
Effect of season on contractile and metabolic properties of desert camel muscle (Camelus...
6
Effect of season on contractile and metabolic properties of desert camel muscle (Camelus dromedarius) O.M.A. Abdelhadi a, ⁎, S.A. Babiker b , B. Picard c , C. Jurie c , R. Jailler c , J.F. Hocquette c , B. Faye d a Dept. of Animal Production, Faculty of Natural Resources & Environmental Studies, University of Kordofan, Sudan b Dept. Meat Production, Faculty of Animal Production, University of Khartoum, Sudan c INRA, UR1213, Herbivore Research Unit, 63122 Theix, France d CIRAD, UR 18, Campus International de Baillarguet, 34398 Montpellier cedex 5, France abstract article info Article history: Received 17 June 2010 Received in revised form 25 May 2011 Accepted 10 June 2011 Keywords: Desert camel Metabolic activity Myosin heavy chain isoforms Season Thirty fattened one humped desert camels were used to examine the effect of season on contractile and metabolic properties of Longissimus thoracis (LT) muscle. Ten camels were slaughtered according to seasons of the year (winter, summer and autumn). Season significantly influenced muscle chemical composition, ultimate pH (pHu) and color. Activities of metabolic enzymes were higher during autumn season compared to summer and winter for phosphofructokinase (+64% compared to both seasons) and for isocitrate dehydrogenase (+35% and +145% in autumn vs. summer and winter, respectively). Quantification of muscle myosin heavy chain isoforms by SDS- PAGE electrophoresis showed only presence of type I and type IIa MyHC in camel muscle and indicated high proportion in winter for type I and in autumn for type IIa with respect to other seasons. Several correlations between different MyHC proportions and enzyme activities were reported. These findings indicated that muscle characteristics in camels are influenced by season. © 2011 Elsevier Ltd. All rights reserved. 1. Introduction Camel is a unique animal having the ability to survive and produce with low cost of feeding under harsh conditions compared to other livestock. It is a good source of meat in areas where the climate adversely affects other animal's production efficiency (Kadim et al., 2006). Traditionally, camel meat comes mostly from old males and females that are primarily kept for milk, racing, and transportation rather than for meat production. General consumers' view is that camel meat is unacceptably tough, but in fact meat from young camels has been reported to be comparable in taste and texture to beef (Kurtu, 2004). Carcass characteristics of camels were equal to those of other red meat animal species (Elgasim & Alkanhal, 1992). Chemically camel muscles had been found to have low fat content, high water holding capacity recommending camel meat as a healthy food with good processing properties (Babiker & Yousif, 1990). However, there is evidence of a great demand for fresh camel meat and for camel meat in blended meat products even in societies not herding camels (Morton, 1984; Pérez et al., 2000). Characteristics of camel and cattle meat as well as muscle fiber types had been studied by many authors (Costa et al., 2008; Kadim, Al-Hosni, et al., 2009; Kadim, Mahgoub, et al., 2009; Lefaucheur, 2010; Rose et al., 1992; Vestergaard, Oksbjerg, & Henchel, 2000). Different isoforms of Myosin Heavy Chain (MyHC) in bovine muscle were described: MyHC I, MyHC IIa and MyHC IIx (Picard, Duris, & Jurie, 1998). The MyHC IIb had been reported in some cattle from the Blonde d'Aquitaine French breed (Picard & Cassar-Malek, 2009). Muscle characteristics and meat quality have been known to be affected by growth and breed type (Renand, 1990), age and sex (Monin, 1990) as well as feeding level and diet composition (Geay, Bauchart, Hocquette, & Culioli, 2001). Klont, Brocks, and Eikelenboom (1998) noted that muscle metabolic and contractile types are adaptable and may be modified in living animals by environmental conditions and genetic selection. They indicated that in cattle, environmental effects were more important due to the large variations in production methods. To our knowledge, the effect of season on camel meat characteristics has not been reported. The present work aimed to investigate the effect of seasons on muscle characteristics of one humped desert camel (Camelus dromedarius). 2. Materials and methods 2.1. Sample collection Thirty fattened, intact males of the one hump desert camel, 2–3 years of age were used in this study. Ten camels were slaughtered at each season of the year: 2008 winter (Feb.–Mar.), summer (May–June) and autumn (Aug.–Sept.). Slaughter was performed following similar routine as described in Yousif and Babiker (1989). Average live weights were 319.4, 272.7 and 232.6 kg in winter, summer and autumn, respectively. Meat Science 90 (2012) 139–144 ⁎ Corresponding author at: P.O. Box: 716, Khartoum, Sudan. Tel.: + 249 9122 56 401. E-mail address: [email protected] (O.M.A. Abdelhadi). 0309-1740/$ – see front matter © 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.meatsci.2011.06.012 Contents lists available at ScienceDirect Meat Science journal homepage: www.elsevier.com/locate/meatsci
-
Upload
nationalagriculturalresearchinra -
Category
Documents
-
view
6 -
download
0
Transcript of Effect of season on contractile and metabolic properties of desert camel muscle (Camelus...

Meat Science 90 (2012) 139–144
Contents lists available at ScienceDirect
Meat Science
j ourna l homepage: www.e lsev ie r.com/ locate /meatsc i
Effect of season on contractile and metabolic properties of desert camel muscle(Camelus dromedarius)
O.M.A. Abdelhadi a,⁎, S.A. Babiker b, B. Picard c, C. Jurie c, R. Jailler c, J.F. Hocquette c, B. Faye d
a Dept. of Animal Production, Faculty of Natural Resources & Environmental Studies, University of Kordofan, Sudanb Dept. Meat Production, Faculty of Animal Production, University of Khartoum, Sudanc INRA, UR1213, Herbivore Research Unit, 63122 Theix, Franced CIRAD, UR 18, Campus International de Baillarguet, 34398 Montpellier cedex 5, France
⁎ Corresponding author at: P.O. Box: 716, Khartoum, SE-mail address: [email protected] (O.M.A
0309-1740/$ – see front matter © 2011 Elsevier Ltd. Aldoi:10.1016/j.meatsci.2011.06.012
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 June 2010Received in revised form 25 May 2011Accepted 10 June 2011
Keywords:Desert camelMetabolic activityMyosin heavy chain isoformsSeason
Thirty fattenedonehumpeddesert camelswere used to examine the effect of season oncontractile andmetabolicproperties of Longissimus thoracis (LT) muscle. Ten camels were slaughtered according to seasons of the year(winter, summer and autumn). Season significantly influencedmuscle chemical composition, ultimate pH (pHu)and color. Activities of metabolic enzymes were higher during autumn season compared to summer and winterfor phosphofructokinase (+64% compared to both seasons) and for isocitrate dehydrogenase (+35% and+145%in autumn vs. summer and winter, respectively). Quantification of muscle myosin heavy chain isoforms by SDS-PAGE electrophoresis showed only presence of type I and type IIa MyHC in camel muscle and indicated highproportion in winter for type I and in autumn for type IIa with respect to other seasons. Several correlationsbetween different MyHC proportions and enzyme activities were reported. These findings indicated thatmusclecharacteristics in camels are influenced by season.
udan. Tel.: +249 9122 56 401.. Abdelhadi).
l rights reserved.
© 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Camel is a unique animal having the ability to survive and producewith low cost of feeding under harsh conditions compared to otherlivestock. It is a good source of meat in areas where the climateadversely affects other animal's production efficiency (Kadim et al.,2006). Traditionally, camel meat comes mostly from old males andfemales that are primarily kept for milk, racing, and transportationrather than for meat production. General consumers' view is thatcamelmeat is unacceptably tough, but in fact meat from young camelshas been reported to be comparable in taste and texture to beef(Kurtu, 2004). Carcass characteristics of camels were equal to those ofother redmeat animal species (Elgasim & Alkanhal, 1992). Chemicallycamel muscles had been found to have low fat content, high waterholding capacity recommending camel meat as a healthy food withgood processing properties (Babiker & Yousif, 1990). However, thereis evidence of a great demand for fresh camelmeat and for camelmeatin blended meat products even in societies not herding camels(Morton, 1984; Pérez et al., 2000).
Characteristics of camel and cattle meat as well as muscle fibertypes had been studied by many authors (Costa et al., 2008; Kadim,Al-Hosni, et al., 2009; Kadim, Mahgoub, et al., 2009; Lefaucheur, 2010;Rose et al., 1992; Vestergaard, Oksbjerg, & Henchel, 2000). Different
isoforms of Myosin Heavy Chain (MyHC) in bovine muscle weredescribed: MyHC I, MyHC IIa and MyHC IIx (Picard, Duris, & Jurie,1998). The MyHC IIb had been reported in some cattle from theBlonde d'Aquitaine French breed (Picard & Cassar-Malek, 2009).Muscle characteristics and meat quality have been known to beaffected by growth and breed type (Renand, 1990), age and sex(Monin, 1990) as well as feeding level and diet composition (Geay,Bauchart, Hocquette, & Culioli, 2001). Klont, Brocks, and Eikelenboom(1998) noted that muscle metabolic and contractile types areadaptable and may be modified in living animals by environmentalconditions and genetic selection. They indicated that in cattle,environmental effects were more important due to the largevariations in production methods. To our knowledge, the effect ofseason on camel meat characteristics has not been reported. Thepresent work aimed to investigate the effect of seasons on musclecharacteristics of one humped desert camel (Camelus dromedarius).
2. Materials and methods
2.1. Sample collection
Thirty fattened, intactmales of the one humpdesert camel, 2–3 yearsof age were used in this study. Ten camels were slaughtered at eachseason of the year: 2008 winter (Feb.–Mar.), summer (May–June) andautumn(Aug.–Sept.). Slaughterwasperformed followingsimilar routineas described in Yousif and Babiker (1989). Average live weights were319.4, 272.7 and 232.6 kg in winter, summer and autumn, respectively.

Table 1Effect of season on chemical composition of Longissimus thoracismuscle of desert camel.
Parameter (%) Season S.E Overallmean
Effect ofseason (P-value)
Winter Summer Autumn
Moisture 76.7 76.8 77.3 0.26 76.9 0.82Dry matter 23.3 23.2 22.7 0.27 23.1 0.82Crude protein 16.8b 17.3a 17.5a 0.08 17.2 0.0002Intramuscular fat 2.7 2.5 2.6 0.17 2.6 0.87Ash 1.4b 1.5b 1.8a 0.05 1.6 0.0002
SEM = Standard error of the mean and Overall = Overall mean of the three seasons.a,b means with different superscripts within a row are significantly different (Pb0.05).
140 O.M.A. Abdelhadi et al. / Meat Science 90 (2012) 139–144
Mean values of temperature (°C) and relative humidity (%) were(26.5 °C, 22.5%), (35 °C, 24.5%) and (32 °C, 44.5%) inwinter, summer andautumn, respectively.
Muscle samples were collected immediately post-slaughter fromLongissimus thoracis (LT) of the right carcass side between the 5th and8th ribs. Samples were placed in plastic bags and transported within60 min of slaughter to the laboratory in an insulated box filled with ice.Visible fat was trimmed from the muscles and each muscle was sub-sampled (approximately 5 g sample weight) at the 5th rib. Sample waschopped into small pieces (0.5 cm), immersed in liquid nitrogen andstored in well tight plastic tubes at −18 °C for a week beforetransportation to INRA laboratories, France where samples were storedat−80 °C awaiting analysis ofmetabolic and contractile characteristics.Samples were also taken at the 6th rib for the determination of freshmuscle ultimate pH (pHu) and chemical composition. The remainder ofthe muscle was packed under vacuum and stored at −18 °C for themeasurements of frozen meat color and pHu.
2.2. Chemical composition
Muscle samples were ground to a homogenous mass in a grinderthen used for chemical analyses. Chemical composition of the muscletissue was measured according to standard methods of AOAC (1990).Crude protein was determined using a Foss Tecator Kjeltec 2300Nitrogen/Protein Analyzer. Fat was determined by Soxhlet extraction ofthe dry sample, using petroleum ether. Ash content was determined byashing samples in a muffle furnace at 500 °C for 24 h.
2.3. Meat color and pH
Muscle color co-ordinates (L*, a* and b*) were determined afterthawing of frozen samples to 2 °C as described by Allais et al. (2010)since it was not possible to measure meat color on freshmeat in Sudan.The L* value relates to lightness, the a * value to red–green hue where apositive value relates to the red intensity; and the b* value to theyellow–blue,where a positive value relates to yellow intensity. Ultimatemuscle pH (pHu) was measured using a portable pH meter (Hannawaterproof pH meter, Model H I 9025, Italy) and a temperatureadjusting probe. Readings were recorded in triplicates for eachmeasurement; the pH probe and the thermometer were inserted intomuscles to a similar depth (1.5 cm).
2.4. Metabolic enzyme activities
The metabolic muscle type was determined by measuring enzymeactivities. Glycolytic enzyme activities: phosphofructokinase (PFK, EC2.7.1.11) and lactate dehydrogenase (LDH, EC 1.1.1.27), and oxidativeenzyme activities: isocitrate dehydrogenase (ICDH, EC 1.1.1.42) andcytochrome-c oxidase (COX, EC 1.9.3.1) were measured spectropho-tometrically, according to the methods and the detailed protocolscited by Jurie, Ortigues-Marty, Picard, Micol, and Hocquette (2006).One unit of the enzyme was defined as the amount which catalyzesper min the disappearance of 1 μmol of NADH for PFK and LDH, thereduction of 1 μmol of NADP for ICDH and the oxidation of 1 μmol ofcytochrome-c for COX. Enzyme activities (means of triplicate) wereexpressed in μmol per min per gram of wet muscle (μmol/min per gmuscle).
2.5. Contractile characteristics of LT muscle
The contractile muscle type was determined by quantifying thedifferent myosin heavy chain (MyHC) isoforms by electrophoresisSDS-PAGE according to the method of Talmadge and Roy (1993) andadapted for bovine muscle by Picard, Barboiron, Chadeyron, and Jurie(2007). Each hole of gel was loaded with 4–5 μg of myofibrillarprotein and electrophoresis was performed at a constant voltage of
70 V for 30 h at 4 °C. The relative proportions of slow isoforms (%MyHC I) and fast isoforms (% MyHC IIa, IIx and IIb) were determinedafter staining of gels in a solution of Coomassie Blue R250 andquantifying using Image Quant TL v2003.
2.6. Statistical analyses
Data were analyzed using Statistical Analysis Systems package (SAS,2001) to evaluate the effect of seasons on muscle characteristics ofdesert camels (Camelus dromedaries). Significant differences betweenmeans were assessed using DUNCAN multiple range test. PrincipalComponent Analysis (PCA) was used to interpret the relationshipbetween different parameters as previously used in cattle for carcasscomposition (Albertí et al., 2008; Hocquette, Bas, Bauchart, Vermorel, &Geay, 1999) and meat quality (Destefanis, Barge, Brugiapaglia, &Tassone, 2000).
3. Results and discussion
3.1. Chemical composition
The overall chemical composition of Longissimus thoracis muscleshowed mean values of 76.9, 23.1, 17.2, 2.6 and 1.6% for moisture, drymatter, crude protein, intramuscular fat and ash content, respectively.Generally themean values of chemical composition in the present workwerewithin the range reportedpreviously formoisture (70–77%), crudeprotein (20–23%), fat (0.5–9.8%) and ash (1–1.3%), for LT muscle of1–3 years old camels (Al-Ani, 2004; Al-Owaimer, 2000; Al-Sheddy, Al-Dagal, & Bazaraa, 1999; Babiker & Yousif, 1990; Dawood & Alkanhal,1995; El-Faer, Rawdah, Attar, & Dawson, 1991; Kadim et al., 2006;Kadim & Mahgoub, 2006; Kadim, Mahgoub, et al., 2009; Kadim,Mahgoub, Al-Marzooqi, Al-Maqbali, & Al-Lawati, 2008; Kamoun, 1995).
No significant differences were found among seasons for moisture,dry matter and intramuscular fat content in camel LT muscle(Table 1). However during winter season both crude protein andash were either lower or significantly lower than values for summerand autumn seasons. The decrease in crude protein and ash duringwintermight be due to nutritional factors as winter feed requirementsare higher than that of the other seasons. Crude protein values ofcamel LT muscles during the different seasons of the year wereslightly lower (17.2%) than values earlier reported by Babiker andYousif (1990) for desert camels in the same region (21.6%), as well asDawood and Alkanhal (1995), Kamoun (1995), Al-Sheddy et al.(1999), Kadim et al. (2006) and Kadim, Mahgoub, et al. (2009) gavecrude protein values of the camel LT muscles of 20.0, 18.7, 21.3, 22.7and 21.6%, respectively. Ash content was significantly higher inautumn than in summer and winter (1.8 vs. 1.5 and 1.4%, Table 1) andalmost similar to that reported by Babiker and Yousif (1990) andKadim, Mahgoub, et al. (2009).
In general, camel meat is high in moisture content, moderate inprotein and ash, but low in fat, which clearly confirms the fact thatcamel meat is healthier due to its low intramuscular fat contentcompared to other species (for review, see Hocquette et al., 2010) likecattle (Chambaz, Scheeder, Kreuzer, & Dufey, 2003; Delgado et al.,

AutumnSummerWinter
A
B C
IIb
IIx
IIa
I
Fig. 1. Separation of different myosin heavy chain (MyHC) isoforms of Longissimusthoracismuscle of the camel and bovine by electrophoresis SDS-PAGE. A = Longissimusthoracis of camels, B = Longissimus thoracis of bovine, and C = Diaphragm of bovine. I,IIa, IIx and IIb are myosin heavy chain isoforms.
141O.M.A. Abdelhadi et al. / Meat Science 90 (2012) 139–144
2005; Sasaki, Mitsumoto, & Kawabata, 2001), sheep (Farid, 1991; Sen,Santra, & Kadim, 2004) and goats (Gaili & Ali, 1985; Mahgoub, Kadim,Al-Saqry, & Al-Busaidi, 2004; Sen et al., 2004).
3.2. Muscle pH and color
Table 2 shows the effect of season onmuscle pHu and color of camelLTmuscle. Muscle pHu values determined in fresh and thawed sampleswere almost identical, only the later were included in the table.The mean value pHu was 5.8 which falls within the pHu range value of5.4–6.0 reported by Kadim and Mahgoub (2006), Shariatmadari andKadivar (2006), Kadim et al. (2006, 2008) and Kadim, Mahgoub, et al.(2009) for camel Longissimus dorsi muscle at 1–3 years of age. MusclepHu in the present work was significantly higher in autumn than inwinter or summer respectively (6.03 vs. 5.7 and 5.8, P=0.002). Thissuggests that the muscle tissue may have low stored muscle glycogenduring autumn due to a slight higher (but not significant) moisturecontent of themeat during this season. Soltanizadeh, Kadivar, Keramat,and Fazilati (2008) and Kadim et al. (2006) indicated that camels havehigher pH values compared to bovine especially in young ages due toslow/or low level of stored glycogen.
The overall mean values of L*, a* and b* were 34.0, 13.2 and 11.2,respectively. The L*, a* values were lower while the b* values werehigher than that reported byKadim et al. (2006), Kadim,Mahgoub, et al.(2009) and Kadim, Al-Hosni, et al. (2009) for camel LT muscle. Thesedifferences could be attributed to breed differences. Between seasons,significant differences were observed in a* (P=0.03) and b* values(P=0.0001), while, no significant differences were observed in L*values. The b* values were significantly lower in autumn than insummer and winter (9.4 vs. 12.9 and 11.3), and a* values weresignificantly higher in autumn (14.6) than summer (11.8). Thesefindings indicated that camel meat color was variable between seasons.It was significantly darker red in autumn than in summer and lessyellow in autumn than in summer and winter. Young, Seok, Young, andSung (2003) found significant effect of seasons on color and pH of LTmuscle in cattle: The L*, a* and b* values were significantly lower(Pb0.05) for cattle slaughtered inwinter compared to summer, autumnand spring. In contrast, Kadim et al. (2004) found significant increase inL*, a* and b* values in bovineduring cold season compared tohot season.They concluded that season had a significant effect on meat qualitycharacteristics of beef LT muscle: ambient temperatures of approxi-mately 35°C had increased the ultimate pH and resulted in darker meatcolor (L*, a*and b* reduced). Camel LT muscle is characterized by moreredness and less yellowness in autumn than in the other seasons of theyear which make the meat dark red in color during this season.
3.3. Contractile characteristics of LT muscle
The separation of different MyHC isoforms by electrophoresis SDS-PAGE revealed twoMyHC isoforms (MyHC I andMyHC IIa) in camel LTmuscle (Fig. 1). The fast isoforms (MyHC IIx and MyHC IIb), describedin bovine muscle (Picard & Cassar-Malek, 2009) and found in bovine
Table 2Effect of season onmuscle pHu and color values of Longissimus thoracismuscle of desertcamels.
Parameter Season S.E. Overallmean
Effect ofseason (P-value)
Winter Summer Autumn
pHu 5.7b 5.8b 6.03a 0.24 5.8 0.002Muscle color
L* lightness 34.6 34.9 32.6 0.45 34 0.32a* redness 13.3ab 11.8b 14.6a 0.68 13.2 0.03b* yellowness 11.3a 12.9a 9.4b 0.35 11.2 0.0001
SEM = Standard error of the mean, Overall = Overall mean of the three seasons.a,b means with different superscripts within a row are significantly different (Pb0.05).
LT muscle were taken as a reference for our electrophoresis (Fig. 1),but they were not revealed in any of the camel LT samples. This resultfits with the absence of IIX (fast glycolytic) fibers in camel LT in thepresent study.
The proportions of the MyHC I and MyHC IIa isoforms weresignificantlymodifiedwith season (P=0.0001), (Table 3). TheMyHC Ipercentage was the highest in winter (85.2), least in autumn (52.9)and intermediate in summer (65.4), and inversely the MyHC IIapercentage was least in winter (14.8), highest in autumn (47.1) andintermediate in summer (34.6). The fiber types differ in MyHCisoforms among muscles. The major MyHC isoforms (slow MyHC I orfast MyHC IIa, IIx and IIb) determine the contractile activity of musclefibers. So slow MyCH I and fast MyHC IIa, IIx and IIb were reported tobe expressed in type I and type IIA, and type IIX and IIB fibers,respectively. In addition, the types of muscle fibers can be classified onthe basis of their contractile and metabolic activities. The commonclassification involves slow twitch oxidative (SO, type I), fast twitchoxidative glycolytic (FOG, type IIA) and fast twitch glycolytic (FG, typeIIX and type IIB) fiber types. Consequently slow fibers have a higheroxidative activity as they are able to use mainly fatty acids as energy-yielding nutrients.
Unlike this study, the presence of type IIB muscle fibers (or FGfibers) has been reported by Kadim,Mahgoub, et al. (2009) and Kadimet al. (2004) in LT muscle of camels. This discrepancy can be explainedby the fact that these authors used histochemistry to assess both thecontractile and metabolic properties of muscle fibers unlike in ourstudy in which we assessed myosin heavy chain proportions only. Arecent work indicated that a classification based on ATPase and SDHactivities may not be always appropriate, since FOG fibers may haveno SDH activity and may be confused with FG fibers (Oury, Dumont,Jurie, Hocquette, & Picard, 2010). The overall mean percentages oftype I and type IIa fibers in the present study were 67.8 and 32.2%,respectively. In the present work, it was observed that the proportionof type I fibers were higher than type IIa among seasons, which isgenerally in line with the results of Kadim, Mahgoub, et al. (2009)who observed a higher proportion of type I (33.1%) compared to typeIIa (25.2%). Rose et al. (1992) also reported a higher proportion of typeI compared to type IIa muscle fibers in racing camels during restingtime with an average of 70.3 and 28.0%, respectively.
In contrast, Kadim, Al-Hosni, et al. (2009) found a low proportionof type I vs. types IIa and IIb (29.9 vs. 40.1 and 29.9)%, respectively, inLT muscle of 1–3 years old camels, while Kassem, El-Sayed, andAhmed (2004) reported mean percentages of 14.5%, 46.7% and 38.8%for types I, IIa and IIb fibers in LT muscle of 2 years old camels,respectively. In addition to breed differences, genotype and seasonalambient temperature were reported to affect muscle biochemicalcharacteristics (Gondret, Combes, Lefaucheur, & Lebret, 2005;Gunning & Hardeman, 1991; Pette & Staron, 2001). The decrease inthe proportion of type I MyHC isoforms (from 85.2 in winter to 52.9%in autumn) is associated with an increase in type IIa MyHC isoforms(from 14.8 in winter to 47.1% in autumn) concomitant to high values
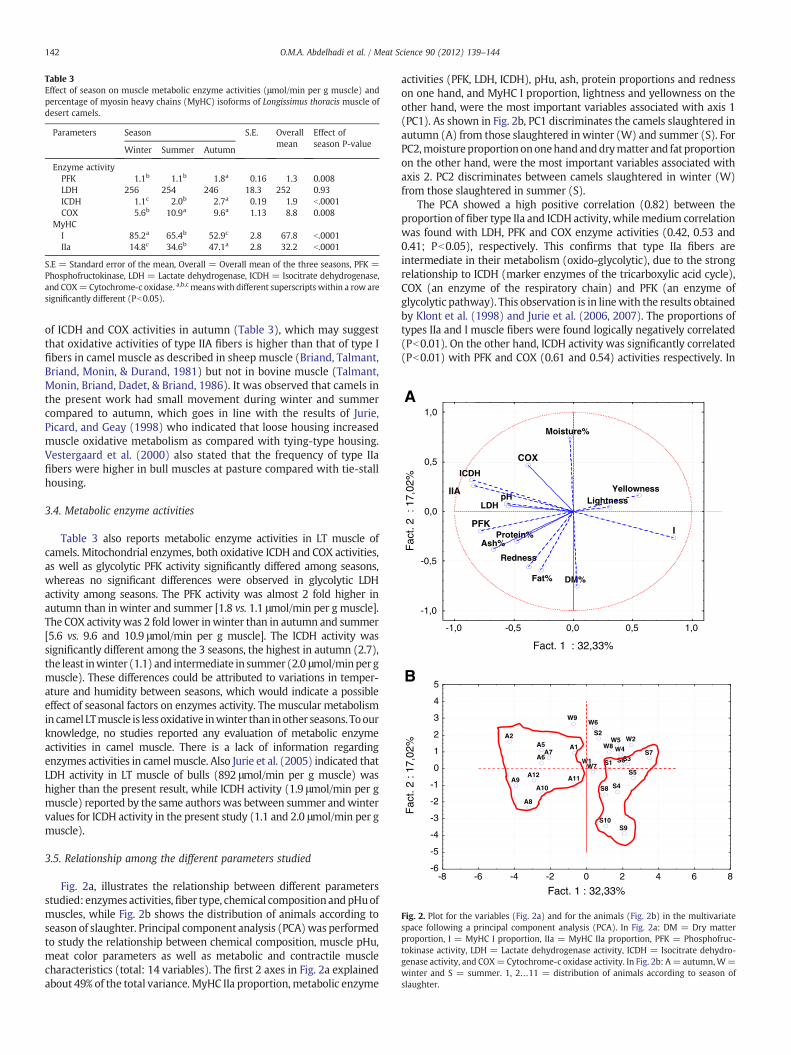
Table 3Effect of season on muscle metabolic enzyme activities (μmol/min per g muscle) andpercentage of myosin heavy chains (MyHC) isoforms of Longissimus thoracis muscle ofdesert camels.
Parameters Season S.E. Overallmean
Effect ofseason P-value
Winter Summer Autumn
Enzyme activityPFK 1.1b 1.1b 1.8a 0.16 1.3 0.008LDH 256 254 246 18.3 252 0.93ICDH 1.1c 2.0b 2.7a 0.19 1.9 b.0001COX 5.6b 10.9a 9.6a 1.13 8.8 0.008
MyHCI 85.2a 65.4b 52.9c 2.8 67.8 b.0001IIa 14.8c 34.6b 47.1a 2.8 32.2 b.0001
S.E = Standard error of the mean, Overall = Overall mean of the three seasons, PFK =Phosphofructokinase, LDH = Lactate dehydrogenase, ICDH = Isocitrate dehydrogenase,and COX=Cytochrome-c oxidase. a,b,c meanswith different superscripts within a row aresignificantly different (Pb0.05).
Moisture%
DM%
Protein%
Fat%
Ash%
Lightness
Redness
YellownesspH
ICDH
LDH
PFK
COX
IIA
I
-1,0 -0,5 0,0 0,5 1,0
Fact. 1 : 32,33%
-1,0
-0,5
0,0
0,5
1,0F
act.
2 :
17,0
2%
A1
A2A5
A6A7
A8
A9A10
A11A12
W1
W2
W4W5
W6
W7
W8
W9
S1
S2
S3
S4
S5
S6S7
S8
S9S10
-8 -6 -4 -2 0 2 4 6 8
Fact. 1 : 32,33%
-6
-5
-4
-3
-2
-1
0
1
2
3
4
5
Fac
t. 2
: 17,
02%
A
B
Fig. 2. Plot for the variables (Fig. 2a) and for the animals (Fig. 2b) in the multivariatespace following a principal component analysis (PCA). In Fig. 2a: DM = Dry matterproportion, I = MyHC I proportion, IIa = MyHC IIa proportion, PFK = Phosphofruc-tokinase activity, LDH = Lactate dehydrogenase activity, ICDH = Isocitrate dehydro-genase activity, and COX=Cytochrome-c oxidase activity. In Fig. 2b: A= autumn,W=winter and S = summer. 1, 2…11 = distribution of animals according to season ofslaughter.
142 O.M.A. Abdelhadi et al. / Meat Science 90 (2012) 139–144
of ICDH and COX activities in autumn (Table 3), which may suggestthat oxidative activities of type IIA fibers is higher than that of type Ifibers in camel muscle as described in sheep muscle (Briand, Talmant,Briand, Monin, & Durand, 1981) but not in bovine muscle (Talmant,Monin, Briand, Dadet, & Briand, 1986). It was observed that camels inthe present work had small movement during winter and summercompared to autumn, which goes in line with the results of Jurie,Picard, and Geay (1998) who indicated that loose housing increasedmuscle oxidative metabolism as compared with tying-type housing.Vestergaard et al. (2000) also stated that the frequency of type IIafibers were higher in bull muscles at pasture compared with tie-stallhousing.
3.4. Metabolic enzyme activities
Table 3 also reports metabolic enzyme activities in LT muscle ofcamels. Mitochondrial enzymes, both oxidative ICDH and COX activities,as well as glycolytic PFK activity significantly differed among seasons,whereas no significant differences were observed in glycolytic LDHactivity among seasons. The PFK activity was almost 2 fold higher inautumn than in winter and summer [1.8 vs. 1.1 μmol/min per g muscle].The COX activity was 2 fold lower inwinter than in autumn and summer[5.6 vs. 9.6 and 10.9 μmol/min per g muscle]. The ICDH activity wassignificantly different among the 3 seasons, the highest in autumn (2.7),the least inwinter (1.1) and intermediate in summer (2.0 μmol/minper gmuscle). These differences could be attributed to variations in temper-ature and humidity between seasons, which would indicate a possibleeffect of seasonal factors on enzymes activity. The muscular metabolismin camel LTmuscle is less oxidative inwinter than inother seasons. Toourknowledge, no studies reported any evaluation of metabolic enzymeactivities in camel muscle. There is a lack of information regardingenzymes activities in camelmuscle. Also Jurie et al. (2005) indicated thatLDH activity in LT muscle of bulls (892 μmol/min per g muscle) washigher than the present result, while ICDH activity (1.9 μmol/min per gmuscle) reported by the same authors was between summer andwintervalues for ICDH activity in the present study (1.1 and 2.0 μmol/min per gmuscle).
3.5. Relationship among the different parameters studied
Fig. 2a, illustrates the relationship between different parametersstudied: enzymes activities,fiber type, chemical compositionandpHuofmuscles, while Fig. 2b shows the distribution of animals according toseason of slaughter. Principal component analysis (PCA)was performedto study the relationship between chemical composition, muscle pHu,meat color parameters as well as metabolic and contractile musclecharacteristics (total: 14 variables). The first 2 axes in Fig. 2a explainedabout 49% of the total variance. MyHC IIa proportion, metabolic enzyme
activities (PFK, LDH, ICDH), pHu, ash, protein proportions and rednesson one hand, and MyHC I proportion, lightness and yellowness on theother hand, were the most important variables associated with axis 1(PC1). As shown in Fig. 2b, PC1 discriminates the camels slaughtered inautumn (A) from those slaughtered in winter (W) and summer (S). ForPC2,moistureproportionononehandanddrymatter and fat proportionon the other hand, were the most important variables associated withaxis 2. PC2 discriminates between camels slaughtered in winter (W)from those slaughtered in summer (S).
The PCA showed a high positive correlation (0.82) between theproportion of fiber type IIa and ICDH activity, while medium correlationwas found with LDH, PFK and COX enzyme activities (0.42, 0.53 and0.41; Pb0.05), respectively. This confirms that type IIa fibers areintermediate in their metabolism (oxido-glycolytic), due to the strongrelationship to ICDH (marker enzymes of the tricarboxylic acid cycle),COX (an enzyme of the respiratory chain) and PFK (an enzyme ofglycolytic pathway). This observation is in linewith the results obtainedby Klont et al. (1998) and Jurie et al. (2006, 2007). The proportions oftypes IIa and I muscle fibers were found logically negatively correlated(Pb0.01). On the other hand, ICDH activity was significantly correlated(Pb0.01) with PFK and COX (0.61 and 0.54) activities respectively. In

143O.M.A. Abdelhadi et al. / Meat Science 90 (2012) 139–144
addition, activities of PFK and LDH (another enzyme of the glycolyticpathway) were also positively correlated (r=0.53; Pb0.01, Fig. 2a). Onaverage, this analyses indicates that type IIa muscle fibers have a highermetabolic activity (oxidative and glycolytic) than type I fibers.
The relationship between PFK and muscle pHu has been reportedearlier by Trivedi and Danforth (1966), Pettigrew and Frieden (1979)and Ichihara and Abiko (1982) who indicated that PFK activity ishighly sensitive and inhibited by low muscle pH in situ in rabbit, frogand canine muscles. Rhoades et al. (2005) indicated that PFK activityin bovine muscles was depressed in low pHu. These findingscontrasted the present results that no significant correlation coeffi-cient (PN0.05) was observed between PFK enzyme and muscle pHu,although higher pHu value and PFK activity were observed in autumncompared to other seasons (Pb0.001). Significant correlation(Pb0.01) was found between pHu and ICDH (0.59).
No significant relationship was observed in the present findingsbetween pHu, or LDH with color (L*, a*, and b*) values. This contrastswith the results of Mancini, Kim, Hunt, and Lawrence (2004) whoreported that LDH is involved in regeneration of postmortem NADHfrom lactate and, consequently, in metmyoglobin reduction whichdiminishes redness of meat. However, the correlations between PFKand L* on one hand, and a* values were −0.26 and +0.43,respectively, which indicated that camel meat color was brighterand less red when PFK activity (i.e. glycolytic metabolism) was high.This fits with the findings reported by Cuvelier et al. (2006) in bovineLT muscle who indicated that bovine meat was redder when COXactivity (i.e. oxidative metabolism) increased and brighter when LDHactivity (i.e. glycolytic metabolism) was high. Significant correlationcoefficient was observed between pHu and ICDH (0.59, Pb0.01). Theb* value was also negatively correlated (Pb0.05) with ICDH (−0.39).
In conclusion, the results from this study would indicate thatmuscle fiber characteristics of desert camel's Longissimus thoracismuscle as well as enzymes activities could be influenced by seasonalfactors. The study also revealed that the season has a significant effecton chemical composition of LT muscle (ash and protein), as well asmeat color in terms of a* and b* values.
Acknowledgments
The chemical analysis at INRA-Theix, France was funded by theFrench embassy at Khartoum, Sudan. Thanks are extended toChristiane Barboiron and David Chadeyron from Herbivore ResearchUnit (INRA-Theix, France) for their assistance in the lab.
References
Al-Ani, F. K. (2004). Use and production of camels. In F. K. Al-Ani (Ed.), Camelmanagement and diseases (pp. 91–114). (1st ed.). : Al-Sharq Printing Press.
Albertí, P., Panea, B., Sanudo, C., Olleta, J. L., Ripoll, G., Ertbjerg, P., et al. (2008). Liveweight, body size and carcass characteristics of young bulls of fifteen Europeanbreeds. Livestock Science, 114, 19–30.
Allais, S., Leveziel, H., Payet, N., Hocquette, J. F., Lepetit, J., Rousset, S., et al. (2010). Thetwo mutations Q204X and nt821 of the myostatin gene affect carcass and meatquality in heterozygous young bulls of three French beef breeds. Journal of AnimalScience, 88, 446–454.
Al-Owaimer, A. N. (2000). Effect of dietary Halophyte Salicornia bigelovii Torr on carcasscharacteristics, minerals, fatty acids and amino acids profile of camel meat. AppliedAnimal Research, 18, 185–192.
Al-Sheddy, I., Al-Dagal, M., & Bazaraa, W. A. (1999). Microbial and sensory quality offresh camel meat treated with organic acid salts and/or bifidobacteria. Journal ofFood Science, 64, 336–339.
AOAC (1990). Official methods of analysis (15th Ed.). Arlington, VA, USA: Associationofficial Analytical Chemists.
Babiker, S. A., & Yousif, O. Kh. (1990). Chemical composition and quality of camel meat.Meat Science, 27(4), 283–287.
Briand, M., Talmant, A., Briand, Y., Monin, G., & Durand, R. (1981). Metabolic types ofmuscle in the sheep. 1. Myosin atpase, glycolytic, and mitochondrial enzyme-activities. European Journal of Applied Physiology and Occupational Physiology, 46,347–358.
Chambaz, A., Scheeder, M. R. L., Kreuzer, M., & Dufey, P. A. (2003).Meat quality of Angus,Simmental, Charolais and Limousin steers compared at the same fat content.
Costa, P., Roseiro, L. C., Bessa, R. J. B., Padilha, M., Partidario, A., Marques, J., et al. (2008).Musclefibre and fatty acid profile ofMertolenga-PODmeat.Meat Science, 78, 502–512.
Cuvelier, C., Cabaraux, J. F., Dufrasne, I., Clinquart, A., Hocquette, J. F., Istasse, L., et al.(2006). Performance, slaughter characteristics and meat quality of young bullsfrom Belgian Blue, Limousin and Aberdeen Angus breeds fattenedwith a sugar-beetpulp or a cereal-based diet. Animal Science, 82, 125–132.
Dawood, A., & Alkanhal, M. A. (1995). Nutrient composition of Najidi Camel Meat.MeatScience, 39, 71–78.
Delgado, E. J., Rubio, M. S., Iturbe, F. A., Mendez, R. D., Cassis, L., & Rosiles, R. (2005).Composition and quality of Mexican and imported retail beef in Mexico. MeatScience, 69, 465–471.
Destefanis, G., Barge, M. T., Brugiapaglia, A., & Tassone, S. (2000). The use of principalcomponent analysis (PCA) to characterize beef. Meat Science, 56, 255–259.
El-Faer, M. Z., Rawdah, T. N., Attar, K. M., & Dawson, M. V. (1991). Mineral andproximate composition of the meat of the one-humped camel (Camelusdromedaries). Food Chemistry, 42, 139–143.
Elgasim, E. A., & Alkanhal, M. A. (1992). Proximate composition, amino acids andinorganic mineral content of Arabian camel meat. Comparative study. FoodChemistry, 45, 1–4.
Farid, A. (1991). Carcass physical and chemical composition of three fat-tail breeds ofsheep. Meat Science, 29, 109–120.
Gaili, E. S., & Ali, A. E. (1985). Meat from Sudan desert sheep and goats: Part 2-composition of the muscular fatty tissues. Meat Science, 13, 229–236.
Geay, Y., Bauchart, D., Hocquette, J. F., & Culioli, J. (2001). Effect of nutritional factors onbiochemical, structural and metabolic characteristics of muscles in ruminants;consequences on dietetic value and sensorial qualities of meat. ReproductionNutrition Development, 41, 1–26 Erratum, 41, 377.
Gondret, F., Combes, S., Lefaucheur, L., & Lebret, B. (2005). Effects of exercise duringgrowth and alternative rearing systems on muscle fibres and collagen properties.Reproduction Nutrition Development, 45, 69–86.
Gunning, P., & Hardeman, E. (1991). Multiple mechanisms regulate muscle fibrediversity. The FASEB Journal, 5, 3064–3070.
Hocquette, J. F., Bas, P., Bauchart, D., Vermorel, M., & Geay, Y. (1999). Fat partitioningand biochemical characteristics of fatty tissues in relation to plasma metabolitesand hormones in normal and double-muscled young growing bulls. ComparativeBiochemistry and physiology A, 122, 127–138 Erratum, 1999. ComparativeBiochemistry and physiology, 123, 311–312.
Hocquette, J. F., Gondret, F., Baéza, E., Médale, F., Jurie, C., & Pethick, D. W. (2010).Intramuscular fat content in meat-producing animals: development, genetic andnutritional control, identification of putative markers. Animal, 4, 303–319.
Ichihara, K., & Abiko, Y. (1982). Crossover plot study of glycolytic intermediates in theischemic canine heart. Japanese Heart Journal, 23(5), 817–828.
Jurie, C., Cassar-Malek, I., Bonnet, M., Leroux, C., Bauchart, D., Boulesteix, P., et al. (2007).Adipocyte fatty acid-binding protein and mitochondrial enzyme activities inmuscles as relevant indicators of marbling in cattle. Journal of Animal Science, 85,2660–2669.
Jurie, C., Martin, J. F., Listrat, A., Jailler, R., Culioli, J., & Picard, B. (2005). Effect of age andbreed of beef bulls on growth parameters, carcass and muscle characteristics.Animal Science, 80(3), 257–263.
Jurie, C., Ortigues-Marty, I., Picard, B., Micol, D., & Hocquette, J. F. (2006). The separateeffect of the nature of diet and grazing mobility on metabolic potential of musclesfrom Charolais steer. Livestock Science, 104, 182–192.
Jurie, C., Picard, B., & Geay, G. (1998). Influences of the method of housing bulls on theirbody composition and muscle fibre types. Meat Science, 50, 457–469.
Kadim, I. T., Al-Hosni, Y., Mahgoub, O., Al-Marzooqi, W., Khalaf, S. K., Al-Maqbaly, R. S.,et al. (2009). Effect of low voltage electrical stimulation on biochemical and qualitycharacteristics of Longissimus thoracis muscle from one-humped camel (Camelusdromedaries). Meat Science, 82, 77–85.
Kadim, I. T., & Mahgoub, O. (2006). Meat quality and composition of longissimusthoracis from Arabian camel (Camelus dromedarius and Omani beef): A compar-ative study. 1st conference of International Society of Camelid Research andDevelopment (ISOCARD), Al Ain Rotana Hotel, April 15–17 (pp. 161).
Kadim, I. T., Mahgoub, O., Al-Ajmi, D. S., Al-Maqbaly, R. S., Al-Mugheiry, S. M., &Bartolome, D. Y. (2004). The influence of season on quality characteristics of hot-boned beef m. longissimus thoracis. Meat Science, 66, 831–836.
Kadim, I. T., Mahgoub, O., Al-Marzooqi, W., Al-Maqbali, R. S., & Al-Lawati, S. M. (2008).The influence of seasonal temperatures on meat quality characteristics of hot-boned, m. psoas major and minor from goats and sheep.Meat Science, 80, 210–215.
Kadim, I. T., Mahgoub, O., Al-Marzooqi, W., Al-Zadjali, S., Annamalai, K., & Mansour, M. H.(2006). Effects of age on composition and quality of muscle Longissimus thoracis ofthe Omani Arabian camel (Camelus dromedaries). Meat Science, 73(4), 619–662.
Kadim, I. T., Mahgoub, O., Al-Marzooqi, W., Khalf, S. K., Mansour, M. H., Al-Sinani, S. S. H.,et al. (2009). Effects of electrical stimulation on histochemical muscle fibrestaining, quality and composition of camel and cattle longissimus thoracis muscles.Journal of Food Science, 74(1), S44–S52.
Kamoun,M. (1995).Dromedarymeat:production, qualitativeaspects andacceptability fortransformation. Option Mediterraneennes Serie B, Etudes et Recherches, 13, 105–130.
Kassem, M. T., El-Sayed, M. T., & Ahmed, A. (2004). Micostructural characteristics ofArabian camel meat. Journal of Camel Science, 1, 86–95.
Klont, R. E., Brocks, L., & Eikelenboom, G. (1998). Muscle fibre type and meat quality.Meat Science, 49(1), S219–S229.
Kurtu, M. Y. (2004). An assessment of the productivity for meat and carcass yield ofcamel (Camelus dromedarius) and the consumption o f camel meat in the Easternregion of Ethiopia. Tropical Animal Health and Production, 36, 65–76.
Lefaucheur, L. (2010). A second look into fibre typing—Relating to meat quality. MeatScience, 84(2), 257–270.

144 O.M.A. Abdelhadi et al. / Meat Science 90 (2012) 139–144
Mahgoub, O., Kadim, I. T., Al-Saqry, N. M., & Al-Busaidi, R. M. (2004). Effects of bodyweight and sex on carcass tissue distribution in goats. Meat Science, 67, 577–585.
Mancini, R. A., Kim, Y. H., Hunt, M. C., & Lawrence, T. E. (2004). How does lactateenhancement improve beef color stability? Proceedings 50th international congressof meat science and technology, 8–13 August 2004, Helsinki, Finland.
Monin, G. (1990). Facteurs biologiques de qualités de la viande. In R. G. Guillermet, & Y.Geay (Eds.), Croissance des Bovins et Quqlité de la Viande (pp. 177–196). Rennes:ENSAR.
Morton, R. H. (1984). Camels for meat andmilk production in sub-Sahara Africa. Journalof Dairy Science, 67, 1548–1553.
Oury, M. P., Dumont, R., Jurie, C., Hocquette, J. F., & Picard, B. (2010). Specificities of fibrecomposition and metabolism of the rectus abdominis muscle of bovine Charolaiscattle. BMC Biochemistry, 11, 12.
Pérez, P., Maino, M., Guzmgn, R., Vaquero, A., Kobrich, C., & Pokniak, J. (2000). Carcasscharacteristics of Ilamas (Lama glama) reared in Central Chile. Small RuminantResearch, 37, 93–97.
Pette, D., & Staron, R. S. (2001). Transitions of muscle fibre genotypic profile.Histochemistry and Cell Biology, 115, 359–372.
Pettigrew, D. W., & Frieden, C. (1979). Binding of regulatory ligands to rabbit musclephosphofructokinase. A model for nucleotide binding as a function of temperatureand pH. Journal of Biological Chemistry, 254, 1887–1895.
Picard, B., Barboiron, C., Chadeyron, D., & Jurie, C. (2007). Une technique d'électro-phorèse appliquée à la séparation des isoformes de chaînes lourdes de myosine dumuscle squelettique de bovin. Cahier des techniques INRA, 62, 17–24.
Picard, B., & Cassar-Malek, I. (2009). Evidence for expression of IIb myosin heavy chainisoform in some skeletal muscles of Blonde d'Aquitaine bulls.Meat Science, 82, 30–36.
Picard, B., Duris, M. P., & Jurie, C. (1998). Classification of bovine muscle fibres bydifferent histochemical techniques. Histochemistry Journal, 30(7), 473–479.
Renand, G. (1990). Détermination génétique de caractéristiques de carcasses et desviandes. In R. G. Guillermet, & Y. Geay (Eds.), Croissance des Bovins et Qualité de laViande (pp. 31–41). Rennes: ENSAR.
Rhoades, R. D., Andy King, D., Jenschke, B. E., Behrends, J. M., Hively, T. S., & Smith, S. B.(2005). Postmortem regulation of glycolysis by 6-phosphofructokinase in bovineM. Sternocephalicus pars mandibularis. Meat Science, 70, 621–626.
Rose, R. J., Evans, D. L., Knight, P. K., Henckel, P., Cluer, D., & Saltin, B. (1992). Proceedingsof the First International Camel Conference. 2nd–6th Feb. UAE.
Sasaki, K., Mitsumoto, M., & Kawabata, K. (2001). Relation between lipid peroxidationand fat content in Japanese Black beef Longissimus muscle during storage. MeatScience, 59, 407–410.
Sen, A. R., Santra, A., & Kadim, S. A. (2004). Carcass yield, composition and meat qualityattribute of sheep and goat under semiarid conditions. Meat Science, 66, 757–763.
Shariatmadari, R., & Kadivar, M. (2006). Postmortem ageing and freezing of camel meat(a comparative study). 52nd Inter Cong Meat Science (pp. 673–674).
Soltanizadeh, N., Kadivar, M., Keramat, J., & Fazilati, M. (2008). Comparison of fresh beefand camel meat proteolysis during cold storage. Meat Science, 80, 892–895.
Statistical Analysis Systems Institute Inc. (2001). SAS user's guide: Statistics, version 8.2.Cary, NC, USA: SAS Institute Inc.
Talmadge, R. J., & Roy, R. R. (1993). Electrophoretic separation of rat skeletal musclemyosin heavy-chain isoforms. The American Physiological Society, 75, 2337–2340.
Talmant, A., Monin, G., Briand, M., Dadet, M., & Briand, Y. (1986). Activities of metabolicand contractile enzymes in 18 bovine muscles. Meat Science, 18, 23–40.
Trivedi, B., & Danforth, W. H. (1966). Effect of pH on the kinetics of frog musclephosphofructokinase. Journal of Biological Chemistry, 241, 4110–4114.
Vestergaard, M., Oksbjerg, N., & Henchel, P. (2000). Influence of feeding intensity,grazing and finishing feeding on muscle fibre characteristics and meat color ofsemitendinosus, longissimus dorsi and supraspinatus muscle of young bulls. MeatScience, 54(2), 177–185.
Young, S. K., Seok, K. Y., Young, H. S., & Sung, K. L. (2003). Effect of season on color ofHanwoo (Korean native cattle) beef. Meat Science, 63, 509–513.
Yousif, O. Kh., & Babiker, S. A. (1989). The desert camel as a meat animal. Meat Science,26(4), 245–254.




















