
Antimicrobial and cytotoxic knotwood extracts and related pure compounds and their effects on...
9
Antimicrobial and cytotoxic knotwood extracts and related pure compounds and their effects on food-associated microorganisms Anna-Liisa Välimaa a,1 , Ulla Honkalampi-Hämäläinen b,1 , Suvi Pietarinen c , Stefan Willför c , Bjarne Holmbom c , Atte von Wright b, ⁎ a Tampere University of Technology, Institute of Environmental Engineering and Biotechnology, P.O. Box 541, FIN-33101 Tampere, Finland b University of Kuopio, Institute of Applied Biotechnology, P.O. Box 1627, FIN-70211 Kuopio, Finland c Åbo Akademi University, Process Chemistry Centre, c/o Laboratory of Wood and Paper Chemistry, Porthansgatan 3, FIN-20500 Turku/Åbo, Finland Received 25 April 2006; received in revised form 10 August 2006; accepted 26 October 2006 Abstract Knotwood or bark extracts prepared from 30 species of hard and soft wood trees as well as selected pure compounds (lignans, stilbenes and flavonoids) were assayed for their antimicrobial activity against a battery of both gram positive and negative bacteria, yeasts, and filamentous fungi (Bacillus cereus, Staphylococcus aureus, Listeria monocytogenes, Lactobacillus plantarum, Escherichia coli, Salmonella infantis, Pseu- domonas fluorescens, Candida albicans, Saccharomyces cerevisiae, Aspergillus fumigatus and Penicillium brevicompactum). By far the most consistent antibacterial and antifungal properties were associated with extracts of Pinus species. These extracts showed also cytotoxicity against a mouse hepatoma cell line. Both antimicrobial and cytotoxic properties correlated with the stilbene content of the extracts. Purified stilbenes showed the most consistent antimicrobial and cytotoxic activities, while purified lignans had marginal effects, only. The results suggest that stilbenes account both for the antimicrobial and cytotoxic properties of Pinus knotwood extracts. © 2006 Elsevier B.V. All rights reserved. Keywords: Knotwood; Pinus; Food microorganisms; Antibacterial activity; Antifungal activity; Cytotoxicity; Stilbenes; Pinosylvin 1. Introduction Plant-associated phenolic compounds have attracted atten- tion as potential antimicrobials for food protection or even for therapeutic purposes. The antimicrobial screening of 13 pure substances (tannins and flavonoids) and 29 plant extracts (mostly from food plants and herbs, but also from birch, Norway spruce and Scots pine) revealed a variable range of antibacterial properties. Sensitive bacterial species included staphylococci (Staphylococcus aureus and S. epidermidis), Bacillus subtilis, Micrococcus luteus, Escherichia coli and Pseudomonas aeruginosa. Antifungal properties were generally not observed (Rauha et al., 2000). In a recent study with berry-associated phenolics (ellagitan- nins, anthocyanins, cathecins and proanthocyanidins) marked antibacterial effects were detected against Salmonella enterica serovars and against S. aureus, but not against Listeria monocytogenes (with the exception of cranberry extracts) or Lactobacillus rhamnosus (Puupponen-Pimiä et al., 2005). Trees are rich in polyphenolic compounds, and might serve as a source of potentially useful substances, if their antimicro- bial properties and safety can be established. Knotwood, or the branch stub embedded in a tree stem, could be particularly promising in this respect. Knotwood of several softwoods, more specifically different Abies (Willför et al., 2004a), Picea (Willför et al., 2004b), and Pinus (Willför et al., 2003a; Lindberg et al., 2004) species, as well as hardwood species such as Populus grandidentata, P. tremuloides, P. tremula (Pietarinen et al., in press), and Acacia mangium and A. crassicarpa (Pietarinen et al., 2004) have been found to contain exception- ally large amounts of extractable polyphenols. Softwood knots International Journal of Food Microbiology 115 (2007) 235 – 243 www.elsevier.com/locate/ijfoodmicro ⁎ Corresponding author. Tel.: +358 17 162087; fax: +358 17 163322. E-mail address: [email protected] (A. von Wright). 1 These authors have equally contributed to the present paper. 0168-1605/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.ijfoodmicro.2006.10.031
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Antimicrobial and cytotoxic knotwood extracts and related pure compounds and their effects on...

biology 115 (2007) 235–243www.elsevier.com/locate/ijfoodmicro
International Journal of Food Micro
Antimicrobial and cytotoxic knotwood extracts and related pure compoundsand their effects on food-associated microorganisms
Anna-Liisa Välimaa a,1, Ulla Honkalampi-Hämäläinen b,1, Suvi Pietarinen c,Stefan Willför c, Bjarne Holmbom c, Atte von Wright b,⁎
a Tampere University of Technology, Institute of Environmental Engineering and Biotechnology, P.O. Box 541, FIN-33101 Tampere, Finlandb University of Kuopio, Institute of Applied Biotechnology, P.O. Box 1627, FIN-70211 Kuopio, Finland
c Åbo Akademi University, Process Chemistry Centre, c/o Laboratory of Wood and Paper Chemistry, Porthansgatan 3, FIN-20500 Turku/Åbo, Finland
Received 25 April 2006; received in revised form 10 August 2006; accepted 26 October 2006
Abstract
Knotwood or bark extracts prepared from 30 species of hard and soft wood trees as well as selected pure compounds (lignans, stilbenes andflavonoids) were assayed for their antimicrobial activity against a battery of both gram positive and negative bacteria, yeasts, and filamentousfungi (Bacillus cereus, Staphylococcus aureus, Listeria monocytogenes, Lactobacillus plantarum, Escherichia coli, Salmonella infantis, Pseu-domonas fluorescens, Candida albicans, Saccharomyces cerevisiae, Aspergillus fumigatus and Penicillium brevicompactum). By far the mostconsistent antibacterial and antifungal properties were associated with extracts of Pinus species. These extracts showed also cytotoxicity against amouse hepatoma cell line. Both antimicrobial and cytotoxic properties correlated with the stilbene content of the extracts. Purified stilbenesshowed the most consistent antimicrobial and cytotoxic activities, while purified lignans had marginal effects, only. The results suggest thatstilbenes account both for the antimicrobial and cytotoxic properties of Pinus knotwood extracts.© 2006 Elsevier B.V. All rights reserved.
Keywords: Knotwood; Pinus; Food microorganisms; Antibacterial activity; Antifungal activity; Cytotoxicity; Stilbenes; Pinosylvin
1. Introduction
Plant-associated phenolic compounds have attracted atten-tion as potential antimicrobials for food protection or even fortherapeutic purposes. The antimicrobial screening of 13 puresubstances (tannins and flavonoids) and 29 plant extracts(mostly from food plants and herbs, but also from birch,Norway spruce and Scots pine) revealed a variable range ofantibacterial properties. Sensitive bacterial species includedstaphylococci (Staphylococcus aureus and S. epidermidis),Bacillus subtilis, Micrococcus luteus, Escherichia coli andPseudomonas aeruginosa. Antifungal properties were generallynot observed (Rauha et al., 2000).
⁎ Corresponding author. Tel.: +358 17 162087; fax: +358 17 163322.E-mail address: [email protected] (A. von Wright).
1 These authors have equally contributed to the present paper.
0168-1605/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.ijfoodmicro.2006.10.031
In a recent study with berry-associated phenolics (ellagitan-nins, anthocyanins, cathecins and proanthocyanidins) markedantibacterial effects were detected against Salmonella entericaserovars and against S. aureus, but not against Listeriamonocytogenes (with the exception of cranberry extracts) orLactobacillus rhamnosus (Puupponen-Pimiä et al., 2005).
Trees are rich in polyphenolic compounds, and might serveas a source of potentially useful substances, if their antimicro-bial properties and safety can be established. Knotwood, or thebranch stub embedded in a tree stem, could be particularlypromising in this respect. Knotwood of several softwoods, morespecifically different Abies (Willför et al., 2004a), Picea(Willför et al., 2004b), and Pinus (Willför et al., 2003a;Lindberg et al., 2004) species, as well as hardwood species suchas Populus grandidentata, P. tremuloides, P. tremula (Pietarinenet al., in press), and Acacia mangium and A. crassicarpa(Pietarinen et al., 2004) have been found to contain exception-ally large amounts of extractable polyphenols. Softwood knots

236 A.-L. Välimaa et al. / International Journal of Food Microbiology 115 (2007) 235–243
contain mostly lignans, oligolignans, and stilbenes, whileflavonoids dominate in hardwood knots. For instance, theamount of polyphenols can be even up to 30% in knotwoodof Picea abies (Willför et al., 2003b). The stem heartwood ofPinus sylvestris can contain around 1% (w/w) whereas theknotwood contains around 1–7% of stilbenes alone (Willföret al., 2003a,b,c).
Polyphenols are enriched in knotwood probably due to theirstrong antioxidant (Willför et al., 2003b; Pietarinen et al., 2006)and antimicrobial (Lindberg et al., 2004) effects. For instance, thestilbene pinosylvin is known as both a fungicide and as abactericide (Bergman et al., 1965; Rennerfelt, 1943; Erdtman andRennerfelt, 1944). Furthermore, pinosylvin also has antioxidantproperties (Stojanovic and Brede, 2002). These properties areneeded most in branch knots, since an open wound is formedwhen a branch is broken close to the stem, making the treesusceptible to attack by microorganisms. Malterud et al. (1985)studied the antimicrobial properties of flavonoids present in
Table 1Main identified compounds in the tested knotwood and bark extracts
Tree species Main identified compound
Knotwood extractsAbies balsamea Lignans, oligolignans, juvAbies lasiocarpa Lignans, oligolignans, juvAbies alba Lignans, oligolignans, juvAbies sibirica Lignans, oligolignans, juvLarix decidua Lignans, oligolignans, flavLarix lariciana Lignans, flavonoids, oligoLarix sibirica Lignans, oligolignans, flavPicea abies Lignans, oligolignansPicea glauca Lignans, oligolignansPicea mariana Lignans, oligolignansPicea sitchensis Oligolignans, lignansPinus banksiana Lignans, flavonoids, pinosPinus contorta Flavonoids, pinosylvins, lPinus resinosa Pinosylvins, lignansPinus sibirica Pinosylvins, lignans, flavoPinus strobus Pinosylvins, flavonoids, liPinus sylvestris Pinosylvins, lignans, oligoPseudotsuga menziesii Flavonoids, lignans, oligoTsuga canadensis Lignans, oligolignansTsuga heterophylla Lignans, oligolignans, flavThuja occidentalis Thujalignans (related to thThuja plicata Thujalignans (related to thBetula pendula Polyphenolic unidentifiedPopulus grandidentata Flavonoids, flavonoid glyPopulus tremula Flavonoids, flavonoid glyPopulus tremuloides Flavonoid glycosides, flavAcacia crassicarpa Flavonoids, biflavonoidsAcacia mangium Flavonoids, biflavonoidsEucalyptus globulus Tannins, tannin monomersFagus sylvatica Flavonoids
Bark extractsBetula pendula Betuloside, flavonoids, suPinus banksiana Sugars, flavonoids, tanninThuja occidentalis Sugars, stilbene glucosidePseudotsuga menziesii Sugars, flavonoids, tanninPopulus tremula Unidentified glycosides, tPicea abies Stilbene glycosides, tanninAcacia lasiocarpa Sugars, resin acids, tanninPicea maritima Sugars, cathechin, tannins
Salix caprea and found that six identified flavonoids (dihydro-kaempferol, naringenin, taxifolin, prunin, and cathechin) wereeffective against both microbes and fungi. Naringenin, dihydro-kaempferol, and taxifolin have also been identified in P.tremuloides black galls, which are a certain type of plant tumoursobserved in trees resistant to certain fungal infections (Pausler etal., 1995). The predominant flavonoids in A. mangium branchknots, teracacidin and 3,4′,7,8-tetrahydroxyflavanone, alsopossess antifungal properties (Mihara et al., 2005).
In addition to antimicrobial effects stilbenes are also knownto have cytotoxic and apoptosis inducing effects on tumour cells.Particularly resveratrol (trans-3,4′,5-trihydroxystilbene) andrelated compounds have attracted attention due to theirantimicrobial effects against various bacteria and dermatophyticfungi (Man-Ying Chan, 2002). Resveratrol has also beenassociated with cancer prevention having shown in bioassayswith cancer cell lines and in animal studies interference in themajor steps of the carcinogenesis, initiation, promotion and
s Reference
abiones Willför et al. (2003c)abiones Pietarinen et al. (2006)abiones Pietarinen et al. (2006)abiones Willför et al. (2003c)onoids Willför et al. (2003c)lignans Pietarinen et al. (2006)onoids Pietarinen et al. (2006)
Willför et al. (2003c)Pietarinen et al. (2006)Pietarinen et al. (2006)Pietarinen et al. (2006)
ylvins Pietarinen et al. (2006)ignans Willför et al. (2003c)
Pietarinen et al. (2006)noids, oligolignans Willför et al. (2003c)gnans Pietarinen et al. (2006)lignans Willför et al. (2003)mers Willför et al. (2003c)
Pietarinen et al. (2006)onoids Pietarinen et al. (2006)ujaplicatin) Willför et al. (2003c)ujaplicatin) Pietarinen et al. (2006)compounds Willför et al. (2003c)cosides Pietarinen et al. (2006)cosides Pietarinen et al. (2006)onoids Pietarinen et al. (2006)
Pietarinen et al. (2006)Pietarinen et al. (2006)Pietarinen et al. (2006)Pietarinen et al. (2006)
gars, tannins Pietarinen et al. (2006)s Pietarinen et al. (2006)s, flavonoids, tannins Pietarinen et al. (2006)s Pietarinen et al. (2006)annins Pietarinen et al. (2006)s Pietarinen et al. (2006)s Pietarinen et al. (2006)
Pietarinen et al. (2006)

237A.-L. Välimaa et al. / International Journal of Food Microbiology 115 (2007) 235–243
progression (Jang et al., 1997; Boissy et al., 2005; Tolomeoet al., 2005).
In this study, the antimicrobial potencies of 38 hydrophilicknotwood extracts and ten pure polyphenols isolated fromknotwood extracts were screened using a variety of microorgan-isms associated with food. The cytotoxic properties of the mostpotent antimicrobial extracts and compounds against tumourcells were also assessed with a murine hepatic cell line. The aimwas to find out, whether tree-associated polyphenols, which areabundant in the side streams of wood processing industry, couldhave potential as antimicrobials in food applications or even asfunctional, antineoplastic food components.
2. Materials and methods
2.1. Preparation of knotwood extracts and individual purecompounds
Hydrophilic knotwood and bark extracts from 30 species oftrees (Table 1) were prepared according to Willför et al. (2003b)using an accelerated solvent extractor (ASE) apparatus (DionexCorp., Sunnyvale, CA). The lipophilic extractives were firstextracted with hexane, and thereafter the hydrophilic extractiveswere extractedwith an acetone/water (95:5 v/v)mixture. The barkextracts were from Abies lasiocarpa, Betula pendula, P. abies,Pinus banksiana, Pinus maritima, Populus tremula, Pseudot-sugamenziesii, and Thuja occidentalis. All the extracts have beencharacterised earlier and their chemical composition has beenreported byWillför et al. (2003c) and Pietarinen et al. (2006). Themain identified compounds in the extracts are given in Table 1.
Hydroxymatairesinol from P. abies knotwood, secoisolar-iciresinol from Araucaria angustifolia knotwood, nortrachelo-genin from P. sylvestris knotwood, and lariciresinol from Abiesbalsamea knotwood were prepared as described byWillför et al.(2003b). Matairesinol was synthesized from pure hydroxyma-tairesinol according to Eklund et al. (2003).
Pinocembrin was prepared from Pinus cembra knotwoodwith sequential extraction by ASE. Pinocembrin was fraction-ated from the hydrophilic extract by flash chromatography onnormal-phase silica gel columns, using dichloromethane aseluent (Neacsu et al., in press).
Pinosylvin (PS) and pinosylvin monomethyl ether (PSMME)were prepared from P. sylvestris knotwood by extraction, firstwith hexane and then with acetone in a large Soxhlet apparatus.Pure compounds were fractionated from the hydrophilic extractby flash chromatography on normal-phase silica gel columns,using dichloromethane as eluent. The purities of the compoundswere N95%.
Dihydropinosylvin monomethyl ether (DHPSMME) wasprepared from Pinus strobus knotwood by extraction, first withhexane and then with acetone, in a large Soxhlet apparatus.DHPSMME was fractionated from the hydrophilic extract byflash chromatography on normal-phase silica gel columns asdescribed above. The purity of the compound was N95%.
For the bioassays the knotwood and bark extractives weredissolved in 80% ethanol (v/v). The solvent for stilbenes,lignans and pinocembrin was acetone/water (95:5 v/v).
2.2. Analysis by gas chromatography (GC) and GC–massspectrometry (MS)
Lignans, stilbenes and flavonoids were analyzed on a 25-m�0.20-mm i.d., 0.11-μmHP-1 capillary column coated with cross-linked methyl polysiloxane (Agilent Technologies, Palo Alto,CA, USA). The gas chromatograph was a Perkin-Elmer Auto-System XL instrument (Perkin-Elmer, Boston, MA).
Other conditions were: column oven, 120 °C–6 °C/min–300 °C (10 min); carrier gas, H2 (20 ml/min); split injector,(1:20) 260 °C; FID detector, 300 °C; injection volume, 1 μl. Theethanol was evaporated and the extractives were silylated byaddition of 80 μl of bis-(trimethylsilyl)-trifluoroacetamide,20 μl of trimethylchlorosilane, and 20 μl of pyridine. Thereaction was completed by keeping the test tubes in an oven at70 °C for 1 h. Heneicosanoic acid and betulinol were used asinternal standards. A correction factor of 1.2 was used for thelignans that were calculated against betulinol (Willför et al.,2003b). Oligolignans, flavonoid glycosides, and biflavonoidswere quantified on a short 6-m � 0.53-mm i.d., 0.15-μm HP-1column, using cholesteryl heptadecanoate and 1,3-dipalmitoyl-2-oleyl glycerol as internal standards (Willför et al., 2004c;Pietarinen et al., 2004). The gas chromatograph was a Varian3400 instrument (Varian Inc., Palo Alto, CA, USA). Otherconditions were: column oven, 100 °C (1.5 min), 12 °C/min–340 °C (5 min); carrier gas, H2 (20 ml/min); SPI (septumequipped programmable injector), 80 °C (0.5 min)–200 °C/min–340 °C (18 min); FID detector, 340 °C; injection volume, 0.4 μl.Identification of individual components was performed by GC–MS analysis of the silylated components with an HP 6890-5973GC-quadrupole-MSD instrument (Agilent (Hewlett Packard),Warbrunn, Germany). A column similar as above and a 15-m ξ0.25-mm i.d., 0.1-μmMXT-65TG column (Restek Corp., USA),which allowed elution of the silylated oligolignans, flavonoidglycosides, and biflavonoids (Willför et al., 2004b; Pietarinenet al., 2004), were used.
2.3. Microbial strains and culture conditions
The microbial strains and media are listed in Table 2.Saccharomyces cerevisiae and Lactobacillus plantarum werefrom the Technical Research Centre of Finland (VTT) straincollection (Finland). ATCC strains were received from theAmerican Type Culture Collection (USA) and EELA strainsfrom The National Veterinary and Food Research Institute(Finland). The Pseudomonas fluorescens was freshly isolatedfrom fresh rainbow trout (Oncorhynchus mykiss) fillet atUniversity of Kuopio (Finland), and Aspergillus fumigatus HT30 was obtained from the National Public Health Institute(Finland).
S. aureus, Bacillus cereus, L. monocytogenes, E. coli andSalmonella infantis were grown at 37 °C for 24 h as Petri platecultures or for 16 h in broth. L. plantarum was culturedaerobically on plates for 48 h at 30 °C or in broth for 24–48 h. P.fluorescens was grown at 25 °C for 48 h both on agar and inbroth. Candida albicans and S. cerevisiae were grown for 24–48 h both on plates or in broth, the former at 37 °C and the latter

Table 2The microbial strains and isolates and the media used
Strain Growth medium
Gram positive bacteriaBacillus cereus EELA 37 Tryptone soy agar or broth
(Labema, UK)Listeria monocytogenes, EELA 626 Tryptone soy agar or broth
(Labema, UK)Lactobacillus plantarum E 71034 MRS agar or broth (Difco, USA)Staphylococcus aureus ATCC 25923 Tryptone soy agar or broth
(Labema, UK)
Gram negative bacteriaEscherichia coli ATCC 25922 Tryptone soy agar or broth
(Labema, UK)Pseudomonas fluorescens REB2 (freshisolate from fish material)
Nutrient agar or broth (Difco, USA)
Salmonella infantis EELA 72 Tryptone soy agar or broth(Labema, UK)
YeastsCandida albicans EELA 188 Potato dextrose agar or broth
(Labema, UK)Saccharomyces cerevisiae ALKO 743 Yeast–peptone–glucose agar or broth
(Difco, USA)
Filamentous fungiAspergillus fumigatus HT 30 Potato dextrose agar (Labema, UK)Penicillium brevicompactum EELA 450 Potato dextrose agar (Labema, UK)
238 A.-L. Välimaa et al. / International Journal of Food Microbiology 115 (2007) 235–243
at 30 °C. A. fumigatus and Penicillium brevicompactum werecultured on plates at 30 °C for 2–3 days and at 25 °C for 3–4 days, respectively.
2.4. Antimicrobial effects against bacteria and yeasts
The antimicrobial activities of test extracts towards bacteriaand yeasts were determined using an automated incubator and aturbidity reader (Bioscreen C, ThermoLabsystems, Helsinki,Finland), which enables the simultaneous testing of 200 samplesand monitoring of bacterial growth in real time during the test.
The growth inhibition tests were performed using brothsubcultures. The cells were harvested from stationary phaseculture by centrifugation and suspended in 5–10 ml of sterile0.9% (w/v) sodium chloride solution. The cell densities in thesuspensions were adjusted to 108 cfu/ml (bacteria) and 105 cfu/ml (yeast) by diluting with sterile 0.9% sodium chloridesolution. The cfu counts were based on OD600 values, whichhad been calibrated against plate counts. This suspension wasfurther hundredfold diluted in the corresponding culture broth toobtain the suspension to be used in the assays.
The growth inhibition was measured using 100-well micro-plates (Honeycomb, ThermoLabsystems, Helsinki, Finland).Each test extract was tested in three parallel wells by pipetting296 μl of assay medium, 100 μl of microbial cell suspension and4 μl of test extract (10 mg extractives/ml). The plates were thenincubated (at temperatures indicated in 2.3) until the stationaryphase was reached (12–36 h). The turbidity of each well wasmeasured every 15 min using a wide-band filter (420–580 nm).The plates were shaken continuously (yeasts and P. fluorescens)
or for 10 s (the other test microbes) prior to the measurement toachieve a homogenous suspension. The data obtained werecalculated as average areas under the growth curves of threeparallel wells and compared to data from solvent controls (thesolvent concentrations were non-inhibitory).
Bioassays for the antibacterial and anti-yeast activities wereperformed once to all extracts, purified lignans and pinocembrinas a preliminary screen. The extracts and pure compoundsshowing the strongest activity were chosen for a second roundof testing using three replicates per extract (with again threeparallel wells per measurement). Furthermore, purified stilbeneswere also tested. The concentrations of pure compounds in thereaction mixtures were adjusted to 0.5 mM.
2.5. Activity against filamentous fungi
The ability of the extracts to inhibit the germination andgrowth ofA. fumigatus andP. brevicompactumwas tested in plateassays by measuring the inhibition zones in the confluent fungalgrowth around wells containing the sample extract/solution. Thespore suspensions for the inoculation of the plates were preparedby cultivating the strains on PDA plates until sporulation (3–4 days). The plates were then rinsed with 8 ml of sterile distilledwater and filtered through sterile absorbent cotton.
Aliquots of 100 μl of spore suspension were used to seed2 ml tubes of 0.6% agar which were then poured on standardPDA plates to form a soft agar overlay. Wells were made in theplates using a sterile cork borer (∅ 4.5 mm). Aliquots of 30 μlof either the test substances (10 mg extractives/ml, or 50 mMsolutions of pure compounds), solvent controls [80% ethanoland acetone/water (95:5 v/v) mixture] or positive controlNatamycin (0.2 mg/ml, Abtek Biologicals, UK) were thenpipetted into the wells. The A. fumigatus test plates wereincubated at 30 °C for 2 days and those of P. brevicompactum at25 °C for 3 days. After incubation, the mean diameters of theinhibition zones around three wells were recorded.
2.6. Cytotoxicity tests with the mouse hepatoma cell line Hepa-1
Extracts and pure compounds showing marked antimicrobialproperties aswell as selected extractswith low activitywere testedfor their cytotoxicity towards a murine hepatic cell line. Thetoxicological endpoint of this cytotoxicity assay is cell viability,which is determined as relative total protein concentration(TPC) of the samples. A protein assay, based on the rapid reac-tion of fluorescamine (4-phenyl-spiro[furan-2(3H),1′-phthalan]-3,3′-dione) (Sigma, F-9015), with substances containing primaryamino groups to produce fluorescent products, was used.Measurements of fluorescence were made with a fluorescenceplate reader (Victor II, Perkin-Elmer, Wallac, Turku, Finland).
Cells of the subclone Hepa-1c1c7 of the mouse hepatoma celllineHepa-1 (Hankinson, 1979)were grown asmonolayer at 37 °Cin 5% CO2 atmosphere (HeraCell CO2 incubator, HeraeussInstruments GmbH and CoKG, Hanau Germany) in AlphaMEMmedium (BioWhittaker BE12-169F, MD, USA) supplementedwith 1% glutamine (Sigma G-7513), 10% foetal calf serum(CELLect GOLD, MP Biomedicals Inc., OH, USA) and 1%
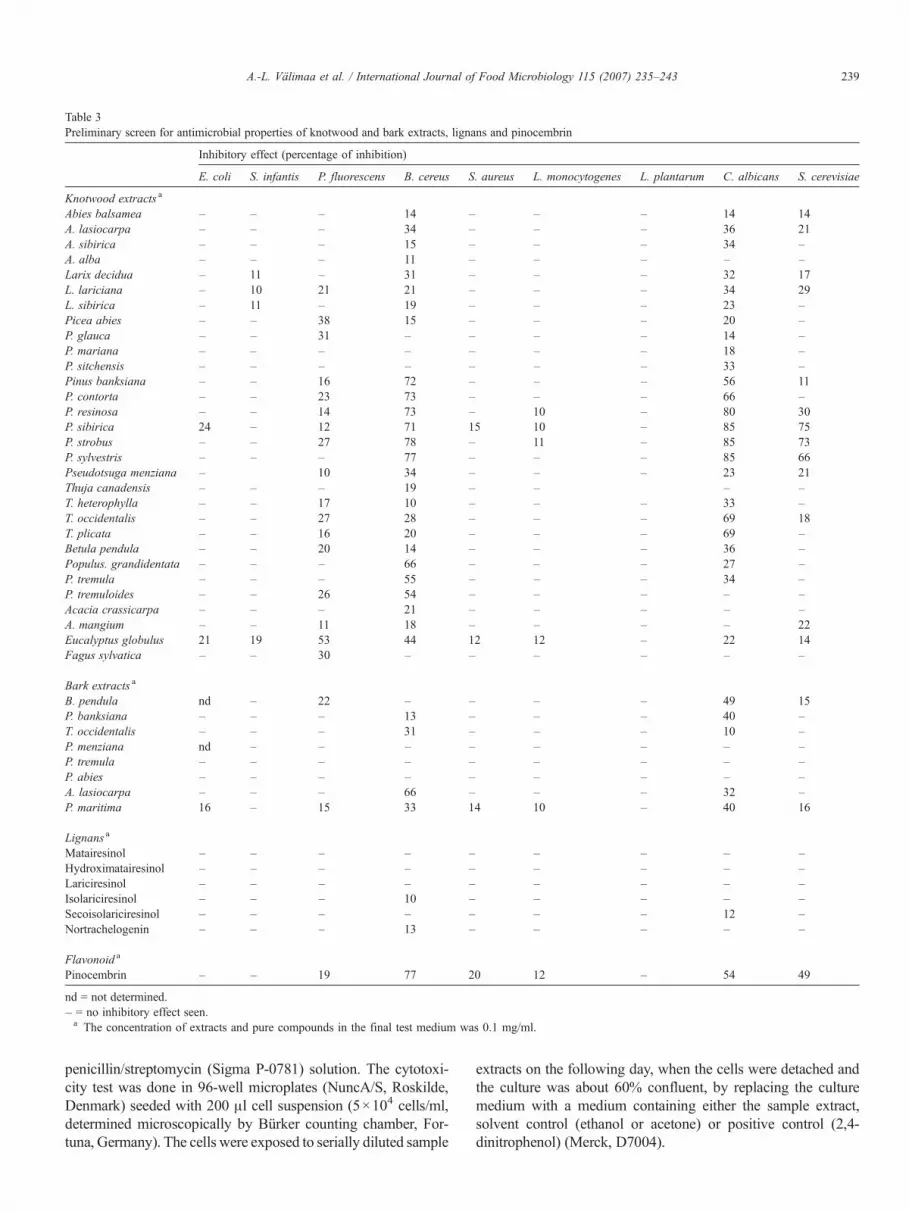
Table 3Preliminary screen for antimicrobial properties of knotwood and bark extracts, lignans and pinocembrin
Inhibitory effect (percentage of inhibition)
E. coli S. infantis P. fluorescens B. cereus S. aureus L. monocytogenes L. plantarum C. albicans S. cerevisiae
Knotwood extracts a
Abies balsamea – – – 14 – – – 14 14A. lasiocarpa – – – 34 – – – 36 21A. sibirica – – – 15 – – – 34 –A. alba – – – 11 – – – – –Larix decidua – 11 – 31 – – – 32 17L. lariciana – 10 21 21 – – – 34 29L. sibirica – 11 – 19 – – – 23 –Picea abies – – 38 15 – – – 20 –P. glauca – – 31 – – – – 14 –P. mariana – – – – – – – 18 –P. sitchensis – – – – – – – 33 –Pinus banksiana – – 16 72 – – – 56 11P. contorta – – 23 73 – – – 66 –P. resinosa – – 14 73 – 10 – 80 30P. sibirica 24 – 12 71 15 10 – 85 75P. strobus – – 27 78 – 11 – 85 73P. sylvestris – – – 77 – – – 85 66Pseudotsuga menziana – 10 34 – – – 23 21Thuja canadensis – – – 19 – – – –T. heterophylla – – 17 10 – – – 33 –T. occidentalis – – 27 28 – – – 69 18T. plicata – – 16 20 – – – 69 –Betula pendula – – 20 14 – – – 36 –Populus. grandidentata – – – 66 – – – 27 –P. tremula – – – 55 – – – 34 –P. tremuloides – – 26 54 – – – – –Acacia crassicarpa – – – 21 – – – – –A. mangium – – 11 18 – – – – 22Eucalyptus globulus 21 19 53 44 12 12 – 22 14Fagus sylvatica – – 30 – – – – – –
Bark extracts a
B. pendula nd – 22 – – – – 49 15P. banksiana – – – 13 – – – 40 –T. occidentalis – – – 31 – – – 10 –P. menziana nd – – – – – – – –P. tremula – – – – – – – – –P. abies – – – – – – – – –A. lasiocarpa – – – 66 – – – 32 –P. maritima 16 – 15 33 14 10 – 40 16
Lignans a
Matairesinol – – – – – – – – –Hydroximatairesinol – – – – – – – – –Lariciresinol – – – – – – – – –Isolariciresinol – – – 10 – – – – –Secoisolariciresinol – – – – – – – 12 –Nortrachelogenin – – – 13 – – – – –
Flavonoid a
Pinocembrin – – 19 77 20 12 – 54 49
nd = not determined.– = no inhibitory effect seen.a The concentration of extracts and pure compounds in the final test medium was 0.1 mg/ml.
239A.-L. Välimaa et al. / International Journal of Food Microbiology 115 (2007) 235–243
penicillin/streptomycin (Sigma P-0781) solution. The cytotoxi-city test was done in 96-well microplates (NuncA/S, Roskilde,Denmark) seeded with 200 μl cell suspension (5×104 cells/ml,determined microscopically by Bürker counting chamber, For-tuna, Germany). The cells were exposed to serially diluted sample
extracts on the following day, when the cells were detached andthe culture was about 60% confluent, by replacing the culturemedium with a medium containing either the sample extract,solvent control (ethanol or acetone) or positive control (2,4-dinitrophenol) (Merck, D7004).
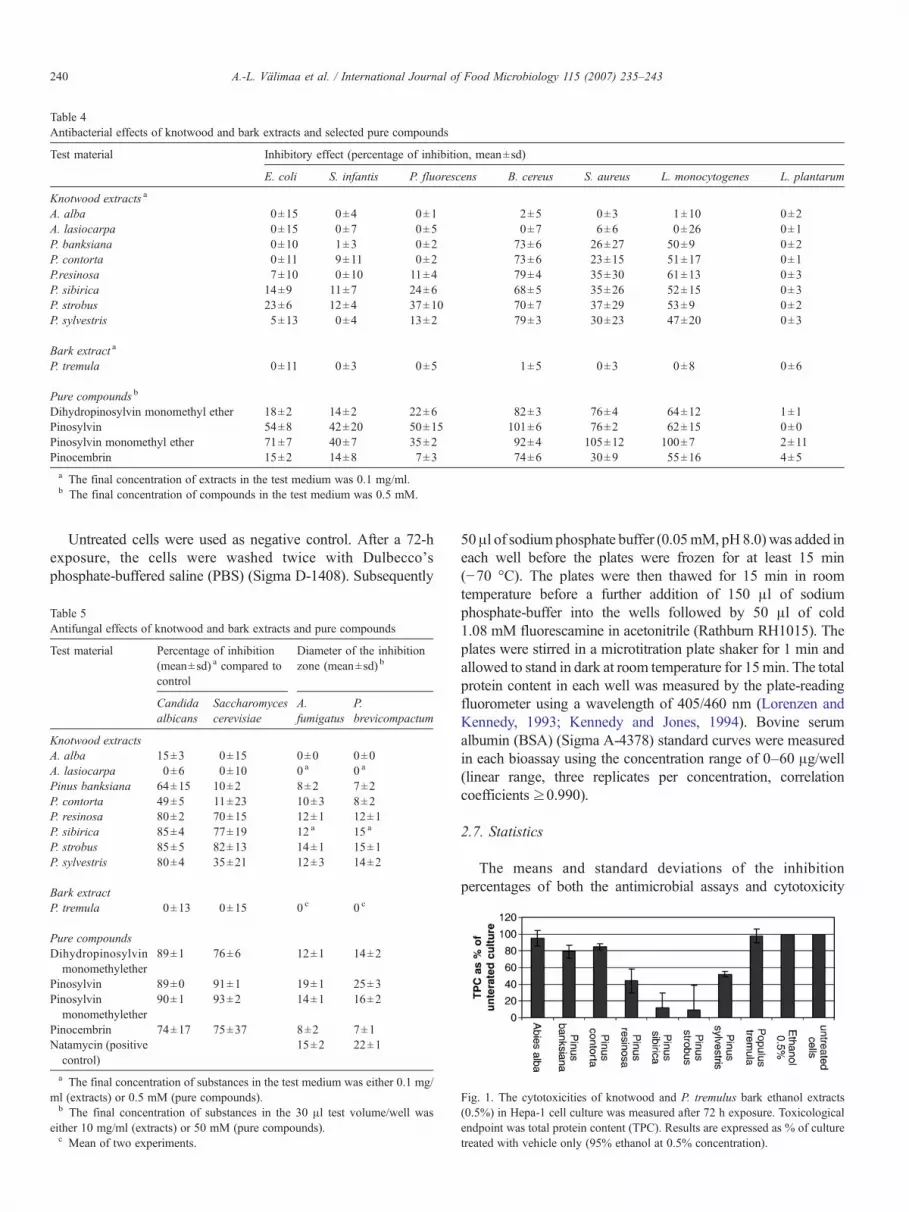
Table 4Antibacterial effects of knotwood and bark extracts and selected pure compounds
Test material Inhibitory effect (percentage of inhibition, mean±sd)
E. coli S. infantis P. fluorescens B. cereus S. aureus L. monocytogenes L. plantarum
Knotwood extracts a
A. alba 0±15 0±4 0±1 2±5 0±3 1±10 0±2A. lasiocarpa 0±15 0±7 0±5 0±7 6±6 0±26 0±1P. banksiana 0±10 1±3 0±2 73±6 26±27 50±9 0±2P. contorta 0±11 9±11 0±2 73±6 23±15 51±17 0±1P.resinosa 7±10 0±10 11±4 79±4 35±30 61±13 0±3P. sibirica 14±9 11±7 24±6 68±5 35±26 52±15 0±3P. strobus 23±6 12±4 37±10 70±7 37±29 53±9 0±2P. sylvestris 5±13 0±4 13±2 79±3 30±23 47±20 0±3
Bark extract a
P. tremula 0±11 0±3 0±5 1±5 0±3 0±8 0±6
Pure compounds b
Dihydropinosylvin monomethyl ether 18±2 14±2 22±6 82±3 76±4 64±12 1±1Pinosylvin 54±8 42±20 50±15 101±6 76±2 62±15 0±0Pinosylvin monomethyl ether 71±7 40±7 35±2 92±4 105±12 100±7 2±11Pinocembrin 15±2 14±8 7±3 74±6 30±9 55±16 4±5a The final concentration of extracts in the test medium was 0.1 mg/ml.b The final concentration of compounds in the test medium was 0.5 mM.
240 A.-L. Välimaa et al. / International Journal of Food Microbiology 115 (2007) 235–243
Untreated cells were used as negative control. After a 72-hexposure, the cells were washed twice with Dulbecco'sphosphate-buffered saline (PBS) (Sigma D-1408). Subsequently
Table 5Antifungal effects of knotwood and bark extracts and pure compounds
Test material Percentage of inhibition(mean±sd) a compared tocontrol
Diameter of the inhibitionzone (mean±sd) b
Candidaalbicans
Saccharomycescerevisiae
A.fumigatus
P.brevicompactum
Knotwood extractsA. alba 15±3 0±15 0±0 0±0A. lasiocarpa 0±6 0±10 0 a 0 a
Pinus banksiana 64±15 10±2 8±2 7±2P. contorta 49±5 11±23 10±3 8±2P. resinosa 80±2 70±15 12±1 12±1P. sibirica 85±4 77±19 12 a 15 a
P. strobus 85±5 82±13 14±1 15±1P. sylvestris 80±4 35±21 12±3 14±2
Bark extractP. tremula 0±13 0±15 0 c 0 c
Pure compoundsDihydropinosylvinmonomethylether
89±1 76±6 12±1 14±2
Pinosylvin 89±0 91±1 19±1 25±3Pinosylvinmonomethylether
90±1 93±2 14±1 16±2
Pinocembrin 74±17 75±37 8±2 7±1Natamycin (positivecontrol)
15±2 22±1
a The final concentration of substances in the test medium was either 0.1 mg/ml (extracts) or 0.5 mM (pure compounds).b The final concentration of substances in the 30 μl test volume/well was
either 10 mg/ml (extracts) or 50 mM (pure compounds).c Mean of two experiments.
50μl of sodiumphosphate buffer (0.05mM, pH8.0)was added ineach well before the plates were frozen for at least 15 min(−70 °C). The plates were then thawed for 15 min in roomtemperature before a further addition of 150 μl of sodiumphosphate-buffer into the wells followed by 50 μl of cold1.08 mM fluorescamine in acetonitrile (Rathburn RH1015). Theplates were stirred in a microtitration plate shaker for 1 min andallowed to stand in dark at room temperature for 15min. The totalprotein content in each well was measured by the plate-readingfluorometer using a wavelength of 405/460 nm (Lorenzen andKennedy, 1993; Kennedy and Jones, 1994). Bovine serumalbumin (BSA) (Sigma A-4378) standard curves were measuredin each bioassay using the concentration range of 0–60 μg/well(linear range, three replicates per concentration, correlationcoefficients ≥0.990).
2.7. Statistics
The means and standard deviations of the inhibitionpercentages of both the antimicrobial assays and cytotoxicity
Fig. 1. The cytotoxicities of knotwood and P. tremulus bark ethanol extracts(0.5%) in Hepa-1 cell culture was measured after 72 h exposure. Toxicologicalendpoint was total protein content (TPC). Results are expressed as % of culturetreated with vehicle only (95% ethanol at 0.5% concentration).
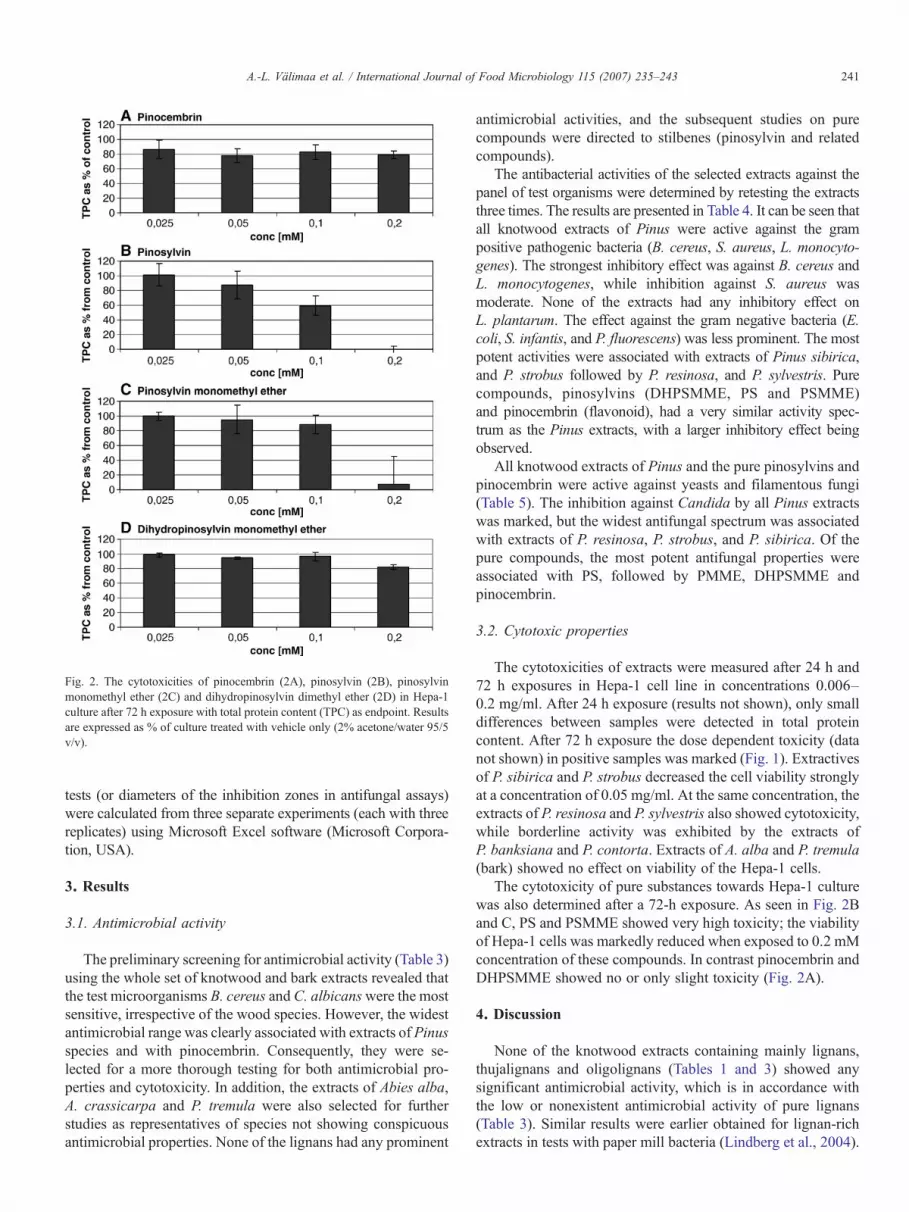
Fig. 2. The cytotoxicities of pinocembrin (2A), pinosylvin (2B), pinosylvinmonomethyl ether (2C) and dihydropinosylvin dimethyl ether (2D) in Hepa-1culture after 72 h exposure with total protein content (TPC) as endpoint. Resultsare expressed as % of culture treated with vehicle only (2% acetone/water 95/5v/v).
241A.-L. Välimaa et al. / International Journal of Food Microbiology 115 (2007) 235–243
tests (or diameters of the inhibition zones in antifungal assays)were calculated from three separate experiments (each with threereplicates) using Microsoft Excel software (Microsoft Corpora-tion, USA).
3. Results
3.1. Antimicrobial activity
The preliminary screening for antimicrobial activity (Table 3)using the whole set of knotwood and bark extracts revealed thatthe test microorganisms B. cereus and C. albicanswere the mostsensitive, irrespective of the wood species. However, the widestantimicrobial range was clearly associated with extracts of Pinusspecies and with pinocembrin. Consequently, they were se-lected for a more thorough testing for both antimicrobial pro-perties and cytotoxicity. In addition, the extracts of Abies alba,A. crassicarpa and P. tremula were also selected for furtherstudies as representatives of species not showing conspicuousantimicrobial properties. None of the lignans had any prominent
antimicrobial activities, and the subsequent studies on purecompounds were directed to stilbenes (pinosylvin and relatedcompounds).
The antibacterial activities of the selected extracts against thepanel of test organisms were determined by retesting the extractsthree times. The results are presented in Table 4. It can be seen thatall knotwood extracts of Pinus were active against the grampositive pathogenic bacteria (B. cereus, S. aureus, L. monocyto-genes). The strongest inhibitory effect was against B. cereus andL. monocytogenes, while inhibition against S. aureus wasmoderate. None of the extracts had any inhibitory effect onL. plantarum. The effect against the gram negative bacteria (E.coli, S. infantis, and P. fluorescens) was less prominent. The mostpotent activities were associated with extracts of Pinus sibirica,and P. strobus followed by P. resinosa, and P. sylvestris. Purecompounds, pinosylvins (DHPSMME, PS and PSMME)and pinocembrin (flavonoid), had a very similar activity spec-trum as the Pinus extracts, with a larger inhibitory effect beingobserved.
All knotwood extracts of Pinus and the pure pinosylvins andpinocembrin were active against yeasts and filamentous fungi(Table 5). The inhibition against Candida by all Pinus extractswas marked, but the widest antifungal spectrum was associatedwith extracts of P. resinosa, P. strobus, and P. sibirica. Of thepure compounds, the most potent antifungal properties wereassociated with PS, followed by PMME, DHPSMME andpinocembrin.
3.2. Cytotoxic properties
The cytotoxicities of extracts were measured after 24 h and72 h exposures in Hepa-1 cell line in concentrations 0.006–0.2 mg/ml. After 24 h exposure (results not shown), only smalldifferences between samples were detected in total proteincontent. After 72 h exposure the dose dependent toxicity (datanot shown) in positive samples was marked (Fig. 1). Extractivesof P. sibirica and P. strobus decreased the cell viability stronglyat a concentration of 0.05 mg/ml. At the same concentration, theextracts of P. resinosa and P. sylvestris also showed cytotoxicity,while borderline activity was exhibited by the extracts ofP. banksiana and P. contorta. Extracts of A. alba and P. tremula(bark) showed no effect on viability of the Hepa-1 cells.
The cytotoxicity of pure substances towards Hepa-1 culturewas also determined after a 72-h exposure. As seen in Fig. 2Band C, PS and PSMME showed very high toxicity; the viabilityof Hepa-1 cells was markedly reduced when exposed to 0.2 mMconcentration of these compounds. In contrast pinocembrin andDHPSMME showed no or only slight toxicity (Fig. 2A).
4. Discussion
None of the knotwood extracts containing mainly lignans,thujalignans and oligolignans (Tables 1 and 3) showed anysignificant antimicrobial activity, which is in accordance withthe low or nonexistent antimicrobial activity of pure lignans(Table 3). Similar results were earlier obtained for lignan-richextracts in tests with paper mill bacteria (Lindberg et al., 2004).

242 A.-L. Välimaa et al. / International Journal of Food Microbiology 115 (2007) 235–243
Lignans and oligolignans are strong antioxidants and radicalscavengers (Willför et al., 2003c; Pietarinen et al., 2006; Eklundet al., 2005), but these mechanisms are obviously not directlyassociated with antimicrobial activity.
Knotwood extracts containing substantial amounts offlavonoids also did not exhibit any significant antimicrobialactivity. Against this background the relatively strong antifun-gal activity of the flavonoid pinocembrin is exceptional, and themechanism of action remains to be solved. It is noteworthy thatpinocembrin had no marked cytotoxic effects on Hepa-1 cellline, in contrast to pinosylvins.
The knotwood extract of Eucalyptus globulus showed someantimicrobial activity, which probably can be attributed to thepresence of tannins and tannin precursors (ellagic and tannicacids), which are known to have antimicrobial properties(Shimizu et al., 2002; Puupponen-Pimiä et al., 2005). However,among the bark extracts only the P. maritima bark extractshowed any broader antimicrobial activity despite the observa-tion that all bark extracts contained tannins.
The antimicrobial and cytotoxic activity of the differentPinus knotwood extracts is probably explained by the pre-sence of pinosylvins, which were most active when tested aspure compounds, and which are commonly found in knot-wood extracts of the Pinus family (Table 1). For example thetotal concentration of PS and PSMME in P. strobus,P. resinosa and P. sylvestris knotwood extracts was approx-imately 38% (Willför et al., 2003a,b,c), and 28% in extract ofP. sibirica (Pietarinen et al., 2006). The results with woodextracts had a relatively high variation in comparison to resultsobtained with pure compounds. This is probably due to chanceeffects, when substances are present at concentrations neartheir effective dose. The further testing of the antibacterial andantifungal properties of purified pinosylvins (Tables 3 and 4)corroborated the initial conclusions on the effects ofpinosylvin-type compounds. Despite some variation betweenmicrobial strains and tests, it is clear that PS and PSMME hadthe highest antimicrobial activities of the pure compounds,closely followed by DHPSMME and pinocembrin. The resultsare in accordance with various earlier reports on antibacterialand antifungal properties of different stilbenes (Lindberg et al.,2004; Nitta et al., 2002; Ali and Debnath, 1997; Frykholm,1945). Pinosylvins in particular have been shown to exhibitantifungal activities (Rennerfelt, 1943; Erdtman and Renner-felt, 1944; Bois et al., 1999; Celimene et al., 1999).
The antimicrobial range of pinosylvins and Pinus extractsappears wider than that of the plant or berry extracts (Rauhaet al., 2000; Puupponen-Pimiä et al., 2005). Particularly theaction against Listeria is noteworthy, since this activity has notbeen previously reported with phenolic plant extracts. The factthat even pure pinosylvins did not inhibit the growth ofL. plantarum provokes the question, whether lactic acid bacteriaare generally tolerant to these compounds, and whether they canbe utilised in controlling food-related fermentations.
The antimicrobial properties of pinosylvins correlated veryclosely with their cytotoxicity (Tables 3–5, Figs. 1 and 2). Thissuggests a common mechanism of action for these properties, inwhich two hydroxyl groups in meta position in one of the
aromatic rings in the stilbene-structure may play an importantrole. In previous studies, stilbenes with the 3,5-dimethoxy motifhas been shown to be biologically even more active thancorresponding 3,5-dihydroxy compounds (Tolomeo et al.,2005). The double bond in the carbon chain between the twoaromatic rings also seems to play a role, since DHPSMMEshowed lower toxicity than PSMME. This may be due to thelost electron transfer capability between the two aromatic ringsin DHPSMME compared to PSMME. However, more in-depthstudies should be conducted on this matter in the future.
Regarding the potential food applications both safety andefficacy at concentrations that can be applied are crucial.Stilbenes occur naturally in many fruits and berries traditionallyused as human food, resveratrol containing grapes being themost well known example. However, also the berries of thegenus Vaccinium, such as lingonberry (V. vitis-idae) andbilberry (V. myrtillus) have been found in an American–Canadian study to contain considerable amounts (768 ng/g and5884 ng/g, respectively) of resveratrol, the content inlingonberries approaching that of grapes (Rimando et al.,2004). Although the history of apparent safe use is not anabsolute proof of safety, it is an indication of relativeharmlessness.
The stilbene concentrations used in this study wereapproximately 50 times higher than those occurring naturallyin food products, and it could be questioned, whether theobserved antimicrobial effects are strong enough to have anypractical relevance. However, also the microbial challenge inthe experimental settings was more severe than could be usuallyexpected in foods, and the effective concentrations andtreatment conditions should be tested with actual foodstuffs.As an example of potential applications Montero et al. (2003)showed that a submersion for a few seconds in 0.16 mM watersolution of resveratrol was effective in the prevention of postharvest fungal spoilage of grapes.
Since the antimicrobial action mechanisms of wood extractsor the wood associated stilbenes are still unknown, the potentialof resistance development among spoilage organisms orpathogens cannot be assessed at the moment. The same appliesto possible interactions with different food components and thestabilities of these compounds in different food processes.These are aspects that should be assessed during the eventualdevelopment of practical applications.
5. Conclusions
In conclusion, pure pinosylvin-type compounds and extractsrich in pinosylvins are potent antimicrobial and cytotoxicagents, and in this respect resemble closely a related compound,resveratrol, which has gained attention not only as a potentialnatural antimicrobial but is also thought to have functional andeven therapeutic uses. The use of pure pinosylvins or extractscontaining large amounts of pinosylvins should be tested andevaluated in different applications needing antimicrobialproperties or cytostatic activities. The raw material for purepinosylvins or natural extracts could be obtained from theknotwoods and/or stemwood of Pinus trees.

243A.-L. Välimaa et al. / International Journal of Food Microbiology 115 (2007) 235–243
Acknowledgement
This research was partially supported by the TechnologyDevelopment centre (TEKES). The skillful technical assistanceof Riitta Venäläinen is gratefully acknowledged.
References
Ali, M.A., Debnath, D.C., 1997. Isolation and characterization of antibacterialconstituent from devdaru (lignum of Polyalthia longifolia L.). BangladeshJournal of Scientific and Industrial Research 32, 20–24.
Bergman, J., Lindgren, B.O., Svahn, C.M., 1965. Triterpenes and 4-alpha-methylsterols in birch wood. Acta Chemica Scandinavica 19, 1661–1666.
Bois, E., Lieutier, F., Yart, A., 1999. Bioassays on Leptographium wingfieldii, abark beetle associated fungus, with phenolic compounds of Scots pinephloem. European Journal of Plant Pathology 105, 51–60.
Boissy, P., Andersen, T.L., Abdallah, B.A., Kassem,M., Plesner, T., Delaissé, J.-M.,2005. Resveratrol inhibits myeloma cell growth, prevents osteoclast formationand promotes osteoblast differentiation. Cancer Research 65, 9943–9952.
Celimene, C.C., Micales, J.A., Ferge, L., Young, R.A., 1999. Efficacy ofpinosylvins against white-rot and brown-rot fungi. Holzforschung 53,491–497.
Eklund, P., Lindholm, A., Mikkola, J.-P., Smeds, A., Lehtilä, R., Sjöholm, R.,2003. Synthesis of (−)-matairesinol, (−)-enterolactone, and (−)-enterodiolfrom the natural lignan hydroxymatairesinol. Organic Letters 5, 491–493.
Eklund, P.C., Långvik, O.K., Wärnå, J.P., Salmi, T.O., Willför, S., Sjöholm, R.E.,2005. Chemical studies on antioxidant mechanisms and free radicalscavenging properties of lignans. Organic and Biomolecular Chemistry 3(18), 3336–3347.
Erdtman, H., Rennerfelt, E., 1944. The pinosylvin–phenolic content of pineheartwood, its determination and its antiseptic action towards wood-destroying fungi. Svensk Papperstidning 47, 45–56.
Frykholm, K.O., 1945. Bacteriological studies of pinosylvine and itsmonomethyl and dimethyl ethers, and toxicologic studies of pinosylvine.Nature 155, 454–455.
Hankinson, O., 1979. Single step selection of clones of a mouse hepatoma linedeficient in aryl hydrocarbon hydroxylase. Proceedings of the NationalAcademy of Sciences USA 76, 373–376.
Jang, M., Cai, L., Udeani, G.O., Slowing, K.V., Thomas, C.F., Beecher, C.W.W.,Fong, H.H.S., Farnsworth, N.R., Kinghorn, A.D., Mehta, R.G., Moon, R.C.,Pezzuto, J.M., 1997. Cancer chemopreventive action of resveratrol, a naturalproduct derived from grapes. Science 275, 218–220.
Kennedy, S.W., Jones, P.S., 1994. Simultaneous measurement of cytochromeP4501A catalytic activity and total protein concentration with a fluorescenceplate reader. Analytical Biochemistry 222, 217–223.
Lindberg, L., Willför, S., Hemming, J., Holmbom, B., 2004. Antibacterialeffects of hydrophilic knotwood extracts on papermill bacteria. Journal ofIndustrial Microbiology and Biotechnology 31, 137–147.
Lorenzen, A., Kennedy, S.W., 1993. A Fluorescence based protein assay for usewith a microplate reader. Analytical Biochemistry 214, 346–348.
Malterud, K.E., Bremnes, T.E., Faegri, A., Moe, T., Sandanger Dugstad, E.K.,Anthonsen, T., Henriksen, L.M., 1985. Flavonoids from the wood of Salixcaprea as inhibitors of wood-destroying fungi. Journal of Natural Products40, 559–563.
Man-Ying Chan, M., 2002. Antimicrobial effect of resveratrol on dermatophytesand bacterial pathogens of the skin. Biochemical Pharmacology 63, 99–104.
Mihara, R., Barry, K., Mohammed, C., Mitsunaga, T., 2005. Comparison ofantifungal and antioxidant properties of Acacia mangium and A. auriculi-formis heartwood extracts. Journal of Chemical Ecology 4, 789–804.
Montero, C., Cristescu, S.M., Jiménez, J.B., Orea, J.M., te Lintel Hekkert, S.,Harren, F.J.M., Gonzaléz Ureña, A., 2003. trans-Resveratrol and grapedisease resistance. A dynamical study by high resolution laser-basedtechniques. Plant Physiology 131, 129–138.
Neacsu, M., Eklund, P.C., Sjöholm, R.E., Pietarinen, S.P., Ahotupa, M.O.,Holmbom, B.R., Willför, S.M., in press. Antioxidant flavonoids from
knotwood of Jack pine and European aspen. Holz als Roh und Werkstoff.doi:10.1007/s00107-006-0121-0.
Nitta, T., Arai, T., Takamatsu, H., Inatomi, Y., Murata, H., Iinuma, M., Tanaka,T., Ito, T., Asai, F., Ibrahim, I., Nakanishi, T., Watabe, K., 2002.Antibacterial activity of extracts prepared from tropical and subtropicalplants on methicillin-resistant Staphylococcus aureus. Journal of HealthScience 48, 273–276.
Pausler, M.G., Ayer, W.A., Hirazuka, Y., 1995. Benzoic acid, salicylic acid andthe role of black galls on aspen in protection against decay. Canadian Journalof Forest Research 25, 1479–1483.
Pietarinen, S., Willför, S., Sjöholm, R., Holmbom, B., 2004. Bioactive phenolicsubstances in important tree species. Part 3. Knots and stemwood of Acaciacrassicarpa and A. mangium. Holzforschung 59, 94–101.
Pietarinen, S., Willför, S., Ahotupa, M., Hemming, J., Holmbom, B., 2006.Knotwood and bark extracts: strong antioxidants from waste materials.Journal of Wood Science. doi:10.1007/s10086-005-0780-1 (publishedonline first:).
Pietarinen, S.P., Willför, S.M., Vikström, F.A., Holmbom, B.R., in press. Aspenknots, a rich source of flavonoids. Journal ofWoodChemistry and Technology.
Puupponen-Pimiä, R., Nohynek, L., Hartman-Schmidlin, S., Kähkönen, M.,Heinonen, M., Määttä-Riihinen, K., Oksman-Caldentey, K.-M., 2005. Berryphenolics selectively inhibit the growth of intestinal pathogens. Journal ofApplied Microbiology 98, 991–1000.
Rauha, J.-P., Remes, S., Heinonen, M., Hopia, A., Kähkönen, M., Kujala, T.,Pihljaj, K., Vuorela, H., Vuorela, P., 2000. Antimicrobial effects of Finnishplant extracts containing flavonoids and other phenolic compounds.International Journal of Food Microbiology 56, 3–12.
Rennerfelt, E., 1943. The toxicity of phenolic extractives of pine toward severaldecay-producing fungi. Svensk Botaniska Tidskrift 83–92.
Rimando, A.M., Kalt, W., Magee, J.B., Devey, J., Ballington, J.R., 2004.Resveratrol, pterostilbene and piceatannol in Vaccinium berries. Journal ofAgricultural and Food Chemistry 52, 4713–4719.
Shimizu, K., Kondo, R., Sakai, K., 2002. Antioxidant activity of heartwoodextracts of Papua New Guinean woods. Journal of Wood Science 48,446–450.
Stojanovic, S., Brede, O., 2002. Elementary reactions of the antioxidant actionof trans-stilbene derivatives: resveratrol, pinosylvin and 4-hydroxystilbene.Physical Chemistry Chemical Physics 4, 757–764.
Tolomeo, M., Grimaudo, S., Di Critina, A., Roberti, M., Pizzirani, D., Meli, M.,Dusconchet, L., Gebbia, N., Abbadessa, V., Crosta, L., Barucchello, R.,Grisolia, G., Invidiata, F., Simoni, D., 2005. Pterostilbene and 3′-hydroxypterostilbene are effective apoptosis-inducing agents in MDR andBCR-ABL-expressing leukemia cells. International Journal of Biochemistryand Cell Biology 37, 1709–1726.
Willför, S., Hemming, J., Reunanen, M., Holmbom, B., 2003a. Phenolic andlipophilic extractives in Scots pine knots and stemwood. Holzforschung 57,359–372.
Willför, S., Hemming, J., Reunanen, M., Eckerman, C., Holmbom, B., 2003b.Lignans and lipophilic extractives in Norway spruce knots and stemwood.Holzforschung 57, 27–36.
Willför, S.M., Ahotupa, M.O., Hemming, J.E., Reunanen, M.H., Eklund, P.C.,Sjöholm, R.E., Eckerman, C.S., Pohjamo, S.P., Holmbom, B.R., 2003c.Antioxidant activity of knotwood extractives and phenolic compounds ofselected tree species. Journal of Agricultural and Food Chemistry 51,7600–7606.
Willför, S., Nisula, L., Hemming, J., Reunanen, M., Holmbom, B., 2004a.Bioactive phenolic substances in important tree species. Part 2. Knots andstemwood of fir species. Holzforschung 58, 650–659.
Willför, S., Nisula, L., Hemming, J., Reunanen, M., Holmbom, B., 2004b.Bioactive phenolic substances in industrially important tree species. Part 1.Knots and stemwood of different spruce species. Holzforschung 58, 335–344.
Willför, S., Reunanen, M., Eklund, P., Sjöholm, R., Kronberg, L., Fardim, P.,Pietarinen, S., Holmbom, B., 2004c. Oligolignans in Norway spruce and Scotspine knots and Norway spruce stemwood. Holzforschung 58, 345–354.




















