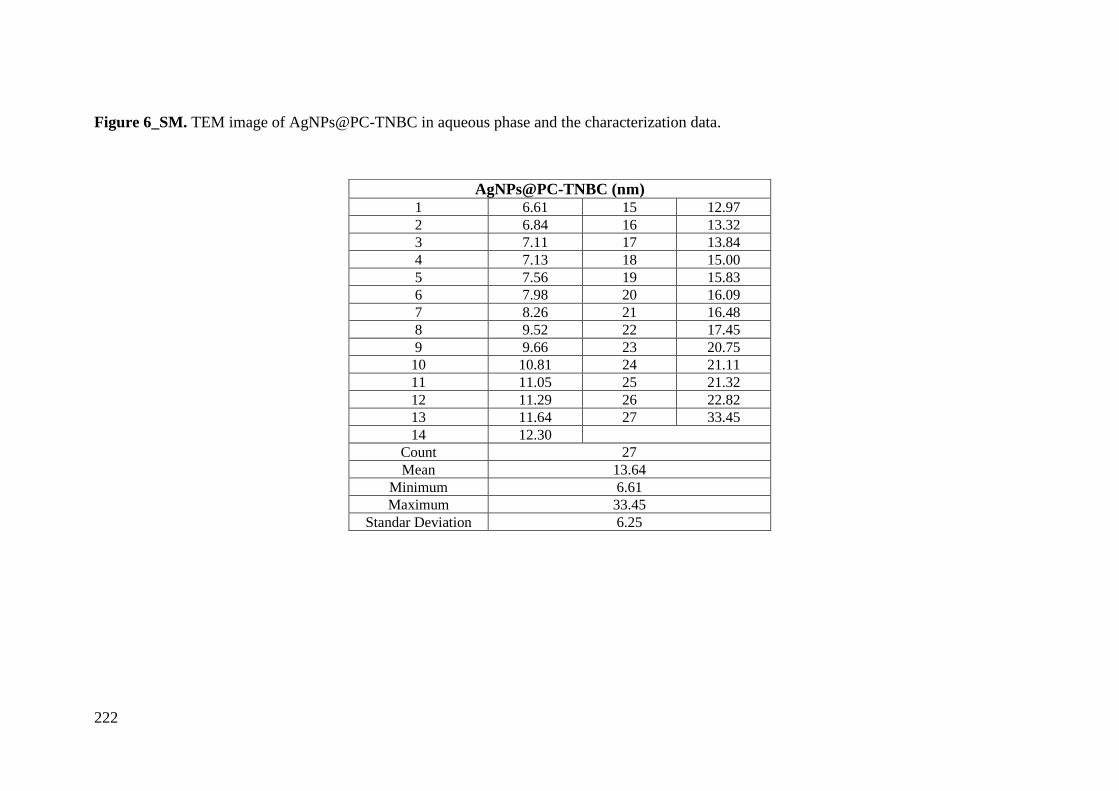
222
Figure 6_SM. TEM image of AgNPs@PC-TNBC in aqueous phase and the characterization data.
AgNPs@PC-TNBC (nm) 1 6.61 15 12.97
2 6.84 16 13.32
3 7.11 17 13.84
4 7.13 18 15.00
5 7.56 19 15.83
6 7.98 20 16.09
7 8.26 21 16.48
8 9.52 22 17.45
9 9.66 23 20.75
10 10.81 24 21.11
11 11.05 25 21.32
12 11.29 26 22.82
13 11.64 27 33.45
14 12.30
Count 27
Mean 13.64
Minimum 6.61
Maximum 33.45
Standar Deviation 6.25
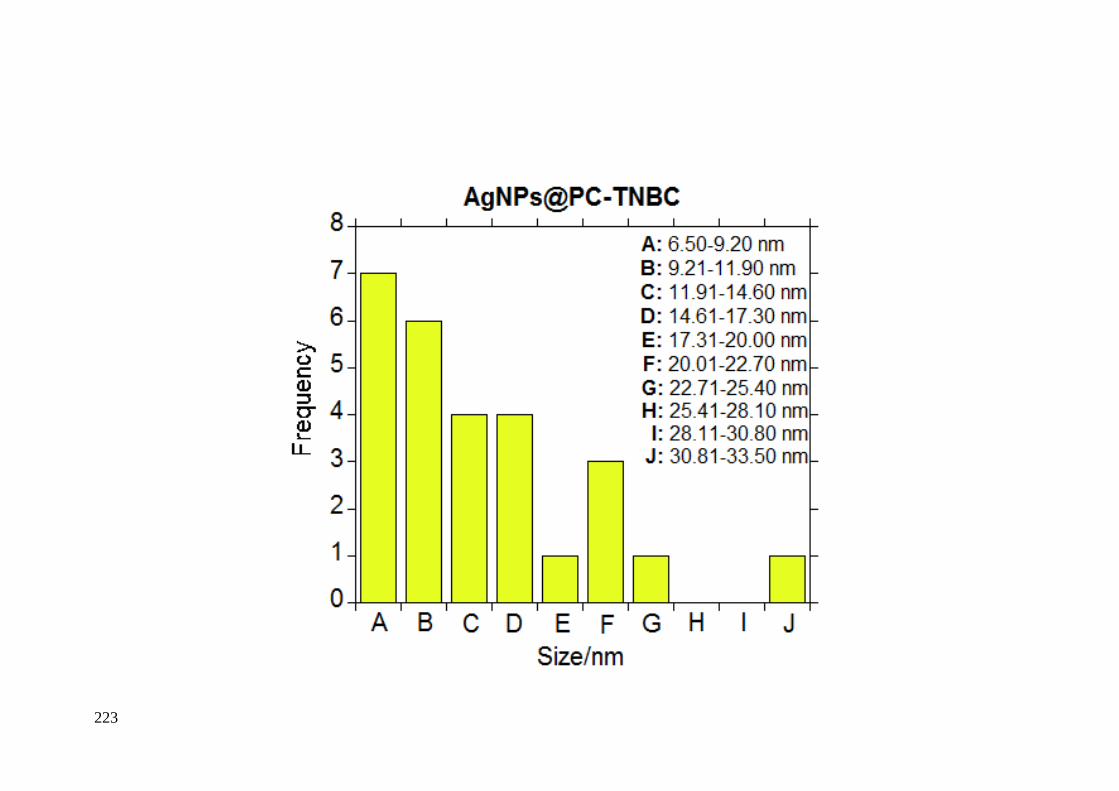
223
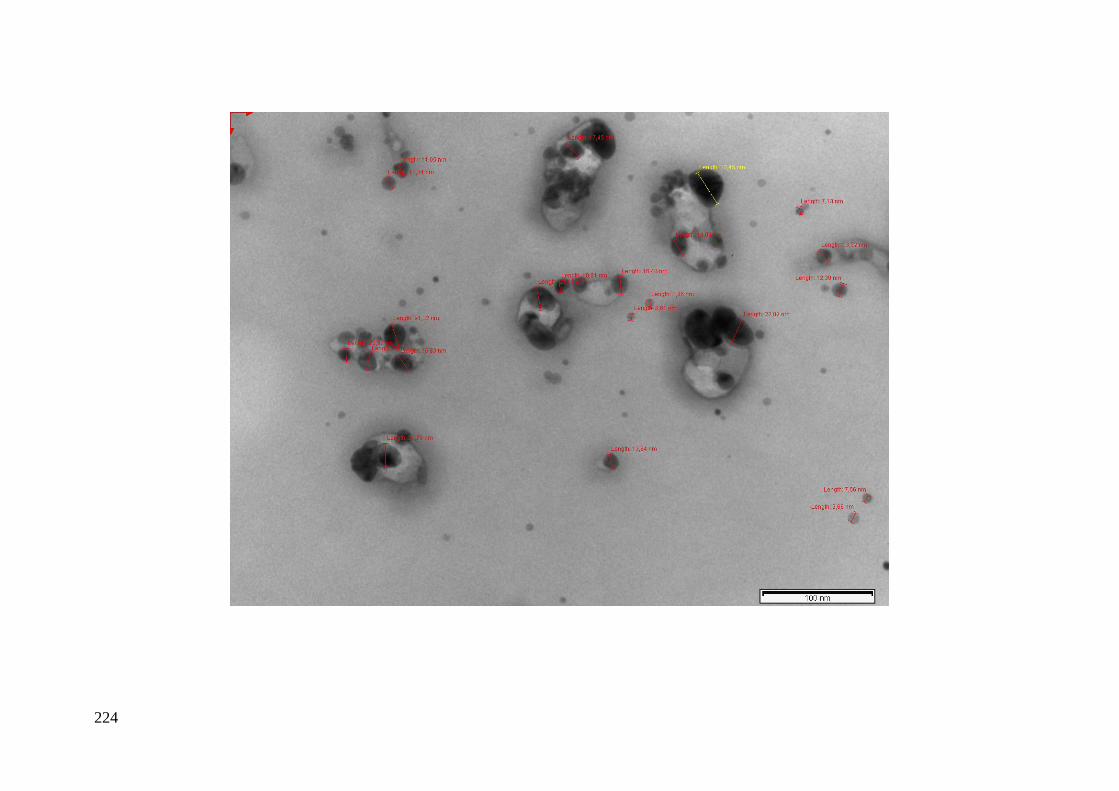
224
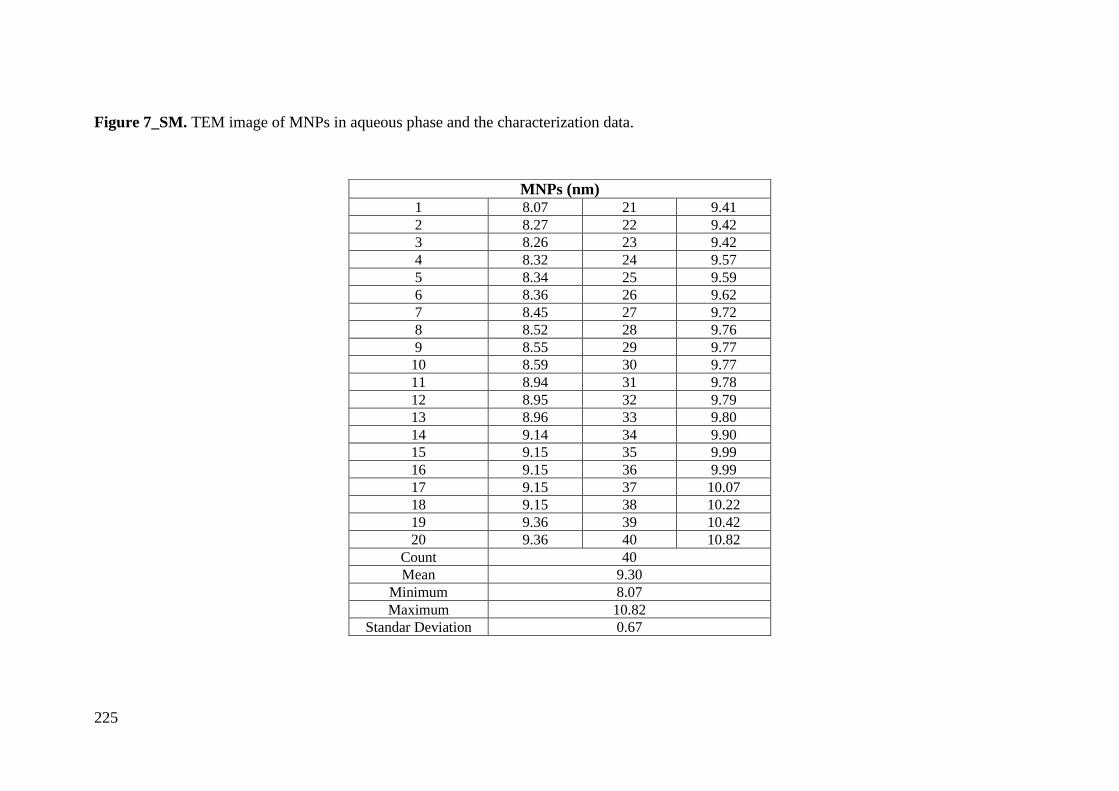
225
Figure 7_SM. TEM image of MNPs in aqueous phase and the characterization data.
MNPs (nm) 1 8.07 21 9.41
2 8.27 22 9.42
3 8.26 23 9.42
4 8.32 24 9.57
5 8.34 25 9.59
6 8.36 26 9.62
7 8.45 27 9.72
8 8.52 28 9.76
9 8.55 29 9.77
10 8.59 30 9.77
11 8.94 31 9.78
12 8.95 32 9.79
13 8.96 33 9.80
14 9.14 34 9.90
15 9.15 35 9.99
16 9.15 36 9.99
17 9.15 37 10.07
18 9.15 38 10.22
19 9.36 39 10.42
20 9.36 40 10.82
Count 40
Mean 9.30
Minimum 8.07
Maximum 10.82
Standar Deviation 0.67
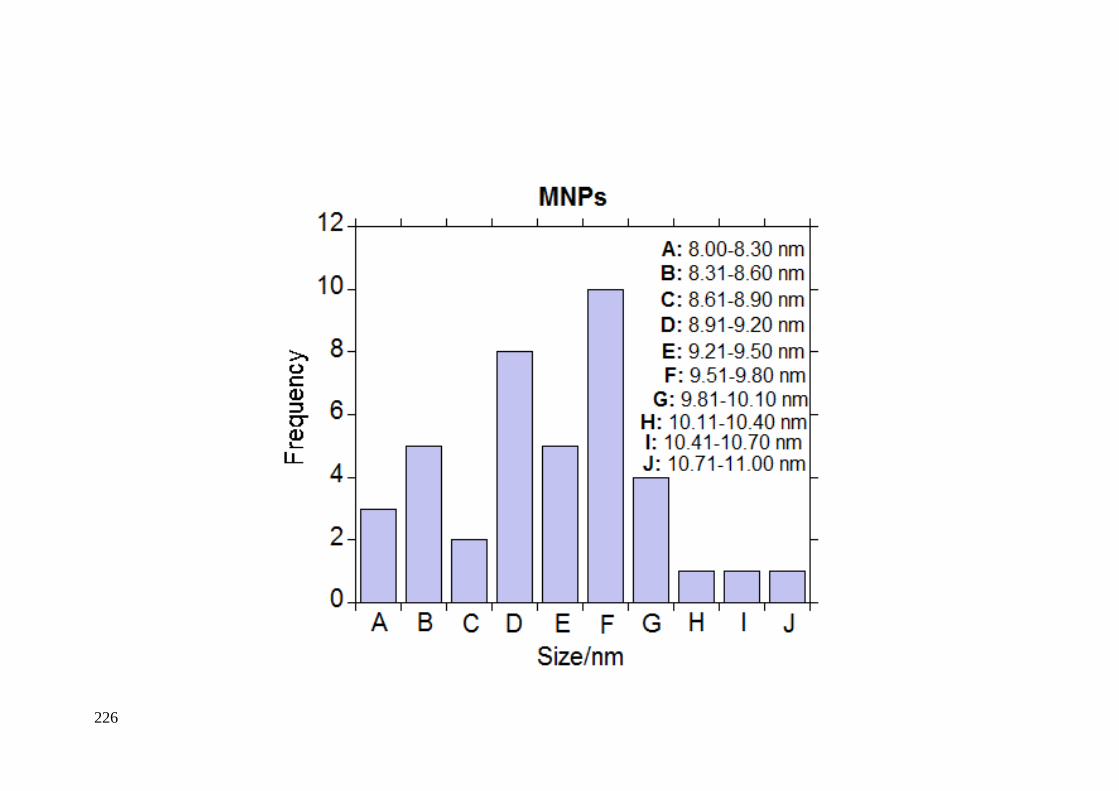
226

227

228
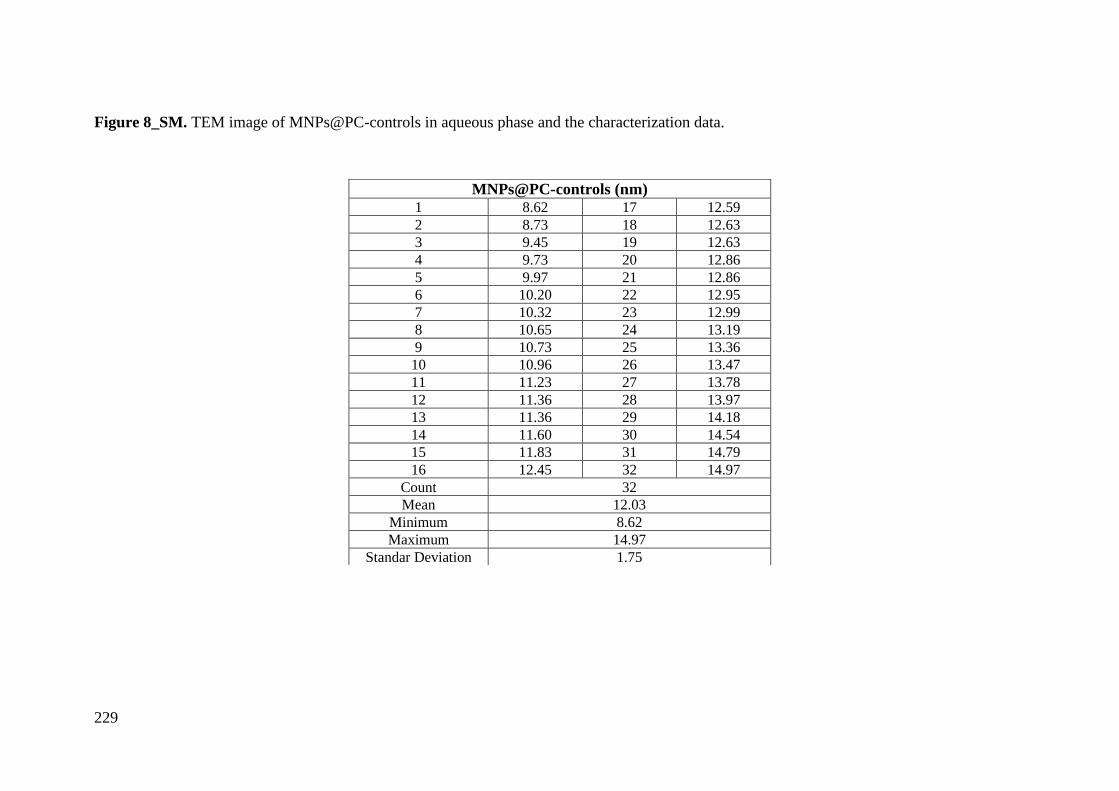
229
Figure 8_SM. TEM image of MNPs@PC-controls in aqueous phase and the characterization data.
MNPs@PC-controls (nm) 1 8.62 17 12.59
2 8.73 18 12.63
3 9.45 19 12.63
4 9.73 20 12.86
5 9.97 21 12.86
6 10.20 22 12.95
7 10.32 23 12.99
8 10.65 24 13.19
9 10.73 25 13.36
10 10.96 26 13.47
11 11.23 27 13.78
12 11.36 28 13.97
13 11.36 29 14.18
14 11.60 30 14.54
15 11.83 31 14.79
16 12.45 32 14.97
Count 32
Mean 12.03
Minimum 8.62
Maximum 14.97
Standar Deviation 1.75
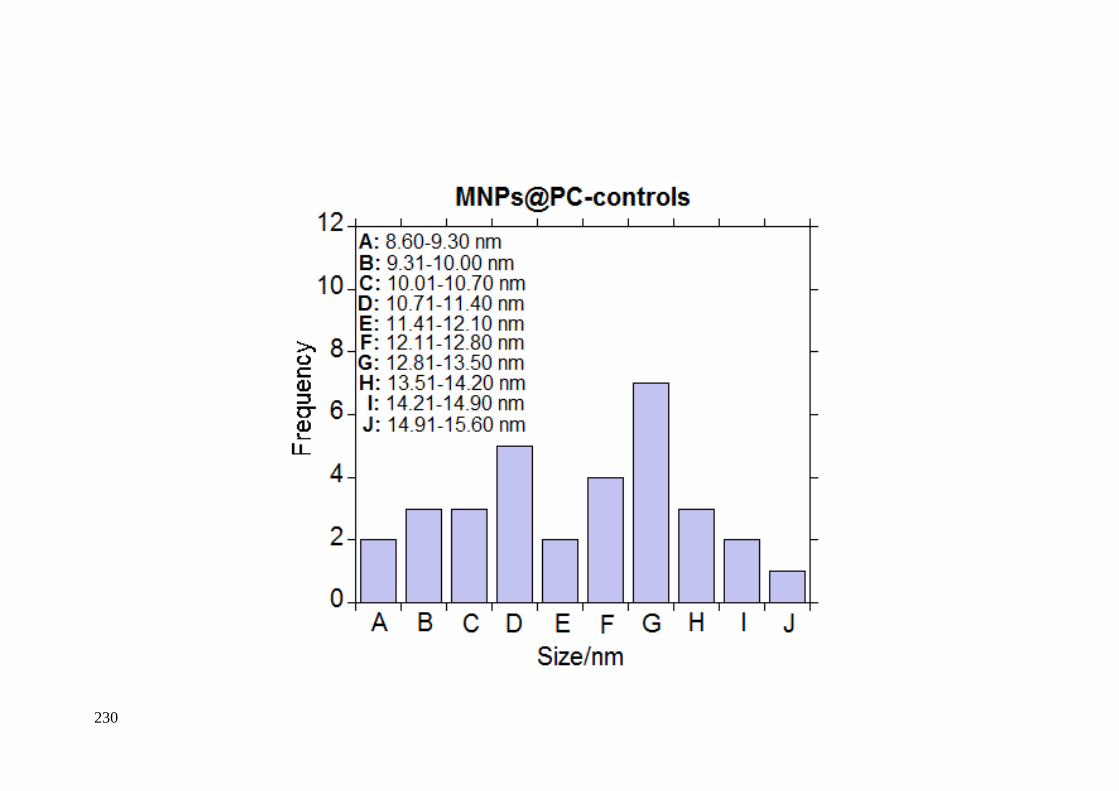
230

231

232
Figure 9_SM. TEM image of MNPs@PC-TNBC in aqueous phase and the characterization data.
MNPs@PC-TNBC (nm) 1 7.63 16 12.56
2 8.49 17 12.66
3 9.57 18 12.71
4 10.65 19 12.74
5 11.13 20 12.75
6 11.26 21 13.2
7 11.36 22 13.34
8 11.82 23 13.42
9 11.83 24 13.76
10 11.89 25 14
11 11.96 26 14.09
12 11.97 27 14.61
13 12.15 28 14.76
14 12.17 29 15.01
15 12.55 30 15.54
Count 30
Mean 12.39
Minimum 7.63
Maximum 15.54
Standar Deviation 1.78
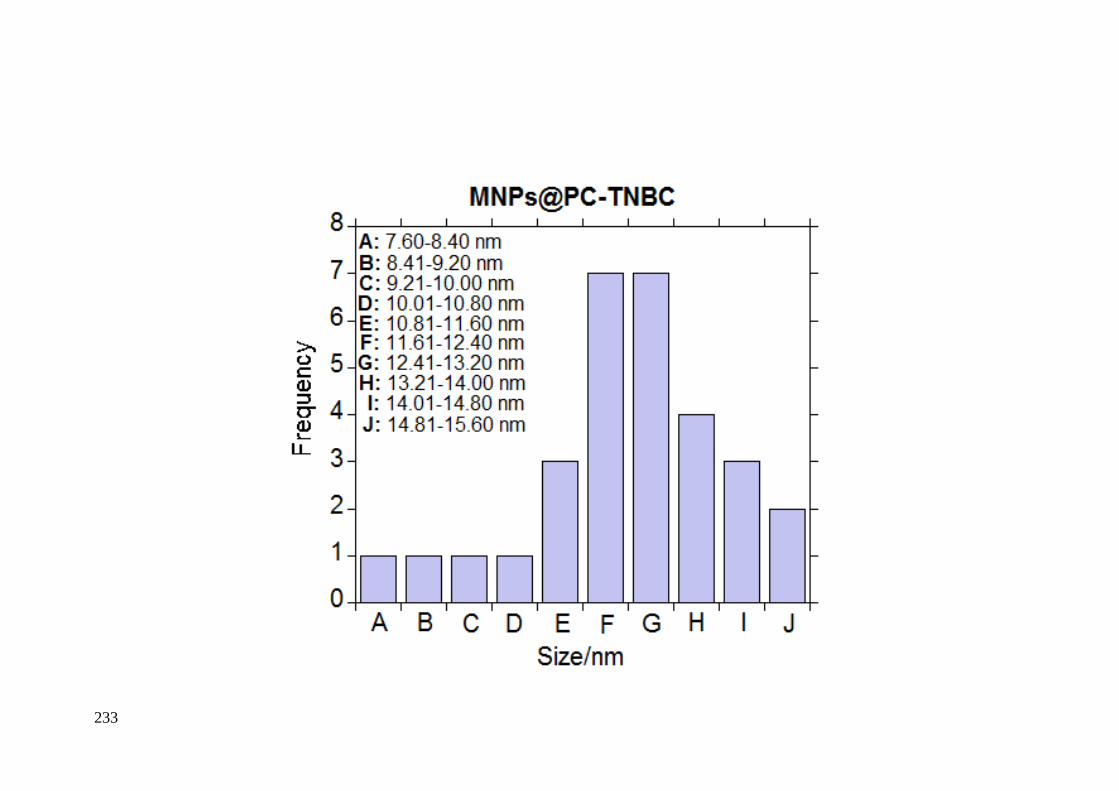
233

234
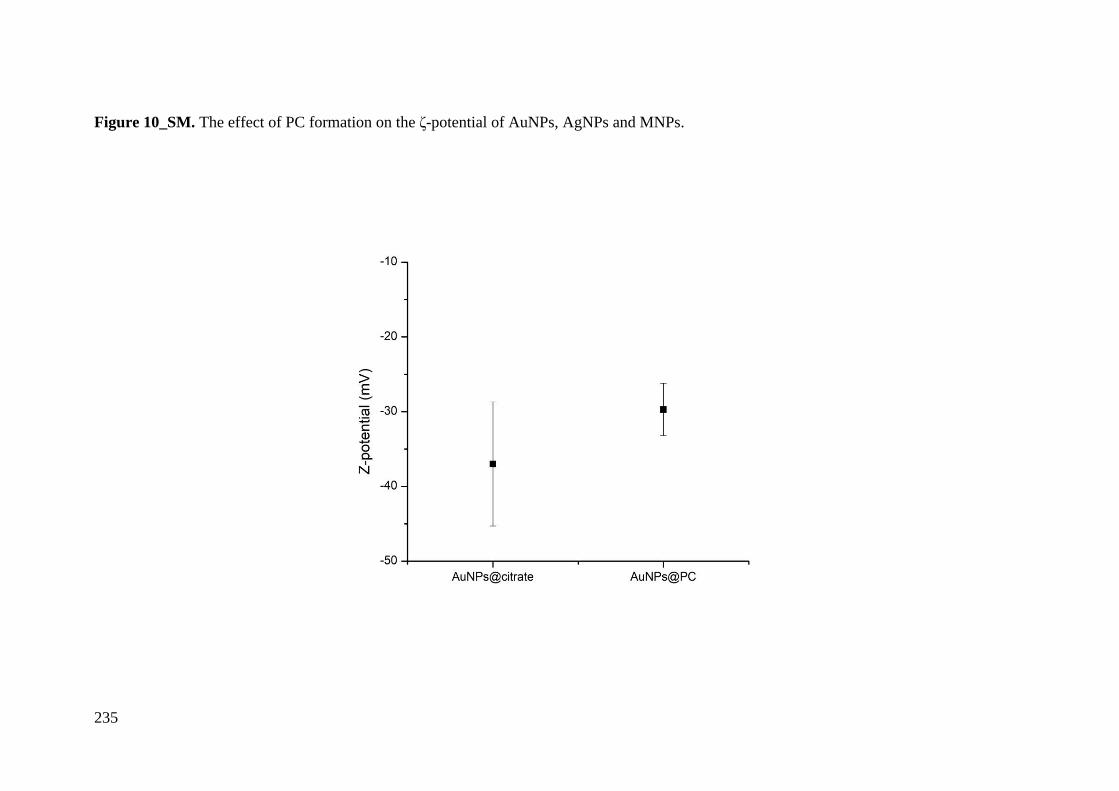
235
Figure 10_SM. The effect of PC formation on the ζ-potential of AuNPs, AgNPs and MNPs.
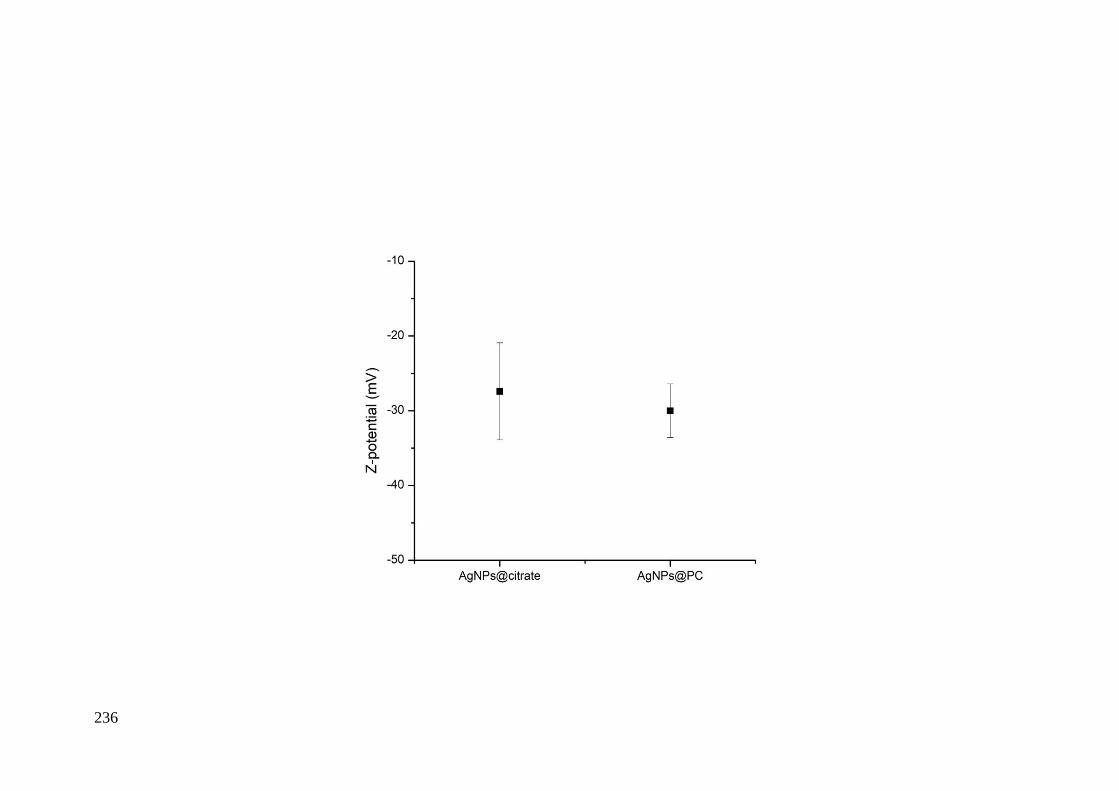
236
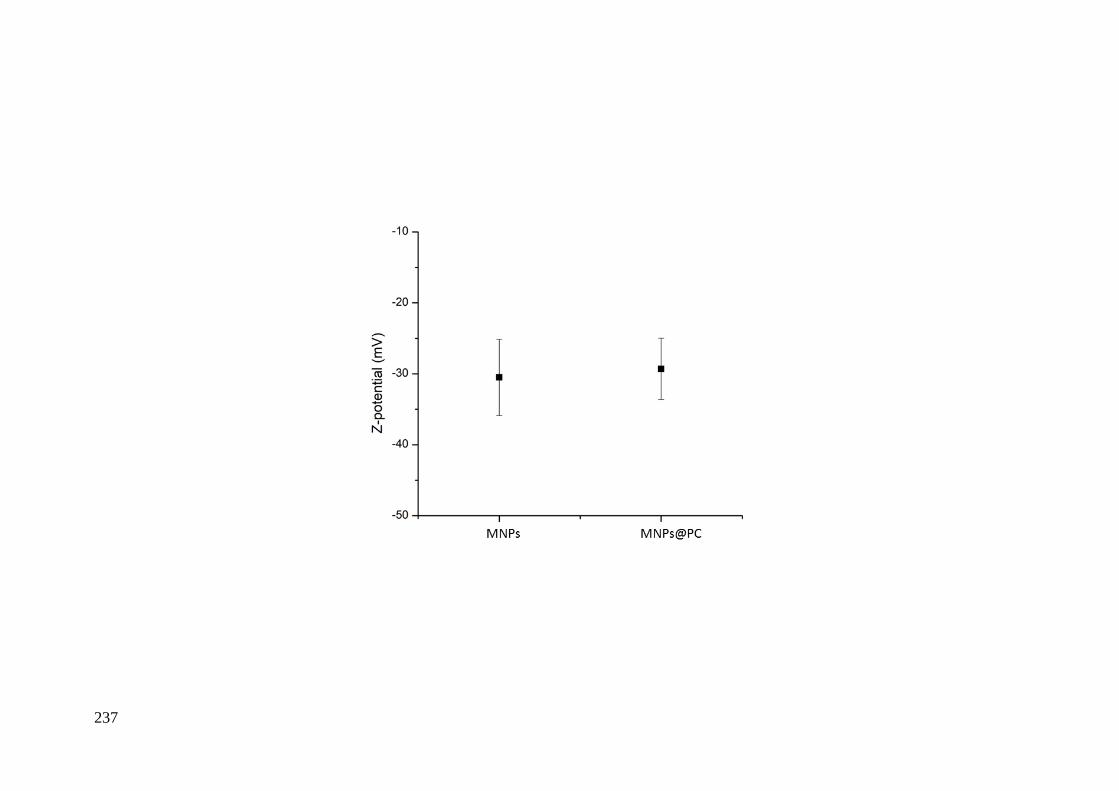
237
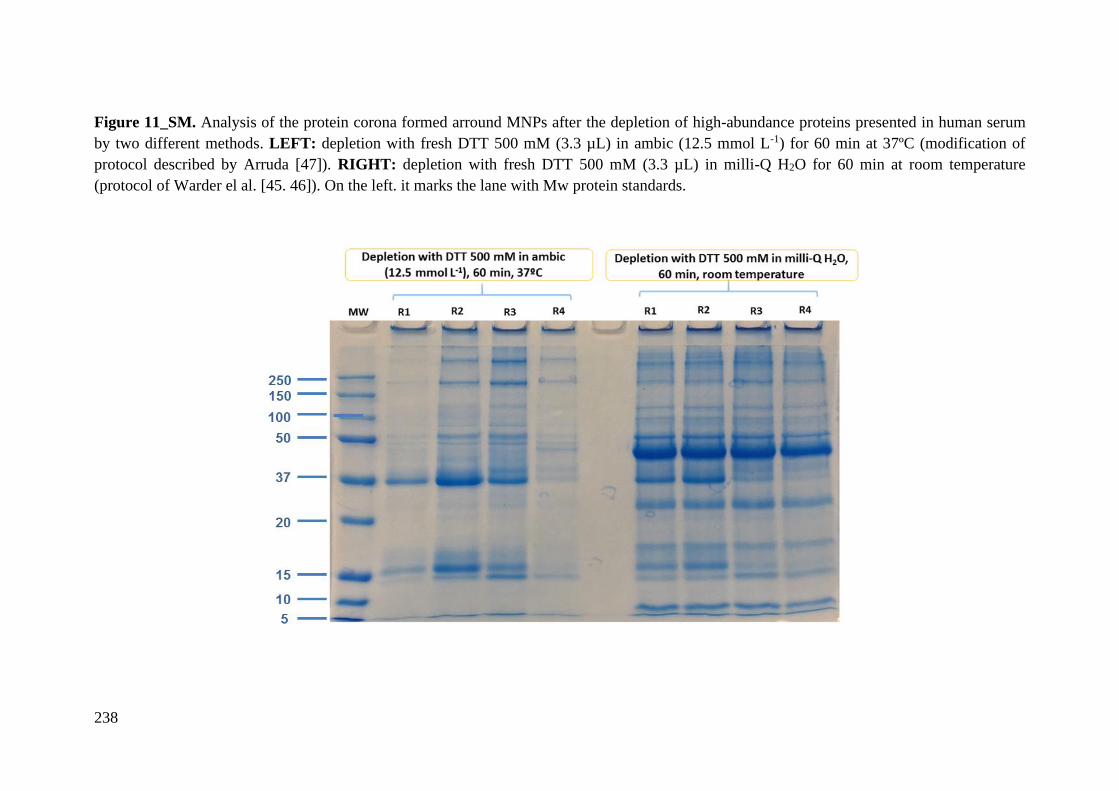
238
Figure 11_SM. Analysis of the protein corona formed arround MNPs after the depletion of high-abundance proteins presented in human serum
by two different methods. LEFT: depletion with fresh DTT 500 mM (3.3 µL) in ambic (12.5 mmol L-1) for 60 min at 37ºC (modification of
protocol described by Arruda [47]). RIGHT: depletion with fresh DTT 500 mM (3.3 µL) in milli-Q H2O for 60 min at room temperature
(protocol of Warder el al. [45. 46]). On the left. it marks the lane with Mw protein standards.
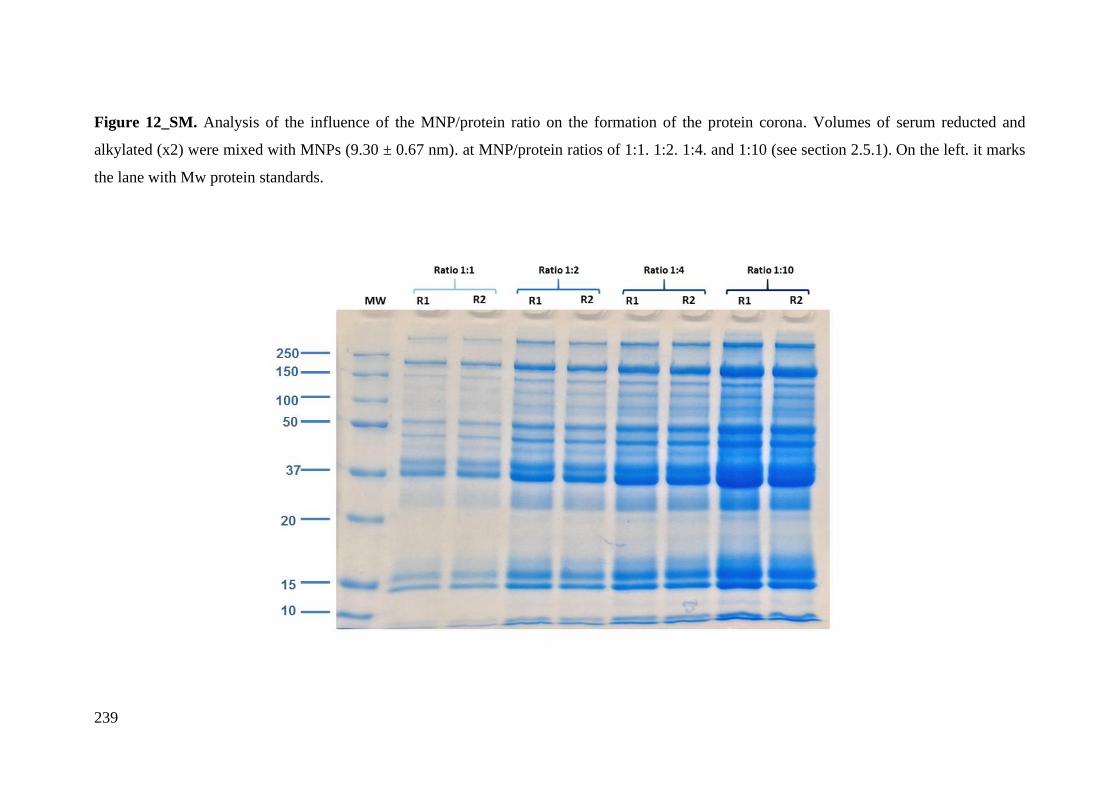
239
Figure 12_SM. Analysis of the influence of the MNP/protein ratio on the formation of the protein corona. Volumes of serum reducted and
alkylated (x2) were mixed with MNPs (9.30 ± 0.67 nm). at MNP/protein ratios of 1:1. 1:2. 1:4. and 1:10 (see section 2.5.1). On the left. it marks
the lane with Mw protein standards.

240
Table 1_SM. Analysis of the protein corona formed around AuNPs (10.02 ± 0.91 nm). AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm)
after their incubation with serum aliquots (x2) from eight different healthy controls (6 protein samples per individual: 2 treated with AuNPs. 2
with AgNPs and 2 with MNPs). The accession number. gene name. species (Human). molecular weight and protein function were reported. Grey
color: 149 proteins commonly found in the protein corona of AuNPs (10.02 ± 0.91 nm). AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm).
Pink color: 71 identified corona proteins exclusively bound to the AuNPs (10.02 ± 0.91 nm). Yellow color: 85 identified corona proteins
exclusively bound to the AgNPs (9.73 ± 1.70 nm). Blue: 46 identified corona proteins exclusively bound to the MNPs (9.30 ± 0.67 nm). Orange
color: 56 identified corona proteins commonly bound to the AuNPs (10.02 ± 0.91 nm) and AgNPs (9.73 ± 1.70 nm). Violet color: 9 identified
corona proteins commonly bound to the AuNPs (10.02 ± 0.91 nm) and MNPs (9.30 ± 0.67 nm). Green color: 2 identified corona proteins
commonly bound to the AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm).

241
Protein Name UniProt Name Entry Name Gene AuNPs AgNPs MNPs Mass (kDa) Function
Acid-sensing ion channel 2 ASIC2_HUMAN Q16515 ASIC2 X 57.7
Cation channel with high affinity for
sodium. which is gated by
extracellular protons and inhibited by
the diuretic amiloride
Acrosomal protein KIAA1210 K1210_HUMAN Q9ULL0 KIAA1210 X 187.0 -
Adenylate kinase isoenzyme 6 KAD6_HUMAN Q9Y3D8 AK6 X 20.0 Catalytic activity
Adhesion G protein-coupled
receptor L4 AGRL4_HUMAN Q9HBW9 ADGRL4 X 77.8
Endothelial orphan receptor that acts
as a key regulator of angiogenesis
Adipocyte plasma membrane-
associated protein APMAP_HUMAN Q9HDC9 APMAP X 46.5
May play a role in adipocyte
differentiation
Adiponectin ADIPO_HUMAN Q15848 ADIPOQ X 26.4
Important adipokine involved in the
control of fat metabolism and insulin
sensitivity. with direct anti-diabetic.
anti-atherogenic and anti-
inflammatory activities
Afamin AFAM_HUMAN P43652 AFM X X X 69.1 Transporter activity
Alpha-1-acid glycoprotein 1 A1AG1_HUMAN P02763 ORM1 X X X 23.5 Transporter activity
Alpha-1-acid glycoprotein 2 A1AG2_HUMAN P19652 ORM2 X X X 23.6 Transporter activity
Alpha-1-antichymotrypsin AACT_HUMAN P01011 SERPINA3 X X X 47.6 Protease inhibitor
Alpha-1-antitrypsin A1AT_HUMAN P01009 SERPINA1 X X X 46.7 Protease inhibitor
Alpha-1B-glycoprotein A1BG_HUMAN P04217 A1BG X X X 54.3 Neutrophil and platelet degranulation
Alpha-2-antiplasmin A2AP_HUMAN P08697 SERPINF2 X X X 54.6 Protease inhibitor
Alpha-2-HS-glycoprotein FETUA_HUMAN P02765 AHSG X X X 39.3
Promotes endocitosis. brain
development and the formation of
bone tissue
Alpha-2-macroglobulin A2MG_HUMAN P01023 A2M X X X 163.3 Immune response
Angiotensinogen ANGT_HUMAN P01019 AGT X X X 53.1 Regulator of blood pressure. body
fluid and electrolyte homeostasis
Ankyrin repeat domain-
containing protein 33B AN33B_HUMAN A6NCL7
ANKRD33
B X 53.9 -
Ankyrin repeat domain-
containing protein SOWAHC SWAHC_HUMAN Q53LP3 SOWAHC X 55.7 -
Annexin A6 ANXA6_HUMAN P08133 ANXA6 X 75.9 May associate with CD21
Antithrombin-III ANT3_HUMAN P01008 SERPINC1 X X X 52.6 Protease inhibitor
AP-3 complex subunit beta-2 AP3B2_HUMAN Q13367 AP3B2 X X 119.1 Transporter activity
Apolipoprotein A-I APOA1_HUMAN P02647 APOA1 X X X 30.8 Metabolism
Apolipoprotein A-II APOA2_HUMAN P02652 APOA2 X X X 11.2 Transport
Apolipoprotein A-IV APOA4_HUMAN P06727 APOA4 X X X 45.4 Hepatic transcellukar lipid transport

242
Apolipoprotein B-100 APOB_HUMAN P04114 APOB X X X 515.6
Recognition signal for the cellular
binding and internalization of LDL
particles by the apoB/E receptor
Apolipoprotein C-I APOC1_HUMAN P02654 APOC1 X X X 9.3
Inhibitor of lipoprotein binding to the
low density lipoprotein (LDL)
receptor
Apolipoprotein C-II APOC2_HUMAN P02655 APOC2 X X X 11.3 Enzyme regulator activity
(lipoprotein lipase activator)
Apolipoprotein C-III APOC3_HUMAN P02656 APOC3 X X X 10.8 Enzyme regulator activity
(lipoprotein lipase inhibitor)
Apolipoprotein C-IV APOC4_HUMAN P55056 APOC4 X 14.5 Biological regulation
Apolipoprotein D APOD_HUMAN P05090 APOD X X X 21.3 Transporter activity
Apolipoprotein E APOE_HUMAN P02649 APOE X X X 36.1 Transporter activity and cell uptake
Apolipoprotein F APOF_HUMAN Q13790 APOF X X X 35.4 Transporter activity
Apolipoprotein L1 APOL1_HUMAN O14791 APOL1 X X X 43.9 Transporter activity
Apolipoprotein M APOM_HUMAN O95445 APOM X X X 21.2 Transporter activity
Apolipoprotein(a) APOA_HUMAN P08519 LPA X X X 501.3 Catalytic activity
Arf-GAP with SH3 domain.
ANK repeat and PH domain-
containing protein 1
ASAP1_HUMAN Q9ULH1 ASAP1 X 125.5 Plays a role in ciliogenesis
Armadillo repeat-containing X-
linked protein 5 ARMX5_HUMAN Q6P1M9 ARMCX5 X 62.3 -
Arrestin-C ARRC_HUMAN P36575 ARR3 X 42.8 May play a role in an as yet undefined
retina-specific signal transduction
ATP-binding cassette sub-family
B member 5 ABCB5_HUMAN Q2M3G0 ABCB5 X 138.6 Transporter activity
ATP-binding cassette sub-family
C member 8 ABCC8_HUMAN Q09428 ABCC8 X 176.9
Regulator of ATP-sensitive K+
channels and insulin release
ATP-binding cassette sub-family
F member 1 ABCF1_HUMAN Q8NE71 ABCF1 X X X 95.9 Transporter activity
Attractin ATRN_HUMAN O75882 ATRN X X X 158.5
Involved in the initial immune cell
clustering during inflammatory
response and may regulate
chemotactic activity of chemokines
Beta-Ala-His dipeptidase CNDP1_HUMAN Q96KN2 CNDP1 X X 56.7 Catalytic activity
Beta-2-glycoprotein 1 APOH_HUMAN P02749 APOH X X X 38.3 Transporter activity
Beta-2-microglobulin B2MG_HUMAN P61769 B2M X 13.7
Component of the class I major
histocompatibility complex (MHC).
Involved in the presentation of
peptide antigens to the immune

243
system.
Beta-ureidopropionase BUP1_HUMAN Q9UBR1 UPB1 X 43.1 Catalytic activity
Biotinidase BTD_HUMAN P43251 BTD X X 61.1 Catalytic activity
Bone morphogenetic protein 10 BMP10_HUMAN O95393 BMP10 X 48.0
Inhibits endothelial cell migration and
growth. May reduce cell migration
and cell matrix adhesion in breast
cancer cell lines.
Carbonic anhydrase 1 CAH1_HUMAN P00915 CA1 X 28.9 Catalytic activity
Carboxypeptidase B2 CBPB2_HUMAN Q96IY4 CPB2 X X X 48.4 Catalytic activity
Carboxypeptidase N catalytic
chain CBPN_HUMAN P15169 CPN1 X X X 52.3 Catalytic activity
Carboxypeptidase N subunit 2 CPN2_HUMAN P22792 CPN2 X X X 60.6 Enzyme regulator activity
Carboxypeptidase Q CBPQ_HUMAN Q9Y646 CPQ X 51.8 Catalytic activity
Cardiomyopathy-associated
protein 5 CMYA5_HUMAN Q8N3K9 CMYA5 X 449.2
May serve as an anchoring protein
that mediates the subcellular
compartmentation of protein kinase A
(PKA) via binding to PRKAR2A
Cartilage oligomeric matrix
protein COMP_HUMAN P49747 COMP X X 82.9
May play a role in the structural
integrity of cartilage via its
interaction with other extracellular
matrix proteins such as the collagens
and fibronectin
CASP8-associated protein 2 C8AP2_HUMAN Q9UKL3 CASP8AP2 X 222.6
Involved in TNF-alpha-induced
activation of NF-kappa-B via a
TRAF2-dependent pathway
Cathelicidin antimicrobial
peptide CAMP_HUMAN P49913 CAMP X 19.3 Antibacterial activity
C4b-binding protein alpha chain C4BPA_HUMAN P04003 C4BPA X X X 67.1 Enzyme regulator activity (inhibitor)
C4b-binding protein beta chain C4BPB_HUMAN P20851 C4BPB X X X 28.4 Enzyme regulator activity (inhibitor)
CCR4-NOT transcription
complex subunit 4 CNOT4_HUMAN O95628 CNOT4 X 63.5 Catalytic activity
CDK5 regulatory subunit-
associated protein 2 CK5P2_HUMAN Q96SN8 CDK5RAP2 X 215.0
Involved in regulation of mitotic
spindle orientation
CD5 antigen-like CD5L_HUMAN O43866 CD5L X X X 38.1 Regulator of lipid synthesis
CD44 antigen CD44_HUMAN P16070 CD44 X X 81.5
Mediates cell-cell and cell-matrix
interactions through its affinity for
hyaluronic acid (HA)
CD83 antigen CD83_HUMAN Q01151 CD83 X 23.0
May play a significant role in antigen
presentation or the cellular
interactions that follow lymphocyte
244
activation
Centrosome-associated protein
350 CE350_HUMAN Q5VT06 CEP350 X 350.9 Structural
Ceruloplasmin CERU_HUMAN P00450 CP X X X 122.2 Catalytic activity
Chloride intracellular channel
protein 1 CLIC1_HUMAN O00299 CLIC1 X 26.9
Involved in regulation of the cell
cycle
Cholesteryl ester transfer protein CETP_HUMAN P11597 CETP X X 54.8 Transporter activity
Cholinesterase CHLE_HUMAN P06276 BCHE X 68.4 Catalytic activity
Cip1-interacting zinc finger
protein CIZ1_HUMAN Q9ULV3 CIZ1 X 100.0
May regulate the subcellular
localization of CIP/WAF1
Clusterin CLUS_HUMAN P10909 CLU X X X 52.5 Regulation of cell proliferation
Coagulation factor XIII A chain F13A_HUMAN P00488 F13A1 X X 83.3 Catalytic activity
Coagulation factor XIII B chain F13B_HUMAN P05160 F13B X X X 75.5 Enzyme regulator activity
Coagulation factor IX FA9_HUMAN P00740 F9 X X X 51.8 Catalytic activity
Coagulation factor V FA5_HUMAN P12259 F5 X X X 251.7 Enzyme regulator activity
Coagulation factor XII FA12_HUMAN P00748 F12 X X X 67.8 Catalytic activity
Coiled-coil domain-containing
protein 90B. mitochondrial CC90B_HUMAN Q9GZT6 CCDC90B X 29.5 -
Coiled-coil domain-containing
protein 127 CC127_HUMAN Q96BQ5 CCDC127 X 30.8 -
Coiled-coil domain-containing
protein 146 CC146_HUMAN Q8IYE0 CCDC146 X 112.8 -
Collectin-11 COL11_HUMAN Q9BWP8 COLEC11 X 28.7 Plays a role in innate immunity.
apoptosis and embryogenesis
Complement C1q subcomponent
subunit A C1QA_HUMAN P02745 C1QA X X X 26.1 Enzyme regulator activity
Complement C1q subcomponent
subunit B C1QB_HUMAN P02746 C1QB X X X 26.7 Enzyme regulator activity
Complement C1q subcomponent
subunit C C1QC_HUMAN P02747 C1QC X X X 25.8 Enzyme regulator activity
Complement C1r subcomponent C1R_HUMAN P00736 C1R X X X 80.1 Catalytic activity
Complement C1r subcomponent-
like protein C1RL_HUMAN Q9NZP8 C1RL X X X 53.5 Enzyme regulator activity
Complement C1s subcomponent C1S_HUMAN P09871 C1S X X X 76.7 Catalytic activity
Complement C2 CO2_HUMAN P06681 C2 X X X 83.3 Catalytic activity
Complement C3 CO3_HUMAN P01024 C3 X X X 187.1 Immune response
Complement C4-A CO4A_HUMAN P0C0L4 C4A X X X 192.8 Inflammatory response
Complement C4-B CO4B_HUMAN P0C0L5 C4B X X X 192.7 Enzyme regulator activity
Complement C5 CO5_HUMAN P01031 C5 X X X 188.3 Enzyme regulator activity

245
Complement component C6 CO6_HUMAN P13671 C6 X X X 104.8 Structural
Complement component C7 CO7_HUMAN P10643 C7 X X X 93.5 Structural
Complement component C8
alpha chain CO8A_HUMAN P07357 C8A X X X 65.2 Structural
Complement component C8 beta
chain CO8B_HUMAN P07358 C8B X X X 67.1 Structural
Complement component C8
gamma chain CO8G_HUMAN P07360 C8G X X X 22.3 Structural
Complement component C9 CO9_HUMAN P02748 C9 X X X 63.2 Structural
Complement factor B CFAB_HUMAN P00751 CFB X X X 85.5 Immune response
Complement factor D CFAD_HUMAN P00746 CFD X X X 27.1 Catalytic activity
Complement factor H CFAH_HUMAN P08603 CFH X X X 139.1 Enzyme regulator activity
Complement factor H-related
protein 1 FHR1_HUMAN Q03591 CFHR1 X X X 37.6 Plays a role in lipid metabolism
Complement factor H-related
protein 2 FHR2_HUMAN P36980 CFHR2 X X X 30.6 Plays a role in lipid metabolism
Complement factor H-related
protein 3 FHR3_HUMAN Q02985 CFHR3 X 37.3 Involved in complement regulation
Complement factor H-related
protein 4 FHR4_HUMAN Q92496 CFHR4 X 65.3 Plays a role in lipid metabolism
Complement factor I CFAI_HUMAN P05156 CFI X X X 65.7 Catalytic activity
Conserved oligomeric Golgi
complex subunit 1 COG1_HUMAN Q8WTW3 COG1 X 108.9 Required for normal Golgi function
Contactin-1 CNTN1_HUMAN Q12860 CNTN1 X X 113.3 Mediates cell surface interactions
during nervous system development.
Copine-1 CPNE1_HUMAN Q99829 CPNE1 X 59.1
Calcium-dependent phospholipid-
binding protein that plays a role in
calcium-mediated intracellular
processes
Corticosteroid-binding globulin CBG_HUMAN P08185 SERPINA6 X X 45.1 Transporter activity
C-reactive protein CRP_HUMAN P02741 CRP X 25.0 Displays several functions associated
with host defense
Cysteine-rich secretory protein 3 CRIS3_HUMAN P54108 CRISP3 X 27.6 Defense response
Cytoplasmic polyadenylation
element-binding protein 1 CPEB1_HUMAN Q9BZB8 CPEB1 X 62.6
Sequence-specific RNA-binding
protein that regulates mRNA
cytoplasmic polyadenylation and
translation initiation during oocyte
maturation. early development and at
postsynapse sites of neurons

246
Death-associated protein kinase 2 DAPK2_HUMAN Q9UIK4 DAPK2 X 42.9 Catalyctic activity
Disheveled-associated activator
of morphogenesis 2 DAAM2_HUMAN Q86T65 DAAM2 X 123.5
Key regulator of the Wnt signaling
pathway
DNA polymerase eta POLH_HUMAN Q9Y253 POLH X 46.3 Catalyctic activity
Dolichol-phosphate
mannosyltransferase subunit 1 DPM1_HUMAN O60762 DPM1 X 29.6 Catalyctic activity
Dopamine beta-hydroxylase DOPO_HUMAN P09172 DBH X X 69.1 Catalyctic activity
Double-stranded RNA-specific
editase 1 RED1_HUMAN P78563 ADARB1 X 80.8 Catalyctic activity
EF-hand calcium-binding
domain-containing protein 7 EFCB7_HUMAN A8K855 EFCAB7 X 71.9
Component of the EvC complex that
positively regulates ciliary Hedgehog
(Hh) signaling
EGF-containing fibulin-like
extracellular matrix protein 1 FBLN3_HUMAN Q12805 EFEMP1 X 54.6
Binds EGFR. the EGF receptor.
inducing EGFR autophosphorylation
and the activation of downstream
signaling pathways. May play a role
in cell adhesion and migration
Electron transfer flavoprotein
subunit beta ETFB_HUMAN P38117 ETFB X 27.8
Required for normal mitochondrial
fatty acid oxidation and normal amino
acid metabolism
Endophilin-A3 SH3G3_HUMAN Q99963 SH3GL3 X 39.3
Implicated in endocytosis. May
recruit other proteins to membranes
with high curvature
ERC protein 2 ERC2_HUMAN O15083 ERC2 X 110.5
Involved in the organization of the
cytomatrix at the nerve terminals
active zone (CAZ) which regulates
neurotransmitter release
E3 ubiquitin-protein ligase
HUWE1 HUWE1_HUMAN Q7Z6Z7 HUWE1 X 481.9 Catalytic activity
Eukaryotic translation initiation
factor 3 subunit C EIF3C_HUMAN Q99613 EIF3C X 105.3
Component of the eukaryotic
translation initiation factor 3 (eIF-3)
complex. which is required for
several steps in the initiation of
protein synthesis
Extracellular matrix protein 1 ECM1_HUMAN Q16610 ECM1 X X X 60.7 Enzyme regulator inhibitor
Fas-binding factor 1 FBF1_HUMAN Q8TES7 FBF1 X 125.4 Keratin-binding protein required for
epithelial cell polarization
FERM. ARHGEF and pleckstrin
domain-containing protein 1 FARP1_HUMAN Q9Y4F1 FARP1 X 118.6
Functions as guanine nucleotide
exchange factor for RAC1
FERM and PDZ domain- FRPD3_HUMAN Q5JV73 FRMPD3 X 199.2 Neutrophil degranulation
247
containing protein 3
Fetuin-B FETUB_HUMAN FETUB FETUB X X X 42.1 Catalytic activity
Fibronectin FINC_HUMAN P02751 FN1 X X X 262.6 Growth. migration and differenciation
Fibrinogen alpha chain FIBA_HUMAN P02671 FGA X X X 94.9 Coagulation. immune response
Ficolin-2 FCN2_HUMAN Q15485 FCN2 X X X 34.0 Immunity response
Ficolin-3 FCN3_HUMAN O75636 FCN3 X X X 32.9 Immunity response
Fibulin-1 FBLN1_HUMAN P23142 FBLN1 X X 77.2 Structural
FYVE. RhoGEF and PH domain-
containing protein 5 FGD5_HUMAN Q6ZNL6 FGD5 X 159.9
Activates CDC42. a member of the
Ras-like family of Rho- and Rac
proteins
Galectin-3-binding protein LG3BP_HUMAN Q08380 LGALS3BP X X 65.3 Stimulate host defense against viruses
and tumor cells
Gelsolin GELS_HUMAN P06396 GSN X X X 85.7 Actin regulation (assembly)
Glutathione peroxidase 3 GPX3_HUMAN P22352 GPX3 X 25.5 Catalytic activity
Haptoglobin HPT_HUMAN P00738 HP X X X 45.2
Prevents loss of iron through the
kidneys. protecting them from
damage by hemoglobin
Haptoglobin-related protein HPTR_HUMAN P00739 HPR X X X 39 Binds hemoglobin as efficiently as
haptoglobin
HEAT repeat-containing protein
4 HEAT4_HUMAN Q86WZ0 HEATR4 X 117.2 -
Heat shock 70 kDa protein 1-like HS71L_HUMAN P34931 HSPA1L X 70.4
Implicates in the protection of the
proteome from stress. folding and
transport of newly synthesized
polypeptides. activation of proteolysis
of misfolded proteins and the
formation and dissociation of protein
complexes
Hemoglobin subunit alpha HBA_HUMAN P69905 HBA1 X X X 15.3 Transporter activity
Hemoglobin subunit beta HBB_HUMAN P68871 HBB X X X 15.9 Transporter activity
Hemopexin HEMO_HUMAN P02790 HPX X X X 51.7 Transporter activity
Heparan sulfate glucosamine 3-
O-sulfotransferase 5 HS3S5_HUMAN Q8IZT8 HS3ST5 X 40.4 Catalytic activity
Heparin cofactor 2 HEP2_HUMAN P05546 SERPIND1 X X X 57.1 Enzyme regulator activity (inhibitor)
Hepatocyte growth factor
activator HGFA_HUMAN Q04756 HGFAC X 70.7 Enzyme regulator activity (activator)
Hepatocyte growth factor-like
protein HGFL_HUMAN P26927 MST1 X 80.3 Catalytic activity
Histidine-rich glycoprotein HRG_HUMAN P04196 HRG X X X 59.6 Inhibits fibrinolysis and reduces
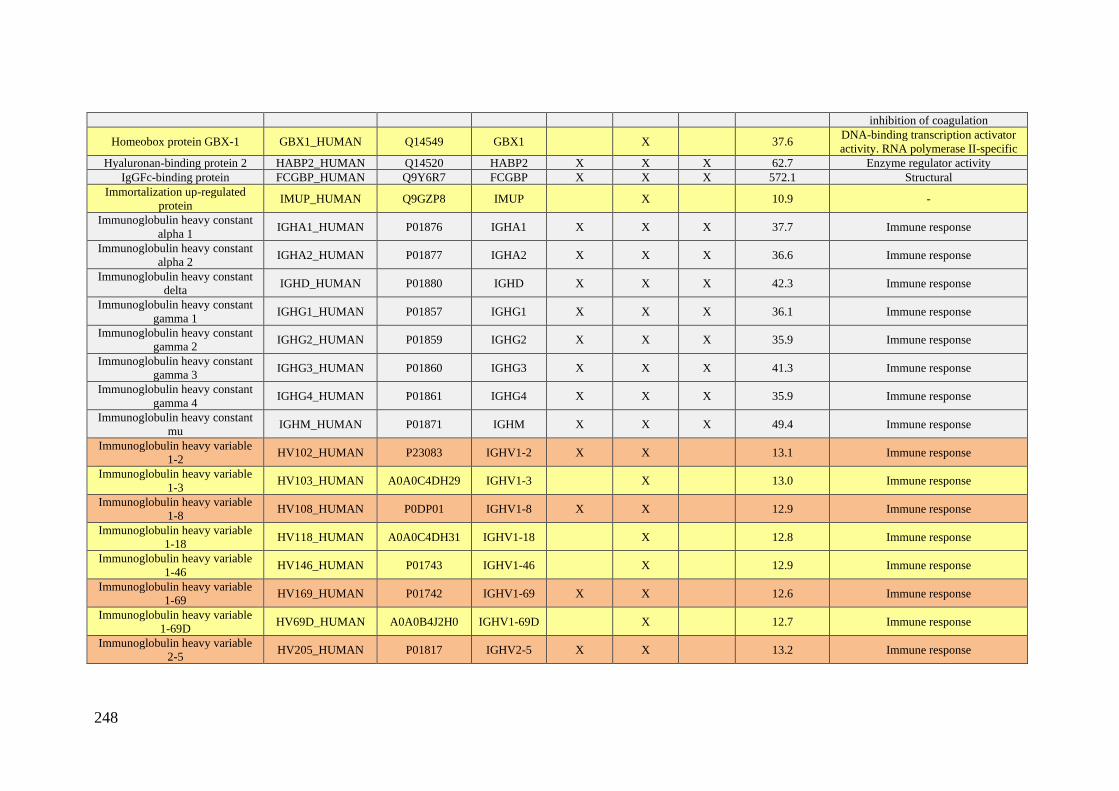
248
inhibition of coagulation
Homeobox protein GBX-1 GBX1_HUMAN Q14549 GBX1 X 37.6 DNA-binding transcription activator
activity. RNA polymerase II-specific
Hyaluronan-binding protein 2 HABP2_HUMAN Q14520 HABP2 X X X 62.7 Enzyme regulator activity
IgGFc-binding protein FCGBP_HUMAN Q9Y6R7 FCGBP X X X 572.1 Structural
Immortalization up-regulated
protein IMUP_HUMAN Q9GZP8 IMUP X 10.9 -
Immunoglobulin heavy constant
alpha 1 IGHA1_HUMAN P01876 IGHA1 X X X 37.7 Immune response
Immunoglobulin heavy constant
alpha 2 IGHA2_HUMAN P01877 IGHA2 X X X 36.6 Immune response
Immunoglobulin heavy constant
delta IGHD_HUMAN P01880 IGHD X X X 42.3 Immune response
Immunoglobulin heavy constant
gamma 1 IGHG1_HUMAN P01857 IGHG1 X X X 36.1 Immune response
Immunoglobulin heavy constant
gamma 2 IGHG2_HUMAN P01859 IGHG2 X X X 35.9 Immune response
Immunoglobulin heavy constant
gamma 3 IGHG3_HUMAN P01860 IGHG3 X X X 41.3 Immune response
Immunoglobulin heavy constant
gamma 4 IGHG4_HUMAN P01861 IGHG4 X X X 35.9 Immune response
Immunoglobulin heavy constant
mu IGHM_HUMAN P01871 IGHM X X X 49.4 Immune response
Immunoglobulin heavy variable
1-2 HV102_HUMAN P23083 IGHV1-2 X X 13.1 Immune response
Immunoglobulin heavy variable
1-3 HV103_HUMAN A0A0C4DH29 IGHV1-3 X 13.0 Immune response
Immunoglobulin heavy variable
1-8 HV108_HUMAN P0DP01 IGHV1-8 X X 12.9 Immune response
Immunoglobulin heavy variable
1-18 HV118_HUMAN A0A0C4DH31 IGHV1-18 X 12.8 Immune response
Immunoglobulin heavy variable
1-46 HV146_HUMAN P01743 IGHV1-46 X 12.9 Immune response
Immunoglobulin heavy variable
1-69 HV169_HUMAN P01742 IGHV1-69 X X 12.6 Immune response
Immunoglobulin heavy variable
1-69D HV69D_HUMAN A0A0B4J2H0 IGHV1-69D X 12.7 Immune response
Immunoglobulin heavy variable
2-5 HV205_HUMAN P01817 IGHV2-5 X X 13.2 Immune response
249
Immunoglobulin heavy variable
2-26 HV226_HUMAN A0A0B4J1V2 IGHV2-26 X 13.2 Immune response
Immunoglobulin heavy variable
2-70D HV70D_HUMAN A0A0C4DH43 IGHV2-70D X X 13.3 Immune response
Immunoglobulin heavy variable
3-7 HV307_HUMAN P01780 IGHV3-7 X X X 12.9 Immune response
Immunoglobulin heavy variable
3-9 HV309_HUMAN P01782 IGHV3-9 X 12.9 Immune response
Immunoglobulin heavy variable
3-11 HV311_HUMAN P01762 IGHV3-11 X 12.9 Immune response
Immunoglobulin heavy variable
3-15 HV315_HUMAN A0A0B4J1V0 IGHV3-15 X X 12.9 Immune response
Immunoglobulin heavy variable
3-20 HV320_HUMAN A0A0C4DH32 IGHV3-20 X 12.7 Immune response
Immunoglobulin heavy variable
3-21 HV321_HUMAN A0A0B4J1V1 IGHV3-21 X 12.8 Immune response
Immunoglobulin heavy variable
3-23 HV323_HUMAN P01764 IGHV3-23 X 12.6 Immune response
Immunoglobulin heavy variable
3-30-3 HVC33_HUMAN P0DP02
IGHV3-30-
3 X X X 13 Immune response
Immunoglobulin heavy variable
3-30-5 HV335_HUMAN P0DP03
IGHV3-30-
5 X X X 12.9 Immune response
Immunoglobulin heavy variable
3-43D HV43D_HUMAN P0DP04 IGHV3-43D X X 13.1 Immune response
Immunoglobulin heavy variable
3-48 HV348_HUMAN P01763 IGHV3-48 X 12.8 Immune response
Immunoglobulin heavy variable
3-49 HV349_HUMAN A0A0A0MS15 IGHV3-49 X X 13.0 Immune response
Immunoglobulin heavy variable
3-53 HV353_HUMAN P01767 IGHV3-53 X 12.8 Immune response
Immunoglobulin heavy variable
3-64D HV64D_HUMAN A0A0J9YX35 IGHV3-64D X X 12.8 Immune response
Immunoglobulin heavy variable
3-66 HV366_HUMAN A0A0C4DH42 IGHV3-66 X X 12.7 Immune response
Immunoglobulin heavy variable
3-72 HV372_HUMAN A0A0B4J1Y9 IGHV3-72 X X X 13.2 Immune response
Immunoglobulin heavy variable
3-73 HV373_HUMAN A0A0B4J1V6 IGHV3-73 X 12.8 Immune response
Immunoglobulin heavy variable
3-74 HV374_HUMAN A0A0B4J1X5 IGHV3-74 X X X 12.8 Immune response
250
Immunoglobulin heavy variable
4-4 HV404_HUMAN A0A075B6R2 IGHV4-4 X X 12.8 Immune response
Immunoglobulin heavy variable
4-28 HV428_HUMAN A0A0C4DH34 IGHV4-28 X X 13.1 Immune response
Immunoglobulin heavy variable
4-30-2 HV432_HUMAN A0A087WSY4
IGHV4-30-
2 X 13.0 Immune response
Immunoglobulin heavy variable
4-34 HV434_HUMAN P06331 IGHV4-34 X X 13.8 Immune response
Immunoglobulin heavy variable
4-38-2 HVD82_HUMAN P0DP08
IGHV4-38-
2 X X X 13.1 Immune response
Immunoglobulin heavy variable
4-39 HV439_HUMAN P01824 IGHV4-39 X 13.9 Immune response
Immunoglobulin heavy variable
4-59 HV459_HUMAN P01825 IGHV4-59 X 12.9 Immune response
Immunoglobulin heavy variable
5-10-1 HV5X1_HUMAN A0A0J9YXX1
IGHV5-10-
1 X X 12.8 Imuune response
Immunoglobulin heavy variable
5-51 HV551_HUMAN A0A0C4DH38 IGHV5-51 X X X 12.7 Immune response
Immunoglobulin heavy variable
6-1 HV601_HUMAN A0A0B4J1U7 IGHV6-1 X X 13.5 Imuune response
Immunoglobulin J chain IGJ_HUMAN P01591 JCHAIN X X X 18.1 Immune response
Immunoglobulin kappa constant IGKC_HUMAN P01834 IGKC X X X 11.8 Immune response
Immunoglobulin kappa variable
1D-8 KVD08_HUMAN A0A087WSZ0 IGKV1D-8 X 12.8 Immune response
Immunoglobulin kappa variable
1D-12 KVD12_HUMAN P01611 IGKV1D-12 X 12.6 Immune response
Immunoglobulin kappa variable
1-5 KV105_HUMAN P01602 IGKV1-5 X X 12.8 Immune response
Immunoglobulin kappa variable
1-6 KV106_HUMAN A0A0C4DH72 IGKV1-6 X 12.7 Immune response
Immunoglobulin kappa variable
1-8 KV108_HUMAN A0A0C4DH67 IGKV1-8 X X X 12.5 Immune response
Immunoglobulin kappa variable
1-9 KV109_HUMAN A0A0C4DH69 IGKV1-9 X X 12.7 Immune response
Immunoglobulin kappa variable
1-13 KV113_HUMAN P0DP09 IGKV1-13 X X 12.5 Immune response
Immunoglobulin kappa variable
1-16 KV116_HUMAN P04430 IGKV1-16 X 12.6 Immune response
Immunoglobulin kappa variable
1-17 KV117_HUMAN P01599 IGKV1-17 X X 12.8 Immune response
251
Immunoglobulin kappa variable
1-27 KV127_HUMAN A0A075B6S5 IGKV1-27 X X X 12.7 Immune response
Immunoglobulin kappa variable
1-33 KV133_HUMAN P01594 IGKV1-33 X X X 12.8 Immune response
Immunoglobulin kappa variable
1D-16 KVD16_HUMAN P01601 IGKV1D-16 X 12.7 Immune response
Immunoglobulin kappa variable
1D-39 KVD39_HUMAN P04432 IGKV1D-39 X X X 12.7 Immune response
Immunoglobulin kappa variable
2-24 KV224_HUMAN A0A0C4DH68 IGKV2-24 X X 13.1 Immune response
Immunoglobulin kappa variable
2-29 KV229_HUMAN A2NJV5 IGKV2-29 X X 13.1 Immune response
Immunoglobulin kappa variable
2-30 KV230_HUMAN P06310 IGKV2-30 X X X 13.2 Immune response
Immunoglobulin kappa variable
2D-28 KVD28_HUMAN P01615 IGKV2D-28 X X X 12.9 Immune response
Immunoglobulin kappa variable
2D-29 KVD29_HUMAN A0A075B6S2 IGKV2D-29 X 13.1 Immune response
Immunoglobulin kappa variable
3D-15 KVD15_HUMAN A0A087WSY6 IGKV3D-15 X 12.5 Immune response
Immunoglobulin kappa variable
3-11 KV311_HUMAN P04433 IGKV3-11 X X X 12.6 Immune response
Immunoglobulin kappa variable
3-15 KV315_HUMAN P01624 IGKV3-15 X X X 12.5 Immune response
Immunoglobulin kappa variable
3-20 KV320_HUMAN P01619 IGKV3-20 X X X 12.6 Immune response
Immunoglobulin kappa variable
3D-20 KVD20_HUMAN A0A0C4DH25 IGKV3D-20 X X X 12.5 Immune response
Immunoglobulin kappa variable
4-1 KV401_HUMAN P06312 IGKV4-1 X X X 13.4 Immune response
Immunoglobulin kappa variable
6-21 KV621_HUMAN A0A0C4DH24 IGKV6-21 X 12.4 Immune response
Immunoglobulin kappa variable
6D-21 KVD21_HUMAN A0A0A0MT36 IGKV6D-21 X X 12.3 Immune response
Immunoglobulin lambda constant
2 IGLC2_HUMAN P0DOY2 IGLC2 X X X 11.3 Immune response
Immunoglobulin lambda constant
3 IGLC3_HUMAN P0DOY3 IGLC3 X X X 11.3 Immune response
Immunoglobulin lambda constant
7 IGLC7_HUMAN A0M8Q6 IGLC7 X 11.2 Immune response

252
Immunoglobulin lambda-like
polypeptide 1 IGLL1_HUMAN P15814 IGLL1 X X 22.9 Immune response
Immunoglobulin lambda variable
1-36 LV136_HUMAN A0A0B4J1U3 IGLV1-36 X 12.5 Immune response
Immunoglobulin lambda variable
1-40 LV140_HUMAN P01703 IGLV1-40 X 12.3 Immune response
Immunoglobulin lambda variable
1-47 LV147_HUMAN P01700 IGLV1-47 X X X 12.3 Immune response
Immunoglobulin lambda variable
1-51 LV151_HUMAN P01701 IGLV1-51 X X 12.2 Immune response
Immunoglobulin lambda variable
2-8 LV208_HUMAN P01709 IGLV2-8 X 12.4 Immune response
Immunoglobulin lambda variable
2-11 LV211_HUMAN P01706 IGLV2-11 X 12.6 Immune response
Immunoglobulin lambda variable
2-14 LV214_HUMAN P01704 IGLV2-14 X 12.6 Immune response
Immunoglobulin lambda variable
2-18 LV218_HUMAN A0A075B6J9 IGLV2-18 X 12.4 Immune response
Immunoglobulin lambda variable
2-23 LV223_HUMAN P01705 IGLV2-23 X 11.9 Immune response
Immunoglobulin lambda variable
3-9 LV39_HUMAN A0A075B6K5 IGLV3-9 X 12.3 Immune response
Immunoglobulin lambda variable
3-19 LV319_HUMAN P01714 IGLV3-19 X X 12.0 Immune response
Immunoglobulin lambda variable
3-21 LV321_HUMAN P80748 IGLV3-21 X X X 12.4 Immune response
Immunoglobulin lambda variable
3-25 LV325_HUMAN P01717 IGLV3-25 X X 12.0 Immune response
Immunoglobulin lambda variable
4-60 LV460_HUMAN A0A075B6I1 IGLV4-60 X 12.9 Immune response
Immunoglobulin lambda variable
4-69 LV469_HUMAN A0A075B6H9 IGLV4-69 X X 12.8 Immune response
Immunoglobulin lambda variable
5-45 LV545_HUMAN A0A087WSX0 IGLV5-45 X 13.2 Immune response
Immunoglobulin lambda variable
6-57 LV657_HUMAN P01721 IGLV6-57 X 12.6 Immune response
Immunoglobulin lambda variable
7-43 LV743_HUMAN P04211 IGLV7-43 X X 12.4 Immune response
Immunoglobulin lambda variable
7-46 LV746_HUMAN A0A075B6I9 IGLV7-46 X X 12.5 Immune response

253
Immunoglobulin lambda variable
8-61 LV861_HUMAN A0A075B6I0 IGLV8-61 X X 12.8 Immune response
Immunoglobulin lambda variable
9-49 LV949_HUMAN A0A0B4J1Y8 IGLV9-49 X 13.0 Immune response
Immunoglobulin lambda variable
10-54 LVX54_HUMAN A0A075B6I4 IGLV10-54 X 12.4 Immune response
Immunoglobulin lambda-like
polypeptide 5 IGLL5_HUMAN B9A064 IGLL5 X X X 23.1 Immune response
Interleukin-22 IL22_HUMAN Q9GZX6 IL22 X 20.0 Cytokine that contributes to the
inflammatory response in vivo.
Importin-4 IPO4_HUMAN Q8TEX9 IPO4 X 118.7 Functions in nuclear protein import as
nuclear transport receptor
Inositol polyphosphate 1-
phosphatase INPP_HUMAN P49441 INPP1 X 43.9 Catalytic activity
Insulin-like growth factor-
binding protein 3 IBP3_HUMAN P17936 IGFBP3 X 31.7
IGF-binding proteins prolong the
half-life of the IGFs
Insulin-like growth factor-
binding protein complex acid
labile subunit
ALS_HUMAN P35858 IGFALS X X X 66.1
Binds insulin-like growth factors.
increasing their half-life and their
vascular localization
Inter-alpha-trypsin inhibitor
heavy chain H1 ITIH1_HUMAN P19827 ITIH1 X X X 101.4 Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H2 ITIH2_HUMAN P19823 ITIH2 X X X 106.5 Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H3 ITIH3_HUMAN Q06033 ITIH3 X X X 99.9 Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H4 ITIH4_HUMAN Q14624 ITIH4 X X X 103.4 Inflammatory response
Kallistatin KAIN_HUMAN P29622 SERPINA4 X X 48.5 Enzyme regulator inhibitor
Kelch-like protein 38 KLH38_HUMAN Q2WGJ6 KLHL38 X 65.5 -
Keratin. type I cytoskeletal 9 K1C9_HUMAN P35527 KRT9 X X 62.1 Structural
Keratin. type I cytoskeletal 10 K1C10_HUMAN P13645 KRT10 X X X 58.8 Structural
Keratin. type II cytoskeletal 1 K2C1_HUMAN P04264 KRT1 X X X 66.1 Enzyme regulator activity
Keratin. type II cytoskeletal 2
epidermal K22E_HUMAN P35908 KRT2 X X X 65.4 Structural
Keratin. type II cuticular Hb1 KRT81_HUMAN Q14533 KRT81 X 54.9 Structural
Kinesin heavy chain isoform 5A KIF5A_HUMAN Q12840 KIF5A X 117.4
Microtubule-dependent motor
required for slow axonal transport of
neurofilament proteins (NFH. NFM
and NFL)
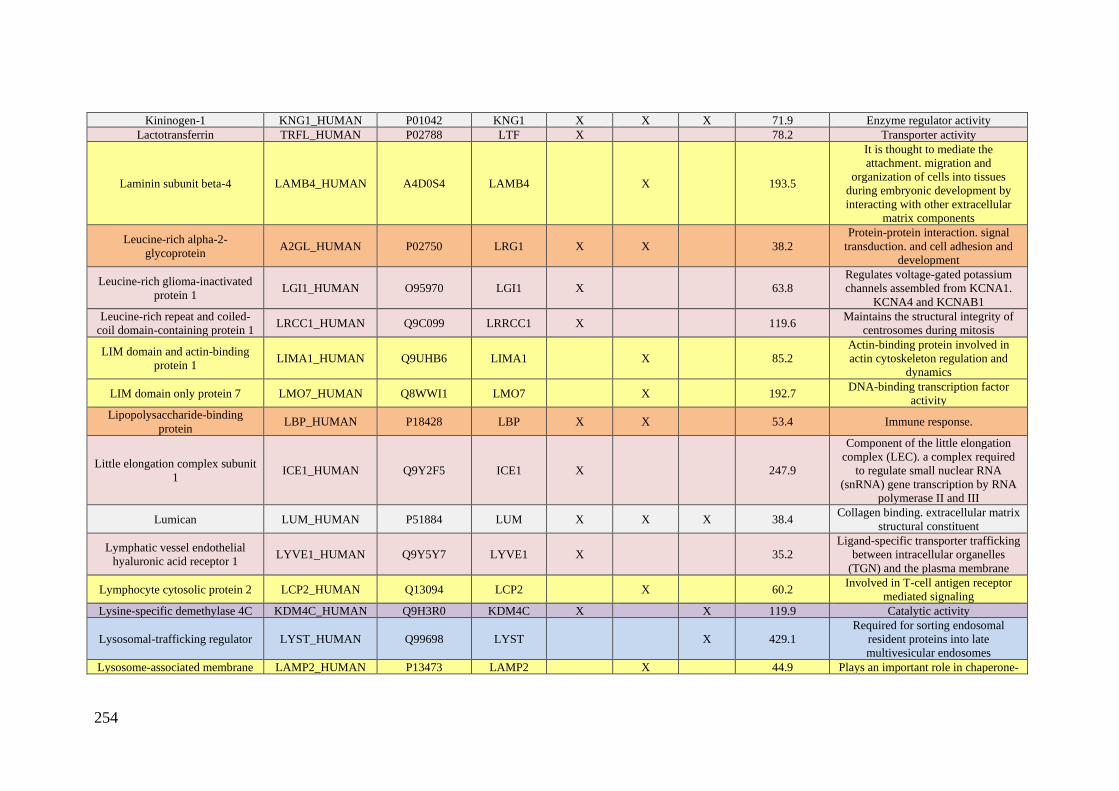
254
Kininogen-1 KNG1_HUMAN P01042 KNG1 X X X 71.9 Enzyme regulator activity
Lactotransferrin TRFL_HUMAN P02788 LTF X 78.2 Transporter activity
Laminin subunit beta-4 LAMB4_HUMAN A4D0S4 LAMB4 X 193.5
It is thought to mediate the
attachment. migration and
organization of cells into tissues
during embryonic development by
interacting with other extracellular
matrix components
Leucine-rich alpha-2-
glycoprotein A2GL_HUMAN P02750 LRG1 X X 38.2
Protein-protein interaction. signal
transduction. and cell adhesion and
development
Leucine-rich glioma-inactivated
protein 1 LGI1_HUMAN O95970 LGI1 X 63.8
Regulates voltage-gated potassium
channels assembled from KCNA1.
KCNA4 and KCNAB1
Leucine-rich repeat and coiled-
coil domain-containing protein 1 LRCC1_HUMAN Q9C099 LRRCC1 X 119.6
Maintains the structural integrity of
centrosomes during mitosis
LIM domain and actin-binding
protein 1 LIMA1_HUMAN Q9UHB6 LIMA1 X 85.2
Actin-binding protein involved in
actin cytoskeleton regulation and
dynamics
LIM domain only protein 7 LMO7_HUMAN Q8WWI1 LMO7 X 192.7 DNA-binding transcription factor
activity
Lipopolysaccharide-binding
protein LBP_HUMAN P18428 LBP X X 53.4 Immune response.
Little elongation complex subunit
1 ICE1_HUMAN Q9Y2F5 ICE1 X 247.9
Component of the little elongation
complex (LEC). a complex required
to regulate small nuclear RNA
(snRNA) gene transcription by RNA
polymerase II and III
Lumican LUM_HUMAN P51884 LUM X X X 38.4 Collagen binding. extracellular matrix
structural constituent
Lymphatic vessel endothelial
hyaluronic acid receptor 1 LYVE1_HUMAN Q9Y5Y7 LYVE1 X 35.2
Ligand-specific transporter trafficking
between intracellular organelles
(TGN) and the plasma membrane
Lymphocyte cytosolic protein 2 LCP2_HUMAN Q13094 LCP2 X 60.2 Involved in T-cell antigen receptor
mediated signaling
Lysine-specific demethylase 4C KDM4C_HUMAN Q9H3R0 KDM4C X X 119.9 Catalytic activity
Lysosomal-trafficking regulator LYST_HUMAN Q99698 LYST X 429.1
Required for sorting endosomal
resident proteins into late
multivesicular endosomes
Lysosome-associated membrane LAMP2_HUMAN P13473 LAMP2 X 44.9 Plays an important role in chaperone-
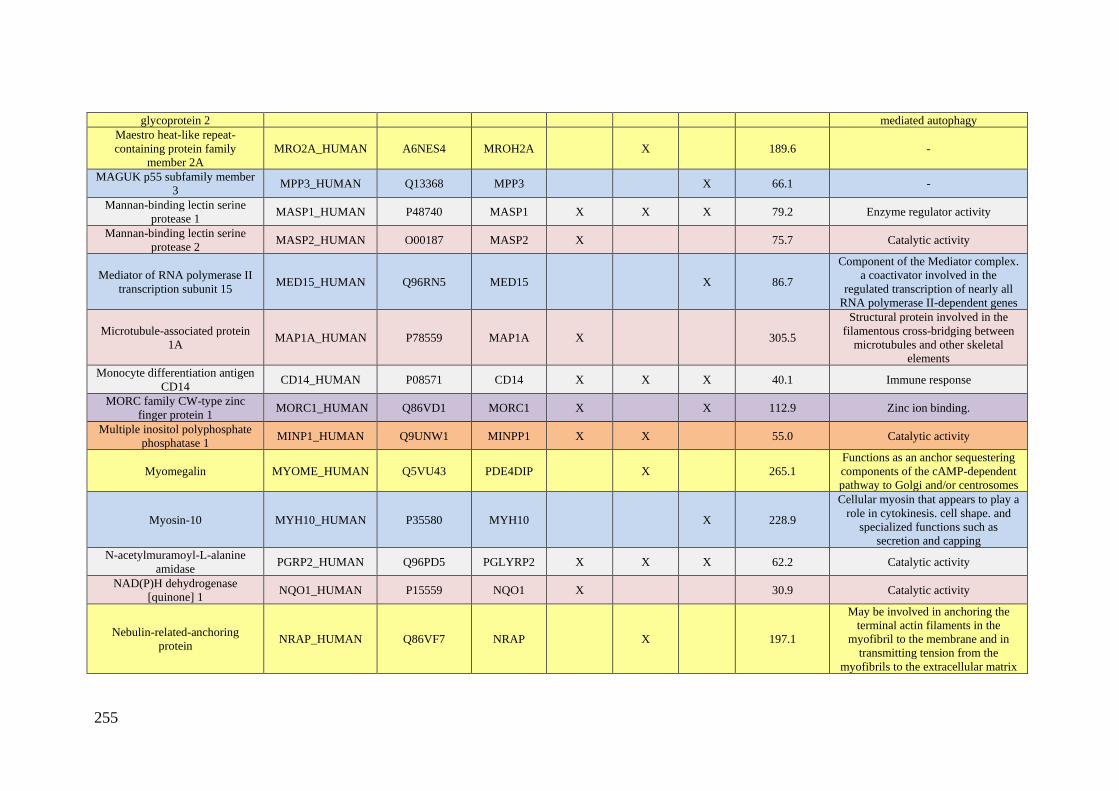
255
glycoprotein 2 mediated autophagy
Maestro heat-like repeat-
containing protein family
member 2A
MRO2A_HUMAN A6NES4 MROH2A X 189.6 -
MAGUK p55 subfamily member
3 MPP3_HUMAN Q13368 MPP3 X 66.1 -
Mannan-binding lectin serine
protease 1 MASP1_HUMAN P48740 MASP1 X X X 79.2 Enzyme regulator activity
Mannan-binding lectin serine
protease 2 MASP2_HUMAN O00187 MASP2 X 75.7 Catalytic activity
Mediator of RNA polymerase II
transcription subunit 15 MED15_HUMAN Q96RN5 MED15 X 86.7
Component of the Mediator complex.
a coactivator involved in the
regulated transcription of nearly all
RNA polymerase II-dependent genes
Microtubule-associated protein
1A MAP1A_HUMAN P78559 MAP1A X 305.5
Structural protein involved in the
filamentous cross-bridging between
microtubules and other skeletal
elements
Monocyte differentiation antigen
CD14 CD14_HUMAN P08571 CD14 X X X 40.1 Immune response
MORC family CW-type zinc
finger protein 1 MORC1_HUMAN Q86VD1 MORC1 X X 112.9 Zinc ion binding.
Multiple inositol polyphosphate
phosphatase 1 MINP1_HUMAN Q9UNW1 MINPP1 X X 55.0 Catalytic activity
Myomegalin MYOME_HUMAN Q5VU43 PDE4DIP X 265.1
Functions as an anchor sequestering
components of the cAMP-dependent
pathway to Golgi and/or centrosomes
Myosin-10 MYH10_HUMAN P35580 MYH10 X 228.9
Cellular myosin that appears to play a
role in cytokinesis. cell shape. and
specialized functions such as
secretion and capping
N-acetylmuramoyl-L-alanine
amidase PGRP2_HUMAN Q96PD5 PGLYRP2 X X X 62.2 Catalytic activity
NAD(P)H dehydrogenase
[quinone] 1 NQO1_HUMAN P15559 NQO1 X 30.9 Catalytic activity
Nebulin-related-anchoring
protein NRAP_HUMAN Q86VF7 NRAP X 197.1
May be involved in anchoring the
terminal actin filaments in the
myofibril to the membrane and in
transmitting tension from the
myofibrils to the extracellular matrix
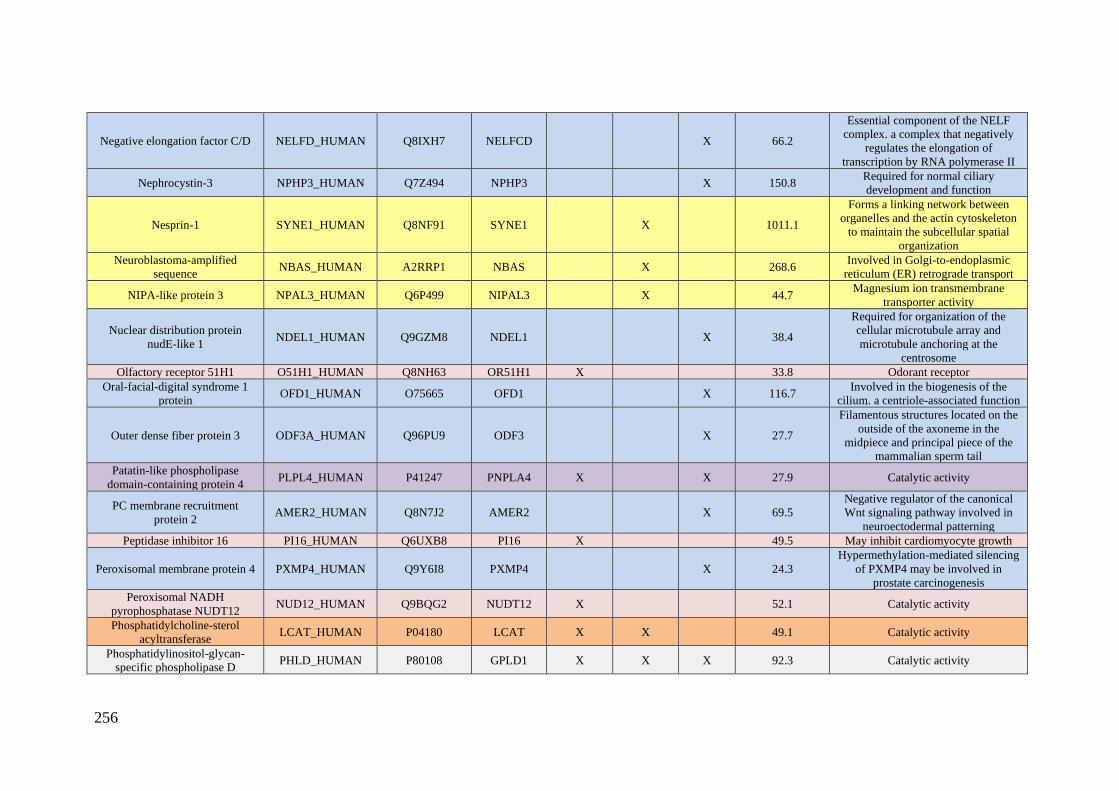
256
Negative elongation factor C/D NELFD_HUMAN Q8IXH7 NELFCD X 66.2
Essential component of the NELF
complex. a complex that negatively
regulates the elongation of
transcription by RNA polymerase II
Nephrocystin-3 NPHP3_HUMAN Q7Z494 NPHP3 X 150.8 Required for normal ciliary
development and function
Nesprin-1 SYNE1_HUMAN Q8NF91 SYNE1 X 1011.1
Forms a linking network between
organelles and the actin cytoskeleton
to maintain the subcellular spatial
organization
Neuroblastoma-amplified
sequence NBAS_HUMAN A2RRP1 NBAS X 268.6
Involved in Golgi-to-endoplasmic
reticulum (ER) retrograde transport
NIPA-like protein 3 NPAL3_HUMAN Q6P499 NIPAL3 X 44.7 Magnesium ion transmembrane
transporter activity
Nuclear distribution protein
nudE-like 1 NDEL1_HUMAN Q9GZM8 NDEL1 X 38.4
Required for organization of the
cellular microtubule array and
microtubule anchoring at the
centrosome
Olfactory receptor 51H1 O51H1_HUMAN Q8NH63 OR51H1 X 33.8 Odorant receptor
Oral-facial-digital syndrome 1
protein OFD1_HUMAN O75665 OFD1 X 116.7
Involved in the biogenesis of the
cilium. a centriole-associated function
Outer dense fiber protein 3 ODF3A_HUMAN Q96PU9 ODF3 X 27.7
Filamentous structures located on the
outside of the axoneme in the
midpiece and principal piece of the
mammalian sperm tail
Patatin-like phospholipase
domain-containing protein 4 PLPL4_HUMAN P41247 PNPLA4 X X 27.9 Catalytic activity
PC membrane recruitment
protein 2 AMER2_HUMAN Q8N7J2 AMER2 X 69.5
Negative regulator of the canonical
Wnt signaling pathway involved in
neuroectodermal patterning
Peptidase inhibitor 16 PI16_HUMAN Q6UXB8 PI16 X 49.5 May inhibit cardiomyocyte growth
Peroxisomal membrane protein 4 PXMP4_HUMAN Q9Y6I8 PXMP4 X 24.3
Hypermethylation-mediated silencing
of PXMP4 may be involved in
prostate carcinogenesis
Peroxisomal NADH
pyrophosphatase NUDT12 NUD12_HUMAN Q9BQG2 NUDT12 X 52.1 Catalytic activity
Phosphatidylcholine-sterol
acyltransferase LCAT_HUMAN P04180 LCAT X X 49.1 Catalytic activity
Phosphatidylinositol-glycan-
specific phospholipase D PHLD_HUMAN P80108 GPLD1 X X X 92.3 Catalytic activity
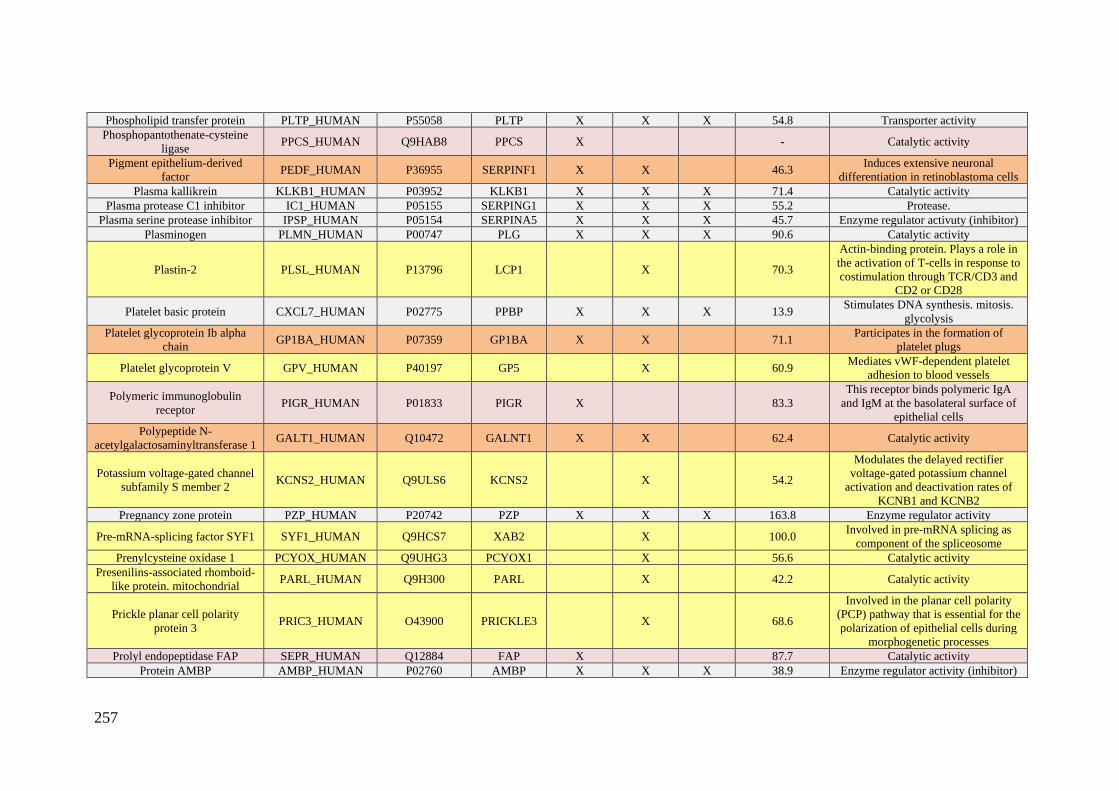
257
Phospholipid transfer protein PLTP_HUMAN P55058 PLTP X X X 54.8 Transporter activity
Phosphopantothenate-cysteine
ligase PPCS_HUMAN Q9HAB8 PPCS X - Catalytic activity
Pigment epithelium-derived
factor PEDF_HUMAN P36955 SERPINF1 X X 46.3
Induces extensive neuronal
differentiation in retinoblastoma cells
Plasma kallikrein KLKB1_HUMAN P03952 KLKB1 X X X 71.4 Catalytic activity
Plasma protease C1 inhibitor IC1_HUMAN P05155 SERPING1 X X X 55.2 Protease.
Plasma serine protease inhibitor IPSP_HUMAN P05154 SERPINA5 X X X 45.7 Enzyme regulator activuty (inhibitor)
Plasminogen PLMN_HUMAN P00747 PLG X X X 90.6 Catalytic activity
Plastin-2 PLSL_HUMAN P13796 LCP1 X 70.3
Actin-binding protein. Plays a role in
the activation of T-cells in response to
costimulation through TCR/CD3 and
CD2 or CD28
Platelet basic protein CXCL7_HUMAN P02775 PPBP X X X 13.9 Stimulates DNA synthesis. mitosis.
glycolysis
Platelet glycoprotein Ib alpha
chain GP1BA_HUMAN P07359 GP1BA X X 71.1
Participates in the formation of
platelet plugs
Platelet glycoprotein V GPV_HUMAN P40197 GP5 X 60.9 Mediates vWF-dependent platelet
adhesion to blood vessels
Polymeric immunoglobulin
receptor PIGR_HUMAN P01833 PIGR X 83.3
This receptor binds polymeric IgA
and IgM at the basolateral surface of
epithelial cells
Polypeptide N-
acetylgalactosaminyltransferase 1 GALT1_HUMAN Q10472 GALNT1 X X 62.4 Catalytic activity
Potassium voltage-gated channel
subfamily S member 2 KCNS2_HUMAN Q9ULS6 KCNS2 X 54.2
Modulates the delayed rectifier
voltage-gated potassium channel
activation and deactivation rates of
KCNB1 and KCNB2
Pregnancy zone protein PZP_HUMAN P20742 PZP X X X 163.8 Enzyme regulator activity
Pre-mRNA-splicing factor SYF1 SYF1_HUMAN Q9HCS7 XAB2 X 100.0 Involved in pre-mRNA splicing as
component of the spliceosome
Prenylcysteine oxidase 1 PCYOX_HUMAN Q9UHG3 PCYOX1 X 56.6 Catalytic activity
Presenilins-associated rhomboid-
like protein. mitochondrial PARL_HUMAN Q9H300 PARL X 42.2 Catalytic activity
Prickle planar cell polarity
protein 3 PRIC3_HUMAN O43900 PRICKLE3 X 68.6
Involved in the planar cell polarity
(PCP) pathway that is essential for the
polarization of epithelial cells during
morphogenetic processes
Prolyl endopeptidase FAP SEPR_HUMAN Q12884 FAP X 87.7 Catalytic activity
Protein AMBP AMBP_HUMAN P02760 AMBP X X X 38.9 Enzyme regulator activity (inhibitor)
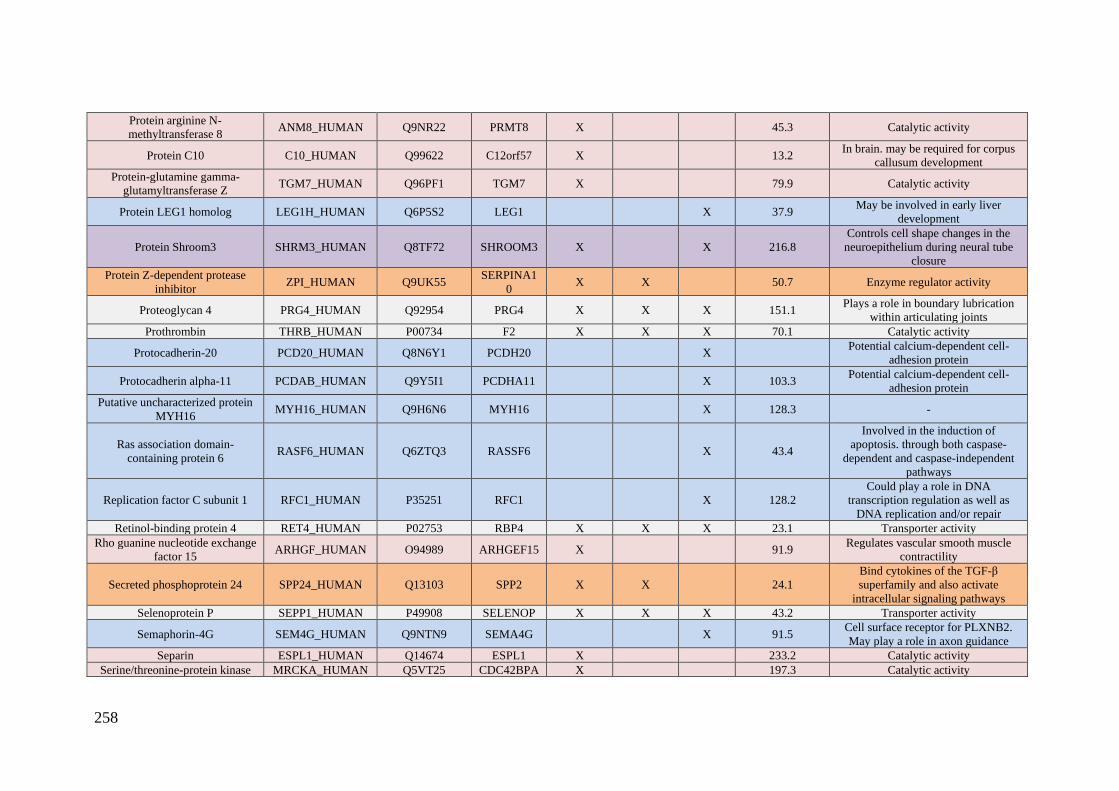
258
Protein arginine N-
methyltransferase 8 ANM8_HUMAN Q9NR22 PRMT8 X 45.3 Catalytic activity
Protein C10 C10_HUMAN Q99622 C12orf57 X 13.2 In brain. may be required for corpus
callusum development
Protein-glutamine gamma-
glutamyltransferase Z TGM7_HUMAN Q96PF1 TGM7 X 79.9 Catalytic activity
Protein LEG1 homolog LEG1H_HUMAN Q6P5S2 LEG1 X 37.9 May be involved in early liver
development
Protein Shroom3 SHRM3_HUMAN Q8TF72 SHROOM3 X X 216.8
Controls cell shape changes in the
neuroepithelium during neural tube
closure
Protein Z-dependent protease
inhibitor ZPI_HUMAN Q9UK55
SERPINA1
0 X X 50.7 Enzyme regulator activity
Proteoglycan 4 PRG4_HUMAN Q92954 PRG4 X X X 151.1 Plays a role in boundary lubrication
within articulating joints
Prothrombin THRB_HUMAN P00734 F2 X X X 70.1 Catalytic activity
Protocadherin-20 PCD20_HUMAN Q8N6Y1 PCDH20 X Potential calcium-dependent cell-
adhesion protein
Protocadherin alpha-11 PCDAB_HUMAN Q9Y5I1 PCDHA11 X 103.3 Potential calcium-dependent cell-
adhesion protein
Putative uncharacterized protein
MYH16 MYH16_HUMAN Q9H6N6 MYH16 X 128.3 -
Ras association domain-
containing protein 6 RASF6_HUMAN Q6ZTQ3 RASSF6 X 43.4
Involved in the induction of
apoptosis. through both caspase-
dependent and caspase-independent
pathways
Replication factor C subunit 1 RFC1_HUMAN P35251 RFC1 X 128.2
Could play a role in DNA
transcription regulation as well as
DNA replication and/or repair
Retinol-binding protein 4 RET4_HUMAN P02753 RBP4 X X X 23.1 Transporter activity
Rho guanine nucleotide exchange
factor 15 ARHGF_HUMAN O94989 ARHGEF15 X 91.9
Regulates vascular smooth muscle
contractility
Secreted phosphoprotein 24 SPP24_HUMAN Q13103 SPP2 X X 24.1
Bind cytokines of the TGF-β
superfamily and also activate
intracellular signaling pathways
Selenoprotein P SEPP1_HUMAN P49908 SELENOP X X X 43.2 Transporter activity
Semaphorin-4G SEM4G_HUMAN Q9NTN9 SEMA4G X 91.5 Cell surface receptor for PLXNB2.
May play a role in axon guidance
Separin ESPL1_HUMAN Q14674 ESPL1 X 233.2 Catalytic activity
Serine/threonine-protein kinase MRCKA_HUMAN Q5VT25 CDC42BPA X 197.3 Catalytic activity
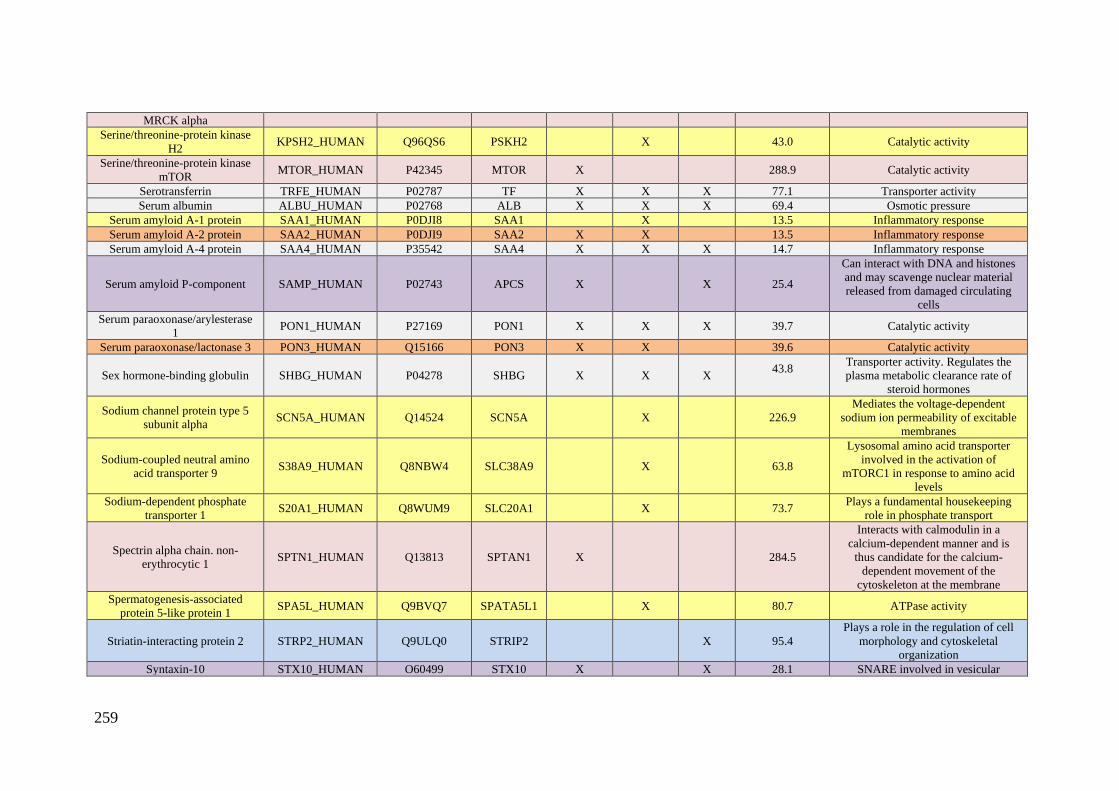
259
MRCK alpha
Serine/threonine-protein kinase
H2 KPSH2_HUMAN Q96QS6 PSKH2 X 43.0 Catalytic activity
Serine/threonine-protein kinase
mTOR MTOR_HUMAN P42345 MTOR X 288.9 Catalytic activity
Serotransferrin TRFE_HUMAN P02787 TF X X X 77.1 Transporter activity
Serum albumin ALBU_HUMAN P02768 ALB X X X 69.4 Osmotic pressure
Serum amyloid A-1 protein SAA1_HUMAN P0DJI8 SAA1 X 13.5 Inflammatory response
Serum amyloid A-2 protein SAA2_HUMAN P0DJI9 SAA2 X X 13.5 Inflammatory response
Serum amyloid A-4 protein SAA4_HUMAN P35542 SAA4 X X X 14.7 Inflammatory response
Serum amyloid P-component SAMP_HUMAN P02743 APCS X X 25.4
Can interact with DNA and histones
and may scavenge nuclear material
released from damaged circulating
cells
Serum paraoxonase/arylesterase
1 PON1_HUMAN P27169 PON1 X X X 39.7 Catalytic activity
Serum paraoxonase/lactonase 3 PON3_HUMAN Q15166 PON3 X X 39.6 Catalytic activity
Sex hormone-binding globulin SHBG_HUMAN P04278 SHBG X X X 43.8
Transporter activity. Regulates the
plasma metabolic clearance rate of
steroid hormones
Sodium channel protein type 5
subunit alpha SCN5A_HUMAN Q14524 SCN5A X 226.9
Mediates the voltage-dependent
sodium ion permeability of excitable
membranes
Sodium-coupled neutral amino
acid transporter 9 S38A9_HUMAN Q8NBW4 SLC38A9 X 63.8
Lysosomal amino acid transporter
involved in the activation of
mTORC1 in response to amino acid
levels
Sodium-dependent phosphate
transporter 1 S20A1_HUMAN Q8WUM9 SLC20A1 X 73.7
Plays a fundamental housekeeping
role in phosphate transport
Spectrin alpha chain. non-
erythrocytic 1 SPTN1_HUMAN Q13813 SPTAN1 X 284.5
Interacts with calmodulin in a
calcium-dependent manner and is
thus candidate for the calcium-
dependent movement of the
cytoskeleton at the membrane
Spermatogenesis-associated
protein 5-like protein 1 SPA5L_HUMAN Q9BVQ7 SPATA5L1 X 80.7 ATPase activity
Striatin-interacting protein 2 STRP2_HUMAN Q9ULQ0 STRIP2 X 95.4
Plays a role in the regulation of cell
morphology and cytoskeletal
organization
Syntaxin-10 STX10_HUMAN O60499 STX10 X X 28.1 SNARE involved in vesicular

260
transport from the late endosomes to
the trans-Golgi network
Taste receptor type 2 member 39 T2R39_HUMAN P59534 TAS2R39 X X 38.6
Receptor that may play a role in the
perception of bitterness and is
gustducin-linked
T-complex-associated testis-
expressed protein 1 TCTE1_HUMAN - - X - -
Telomere length regulation
protein TEL2 homolog TELO2_HUMAN Q9Y4R8 TELO2 X 91.7
Regulator of the DNA damage
response (DDR)
Teneurin-3 TEN3_HUMAN Q9P273 TENM3 X 300.9
Involved in neural development by
regulating the establishment of proper
connectivity within the nervous
system
Tetranectin TETN_HUMAN P05452 CLEC3B X X X 22.5 Involved in the packaging of
molecules destined for exocytosis
Thrombospondin-1 TSP1_HUMAN P07996 THBS1 X X X 129.4 Immune response
Thyroxine-binding globulin THBG_HUMAN P05543 SERPINA7 X X 46.3 Transporter activity
TOM1-like protein 1 TM1L1_HUMAN O75674 TOM1L1 X 52.9 Adapter protein involved in signaling
pathways
Transcobalamin-1 TCO1_HUMAN P20061 TCN1 X 48.2
Binds vitamin B12 with femtomolar
affinity and protects it from the acidic
environment of the stomach
Transcription factor AP-2-epsilon AP2E_HUMAN Q6VUC0 TFAP2E X 46.2
Sequence-specific DNA-binding
protein that interacts with inducible
viral and cellular enhancer elements
to regulate transcription of selected
genes
Transcription factor HES-1 HES1_HUMAN Q14469 HES1 X 29.5
Transcriptional repressor of genes
that require a bHLH protein for their
transcription.
Transferrin receptor protein 1 TFR1_HUMAN P02786 TFRC X X 84.9 Transporter activity
Transient receptor potential
cation channel subfamily V
member 5
TRPV5_HUMAN Q9NQA5 TRPV5 X 82.5
Constitutively active calcium
selective cation channel thought to be
involved in Ca2+ reabsorption in
kidney and intestine
Transmembrane protein 198 TM198_HUMAN Q66K66 TMEM198 X X X 39.4
Promotes low-density lipoprotein
receptor-related protein 6 (LRP6)
phosphorylation
Transthyretin TTHY_HUMAN P02766 TTR X X 15.8 Transporter activity
Tyrosine--tRNA ligase. SYYC_HUMAN P54577 YARS X 59.1 Catalytic activity
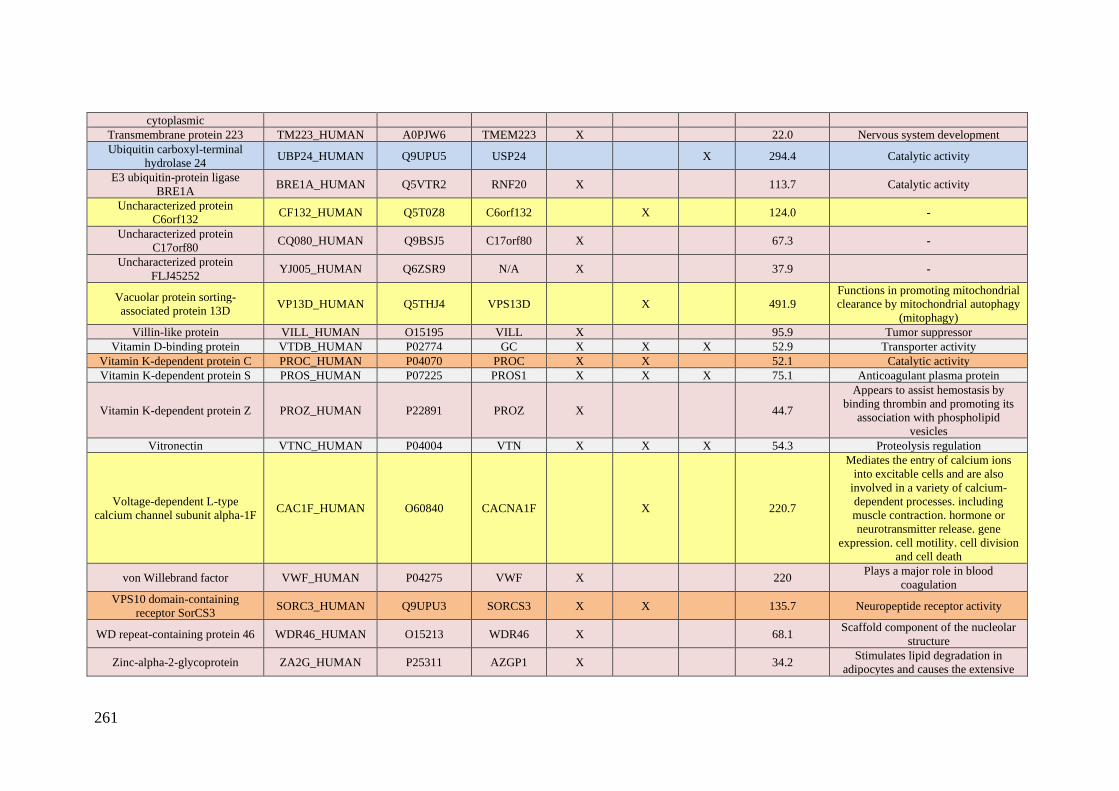
261
cytoplasmic
Transmembrane protein 223 TM223_HUMAN A0PJW6 TMEM223 X 22.0 Nervous system development
Ubiquitin carboxyl-terminal
hydrolase 24 UBP24_HUMAN Q9UPU5 USP24 X 294.4 Catalytic activity
E3 ubiquitin-protein ligase
BRE1A BRE1A_HUMAN Q5VTR2 RNF20 X 113.7 Catalytic activity
Uncharacterized protein
C6orf132 CF132_HUMAN Q5T0Z8 C6orf132 X 124.0 -
Uncharacterized protein
C17orf80 CQ080_HUMAN Q9BSJ5 C17orf80 X 67.3 -
Uncharacterized protein
FLJ45252 YJ005_HUMAN Q6ZSR9 N/A X 37.9 -
Vacuolar protein sorting-
associated protein 13D VP13D_HUMAN Q5THJ4 VPS13D X 491.9
Functions in promoting mitochondrial
clearance by mitochondrial autophagy
(mitophagy)
Villin-like protein VILL_HUMAN O15195 VILL X 95.9 Tumor suppressor
Vitamin D-binding protein VTDB_HUMAN P02774 GC X X X 52.9 Transporter activity
Vitamin K-dependent protein C PROC_HUMAN P04070 PROC X X 52.1 Catalytic activity
Vitamin K-dependent protein S PROS_HUMAN P07225 PROS1 X X X 75.1 Anticoagulant plasma protein
Vitamin K-dependent protein Z PROZ_HUMAN P22891 PROZ X 44.7
Appears to assist hemostasis by
binding thrombin and promoting its
association with phospholipid
vesicles
Vitronectin VTNC_HUMAN P04004 VTN X X X 54.3 Proteolysis regulation
Voltage-dependent L-type
calcium channel subunit alpha-1F CAC1F_HUMAN O60840 CACNA1F X 220.7
Mediates the entry of calcium ions
into excitable cells and are also
involved in a variety of calcium-
dependent processes. including
muscle contraction. hormone or
neurotransmitter release. gene
expression. cell motility. cell division
and cell death
von Willebrand factor VWF_HUMAN P04275 VWF X 220 Plays a major role in blood
coagulation
VPS10 domain-containing
receptor SorCS3 SORC3_HUMAN Q9UPU3 SORCS3 X X 135.7 Neuropeptide receptor activity
WD repeat-containing protein 46 WDR46_HUMAN O15213 WDR46 X 68.1 Scaffold component of the nucleolar
structure
Zinc-alpha-2-glycoprotein ZA2G_HUMAN P25311 AZGP1 X 34.2 Stimulates lipid degradation in
adipocytes and causes the extensive

262
fat losses associated with some
advanced cancers
Zinc finger and BTB domain-
containing protein 4 ZBTB4_HUMAN Q9P1Z0 ZBTB4 X 105.1
Transcriptional repressor with
bimodal DNA-binding specificity
Zinc finger CCCH domain-
containing protein 4 ZC3H4_HUMAN Q9UPT8 ZC3H4 X 140.2
DNA-binding transcription factor
activity
Zinc finger protein 350 ZN350_HUMAN Q9GZX5 ZNF350 X 60.0 Transcriptional repressor
Zinc finger protein 570 ZN570_HUMAN Q96NI8 ZNF570 X 62.3 May be involved in transcriptional
regulation
Zinc finger protein 700 ZN700_HUMAN Q9H0M5 ZNF700 X 86.2 May be involved in transcriptional
regulation

263
Table 2_SM. Analysis of the protein corona formed around AuNPs (10.02 ± 0.91 nm). AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm)
after their incubation with serum aliquots (x2) from eight different triple negative breast cancer patients (6 protein samples per individual: 2
treated with AuNPs. 2 with AgNPs and 2 with MNPs). The accession number. gene name. species (Human). molecular weight and protein
function were reported. Grey color: 138 proteins commonly found in the protein corona of AuNPs (10.02 ± 0.91 nm). AgNPs (9.73 ± 1.70 nm)
and MNPs (9.30 ± 0.67 nm). Pink color: 56 identified corona proteins exclusively bound to the AuNPs (10.02 ± 0.91 nm). Yellow color: 33
identified corona proteins exclusively bound to the AgNPs (9.73 ± 1.70 nm). Blue: 53 identified corona proteins exclusively bound to the MNPs
(9.30 ± 0.67 nm). Orange color: 30 identified corona proteins commonly bound to the AuNPs (10.02 ± 0.91 nm) and AgNPs (9.73 ± 1.70 nm).
Violet color: 7 identified corona proteins commonly bound to the AuNPs (10.02 ± 0.91 nm) and MNPs (9.30 ± 0.67 nm). Green color: 5
identified corona proteins commonly bound to the AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm).
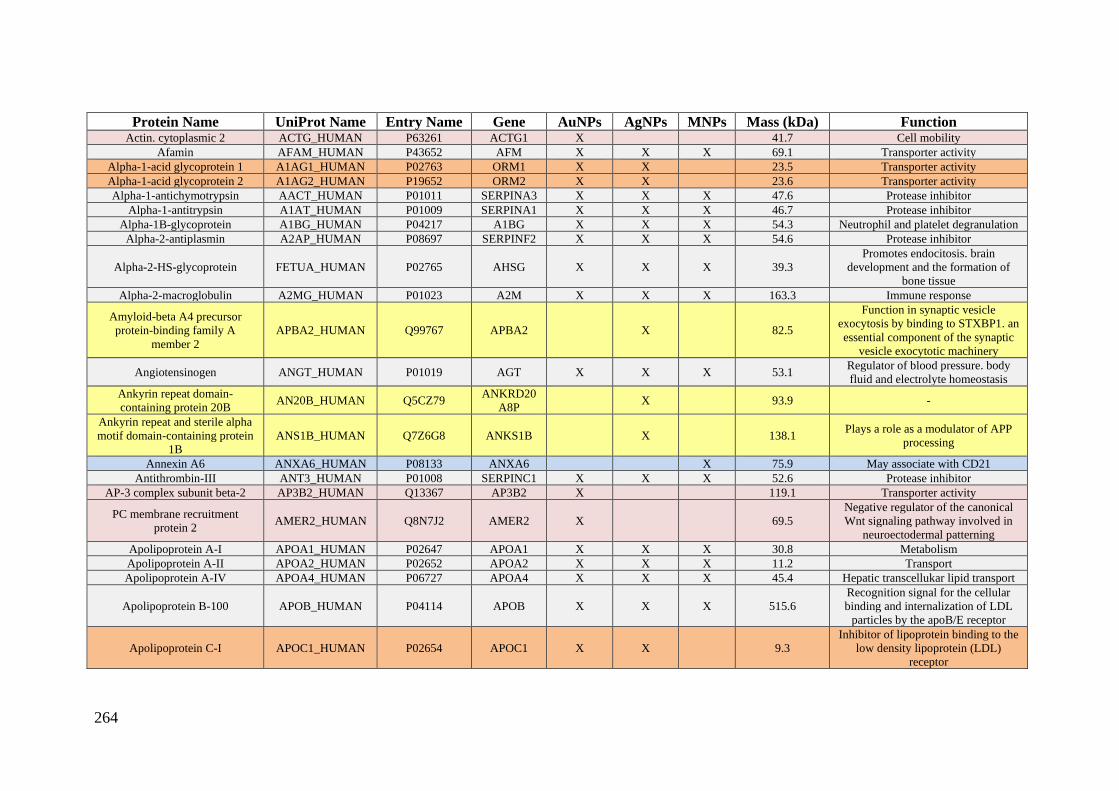
264
Protein Name UniProt Name Entry Name Gene AuNPs AgNPs MNPs Mass (kDa) Function Actin. cytoplasmic 2 ACTG_HUMAN P63261 ACTG1 X 41.7 Cell mobility
Afamin AFAM_HUMAN P43652 AFM X X X 69.1 Transporter activity
Alpha-1-acid glycoprotein 1 A1AG1_HUMAN P02763 ORM1 X X 23.5 Transporter activity
Alpha-1-acid glycoprotein 2 A1AG2_HUMAN P19652 ORM2 X X 23.6 Transporter activity
Alpha-1-antichymotrypsin AACT_HUMAN P01011 SERPINA3 X X X 47.6 Protease inhibitor
Alpha-1-antitrypsin A1AT_HUMAN P01009 SERPINA1 X X X 46.7 Protease inhibitor
Alpha-1B-glycoprotein A1BG_HUMAN P04217 A1BG X X X 54.3 Neutrophil and platelet degranulation
Alpha-2-antiplasmin A2AP_HUMAN P08697 SERPINF2 X X X 54.6 Protease inhibitor
Alpha-2-HS-glycoprotein FETUA_HUMAN P02765 AHSG X X X 39.3
Promotes endocitosis. brain
development and the formation of
bone tissue
Alpha-2-macroglobulin A2MG_HUMAN P01023 A2M X X X 163.3 Immune response
Amyloid-beta A4 precursor
protein-binding family A
member 2
APBA2_HUMAN Q99767 APBA2 X 82.5
Function in synaptic vesicle
exocytosis by binding to STXBP1. an
essential component of the synaptic
vesicle exocytotic machinery
Angiotensinogen ANGT_HUMAN P01019 AGT X X X 53.1 Regulator of blood pressure. body
fluid and electrolyte homeostasis
Ankyrin repeat domain-
containing protein 20B AN20B_HUMAN Q5CZ79
ANKRD20
A8P X 93.9 -
Ankyrin repeat and sterile alpha
motif domain-containing protein
1B
ANS1B_HUMAN Q7Z6G8 ANKS1B X 138.1 Plays a role as a modulator of APP
processing
Annexin A6 ANXA6_HUMAN P08133 ANXA6 X 75.9 May associate with CD21
Antithrombin-III ANT3_HUMAN P01008 SERPINC1 X X X 52.6 Protease inhibitor
AP-3 complex subunit beta-2 AP3B2_HUMAN Q13367 AP3B2 X 119.1 Transporter activity
PC membrane recruitment
protein 2 AMER2_HUMAN Q8N7J2 AMER2 X 69.5
Negative regulator of the canonical
Wnt signaling pathway involved in
neuroectodermal patterning
Apolipoprotein A-I APOA1_HUMAN P02647 APOA1 X X X 30.8 Metabolism
Apolipoprotein A-II APOA2_HUMAN P02652 APOA2 X X X 11.2 Transport
Apolipoprotein A-IV APOA4_HUMAN P06727 APOA4 X X X 45.4 Hepatic transcellukar lipid transport
Apolipoprotein B-100 APOB_HUMAN P04114 APOB X X X 515.6
Recognition signal for the cellular
binding and internalization of LDL
particles by the apoB/E receptor
Apolipoprotein C-I APOC1_HUMAN P02654 APOC1 X X 9.3
Inhibitor of lipoprotein binding to the
low density lipoprotein (LDL)
receptor

265
Apolipoprotein C-II APOC2_HUMAN P02655 APOC2 X X X 11.3 Enzyme regulator activity
(lipoprotein lipase activator)
Apolipoprotein C-III APOC3_HUMAN P02656 APOC3 X X X 10.8 Enzyme regulator activity
(lipoprotein lipase inhibitor)
Apolipoprotein C-IV APOC4_HUMAN P55056 APOC4 X 14.5 Biological regulation
Apolipoprotein D APOD_HUMAN P05090 APOD X X X 21.3 Transporter activity
Apolipoprotein E APOE_HUMAN P02649 APOE X X X 36.1 Transporter activity and cell uptake
Apolipoprotein F APOF_HUMAN Q13790 APOF X X X 35.4 Transporter activity
Apolipoprotein L1 APOL1_HUMAN O14791 APOL1 X X X 43.9 Transporter activity
Apolipoprotein M APOM_HUMAN O95445 APOM X X X 21.2 Transporter activity
Apolipoprotein(a) APOA_HUMAN P08519 LPA X X X 501.3 Catalytic activity
ATP-binding cassette sub-family
B member 5 ABCB5_HUMAN Q2M3G0 ABCB5 X X 138.6 Transporter activity
ATP-binding cassette sub-family
F member 1 ABCF1_HUMAN Q8NE71 ABCF1 X X X
95.9
Transporter activity
Attractin ATRN_HUMAN O75882 ATRN X X X 158.5
Involved in the initial immune cell
clustering during inflammatory
response and may regulate
chemotactic activity of chemokines
Atypical kinase COQ8B.
mitochondrial COQ8B_HUMAN Q96D53 COQ8B X 60.1
Involved in the biosynthesis of
coenzyme Q
Beta-2-glycoprotein 1 APOH_HUMAN P02749 APOH X X X 38.3 Transporter activity
Biotinidase BTD_HUMAN P43251 BTD X X 61.1 Catalytic activity
Beta-ureidopropionase BUP1_HUMAN Q9UBR1 UPB1 X 43.1 Catalytic activity
Bile salt-activated lipase CEL_HUMAN P19835 CEL X 79.3 Catalytic activity
Bone morphogenetic protein 10 BMP10_HUMAN O95393 BMP10 X 48.0
Inhibits endothelial cell migration and
growth. May reduce cell migration
and cell matrix adhesion in breast
cancer cell lines.
Brefeldin A-inhibited guanine
nucleotide-exchange protein 3 BIG3_HUMAN Q5TH69 ARFGEF3 X 240.6
Participates in the regulation of
systemic glucose homeostasis
Bromodomain adjacent to zinc
finger domain protein 2A BAZ2A_HUMAN Q9UIF9 BAZ2A X 211.2
Essential component of the NoRC
(nucleolar remodeling complex)
complex that mediates silencing of a
fraction of rDNA
C4b-binding protein alpha chain C4BPA_HUMAN P04003 C4BPA X X X 67.1 Enzyme regulator activity (inhibitor)
C4b-binding protein beta chain C4BPB_HUMAN P20851 C4BPB X X X 28.4 Enzyme regulator activity (inhibitor)
85/88 kDa calcium-independent
phospholipase A2 PLPL9_HUMAN O60733 PLA2G6 X 89.9 Catalytic activity
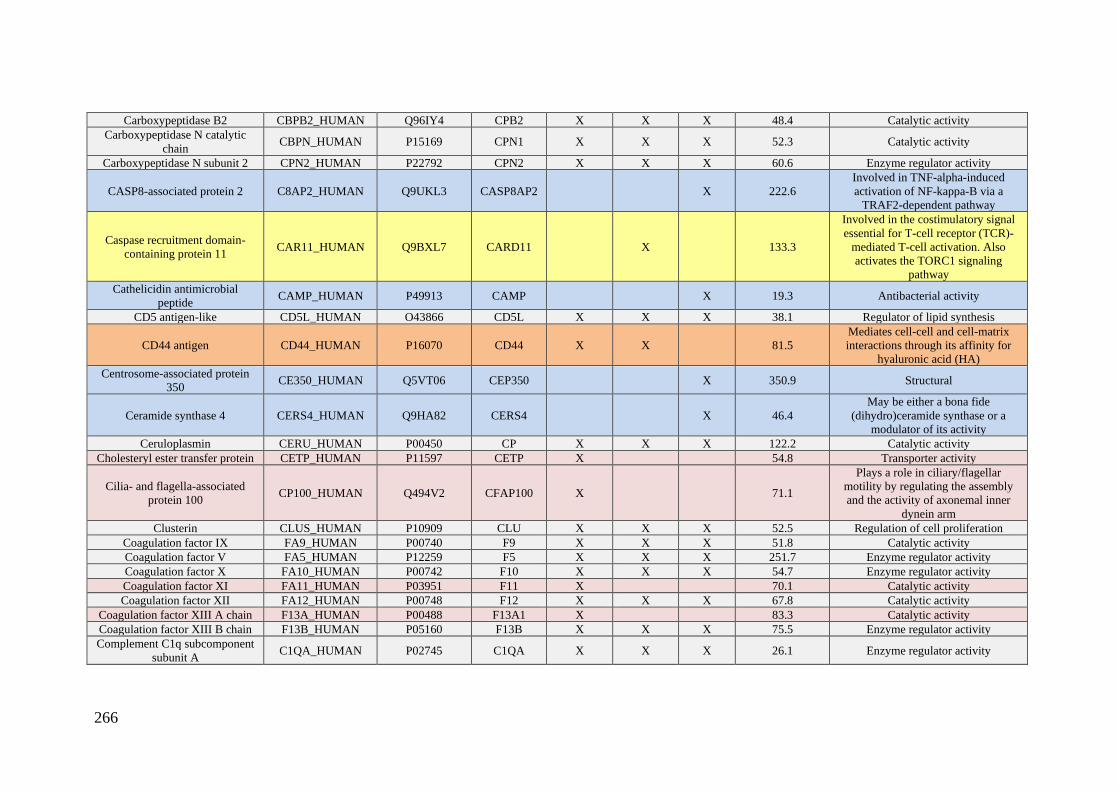
266
Carboxypeptidase B2 CBPB2_HUMAN Q96IY4 CPB2 X X X 48.4 Catalytic activity
Carboxypeptidase N catalytic
chain CBPN_HUMAN P15169 CPN1 X X X 52.3 Catalytic activity
Carboxypeptidase N subunit 2 CPN2_HUMAN P22792 CPN2 X X X 60.6 Enzyme regulator activity
CASP8-associated protein 2 C8AP2_HUMAN Q9UKL3 CASP8AP2 X 222.6
Involved in TNF-alpha-induced
activation of NF-kappa-B via a
TRAF2-dependent pathway
Caspase recruitment domain-
containing protein 11 CAR11_HUMAN Q9BXL7 CARD11 X 133.3
Involved in the costimulatory signal
essential for T-cell receptor (TCR)-
mediated T-cell activation. Also
activates the TORC1 signaling
pathway
Cathelicidin antimicrobial
peptide CAMP_HUMAN P49913 CAMP X 19.3 Antibacterial activity
CD5 antigen-like CD5L_HUMAN O43866 CD5L X X X 38.1 Regulator of lipid synthesis
CD44 antigen CD44_HUMAN P16070 CD44 X X 81.5
Mediates cell-cell and cell-matrix
interactions through its affinity for
hyaluronic acid (HA)
Centrosome-associated protein
350 CE350_HUMAN Q5VT06 CEP350 X 350.9 Structural
Ceramide synthase 4 CERS4_HUMAN Q9HA82 CERS4 X 46.4
May be either a bona fide
(dihydro)ceramide synthase or a
modulator of its activity
Ceruloplasmin CERU_HUMAN P00450 CP X X X 122.2 Catalytic activity
Cholesteryl ester transfer protein CETP_HUMAN P11597 CETP X 54.8 Transporter activity
Cilia- and flagella-associated
protein 100 CP100_HUMAN Q494V2 CFAP100 X 71.1
Plays a role in ciliary/flagellar
motility by regulating the assembly
and the activity of axonemal inner
dynein arm
Clusterin CLUS_HUMAN P10909 CLU X X X 52.5 Regulation of cell proliferation
Coagulation factor IX FA9_HUMAN P00740 F9 X X X 51.8 Catalytic activity
Coagulation factor V FA5_HUMAN P12259 F5 X X X 251.7 Enzyme regulator activity
Coagulation factor X FA10_HUMAN P00742 F10 X X X 54.7 Enzyme regulator activity
Coagulation factor XI FA11_HUMAN P03951 F11 X 70.1 Catalytic activity
Coagulation factor XII FA12_HUMAN P00748 F12 X X X 67.8 Catalytic activity
Coagulation factor XIII A chain F13A_HUMAN P00488 F13A1 X 83.3 Catalytic activity
Coagulation factor XIII B chain F13B_HUMAN P05160 F13B X X X 75.5 Enzyme regulator activity
Complement C1q subcomponent
subunit A C1QA_HUMAN P02745 C1QA X X X 26.1 Enzyme regulator activity

267
Complement C1q subcomponent
subunit B C1QB_HUMAN P02746 C1QB X X X 26.7 Enzyme regulator activity
Complement C1q subcomponent
subunit C C1QC_HUMAN P02747 C1QC X X X 25.8 Enzyme regulator activity
Complement C1r subcomponent C1R_HUMAN P00736 C1R X X X 80.1 Catalytic activity
Complement C1r subcomponent-
like protein C1RL_HUMAN Q9NZP8 C1RL X X X 53.5 Enzyme regulator activity
Complement C1s subcomponent C1S_HUMAN P09871 C1S X X X 76.7 Catalytic activity
Complement C2 CO2_HUMAN P06681 C2 X X X 83.3 Catalytic activity
Complement C3 CO3_HUMAN P01024 C3 X X X 187.1 Immune response
Complement C4-A CO4A_HUMAN P0C0L4 C4A X X X 192.8 Inflammatory response
Complement C4-B CO4B_HUMAN P0C0L5 C4B X X X 192.7 Enzyme regulator activity
Complement C5 CO5_HUMAN P01031 C5 X X X 188.3 Enzyme regulator activity
Complement component C6 CO6_HUMAN P13671 C6 X X 104.8 Structural
Complement component C7 CO7_HUMAN P10643 C7 X X X 93.5 Structural
Complement component C8
alpha chain CO8A_HUMAN P07357 C8A X X 65.2 Structural
Complement component C8 beta
chain CO8B_HUMAN P07358 C8B X X X 67.1 Structural
Complement component C8
gamma chain CO8G_HUMAN P07360 C8G X X X 22.3 Structural
Complement component C9 CO9_HUMAN P02748 C9 X X X 63.2 Structural
Complement factor B CFAB_HUMAN P00751 CFB X X X 85.5 Immune response
Complement factor D CFAD_HUMAN P00746 CFD X X X 27.1 Catalytic activity
Complement factor H CFAH_HUMAN P08603 CFH X X X 139.1 Enzyme regulator activity
Complement factor H-related
protein 1 FHR1_HUMAN Q03591 CFHR1 X X X 37.6 Plays a role in lipid metabolism
Complement factor H-related
protein 2 FHR2_HUMAN P36980 CFHR2 X X X 30.6 Plays a role in lipid metabolism
Complement factor H-related
protein 3 FHR3_HUMAN Q02985 CFHR3 X X 37.3 Involved in complement regulation
Complement factor H-related
protein 4 FHR4_HUMAN Q92496 CFHR4 X X 65.3 Plays a role in lipid metabolism
Complement factor I CFAI_HUMAN P05156 CFI X X X 65.7 Catalytic activity
Contactin-1 CNTN1_HUMAN Q12860 CNTN1 X 113.3 Mediates cell surface interactions
during nervous system development.
Contactin-associated protein-like
2 CNTP2_HUMAN Q9UHC6 CNTNAP2 X 148.2
Plays a role in the formation of
functional distinct domains critical for
saltatory conduction of nerve
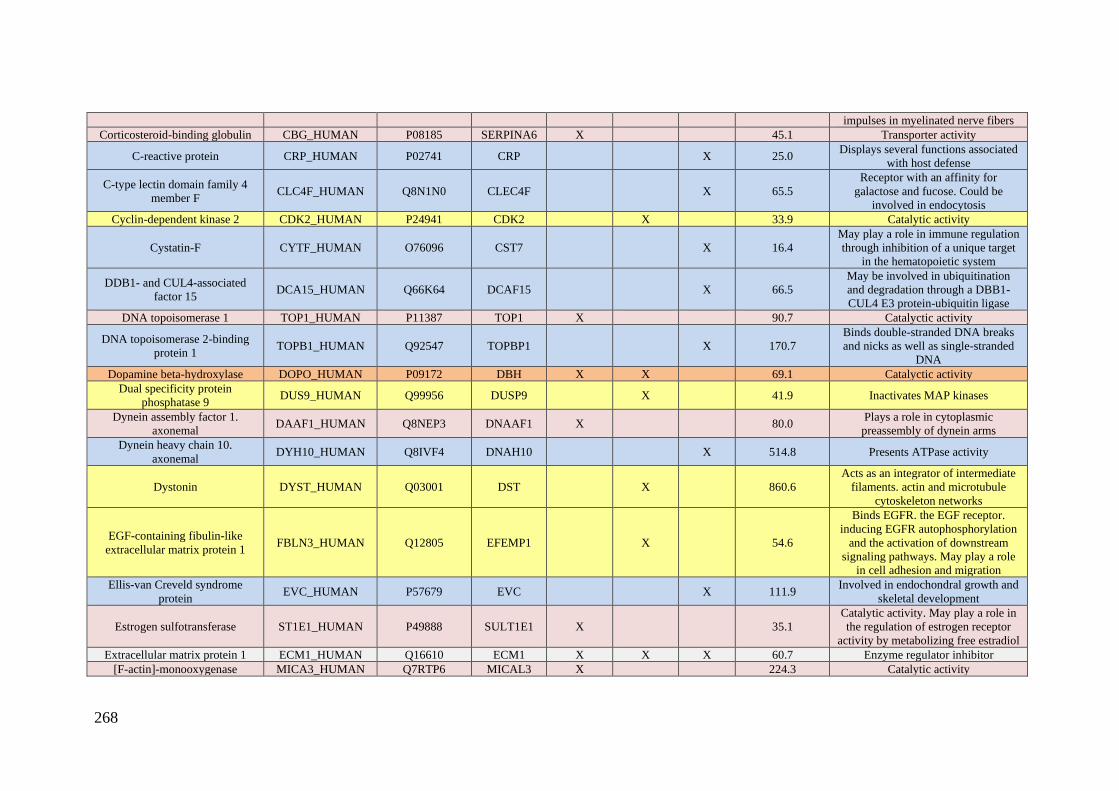
268
impulses in myelinated nerve fibers
Corticosteroid-binding globulin CBG_HUMAN P08185 SERPINA6 X 45.1 Transporter activity
C-reactive protein CRP_HUMAN P02741 CRP X 25.0 Displays several functions associated
with host defense
C-type lectin domain family 4
member F CLC4F_HUMAN Q8N1N0 CLEC4F X 65.5
Receptor with an affinity for
galactose and fucose. Could be
involved in endocytosis
Cyclin-dependent kinase 2 CDK2_HUMAN P24941 CDK2 X 33.9 Catalytic activity
Cystatin-F CYTF_HUMAN O76096 CST7 X 16.4
May play a role in immune regulation
through inhibition of a unique target
in the hematopoietic system
DDB1- and CUL4-associated
factor 15 DCA15_HUMAN Q66K64 DCAF15 X 66.5
May be involved in ubiquitination
and degradation through a DBB1-
CUL4 E3 protein-ubiquitin ligase
DNA topoisomerase 1 TOP1_HUMAN P11387 TOP1 X 90.7 Catalyctic activity
DNA topoisomerase 2-binding
protein 1 TOPB1_HUMAN Q92547 TOPBP1 X 170.7
Binds double-stranded DNA breaks
and nicks as well as single-stranded
DNA
Dopamine beta-hydroxylase DOPO_HUMAN P09172 DBH X X 69.1 Catalyctic activity
Dual specificity protein
phosphatase 9 DUS9_HUMAN Q99956 DUSP9 X 41.9 Inactivates MAP kinases
Dynein assembly factor 1.
axonemal DAAF1_HUMAN Q8NEP3 DNAAF1 X 80.0
Plays a role in cytoplasmic
preassembly of dynein arms
Dynein heavy chain 10.
axonemal DYH10_HUMAN Q8IVF4 DNAH10 X 514.8 Presents ATPase activity
Dystonin DYST_HUMAN Q03001 DST X 860.6
Acts as an integrator of intermediate
filaments. actin and microtubule
cytoskeleton networks
EGF-containing fibulin-like
extracellular matrix protein 1 FBLN3_HUMAN Q12805 EFEMP1 X 54.6
Binds EGFR. the EGF receptor.
inducing EGFR autophosphorylation
and the activation of downstream
signaling pathways. May play a role
in cell adhesion and migration
Ellis-van Creveld syndrome
protein EVC_HUMAN P57679 EVC X 111.9
Involved in endochondral growth and
skeletal development
Estrogen sulfotransferase ST1E1_HUMAN P49888 SULT1E1 X 35.1
Catalytic activity. May play a role in
the regulation of estrogen receptor
activity by metabolizing free estradiol
Extracellular matrix protein 1 ECM1_HUMAN Q16610 ECM1 X X X 60.7 Enzyme regulator inhibitor
[F-actin]-monooxygenase MICA3_HUMAN Q7RTP6 MICAL3 X 224.3 Catalytic activity
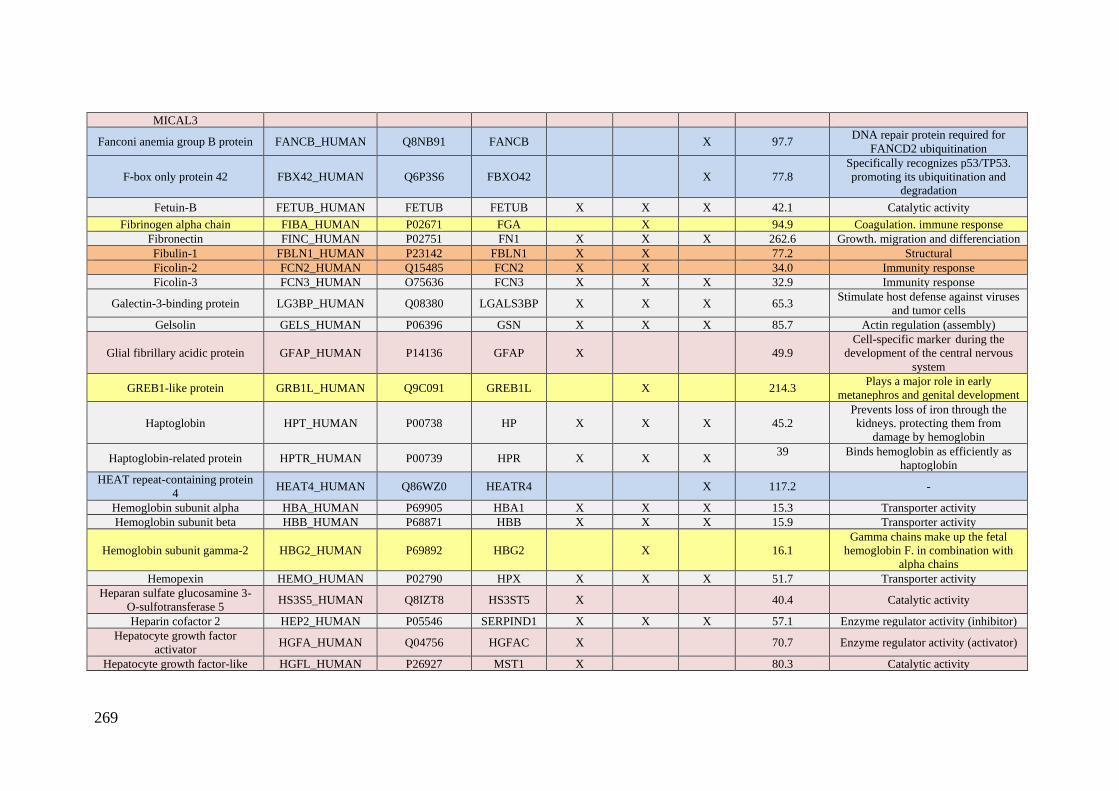
269
MICAL3
Fanconi anemia group B protein FANCB_HUMAN Q8NB91 FANCB X 97.7 DNA repair protein required for
FANCD2 ubiquitination
F-box only protein 42 FBX42_HUMAN Q6P3S6 FBXO42 X 77.8
Specifically recognizes p53/TP53.
promoting its ubiquitination and
degradation
Fetuin-B FETUB_HUMAN FETUB FETUB X X X 42.1 Catalytic activity
Fibrinogen alpha chain FIBA_HUMAN P02671 FGA X 94.9 Coagulation. immune response
Fibronectin FINC_HUMAN P02751 FN1 X X X 262.6 Growth. migration and differenciation
Fibulin-1 FBLN1_HUMAN P23142 FBLN1 X X 77.2 Structural
Ficolin-2 FCN2_HUMAN Q15485 FCN2 X X 34.0 Immunity response
Ficolin-3 FCN3_HUMAN O75636 FCN3 X X X 32.9 Immunity response
Galectin-3-binding protein LG3BP_HUMAN Q08380 LGALS3BP X X X 65.3 Stimulate host defense against viruses
and tumor cells
Gelsolin GELS_HUMAN P06396 GSN X X X 85.7 Actin regulation (assembly)
Glial fibrillary acidic protein GFAP_HUMAN P14136 GFAP X 49.9
Cell-specific marker during the
development of the central nervous
system
GREB1-like protein GRB1L_HUMAN Q9C091 GREB1L X 214.3 Plays a major role in early
metanephros and genital development
Haptoglobin HPT_HUMAN P00738 HP X X X 45.2
Prevents loss of iron through the
kidneys. protecting them from
damage by hemoglobin
Haptoglobin-related protein HPTR_HUMAN P00739 HPR X X X 39
Binds hemoglobin as efficiently as
haptoglobin
HEAT repeat-containing protein
4 HEAT4_HUMAN Q86WZ0 HEATR4 X 117.2 -
Hemoglobin subunit alpha HBA_HUMAN P69905 HBA1 X X X 15.3 Transporter activity
Hemoglobin subunit beta HBB_HUMAN P68871 HBB X X X 15.9 Transporter activity
Hemoglobin subunit gamma-2 HBG2_HUMAN P69892 HBG2 X 16.1
Gamma chains make up the fetal
hemoglobin F. in combination with
alpha chains
Hemopexin HEMO_HUMAN P02790 HPX X X X 51.7 Transporter activity
Heparan sulfate glucosamine 3-
O-sulfotransferase 5 HS3S5_HUMAN Q8IZT8 HS3ST5 X 40.4 Catalytic activity
Heparin cofactor 2 HEP2_HUMAN P05546 SERPIND1 X X X 57.1 Enzyme regulator activity (inhibitor)
Hepatocyte growth factor
activator HGFA_HUMAN Q04756 HGFAC X 70.7 Enzyme regulator activity (activator)
Hepatocyte growth factor-like HGFL_HUMAN P26927 MST1 X 80.3 Catalytic activity
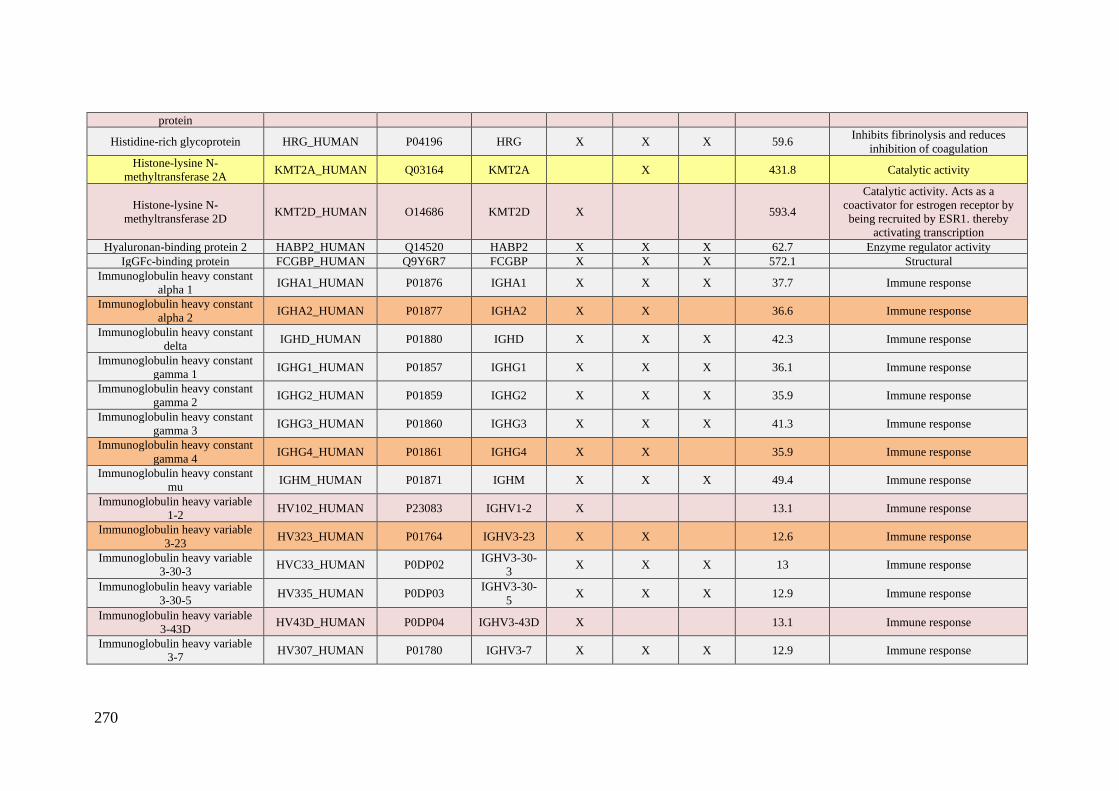
270
protein
Histidine-rich glycoprotein HRG_HUMAN P04196 HRG X X X 59.6 Inhibits fibrinolysis and reduces
inhibition of coagulation
Histone-lysine N-
methyltransferase 2A KMT2A_HUMAN Q03164 KMT2A X 431.8 Catalytic activity
Histone-lysine N-
methyltransferase 2D KMT2D_HUMAN O14686 KMT2D X 593.4
Catalytic activity. Acts as a
coactivator for estrogen receptor by
being recruited by ESR1. thereby
activating transcription
Hyaluronan-binding protein 2 HABP2_HUMAN Q14520 HABP2 X X X 62.7 Enzyme regulator activity
IgGFc-binding protein FCGBP_HUMAN Q9Y6R7 FCGBP X X X 572.1 Structural
Immunoglobulin heavy constant
alpha 1 IGHA1_HUMAN P01876 IGHA1 X X X 37.7 Immune response
Immunoglobulin heavy constant
alpha 2 IGHA2_HUMAN P01877 IGHA2 X X 36.6 Immune response
Immunoglobulin heavy constant
delta IGHD_HUMAN P01880 IGHD X X X 42.3 Immune response
Immunoglobulin heavy constant
gamma 1 IGHG1_HUMAN P01857 IGHG1 X X X 36.1 Immune response
Immunoglobulin heavy constant
gamma 2 IGHG2_HUMAN P01859 IGHG2 X X X 35.9 Immune response
Immunoglobulin heavy constant
gamma 3 IGHG3_HUMAN P01860 IGHG3 X X X 41.3 Immune response
Immunoglobulin heavy constant
gamma 4 IGHG4_HUMAN P01861 IGHG4 X X 35.9 Immune response
Immunoglobulin heavy constant
mu IGHM_HUMAN P01871 IGHM X X X 49.4 Immune response
Immunoglobulin heavy variable
1-2 HV102_HUMAN P23083 IGHV1-2 X 13.1 Immune response
Immunoglobulin heavy variable
3-23 HV323_HUMAN P01764 IGHV3-23 X X 12.6 Immune response
Immunoglobulin heavy variable
3-30-3 HVC33_HUMAN P0DP02
IGHV3-30-
3 X X X 13 Immune response
Immunoglobulin heavy variable
3-30-5 HV335_HUMAN P0DP03
IGHV3-30-
5 X X X 12.9 Immune response
Immunoglobulin heavy variable
3-43D HV43D_HUMAN P0DP04 IGHV3-43D X 13.1 Immune response
Immunoglobulin heavy variable
3-7 HV307_HUMAN P01780 IGHV3-7 X X X 12.9 Immune response
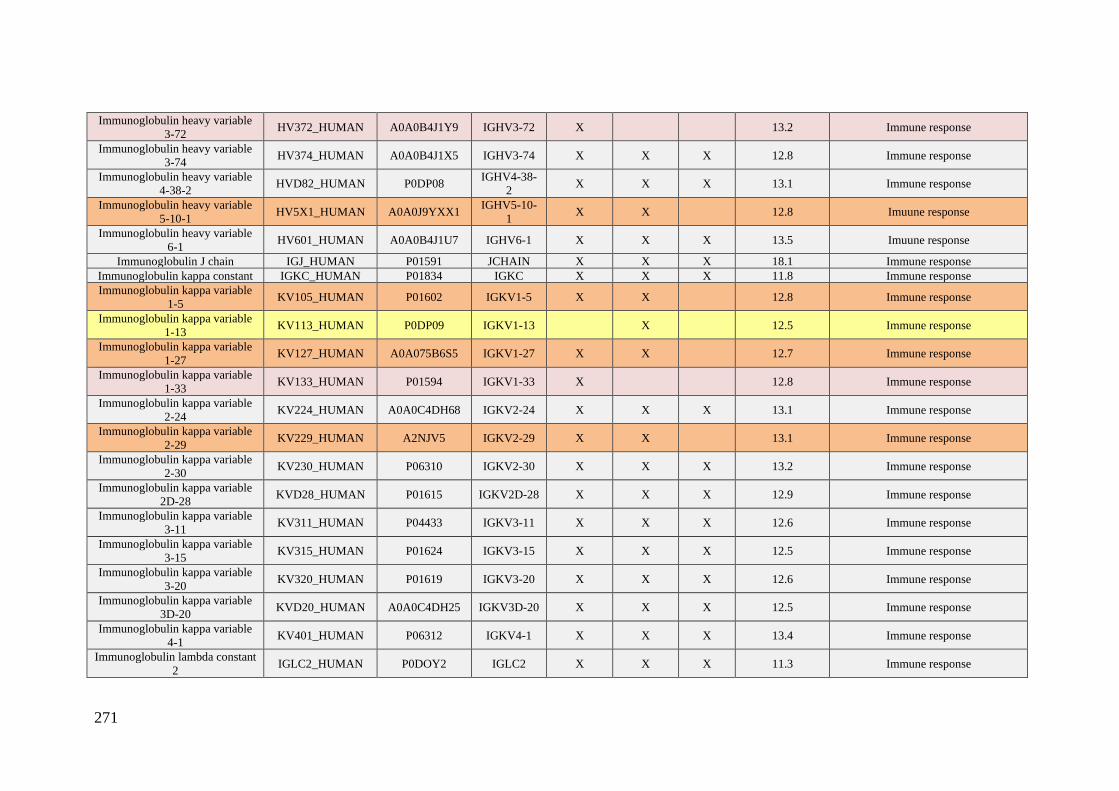
271
Immunoglobulin heavy variable
3-72 HV372_HUMAN A0A0B4J1Y9 IGHV3-72 X 13.2 Immune response
Immunoglobulin heavy variable
3-74 HV374_HUMAN A0A0B4J1X5 IGHV3-74 X X X 12.8 Immune response
Immunoglobulin heavy variable
4-38-2 HVD82_HUMAN P0DP08
IGHV4-38-
2 X X X 13.1 Immune response
Immunoglobulin heavy variable
5-10-1 HV5X1_HUMAN A0A0J9YXX1
IGHV5-10-
1 X X 12.8 Imuune response
Immunoglobulin heavy variable
6-1 HV601_HUMAN A0A0B4J1U7 IGHV6-1 X X X 13.5 Imuune response
Immunoglobulin J chain IGJ_HUMAN P01591 JCHAIN X X X 18.1 Immune response
Immunoglobulin kappa constant IGKC_HUMAN P01834 IGKC X X X 11.8 Immune response
Immunoglobulin kappa variable
1-5 KV105_HUMAN P01602 IGKV1-5 X X 12.8 Immune response
Immunoglobulin kappa variable
1-13 KV113_HUMAN P0DP09 IGKV1-13 X 12.5 Immune response
Immunoglobulin kappa variable
1-27 KV127_HUMAN A0A075B6S5 IGKV1-27 X X 12.7 Immune response
Immunoglobulin kappa variable
1-33 KV133_HUMAN P01594 IGKV1-33 X 12.8 Immune response
Immunoglobulin kappa variable
2-24 KV224_HUMAN A0A0C4DH68 IGKV2-24 X X X 13.1 Immune response
Immunoglobulin kappa variable
2-29 KV229_HUMAN A2NJV5 IGKV2-29 X X 13.1 Immune response
Immunoglobulin kappa variable
2-30 KV230_HUMAN P06310 IGKV2-30 X X X 13.2 Immune response
Immunoglobulin kappa variable
2D-28 KVD28_HUMAN P01615 IGKV2D-28 X X X 12.9 Immune response
Immunoglobulin kappa variable
3-11 KV311_HUMAN P04433 IGKV3-11 X X X 12.6 Immune response
Immunoglobulin kappa variable
3-15 KV315_HUMAN P01624 IGKV3-15 X X X 12.5 Immune response
Immunoglobulin kappa variable
3-20 KV320_HUMAN P01619 IGKV3-20 X X X 12.6 Immune response
Immunoglobulin kappa variable
3D-20 KVD20_HUMAN A0A0C4DH25 IGKV3D-20 X X X 12.5 Immune response
Immunoglobulin kappa variable
4-1 KV401_HUMAN P06312 IGKV4-1 X X X 13.4 Immune response
Immunoglobulin lambda constant
2 IGLC2_HUMAN P0DOY2 IGLC2 X X X 11.3 Immune response
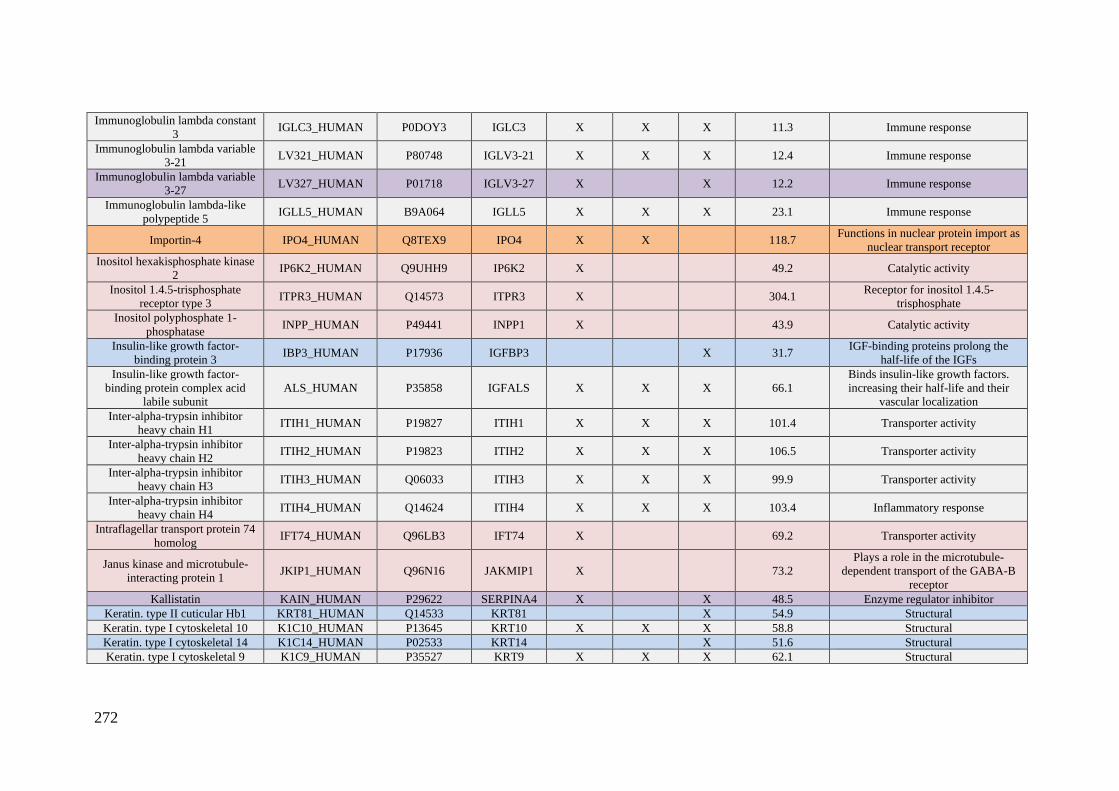
272
Immunoglobulin lambda constant
3 IGLC3_HUMAN P0DOY3 IGLC3 X X X 11.3 Immune response
Immunoglobulin lambda variable
3-21 LV321_HUMAN P80748 IGLV3-21 X X X 12.4 Immune response
Immunoglobulin lambda variable
3-27 LV327_HUMAN P01718 IGLV3-27 X X 12.2 Immune response
Immunoglobulin lambda-like
polypeptide 5 IGLL5_HUMAN B9A064 IGLL5 X X X 23.1 Immune response
Importin-4 IPO4_HUMAN Q8TEX9 IPO4 X X 118.7 Functions in nuclear protein import as
nuclear transport receptor
Inositol hexakisphosphate kinase
2 IP6K2_HUMAN Q9UHH9 IP6K2 X 49.2 Catalytic activity
Inositol 1.4.5-trisphosphate
receptor type 3 ITPR3_HUMAN Q14573 ITPR3 X 304.1
Receptor for inositol 1.4.5-
trisphosphate
Inositol polyphosphate 1-
phosphatase INPP_HUMAN P49441 INPP1 X 43.9 Catalytic activity
Insulin-like growth factor-
binding protein 3 IBP3_HUMAN P17936 IGFBP3 X 31.7
IGF-binding proteins prolong the
half-life of the IGFs
Insulin-like growth factor-
binding protein complex acid
labile subunit
ALS_HUMAN P35858 IGFALS X X X 66.1
Binds insulin-like growth factors.
increasing their half-life and their
vascular localization
Inter-alpha-trypsin inhibitor
heavy chain H1 ITIH1_HUMAN P19827 ITIH1 X X X 101.4 Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H2 ITIH2_HUMAN P19823 ITIH2 X X X 106.5 Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H3 ITIH3_HUMAN Q06033 ITIH3 X X X 99.9 Transporter activity
Inter-alpha-trypsin inhibitor
heavy chain H4 ITIH4_HUMAN Q14624 ITIH4 X X X 103.4 Inflammatory response
Intraflagellar transport protein 74
homolog IFT74_HUMAN Q96LB3 IFT74 X 69.2 Transporter activity
Janus kinase and microtubule-
interacting protein 1 JKIP1_HUMAN Q96N16 JAKMIP1 X 73.2
Plays a role in the microtubule-
dependent transport of the GABA-B
receptor
Kallistatin KAIN_HUMAN P29622 SERPINA4 X X 48.5 Enzyme regulator inhibitor
Keratin. type II cuticular Hb1 KRT81_HUMAN Q14533 KRT81 X 54.9 Structural
Keratin. type I cytoskeletal 10 K1C10_HUMAN P13645 KRT10 X X X 58.8 Structural
Keratin. type I cytoskeletal 14 K1C14_HUMAN P02533 KRT14 X 51.6 Structural
Keratin. type I cytoskeletal 9 K1C9_HUMAN P35527 KRT9 X X X 62.1 Structural

273
Keratin. type II cytoskeletal 1 K2C1_HUMAN P04264 KRT1 X X X 66.1 Enzyme regulator activity
Keratin. type II cytoskeletal 2
epidermal K22E_HUMAN P35908 KRT2 X X X 65.4 Structural
Kininogen-1 KNG1_HUMAN P01042 KNG1 X X X 71.9 Enzyme regulator activity
Lactotransferrin TRFL_HUMAN P02788 LTF X 78.2 Transporter activity
Lebercilin LCA5_HUMAN Q86VQ0 LCA5 X X 80.5 Transporter activity
Leucine-rich alpha-2-
glycoprotein A2GL_HUMAN P02750 LRG1 X X 38.2
Protein-protein interaction. signal
transduction. and cell adhesion and
development
LINE-1 type transposase domain-
containing protein 1 LITD1_HUMAN Q5T7N2 L1TD1 X X 98.8 Single-stranded RNA binding
Lipopolysaccharide-binding
protein LBP_HUMAN P18428 LBP X X X 53.4 Immune response.
LRP chaperone MESD MESD_HUMAN Q14696 MESD X 26.1
Assisting the folding of beta-
propeller/EGF modules within the
family of low-density lipoprotein
receptors (LDLRs)
Lumican LUM_HUMAN P51884 LUM X X X 38.4 Collagen binding. extracellular matrix
structural constituent
Lysine-specific demethylase 4C KDM4C_HUMAN Q9H3R0 KDM4C X X 119.9 Catalytic activity
Lysosomal-trafficking regulator LYST_HUMAN Q99698 LYST X X 429.1
Required for sorting endosomal
resident proteins into late
multivesicular endosomes
MaFF-interacting protein MAFIP_HUMAN Q8WZ33 MAFIP X 13.9 Inhibits cell growth and colony-
forming efficiency
Mannan-binding lectin serine
protease 1 MASP1_HUMAN P48740 MASP1 X X X 79.2 Enzyme regulator activity
Mannan-binding lectin serine
protease 2 MASP2_HUMAN O00187 MASP2 X 75.7 Catalytic activity
MANSC domain-containing
protein 1 MANS1_HUMAN Q9H8J5 MANSC1 X 46.8 -
Matrix metalloproteinase-9 MMP9_HUMAN P14780 MMP9 X X 78.5 Catalytic activity
Meckelin MKS3_HUMAN Q5HYA8 TMEM67 X 111.7 Required for ciliary structure and
function
Microtubule-associated protein
1A MAP1A_HUMAN P78559 MAP1A X X 305.5
Structural protein involved in the
filamentous cross-bridging between
microtubules and other skeletal
elements
Microtubule-associated protein 2 MTAP2_HUMAN P11137 MAP2 X 199.5 Stabilizes the microtubules against

274
depolymerization
Mitochondrial 2-
oxoglutarate/malate carrier
protein
M2OM_HUMAN Q02978 SLC25A11 X 34.1 Catalytic and transporter activities
Monocyte differentiation antigen
CD14 CD14_HUMAN P08571 CD14 X X X 40.1 Immune response
MORC family CW-type zinc
finger protein 1 MORC1_HUMAN Q86VD1 MORC1 X 112.9 Zinc ion binding.
Mucolipin-3 MCLN3_HUMAN Q8TDD5 MCOLN3 X 64.2 Pays a role in the regulation of
membrane trafficking events
Multidrug resistance protein 1 MDR1_HUMAN P08183 ABCB1 X 141.5
Responsible for decreased drug
accumulation in multidrug-resistant
cells
NAD(P)H dehydrogenase
[quinone] 1 NQO1_HUMAN P15559 NQO1 X 30.9 Catalytic activity
N-acetyllactosaminide beta-1.3-
N-acetylglucosaminyltransferase
2
B3GN2_HUMAN Q9NY97 B3GNT2 X 46.0 Catalytic activity
N-acetylmuramoyl-L-alanine
amidase PGRP2_HUMAN Q96PD5 PGLYRP2 X X X 62.2 Catalytic activity
Nck-associated protein 1 NCKP1_HUMAN Q9Y2A7 NCKAP1 X 128.8
As component of the WAVE1
complex. required for BDNF-NTRK2
endocytic trafficking and signaling
from early endosomes
Nebulette NEBL_HUMAN O76041 NEBL X 116.4 Plays an important role in the
assembly of the Z-disk
Nesprin-1 SYNE1_HUMAN Q8NF91 SYNE1 X 1011.1
Forms a linking network between
organelles and the actin cytoskeleton
to maintain the subcellular spatial
organization
Nuclear distribution protein
nudE-like 1 NDEL1_HUMAN Q9GZM8 NDEL1 X X 38.4
Required for organization of the
cellular microtubule array and
microtubule anchoring at the
centrosome
Nuclear receptor coactivator 6 NCOA6_HUMAN Q14686 NCOA6 X 219.1 Involved in the coactivation of the
NF-kappa-B pathway
Nucleoporin NUP188 homolog NU188_HUMAN Q5SRE5 NUP188 X 196.1 May function as a component of the
nuclear pore complex (NPC)
PAN2-PAN3 deadenylation
complex subunit PAN3 PAN3_HUMAN Q58A45 PAN3 X 95.6
Regulatory subunit of the poly(A)-
nuclease (PAN) deadenylation
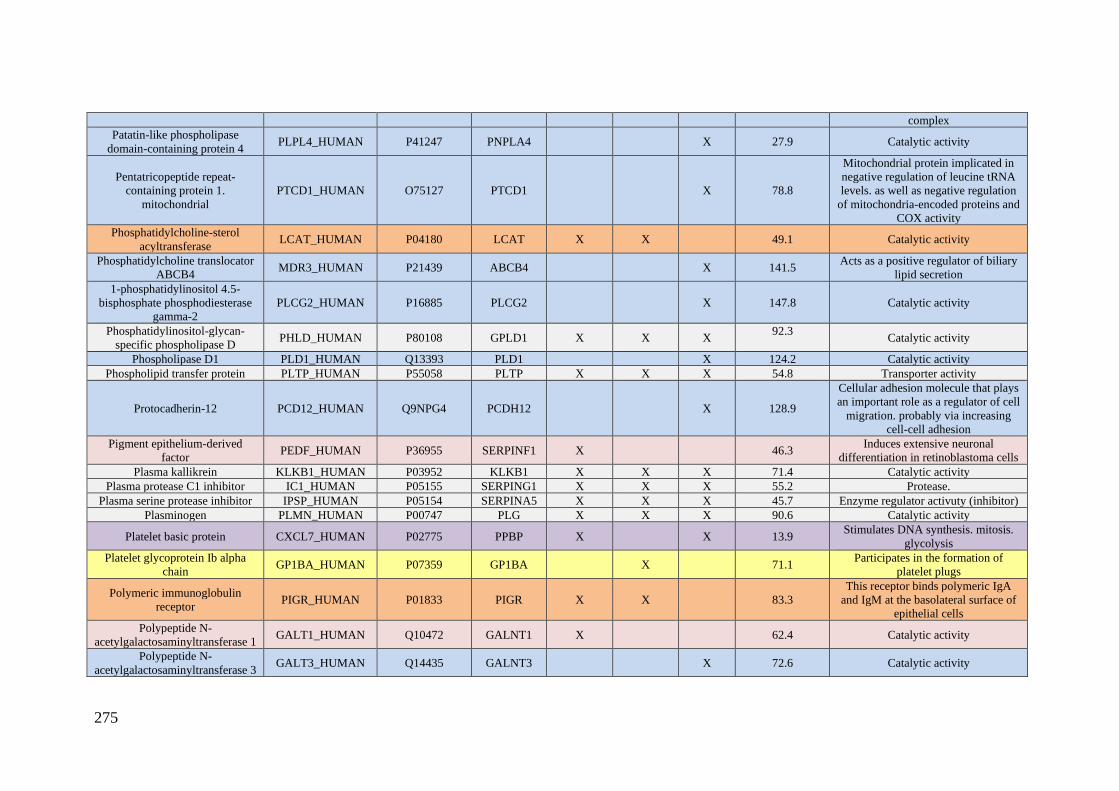
275
complex
Patatin-like phospholipase
domain-containing protein 4 PLPL4_HUMAN P41247 PNPLA4 X 27.9 Catalytic activity
Pentatricopeptide repeat-
containing protein 1.
mitochondrial
PTCD1_HUMAN O75127 PTCD1 X 78.8
Mitochondrial protein implicated in
negative regulation of leucine tRNA
levels. as well as negative regulation
of mitochondria-encoded proteins and
COX activity
Phosphatidylcholine-sterol
acyltransferase LCAT_HUMAN P04180 LCAT X X 49.1 Catalytic activity
Phosphatidylcholine translocator
ABCB4 MDR3_HUMAN P21439 ABCB4 X 141.5
Acts as a positive regulator of biliary
lipid secretion
1-phosphatidylinositol 4.5-
bisphosphate phosphodiesterase
gamma-2
PLCG2_HUMAN P16885 PLCG2 X 147.8 Catalytic activity
Phosphatidylinositol-glycan-
specific phospholipase D PHLD_HUMAN P80108 GPLD1 X X X
92.3
Catalytic activity
Phospholipase D1 PLD1_HUMAN Q13393 PLD1 X 124.2 Catalytic activity
Phospholipid transfer protein PLTP_HUMAN P55058 PLTP X X X 54.8 Transporter activity
Protocadherin-12 PCD12_HUMAN Q9NPG4 PCDH12 X 128.9
Cellular adhesion molecule that plays
an important role as a regulator of cell
migration. probably via increasing
cell-cell adhesion
Pigment epithelium-derived
factor PEDF_HUMAN P36955 SERPINF1 X 46.3
Induces extensive neuronal
differentiation in retinoblastoma cells
Plasma kallikrein KLKB1_HUMAN P03952 KLKB1 X X X 71.4 Catalytic activity
Plasma protease C1 inhibitor IC1_HUMAN P05155 SERPING1 X X X 55.2 Protease.
Plasma serine protease inhibitor IPSP_HUMAN P05154 SERPINA5 X X X 45.7 Enzyme regulator activuty (inhibitor)
Plasminogen PLMN_HUMAN P00747 PLG X X X 90.6 Catalytic activity
Platelet basic protein CXCL7_HUMAN P02775 PPBP X X 13.9 Stimulates DNA synthesis. mitosis.
glycolysis
Platelet glycoprotein Ib alpha
chain GP1BA_HUMAN P07359 GP1BA X 71.1
Participates in the formation of
platelet plugs
Polymeric immunoglobulin
receptor PIGR_HUMAN P01833 PIGR X X 83.3
This receptor binds polymeric IgA
and IgM at the basolateral surface of
epithelial cells
Polypeptide N-
acetylgalactosaminyltransferase 1 GALT1_HUMAN Q10472 GALNT1 X 62.4 Catalytic activity
Polypeptide N-
acetylgalactosaminyltransferase 3 GALT3_HUMAN Q14435 GALNT3 X 72.6 Catalytic activity
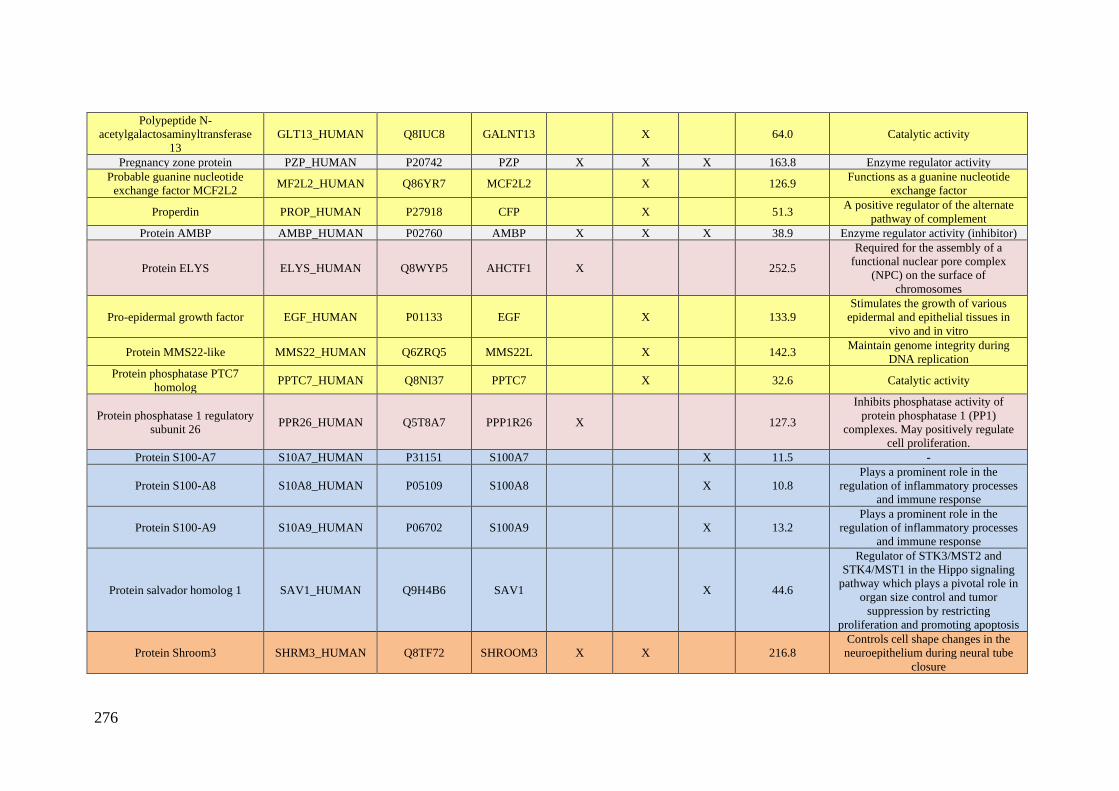
276
Polypeptide N-
acetylgalactosaminyltransferase
13
GLT13_HUMAN Q8IUC8 GALNT13 X 64.0 Catalytic activity
Pregnancy zone protein PZP_HUMAN P20742 PZP X X X 163.8 Enzyme regulator activity
Probable guanine nucleotide
exchange factor MCF2L2 MF2L2_HUMAN Q86YR7 MCF2L2 X 126.9
Functions as a guanine nucleotide
exchange factor
Properdin PROP_HUMAN P27918 CFP X 51.3 A positive regulator of the alternate
pathway of complement
Protein AMBP AMBP_HUMAN P02760 AMBP X X X 38.9 Enzyme regulator activity (inhibitor)
Protein ELYS ELYS_HUMAN Q8WYP5 AHCTF1 X 252.5
Required for the assembly of a
functional nuclear pore complex
(NPC) on the surface of
chromosomes
Pro-epidermal growth factor EGF_HUMAN P01133 EGF X 133.9
Stimulates the growth of various
epidermal and epithelial tissues in
vivo and in vitro
Protein MMS22-like MMS22_HUMAN Q6ZRQ5 MMS22L X 142.3 Maintain genome integrity during
DNA replication
Protein phosphatase PTC7
homolog PPTC7_HUMAN Q8NI37 PPTC7 X 32.6 Catalytic activity
Protein phosphatase 1 regulatory
subunit 26 PPR26_HUMAN Q5T8A7 PPP1R26 X 127.3
Inhibits phosphatase activity of
protein phosphatase 1 (PP1)
complexes. May positively regulate
cell proliferation.
Protein S100-A7 S10A7_HUMAN P31151 S100A7 X 11.5 -
Protein S100-A8 S10A8_HUMAN P05109 S100A8 X 10.8
Plays a prominent role in the
regulation of inflammatory processes
and immune response
Protein S100-A9 S10A9_HUMAN P06702 S100A9 X 13.2
Plays a prominent role in the
regulation of inflammatory processes
and immune response
Protein salvador homolog 1 SAV1_HUMAN Q9H4B6 SAV1 X 44.6
Regulator of STK3/MST2 and
STK4/MST1 in the Hippo signaling
pathway which plays a pivotal role in
organ size control and tumor
suppression by restricting
proliferation and promoting apoptosis
Protein Shroom3 SHRM3_HUMAN Q8TF72 SHROOM3 X X 216.8
Controls cell shape changes in the
neuroepithelium during neural tube
closure
277
Protein Z-dependent protease
inhibitor ZPI_HUMAN Q9UK55
SERPINA1
0 X 50.7 Enzyme regulator activity
Protein ZGRF1 ZGRF1_HUMAN Q86YA3 ZGRF1 X X X 236.6
Zinc ion binding that inhibits factors
Xa and XIa of the coagulation
cascade
Proteoglycan 4 PRG4_HUMAN Q92954 PRG4 X X X 151.1 Plays a role in boundary lubrication
within articulating joints
Prothrombin THRB_HUMAN P00734 F2 X X X 70.1 Catalytic activity
Putative solute carrier organic
anion transporter family member
1B7
SO1B7_HUMAN G3V0H7 SLCO1B7 X 71.2 May encode a non-functional
truncated protein
Prickle planar cell polarity
protein 3 PRIC3_HUMAN O43900 PRICKLE3 X 68.6
Involved in the planar cell polarity
(PCP) pathway that is essential for the
polarization of epithelial cells during
morphogenetic processes
Putative transmembrane protein
encoded by LINC00477 CL067_HUMAN Q96M19 LINC00477 X 18.2
Product of a dubious CDS prediction.
May be a non-coding RNA
Ras-interacting protein 1 RAIN_HUMAN Q5U651 RASIP1 X 103.4
Acts as a critical and vascular-
specific regulator of GTPase
signaling. cell architecture. and
adhesion
Retinol-binding protein 4 RET4_HUMAN P02753 RBP4 X X X 23.1 Transporter activity
Ribosomal protein S6 kinase
alpha-1 KS6A1_HUMAN Q15418 RPS6KA1 X 82.7 Catalytic activity
Roundabout homolog 4 ROBO4_HUMAN Q8WZ75 ROBO4 X 107.4
Mediates the inhibition of primary
endothelial cell migration by Slit
proteins
Secreted phosphoprotein 24 SPP24_HUMAN Q13103 SPP2 X 24.1
Bind cytokines of the TGF-β
superfamily and also activate
intracellular signaling pathways
Selenoprotein P SEPP1_HUMAN P49908 SELENOP X X X 43.2 Transporter activity
Serine protease 33 PRS33_HUMAN PRSS33 PRSS33 X 29.8 Catalytic activity
Serine-protein kinase ATM ATM_HUMAN Q13315 ATM X 350.7
Activates checkpoint signaling upon
double strand breaks (DSBs).
apoptosis and genotoxic stresses
Serotransferrin TRFE_HUMAN P02787 TF X X X 77.1 Transporter activity
Serum albumin ALBU_HUMAN P02768 ALB X X X 69.4 Osmotic pressure
Serum amyloid A-1 protein SAA1_HUMAN P0DJI8 SAA1 X X 13.5 Inflammatory response
Serum amyloid A-2 protein SAA2_HUMAN P0DJI9 SAA2 X X 13.5 Inflammatory response
Serum amyloid A-4 protein SAA4_HUMAN P35542 SAA4 X X X 14.7 Inflammatory response

278
Serum amyloid P-component SAMP_HUMAN P02743 APCS X 25.4
Can interact with DNA and histones
and may scavenge nuclear material
released from damaged circulating
cells
Serum paraoxonase/arylesterase
1 PON1_HUMAN P27169 PON1 X X X 39.7 Catalytic activity
Sex hormone-binding globulin SHBG_HUMAN P04278 SHBG X X X 43.8
Transporter activity. Regulates the
plasma metabolic clearance rate of
steroid hormones
Short coiled-coil protein SCOC_HUMAN Q9UIL1 SCOC X X 18.0 Positive regulator of amino acid
starvation-induced autophagy
Sodium-dependent noradrenaline
transporter SC6A2_HUMAN P23975 SLC6A2 X 69.3 Transporter activity
Structural maintenance of
chromosomes protein 6 SMC6_HUMAN Q96SB8 SMC6 X X 126.3 Structural
Supervillin SVIL_HUMAN O95425 SVIL X 247.7 Structural
Suppressor of tumorigenicity 7
protein ST7_HUMAN Q9NRC1 ST7 X 67.2 Acts as a tumor suppressor
Synaptotagmin-5 SYT5_HUMAN O00445 SYT5 X 42.9 May be involved in Ca2+-dependent
exocytosis of secretory vesicles
Syncoilin SYNCI_HUMAN Q9H7C4 SYNC X 55.3
Plays a supportive role in the efficient
coupling of mechanical stress
between the myofibril and fiber
exterior
TBC1 domain family member 9 TBCD9_HUMAN Q6ZT07 TBC1D9 X 143.2 Acts as a GTPase-activating protein
for Rab family protein(s)
Tetranectin TETN_HUMAN P05452 CLEC3B X X X 22.5 Involved in the packaging of
molecules destined for exocytosis
Thrombospondin-1 TSP1_HUMAN P07996 THBS1 X X X 129.4 Immune response
Thyroxine-binding globulin THBG_HUMAN P05543 SERPINA7 X 46.3 Transporter activity
Transferrin receptor protein 1 TFR1_HUMAN P02786 TFRC X X 84.9 Transporter activity
Transmembrane protein 198 TM198_HUMAN Q66K66 TMEM198 X 39.4
Promotes low-density lipoprotein
receptor-related protein 6 (LRP6)
phosphorylation
Transportin-1 TNPO1_HUMAN Q92973 TNPO1 X 102.3 Transporter activity
Transthyretin TTHY_HUMAN P02766 TTR X X 15.8 Transporter activity
Tudor domain-containing protein
1 TDRD1_HUMAN Q9BXT4 TDRD1 X 132.0
Acts via the piRNA metabolic
process. which mediates the
repression of transposable elements
during meiosis
279
Tudor domain-containing protein
5 TDRD5_HUMAN Q8NAT2 TDRD5 X 109.7
Acts via the piRNA metabolic
process. which mediates the
repression of transposable elements
during meiosis
Tyrosine--tRNA ligase.
cytoplasmic SYYC_HUMAN P54577 YARS X 59.1 Catalytic activity
Ubiquitin-like modifier-
activating enzyme 1 UBA1_HUMAN P22314 UBA1 X 117.8 Catalytic activity
E3 ubiquitin-protein ligase MSL2 MSL2_HUMAN Q9HCI7 MSL2 X 62.5 Component of histone
acetyltransferase complex
E3 ubiquitin-protein ligase
SHPRH SHPRH_HUMAN Q149N8 SHPRH X 193.1 Enzyme involved in DNA repair
Vasorin VASN_HUMAN Q6EMK4 VASN X 71.7 May act as an inhibitor of TGF-beta
signaling
Vigilin VIGLN_HUMAN Q00341 HDLBP X 141.4 Protect cells from over-accumulation
of cholesterol
Villin-like protein VILL_HUMAN O15195 VILL X 95.9 Tumor suppressor
Vitamin D-binding protein VTDB_HUMAN P02774 GC X X X 52.9 Transporter activity
Vitamin K-dependent protein C PROC_HUMAN P04070 PROC X X X 52.1 Catalytic activity
Vitamin K-dependent protein S PROS_HUMAN P07225 PROS1 X X X 75.1 Anticoagulant plasma protein
Vitronectin VTNC_HUMAN P04004 VTN X X X 54.3 Proteolysis regulation
von Willebrand factor VWF_HUMAN P04275 VWF X X 220 Plays a major role in blood
coagulation
VPS10 domain-containing
receptor SorCS3 SORC3_HUMAN Q9UPU3 SORCS3 X 135.7 Neuropeptide receptor activity
Wee1-like protein kinase WEE1_HUMAN P30291 WEE1 X 71.6 Acts as a negative regulator of entry
into mitosis
Zinc finger protein 99 ZNF99_HUMAN A8MXY4 ZNF99 X 100.8 May be involved in transcriptional
regulation
Zinc finger protein 114 ZN114_HUMAN Q8NC26 ZNF114 X 47.7 May be involved in transcriptional
regulation.
Zinc finger RNA-binding protein ZFR_HUMAN Q96KR1 ZFR X 117.0 Involved in the nucleocytoplasmic
shuttling of STAU2
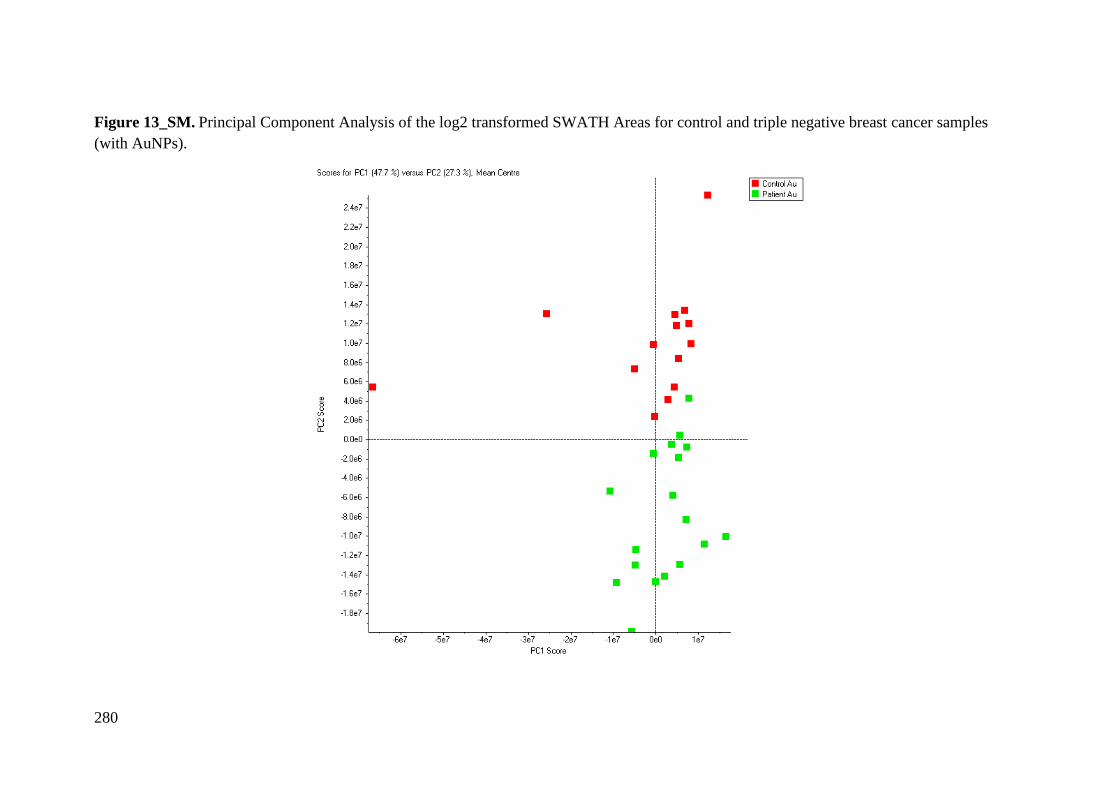
280
Figure 13_SM. Principal Component Analysis of the log2 transformed SWATH Areas for control and triple negative breast cancer samples
(with AuNPs).
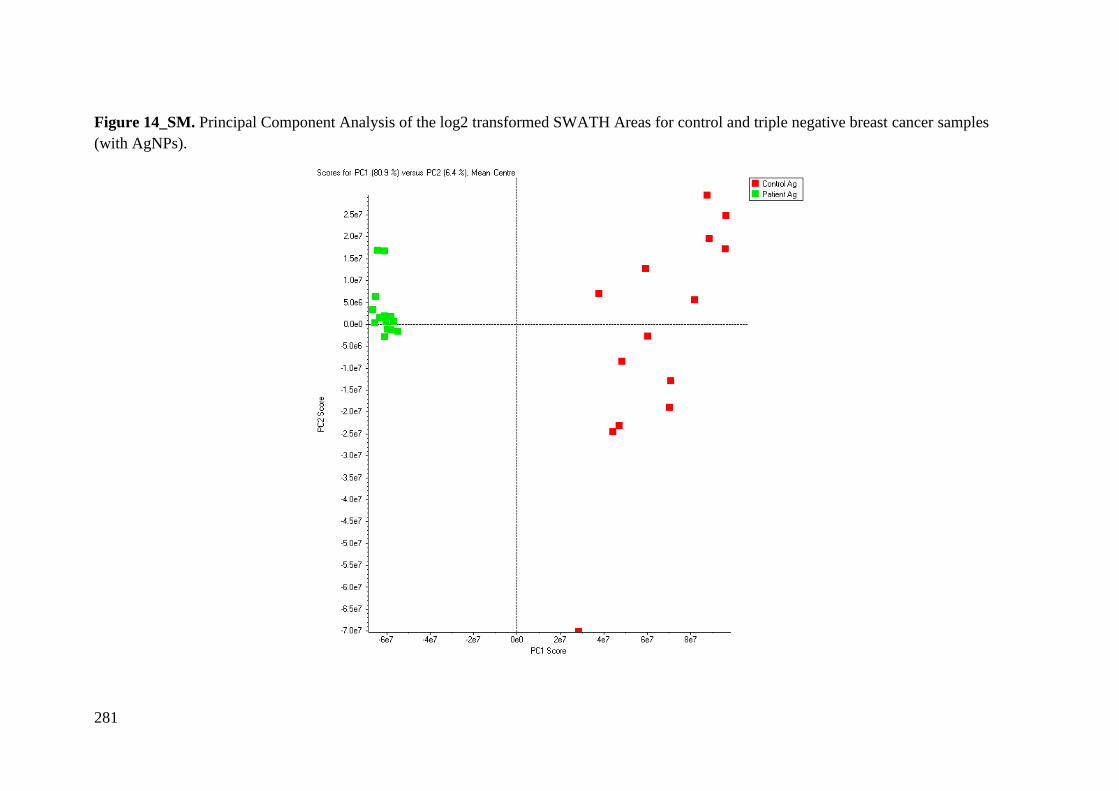
281
Figure 14_SM. Principal Component Analysis of the log2 transformed SWATH Areas for control and triple negative breast cancer samples
(with AgNPs).

282
Figure 15_SM. Principal Component Analysis of the log2 transformed SWATH Areas for control and triple negative breast cancer samples
(with MNPs).

ANNEX C
Supplemental Material Chapter 3

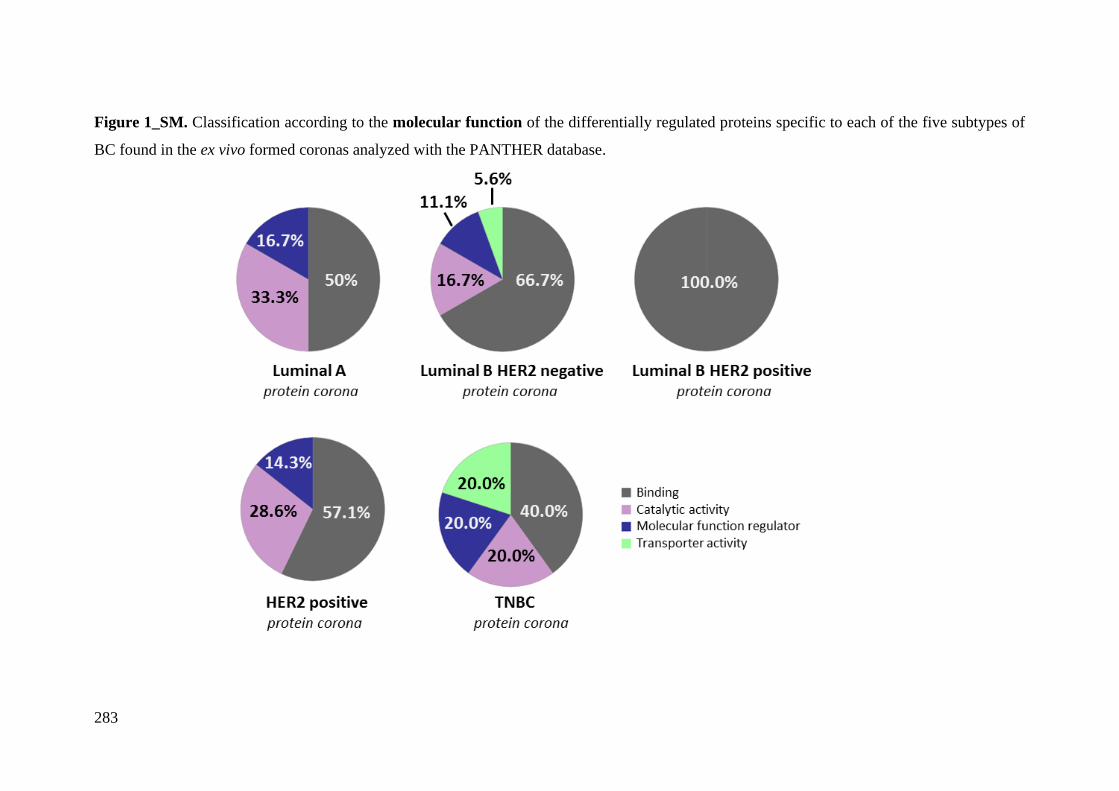
283
Figure 1_SM. Classification according to the molecular function of the differentially regulated proteins specific to each of the five subtypes of
BC found in the ex vivo formed coronas analyzed with the PANTHER database.
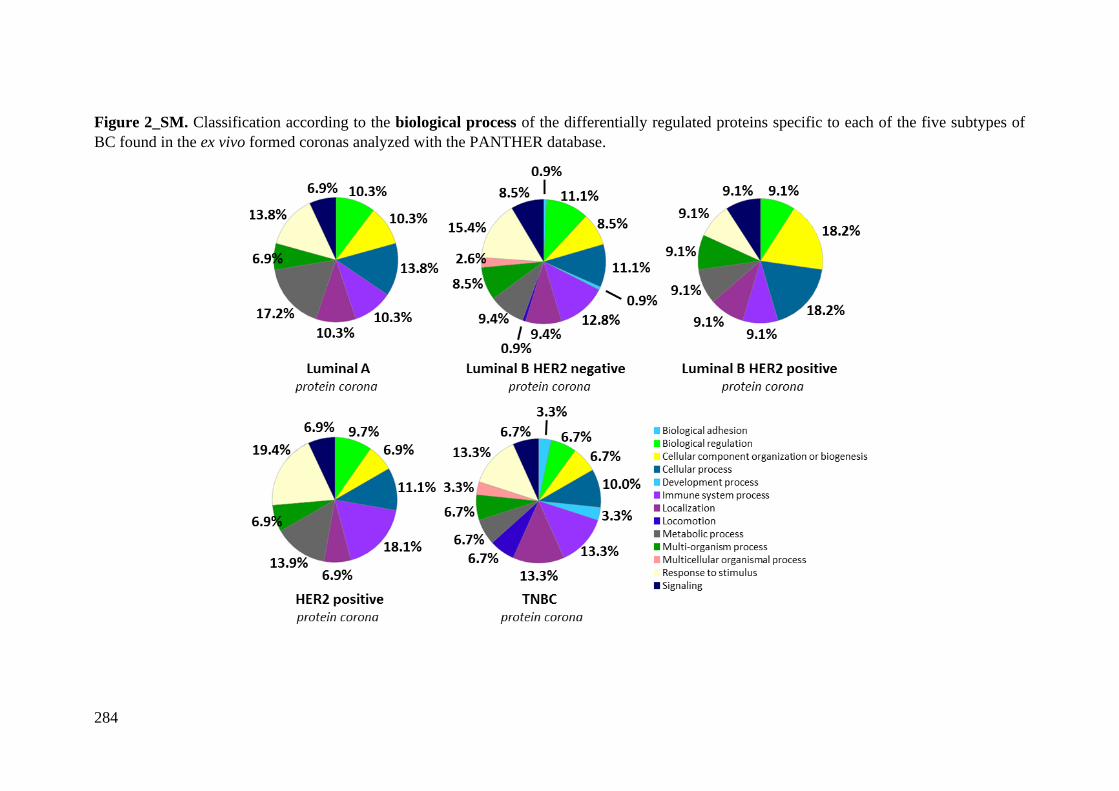
284
Figure 2_SM. Classification according to the biological process of the differentially regulated proteins specific to each of the five subtypes of
BC found in the ex vivo formed coronas analyzed with the PANTHER database.
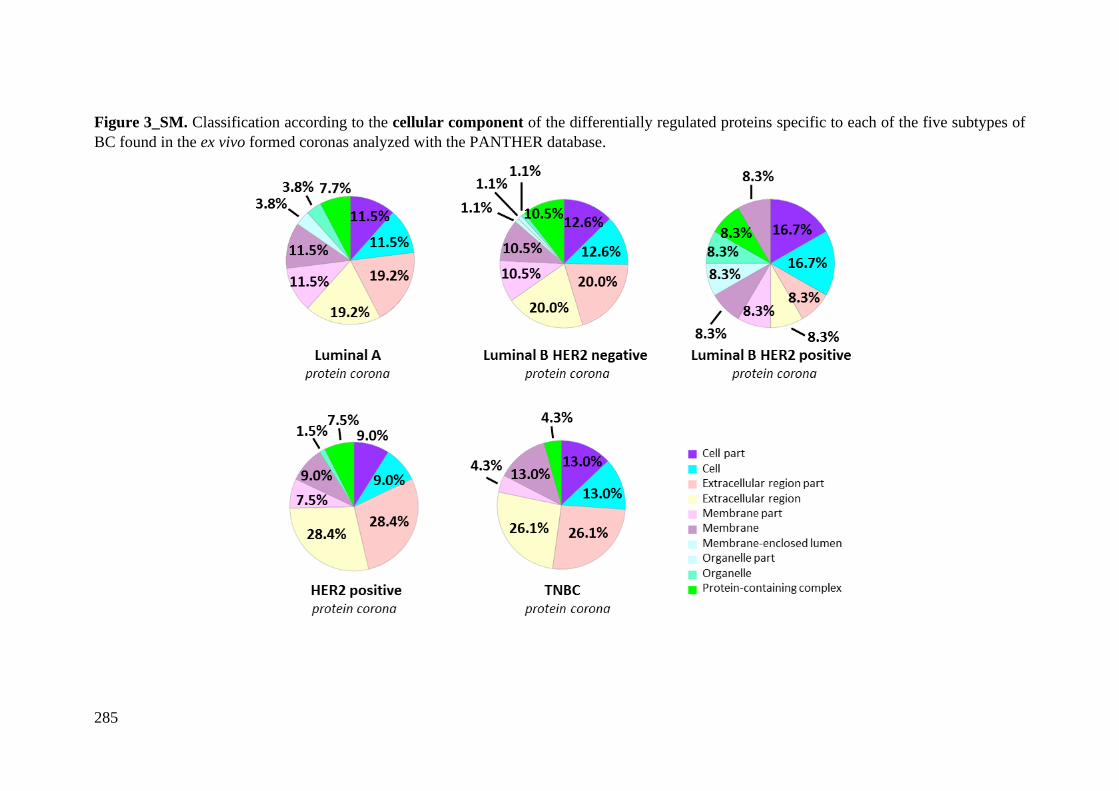
285
Figure 3_SM. Classification according to the cellular component of the differentially regulated proteins specific to each of the five subtypes of
BC found in the ex vivo formed coronas analyzed with the PANTHER database.
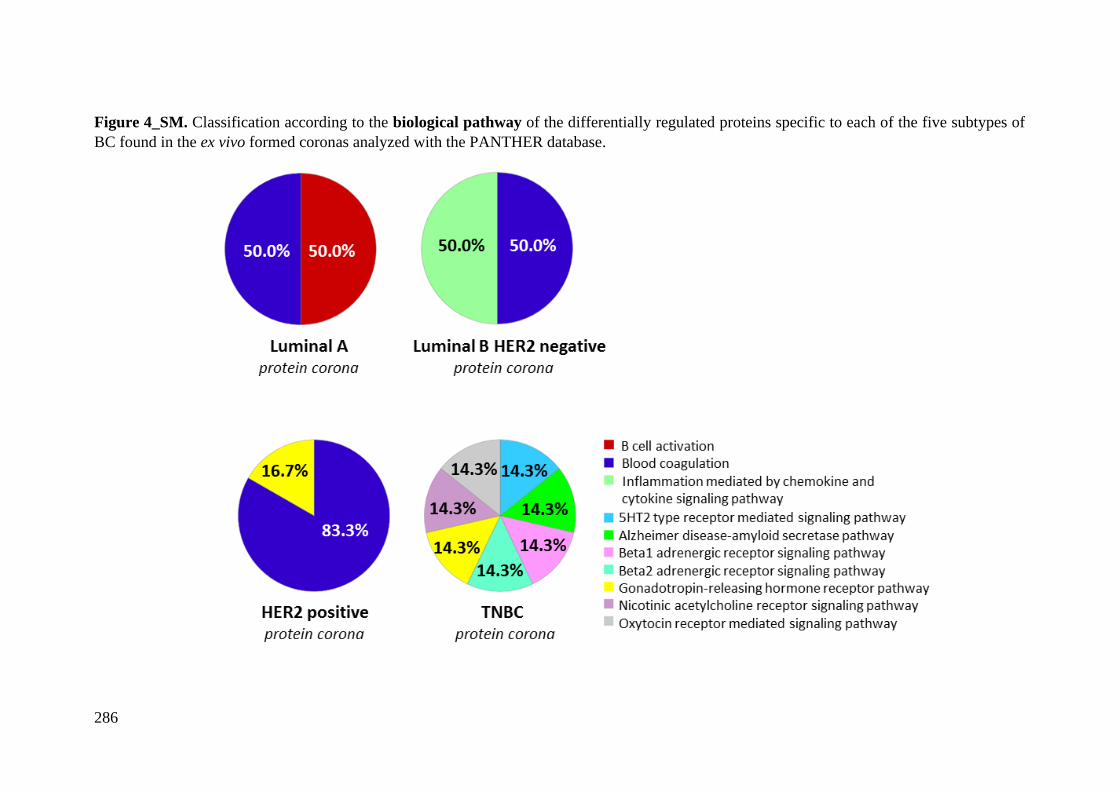
286
Figure 4_SM. Classification according to the biological pathway of the differentially regulated proteins specific to each of the five subtypes of
BC found in the ex vivo formed coronas analyzed with the PANTHER database.
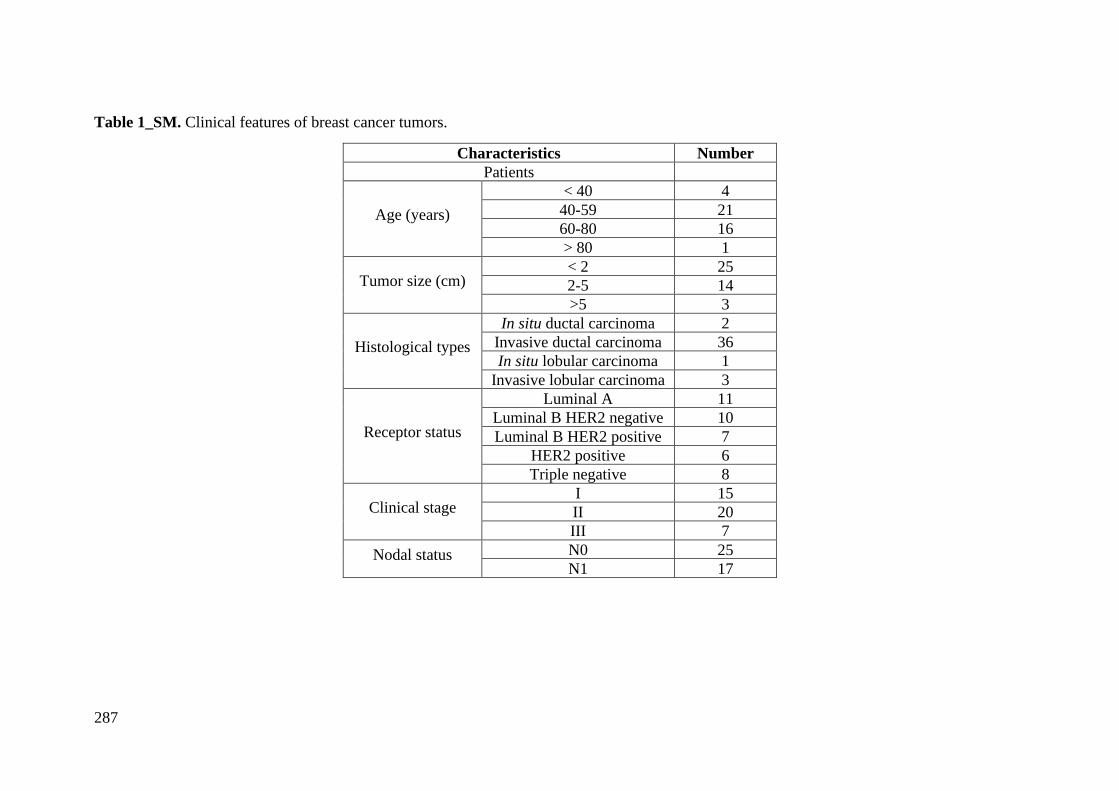
287
Table 1_SM. Clinical features of breast cancer tumors.
Characteristics Number
Patients
Age (years)
< 40 4
40-59 21
60-80 16
> 80 1
Tumor size (cm) < 2 25
2-5 14
>5 3
Histological types
In situ ductal carcinoma 2
Invasive ductal carcinoma 36
In situ lobular carcinoma 1
Invasive lobular carcinoma 3
Receptor status
Luminal A 11
Luminal B HER2 negative 10
Luminal B HER2 positive 7
HER2 positive 6
Triple negative 8
Clinical stage I 15
II 20
III 7
Nodal status N0 25
N1 17
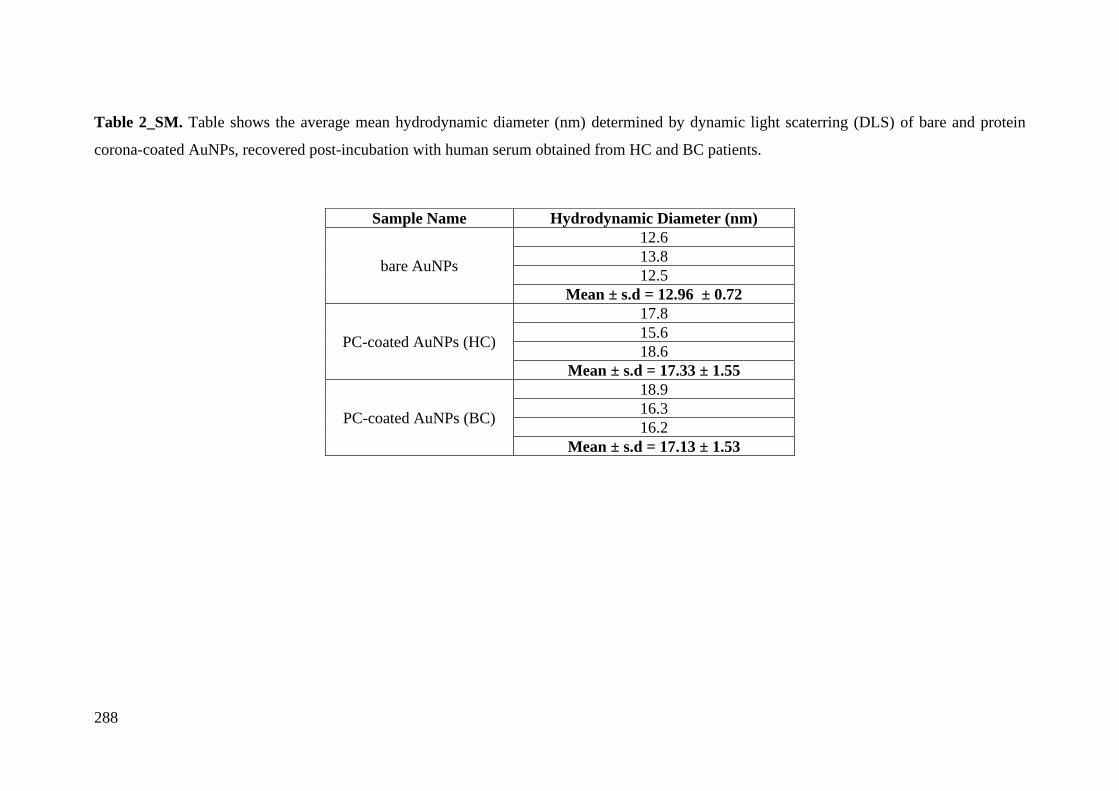
288
Table 2_SM. Table shows the average mean hydrodynamic diameter (nm) determined by dynamic light scaterring (DLS) of bare and protein
corona-coated AuNPs, recovered post-incubation with human serum obtained from HC and BC patients.
Sample Name Hydrodynamic Diameter (nm)
bare AuNPs
12.6
13.8
12.5
Mean ± s.d = 12.96 ± 0.72
PC-coated AuNPs (HC)
17.8
15.6
18.6
Mean ± s.d = 17.33 ± 1.55
PC-coated AuNPs (BC)
18.9
16.3
16.2
Mean ± s.d = 17.13 ± 1.53
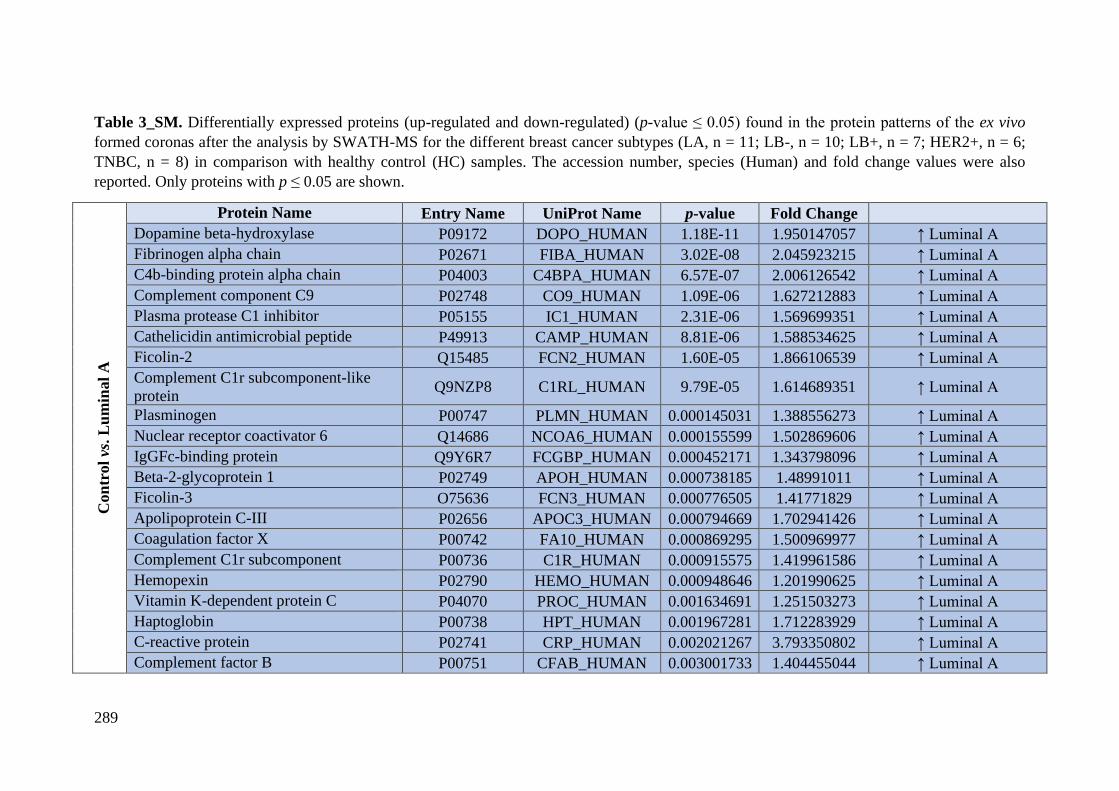
289
Table 3_SM. Differentially expressed proteins (up-regulated and down-regulated) (p-value ≤ 0.05) found in the protein patterns of the ex vivo
formed coronas after the analysis by SWATH-MS for the different breast cancer subtypes (LA, n = 11; LB-, n = 10; LB+, n = 7; HER2+, n = 6;
TNBC, n = 8) in comparison with healthy control (HC) samples. The accession number, species (Human) and fold change values were also
reported. Only proteins with p ≤ 0.05 are shown.
Con
trol vs
. L
um
inal
A
Protein Name Entry Name UniProt Name p-value Fold Change
Dopamine beta-hydroxylase P09172 DOPO_HUMAN 1.18E-11 1.950147057 ↑ Luminal A
Fibrinogen alpha chain P02671 FIBA_HUMAN 3.02E-08 2.045923215 ↑ Luminal A
C4b-binding protein alpha chain P04003 C4BPA_HUMAN 6.57E-07 2.006126542 ↑ Luminal A
Complement component C9 P02748 CO9_HUMAN 1.09E-06 1.627212883 ↑ Luminal A
Plasma protease C1 inhibitor P05155 IC1_HUMAN 2.31E-06 1.569699351 ↑ Luminal A
Cathelicidin antimicrobial peptide P49913 CAMP_HUMAN 8.81E-06 1.588534625 ↑ Luminal A
Ficolin-2 Q15485 FCN2_HUMAN 1.60E-05 1.866106539 ↑ Luminal A
Complement C1r subcomponent-like
protein Q9NZP8 C1RL_HUMAN 9.79E-05 1.614689351 ↑ Luminal A
Plasminogen P00747 PLMN_HUMAN 0.000145031 1.388556273 ↑ Luminal A
Nuclear receptor coactivator 6 Q14686 NCOA6_HUMAN 0.000155599 1.502869606 ↑ Luminal A
IgGFc-binding protein Q9Y6R7 FCGBP_HUMAN 0.000452171 1.343798096 ↑ Luminal A
Beta-2-glycoprotein 1 P02749 APOH_HUMAN 0.000738185 1.48991011 ↑ Luminal A
Ficolin-3 O75636 FCN3_HUMAN 0.000776505 1.41771829 ↑ Luminal A
Apolipoprotein C-III P02656 APOC3_HUMAN 0.000794669 1.702941426 ↑ Luminal A
Coagulation factor X P00742 FA10_HUMAN 0.000869295 1.500969977 ↑ Luminal A
Complement C1r subcomponent P00736 C1R_HUMAN 0.000915575 1.419961586 ↑ Luminal A
Hemopexin P02790 HEMO_HUMAN 0.000948646 1.201990625 ↑ Luminal A
Vitamin K-dependent protein C P04070 PROC_HUMAN 0.001634691 1.251503273 ↑ Luminal A
Haptoglobin P00738 HPT_HUMAN 0.001967281 1.712283929 ↑ Luminal A
C-reactive protein P02741 CRP_HUMAN 0.002021267 3.793350802 ↑ Luminal A
Complement factor B P00751 CFAB_HUMAN 0.003001733 1.404455044 ↑ Luminal A
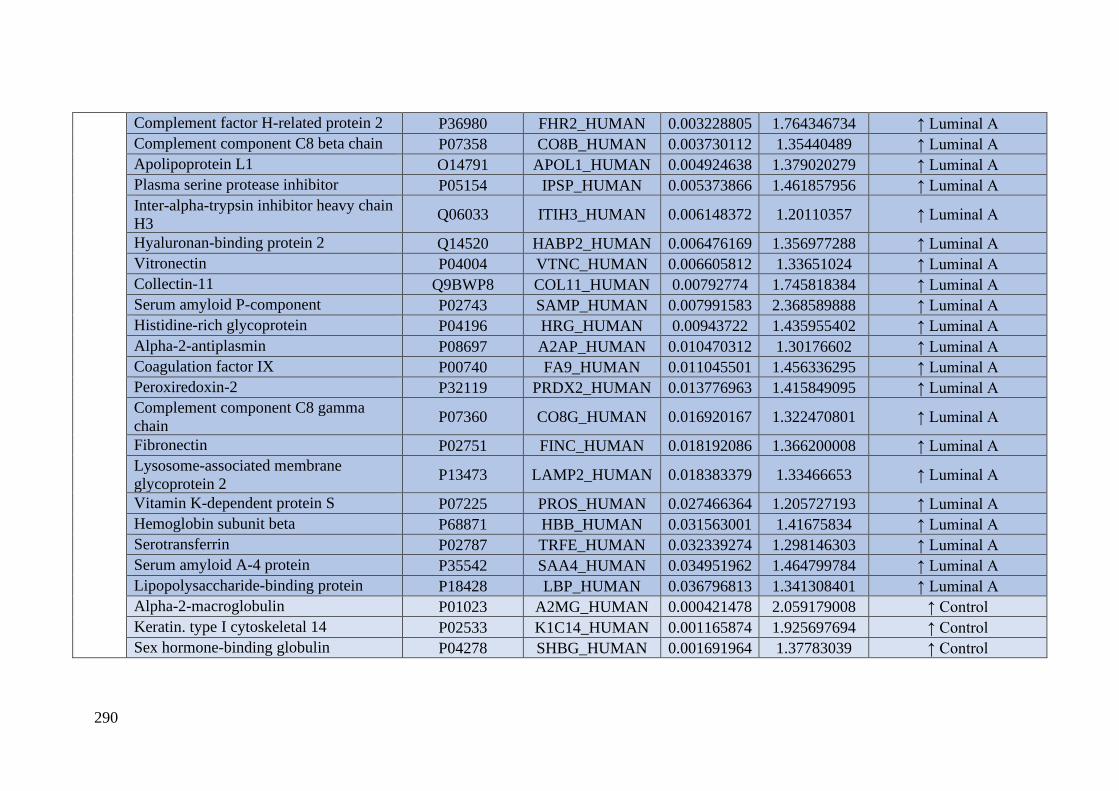
290
Complement factor H-related protein 2 P36980 FHR2_HUMAN 0.003228805 1.764346734 ↑ Luminal A
Complement component C8 beta chain P07358 CO8B_HUMAN 0.003730112 1.35440489 ↑ Luminal A
Apolipoprotein L1 O14791 APOL1_HUMAN 0.004924638 1.379020279 ↑ Luminal A
Plasma serine protease inhibitor P05154 IPSP_HUMAN 0.005373866 1.461857956 ↑ Luminal A
Inter-alpha-trypsin inhibitor heavy chain
H3 Q06033 ITIH3_HUMAN 0.006148372 1.20110357 ↑ Luminal A
Hyaluronan-binding protein 2 Q14520 HABP2_HUMAN 0.006476169 1.356977288 ↑ Luminal A
Vitronectin P04004 VTNC_HUMAN 0.006605812 1.33651024 ↑ Luminal A
Collectin-11 Q9BWP8 COL11_HUMAN 0.00792774 1.745818384 ↑ Luminal A
Serum amyloid P-component P02743 SAMP_HUMAN 0.007991583 2.368589888 ↑ Luminal A
Histidine-rich glycoprotein P04196 HRG_HUMAN 0.00943722 1.435955402 ↑ Luminal A
Alpha-2-antiplasmin P08697 A2AP_HUMAN 0.010470312 1.30176602 ↑ Luminal A
Coagulation factor IX P00740 FA9_HUMAN 0.011045501 1.456336295 ↑ Luminal A
Peroxiredoxin-2 P32119 PRDX2_HUMAN 0.013776963 1.415849095 ↑ Luminal A
Complement component C8 gamma
chain P07360 CO8G_HUMAN 0.016920167 1.322470801 ↑ Luminal A
Fibronectin P02751 FINC_HUMAN 0.018192086 1.366200008 ↑ Luminal A
Lysosome-associated membrane
glycoprotein 2 P13473 LAMP2_HUMAN 0.018383379 1.33466653 ↑ Luminal A
Vitamin K-dependent protein S P07225 PROS_HUMAN 0.027466364 1.205727193 ↑ Luminal A
Hemoglobin subunit beta P68871 HBB_HUMAN 0.031563001 1.41675834 ↑ Luminal A
Serotransferrin P02787 TRFE_HUMAN 0.032339274 1.298146303 ↑ Luminal A
Serum amyloid A-4 protein P35542 SAA4_HUMAN 0.034951962 1.464799784 ↑ Luminal A
Lipopolysaccharide-binding protein P18428 LBP_HUMAN 0.036796813 1.341308401 ↑ Luminal A
Alpha-2-macroglobulin P01023 A2MG_HUMAN 0.000421478 2.059179008 ↑ Control
Keratin. type I cytoskeletal 14 P02533 K1C14_HUMAN 0.001165874 1.925697694 ↑ Control
Sex hormone-binding globulin P04278 SHBG_HUMAN 0.001691964 1.37783039 ↑ Control
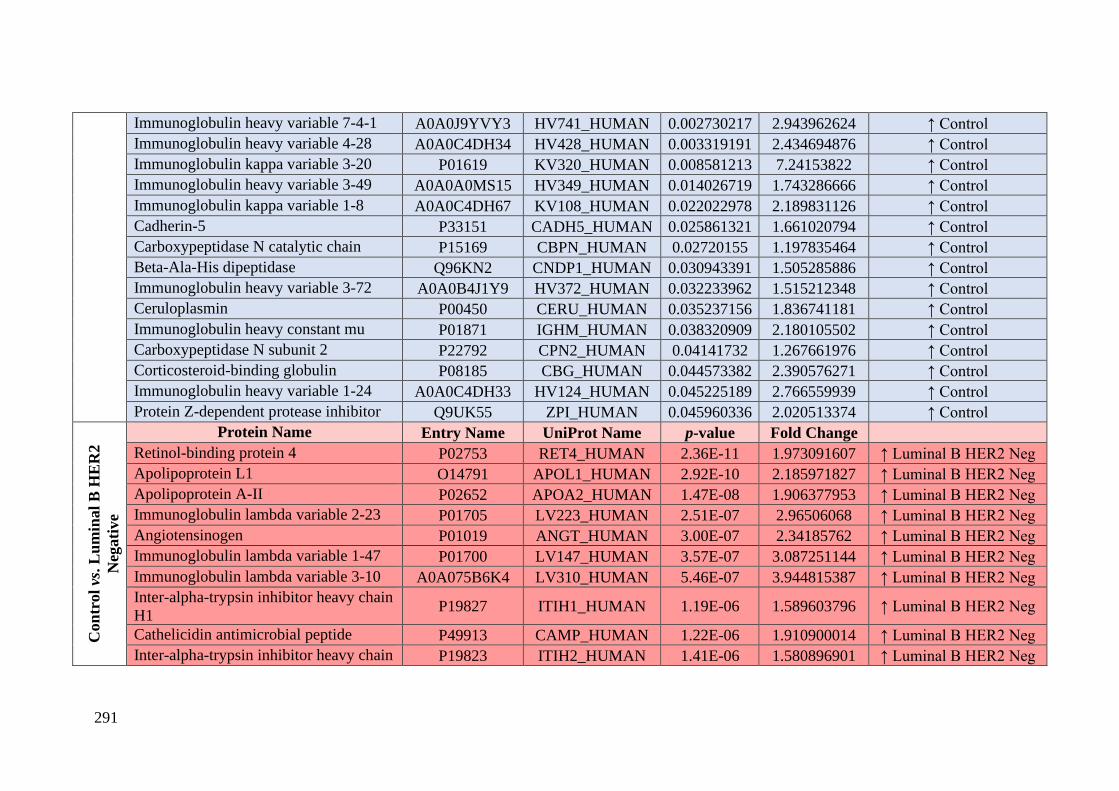
291
Immunoglobulin heavy variable 7-4-1 A0A0J9YVY3 HV741_HUMAN 0.002730217 2.943962624 ↑ Control
Immunoglobulin heavy variable 4-28 A0A0C4DH34 HV428_HUMAN 0.003319191 2.434694876 ↑ Control
Immunoglobulin kappa variable 3-20 P01619 KV320_HUMAN 0.008581213 7.24153822 ↑ Control
Immunoglobulin heavy variable 3-49 A0A0A0MS15 HV349_HUMAN 0.014026719 1.743286666 ↑ Control
Immunoglobulin kappa variable 1-8 A0A0C4DH67 KV108_HUMAN 0.022022978 2.189831126 ↑ Control
Cadherin-5 P33151 CADH5_HUMAN 0.025861321 1.661020794 ↑ Control
Carboxypeptidase N catalytic chain P15169 CBPN_HUMAN 0.02720155 1.197835464 ↑ Control
Beta-Ala-His dipeptidase Q96KN2 CNDP1_HUMAN 0.030943391 1.505285886 ↑ Control
Immunoglobulin heavy variable 3-72 A0A0B4J1Y9 HV372_HUMAN 0.032233962 1.515212348 ↑ Control
Ceruloplasmin P00450 CERU_HUMAN 0.035237156 1.836741181 ↑ Control
Immunoglobulin heavy constant mu P01871 IGHM_HUMAN 0.038320909 2.180105502 ↑ Control
Carboxypeptidase N subunit 2 P22792 CPN2_HUMAN 0.04141732 1.267661976 ↑ Control
Corticosteroid-binding globulin P08185 CBG_HUMAN 0.044573382 2.390576271 ↑ Control
Immunoglobulin heavy variable 1-24 A0A0C4DH33 HV124_HUMAN 0.045225189 2.766559939 ↑ Control
Protein Z-dependent protease inhibitor Q9UK55 ZPI_HUMAN 0.045960336 2.020513374 ↑ Control
Con
trol vs
. L
um
inal
B H
ER
2
Neg
ati
ve
Protein Name Entry Name UniProt Name p-value Fold Change
Retinol-binding protein 4 P02753 RET4_HUMAN 2.36E-11 1.973091607 ↑ Luminal B HER2 Neg
Apolipoprotein L1 O14791 APOL1_HUMAN 2.92E-10 2.185971827 ↑ Luminal B HER2 Neg
Apolipoprotein A-II P02652 APOA2_HUMAN 1.47E-08 1.906377953 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 2-23 P01705 LV223_HUMAN 2.51E-07 2.96506068 ↑ Luminal B HER2 Neg
Angiotensinogen P01019 ANGT_HUMAN 3.00E-07 2.34185762 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 1-47 P01700 LV147_HUMAN 3.57E-07 3.087251144 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 3-10 A0A075B6K4 LV310_HUMAN 5.46E-07 3.944815387 ↑ Luminal B HER2 Neg
Inter-alpha-trypsin inhibitor heavy chain
H1 P19827 ITIH1_HUMAN 1.19E-06 1.589603796 ↑ Luminal B HER2 Neg
Cathelicidin antimicrobial peptide P49913 CAMP_HUMAN 1.22E-06 1.910900014 ↑ Luminal B HER2 Neg
Inter-alpha-trypsin inhibitor heavy chain P19823 ITIH2_HUMAN 1.41E-06 1.580896901 ↑ Luminal B HER2 Neg
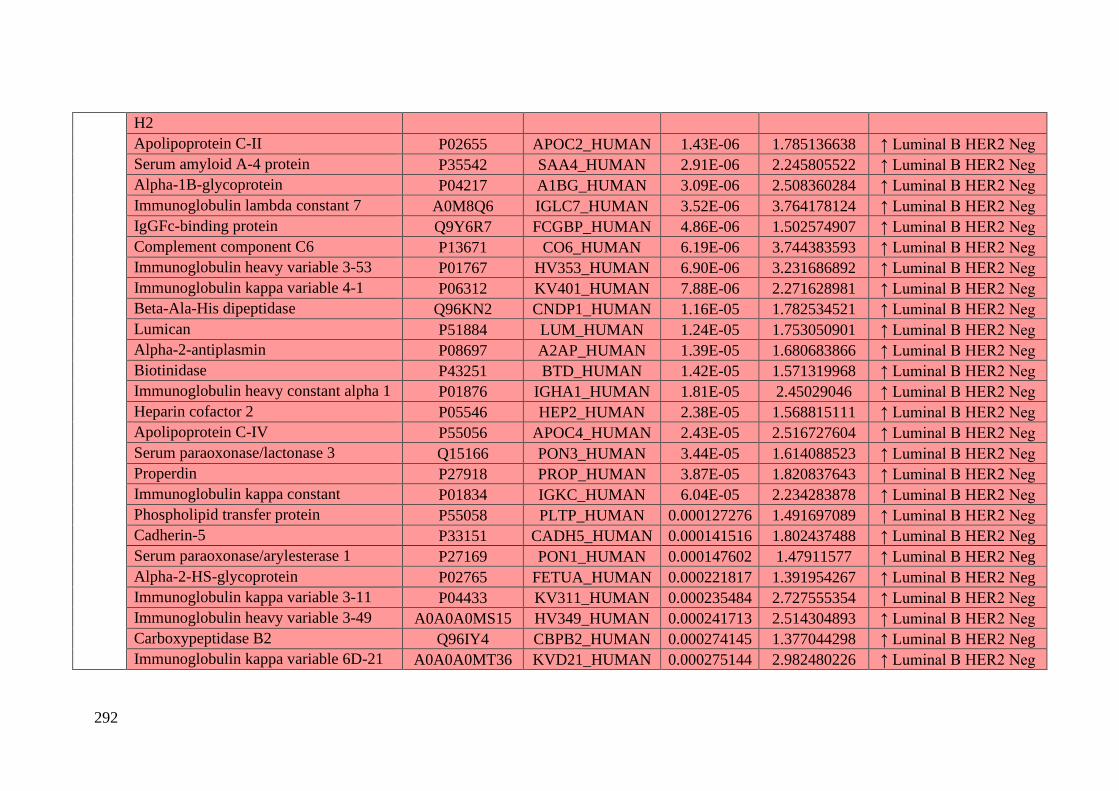
292
H2
Apolipoprotein C-II P02655 APOC2_HUMAN 1.43E-06 1.785136638 ↑ Luminal B HER2 Neg
Serum amyloid A-4 protein P35542 SAA4_HUMAN 2.91E-06 2.245805522 ↑ Luminal B HER2 Neg
Alpha-1B-glycoprotein P04217 A1BG_HUMAN 3.09E-06 2.508360284 ↑ Luminal B HER2 Neg
Immunoglobulin lambda constant 7 A0M8Q6 IGLC7_HUMAN 3.52E-06 3.764178124 ↑ Luminal B HER2 Neg
IgGFc-binding protein Q9Y6R7 FCGBP_HUMAN 4.86E-06 1.502574907 ↑ Luminal B HER2 Neg
Complement component C6 P13671 CO6_HUMAN 6.19E-06 3.744383593 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 3-53 P01767 HV353_HUMAN 6.90E-06 3.231686892 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 4-1 P06312 KV401_HUMAN 7.88E-06 2.271628981 ↑ Luminal B HER2 Neg
Beta-Ala-His dipeptidase Q96KN2 CNDP1_HUMAN 1.16E-05 1.782534521 ↑ Luminal B HER2 Neg
Lumican P51884 LUM_HUMAN 1.24E-05 1.753050901 ↑ Luminal B HER2 Neg
Alpha-2-antiplasmin P08697 A2AP_HUMAN 1.39E-05 1.680683866 ↑ Luminal B HER2 Neg
Biotinidase P43251 BTD_HUMAN 1.42E-05 1.571319968 ↑ Luminal B HER2 Neg
Immunoglobulin heavy constant alpha 1 P01876 IGHA1_HUMAN 1.81E-05 2.45029046 ↑ Luminal B HER2 Neg
Heparin cofactor 2 P05546 HEP2_HUMAN 2.38E-05 1.568815111 ↑ Luminal B HER2 Neg
Apolipoprotein C-IV P55056 APOC4_HUMAN 2.43E-05 2.516727604 ↑ Luminal B HER2 Neg
Serum paraoxonase/lactonase 3 Q15166 PON3_HUMAN 3.44E-05 1.614088523 ↑ Luminal B HER2 Neg
Properdin P27918 PROP_HUMAN 3.87E-05 1.820837643 ↑ Luminal B HER2 Neg
Immunoglobulin kappa constant P01834 IGKC_HUMAN 6.04E-05 2.234283878 ↑ Luminal B HER2 Neg
Phospholipid transfer protein P55058 PLTP_HUMAN 0.000127276 1.491697089 ↑ Luminal B HER2 Neg
Cadherin-5 P33151 CADH5_HUMAN 0.000141516 1.802437488 ↑ Luminal B HER2 Neg
Serum paraoxonase/arylesterase 1 P27169 PON1_HUMAN 0.000147602 1.47911577 ↑ Luminal B HER2 Neg
Alpha-2-HS-glycoprotein P02765 FETUA_HUMAN 0.000221817 1.391954267 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 3-11 P04433 KV311_HUMAN 0.000235484 2.727555354 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 3-49 A0A0A0MS15 HV349_HUMAN 0.000241713 2.514304893 ↑ Luminal B HER2 Neg
Carboxypeptidase B2 Q96IY4 CBPB2_HUMAN 0.000274145 1.377044298 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 6D-21 A0A0A0MT36 KVD21_HUMAN 0.000275144 2.982480226 ↑ Luminal B HER2 Neg

293
Complement C4-A P0C0L4 CO4A_HUMAN 0.000311066 1.582264414 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 3-9 P01782 HV309_HUMAN 0.000418498 2.709287474 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 4-31 P0DP07 HV431_HUMAN 0.000447522 4.063279859 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 3-9 A0A075B6K5 LV39_HUMAN 0.000527457 2.179105185 ↑ Luminal B HER2 Neg
Tetranectin P05452 TETN_HUMAN 0.000529868 1.375681621 ↑ Luminal B HER2 Neg
Alpha-mannosidase 2 Q16706 MA2A1_HUMAN 0.000577191 2.094925838 ↑ Luminal B HER2 Neg
Peroxiredoxin-2 P32119 PRDX2_HUMAN 0.000663743 1.658367127 ↑ Luminal B HER2 Neg
Cholinesterase P06276 CHLE_HUMAN 0.000739103 2.443938708 ↑ Luminal B HER2 Neg
Immunoglobulin heavy constant gamma
1 P01857 IGHG1_HUMAN 0.000838666 1.832731289 ↑ Luminal B HER2 Neg
Galectin-3-binding protein Q08380 LG3BP_HUMAN 0.000924525 1.669562645 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 1-40 P01703 LV140_HUMAN 0.00099044 2.058275359 ↑ Luminal B HER2 Neg
Kallistatin P29622 KAIN_HUMAN 0.001372229 1.767108231 ↑ Luminal B HER2 Neg
Alpha-1-acid glycoprotein 1 P02763 A1AG1_HUMAN 0.001636216 2.215227977 ↑ Luminal B HER2 Neg
Apolipoprotein B-100 P04114 APOB_HUMAN 0.001926757 1.618943103 ↑ Luminal B HER2 Neg
Immunoglobulin heavy constant alpha 2 P01877 IGHA2_HUMAN 0.00207989 2.137794634 ↑ Luminal B HER2 Neg
Basement membrane-specific heparan
sulfate proteoglycan core protein P98160 PGBM_HUMAN 0.002185804 2.617271104 ↑ Luminal B HER2 Neg
Cholesteryl ester transfer protein P11597 CETP_HUMAN 0.002360349 1.663599851 ↑ Luminal B HER2 Neg
Phosphatidylcholine-sterol
acyltransferase P04180 LCAT_HUMAN 0.002466447 2.154281745 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 1-16 P04430 KV116_HUMAN 0.002548994 2.530617013 ↑ Luminal B HER2 Neg
Pregnancy zone protein P20742 PZP_HUMAN 0.002663618 4.343426266 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 1-46 P01743 HV146_HUMAN 0.00286485 3.896313023 ↑ Luminal B HER2 Neg
Apolipoprotein A-I P02647 APOA1_HUMAN 0.002969507 1.292738115 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 3-19 P01714 LV319_HUMAN 0.003020851 1.830912469 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 1-69D A0A0B4J2H0 HV69D_HUMAN 0.00307313 2.762019962 ↑ Luminal B HER2 Neg
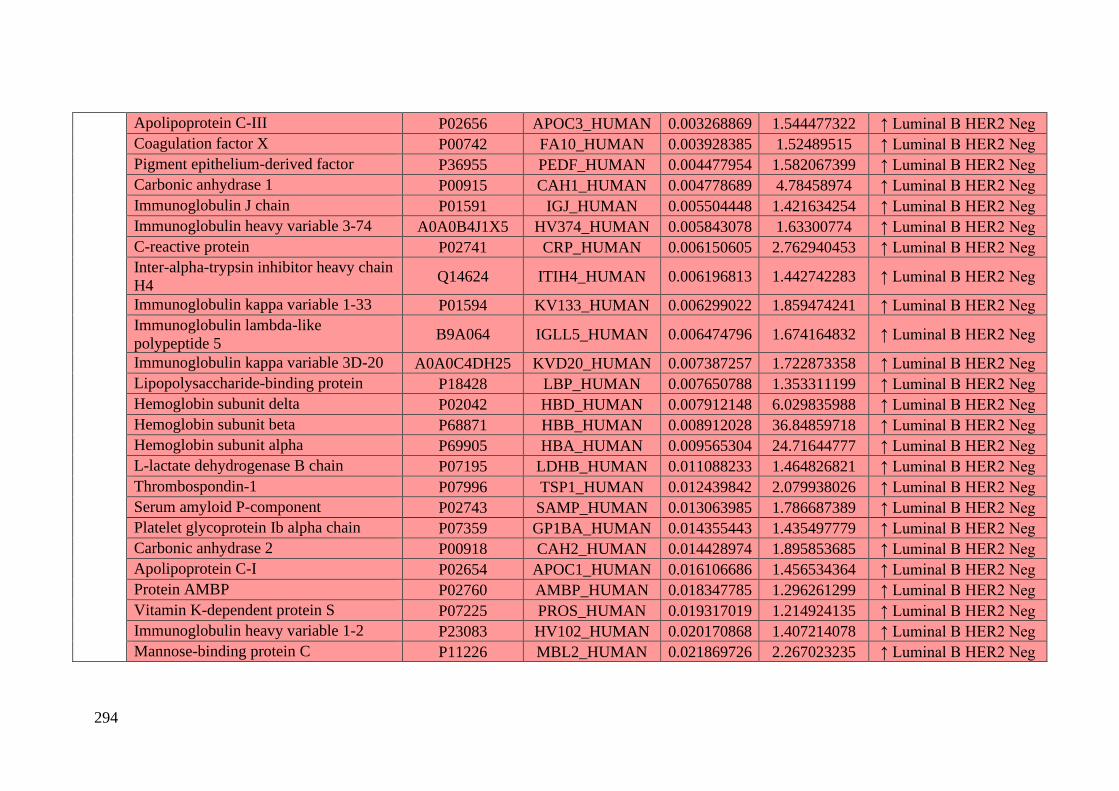
294
Apolipoprotein C-III P02656 APOC3_HUMAN 0.003268869 1.544477322 ↑ Luminal B HER2 Neg
Coagulation factor X P00742 FA10_HUMAN 0.003928385 1.52489515 ↑ Luminal B HER2 Neg
Pigment epithelium-derived factor P36955 PEDF_HUMAN 0.004477954 1.582067399 ↑ Luminal B HER2 Neg
Carbonic anhydrase 1 P00915 CAH1_HUMAN 0.004778689 4.78458974 ↑ Luminal B HER2 Neg
Immunoglobulin J chain P01591 IGJ_HUMAN 0.005504448 1.421634254 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 3-74 A0A0B4J1X5 HV374_HUMAN 0.005843078 1.63300774 ↑ Luminal B HER2 Neg
C-reactive protein P02741 CRP_HUMAN 0.006150605 2.762940453 ↑ Luminal B HER2 Neg
Inter-alpha-trypsin inhibitor heavy chain
H4 Q14624 ITIH4_HUMAN 0.006196813 1.442742283 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 1-33 P01594 KV133_HUMAN 0.006299022 1.859474241 ↑ Luminal B HER2 Neg
Immunoglobulin lambda-like
polypeptide 5 B9A064 IGLL5_HUMAN 0.006474796 1.674164832 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 3D-20 A0A0C4DH25 KVD20_HUMAN 0.007387257 1.722873358 ↑ Luminal B HER2 Neg
Lipopolysaccharide-binding protein P18428 LBP_HUMAN 0.007650788 1.353311199 ↑ Luminal B HER2 Neg
Hemoglobin subunit delta P02042 HBD_HUMAN 0.007912148 6.029835988 ↑ Luminal B HER2 Neg
Hemoglobin subunit beta P68871 HBB_HUMAN 0.008912028 36.84859718 ↑ Luminal B HER2 Neg
Hemoglobin subunit alpha P69905 HBA_HUMAN 0.009565304 24.71644777 ↑ Luminal B HER2 Neg
L-lactate dehydrogenase B chain P07195 LDHB_HUMAN 0.011088233 1.464826821 ↑ Luminal B HER2 Neg
Thrombospondin-1 P07996 TSP1_HUMAN 0.012439842 2.079938026 ↑ Luminal B HER2 Neg
Serum amyloid P-component P02743 SAMP_HUMAN 0.013063985 1.786687389 ↑ Luminal B HER2 Neg
Platelet glycoprotein Ib alpha chain P07359 GP1BA_HUMAN 0.014355443 1.435497779 ↑ Luminal B HER2 Neg
Carbonic anhydrase 2 P00918 CAH2_HUMAN 0.014428974 1.895853685 ↑ Luminal B HER2 Neg
Apolipoprotein C-I P02654 APOC1_HUMAN 0.016106686 1.456534364 ↑ Luminal B HER2 Neg
Protein AMBP P02760 AMBP_HUMAN 0.018347785 1.296261299 ↑ Luminal B HER2 Neg
Vitamin K-dependent protein S P07225 PROS_HUMAN 0.019317019 1.214924135 ↑ Luminal B HER2 Neg
Immunoglobulin heavy variable 1-2 P23083 HV102_HUMAN 0.020170868 1.407214078 ↑ Luminal B HER2 Neg
Mannose-binding protein C P11226 MBL2_HUMAN 0.021869726 2.267023235 ↑ Luminal B HER2 Neg
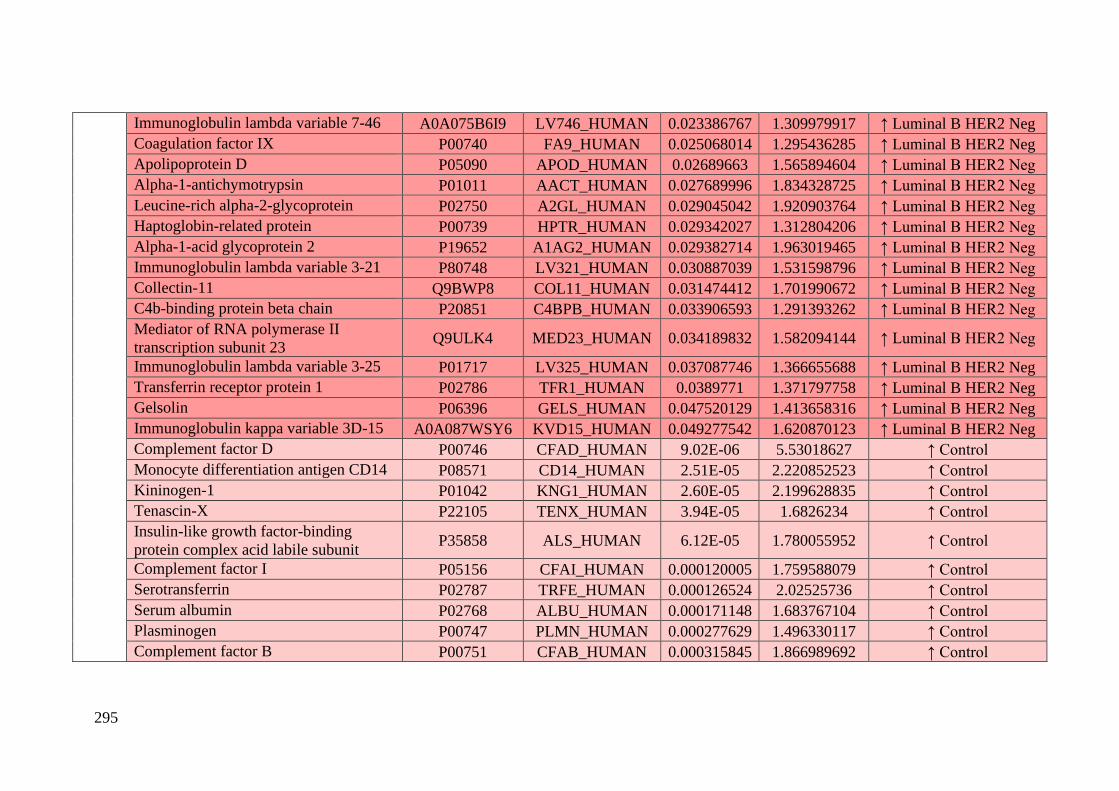
295
Immunoglobulin lambda variable 7-46 A0A075B6I9 LV746_HUMAN 0.023386767 1.309979917 ↑ Luminal B HER2 Neg
Coagulation factor IX P00740 FA9_HUMAN 0.025068014 1.295436285 ↑ Luminal B HER2 Neg
Apolipoprotein D P05090 APOD_HUMAN 0.02689663 1.565894604 ↑ Luminal B HER2 Neg
Alpha-1-antichymotrypsin P01011 AACT_HUMAN 0.027689996 1.834328725 ↑ Luminal B HER2 Neg
Leucine-rich alpha-2-glycoprotein P02750 A2GL_HUMAN 0.029045042 1.920903764 ↑ Luminal B HER2 Neg
Haptoglobin-related protein P00739 HPTR_HUMAN 0.029342027 1.312804206 ↑ Luminal B HER2 Neg
Alpha-1-acid glycoprotein 2 P19652 A1AG2_HUMAN 0.029382714 1.963019465 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 3-21 P80748 LV321_HUMAN 0.030887039 1.531598796 ↑ Luminal B HER2 Neg
Collectin-11 Q9BWP8 COL11_HUMAN 0.031474412 1.701990672 ↑ Luminal B HER2 Neg
C4b-binding protein beta chain P20851 C4BPB_HUMAN 0.033906593 1.291393262 ↑ Luminal B HER2 Neg
Mediator of RNA polymerase II
transcription subunit 23 Q9ULK4 MED23_HUMAN 0.034189832 1.582094144 ↑ Luminal B HER2 Neg
Immunoglobulin lambda variable 3-25 P01717 LV325_HUMAN 0.037087746 1.366655688 ↑ Luminal B HER2 Neg
Transferrin receptor protein 1 P02786 TFR1_HUMAN 0.0389771 1.371797758 ↑ Luminal B HER2 Neg
Gelsolin P06396 GELS_HUMAN 0.047520129 1.413658316 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 3D-15 A0A087WSY6 KVD15_HUMAN 0.049277542 1.620870123 ↑ Luminal B HER2 Neg
Complement factor D P00746 CFAD_HUMAN 9.02E-06 5.53018627 ↑ Control
Monocyte differentiation antigen CD14 P08571 CD14_HUMAN 2.51E-05 2.220852523 ↑ Control
Kininogen-1 P01042 KNG1_HUMAN 2.60E-05 2.199628835 ↑ Control
Tenascin-X P22105 TENX_HUMAN 3.94E-05 1.6826234 ↑ Control
Insulin-like growth factor-binding
protein complex acid labile subunit P35858 ALS_HUMAN 6.12E-05 1.780055952 ↑ Control
Complement factor I P05156 CFAI_HUMAN 0.000120005 1.759588079 ↑ Control
Serotransferrin P02787 TRFE_HUMAN 0.000126524 2.02525736 ↑ Control
Serum albumin P02768 ALBU_HUMAN 0.000171148 1.683767104 ↑ Control
Plasminogen P00747 PLMN_HUMAN 0.000277629 1.496330117 ↑ Control
Complement factor B P00751 CFAB_HUMAN 0.000315845 1.866989692 ↑ Control
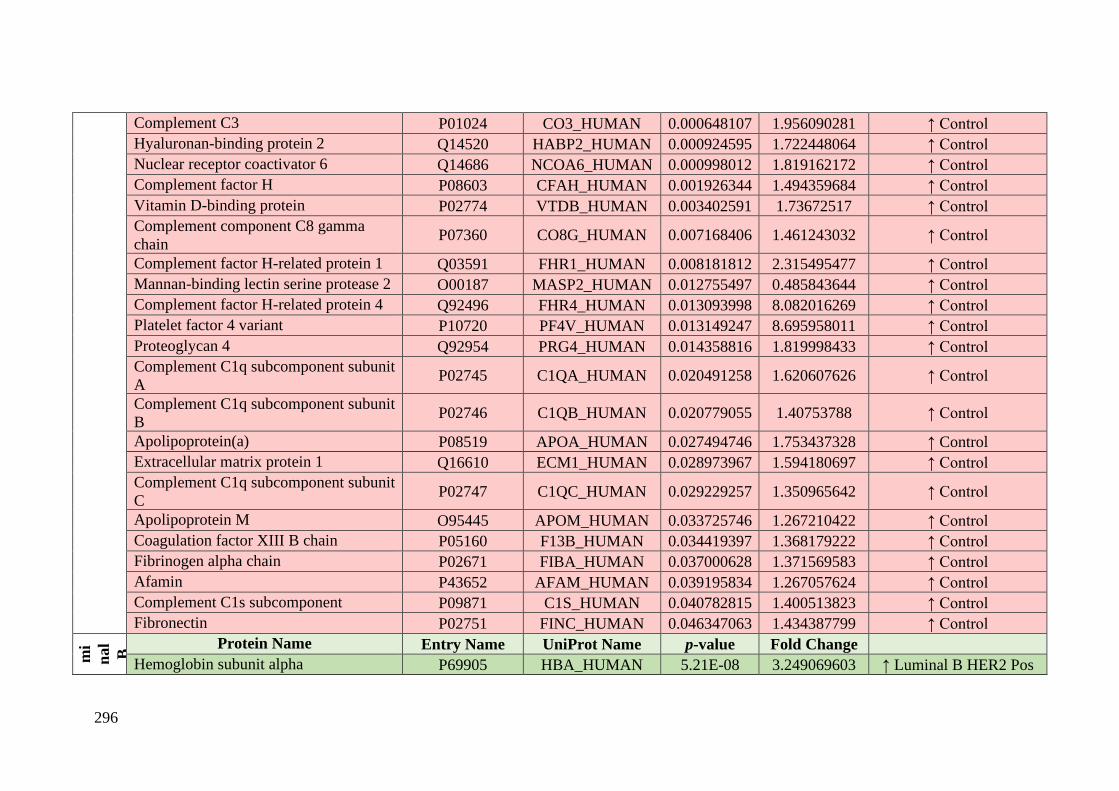
296
Complement C3 P01024 CO3_HUMAN 0.000648107 1.956090281 ↑ Control
Hyaluronan-binding protein 2 Q14520 HABP2_HUMAN 0.000924595 1.722448064 ↑ Control
Nuclear receptor coactivator 6 Q14686 NCOA6_HUMAN 0.000998012 1.819162172 ↑ Control
Complement factor H P08603 CFAH_HUMAN 0.001926344 1.494359684 ↑ Control
Vitamin D-binding protein P02774 VTDB_HUMAN 0.003402591 1.73672517 ↑ Control
Complement component C8 gamma
chain P07360 CO8G_HUMAN 0.007168406 1.461243032 ↑ Control
Complement factor H-related protein 1 Q03591 FHR1_HUMAN 0.008181812 2.315495477 ↑ Control
Mannan-binding lectin serine protease 2 O00187 MASP2_HUMAN 0.012755497 0.485843644 ↑ Control
Complement factor H-related protein 4 Q92496 FHR4_HUMAN 0.013093998 8.082016269 ↑ Control
Platelet factor 4 variant P10720 PF4V_HUMAN 0.013149247 8.695958011 ↑ Control
Proteoglycan 4 Q92954 PRG4_HUMAN 0.014358816 1.819998433 ↑ Control
Complement C1q subcomponent subunit
A P02745 C1QA_HUMAN 0.020491258 1.620607626 ↑ Control
Complement C1q subcomponent subunit
B P02746 C1QB_HUMAN 0.020779055 1.40753788 ↑ Control
Apolipoprotein(a) P08519 APOA_HUMAN 0.027494746 1.753437328 ↑ Control
Extracellular matrix protein 1 Q16610 ECM1_HUMAN 0.028973967 1.594180697 ↑ Control
Complement C1q subcomponent subunit
C P02747 C1QC_HUMAN 0.029229257 1.350965642 ↑ Control
Apolipoprotein M O95445 APOM_HUMAN 0.033725746 1.267210422 ↑ Control
Coagulation factor XIII B chain P05160 F13B_HUMAN 0.034419397 1.368179222 ↑ Control
Fibrinogen alpha chain P02671 FIBA_HUMAN 0.037000628 1.371569583 ↑ Control
Afamin P43652 AFAM_HUMAN 0.039195834 1.267057624 ↑ Control
Complement C1s subcomponent P09871 C1S_HUMAN 0.040782815 1.400513823 ↑ Control
Fibronectin P02751 FINC_HUMAN 0.046347063 1.434387799 ↑ Control
Co
ntr ol
vs.
Lu
mi
nal
B
HE
R2
Pos
itiv e
Protein Name Entry Name UniProt Name p-value Fold Change
Hemoglobin subunit alpha P69905 HBA_HUMAN 5.21E-08 3.249069603 ↑ Luminal B HER2 Pos

297
Hemoglobin subunit beta P68871 HBB_HUMAN 4.10E-07 3.803126189 ↑ Luminal B HER2 Pos
Pigment epithelium-derived factor P36955 PEDF_HUMAN 5.54E-07 3.00054601 ↑ Luminal B HER2 Pos
Immunoglobulin heavy variable 3-49 A0A0A0MS15 HV349_HUMAN 1.97E-06 2.863927632 ↑ Luminal B HER2 Pos
Apolipoprotein C-I P02654 APOC1_HUMAN 2.13E-05 2.739130492 ↑ Luminal B HER2 Pos
Hemoglobin subunit delta P02042 HBD_HUMAN 2.67E-05 3.306697207 ↑ Luminal B HER2 Pos
Immunoglobulin kappa variable 6D-21 A0A0A0MT36 KVD21_HUMAN 2.68E-05 3.798498309 ↑ Luminal B HER2 Pos
Apolipoprotein C-II P02655 APOC2_HUMAN 4.39E-05 1.846194109 ↑ Luminal B HER2 Pos
Immunoglobulin lambda variable 1-47 P01700 LV147_HUMAN 5.77E-05 2.850504061 ↑ Luminal B HER2 Pos
Serum amyloid A-4 protein P35542 SAA4_HUMAN 6.36E-05 2.160815047 ↑ Luminal B HER2 Pos
Immunoglobulin heavy variable 4-31 P0DP07 HV431_HUMAN 7.03E-05 4.89656649 ↑ Luminal B HER2 Pos
Apolipoprotein L1 O14791 APOL1_HUMAN 8.28E-05 1.900750537 ↑ Luminal B HER2 Pos
C4b-binding protein beta chain P20851 C4BPB_HUMAN 0.00010547 2.155401464 ↑ Luminal B HER2 Pos
Immunoglobulin heavy variable 4-28 A0A0C4DH34 HV428_HUMAN 0.000139308 2.192608342 ↑ Luminal B HER2 Pos
Serum amyloid P-component P02743 SAMP_HUMAN 0.000181209 2.454265354 ↑ Luminal B HER2 Pos
Immunoglobulin lambda constant 7 A0M8Q6 IGLC7_HUMAN 0.000665214 3.191779846 ↑ Luminal B HER2 Pos
Gelsolin P06396 GELS_HUMAN 0.000974323 1.951968831 ↑ Luminal B HER2 Pos
Apolipoprotein A-IV P06727 APOA4_HUMAN 0.001025945 1.544141914 ↑ Luminal B HER2 Pos
Apolipoprotein A-II P02652 APOA2_HUMAN 0.001171293 1.776849765 ↑ Luminal B HER2 Pos
Plastin-2 P13796 PLSL_HUMAN 0.002018957 4.501473239 ↑ Luminal B HER2 Pos
Kallistatin P29622 KAIN_HUMAN 0.002130037 1.810271897 ↑ Luminal B HER2 Pos
Polymeric immunoglobulin receptor P01833 PIGR_HUMAN 0.002912228 1.841450361 ↑ Luminal B HER2 Pos
Apolipoprotein C-III P02656 APOC3_HUMAN 0.003035205 2.024844997 ↑ Luminal B HER2 Pos
Immunoglobulin lambda variable 3-10 A0A075B6K4 LV310_HUMAN 0.003668828 3.284821215 ↑ Luminal B HER2 Pos
Alpha-1-acid glycoprotein 1 P02763 A1AG1_HUMAN 0.005475644 2.208206222 ↑ Luminal B HER2 Pos
Galectin-3-binding protein Q08380 LG3BP_HUMAN 0.006686721 1.692847346 ↑ Luminal B HER2 Pos
Complement component C9 P02748 CO9_HUMAN 0.006887579 1.41639914 ↑ Luminal B HER2 Pos
Immunoglobulin lambda variable 3-19 P01714 LV319_HUMAN 0.007858626 1.985710229 ↑ Luminal B HER2 Pos

298
Immunoglobulin kappa variable 1-13 P0DP09 KV113_HUMAN 0.008612844 2.555407574 ↑ Luminal B HER2 Pos
C-reactive protein P02741 CRP_HUMAN 0.008678988 3.370377857 ↑ Luminal B HER2 Pos
Complement C4-A P0C0L4 CO4A_HUMAN 0.010923185 1.488032623 ↑ Luminal B HER2 Pos
Corticosteroid-binding globulin P08185 CBG_HUMAN 0.011580639 1.994591255 ↑ Luminal B HER2 Pos
Xaa-Pro dipeptidase P12955 PEPD_HUMAN 0.012031361 2.165189136 ↑ Luminal B HER2 Pos
Immunoglobulin lambda variable 2-11 P01706 LV211_HUMAN 0.01295552 1.570173745 ↑ Luminal B HER2 Pos
Leucine-rich alpha-2-glycoprotein P02750 A2GL_HUMAN 0.013102391 2.443411846 ↑ Luminal B HER2 Pos
Keratin. type II cytoskeletal 2 epidermal P35908 K22E_HUMAN 0.01347707 1.499797459 ↑ Luminal B HER2 Pos
Complement component C6 P13671 CO6_HUMAN 0.013979583 2.568266773 ↑ Luminal B HER2 Pos
Cholinesterase P06276 CHLE_HUMAN 0.015593089 2.140328724 ↑ Luminal B HER2 Pos
Immunoglobulin heavy variable 4-61 A0A0C4DH41 HV461_HUMAN 0.015895648 2.015912108 ↑ Luminal B HER2 Pos
Transthyretin P02766 TTHY_HUMAN 0.015961518 1.320554649 ↑ Luminal B HER2 Pos
Immunoglobulin lambda variable 1-40 P01703 LV140_HUMAN 0.016785314 1.847853806 ↑ Luminal B HER2 Pos
Immunoglobulin lambda variable 3-9 A0A075B6K5 LV39_HUMAN 0.017837178 1.884838259 ↑ Luminal B HER2 Pos
Complement component C8 alpha chain P07357 CO8A_HUMAN 0.01851134 1.790235827 ↑ Luminal B HER2 Pos
Immunoglobulin heavy variable 2-70D A0A0C4DH43 HV70D_HUMAN 0.019383958 2.403651553 ↑ Luminal B HER2 Pos
Plasma protease C1 inhibitor P05155 IC1_HUMAN 0.0196862 1.276319883 ↑ Luminal B HER2 Pos
Immunoglobulin heavy variable 6-1 A0A0B4J1U7 HV601_HUMAN 0.020191442 1.539338405 ↑ Luminal B HER2 Pos
Alpha-1-antichymotrypsin P01011 AACT_HUMAN 0.026362641 2.102965709 ↑ Luminal B HER2 Pos
Transferrin receptor protein 1 P02786 TFR1_HUMAN 0.02753498 1.498502068 ↑ Luminal B HER2 Pos
Immunoglobulin lambda variable 3-25 P01717 LV325_HUMAN 0.028817061 1.44474454 ↑ Luminal B HER2 Pos
Serum paraoxonase/arylesterase 1 P27169 PON1_HUMAN 0.029928679 1.297670132 ↑ Luminal B HER2 Pos
Immunoglobulin kappa variable 1-33 P01594 KV133_HUMAN 0.030577751 1.791455428 ↑ Luminal B HER2 Pos
Prenylcysteine oxidase 1 Q9UHG3 PCYOX_HUMAN 0.030672346 1.304466122 ↑ Luminal B HER2 Pos
Vitronectin P04004 VTNC_HUMAN 0.035879886 1.313506051 ↑ Luminal B HER2 Pos
Immunoglobulin kappa variable 3D-15 A0A087WSY6 KVD15_HUMAN 0.04021589 1.814796476 ↑ Luminal B HER2 Pos
Mannose-binding protein C P11226 MBL2_HUMAN 0.040341289 2.238378752 ↑ Luminal B HER2 Pos
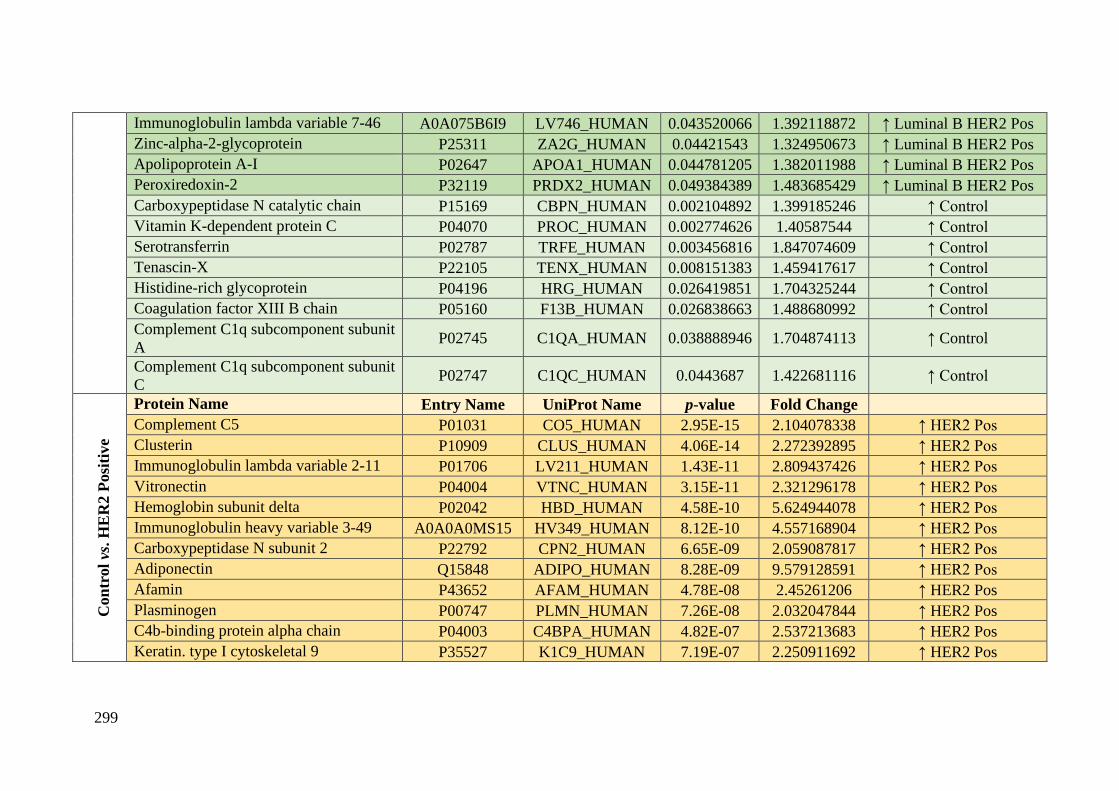
299
Immunoglobulin lambda variable 7-46 A0A075B6I9 LV746_HUMAN 0.043520066 1.392118872 ↑ Luminal B HER2 Pos
Zinc-alpha-2-glycoprotein P25311 ZA2G_HUMAN 0.04421543 1.324950673 ↑ Luminal B HER2 Pos
Apolipoprotein A-I P02647 APOA1_HUMAN 0.044781205 1.382011988 ↑ Luminal B HER2 Pos
Peroxiredoxin-2 P32119 PRDX2_HUMAN 0.049384389 1.483685429 ↑ Luminal B HER2 Pos
Carboxypeptidase N catalytic chain P15169 CBPN_HUMAN 0.002104892 1.399185246 ↑ Control
Vitamin K-dependent protein C P04070 PROC_HUMAN 0.002774626 1.40587544 ↑ Control
Serotransferrin P02787 TRFE_HUMAN 0.003456816 1.847074609 ↑ Control
Tenascin-X P22105 TENX_HUMAN 0.008151383 1.459417617 ↑ Control
Histidine-rich glycoprotein P04196 HRG_HUMAN 0.026419851 1.704325244 ↑ Control
Coagulation factor XIII B chain P05160 F13B_HUMAN 0.026838663 1.488680992 ↑ Control
Complement C1q subcomponent subunit
A P02745 C1QA_HUMAN 0.038888946 1.704874113 ↑ Control
Complement C1q subcomponent subunit
C P02747 C1QC_HUMAN 0.0443687 1.422681116 ↑ Control
Con
trol vs
. H
ER
2 P
osi
tive
Protein Name Entry Name UniProt Name p-value Fold Change
Complement C5 P01031 CO5_HUMAN 2.95E-15 2.104078338 ↑ HER2 Pos
Clusterin P10909 CLUS_HUMAN 4.06E-14 2.272392895 ↑ HER2 Pos
Immunoglobulin lambda variable 2-11 P01706 LV211_HUMAN 1.43E-11 2.809437426 ↑ HER2 Pos
Vitronectin P04004 VTNC_HUMAN 3.15E-11 2.321296178 ↑ HER2 Pos
Hemoglobin subunit delta P02042 HBD_HUMAN 4.58E-10 5.624944078 ↑ HER2 Pos
Immunoglobulin heavy variable 3-49 A0A0A0MS15 HV349_HUMAN 8.12E-10 4.557168904 ↑ HER2 Pos
Carboxypeptidase N subunit 2 P22792 CPN2_HUMAN 6.65E-09 2.059087817 ↑ HER2 Pos
Adiponectin Q15848 ADIPO_HUMAN 8.28E-09 9.579128591 ↑ HER2 Pos
Afamin P43652 AFAM_HUMAN 4.78E-08 2.45261206 ↑ HER2 Pos
Plasminogen P00747 PLMN_HUMAN 7.26E-08 2.032047844 ↑ HER2 Pos
C4b-binding protein alpha chain P04003 C4BPA_HUMAN 4.82E-07 2.537213683 ↑ HER2 Pos
Keratin. type I cytoskeletal 9 P35527 K1C9_HUMAN 7.19E-07 2.250911692 ↑ HER2 Pos

300
Immunoglobulin heavy variable 3-73 A0A0B4J1V6 HV373_HUMAN 1.67E-06 15.87006684 ↑ HER2 Pos
Vitamin D-binding protein P02774 VTDB_HUMAN 1.96E-06 2.050789715 ↑ HER2 Pos
Coagulation factor XII P00748 FA12_HUMAN 2.50E-06 4.48323521 ↑ HER2 Pos
Complement C3 P01024 CO3_HUMAN 2.61E-06 2.060113843 ↑ HER2 Pos
Immunoglobulin heavy variable 4-61 A0A0C4DH41 HV461_HUMAN 4.13E-06 5.17636397 ↑ HER2 Pos
Ficolin-2 Q15485 FCN2_HUMAN 7.19E-06 2.289548422 ↑ HER2 Pos
Fibrinogen alpha chain P02671 FIBA_HUMAN 1.19E-05 1.946313958 ↑ HER2 Pos
Plasma kallikrein P03952 KLKB1_HUMAN 2.05E-05 2.839283512 ↑ HER2 Pos
Transthyretin P02766 TTHY_HUMAN 2.53E-05 1.710033019 ↑ HER2 Pos
Hemoglobin subunit alpha P69905 HBA_HUMAN 3.18E-05 1.915979149 ↑ HER2 Pos
Serum albumin P02768 ALBU_HUMAN 6.15E-05 1.617918878 ↑ HER2 Pos
Immunoglobulin heavy variable 3-23 P01764 HV323_HUMAN 8.83E-05 2.952483084 ↑ HER2 Pos
Prenylcysteine oxidase 1 Q9UHG3 PCYOX_HUMAN 9.23E-05 2.137601432 ↑ HER2 Pos
Fibronectin P02751 FINC_HUMAN 0.000101285 1.917381182 ↑ HER2 Pos
Coagulation factor IX P00740 FA9_HUMAN 0.00012729 1.600811451 ↑ HER2 Pos
Serum amyloid P-component P02743 SAMP_HUMAN 0.000144697 4.818702495 ↑ HER2 Pos
Complement component C8 alpha chain P07357 CO8A_HUMAN 0.0001513 1.959771044 ↑ HER2 Pos
Complement factor H P08603 CFAH_HUMAN 0.000175819 1.83792215 ↑ HER2 Pos
Complement factor H-related protein 4 Q92496 FHR4_HUMAN 0.000204336 3.107198257 ↑ HER2 Pos
Glutathione peroxidase 3 P22352 GPX3_HUMAN 0.000266714 2.917862479 ↑ HER2 Pos
Complement factor D P00746 CFAD_HUMAN 0.00027027 2.477435187 ↑ HER2 Pos
Immunoglobulin lambda variable 1-51 P01701 LV151_HUMAN 0.000382645 2.419267666 ↑ HER2 Pos
Immunoglobulin heavy variable 3-64 A0A075B6Q5 HV364_HUMAN 0.000401774 2.217759109 ↑ HER2 Pos
Hemoglobin subunit beta P68871 HBB_HUMAN 0.000414991 2.238927783 ↑ HER2 Pos
Proteoglycan 4 Q92954 PRG4_HUMAN 0.000502566 2.732642249 ↑ HER2 Pos
Selenoprotein P P49908 SEPP1_HUMAN 0.000539614 3.796988329 ↑ HER2 Pos
Insulin-like growth factor-binding P35858 ALS_HUMAN 0.000815729 1.776314681 ↑ HER2 Pos
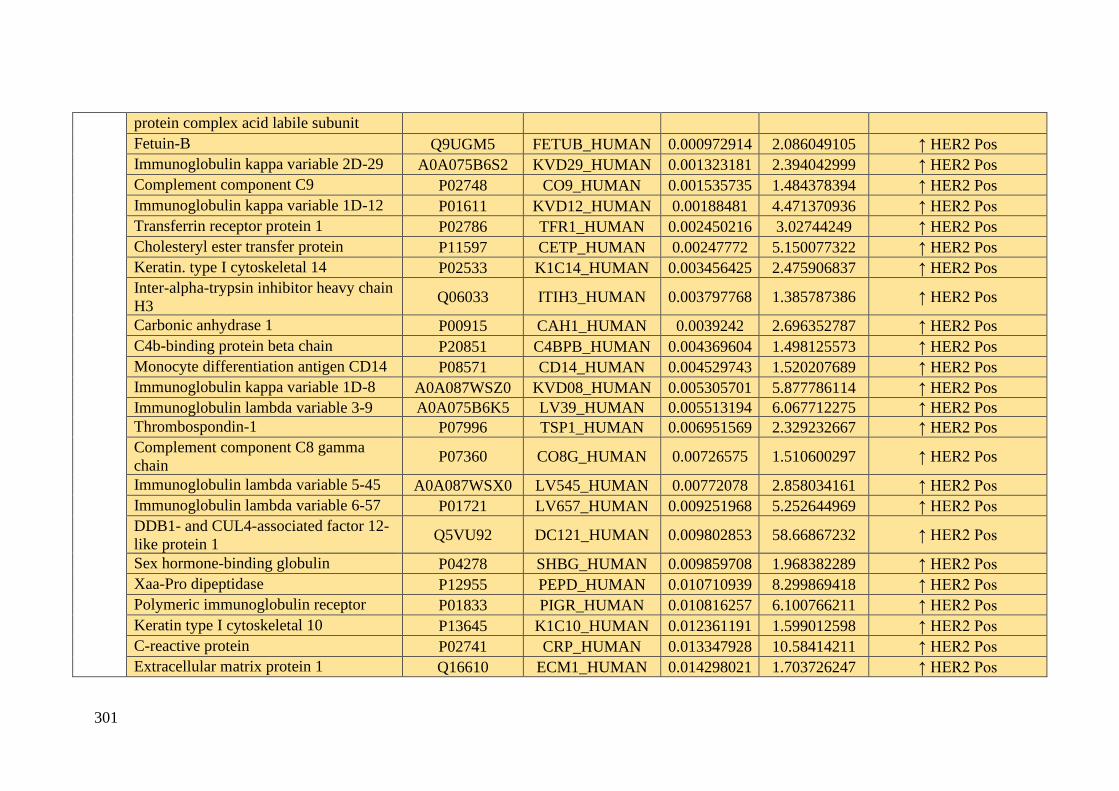
301
protein complex acid labile subunit
Fetuin-B Q9UGM5 FETUB_HUMAN 0.000972914 2.086049105 ↑ HER2 Pos
Immunoglobulin kappa variable 2D-29 A0A075B6S2 KVD29_HUMAN 0.001323181 2.394042999 ↑ HER2 Pos
Complement component C9 P02748 CO9_HUMAN 0.001535735 1.484378394 ↑ HER2 Pos
Immunoglobulin kappa variable 1D-12 P01611 KVD12_HUMAN 0.00188481 4.471370936 ↑ HER2 Pos
Transferrin receptor protein 1 P02786 TFR1_HUMAN 0.002450216 3.02744249 ↑ HER2 Pos
Cholesteryl ester transfer protein P11597 CETP_HUMAN 0.00247772 5.150077322 ↑ HER2 Pos
Keratin. type I cytoskeletal 14 P02533 K1C14_HUMAN 0.003456425 2.475906837 ↑ HER2 Pos
Inter-alpha-trypsin inhibitor heavy chain
H3 Q06033 ITIH3_HUMAN 0.003797768 1.385787386 ↑ HER2 Pos
Carbonic anhydrase 1 P00915 CAH1_HUMAN 0.0039242 2.696352787 ↑ HER2 Pos
C4b-binding protein beta chain P20851 C4BPB_HUMAN 0.004369604 1.498125573 ↑ HER2 Pos
Monocyte differentiation antigen CD14 P08571 CD14_HUMAN 0.004529743 1.520207689 ↑ HER2 Pos
Immunoglobulin kappa variable 1D-8 A0A087WSZ0 KVD08_HUMAN 0.005305701 5.877786114 ↑ HER2 Pos
Immunoglobulin lambda variable 3-9 A0A075B6K5 LV39_HUMAN 0.005513194 6.067712275 ↑ HER2 Pos
Thrombospondin-1 P07996 TSP1_HUMAN 0.006951569 2.329232667 ↑ HER2 Pos
Complement component C8 gamma
chain P07360 CO8G_HUMAN 0.00726575 1.510600297 ↑ HER2 Pos
Immunoglobulin lambda variable 5-45 A0A087WSX0 LV545_HUMAN 0.00772078 2.858034161 ↑ HER2 Pos
Immunoglobulin lambda variable 6-57 P01721 LV657_HUMAN 0.009251968 5.252644969 ↑ HER2 Pos
DDB1- and CUL4-associated factor 12-
like protein 1 Q5VU92 DC121_HUMAN 0.009802853 58.66867232 ↑ HER2 Pos
Sex hormone-binding globulin P04278 SHBG_HUMAN 0.009859708 1.968382289 ↑ HER2 Pos
Xaa-Pro dipeptidase P12955 PEPD_HUMAN 0.010710939 8.299869418 ↑ HER2 Pos
Polymeric immunoglobulin receptor P01833 PIGR_HUMAN 0.010816257 6.100766211 ↑ HER2 Pos
Keratin type I cytoskeletal 10 P13645 K1C10_HUMAN 0.012361191 1.599012598 ↑ HER2 Pos
C-reactive protein P02741 CRP_HUMAN 0.013347928 10.58414211 ↑ HER2 Pos
Extracellular matrix protein 1 Q16610 ECM1_HUMAN 0.014298021 1.703726247 ↑ HER2 Pos
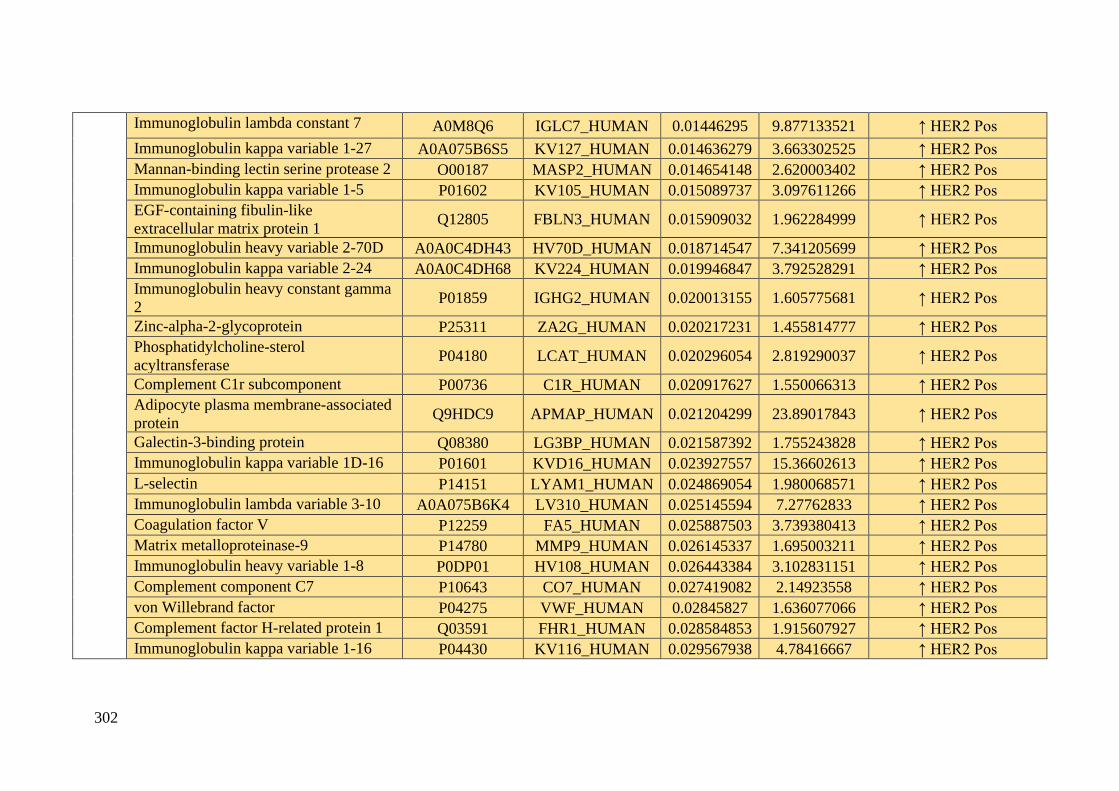
302
Immunoglobulin lambda constant 7 A0M8Q6 IGLC7_HUMAN 0.01446295 9.877133521 ↑ HER2 Pos
Immunoglobulin kappa variable 1-27 A0A075B6S5 KV127_HUMAN 0.014636279 3.663302525 ↑ HER2 Pos
Mannan-binding lectin serine protease 2 O00187 MASP2_HUMAN 0.014654148 2.620003402 ↑ HER2 Pos
Immunoglobulin kappa variable 1-5 P01602 KV105_HUMAN 0.015089737 3.097611266 ↑ HER2 Pos
EGF-containing fibulin-like
extracellular matrix protein 1 Q12805 FBLN3_HUMAN 0.015909032 1.962284999 ↑ HER2 Pos
Immunoglobulin heavy variable 2-70D A0A0C4DH43 HV70D_HUMAN 0.018714547 7.341205699 ↑ HER2 Pos
Immunoglobulin kappa variable 2-24 A0A0C4DH68 KV224_HUMAN 0.019946847 3.792528291 ↑ HER2 Pos
Immunoglobulin heavy constant gamma
2 P01859 IGHG2_HUMAN 0.020013155 1.605775681 ↑ HER2 Pos
Zinc-alpha-2-glycoprotein P25311 ZA2G_HUMAN 0.020217231 1.455814777 ↑ HER2 Pos
Phosphatidylcholine-sterol
acyltransferase P04180 LCAT_HUMAN 0.020296054 2.819290037 ↑ HER2 Pos
Complement C1r subcomponent P00736 C1R_HUMAN 0.020917627 1.550066313 ↑ HER2 Pos
Adipocyte plasma membrane-associated
protein Q9HDC9 APMAP_HUMAN 0.021204299 23.89017843 ↑ HER2 Pos
Galectin-3-binding protein Q08380 LG3BP_HUMAN 0.021587392 1.755243828 ↑ HER2 Pos
Immunoglobulin kappa variable 1D-16 P01601 KVD16_HUMAN 0.023927557 15.36602613 ↑ HER2 Pos
L-selectin P14151 LYAM1_HUMAN 0.024869054 1.980068571 ↑ HER2 Pos
Immunoglobulin lambda variable 3-10 A0A075B6K4 LV310_HUMAN 0.025145594 7.27762833 ↑ HER2 Pos
Coagulation factor V P12259 FA5_HUMAN 0.025887503 3.739380413 ↑ HER2 Pos
Matrix metalloproteinase-9 P14780 MMP9_HUMAN 0.026145337 1.695003211 ↑ HER2 Pos
Immunoglobulin heavy variable 1-8 P0DP01 HV108_HUMAN 0.026443384 3.102831151 ↑ HER2 Pos
Complement component C7 P10643 CO7_HUMAN 0.027419082 2.14923558 ↑ HER2 Pos
von Willebrand factor P04275 VWF_HUMAN 0.02845827 1.636077066 ↑ HER2 Pos
Complement factor H-related protein 1 Q03591 FHR1_HUMAN 0.028584853 1.915607927 ↑ HER2 Pos
Immunoglobulin kappa variable 1-16 P04430 KV116_HUMAN 0.029567938 4.78416667 ↑ HER2 Pos

303
Cysteine-rich secretory protein 3 P54108 CRIS3_HUMAN 0.034870563 3.338300324 ↑ HER2 Pos
Keratin. type II cytoskeletal 2 epidermal P35908 K22E_HUMAN 0.035300084 1.805122667 ↑ HER2 Pos
Immunoglobulin heavy variable 3-33 P01772 HV333_HUMAN 0.038394512 8.344444793 ↑ HER2 Pos
Properdin P27918 PROP_HUMAN 0.038520124 1.768625617 ↑ HER2 Pos
Complement C1q subcomponent subunit
A P02745 C1QA_HUMAN 0.039735285 1.871918365 ↑ HER2 Pos
Pigment epithelium-derived factor P36955 PEDF_HUMAN 0.042658539 1.861759712 ↑ HER2 Pos
Immunoglobulin heavy variable 1-46 P01743 HV146_HUMAN 0.043543848 4.597946701 ↑ HER2 Pos
Immunoglobulin kappa variable 1-8 A0A0C4DH67 KV108_HUMAN 0.044140149 1.81212033 ↑ HER2 Pos
Immunoglobulin heavy variable 3-72 A0A0B4J1Y9 HV372_HUMAN 0.047935423 1.687200865 ↑ HER2 Pos
Ficolin-3 O75636 FCN3_HUMAN 1.60E-07 5.889667169 ↑ Control
Apolipoprotein A-I P02647 APOA1_HUMAN 6.44E-07 3.362021938 ↑ Control
Nuclear receptor coactivator 6 Q14686 NCOA6_HUMAN 8.47E-07 8.441443914 ↑ Control
Heparin cofactor 2 P05546 HEP2_HUMAN 1.00E-05 3.661036082 ↑ Control
Kininogen-1 P01042 KNG1_HUMAN 3.73E-05 3.357349607 ↑ Control
Vitamin K-dependent protein S P07225 PROS_HUMAN 9.45E-05 1.853196173 ↑ Control
Histidine-rich glycoprotein P04196 HRG_HUMAN 0.000112488 5.940948958 ↑ Control
Immunoglobulin J chain P01591 IGJ_HUMAN 0.000238757 2.91931338 ↑ Control
N-acetylmuramoyl-L-alanine amidase Q96PD5 PGRP2_HUMAN 0.000321687 1.799194526 ↑ Control
Angiotensinogen P01019 ANGT_HUMAN 0.00043286 3.016543479 ↑ Control
Inter-alpha-trypsin inhibitor heavy chain
H2 P19823 ITIH2_HUMAN 0.00074408 1.923949583 ↑ Control
Apolipoprotein M O95445 APOM_HUMAN 0.000850537 1.771464702 ↑ Control
Haptoglobin-related protein P00739 HPTR_HUMAN 0.000868695 2.12707613 ↑ Control
Alpha-1-antitrypsin P01009 A1AT_HUMAN 0.001111283 4.959761437 ↑ Control
Inter-alpha-trypsin inhibitor heavy chain
H1 P19827 ITIH1_HUMAN 0.001316207 1.682469513 ↑ Control

304
Beta-2-glycoprotein 1 P02749 APOH_HUMAN 0.001791485 2.272364551 ↑ Control
Alpha-2-HS-glycoprotein P02765 FETUA_HUMAN 0.001836174 1.586208463 ↑ Control
Apolipoprotein C-IV P55056 APOC4_HUMAN 0.001950165 6.335750729 ↑ Control
Apolipoprotein A-II P02652 APOA2_HUMAN 0.002133097 2.248237048 ↑ Control
Trypsin-1 P07477 TRY1_HUMAN 0.002454431 4.217786063 ↑ Control
Apolipoprotein C-III P02656 APOC3_HUMAN 0.002567875 3.440587523 ↑ Control
Plasma protease C1 inhibitor P05155 IC1_HUMAN 0.00372715 1.579476004 ↑ Control
Apolipoprotein F Q13790 APOF_HUMAN 0.005336626 7.55893037 ↑ Control
Carboxypeptidase B2 Q96IY4 CBPB2_HUMAN 0.009929161 1.517845486 ↑ Control
Antithrombin-III P01008 ANT3_HUMAN 0.018612975 1.308597079 ↑ Control
Complement C1q subcomponent subunit
B P02746 C1QB_HUMAN 0.020285082 1.835321411 ↑ Control
Inter-alpha-trypsin inhibitor heavy chain
H4 Q14624 ITIH4_HUMAN 0.02171345 1.978217456 ↑ Control
Alpha-2-macroglobulin P01023 A2MG_HUMAN 0.021759431 1.818611048 ↑ Control
Ceruloplasmin P00450 CERU_HUMAN 0.023271461 3.101927554 ↑ Control
Alpha-1B-glycoprotein P04217 A1BG_HUMAN 0.030481015 3.465086804 ↑ Control
Haptoglobin P00738 HPT_HUMAN 0.032119775 1.834688417 ↑ Control
Serum amyloid A-4 protein P35542 SAA4_HUMAN 0.033032185 2.177960496 ↑ Control
Complement C1q subcomponent subunit
C P02747 C1QC_HUMAN 0.034179123 1.548970133 ↑ Control
Serotransferrin P02787 TRFE_HUMAN 0.034307125 1.559639179 ↑ Control
Apolipoprotein(a) P08519 APOA_HUMAN 0.046687987 2.207374657 ↑ Control
Con
trol
vs. T
rip
le
Neg
ati
ve Protein Name Entry Name UniProt Name p-value Fold Change
Immunoglobulin heavy variable 4-31 P0DP07 HV431_HUMAN 3.40E-06 6.336923351 ↑ Triple Negative
Hemoglobin subunit beta P68871 HBB_HUMAN 4.66E-06 1.856970774 ↑ Triple Negative
Beta-2-glycoprotein 1 P02749 APOH_HUMAN 9.79E-06 1.826994929 ↑ Triple Negative
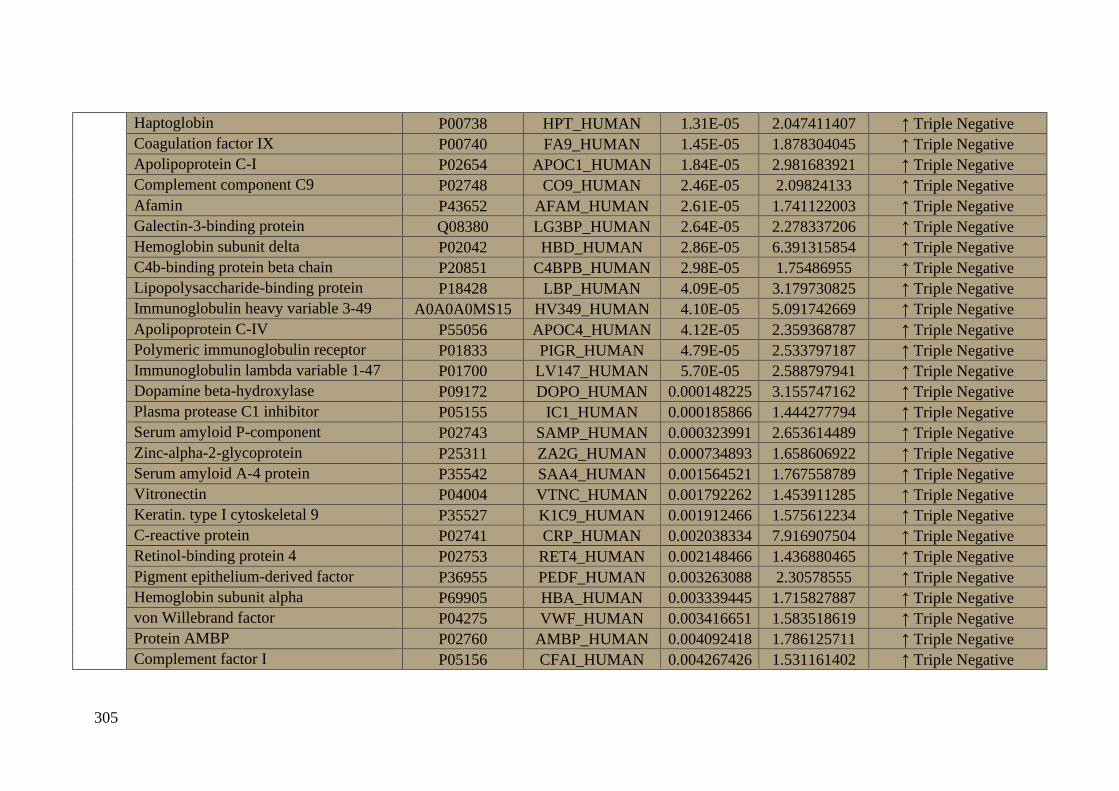
305
Haptoglobin P00738 HPT_HUMAN 1.31E-05 2.047411407 ↑ Triple Negative
Coagulation factor IX P00740 FA9_HUMAN 1.45E-05 1.878304045 ↑ Triple Negative
Apolipoprotein C-I P02654 APOC1_HUMAN 1.84E-05 2.981683921 ↑ Triple Negative
Complement component C9 P02748 CO9_HUMAN 2.46E-05 2.09824133 ↑ Triple Negative
Afamin P43652 AFAM_HUMAN 2.61E-05 1.741122003 ↑ Triple Negative
Galectin-3-binding protein Q08380 LG3BP_HUMAN 2.64E-05 2.278337206 ↑ Triple Negative
Hemoglobin subunit delta P02042 HBD_HUMAN 2.86E-05 6.391315854 ↑ Triple Negative
C4b-binding protein beta chain P20851 C4BPB_HUMAN 2.98E-05 1.75486955 ↑ Triple Negative
Lipopolysaccharide-binding protein P18428 LBP_HUMAN 4.09E-05 3.179730825 ↑ Triple Negative
Immunoglobulin heavy variable 3-49 A0A0A0MS15 HV349_HUMAN 4.10E-05 5.091742669 ↑ Triple Negative
Apolipoprotein C-IV P55056 APOC4_HUMAN 4.12E-05 2.359368787 ↑ Triple Negative
Polymeric immunoglobulin receptor P01833 PIGR_HUMAN 4.79E-05 2.533797187 ↑ Triple Negative
Immunoglobulin lambda variable 1-47 P01700 LV147_HUMAN 5.70E-05 2.588797941 ↑ Triple Negative
Dopamine beta-hydroxylase P09172 DOPO_HUMAN 0.000148225 3.155747162 ↑ Triple Negative
Plasma protease C1 inhibitor P05155 IC1_HUMAN 0.000185866 1.444277794 ↑ Triple Negative
Serum amyloid P-component P02743 SAMP_HUMAN 0.000323991 2.653614489 ↑ Triple Negative
Zinc-alpha-2-glycoprotein P25311 ZA2G_HUMAN 0.000734893 1.658606922 ↑ Triple Negative
Serum amyloid A-4 protein P35542 SAA4_HUMAN 0.001564521 1.767558789 ↑ Triple Negative
Vitronectin P04004 VTNC_HUMAN 0.001792262 1.453911285 ↑ Triple Negative
Keratin. type I cytoskeletal 9 P35527 K1C9_HUMAN 0.001912466 1.575612234 ↑ Triple Negative
C-reactive protein P02741 CRP_HUMAN 0.002038334 7.916907504 ↑ Triple Negative
Retinol-binding protein 4 P02753 RET4_HUMAN 0.002148466 1.436880465 ↑ Triple Negative
Pigment epithelium-derived factor P36955 PEDF_HUMAN 0.003263088 2.30578555 ↑ Triple Negative
Hemoglobin subunit alpha P69905 HBA_HUMAN 0.003339445 1.715827887 ↑ Triple Negative
von Willebrand factor P04275 VWF_HUMAN 0.003416651 1.583518619 ↑ Triple Negative
Protein AMBP P02760 AMBP_HUMAN 0.004092418 1.786125711 ↑ Triple Negative
Complement factor I P05156 CFAI_HUMAN 0.004267426 1.531161402 ↑ Triple Negative

306
Apolipoprotein E P02649 APOE_HUMAN 0.005108321 1.297484301 ↑ Triple Negative
Peroxiredoxin-2 P32119 PRDX2_HUMAN 0.005462251 1.56831661 ↑ Triple Negative
IgGFc-binding protein Q9Y6R7 FCGBP_HUMAN 0.006429638 1.543604283 ↑ Triple Negative
Transferrin receptor protein 1 P02786 TFR1_HUMAN 0.006576023 2.659048849 ↑ Triple Negative
Complement component C6 P13671 CO6_HUMAN 0.006913895 3.211027998 ↑ Triple Negative
Matrix metalloproteinase-9 P14780 MMP9_HUMAN 0.007583113 2.1547884 ↑ Triple Negative
Immunoglobulin heavy variable 1-8 P0DP01 HV108_HUMAN 0.007621133 2.188587398 ↑ Triple Negative
Gelsolin P06396 GELS_HUMAN 0.008449427 1.600858701 ↑ Triple Negative
Cadherin-5 P33151 CADH5_HUMAN 0.008460741 1.74346858 ↑ Triple Negative
Plasma serine protease inhibitor P05154 IPSP_HUMAN 0.008634823 1.750498274 ↑ Triple Negative
Immunoglobulin heavy variable 1-2 P23083 HV102_HUMAN 0.008919673 1.559197168 ↑ Triple Negative
Fetuin-B Q9UGM5 FETUB_HUMAN 0.009781609 1.67811206 ↑ Triple Negative
Voltage-dependent L-type calcium
channel subunit alpha-1F O60840 CAC1F_HUMAN 0.010760078 3.330169664 ↑ Triple Negative
Properdin P27918 PROP_HUMAN 0.010990918 2.859498176 ↑ Triple Negative
Heparin cofactor 2 P05546 HEP2_HUMAN 0.011416361 1.356715555 ↑ Triple Negative
Immunoglobulin lambda variable 1-40 P01703 LV140_HUMAN 0.011566545 2.233258232 ↑ Triple Negative
Transthyretin P02766 TTHY_HUMAN 0.012616261 1.557894268 ↑ Triple Negative
Apolipoprotein A-IV P06727 APOA4_HUMAN 0.013273745 1.393597405 ↑ Triple Negative
Immunoglobulin kappa variable 2D-29 A0A075B6S2 KVD29_HUMAN 0.014548683 1.976553582 ↑ Triple Negative
Immunoglobulin lambda constant 7 A0M8Q6 IGLC7_HUMAN 0.015204337 2.30606418 ↑ Triple Negative
Hyaluronan-binding protein 2 Q14520 HABP2_HUMAN 0.017778041 1.837364492 ↑ Triple Negative
Complement component C7 P10643 CO7_HUMAN 0.018680554 2.247802867 ↑ Triple Negative
Immunoglobulin lambda variable 3-9 A0A075B6K5 LV39_HUMAN 0.018777301 5.992460294 ↑ Triple Negative
Apolipoprotein C-III P02656 APOC3_HUMAN 0.020727463 1.580436985 ↑ Triple Negative
Complement factor H-related protein 4 Q92496 FHR4_HUMAN 0.021147868 2.333186411 ↑ Triple Negative
Hemopexin P02790 HEMO_HUMAN 0.021507045 1.152784299 ↑ Triple Negative
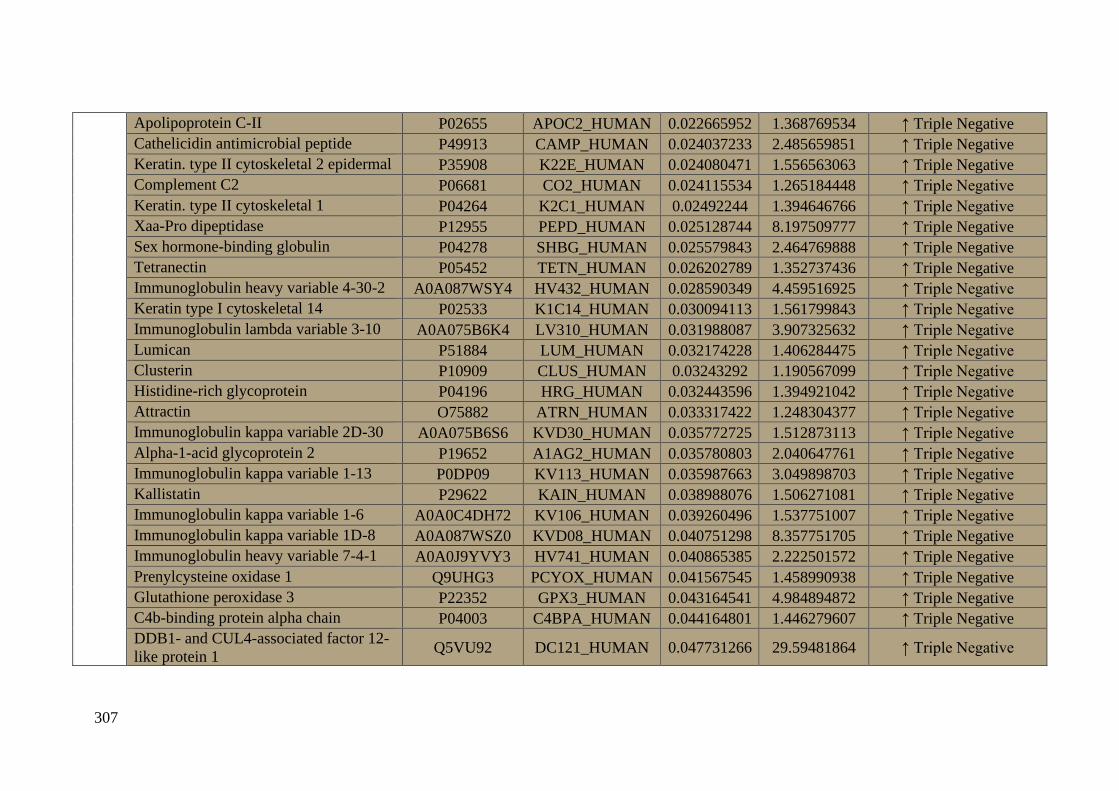
307
Apolipoprotein C-II P02655 APOC2_HUMAN 0.022665952 1.368769534 ↑ Triple Negative
Cathelicidin antimicrobial peptide P49913 CAMP_HUMAN 0.024037233 2.485659851 ↑ Triple Negative
Keratin. type II cytoskeletal 2 epidermal P35908 K22E_HUMAN 0.024080471 1.556563063 ↑ Triple Negative
Complement C2 P06681 CO2_HUMAN 0.024115534 1.265184448 ↑ Triple Negative
Keratin. type II cytoskeletal 1 P04264 K2C1_HUMAN 0.02492244 1.394646766 ↑ Triple Negative
Xaa-Pro dipeptidase P12955 PEPD_HUMAN 0.025128744 8.197509777 ↑ Triple Negative
Sex hormone-binding globulin P04278 SHBG_HUMAN 0.025579843 2.464769888 ↑ Triple Negative
Tetranectin P05452 TETN_HUMAN 0.026202789 1.352737436 ↑ Triple Negative
Immunoglobulin heavy variable 4-30-2 A0A087WSY4 HV432_HUMAN 0.028590349 4.459516925 ↑ Triple Negative
Keratin type I cytoskeletal 14 P02533 K1C14_HUMAN 0.030094113 1.561799843 ↑ Triple Negative
Immunoglobulin lambda variable 3-10 A0A075B6K4 LV310_HUMAN 0.031988087 3.907325632 ↑ Triple Negative
Lumican P51884 LUM_HUMAN 0.032174228 1.406284475 ↑ Triple Negative
Clusterin P10909 CLUS_HUMAN 0.03243292 1.190567099 ↑ Triple Negative
Histidine-rich glycoprotein P04196 HRG_HUMAN 0.032443596 1.394921042 ↑ Triple Negative
Attractin O75882 ATRN_HUMAN 0.033317422 1.248304377 ↑ Triple Negative
Immunoglobulin kappa variable 2D-30 A0A075B6S6 KVD30_HUMAN 0.035772725 1.512873113 ↑ Triple Negative
Alpha-1-acid glycoprotein 2 P19652 A1AG2_HUMAN 0.035780803 2.040647761 ↑ Triple Negative
Immunoglobulin kappa variable 1-13 P0DP09 KV113_HUMAN 0.035987663 3.049898703 ↑ Triple Negative
Kallistatin P29622 KAIN_HUMAN 0.038988076 1.506271081 ↑ Triple Negative
Immunoglobulin kappa variable 1-6 A0A0C4DH72 KV106_HUMAN 0.039260496 1.537751007 ↑ Triple Negative
Immunoglobulin kappa variable 1D-8 A0A087WSZ0 KVD08_HUMAN 0.040751298 8.357751705 ↑ Triple Negative
Immunoglobulin heavy variable 7-4-1 A0A0J9YVY3 HV741_HUMAN 0.040865385 2.222501572 ↑ Triple Negative
Prenylcysteine oxidase 1 Q9UHG3 PCYOX_HUMAN 0.041567545 1.458990938 ↑ Triple Negative
Glutathione peroxidase 3 P22352 GPX3_HUMAN 0.043164541 4.984894872 ↑ Triple Negative
C4b-binding protein alpha chain P04003 C4BPA_HUMAN 0.044164801 1.446279607 ↑ Triple Negative
DDB1- and CUL4-associated factor 12-
like protein 1 Q5VU92 DC121_HUMAN 0.047731266 29.59481864 ↑ Triple Negative
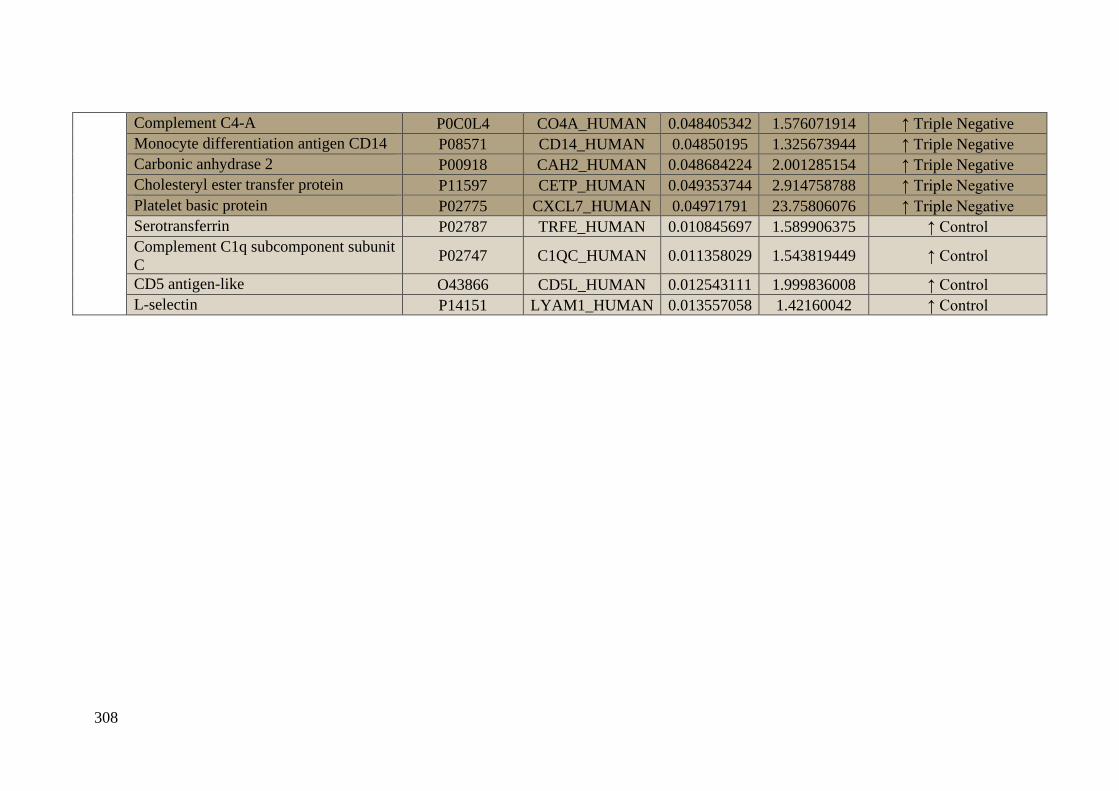
308
Complement C4-A P0C0L4 CO4A_HUMAN 0.048405342 1.576071914 ↑ Triple Negative
Monocyte differentiation antigen CD14 P08571 CD14_HUMAN 0.04850195 1.325673944 ↑ Triple Negative
Carbonic anhydrase 2 P00918 CAH2_HUMAN 0.048684224 2.001285154 ↑ Triple Negative
Cholesteryl ester transfer protein P11597 CETP_HUMAN 0.049353744 2.914758788 ↑ Triple Negative
Platelet basic protein P02775 CXCL7_HUMAN 0.04971791 23.75806076 ↑ Triple Negative
Serotransferrin P02787 TRFE_HUMAN 0.010845697 1.589906375 ↑ Control
Complement C1q subcomponent subunit
C P02747 C1QC_HUMAN 0.011358029 1.543819449 ↑ Control
CD5 antigen-like O43866 CD5L_HUMAN 0.012543111 1.999836008 ↑ Control
L-selectin P14151 LYAM1_HUMAN 0.013557058 1.42160042 ↑ Control

309
Table 4_SM. Specific or unique differentially expressed proteins (up-regulated and down-regulated) (p-value ≤ 0.05) found in the protein
patterns of the ex vivo formed coronas after the analysis by SWATH-MS for the different breast cancer subtypes (LA, n = 11; LB-, n = 10; LB+,
n = 7; HER2+, n = 6; TNBC, n = 8) in comparison with healthy control (HC) samples.
Luminal A
Entry name Statistically significant
Luminal B-HER negative
Entry name
Statistically significant
Luminal B-HER positive Entry name
Statistically significant
HER2+
Entry name
Statistically significant
TNBC
Entry name
Statistically significant
up-regulated
n = 42
down-
regulated
n = 18
up-regulated
n = 100
down-
regulated
n = 32
up-regulated
n = 59
down-
regulated
n = 8
up-regulated
n = 95
down-
regulated
n = 35
up-regulated
n = 87
down-
regulated
n = 4
uniques = 4 uniques =
4
uniques = 25 uniques = 2 uniques = 2 uniques =
0
uniques = 23 uniques = 5 uniques = 9 uniques = 1
A2AP ↑ A2MG ↓ A1AG1 ↑ IGHA1 ↑ AFAM ↓ A1AG1 ↑ K22E ↑ C1QA ↓ ADIPO ↑ HV461 ↑ A1AT ↓ A1AG2 ↑ HV102 ↑ C1QC ↓
APOC3 ↑ CADH5 ↓ A1AG2 ↑ IGHA2 ↑ ALBU ↓ A2GL ↑ KAIN ↑ C1QC ↓ AFAM ↑ HV70D ↑ A1BG ↓ AFAM ↑ HV108 ↑ CD5L ↓
APOH ↑ CBG ↓ A1BG ↑ IGHG1 ↑ ALS ↓ AACT ↑ KV113 ↑ CBPN ↓ ALBU ↑ IGHG2 ↑ A2MG ↓ AMBP ↑ HV349 ↑ LYAM1 ↓
APOL1 ↑ CBPN ↓ A2AP ↑ IGJ ↑ APOA ↓ APOA1 ↑ KV133 ↑ F13B ↓ ALS ↑ IGLC7 ↑ ANGT ↓ APOA4 ↑ HV431 ↑ TRFE ↓
C1R ↑ CERU ↓ A2GL ↑ IGKC↑ APOM ↓ APOA2 ↑ KVD15 ↑ HRG ↓ APMAP ↑ ITIH3 ↑ ANT3 ↓ APOC1↑ HV432 ↑
C1RL ↑ CNDP1 ↓ AACT ↑ IGLC7 ↑ C1QA ↓ APOA4 ↑ KVD21 ↑ PROC ↓ C1QA ↑ K1C10 ↑ APOA ↓ APOC2 ↑ HV741 ↑
C4BPA ↑ CPN2 ↓ AMBP ↑ IGLL5 ↑ C1QB ↓ APOC1 ↑ LG3BP ↑ TENX ↓ C1R ↑ K1C14 ↑ APOA1 ↓ APOC3 ↑ IC1 ↑
CAMP ↑ HV124 ↓ ANGT ↑ ITIH1 ↑ C1QC↓ APOC2 ↑ LV140 ↑ TRFE ↓ C4BPA ↑ K1C9 ↑ APOA2 ↓ APOC4 ↑ IGLC7 ↑
CFAB ↑ HV349 ↓ APOA1 ↑ ITIH2 ↑ C1S ↓ APOC3 ↑ LV147 ↑ C4BPB ↑ K22E ↑ APOC3 ↓ APOE ↑ IPSP ↑
CO8B ↑ HV372 ↓ APOA2 ↑ ITIH4 ↑ CD14 ↓ APOL1 ↑ LV211 ↑ CAH1 ↑ KLKB1 ↑ APOC4 ↓ APOH ↑ K1C14 ↑
CO8G ↑ HV428 ↓ APOB ↑ KAIN ↑ CFAB ↓ C4BPB ↑ LV310 ↑ CD14 ↑ KV105 ↑ APOF ↓ ATRN ↑ K1C9 ↑
CO9 ↑ HV741 ↓ APOC1 ↑ KV116 ↑ CFAD ↓ CBG ↑ LV319 ↑ CETP ↑ KV108 ↑ APOH ↓ C4BPA ↑ K22E ↑
COL11 ↑ IGHM ↓ APOC2 ↑ KV133 ↑ CFAH ↓ CHLE ↑ LV325 ↑ CFAD ↑ KV116 ↑ APOM ↓ C4BPB ↑ K2C1 ↑
CRP ↑ K1C14 ↓ APOC3 ↑ KV311 ↑ CFAI ↓ CO4A ↑ LV39 ↑ CFAH ↑ KV127 ↑ C1QB ↓ CAC1F ↑ KAIN ↑
DOPO ↑ KV108 ↓ APOC4 ↑ KV401 ↑ CO3 ↓ CO6 ↑ LV746 CLUS ↑ KV224 ↑ C1QC ↓ CADH5 ↑ KV106 ↑
FA10 ↑ KV320 ↓ APOD ↑ KVD15 ↑ CO8G ↓ CO8A ↑ MBL2 ↑ CO3 ↑ KVD08 ↑ CBPB2 ↓ CAH2 ↑ KV113 ↑
FA9 ↑ SHBG ↓ APOL1 ↑ KVD20 ↑ ECM1↓ CO9 ↑ PCYOX ↑ CO5 ↑ KVD12 ↑ CERU ↓ CAMP ↑ KVD08 ↑
FCGBP ↑ ZPI ↓ BTD ↑ KVD21↑ F13B ↓ CRP ↑ PEDF ↑ CO7 ↑ KVD16 ↑ FCN3 ↓ CD14 ↑ KVD29 ↑
FCN2 ↑ C4BPB ↑ LBP ↑ FHR1 ↓ GELS PEPD ↑ CO8A ↑ KVD29 ↑ FETUA ↓ CETP ↑ KVD30 ↑
FCN3 ↑ CADH5 ↑ LCAT ↑ FHR4 ↓ HBA ↑ PIGR ↑ CO8G ↑ LCAT ↑ HEP2 ↓ CFAI ↑ LBP ↑
FHR2 ↑ CAH1 ↑ LDHB ↑ FIBA ↓ HBB ↑ PLSL ↑ CO9 ↑ LG3BP ↑ HPT ↓ CLUS ↑ LG3BP ↑
FIBA ↑ CAH2 ↑ LG3BP ↑ FINC ↓ HBD ↑ PON1 ↑ CPN2 ↑ LV151 ↑ HPTR ↓ CO2 ↑ LUM ↑
FINC ↑ CAMP ↑ LUM ↑ HABP2 ↓ HV349 ↑ PRDX2 ↑ CRIS3 ↑ LV211 ↑ HRG ↓ CO4A ↑ LV140 ↑
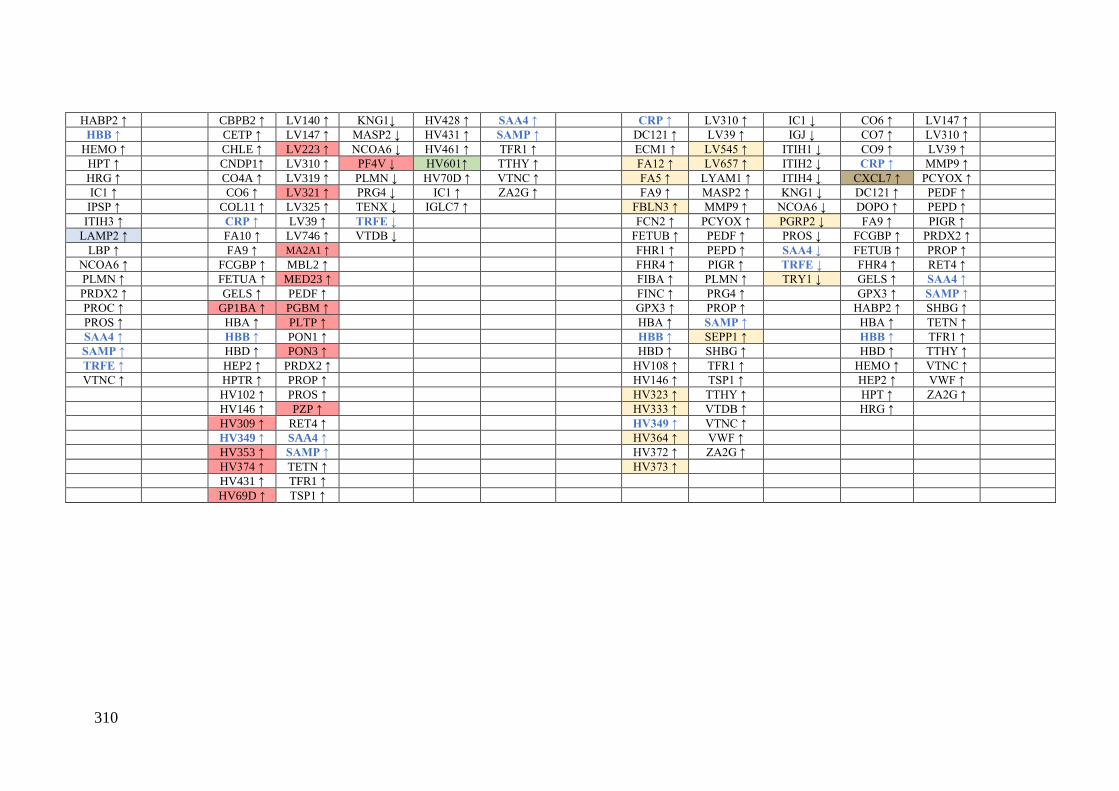
310
HABP2 ↑ CBPB2 ↑ LV140 ↑ KNG1↓ HV428 ↑ SAA4 ↑ CRP ↑ LV310 ↑ IC1 ↓ CO6 ↑ LV147 ↑
HBB ↑ CETP ↑ LV147 ↑ MASP2 ↓ HV431 ↑ SAMP ↑ DC121 ↑ LV39 ↑ IGJ ↓ CO7 ↑ LV310 ↑
HEMO ↑ CHLE ↑ LV223 ↑ NCOA6 ↓ HV461 ↑ TFR1 ↑ ECM1 ↑ LV545 ↑ ITIH1 ↓ CO9 ↑ LV39 ↑
HPT ↑ CNDP1↑ LV310 ↑ PF4V ↓ HV601↑ TTHY ↑ FA12 ↑ LV657 ↑ ITIH2 ↓ CRP ↑ MMP9 ↑
HRG ↑ CO4A ↑ LV319 ↑ PLMN ↓ HV70D ↑ VTNC ↑ FA5 ↑ LYAM1 ↑ ITIH4 ↓ CXCL7 ↑ PCYOX ↑
IC1 ↑ CO6 ↑ LV321 ↑ PRG4 ↓ IC1 ↑ ZA2G ↑ FA9 ↑ MASP2 ↑ KNG1 ↓ DC121 ↑ PEDF ↑
IPSP ↑ COL11 ↑ LV325 ↑ TENX ↓ IGLC7 ↑ FBLN3 ↑ MMP9 ↑ NCOA6 ↓ DOPO ↑ PEPD ↑
ITIH3 ↑ CRP ↑ LV39 ↑ TRFE ↓ FCN2 ↑ PCYOX ↑ PGRP2 ↓ FA9 ↑ PIGR ↑
LAMP2 ↑ FA10 ↑ LV746 ↑ VTDB ↓ FETUB ↑ PEDF ↑ PROS ↓ FCGBP ↑ PRDX2 ↑
LBP ↑ FA9 ↑ MA2A1 ↑ FHR1 ↑ PEPD ↑ SAA4 ↓ FETUB ↑ PROP ↑
NCOA6 ↑ FCGBP ↑ MBL2 ↑ FHR4 ↑ PIGR ↑ TRFE ↓ FHR4 ↑ RET4 ↑
PLMN ↑ FETUA ↑ MED23 ↑ FIBA ↑ PLMN ↑ TRY1 ↓ GELS ↑ SAA4 ↑
PRDX2 ↑ GELS ↑ PEDF ↑ FINC ↑ PRG4 ↑ GPX3 ↑ SAMP ↑
PROC ↑ GP1BA ↑ PGBM ↑ GPX3 ↑ PROP ↑ HABP2 ↑ SHBG ↑
PROS ↑ HBA ↑ PLTP ↑ HBA ↑ SAMP ↑ HBA ↑ TETN ↑
SAA4 ↑ HBB ↑ PON1 ↑ HBB ↑ SEPP1 ↑ HBB ↑ TFR1 ↑
SAMP ↑ HBD ↑ PON3 ↑ HBD ↑ SHBG ↑ HBD ↑ TTHY ↑
TRFE ↑ HEP2 ↑ PRDX2 ↑ HV108 ↑ TFR1 ↑ HEMO ↑ VTNC ↑
VTNC ↑ HPTR ↑ PROP ↑ HV146 ↑ TSP1 ↑ HEP2 ↑ VWF ↑
HV102 ↑ PROS ↑ HV323 ↑ TTHY ↑ HPT ↑ ZA2G ↑
HV146 ↑ PZP ↑ HV333 ↑ VTDB ↑ HRG ↑
HV309 ↑ RET4 ↑ HV349 ↑ VTNC ↑
HV349 ↑ SAA4 ↑ HV364 ↑ VWF ↑
HV353 ↑ SAMP ↑ HV372 ↑ ZA2G ↑
HV374 ↑ TETN ↑ HV373 ↑
HV431 ↑ TFR1 ↑
HV69D ↑ TSP1 ↑
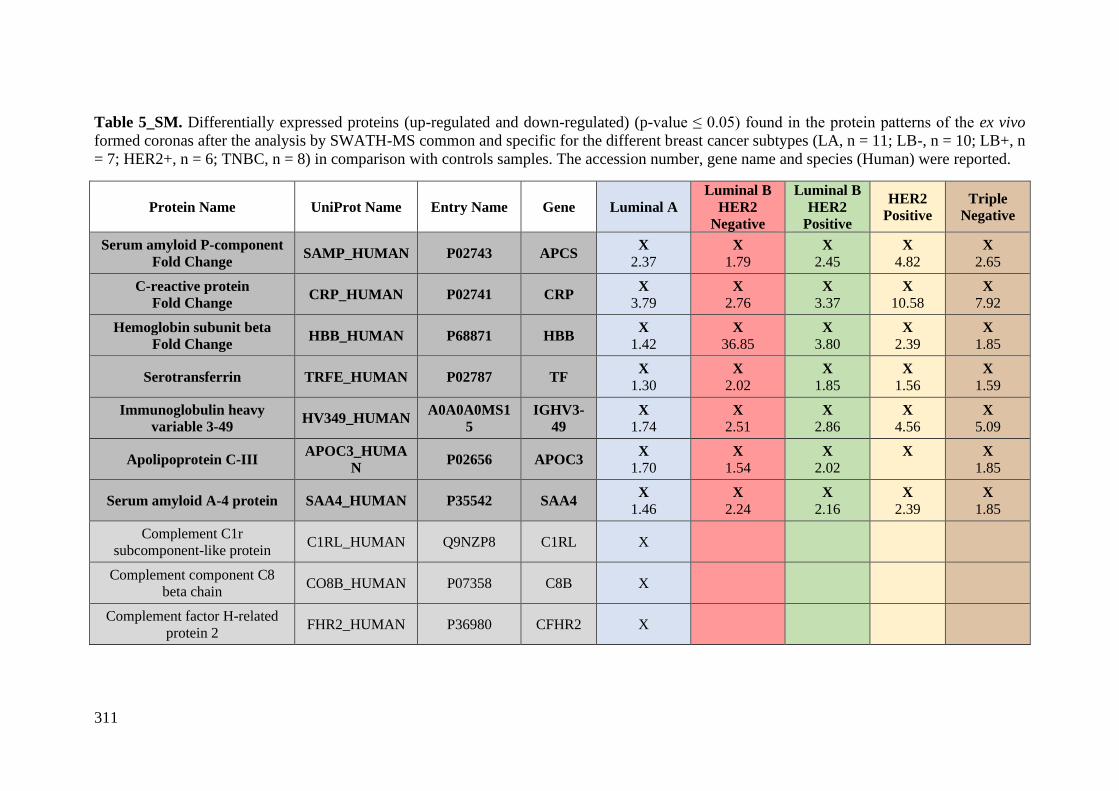
311
Table 5_SM. Differentially expressed proteins (up-regulated and down-regulated) (p-value ≤ 0.05) found in the protein patterns of the ex vivo
formed coronas after the analysis by SWATH-MS common and specific for the different breast cancer subtypes (LA, n = 11; LB-, n = 10; LB+, n
= 7; HER2+, n = 6; TNBC, n = 8) in comparison with controls samples. The accession number, gene name and species (Human) were reported.
Protein Name UniProt Name Entry Name Gene Luminal A
Luminal B
HER2
Negative
Luminal B
HER2
Positive
HER2
Positive
Triple
Negative
Serum amyloid P-component
Fold Change SAMP_HUMAN P02743 APCS
X
2.37
X
1.79
X
2.45
X
4.82
X
2.65
C-reactive protein
Fold Change CRP_HUMAN P02741 CRP
X
3.79
X
2.76
X
3.37
X
10.58
X
7.92
Hemoglobin subunit beta
Fold Change HBB_HUMAN P68871 HBB
X
1.42
X
36.85
X
3.80
X
2.39
X
1.85
Serotransferrin TRFE_HUMAN P02787 TF X
1.30
X
2.02
X
1.85
X
1.56
X
1.59
Immunoglobulin heavy
variable 3-49 HV349_HUMAN
A0A0A0MS1
5
IGHV3-
49
X
1.74
X
2.51
X
2.86
X
4.56
X
5.09
Apolipoprotein C-III APOC3_HUMA
N P02656 APOC3
X
1.70
X
1.54
X
2.02
X
X
1.85
Serum amyloid A-4 protein SAA4_HUMAN P35542 SAA4 X
1.46
X
2.24
X
2.16
X
2.39
X
1.85
Complement C1r
subcomponent-like protein C1RL_HUMAN Q9NZP8 C1RL X
Complement component C8
beta chain CO8B_HUMAN P07358 C8B X
Complement factor H-related
protein 2 FHR2_HUMAN P36980 CFHR2 X
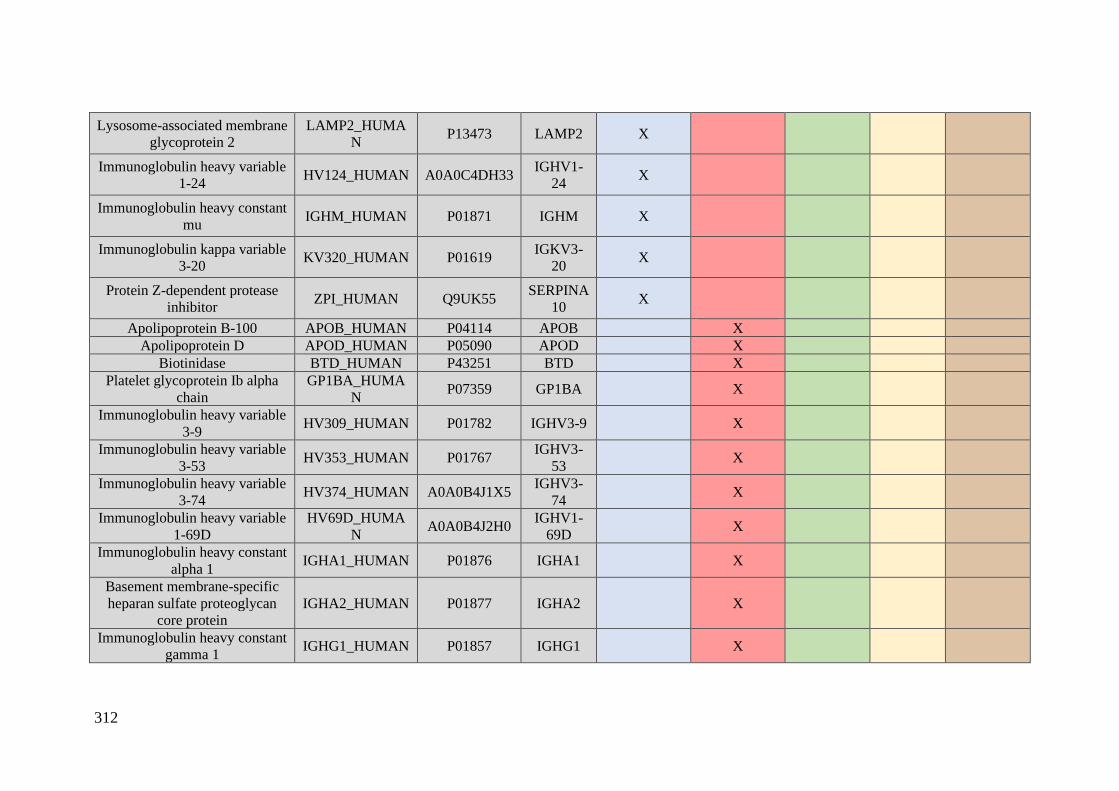
312
Lysosome-associated membrane
glycoprotein 2
LAMP2_HUMA
N P13473 LAMP2 X
Immunoglobulin heavy variable
1-24 HV124_HUMAN A0A0C4DH33
IGHV1-
24 X
Immunoglobulin heavy constant
mu IGHM_HUMAN P01871 IGHM X
Immunoglobulin kappa variable
3-20 KV320_HUMAN P01619
IGKV3-
20 X
Protein Z-dependent protease
inhibitor ZPI_HUMAN Q9UK55
SERPINA
10 X
Apolipoprotein B-100 APOB_HUMAN P04114 APOB X
Apolipoprotein D APOD_HUMAN P05090 APOD X
Biotinidase BTD_HUMAN P43251 BTD X
Platelet glycoprotein Ib alpha
chain
GP1BA_HUMA
N P07359 GP1BA X
Immunoglobulin heavy variable
3-9 HV309_HUMAN P01782 IGHV3-9 X
Immunoglobulin heavy variable
3-53 HV353_HUMAN P01767
IGHV3-
53 X
Immunoglobulin heavy variable
3-74 HV374_HUMAN A0A0B4J1X5
IGHV3-
74 X
Immunoglobulin heavy variable
1-69D
HV69D_HUMA
N A0A0B4J2H0
IGHV1-
69D X
Immunoglobulin heavy constant
alpha 1 IGHA1_HUMAN P01876 IGHA1 X
Basement membrane-specific
heparan sulfate proteoglycan
core protein
IGHA2_HUMAN P01877 IGHA2 X
Immunoglobulin heavy constant
gamma 1 IGHG1_HUMAN P01857 IGHG1 X

313
Immunoglobulin kappa constant IGKC_HUMAN P01834 IGKC X
Immunoglobulin lambda-like
polypeptide 5 IGLL5_HUMAN B9A064 IGLL5 X
Immunoglobulin kappa variable
3-11 KV311_HUMAN P04433
IGKV3-
11 X
Immunoglobulin kappa variable
4-1 KV401_HUMAN P06312 IGKV4-1 X
Immunoglobulin kappa variable
3D-20
KVD20_HUMA
N A0A0C4DH25
IGKV3D-
20 X
L-lactate dehydrogenase B
chain LDHB_HUMAN P07195 LDHB X
Immunoglobulin lambda
variable 2-23 LV223_HUMAN P01705
IGLV2-
23 X
Immunoglobulin lambda
variable 3-21 LV321_HUMAN P80748
IGLV3-
21 X
Alpha-mannosidase 2 MA2A1_HUMA
N Q16706 MAN2A1 X
Mediator of RNA polymerase II
transcription subunit 23
MED23_HUMA
N Q9ULK4 MED23 X
Basement membrane-specific
heparan sulfate proteoglycan
core protein
PGBM_HUMAN P98160 HSPG2 X
Phospholipid transfer protein PLTP_HUMAN P55058 PLTP X
Serum paraoxonase/lactonase 3 PON3_HUMAN Q15166 PON3 X
Pregnancy zone protein PZP_HUMAN P20742 PZP X
Complement C1s
subcomponent C1S_HUMAN P09871 C1S X
Platelet factor 4 variant PF4V_HUMAN P10720 PF4V1 X
Immunoglobulin heavy variable
6-1 HV601_HUMAN A0A0B4J1U7 IGHV6-1 X
Plastin-2 PLSL_HUMAN P13796 LCP1 X
Adiponectin ADIPO_HUMAN Q15848 ADIPOQ X
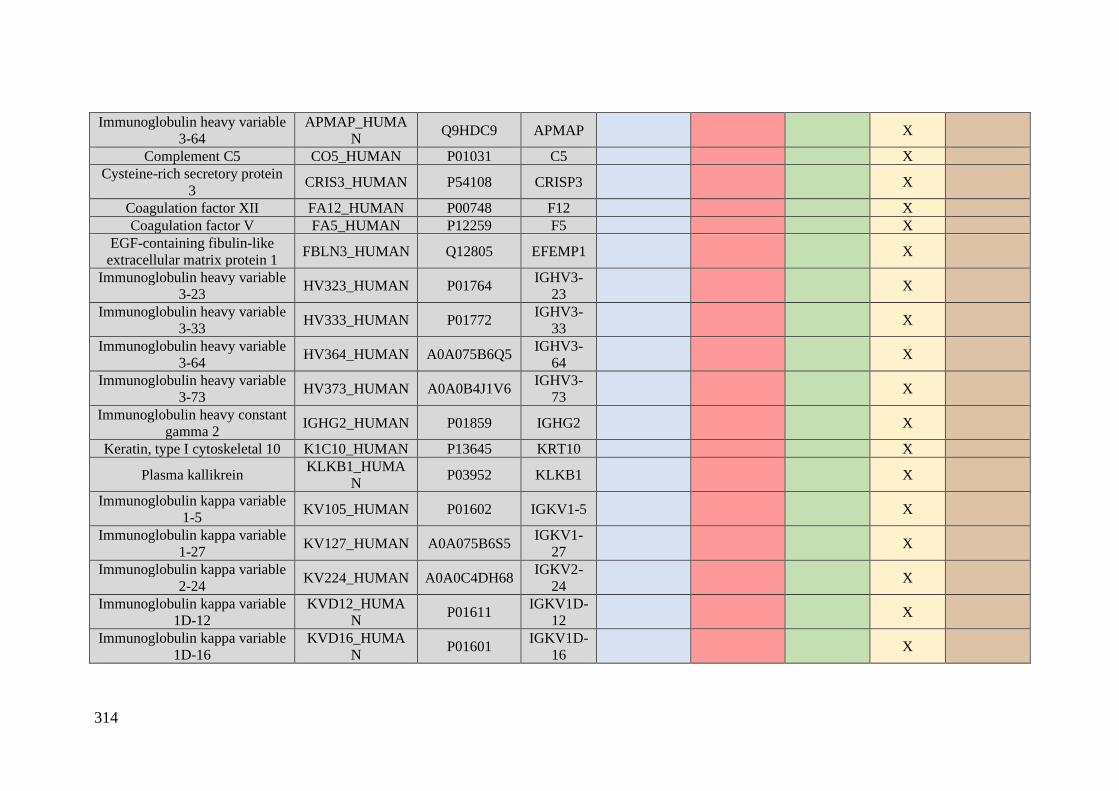
314
Immunoglobulin heavy variable
3-64
APMAP_HUMA
N Q9HDC9 APMAP X
Complement C5 CO5_HUMAN P01031 C5 X
Cysteine-rich secretory protein
3 CRIS3_HUMAN P54108 CRISP3 X
Coagulation factor XII FA12_HUMAN P00748 F12 X
Coagulation factor V FA5_HUMAN P12259 F5 X
EGF-containing fibulin-like
extracellular matrix protein 1 FBLN3_HUMAN Q12805 EFEMP1 X
Immunoglobulin heavy variable
3-23 HV323_HUMAN P01764
IGHV3-
23 X
Immunoglobulin heavy variable
3-33 HV333_HUMAN P01772
IGHV3-
33 X
Immunoglobulin heavy variable
3-64 HV364_HUMAN A0A075B6Q5
IGHV3-
64 X
Immunoglobulin heavy variable
3-73 HV373_HUMAN A0A0B4J1V6
IGHV3-
73 X
Immunoglobulin heavy constant
gamma 2 IGHG2_HUMAN P01859 IGHG2 X
Keratin, type I cytoskeletal 10 K1C10_HUMAN P13645 KRT10 X
Plasma kallikrein KLKB1_HUMA
N P03952 KLKB1 X
Immunoglobulin kappa variable
1-5 KV105_HUMAN P01602 IGKV1-5 X
Immunoglobulin kappa variable
1-27 KV127_HUMAN A0A075B6S5
IGKV1-
27 X
Immunoglobulin kappa variable
2-24 KV224_HUMAN A0A0C4DH68
IGKV2-
24 X
Immunoglobulin kappa variable
1D-12
KVD12_HUMA
N P01611
IGKV1D-
12 X
Immunoglobulin kappa variable
1D-16
KVD16_HUMA
N P01601
IGKV1D-
16 X
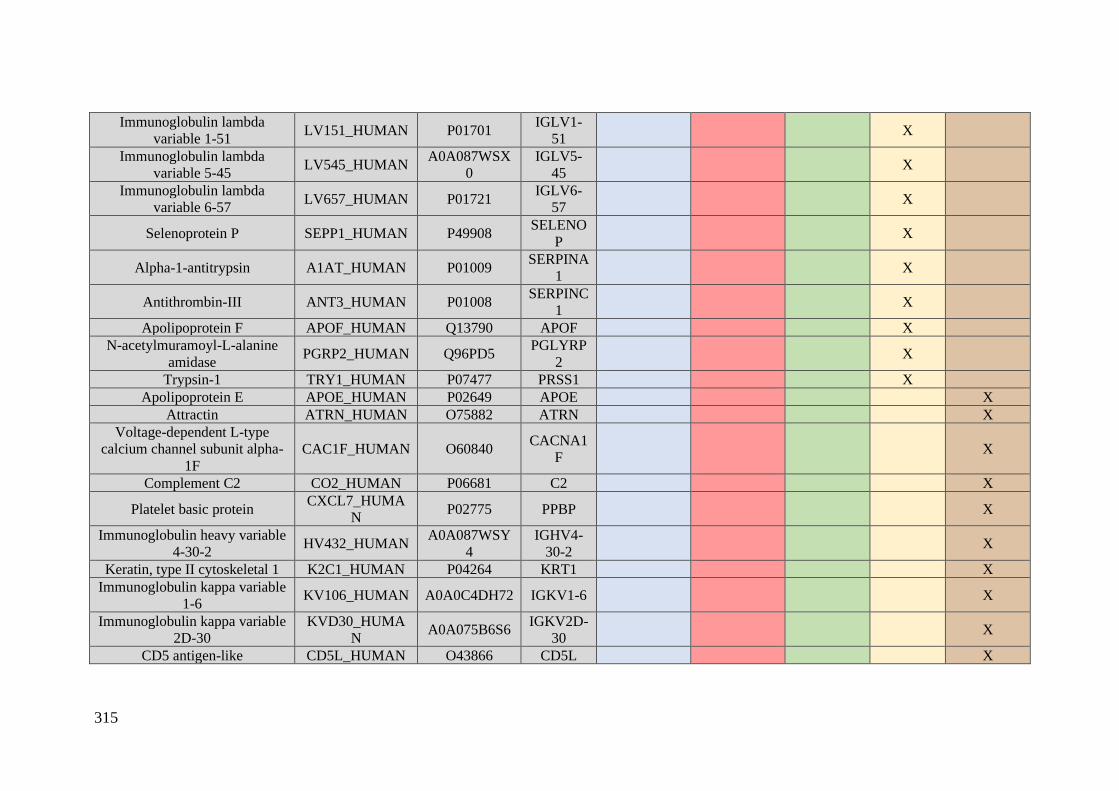
315
Immunoglobulin lambda
variable 1-51 LV151_HUMAN P01701
IGLV1-
51 X
Immunoglobulin lambda
variable 5-45 LV545_HUMAN
A0A087WSX
0
IGLV5-
45 X
Immunoglobulin lambda
variable 6-57 LV657_HUMAN P01721
IGLV6-
57 X
Selenoprotein P SEPP1_HUMAN P49908 SELENO
P X
Alpha-1-antitrypsin A1AT_HUMAN P01009 SERPINA
1 X
Antithrombin-III ANT3_HUMAN P01008 SERPINC
1 X
Apolipoprotein F APOF_HUMAN Q13790 APOF X
N-acetylmuramoyl-L-alanine
amidase PGRP2_HUMAN Q96PD5
PGLYRP
2 X
Trypsin-1 TRY1_HUMAN P07477 PRSS1 X
Apolipoprotein E APOE_HUMAN P02649 APOE X Attractin ATRN_HUMAN O75882 ATRN X
Voltage-dependent L-type
calcium channel subunit alpha-
1F
CAC1F_HUMAN O60840 CACNA1
F X
Complement C2 CO2_HUMAN P06681 C2 X
Platelet basic protein CXCL7_HUMA
N P02775 PPBP X
Immunoglobulin heavy variable
4-30-2 HV432_HUMAN
A0A087WSY
4
IGHV4-
30-2 X
Keratin, type II cytoskeletal 1 K2C1_HUMAN P04264 KRT1 X Immunoglobulin kappa variable
1-6 KV106_HUMAN A0A0C4DH72 IGKV1-6 X
Immunoglobulin kappa variable
2D-30
KVD30_HUMA
N A0A075B6S6
IGKV2D-
30 X
CD5 antigen-like CD5L_HUMAN O43866 CD5L X
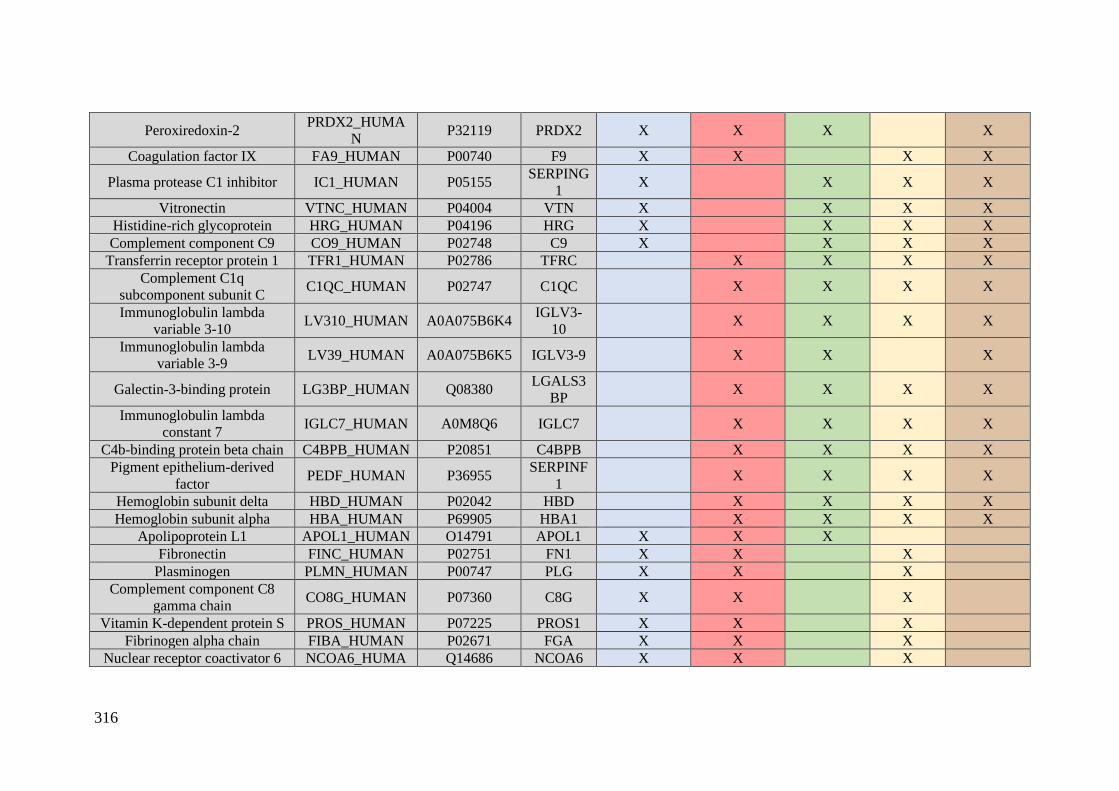
316
Peroxiredoxin-2 PRDX2_HUMA
N P32119 PRDX2 X X X X
Coagulation factor IX FA9_HUMAN P00740 F9 X X X X
Plasma protease C1 inhibitor IC1_HUMAN P05155 SERPING
1 X X X X
Vitronectin VTNC_HUMAN P04004 VTN X X X X
Histidine-rich glycoprotein HRG_HUMAN P04196 HRG X X X X
Complement component C9 CO9_HUMAN P02748 C9 X X X X
Transferrin receptor protein 1 TFR1_HUMAN P02786 TFRC X X X X
Complement C1q
subcomponent subunit C C1QC_HUMAN P02747 C1QC X X X X
Immunoglobulin lambda
variable 3-10 LV310_HUMAN A0A075B6K4
IGLV3-
10 X X X X
Immunoglobulin lambda
variable 3-9 LV39_HUMAN A0A075B6K5 IGLV3-9 X X X
Galectin-3-binding protein LG3BP_HUMAN Q08380 LGALS3
BP X X X X
Immunoglobulin lambda
constant 7 IGLC7_HUMAN A0M8Q6 IGLC7 X X X X
C4b-binding protein beta chain C4BPB_HUMAN P20851 C4BPB X X X X
Pigment epithelium-derived
factor PEDF_HUMAN P36955
SERPINF
1 X X X X
Hemoglobin subunit delta HBD_HUMAN P02042 HBD X X X X
Hemoglobin subunit alpha HBA_HUMAN P69905 HBA1 X X X X
Apolipoprotein L1 APOL1_HUMAN O14791 APOL1 X X X
Fibronectin FINC_HUMAN P02751 FN1 X X X
Plasminogen PLMN_HUMAN P00747 PLG X X X
Complement component C8
gamma chain CO8G_HUMAN P07360 C8G X X X
Vitamin K-dependent protein S PROS_HUMAN P07225 PROS1 X X X
Fibrinogen alpha chain FIBA_HUMAN P02671 FGA X X X
Nuclear receptor coactivator 6 NCOA6_HUMA Q14686 NCOA6 X X X
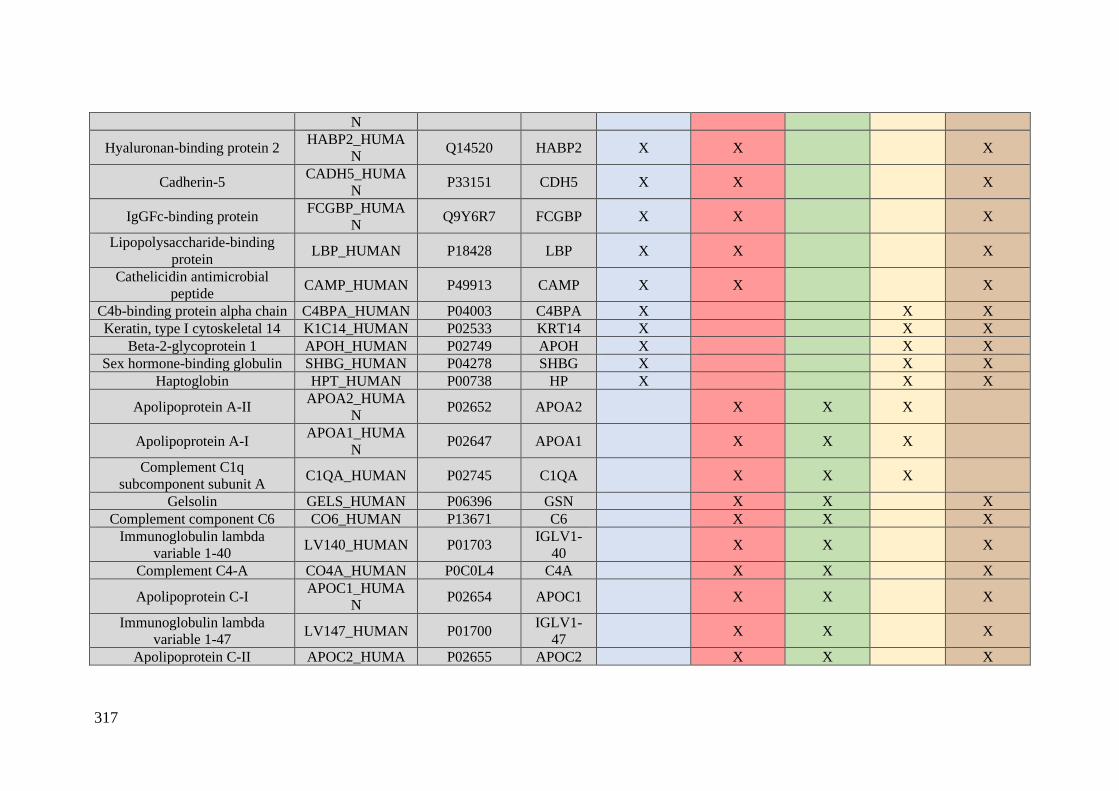
317
N
Hyaluronan-binding protein 2 HABP2_HUMA
N Q14520 HABP2 X X X
Cadherin-5 CADH5_HUMA
N P33151 CDH5 X X X
IgGFc-binding protein FCGBP_HUMA
N Q9Y6R7 FCGBP X X X
Lipopolysaccharide-binding
protein LBP_HUMAN P18428 LBP X X X
Cathelicidin antimicrobial
peptide CAMP_HUMAN P49913 CAMP X X X
C4b-binding protein alpha chain C4BPA_HUMAN P04003 C4BPA X X X
Keratin, type I cytoskeletal 14 K1C14_HUMAN P02533 KRT14 X X X
Beta-2-glycoprotein 1 APOH_HUMAN P02749 APOH X X X
Sex hormone-binding globulin SHBG_HUMAN P04278 SHBG X X X
Haptoglobin HPT_HUMAN P00738 HP X X X
Apolipoprotein A-II APOA2_HUMA
N P02652 APOA2 X X X
Apolipoprotein A-I APOA1_HUMA
N P02647 APOA1 X X X
Complement C1q
subcomponent subunit A C1QA_HUMAN P02745 C1QA X X X
Gelsolin GELS_HUMAN P06396 GSN X X X
Complement component C6 CO6_HUMAN P13671 C6 X X X
Immunoglobulin lambda
variable 1-40 LV140_HUMAN P01703
IGLV1-
40 X X X
Complement C4-A CO4A_HUMAN P0C0L4 C4A X X X
Apolipoprotein C-I APOC1_HUMA
N P02654 APOC1 X X X
Immunoglobulin lambda
variable 1-47 LV147_HUMAN P01700
IGLV1-
47 X X X
Apolipoprotein C-II APOC2_HUMA P02655 APOC2 X X X
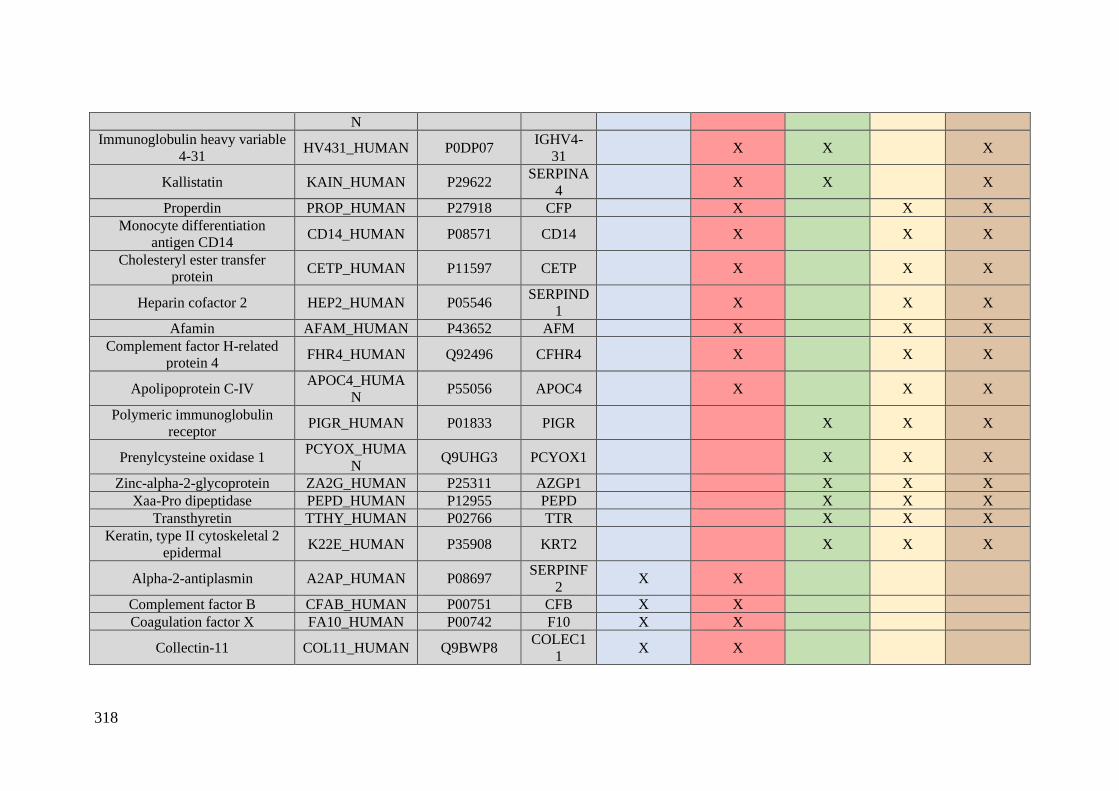
318
N
Immunoglobulin heavy variable
4-31 HV431_HUMAN P0DP07
IGHV4-
31 X X X
Kallistatin KAIN_HUMAN P29622 SERPINA
4 X X X
Properdin PROP_HUMAN P27918 CFP X X X
Monocyte differentiation
antigen CD14 CD14_HUMAN P08571 CD14 X X X
Cholesteryl ester transfer
protein CETP_HUMAN P11597 CETP X X X
Heparin cofactor 2 HEP2_HUMAN P05546 SERPIND
1 X X X
Afamin AFAM_HUMAN P43652 AFM X X X
Complement factor H-related
protein 4 FHR4_HUMAN Q92496 CFHR4 X X X
Apolipoprotein C-IV APOC4_HUMA
N P55056 APOC4 X X X
Polymeric immunoglobulin
receptor PIGR_HUMAN P01833 PIGR X X X
Prenylcysteine oxidase 1 PCYOX_HUMA
N Q9UHG3 PCYOX1 X X X
Zinc-alpha-2-glycoprotein ZA2G_HUMAN P25311 AZGP1 X X X
Xaa-Pro dipeptidase PEPD_HUMAN P12955 PEPD X X X
Transthyretin TTHY_HUMAN P02766 TTR X X X
Keratin, type II cytoskeletal 2
epidermal K22E_HUMAN P35908 KRT2 X X X
Alpha-2-antiplasmin A2AP_HUMAN P08697 SERPINF
2 X X
Complement factor B CFAB_HUMAN P00751 CFB X X
Coagulation factor X FA10_HUMAN P00742 F10 X X
Collectin-11 COL11_HUMAN Q9BWP8 COLEC1
1 X X
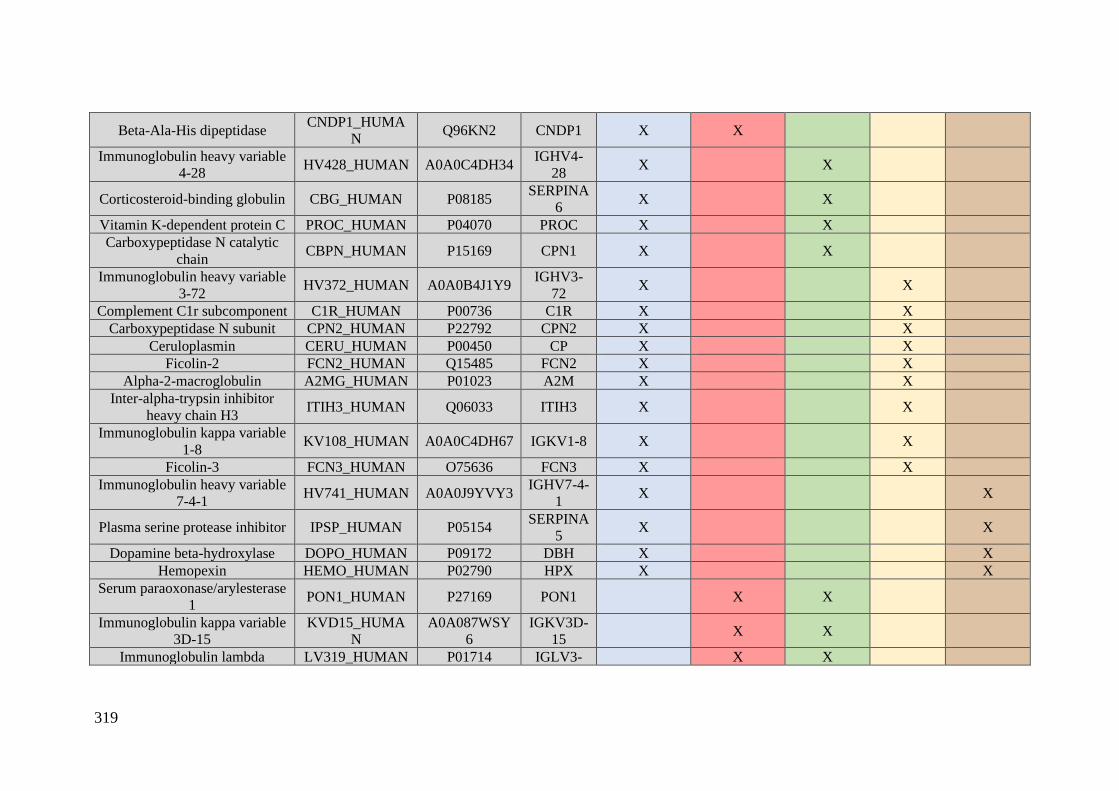
319
Beta-Ala-His dipeptidase CNDP1_HUMA
N Q96KN2 CNDP1 X X
Immunoglobulin heavy variable
4-28 HV428_HUMAN A0A0C4DH34
IGHV4-
28 X X
Corticosteroid-binding globulin CBG_HUMAN P08185 SERPINA
6 X X
Vitamin K-dependent protein C PROC_HUMAN P04070 PROC X X
Carboxypeptidase N catalytic
chain CBPN_HUMAN P15169 CPN1 X X
Immunoglobulin heavy variable
3-72 HV372_HUMAN A0A0B4J1Y9
IGHV3-
72 X X
Complement C1r subcomponent C1R_HUMAN P00736 C1R X X
Carboxypeptidase N subunit CPN2_HUMAN P22792 CPN2 X X
Ceruloplasmin CERU_HUMAN P00450 CP X X
Ficolin-2 FCN2_HUMAN Q15485 FCN2 X X
Alpha-2-macroglobulin A2MG_HUMAN P01023 A2M X X
Inter-alpha-trypsin inhibitor
heavy chain H3 ITIH3_HUMAN Q06033 ITIH3 X X
Immunoglobulin kappa variable
1-8 KV108_HUMAN A0A0C4DH67 IGKV1-8 X X
Ficolin-3 FCN3_HUMAN O75636 FCN3 X X
Immunoglobulin heavy variable
7-4-1 HV741_HUMAN A0A0J9YVY3
IGHV7-4-
1 X X
Plasma serine protease inhibitor IPSP_HUMAN P05154 SERPINA
5 X X
Dopamine beta-hydroxylase DOPO_HUMAN P09172 DBH X X
Hemopexin HEMO_HUMAN P02790 HPX X X
Serum paraoxonase/arylesterase
1 PON1_HUMAN P27169 PON1 X X
Immunoglobulin kappa variable
3D-15
KVD15_HUMA
N
A0A087WSY
6
IGKV3D-
15 X X
Immunoglobulin lambda LV319_HUMAN P01714 IGLV3- X X
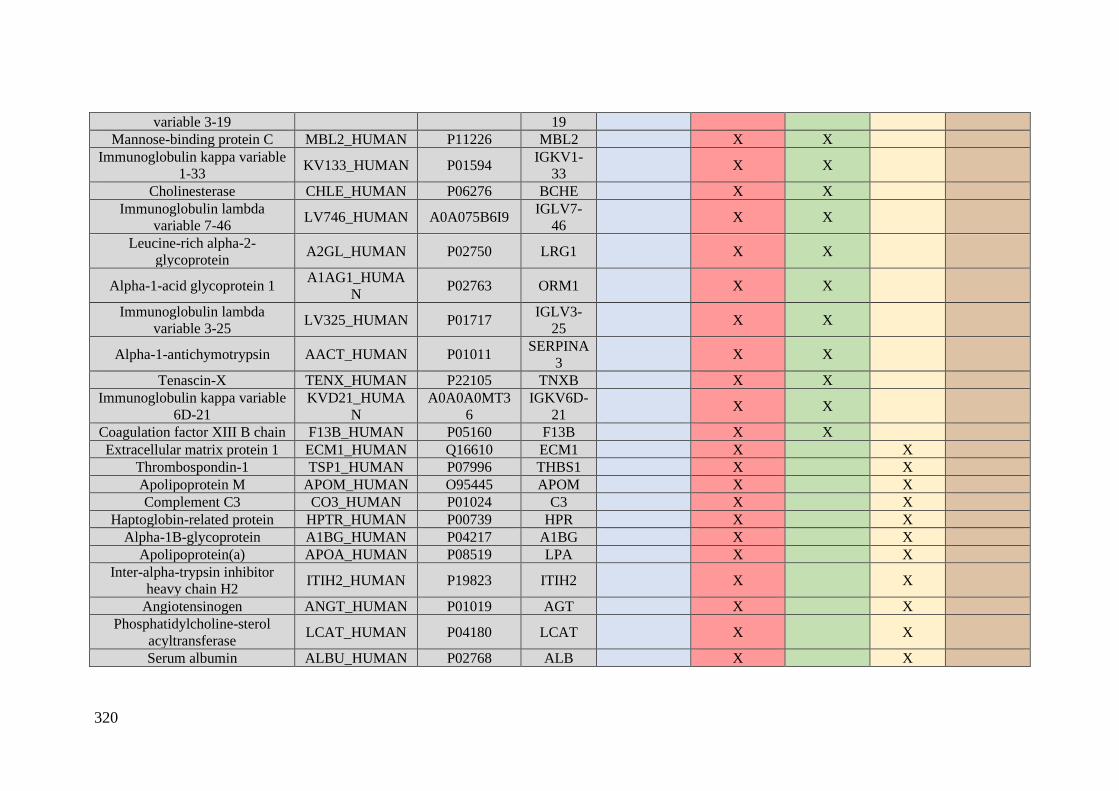
320
variable 3-19 19
Mannose-binding protein C MBL2_HUMAN P11226 MBL2 X X
Immunoglobulin kappa variable
1-33 KV133_HUMAN P01594
IGKV1-
33 X X
Cholinesterase CHLE_HUMAN P06276 BCHE X X
Immunoglobulin lambda
variable 7-46 LV746_HUMAN A0A075B6I9
IGLV7-
46 X X
Leucine-rich alpha-2-
glycoprotein A2GL_HUMAN P02750 LRG1 X X
Alpha-1-acid glycoprotein 1 A1AG1_HUMA
N P02763 ORM1 X X
Immunoglobulin lambda
variable 3-25 LV325_HUMAN P01717
IGLV3-
25 X X
Alpha-1-antichymotrypsin AACT_HUMAN P01011 SERPINA
3 X X
Tenascin-X TENX_HUMAN P22105 TNXB X X
Immunoglobulin kappa variable
6D-21
KVD21_HUMA
N
A0A0A0MT3
6
IGKV6D-
21 X X
Coagulation factor XIII B chain F13B_HUMAN P05160 F13B X X
Extracellular matrix protein 1 ECM1_HUMAN Q16610 ECM1 X X
Thrombospondin-1 TSP1_HUMAN P07996 THBS1 X X
Apolipoprotein M APOM_HUMAN O95445 APOM X X
Complement C3 CO3_HUMAN P01024 C3 X X
Haptoglobin-related protein HPTR_HUMAN P00739 HPR X X
Alpha-1B-glycoprotein A1BG_HUMAN P04217 A1BG X X
Apolipoprotein(a) APOA_HUMAN P08519 LPA X X
Inter-alpha-trypsin inhibitor
heavy chain H2 ITIH2_HUMAN P19823 ITIH2 X X
Angiotensinogen ANGT_HUMAN P01019 AGT X X
Phosphatidylcholine-sterol
acyltransferase LCAT_HUMAN P04180 LCAT X X
Serum albumin ALBU_HUMAN P02768 ALB X X
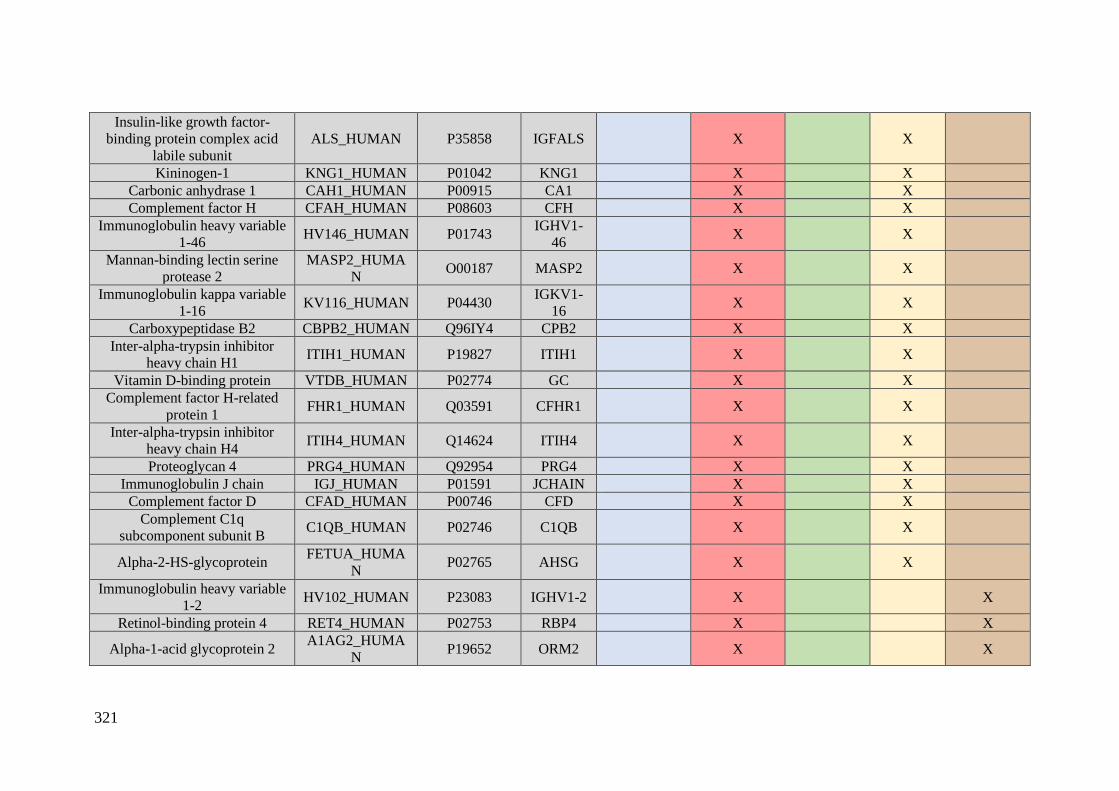
321
Insulin-like growth factor-
binding protein complex acid
labile subunit
ALS_HUMAN P35858 IGFALS X X
Kininogen-1 KNG1_HUMAN P01042 KNG1 X X
Carbonic anhydrase 1 CAH1_HUMAN P00915 CA1 X X
Complement factor H CFAH_HUMAN P08603 CFH X X
Immunoglobulin heavy variable
1-46 HV146_HUMAN P01743
IGHV1-
46 X X
Mannan-binding lectin serine
protease 2
MASP2_HUMA
N O00187 MASP2 X X
Immunoglobulin kappa variable
1-16 KV116_HUMAN P04430
IGKV1-
16 X X
Carboxypeptidase B2 CBPB2_HUMAN Q96IY4 CPB2 X X
Inter-alpha-trypsin inhibitor
heavy chain H1 ITIH1_HUMAN P19827 ITIH1 X X
Vitamin D-binding protein VTDB_HUMAN P02774 GC X X
Complement factor H-related
protein 1 FHR1_HUMAN Q03591 CFHR1 X X
Inter-alpha-trypsin inhibitor
heavy chain H4 ITIH4_HUMAN Q14624 ITIH4 X X
Proteoglycan 4 PRG4_HUMAN Q92954 PRG4 X X
Immunoglobulin J chain IGJ_HUMAN P01591 JCHAIN X X
Complement factor D CFAD_HUMAN P00746 CFD X X
Complement C1q
subcomponent subunit B C1QB_HUMAN P02746 C1QB X X
Alpha-2-HS-glycoprotein FETUA_HUMA
N P02765 AHSG X X
Immunoglobulin heavy variable
1-2 HV102_HUMAN P23083 IGHV1-2 X X
Retinol-binding protein 4 RET4_HUMAN P02753 RBP4 X X
Alpha-1-acid glycoprotein 2 A1AG2_HUMA
N P19652 ORM2 X X
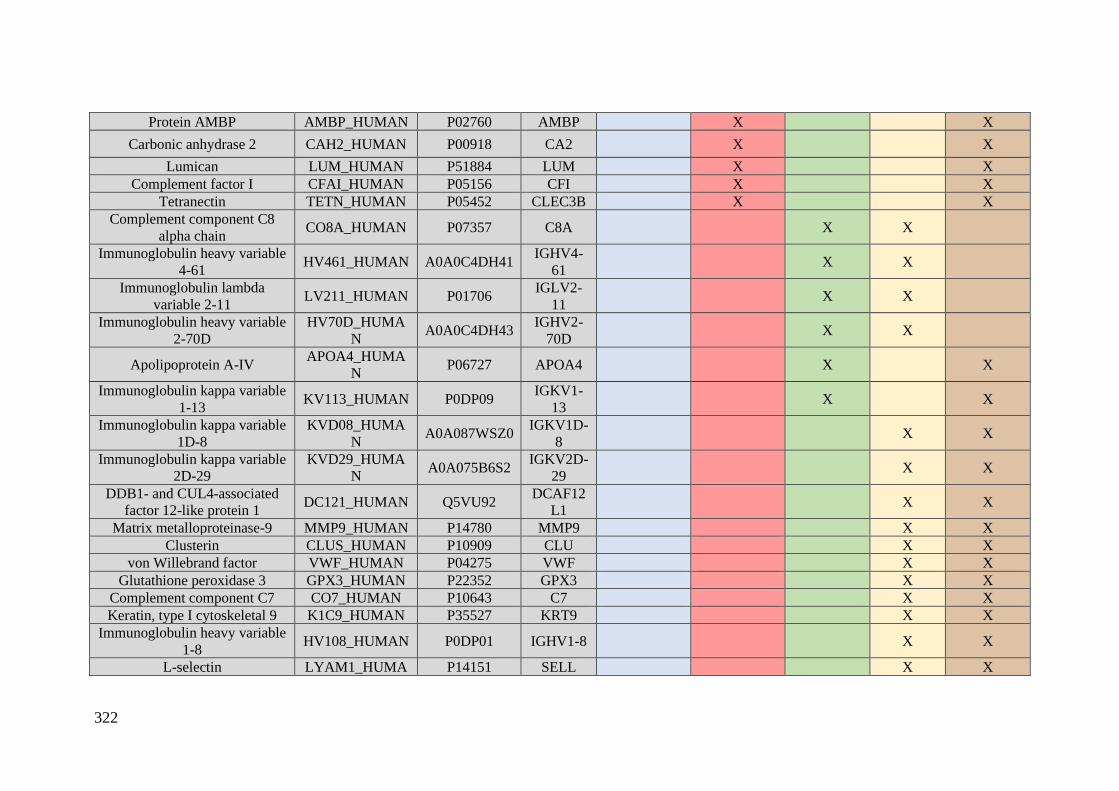
322
Protein AMBP AMBP_HUMAN P02760 AMBP X X
Carbonic anhydrase 2 CAH2_HUMAN P00918 CA2 X X
Lumican LUM_HUMAN P51884 LUM X X Complement factor I CFAI_HUMAN P05156 CFI X X
Tetranectin TETN_HUMAN P05452 CLEC3B X X Complement component C8
alpha chain CO8A_HUMAN P07357 C8A X X
Immunoglobulin heavy variable
4-61 HV461_HUMAN A0A0C4DH41
IGHV4-
61 X X
Immunoglobulin lambda
variable 2-11 LV211_HUMAN P01706
IGLV2-
11 X X
Immunoglobulin heavy variable
2-70D
HV70D_HUMA
N A0A0C4DH43
IGHV2-
70D X X
Apolipoprotein A-IV APOA4_HUMA
N P06727 APOA4 X X
Immunoglobulin kappa variable
1-13 KV113_HUMAN P0DP09
IGKV1-
13 X X
Immunoglobulin kappa variable
1D-8
KVD08_HUMA
N A0A087WSZ0
IGKV1D-
8 X X
Immunoglobulin kappa variable
2D-29
KVD29_HUMA
N A0A075B6S2
IGKV2D-
29 X X
DDB1- and CUL4-associated
factor 12-like protein 1 DC121_HUMAN Q5VU92
DCAF12
L1 X X
Matrix metalloproteinase-9 MMP9_HUMAN P14780 MMP9 X X
Clusterin CLUS_HUMAN P10909 CLU X X
von Willebrand factor VWF_HUMAN P04275 VWF X X
Glutathione peroxidase 3 GPX3_HUMAN P22352 GPX3 X X
Complement component C7 CO7_HUMAN P10643 C7 X X
Keratin, type I cytoskeletal 9 K1C9_HUMAN P35527 KRT9 X X
Immunoglobulin heavy variable
1-8 HV108_HUMAN P0DP01 IGHV1-8 X X
L-selectin LYAM1_HUMA P14151 SELL X X
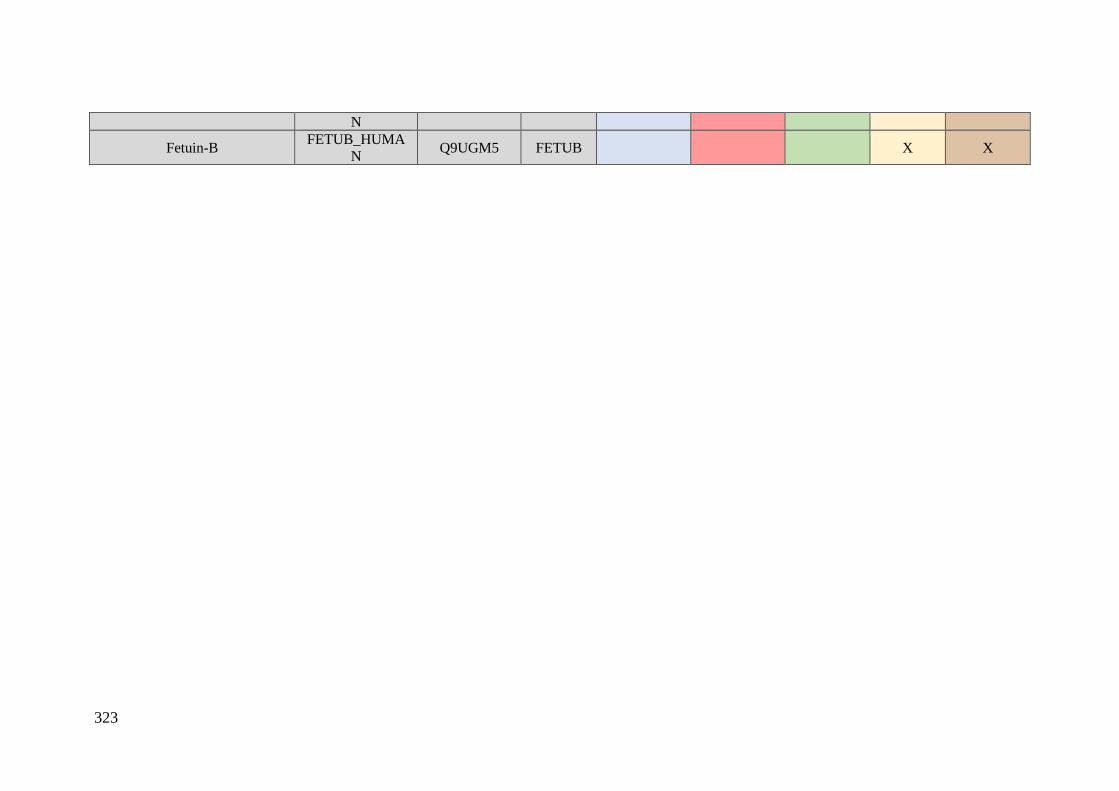
323
N
Fetuin-B FETUB_HUMA
N Q9UGM5 FETUB X X

324

ANNEX D
Extended abstract


325
Resumen extendido
El cáncer de mama es uno de los tipos de cáncer más común en mujeres y supone
aproximadamente el 14% de las muertes relacionadas con el cáncer en mujeres de todo el
mundo. La detección precoz del cáncer de mama juega un papel determinante en el
pronóstico y la tasa de supervivencia de dicha enfermedad. Hoy en día, la mamografía es
la técnica que se emplea para el diagnóstico precoz del cáncer de mama, sin embargo,
esta técnica presenta una baja sensibilidad (25-59%) para el diagnóstico en las mujeres
jóvenes.
En la práctica clínica habitual del cáncer de mama, las pruebas pronósticas
requieren la obtención de tejido tumoral mediante una biopsia u otros abordajes
quirúrgicos. Con el objetivo de minimizar dichos procedimientos invasivos, los
biomarcadores presentes en el suero/plasma sanguíneo abren una nueva vía para el
diagnóstico precoz del cáncer de mama en mujeres asintomáticas, un pronóstico más
preciso y la predicción de la respuesta al tratamiento.
A pesar de que en las última tres décadas se han evaluado varios biomarcadores
séricos, éstos carecen de la sensibilidad suficiente para detectar el cáncer de mama en una
fase inicial de la enfermedad y ninguno de estos posee la precisión suficiente para predecir
la recurrencia. Por lo tanto, es imprescindible encontrar nuevos biomarcadores
sanguíneos relacionados con el cáncer de mama.
La proteómica es una ciencia que permite identificar y cuantificar las
proteínas/péptidos presentes en una determinada muestra biológica (suero, plasma, orina),
empleando para ello la técnica de espectrometría de masas. Partiendo de que la detección
del cáncer en una fase temprana se basa en la hipótesis de que la enfermedad se desarrolla
al producirse una desviación del estado normal, la comparación de los perfiles
proteómicos de los pacientes con cáncer y los individuos sanos (controles) nos permitirá
localizar distintos biomarcadores. Además, el análisis proteómico es complementario de
los estudios genéticos que se desarrollan para evaluar el pronóstico del cáncer de mama.
Concretamente, centraremos esta introducción en los estudios proteómicos llevados a
cabo en el cáncer de mama para la detección precoz, el pronóstico y la evaluación de la
respuesta al tratamiento en este tipo de pacientes.

326
Existen biomarcadores tumorales circulantes (en la sangre) capaces de detectar la
existencia de malignidad en etapas previas al diagnóstico clínico y actualmente se están
evaluando en ensayos clínicos con ciertos tipos de cáncer; por ejemplo, CA125 en la
detección del cáncer de ovario. Sin embargo, actualmente no hay biomarcadores
circulantes (en la sangre) adecuados para el diagnóstico o la detección del cáncer de
mama. Aunque existen candidatos, como el antígeno carcinoembrionario (CEA), la forma
soluble de la proteína MUC1 (CA15-3, CA27.29), la proteína oncogénica RS / DJ-1, el
factor de crecimiento epidérmico humano receptor-2 (HER2) o los fragmentos circulantes
de citoqueratina (TPA, TPS y CYFRA 21-1), estos presentan algunas limitaciones tanto
en su sensibilidad como en su especificidad para el diagnóstico precoz del cáncer de
mama. Por lo tanto, en la práctica clínica estos biomarcadores se usan principalmente para
predecir la respuesta a la terapia, la monitorización después de la terapia primaria, o como
indicadores pronósticos.
De entre todos, el antígeno carbohidrato 15-3 (CA 15-3) es el marcador sérico más
utilizado en las pacientes con cáncer de mama. CA 15-3 se utiliza para el cribado de rutina
del cáncer de mama, así como para el control y el seguimiento de pacientes [Clin. Chim.
Acta 411 (2010) 1869-1874]. El valor medio de CA 15-3 es de 17 U/mL (rango 3.9-99.5
U/mL) en pacientes con cáncer de mama primario no tratado.
Con el fin de desarrollar estrategias novedosas para la detección ultrasensible de
CA 15-3, se desarrolló un inmunosensor electroquímico nanoestructurado basado en
óxido de grafeno y funcionalizado no covalentemente (GO/Py-COOH), así como
nanotubos de carbono de pared múltiple (MWCNTs). Este método permitió la detección
de CA 15-3 en suero sanguíneo a niveles tan bajos como 0.01 ± 0.07 U/mL. Las ventajas
de este nanosistema es que es altamente selectivo y se puede regenerar para ser utilizado
múltiples veces, lo que ofrece un gran potencial para su uso en el futuro como método de
diagnóstico del cáncer de mama. En esta línea, el ácido perclórico (PCA) se usó para
mejorar la detección de proteínas O-glicosiladas en suero (como CA 27.29 y CA 15-3)
usando un ensayo de lectina enlazada con enzimas sandwich (ELLA). Al analizar
mediante electroforesis en gel de poliacrilamida (SDS-PAGE) las fracciones de
glicoproteína capturadas con la lectina champedak galactose binding (CGB) procedentes
de suero de las pacientes con cáncer de mama en estadio 0 (n = 31), estadio I (n = 48) y
controles (n = 105), se detectó una abundancia alterada inversa sustancial del inhibidor

327
de la proteasa plasmática C1 y del proteoglicano 4 en estos dos primeros estadios en
relación con el grupo control. Aunque es necesario la validación del método empleando
poblaciones clínicamente representativas, la ratio de expresión de proteoglicano 4 frente
al inhibidor de la proteasa C1 se puede explotar para la detección temprana del cáncer de
mama usando muestras de suero sanguíneo.
Además, se exploró el potencial de diferentes paneles de biomarcadores que
contienen CA15-3 para el diagnóstico temprano del cáncer de mama. Se descubrió que
un conjunto de diez posibles biomarcadores séricos de cáncer de mama y antígenos de
cáncer (haptoglobina, osteopontina (OPN), CA15-3, antígeno de cáncer 125 (CA-125),
antígeno de cáncer 19-9 (CA19-9), el antígeno carcinoembrionario (CEA), la prolactina,
la α- fetoproteína (AFP), la leptina y el factor inhibidor de la migración (MIF)) no fue útil
para predecir el cáncer de mama en fases tempranas de la enfermedad. Otro estudio
proporciona resultados similares, en el que ninguno de los 9 marcadores candidatos
utilizados (CA15-3, RANTES/CCL5 (regulados por activación de células T normales
expresadas y secretadas/ el ligando 5 de quimiocina (motivo CC)), OPN (osteopontina) ,
PAI-1 (inhibidor del activador del plasminógeno-1), SLPI (inhibidor de la proteasa
leucocitaria secretora), HSP90A (proteína de choque térmico 90A), IGFBP3 (proteína de
unión al factor de crecimiento similar a la insulina 3), APOC1 (apolipoproteína CI) y
PAPPA (pappalysin -1), o sus combinaciones, resultaron útiles para detectar el cáncer de
mama; además se encontraron vínculos entre los candidatos CA15-3, HSP90A y PAI-1
con aspectos clínico-patológicos, que, a su vez, se correlacionaban con el pronóstico.
Sin embargo, un panel de antígenos compuesto por B11 (LGALS3), B18 (PHB2),
B119 (MUC1) y B130 (GK2), junto con CA15-3, mostró un aumento significativo de la
sensibilidad (87%), la especificidad (76%) y en la supervivencia global (OS) (82,7%)
para el diagnóstico del cáncer de mama en fase T1N0M0, en comparación con la utilización
de CA15-3 sólo. Aunque este panel de antígenos requiere de ser validado utilizando más
muestras de cáncer de mama, se presenta como un procedimiento prometedor para
detectar el cáncer de mama en una fase temprana. Además, CA15-3 también se incluyó
en el panel de diagnóstico constituido por los 4 picos de un conjunto de proteínas
[ m/z 3,972, 6,850 y 8,115 (BC2) y 8,949 (BC3)] utilizadas para distinguir a 62 pacientes
con cáncer de mama con carcinoma ductal invasivo de 16 controles sanos (HCs) y 31
pacientes con enfermedades mamarias benignas (BBDs). Se demostró que este panel de
4 picos junto con CA15-3 ofrecía una buena sensibilidad y especificidad para el
diagnóstico de cáncer de mama. Cierto es que, como mencionamos anteriormente, se

328
debería realizar una investigación adicional utilizando un tamaño de muestra más grande
para poder verificar estos resultados.
También se exploró el potencial de CA15-3 en relación con el diagnóstico precoz
del cáncer de mama metastásico. Así, se investigó la sensibilidad de CA 15-3, CEA y
HER2, descubriéndose que la combinación de dos de estos marcadores tumorales
mejoraba la sensibilidad para la detección del cáncer de mama metastásico, y,
curiosamente, la determinación conjunta de los tres marcadores tumorales sólo mejoraba
levemente la sensibilidad. Los autores propusieron el uso combinado de CEA y HER2 en
tumores del subtipo HER2+ y la combinación de CA 15-3 y CEA en tumores del subtipo
HER2−. De nuevo, se requieren ensayos clínicos aleatorios prospectivos para explorar
los beneficios clínicos de la detección temprana y el tratamiento de la enfermedad
metastásica.
También se evaluó la eficacia de los biomarcadores séricos Bc1, Bc2 y Bc3 en la
detección temprana del cáncer de mama, siendo sólo la expresión de Bc2 estadísticamente
significativa entre el grupo de enfermedad maligna, el grupo control y el grupo de
enfermedad benigna.
El cáncer de mama es una enfermedad heterogénea en la que las células
cancerígenas pueden expresar una variedad de proteínas aberrantes (los denominados
antígenos asociados a tumores: TAAs) que son capaces de provocar una respuesta inmune
y producir anticuerpos. Curiosamente, esta respuesta inmune aparece meses y hasta años
antes del diagnóstico clínico de la neoplasia maligna. Así, la identificación de los TAAs
y sus anticuerpos específicos podrían ofrecer una amplificación in vivo de una señal
carcinogénica en fases muy tempranas, lo que posiblemente permita una detección mucho
más precoz del cáncer que con los métodos disponibles actualmente.
En particular, en el suero se encuentran varios antígenos y anticuerpos circulantes
que pueden relacionarse con la progresión y el desarrollo del cáncer. Así, se se han
evaluado en cáncer de mama la presencia de autoanticuerpos en suero contra antígenos
tumorales como son p53, antígenos antineurales/antinucleares, o proteínas neurales
embrionarias.
Los antígenos del cáncer han demostrado ser de una importancia extraordinaria
para el cribado, así como ser indicadores pronósticos. En particular, podemos citar a las
proteínas de choque térmico (HSPs), que se encuentran sobreexpresadas en una amplia
gama de cánceres humanos y son capaces de causar la estimulación del sistema inmune
y, en consecuencia, generar una concentración elevada de autoanticuerpos anti-HSP.

329
Estos autoanticuerpos se han asociado a metástasis tumorales en pacientes con cáncer de
mama, y, en consecuencia, su detección podría tener utilidad en el pronóstico y
diagnóstico. De esta manera, L. Shi et al. inmovilizaron siete proteínas pertenecientes a
la familia de proteínas de choque térmico (HSPB1, HSPD1, HSP70, HSP90, HSPA5,
HSP90B1) y una oncoproteína, P53, a seis superficies químicas diferentes. Se emplearon
dos superficies químicas (COOH y quitosano) para detectar autoanticuerpos antigénicos
antitumorales en 26 muestras de suero pertenecientes a donantes sanos y 50 muestras a
pacientes con cáncer de mama. La detección de un único autoanticuerpo no permitió
discriminar significativamente los sueros pertenecientes a las pacientes con cáncer de
mama de los sueros de los donantes sanos, pero la combinación de los siete
autoanticuerpos (autoanticuerpos contra HSPB1, HSPD1, HSP70, HSP90, HSPA5,
HSP90B1 y P53) aumentó la especificidad y la sensibilidad de la prueba (con una
especificidad del 100% y una sensibilidad del 86%). Este estudio ha demostrado que los
microarrays de proteínas personalizados pueden ser herramientas efectivas para la
detección rápida de miles de biomarcadores paralelamente ofreciendo un alto
rendimiento. Por supuesto, este rendimiento del microarray de proteínas estaría
influenciado por muchos parámetros, como el tampón de detección que se use, las
características químicas de la superficie utilizada, o la concentración de proteínas, lo que
requiere validar exhaustivamente los procedimientos; además, se necesitarán cohortes
más grandes de pacientes con cáncer de mama y de donantes sanos para validar su utilidad
a gran escala.
El enfoque proporcionado por la inmuno-proteómica también apoya el uso de
autoanticuerpos. Así, la presencia de autoanticuerpos en suero contra la alfa-2-
glicoproteína-HS (AHSG) podría ser útil en el diagnóstico y la detección mínimamente
invasiva del cáncer de mama en una fase temprana. Sin embargo, la determinación de
AHSG deberá ser probada y validada aún por múltiples estudios independientes que
utilicen un tamaño muestral adecuado y un conjunto de muestras de suero de pacientes
con cáncer de mama en una fase muy temprana. Además, la verificación adicional con
muestras de pacientes con carcinoma ductal in situ y cáncer de mama en estadios III y IV
ayudaría a confirmar la especificidad de los autoanticuerpos AHSG en este subconjunto
de pacientes con cáncer de mama. Cierto es que esta investigación ha proporcionado datos
preliminares de interés sobre la ventaja potencial que supone la medición de
autoanticuerpos para detectar tumores pequeños en etapas tempranas, ya que hasta el

330
momento estos autoanticuerpos solo se habían empleado en el estudio del cáncer de mama
en etapas más tardías.
Se descubrió que el estado de los tumores de mama podía relacionarse con
cambios sistémicos en los niveles de biomarcadores de proteínas séricas (SPB) y de
autoanticuerpos asociados a tumores (TAAb). Meredith C. Henderson y col. evaluaron
por primera vez la contribución independiente y combinatoria de los datos de expresión
SPB y TAAb para identificar cáncer de mama utilizando una cohorte retrospectiva de
muestras de suero obtenidas en prebiopsia de 18 participantes que no mostraban evidencia
de enfermedad mamaria (ND), de 92 participantes diagnosticadas con enfermedad mama
benigna (BBD) y de 100 participantes diagnosticados con cáncer de mama, incluido
DCIS. Es importante mencionar que al modelar los datos integrados de SPB y TAAb, la
sensibilidad clínica y la especificidad para la detección del cáncer de mama mejoraron a
81.0% y 78.8%, respectivamente. Estos datos mostraron la ventaja que supone combinar
SPB y TAAb y son un argumento claro para defender el futuro desarrollo de otros
enfoques proteómicos combinatorios similares.
M. Pla-Roca et al. introdujeron un nuevo concepto para la detección multiplex
mediante un microarray de colocalización de anticuerpos (ACM). Esta técnica se validó
determinando 32 proteínas en el suero de: (i) 11 individuos control de edad similar que
se sometieron a mamoplastias de reducción, y (ii) 15 pacientes con cáncer de mama
primario que sobreexpresa el receptor de estrógenos (ER) en el tumor primario (subtipo
ER+). Se encontró una asociación entre seis proteínas (ENG, LEP, OPN, IL-1B, TNF- α
y uPAR) y el grado de cáncer de las pacientes. Los biomarcadores candidatos
identificados coinciden con los hallazgos de estudios preliminares, en los que había
mayores concentraciones de uPAR, TNF-RII, IL-1B y ENG. Sin embargo, repetimos,
todos ellos deben verificarse en estudios de seguimiento con más pacientes y controles.
Reconocer y caracterizar las diferentes formas de una proteína (isoformas) es
fundamental para el estudio de los mecanismos moleculares y la detección temprana de
enfermedades complejas como es el cáncer de mama. De esta manera, F. Zhang et al.
mostraron que usando los péptidos específicos de isoformas eran capaces de diferenciar
el cáncer de mama de una muestra sana, siendo capaces de identificar el 92.5% de las
muestras de cáncer y el 72.5% de las muestras normales en un conjunto independiente de
40 muestras normales y 40 muestras de cáncer de mama. Este trabajo demuestra que el
uso de péptidos obtenidos por mecanismos de splicing alternativo puede proporcionar
biomarcadores de interés para el estudio del cáncer de mama.

331
En un estudio desarrollado por D.L. Wang et al. se utilizó una tecnología
proteómica funcional para evaluar la actividad de proteasas y enzimas metabólicas
(hexoquinasas) a partir de una mezcla de proteínas séricas que se resolvía mediante una
primera separación en un gel bidimensional (2D) modificado y posterior transferencia a
platos de elución de proteínas (PEP) en los que realizar dicha determinación de la
actividad enzimática. Por primera vez, se encontraron diferencias sustanciales entre el
suero de pacientes con cáncer de mama y el suero normal en estas familias de enzimas,
lo que las señalaba como potenciales implicados en el desarrollo del cáncer y el proceso
de metástasis. Esta plataforma de trabajo proporciona excelentes biomarcadores
candidatos para el diagnóstico de cáncer de mama y para el desarrollo de fármacos.
El cáncer de mama es una enfermedad heterogénea con una amplia variedad de
características moleculares y clínicas, así como una variabilidad en la progresión clínica.
Para la elección del tratamiento adecuado, las pacientes con cáncer de mama se clasifican
en distintos subtipos empleando criterios clínico-patológicos que se basan en los niveles
de expresión del oncogén del receptor 2 del factor de crecimiento epidérmico humano
(HER2), la clasificación inmunohistoquímica del receptor de estrógeno (RE) y el receptor
de progesterona (RP), y el índice Ki-67. La clasificación del cáncer de mama en los
distintos subtipos permite un abordaje más personalizado de los tratamientos médicos,
con resultados favorables. Sin embargo, a pesar de esto, casi el 10-15% de estos pacientes
todavía experimentan recidivas locales o distantes en los primeros 5 años tras el
diagnóstico.
Las nuevas herramientas “ómicas” permiten la identificación de nuevos
biomarcadores que respalden el diagnóstico basado en patrones histopatológicos, lo que
se traduce en una mejora en la clasificación del cáncer de mama.
Con esta finalidad se comenzaron a aplicar los nanomateriales en el campo de la
proteómica, dando lugar a una nueva área de investigación denominada nanoproteómica.
La nanoproteómica se basa en que la dispersión de un nanomaterial en un fluido
fisiológico da como resultado la formación de una capa de proteinas denominada
"corona". Esta corona de proteínas varía en función de las características del medio
biológico, las propiedades físicas (tamaño, forma, curvatura) y químicas (composición,
carga superficial/química, hidrofobicidad/hidrofilicidad) del nanomaterial y el tiempo de
incubación.

332
Los biomarcadores asociados a un determinada enfermedad suponen menos del 1%
de las proteínas presentes en el suero sanguíneo. Con la formación de la corona de
proteínas, los nanomateriales actúan como materiales adsorbentes con los que se lleva a
cabo el enriquecimiento de los péptidos/proteínas de baja abundancia presentes en el
suero sanguíneo. El análisis de estas proteínas ancladas a la superficie de los
nanomateriales mediante técnicas de espectrometría de masas permitirá la identificación
de nuevos biomarcadores asociados con una determinada enfermedad, como el cáncer de
mama. Así, mediante este tipo de análisis, se podrán detectar cambios en la concentración
de proteínas en una fase temprana de una enfermedad, tras cualquier tratamiento
(quimioterapia, inmunoterapia) o una intervención quirúrgica. Por lo tanto, la
caracterización de la corona de proteínas que se forma alrededor de los nanomateriales
ofrece distintas ventajas en relación con los análisis proteómicos convencionales, y es
más eficaz a la hora de llevar a cabo la identificación de nuevas dianas moleculares.
El objetivo principal de la presente Tesis Doctoral se centra en descubrir nuevos
biomarcadores proteicos presentes en el suero sanguíneo asociados con los diferentes
subtipos de cáncer de mama empleando una nueva herramienta nanoproteómica. Los
objetivos específicos son:
1. Encontrar las condiciones óptimas de pH, temperatura, relación
[proteína]/[NPs] y tiempo de incubación, para la formación de la corona de proteínas
alrededor de las AuNPs, AgNPs, FeNPs y PtNPs tras su interacción con el suero
sanguíneo.
2. Llevar a cabo un análisis cualitativo y comparar la funcionalidad de las
proteínas presentes en el suero sanguíneo adsorbidas en la superficie de diferentes
nanomateriales funcionalizados con citrato, concretamente AuNPs (10.02 ± 0.91 nm),
AgNPs (9.73 ± 1.70 nm) y PtNPs (2.40 ± 0.30 nm).
3. Identificar nuevos biomarcadores proteicos presentes en el suero
sanguíneo relacionados con el cáncer de mama triple negativo a través del análisis
cualitativo y cuantitativo de la corona de proteínas que se forma alrededor de AuNPs
(10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) y MNPs (9.30 ± 0.67 nm).
4. Identificar nuevos biomarcadores proteicos presentes en el suero
sanguíneo relacionados con los distintos subtipos de cáncer de mama (luminal A, luminal
B HER2 negativo, luminal B HER2 positivo, HER2 positivo y triple negativo) a través

333
del análisis cualitativo y cuantitativo de la corona de proteínas que se forma alrededor de
AuNPs (12.96 ± 0.72 nm).
Las conclusiones principales de esta Tesis Doctoral son:
1. En todos los casos, tras la interacción de AuNPs (10.02 ± 0.91 nm y 12.96 ±
0.72 nm), AgNPs (9.73 ± 1.70 nm), PtNPs (2.40 ± 0.30 nm) y MNPs (9.30 ± 0.67 nm)
con suero sanguíneo, tiene lugar la formación de una corona de proteínas alrededor de la
superficie de los nanomaterials.
2. La formación de la corona de proteínas depende de la composición del
nanomaterial y su tamaño (observándose que las nanopartículas más pequeñas como las
PtNPs tienen una capacidad de adsorción inferior que las de mayor tamaño como las
AuNPs), la relación [proteína]/[NPs], el pH de la muestra y el tiempo de incubación.
3. Se identificaron un total de 215, 215 y 198 proteínas en la corona de proteínas
de AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) y PtNPs (2.40 ± 0.30 nm),
respectivamente, tras el ajuste de las siguientes condiciones óptimas: valor de pH de 5.8
con un tampon de citrato/ácido cítrico, tiempo de incubación igual a 30 minutos, y una
relación [proteína]/[NPs] de 10,7. Del total de proteínas identificadas, 170 son comunes
en los tres tipos de NPs y 52 son proteínas específicas de cada tipo (21 proteínas de las
AuNPs, 17 proteínas de las AgNPs y 14 proteínas de las PtNPs).
4. Del total de proteínas identificadas, la mayoría (un total de 66) participan en la
respuesta immune. Además, tienen otra funcionalidad como la enzimática, estructural,
transportadora, inflamatoria, transducción de la señal y propiedades
antibióticas/antibacterianas.
5. Las proteínas específicas identificadas en la superficie de las AuNPs tienen una
función estructural y están implicadas en la transdución de señal, son proteínas con
propiedades antibióticas/antibacterianas en el caso de las AgNPs, y son proteínas
implicadas en procesos inflamatorios en el caso de las PtNPs.
6. Las condiciones óptimas para la formación de la corona de proteínas alrededor
de las MNPs (9.30 ± 0.67 nm) son una depleción con DTT fresco en agua milli-Q en una
concentración 500 mM durante 60 minutos a temperatura ambiente, una relación
[proteína]/[MNPs] 1:2 y un valor de pH de 5.5 tanto en el paso de incubación como en el
paso final de lavado.

334
7. La combinación de AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) y MNPs
(9.30 ± 0.67 nm) con la espectrometría de masas permitió la identificación de siete
biomarcadores potenciales para el diagnóstico y la evaluación de la progresión del cancer
de mama triple negative: GRF-type zinc finger domain-containing protein 1 (proteína
ZGRF1), metaloproteinasa de matriz 9 (MMP9), lebercilina, immunoglobulina lambda
variable 3-27 (LV327), LINE-1 type transposase domain-containing protein 1 (proteína
LITD1), structural maintenance of chromosomes protein 6 (proteina SMC6) y short
coiled-coil protein (proteína SCOC).
8. La combinación de AuNPs (12.96 ± 0.72 nm) con la técnica de espectrometría
de masas Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS)
permitió analizar cuantitativamente las alteraciones proteómicas presents en el suero
sanguíneo en los diferentes subtipos de cancer de mama (luminal A, luminal B HER2
negativo, luminal B HER2 positivo, HER2 positivo y triple negativo).
9. El análisis cuantitativo de la composición de la corona de proteínas que se forma
alrededor de las AuNPs tras la incubación con las muestras de suero sanguíneo
pertenecientes a pacientes con los distintos subtipos de cancer de mama permitió
identificar 75 potenciales biomarcadores específicos: 8, 27, 2, 28 y 10 proteínas
específicas asociadas a los subtipos LA, LB-, LB+, HER2+ y TNBC, respectivamente.
10. El análisis cuantitativo de la composición de la corona de proteínas que se
forma alrededor de las AuNPs tras la incubación con las muestras de suero sanguíneo
pertenecientes a pacientes con los distintos subtipos de cancer de mama revelaron una
alteración en los niveles de las proteínas de coagulación sanguínea en el caso de los
pacientes en los que hay una sobreexpresion de HER2. Estas proteínas biomarcadoras
están implicadas en la progresión del tumor de mama, incluida la transformación celular,
la proliferación, la supervivencia de las células tumorales y la angiogénesis.

ANNEX E
List of Publications


Contents lists available at ScienceDirect
Colloids and Surfaces B: Biointerfaces
journal homepage: www.elsevier.com/locate/colsurfb
Proteomic analysis of the bio-corona formed on the surface of (Au, Ag, Pt)-nanoparticles in human serum
María del Pilar Chantada-Vázqueza, Antonio Castro Lópezb, Susana B. Bravoc,Sergio Vázquez-Estévezd, Benigno Acea-Nebrile, Cristina Núñeza,⁎
a Research Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde (SERGAS), 27002, Lugo, Spainb Breast Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde (SERGAS), 27002, Lugo, Spainc Proteomic Unit, Instituto de Investigaciones Sanitarias-IDIS, Complejo Hospitalario Universitario de Santiago de Compostela (CHUS), 15706, Santiago de Compostela,SpaindOncology Division, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde (SERGAS), 27002, Lugo, Spaine Department of Surgery, Breast Unit, Complexo Hospitalario Universitario A Coruña (CHUAC), SERGAS, A Coruña, Spain
A R T I C L E I N F O
Keywords:Gold nanoparticles (AuNPs)Silver nanoparticles (AgNPs)Platinum nanoparticles (PtNPs)Human serumProtein coronaPrognostic biomarker
A B S T R A C T
Adsorption of biomolecules onto nanoparticles surface in biological samples led to the formation of a bio-corona,it could modified the “identity” of nanoparticles, contributing to the determination of their toxicity and bio-compatibility.
Gel electrophoresis in combination with liquid chromatography-tandem mass spectrometry (LC–MS/MS) wasemployed to qualitatively analyze and identify the human serum proteins adsorbed on the surface of threedifferent nanomaterials stabilized with citrate: 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nmsilver nanoparticles (AgNPs) and, 2.40 ± 0.30 nm platinum nanoparticles (PtNPs). An exhaustive analysis andclassification of all identified proteins related with their function were also developed.
1. Introduction
In recent years, noble metal NPs have gained great interest becausetheir diverse material properties hold promising potential in advancingcurrent diagnostic and therapeutic technologies. Such properties arefundamentally dependent on their size, shape, and composition [1].
Particularly, gold nanoparticles (AuNPs) have a wide range of ap-plications in the biological and medical fields, including thermo-therapy, biosensors, and molecular imaging [2]. Silver nanoparticles(AgNPs) are known to be potent antimicrobial agents with a broadantimicrobial spectrum and high efficacy against bacteria [3], and theyhave been widely used in consumer products [4]. Platinum nano-particles (PtNPs) have been already proposed as efficient and selectiveradical scavengers for therapies of oxidative stress diseases [5,6].However, their clinical potential has been slowed down by some tox-icological concerns. Nevertheless, the use of PtNPs as additives inconsumer products and cosmetics has been already approved [7]. Thissuggests that PtNPs will likely register an increase shortly in a widerange of applications, including healthcare devices, diagnostics, andcosmetics [8]. Hence, the possibility of living organisms being exposedto Ag/Au/Pt-NPs either directly or indirectly is greatly increasing,
which has resulted in safety concerns [9].NPs can enter the human body in different ways, among which the
main exposure routes include inhalation, oral administration, in-travenous injection, and dermal exposure [10]. Once NPs enter thebody, they contact various biological molecules such as proteins, lipid,polysaccharides, and nucleic acids [11]. Particularly, when NPs areexposed to biological fluids (as serum/plasma), proteins and otherbiomolecules are easily adsorbed onto the surface to form a protein‘corona’ around NPs, which reduces the surface free energy of NPs [12].
Protein corona is divided into “soft corona” and “hard corona”.While soft corona is formed by lower affinity proteins reversibly boundto NPs (which can be further exchangeable), hard corona containshigher affinity proteins on the NP surface that may irreversibly bind toNPs. Protein corona is a dynamic layer in which amount of protein andthe arrangement are changeable according to the conditions of biolo-gical and physicochemical interaction [13].
During the processes, chemical or physical adsorption takes part inthe formation of protein corona. Coordination, hydrogen bonding, vander Waals forces, electrostatic and hydrophobic interactions, sterichindrance, etc. play important roles in driving the binding of proteins toNPs [14,15].
https://doi.org/10.1016/j.colsurfb.2019.01.056Received 11 September 2018; Received in revised form 15 January 2019; Accepted 26 January 2019
⁎ Corresponding author.E-mail address: [email protected] (C. Núñez).
Colloids and Surfaces B: Biointerfaces 177 (2019) 141–148
Available online 29 January 20190927-7765/ © 2019 Elsevier B.V. All rights reserved.
T

Furthermore, protein corona patterns mainly depend on the physi-cochemical properties of NPs (nanomaterial, size, charge, surfacefunctional groups, shape) and exposing environments including im-mersed media components, temperature, pH, dynamic shear stress, andinteraction (or exposing) time [16–19]. Proteins with large quantitiesare first bound to NP surface, and then gradually replaced by higheraffinity proteins (Vroman effect) [20].
When the protein is bound to NPs to form protein corona, proteinsmay reorganize their structures to adapt to surrounding environmentsand NPs surface. The secondary and/or tertiary protein structure ismodified, and this event is known as “conformational changes” [21].Furthermore, the formation of a corona may eliminate the physiologicalfunctions of proteins, which leads to the loss of original targetingcapabilities [22], induces various cellular responses including in-flammatory responses, increased lysosomal permeability, activatedcaspase-related pathways, or even apoptosis [23,24].
Understanding how the different properties of nanoparticles affectthe composition of protein coronas is therefore important. The serumand plasma protein corona compositions of many AuNPs with differentcoatings and different sizes have been identified [25–27]. However, thehigh-throughput protein analyses of the AgNPs’ coronas have beenlimited [26,28–30]. To better understand the relationship betweencorona composition and the nanoparticles’ properties, a recent studywas developed. This analysis showed specific binding patterns for theblood plasma-derived corona composition of AgNPs and AuNPs withthree surface coatings (polyethyleneimine (BPEI), citrate (CIT), andpolyvinylpyrrolidone (PVP)) [31]. To the best of our knowledge, theanalysis of the composition of the serum protein corona formed aroundPtNPs was not carried out until the moment.
On the other hand, a prognostic factor is defined as any parameter,evaluated at diagnosis (or surgery), which is associated with treatmentoutcome (local control, disease-free interval, survival) and may predictpatient outcome independent of treatment. Furthermore, prognosticfactors (clinical or biological) may be defined in any disease stage orsetting.
Blood-based biomarkers can be useful as pre-treatment prognosticmarkers, as they can reflect variations in the tumor microenvironmentand host immune response and can complement biopsy-based bio-markers that evaluate tumor cells directly [32]. As whole blood pro-vides a dynamic representation of physiological and pathological state,serum or plasma represents the most broadly studied biological matrixfor cancer biomarkers. Therefore, analysis of the plasma or serumproteome could be important to achieve accurate diagnosis or prog-nosis.
A great number of proteomics-based studies of plasma and serumhave reported differential peptide/protein ion peaks, either as identi-fied proteins or on the basis of their mass/charge (m/z) values, fordifferent cancer diagnosis or prognosis as, for example, breast cancer[33,34], ovarian cancer [35], head and neck cancer [36], bladdercancer [37], lung cancer [38]; or other diseases as amyotrophic lateralsclerosis [39], ST-segment elevation myocardial infarction [40], severesepsis [41].
Taking into consideration all the aforementioned arguments, withour experimental work of the serum protein corona formed aroundAuNPs, AgNPs and PtNPs, we are providing basic information for tox-icological and immunological risk assessment, as well as informationabout the properties of different nanomaterials for the development ofnovel sensors with potential medical applications.
2. Experimental
2.1. Chemicals and reagents
All reagents and solvents used were HPLC-grade or higher. Sodiumcitrate tribasic dihydrate, tannic acid, chloroplatinic acid (H2PtCl6),sodium borohydride (NaBH4), trypsin, trifluoroacetic acid, DL-
Dithiothreitol(DTT), Iodoacetamide (IAA), acrylamide/bis-acrylamide30% solution (37.5:1), Glycerol 86–88%, Tris-base, Coomassie BrilliantBlue R250 (CBB), sodium carbonate, and the Sigma Marker wide range6.5–200 KDa were purchased from Sigma-Aldrich (St. Louis, MO, USA).Sodium dodecylsulfate (SDS) and formaldehyde were purchased fromPanreac (Barcelona, Spain). β-mercaptoethanol was purchased fromMerck (Hohen-brunn,Germany) and bromophenol-blue was purchasedfrom Riedel-de Haen (Seelze,Germany). Hydrogen tetrachloroaurate(III) hydrate (99.9%-Au) (49%Au) at 10%w/v was purchased fromStrem Chemicals (Newburyport, MA, USA). Ammoniumbicarbonate(AMBIC) and formic acid were purchased from Fluka (Steinheim,Germany).
2.2. Instrumentation
Microscopic characterizations of AuNPs, AgNPs and PtNPs wereperformed by transmission electron microscopy (TEM) using a Jeol JEM1011 microscope. Samples for TEM were prepared by pipetting a dropof the colloidal dispersion onto an ultrathin carbon-coated copper gridand allowing the solvent to evaporate. Power Pac Basic power supplyfrom Bio-Rad (CA, USA) was used for sodium dodecyl sulfate poly-acrylamide gel electrophoresis (SDS-PAGE) protein separation. Proteinquantification was accomplished by measuring the absorbance at280 nm with the use of a Qubit™ 4 Quantitation Starter Kit fromThermo Fisher Scientific. Gel image acquisition was carried out with aUVP PhotoDoc-It™ Imaging System from Analytik Jena.
2.3. Synthesis of inorganic nanoparticles
Gold nanoparticles (AuNPs), silver nanoparticles (AgNPs) and pla-tinum nanoparticles (PtNPs) were synthesized by the citrate reductionmethod in aqueous solution by the method reported by R. López-Cortéset al. [42], V. Puntes et al. [43] and W. Chen et al. [44], respectively(see Supplemental Material).
2.4. Serum samples
Venous blood sample was obtained from five disease-free in-dividuals with the use of VACUETTE® Serum Clot Activator Tubes(10mL). The collected blood samples were allowed to clot for 15min,and then centrifuged for 5min at 4 °C and 1800×g. Sera were trans-ferred into clean plastic tubes (1 mL) and immediately frozen at −80 °Cat Research Unit, Hospital Universitario Lucus Augusti (HULA).
2.5. Depletion of multiple high abundant proteins
Serum aliquots (×3) were filtered with Miller-GP® Filter Unit(Millipore) with a size of 0.22 μm. Each aliquot of human serum (30 μL)was depleted with dithiothreitol (DTT) according to the protocol de-scribed by Warder el al. [45,46]. Fresh DTT 500mM (3.3 μL) was mixedwith 30 μL of human serum and vortex briefly. Samples were then in-cubated until a viscous white precipitate persisted (60min), followedby centrifugation at 18,840×g for 20min. Supernatants were trans-ferred to a clean tube prior to protein alkylation and nanoparticles(NPs) fractionation.
2.6. NPs protein alkylation and fractionation
After protein depletion, the reduced SH-groups were alkylated withiodoacetic acid (IAA) for 45min at room temperature and protectedfrom light. After protein reduction and alkylation, 75 μL of AuNPs(10.02 ± 0.91 nm), 75 μL of AgNPs (9.73 ± 1.70 nm) and 75 μL ofPtNPs (2.40 ± 0.30 nm) were added to each different serum aliquots(×3) belonging to the five disease-free individuals (9 aliquots per in-dividual, 3 with each nanoparticle type), followed by the addition of40 μL of citrate/citric acid buffer to a final pH of 5.8, as described by
M. del Pilar Chantada-Vázquez et al. Colloids and Surfaces B: Biointerfaces 177 (2019) 141–148
142

López-Cortés et al [42]. Then, all NPs-serum solutions were incubatedat 37 °C with shaking in a thermostatic bath during 30min.
Pellets were harvested by centrifugation at 18,840×g (AuNPs andAgNPs) and 24,610×g (PtNPs) during 30min. In all cases, pelletscontaining proteins bound to nanoparticles were washed three timeswith 25 μL citrate/citric acid buffer and harvested again by cen-trifugation at 18,840×g (AuNPs and AgNPs) and 24,610×g (PtNPs)during 30min to remove unbound proteins.
All pellets were reconstituted in 10 μL of buffer with 0.2 MTris−HCl, 2% w/v SDS and 20% v/v glycerol. This 10 μL was mixedwith 4 μL of SDS-PAGE loading buffer (10% w/v SDS, Tris-Base 40mM,pH 6.8, 50% v/v glycerol, 0.1% v/v bromophenol blue, 10% v/v β-mercaptoethanol) in a final volume of 20 μL. Then, all samples weredenatured by heating at 100 °C for 5min and loaded into a 10% acry-lamide/bis-acrylamide, stacking gel /12.5% acrylamide/bis-acrylamiderunning gel, of 1 mm thickness, and separated at 180 V (constant vol-tage) for 120min. After electrophoresis, the gel was fixed for 30minwith 40% (v/v) ethanol and 10% (v/v) acetic acid and then stainedovernight with Colloidal Coomassie Blue [47]. Gels were rinsed withdistilled water and a 0.5 M sodium chloride solution until a clearbackground was observed. Gel imaging was carried out with a with aUVP PhotoDoc-It™ Imaging System.
2.7. In-gel protein digestion
Protein bands were excised manually and transferred to 2.5-mL Lo-Bind tubes, and then washed twice with water and with 50% (v/v)acetonitrile/ 25mM ammonium bicarbonate (ambic) until the bluecolor disappeared.
Prior to trypsin digestion, gel spots were washed with 25mM ambicand dehydrated with acetonitrile. Then, 30 μL of trypsin (20 ng μL−1 in12.5 mM ambic/2% (v/v) acetonitrile) was added to the gel spots andincubated for 60min at 0 °C.
After this time, gel spots were inspected, trypsin solution not ab-sorbed into the gel was removed, and the gels were covered with 100 μLof 12.5mM ambic. Samples were incubated for 12 h at 37 °C. Then50 μL of 5% (v/v) formic acid was added and the supernatant wastransferred to a new Lo-Bind tube and the peptides were further ex-tracted from the gel twice with 50% (v/v) acetonitrile/0.1% (v/v) tri-fluoroacetic acid (TFA) (×3) and acetonitrile (ACN) (x1). Samples weredried-down and stored at −20 °C [48].
2.8. Protein identification by mass spectrometry (LC–MS/MS) and dataanalysis
Digested peptides of each sample were separated using ReversePhase Chromatography. Gradient was developed using a micro liquidchromatography system (Eksigent Technologies nanoLC 400, SCIEX)coupled to high speed Triple TOF 6600 mass spectrometer (SCIEX) witha micro flow source. The analytical column used was a silica-basedreversed phase column YMC-TRIART C18 150× 0.30mm, 3mm par-ticle size and 120 Å pore size (YMC Technologies, Teknokroma). Thetrap column was a YMC-TRIART C18 (YMC Technologies, Teknokromawith a 3mm particle size and 120 Å pore size, switched on-line with theanalytical column. The loading pump delivered a solution of 0.1%formic acid in water at 10 μL/min. The micro-pump provided a flow-rate of 5 μL/min and was operated under gradient elution conditions,using 0.1% formic acid in water as mobile phase A, and 0.1% formicacid in acetonitrile as mobile phase B. Peptides were separated using a25min gradient ranging from 2% to 90% mobile phase B (mobile phaseA: 2% acetonitrile, 0.1% formic acid; mobile phase B: 100% acetoni-trile, 0.1% formic acid). Injection volume was 4 μL.
Data acquisition was carried out in a TripleTOF 6600 System(SCIEX, Foster City, CA) using a Data dependent workflow. Source andinterface conditions were as follows: ionspray voltage floating (ISVF)5500 V, curtain gas (CUR) 25, collision energy (CE) 10 and ion source
gas 1 (GS1) 25. Instrument was operated with Analyst TF 1.7.1 software(SCIEX, USA). Switching criteria was set to ions greater than mass tocharge ratio (m/z) 350 and smaller than m/z 1400 with charge state of2–5, mass tolerance 250 ppm and an abundance threshold of more than200 counts (cps). Former target ions were excluded for 15 s. Instrumentwas automatically calibrated every 4 h using as external calibranttryptic peptides from PepcalMix (Sciex).
2.9. Data analysis
After MS/MS analysis, data files were processed usingProteinPilotTM 5.0.1 software from Sciex which uses the algorithmParagonTM for database search and ProgroupTM for data grouping.Data were searched using a Human specific Uniprot database. Falsediscovery rate was performed using a non-lineal fitting method dis-playing only those results that reported a 1% Global false discovery rateor better [49,50].
3. Results and discussion
3.1. Serum fraction preparation and protein corona purification
Following the synthetic methods described by R. López-Cortés [42],V. Puntes et al. [43] and W. Chen et al. [44], AuNPs(10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and PtNPs(2.40 ± 0.30 nm) were successfully obtained and characterized (seeFigs. 1_SM to 3_SM).
High abundance serum proteins were first depleted using DTT andseparated from low abundance serum proteins [42,43]. Low abundanceproteins were further processed as described in the experimental sec-tion. Afterward, AuNPs (10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm)and PtNPs (2.40 ± 0.30 nm) were mixed with serum aliquots (×3)from five different individuals (9 protein samples per individual: 3treated with AuNPs, 3 with AgNPs and 3 with PtNPs).
It is well known that some variables influence the protein-captureefficiency, namely (i) the NPs/protein ratio, (ii) sample pH and (iii)incubation time [42]. Particularly, the pH value is an important para-meter because influences the charge state of proteins, and a maximumbinding capacity between NPs and proteins takes place at the pH near tothe protein pI [51,52]. The optimum conditions found in preliminaryexperiments with AuNPs [42], were used in this case to reduce thecomplexity and large dynamic range of the human serum. To this aim,the pH was adjusted to 5.8 with citrate/citric acid buffer and threeincubation times (30, 60 and 90min) were tested. After the incubation,the pellets and the supernatants were separated via centrifugation andthe respective protein content was assessed through the use of 1D gelelectrophoresis. Similar results to that obtained after 30min were foundwith incubation times of 60 and 90min (Figs. 1_SM to 3_SM), sug-gesting that 30min was enough to achieve a good separation efficiencyand it was selected as the optimum incubation time.
To investigate the influence of the amount of each type of NPsduring the separation process, three volumes of each type of nano-particles were explored: 75 μL, 100 μL and 125 μL, to get the followingprotein/NPs ratios: 10.7, 8.6 and 6.5. Fig. 2 and Fig. 4_SM to Fig. 6_SMshow the result of this set of experiments, which suggest good separa-tion efficiency, even for the lowest amounts of each NPs tested (75 μL).
As it is shown in Fig. 2, for an incubation time of 30min, differencesin the protein corona formed around the three different NPs werevisible after Coomassie staining. When the gel profiles of each fraction(AuNPs-protein corona, AgNPs-protein corona, PtNPs-protein corona)were compared, it was easily noted that there is a different in the in-tensity of the bands. However, no conclusion can be drawn unless theproteins were identified. However, the supernatants were very similarin all cases, independently of the type of nanoparticle employed and thequantity of each one (Fig. 7_SM).
After that, gel bands corresponding to the fractions AuNPs-protein
M. del Pilar Chantada-Vázquez et al. Colloids and Surfaces B: Biointerfaces 177 (2019) 141–148
143

corona, AgNPs-protein corona, PtNPs-protein corona formed after theaddition of 75 μL, were excised and submitted to the sample treatmentdescribed in the experimental section. The resulting pools of peptideswere then analyzed by mass spectrometry (LC–MS/MS) for proteinidentification.
A similar number of proteins were identified from the proteincorona formed around AuNPs (10.02 ± 0.91 nm), AgNPs(9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm) after their incubationwith serum aliquots (×3) from five different individuals (9 proteinsamples per individual: 3 treated with AuNPs, 3 with AgNPs and 3 withPtNPs) (see Table 1_SM). Importantly, 215, 215 and 198 were com-monly found in all cases in the surface of AuNPs (10.02 ± 0.91 nm),AgNPs (9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm) (see Table 2_SM). From them, 170 proteins were commonly detected in the proteincorona of all three different types of NPs (see Fig. 1). One of this pro-teins was serum albumin, the most abundant protein in the blood.
However, 52 different proteins were found on the three different NPssurface (see Fig. 1): 21 different proteins on the 10.02 ± 0.91 nmAuNPs (see Table 1), 17 on the 9.73 ± 1.70 nm AgNPs (see Table 2)and 14 individual proteins on the 2.40 ± 0.30 nm PtNPs (see Table 3).
Then, an analysis of the protein corona formed around the threenanoparticles (AuNPs, AgNPs and PtNPs) will be carried out in relationto the different functionality of the proteins identified.
3.2. Proteins implicated in the immnune response
A total of 66 proteins implicated in the immune response (55 ofthem are immunoglobulins) were identified from the protein coronaformed around the AuNPs, AgNPs, and PtNPs after their incubationwith serum of all individuals. From them, 48 common proteins werefound on the surface of the three different nanoparticles (seeTable 1_SM). Particularly, two of these common immune-related
Fig. 1. Number of identified proteins found in the protein corona of 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs) and2.40 ± 0.30 nm platinum nanoparticles (PtNPs); and Venn diagram showing the common proteins in the three different nanoparticles surfaces.
Fig. 2. 1D-SDS-PAGE of protein coronas formed around10.02 ± 0.91 nm gold nanoparticles (AuNPs),9.73 ± 1.70 nm silver nanoparticles (AgNPs) and2.40 ± 0.30 nm platinum nanoparticles (PtNPs) in humanserum (incubation time: 30min; volumes of each nano-particles solution: 75 μL, 100 μL and 125 μL, to get the fol-lowing protein/NPs ratios: 10.7, 8.6 and 6.5, respectively). Onthe left, it marks the lane with Mw protein standards withmolecular weights in kDa.
M. del Pilar Chantada-Vázquez et al. Colloids and Surfaces B: Biointerfaces 177 (2019) 141–148
144

proteins were complement C3 (187.1 kDa) and complement factor B(85.5 kDa). The according complement system is involved in an effec-tive strategy called opsonization for labeling pathogens for removal byphagocytic cells from the circulation. To minimize opsonization ofAuNP, AgNPs and PtNPs in pharmaceutical applications (since thedrug-loaded NP is cleared by the organism and cannot release its drugat the desired site) binding of the NP with complement factors shouldbe avoided [49].
An example of a protein implicated in the immune response thatwas only found on the surface of AgNPs was fibrinogen alpha chain(94.9 kDa), involved in blood coagulation but also opsonizes surfaces offoreign bodies for immune cell recognition. Furthermore, AuNPs na-noparticles exhibited selective adsorption towards immunoglobulinheavy constant gamma 1 (36.1 kDa) (IGHG1), similar to that reportedpreviously in by zeolites in plasma [53].
3.3. Proteins with a transport function
A total of 29 proteins with a transporter function (7 of them apo-lipoproteins) were identified from the protein corona formed aroundthe AuNPs, AgNPs, and PtNPs. From them, 25 common proteins werefound on the surface of the three different nanoparticles (seeTable 2_SM). Among them, the most interesting protein regardingtoxicity and drug delivery potential is the ApoE. This transport proteinis known to mediate transcytosis across biological barriers, e.g., theblood-brain barrier [54,55].
AuNPs and AgNPs exhibited selective adsorption towards two pro-teins that belong to the ATP-binding cassette (ABC) transporter super-family of integral membrane proteins: ATP-binding cassette sub-familyB member 5 (ABCB5) (138.6 kDa) and ATP-binding cassette sub-familyF member 1 (ABCF1) (95.9 kDa), respectively. Both proteins participatein the ATP-dependent transmembrane transport of structurally diverse
Table 1Selection of identified single-detected corona proteins bound to the 10.02 ± 0.91 nm AuNPs after 30min incubation and subsequent washing. The accessionnumber, gene name, species (Human), molecular weight (kDa) and protein function were reported.
Protein Name UniProt Name Entry Name Gene Mass(kDa)
Function
ATP-binding cassette sub-family B member 5 ABCB5_HUMAN Q2M3G0 ABCB5 138.6 Transporter activityBasement membrane-specific heparan sulfate proteoglycan
core proteinPGBM_HUMAN P98160 HSPG2 468.8 Structural
Cadherin-5 CADH5_HUMAN P33151 CDH5 87.5 Controls the cohesion and organization of theintercellular junctions
Centlein CNTLN_HUMAN Q9NXG0 CNTLN 161.6 StructuralComplement component C8 alpha chain CO8A_HUMAN P07357 C8A 65.2 StructuralComplement factor D CFAD_HUMAN P00746 CFD 27.1 Catalytic activityDENN domain-containing protein 5B DEN5B_HUMAN Q6ZUT9 DENND5B 145.1 Enzyme regulator activityF-box only protein 3 FBX3_HUMAN Q9UK99 FBXO3 54.6 Enzyme regulator activityFilaggrin-2 FILA2_HUMAN Q5D862 FLG2 248.1 _Glyceraldehyde-3-phosphate dehydrogenase G3P_HUMAN P04406 GAPDH 36.1 Catalytic activityHAUS augmin-like complex subunit 8 HAUS8_HUMAN Q9BT25 HAUS8 44.9 Involved in microtubule generation within the mitotic
spindleHepatocyte growth factor activator HGFA_HUMAN Q04756 HGFAC 70.7 Enzyme regulator activity (activator)Immunoglobulin heavy constant gamma 1 IGHG1_HUMAN P01857 IGHG1 36.1 Immune responseImmunoglobulin heavy variable 4-39 HV439_HUMAN P01824 IGHV4-39 13.9 Immune responseImmunoglobulin lambda-like polypeptide 5 IGLL5_HUMAN B9A064 IGLL5 23.1 Immune responseKeratin, type I cytoskeletal 15 K1C15_HUMAN P19012 KRT15 49.2 StructuralMaestro heat-like repeat-containing protein family member
2AMRO2A_HUMAN A6NES4 MROH2A 189.6 _
Protein ENL ENL_HUMAN Q03111 MLLT1 62.1 Enzyme regulator activityRegulator of G-protein signaling 20 RGS20_HUMAN O76081 RGS20 31.5 Inhibits signal transductionTestis-specific gene 10 protein TSG10_HUMAN Q9BZW7 TSGA10 81.4 Plays a role in the sperm tail fibrous sheath, a major
sperm tail structureZinc-alpha-2-glycoprotein ZA2G_HUMAN P25311 AZGP1 34.3 Stimulates lipid degradation in adipocytes
Table 2Selection of identified single-detected corona proteins bound to the 9.73 ± 1.70 nm AgNPs after 30min incubation and subsequent washing. The accession number,gene name, species (Human), molecular weight (kDa) and protein function were reported.
Protein Name UniProt Name Entry Name Gene Mass (kDa) Function
Angiotensin-converting enzyme ACE_HUMAN P12821 ACE 149.7 Catalytic activityATP-binding cassette sub-family F member 1 ABCF1_HUMAN Q8NE71 ABCF1 95.9 Transporter activityCathelicidin antimicrobial peptide CAMP_HUMAN P49913 CAMP 19.3 Antibacterial activityCMP-N-acetylneuraminate-poly-alpha-2,8-sialyltransferase SIA8D_HUMAN Q92187 ST8SIA4 41.3 Catalytic activityDedicator of cytokinesis protein 3 DOCK3_HUMAN Q8IZD9 DOCK3 233.1 Enzyme regulator activityDopamine beta-hydroxylase DOPO_HUMAN P09172 DBH 69.1 Catalyctic activityEukaryotic translation elongation factor 1 epsilon-1 MCA3_HUMAN O43324 EEF1E1 19.8 Positive modulator of ATM response to DNA damage.Fibrinogen alpha chain FIBA_HUMAN P02671 FGA 94.9 Coagulation, immune responseImmunoglobulin heavy variable 3-11 HV311_HUMAN P01762 IGHV3-11 12.9 Immune responseImmunoglobulin heavy variable 3-30-3 HVC33_HUMAN P0DP02 IGHV3-30-3 13 Immune responseImmunoglobulin kappa variable 1D-12 KVD12_HUMAN P01611 IGKV1D-12 12.6 Immune responseImmunoglobulin lambda variable 3-27 LV327_HUMAN P01718 IGLV3-27 12.2 Immune responseL-lactate dehydrogenase B chain LDHB_HUMAN P07195 LDHB 36.6 Catalytic activityMultiple inositol polyphosphate phosphatase 1 MINP1_HUMAN Q9UNW1 MINPP1 55.1 Catalytic activityNeutrophil cytosol factor 4 NCF4_HUMAN Q15080 NCF4 39.1 Component of the NADPH-oxidasePleckstrin homology domain-containing family G member 6 PKHG6_HUMAN Q3KR16 PLEKHG6 88.9 Enzyme regulator activityUnconventional myosin-If MYO1F_HUMAN O00160 MYO1F 124.8 Catalytic activity (ATPase)
M. del Pilar Chantada-Vázquez et al. Colloids and Surfaces B: Biointerfaces 177 (2019) 141–148
145
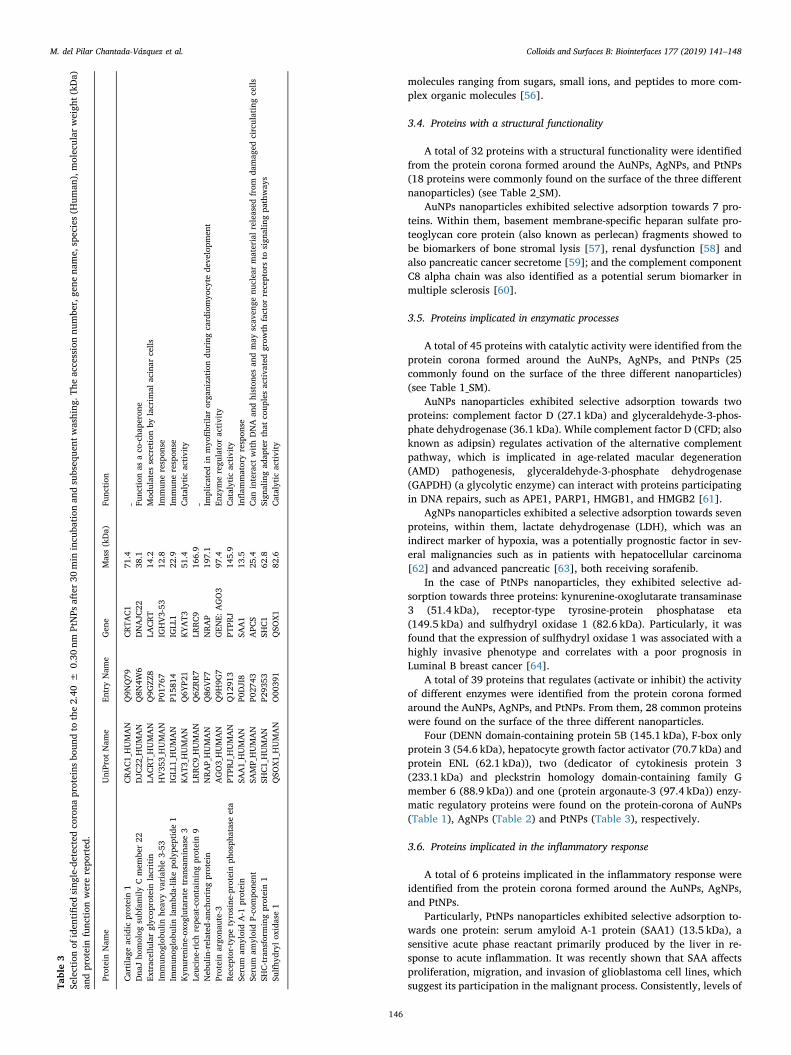
molecules ranging from sugars, small ions, and peptides to more com-plex organic molecules [56].
3.4. Proteins with a structural functionality
A total of 32 proteins with a structural functionality were identifiedfrom the protein corona formed around the AuNPs, AgNPs, and PtNPs(18 proteins were commonly found on the surface of the three differentnanoparticles) (see Table 2_SM).
AuNPs nanoparticles exhibited selective adsorption towards 7 pro-teins. Within them, basement membrane-specific heparan sulfate pro-teoglycan core protein (also known as perlecan) fragments showed tobe biomarkers of bone stromal lysis [57], renal dysfunction [58] andalso pancreatic cancer secretome [59]; and the complement componentC8 alpha chain was also identified as a potential serum biomarker inmultiple sclerosis [60].
3.5. Proteins implicated in enzymatic processes
A total of 45 proteins with catalytic activity were identified from theprotein corona formed around the AuNPs, AgNPs, and PtNPs (25commonly found on the surface of the three different nanoparticles)(see Table 1_SM).
AuNPs nanoparticles exhibited selective adsorption towards twoproteins: complement factor D (27.1 kDa) and glyceraldehyde-3-phos-phate dehydrogenase (36.1 kDa). While complement factor D (CFD; alsoknown as adipsin) regulates activation of the alternative complementpathway, which is implicated in age-related macular degeneration(AMD) pathogenesis, glyceraldehyde-3-phosphate dehydrogenase(GAPDH) (a glycolytic enzyme) can interact with proteins participatingin DNA repairs, such as APE1, PARP1, HMGB1, and HMGB2 [61].
AgNPs nanoparticles exhibited a selective adsorption towards sevenproteins, within them, lactate dehydrogenase (LDH), which was anindirect marker of hypoxia, was a potentially prognostic factor in sev-eral malignancies such as in patients with hepatocellular carcinoma[62] and advanced pancreatic [63], both receiving sorafenib.
In the case of PtNPs nanoparticles, they exhibited selective ad-sorption towards three proteins: kynurenine-oxoglutarate transaminase3 (51.4 kDa), receptor-type tyrosine-protein phosphatase eta(149.5 kDa) and sulfhydryl oxidase 1 (82.6 kDa). Particularly, it wasfound that the expression of sulfhydryl oxidase 1 was associated with ahighly invasive phenotype and correlates with a poor prognosis inLuminal B breast cancer [64].
A total of 39 proteins that regulates (activate or inhibit) the activityof different enzymes were identified from the protein corona formedaround the AuNPs, AgNPs, and PtNPs. From them, 28 common proteinswere found on the surface of the three different nanoparticles.
Four (DENN domain-containing protein 5B (145.1 kDa), F-box onlyprotein 3 (54.6 kDa), hepatocyte growth factor activator (70.7 kDa) andprotein ENL (62.1 kDa)), two (dedicator of cytokinesis protein 3(233.1 kDa) and pleckstrin homology domain-containing family Gmember 6 (88.9 kDa)) and one (protein argonaute-3 (97.4 kDa)) enzy-matic regulatory proteins were found on the protein-corona of AuNPs(Table 1), AgNPs (Table 2) and PtNPs (Table 3), respectively.
3.6. Proteins implicated in the inflammatory response
A total of 6 proteins implicated in the inflammatory response wereidentified from the protein corona formed around the AuNPs, AgNPs,and PtNPs.
Particularly, PtNPs nanoparticles exhibited selective adsorption to-wards one protein: serum amyloid A-1 protein (SAA1) (13.5 kDa), asensitive acute phase reactant primarily produced by the liver in re-sponse to acute inflammation. It was recently shown that SAA affectsproliferation, migration, and invasion of glioblastoma cell lines, whichsuggest its participation in the malignant process. Consistently, levels ofTa
ble3
Selectionof
iden
tified
sing
le-detectedco
rona
proteins
boun
dto
the2.40
±0.30
nmPtNPs
after30
min
incu
bation
andsubseq
uent
washing
.The
accessionnu
mbe
r,ge
nena
me,
species(H
uman
),molecular
weigh
t(kD
a)an
dproteinfunc
tion
wererepo
rted
.
ProteinNam
eUniProt
Nam
eEn
tryNam
eGen
eMass(kDa)
Func
tion
Cartilage
acidic
protein1
CRAC1_HUMAN
Q9N
Q79
CRTA
C1
71.4
_Dna
Jho
molog
subfam
ilyCmem
ber22
DJC
22_H
UMAN
Q8N
4W6
DNAJC
2238
.1Fu
nction
asaco
-cha
perone
Extracellularglycop
rotein
lacritin
LACRT_HUMAN
Q9G
ZZ8
LACRT
14.2
Mod
ulates
secretionby
lacrim
alacinar
cells
Immun
oglobu
linhe
avyva
riab
le3-53
HV35
3_HUMAN
P017
67IG
HV3-53
12.8
Immun
erespon
seIm
mun
oglobu
linlambd
a-lik
epo
lype
ptide1
IGLL
1_HUMAN
P158
14IG
LL1
22.9
Immun
erespon
seKyn
uren
ine-ox
oglutarate
tran
saminase3
KAT3
_HUMAN
Q6Y
P21
KYAT3
51.4
Catalytic
activity
Leuc
ine-rich
repe
at-con
tainingprotein9
LRRC9_HUMAN
Q6Z
RR7
LRRC9
166.9
_Neb
ulin-related
-anc
horing
protein
NRAP_HUMAN
Q86
VF7
NRAP
197.1
Implicated
inmyo
fibrila
rorga
nization
during
cardiomyo
cyte
deve
lopm
ent
Proteinargo
naute-3
AGO3_HUMAN
Q9H
9G7
GEN
E:AGO3
97.4
Enzymeregu
latoractivity
Recep
tor-type
tyrosine
-protein
phosph
ataseeta
PTPR
J_HUMAN
Q12
913
PTPR
J14
5.9
Catalytic
activity
Serum
amyloidA-1
protein
SAA1_HUMAN
P0DJI8
SAA1
13.5
Inflam
matoryrespon
seSe
rum
amyloidP-co
mpo
nent
SAMP_HUMAN
P027
43APC
S25
.4Can
interact
withDNA
andhiston
esan
dmay
scav
enge
nuclearmaterialreleased
from
damag
edcirculatingcells
SHC-transform
ingprotein1
SHC1_HUMAN
P293
53SH
C1
62.8
Sign
alingad
apterthat
coup
lesactiva
tedgrow
thfactor
receptorsto
sign
alingpa
thway
sSu
lfhy
dryl
oxidase1
QSO
X1_HUMAN
O00
391
QSO
X1
82.6
Catalytic
activity
M. del Pilar Chantada-Vázquez et al. Colloids and Surfaces B: Biointerfaces 177 (2019) 141–148
146

SAA have been used as a non-invasive biomarker for the prognosis ofmany cancers and, particularly, serum amyloid A1 was found to beupregulated in human glioblastoma [65].
3.7. Proteins with antibiotic activity
Dermcidin (11.3 kDa), a protein with antimicrobial activity, wasidentified from the protein corona formed around the AuNPs, AgNPs,and PtNPs. Dermcidin (DCD), an antimicrobial peptide with a broadspectrum of activity against bacteria such as Propionibacterium acnes, isexpressed constitutively in sweat in the absence of stimulation due toinjury or inflammation. It was found that reduced DCD concentration insweat in patients with inflammatory acne may permit proliferation of P.acnes in pilosebaceous units, resulting in progression of inflammatoryacne [66].
However, only AgNPs nanoparticles exhibited selective adsorptiontowards one protein with antibacterial activity: cathelicidin anti-microbial peptide (19.3 kDa), responsible for protecting the urinarytract against invasive bacterial infection [67].
3.8. Proteins implicated in signal transduction
There were identified three proteins implicated in the signal trans-duction from the protein corona formed around the three nanosystems.While protein leucine-rich alpha-2-glycoprotein (38.2 kDa) (implicatedin the protein-protein interaction, signal transduction and cell adhesionand development) was presented in the protein corona of the threenanoparticles, the protein regulator of G-protein signaling 20(31.5 kDa) (that inhibits signal transduction) and SHC-transformingprotein 1 (62.8 kDa) were only found in the surface of AuNPs andPtNPs, respectively.
In general, in relation with the total number of proteins identifiedfrom the protein corona surfaces (see Fig. 3), the most abundant groupsare constituted by the proteins implicated in the immune response,followed by proteins with and enzymatic function, structural, trans-porter, inflammatory, signal transduction and with antibiotic/anti-bacterial properties.
However, we found exclusive proteins with structural function andimplicated in the signal transduction only in the protein corona of10.02 ± 0.91 nm AuNPs (see Table 1), with antibiotic/antibacterialproperties in 9.73 ± 1.70 nm AgNPs (see Table 2) and implicated oninflammatory processes only in 2.40 ± 0.30 nm PtNPs (see Table 3).
4. Conclusions
In this study, it was shown that the interaction of AuNPs(10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and PtNPs(2.40 ± 0.30 nm) with human serum withstands the formation of aprotein corona enveloping the nanoparticle, in all cases.
The formation of this protein corona depends on the composition ofthe nanoparticle (core material) and its size. In that case, it was ob-served that smaller NPs (2.40 ± 0.30 nm PtNPs) have lower proteinadsorption (198 proteins) than larger NPs (10.02 ± 0.91 nm AuNPsand 9.73 ± 1.70 nm AgNPs) (215 proteins). A total of 170 proteinswith different functionality were detected in the protein corona of allthree different types of NPs. However, 21, 14 and 14 different proteinswere found on the AuNPs (10.02 ± 0.91 nm), AgNPs(9.73 ± 1.70 nm) and PtNPs (2.40 ± 0.30 nm), respectively.
In general, the function of all identified proteins ranges from pro-teins implicated in the immune response, followed by proteins with anenzymatic function, structural, transporter, inflammatory, signaltransduction and with antibiotic/antibacterial properties, being themajority group the first one with 66 proteins identified implicated inthe immune response. However, if we observed the different proteinson the corona of the three different NPs, we found exclusive proteinswith structural function and implicated in the signal transduction onlyin the protein corona of AuNPs (10.02 ± 0.91 nm), with antibiotic/antibacterial properties in AgNPs (9.73 ± 1.70 nm) and implicated oninflammatory processes only in PtNPs (2.40 ± 0.30 nm).
This has implications for immune safety, biocompatibility and in-formation for developing novel nanomaterials with high specificity andselectivity towards proteins with an important biological function(prognostic and diagnostic protein biomarkers). However, it is im-portant to mention that the interaction of these nanoparticles withhuman serum could drive to the formation of a different protein coronaat a pH dissimilar of 5.8, for example, under physiological conditions.
Conflict of interest
The authors declare that they have no conflict of interest.
Acknowledgments
All authors acknowledge Miguel Servet I Programme (CP16/00139)from the “Instituto de Salud Carlos III” (Plan Estatal de I+D+i 2013-2016 and European Development Regional Fund) of the SpanishMinistry of Economy and Competitiveness. M. P. Chantada-Vázquezand A. Castro López contributed equally to this work.
Appendix A. Supplementary data
Supplementary material related to this article can be found, in theonline version, at doi:https://doi.org/10.1016/j.colsurfb.2019.01.056.
References
[1] J. Zheng, C. Zhou, M. Yu, J. Liu, Nanoscale 4 (14) (2012) 4073–4083.[2] Z. Miao, Z. Gao, R. Chen, X. Yu, Z. Su, G. Wei, Curr. Med. Chem. 25 (16) (2018)
1920–1944.[3] N. Duran, M. Duran, M.B. de Jesus, A.B. Seabra, W.J. Favaro, G. Nakazato,
Nanomedicine 12 (3) (2016) 789–799.[4] K.W. Lem, A. Choudhury, A.A. Lakhani, P. Kuyate, J.R. Haw, D.S. Lee, Z. Iqbal,
C.J. Brumlik, Recent Pat. Nanotechnol. 6 (2012) 60–72.[5] M. Moglianetti, E. De Luca, D. Pedone, R. Marotta, T. Catelani, B. Sartori,
H. Amenitsch, S.F. Retta, P.P. Pompa, Nanoscale 8 (2016) 3739–3752.[6] H. Hosaka, R. Haruki, K. Yamada, C. Böttcher, T. Komatsu, PLoS One 9 (2014)
e110541.[7] S. Onizawa, K. Aoshiba, M. Kajita, Y. Miyamoto, A. Nagai, Pulm. Pharmacol. Ther.
22 (2009) 340–349.[8] D. Pedone, M. Moglianetti, E. De Luca, G. Bardi, P.P. Pompa, Chem. Soc. Rev. 46
(2017) 4951–4975.[9] T. Zhang, L. Wang, Q. Chen, C. Chen, Yonsei Med. J. 55 (2014) 283–291.
Fig. 3. Total number of identified proteins found on the surface of the threedifferent nanoparticles (color: black) and commonly found on the surface ofAuNPs, AgNPs and PtNPs (color: grey) with a different function. (For inter-pretation of the references to colour in this figure legend, the reader is referredto the web version of this article).
M. del Pilar Chantada-Vázquez et al. Colloids and Surfaces B: Biointerfaces 177 (2019) 141–148
147

[10] B. Wang, X. He, Z.Y. Zhang, Y.L. Zhao, W.Y. Feng, Acc. Chem. Res. 46 (2012)761–769.
[11] M. Mahmoudi, I. Lynch, M.R. Ejtehadi, M.P. Monopoli, F.B. Bombelli, S. Laurent,Chem. Rev. 111 (2011) 5610–5637.
[12] E. Casals, T. Pfaller, A. Duschl, G.J. Oostingh, V.F. Puntes, Small 7 (2011)3479–3486.
[13] S. Milani, F. Baldelli Bombelli, A.S. Pitek, K.A. Dawson, J. Rädler, ACS Nano 6(2012) 2532–2541.
[14] L. Treuel, S. Brandholt, P. Maffre, S. Wiegele, L. Shang, G.U. Nienhaus, ACS Nano 8(2014) 503–513.
[15] L.M. Wang, J.Y. Li, J. Pan, X.M. Jiang, Y.L. Ji, Y.F. Li, Y. Qu, Y.L. Zhao, X.C. Wu, J.Am. Chem. Soc. 135 (2013) 17359–17368.
[16] M.P. Monopoli, C. Aberg, A. Salvati, K.A. Dawson, Nat. Nanotechnol. 7 (2012)779–786.
[17] M. Lundqvist, J. Stigler, G. Elia, I. Lynch, T. Cedervall, K.A. Dawson, PNAS 105 (38)(2008) 14265–14270.
[18] S. Tenzer, D. Docter, S. Rosfa, A. Wlodarski, J. Kuharev, A. Rekik, S.K. Knauer,C. Bantz, T. Nawroth, C. Bier, J. Sirirattanapan, W. Mann, L. Treuel, R. Zellner,M. Maskos, H. Schild, R.H. Stauber, ACS Nano 5 (9) (2011) 7155–7167.
[19] J. Wolfram, Y. Yang, J. Shen, A. Moten, C. Chen, H. Shen, M. Ferrari, Y. Zhao,Colloids Surf. B Biointerfaces 124 (2014) 17–24.
[20] L. Vroman, A. Adams, G.C. Fischer, P.C. Munoz, Blood 55 (1) (1980) 156–159.[21] I. Lynch, K.A. Dawson, Nano Today 3 (1) (2008) 40–47.[22] A. Salvati, A.S. Pitek, M.P. Monopoli, K. Prapainop, F.B. Bombelli, D.R. Hristov,
P.M. Kelly, C. Åberg, E. Mahon, K.A. Dawson, Nat. Nanotechnol. 8 (2013) 137–143.[23] C. Paviolo, J.W. Haycock, J. Yong, A. Yu, S.L. McArthur, P.R. Stoddart, Proc. SPIE
8579 (2013) 85790C.[24] J.Y. Lee, W. Park, D.K. Yi, Toxicol. Lett. 209 (2012) 51–57.[25] M. Schaffler, M. Semmler-Behnke, H. Sarioglu, S. Takenaka, A. Wenk, C. Schleh,
S.M. Hauck, B.D. Johnston, W.G. Kreyling, Nanotechnology 24 (2013) 265103.[26] C.D. Walkey, J.B. Olsen, F. Song, R. Liu, H. Guo, D.W. Olsen, Y. Cohen, A. Emili,
W.C. Chan, ACS Nano 8 (2014) 2439–2455.[27] M.A. Dobrovolskaia, B.W. Neun, S. Man, X. Ye, M. Hansen, A.K. Patri, R.M. Crist,
S.E. McNeil, Nanomedicine 10 (2014) 1453–1463.[28] N. Duran, C.P. Silveira, M. Duran, D.S. Martínez, J. Nanobiotechnology 13
(2015) 55.[29] J.H. Shannahan, X. Lai, P.C. Ke, R. Podila, J.M. Brown, F.A. Witzmann, PLoS One 8
(9) (2013) e74001.[30] H. Huang, W. Lai, M. Cui, L. Liang, Y. Lin, Q. Fang, Y. Liu, L. Xie, Sci. Rep. 6 (2016)
25518.[31] W. Lai, Q. Wang, L. Li, Z. Hu, J. Chen, Q. Fang, Colloids Surf. B Biointerfaces 152
(2017) 317–325.[32] L. Brøndum, J.G. Eriksen, B.S. Sørensen, L.S. Mortensen, K. Toustrup, J. Overgaard,
J. Alsner, Clin. Trans. Rad. Oncol. 2 (2017) 46–52.[33] A. Kazarian, O. Blyuss, G. Metodieva, A. Gentry-Maharaj, A. Ryan, E.M. Kiseleva,
O.M. Prytomanova, I.J. Jacobs, M. Widschwendter, U. Menon, J.F. Timms, Br. J.Cancer 116 (2017) 501–508.
[34] D. Song, L. Yue, J. Zhang, S. Ma, W. Zhao, F. Guo, Y. Fan, H. Yang, Q. Liu, D. Zhang,Z. Xia, P. Qin, J. Jia, M. Yue, J. Yu, S. Zheng, F. Yang, J. Wang, Cancer Biol. Ther. 17(6) (2016) 635–647.
[35] T. Muinao, H.P. Deka Boruah, M. Pal, Exp. Cell Res. 362 (1) (2018) 1–10.[36] H.-S. Lin, F. Siddiq, H.S. Talwar, W. Chen, C. Voichita, S. Draghici, G. Jeyapalan,
M. Chatterjee, A. Fribley, G.H. Yoo, S. Sethi, H. Kim, A. Sukari, A.J. Folbe,M.A. Tainsky, Laryngoscope 124 (8) (2014) 1819–1826.
[37] K. Schwamborn, N.T. Gaisa, C. Henkel, Expert Rev. Proteomics 27 (6) (2010)897–906.
[38] T. Targowski, K. Jahnz-Rozyk, W. Owczarek, A. Raczka, P. Janda, T. Szkoda,T. Płusa, Respir. Med. 104 (9) (2010) 1356–1361.
[39] C. Lunetta, A. Lizio, E. Maestri, V.A. Sansone, G. Mora, R.G. Miller, S.H. Appel,A. Chiò, JAMA Neurol. 74 (6) (2017) 660–667.
[40] S. Baumann, A. Huseynov, J. Koepp, C. Jabbour, M. Behnes, T. Becher, M. Renker,S. Lang, M. Borggrefe, R. Lehmann, I. Akin, Angiology 37 (3) (2016) 272–280.
[41] J. Szederjesi, E. Almasy, A. Lazar, A. Huţanu, I. Badea, A. Georgescu, J. Crit. Care
Med. 1 (4) (2015) 147–153.[42] R. Lopez-Cortés, E. Oliveira, C. Núñez, C. Lodeiro, María Páez dela Cadena,
Florentino Fdez-Riverola, H. López-Fernández, Miguel Reboiro-Jato, Daniel Glez-Peña, José Luis Capelo, Hugo M. Santos, Talanta 100 (2012) 239–245.
[43] N.G. Bastus, F. Merkoci, J. Piella, V. Puntes, Chem. Mater. 26 (9) (2014)2836–2846.
[44] G.-W. Wu, S.-B. He, H. Peng, H.-H. Deng, A.-L. Liu, X.-H. Lin, X.-h. Xia, W. Chen,Anal. Chem. 86 (21) (2014) 10955–10960.
[45] S.E. Warder, L.A. Tucker, T.J. Strelitzer, E.M. McKeegan, J.L. Meuth, P.M. Jung,A. Saraf, B. Singh, J. Lai-Zhang, G. Gagne, J.C. Rogers, Anal. Biochem. 387 (2)(2009) 184–193.
[46] C. Fernández, H.M. Santos, C. Ruíz-Romero, F.J. Blanco, J.L. Capelo-Martínez,Electrophoresis 32 (21) (2011) 2966–2974.
[47] G. Candiano, M. Bruschi, L. Musante, L. Santucci, G.M. Ghiggeri, B. Carnemolla,P. Orecchia, L. Zardi, P.G. Righetti, Electrophoresis 25 (9) (2004) 1327–1333.
[48] E. Oliveira, J.E. Araújo, S. Gómez-Meire, C. Lodeiro, C. Perez-Melon, E. Iglesias-Lamas, A. Otero-Glez, J.L. Capelo, H.M. Santos, Clin. Proteomics 11 (1) (2014) 17.
[49] I.V. Shilov, S.L. Seymour, A.A. Patel, A. Loboda, W.H. Tang, S.P. Keating,C.L. Hunter, L.M. Nuwaysir, D.A. Schaeffer, Mol. Cell Proteomics 6 (2007)1638–1655.
[50] W.H. Tang, I.V. Shilov, S.L. Seymour, J. Proteome Res. 7 (2008) 3661–3667.[51] Z.G. Peng, K. Hidajat, M.S. Uddin, J. Colloid Interface Sci. 271 (2004) 277–283.[52] J.R. de Jesús, R. da Silva Fernandes, Gde Souza Pessôa, I.M. Raimundo Jr.,
M.A.Z. Arruda, Talanta 170 (2017) 199–209.[53] M. Rahimi, E.P. Ng, K. Bakhtiari, M. Vinciguerra, H. Ali Ahmad, H. Awala,
S. Mintova, M. Daghighi, F. Bakhshandeh Rostami, M. de Vries, M.M. Motazacker,M.P. Peppelenbosch, M. Mahmoudi, F. Rezaee, Sci. Rep. 5 (2015) 17259.
[54] J. Kreuter, D. Shamenkov, V. Petrov, P. Ramge, K. Cychutek, C. Koch-Brandt,R. Alyautdin, J. Drug Target. 10 (4) (2002) 317–325.
[55] S. Wagner, A. Zensi, S.L. Wien, S.E. Tschickardt, W. Maier, T. Vogel, F. Worek,C.U. Pietrzik, J. Kreuter, H. von Briesen, PLoS One 7 (3) (2012) e32568.
[56] K.G. Chen, G. Szakács, J.P. Annereau, F. Rouzaud, X.J. Liang, J.C. Valencia,C.N. Nagineni, J.J. Hooks, V.J. Hearing, M.M. Gottesman, Pigment Cell Res. 18 (2)(2005) 102–112.
[57] Patent No.: US 8.481,273 B2. Inventor: Mary C. Farach-Carson. Date of Patent: Jul.9, 2013.
[58] Patent No.: US 9,465,039 B2. Inventors: Koen Kas, Schilde (BE); Griet Vanpoucke,Ingooigem (BE); Piet Moerman, Deurle (BE). Date of Patent: Oct. 11, 2016.
[59] M. Grønborg, T.Z. Kristiansen, A. Iwahori, R. Chang, R. Reddy, N. Sato, H. Molina,O.N. Jensen, R.H. Hruban, M.G. Goggins, A. Maitra, A. Pandey, Mol. CellProteomics 5 (1) (2006) 157–171.
[60] B. Amin, A. Maurer, W. Voelter, A. Melms, H. Kalbacher, Curr. Med. Chem. 21 (13)(2014) 1544–1556.
[61] A.A. Kosova, S.N. Khodyreva, O.I. Lavrik, Biochemistry (Mosc) 82 (6) (2017)643–654.
[62] L. Mu-xing, Z. Hong, B. Xin-yu, L. Zhi-yu, Y. Xue-song, L. Huai, H. Zhen, H. Yue,Z. Jian-guo, Z. Jian-jun, Z. Ye-fan, Z. Dong-bin, C. Jian-qiang, Oncotarget 7 (52)(2016) 86630–86647.
[63] L. Faloppi, M. Bianconi, R. Giampieri, A. Sobrero, R. Labianca, D. Ferrari, S. Barni,E. Aitini, A. Zaniboni, C. Boni, F. Caprioni, S. Mosconi, S. Fanello, R. Berardi,A. Bittoni, K. Andrikou, M. Cinquini, V. Torri, M. Scartozzi, S. Cascinu, Italian groupfor the study of digestive tract Cancer (GISCAD), Oncotarget 6 (33) (2015)35087–35094.
[64] B.A. Katchman, I.T. Ocal, H.E. Cunliffe, Y.-H. Chang, G. Hostetter, A. Watanabe,J. LoBello, D.F. Lake, Breast Cancer Res. 15 (2) (2013) R28.
[65] F.H. Knebel, M. Uno, T.F. Galatro, L.P. Bellé, S.M. Oba-Shinjo, S.K.N. Marie,A. Campa, J. Neurooncol. 132 (3) (2017) 383–391.
[66] T. Nakano, T. Yoshino, T. Fujimura, S. Arai, A. Mukuno, N. Sato, K. Katsuoka, ActaDerm. Venereol. 95 (7) (2015) 783–786.
[67] M. Chromek, Z. Slamová, P. Bergman, L. Kovács, L. Podracká, I. Ehrén, T. Hökfelt,G.H. Gudmundsson, R.L. Gallo, B. Agerberth, A. Brauner, Nat. Med. 12 (2016)636–641.
M. del Pilar Chantada-Vázquez et al. Colloids and Surfaces B: Biointerfaces 177 (2019) 141–148
148

Contents lists available at ScienceDirect
Journal of Proteomics
journal homepage: www.elsevier.com/locate/jprot
Proteomic investigation on bio-corona of Au, Ag and Fe nanoparticles for thediscovery of triple negative breast cancer serum protein biomarkersMaría del Pilar Chantada-Vázqueza, Antonio Castro Lópezb, María García Vencec,Sergio Vázquez-Estévezd, Benigno Acea-Nebrile, David G. Calatayudf, Teresa Jardielf,Susana B. Bravoc,⁎, Cristina Núñeza,⁎
a Research Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde (SERGAS), 27002 Lugo, Spainb Breast Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde (SERGAS), 27002 Lugo, Spainc Proteomic Unit, Instituto de Investigaciones Sanitarias-IDIS, Complejo Hospitalario Universitario de Santiago de Compostela (CHUS), 15706 Santiago de Compostela,Spaind Oncology Division, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde (SERGAS), 27002 Lugo, Spaine Department of Surgery, Breast Unit, Complexo Hospitalario Universitario A Coruña (CHUAC), SERGAS, A Coruña, Spainf Department of Electroceramics, Instituto de Cerámica y Vidrio-CSIC, Kelsen 5, Campus de Cantoblanco, 28049 Madrid, Spain
A R T I C L E I N F O
Keywords:Triple negative breast cancer (TNBC)ProteomicsNanoparticlesBiomarkersSWATH-MSMass spectrometry (MS)
A B S T R A C T
Nowadays, there are no targeted therapeutic modalities for triple negative breast cancer (TNBC). This disease isassociated with poor prognosis and worst clinical outcome because of the aggressive nature of the tumor, de-layed diagnosis, and non-specific symptoms in the early stages. Therefore, identification of novel specific TNBCserum biomarkers for screening and therapeutic purposes remains an urgent clinical requirement.
New user-friendly and cheap methods for biomarker identification are needed, and nanotechnology offersnew opportunities. When dispersed in blood, nanoparticles (NPs) are covered by a protein shell termed “proteincorona” (PC). While alterations in protein patterns are challeging to detect by conventional blood analyses, PCacts as a “nano-concentrator” of serum proteins with affinity for NPs’ surface. So, the characterization of PCcould allow the detection of otherwise undetectable changes in protein concentration at an early stage of thedisease or after chemotherapy or surgery.
To explore this research idea, serum samples from 8 triple negative breast cancer (TNBC) patients and 8patients without malignancy were allowed to interact with gold nanoparticles (AuNPs: 10.02 ± 0.91 nm), silvernanoparticles (AgNPs: 9.73 ± 1.70 nm) and magnetic nanoparticles (MNPs: (9.30 ± 0.67 nm). Here, in orderto identify biomarker candidates in serum of TNBC patients, these nanomaterials were combined with elec-trophoretic separation (SDS-PAGE) to performed qualitative and quantitative comparisons of the serum pro-teomes of TNBC patients (n = 8) and healthy controls (n = 8) by liquid chromatography tandem-mass spec-trometry (LC-MS/MS) analysis. The results were validated through a sequential window acquisition of alltheoretical mass spectra (SWATH) analysis, performed in total serum samples (patients and controls) using thisapproach as a multiple reaction monitoring (MRM) analysis.Significance: It is well known that several proteins presented in human serum are important biomarkers for thediagnosis or prognosis of different diseases, as triple negative breast cancer (TNBC). Determining how nano-materials as gold nanoparticles (AuNPs: 10.02 ± 0.91 nm), silver nanoparticles (AgNPs: 9.73 ± 1.70 nm) andmagnetic nanoparticles (MNPs: (9.30 ± 0.67 nm) interact with human serum will assist not only in under-standing their effects on the biological system (biocompability and toxicity), but also to obtain information fordeveloping novel nanomaterials with high specificity and selectivity towards proteins with an important bio-logical function (prognostic and diagnostic protein biomarkers).
https://doi.org/10.1016/j.jprot.2019.103581Received 24 June 2019; Received in revised form 14 October 2019; Accepted 7 November 2019
⁎ Corresponding authors.E-mail addresses: [email protected] (S.B. Bravo), [email protected] (C. Núñez).
Journal of Proteomics 212 (2020) 103581
Available online 12 November 20191874-3919/ © 2019 Published by Elsevier B.V.
T

1. Introduction
Breast cancer (BC) is the most frequently diagnosed cancer and theleading cause of cancer death in women worldwide, accounting for 23%of total new cancer cases [1].
Mainly, triple negative breast cancer (TNBC) is a heterogeneousdisease that is characterized by a lack of estrogen receptor/proges-terone receptor (ER/PgR) expression and absence of human epidermalgrowth factor receptor 2 (HER2) overexpression or amplification. Thissubgroup accounts for 12–15% of all types of breast cancer and exhibitsa distinct molecular profile, clinical behavior, and response to therapy[2]. Notably, triple negative tumors are usually high grade and exhibitincreased aggressiveness, poor prognosis, and worst clinical outcome[3]. Because hormonal (tamoxifen) and HER2-directed (trastuzumab)therapies are not effective, TNBC patients are managed with standardchemotherapy; however, a high rate of local and systemic relapse isfrequently associated with treatment. Unfortunately, no useful bio-markers neither targeted therapeutic modalities exist for this breastcancer subtype [4].
It is well known that proteins secreted from tumor tissues have ahigher likelihood of reaching the systemic circulation and may, there-fore, serve as potential biomarkers for early detection [5]. Serum pro-teomics is a valuable tool that can facilitate comprehensive and sys-tematic elucidation of the serum proteome under both healthy anddisease conditions as well as identification of serum protein markersused for disease diagnosis and prognosis, particularly for identifyingbreast cancer-specific markers [5].
Current proteomic technologies that promote large-scale samplescreening and facilitate the identification of proteins associated withdisease and treatment are developing rapidly [6]. Mass spectrometry(MS), a powerful proteomics tool, has evolved to a high-throughputlevel, allowing rapid and accurate analysis of several thousand proteinsin a single study [7]. Several studies have addressed the possibility ofapplying MS proteome analysis to diagnostics of TNBC, revealing pro-tein patterns specific for patients with TNBC at either early or lateclinical stages [8]. The peptide markers identified with differentiatingpatterns include glycolytic enzymes (as for example MDH2, PGK1, TKT,Aldolase1), cytokeratins (CK7, 8, 9, 14, 17, 19), further structure pro-teins (vimentin, fibronectin, L-plastin), for NME1-NME2, lactoferrin,and members of the Annexin family, among others [9].
SWATH-MS is an emerging technique that combines deep proteomecoverage capabilities with quantitative consistency and accuracy [10].Mainly, SWATH-MS analysis offers several advantages, including highreproducibility and reliability of quantitative information, in discoveryproteomics [11]. Furthermore, SWATH-MS methods can be inter-changed to MRM approaches focused on the validated biomarkers.Therefore, SWATH-MS is an important tool not only for the biomarkerdiscovery but also for the development of preliminary validation studies[12].
However, currently available proteomic tests detect only a tinyfraction of potential biomarkers due to their deficient concentration inbiofluids, in addition to the ‘swamping’ effect, caused by non-specifichighly abundant molecules. The issue of signal-to-noise exceeds thecurrent capability of proteomic analysis and therefore limits the diag-nostic information that can be obtained [13].
To overcome these challenges, several approaches have been de-veloped, comprising depletion of high-abundance proteins that maskless abundant proteins [14,15], chromatographic or electrophoreticsample prefractionation, to decrease complexity before mass spectro-metric analyses [16,17], and direct isolation of preferred proteins [18].Nevertheless, none of the methods can provide a standard solution tobiomarker discovery or can give a reproducible diagnostic platform forestablishing biomarker guides. In this way, one promising way taken isthe use of nanoscale materials [19].
Nanotechnology-based platforms hold great promise in addressingthe above fundamental and technical issues of biomarker discovery to
overcome persistent deficiencies of conventional methods. Currently, itis well known that the surfaces of nanoparticles (NPs) are rapidlycovered by different types of biomolecules when they contact biologicalmedia called protein corona (PC) [20].
The protein composition and content in the corona depend onseveral parameters, including: i) physicochemical properties of the NPs(i.e. composition, size, shape, curvature, surface chemistry and surfacecharge, hydrophobicity/hydrophilicity) [21,22]; ii) characteristics ofbiological media (i.e. protein source, and temperature) [23,24]); iii)incubation time [25].
Notably, the composition of the protein corona varies amonghealthy individuals, as well as among patients with various diseases/medical conditions. Thus, the same NPs may have different proteincoronas in different individuals. These alterations are often small andchallenging to be detected by conventional blood analyses.
On the other side, the protein corona can act as a “nano-con-centrator” [26] of those serum proteins with affinity for the NP surface.Therefore, characterization of protein corona could allow detectingminor changes in protein concentration at the very early stages ofdisease development or even after chemotherapy or surgery (i.e., whenan alteration in circulating level of proteins could be undetectable byblood tests).
Keeping in mind that each disease is characterized by differentplasma/serum proteomes, inducing the formation of different PCs onthe same nanomaterial, M. Mahmoudi, et al. introduced the novelconcept of “personalized protein corona” (PPC) [27]. More specifically,depending on the type, period and severity of the disease (which de-termines the serum/plasma alterations), each patient may have a per-sonalized protein corona.
In the present study, gold nanoparticles (AuNPs: 10.02 ± 0.91 nm),silver nanoparticles (AgNPs: 9.73 ± 1.70 nm) and magnetic nano-particles (MNPs: (9.30 ± 0.67 nm) were used to pre-concentrate andseparate proteins from sera samples of eight patients with TNBC as wellas from eight healthy people. For protein biomarkers identification andquantification, the proteome map changes between both groups weredetected using a proteomic approach based on electrophoretic separa-tion (SDS-PAGE) and mass spectrometry (nLC-MS/MS).
2. Materials and methods
2.1. Chemicals and reagents
All reagents and solvents used were HPLC-grade or higher. Sodiumcitrate tribasic dihydrate, tannic acid, silver nitrate, ammonium hy-droxide,
iron(III) chloride hexahydrate and iron(II) sulfate heptahydrate,sodium borohydride (NaBH4), trypsin, trifluoroacetic acid, DL-Dithiothreitol (DTT), Iodoacetamide (IAA), acrylamide/bis-acrylamide30% solution (37.5:1), Glycerol 86–88%, Tris-base, Coomassie BrilliantBlue R250 (CBB), sodium carbonate, and the Sigma Marker wide range6.5–200 kDa were purchased from Sigma-Aldrich (St. Louis, MO, USA).Sodium dodecyl sulfate (SDS) and formaldehyde were purchased fromPanreac (Barcelona, Spain). β-mercaptoethanol was purchased fromMerck (Hohen-Brunn, Germany), and bromophenol-blue was purchasedfrom Riedel-de Haen (Seelze, Germany). Hydrogen tetrachloroaurate(III) hydrate (99.9%-Au) (49%Au) at 10%w/v was purchased fromStrem Chemicals (Newburyport, MA, USA). Ammonium bicarbonate(ambic) and formic acid were purchased from Fluka (Steinheim,Germany).
2.2. Instrumentation
Microscopic characterizations of AuNPs, AgNPs, and MNPs wereperformed by transmission electron microscopy (TEM) using a Jeol JEM1011 microscope. Samples for TEM were prepared by pipetting a dropof the colloidal dispersion onto an ultrathin carbon-coated copper grid
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
2

and allowing the solvent to evaporate. AuNPs, AgNPs and MNPs ζ-po-tentials were measured at 25°C before and after protein corona for-mation using a Malvern Zetasizer Nano ZS instrument. For ζ-potentialmeasurements samples were diluted in 1 mL milli-Q water and placed inZetasizer disposable cuvettes. A minimum of 3 measurements persample were made.
Power Pac Basic power supply from Bio-Rad (CA, USA) was used forsodium dodecyl sulfate- polyacrylamide gel electrophoresis (SDS-PAGE)protein separation. Protein quantification was accomplished by mea-suring the absorbance at 280 nm with the use of a Qubit™ 4Quantitation Starter Kit from Thermo Fisher Scientific. Gel image ac-quisition was carried out with a UVP PhotoDoc-ItTM Imaging Systemfrom Analytik Jena.
2.3. Synthesis of inorganic nanoparticles
2.3.1. Synthesis of citrate-gold nanoparticles (10.02 ± 0.91 nm)Gold nanoparticles (AuNPs) were synthesized by the citrate reduc-
tion method in aqueous solution [28]. Briefly, 60 ml of sodium citratetribasic solution (0.075% w/v) was heated to 100 °C, and then gold wasadded as 54 μL of 10% w/v of hydrogen tetrachloroaurate (III) hydratesolution. The reaction mixture was kept under reflux until a deep redcolor was detected. The solution of nanoparticles is chilled at roomtemperature and stored at 4 °C for a maximum of one month.
2.3.2. Synthesis of citrate-silver nanoparticles (9.73 ± 1.70 nm)Silver nanoparticles (AgNPs) were synthesized by the citrate re-
duction method in aqueous solution by the method reported by V.Puntes et al. [29]. A 100 mL volume of an aqueous solution containingsodium citrate (SC) (5 mM) and tannic acid (TA) (0.025 mM) was pre-pared and heated with a heating mantle in a three-neck round-bot-tomed flask for 15 min under vigorous stirring. A condenser was used toprevent the evaporation of the solvent. After boiling had commenced,1 mL of AgNO3 (25 mM) was injected into this solution. The solutionbecame bright yellow immediately. Resultant Ag NPs were purified bycentrifugation at 18,000×g to remove the excess of TA and furtherredispersed in Milli-Q-water before sample characterization.
2.3.3. Synthesis of Fe3O4 magnetic nanoparticles (9.30 ± 0.67 nm)The synthesis of magnetic nanoparticles was performed according to
a previously described procedure [30]. Briefly, 6 mL of concentratedammonium hydroxide and 4 mL of water saturated with N2 were mixedin a round-bottom flask under an inert atmosphere. Subsequently, inanother vessel, 1 g of FeCl3.6H2O and 0.25–0.5 g of FeSO4.7H2O weredissolved in 10 mL of water saturated with N2. After mixing both so-lutions, the system was kept under constant stirring for 80 min at 80 °C.The final reaction product was washed three times with deionizedwater and magnetically separated for 24 h at room temperature.
2.4. Sample resources
Venous blood samples were obtained from eight females with triplenegative breast cancer (TNBC) and eight disease-free individuals withthe use of VACUETTE® Serum Clot Activator Tubes (10 mL). The collectedblood samples were allowed to clot for 15 min and then centrifuged for5 min at 4°C and 1,800×g. Sera were transferred into clean plastictubes (1 mL) and immediately frozen at -80°C at Research Unit, HospitalUniversitario Lucus Augusti (HULA). Clinical features of TNBC tumors,including tumor size, histology, receptor status, clinical stage, andnodal status, are summarized in Table 1.
2.5. Sample preparation
2.5.1. Optimization of protein corona formation in serum using magneticnanoparticles
A series of serum sample aliquots were used for checking the effects
of sample pH and temperature on the high-abundance proteins deple-tion using DTT, MNP/protein ratios, and pH of the medium on thewashing steps.
2.5.1.1. Depletion of multiple high abundant proteins. Human serumaliquots (x8) were filtered with Miller-GP® Filter Unit (Millipore)with a size of 0.22 μm. Four aliquots of human serum (30 μL) weredepleted with dithiothreitol (DTT) according to the protocol describedby Warder el al. [31,32]. Briefly, fresh DTT 500 mM (3.3 μL) in milli-Qwater was mixed with 30 μL of human serum and vortex quickly.Samples were then incubated at room temperature until a viscous whiteprecipitate persisted (60 min), followed by centrifugation at 18,840 ×gfor 20 min. Supernatants were transferred to a clean tube before theprotein alkylation and nanoparticles (NPs) fractionation.
To evaluate the effects of sample pH and temperature on the high-abundance proteins depletion with DTT, four aliquots of human serum(30 μL) were depleted with dithiothreitol (DTT) following a modifica-tion of the protocol previously published by Arruda et al. [33]. Fouraliquots of 30 μL of human serum were mixed fresh DTT 500 mM(3.3 μL) in ambic (12.5 mmol L−1)] and vortexed. Samples were in-cubated for 60 min at 37°C, centrifuged at 13,000 ×g for 40 min toseparate supernatants to be alkylated and fractionated with nano-particles (NPs).
2.5.1.2. NPs protein alkylation and fractionation. After proteindepletion, the reduced SH-groups were alkylated with iodoacetic acid(IAA) for 45 min at room temperature and protected from light.Volumes of serum reduced and alkylated, were diluted to a finalvolume of 100 μL in Tris-HCl (0.1 mol L−1, pH 5.5), and mixed withMNPs (5 μg) to obtain the following MNP/protein ratios: 1:1, 1:2, 1:4,1:10. Then, all NPs-serum solutions were incubated at 25 °C withshaking (300 rpm) in a thermostatic bath for 30 min and then pelletswere harvested by centrifugation at 20,186 ×g for 30 min. To evaluatethe effects of sample pH on the stabilization of the protein corona in thewashing steps, a fraction of pellets were washed (x3) with 50 μL of Tris-HCl (0.1 mol L−1, pH 5.5) and another fraction with 50 μL of milli-Qwater (x3). In both cases, pellets were harvested again by centrifugationat 20,186 ×g for 30 min to remove unbound proteins.
2.5.2. Incubation of nanoparticles with serum samplesSerum aliquots (x2) belonging to the eight disease-free individuals
and eight triple negative breast cancer patients were depleted withdithiothreitol (DTT) following the method of Warder el al. [33,34].After that, the reduced SH-groups were alkylated with iodoacetic acid(IAA) at room temperature (45 min in the dark).
After protein reduction and alkylation, serum aliquots (x2) be-longing to the eight disease-free individuals and eight triple negativebreast cancer patients were incubated with AuNPs (10.02 ± 0.91 nm)
Table 1Clinical features of triple negative breast cancer tumors.
Characteristics Number
Patients 8Age (years) < 40 1
40–70 4> 70 3
Tumor size (cm) < 2 32–5 4> 5 1
Histological types Ductal invasive carcinoma 8Receptor status Triple negative 8Clinical stage I 1
II 4III 3
Nodal status N0 7N1 1
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
3

and AgNPs (9.73 ± 1.70 nm) (4 aliquots per individual, 2 with eachnanoparticle type) following the method described by C. Núñez et al.[34]. Briefly, 75 μL of AuNPs (10.02 ± 0.91 nm) and 75 μL of AgNPs(9.73 ± 1.70 nm) were added to each different serum aliquots (×2)belonging to the eight disease-free individuals and eight triple negativebreast cancer patients (4 aliquots per individual, 2 with each nano-particle type), followed by the addition of 40 μL of citrate/citric acidbuffer to a final pH of 5.8. Then, all NPs-serum solutions were in-cubated at 37 °C with shaking in a thermostatic bath for 30 min. Pelletswere harvested by centrifugation at 18,840 ×g for 30 min. In all cases,pellets containing proteins bound to nanoparticles were washed threetimes with 25 μL citrate/citric acid buffer and harvested again by cen-trifugation at 18,840 ×g for 30 min to remove unbound proteins.
In the particular case of magnetic nanoparticles, each different re-duced and alkylated serum aliquots (x2) from disease-free individuals(n = 8) and negative breast cancer patients (n = 8) were incubated(shaking at 300 rpm, 25 °C, 30 min) with 5 μL of MNPs(9.30 ± 0.67 nm) after the addition of 87 μL of Tris-HCl (0.1 mol L−1,pH 5.5).
After centrifuging (15,000 ×g, 30 min), pellets were separated andwashed (x3) with 50 μL of Tris-HCl (0.1 mol L−1, pH 5.5) and cen-trifuged again (20,186×g, 30 min).
2.5.3. Gel electrophoresisAfter that, pellets were reconstituted in 10 μL of a buffer with 0.2 M
Tris-HCl, 2% w/v SDS and 20% v/v glycerol. This 10 μL was mixed with4 μL of SDS-PAGE loading buffer (10% w/v SDS, Tris-Base 40 mM, pH6.8, 50% v/v glycerol, 0.1% v/v bromophenol blue, 10% v/v β-mer-captoethanol) in a final volume of 20 μL. Then, all samples were de-natured by heating at 100°C for 5 min and loaded into a 10% acryla-mide/bis-acrylamide, stacking gel/12.5% acrylamide/bis-acrylamiderunning gel, of 1 mm thickness, and separated at 180 V (constant vol-tage) for 120 min. After electrophoresis, the gel was fixed for 30 min-utes with 40% (v/v) ethanol and 10% (v/v) acetic acid and then stainedovernight with Colloidal Coomassie Blue [35]. Gels were rinsed withdistilled water and a 0.5 M sodium chloride solution until a clearbackground was observed. Gel imaging was carried out with a UVPPhotoDoc-ItTM Imaging System.
2.5.4. In-gel protein digestionProtein bands were excised manually and transferred to 2.5-mL Lo-
Bind tubes, and then washed twice with water and with 50% (v/v)acetonitrile/25 mM ammonium bicarbonate (ambic) until the bluecolor disappeared.
Before the trypsin digestion, gel spots were washed with 25 mMambic and dehydrated with acetonitrile. Then, 30 μL of trypsin(20 ng μL−1 in 12.5 mM ambic/2% (v/v) acetonitrile) was added to thegel spots and incubated for 60 min at 0°C.
After this time, gel spots were inspected, trypsin solution not ab-sorbed into the gel was removed, and the gels were covered with 100 μLof 12.5 mM ambic. Samples were incubated for 12 h at 37°C. Then 50 μLof 5% (v/v) formic acid was added, and the supernatant was transferredto a new Lo-Bind tube and the peptides were further extracted from thegel twice with 50% (v/v) acetonitrile/0.1% (v/v) trifluoroacetic acid(TFA) (x3) and acetonitrile (ACN) (x1). Samples were dried-down andstored at -20 °C [36].
2.6. Protein identification by mass spectrometry (LC-MS/MS) and dataanalysis
Digested peptides of each sample were separated using ReversePhase Chromatography. The gradient was developed using a micro li-quid chromatography system (Eksigent Technologies nanoLC 400,SCIEX) coupled to high-speed Triple TOF 6600 mass spectrometer(SCIEX) with a microflow source. The analytical column used was asilica-based reversed phase column Chrom XP C18 150 × 0.30 mm,
3 mm particle size and 120 Å pore size (Eksigen, SCIEX). The trapcolumn was a YMC-TRIART C18 (YMC Technologies, Teknokroma witha 3 mm particle size and 120 Å pore size, switched on-line with theanalytical column. The loading pump delivered a solution of 0.1%formic acid in water at 10 μL/min. The micro-pump provided a flow-rate of 5 μL/min and was operated under gradient elution conditions,using 0.1% formic acid in water as mobile phase A, and 0.1% formicacid in acetonitrile as mobile phase B. Peptides were separated using a25 minutes gradient ranging from 2% to 90% mobile phase B (mobilephase A: 2% acetonitrile, 0.1% formic acid; mobile phase B: 100%acetonitrile, 0.1% formic acid). The injection volume was 4 μL.
Data acquisition was carried out in a TripleTOF 6600 System(SCIEX, Foster City, CA) using a Data dependent workflow. Source andinterface conditions were as follows: ion spray voltage floating (ISVF)5500 V, curtain gas (CUR) 25, collision energy (CE) 10 and ion sourcegas 1 (GS1) 25. The instrument was operated with Analyst TF 1.7.1software (SCIEX, USA). Switching criteria were set to ions greater thanmass to charge ratio (m/z) 350 and smaller than m/z 1400 with acharge state of 2–5, mass tolerance 250 ppm and an abundancethreshold of more than 200 counts (cps). Former target ions were ex-cluded for 15s. The instrument was automatically calibrated every4 hours using as external calibrant tryptic peptides from PepcalMix(Sciex).
After MS/MS analysis, data files were processed usingProteinPilotTM 5.0.1 software from Sciex, which uses the algorithmParagonTM for database search and ProgroupTM for data grouping. Datawere searched using a Human-specific UniProt database. False dis-covery rate was performed using a non-linear fitting method displayingonly those results that reported a 1% Global false discovery rate orbetter [37,38].
2.7. Protein quantification by SWATH (Sequential Window Acquisition ofall Theoretical Mass Spectra)
2.7.1. Creation of the spectral libraryTo construct the MS/MS spectral libraries, the peptide solutions
were analyzed by a shotgun data-dependent acquisition (DDA) ap-proach by micro-LC-MS/MS. To get a good representation of the pep-tides and proteins present in all samples, pooled vials of samples fromeach group (control and triple negative breast cancer patients) wereprepared using equal mixtures of the original samples. 4 μL (4 mg) ofeach pool was separated into a micro-LC system Ekspert nLC425(Eksigen. Dublin. CA. USA) using a column Chrom XP C18150 × 0.30 mm. 3 mm particle size and 120 Å pore size (Eksigent,Sciex) at a flow rate of 5 μL/min. Water and ACN, both containing 0.1%formic acid, were used as solvents A and B, respectively. The gradientrun consisted of 5–95% B for 30 min, 5 min at 90% B and finally 5 minat 5% B for column equilibration, for a total run time of 40 min. Whenthe peptides eluted, they were directly injected into a hybrid quadru-pole-TOF mass spectrometer Triple TOF 6600 (Sciex, Redwood City.CA. USA) operated with a data-dependent acquisition system in positiveion mode. A Micro source (Sciex) was used for the interface betweenmicroLC and MS, with an application of 2600 V voltage. The acquisitionmode consisted of a 250 ms survey MS scan from 400 to 1250 m/zfollowed by an MS/MS scan from 100 to 1500 m/z (25 ms acquisitiontime) of the top 65 precursor ions from the survey scan, for a total cycletime of 2.8 s. The fragmented precursors were then added to a dynamicexclusion list for 15 s; any singly charged ions were excluded from theMS/MS analysis.
The peptide and protein identifications were performed usingProtein Pilot software (version 5.0.1. Sciex) with a Data were searchedusing a Human-specific UniProt database, specifying iodoacetamide asCys alkylation. The false discovery rate (FDR) was set to 1 for bothpeptides and proteins. The MS/MS spectra of the identified peptideswere then used to generate the spectral library for SWATH peak ex-traction using the add-in for PeakView Software (version 2.2. Sciex)
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
4

MS/MSALL with SWATH Acquisition MicroApp (version 2.0. Sciex).Peptides with a confidence score above 99% (as obtained from ProteinPilot database search) were included in the spectral library).
2.7.2. Relative quantification by SWATH acquisitionSWATH-MS (Sequential Window Acquisition of all Theoretical Mass
Spectra) acquisition was performed on a TripleTOF® 6600 LC-MS/MSsystem (Sciex). Samples from control and triple negative breast cancerpatients were analyzed using data-independent acquisition (DIA)method (30 total samples). Each sample (4 μL (from a mg/ml solution)was analyzed using the LC-MS equipment and LC gradient describedabove for building the spectral library but instead using the SWATH-MSacquisition method. The method consisted of repeating a cycle thatconsisted of the acquisition of 65 TOF MS/MS scans (400–1500 m/z,high sensitivity mode, 50 ms acquisition time) of overlapping sequen-tial precursor isolation windows of variable width (1 m/z overlap)covering the 400–1250 m/z mass range with a previous TOF MS scan(400–1500 m/z. 50 ms acquisition time) for each cycle. The total cycletime was 6.3 s. For each sample set, the width of the 100 variablewindows was optimized according to the ion density found in the DDAruns using a SWATH variable window calculator worksheet from Sciex.
2.7.3. Data analysisThe targeted data extraction of the fragment ion chromatogram
traces from the SWATH runs was performed by PeakView (version 2.2)using the SWATH Acquisition MicroApp (version 2.0). This applicationprocessed the data using the spectral library created from the shotgundata. Up to ten peptides per protein and seven fragments per peptidewere selected, based on signal intensity; any shared and modifiedpeptides were excluded from the processing. Five-minute windows and30 ppm widths were used to extract the ion chromatograms; SWATHquantitation was attempted for all proteins in the ion library that wereidentified by ProteinPilot with an FDR below 1%.
The retention times from the peptides that were selected for eachprotein were realigned in each run according to the iRT peptides spikedin each sample and eluted along the whole-time axis. The extracted ionchromatograms were then generated for each selected fragment ion; thepeak areas for the peptides were obtained by summing the peak areasfrom the corresponding fragment ions. PeakView computed an FDR anda score for each assigned peptide according to the chromatographic andspectral components; only peptides with an FDR below 1% were usedfor protein quantitation. Protein quantitation was calculated by addingthe peak areas of the corresponding peptides.
The integrated peak areas (processed, mrkvw files from PeakView)were directly exported to the MarkerView software (Sciex) for relativequantitative analysis. The export will generate three files containingquantitative information about individual ions, the summed intensity ofdifferent ions for a particular peptide, and the summed intensity ofdifferent peptides for a specific protein. MarkerView has been used forthe analysis of SWATH-MS data reported in other proteomics studies[39,40] because of its data-independent method of quantitation. Mar-kerView uses processing algorithms that accurately find chromato-graphic and spectral peaks direct from the raw SWATH data. Dataalignment by MarkerView compensates for minor variations in bothmass and retention time values, ensuring that identical compounds indifferent samples are accurately compared to one another.
To control for possible uneven sample loss across the differentsamples during the sample preparation process, we performed a globalnormalization based on the total sum of all the peak areas extractedfrom all the peptides and transitions across the replicates of eachsample. Unsupervised multivariate statistical analysis using principalcomponent analysis (PCA) was performed to compare the data acrossthe samples. The average MS peak area of each protein was derivedfrom the biological replicates of the SWATH-MS of each sample fol-lowed by Student’s t-test analysis using the MarkerView software forcomparison among the samples based on the averaged area sums of all
the transitions derived for each protein. The t-test will indicate howwell each variable distinguishes the two groups, reported as a p-value.To set of differentially expressed proteins (p-value < 0.05) with a 1.5fold in- or decrease was selected.
Functional analysis was performed by FunRich open access software(Functional Enrichment analysis tool) for functional enrichment andinteraction network analysis (http://funrich.org/index.html).
2.8. TNBC biomarkers validation
A SWATH-MS analysis was performed using the same conditionsdescribed in Section 2.7. In the validation phase, total serum samplesfrom control and triple negative breast cancer patients previously de-pleted with DTT were used.
To perform the biomarker validation, the SWATH library was per-formed using not only serum pools from incubation with the differentnanoparticles but also pools of total serum samples. Therefore, weimprove our library and perform better protein quantification toachieve biomarker validation.
3. Results
Following the synthetic methods described by R. López-Cortés [30],V. Puntes et al. [31] and F. Schüth et al. [32], AuNPs(10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and MNPs(9.30 ± 0.67 nm) were successfully obtained, respectively.
The sizes and ζ-potential of AuNPs (10.02 ± 0.91 nm), AgNPs(9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm) were examined beforeand after their incubation with two pools of human blood serum fromhealthy individuals and triple negative breast cancer patients, followingthe conditions described in Section 2.5.2.
TEM and ζ-potential measurements after the incubation of AuNPs(10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and MNPs(9.30 ± 0.67 nm) with serum demonstrated that, in all cases, the sizedid not change significantly and the surface charge remained negative(see Figs. S1–S10). Upon serum incubation, the mean particle surfacecharge of the AuNPs (10.02 ± 0.91 nm) increased (became less nega-tive) from -37.0 mV to -29.7 mV, and the same measurement for theMNPs (9.30 ± 0.67 nm) increased from -30.5 mV to -29.3 mV.However, upon serum incubation the mean particle surface charge ofthe AgNPs (9.73 ± 1.70 nm) decreased (became more negative) from-27.4 mV to -30.0 mV. These results are in agreement with previousstudies suggesting that negatively charged NPs do not exclusively in-teract with positively charged proteins, as electrostatic interactions arenot the only driving force behind NP-corona interactions [41,42]. In-terestingly, for AuNPs (10.02 ± 0.91 nm) and MNPs(9.30 ± 0.67 nm), where the ζ-potential was shifted toward less ne-gative values, it could be suggested preferential interaction with posi-tively charged proteins. The presence of negatively charged proteinscan be explained by a sequential model of protein binding, in whichpositively charged proteins initially bind the NP, followed by negativelycharged ones [45,46].
3.1. Optimization of parameters for the protein corona formation in serumusing MNPs
As mentioned above, a great number of variables could influencethe efficiency of protein adsorption on the MNPs surface [43]. For thisreason, three parameters were evaluated: (i) the effects of sample pHand temperature on the depletion of high-abundance proteins presentedin serum using DTT, (ii) MNP/protein ratios; (iii) and pH of the mediumon the washing steps. For this study, shaking and incubation tempera-ture were previously defined as 300 rpm and 25 °C, respectively.
In order to evaluate the effects of sample pH and temperature on thedepletion of high-abundance proteins, four human serum samples (x2)(30 μL) were depleted with fresh DTT 500 mM (3.3 μL) in milli-Q H2O
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
5

for 60 min at room temperature (protocol of Warder el al. [33,34]), andfour human serum samples (x2) (30 μL) were depleted with fresh DTT500 mM (3.3 μL) in ambic (12.5 mmol L−1) for 60 min at 37°C (mod-ification of protocol described by Arruda [35]). In both cases, after theincubation and centrifugation, supernatants were transferred to a cleantube before the protein alkylation and nanoparticles (NPs) fractiona-tion. Depletion with fresh DTT 500 mM in milli-Q for 60 min at roomtemperature showed more reproducible results (see Fig. S4).
Protein concentration is another critical parameter that may affectthe capacity and kinetics of protein adsorption. To investigate the in-fluence of the MNP/protein ratio on the formation of the proteincorona, volumes of serum reduced and alkylated (x2) were mixed withMNPs (9.30 ± 0.67 nm), at MNP/protein ratios of 1:1, 1:2, 1:4, and1:10 (see Section 2.5.1). Maintaining the amount of adsorbent (i.e.,MNPs) constant and increasing the protein concentration, would beexpected to lead to a decrease of available adsorption sites, reducing theefficiency of protein removal [44]. As a compromise between MNPs andprotein corona formation, the 1:2 ratio (MNP/protein) was then se-lected for future experiments (see Fig. S5).
The pH value is an essential parameter because it influences thecharge state of proteins, therefore influencing their interaction withMNPs [45]. To evaluate the effects of sample pH on the stabilization ofthe protein corona in the washing steps, a fraction of pellets were wa-shed three times with 50 μL Tris-HCl (0.1 mol L−1, pH 5.5) and anotherfraction was washed three times with milli-Q water. The first one wasselected as the preferred method, because the washes with milli-Qwater promoted the destabilization of the protein corona formedaround the MNPs, due to the modifications of the pH (data not shown).
3.2. Serum fraction preparation and protein corona purification (patientsvs. controls)
Serum aliquots (x2) belonging to the eight disease-free individualsand serum samples from eight triple negative breast cancer patientswere depleted with dithiothreitol (DTT) according to the protocol de-scribed by Warder el al. [33,34]. After that, the reduced SH-groupswere alkylated with iodoacetic acid (IAA) for 45 min at room tem-perature and protected from light.
After protein reduction and alkylation, serum aliquots (x2) be-longing to the eight disease-free individuals and eight triple negativebreast cancer patients were incubated with AuNPs (10.02 ± 0.91 nm),AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm) (6 aliquots perindividual, 2 with each nanoparticle type) and further processed asdescribed in Section 2.5.2.
Two protein fractions were thus obtained in each case, one in thesupernatant and the second one attached to the surface of each nano-particles types (protein corona). Then, protein fractions (AuNPs-proteincorona, AgNPs-protein corona, MNPs-protein corona) were separatelyloaded onto a 1D-SDS-PAGE. Proteins were separated and, afterstaining, gel bands were excised and submitted to the sample incuba-tion described in Section 2.5.4. The resulting peptides were then ana-lyzed by mass spectrometry (LC-MS/MS) for protein identification.
Fig. 1 shows the 1D gels for the protein corona formed around thethree different NPs (AuNPs-protein corona, AgNPs-protein corona,MNPs-protein corona) visible after Coomassie staining. As may be seen,it is quickly noted that there is a difference in the intensity of the bandson the gel profiles for the healthy controls (from C1 to C8) and thepatients (from P1 to P8) for each type of nanoparticle. However, noconclusion can be drawn unless the proteins are identified.
As Fig. 2 shows, 192, 161 and 142 proteins were commonly de-tected in the protein corona formed around AuNPs, AgNPs, and MNPs,after their incubation with serum samples of the eight triple negativebreast cancer patients and eight healthy controls, respectively.
In the particular case of all serum samples from healthy controls,285, 292 and 206 were identified on the surface of AuNPs(10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and MNPs
(9.30 ± 0.67 nm), respectively. From them, 149 proteins were com-monly detected in the protein corona of all three different types of NPs.However, 202 different proteins were found on the three distinct NPssurface: 71 different proteins on the AuNPs, 85 on the AgNPs, and 46individual proteins on the MNPs (see Table S1).
Furthermore, 231, 206 and 203 were found in the surface of AuNPs(10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and MNPs(9.30 ± 0.67 nm) after their incubation with all serum aliquots (x2)from eight triple negative breast cancer patients (6 protein samples perindividual: 2 incubated with AuNPs, 2 with AgNPs and 2 with MNPs)(see Table S2). A total of 138 proteins were commonly found in theprotein corona of all three different types of NPs. However, 142 dif-ferent proteins were found in the three different NPs surface: 56 dif-ferent proteins on the AuNPs, 33 on the AgNPs and 53 individualproteins on the MNPs.
Fractionation of the proteome using AuNPs, AgNPs, and MNPs al-lows for the identification of 39 (see Table 2), 45 (see Table 3) and 61(see Table 4) protein biomarkers in the pellet of all patient samples,respectively (see Fig. 3). Remarkably, these proteins were not identifiedin the healthy control. These potential biomarkers came from differentcellular components; most of them from the nucleus and cytoplasm (seeFig. 4). Protein biomarkers also showed different functionality and areconstituted by proteins implicated in the immune response, followed byproteins with an enzymatic function, structural, transporter, in-flammatory, signal transduction, and with antibiotic/antibacterialproperties.
Importantly, the GRF-type zinc finger domain-containing protein 1(protein ZGRF1) was identified in the protein corona of AuNPs, AgNPsand MNPs after their incubation with the serum samples of all triplenegative breast cancer patients. Zinc-finger proteins (ZNFs) are one ofthe most abundant groups of proteins and have a wide range of mole-cular functions. Given the wide variety of zinc-finger domains, ZNFscan interact with DNA, RNA, PAR (poly-ADP-ribose) and other proteins.Thus, ZNFs are involved in the regulation of several cellular processes.ZNFs are implicated in transcriptional regulation, ubiquitin-mediatedprotein degradation, signal transduction, actin targeting, DNA repair,cell migration, and numerous other processes [46]. Notably, over-expression of similar zinc finger proteins has been shown to promotecell growth and metastasis in laryngeal squamous cell carcinoma,glioma, non-small cell lung cancer, gastric cancer, oral squamous cellcarcinoma, gallbladder cancer, and breast cancer [47], and also in triplenegative breast cancers [48].
Matrix metalloproteinase-9 (MMP9) was identified in the proteincorona of AuNPs and AgNPs after their incubation with the serumsamples of all triple negative breast cancer patients. Members of thematrix metalloproteinase (MMP) family have been identified as poorprognosis markers for breast cancer patients and as drivers of manyfacets of the tumor phenotype in experimental models [49]. Studies ofthe pathological processes involved in tumor progression and metas-tasis revealed matrix metalloproteinases (MMPs) as prominent mole-cules engaged in shaping the tumor microenvironment and drivingcancer progression and metastasis [50,51]. Mainly, MMP9 was in-vestigated as a potential tumor marker in breast cancer [52]. MMP-9 isone of 70 genes in the Rosetta poor prognosis signature for breastcancer patients [53], the basis for the clinically implemented Mam-maprint prognostic assay (Agendia Inc., Irvine, CA). MMP-9 was alsohighly expressed in node-positive tumors and the preoperative bloodserum of patients, but MMP-9 activity was appreciably inhibited inblood serum samples collected after surgery.
Lebercilin and Immunoglobulin lambda variable 3-27 (LV327) wereidentified in the protein corona of AuNPs and MNPs after their incuba-tion with the serum samples of all triple negative breast cancer patients.While the protein expression of lebercilin was observed in several tissuecancers like colorectal cancer, breast cancer, prostate cancer, lung cancer(The Human Protein Atlas, https://www.proteinatlas.org/ENSG00000157578-LCA5L/pathology), immunoglobulin free light
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
6
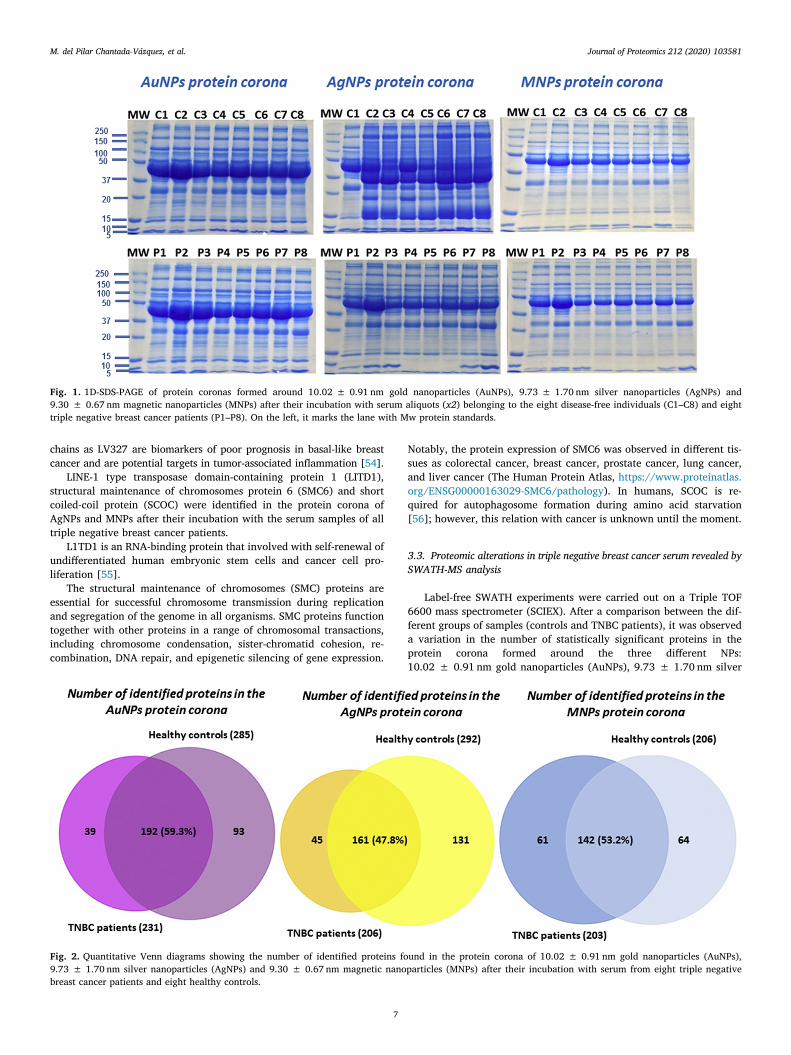
chains as LV327 are biomarkers of poor prognosis in basal-like breastcancer and are potential targets in tumor-associated inflammation [54].
LINE-1 type transposase domain-containing protein 1 (LITD1),structural maintenance of chromosomes protein 6 (SMC6) and shortcoiled-coil protein (SCOC) were identified in the protein corona ofAgNPs and MNPs after their incubation with the serum samples of alltriple negative breast cancer patients.
L1TD1 is an RNA-binding protein that involved with self-renewal ofundifferentiated human embryonic stem cells and cancer cell pro-liferation [55].
The structural maintenance of chromosomes (SMC) proteins areessential for successful chromosome transmission during replicationand segregation of the genome in all organisms. SMC proteins functiontogether with other proteins in a range of chromosomal transactions,including chromosome condensation, sister-chromatid cohesion, re-combination, DNA repair, and epigenetic silencing of gene expression.
Notably, the protein expression of SMC6 was observed in different tis-sues as colorectal cancer, breast cancer, prostate cancer, lung cancer,and liver cancer (The Human Protein Atlas, https://www.proteinatlas.org/ENSG00000163029-SMC6/pathology). In humans, SCOC is re-quired for autophagosome formation during amino acid starvation[56]; however, this relation with cancer is unknown until the moment.
3.3. Proteomic alterations in triple negative breast cancer serum revealed bySWATH-MS analysis
Label-free SWATH experiments were carried out on a Triple TOF6600 mass spectrometer (SCIEX). After a comparison between the dif-ferent groups of samples (controls and TNBC patients), it was observeda variation in the number of statistically significant proteins in theprotein corona formed around the three different NPs:10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nm silver
Fig. 1. 1D-SDS-PAGE of protein coronas formed around 10.02 ± 0.91 nm gold nanoparticles (AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs) and9.30 ± 0.67 nm magnetic nanoparticles (MNPs) after their incubation with serum aliquots (x2) belonging to the eight disease-free individuals (C1–C8) and eighttriple negative breast cancer patients (P1–P8). On the left, it marks the lane with Mw protein standards.
Fig. 2. Quantitative Venn diagrams showing the number of identified proteins found in the protein corona of 10.02 ± 0.91 nm gold nanoparticles (AuNPs),9.73 ± 1.70 nm silver nanoparticles (AgNPs) and 9.30 ± 0.67 nm magnetic nanoparticles (MNPs) after their incubation with serum from eight triple negativebreast cancer patients and eight healthy controls.
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
7

Table2
Sele
ctio
nof
iden
tified
sing
le-d
etec
ted
coro
napr
otei
nsbo
und
toth
e10
.02
±0.
91nm
AuN
Psaf
ter
30m
inin
cuba
tion
and
subs
eque
ntw
ashi
ng.
Prot
ein
nam
eU
niPr
otna
me
Entr
yna
me
Gen
eM
ass
(kD
a)Fu
nctio
n
Act
in,c
ytop
lasm
ic2
ACT
G_H
UM
AN
P632
61A
CTG
141
.7Ce
llm
obili
tyA
typi
calk
inas
eCO
Q8B
,mito
chon
dria
lCO
Q8B
_HU
MA
NQ
96D
53CO
Q8B
60.1
Invo
lved
inth
ebi
osyn
thes
isof
coen
zym
eQ
Beta
-ure
idop
ropi
onas
eBU
P1_H
UM
AN
Q9U
BR1
UPB
143
.1Ca
taly
ticac
tivity
Bref
eldi
nA
-inhi
bite
dgu
anin
enu
cleo
tide-
exch
ange
prot
ein
3BI
G3_
HU
MA
NQ
5TH
69A
RFG
EF3
240.
6Pa
rtic
ipat
esin
the
regu
latio
nof
syst
emic
gluc
ose
hom
eost
asis
85/8
8kD
aca
lciu
m-in
depe
nden
tpho
spho
lipas
eA
2PL
PL9_
HU
MA
NO
6073
3PL
A2G
689
.9Ca
taly
ticac
tivity
Cilia
-and
flage
lla-a
ssoc
iate
dpr
otei
n10
0CP
100_
HU
MA
NQ
494V
2CF
AP1
0071
.1Pl
ays
aro
lein
cilia
ry/fl
agel
lar
mot
ility
byre
gula
ting
the
asse
mbl
yan
dth
eac
tivity
ofax
onem
alin
ner
dyne
inar
mCo
agul
atio
nfa
ctor
XIFA
11_H
UM
AN
P039
51F1
170
.1Ca
taly
ticac
tivity
Cont
actin
-ass
ocia
ted
prot
ein-
like
2CN
TP2_
HU
MA
NQ
9UH
C6CN
TNA
P214
8.2
Play
sa
role
inth
efo
rmat
ion
offu
nctio
nald
istin
ctdo
mai
nscr
itica
lfor
salta
tory
cond
uctio
nof
nerv
eim
puls
esin
mye
linat
edne
rve
fiber
sD
NA
topo
isom
eras
e1
TOP1
_HU
MA
NP1
1387
TOP1
90.7
Cata
lyct
icac
tivity
Dyn
ein
asse
mbl
yfa
ctor
1,ax
onem
alD
AA
F1_H
UM
AN
Q8N
EP3
DN
AA
F180
.0Pl
ays
aro
lein
cyto
plas
mic
prea
ssem
bly
ofdy
nein
arm
sE3
ubiq
uitin
-pro
tein
ligas
eSH
PRH
SHPR
H_H
UM
AN
Q14
9N8
SHPR
H19
3.1
Enzy
me
invo
lved
inD
NA
repa
irEs
trog
ensu
lfotr
ansf
eras
eST
1E1_
HU
MA
NP4
9888
SULT
1E1
35.1
Cata
lytic
activ
ity.M
aypl
aya
role
inth
ere
gula
tion
ofes
trog
enre
cept
orac
tivity
bym
etab
oliz
ing
free
estr
adio
l[F
-act
in]-
mon
ooxy
gena
seM
ICA
L3M
ICA
3_H
UM
AN
Q7R
TP6
MIC
AL3
224.
3Ca
taly
ticac
tivity
Glia
lfibr
illar
yac
idic
prot
ein
GFA
P_H
UM
AN
P141
36G
FAP
49.9
Cell-
spec
ific
mar
ker
duri
ngth
ede
velo
pmen
tof
the
cent
raln
ervo
ussy
stem
His
tone
-lysi
neN
-met
hyltr
ansf
eras
e2D
KMT2
D_H
UM
AN
O14
686
KMT2
D59
3.4
Cata
lytic
activ
ity.A
cts
asa
coac
tivat
orfo
res
trog
enre
cept
orby
bein
gre
crui
ted
byES
R1,t
here
byac
tivat
ing
tran
scri
ptio
nIm
mun
oglo
bulin
lam
bda
vari
able
3-27
LV32
7_H
UM
AN
P017
18IG
LV3-
2712
.2Im
mun
ere
spon
seIn
osito
lhex
akis
phos
phat
eki
nase
2IP
6K2_
HU
MA
NQ
9UH
H9
IP6K
249
.2Ca
taly
ticac
tivity
Inos
itol1
,4,5
-tris
phos
phat
ere
cept
orty
pe3
ITPR
3_H
UM
AN
Q14
573
ITPR
330
4.1
Rece
ptor
for
inos
itol1
,4,5
-tris
phos
phat
eIn
trafl
agel
lar
tran
spor
tpro
tein
74ho
mol
ogIF
T74_
HU
MA
NQ
96LB
3IF
T74
69.2
Tran
spor
ter
activ
ityJa
nus
kina
sean
dm
icro
tubu
le-in
tera
ctin
gpr
otei
n1
JKIP
1_H
UM
AN
Q96
N16
JAKM
IP1
73.2
Play
sa
role
inth
em
icro
tubu
le-d
epen
dent
tran
spor
tof
the
GA
BA-B
rece
ptor
Lebe
rcili
nLC
A5_
HU
MA
NQ
86VQ
0LC
A5
80.5
Tran
spor
ter
activ
ityM
aFF-
inte
ract
ing
prot
ein
MA
FIP_
HU
MA
NQ
8WZ3
3M
AFI
P13
.9In
hibi
tsce
llgr
owth
and
colo
ny-fo
rmin
geffi
cien
cyM
atri
xm
etal
lopr
otei
nase
-9M
MP9
_HU
MA
NP1
4780
MM
P978
.5Ca
taly
ticac
tivity
Mic
rotu
bule
-ass
ocia
ted
prot
ein
2M
TAP2
_HU
MA
NP1
1137
MA
P219
9.5
Stab
ilize
sth
em
icro
tubu
les
agai
nstd
epol
ymer
izat
ion
Muc
olip
in-3
MCL
N3_
HU
MA
NQ
8TD
D5
MCO
LN3
64.2
Pays
aro
lein
the
regu
latio
nof
mem
bran
etr
affick
ing
even
tsM
ultid
rug
resi
stan
cepr
otei
n1
MD
R1_H
UM
AN
P081
83A
BCB1
141.
5Re
spon
sibl
efo
rde
crea
sed
drug
accu
mul
atio
nin
mul
tidru
g-re
sist
antc
ells
Neb
ulet
teN
EBL_
HU
MA
NO
7604
1N
EBL
116.
4Pl
ays
anim
port
ant
role
inth
eas
sem
bly
ofth
eZ-
disk
Nuc
leop
orin
NU
P188
hom
olog
NU
188_
HU
MA
NQ
5SRE
5N
UP1
8819
6.1
May
func
tion
asa
com
pone
ntof
the
nucl
ear
pore
com
plex
(NPC
)PC
mem
bran
ere
crui
tmen
tpr
otei
n2
AM
ER2_
HU
MA
NQ
8N7J
2A
MER
269
.5N
egat
ive
regu
lato
rof
the
cano
nica
lWnt
sign
alin
gpa
thw
ayin
volv
edin
neur
oect
oder
mal
patt
erni
ngPr
otei
nEL
YSEL
YS_H
UM
AN
Q8W
YP5
AH
CTF1
252.
5Re
quir
edfo
rth
eas
sem
bly
ofa
func
tiona
lnuc
lear
pore
com
plex
(NPC
)on
the
surf
ace
ofch
rom
osom
esPr
otei
nph
osph
atas
e1
regu
lato
rysu
buni
t26
PPR2
6_H
UM
AN
Q5T
8A7
PPP1
R26
127.
3In
hibi
tsph
osph
atas
eac
tivity
ofpr
otei
nph
osph
atas
e1
(PP1
)co
mpl
exes
.May
posi
tivel
yre
gula
tece
llpr
olife
ratio
n.Pr
otei
nZG
RF1
ZGRF
1_H
UM
AN
Q86
YA3
ZGRF
123
6.6
Zinc
ion
bind
ing
that
inhi
bits
fact
ors
Xaan
dXI
aof
the
coag
ulat
ion
casc
ade
Puta
tive
tran
smem
bran
epr
otei
nen
code
dby
LIN
C004
77CL
067_
HU
MA
NQ
96M
19LI
NC0
0477
18.2
Prod
uct
ofa
dubi
ous
CDS
pred
ictio
n.M
aybe
ano
n-co
ding
RNA
Seri
nepr
otea
se33
PRS3
3_H
UM
AN
PRSS
33PR
SS33
29.8
Cata
lytic
activ
itySe
rine
-pro
tein
kina
seA
TMA
TM_H
UM
AN
Q13
315
ATM
350.
7A
ctiv
ates
chec
kpoi
ntsi
gnal
ing
upon
doub
lest
rand
brea
ks(D
SBs)
,apo
ptos
isan
dge
noto
xic
stre
sses
Seru
mam
yloi
dA
-1pr
otei
nSA
A1_
HU
MA
NP0
DJI
8SA
A1
13.5
Infla
mm
ator
yre
spon
seSo
dium
-dep
ende
ntno
radr
enal
ine
tran
spor
ter
SC6A
2_H
UM
AN
P239
75SL
C6A
269
.3Tr
ansp
orte
rac
tivity
Supp
ress
orof
tum
orig
enic
ity7
prot
ein
ST7_
HU
MA
NQ
9NRC
1ST
767
.2A
cts
asa
tum
orsu
ppre
ssor
Vaso
rin
VASN
_HU
MA
NQ
6EM
K4VA
SN71
.7M
ayac
tas
anin
hibi
tor
ofTG
F-be
tasi
gnal
ing
The
acce
ssio
nnu
mbe
r,ge
nena
me,
spec
ies
(Hum
an),
mol
ecul
arw
eigh
t(kD
a)an
dpr
otei
nfu
nctio
nw
ere
repo
rted
.
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
8

Table3
Sele
ctio
nof
iden
tified
sing
le-d
etec
ted
coro
napr
otei
nsbo
und
toth
e9.
73±
1.70
nmA
gNPs
afte
r30
min
incu
batio
nan
dsu
bseq
uent
was
hing
.
Prot
ein
nam
eU
niPr
otna
me
Entr
yna
me
Gen
eM
ass
(kD
a)Fu
nctio
n
Am
yloi
d-be
taA
4pr
ecur
sor
prot
ein-
bind
ing
fam
ilyA
mem
ber
2A
PBA
2_H
UM
AN
Q99
767
APB
A2
82.5
Func
tion
insy
napt
icve
sicl
eex
ocyt
osis
bybi
ndin
gto
STXB
P1,a
nes
sent
ialc
ompo
nent
ofth
esy
napt
icve
sicl
eex
ocyt
otic
mac
hine
ryA
nkyr
inre
peat
dom
ain-
cont
aini
ngpr
otei
n20
BA
N20
B_H
UM
AN
Q5C
Z79
AN
KRD
20A
8P93
.9–
Ank
yrin
repe
atan
dst
erile
alph
am
otif
dom
ain-
cont
aini
ngpr
otei
n1B
AN
S1B_
HU
MA
NQ
7Z6G
8A
NKS
1B13
8.1
Play
sa
role
asa
mod
ulat
orof
APP
proc
essi
ngBo
nem
orph
ogen
etic
prot
ein
10BM
P10_
HU
MA
NO
9539
3BM
P10
48.0
Inhi
bits
endo
thel
ialc
ellm
igra
tion
and
grow
th.M
ayre
duce
cell
mig
ratio
nan
dce
llm
atri
xad
hesi
onin
brea
stca
ncer
cell
lines
.Br
omod
omai
nad
jace
ntto
zinc
finge
rdo
mai
npr
otei
n2A
BAZ2
A_H
UM
AN
Q9U
IF9
BAZ2
A21
1.2
Esse
ntia
lcom
pone
ntof
the
NoR
C(n
ucle
olar
rem
odel
ing
com
plex
)co
mpl
exth
atm
edia
tes
sile
ncin
gof
afr
actio
nof
rDN
ACa
spas
ere
crui
tmen
tdo
mai
n-co
ntai
ning
prot
ein
11CA
R11_
HU
MA
NQ
9BXL
7CA
RD11
133.
3In
volv
edin
the
cost
imul
ator
ysi
gnal
esse
ntia
lfor
T-ce
llre
cept
or(T
CR)-
med
iate
dT-
cell
activ
atio
n.A
lso
activ
ates
the
TORC
1si
gnal
ing
path
way
Com
plem
ent
fact
orH
-rel
ated
prot
ein
3FH
R3_H
UM
AN
Q02
985
CFH
R337
.3In
volv
edin
com
plem
ent
regu
latio
nCo
mpl
emen
tfa
ctor
H-r
elat
edpr
otei
n4
FHR4
_HU
MA
NQ
9249
6CF
HR4
65.3
Play
sa
role
inlip
idm
etab
olis
mCy
clin
-dep
ende
ntki
nase
2CD
K2_H
UM
AN
P249
41CD
K233
.9Ca
taly
ticac
tivity
Dua
lspe
cific
itypr
otei
nph
osph
atas
e9
DU
S9_H
UM
AN
Q99
956
DU
SP9
41.9
Inac
tivat
esM
AP
kina
ses
Dys
toni
nD
YST_
HU
MA
NQ
0300
1D
ST86
0.6
Act
sas
anin
tegr
ator
ofin
term
edia
tefil
amen
ts,a
ctin
and
mic
rotu
bule
cyto
skel
eton
netw
orks
EGF-
cont
aini
ngfib
ulin
-like
extr
acel
lula
rm
atri
xpr
otei
n1
FBLN
3_H
UM
AN
Q12
805
EFEM
P154
.6Bi
nds
EGFR
,the
EGF
rece
ptor
,ind
ucin
gEG
FRau
toph
osph
oryl
atio
nan
dth
eac
tivat
ion
ofdo
wns
trea
msi
gnal
ing
path
way
s.M
aypl
aya
role
ince
llad
hesi
onan
dm
igra
tion
GRE
B1-li
kepr
otei
nG
RB1L
_HU
MA
NQ
9C09
1G
REB1
L21
4.3
Play
sa
maj
orro
lein
earl
ym
etan
ephr
osan
dge
nita
ldev
elop
men
tH
emog
lobi
nsu
buni
tga
mm
a-2
HBG
2_H
UM
AN
P698
92H
BG2
16.1
Gam
ma
chai
nsm
ake
upth
efe
talh
emog
lobi
nF,
inco
mbi
natio
nw
ithal
pha
chai
nsH
isto
ne-ly
sine
N-m
ethy
ltran
sfer
ase
2AKM
T2A
_HU
MA
NQ
0316
4KM
T2A
431.
8Ca
taly
ticac
tivity
Imm
unog
lobu
linhe
avy
vari
able
3-23
HV3
23_H
UM
AN
P017
64IG
HV3
-23
12.6
Imm
une
resp
onse
Impo
rtin
-4IP
O4_
HU
MA
NQ
8TEX
9IP
O4
118.
7Fu
nctio
nsin
nucl
ear
prot
ein
impo
rtas
nucl
ear
tran
spor
trec
epto
rKe
ratin
,typ
eIc
ytos
kele
tal9
K1C9
_HU
MA
NP3
5527
KRT9
62.1
Stru
ctur
alLI
NE-
1ty
petr
ansp
osas
edo
mai
n-co
ntai
ning
prot
ein
1LI
TD1_
HU
MA
NQ
5T7N
2L1
TD1
98.8
Sing
le-s
tran
ded
RNA
bind
ing
Lyso
som
al-tr
affick
ing
regu
lato
rLY
ST_H
UM
AN
Q99
698
LYST
429,
1Re
quir
edfo
rso
rtin
gen
doso
mal
resi
dent
prot
eins
into
late
mul
tives
icul
aren
doso
mes
Mat
rix
met
allo
prot
eina
se-9
MM
P9_H
UM
AN
P147
80M
MP9
78.5
Cata
lytic
activ
ityM
icro
tubu
le-a
ssoc
iate
dpr
otei
n1A
MA
P1A
_HU
MA
NP7
8559
MA
P1A
305.
5St
ruct
ural
prot
ein
invo
lved
inth
efil
amen
tous
cros
s-br
idgi
ngbe
twee
nm
icro
tubu
les
and
othe
rsk
elet
alel
emen
tsM
ORC
fam
ilyCW
-type
zinc
finge
rpr
otei
n1
MO
RC1_
HU
MA
NQ
86VD
1M
ORC
111
2.9
Zinc
ion
bind
ing.
Nuc
lear
dist
ribu
tion
prot
ein
nudE
-like
1N
DEL
1_H
UM
AN
Q9G
ZM8
ND
EL1
38.4
Requ
ired
for
orga
niza
tion
ofth
ece
llula
rm
icro
tubu
lear
ray
and
mic
rotu
bule
anch
orin
gat
the
cent
roso
me
Poly
mer
icim
mun
oglo
bulin
rece
ptor
PIG
R_H
UM
AN
P018
33PI
GR
83.3
This
rece
ptor
bind
spo
lym
eric
IgA
and
IgM
atth
eba
sola
tera
lsur
face
ofep
ithel
ialc
ells
Poly
pept
ide
N-a
cety
lgal
acto
sam
inyl
tran
sfer
ase
13G
LT13
_HU
MA
NQ
8IU
C8G
ALN
T13
64.0
Cata
lytic
activ
ityPr
obab
legu
anin
enu
cleo
tide
exch
ange
fact
orM
CF2L
2M
F2L2
_HU
MA
NQ
86YR
7M
CF2L
212
6.9
Func
tions
asa
guan
ine
nucl
eotid
eex
chan
gefa
ctor
Pro-
epid
erm
algr
owth
fact
orEG
F_H
UM
AN
P011
33EG
F13
3.9
Stim
ulat
esth
egr
owth
ofva
riou
sep
ider
mal
and
epith
elia
ltis
sues
invi
voan
din
vitr
oPr
oper
din
PRO
P_H
UM
AN
P279
18CF
P51
.3A
posi
tive
regu
lato
rof
the
alte
rnat
epa
thw
ayof
com
plem
ent
Prot
ein
MM
S22-
like
MM
S22_
HU
MA
NQ
6ZRQ
5M
MS2
2L14
2.3
Mai
ntai
nge
nom
ein
tegr
itydu
ring
DN
Are
plic
atio
nPr
otei
nph
osph
atas
ePT
C7ho
mol
ogPP
TC7_
HU
MA
NQ
8NI3
7PP
TC7
32.6
Cata
lytic
activ
ityPr
otei
nSh
room
3SH
RM3_
HU
MA
NQ
8TF7
2SH
ROO
M3
216.
8Co
ntro
lsce
llsh
ape
chan
ges
inth
ene
uroe
pith
eliu
mdu
ring
neur
altu
becl
osur
ePr
otei
nZG
RF1
ZGRF
1_H
UM
AN
Q86
YA3
ZGRF
123
6.6
Zinc
ion
bind
ing
that
inhi
bits
fact
ors
Xaan
dXI
aof
the
coag
ulat
ion
casc
ade
Puta
tive
solu
teca
rrie
rorg
anic
anio
ntr
ansp
orte
rfam
ilym
embe
r1B7
SO1B
7_H
UM
AN
G3V
0H7
SLCO
1B7
71.2
May
enco
dea
non-
func
tiona
ltru
ncat
edpr
otei
nRa
s-in
tera
ctin
gpr
otei
n1
RAIN
_HU
MA
NQ
5U65
1RA
SIP1
103.
4A
cts
asa
criti
cala
ndva
scul
ar-s
peci
ficre
gula
tor
ofG
TPas
esi
gnal
ing,
cell
arch
itect
ure,
and
adhe
sion
Ribo
som
alpr
otei
nS6
kina
seal
pha-
1KS
6A1_
HU
MA
NQ
1541
8RP
S6KA
182
.7Ca
taly
ticac
tivity
Shor
tco
iled-
coil
prot
ein
SCO
C_H
UM
AN
Q9U
IL1
SCO
C18
.0Po
sitiv
ere
gula
tor
ofam
ino
acid
star
vatio
n-in
duce
dau
toph
agy
Stru
ctur
alm
aint
enan
ceof
chro
mos
omes
prot
ein
6SM
C6_H
UM
AN
Q96
SB8
SMC6
126.
3St
ruct
ural
Syna
ptot
agm
in-5
SYT5
_HU
MA
NO
0044
5SY
T542
.9M
aybe
invo
lved
inCa
2+-d
epen
dent
exoc
ytos
isof
secr
etor
yve
sicl
esTu
dor
dom
ain-
cont
aini
ngpr
otei
n1
TDRD
1_H
UM
AN
Q9B
XT4
TDRD
113
2.0
Act
svi
ath
epi
RNA
met
abol
icpr
oces
s,w
hich
med
iate
sth
ere
pres
sion
oftr
ansp
osab
leel
emen
tsdu
ring
mei
osis
Tyro
sine
–tRN
Alig
ase,
cyto
plas
mic
SYYC
_HU
MA
NP5
4577
YARS
59.1
Cata
lytic
activ
ityVi
gilin
VIG
LN_H
UM
AN
Q00
341
HD
LBP
141.
4Pr
otec
tcel
lsfr
omov
er-a
ccum
ulat
ion
ofch
oles
tero
lvo
nW
illeb
rand
fact
orVW
F_H
UM
AN
P042
75VW
F22
0Pl
ays
am
ajor
role
inbl
ood
coag
ulat
ion
Wee
1-lik
epr
otei
nki
nase
WEE
1_H
UM
AN
P302
91W
EE1
71.6
Act
sas
ane
gativ
ere
gula
tor
ofen
try
into
mito
sis
Zinc
finge
rpr
otei
n11
4ZN
114_
HU
MA
NQ
8NC2
6ZN
F114
47.7
May
bein
volv
edin
tran
scri
ptio
nalr
egul
atio
n.
The
acce
ssio
nnu
mbe
r,ge
nena
me,
spec
ies
(Hum
an),
mol
ecul
arw
eigh
t(kD
a)an
dpr
otei
nfu
nctio
nw
ere
repo
rted
.
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
9

Table4
Sele
ctio
nof
iden
tified
sing
le-d
etec
ted
coro
napr
otei
nsbo
und
toth
e9.
30±
0.67
nmM
NPs
afte
r30
min
incu
batio
nan
dsu
bseq
uent
was
hing
.
Prot
ein
nam
eU
niPr
otna
me
Entr
yna
me
Gen
eM
ass
(kD
a)Fu
nctio
n
ATP
-bin
ding
cass
ette
sub-
fam
ilyB
mem
ber
5A
BCB5
_HU
MA
NQ
2M3G
0A
BCB5
138.
6Tr
ansp
orte
rac
tivity
Bile
salt-
activ
ated
lipas
eCE
L_H
UM
AN
P198
35CE
L79
.3Ca
taly
ticac
tivity
Cath
elic
idin
antim
icro
bial
pept
ide
CAM
P_H
UM
AN
P499
13CA
MP
19.3
Ant
ibac
teri
alac
tivity
Cera
mid
esy
ntha
se4
CERS
4_H
UM
AN
Q9H
A82
CERS
446
.4M
aybe
eith
era
bona
fide
(dih
ydro
)cer
amid
esy
ntha
seor
am
odul
ator
ofits
activ
ityC-
reac
tive
prot
ein
CRP_
HU
MA
NP0
2741
CRP
25.0
Dis
play
sse
vera
lfun
ctio
nsas
soci
ated
with
host
defe
nse
C-ty
pele
ctin
dom
ain
fam
ily4
mem
ber
FCL
C4F_
HU
MA
NQ
8N1N
0CL
EC4F
65.5
Rece
ptor
with
anaffi
nity
for
gala
ctos
ean
dfu
cose
.Cou
ldbe
invo
lved
inen
docy
tosi
sCy
stat
in-F
CYTF
_HU
MA
NO
7609
6CS
T716
.4M
aypl
aya
role
inim
mun
ere
gula
tion
thro
ugh
inhi
bitio
nof
aun
ique
targ
etin
the
hem
atop
oiet
icsy
stem
DD
B1-a
ndCU
L4-a
ssoc
iate
dfa
ctor
15D
CA15
_HU
MA
NQ
66K6
4D
CAF1
566
.5M
aybe
invo
lved
inub
iqui
tinat
ion
and
degr
adat
ion
thro
ugh
aD
BB1-
CUL4
E3pr
otei
n-ub
iqui
tinlig
ase
DN
Ato
pois
omer
ase
2-bi
ndin
gpr
otei
n1
TOPB
1_H
UM
AN
Q92
547
TOPB
P117
0.7
Bind
sdo
uble
-str
ande
dD
NA
brea
ksan
dni
cks
asw
ella
ssi
ngle
-str
ande
dD
NA
Dyn
ein
heav
ych
ain
10,a
xone
mal
DYH
10_H
UM
AN
Q8I
VF4
DN
AH
1051
4.8
Pres
ents
ATP
ase
activ
ityEl
lis-v
anCr
evel
dsy
ndro
me
prot
ein
EVC_
HU
MA
NP5
7679
EVC
111.
9In
volv
edin
endo
chon
dral
grow
than
dsk
elet
alde
velo
pmen
tFa
ncon
iane
mia
grou
pB
prot
ein
FAN
CB_H
UM
AN
Q8N
B91
FAN
CB97
.7D
NA
repa
irpr
otei
nre
quir
edfo
rFA
NCD
2ub
iqui
tinat
ion
F-bo
xon
lypr
otei
n42
FBX4
2_H
UM
AN
Q6P
3S6
FBXO
4277
.8Sp
ecifi
cally
reco
gniz
esp5
3/TP
53,p
rom
otin
gits
ubiq
uitin
atio
nan
dde
grad
atio
nG
alec
tin-3
-bin
ding
prot
ein
LG3B
P_H
UM
AN
Q08
380
LGA
LS3B
P65
.3St
imul
ate
host
defe
nse
agai
nstv
irus
esan
dtu
mor
cells
HEA
Tre
peat
-con
tain
ing
prot
ein
4H
EAT4
_HU
MA
NQ
86W
Z0H
EATR
411
7.2
–Im
mun
oglo
bulin
heav
yva
riab
le6-
1H
V601
_HU
MA
NA
0A0B
4J1U
7IG
HV6
-113
.5Im
uune
resp
onse
Imm
unog
lobu
linka
ppa
vari
able
2-24
KV22
4_H
UM
AN
A0A
0C4D
H68
IGKV
2-24
13.1
Imm
une
resp
onse
Imm
unog
lobu
linla
mbd
ava
riab
le3-
27LV
327_
HU
MA
NP0
1718
IGLV
3-27
12.2
Imm
une
resp
onse
Kalli
stat
inKA
IN_H
UM
AN
P296
22SE
RPIN
A4
48.5
Enzy
me
regu
lato
rin
hibi
tor
Kera
tin,t
ype
Icyt
oske
leta
l14
K1C1
4_H
UM
AN
P025
33KR
T14
51.6
Stru
ctur
alKe
ratin
,typ
eII
cutic
ular
Hb1
KRT8
1_H
UM
AN
Q14
533
KRT8
154
.9St
ruct
ural
Lebe
rcili
nLC
A5_
HU
MA
NQ
86VQ
0LC
A5
80.5
Tran
spor
ter
activ
ityLI
NE-
1ty
petr
ansp
osas
edo
mai
n-co
ntai
ning
prot
ein
1LI
TD1_
HU
MA
NQ
5T7N
2L1
TD1
98.8
Sing
le-s
tran
ded
RNA
bind
ing
Lipo
poly
sacc
hari
de-b
indi
ngpr
otei
nLB
P_H
UM
AN
P184
28LB
P53
.4Im
mun
ere
spon
seLR
Pch
aper
one
MES
DM
ESD
_HU
MA
NQ
1469
6M
ESD
26.1
Ass
istin
gth
efo
ldin
gof
beta
-pro
pelle
r/EG
Fm
odul
esw
ithin
the
fam
ilyof
low
-den
sity
lipop
rote
inre
cept
ors
(LD
LRs)
MA
NSC
dom
ain-
cont
aini
ngpr
otei
n1
MA
NS1
_HU
MA
NQ
9H8J
5M
AN
SC1
46.8
–M
ecke
linM
KS3_
HU
MA
NQ
5HYA
8TM
EM67
111.
7Re
quir
edfo
rci
liary
stru
ctur
ean
dfu
nctio
nM
itoch
ondr
ial2
-oxo
glut
arat
e/m
alat
eca
rrie
rpr
otei
nM
2OM
_HU
MA
NQ
0297
8SL
C25A
1134
.1Ca
taly
tican
dtr
ansp
orte
rac
tiviti
esN
AD
(P)H
dehy
drog
enas
e[q
uino
ne]
1N
QO
1_H
UM
AN
P155
59N
QO
130
.9Ca
taly
ticac
tivity
N-a
cety
llact
osam
inid
ebe
ta-1
,3-N
-ace
tylg
luco
sam
inyl
tran
sfer
ase
2B3
GN
2_H
UM
AN
Q9N
Y97
B3G
NT2
46.0
Cata
lytic
activ
ity
Nck
-ass
ocia
ted
prot
ein
1N
CKP1
_HU
MA
NQ
9Y2A
7N
CKA
P112
8.8
As
com
pone
ntof
the
WA
VE1
com
plex
,req
uire
dfo
rBD
NF-
NTR
K2en
docy
tictr
affick
ing
and
sign
alin
gfr
omea
rly
endo
som
esN
espr
in-1
SYN
E1_H
UM
AN
Q8N
F91
SYN
E110
11.1
Form
sa
linki
ngne
twor
kbe
twee
nor
gane
lles
and
the
actin
cyto
skel
eton
tom
aint
ain
the
subc
ellu
lar
spat
ial
orga
niza
tion
Nuc
lear
rece
ptor
coac
tivat
or6
NCO
A6_
HU
MA
NQ
1468
6N
COA
621
9.1
Invo
lved
inth
eco
activ
atio
nof
the
NF-
kapp
a-B
path
way
PAN
2-PA
N3
dead
enyl
atio
nco
mpl
exsu
buni
tPA
N3
PAN
3_H
UM
AN
Q58
A45
PAN
395
.6Re
gula
tory
subu
nit
ofth
epo
ly(A
)-nu
clea
se(P
AN
)de
aden
ylat
ion
com
plex
Pent
atri
cope
ptid
ere
peat
-con
tain
ing
prot
ein
1,m
itoch
ondr
ial
PTCD
1_H
UM
AN
O75
127
PTCD
178
.8M
itoch
ondr
ialp
rote
inim
plic
ated
inne
gativ
ere
gula
tion
ofle
ucin
etR
NA
leve
ls,a
sw
ella
sne
gativ
ere
gula
tion
ofm
itoch
ondr
ia-e
ncod
edpr
otei
nsan
dCO
Xac
tivity
Phos
phat
idyl
chol
ine
tran
sloc
ator
ABC
B4M
DR3
_HU
MA
NP2
1439
ABC
B414
1.5
Act
sas
apo
sitiv
ere
gula
tor
ofbi
liary
lipid
secr
etio
n1-
phos
phat
idyl
inos
itol4
,5-b
isph
osph
ate
phos
phod
iest
eras
ega
mm
a-2
PLCG
2_H
UM
AN
P168
85PL
CG2
147.
8Ca
taly
ticac
tivity
Phos
phol
ipas
eD
1PL
D1_
HU
MA
NQ
1339
3PL
D1
124.
2Ca
taly
ticac
tivity
Poly
pept
ide
N-a
cety
lgal
acto
sam
inyl
tran
sfer
ase
3G
ALT
3_H
UM
AN
Q14
435
GA
LNT3
72.6
Cata
lytic
activ
ityPr
otei
nsa
lvad
orho
mol
og1
SAV1
_HU
MA
NQ
9H4B
6SA
V144
.6Re
gula
tor
ofST
K3/M
ST2
and
STK4
/MST
1in
the
Hip
posi
gnal
ing
path
way
whi
chpl
ays
api
vota
lrol
ein
orga
nsi
zeco
ntro
land
tum
orsu
ppre
ssio
nby
rest
rict
ing
prol
ifera
tion
and
prom
otin
gap
opto
sis
Prot
ein
S100
-A7
S10A
7_H
UM
AN
P311
51S1
00A
711
.5–
Prot
ein
S100
-A8
S10A
8_H
UM
AN
P051
09S1
00A
810
.8Pl
ays
apr
omin
ent
role
inth
ere
gula
tion
ofin
flam
mat
ory
proc
esse
san
dim
mun
ere
spon
sePr
otei
nS1
00-A
9S1
0A9_
HU
MA
NP0
6702
S100
A9
13.2
Play
sa
prom
inen
tro
lein
the
regu
latio
nof
infla
mm
ator
ypr
oces
ses
and
imm
une
resp
onse
Prot
ein
Z-de
pend
ent
prot
ease
inhi
bito
rZP
I_HU
MA
NQ
9UK5
5SE
RPIN
A10
50.7
Enzy
me
regu
lato
rac
tivity
Prot
ein
ZGRF
1ZG
RF1_
HU
MA
NQ
86YA
3ZG
RF1
236.
6Zi
ncio
nbi
ndin
gth
atin
hibi
tsfa
ctor
sXa
and
XIa
ofth
eco
agul
atio
nca
scad
ePr
otoc
adhe
rin-
12PC
D12
_HU
MA
NQ
9NPG
4PC
DH
1212
8.9
Cellu
lar
adhe
sion
mol
ecul
eth
atpl
ays
anim
port
ant
role
asa
regu
lato
rof
cell
mig
ratio
n,pr
obab
lyvi
ain
crea
sing
cell-
cell
adhe
sion
(continuedon
next
page
)
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
10
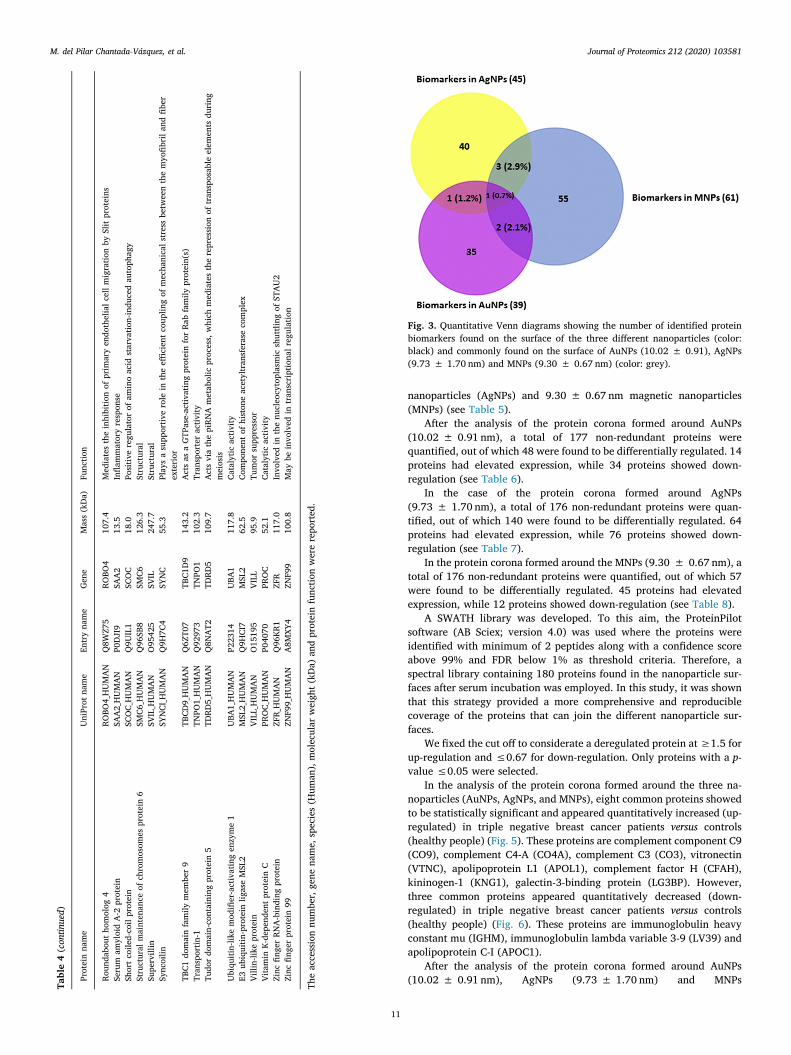
nanoparticles (AgNPs) and 9.30 ± 0.67 nm magnetic nanoparticles(MNPs) (see Table 5).
After the analysis of the protein corona formed around AuNPs(10.02 ± 0.91 nm), a total of 177 non-redundant proteins werequantified, out of which 48 were found to be differentially regulated. 14proteins had elevated expression, while 34 proteins showed down-regulation (see Table 6).
In the case of the protein corona formed around AgNPs(9.73 ± 1.70 nm), a total of 176 non-redundant proteins were quan-tified, out of which 140 were found to be differentially regulated. 64proteins had elevated expression, while 76 proteins showed down-regulation (see Table 7).
In the protein corona formed around the MNPs (9.30 ± 0.67 nm), atotal of 176 non-redundant proteins were quantified, out of which 57were found to be differentially regulated. 45 proteins had elevatedexpression, while 12 proteins showed down-regulation (see Table 8).
A SWATH library was developed. To this aim, the ProteinPilotsoftware (AB Sciex; version 4.0) was used where the proteins wereidentified with minimum of 2 peptides along with a confidence scoreabove 99% and FDR below 1% as threshold criteria. Therefore, aspectral library containing 180 proteins found in the nanoparticle sur-faces after serum incubation was employed. In this study, it was shownthat this strategy provided a more comprehensive and reproduciblecoverage of the proteins that can join the different nanoparticle sur-faces.
We fixed the cut off to considerate a deregulated protein at ≥1.5 forup-regulation and ≤0.67 for down-regulation. Only proteins with a p-value ≤0.05 were selected.
In the analysis of the protein corona formed around the three na-noparticles (AuNPs, AgNPs, and MNPs), eight common proteins showedto be statistically significant and appeared quantitatively increased (up-regulated) in triple negative breast cancer patients versus controls(healthy people) (Fig. 5). These proteins are complement component C9(CO9), complement C4-A (CO4A), complement C3 (CO3), vitronectin(VTNC), apolipoprotein L1 (APOL1), complement factor H (CFAH),kininogen-1 (KNG1), galectin-3-binding protein (LG3BP). However,three common proteins appeared quantitatively decreased (down-regulated) in triple negative breast cancer patients versus controls(healthy people) (Fig. 6). These proteins are immunoglobulin heavyconstant mu (IGHM), immunoglobulin lambda variable 3-9 (LV39) andapolipoprotein C-I (APOC1).
After the analysis of the protein corona formed around AuNPs(10.02 ± 0.91 nm), AgNPs (9.73 ± 1.70 nm) and MNPsTa
ble4
(continued)
Prot
ein
nam
eU
niPr
otna
me
Entr
yna
me
Gen
eM
ass
(kD
a)Fu
nctio
n
Roun
dabo
utho
mol
og4
ROBO
4_H
UM
AN
Q8W
Z75
ROBO
410
7.4
Med
iate
sth
ein
hibi
tion
ofpr
imar
yen
doth
elia
lcel
lmig
ratio
nby
Slit
prot
eins
Seru
mam
yloi
dA
-2pr
otei
nSA
A2_
HU
MA
NP0
DJI
9SA
A2
13.5
Infla
mm
ator
yre
spon
seSh
ort
coile
d-co
ilpr
otei
nSC
OC_
HU
MA
NQ
9UIL
1SC
OC
18.0
Posi
tive
regu
lato
rof
amin
oac
idst
arva
tion-
indu
ced
auto
phag
ySt
ruct
ural
mai
nten
ance
ofch
rom
osom
espr
otei
n6
SMC6
_HU
MA
NQ
96SB
8SM
C612
6.3
Stru
ctur
alSu
perv
illin
SVIL
_HU
MA
NO
9542
5SV
IL24
7.7
Stru
ctur
alSy
ncoi
linSY
NCI
_HU
MA
NQ
9H7C
4SY
NC
55.3
Play
sa
supp
ortiv
ero
lein
the
effici
ent
coup
ling
ofm
echa
nica
lstr
ess
betw
een
the
myo
fibri
land
fiber
exte
rior
TBC1
dom
ain
fam
ilym
embe
r9
TBCD
9_H
UM
AN
Q6Z
T07
TBC1
D9
143.
2A
cts
asa
GTP
ase-
activ
atin
gpr
otei
nfo
rRa
bfa
mily
prot
ein(
s)Tr
ansp
ortin
-1TN
PO1_
HU
MA
NQ
9297
3TN
PO1
102.
3Tr
ansp
orte
rac
tivity
Tudo
rdo
mai
n-co
ntai
ning
prot
ein
5TD
RD5_
HU
MA
NQ
8NA
T2TD
RD5
109.
7A
cts
via
the
piRN
Am
etab
olic
proc
ess,
whi
chm
edia
tes
the
repr
essi
onof
tran
spos
able
elem
ents
duri
ngm
eios
isU
biqu
itin-
like
mod
ifier
-act
ivat
ing
enzy
me
1U
BA1_
HU
MA
NP2
2314
UBA
111
7.8
Cata
lytic
activ
ityE3
ubiq
uitin
-pro
tein
ligas
eM
SL2
MSL
2_H
UM
AN
Q9H
CI7
MSL
262
.5Co
mpo
nent
ofhi
ston
eac
etyl
tran
sfer
ase
com
plex
Villi
n-lik
epr
otei
nVI
LL_H
UM
AN
O15
195
VILL
95.9
Tum
orsu
ppre
ssor
Vita
min
K-de
pend
ent
prot
ein
CPR
OC_
HU
MA
NP0
4070
PRO
C52
.1Ca
taly
ticac
tivity
Zinc
finge
rRN
A-b
indi
ngpr
otei
nZF
R_H
UM
AN
Q96
KR1
ZFR
117.
0In
volv
edin
the
nucl
eocy
topl
asm
icsh
uttli
ngof
STA
U2
Zinc
finge
rpr
otei
n99
ZNF9
9_H
UM
AN
A8M
XY4
ZNF9
910
0.8
May
bein
volv
edin
tran
scri
ptio
nalr
egul
atio
n
The
acce
ssio
nnu
mbe
r,ge
nena
me,
spec
ies
(Hum
an),
mol
ecul
arw
eigh
t(kD
a)an
dpr
otei
nfu
nctio
nw
ere
repo
rted
.Fig. 3. Quantitative Venn diagrams showing the number of identified proteinbiomarkers found on the surface of the three different nanoparticles (color:black) and commonly found on the surface of AuNPs (10.02 ± 0.91), AgNPs(9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm) (color: grey).
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
11
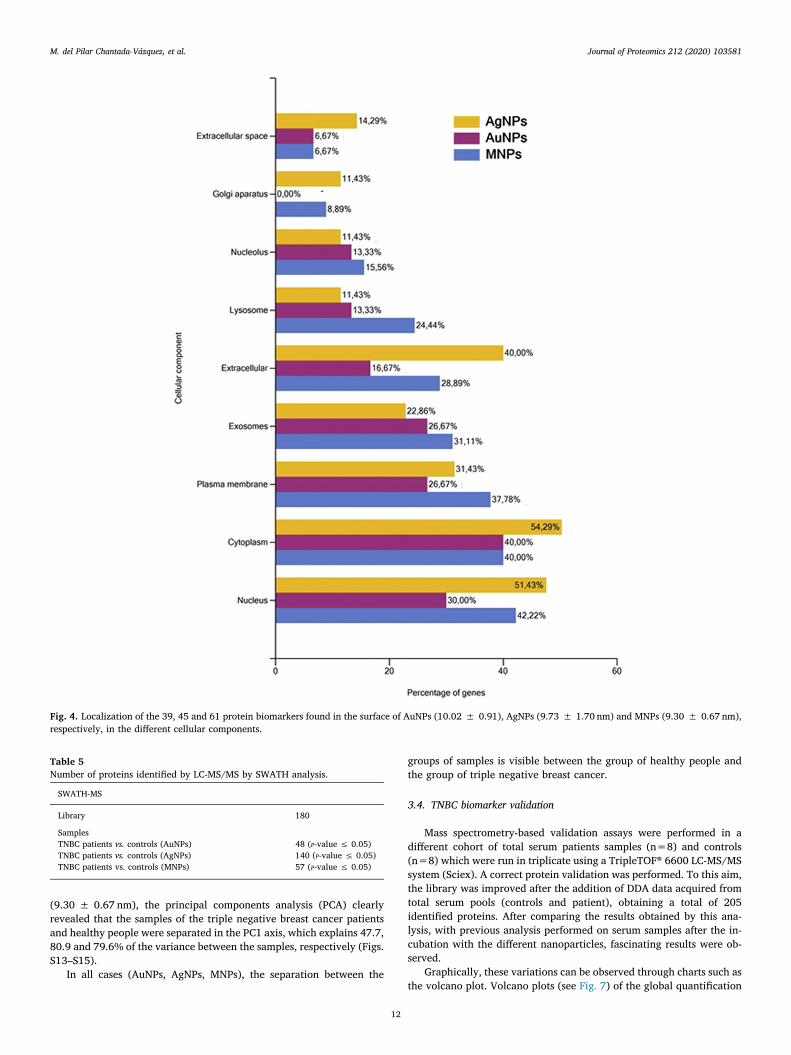
(9.30 ± 0.67 nm), the principal components analysis (PCA) clearlyrevealed that the samples of the triple negative breast cancer patientsand healthy people were separated in the PC1 axis, which explains 47.7,80.9 and 79.6% of the variance between the samples, respectively (Figs.S13–S15).
In all cases (AuNPs, AgNPs, MNPs), the separation between the
groups of samples is visible between the group of healthy people andthe group of triple negative breast cancer.
3.4. TNBC biomarker validation
Mass spectrometry-based validation assays were performed in adifferent cohort of total serum patients samples (n=8) and controls(n=8) which were run in triplicate using a TripleTOF® 6600 LC-MS/MSsystem (Sciex). A correct protein validation was performed. To this aim,the library was improved after the addition of DDA data acquired fromtotal serum pools (controls and patient), obtaining a total of 205identified proteins. After comparing the results obtained by this ana-lysis, with previous analysis performed on serum samples after the in-cubation with the different nanoparticles, fascinating results were ob-served.
Graphically, these variations can be observed through charts such asthe volcano plot. Volcano plots (see Fig. 7) of the global quantification
Fig. 4. Localization of the 39, 45 and 61 protein biomarkers found in the surface of AuNPs (10.02 ± 0.91), AgNPs (9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm),respectively, in the different cellular components.
Table 5Number of proteins identified by LC-MS/MS by SWATH analysis.
SWATH-MS
Library 180
SamplesTNBC patients vs. controls (AuNPs) 48 (P-value ≤ 0.05)TNBC patients vs. controls (AgNPs) 140 (P-value ≤ 0.05)TNBC patients vs. controls (MNPs) 57 (P-value ≤ 0.05)
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
12

of proteins between healthy and triple negative breast cancer patientswith A: AuNPs (10.02 ± 0.91 nm), B: AgNPs (9.73 ± 1.70 nm) and C:MNPs (9.30 ± 0.67 nm) were generated by plotting the log 2-foldchanges for the identified proteins against their corresponding adjustedp-value.
Validation assays verified and confirmed that transthyretin (TTHY)was statistically significant and appeared quantitatively decreased(down-regulated) in triple negative breast cancer patients when com-pared with controls (healthy people). This result was observed after theanalysis of the protein corona formed around AuNPs(10.02 ± 0.91 nm). A similar observation was recently reported,where it was found that transthyretin (TTHY) was predominantly(68.75%) down-regulated (n = 33/48) in the sera of breast cancerpatients [57].
Furthermore, validation assays verified and confirmed that com-plement component C8 gamma chain (CO8G), ficolin-3 (FCN3), retinol-binding protein 4 (RET4), fibronectin (FINC), fetuin-B (FETUB) andapolipoprotein A-IV (APOA4), were up-regulated in triple negativebreast cancer when compared with controls. However, apolipoprotein
C-III (APOC3), immunoglobulin kappa variable 2D-28 (KVD28), im-munoglobulin kappa variable 1-5 (KV105), immunoglobulin kappavariable 4-1 (KV401), immunoglobulin kappa variable 1D-12 (KVD12),immunoglobulin heavy variable 1-46 (HV146), immunoglobulin heavyvariable 3-30-5 (HV335), immunoglobulin heavy constant gamma 2(IGHG2), immunoglobulin heavy constant gamma 3 (IGHG3), and im-munoglobulin heavy constant gamma 4 (IGHG4) were down-regulatedin triple negative breast cancer when compared with controls. Theseresults were observed after the analysis of the protein corona formedaround AgNPs (9.73 ± 1.70 nm).
Recently, similar studies found that elevated serum levels of retinolbinding protein 4 (RBP4) [58] and fibronectin (FINC) [59] were asso-ciated with breast cancer risk and they could be useful markers forpredicting poor prognosis in breast cancer patients.
In the particular case of apolipoprotein A-IV (APOA4), comparativeproteomic profiling of immunodepleted plasma of healthy and of BCindividuals revealed that this protein was also up-regulated in theplasma of the BC individuals. Furthermore, this protein was found to beinvolved in the pathogenesis of cancer and played an important role in
Table 6Significant proteins (p-value < 0.05) in comparisons between triple negative breast cancer and controls after the analysis of the protein corona of AuNPs(10.02 ± 0.91 nm).
Protein UniProt ID p-value Fold change
TNBC patients vs. controls Complement component C6 CO6 0.021032066 3.249007224 ↑ TNBCVitamin D-binding protein VTDB 0.000044522 1.663281358 ↑ TNBCComplement component C9 CO9 0.000172961 1.658309079 ↑ TNBCComplement C4-A CO4A 0.003482887 1.525866959 ↑ TNBCComplement C3 CO3 0.000115208 1.451520626 ↑ TNBCPlasminogen PLMN 0.005392898 1.329388215 ↑ TNBCVitronectin VTNC 0.004766057 1.282037959 ↑ TNBCApolipoprotein L1 APOL1 0.022535013 1.251913563 ↑ TNBCAfamin AFAM 0.020811026 1.248070740 ↑ TNBCComplement factor H CFAH 0.020861177 1.204471284 ↑ TNBCComplement C5 CO5 0.016073885 1.185058578 ↑ TNBCSerum albumin ALBU 0.004803944 1.172992995 ↑ TNBCKininogen-1 KNG1 0.024754757 1.167133709 ↑ TNBCGalectin-3-binding protein LG3BP 0.030403083 1.088223023 ↑ TNBCGlutathione peroxidase 3 GPX3 0.01510168 7.16326536 ↑ CONTROLImmunoglobulin heavy variable 5-51 HV551 0.02782821 4.90904368 ↑ CONTROLImmunoglobulin heavy constant mu IGHM 0.04111454 4.42849357 ↑ CONTROLApolipoprotein C-I APOC1 0.00460538 4.09989381 ↑ CONTROLImmunoglobulin lambda variable 3-9 LV39 0.0394842 3.35634858 ↑ CONTROLApolipoprotein D APOD 0.04050732 3.22034685 ↑ CONTROLImmunoglobulin heavy constant alpha 1 IGHA1 0.00960489 3.19128417 ↑ CONTROLImmunoglobulin kappa variable 2-29 KV229 0.00334596 2.48673552 ↑ CONTROLImmunoglobulin heavy constant alpha 2 IGHA2 0.02785381 2.43044159 ↑ CONTROLInter-alpha-trypsin inhibitor heavy chain H4 ITIH4 0.02521756 2.41781135 ↑ CONTROLImmunoglobulin lambda variable 4-69 LV469 0.00833282 2.37374475 ↑ CONTROLApolipoprotein(a) OS=Homo sapiens APOA 0.03681681 2.12362227 ↑ CONTROLPigment epithelium-derived factor PEDF 0.00858685 1.94114234 ↑ CONTROLAlpha-2-macroglobulin A2MG 0.00477503 1.9190963 ↑ CONTROLHemoglobin subunit beta HBB 0.00500101 1.90068671 ↑ CONTROLCD5 antigen-like CD5L 0.0397185 1.7759815 ↑ CONTROLSerum amyloid A-4 protein SAA4 0.04115058 1.77144695 ↑ CONTROLTetranectin TETN 0.01611908 1.7097494 ↑ CONTROLApolipoprotein A-II APOA2 0.01467561 1.67488772 ↑ CONTROLInter-alpha-trypsin inhibitor heavy chain H2 ITIH2 0.00096553 1.61898708 ↑ CONTROLCarboxypeptidase B2 CBPB2 0.00728968 1.46619249 ↑ CONTROLTransthyretin TTHY 0.04987991 1.45277849 ↑ CONTROLComplement C1r subcomponent-like protein C1RL 0.03887632 1.43784936 ↑ CONTROLInter-alpha-trypsin inhibitor heavy chain H1 ITIH1 0.00031251 1.41502216 ↑ CONTROLSerum paraoxonase/arylesterase 1 PON1 0.00142352 1.40629048 ↑ CONTROLApolipoprotein F APOF 0.03956903 1.39106396 ↑ CONTROLApolipoprotein M APOM 0.00064927 1.36179665 ↑ CONTROLApolipoprotein A-I APOA1 0.02341567 1.34721575 ↑ CONTROLSerotransferrin TRFE 0.0020036 1.31516423 ↑ CONTROLSelenoprotein P SEPP1 0.00426885 1.30905254 ↑ CONTROLMannan-binding lectin serine protease 1 MASP1 0.03643268 1.29206286 ↑ CONTROLCarboxypeptidase N subunit 2 CPN2 0.03700623 1.28514087 ↑ CONTROLProtein AMBP AMBP 0.0084045 1.22749917 ↑ CONTROLPlasma protease C1 inhibitor IC1 0.04106727 1.2095684 ↑ CONTROL
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
13
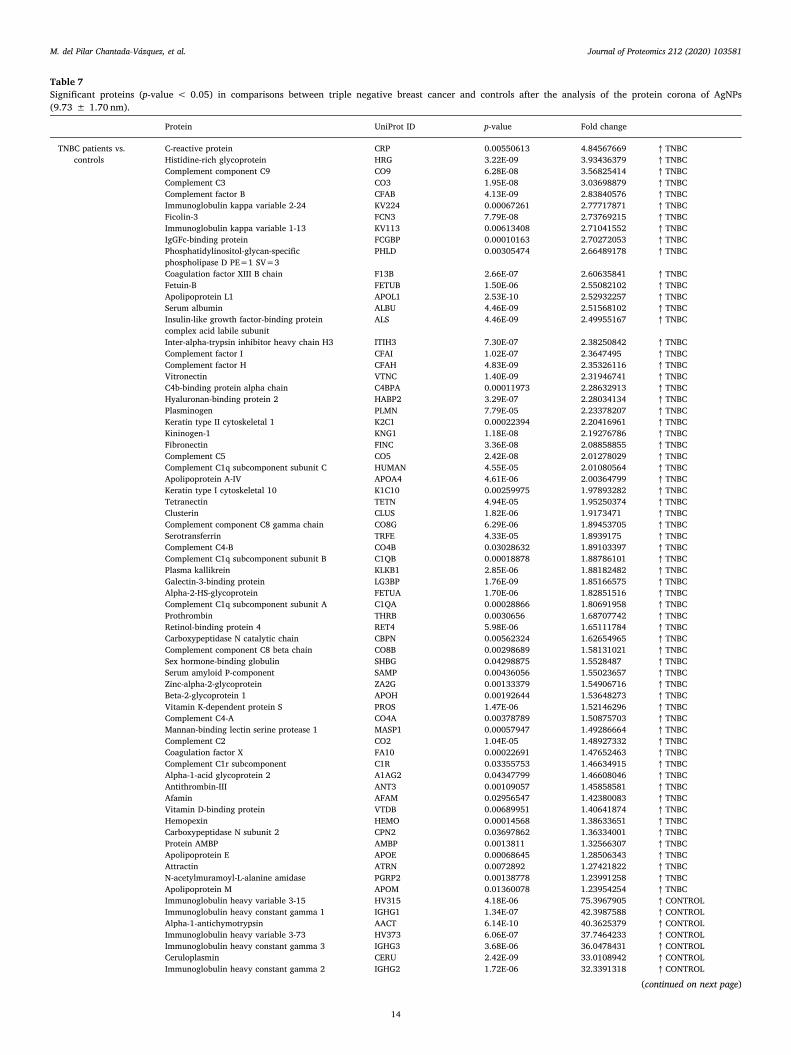
Table 7Significant proteins (p-value < 0.05) in comparisons between triple negative breast cancer and controls after the analysis of the protein corona of AgNPs(9.73 ± 1.70 nm).
Protein UniProt ID p-value Fold change
TNBC patients vs.controls
C-reactive protein CRP 0.00550613 4.84567669 ↑ TNBCHistidine-rich glycoprotein HRG 3.22E-09 3.93436379 ↑ TNBCComplement component C9 CO9 6.28E-08 3.56825414 ↑ TNBCComplement C3 CO3 1.95E-08 3.03698879 ↑ TNBCComplement factor B CFAB 4.13E-09 2.83840576 ↑ TNBCImmunoglobulin kappa variable 2-24 KV224 0.00067261 2.77717871 ↑ TNBCFicolin-3 FCN3 7.79E-08 2.73769215 ↑ TNBCImmunoglobulin kappa variable 1-13 KV113 0.00613408 2.71041552 ↑ TNBCIgGFc-binding protein FCGBP 0.00010163 2.70272053 ↑ TNBCPhosphatidylinositol-glycan-specificphospholipase D PE=1 SV=3
PHLD 0.00305474 2.66489178 ↑ TNBC
Coagulation factor XIII B chain F13B 2.66E-07 2.60635841 ↑ TNBCFetuin-B FETUB 1.50E-06 2.55082102 ↑ TNBCApolipoprotein L1 APOL1 2.53E-10 2.52932257 ↑ TNBCSerum albumin ALBU 4.46E-09 2.51568102 ↑ TNBCInsulin-like growth factor-binding proteincomplex acid labile subunit
ALS 4.46E-09 2.49955167 ↑ TNBC
Inter-alpha-trypsin inhibitor heavy chain H3 ITIH3 7.30E-07 2.38250842 ↑ TNBCComplement factor I CFAI 1.02E-07 2.3647495 ↑ TNBCComplement factor H CFAH 4.83E-09 2.35326116 ↑ TNBCVitronectin VTNC 1.40E-09 2.31946741 ↑ TNBCC4b-binding protein alpha chain C4BPA 0.00011973 2.28632913 ↑ TNBCHyaluronan-binding protein 2 HABP2 3.29E-07 2.28034134 ↑ TNBCPlasminogen PLMN 7.79E-05 2.23378207 ↑ TNBCKeratin type II cytoskeletal 1 K2C1 0.00022394 2.20416961 ↑ TNBCKininogen-1 KNG1 1.18E-08 2.19276786 ↑ TNBCFibronectin FINC 3.36E-08 2.08858855 ↑ TNBCComplement C5 CO5 2.42E-08 2.01278029 ↑ TNBCComplement C1q subcomponent subunit C HUMAN 4.55E-05 2.01080564 ↑ TNBCApolipoprotein A-IV APOA4 4.61E-06 2.00364799 ↑ TNBCKeratin type I cytoskeletal 10 K1C10 0.00259975 1.97893282 ↑ TNBCTetranectin TETN 4.94E-05 1.95250374 ↑ TNBCClusterin CLUS 1.82E-06 1.9173471 ↑ TNBCComplement component C8 gamma chain CO8G 6.29E-06 1.89453705 ↑ TNBCSerotransferrin TRFE 4.33E-05 1.8939175 ↑ TNBCComplement C4-B CO4B 0.03028632 1.89103397 ↑ TNBCComplement C1q subcomponent subunit B C1QB 0.00018878 1.88786101 ↑ TNBCPlasma kallikrein KLKB1 2.85E-06 1.88182482 ↑ TNBCGalectin-3-binding protein LG3BP 1.76E-09 1.85166575 ↑ TNBCAlpha-2-HS-glycoprotein FETUA 1.70E-06 1.82851516 ↑ TNBCComplement C1q subcomponent subunit A C1QA 0.00028866 1.80691958 ↑ TNBCProthrombin THRB 0.0030656 1.68707742 ↑ TNBCRetinol-binding protein 4 RET4 5.98E-06 1.65111784 ↑ TNBCCarboxypeptidase N catalytic chain CBPN 0.00562324 1.62654965 ↑ TNBCComplement component C8 beta chain CO8B 0.00298689 1.58131021 ↑ TNBCSex hormone-binding globulin SHBG 0.04298875 1.5528487 ↑ TNBCSerum amyloid P-component SAMP 0.00436056 1.55023657 ↑ TNBCZinc-alpha-2-glycoprotein ZA2G 0.00133379 1.54906716 ↑ TNBCBeta-2-glycoprotein 1 APOH 0.00192644 1.53648273 ↑ TNBCVitamin K-dependent protein S PROS 1.47E-06 1.52146296 ↑ TNBCComplement C4-A CO4A 0.00378789 1.50875703 ↑ TNBCMannan-binding lectin serine protease 1 MASP1 0.00057947 1.49286664 ↑ TNBCComplement C2 CO2 1.04E-05 1.48927332 ↑ TNBCCoagulation factor X FA10 0.00022691 1.47652463 ↑ TNBCComplement C1r subcomponent C1R 0.03355753 1.46634915 ↑ TNBCAlpha-1-acid glycoprotein 2 A1AG2 0.04347799 1.46608046 ↑ TNBCAntithrombin-III ANT3 0.00109057 1.45858581 ↑ TNBCAfamin AFAM 0.02956547 1.42380083 ↑ TNBCVitamin D-binding protein VTDB 0.00689951 1.40641874 ↑ TNBCHemopexin HEMO 0.00014568 1.38633651 ↑ TNBCCarboxypeptidase N subunit 2 CPN2 0.03697862 1.36334001 ↑ TNBCProtein AMBP AMBP 0.0013811 1.32566307 ↑ TNBCApolipoprotein E APOE 0.00068645 1.28506343 ↑ TNBCAttractin ATRN 0.0072892 1.27421822 ↑ TNBCN-acetylmuramoyl-L-alanine amidase PGRP2 0.00138778 1.23991258 ↑ TNBCApolipoprotein M APOM 0.01360078 1.23954254 ↑ TNBCImmunoglobulin heavy variable 3-15 HV315 4.18E-06 75.3967905 ↑ CONTROLImmunoglobulin heavy constant gamma 1 IGHG1 1.34E-07 42.3987588 ↑ CONTROLAlpha-1-antichymotrypsin AACT 6.14E-10 40.3625379 ↑ CONTROLImmunoglobulin heavy variable 3-73 HV373 6.06E-07 37.7464233 ↑ CONTROLImmunoglobulin heavy constant gamma 3 IGHG3 3.68E-06 36.0478431 ↑ CONTROLCeruloplasmin CERU 2.42E-09 33.0108942 ↑ CONTROLImmunoglobulin heavy constant gamma 2 IGHG2 1.72E-06 32.3391318 ↑ CONTROL
(continued on next page)
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
14
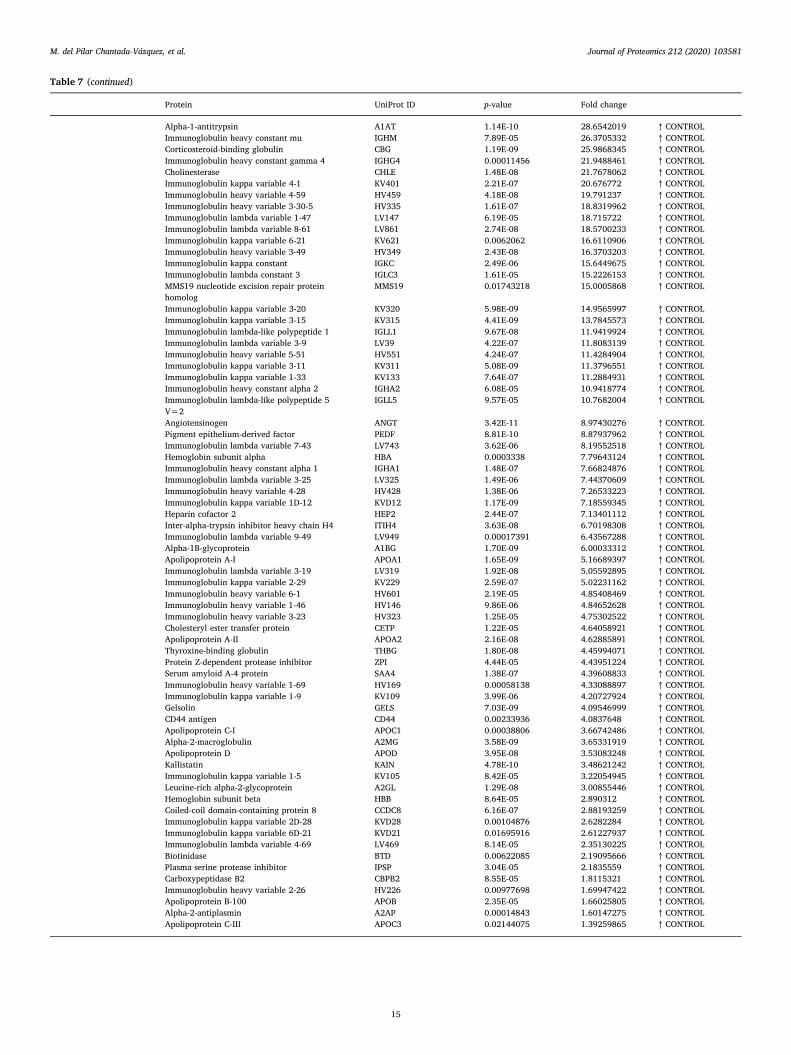
Table 7 (continued)
Protein UniProt ID p-value Fold change
Alpha-1-antitrypsin A1AT 1.14E-10 28.6542019 ↑ CONTROLImmunoglobulin heavy constant mu IGHM 7.89E-05 26.3705332 ↑ CONTROLCorticosteroid-binding globulin CBG 1.19E-09 25.9868345 ↑ CONTROLImmunoglobulin heavy constant gamma 4 IGHG4 0.00011456 21.9488461 ↑ CONTROLCholinesterase CHLE 1.48E-08 21.7678062 ↑ CONTROLImmunoglobulin kappa variable 4-1 KV401 2.21E-07 20.676772 ↑ CONTROLImmunoglobulin heavy variable 4-59 HV459 4.18E-08 19.791237 ↑ CONTROLImmunoglobulin heavy variable 3-30-5 HV335 1.61E-07 18.8319962 ↑ CONTROLImmunoglobulin lambda variable 1-47 LV147 6.19E-05 18.715722 ↑ CONTROLImmunoglobulin lambda variable 8-61 LV861 2.74E-08 18.5700233 ↑ CONTROLImmunoglobulin kappa variable 6-21 KV621 0.0062062 16.6110906 ↑ CONTROLImmunoglobulin heavy variable 3-49 HV349 2.43E-08 16.3703203 ↑ CONTROLImmunoglobulin kappa constant IGKC 2.49E-06 15.6449675 ↑ CONTROLImmunoglobulin lambda constant 3 IGLC3 1.61E-05 15.2226153 ↑ CONTROLMMS19 nucleotide excision repair proteinhomolog
MMS19 0.01743218 15.0005868 ↑ CONTROL
Immunoglobulin kappa variable 3-20 KV320 5.98E-09 14.9565997 ↑ CONTROLImmunoglobulin kappa variable 3-15 KV315 4.41E-09 13.7845573 ↑ CONTROLImmunoglobulin lambda-like polypeptide 1 IGLL1 9.67E-08 11.9419924 ↑ CONTROLImmunoglobulin lambda variable 3-9 LV39 4.22E-07 11.8083139 ↑ CONTROLImmunoglobulin heavy variable 5-51 HV551 4.24E-07 11.4284904 ↑ CONTROLImmunoglobulin kappa variable 3-11 KV311 5.08E-09 11.3796551 ↑ CONTROLImmunoglobulin kappa variable 1-33 KV133 7.64E-07 11.2884931 ↑ CONTROLImmunoglobulin heavy constant alpha 2 IGHA2 6.08E-05 10.9418774 ↑ CONTROLImmunoglobulin lambda-like polypeptide 5V=2
IGLL5 9.57E-05 10.7682004 ↑ CONTROL
Angiotensinogen ANGT 3.42E-11 8.97430276 ↑ CONTROLPigment epithelium-derived factor PEDF 8.81E-10 8.87937962 ↑ CONTROLImmunoglobulin lambda variable 7-43 LV743 3.62E-06 8.19552518 ↑ CONTROLHemoglobin subunit alpha HBA 0.0003338 7.79643124 ↑ CONTROLImmunoglobulin heavy constant alpha 1 IGHA1 1.48E-07 7.66824876 ↑ CONTROLImmunoglobulin lambda variable 3-25 LV325 1.49E-06 7.44370609 ↑ CONTROLImmunoglobulin heavy variable 4-28 HV428 1.38E-06 7.26533223 ↑ CONTROLImmunoglobulin kappa variable 1D-12 KVD12 1.17E-09 7.18559345 ↑ CONTROLHeparin cofactor 2 HEP2 2.44E-07 7.13401112 ↑ CONTROLInter-alpha-trypsin inhibitor heavy chain H4 ITIH4 3.63E-08 6.70198308 ↑ CONTROLImmunoglobulin lambda variable 9-49 LV949 0.00017391 6.43567288 ↑ CONTROLAlpha-1B-glycoprotein A1BG 1.70E-09 6.00033312 ↑ CONTROLApolipoprotein A-I APOA1 1.65E-09 5.16689397 ↑ CONTROLImmunoglobulin lambda variable 3-19 LV319 1.92E-08 5.05592895 ↑ CONTROLImmunoglobulin kappa variable 2-29 KV229 2.59E-07 5.02231162 ↑ CONTROLImmunoglobulin heavy variable 6-1 HV601 2.19E-05 4.85408469 ↑ CONTROLImmunoglobulin heavy variable 1-46 HV146 9.86E-06 4.84652628 ↑ CONTROLImmunoglobulin heavy variable 3-23 HV323 1.25E-05 4.75302522 ↑ CONTROLCholesteryl ester transfer protein CETP 1.22E-05 4.64058921 ↑ CONTROLApolipoprotein A-II APOA2 2.16E-08 4.62885891 ↑ CONTROLThyroxine-binding globulin THBG 1.80E-08 4.45994071 ↑ CONTROLProtein Z-dependent protease inhibitor ZPI 4.44E-05 4.43951224 ↑ CONTROLSerum amyloid A-4 protein SAA4 1.38E-07 4.39608833 ↑ CONTROLImmunoglobulin heavy variable 1-69 HV169 0.00058138 4.33088897 ↑ CONTROLImmunoglobulin kappa variable 1-9 KV109 3.99E-06 4.20727924 ↑ CONTROLGelsolin GELS 7.03E-09 4.09546999 ↑ CONTROLCD44 antigen CD44 0.00233936 4.0837648 ↑ CONTROLApolipoprotein C-I APOC1 0.00038806 3.66742486 ↑ CONTROLAlpha-2-macroglobulin A2MG 3.58E-09 3.65331919 ↑ CONTROLApolipoprotein D APOD 3.95E-08 3.53083248 ↑ CONTROLKallistatin KAIN 4.78E-10 3.48621242 ↑ CONTROLImmunoglobulin kappa variable 1-5 KV105 8.42E-05 3.22054945 ↑ CONTROLLeucine-rich alpha-2-glycoprotein A2GL 1.29E-08 3.00855446 ↑ CONTROLHemoglobin subunit beta HBB 8.64E-05 2.890312 ↑ CONTROLCoiled-coil domain-containing protein 8 CCDC8 6.16E-07 2.88193259 ↑ CONTROLImmunoglobulin kappa variable 2D-28 KVD28 0.00104876 2.6282284 ↑ CONTROLImmunoglobulin kappa variable 6D-21 KVD21 0.01695916 2.61227937 ↑ CONTROLImmunoglobulin lambda variable 4-69 LV469 8.14E-05 2.35130225 ↑ CONTROLBiotinidase BTD 0.00622085 2.19095666 ↑ CONTROLPlasma serine protease inhibitor IPSP 3.04E-05 2.1835559 ↑ CONTROLCarboxypeptidase B2 CBPB2 8.55E-05 1.8115321 ↑ CONTROLImmunoglobulin heavy variable 2-26 HV226 0.00977698 1.69947422 ↑ CONTROLApolipoprotein B-100 APOB 2.35E-05 1.66025805 ↑ CONTROLAlpha-2-antiplasmin A2AP 0.00014843 1.60147275 ↑ CONTROLApolipoprotein C-III APOC3 0.02144075 1.39259865 ↑ CONTROL
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
15

the regulation of metastasis [60].Validation assays verified and confirmed that pregnancy zone pro-
tein (PZP), coagulation factor V (FA5), protein Z-dependent proteaseinhibitor (ZPI), alpha-1-acid glycoprotein 1 (A1AG1) and inter-alpha-trypsin inhibitor heavy chain H1 (ITIH1), were statistically significantand appeared quantitatively up-regulated in triple negative breastcancer patients when compared with controls (healthy people). Thisresult was observed after the analysis of the protein corona formedaround MNPs (9.30 ± 0.67 nm).
Mainly, the pregnancy zone protein (PZP) was found to be im-plicated the pathogenesis of breast cancer [61].
It is well known that cancer is associated with hypercoagulability,and circumstantial evidence suggests that tumor-expressed coagulationfactors actively support cancer pathogenesis and progression. Notably,single nucleotide polymorphisms (SNPs) in F5, encoding coagulationfactor V (FA5), have been found associated with breast cancer. In 2018,M. Tinholt et al. [62] found that FA5 expression was higher in breasttumors compared to normal tissue. Importantly, FA5 expression wassignificantly increased in patients with tumors of aggressive nature(hormone receptor negative-, triple negative-, HER2 enriched-, andbasal-like tumors). These authors also suggested that tumor-expressedFA5 could be a possible marker of aggressive breast cancer, and it could
Table 8Significant proteins (p-value < 0.05) in comparisons between triple negative breast cancer and controls after the analysis of the protein corona of MNPs(9.30 ± 0.67 nm).
Protein UniProt ID p-value Fold change
TNBC patients vs. control C-reactive protein CRP 0.00169149 3.40849571 ↑ TNBCLipopolysaccharide-binding protein LBP 0.00837847 2.93851762 ↑ TNBCImmunoglobulin kappa variable 2-24 KV224 0.00583062 2.27733934 ↑ TNBCSerum amyloid P-component SAMP 0.00010388 2.20872021 ↑ TNBCKeratin. type II cytoskeletal 2 epidermal K22E 0.01682722 2.10615702 ↑ TNBCComplement C4-B CO4B 0.04180946 1.9800469 ↑ TNBCComplement component C9 CO9 5.75E-06 1.90848624 ↑ TNBCIgGFc-binding protein FCGBP 0.01481938 1.86546782 ↑ TNBCProtein Z-dependent protease inhibitor ZPI 0.00252121 1.78800298 ↑ TNBCComplement C1q subcomponent subunit B C1QB 0.00199072 1.78457812 ↑ TNBCPhosphatidylinositol-glycan-specific phospholipase D PHLD 0.00435099 1.73844701 ↑ TNBCProthrombin THRB 0.00750791 1.72802617 ↑ TNBCComplement C4-A CO4A 0.00510952 1.67059345 ↑ TNBCAlpha-1-acid glycoprotein 1 A1AG1 0.02552784 1.64048407 ↑ TNBCApolipoprotein C-III APOC3 0.03298589 1.61371634 ↑ TNBCKeratin. type II cytoskeletal 1 K2C1 0.01154011 1.60905471 ↑ TNBCSex hormone-binding globulin SHBG 0.02523265 1.59168855 ↑ TNBCInter-alpha-trypsin inhibitor heavy chain H3 ITIH3 0.00066933 1.55061894 ↑ TNBCComplement C3 CO3 6.41E-06 1.53090904 ↑ TNBCKeratin. type I cytoskeletal 10 K1C10 0.0384444 1.52567319 ↑ TNBCC4b-binding protein alpha chain C4BPA 0.00027605 1.51193215 ↑ TNBCComplement factor H CFAH 2.79E-05 1.46546396 ↑ TNBCPlasma serine protease inhibitor IPSP 0.00973255 1.4623027 ↑ TNBCHistidine-rich glycoprotein HRG 0.01164428 1.4616738 ↑ TNBCComplement factor B CFAB 0.00061903 1.45925143 ↑ TNBCPregnancy zone protein PZP 0.00155229 1.44791482 ↑ TNBCComplement C1q subcomponent subunit A C1QA 0.02763702 1.4062295 ↑ TNBCCoagulation factor V FA5 0.01312931 1.40131004 ↑ TNBCComplement factor I CFAI 7.52E-05 1.40080904 ↑ TNBCComplement C2 CO2 0.00061386 1.33762187 ↑ TNBCHemopexin HEMO 3.66E-05 1.29733733 ↑ TNBCCoagulation factor XIII B chain F13B 0.01256843 1.28491873 ↑ TNBCComplement C1q subcomponent subunit C C1QC 0.03780247 1.26491165 ↑ TNBCKininogen-1 KNG1 0.00429848 1.26291248 ↑ TNBCBeta-2-glycoprotein 1 APOH 0.04746608 1.24963502 ↑ TNBCApolipoprotein L1 APOL1 0.00237944 1.23613552 ↑ TNBCAntithrombin-III ANT3 0.01869897 1.22888805 ↑ TNBCProtein AMBP AMBP 0.01434176 1.22857819 ↑ TNBCVitronectin VTNC 0.01666796 1.21690616 ↑ TNBCInter-alpha-trypsin inhibitor heavy chain H1 ITIH1 0.01847152 1.20880937 ↑ TNBCInsulin-like growth factor-binding protein complex acid labile subunit ALS 0.03555036 1.20445695 ↑ TNBCHyaluronan-binding protein 2 HABP2 0.01352977 1.18411135 ↑ TNBCClusterin CLUS 0.04346683 1.1756767 ↑ TNBCAlpha-2-HS-glycoprotein FETUA 0.02891723 1.16730605 ↑ TNBCGalectin-3-binding protein LG3BP 0.00292722 1.116735 ↑ TNBCApolipoprotein C-I APOC1 0.02222358 2.50213212 ↑ CONTROLImmunoglobulin heavy constant mu IGHM 0.00022527 2.032717896 ↑ CONTROLImmunoglobulin lambda variable 8-61 LV861 0.00063688 2.023228572 ↑ CONTROLImmunoglobulin lambda variable 3-19 LV319 0.0082843 1.73294072 ↑ CONTROLCD5 antigen-like CD5L 0.02066785 1.731935965 ↑ CONTROLImmunoglobulin lambda variable 3-9 LV39 0.02497423 1.576244295 ↑ CONTROLApolipoprotein F APOF 0.02131359 1.556435198 ↑ CONTROLImmunoglobulin kappa variable 6-21 KV621 0.00608326 1.473240834 ↑ CONTROLImmunoglobulin heavy variable 3-23 HV323 0.02960462 1.38352045 ↑ CONTROLImmunoglobulin heavy variable 4-28 HV428 0.02327515 1.304707447 ↑ CONTROLPlasma kallikrein KLKB1 0.04204343 1.23839695 ↑ CONTROLApolipoprotein M APOM 0.01063485 1.232467766 ↑ CONTROL
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
16
emerge as a promising tumor suppressor candidate.Finally, protein Z-dependent protease inhibitor (ZPI) were identified
by immunochemistry in breast cancer cells, whereas they were absentfrom normal breast tissue [63], and alpha-1-acid glycoprotein (AGP)was also found to be a potential biomarker for breast cancer in 'at risk'individuals, particularly, TNBC patients [64].
4. Discussion
In this study, we provided by the first time the application of acombined proteomic approach as DDA and SWATH-MS to develop thecharacterization and quantification of triple negative breast cancerproteins after bio-corona nanoparticle protein pre-concentration.
Gold nanoparticles (AuNPs: 10.02 ± 0.91 nm), silver nanoparticles(AgNPs: 9.73 ± 1.70 nm) and magnetic nanoparticles (MNPs:(9.30 ± 0.67 nm) were assessed in biomarker discovery as a tool forthe pre-concentration and separation of proteins from complex pro-teomes. To this end, sera from eight healthy individuals were comparedwith sera from eight patients diagnosed with triple negative breastcancer. The application of these nanomaterials, combined with massspectrometry, has allowed the identification of seven potential bio-markers for the diagnostic and control of TNBC progression: GRF-typezinc finger domain-containing protein 1 (protein ZGRF1), Matrix me-talloproteinase-9 (MMP9), Lebercilin and Immunoglobulin lambdavariable 3-27 (LV327) and LINE-1 type transposase domain-containingprotein 1 (LITD1), structural maintenance of chromosomes protein 6(SMC6) and short coiled-coil protein (SCOC).
After performing over these samples, a SWATH analysis to quantifythe protein changes, the separation between the group of healthypeople and the group of triple negative breast cancer patients was ob-served. Moreover, a lot of deregulated proteins among both groupswere observed. However, these proteins are not among the alteredproteins find by the qualitative DDA assay. The proteomic methodsused in this study are complementary and allow improving character-ization studies. The fact that they are complementary and not ne-cessarily identify the same proteins is due to the search methods DDA(qualitative), and IDA (SWATH-quantitative) analysis is different.
So, in the analysis of the protein corona formed around the threenanoparticles (AuNPs, AgNPs, and MNPs), eight common proteinsshowed to be statistically significant and appeared quantitatively in-creased (up-regulated) in triple negative breast cancer patients versuscontrols (healthy people). These proteins are complement componentC9 (CO9), complement C4-A (CO4A), complement C3 (CO3), vi-tronectin (VTNC), apolipoprotein L1 (APOL1), complement factor H(CFAH), kininogen-1 (KNG1), galectin-3-binding protein (LG3BP). Andthree common proteins appeared quantitatively decreased (down-regulated) in triple negative breast cancer patients versus controls(healthy people). These proteins are immunoglobulin heavy constantmu (IGHM), immunoglobulin lambda variable 3-9 (LV39), and apoli-poprotein C-I (APOC1).
Moreover, a lot of deregulated proteins not common to all samplesincubated with the different nanoparticles were found. Therefore,protein corona formed around AuNPs in breast cancer patients showed14 proteins up-regulated and 34 proteins down-regulated, in compar-ison with the protein corona formed around AuNPs in healthy people.In the case of the protein corona formed around AgNPs, 64 proteinswere found to be up-regulated, and 76 proteins down-regulated.Finally, in the protein corona formed around the MNPs, 45 proteins hadelevated expression, while 12 proteins showed down-regulation. Fromthis point of view, AgNPs seem to be more suitable for a clinicaltranslational purpose, because the analysis of the protein coronaformed around these systems allows better differentiation between bothgroups of study: healthy and diseased individuals.
All these proteins can be considered potential triple negative breastcancer biomarker candidates; however, these proteins were found whenserum samples were concentrated using a bio-corona nanoparticle.
Fig. 5. Quantitative Venn diagrams showing the number of up-regulated pro-teins found in the protein corona of 10.02 ± 0.91 nm gold nanoparticles(AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs) and 9.30 ± 0.67 nmmagnetic nanoparticles (MNPs) after their incubation with serum from eighttriple negative breast cancer patients and eight healthy controls.
Fig. 6. Quantitative Venn diagrams showing the number of down-regulatedproteins found in the protein corona of 10.02 ± 0.91 nm gold nanoparticles(AuNPs), 9.73 ± 1.70 nm silver nanoparticles (AgNPs) and 9.30 ± 0.67 nmmagnetic nanoparticles (MNPs) after their incubation with serum from eighttriple negative breast cancer patients and eight healthy controls.
Fig. 7. Volcano plots of the SWATH analysis of proteins between healthy andtriple negative breast cancer patients with AuNPs (10.02 ± 0.91 nm), AgNPs(9.73 ± 1.70 nm) and MNPs (9.30 ± 0.67 nm). X-axis shows log(2)-foldchange and Y-axis the statistical significance through -log(10)-pvalue. The dotlines represent the cut off (p value ≤0.05). Proteins biomarkers are those thatwere common and statistical significative in all the nanoparticles.
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
17

Thus, it was thought that the best validation could be finding theseproteins in total serum samples. To this aim, it was performed a newSWATH-MS analysis improving the library, and it allowed us to findseveral validated proteins.
When it was compared total serum SWATH with concentratedproteins in AgNPs, we found again complement component C8 (CO8G),ficolin-3 (FCN3), retinol-binding protein 4 (RET4), fibronectin (FINC),fetuin-B (FETUB) and apolipoprotein A-IV (APOA4), up-regulated; andimmunoglobulin heavy constant gamma 4 (IGHG4), immunoglobulinkappa variable 2D-28 (KVD28), immunoglobulin kappa variable 1-5(KV105), apolipoprotein C-III (APOC3), immunoglobulin heavy vari-able 1-46 (HV146), immunoglobulin heavy constant gamma 2(IGHG2), immunoglobulin heavy constant gamma 3 (IGHG3), im-munoglobulin kappa variable 4-1 (KV401), immunoglobulin kappavariable 1D-12 (KVD12) and immunoglobulin heavy variable 3-30-5(HV335), down-regulated. It is consistent with data from previousstudies, and these proteins can be considered validated. In the com-parison between total serum SWATH and MNPs only found 5 up-regulated proteins pregnancy zone protein (PZP), coagulation factor V(FA5), protein Z-dependent protease inhibitor (ZPI), alpha-1-acid gly-coprotein 1 (A1AG1) and inter-alpha-trypsin inhibitor heavy chain H1(ITIH1). And finally, in the comparison between total serum SWATHand AuNPs, only one down-regulated protein was validated: trans-thyretin (TTHY).
This study shows that serum proteomics is a valuable tool that canfacilitate comprehensive and systematic identification of the serumproteome under both healthy and disease conditions. Thus, serumproteomics could be used for disease diagnosis and prognosis. In ourcase, we found several breast cancer-specific markers that can be usedin the diagnosis. However, due to we found a lot of differentiatedproteins, it is necessary complementary assays to reduce the number ofprotein biomarkers.
Declaration of competing interest
The authors declare that they have no conflict of interest.
Acknowledgment
All authors acknowledge Miguel Servet I Programme (CP16/00139)from the “Instituto de Salud Carlos III” (Plan Estatal de I+D+i 2013-2016 and European Development Regional Fund) of the SpanishMinistry of Science, Innovation and Universities. A. Castro López andM.P. Chantada-Vázquez contributed equally to this work.
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jprot.2019.103581.
References
[1] A. Jemal, F. Bray, M.M. Center, J. Ferlay, E. Ward, D. Forman, Global cancer sta-tistics, CA Cancer J. Clin. 61 (2) (2011) 69–90.
[2] M. Arnedos, C. Bihan, S. Delaloge, F. Andre, Triple-negative breast cancer: are wemaking headway at least? Ther. Adv. Med. Oncol. 4 (4) (2012) 195–210.
[3] F.C. Geyer, F. Pareja, B. Weigelt, E. Rakha, I.O. Ellis, S.J. Schnitt, J.S. Reis-Filho,The spectrum of triple-negative breast disease, high- and low-grade lesions, Am. J.Pathol. 187 (10) (2017) 2139–2151.
[4] S. Cleator, W. Heller, R.C. Coombes, Triple-negative breast cancer: therapeuticoptions, Lancet Oncol. 8 (3) (2007) 235–244.
[5] C. Núñez, Blood-based protein biomarkers in breast cancer, Clin. Chim. Acta 490(2019) 113–127.
[6] Y. He, A. Mohamedali, C. Huang, M.S. Baker, E.C. Nice, Oncoproteomics: currentstatus and future opportunities, Clin. Chim. Acta 495 (2019) 611–624.
[7] M. Wilhelm, J. Schlegl, H. Hahne, A. Moghaddas Gholami, M. Lieberenz,M.M. Savitski, E. Ziegler, L. Butzmann, S. Gessulat, H. Marx, T. Mathieson,S. Lemeer, K. Schnatbaum, U. Reimer, H. Wenschuh, M. Mollenhauer, J. Slotta-Huspenina, J.-H. Boese, M. Bantscheff, A. Gerstmair, F. Faerber, B. Kuster, Massspectrometry-based draft of the human proteome, Nature 509 (2014) 582–587.
[8] S. Miah, C.A.S. Banks, M.K. Adams, L. Florens, K.E. Lukong, M.P. Washburn,Advancement of mass spectrometry-based proteomics technologies to explore triplenegative breast cancer, Mol. BioSyst. 13 (1) (2016) 42–55.
[9] D.M. Schulz, C. Böllner, G. Thomas, M. Atkinson, I. Esposito, H. Höfler, M. Aubele,Identification of differentially expressed proteins in triple-negative breast carci-nomas using DIGE and mass spectrometry, J. Proteome Res. 8 (7) (2009)3430–3438.
[10] C. Ludwig, L. Gillet, G. Rosenberger, S. Amon, B.C. Collins, R. Aebersold, Data-independent acquisition-based SWATH-MS for quantitative proteomics: a tutorial,Mol. Syst. Biol. 14 (8) (2018) e8126.
[11] J.G. Meyer, B. Schilling, Clinical applications of quantitative proteomics usingtargeted and untargeted data-independent acquisition techniques, Expert Rev.Proteomics 14 (5) (2017) 419–429.
[12] M. Narasimhan, S. Kannan, A. Chawade, A. Bhattacharjee, R. Govekar, Clinicalbiomarker discovery by SWATH-MS based label-free quantitative proteomics: im-pact of criteria for identification of differentiators and data normalization method,J. Transl. Med. 17 (1) (2019) 184.
[13] N. Rifai, M.A. Gillette, S.A. Carr, Protein biomarker discovery and validation: thelong and uncertain path to clinical utility, Nat. Biotechnol. 24 (8) (2006) 971–983.
[14] R. Millioni, S. Tolin, L. Puricelli, S. Sbrignadello, G.P. Fadini, P. Tessari, G. Arrigoni,High abundance proteins depletion vs low abundance proteins enrichment: com-parison of methods to reduce the plasma proteome complexity, PLoS One 6 (5)(2011) e19603.
[15] E. Bellei, S. Bergamini, E. Monari, L.I. Fantoni, A. Cuoghi, T. Ozben, A. Tomasi,High-abundance proteins depletion for serum proteomic analysis: concomitant re-moval of non-targeted proteins, Amino Acids 40 (1) (2011) 145–156.
[16] S. Chutipongtanate, S. Chatchen, J. Svasti, Plasma prefractionation methods forproteomic analysis and perspectives in clinical applications, Proteomics Clin. Appl.11 (7-8) (2017).
[17] C.P. Albuquerque, M.B. Smolka, S.H. Payne, V. Bafna, J. Eng, H. Zhou, A multi-dimensional chromatography technology for in-depth phosphoproteome analysis,Mol. Cell. Proteomics 7 (7) (2008) 1389–1396.
[18] D.-G. Liu, L. Sun, Direct isolation of specific RNA-interacting proteins using a novelaffinity médium, Nucleic Acids Res. 33 (15) (2005) e132.
[19] P.D. Rakowska, M.G. Ryadnov, Nano-enabled biomarker discovery and detection,Biomark. Med. 5 (3) (2011) 387–396.
[20] N. Phogat, M. Kohl, I. Uddin, A. Jahan, Chapter 11 – Interaction of nanoparticleswith biomolecules, protein, enzymes, and its applications, in: Hans-Peter Deigner,Matthias Kohl (Eds.), Precision Medicine, Tools and Quantitative Approaches,Academic Press, 2018, pp. 253–276.
[21] S. Tenzer, D. Docter, S. Rosfa, A. Wlodarski, J. Kuharev, A. Rekik, S.K. Knauer,C. Bantz, T. Nawroth, C. Bier, J. Sirirattanapan, W. Mann, L. Treuel, R. Zellner,M. Maskos, H. Schild, R.H. Stauber, Nanoparticle size is a critical physicochemicaldeterminant of the human blood plasma corona: a comprehensive quantitativeproteomic analysis, ACS Nano 5 (2011) 7155–7167.
[22] M.P. Monopoli, D. Walczyk, A. Campbell, G. Elia, I. Lynch, F.B. Bombelli,K.A. Dawson, Physical-chemical aspects of protein corona: relevance to in vitro andin vivo biological impacts of nanoparticles, J. Am. Chem. Soc. 133 (2011)2525–2534.
[23] S. Laurent, C. Burtea, C. Thirifays, F. Rezaee, M. Mahmoudi, Significance of cell“observer” and protein source in nanobiosciences, J. Colloid Interface Sci. 392(2013) 431–445.
[24] M. Mahmoudi, A.M. Abdelmonem, S. Behzadi, J.H. Clement, S. Dutz, M.R. Ejtehadi,R. Hartmann, K. Kantner, U. Linne, P. Maffre, S. Metzler, M.K. Moghadam,C. Pfeiffer, M. Rezaei, P. Ruiz-Lozano, V. Serpooshan, M.A. Shokrgozar,G.U. Nienhaus, W.J. Parak, Temperature: the “ignored” factor at the NanoBio in-terface, ACS Nano 7 (2013) 6555–6562.
[25] S. Tenzer, D. Docter, J. Kuharev, A. Musyanovych, V. Fetz, R. Hecht, F. Schlenk,D. Fischer, K. Kiouptsi, C. Reinhardt, K. Landfester, H. Schild, M. Maskos,S.K. Knauer, R.H. Stauber, Rapid formation of plasma protein corona critically af-fects nanoparticle pathophysiology, Nat. Nanotechnol. 8 (2013) 772–781.
[26] T. Zheng, N. Pierre-Pierre, X. Yan, Q. Huo, A.J. Almodovar, F. Valerio, I. Rivera-Ramirez, E. Griffith, D.D. Decker, S. Chen, Gold nanoparticle-enabled blood test forearly stage cancer detection and risk assessment, ACS Appl. Mater. Interfaces 7 (12)(2015) 6819–6827.
[27] C. Corbo, R. Molinaro, M. Tabatabaei, O.C. Farokhzad, M. Mahmoudi, Personalizedprotein corona on nanoparticles and its clinical implications, Biomater. Sci. 5 (3)(2017) 378–387.
[28] R. Lopez-Cortés, E. Oliveira, C. Núñez, C. Lodeiro, M. Páez de la Cadena, F. Fdez-Riverola, H. López-Fernández, M. Reboiro-Jato, D. Glez-Peña, J.L. Capelo,H.M. Santos, Fast human serum profiling through chemical depletion coupled togold-nanoparticle-assisted protein separation, Talanta 100 (2012) 239–245.
[29] N.G. Bastus, F. Merkoci, J. Piella, V. Puntes, Synthesis of highly monodisperse ci-trate-stabilized silver nanoparticles of up to 200 nm: kinetic control and catalyticproperties, Chem. Mater. 26 (9) (2014) 2836–2846.
[30] A.H. Lu, E.L. Salabas, F. Schüth, Magnetic nanoparticles: synthesis, protection,functionalization, and application, Angew. Chemie-Int. Ed. 46 (2007) 1222–1244.
[31] S.E. Warder, L.A. Tucker, T.J. Strelitzer, E.M. McKeegan, J.L. Meuth, P.M. Jung,A. Saraf, B. Singh, J. Lai-Zhang, G. Gagne, J.C. Rogers, Reducing agent-mediatedprecipitation of high-abundance plasma proteins, Anal. Biochem. 387 (2) (2009)184–193.
[32] C. Fernández, H.M. Santos, C. Ruíz-Romero, F.J. Blanco, J.L. Capelo-Martínez, Acomparison of depletion versus equalization for reducing high-abundance proteinsin human serum, Electrophoresis 32 (21) (2011) 2966–2974.
[33] J.R. de Jesus, R. da Silva Fernandes, G. de Souza Pessôa, I.M. Raimundo Jr.,M.A.Z. Arruda, Depleting high-abundant and enriching low-abundant proteins in
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
18

human serum: an evaluation of sample preparation methods using magnetic na-noparticle, chemical depletion and immunoaffinity techniques, Talanta 170 (2017)199–209.
[34] M.P. Chantada-Vázquez, A. Castro López, S.B. Bravo, S. Vázquez-Estévez, B. Acea-Nebril, C. Núñez, Proteomic analysis of the bio-corona formed on the surface of (Au,Ag, Pt)-nanoparticles in human serum, Colloids Surf. B Biointerfaces 177 (2019)141–148.
[35] G. Candiano, M. Bruschi, L. Musante, L. Santucci, G.M. Ghiggeri, B. Carnemolla,P. Orecchia, L. Zardi, P.G. Righetti, Blue silver: a very sensitive colloidal CoomassieG-250 staining for proteome analysis, Electrophoresis 25 (9) (2004) 1327–1333.
[36] E. Oliveira, J.E. Araújo, S. Gómez-Meire, C. Lodeiro, C. Perez-Melon, E. Iglesias-Lamas, A. Otero-Glez, J.L. Capelo, H.M. Santos, Proteomics analysis of the perito-neal dialysate effluent reveals the presence of calcium-regulation proteins and acuteinflammatory response, Clin. Proteomics 11 (1) (2014) 17.
[37] I.V. Shilov, S.L. Seymour, A.A. Patel, A. Loboda, W.H. Tang, S.P. Keating,C.L. Hunter, L.M. Nuwaysir, D.A. Schaeffer, The paragon algorithm, a next gen-eration search engine that uses sequence temperature values and feature prob-abilities to identify peptides from tandem mass spectra, Mol. Cell. Proteomics 6(2007) 1638–1655.
[38] W.H. Tang, I.V. Shilov, S.L. Seymour, Nonlinear fitting method for determininglocal false discovery rates from decoy database searches, J. Proteome Res. 7 (2008)3661–3667.
[39] I. Ortea, I. Ruiz-Sánchez, R. Cañete, J. Caballero-Villarraso, M.D. Cañete,Identification of candidate serum biomarkers of childhood-onset growth hormonedeficiency using SWATH-MS and feature selection, J. Proteome 175 (2018)105–113.
[40] H.T. Tan, M.C.M. Chung, Label-free quantitative phosphoproteomics reveals reg-ulation of vasodilator-stimulated phosphoprotein upon stathmin-1 silencing in apair of isogenic colorectal cancer cell lines, Proteomics 18 (8) (2018) e1700242.
[41] M.A. Dobrovolskaia, A.K. Patri, J. Zheng, J.D. Clogston, N. Ayub, P. Aggarwal,B.W. Neun, J.B. Hall, S.E. McNeil, Interaction of colloidal gold nanoparticles withhuman blood: effects on particle size and analysis of plasma protein binding pro-files, Nanomedicine 5 (2009) 106–117.
[42] R. García-Álvarez, M. Hadjidemetriou, A. Sánchez-Iglesias, L.M. Liz-Marzán,K. Kostarelos, In vivo formation of protein corona on gold nanoparticles. The effectof size and shape, Nanoscale 10 (3) (2018) 1256–1264.
[43] S.T. Yang, Y. Liu, Y.W. Wang, A. Cao, Biosafety and bioapplication of nanomaterialsby designing protein-nanoparticle interactions, Small 9 (2013) 1635–1653.
[44] J. Chen, Y. Wang, X. Ding, Y. Huang, K. Xu, Magnetic solid-phase extraction ofproteins based on hydroxy functional ionic liquid-modified magnetic nanoparticles,Anal. Methods 6 (2014) 8358–8367.
[45] L. Treuel, D. Docter, M. Maskos, R.H. Stauber, R.H. Stauber, Protein corona – frommolecular adsorption to physiological complexity, Beilstein J. Nanotechnol. 6(2015) 857–873.
[46] M. Cassandri, A. Smirnov, F. Novelli, C. Pitolli, M. Agostini, M. Malewicz,G. Melino, G. Raschellà, Zinc-finger proteins in health and disease, Cell Death Dis. 3(2017) 17071.
[47] J. Jen, Y.-C. Wang, Zinc finger proteins in cancer progression, J. Biomed. Sci. 23(2016) 53.
[48] B.I. Bassey-Archibong, J.M. Kwiecien, S.B. Milosavljevic, R.M. Hallett, L.G. Rayner,M.J. Erb, C.J. Crawford-Brown, K.B. Stephenson, P.A. Bédard, J.A. Hassell,J.M. Daniel, Kaiso depletion attenuates transforming growth factor-β signaling and
metastatic activity of triple-negative breast cancer cells, Oncogenesis 5 (2016)e208.
[49] E.S. Radisky, D.C. Radisky, Matrix metalloproteinases as breast cancer drivers andtherapeutic targets, Front. Biosci. (Landmark Ed.) 20 (2015) 1144–1163.
[50] K. Kessenbrock, V. Plaks, Z. Werb, Matrix metalloproteinases: regulators of thetumor microenvironment, Cell 141 (1) (2010) 52–67.
[51] E.S. Radisky, D.C. Radisky, Matrix metalloproteinase-induced epithelial-mesench-ymal transition in breast cancer, J. Mammary Gland Biol. Neoplasia 15 (2) (2010)201–212.
[52] D.P. Nanda, H. Sil, S. Moulik, J. Biswas, S.S. Mandal, A. Chatterjee, Matrix me-talloproteinase-9 as a potential tumor marker in breast cancer, J. Environ. Pathol.Toxicol. Oncol. 32 (2) (2013) 115–129.
[53] L.J. van 't Veer, H. Dai, M.J. van de Vijver, Y.D. He, A.A. Hart, M. Mao, H.L. Peterse,K. van der Kooy, M.J. Marton, A.T. Witteveen, G.J. Schreiber, R.M. Kerkhoven,C. Roberts, P.S. Linsley, R. Bernards, S.H. Friend, Gene expression profiling predictsclinical outcome of breast cancer, Nature 415 (6871) (2002) 530–536.
[54] T.G. Kormelink, D.G. Powe, S.A. Kuijpers, A. Abudukelimu, M.H.A.M. Fens,E.H.E. Pieters, W.W. Kassing-van der Ven, H.O. Habashy, I.O. Ellis, B.R. Blokhuis,M. Thio, W.E. Hennink, G. Storm, F.A. Redegeld, R.M. Schiffelers, Immunoglobulinfree light chains are biomarkers of poor prognosis in basal-like breast cancer and arepotential targets in tumor-associated inflammation, Oncotarget 5 (10) (2014)3159–3167.
[55] S.L. Anwar, W. Wulaningsih, U. Lehmann, Transposable elements in human cancer:causes and consequences of deregulation, Int. J. Mol. Sci. 18 (5) (2017) pii: E974.
[56] C. Behrens, B. Binotti, C. Schmidt, C.V. Robinson, J.J. Chua, K. Kühnel, Crystalstructure of the human short coiled coil protein and insights into SCOC-FEZ1complex formation, PLoS One 8 (10) (2013) e76355.
[57] F.U. Nasim, S. Ejaz, M. Ashraf, A.R. Asif, M. Oellerich, G. Ahmad, G.A. Malik, Attiq-Ur- Rehman, Potential biomarkers in the sera of breast cancer patients fromBahawalpur, Pakistan, Biomark Cancer 4 (2012) 19–34.
[58] C. Jiao, L. Cui, A. Ma, N. Li, H. Si, Elevated serum levels of retinol-binding protein 4are associated with breast cancer risk: a case-control study, PLoS One 11 (12)(2016) e0167498.
[59] Y.K. Bae, A. Kim, M.K. Kim, J.E. Choi, S.H. Kang, S.J. Lee, Fibronectin expression incarcinoma cells correlates with tumor aggressiveness and poor clinical outcome inpatients with invasive breast cancer, Hum. Pathol. 44 (2013) 2028–2037.
[60] D. Arora, Z. Mahmood, J. George, A.K. Yadav, S. Kumar, U.S. Singh, Y. Shukla,Plasma protein profiling of breast cancer patients of North Indian population: apotential approach to early detection, J. Proteomics Bioinform. 6 (2013) 5.
[61] R. Kumar, E. Kuligina, A. Sokolenko, I. Bizin, A.K. Varma, S.K. Hasan, Identificationof Pregnancy Zone Protein in the Pathogenesis of Breast Cancer, (December 2018),https://doi.org/10.13140/RG.2.2.15317.06887.
[62] M. Tinholt, Ø. Garred, E. Borgen, E. Beraki, M. Sletten, K. Kleivi Sahlberg,P.M. Sandset, N. Iversen, Coagulation factor V is expressed in tumors and predictsfavorable outcome in aggressive breast cancer, Thromb. Res. 164 (2018)S183–S184.
[63] E. Sierko, M.Z. Wojtukiewicz, K. Ostrowska-Cichocka, L. Zimnoch, Protein Z-de-pendent protease inhibitor (ZPI) is present in loco in human breast cancer tissue,Thromb. Haemost. 104 (1) (2010) 183–185.
[64] K.D. Smith, J. Behan, G. Matthews-Smith, A.M. Magliocco, Alpha-1-acid glycopro-tein (AGP) as a potential biomarker for breast cancer, Glycosylation (2012),https://doi.org/10.5772/48177.
M. del Pilar Chantada-Vázquez, et al. Journal of Proteomics 212 (2020) 103581
19

International Journal of
Molecular Sciences
Article
Protein Corona Gold Nanoparticles FingerprintingReveals a Profile of Blood Coagulation Proteins in theSerum of HER2-Overexpressing BreastCancer Patients
María del Pilar Chantada-Vázquez 1,2, Antonio Castro López 3, María García-Vence 2,Benigno Acea-Nebril 4 , Susana B. Bravo 2,* and Cristina Núñez 1,*
1 Research Unit, Lucus Augusti University Hospital (HULA), Servizo Galego de Saúde (SERGAS),27002 Lugo, Spain; [email protected]
2 Proteomic Unit, Health Research Institute of Santiago de Compostela (IDIS), University Clinical Hospital ofSantiago de Compostela (CHUS), 15706 Santiago de Compostela, Spain; [email protected]
3 Breast Unit, Hospital Universitario Lucus Augusti (HULA), Servizo Galego de Saúde (SERGAS), 27002 Lugo,Spain; [email protected]
4 Department of Surgery, Breast Unit, Complexo Hospitalario Universitario A Coruña (CHUAC),Servizo Galego de Saúde (SERGAS), 15006 A Coruña, Spain; [email protected]
* Correspondence: [email protected] (S.B.B.); [email protected] (C.N.)
Received: 15 October 2020; Accepted: 9 November 2020; Published: 10 November 2020�����������������
Abstract: Breast cancer (BC) is a molecularly heterogeneous disease that encompasses five majormolecular subtypes (luminal A (LA), luminal B HER2 negative (LB-), luminal B HER2 positive (LB+),HER2 positive (HER2+) and triple negative breast cancer (TNBC)). BC treatment mainly dependson the identification of the specific subtype. Despite the correct identification, therapies could failin some patients. Thus, further insights into the genetic and molecular status of the different BCsubtypes could be very useful to improve the response of BC patients to the range of availabletherapies. In this way, we used gold nanoparticles (AuNPs, 12.96 ± 0.72 nm) as a scavenging toolin combination with Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS)to quantitatively analyze the serum proteome alterations in the different breast cancer intrinsicsubtypes. The differentially regulated proteins specific of each subtype were further analyzed withthe bioinformatic tools STRING and PANTHER to identify the major molecular function, biologicalprocesses, cellular origin, protein class and biological pathways altered due to the heterogeneityin proteome of the different BC subtypes. Importantly, a profile of blood coagulation proteins wasidentified in the serum of HER2-overexpressing BC patients.
Keywords: protein corona (PC); gold nanoparticles (AuNPs); breast cancer (BC); fingerprinting;SWATH-MS; HER2+
1. Introduction
Breast cancer (BC) is a heterogeneous disease that presents a wide variety of molecular and clinicalcharacteristics, as well as variability in clinical progression [1]. For the treatment choice, patients areclassified according to intrinsic biological subtypes within the BC spectrum, using clinical-pathologicalcriteria, i.e., the recognition of amplification and/or overexpression of the human epidermal growthfactor receptor 2 (HER2) oncogene, the immunohistochemical classification of the estrogen receptor(ER) and the progesterone receptor (PR) and Ki-67 labeling index [2]. This classification allows fora more personalized approach to medical treatments, with favorable results. However, despite thatalmost 10–15% of these patients still experience local or distant recurrences in the first five years
Int. J. Mol. Sci. 2020, 21, 8449; doi:10.3390/ijms21228449 www.mdpi.com/journal/ijms

Int. J. Mol. Sci. 2020, 21, 8449 2 of 18
from diagnosis [3]. Particularly, HER2-positive BC, defined by the overexpression of HER2 protein,represents 15–20% of BC cases [4,5] and is correlated with poor prognosis, high rates of recurrence andshort survival [6].
Classification of BC might be markedly improved if new biomarkers identified with the use ofhigh-throughput “omics” approaches could support diagnosis based on histopathological patterns [7–9].Nanomaterials have been introduced into the field of proteomics to establish a new and rapidly evolvingresearch area termed nanoproteomics [10].
It is well known that the dispersion of a nanomaterial in physiological fluid results in the formationof a protein shell named “protein corona” (PC). PC varies depending on the characteristics of thebiological media, the physical (size, shape, curvature) and chemical properties (composition, surfacecharge/chemistry and hydrophobicity/hydrophilicity) of the nanomaterial and the incubation time [11].Disease-associated biomarkers comprise less than 1% of serum proteins. In this way, through theformation of the PC, nanoparticles could act as sorbent materials for the enrichment of low-abundancepeptides/proteins presented in serum samples before the biomarker identification by mass spectrometry(MS) analysis [12–14]. Importantly, otherwise undetectable changes in protein concentration at anearly stage of the disease (as breast cancer), after any treatment (chemotherapy, immunotherapy) orsurgery could be detected analyzing the PC composition [15]. Thus, characterization of the PC aroundNPs offers distinct advantages over sole proteomic approaches and increases the success of identifyingmolecular targets [16].
Particularly, AuNPs present some properties to be used as suiTable Sorbent nanomaterials: highsurface-area-to-volume ratio, colloidal stability and the ability to conjugate with biomolecules [17].Here, the interaction of AuNPs (12.96 ± 0.72 nm) with the sera of disease-free women (healthy controls,HC) (n = 42) and BC patients (n = 42) allowed the pre-concentration of the low-abundance proteinsthorough the PC formation. Then, an exhaustive quantitative analysis of the PCs by SWATH-MSwas carried out to identify novel molecular targets associated to the different BC intrinsic subtypes(see Figure 1).
Figure 1. Simplified representation of the experimental procedure. AuNPs (12.96 ± 0.72 nm) wereincubated ex vivo with human serum samples obtained from HC (n = 42) and BC patients (n = 42).Ex vivo corona-coated AuNPs were recovered and purified from unbound proteins by centrifugation.The formed PCs were quantitatively characterized, analyzed by SWATH-MS and compared betweenthe groups.

Int. J. Mol. Sci. 2020, 21, 8449 3 of 18
2. Results
2.1. Incubation of AuNPs (12.96 ± 0.72 nm) with Human Serum Samples: Ex Vivo Protein Corona Formationand Characterization
Human serum samples from n = 42 healthy controls (HC) and n = 42 breast cancer (BC) patients(n = 42) were recruited, handled and analyzed in the same way as further detailed in Figure 2. The groupof BC patients was divided into the following biological subtypes: n = 11 patients with the luminal A(LA) subtype, n = 10 patients with the luminal B HER2 negative (LB-) subtype, n = 7 patients with theluminal B HER2 positive (LB+) subtype, n = 6 patients with the HER2 positive (HER2+) subtype andn = 8 patients with the triple negative breast cancer (TNBC) subtype. Patient clinical characteristics aresummarized in Table S1.
Figure 2. Flowchart depicting serum samples pretreatment and protein corona formation.
AuNPs with a size of 12.96 ± 0.72 nm were prepared by a chemical reduction method [12–14].As Figure 2 shows, proteins presented in serum samples (×2) were chemically reduced withdithiothreitol (DTT) and alkylated with iodoacetamide (IAA) before their ex vivo incubation withAuNPs (12.96 ± 0.72 nm) to get the formation of the PCs [12–14].
After the ex vivo incubation of AuNPs with human serum samples of HC (n = 42) and BC patients(n = 42), the resultant protein corona-coated AuNPs were centrifugated and structurally characterizedby dynamic light scattering (DLS) and negative stain transmission electron microscopy (TEM). DLSmeasurements showed that the interaction of serum proteins with the surface of AuNPs resulted in anincrease of the size of the AuNPs, from 12.96 ± 0.72 to 17.33 ± 1.55 (HC) and 17.13 ± 1.53 nm (BC) (seeTable S2). Probably, the preferential interaction of positively charged proteins with the AuNPs surfacepromoted the increase of the mean particle surface charge from −38.3 (bare AuNPs) to −30.5 (HC) and−30.3 mV (BC) [18,19]. TEM imaging revealed a well-dispersed nanoparticles population corroboratingthe PC formation around AuNPs (see Figure 3).

Int. J. Mol. Sci. 2020, 21, 8449 4 of 18
Figure 3. Negative stain TEM imaging of bare (A) and protein corona-coated AuNPs, recoveredpost-incubation with human serum obtained from HC (B) and BC patients (C). All scale bars are 50 nm.

Int. J. Mol. Sci. 2020, 21, 8449 5 of 18
2.2. Quantitative Analysis of the Protein Corona-Coated AuNPs by SWATH-MS
Corona proteins associated with AuNPs were separated by sodium dodecyl sulfate–polyacrylamidegel electrophoresis (SDS-PAGE). After a staining step, gels were processed following the methoddescribed in Section 4.5. The resulting peptides were then quantitatively analyzed by the emergingproteomic platform for label-free quantification SWATH-MS.
The comparison of the protein patterns of the ex vivo formed PCs allowed the identification of thedifferentially expressed proteins between HC and the different BC subtypes. The results were filteredto present a p-value ≤ 0.05, and, interestingly, n = 60 proteins were found to be differentially expressed,of which n = 42 were upregulated and n = 18 downregulated in BC patients for the LA subtype; n = 132were found to be differentially expressed (n = 100 upregulated and n = 32 downregulated) for theLB- subtype; n = 67 proteins were found to be differentially expressed (n = 59 upregulated and n = 8downregulated) for the LB+ subtype; n = 130 proteins were found to be differentially expressed (n = 95upregulated and n = 35 downregulated) for the HER2+ subtype; and n = 91 proteins were found tobe differentially expressed (n = 87 upregulated and n = 4 downregulated) for the TNBC subtype (seeTable 1). The full list of candidate protein biomarkers identified to be upregulated or downregulatedin each different BC subtypes in comparison to healthy controls (HC) with the fold-change values isshown in Table S3.
Table 1. Number of differentially expressed proteins (up- and downregulated) (p-value ≤ 0.05) foundin the protein patterns of the ex vivo formed coronas after the analysis by SWATH-MS for the differentbreast cancer subtypes (LA LB-, LB+, HER2+ and TNBC) in comparison with healthy controls (HC)samples. The number of differentially expressed proteins (up- and downregulated) specific to each ofthe five subtypes of BC found in the ex vivo formed coronas is also indicated.
SWATH-MS Analysis
ComparisonProtein Number (p-Value ≤ 0.05)
Total Upregulated Downregulated Specific Upregulated Downregulated
Controls vs. LA 60 42 18 8 4 4
Controls vs. LB- 132 100 32 27 25 2
Controls vs. LB+ 67 59 8 2 2 0
Controls vs. HER2+ 130 95 35 28 23 5
Controls vs. TNBC 91 87 4 10 9 1
The Venn diagram of statistically significant (up- and down)regulated proteins shows that sevenproteins were found to be commonly altered in all BC subtypes (see Figure 4): apolipoprotein C-III(APOC3), c-reactive protein (CRP), hemoglobin subunit beta (HBB), immunoglobulin heavy variable3–49 (IGHV3–49), serum amyloid A-4 protein (SAA4), serum amyloid P-component (APCS) andserotransferrin (TF) (Tables S4 and S5).
As mentioned above, subtype-specific unique proteins were also identified using SWATH-MS (seeFigure 4 and Table 2). Overall, n = 8 proteins were specifically associated in LA (of which n = 4 wereupregulated and n = 4 showed downregulation). For LB-, n = 27 proteins were found to be specificto this subtype (n = 25 with increased expression and n = 2 with decreased expression). In the LB+
subtype, only n = 2 specific proteins were found to be upregulated. In the HER2+ subtype, n = 28specific proteins were found (n = 23 upregulated and n = 5 downregulated). The TNBC subtypecomprised of n = 10 specific proteins (n = 9 upregulated and n = 1 downregulated).
Int. J. Mol. Sci. 2020, 21, 8449 6 of 18
Figure 4. Venn diagram showing the number of shared and specific (or unique) deregulated proteinsidentified in the PCs formed after the interaction of AuNPs (12.96 ± 0.72 nm) with serum samples ofthe different BC subtypes (LA, LB-, LB+, HER2+ and TNBC).
Table 2. Differentially expressed proteins (up- and downregulated) (p-value ≤ 0.05) found in theprotein patterns of the ex vivo formed coronas after the analysis by SWATH-MS specific (or unique) forthe different breast cancer subtypes (LA, LB-, LB+, HER2+ and TNBC) in comparison with healthycontrol (HC) samples. The fold change ratio was calculated as the ratio of geometric means of thesample replicates, which corresponds to calculating the normal arithmetic ratio of log-transformedareas and back-transforming.
Protein Name Gene p-Value Fold ChangeComplement C1r
subcomponent-like protein C1RL 0.0000979 1.614689351 ↑ Luminal A
Complement factor H-related protein 2 CFHR2 0.003228805 1.764346734 ↑ Luminal AComplement component C8 beta chain C8B 0.003730112 1.35440489 ↑ Luminal A
Lysosome-associated membraneglycoprotein 2 LAMP2 0.018383379 1.33466653 ↑ Luminal A
Immunoglobulin kappa variable 3–20 IGKV3–20 0.008581213 7.24153822 ↓ Luminal AImmunoglobulin heavy constant mu IGHM 0.038320909 2.180105502 ↓ Luminal AImmunoglobulin heavy variable 1–24 IGHV1–24 0.045225189 2.766559939 ↓ Luminal A
Protein Z-dependent protease inhibitor SERPINA10 0.045960336 2.020513374 ↓ Luminal AImmunoglobulin lambda variable 2–23 IGLV2–23 0.000000251 2.96506068 ↑ Luminal B HER2 NegImmunoglobulin heavy variable 3–53 IGHV3–53 0.00000690 3.231686892 ↑ Luminal B HER2 NegImmunoglobulin kappa variable 4–1 IGKV4–1 0.00000788 2.271628981 ↑ Luminal B HER2 Neg
Biotinidase BTD 0.0000142 1.571319968 ↑ Luminal B HER2 NegImmunoglobulin heavy constant alpha 1 IGHA1 0.0000181 2.45029046 ↑ Luminal B HER2 Neg
Serum paraoxonase/lactonase 3 PON3 0.0000344 1.614088523 ↑ Luminal B HER2 NegImmunoglobulin kappa constant IGKC 0.0000604 2.234283878 ↑ Luminal B HER2 Neg
Phospholipid transfer protein PLTP 0.000127276 1.491697089 ↑ Luminal B HER2 NegImmunoglobulin kappa variable 3–11 IGKV3–11 0.000235484 2.727555354 ↑ Luminal B HER2 NegImmunoglobulin heavy variable 3–9 IGHV3–9 0.000418498 2.709287474 ↑ Luminal B HER2 Neg
Alpha-mannosidase 2 MAN2A1 0.000577191 2.094925838 ↑ Luminal B HER2 NegImmunoglobulin heavy constant gamma 1 IGHG1 0.000838666 1.832731289 ↑ Luminal B HER2 Neg
Apolipoprotein B-100 APOB 0.001926757 1.618943103 ↑ Luminal B HER2 NegImmunoglobulin heavy constant alpha 2 IGHA2 0.00207989 2.137794634 ↑ Luminal B HER2 Neg
Basement membrane-specific heparansulfate proteoglycan core protein HSPG2 0.002185804 2.617271104 ↑ Luminal B HER2 Neg
Pregnancy zone protein PZP 0.002663618 4.343426266 ↑ Luminal B HER2 NegImmunoglobulin heavy variable 1–69D IGHV1–69D 0.00307313 2.762019962 ↑ Luminal B HER2 NegImmunoglobulin heavy variable 3–74 IGHV3–74 0.005843078 1.63300774 ↑ Luminal B HER2 Neg
Immunoglobulin lambda-likepolypeptide 5 IGLL5 0.006474796 1.674164832 ↑ Luminal B HER2 Neg
Immunoglobulin kappa variable 3D-20 IGKV3D-20 0.007387257 1.722873358 ↑ Luminal B HER2 NegL-lactate dehydrogenase B chain LDHB 0.011088233 1.464826821 ↑ Luminal B HER2 Neg
Platelet glycoprotein Ib alpha chain GP1BA 0.014355443 1.435497779 ↑ Luminal B HER2 NegApolipoprotein D APOD 0.02689663 1.565894604 ↑ Luminal B HER2 Neg
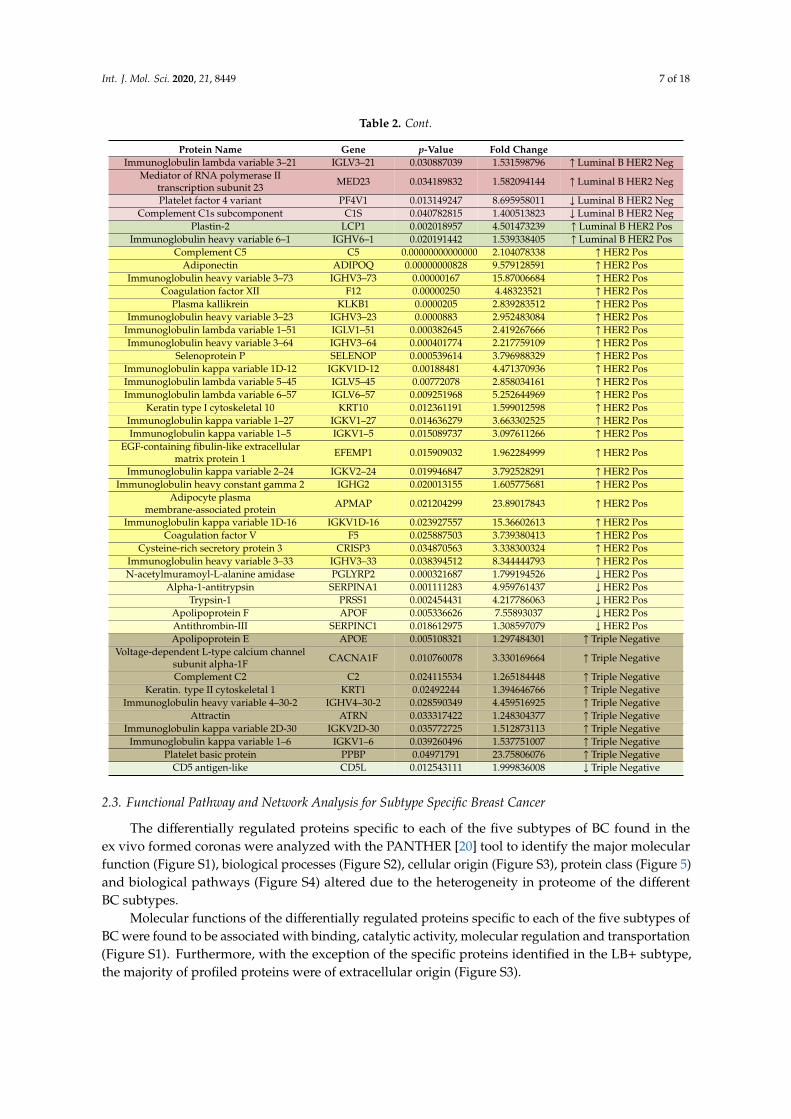
Int. J. Mol. Sci. 2020, 21, 8449 7 of 18
Table 2. Cont.
Protein Name Gene p-Value Fold ChangeImmunoglobulin lambda variable 3–21 IGLV3–21 0.030887039 1.531598796 ↑ Luminal B HER2 Neg
Mediator of RNA polymerase IItranscription subunit 23 MED23 0.034189832 1.582094144 ↑ Luminal B HER2 Neg
Platelet factor 4 variant PF4V1 0.013149247 8.695958011 ↓ Luminal B HER2 NegComplement C1s subcomponent C1S 0.040782815 1.400513823 ↓ Luminal B HER2 Neg
Plastin-2 LCP1 0.002018957 4.501473239 ↑ Luminal B HER2 PosImmunoglobulin heavy variable 6–1 IGHV6–1 0.020191442 1.539338405 ↑ Luminal B HER2 Pos
Complement C5 C5 0.000000000000002952.104078338 ↑ HER2 PosAdiponectin ADIPOQ 0.00000000828 9.579128591 ↑ HER2 Pos
Immunoglobulin heavy variable 3–73 IGHV3–73 0.00000167 15.87006684 ↑ HER2 PosCoagulation factor XII F12 0.00000250 4.48323521 ↑ HER2 Pos
Plasma kallikrein KLKB1 0.0000205 2.839283512 ↑ HER2 PosImmunoglobulin heavy variable 3–23 IGHV3–23 0.0000883 2.952483084 ↑ HER2 Pos
Immunoglobulin lambda variable 1–51 IGLV1–51 0.000382645 2.419267666 ↑ HER2 PosImmunoglobulin heavy variable 3–64 IGHV3–64 0.000401774 2.217759109 ↑ HER2 Pos
Selenoprotein P SELENOP 0.000539614 3.796988329 ↑ HER2 PosImmunoglobulin kappa variable 1D-12 IGKV1D-12 0.00188481 4.471370936 ↑ HER2 PosImmunoglobulin lambda variable 5–45 IGLV5–45 0.00772078 2.858034161 ↑ HER2 PosImmunoglobulin lambda variable 6–57 IGLV6–57 0.009251968 5.252644969 ↑ HER2 Pos
Keratin type I cytoskeletal 10 KRT10 0.012361191 1.599012598 ↑ HER2 PosImmunoglobulin kappa variable 1–27 IGKV1–27 0.014636279 3.663302525 ↑ HER2 PosImmunoglobulin kappa variable 1–5 IGKV1–5 0.015089737 3.097611266 ↑ HER2 Pos
EGF-containing fibulin-like extracellularmatrix protein 1 EFEMP1 0.015909032 1.962284999 ↑ HER2 Pos
Immunoglobulin kappa variable 2–24 IGKV2–24 0.019946847 3.792528291 ↑ HER2 PosImmunoglobulin heavy constant gamma 2 IGHG2 0.020013155 1.605775681 ↑ HER2 Pos
Adipocyte plasmamembrane-associated protein APMAP 0.021204299 23.89017843 ↑ HER2 Pos
Immunoglobulin kappa variable 1D-16 IGKV1D-16 0.023927557 15.36602613 ↑ HER2 PosCoagulation factor V F5 0.025887503 3.739380413 ↑ HER2 Pos
Cysteine-rich secretory protein 3 CRISP3 0.034870563 3.338300324 ↑ HER2 PosImmunoglobulin heavy variable 3–33 IGHV3–33 0.038394512 8.344444793 ↑ HER2 PosN-acetylmuramoyl-L-alanine amidase PGLYRP2 0.000321687 1.799194526 ↓ HER2 Pos
Alpha-1-antitrypsin SERPINA1 0.001111283 4.959761437 ↓ HER2 PosTrypsin-1 PRSS1 0.002454431 4.217786063 ↓ HER2 Pos
Apolipoprotein F APOF 0.005336626 7.55893037 ↓ HER2 PosAntithrombin-III SERPINC1 0.018612975 1.308597079 ↓ HER2 PosApolipoprotein E APOE 0.005108321 1.297484301 ↑ Triple Negative
Voltage-dependent L-type calcium channelsubunit alpha-1F CACNA1F 0.010760078 3.330169664 ↑ Triple Negative
Complement C2 C2 0.024115534 1.265184448 ↑ Triple NegativeKeratin. type II cytoskeletal 1 KRT1 0.02492244 1.394646766 ↑ Triple Negative
Immunoglobulin heavy variable 4–30-2 IGHV4–30-2 0.028590349 4.459516925 ↑ Triple NegativeAttractin ATRN 0.033317422 1.248304377 ↑ Triple Negative
Immunoglobulin kappa variable 2D-30 IGKV2D-30 0.035772725 1.512873113 ↑ Triple NegativeImmunoglobulin kappa variable 1–6 IGKV1–6 0.039260496 1.537751007 ↑ Triple Negative
Platelet basic protein PPBP 0.04971791 23.75806076 ↑ Triple NegativeCD5 antigen-like CD5L 0.012543111 1.999836008 ↓ Triple Negative
2.3. Functional Pathway and Network Analysis for Subtype Specific Breast Cancer
The differentially regulated proteins specific to each of the five subtypes of BC found in theex vivo formed coronas were analyzed with the PANTHER [20] tool to identify the major molecularfunction (Figure S1), biological processes (Figure S2), cellular origin (Figure S3), protein class (Figure 5)and biological pathways (Figure S4) altered due to the heterogeneity in proteome of the differentBC subtypes.
Molecular functions of the differentially regulated proteins specific to each of the five subtypes ofBC were found to be associated with binding, catalytic activity, molecular regulation and transportation(Figure S1). Furthermore, with the exception of the specific proteins identified in the LB+ subtype,the majority of profiled proteins were of extracellular origin (Figure S3).
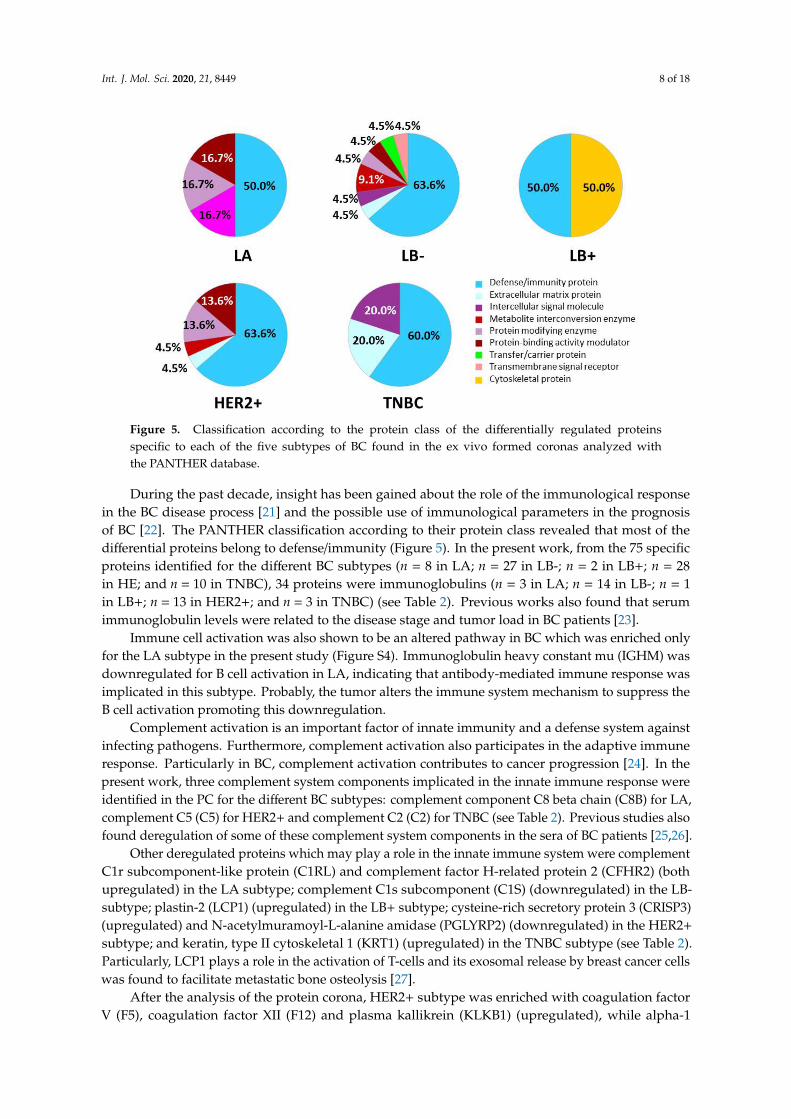
Int. J. Mol. Sci. 2020, 21, 8449 8 of 18
Figure 5. Classification according to the protein class of the differentially regulated proteinsspecific to each of the five subtypes of BC found in the ex vivo formed coronas analyzed withthe PANTHER database.
During the past decade, insight has been gained about the role of the immunological responsein the BC disease process [21] and the possible use of immunological parameters in the prognosisof BC [22]. The PANTHER classification according to their protein class revealed that most of thedifferential proteins belong to defense/immunity (Figure 5). In the present work, from the 75 specificproteins identified for the different BC subtypes (n = 8 in LA; n = 27 in LB-; n = 2 in LB+; n = 28in HE; and n = 10 in TNBC), 34 proteins were immunoglobulins (n = 3 in LA; n = 14 in LB-; n = 1in LB+; n = 13 in HER2+; and n = 3 in TNBC) (see Table 2). Previous works also found that serumimmunoglobulin levels were related to the disease stage and tumor load in BC patients [23].
Immune cell activation was also shown to be an altered pathway in BC which was enriched onlyfor the LA subtype in the present study (Figure S4). Immunoglobulin heavy constant mu (IGHM) wasdownregulated for B cell activation in LA, indicating that antibody-mediated immune response wasimplicated in this subtype. Probably, the tumor alters the immune system mechanism to suppress theB cell activation promoting this downregulation.
Complement activation is an important factor of innate immunity and a defense system againstinfecting pathogens. Furthermore, complement activation also participates in the adaptive immuneresponse. Particularly in BC, complement activation contributes to cancer progression [24]. In thepresent work, three complement system components implicated in the innate immune response wereidentified in the PC for the different BC subtypes: complement component C8 beta chain (C8B) for LA,complement C5 (C5) for HER2+ and complement C2 (C2) for TNBC (see Table 2). Previous studies alsofound deregulation of some of these complement system components in the sera of BC patients [25,26].
Other deregulated proteins which may play a role in the innate immune system were complementC1r subcomponent-like protein (C1RL) and complement factor H-related protein 2 (CFHR2) (bothupregulated) in the LA subtype; complement C1s subcomponent (C1S) (downregulated) in the LB-subtype; plastin-2 (LCP1) (upregulated) in the LB+ subtype; cysteine-rich secretory protein 3 (CRISP3)(upregulated) and N-acetylmuramoyl-L-alanine amidase (PGLYRP2) (downregulated) in the HER2+
subtype; and keratin, type II cytoskeletal 1 (KRT1) (upregulated) in the TNBC subtype (see Table 2).Particularly, LCP1 plays a role in the activation of T-cells and its exosomal release by breast cancer cellswas found to facilitate metastatic bone osteolysis [27].
After the analysis of the protein corona, HER2+ subtype was enriched with coagulation factorV (F5), coagulation factor XII (F12) and plasma kallikrein (KLKB1) (upregulated), while alpha-1

Int. J. Mol. Sci. 2020, 21, 8449 9 of 18
-antitrypsin (SERPINA1), trypsin-1 (PRSS1) and antithrombin-III (SERPINC1) (downregulated) forblood coagulation pathway (see Tables 2 and 3, and Figure S4).
Table 3. Candidate deregulated blood coagulation biomarkers in HER2-overexpressing BC patientsfound after the proteomic analysis of the ex vivo corona-coated AuNPs. On the bottom, a cluster ofblood coagulation proteins found in the protein–protein interaction network map of the genes encodeddifferentially regulated proteins for the HER2-overexpressing BC patients found after the proteomicanalysis of the ex vivo corona-coated AuNPs.
Protein Name Gene p-Value Fold Change Control vs. HER2 PositiveCoagulation factor XII F12 0.00000250 4.48323521 ↑ HER2 Pos
Plasma kallikrein KLKB1 0.0000205 2.839283512 ↑ HER2 PosCoagulation factor V F5 0.025887503 3.739380413 ↑ HER2 PosAlpha-1-antitrypsin SERPINA1 0.001111283 4.959761437 ↓ HER2 Pos
Trypsin-1 PRSS1 0.002454431 4.217786063 ↓ HER2 PosAntithrombin-III SERPINC1 0.018612975 1.308597079 ↓ HER2 Pos
Platelets, as small cell fragments, are not only important coagulation-related factors, also playa vital role in tumor progression [28]. Particularly, platelet factor 4 (PF4) or CXCL4, a member ofCXC chemokine family, acts as an angiogenesis inhibitor which may contribute to prevent tumormetastasis [29]. In the present work, it was observed that the inflammation mediated by chemokine andcytokine signaling pathway was enriched specifically for LB- subtype with the platelet factor 4 variant(PF4V1) being downregulated (Figure S4). Another protein of the family of platelets, the platelet basicprotein (PPBP), was upregulated in the TNBC subtype (see Table 2).
The gonadotropin-releasing hormone receptor pathway was found to be enriched with adiponectin(ADIPOQ) (upregulated) and voltage-dependent L-type calcium channel subunit alpha-1F (CACNA1F)(upregulated) in the HER2+ and TNBC subtypes, respectively (Figure S4).
In the present work, a group of proteins implicated in the combination and transportationof lipids, apolipoproteins, were also found to be deregulated in the LB- (apolipoprotein B-100(APOB), apolipoprotein D (APOD) and phospholipid transfer protein (PLTP); upregulated), HER2+
(apolipoprotein F (APOF); downregulated) and TNBC subtypes (apolipoprotein E (APOE); upregulated)(see Table 2).
Another family of potential molecular targets that was found to be deregulated in the present studyincludes some glycoproteins: lysosome-associated membrane glycoprotein 2 (LAMP2) (upregulated)in the LA subtype; platelet glycoprotein Ib alpha chain (GP1BA) and basement membrane-specificheparan sulfate proteoglycan core protein (HSPG2) (upregulated) in the LB- subtype; EGF-containingfibulin-like extracellular matrix protein 1 (EFEMP1) and selenoprotein P (SELENOP) (upregulated) inthe HER2+ subtype; and CD5 antigen-like (CD5L) (downregulated) in the TNBC subtype (see Table 2).
While some proteins with an enzymatic functionality such as biotinidase (BTD), serumparaoxonase/lactonase 3 (PON3), L-lactate dehydrogenase B chain (LDHB) and alpha-mannosidase 2(MAN2A1) were found to be the upregulated in LB- subtype, protein Z-dependent protease inhibitor(SERPINA10) were downregulated in the LA subtype (see Table 2).

Int. J. Mol. Sci. 2020, 21, 8449 10 of 18
3. Discussion
Nowadays, the different breast cancer intrinsic subtypes (LA, LB-, LB+, HER2+ and TNBC) guidethe therapy selection [2]. In some patients, therapies could fail for different reasons such as cancerrecurrence, therapy resistance and/or metastasis. Thus, therapy response in breast cancer patientscould be improved with the study of the molecular alterations at the subtype level.
The application of different omics approaches to deep insight the different BC subtypes allowedthe refinement of the complexity of tumor heterogeneity. In this way, a variety of quantitative proteomicstudies were developed to identify potential signatures for breast cancer using clinical samples such assaliva, the ductal lavage fluids, nipple aspirate fluids (NAFs), urine and tissue [30]. Importantly, bloodmay be a suiTable Sample source for studying the proteomic deregulation in the different BC subtypeswith minimally invasive collection procedures. Thus, the analysis of blood-based markers at thesubtype level in biological fluids such as plasma and serum allowed finding different protein markersrelated with the tumor microenvironment and the subtype-specific changes. Following this researchline, the proteomic alterations in blood plasma [7] and blood serum [8] of BC subtypes were explored.
Potential biomarkers are presented in very low concentrations in blood (less than 1% of bloodproteins). Thus, the isolation and enrichment of low-abundance peptides/proteins from complexmixtures is a mandatory step in the proteomic biomarkers pipeline, and nanoparticles represent anideal alternative [31].
In the present work, an exhaustive quantitative analysis of the PCs formed around AuNPs aftertheir incubation in serum samples was developed by SWATH-MS to identify novel molecular targetsassociated to the different BC intrinsic subtypes (see Figure 1).
Seven proteins were found to be commonly altered in all BC subtypes, namely APOC3, CRP,HBB, IGHV3–49, SAA4, APCS and TR. CRP and SAA4 are acute-phase proteins (APPs), a class ofproteins whose serum concentrations increase or decrease in response to inflammation. Particularly,a significant association of state of inflammation with stage of BC was previously described [32].A recent study found that elevated serum levels of CRP were associated considerably with a high riskof BC and poor outcome, including metastasis and recurrence [33]. In addition, the concentrations ofSAA4 increased gradually with tumor progression and the severity of BC stages [34]. Thus, CRP andSAA4 may be good candidate markers for the staging and prognosis of BC. The expression of HBB andTR, members of the globin family was also found to be associated with BC cells aggressiveness andpoor prognosis, indicating HBB and TR as novel biomarker for BC progression [35,36].
On the other hand, several studies point out that blood coagulation proteins develop an importantrole in tumor progression [37]. These works discussed the impact of the activation of the blood clottingcascade on primary tumor growth [38], tumor metastasis and cancer-associated thrombosis [39] andantitumor therapies that target blood-coagulation-associated proteins [40].
In the particular case of BC, different reports support blood coagulation proteins as an importantpatient factor that facilitates the metastatic potential [41]. Particularly, metastatic patients exhibitedsignificantly higher D-dimer values when compared with early breast cancer patients [42]. Furthermore,high plasma fibrinogen was found to be correlated with poor response to trastuzumab treatment inHER2 positive BC patients [43] and circulating levels of factor VIII (FVIII) were significantly associatedwith axillary lymph node involvement, number of metastatic nodes and HER2 status [44]. These studiessupport that the measurement of some coagulation-related biomarkers could provide additional datafor the evaluation of HER2 positive BC patients’ prognosis and could be novel molecular targets.
The present quantitative proteomic analysis revealed a profile of blood coagulation proteins forthe HER2+ subtype, namely F5, F12 and KLKB1 (upregulated), and SERPINA1, PRSS1 and SERPINC1)(downregulated). While F5 is expressed in tumors and indicates favorable outcome in aggressiveBC [45], F12 is involved in the pathogenesis of thrombosis through the induction and amplification ofthrombin generation [46].
KLKB1 (up), SERPINA1 (down) and SERPINC1 (down) are serine proteases. Particularly,SERPINA1 and SERPINC1 are serine proteases inhibitors (serpins) which belong to the protease inhibitor

Int. J. Mol. Sci. 2020, 21, 8449 11 of 18
family. Members of the kallikrein family, such as KLKB, were also found to be deregulated duringmalignant transformation [47]. Nevertheless, the variations in expression (downregulation/upregulation),activation and secretion are not substantial enough to consider them as suitable biomarkers for follow-updisease progression. SERPINA1 is synthesized and released by tumor cells and plays major rolesin physiologic and pathologic processes such as angiogenesis, tumor invasion and metastasis [48].It was found that that high expression of SERPINA1 could be predictive for a better clinical outcomeof ER+ and ER+/HER2+ patients. Thus, SERPINA1 was found to be a direct ER target gene and apredictor of survival in BC patients [49]. SERPINC1, an antithrombin, develops an important role asan inhibitor of the coagulation cascade. Furthermore, SERPINC1 also functions as an anti-angiogenic,anti-inflammatory, anti-viral and anti-apoptotic protein. The mechanism by which antithrombincontrols invasion, tumor migration and angiogenesis is by inhibition of enteropeptidase. This inhibitionshowed to be a double anti-tumor effect through producing an anti-angiogenic molecule and inhibitinga protease implicated in metastasis [50].
In the present work, the gonadotropin-releasing hormone receptor pathway was found to beenriched with adiponectin (ADIPOQ) in the HER2+ subtype. Although different studies reportedcontroversial findings in the association between ADIPOQ and BC, a recent meta-analysis suggests thatlow serum adiponectin concentration may be associated with an increased BC risk in premenopausaland postmenopausal women [51]. A negative correlation has been also demonstrated between ADIPOQlevels and tumor size and grade. Interestingly, the correlation between ADIPOQ and BC seems to bemore prominent in estrogen-negative and progesterone-negative BC [52]. Therefore, it seems theremay be a set group of BC patients that are most susceptible to the effects of ADIPOQ and would benefitmost from a potential treatment. ADIPOQ may serve as a biomarker of BC risk and help to identifysubjects at high risk for BC development.
Numerous research articles have accumulated solid evidence that lipoproteins are closely relatedto various types of tumorigenesis, as BC [53]. Apolipoproteins in the blood transfer lipids to cancercells to provide energy for cancer cell proliferation and invasion. Apolipoproteins also function asimportant factors in cellular signal transduction. In the present work, different apolipoproteins werefound to be deregulated in the different BC subtypes. Particularly, APOB, APOD and APOE in serumwere found to function as a risk factor for BC, being APOD and APOB involved in BC metastasis [54].Particularly, a recent study found that apolipoprotein B is a risk factor for development of intraocularmetastasis (IOM) in patients with BC [55].
Within the group of glycoproteins found to be deregulated in the present study, it was foundthat LAMP2 overexpression in breast tumors promotes cancer cell survival via chaperone-mediatedautophagy (CMA) [56]. Thus, inhibiting CMA activity in breast tumor cells (with a chemotherapeuticdrug, for example) can be exploited as a potential therapeutic application in the treatment of BC.HSPG2, also known as perlecan, is a heavily glycosylated protein component of the extra-cellularmatrix (ECM) that plays essential roles in tumor vascularization, that is closely related to tumorgrowth and metastasis [57]. Although HSPG2 expression in BC has not been examined in detail,a recent study investigated the expression of HSPG2 in human TNBC and the ability of anti-HSPG2antibodies to specifically target and inhibit tumor growth in a mouse xenograft model [58], showingthat HSPG2 is a promising therapeutic target in TNBC. EFEMP1, also known as fibulin 3, may have apotential cancer-promoting function in BC [59]. EFEMP1 expression decreases during BC progression,with low EFEMP1 levels correlating with a poorer prognosis. Functionally, high EFEMP1 levelsinhibited TGF-β-induced EMT, migration, invasion and endothelial permeability, while loss of EFEMP1expression/function promoted these TGF-β-mediated effects. Further, restoring EFEMP1 expression inbreast cancer cells inhibited TGF-β signaling, breast cancer cell EMT, invasion and metastasis in vivo.Although the role of CD5L in the oncogenesis of BC is not fully understood, a recent work found thatCD5L is upregulated in hepatocellular carcinoma and promotes liver cancer cell proliferation andantiapoptotic responses [60].

Int. J. Mol. Sci. 2020, 21, 8449 12 of 18
The enzyme LDHB, which was found to be upregulated in the LB- subtype, may help identifybreast cancers most likely to respond to neoadjuvant chemotherapy as well as those with the highestrisk of relapse that may benefit from additional adjuvant therapy [61].
All these novel molecular targets found in the serum of BC patients could detect a missing invasion,be performed in ambulatory settings, be repeatedly checked, and be applicable for BC diagnosis,the assessment of prognosis and selection of treatment.
Importantly, further insights exploring the deregulated blood coagulation proteins as potentialeffective prognosis biomarkers and targets for novel therapeutic approaches could have a great impactin the management of HER2-overexpressing BC patients.
4. Materials and Methods
4.1. Chemicals
Acrylamide/bis-acrylamide 30% solution (37.5:1), β-mercaptoethanol (molecular biology grade),Coomassie Brilliant Blue R250 (CBB), DL-dithiothreitol (HSCH2CH(OH)CH(OH)CH2SH, 99%) (DTT),glycerol (HOCH2CH(OH)CH2OH, 86–88%), iodoacetamide (IAA, ICH2CONH2, 99%), sodium citratetribasic dehydrate (HOC(COONa)(CH2COONa)2·2H2O, 99%), sodium carbonate (Na2CO3, 99%),tris-base (NH2C(CH2OH)3), trifluoroacetic acid (CF3COOH, 99%), trypsin from bovine pancreas andthe Sigma Marker wide range 6.5–200 kDa were purchased from Sigma-Aldrich. Formaldehydefor molecular biology (36.5–38% in H2O) and sodium dodecyl sulfate (SDS, CH3(CH2)11SO4Na)were purchased from Panreac. Bromophenol-blue was purchased from Riedel-de Haen. Hydrogentetrachloroaurate (III) hydrate (HAuCl4·xH2O) (99.9%-Au) (49% Au) at 10% w/v was purchased fromStrem Chemicals. Ammoniumbicarbonate (AMBIC, NH4HCO3, 99.5%) and formic acid (HCOOH,95%) were purchased from Fluka.
4.2. Biological Samples
Blood samples were collected from n = 42 newly diagnosed BC patients with the five differentbreast cancer subtypes: n = 11 patients with the luminal A subtype (ER positive, HER2 negative, Ki-67low, and PR high), n = 10 patients with the luminal B-HER2 negative subtype (ER positive, HER2negative, and either Ki-67 high or PR low), n = 7 patients with the luminal B-HER2 positive subtype(ER positive, HER2 overexpressed or amplified, any Ki-67 and any PR), n = 6 patients with the HER2positive subtype (HER2 over-expressed or amplified, ER and PR absent) and n = 8 patients with thetriple negative subtype (ER and PR absent and HER2 negative).
Blood samples were also collected from n = 42 age-matched and gender-matched healthy women(controls). In all cases, venous blood samples were collected in VACUETTE® Serum Clot ActivatorTubes (10 mL).
The experiment was conducted in conformity with the declaration of Helsinki and approved bythe Clinical Research Ethics Committees (CEIC) of Galicia (Spain) with approval number 2017-021.All participants from Lucus Augusti University Hospital (Spain) gave written informed consent priorto their participation.
4.3. Synthesis of Citrate-Gold Nanoparticles (AuNPs, 12.96 ± 0.72 nm)
Colloidal AuNPs with a size of 12.96 ± 0.72 nm were prepared by chemical reduction methodas per the protocol developed previously [12–14]. In short, sodium citrate tribasic solution (0.075%w/v) was dissolved in 60 mL warm distilled water under constant magnetic stirring. To this, 54 µLof 10% w/v of hydrogen tetrachloroaurate (III) hydrate solution was then added drop-wise and thereaction was heated to 100 ◦C under constant magnetic stirring. Reaction was allowed to proceedfurther the color of the solution changes from yellow to deep red indicating the reduction of Au3+ toAu0, which spontaneously aggregates to form colloidal AuNPs. This colloidal dispersion of AuNPswas cooled to room temperature and preserved at 4 ◦C for further analysis.

Int. J. Mol. Sci. 2020, 21, 8449 13 of 18
4.4. Incubation of AuNPs with Human Serum Samples: Ex Vivo Protein Corona Formation
Firstly, collected blood samples were allowed to clot for 15 min. After that, samples werecentrifuged for 5 min at 4 ◦C and 1800× g. Resultant serum were transferred to sterile cryovials, frozenand stored at −80 ◦C until further use at Research Unit, Lucus Augusti University Hospital (HULA).The formation of the ex vivo PC was achieved following the steps shown in Figure 2 [12–14]. Afterthat, unbound serum proteins from the surface of AuNPs were removed by centrifugation at 18,840× gfor 30 min.
4.5. Characterization of Colloidal AuNPs
The morphology of the AuNPs was investigated by transmission electron microscopy (TEM)with a JEM 1011, JEOL instrument. The size and ζ-potentials of colloidal AuNPs were measured(3 determinations per sample) with a Malvern Zetasizer Nano ZS instrument at 25 ◦C.
Protein quantification and protein separation by SDS-PAGE were carried out with the use of aQubit™ 4 Quantitation Starter Kit (Thermo Fisher Scientific) and a PowerPacTM Basic Power Supply(Bio-Rad), respectively.
4.6. Separation and Digestion of the Proteins Presented in the Corona-Coated AuNPs
Corona proteins associated with AuNPs were separated by sodium dodecyl sulfate–polyacrylamidegel electrophoresis (SDS-PAGE) and digested following the scheme represented in Figure 6.
Figure 6. Flowchart depicting the separation and digestion of the corona proteins associatedwith AuNPs.
The digestion was stopped with the addition of 50 µL of 5% (v/v) formic acid. After that,the extraction of the peptides from the gel was carried out with a solution of 50% (v/v) acetonitrile/0.1%(v/v) trifluoroacetic acid (TFA) (×3) and acetonitrile (ACN) (×1). Samples were dried and stored at−20 ◦C until their further use [62].
4.7. Protein Quantification by SWATH-MS
SWATH/MS experiments were carried out following the instrumental parameters describedelsewhere [13]. Briefly, two biological replicates of LA, LB-, LB+, HER+, TNBC and HC samples wereused to get extensive quantitative data by label-free SWATH-MS analysis. Peptides of all samples were

Int. J. Mol. Sci. 2020, 21, 8449 14 of 18
analyzed with a micro-LC system Ekspert nLC425 (Eksigen, Dublin, CA. USA) couplet to a hybridquadrupole-TOF mass spectrometer Triple TOF 6600 (Sciex, Redwood City. CA. USA). One of the firststeps is the construction of the MS/MS spectral libraries. For that purpose, peptide solutions wereanalyzed by a shotgun data-dependent acquisition (DDA) approach by micro-LC-MS/MS. For spectralalignment and peak extraction was employed the Peakview software (version 2.2; AB Sciex) usingthe SWATH Acquisition MicroApp (version 2.0). Parameters used were: number of fragments = 7,number of peptides = 10, peptide confidence = 95%, XIC width = 30 ppm and XIC extraction window= 5 min. Exportation of the SWATH file to the MarkerView software (version 1.3.1; AB Sciex) allowedthe quantitative analysis of ions, peptides and proteins in the different samples. As output result,the summed intensity of ions for the peptide, summed intensity of the peptides for protein and Areaunder Curve (AUC) of the ions were provided. The test set (LA, LB-, LB+, HER+ and TNBC) wascompared with the control (HC) dataset to generate fold change ratios. For protein quantitation,only peptides with a False Discovery Rate (FDR) below 1% were considered. Average MS peak area ofeach protein derived from the analysis of the biological replicates and Student’s t-test analysis amongsamples was developed. t-test indicates the capacity of each variable to distinguish between twogroups, and it was reported as a p-value. The criterion to select differentially expressed proteins was ap-value <0.05 with a 1.5-fold in- or decrease.
4.8. Protein Functional Interaction Network Analysis and Protein Ontology Classification
The informatic tool STRING v.10.0 database (http://string-db.org) was the used to analyze thefunctional interaction networks of the proteins, integrating direct (physical) and indirect protein–proteininteractions (PPI) [63].
Protein ontology classification was performed with the PANTHER classification system (http://www.pantherdb.org/). The differentially expressed proteins in the different breast cancer subtypeswere grouped according to their major molecular function, biological processes, cellular origin, proteinclass and biological pathways.
5. Conclusions
The quantitative comparison of the ex vivo PCs formed upon incubation of AuNPs with serumsamples obtained from BC patients revealed 75 deregulated subtype-specific unique proteins (8, 27, 2,28 and 10 proteins specifically associated to the LA, LB-, LB+, HER2+ and TNBC subtypes, respectively).The analysis of the ex vivo PCs formed onto AuNPs revealed a profile of blood coagulation proteinsin the serum of HER2-overexpressing BC patients that are implicated in breast tumor progression,including cellular transformation, proliferation, tumor cell survival and angiogenesis. Of all BC patients,HER2+ patients have the worst outcome. Further insights exploring these blood coagulation proteinsas potential effective prognosis biomarkers and targets for novel therapeutic approaches could have agreat impact on the management of HER2-overexpressing BC patients.
Supplementary Materials: The following are available online at http://www.mdpi.com/1422-0067/21/22/8449/s1.Figure S1. Classification according to the molecular function of the differentially regulated proteins specific toeach of the five subtypes of BC found in the ex vivo formed coronas analyzed with the PANTHER database,Figure S2. Classification according to the biological process of the differentially regulated proteins specific toeach of the five subtypes of BC found in the ex vivo formed coronas analyzed with the PANTHER database,Figure S3. Classification according to the cellular component of the differentially regulated proteins specific toeach of the five subtypes of BC found in the ex vivo formed coronas analyzed with the PANTHER database,Figure S4. Classification according to the biological pathway of the differentially regulated proteins specific toeach of the five subtypes of BC found in the ex vivo formed coronas analyzed with the PANTHER database,Table S1. Clinical features of breast cancer tumors, Table S2. The average mean hydrodynamic diameter (nm)values of bare and protein corona-coated AuNPs, recovered post-incubation with human serum obtained fromHC and BC patients, Table S3. Differentially expressed proteins (up- and downregulated) (p-value ≤ 0.05) foundin the protein patterns of the ex vivo formed coronas after the analysis by SWATH-MS for the different breastcancer subtypes (LA, n = 11; LB-, n = 10; LB+, n = 7; HER2+, n = 6; TNBC, n = 8) in comparison with healthycontrol (HC) samples. The accession number, species (Human) and fold change values are also reported, Table S4.Differentially expressed proteins (up- and downregulated) (p-value ≤ 0.05) found in the protein patterns of the

Int. J. Mol. Sci. 2020, 21, 8449 15 of 18
ex vivo formed coronas after the analysis by SWATH-MS for the different breast cancer subtypes (LA, n = 11;LB-, n = 10; LB+, n = 7; HER2+, n = 6; TNBC, n = 8) in comparison with healthy control (HC) samples, Table S5.Differentially expressed proteins (up- and downregulated) (p-value ≤ 0.05) found in the protein patterns of theex vivo formed coronas after the analysis by SWATH-MS common and specific for the different breast cancersubtypes (LA, n = 11; LB-, n = 10; LB+, n = 7; HER2+, n = 6; TNBC, n = 8) in comparison with healthy control(HC) samples. The accession number, gene name and species (Human) are reported.
Author Contributions: Conceptualization, C.N.; methodology, M.d.P.C.-V., A.C.L. and C.N.; software, M.d.P.C.-V.,A.C.L., S.B.B. and C.N.; formal analysis, M.d.P.C.-V., A.C.L., S.B.B. and C.N.; investigation, M.d.P.C.-V., A.C.L. andC.N.; resources, S.B.B., M.G.-V., B.A.-N. and C.N.; writing—original draft preparation, C.N.; writing—review andediting, C.N.; visualization, C.N.; supervision, C.N.; project administration, C.N.; and funding acquisition, C.N.All authors have read and agreed to the published version of the manuscript.
Funding: This work was supported by the grant from Instituto de Salud Carlos III (ISCIII) and European RegionalDevelopment Fund (FEDER) (CP16/00139).
Acknowledgments: We would like to acknowledge the essential contribution of the “Unidade de MicroscopíaElectrónica e Confocal” at “Campus Terra, Universidade de Santiago de Compostela, Spain” for the transmission electronmicroscopy (TEM) and ζ-potentials measurements. M.P. Chantada-Vázquez and A. Castro López contributedequally to this work.
Conflicts of Interest: The authors declare no conflict of interest.
Abbreviations
APPs Acute-phase proteinsAuNPs Gold nanoparticlesBC Breast cancerDTT DithiothreitolER Estrogen receptorHC Healthy controlsHER2 Human epidermal growth factor receptor 2IAA Iodoacetic acidLA Luminal ALB Luminal BMS Mass spectrometryPC Protein coronaPR Progesterone receptorSDS-PAGE Sodium dodecyl sulfate–polyacrylamide gel electrophoresisSWATH-MS Sequential Window Acquisition of All Theoretical Mass SpectraTEM Transmission electron microscopyTNBC Triple negative breast cancerTF Serotransferrin
References
1. Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W.; et al.Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics,and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [CrossRef] [PubMed]
2. Inic, Z.; Zegarac, M.; Inic, M.; Markovic, I.; Kozomara, Z.; Djurisic, I.; Inic, I.; Pupic, G.; Jancic, S. Differencebetween Luminal A and Luminal B Subtypes According to Ki-67, Tumor Size, and Progesterone ReceptorNegativity Providing Prognostic Information. Clin. Med. Insights Oncol. 2014, 8, 107–111. [CrossRef][PubMed]
3. Colleoni, M.; Sun, Z.; Price, K.N.; Karlsson, P.; Forbes, J.F.; Thürlimann, B.; Gianni, L.; Castiglione, M.;Gelber, R.D.; Coates, A.S.; et al. Annual Hazard Rates of Recurrence for Breast Cancer During 24 Years ofFollow-Up: Results from the International Breast Cancer Study Group Trials I to V. J. Clin. Oncol. 2016, 34,927–935. [CrossRef] [PubMed]
4. Loibl, S.; Gianni, L. HER2-positive breast cancer. Lancet 2017, 389, 2415–2429. [CrossRef]5. Asif, H.; Sultana, S.; Ahmed, S.; Akhtar, N.; Tariq, M. HER-2 Positive Breast Cance—A Mini-Review. Asian Pac.
J. Cancer Prev. 2016, 17, 1609–1615. [CrossRef]

Int. J. Mol. Sci. 2020, 21, 8449 16 of 18
6. Wolff, A.C.; Hammond, M.E.H.; Allison, K.H.; Harvey, B.E.; Mangu, P.B.; Bartlett, J.M.S.; Bilous, M.; Ellis, I.O.;Fitzgibbons, P.; Hanna, W.; et al. Human epidermal growth factor receptor 2 testing in breast cancer:American society of clinical oncology/college of american pathologists clinical practice guideline focusedupdate. J. Clin. Oncol. 2018, 36, 2105–2122. [CrossRef]
7. Suman, S.; Basak, T.; Gupta, P.; Mishra, S.; Kumar, V.; Sengupta, S.; Shukla, Y. Quantitative Proteomicsrevealed novel proteins associated with molecular subtypes of breast cancer. J. Proteom. 2016, 148, 183–193.[CrossRef]
8. Gajbhiye, A.; Dabhi, R.; Taunk, K.; Jagadeeshaprasad, M.G.; RoyChoudhury, S.; Mane, A.; Bayatigeri, S.;Chaudhury, K.; Santra, M.K.; Rapole, S. Multipronged quantitative proteomics reveals serum proteomealterations in breast cancer intrinsic subtypes. J. Proteom. 2017, 163, 1–13. [CrossRef]
9. Bouchal, P.; Schubert, O.T.; Faktor, J.; Capkova, L.; Imrichova, H.; Zoufalova, K.; Paralova, V.; Hrstka, R.;Liu, Y.; Ebhardt, H.A.; et al. Breast Cancer Classification Based on Proteotypes Obtained by SWATH MassSpectrometry. Cell Rep. 2019, 28, 832–843. [CrossRef]
10. Jia, L.; Lu, Y.; Shao, J.; Liang, X.J.; Xu, Y. Nanoproteomics: A new sprout from emerging links betweennanotechnology and proteomics. Trends Biotechnol. 2013, 31, 99–107. [CrossRef]
11. Pozzi, D.; Caracciolo, G.; Digiacomo, L.; Colapicchioni, V.; Palchetti, S.; Capriotti, A.L.; Cavaliere, C.; ZeneziniChiozzi, R.; Puglisi, A.; Laganà, A. The biomolecular corona of nanoparticles in circulating biological media.Nanoscale 2015, 7, 13958–13966. [CrossRef] [PubMed]
12. Chantada-Vázquez, M.P.; Castro López, A.; Bravo, S.B.; Vázquez-Estévez, S.; Acea-Nebril, B.; Núñez, C.Proteomic analysis of the bio-corona formed on the surface of (Au, Ag, Pt)-nanoparticles in human serum.Colloids Surf. B Biointerfaces 2019, 177, 141–148. [CrossRef] [PubMed]
13. Chantada-Vázquez, M.P.; Castro López, A.; García Vence, M.; Vázquez-Estévez, S.; Acea-Nebril, B.;Calatayud, D.G.; Jardiel, T.; Bravo, S.B.; Núñez, C. Proteomic investigation on bio-corona of Au, Agand Fe nanoparticles for the discovery of triple negative breast cancer serum protein biomarkers. J. Proteom.2020, 212, 103581. [CrossRef] [PubMed]
14. Chantada-Vázquez, M.P.; García-Vence, M.; Vázquez-Estévez, S.; Bravo, S.B.; Núñez, C. Identification of aProfile of Neutrophil-Derived Granule Proteins in the Surface of Gold Nanoparticles after Their Interactionwith Human Breast Cancer Sera. Nanomaterials (Basel) 2020, 10, 1223. [CrossRef] [PubMed]
15. Docter, D.; Westmeier, D.; Markiewicz, M.; Stolte, S.; Knauer, S.K.; Stauber, R.H. The nanoparticle biomoleculecorona: Lessons learned–challenge accepted? Chem. Soc. Rev. 2015, 44, 6094–6121. [CrossRef]
16. Lai, Z.W.; Yan, Y.; Caruso, F.; Nice, E.C. Emerging techniques in proteomics for probing nano-bio interactions.ACS Nano 2012, 6, 10438–10448. [CrossRef]
17. Bai, X.; Wang, Y.; Song, Z.; Feng, Y.; Chen, Y.; Zhang, D.; Feng, L. The Basic Properties of Gold Nanoparticlesand their Applications in Tumor Diagnosis and Treatment. Int. J. Mol. Sci. 2020, 21, 2480. [CrossRef]
18. Dobrovolskaia, M.A.; Patri, A.K.; Zheng, J.; Clogston, J.D.; Ayub, N.; Aggarwal, P.; Neun, B.W.; Hall, J.B.;McNeil, S.E. Interaction of colloidal gold nanoparticles with human blood: Effects on particle size andanalysis of plasma protein binding profiles. Nanomedicine 2009, 5, 106–117. [CrossRef] [PubMed]
19. García-Álvarez, R.; Hadjidemetriou, M.; Sánchez-Iglesias, A.; Liz-Marzán, L.M.; Kostarelos, K. In vivoformation of protein corona on gold nanoparticles. The effect of size and shape. Nanoscale 2018, 10, 1256–1264.[CrossRef]
20. Mi, H.; Muruganujan, A.; Thomas, P.D. PANTHER in 2013: Modeling the evolution of gene function, andother gene attributes, in the context of phylogenetic trees. Nucleic Acids Res. 2013, 41, D377–D386. [CrossRef]
21. Coussens, L.M.; Raymond, W.W.; Bergers, G. Inflammatory mast cells up-regulate angiogenesis duringsquamous epithelial carcinogenesis. Genes Dev. 1999, 13, 1382–1397. [CrossRef] [PubMed]
22. Singh, B.; Berry, J.A.; Shoher, A.; Lucci, A. COX-2 induces IL-11 production in human breast cancer cells.J. Surg. Res. 2006, 131, 267–275. [CrossRef] [PubMed]
23. Singh, R.P.; Singh, V.P.; Udupa, K.N. E-Rosette forming lymphocytes and serum immunoglobulins in breastcancer patients. Mater. Med. Pol. 1991, 23, 179–181. [PubMed]
24. Lu, Y.; Hu, X. C5a stimulates the proliferation of breast cancer cells via Akt-dependent RGC-32 gene activation.Oncol. Rep. 2014, 32, 2817–2823. [CrossRef] [PubMed]
25. Nasim, F.; Ejaz, S.; Ashraf, M.; Asif, A.R.; Oellerich, M.; Ahmad, G.; Malik, G.A.; Rehman, A.-U. PotentialBiomarkers in the Sera of Breast Cancer Patients from Bahawalpur, Pakistan. Biomark. Cancer 2012, 4, 19–34.[CrossRef] [PubMed]

Int. J. Mol. Sci. 2020, 21, 8449 17 of 18
26. Liu, A.; Qu, H.; Yu, C.; Liu, J.; Jiao, A.; Sun, P. Serum biomarkers for lymph node metastasis in patients withtriple-negative breast cancer by proteomics. Int. J. Clin. Exp. Pathol. 2017, 10, 3377–3384.
27. Tiedemann, K.; Sadvakassova, G.; Mikolajewicz, N.; Juhas, M.; Sabirova, Z.; Tabariès, S.; Gettemans, J.;Siegel, P.M.; Komarova, S.V. Exosomal Release of L-Plastin by Breast Cancer Cells Facilitates Metastatic BoneOsteolysis. Transl. Oncol. 2019, 12, 462–474. [CrossRef]
28. Ishikawa, S.; Miyashita, T.; Inokuchi, M.; Hayashi, H.; Oyama, K.; Tajima, H.; Takamura, H.; Ninomiya, I.;Ahmed, A.K.; Harman, J.W.; et al. Platelets surrounding primary tumor cells are related to chemoresistance.Oncol. Rep. 2016, 36, 787. [CrossRef]
29. Goodarzi, A.; Yari, F.; Mohammadipour, M.; Deyhim, M.R.; Naghadeh, H.T. Capability of Platelet Factor 4 toInduce Apoptosis in the Cancerous Cell Lines in Vitro. Int. J. Med. Lab. 2018, 5, 195–207.
30. Pendharkar, N.; Gajbhiye, A.; Taunk, K.; RoyChoudhury, S.; Dhali, S.; Seal, S.; Mane, A.; Abhang, S.;Santra, M.K.; Chaudhury, K.; et al. Quantitative tissue proteomic investigation of invasive ductal carcinomaof breast with luminal B HER2 positive and HER2 enriched subtypes towards potential diagnostic andtherapeutic biomarkers. J. Proteome 2016, 132, 112–130. [CrossRef]
31. Zhang, L.; Lu, H.; Yang, P. Recent developments of nanoparticle-based enrichment methods for massspectrometric analysis in proteomic. Sci. China Chem. 2010, 53, 695–703. [CrossRef]
32. Asegaonkar, S.B.; Takalkar, U.V.; Kodlikeri, P.; Pagdhune, A.; Bonduliya, V.; Thorat, A.P. Serum highsensitivity C-reactive protein in breast cancer patients. Int. J. Res. Med. Sci. 2014, 2, 1408–1411. [CrossRef]
33. Kaur, R.P.; Rubal; Banipal, R.P.S.; Vashistha, R.; Dhiman, M.; Munshi, A. Association of elevated levels ofC-reactive protein with breast cancer, breast cancer subtypes and poor outcome. Curr. Probl. Cancer 2019, 43,123–129. [CrossRef] [PubMed]
34. Zhang, G.; Sun, X.; Lv, H.; Yang, X.; Kang, X. Serum amyloid A: A new potential serum marker correlatedwith the stage of breast cancer. Oncol. Lett. 2012, 3, 940–944. [CrossRef] [PubMed]
35. Ponzetti, M.; Capulli, M.; Angelucci, A.; Ventura, L.; Monache, S.D.; Mercurio, C.; Calgani, A.; Sanita, P.;Teti, A.; Rucci, N. Non-conventional role of haemoglobin beta in breast malignancy. Br. J. Cancer 2017, 117,994–1006. [CrossRef]
36. Vandewalle, B.; Hornez, L.; Revillion, F.; Lefebvre, J. Secretion of transferrin by human breast cancer cells.Biochem. Biophys. Res. Commun. 1989, 163, 149–154. [CrossRef]
37. Lima, L.G.; Monteiro, R.Q. Activation of blood coagulation in cancer: Implications for tumour progression.Biosci. Rep. 2013, 33, e00064. [CrossRef]
38. Metelli, A.; Wu, B.X.; Riesenberg, B.; Guglietta, S.; Huck, J.D.; Mills, C.; Li, A.; Rachidi, S.; Krieg, C.;Rubinstein, M.P.; et al. Thrombin contributes to cancer immune evasion via proteolysis of platelet-boundGARP to activate LTGF-β. Sci. Transl. Med. 2020, 12, eaay4860. [CrossRef]
39. Hisada, Y.; Mackman, N. Cancer-associated pathways and biomarkers of venous thrombosis. Blood 2017,130, 1499–1506. [CrossRef]
40. Ruf, W.; Rothmeier, A.S.; Graf, C. Targeting clotting proteins in cancer therapy—Progress and challenges.Thromb. Res. 2016, 140, S1–S7. [CrossRef]
41. Lal, I.; Dittus, K.; Holmes, C.E. Platelets, coagulation and fibrinolysis in breast cancer progression.Breast Cancer Res. 2013, 15, 207. [CrossRef] [PubMed]
42. Tas, F.; Kilic, L.; Duranyildiz, D. Coagulation tests show significant differences in patients with breast cancer.Tumor Biol. 2014, 35, 5985–5992. [CrossRef]
43. Liu, Y.-L.; Lu, Q.; Liang, J.-W.; Xia, Y.; Zhang, W.; Hu, B.-Q.; Shang, F.-F.; Ji, Y.-R.; Wang, J.; Wang, Q.; et al.High Plasma Fibrinogen is Correlated with Poor Response to Trastuzumab Treatment in HER2 PositiveBreast Cancer. Medicine (Baltimore) 2015, 94, e481. [CrossRef] [PubMed]
44. Yigit, E.; Gönüllü, G.; Yücel, I.; Turgut, M.; Erdem, D.; Cakar, B. Relation between hemostatic parameters andprognostic/predictive factors in breast cancer. Eur. J. Intern. Med. 2008, 19, 602–607. [CrossRef] [PubMed]
45. Tinholt, M.; Garred, Ø.; Borgen, E.; Beraki, E.; Sletten, M.; Kleivi Sahlberg, K.; Sandset, P.M.; Iversen, N.Coagulation factor V is expressed in tumors and predicts favorable outcome in aggressive breast cancer.Thromb. Res. 2011, 164, S183. [CrossRef]
46. Mutch, N.J. Emerging roles for factor XII in vivo. J. Thromb. Haemost. 2011, 9, 1355–1358. [CrossRef][PubMed]
47. Kryza, T.; Silva, M.L.; Loessner, D.; Heuzé-Vourc’h, N.; Clements, J.A. The kallikrein-related peptidase family:Dysregulation and functions during cancer progression. Biochimie 2016, 122, 283–299. [CrossRef] [PubMed]

Int. J. Mol. Sci. 2020, 21, 8449 18 of 18
48. Zelvyte, I.; Sjögren, H.O.; Janciauskiene, S. Effects of native and cleaved forms of alpha1-antitrypsin on ME1477 tumor cell functional activity. Cancer Detect. Prev. 2002, 26, 256–265. [CrossRef]
49. Chan, H.J.; Li, H.; Liu, Z.; Yuan, Y.C.; Mortimer, J.; Chen, S. SERPINA1 is a direct estrogen receptor targetgene and a predictor of survival in breast cancer patients. Oncotarget 2015, 6, 25815–25827. [CrossRef]
50. Luengo-Gil, G.; Calvo, M.I.; Martín-Villar, E.; Águila, S.; Bohdan, N.; Antón, A.I.; Espín, S.; de la Peña, F.A.;Vicente, V.; Corral, J.; et al. Antithrombin controls tumor migration, invasion and angiogenesis by inhibitionof enteropeptidase. Sci. Rep. 2016, 6, 27544. [CrossRef]
51. Gu, L.; Cao, C.; Fu, J.; Li, Q.; Li, D.-H.; Chen, M.-Y. Serum adiponectin in breast cancer. A meta-analysis.Medicine (Baltimore) 2018, 97, e11433. [CrossRef] [PubMed]
52. Oh, S.W.; Park, C.-Y.; Lee, E.S.; Yoon, Y.S.; Lee, E.S.; Park, S.S.; Kim, Y.; Sung, N.J.; Yun, Y.H.; Lee, K.S.; et al.Adipokines, insulin resistance, metabolic syndrome, and breast cancer recurrence: A cohort study.Breast Cancer Res. 2011, 13, R34. [CrossRef] [PubMed]
53. Zhou, Y.; Luo, G. Apolipoproteins, as the carrier proteins for lipids, are involved in the development ofbreast cancer. Clin. Transl. Oncol. 2020, 22, 1952–1962. [CrossRef] [PubMed]
54. Cibeira, G.H.; Giacomazzi, J.; Aguiar, E.; Schneider, S.; Ettrich, B.; De Souza, C.I.; Camey, S.; Caleffi, M.;Weber, B.; Ashton-Prolla, P.; et al. Apolipoprotein E genetic polymorphism, serum lipoprotein levels andbreast cancer risk: A case-control study. Mol. Clin. Oncol. 2014, 2, 1009–1015. [CrossRef] [PubMed]
55. Liu, J.-X.; Yuan, Q.; Min, Y.-L.; He, Y.; Xu, Q.-H.; Li, B.; Shi, W.-Q.; Lin, Q.; Li, Q.-H.; Zhu, P.-W.; et al.Apolipoprotein A1 and B as risk factors for development of intraocular metastasis in patients with breastcancer. Cancer Manag. Res. 2019, 11, 2881–2888. [CrossRef] [PubMed]
56. Saha, T. LAMP2A overexpression in breast tumors promotes cancer cell survival via chaperone-mediatedautophagy. Autophagy 2012, 8, 1643–1656. [CrossRef]
57. Whitelock, J.M.; Graham, L.D.; Melrose, J.; Murdoch, A.D.; Iozzo, R.V.; Underwood, P.A. Human perlecanimmunopurified from different endothelial cell sources has different adhesive properties for vascular cells.Matrix Biol. 1999, 18, 163–178. [CrossRef]
58. Kalscheuer, S.; Khanna, V.; Kim, H.; Li, S.; Sachdev, D.; DeCarlo, A.; Yang, D.; Panyam, J. Discovery of HSPG2(Perlecan) as a Therapeutic Target in Triple Negative Breast Cancer. Sci. Rep. 2019, 9, 12492. [CrossRef]
59. Davidson, B.; Stavnes, H.T.; Holth, A.; Chen, X.; Yang, Y.; Shih, I.-M.; Wang, T.-L. Gene expression signaturesdifferentiate ovarian/peritoneal serous carcinoma from breast carcinoma in effusions. J. Cell. Mol. Med. 2011,15, 535–544. [CrossRef]
60. Aran, G.; Sanjurjo, L.; Bárcena, C.; Simon-Coma, M.; Téllez, E.; Vázquez-Vitali, M.; Garrido, M.; Guerra, L.;Díaz, E.; Ojanguren, I.; et al. CD5L is upregulated in hepatocellular carcinoma and promotes liver cancercell proliferation and antiapoptotic responses by binding to HSPA5 (GRP78). FASEB J. 2018, 32, 3878–3891.[CrossRef]
61. Dennison, J.B.; Molina, J.R.; Mitra, S.; González-Angulo, A.M.; Balko, J.M.; Kuba, M.G.; Sanders, M.E.;Pinto, J.A.; Gómez, H.L.; Arteaga, C.L.; et al. Lactate Dehydrogenase B: A Metabolic Marker of Response toNeoadjuvant Chemotherapy in Breast Cancer. Clin. Cancer Res. 2013, 19, 3703–3713. [CrossRef] [PubMed]
62. Oliveira, E.; Araújo, J.E.; Gómez-Meire, S.; Lodeiro, C.; Pérez-Melón, C.; Iglesias-Lamas, E.; Otero-Glez, A.;Capelo, J.L.; Santos, H.M. Proteomics analysis of the peritoneal dialysate effluent reveals the presence ofcalcium-regulation proteins and acute inflammatory response. Clin. Proteom. 2014, 11, 17. [CrossRef][PubMed]
63. Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.;Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree oflife. Nucleic Acids Res. 2015, 43, D447–D452. [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright © 2022 FDOKUMEN




















