
Erythropoietin directly stimulates osteoclast precursors and induces bone loss
Upload
independentCategory
view
3download
0
524
Angiotensin II Increases SpontaneousContractile Frequency and Stimulates Calcium
Current in Cultured Neonatal Rat HeartMyocytes: Insights into the Underlying
Biochemical Mechanisms
Ira S. Allen, Neri M. Cohen, Ravinder S. Dhallan,
Shirley T. Gaa, W.J. Lederer, and Terry B. Rogers
The effect of angiotensin II on cultured neonatal rat heart myocytes was studied by measuring changesin cell length, the magnitude and kinetics of the calcium current, and changes in cyclic adenosine3',5'-monophosphate (cAMP) and phosphoinositide metabolism. Spontaneous beating frequency ofmulticellular networks was increased by angiotensin II with a maximal increase of 100% above controlvalues at concentrations of 5 nM or greater. The half-maximal response occurred at 0.6 nM angiotensinII. Shortening amplitude, shortening velocity, and relaxation velocity decreased concomitantly withthe increasing contractile rate. In voltage-clamped single myocytes, both steady-state and transientcomponents of the calcium current were increased by the addition of angiotensin II. Angiotensin IIhad no effect on either control or isoproterenol-stimulated adenylate cyclase activity in myocytemembranes. Neither the basal levels nor the isoproterenol-stimulated cAMP accumulation in intactcells was affected by addition of hormone. In myocytes labeled with [3H]inositol, angiotensin IIstimulated the formation of [3H]inositol phosphates. One minute after addition of 5 nM angiotensinII, inositol monophosphate and inositol bisphosphate levels were increased to 73% and 99%,respectively, above control values and remained elevated at 10 minutes. Inositol trisphosphate levelswere not significantly different from control values at either time point. Nifedipine (10 (JLM) had noeffect on angiotensin II-induced increases in [3H]inositol phosphates. We conclude that the increasesin both spontaneous beating rate and calcium current in angiotensin II-stimulated cultured neonatalheart cells are not dependent on cAMP or inositol trisphosphate levels but may involve sustainedphosphoinositide hydrolysis. (Circulation Research 1988; 62:524-534)
Angiotensin II (All) is an important vasoactivepeptide that regulates cellular responses in anumber of tissues.1-2 While the heart has not
traditionally been considered a target tissue for All,this hormone has been reported to increase the strengthof myocardial contraction and modify electrical be-havior in several species.3"5 Although the physiologicalmechanisms by which All brings about its effectsremain uncertain, several important findings have beenreported. 1) There are specific, high-affinity receptorsfor All on both cul tured neonatal rat heart myocytes andsarcolemmal membranes from bovine and rabbitventricle.6"9 2) All increases the slow inward current(Isi) in rabbit papillary muscle, perfused embryonic
From the Departments of Biological Chemistry (I.S.A., R.S.D.,S.T.G., and T.B.R.) and Physiology (N.M.C. and W.J.L.),University of Maryland School of Medicine, Baltimore, Maryland.
This work has been supported in part by United States PublicHealth Service grants HL 28138 and HL 25675, and a researchfellowship award to I.S.A. (HL 07155). R.S.D. is a recipient of anAmerican Heart Association Medical Student Research Fellowship.T.B.R. is a recipient of a Research Career Development Award.
Address for reprints: Dr. Terry B. Rogers, Department ofBiological Chemistry, Howard Hall Room 205, University ofMaryland School of Medicine, Baltimore, MD 21201.
Received April 16, 1987; accepted September 16, 1987.
chick heart, and bovine Purkinje fibers.4-5 3) There isan increase in twitch tension associated with theapplication of All to rabbit papillary muscle and bovinePurkinje fibers.4-5
There is little information on the molecular mech-anisms that couple All receptor binding to changes inboth Isi and contractile behavior in cardiac cells. Incontrast, the calcium mobilizing effects of All innoncardiac tissues are well documented.2 All has beenshown both to inhibit the adenylate cyclase system inthe liver,10 anterior pituitary,11 aorta,12 and adrenalcortex,13 and to stimulate phosphoinositide hydrolysisin the same tissues.14"17
In the present report, the cultured neonatal rat heartmyocyte system was utilized to study the actions ofAIL We report here that All stimulates both contractilefrequency in spontaneously beating cultured myocytesand sarcolemmal calcium channel current (ICa) involtage clamped myocytes. Further, the All-mediatedchanges in ICa and contractile behavior occur througha cyclic adenosine 3',5'-monophosphate (cAMP)-independent mechanism. Finally, we document thatAll stimulates phosphoinositide hydrolysis in cardiacmyocytes, providing new insights into the mechanismof action of All in heart cells.
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from

Allen et al Angiotensin II Stimulation of Cultured Heart Cells 525
Materials and MethodsPreparation of Cultured Rat Heart Myocytes
Cardiac myocytes were isolated from hearts of1-2-day-old Sprague-Dawley rats by a trypsin disag-gregation procedure and cultured as previouslydescribed.6 For most of the experiments described here,cells were grown in either 24-well or 4-well (1.7 cm)tissue culture dishes. The electrophysiological exper-iments were performed on myocytes grown on plasticcoverslips in 35-mm culture dishes.
Preparation of Membranes From Myocyte CulturesA crude microsomal preparation was prepared from
the cultured myocytes for use in the adenylate cyclaseassays. Confluent monolayers of cells in 10-cm culturedishes were harvested on days 4-6 . The dishes wereplaced on ice, and the growth medium was removed.Ten milliliters of ice-cold buffer A (10 mM Tris and250 mM sucrose, pH 7.4) was added to the dish,and the cells were detached by scraping with arubber policeman. The cell suspension was centrifugedat 300g for 10 minutes at 0° C. The cell pellet wasresuspended in 10 ml of buffer A, homogenized in aTeflon glass homogenizer at 0° C, and centrifuged at120,000g for 30 minutes. This total particulate frac-tion was washed with 10 ml of buffer B [50 mM 4-(2-hydroxyethyl)-l-piperazineethanesulfonicacid-Tris(HEPES-Tris), pH 7.4], rehomogenized, and centri-fuged at 120,000g for 30 minutes at 0° C. The pelletwas resuspended in buffer B at an approximate con-centration of 1-2 mg protein/ml. The membranes wererapidly frozen on dry ice and stored at —70° C.Adenylate cyclase activity was stable for at least 2weeks. Protein was determined by the method ofBradford,18 using bovine serum albumin (BSA) as astandard.
Measurement of Contractile ResponsesThe contractile responses of a spontaneously beating
multicellular network were measured using an opticalvideo dimension analysis system as previouslydescribed.6 Shortening and relaxation velocities wereobtained electronically by differentiating the lengthsignal. The output from the video dimension analyzerwas filtered through an eight-pole, low-band pass,active filter with a cutoff frequency of 25 Hz, and thendifferentiated. The signal was calibrated in microns persecond. When the cells were electrically driven,cultures were field-stimulated with pulses of 1-msecduration at 50 volts. The entire culture dish wasmounted on the microscope stage, and a single well(1.7 cm) was superfused with control media (Dul-becco's Modified Eagles Medium [DMEM], 25 mMHEPES, 0.25% BSA, pH 7.4) in a volume of 200 |JL1.The pH was maintained by gassing the bathing mediumwith 95% O2-5% CO2. The temperature was maintainedat 35° C with a variation of less than 0.2° C within eachexperiment. All cultures used in these experiments hada stable beating rate of 45-55 beats/min at 35° C.Superfusion was switched from control media tocontrol media containing All (Peninsula Laboratories,
Belmont, California) or Sar',Ala8-AII (Peninsula Lab-oratories) as indicated. In the dose-response studies,each culture well was exposed to a single concentrationof hormone and then discarded.
Measurement of Calcium Channel CurrentA plastic coverslip with adherent cells was removed
from the culture dish, transferred to the experimentalchamber, and superfused at 1 ml/min with a modifiedTyrode's solution (mM: 145 NaCl, 4 KC1, 1 CaCl7, 1MgCl2, 10 glucose, and 10 HEPES, pH 7.36) con-taining 10 |xM tetrodotoxin (TTX) (Sigma, St. Louis,Missouri). Patch electrodes (BB-CH Mechanex, Gen-eva, Switzerland) were pulled from borosilicate glasscapillaries (1.5 mm outside diameter, World PrecisionInstruments, New Haven, Connecticut) to a tip diam-eter of about 1 (xm. Their resistance was 2-4 MO whenfilled with (mM) 140 Cs-glutamate, 1 NaCl, 0.01EGTA, and 10 HEPES. Voltage errors introduced bypipette series resistance (measured as the time constantof the uncompensated capacity transient divided by thecell capacity [43 ± 6.7 pF]) were compensated by usingdynamic series resistance control compensation toprovide the fastest possible capacity transient withoutringing. The series resistance of the pipette attached toa cell with a ruptured patch was generally twice that ofthe pipette alone. After compensation, there usuallyremained an additional 2-3 MCI of uncompensatedseries resistance. Experiments were performed in the"whole-cell" voltage clamp configuration at 22° C.19
With sodium and potassium channels blocked byexternal TTX and internal cesium, respectively, volt-age clamp pulses (feedback resistor 1 GH; 8900,Dagan, Minneapolis, Minnesota) were applied toinvestigate ICa. Cadmium (0.1 mM) was added be-fore the end of each experiment, and ICa was measuredas the cadmium-sensitive current. Data were storedon videotape (VCR7200, Sanyo, Tokyo, Japan) afterdigitization with a digital audio processor (PCM-501ES, Sony, Park Ridge, New Jersey) mod-ified after Bezanilla20 by Unitrade, Philadelphia,Pennsylvania. Relatively high doses of All were ap-plied to the cells, 100 nM, since BSA had to beeliminated from the medium because it interferedwith the formation of electrode-cell seal. Previousstudies have shown that it is essential to have BSA inthe medium to prevent proteolysis of the hormone inthese cultures6; therefore, it was not possible toaccurately control the effective concentrations of Allin these experiments, but it is certainly lower than100 nM.
Curve Fitting ProcedureThe current records were analyzed off-line using
VACUUM Mkll (M.B.C. Systems, Mount Washing-ton Station, Baltimore, Maryland) software and DataTranslation hardware (DT 2805, Marlborough, Mas-sachusetts). Current records were fit by a nonlinearleast squares method to a single exponential function.21
The amplitude of ICa was estimated at t = 0.The data for the steady-state activation (dx) and
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from

526 Circulation Research Vol 62, No 3, March 1988
inactivation (fx) parameters were fit to the followingBoltzmann equations:
where V is the membrane potential and Vh and k areconstants.
Adenylate Cyclase AssayAdenylate cyclase activity in membranes from
cultured myocytes was determined according to themethod of Salomon et al.22 Briefly, reaction mixtures(final volume 0.1 ml) contained [a-^PJATP (40-70cpm/pmol; ICN Radiochemicals, Irvine, California), 1mM ATP, 1 mM cAMP, 0.25% BSA, 1 mM isobutyl-methylxanthine, 20 mM creatine phosphate, creatinephosphokinase (100 units/ml), adenosine deaminase(3.75 units/ml), myocyte membranes (40-60 \ig), 0.1mM GTP, 80 mM HEPES-Tris (pH 7.4) and MgCl2,0.1mM isoproterenol, and 100 nM All as indicated. Thereaction was initiated by the addition of membranesand allowed to proceed at 35° C for 10 minutes. Thereaction was terminated by adding 100 (xl of stoppingsolution (2% sodium dodecyl sulfate, 40 mM ATP, 1.4mM cAMP, pH 7.4). After addition of [3H]cAMP(about 5,000 cpm/tube to monitor cAMP recovery; ICNRadiochemicals), [32P]cAMP was purified by sequen-tial chromatography on Dowex (Bio-Rad Laboratories,Rockville Center, New York) and alumina columns.Free Mg2* concentrations were calculated by solvingfor multiple equilibria.23
cAMP DeterminationMyocytes, in 1 ml DMEM containing 25 mM
HEPES and 0.25% BSA, pH 7.4, were exposed to All(100 nM) for 5 minutes at 37° C in 95% O2-5% CO2.Isoproterenol (1 p,M) was then added, and the cellswere incubated for an additional 1 minute. Time coursestudies revealed that isoproterenol-stimulated cAMPlevels were maximal by this time. Reactions wereterminated by rapidly aspirating the incubation me-dium and adding 0.5 ml of ice-cold perchloric acid(6%). The acid extracts were neutralized with 40 p.1 of5 M K2CO3. cAMP in the neutral extracts was measuredusing a standard radioimmunoassay procedure (cAMPRadioimmunoassay Kit, New England Nuclear, Bos-ton, Massachusetts).24
Pilot experiments using [3H]cAMP revealed that90 ± 5% of the cAMP was recovered in the neutralizedextracts. For the results reported here, the data wereanalyzed assuming 90% recovery. Protein in experi-mental wells was taken as the mean of the proteindeterminations in three nonassayed wells on the sameculture dish, with a variation of less than 10%. Proteinwas extracted in 0.0125% sodium dodecyl sulfate andmeasured according to the method of Bradford,18 usingBSA as a standard.
Measurement of Inositol PhosphatesMyocytes were grown for at least 4 days and then
incubated an additional 48 hours in DMEM containing
10% fetal calf serum, 1% penicillin, 1% streptomycin,[3H]inositol (20 ^Ci/ml; New England Nuclear), pH7.4. The wells were washed several times with 1 ml ofassay medium (DMEM containing 25 mM HEPES,0.25% BSA, pH 7.4, gassed with 95% O2-5% CO2).Myocytes, in 1 ml of assay medium, were exposed to5 nM All for the indicated times. Control wells weretreated in an identical fashion except that hormone wasnot added. In experiments where nifedipine (10 \LM)was added, cells were incubated with the calciumchannel blocker for 1 minute prior to addition of 5 nMAll. Reactions were terminated by addition of per-chloric acid and extracts were neutralized as describedabove. [3H]Inositol phosphates in the neutral extractswere separated, collected, and counted according to themethod of Downes and Wusteman.25 Statistical signif-icance was determined by analysis of variance(ANOVA).
ResultsEffect of Angiotensin II on MyocyteContractile Behavior
In the initial phase of this study, the effect of All onthe contractile behavior of cultured neonatal rat heartmyocytes was characterized. As shown in Figure 1, Allincreases the spontaneous beating rate and decreasesshortening amplitude. All (5 nM) increases beatingfrequency in a time-dependent fashion and maximalstimulation is achieved approximately 3 minutes afteraddition of hormone (Figure 1A). There is also adecrease in shortening amplitude that develops overtime after cells are exposed to 5 nM All (Figure 1A).The average maximal increase in rate from a numberof experiments is 99 ± 4% above control values (n = 3).The average maximal decrease in amplitude is 49 ± 3%below control values (n = 3). As another measure ofcontractile force, the effect of All on shortening andrelaxation velocities was determined. In response to 5nM All, both shortening velocity and relaxationvelocity decrease over time, reaching a plateau byapproximately 3 minutes (Figure 1A). The averageAll-stimulated maximal decrease in shortening veloc-ity is 46 ± 9% below control values (n = 3). The averagemaximal decrease in relaxation velocity is 49 ± 2 %below control values (n = 3).
The potency of All in stimulating contractile fre-quency is shown in Figure 2. All maximally stimulatesbeating frequency to 100% above control values atconcentrations of 5 nM or greater. The half-maximalchronotropic response occurs at a concentration of 0.6nM All (ECj,,). This response is specific because the Allantagonist Sar',Ala8-AII (40 nM) completely inhibitsthe increase in beating frequency evoked by 5 nM AILFurther, the All response is decreased in the presenceof 1 nM Sar',Ala8-AII at all concentrations examined.
It is possible that the All-mediated decreases in bothshortening velocity and shortening amplitude arefunctions of the increase in contractile frequencyevoked by AIL This issue was addressed by examiningthe force-frequency relation using myocytes that wereelectrically stimulated. Within the frequency range of
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from

Allen et al Angiotensin II Stimulation of Cultured Heart Cells 527
S nM ANGIOTENSIN B
1 M1N
Ba
LJJQ
<
OOw
unit
FIGURE 1. Effect of angiotensin II (AH) on contractile behaviorof cultured neonatal rat heart cells. A: Time courses for effectof AH on myocyte spontaneous beating rate, shortening am-plitude, shortening velocity, and relaxation velocity. In thisrepresentative experiment, superfusion was switched fromcontrol media to media containing 5 nM All for the periodindicated by the solid bar. Each point represents the averagevalue over a 20-second interval. B: Sample records of short-ening amplitude and shortening velocity taken at time (a) and(b) in Panel A.
the All response (55-130 beats/min), both shorteningamplitude and shortening velocity remain relativelyconstant in the absence of All (Figure 3A). Althoughslight decreases in amplitude and contraction velocityare observed when frequency is increased from 50 to120 beats/min, these results reveal that the reduction inthe amplitude and contraction velocity evoked by Allcannot be the result of increases in frequency alone.Further, in the presence of 1 nM AH, the force-frequency curves are shifted downward. Finally, whenmyocytes are electrically driven at a constant beatingrate (160 beats/min), 5 nM All decreases both short-ening amplitude and shortening velocity (Figure 3B).
To further examine force-frequency relations in thesecultured cardiac cells, isoproterenol was used toincrease beating frequency. Isoproterenol (10 nM)increased beating frequency by 30% in 3 minutes.However, in contrast to the responses of the cells to All,isoproterenol resulted in increases in twitch amplitudeand contraction velocity, by 41% and 78%, respec-tively (data not shown). These expected positiveinotropic responses of the cells to a p-agonist furtherunderscore the novel negative inotropic effects of Allin this system.
Effect of Angiotensin II on Calcium Channel CurrentOne explanation for the increase in beating rate
observed with All is that 1^ is elevated by the hormone.Accordingly, 1^ was recorded under control conditionsand in the presence of 100 nM All with Ca2+ (1 mM)as the principal charge carrier. To examine only the roleof "L-type" calcium channels in whole-cell currentrecords, the holding potential was set to - 5 0 mV.26 Ifany "T-type" channels were present in this preparation,they would be inactivated by this holding potential.Depolarizing pulses of 200-msec duration to potentialsbetween - 4 5 and +60 mV were applied at 0.5 Hz.Five features of Ic, were noted (Figure 4): 1) Asteady-state current is observed following activationand inactivation. 2) Both the transient and steady-statecomponents of 1^ show a bell-shaped dependence onvoltage. Ic increases from zero at - 4 0 mV to amaximum at around 0 mV and then decreases as themembrane potential approaches +55 mV. 3) Thetime-dependent component of l^ reverses direction at+ 55 mV. 4) Over the voltage range 0 to +55 mV, therate of inactivation decreases with increasing depolar-
1nM Sar\ Ala* All
40nM Sar'.AIa'AD
10 9 8
Hog [ ANGtOTENSIN II ]
FIGURE 2. Dose-response relation of the angiotensin II ^In-dependent increase in spontaneous beating rate of myocytes.Superfusion was switched from control media to media con-taining indicated concentration of All (•). Each well was usedfor a single concentration of hormone and then discarded. Inexperiments with Sar',Ala'-AII, media containing AH included1 nM Sar',Ala'-AII (o) or cells were superfused for 1 minute withmedia containing 40 nM antagonist prior to switching to mediacontaining 5 nM All and 40 nM antagonist (A). Each pointrepresents average maximal percent increase over control ofthree separate determinations (mean ± SEM).
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from

528 Circulation Research Vol 62, No 3, March 1988
UJQ
tE 2 -
2
11-
7-1
CONTROL
O 50 125
FREQUENCY (beats/min)
200
BUJQ
a!<
IUJ
Control 5nM Al l -2 mln 5nM AII-3 mln
1 jjm
15 sec
15pm/sac
FIGURE 3. Effect of myocyte beating rate on control andangiotensin II (All)-stimulated shortening amplitude and short-ening velocity. A: Time courses of shortening amplitude andshortening velocity for electrically driven cells. In this repre-sentative experiment of three, cells were electrically stimulatedto indicated frequencies with pulses of 1-msec duration at 50volts. Contractions were recorded before (•) and after (o)switching superfusion from control media to media containing1 nMAlI. Each point represents average value over a 20-secondinterval. B: Sample records of shortening amplitude andshortening velocity in cells electrically driven to a constantbeating rate. Myocytes were stimulated with pulses of 1-msecduration at 50 volts to a constant rate of 160 beats/min.
ization. 5) Both the transient and steady-state compo-nents of 4 , are abolished by 0.1 mM Cd.
All modifies several characteristics of i^. Thepeptide (100 nM) increases the magnitude of both thetransient and steady-state components of 1& (Figure4A, 4C, and 4D). Figure 4B shows that the voltagedependence of the inactivation time-constant is notsignificantly affected by 100 nM AIL Further, the shapeof the current-voltage relation of the transient compo-nent of I a is not significantly changed by the additionof 100 nM All (Figure 4C). However, All does increasecurrent over the entire voltage range. Similarly, Figure4D shows nearly identical changes in the voltage de-
pendence of the steady-state component of IQ in thepresence of 100 nM All.
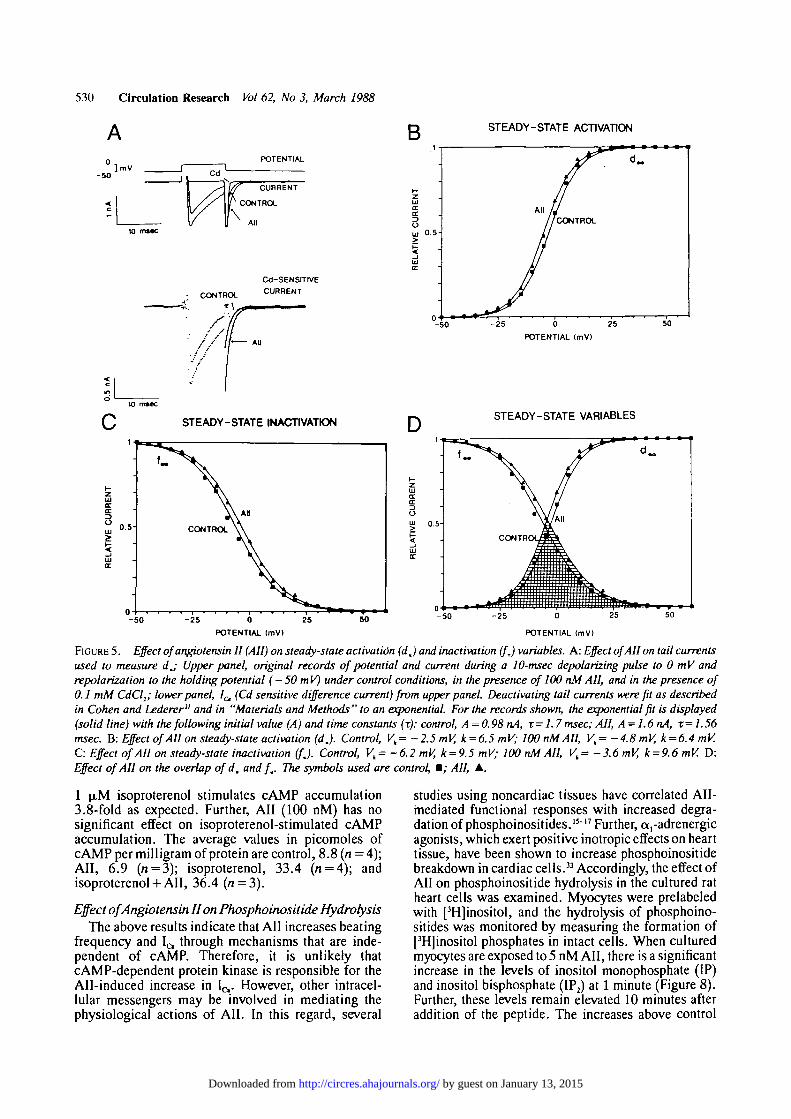
Steady-State Calcium Channel Current (d, and / JTo investigate the steady-state component of 1^
observed in Figure 4D, we measured d. and f I variables(Figure 5). To examine dx, the membrane potential washeld at - 50 mV. The tail current (L ,̂) was measuredupon returning to the holding potential following a10-msec pulse to V ^ (Figure 5A). To control for IQ
"rundown,"27"30 all trials to V M were alternated withtrials to V^(the test potential that gives the largest tailcurrent) and the ratio of the resulting tail currents(\JV^) was determined (Figure 5B). The smoothcurve shows the best-fit line described by a Boltzmannequation.
To investigate f., the membrane potential was heldat - 5 0 mV. Following a 200-msec voltage clamp stepto V^,, the membrane potential was returned to theholding potential for 10 msec and then depolarized tothe evaluation potential ( + 5 mV) for 200 msec. Tocontrol for 1^ "rundown," all trials to Vlcu werealternated with trials to V™ (the test potential that givesthe largest current transient at the evaluation potential),and the ratio of the resulting currents (L^a/I^ wasdetermined (Figure 5C). The smooth curve shows thebest-fit line described by a Boltzmann equation.
In Figure 5D, both d_ and f. have been plotted.These results show a large overlap of the two curves(hatched area). The voltage-dependence of the overlapof d. and f. constitutes a voltage range where signif-icant steady-state IQ should be seen.
Effect of Angiotensin II on dm and fm
To measure the effect of All on d. and f* we usedthe same protocols described above. All produces asmal 1 sh ift of d „ to more negat ive vol tages (Figure 5 B).All decreases Vb without materially affecting k (seeFigure 5 legend). The mean shift of Vh is —2.8 ±1 .1mV (n = 5;p<0.05, two-tailed t test). All produces asmall shift off. to more positive voltages (Figure 5C).All increases Vh without materially affecting k (seeFigure 5). The mean shift of Vh is + 2 . 6 ± 1 . 3 mV(« = 5;p<0.05, two-tailed / test). The effect of All onboth d. and f. is shown in Figure 5D. All evokes aslight increase in the magnitude of the overlap of d.and fx.
To calculate the shape of the current-voltage relationof the transient component of Ic from d. and f., wehave assumed that the kinetics of activation are alwaysmuch faster than the kinetics of inactivation.31 Inaddition, we have adjusted a voltage-independent"relative conductance" term, g,, to give us the best fitline by eye to the data points. The expression used is
( V j = g, x (Vm - EdmJ x [(1 - f.(V J] x d.(VJ(1)
where Vm is the membrane potential and E ^ ^ is thereversal potential for the current. The smooth curvesgenerated by Equation 1, which make use of the
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from

Allen et al Angiotensin II Stimulation of Cultured Heart Cells 529
- 50
D
- 2 5 0 25
POTENTIAL ImV)
STEADY-STATE ICa
- 0 .4 -
-K> 10
POTENTIAL (mVI
-50 -10 10
POTENTIAL (mVI
FIGURE 4. Effect of angiotensin II (All) on calcium channel current (Ia). A: Records of voltage and cadmium-sensitive current. Thecadmium-sensitive current elicited by a 200-msec depolarizing voltage step toOm Vwas measured in the absence (control) and presenceof 100 nM All (see "Materials and Methods "). B: Effect of All on the time constant of inactivation (x) of 1^ C: Effect of AH on thetransient component of the I ̂ -voltage relation. Smooth curves are predicted from measured d. and f. curves (see Figure 5) andEquation 1. Control, gr=22.22 nS; 100 nM All, gr =33.33 nS. D: The effect of All on the steady-state component of the I ^-voltagerelation. Smooth curves are predicted from measured dw and f. curves (see Figure 5) and Equation 2. Control, gr — 24.39 nS; 100nM All, gr=31.25 nS. For Panels B, C, and D, symbols used are control, • ; All, A.
experimentally determined d . (VJ and f,(Vm) curves,are shown in Figure 4C.
The same procedure was used to estimate thevoltage-dependence of the steady-state component of
(VJ = g, x (Vra - x d.(VJ x f.(VJ (2)
The smooth curves generated by Equation 2, whichmake use of the experimentally determined d . (VJ andf , (VJ curves, are shown in Figure 4D.
The results indicate that the g, values used to fit thetransient and steady-state la experimental results arevery similar. In addition, g, increases in the presenceof All. Presumably, g, = W x 7 , ^ ^ x n, where W is aprobability weighting coefficient reflecting the proba-bility that the channel is open independent of d. and fx,7dum*i ' s t n e single channel conductance, and n is thenumber of functional channels. The increase in bothsteady-state and transient components of I,-, in thepresence of All thus reflects alterations in both g, andsteady-state variables.
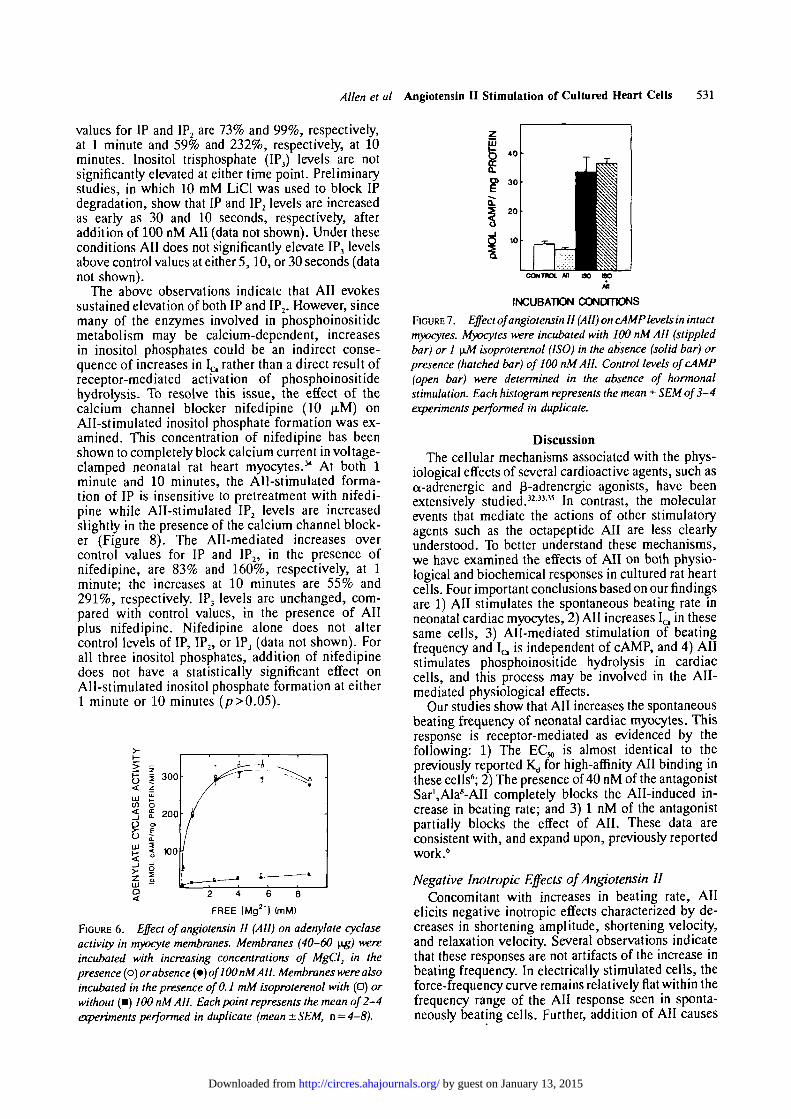
Effect of Angiotensin II on cAMP MetabolismIt is evident from the above data that All increases
IQ in the cultured myocytes. Since it is well known thatelevated cAMP levels increase I a in heart cells,32 theeffect of All on adenylate cyclase activity in myocytemembranes was examined. As shown in Figure 6, All(100 nM) has no effect on Mg2+-dependent adenylatecyclase activity. In contrast, isoproterenol (0.1 mM),which was included as a positive control, increasesadenylate cyclase activity maximally about 13-fold (at3.97 mM free Mg2+). Further, All (100 nM) does notsignificantly affect isoproterenol-stimulated adenylatecyclase activity.
Although All does not stimulate adenylate cyclaseactivity, the peptide could increase cAMP levels inintact cells through mechanisms that are independentof direct receptor-adenylate cyclase interactions (e.g.,inhibition of cAMP degradation). Therefore, the effectof All on cAMP levels in intact cells was determined.All, in doses as high as 100 nM, has no effect on controllevels of cAMP (Figure 7). Under the same conditions,
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from
530 Circulation Research Vol 62, No 3, March 1988
STEADY-STATE ACTIVATION
]mVPOTENTIAL
Cd-SENSrrrveCURRENT
-50 0 25
POTENTIAL (mV)
STEADY-STATE INACTIVATION D STEADY-STATE VARIABLES
LU
0.5-CONTROLS t
MAll
- 5 0 - 2 5 -50 25
POTENTIAL (mV) POTENTIAL (mV)
FIGURE 5. Effect of angiotensin II (All) on steady-state activation (dj and inactivation (fj variables. A: Effect of All on tail currentsused to measure dj Upper panel, original records of potential and current during a 10-msec depolarizing pulse to 0 mV andrepolarization to the holding potential ( — 50mV) under control conditions, in the presence of 100 nM AH, and in the presence of0.1 mM CdCl2; lower panel, I& (Cd sensitive difference current) from upper panel. Deactivating tail currents were fit as describedin Cohen and Lederer" and in "Materials and Methods" to an exponential. For the records shown, the exponential fit is displayed(solid line) with the following initial value (A) and time constants (r): control, A =0.98 nA, x=1.7 msec; All, A=1.6nA, x= 1.56msec. B: Effect ofAH on steady-state activation (dj. Control, Vk= -2.5 mV, k = 6.5 mV; 100 nM AH, Vt= -4.8 mV, k = 6.4 mV.C: Effect of All on steady-stale inactivation (fj. Control, Vh= -6.2 mV, k = 9.5 mV; 100 nM AH, Vk= -3.6 mV, k = 9.6 mV. D:Effect of AH on the overlap of d. and f.. The symbols used are control, • ; AH, A.
1 LIM isoproterenol stimulates cAMP accumulation3.8-fold as expected. Further, All (100 nM) has nosignificant effect on isoproterenol-stimulated cAMPaccumulation. The average values in picomoles ofcAMP per milligram of protein are control, 8.8 (n = 4);All, 6.9 (n = 3); isoproterenol, 33.4 (n = 4); andisoproterenol + All, 36.4 (n = 3).
Effect of Angiotensin II on Phosphoinositide HydrolysisThe above results indicate that All increases beating
frequency and I a through mechanisms that are inde-pendent of cAMP. Therefore, it is unlikely thatcAMP-dependent protein kinase is responsible for theAll-induced increase in IQ. However, other intracel-lular messengers may be involved in mediating thephysiological actions of AN. In this regard, several
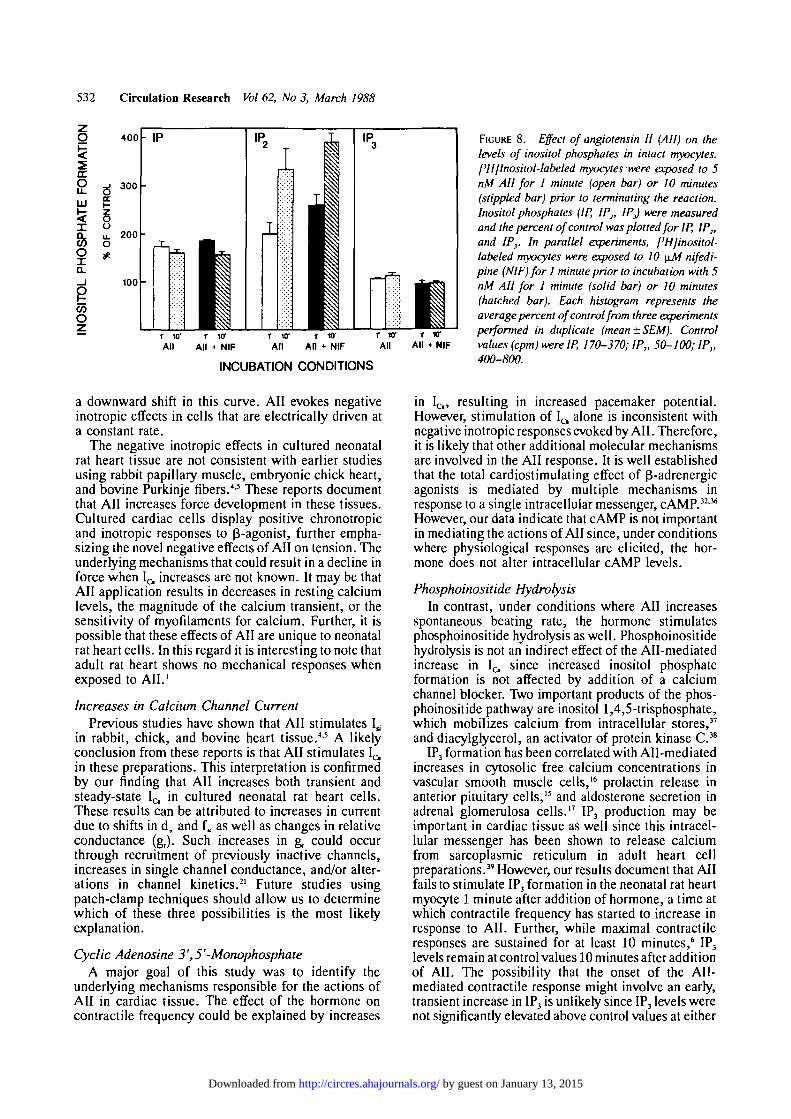
studies using noncardiac tissues have correlated AII-mediated functional responses with increased degra-dation of phosphoinositides.15"17 Further, a,-adrenergicagonists, which exert positive inotropic effects on hearttissue, have been shown to increase phosphoinositidebreakdown in cardiac cells.33 Accordingly, the effect ofAll on phosphoinositide hydrolysis in the cultured ratheart cells was examined. Myocytes were prelabeledwith [3H]inositol, and the hydrolysis of phosphoino-sitides was monitored by measuring the formation of[3H]inositol phosphates in intact cells. When culturedmyocytes are exposed to 5 nM All, there is a significantincrease in the levels of inositol monophosphate (IP)and inositol bisphosphate (IP2) at 1 minute (Figure 8).Further, these levels remain elevated 10 minutes afteraddition of the peptide. The increases above control
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from
Allen et al Angiotensln II Stimulation of Cultured Heart Cells 531
values for IP and IP2 are 73% and 99%, respectively,at 1 minute and 59% and 232%, respectively, at 10minutes. Inositol trisphosphate (IP3) levels are notsignificantly elevated at either time point. Preliminarystudies, in which 10 mM LiCl was used to block IPdegradation, show that IP and IP2 levels are increasedas early as 30 and 10 seconds, respectively, afteraddition of 100 nM AN (data not shown). Under theseconditions All does not significantly elevate IP3 levelsabove control values at either 5,10, or 30 seconds (datanot shown).
The above observations indicate that All evokessustained elevation of both IP and IP2. However, sincemany of the enzymes involved in phosphoinositidemetabolism may be calcium-dependent, increasesin inositol phosphates could be an indirect conse-quence of increases in l^ rather than a direct result ofreceptor-mediated activation of phosphoinositidehydrolysis. To resolve this issue, the effect of thecalcium channel blocker nifedipine (10 ^M) onAll-stimulated inositol phosphate formation was ex-amined. This concentration of nifedipine has beenshown to completely block calcium current in voltage-clamped neonatal rat heart myocytes.3* At both 1minute and 10 minutes, the All-stimulated forma-tion of IP is insensitive to pretreatment with nifedi-pine while All-stimulated IP2 levels are increasedslightly in the presence of the calcium channel block-er (Figure 8). The All-mediated increases overcontrol values for IP and IP2, in the presence ofnifedipine, are 83% and 160%, respectively, at 1minute; the increases at 10 minutes are 55% and291%, respectively. IP3 levels are unchanged, com-pared with control values, in the presence of Allplus nifedipine. Nifedipine alone does not altercontrol levels of IP, IP2, or IP3 (data not shown). Forall three inositol phosphates, addition of nifedipinedoes not have a statistically significant effect onAll-stimulated inositol phosphate formation at either1 minute or 10 minutes (p>0.05).
FREE [Mg2') (mM)
FIGURE 6. Effect of angiotensin II (All) on adenylate cyclaseactivity in myocyte membranes. Membranes (40-60 \ig) wereincubated with increasing concentrations of MgCl2 in thepresence (o) or absence (•) of100 nM All. Membranes were alsoincubated in the presence of 0.1 mM isoproterenol with (•) orwithout (•) 100 nM All. Each point represents the mean of 2-4experiments performed in duplicate (mean ± SEM, n = 4-8).
CONTROL M OO OO
INCUBATION CONDmONS
FIGURE 7. Effect of angiotensin II (All) on cAMP levels in intactmyocytes. Myocytes were incubated with 100 nM All (stippledbar) or 1 \iM isoproterenol (ISO) in the absence (solid bar) orpresence (hatched bar) of 100 nM AIL Control levels ofcAMP(open bar) were determined in the absence of hormonalstimulation. Each histogram represents the mean ± SEM of 3-4experiments performed in duplicate.
DiscussionThe cellular mechanisms associated with the phys-
iological effects of several cardioactive agents, such asa-adrenergic and p-adrenergic agonists, have beenextensively studied.3233-35 In contrast, the molecularevents that mediate the actions of other stimulatoryagents such as the octapeptide All are less clearlyunderstood. To better understand these mechanisms,we have examined the effects of All on both physio-logical and biochemical responses in cultured rat heartcells. Four important conclusions based on our findingsare 1) All stimulates the spontaneous beating rate inneonatal cardiac myocytes, 2) All increases IQ in thesesame cells, 3) All-mediated stimulation of beatingfrequency and I a is independent of cAMP, and 4) Allstimulates phosphoinositide hydrolysis in cardiaccells, and this process may be involved in the All-mediated physiological effects.
Our studies show that All increases the spontaneousbeating frequency of neonatal cardiac myocytes. Thisresponse is receptor-mediated as evidenced by thefollowing: 1) The EC*, is almost identical to thepreviously reported Kj for high-affinity All binding inthese cells6; 2) The presence of 40 nM of the antagonistSar',A]a8-AlI completely blocks the All-induced in-crease in beating rate; and 3) 1 nM of the antagonistpartially blocks the effect of All. These data areconsistent with, and expand upon, previously reportedwork.6
Negative Inotropic Effects of Angiotensin IIConcomitant with increases in beating rate, All
elicits negative inotropic effects characterized by de-creases in shortening amplitude, shortening velocity,and relaxation velocity. Several observations indicatethat these responses are not artifacts of the increase inbeating frequency. In electrically stimulated cells, theforce-frequency curve remains relatively flat within thefrequency range of the All response seen in sponta-neously beating cells. Further, addition of AH causes
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from
532 Circulation Research Vol 62, No 3, March 1988
400
a.P J 300UJ
85oQ.
CO
o
200
100
- IP
INCUBATION CONDITIONS
FIGURE 8. Effect of angiotensin II (All) on thelevels of inositol phosphites in intact myocytes.['HJInositol-labeled myocytes were exposed to 5nM AH for 1 minute (open bar) or 10 minutes(stippled bar) prior to terminating the reaction.Inositol phosphates (IP, IP2, IP) were measuredand the percent of control was plotted for IP, IP2,and IP}. In parallel experiments, ['Hjinositol-labeled myocytes were exposed to 10 \iM nifedi-pine (NIF) for 1 minute prior to incubation with 5nM All for 1 minute (solid bar) or 10 minutes(hatched bar). Each histogram represents theaverage percent of control from three experimentsperformed in duplicate (mean ± SEM). Controlvalues (cpm) were IP, 170-370; IP2, 50-100; IP},400-800.
a downward shift in this curve. All evokes negativeinotropic effects in cells that are electrically driven ata constant rate.
The negative inotropic effects in cultured neonatalrat heart tissue are not consistent with earlier studiesusing rabbit papillary muscle, embryonic chick heart,and bovine Purkinje fibers.0 These reports documentthat All increases force development in these tissues.Cultured cardiac cells display positive chronotropicand inotropic responses to fS-agonist, further empha-sizing the novel negative effects of All on tension. Theunderlying mechanisms that could result in a decline inforce when 1^ increases are not known. It may be thatAll application results in decreases in resting calciumlevels, the magnitude of the calcium transient, or thesensitivity of myofilaments for calcium. Further, it ispossible that these effects of All are unique to neonatalrat heart cells. In this regard it is interesting to note thatadult rat heart shows no mechanical responses whenexposed to AIL'
Increases in Calcium Channel CurrentPrevious studies have shown that All stimulates 1̂
in rabbit, chick, and bovine heart tissue.4-5 A likelyconclusion from these reports is that All stimulates 1^in these preparations. This interpretation is confirmedby our finding that All increases both transient andsteady-state I a in cultured neonatal rat heart cells.These results can be attributed to increases in currentdue to shifts in d, and f. as well as changes in relativeconductance (g,). Such increases in g, could occurthrough recruitment of previously inactive channels,increases in single channel conductance, and/or alter-ations in channel kinetics.21 Future studies usingpatch-clamp techniques should allow us to determinewhich of these three possibilities is the most likelyexplanation.
Cyclic Adenosine 3',5'-MonophosphateA major goal of this study was to identify the
underlying mechanisms responsible for the actions ofAll in cardiac tissue. The effect of the hormone oncontractile frequency could be explained by increases
in b , resulting in increased pacemaker potential.However, stimulation of IQ alone is inconsistent withnegative inotropic responses evoked by All. Therefore,it is likely that other additional molecular mechanismsare involved in the All response. It is well establishedthat the total cardiostimulating effect of P-adrenergicagonists is mediated by multiple mechanisms inresponse to a single intracellular messenger, cAMP.32-36
However, our data indicate that cAMP is not importantin mediating the actions of All since, under conditionswhere physiological responses are elicited, the hor-mone does not alter intracellular cAMP levels.
Phosphoinositide HydrolysisIn contrast, under conditions where AN increases
spontaneous beating rate, the hormone stimulatesphosphoinositide hydrolysis as well. Phosphoinositidehydrolysis is not an indirect effect of the All-mediatedincrease in 1^ since increased inositol phosphateformation is not affected by addition of a calciumchannel blocker. Two important products of the phos-phoinositide pathway are inositol 1,4,5-trisphosphate,which mobilizes calcium from intracellular stores,37
and diacylgrycerol, an activator of protein kinase C.38
IP3 formation has been correlated with All-mediatedincreases in cytosolic free calcium concentrations invascular smooth muscle cells,16 prolactin release inanterior pituitary cells,15 and aldosterone secretion inadrenal glomerulosa cells.17 IP3 production may beimportant in cardiac tissue as well since this intracel-lular messenger has been shown to release calciumfrom sarcoplasmic reticulum in adult heart cellpreparations.3' However, our results document that Allfails to stimulate IP3 formation in the neonatal rat heartmyocyte 1 minute after addition of hormone, a time atwhich contractile frequency has started to increase inresponse to All. Further, while maximal contractileresponses are sustained for at least 10 minutes,6 IP3
levels remain at control values 10 minutes after additionof All. The possibility that the onset of the All-mediated contractile response might involve an early,transient increase in IP3 is unlikely since IP3 levels werenot significantly elevated above control values at either
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from

Allen et al Angiotensln II Stimulation of Cultured Heart Cells 533
5, 10, or 30 seconds after addition of All. Based onthese findings, we conclude that IP3 is probably notinvolved in mediating either early or sustained con-tractile responses evoked by All.
In contrast, All stimulates the formation of IP andIP2 at 1 minute and these levels remain elevated 10minutes after exposure to the peptide. Although wehave not measured diacylglycerol production directly,the sustained elevations of both IP and IP2 reported hereare consistent with a sustained increase in diacylglyc-erol as well. Several recent reports have suggested animportant role for diacylglycerol in mediating sus-tained cellular responses to hormones.'40'" It is inter-esting to note that in both adrenal cortex and vascularsmooth muscle, All stimulates sustained diacylglyc-erol production while IP3 formation is transient.17-42 Ourresults are consistent with the hypothesis that theAll-mediated physiological responses seen in culturedheart cells result from an activation of the diacylgryc-erol/protein kinase C system.
A Possible Role for Protein Kinase C
The activation of protein kinase C could explain boththe negative inotropic effects and the increase in L^evoked by All. Recently, investigators working withelectrically stimulated adult rat myocardial cells43 andrat papillary muscle44 reported negative inotropicresponses evoked by phorbol esters, which mimicdiacylglycerol in their ability to activate protein kinaseC. A 15% decrease in resting intracellular calciumconcentration after phorbol ester treatment has alsobeen reported.44 Other studies using noncardiac tissueshave demonstrated that protein kinase C is involved inthe activation of calcium current and the recruitment ofnew calcium channels,45-46 the phosphorylation ofcontractile and cytoskeletal proteins,39-47 and the reg-ulation of smooth muscle contraction.48 It is also clearthat protein kinase C modulates calcium channelsdifferently in various noncardiac tissues. For example,activation of the enzyme in PC12 pheochromocytomacells and snail neurons results in apparent decreases incalcium current.49-50 Taken together, the above obser-vations, coupled with those reported in the presentstudy, provide a rationale for further study of proteinkinase C as a mediator of All action on heart cells.
References1. Peach MI: Renin-angiotensin system: Biochemistry and mech-
anism of action. Physiol Rev 1977^7:313-3702. Capponi AM, Aguilera G, Fakunding JL, Catt KJ: Angiotensin
II: Receptors and mechanisms of action, in Soffer RL (ed):Biochemical Regulation of Blood Pressure. New York, JohnWiley & Sons, Inc, 1981, pp 205-260
3. Bonnardeaux JL, Regoli D: Action of angiotensin II andanalogues on the heart. Can J Physiol Pharmacol 1974;52:50-60
4. Freer RS, Pappano AJ, Peach MJ, BingKT, McLean MJ, VogelSM, Sperelakis N: Mechanism for the positive inotropic effectof angiotensin II on isolated cardiac muscle. Ore Res 1976;39:178-183
5. Kass RS, Blair ML: Effects of angiotensin II on membranecurrent in cardiac Purkinje fibers. J Mol Cell Cardiol 1981;13:797-809
6. Rogers TB, Gaa ST, Allen IS: Identification and character-
ization of functional angiotensin II receptors on cultured heartmyocytes. / Pharmacol Exp Ther 1986;236:438-444
7. Rogers TB: High affinity angiotensin II receptors in myocardialsarcolemmal membranes. J Biol Chem 1984;259:8106-8114
8. Baker KM, Campanile CP, Trachte GJ, Peach MJ: Identifica-tion and characterization of the rabbit angiotensin II myocardialreceptor. Ore Res 1984;54:286-293
9. Wright GB, Alexander RW, Ekstein LS, Gimbrone MA Jr:Characterization of the rabbit ventricular myocardial receptorfor angiotensin II. Mol Pharmacol 1983;24:213-221
10. Crane JK, Campanile CP, Garrison JC: The hepatic angiotensinII receptor. / Biol Chem 1982;257:4959-4965
11. Marie J, Gaillard RC, Schoenenberg S, Jard S, Bockaert J:Pharmacological characterization of the angiotensin receptornegatively coupled with adenylate cyclase in rat anteriorpituitary gland. Endocrinology 1985;116:1044-1050
12. Anand-Srivastava MB: Angiotensin II receptors negativelycoupled to adenylate cyclase in rat aorta. Biochem Biophys ResCommun 1983;117:420-428
13. Marie J, Jard S: Angiotensin II inhibits adenylate cyclase fromadrenal cortex glomerulosa zone. FEBS Lett 1983; 159:97-101
14. Creba JA, Downes CP, Hawkins PT, Brewster G, Michell RH,Kirk CJ: Rapid breakdown of phosphat idyl inositol 4-phosphateand phosphatidylinositol 4,5-bisphosphate in rat hepatocytesstimulated by vasopressin and other Ca2+-mobilizing hor-mones. Biochem J 1983;212:733-747
15. Canonico PL, MacLeod RM: Angiotensin peptides stimulatephosphoinositide breakdown and prolactin release in anteriorpituitary cells in culture. Endocrinology 1986;118:233-238
16. Alexander RW, Brock TA, Gimbrone MA Jr, Rittenhouse SE:Angiotensin increases inositol trisphosphate and calcium invascular smooth muscle. Hypertension 1985;7:447-451
17. Kojima I, Kojima K, Kreutter D, Rasmussen H: The temporalintegration of the aldosterone secretory response to angiotensinoccurs via two intracellular pathways. J Biol Chem 1984;259:14448-14457
18. Bradford MA: A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal Biochem 1976;72:248-254
19. Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ:Improved patch-clamp techniques for high-resolution currentrecording from cells and cell-free membrane patches. PflugersArch 1981;391:85-100
20. Bezanilla F: A high capacity data recording device based on adigital audio processor and a video cassette recorder. BiophysJ 1985;47:437-441
21. Cohen NM, Lederer WJ: Calcium current in isolated neonatalrat ventricular myocytes. J Physiol 1987;391:169-191
22. Salomon Y, Londos C, Rodbell M: A highly sensitive adenylatecyclase assay. Anal Biochem 1974;58:541-548
23. Fabiato A, Fabiato F: Effects of magnesium on contractileactivation of skinned cardiac cells. J Physiol 1975;249:497-517
24. Harper JS, Brooker GJ: Femtomole sensitive radioimmunoas-say for cAMP and cGMP after 2'0 acetylation by aceticanhydride in aqueous solution. J Cyclic Nucleotide ProteinPhosphor Res 1975;l:207-218
25. Downes CP, Wusteman MM: Breakdown of poryphosphoino-sitides and not phosphatidylinositol accounts for muscarinicagonist-stimulated inositol phospholipid metabolism in ratparotid glands. Biochem J 1983;216:633-640
26. Nilius B, Hess P, Lansman JB, Tsien RW: A novel type ofcardiac calcium channel in ventricular cells. Nature 1985;316:443^46
27. Byerly L, Hagiwara S: Calcium currents in internally perfusednerve cell bodies of Limnea stagnalis. J Physiol (Lond) 1982;322:503-528
28. Fenwick EM, Marty A, Neher E: Sod ium and calcium channelsin bovine chromaffin eel Is. J Physiol (Lond) 1982;331:599-635
29. Bean BP: Nitrendipine block of cardiac calcium channels: Highaffinity binding to the inactivated state. Proc NatlAcad Sci USA1984;81:6388-6392
30. Lee KS, Tsien RW: High selectivity of calcium channels insingle diarysed heart cells of the guinea-pig. J Physiol (Lond)1984;354:253-272
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from

534 Circulation Research Vol 62, No 3, March 1988
31. Sanguinetti MC, Krafte DS, Kass RS: Voltage dependentmodulation of Ca channel current in heart cells by Bay K8644.J Gen Physiol 1986;88:369-392
32. Tsien RW, Bean BP, Hess P, Lansman JB, Nilius B, NowyckyMC: Mechanisms of calcium channel modulation by (}-adrenergic agents and dihydropyridine calcium agonists. JMolCell Cardiol 1986;18:691-710
33. Brown JH, Buxton IL, Brunton LL: o,-Adrenergic and mus-carinic cholinergic stimulation of phosphoinositide hydrolysisin adult rat cardiomyocytes. Ore Res 1985;57:532-537
34. Gurney AM, Nerbonne JM, Lester HA: Photoinduced removalof nifedipine reveals mechanisms of calcium antagonist actionon single heart cells. J Gen Physiol 1985;86:353-379
35. Bruckner R, Mugge A, Scholz H: Existence and functional roleof alpha,-adrenoceptors in the mammalian heart. J Mol CellCardiol 1985;17:639-645
36. Katz AM: Cyclic adenosine monophosphate effects on themyocardium: A man who blows hot and cold with one breath.J Am Coll Cardiol 1983;2:143-149
37. Williamson JR: Role of inositol lipid breakdown in thegeneration of intracellular signals. Hypertension 1986;8(suppl
) I I140II156)38. Nishizuka Y: Studies and perspectives of protein kinase C.
Science 1986;233:305-31239. Fabiato A: Inositol (l,4,5)-trisphosphate-induced release of
Ca2+ from the sarcoplasmic reticulum of skinned cardiac cells(abstract). Biophys J 1986;49:190a
40. Rasmussen H, Barrett PQ: Calcium messenger system: Anintegrated view. Physiol Rev 1984;64:938-984
41. Imai A, Gershengorn MC: Phosphatidylinositol 4,5-bis-phosphate turnover is transient while phosphatidylinositolturnover is persistent in thyrotropin-releasing hormone-stimulated rat pituitary cells. Proc Natl Acad Sci USA 1986;83:8540-8544
42. Griendling KK, Rittenhouse SE, Brock TA, Ekstein LS,Gimbrone MA Jr, Alexander RW: Sustained diacylghycerolformation from inositol phospholipids in angiotensin II-
stimulated vascular smooth muscle cells. J Biol Chem 1986;261:5901-5906
43. Capogrossi MC, Kaku T, Pelto DJ, Filburn C, Hansford RG,Spurgeon H, Lakatta EG: Phorbol ester translocates proteinkinase C and has a negative inotropic effect in rat cardiacmyocytes (abstract). Biophys J 1987;51:112a
44. Uglesity A, Sharma VK, Sheu S-S: Effect of protein kinase Cactivation on the inotropic response induced by a-adrenoceptorstimulation in rat myocardium (abstract). Biophys J 1987;51:264a
45. DeRiemerSA, StrongJA,AlbertKA, Greengard P, KaczmarekLK: Enhancement of calcium current in Aplysia neurons byphorbol ester and protein kinase C. Nature 1985;313:313-316
46. Strong JA, Fox AP, Tsien RW, Kaczmarek LK: Stimulation ofprotein kinase C recruits covert calcium channels in Apryisa bagcell neurons. Nature 1987^325:714-717
47. Naka M, Nishikawa M, Adelstein RS, Hidaka H: Phorbolester-induced activation of human platelets is associated withprotein kinase C phosphorylation of myosin light chains.Nature 1983;3O6:49(M92
48. Menkes H, Baraban JM, Snyder SH: Protein kinase C regulatessmooth muscle tension in guinea-pig trachea and ileum. EurJPharmacol 1986;122:19-27
49. Di Virgilio F, Pozzan T, Wollheim CB, Vicentini LM,Meldolesi J: Tumor promoter phorbol myristate acetate inhibitsCa:+ influx through voltage-gated Ca2+ channels in twosecretory cell lines, PC12 and RINm5F. / Biol Chem 1986;261:32-35
50. Hammond C, Paupardin-Tritsch D, Narin AC, Greengard P,Gerschenfeld HM: Cholecystokinin induces a decrease in Ca2+
current in snail neurons that appears to be mediated by proteinkinase C. Nature 1987;325:809-811
KEY WORDS • angiotensin II • calcium current • cyclicAMP • phosphoinositide hydrolysis • protein kinase C •inositol trisphosphate
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from

I S Allen, N M Cohen, R S Dhallan, S T Gaa, W J Lederer and T B Rogersmechanisms.
in cultured neonatal rat heart myocytes: insights into the underlying biochemical Angiotensin II increases spontaneous contractile frequency and stimulates calcium current
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1988 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.62.3.5241988;62:524-534Circ Res.
http://circres.ahajournals.org/content/62/3/524World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Research Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on January 13, 2015http://circres.ahajournals.org/Downloaded from
Copyright © 2022 FDOKUMEN




















