
Rabbit Nipple-Search Pheromone Versus Rabbit Mammary Pheromone Revisited
Upload
independentCategory
view
6download
0
1 23
Acta Physiologiae Plantarum ISSN 0137-5881Volume 33Number 4 Acta Physiol Plant (2011)33:1399-1409DOI 10.1007/s11738-010-0674-8
Anatomical and physiologicalcharacteristics relating to ionic relationsin some salt tolerant grasses from the SaltRange, Pakistan
Mansoor Hameed, Muhammad Ashraf &Nargis Naz

1 23
Your article is protected by copyright and
all rights are held exclusively by Franciszek
Górski Institute of Plant Physiology, Polish
Academy of Sciences, Kraków. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

ORIGINAL PAPER
Anatomical and physiological characteristics relating to ionicrelations in some salt tolerant grasses from the Salt Range,Pakistan
Mansoor Hameed • Muhammad Ashraf •
Nargis Naz
Received: 11 September 2010 / Revised: 26 October 2010 / Accepted: 7 December 2010 / Published online: 24 December 2010
� Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2010
Abstract Populations of three salt tolerant forage grasses
(Cynodon dactylon, Imperata cylindrica, and Sporobolus
arabicus) were collected from the salt-affected soils of the
Salt Range and normal non-saline soils of the Faisalabad
region to assess their mechanism of adaptation to saline
stress by determining ion relations and some specific ana-
tomical modifications. The population of S. arabicus from
the Salt Range showed increased growth (root and shoot
length, and root and shoot dry weights) under saline con-
ditions. Salt tolerance in this species was related to struc-
tural modifications such as increased area of root, stem,
leaf blade, and leaf sheath for toxic ion accumulation,
increased vesicular hair density in leaves and aerenchyma
formation in leaf sheath for ion exclusion. Uptake of toxic
ions was high in the Salt Range population of C. dactylon
and salt tolerance was related to ion exclusion through
specific leaf structural modifications such as vesicular
hairs. Salt tolerance in the Salt Range population of I. cyl-
indrica was mainly associated with restricted uptake of
toxic Na? and Cl- at root level, and accumulation of toxic
ions via increased succulence in leaf blades and leaf
sheaths in addition to some excretion of toxic ions through
leaf sheath aerenchyma.
Keywords Adaptation � Aerenchyma � Salt tolerance �Succulence � Vesicular hairs
Introduction
Plants growing on naturally salt-affected soils must have
evolved a multitude of morpho-anatomical and physio-
logical adaptive characteristics in view of considerable
length of time they have been exposed to high selection
pressure of the habitat like high salinity and aridity
(Ashraf 1994; Hameed and Ashraf 2008). The Salt
Range in Pakistan is a conspicuous site where soil in
most places is impregnated with high content of sodium
chloride. The full description of the Salt Range has been
given elsewhere (Hameed and Ashraf 2008). However,
the plant species whether dicot or monocot inhabiting
particularly salt-affected soils of the Salt Range are
supposed to be salt tolerant. Of a number of grass species
recorded from the Range, Sporobolus arabicus Boiss,
Cynodon dactylon (L.) Pers, and Imperata cylindrica (L.)
Raeuschel have considerable importance as they provide
valuable forage to sheep and goats being reared in the
Salt Range.
Specific anatomical and physiological modifications in
plants exposed to stressful environments may enable them
to thrive well on such environments. Although plants use a
variety of physiological phenomena to counteract salt
stress, regulation of ion homeostasis is one of the premier
physiological processes operating in plants exposed to salt
stress (Zhu 2003). This includes selective ion uptake
(Flowers and Colmer 2008), accumulation of toxic ions,
partially in terms of increased succulence (Hameed et al.
2009), and excretion of such unwanted toxic ions (Ramadan
and Flowers 2004; Naz et al. 2009).
Communicated by J. Franklin.
M. Hameed (&) � M. Ashraf � N. Naz
Department of Botany, University of Agriculture,
Faisalabad, Pakistan
e-mail: [email protected]
M. Ashraf
Department of Botany and Microbiology,
King Saud University, Riyadh, Saudi Arabia
123
Acta Physiol Plant (2011) 33:1399–1409
DOI 10.1007/s11738-010-0674-8
Author's personal copy

In view of all these, it was hypothesized that the eco-
types of the three grass species inhabiting the salt-affected
soils of the Salt Range differ considerably in taking up and
accumulating toxic inorganic ions such as Na? and Cl- and
excretion of such ions through leaf sheath or leaf surface.
The differences in their salinity tolerance may not be only
due to specific physiological adaptations, but also due to
their structural modifications in root, stem, leaf blade and
leaf sheath. This is perhaps due to some of the specific
anatomical features they have developed so far during
the length of time they have been growing there. Thus, the
major objective of the present study was to elucidate the
mechanism of ion regulation in salt tolerant ecotypes of
three potential grasses and relate these physiological dis-
tinctions with their anatomical adaptations particularly in
their leaves and roots.
Materials and methods
The present investigation was carried out to asses the adapt-
ability in relation to ionic balance of three potential salt tol-
erant grasses from the Salt Range, Pakistan. A population of
C. dactylon (L.) Pers. was collected from the edges of a
natural salt lake, the Uchhali Lake, in the Salt Range, Punjab
(pH 6.62, ECe 19.92 dS m-1). This species was in direct
contact of highly saline water of the lake. A population of
I. cylindrica (L.) Raeuschel was collected from some distance
from the edges of the salt lake, which is seasonally inundated
by diluted less saline waters during the rainy season (pH 7.38,
ECe 15.40 dS m-1). However, that of S. arabicus Boiss was
collected from the valley near the foothills, which was greatly
affected by the salt deposition as a result of dissolved salts
from the exposed hills (pH 8.12, ECe 34.36 dS m-1). The
ecotypes of all three grasses were also collected as checks
(control population) from normal non-saline habitats within
the Faisalabad region (pH 9.20, ECe 1.82 dS m-1 for
C. dactylon, pH 7.70, ECe 2.29 dS m-1 for I. cylindrica, and
pH 8.86, ECe 1.52 dS m-1 for S. arabicus).
The soil taken from the rhizosphere of each grass popu-
lation from each habitat was analyzed for physico-chemical
characteristics. Saturation percentage, pH and electrical
conductivity were determined by preparing saturated soil
paste of each sample. The soil pH and ECe were determined
using a combined pH/electrical conductivity meter. Inor-
ganic ions such as Na?, K? and Ca2? were determined with
a flame photometer and Cl- with a chloride meter.
Populations of both ecotypes of all three grasses were
grown in a mixture of sand, peat and clay mixed in equal
quantities for a period of 6 months under the climatic
conditions of Faisalabad (average day and night tempera-
tures 37 ± 3 and 24 ± 3�C, respectively, photoperiod
11–12 h, relative humidity 45.9–58.6%). Ramets of equal
size with three mature tillers were detached from each
plant and grown in half-strength Hoagland’s nutrient
solution (Hoagland and Arnon 1950) in containers of fiber
glass (25 L capacity) till their establishment in hydropon-
ics. The containers were aerated 12 h daily by air pumps.
Two salt levels, 0 (Hoagland’s nutrient solution with no
salt added) and 150 mM of NaCl in Hoagland’s nutrient
solution, were established in hydroponics. The experiment
was arranged in a completely randomized design (CRD)
with three factors (grasses, ecotypes and salinity levels)
and 12 replicates. After a growth of 60 days, plants were
carefully collected from the hydroponics for determining
physiological and anatomical characteristics.
For determining tissue ionic content of plants, dried
ground plant material (leaves and roots) was digested in
concentrated H2SO4 following Wolf (1982). For the
determination of excreted ions from leaf sheath and leaf
blade surface, 12 samples of the topmost fully expanded
leaf along with leaf sheath from the main tiller of all three
grass species were carefully separated, each sample
immediately washed in 20 ml of deionized water, and leaf
wash determined for the excreted ions. A flame photometer
was used to determine cations (Na?, K? and Ca2?) and a
chloride meter used to determine Cl-.
For studying stem and leaf sheath anatomy, a 2 cm piece
from the base of 3rd internode, for leaf blade anatomy a
2 cm piece from the base of top fully developed leaf, and
for root anatomy a 2 cm piece of the thickest adventitious
root from the root-culm junction were selected. All plant
samples so selected were first fixed in formalin acetic
alcohol fixative (formaldehyde 10%, acetic acid 5%, eth-
anol 50% and distilled water 35%) for 48 h and subse-
quently transferred to acetic alcohol solution (acetic acid
25% and ethanol 75%) for long-term storage. Free-hand
sectioning slides were prepared by a series of dehydrations
in ethanol using a standard double-stained technique of
safranine and fast green stains following Ruzin (1999).
Measurements were taken with a light microscope, using
an ocular micrometer, which was calibrated with a stage
micrometer. Micrographs of stained sections were taken
with a digital camera fitted on a stereo-microscope.
Data for anatomical characteristics were recorded using
all the 12 replicates. The analysis of variance (ANOVA)
for each attribute was computed using the MSTAT Com-
puter Program (MSTAT Development Team, 1989). LSD
at 5% level of probability was calculated to test the dif-
ferences among mean values (Steel et al. 1997).
Results
Ecotypes of all three grass species, C. dactylon, I. cylind-
rica and S. arabicus, responded differently in relation to
1400 Acta Physiol Plant (2011) 33:1399–1409
123
Author's personal copy
both tissue ionic content and their anatomical characteris-
tics to saline stress.
Soil analysis
The Salt Range populations of all three grasses were col-
lected from highly salt-affected soils. However, S. arabicus
was collected from the salt-affected soils with highest
salinity (ECe 34.36 dS m-1), and C. dactylon and I. cyl-
indrica from relatively less saline habitats.
Growth attributes
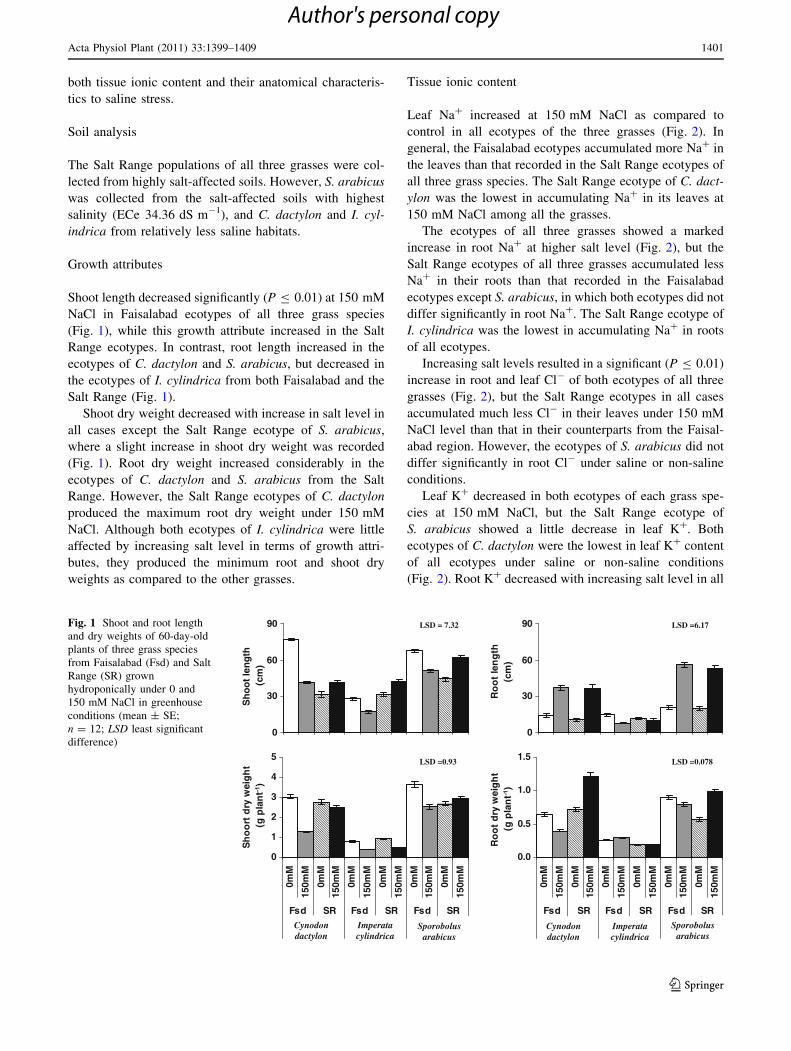
Shoot length decreased significantly (P B 0.01) at 150 mM
NaCl in Faisalabad ecotypes of all three grass species
(Fig. 1), while this growth attribute increased in the Salt
Range ecotypes. In contrast, root length increased in the
ecotypes of C. dactylon and S. arabicus, but decreased in
the ecotypes of I. cylindrica from both Faisalabad and the
Salt Range (Fig. 1).
Shoot dry weight decreased with increase in salt level in
all cases except the Salt Range ecotype of S. arabicus,
where a slight increase in shoot dry weight was recorded
(Fig. 1). Root dry weight increased considerably in the
ecotypes of C. dactylon and S. arabicus from the Salt
Range. However, the Salt Range ecotypes of C. dactylon
produced the maximum root dry weight under 150 mM
NaCl. Although both ecotypes of I. cylindrica were little
affected by increasing salt level in terms of growth attri-
butes, they produced the minimum root and shoot dry
weights as compared to the other grasses.
Tissue ionic content
Leaf Na? increased at 150 mM NaCl as compared to
control in all ecotypes of the three grasses (Fig. 2). In
general, the Faisalabad ecotypes accumulated more Na? in
the leaves than that recorded in the Salt Range ecotypes of
all three grass species. The Salt Range ecotype of C. dact-
ylon was the lowest in accumulating Na? in its leaves at
150 mM NaCl among all the grasses.
The ecotypes of all three grasses showed a marked
increase in root Na? at higher salt level (Fig. 2), but the
Salt Range ecotypes of all three grasses accumulated less
Na? in their roots than that recorded in the Faisalabad
ecotypes except S. arabicus, in which both ecotypes did not
differ significantly in root Na?. The Salt Range ecotype of
I. cylindrica was the lowest in accumulating Na? in roots
of all ecotypes.
Increasing salt levels resulted in a significant (P B 0.01)
increase in root and leaf Cl- of both ecotypes of all three
grasses (Fig. 2), but the Salt Range ecotypes in all cases
accumulated much less Cl- in their leaves under 150 mM
NaCl level than that in their counterparts from the Faisal-
abad region. However, the ecotypes of S. arabicus did not
differ significantly in root Cl- under saline or non-saline
conditions.
Leaf K? decreased in both ecotypes of each grass spe-
cies at 150 mM NaCl, but the Salt Range ecotype of
S. arabicus showed a little decrease in leaf K?. Both
ecotypes of C. dactylon were the lowest in leaf K? content
of all ecotypes under saline or non-saline conditions
(Fig. 2). Root K? decreased with increasing salt level in all
0
30
60
90
Sh
oo
t le
ng
th(c
m)
0
30
60
90
Ro
ot
len
gth
(cm
)
0
1
2
3
4
5
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
Fsd SR Fsd SR Fsd SR
Cda Icy Sar
Sh
oo
rt d
ry w
eig
ht
(g p
lan
t-1)
0.0
0.5
1.0
1.5
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
Fsd SR Fsd SR Fsd SR
Cda Icy Sar
Ro
ot
dry
wei
gh
t(g
pla
nt-1
)
Cynodon dactylon
Cynodon dactylon
Imperata cylindrica
Imperata cylindrica
Sporobolus arabicus
Sporobolus arabicus
71.6=DSL23.7=DSL
870.0=DSL39.0=DSL
Fig. 1 Shoot and root length
and dry weights of 60-day-old
plants of three grass species
from Faisalabad (Fsd) and Salt
Range (SR) grown
hydroponically under 0 and
150 mM NaCl in greenhouse
conditions (mean ± SE;
n = 12; LSD least significant
difference)
Acta Physiol Plant (2011) 33:1399–1409 1401
123
Author's personal copy
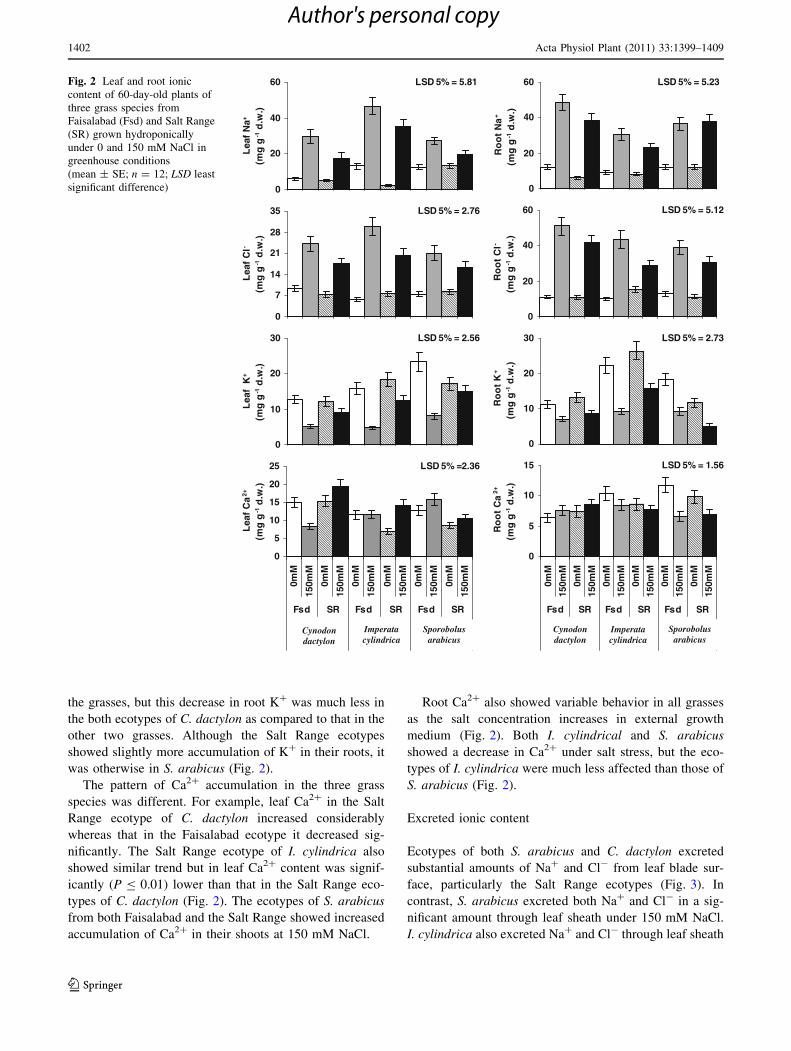
the grasses, but this decrease in root K? was much less in
the both ecotypes of C. dactylon as compared to that in the
other two grasses. Although the Salt Range ecotypes
showed slightly more accumulation of K? in their roots, it
was otherwise in S. arabicus (Fig. 2).
The pattern of Ca2? accumulation in the three grass
species was different. For example, leaf Ca2? in the Salt
Range ecotype of C. dactylon increased considerably
whereas that in the Faisalabad ecotype it decreased sig-
nificantly. The Salt Range ecotype of I. cylindrica also
showed similar trend but in leaf Ca2? content was signif-
icantly (P B 0.01) lower than that in the Salt Range eco-
types of C. dactylon (Fig. 2). The ecotypes of S. arabicus
from both Faisalabad and the Salt Range showed increased
accumulation of Ca2? in their shoots at 150 mM NaCl.
Root Ca2? also showed variable behavior in all grasses
as the salt concentration increases in external growth
medium (Fig. 2). Both I. cylindrical and S. arabicus
showed a decrease in Ca2? under salt stress, but the eco-
types of I. cylindrica were much less affected than those of
S. arabicus (Fig. 2).
Excreted ionic content
Ecotypes of both S. arabicus and C. dactylon excreted
substantial amounts of Na? and Cl- from leaf blade sur-
face, particularly the Salt Range ecotypes (Fig. 3). In
contrast, S. arabicus excreted both Na? and Cl- in a sig-
nificant amount through leaf sheath under 150 mM NaCl.
I. cylindrica also excreted Na? and Cl- through leaf sheath
LSD 5% = 5.23
0
20
40
60
Ro
ot
Na
+
(mg
g-1 d
.w.)
LSD 5% = 5.81
0
20
40
60
Lea
f N
a+
(mg
g-1 d
.w.)
LSD 5% = 5.12
0
20
40
60
Ro
ot
Cl-
(mg
g-1 d
.w.)
LSD 5% = 2.76
0
7
14
21
28
35
Lea
f C
l-
(mg
g-1 d
.w.)
LSD 5% = 2.73
0
10
20
30
Ro
ot
K+
(mg
g-1 d
.w.)
LSD 5% = 2.56
0
10
20
30
Lea
f K
+
(mg
g-1 d
.w.)
LSD 5% = 1.56
0
5
10
15
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
Fsd SR Fsd SR Fsd SR
Cynodondactylon
Imperatacylindrica
Sporobolusarabicus
Ro
ot
Ca
2+
(mg
g-1 d
.w.)
LSD 5% =2.36
0
5
10
15
20
25
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
Fsd SR Fsd SR Fsd SR
Cynodondactylon
Imperatacylindrica
Sporobolusarabicus
Lea
f C
a2+
(mg
g-1 d
.w.)
Cynodon dactylon
Imperata cylindrica
Sporobolus arabicus
Cynodon dactylon
Imperata cylindrica
Sporobolus arabicus
Fig. 2 Leaf and root ionic
content of 60-day-old plants of
three grass species from
Faisalabad (Fsd) and Salt Range
(SR) grown hydroponically
under 0 and 150 mM NaCl in
greenhouse conditions
(mean ± SE; n = 12; LSD least
significant difference)
1402 Acta Physiol Plant (2011) 33:1399–1409
123
Author's personal copy
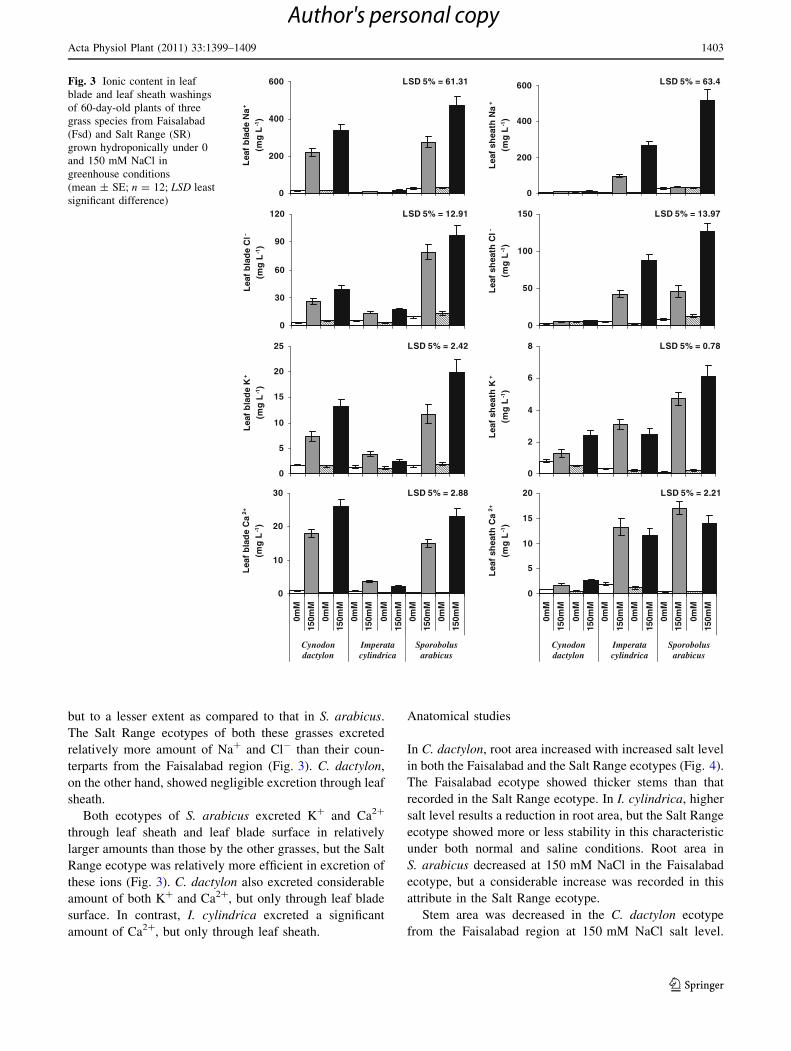
but to a lesser extent as compared to that in S. arabicus.
The Salt Range ecotypes of both these grasses excreted
relatively more amount of Na? and Cl- than their coun-
terparts from the Faisalabad region (Fig. 3). C. dactylon,
on the other hand, showed negligible excretion through leaf
sheath.
Both ecotypes of S. arabicus excreted K? and Ca2?
through leaf sheath and leaf blade surface in relatively
larger amounts than those by the other grasses, but the Salt
Range ecotype was relatively more efficient in excretion of
these ions (Fig. 3). C. dactylon also excreted considerable
amount of both K? and Ca2?, but only through leaf blade
surface. In contrast, I. cylindrica excreted a significant
amount of Ca2?, but only through leaf sheath.
Anatomical studies
In C. dactylon, root area increased with increased salt level
in both the Faisalabad and the Salt Range ecotypes (Fig. 4).
The Faisalabad ecotype showed thicker stems than that
recorded in the Salt Range ecotype. In I. cylindrica, higher
salt level results a reduction in root area, but the Salt Range
ecotype showed more or less stability in this characteristic
under both normal and saline conditions. Root area in
S. arabicus decreased at 150 mM NaCl in the Faisalabad
ecotype, but a considerable increase was recorded in this
attribute in the Salt Range ecotype.
Stem area was decreased in the C. dactylon ecotype
from the Faisalabad region at 150 mM NaCl salt level.
LSD 5% = 61.31
0
200
400
600
Lea
f b
lad
e N
a+
(mg
L-1)
LSD 5% = 63.4
0
200
400
600
Lea
f sh
eath
Na
+
(mg
L-1)
LSD 5% = 12.91
0
30
60
90
120
Lea
f b
lad
e C
l-
(mg
L-1)
LSD 5% = 13.97
0
50
100
150
Lea
f sh
eath
Cl-
(mg
L-1)
LSD 5% = 2.42
0
5
10
15
20
25
Lea
f b
lad
e K
+
(mg
L-1)
LSD 5% = 0.78
0
2
4
6
8
Lea
f sh
eath
K+
(mg
L-1)
LSD 5% = 2.88
0
10
20
30
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
Cynodondactylon
Im peratacylindrica
Sporobolusarabicus
Lea
f b
lad
e C
a2+
(mg
L-1)
LSD 5% = 2.21
0
5
10
15
20
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
0mM
150m
M
Cynodondactylon
Im peratacylindrica
Sporobolusarabicus
Lea
f sh
eath
Ca
2+
(mg
L-1)
Cynodon dactylon
Cynodon dactylon
Imperata cylindrica
Imperata cylindrica
Sporobolus arabicus
Sporobolus arabicus
Fig. 3 Ionic content in leaf
blade and leaf sheath washings
of 60-day-old plants of three
grass species from Faisalabad
(Fsd) and Salt Range (SR)
grown hydroponically under 0
and 150 mM NaCl in
greenhouse conditions
(mean ± SE; n = 12; LSD least
significant difference)
Acta Physiol Plant (2011) 33:1399–1409 1403
123
Author's personal copy
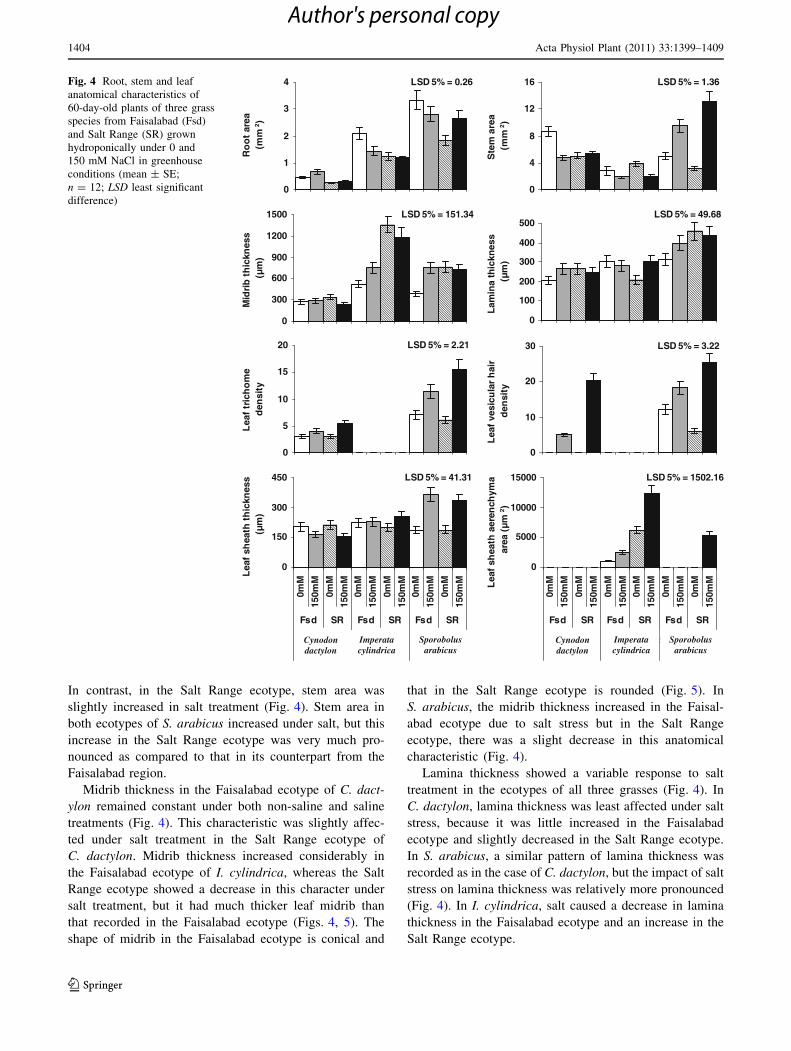
In contrast, in the Salt Range ecotype, stem area was
slightly increased in salt treatment (Fig. 4). Stem area in
both ecotypes of S. arabicus increased under salt, but this
increase in the Salt Range ecotype was very much pro-
nounced as compared to that in its counterpart from the
Faisalabad region.
Midrib thickness in the Faisalabad ecotype of C. dact-
ylon remained constant under both non-saline and saline
treatments (Fig. 4). This characteristic was slightly affec-
ted under salt treatment in the Salt Range ecotype of
C. dactylon. Midrib thickness increased considerably in
the Faisalabad ecotype of I. cylindrica, whereas the Salt
Range ecotype showed a decrease in this character under
salt treatment, but it had much thicker leaf midrib than
that recorded in the Faisalabad ecotype (Figs. 4, 5). The
shape of midrib in the Faisalabad ecotype is conical and
that in the Salt Range ecotype is rounded (Fig. 5). In
S. arabicus, the midrib thickness increased in the Faisal-
abad ecotype due to salt stress but in the Salt Range
ecotype, there was a slight decrease in this anatomical
characteristic (Fig. 4).
Lamina thickness showed a variable response to salt
treatment in the ecotypes of all three grasses (Fig. 4). In
C. dactylon, lamina thickness was least affected under salt
stress, because it was little increased in the Faisalabad
ecotype and slightly decreased in the Salt Range ecotype.
In S. arabicus, a similar pattern of lamina thickness was
recorded as in the case of C. dactylon, but the impact of salt
stress on lamina thickness was relatively more pronounced
(Fig. 4). In I. cylindrica, salt caused a decrease in lamina
thickness in the Faisalabad ecotype and an increase in the
Salt Range ecotype.
LSD 5% = 0.26
0
1
2
3
4
Ro
ot
area
(m
m2 )
LSD 5% = 1.36
0
4
8
12
16
Ste
m a
rea
(mm
2 )
LSD 5% = 49.68
0
100
200
300
400
500
Lam
ina
thic
knes
s (µ
m)
LSD 5% = 151.34
0
300
600
900
1200
1500
Mid
rib
th
ickn
ess
(µm
)
LSD 5% = 2.21
0
5
10
15
20
Lea
f tr
ich
om
e d
ensi
ty
LSD 5% = 3.22
0
10
20
30
Lea
f ve
sicu
lar
hai
r d
ensi
ty
LSD 5% = 41.31
0
150
300
450
0mM
150m
M
0mM
150m
M0m
M
150m
M
0mM
150m
M0m
M
150m
M
0mM
150m
M
Fsd SR Fsd SR Fsd SR
Cynodondactylon
Imperatacylindrica
Sporobolusarabicus
Lea
f sh
eath
th
ickn
ess
(µm
)
LSD 5% = 1502.16
0
5000
10000
15000
0mM
150m
M
0mM
150m
M
0mM
150m
M0m
M
150m
M
0mM
150m
M
0mM
150m
M
Fsd SR Fsd SR Fsd SR
Cynodondactylon
Imperatacylindrica
Sporobolusarabicus
Lea
f sh
eath
aer
ench
yma
area
(µm
2 )
Cynodon dactylon
Cynodon dactylon
Imperata cylindrica
Imperata cylindrica
Sporobolus arabicus
Sporobolus arabicus
Fig. 4 Root, stem and leaf
anatomical characteristics of
60-day-old plants of three grass
species from Faisalabad (Fsd)
and Salt Range (SR) grown
hydroponically under 0 and
150 mM NaCl in greenhouse
conditions (mean ± SE;
n = 12; LSD least significant
difference)
1404 Acta Physiol Plant (2011) 33:1399–1409
123
Author's personal copy

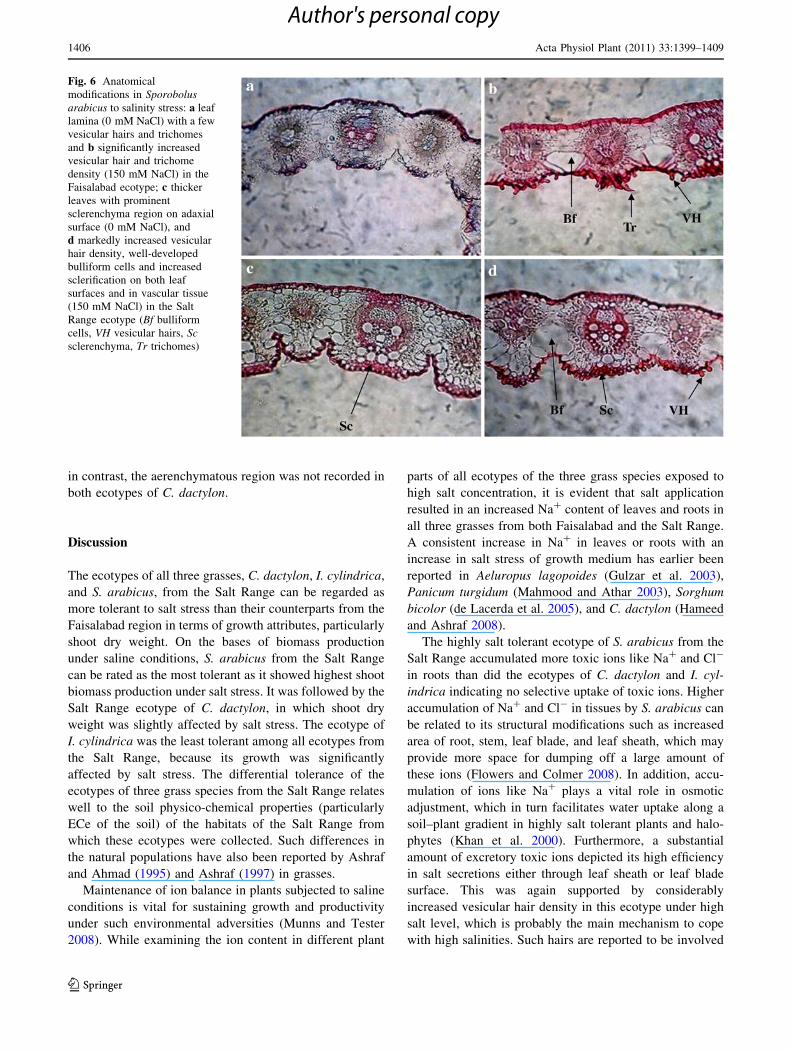
Number of trichomes per unit area increased in both
ecotypes of C. dactylon under salt level (Fig. 4). In the Salt
Range ecotype, salt stress caused a twofold increase in
trichome density as compared to that in non-stress treat-
ment. In S. arabicus, the Faisalabad ecotype showed no
visible change in trichome density under salt stress, but the
Salt Range ecotype showed a marked increase in this
parameter (Figs. 6, 7).
Vesicular (bladder or bicellular microhairs) hairs
increased markedly in both C. dactylon and S. arabicus
under salt stress (Fig. 4). In both ecotypes of C. dactylon,
vesicular hairs were not recorded under non-saline treat-
ment, but their density was increased by about fourfold in
the Salt Range ecotype under salt level as compared to that
in the Faisalabad ecotype (Figs. 4, 8). In I. cylindrica, no
such excretory structures were recorded. In S. arabicus,
vesicular hair density increased greatly in both ecotypes,
but in the Salt Range ecotype, this increase was about
fivefold under salt treatment (Figs. 4, 6).
Leaf sheath thickness decreased in the ecotypes of
C. dactylon from both Faisalabad and the Salt range under
salt stress (Fig. 4). Sheath thickness remained unaffected in
the Faisalabad ecotype of I. cylindrica, but in the Salt
Range ecotype it increased to some extent. However, this
parameter increased greatly in both ecotypes of S. arabicus
due to salt level.
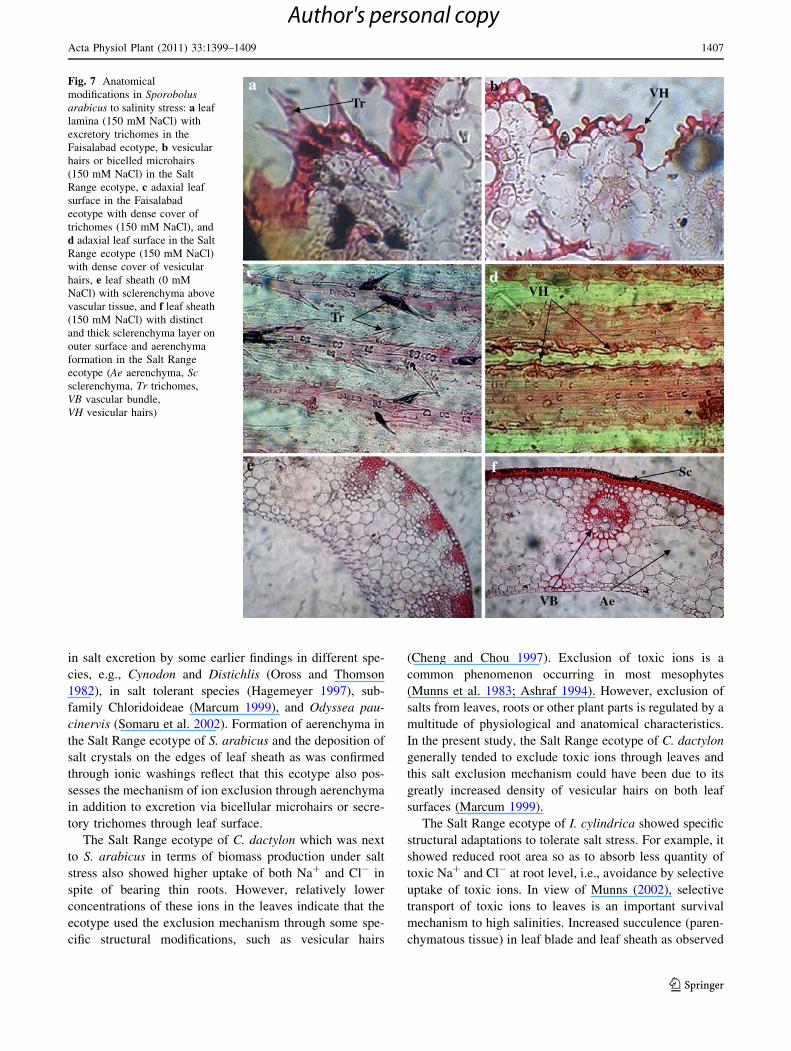
A well-developed aerenchyma was recorded in the leaf
sheaths of both ecotypes of I. cylindrica (Figs. 4, 5). The
Faisalabad ecotype showed slight increase in aerenchy-
matous area under salt stress, but the Salt Range ecotype
showed a twofold increase (Fig. 4), i.e., from 6,172 lm2 at
0 mM to 12,427 lm2 at 150 mM NaCl. In S. arabicus,
aerenchyma formation was recorded in the ecotype from
the Salt Range only under salt stress (Figs. 4, 7). However,
Fig. 5 Anatomical
modifications in Imperatacylindrica to salinity stress:
a thick round and succulent leaf
midrib (0 mM NaCl) and
b increased sclerification in
midrib (150 mM NaCl) in the
Salt Range ecotype; c conical
shape leaf midrib (0 mM NaCl)
and d somewhat round midrib
(150 mM NaCl) in the
Faisalabad ecotype; e leaf
sheath with intact parenchyma
(0 mM NaCl), and f well-
developed aerenchyma
(150 mM NaCl) in leaf sheath
in the Salt Range ecotype
(Ae aerenchyma, P parenchyma,
VB vascular bundles, Scsclerenchyma)
Acta Physiol Plant (2011) 33:1399–1409 1405
123
Author's personal copy
in contrast, the aerenchymatous region was not recorded in
both ecotypes of C. dactylon.
Discussion
The ecotypes of all three grasses, C. dactylon, I. cylindrica,
and S. arabicus, from the Salt Range can be regarded as
more tolerant to salt stress than their counterparts from the
Faisalabad region in terms of growth attributes, particularly
shoot dry weight. On the bases of biomass production
under saline conditions, S. arabicus from the Salt Range
can be rated as the most tolerant as it showed highest shoot
biomass production under salt stress. It was followed by the
Salt Range ecotype of C. dactylon, in which shoot dry
weight was slightly affected by salt stress. The ecotype of
I. cylindrica was the least tolerant among all ecotypes from
the Salt Range, because its growth was significantly
affected by salt stress. The differential tolerance of the
ecotypes of three grass species from the Salt Range relates
well to the soil physico-chemical properties (particularly
ECe of the soil) of the habitats of the Salt Range from
which these ecotypes were collected. Such differences in
the natural populations have also been reported by Ashraf
and Ahmad (1995) and Ashraf (1997) in grasses.
Maintenance of ion balance in plants subjected to saline
conditions is vital for sustaining growth and productivity
under such environmental adversities (Munns and Tester
2008). While examining the ion content in different plant
parts of all ecotypes of the three grass species exposed to
high salt concentration, it is evident that salt application
resulted in an increased Na? content of leaves and roots in
all three grasses from both Faisalabad and the Salt Range.
A consistent increase in Na? in leaves or roots with an
increase in salt stress of growth medium has earlier been
reported in Aeluropus lagopoides (Gulzar et al. 2003),
Panicum turgidum (Mahmood and Athar 2003), Sorghum
bicolor (de Lacerda et al. 2005), and C. dactylon (Hameed
and Ashraf 2008).
The highly salt tolerant ecotype of S. arabicus from the
Salt Range accumulated more toxic ions like Na? and Cl-
in roots than did the ecotypes of C. dactylon and I. cyl-
indrica indicating no selective uptake of toxic ions. Higher
accumulation of Na? and Cl- in tissues by S. arabicus can
be related to its structural modifications such as increased
area of root, stem, leaf blade, and leaf sheath, which may
provide more space for dumping off a large amount of
these ions (Flowers and Colmer 2008). In addition, accu-
mulation of ions like Na? plays a vital role in osmotic
adjustment, which in turn facilitates water uptake along a
soil–plant gradient in highly salt tolerant plants and halo-
phytes (Khan et al. 2000). Furthermore, a substantial
amount of excretory toxic ions depicted its high efficiency
in salt secretions either through leaf sheath or leaf blade
surface. This was again supported by considerably
increased vesicular hair density in this ecotype under high
salt level, which is probably the main mechanism to cope
with high salinities. Such hairs are reported to be involved
Fig. 6 Anatomical
modifications in Sporobolusarabicus to salinity stress: a leaf
lamina (0 mM NaCl) with a few
vesicular hairs and trichomes
and b significantly increased
vesicular hair and trichome
density (150 mM NaCl) in the
Faisalabad ecotype; c thicker
leaves with prominent
sclerenchyma region on adaxial
surface (0 mM NaCl), and
d markedly increased vesicular
hair density, well-developed
bulliform cells and increased
sclerification on both leaf
surfaces and in vascular tissue
(150 mM NaCl) in the Salt
Range ecotype (Bf bulliform
cells, VH vesicular hairs, Scsclerenchyma, Tr trichomes)
1406 Acta Physiol Plant (2011) 33:1399–1409
123
Author's personal copy
in salt excretion by some earlier findings in different spe-
cies, e.g., Cynodon and Distichlis (Oross and Thomson
1982), in salt tolerant species (Hagemeyer 1997), sub-
family Chloridoideae (Marcum 1999), and Odyssea pau-
cinervis (Somaru et al. 2002). Formation of aerenchyma in
the Salt Range ecotype of S. arabicus and the deposition of
salt crystals on the edges of leaf sheath as was confirmed
through ionic washings reflect that this ecotype also pos-
sesses the mechanism of ion exclusion through aerenchyma
in addition to excretion via bicellular microhairs or secre-
tory trichomes through leaf surface.
The Salt Range ecotype of C. dactylon which was next
to S. arabicus in terms of biomass production under salt
stress also showed higher uptake of both Na? and Cl- in
spite of bearing thin roots. However, relatively lower
concentrations of these ions in the leaves indicate that the
ecotype used the exclusion mechanism through some spe-
cific structural modifications, such as vesicular hairs
(Cheng and Chou 1997). Exclusion of toxic ions is a
common phenomenon occurring in most mesophytes
(Munns et al. 1983; Ashraf 1994). However, exclusion of
salts from leaves, roots or other plant parts is regulated by a
multitude of physiological and anatomical characteristics.
In the present study, the Salt Range ecotype of C. dactylon
generally tended to exclude toxic ions through leaves and
this salt exclusion mechanism could have been due to its
greatly increased density of vesicular hairs on both leaf
surfaces (Marcum 1999).
The Salt Range ecotype of I. cylindrica showed specific
structural adaptations to tolerate salt stress. For example, it
showed reduced root area so as to absorb less quantity of
toxic Na? and Cl- at root level, i.e., avoidance by selective
uptake of toxic ions. In view of Munns (2002), selective
transport of toxic ions to leaves is an important survival
mechanism to high salinities. Increased succulence (paren-
chymatous tissue) in leaf blade and leaf sheath as observed
Fig. 7 Anatomical
modifications in Sporobolusarabicus to salinity stress: a leaf
lamina (150 mM NaCl) with
excretory trichomes in the
Faisalabad ecotype, b vesicular
hairs or bicelled microhairs
(150 mM NaCl) in the Salt
Range ecotype, c adaxial leaf
surface in the Faisalabad
ecotype with dense cover of
trichomes (150 mM NaCl), and
d adaxial leaf surface in the Salt
Range ecotype (150 mM NaCl)
with dense cover of vesicular
hairs, e leaf sheath (0 mM
NaCl) with sclerenchyma above
vascular tissue, and f leaf sheath
(150 mM NaCl) with distinct
and thick sclerenchyma layer on
outer surface and aerenchyma
formation in the Salt Range
ecotype (Ae aerenchyma, Scsclerenchyma, Tr trichomes,
VB vascular bundle,
VH vesicular hairs)
Acta Physiol Plant (2011) 33:1399–1409 1407
123
Author's personal copy

in this ecotype might have enhanced the vacuolar volume so
as to provide greater area for dumping off toxic ions such as
Na? and Cl-, as reported by Hameed et al. (2009). Leaf or
stem succulence is an adaptive feature, which contributes to
the regulation of internal ion concentrations in many halo-
phytes (Short and Colmer 1999), although leaf succulence is
a rare phenomenon in monocots (Hameed and Ashraf 2008).
In addition, the ecotype showed an increased aerenchyma in
leaf sheath under saline stress. This could be an important
feature to excrete toxic ions, as well as in overcoming gas-
eous exchange problems during physiological drought
caused by salinity stress.
In conclusion, anatomical modifications to overcome
high salinities were very specific not only in the grass
species but also in the ecotypes. The Salt Range ecotype of
S. arabicus had increased area of root, stem, leaf blade, and
leaf sheath. These characteristics may enhance the ability
of the ecotype to accumulate salts. In addition, increased
vesicular hair density and aerenchyma formation in leaf
sheath of this ecotype under salt stress seem useful in salt
excretion. The Salt Range ecotype of C. dactylon totally
depended on ion exclusion through specific structural
modifications such as vesicular hairs. The Salt Range
ecotype of I. cylindrica showed restricted uptake of toxic
Na? and Cl- at root level in addition to increased succu-
lence in leaf blade and leaf sheath for dumping off the toxic
ions. The most efficient way for maintaining growth and
survival in adverse environmental conditions seems to be
excessive excretion of toxic ions from leaf sheath and leaf
surface, i.e., via microhairs and leaf sheath aerenchyma.
Perhaps, this requires low amount of energy for its survival
and this might be utilized in the growth. On the other hand,
substantial amount of energy is required in selective uptake
of toxic ions and also for ion sequestering in non-meta-
bolically active areas such as vacuoles.
References
Ashraf M (1994) Breeding for salinity tolerance in plants. Crit Rev
Plant Sci 13:17–42
Ashraf M (1997) Changes in soluble carbohydrates and soluble
proteins in three arid-zone grass species under salt stress. J Trop
Agric 74:234–237
Ashraf M, Ahmad H (1995) Response of three arid zone grasses to
salt and waterlogging. Arid Soil Res Rehabil 9:137–154
Cheng KT, Chou CH (1997) Ecotypic variation of Imperatacylindrica populations in Taiwan. I. Morphological and molec-
ular evidences. Bot Bull Acad Sin 38:215–223
de Lacerda CF, Cambraia J, Oliva MA, Ruiz HA (2005) Changes in
growth and in solute concentrations in sorghum leaves and roots
during salt stress recovery. Environ Exp Bot 54:69–76
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New
Phytol 179:945–963
Gulzar S, Khan MA, Ungar IA (2003) Effects of salinity on growth,
ionic content, and plant–water status of Aeluropus lagopoides.
Commun Soil Sci Plant Anal 34:1657–1668
Hagemeyer J (1997) Salt. In: Prasard MNV (ed) Plant ecophysiology.
Wiley, Toronto, ON, pp 173–206
Hameed M, Ashraf M (2008) Physiological and biochemical adap-
tations of Cynodon dactylon (L.) Pers. from the Salt Range
(Pakistan) to salinity stress. Flora 203:683–694
Hameed M, Ashraf M, Naz N (2009) Anatomical adaptations to
salinity in cogon grass [Imperata cylindrica (L.) Raeuschel]
from the Salt Range, Pakistan. Plant Soil 322:229–238
Fig. 8 Anatomical
modifications in Cynodondactylon to salinity stress: a leaf
(0 mM NaCl) in the Salt Range
ecotype, and b increased
vesicular hair density on both
leaf surfaces (150 mM NaCl) in
the Salt Range ecotype, c stem
of the Salt Range ecotype
(0 mM NaCl), and d stem of the
Salt Range ecotype (150 mM
NaCl) with sclerification
(Bf bulliform cells,
Sc sclerenchyma, VB vascular
bundles, VH vesicular hairs)
1408 Acta Physiol Plant (2011) 33:1399–1409
123
Author's personal copy

Hoagland DR, Arnon DI (1950) The water culture method for
growing plants without soil. Circular No. 347. University of
California Agricultural Experimental Station, Berkeley, CA,
pp 1–39
Khan MA, Gul B, Weber DJ (2000) Germination responses to
Salicornia rubra to temperature and salinity. J Arid Environ
45:207–221
Mahmood S, Athar HR (2003) Germination and growth of Panicumturgidum provenance under saline conditions. Pak J Biol Sci
6:164–166
Marcum KB (1999) Salinity tolerance mechanisms of grasses in the
subfamily Chloridoideae. Crop Sci 39:1153–1160
Munns R (2002) Comparative physiology of salt and water stress.
Plant Cell Environ 25:239–250
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu
Rev Plant Biol 59:651–681
Munns R, Greenway H, Kirst GO (1983) Halotolerant eukaryotes. In:
Lange OL, Nobel PS, Osmond CB, Ziegler H, Ziegler H (eds)
Encyclopaedia of plant physiology, vol 12C: Physiological plant
ecology III. Springer, Berlin, pp 59–135
Naz N, Hameed M, Wahid A, Arshad M, Ahmad MSA (2009)
Patterns of ion excretion and survival in two stoloniferous arid
zone grasses. Physiol Plant 135:185–195
Oross WJ, Thomson WW (1982) The ultrastructure of the salt glands
of Cynodon and Distichlis (Poaceae). Am J Bot 69:939–949
Ramadan T, Flowers T (2004) Effects of salinity and benzyl adenine
on development and function of microhairs of Zea mays L.
Planta 219:639–648
Ruzin SE (1999) Plant microtechnique and microscopy. Oxford
University Press, New York
Short DC, Colmer TD (1999) Salt tolerance in the halophyte
Halosarcia pergranulata subsp. Pergranulata. Ann Bot
83:207–213
Somaru R, Naidoo Y, Naidoo G (2002) Morphology and ultrastruc-
ture of the leaf salt glands of Odyssea paucinervis (Stapf)
(Poaceae). Flora 197:67–75
Steel RGD, Torrie JH, Dickie DA (1997) Principles and procedures of
statistics—a biometric approach, 3rd edn. McGraw-Hill Pub-
lishing Company, Toronto
Wolf B (1982) An improved universal extracting solution and its use
for diagnosing soil fertility. Commun Soil Sci Plant Anal
13:1005–1033
Zhu JK (2003) Regulation of ion homeostasis under salt stress. Curr
Opin Plant Biol 6:441–445
Acta Physiol Plant (2011) 33:1399–1409 1409
123
Author's personal copy
Copyright © 2022 FDOKUMEN




















