
Analysis of biomarkers after intramuscular injection of Vaxfectin ®-formulated hCMV gB plasmid DNA
9
Vaccine 27 (2009) 7409–7417 Contents lists available at ScienceDirect Vaccine journal homepage: www.elsevier.com/locate/vaccine Analysis of biomarkers after intramuscular injection of Vaxfectin ® -formulated hCMV gB plasmid DNA Adrián Vilalta ∗ , Mark Shlapobersky, Qun Wei, Rodrick Planchon, Alain Rolland, Sean Sullivan Vical Incorporated, 10390 Pacific Center Ct., San Diego, CA 92121, United States article info Article history: Received 28 April 2009 Received in revised form 10 August 2009 Accepted 21 August 2009 Available online 5 September 2009 Keywords: pDNA vaccine CMV Cationic lipid adjuvant Biomarker abstract Cationic lipids have been used as delivery systems to enhance the performance of vaccines and immunotherapeutics. However, little is known about the effect of administration of cationic lipid- formulated vaccines on gene expression. This study used DNA microarrays (39,000 transcripts) to characterize early changes in gene expression patterns in mouse muscle 1 and 2 days after intramus- cular (IM) injection of a hCMV gB plasmid DNA (pDNA) vaccine formulated with the cationic lipid system Vaxfectin ® ; gene expression profiles were compared to those obtained after IM injection of pDNA in PBS. Analysis of the DNA microarray data indicated that approximately 1% of the represented transcripts were modulated at least 2-fold compared to the PBS samples at both time points. Functional analysis of the modulated genes revealed that transcripts involved in antigen processing and presentation, apoptosis and the Toll-like receptor pathway were significantly enriched. In addition, confirmation of local and systemic modulation of subsets of biomarkers was achieved using Real-Time PCR and Cytometric Bead Assays. Time course and magnitude of cellular infiltration (F4/80+ and CD11b+ cells) to the injection site was changed in response to formulation of hCMVgB pDNA with Vaxfectin ® . Since the expression level of the pDNA-encoded transgene in the muscle was not affected by formulation Vaxfectin ® mechanism of action is expected to rely primarily on modulation of immune pathways and not on an increase in trans- fection of the antigen-encoding pDNA. Taken together, these data help explain the Vaxfectin ® -dependent robust enhancement of immune responses. © 2009 Elsevier Ltd. All rights reserved. 1. Introduction Expression profiling is a promising new approach to under- standing the underlying mechanisms of action of vaccines and adjuvants. A growing body of work [1–9] indicates that the use of DNA arrays can help dissect the influences of antigen, adju- vant and route of administration on vaccine performance. Recent published work by Mosca et al. [9] identified biomarkers asso- ciated with adjuvants MF59, CpG and Alum in the absence of a co-administered antigen. To achieve this, analysis of expression profiles from mouse muscles was conducted after intramuscular injection using whole-mouse genome DNA microarray analysis. The results identified unique profiles of cytokine, cytokine recep- tor as well as other biomarkers consistent with the adjuvant’s biological activity. Work by Purcell et al. [5] provides a compre- hensive example of the molecular response to vaccination with non-adjuvanted plasmid DNA (pDNA) in a commercial pDNA vac- cine against infectious hematopoietic necrosis virus (IHNV). The results showed that intramuscular administration of anti-IHNV ∗ Corresponding author. Tel.: +1 858 646 1108; fax: +1 858 646 1150. E-mail address: [email protected] (A. Vilalta). vaccine in fish results in strong changes in gene expression rel- ative to administration of pDNA with no antigen encoded. The results also showed that the IHNV pDNA vaccine increased type I IFN system across multiple tissues, which is the basis for effective anti-viral immunity. Microarray results from Byon et al. [10] com- pared immune responses for pDNA vs. recombinant protein-based anti-viral hemorrhagic septicemia virus (VHSV) vaccine. The results showed that differences in the nature and magnitude of B- and T-cell responses from vaccination of Japanese flounder fish with either recombinant glycoprotein or pDNA VHSV vaccine correlated with the up-regulation of different sets of stress and immune- related genes. The adjuvant activity of cationic lipid systems has been doc- umented extensively [11–18]. In spite of the large amount of immunological data reported, the mechanism of action of these delivery systems has only been partially elucidated. The experimental results presented here sought to identify expression markers that could provide insight into the biological activity of the cationic lipid system Vaxfectin ® . Characterization of the early, local biological response to intramuscular vaccina- tion with a Vaxfectin ® -formulated pDNA vaccine in mice was achieved using several techniques including DNA microarrays, Real-Time PCR, Cytometric Bead Assays and immunohistochem- 0264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2009.08.075
Transcript of Analysis of biomarkers after intramuscular injection of Vaxfectin ®-formulated hCMV gB plasmid DNA

Ah
AV
a
ARRAA
KpCCB
1
saovpccpiTtbhncr
0d
Vaccine 27 (2009) 7409–7417
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
nalysis of biomarkers after intramuscular injection of Vaxfectin®-formulatedCMV gB plasmid DNA
drián Vilalta ∗, Mark Shlapobersky, Qun Wei, Rodrick Planchon, Alain Rolland, Sean Sullivanical Incorporated, 10390 Pacific Center Ct., San Diego, CA 92121, United States
r t i c l e i n f o
rticle history:eceived 28 April 2009eceived in revised form 10 August 2009ccepted 21 August 2009vailable online 5 September 2009
eywords:DNA vaccineMVationic lipid adjuvantiomarker
a b s t r a c t
Cationic lipids have been used as delivery systems to enhance the performance of vaccines andimmunotherapeutics. However, little is known about the effect of administration of cationic lipid-formulated vaccines on gene expression. This study used DNA microarrays (39,000 transcripts) tocharacterize early changes in gene expression patterns in mouse muscle 1 and 2 days after intramus-cular (IM) injection of a hCMV gB plasmid DNA (pDNA) vaccine formulated with the cationic lipid systemVaxfectin®; gene expression profiles were compared to those obtained after IM injection of pDNA in PBS.Analysis of the DNA microarray data indicated that approximately 1% of the represented transcripts weremodulated at least 2-fold compared to the PBS samples at both time points. Functional analysis of themodulated genes revealed that transcripts involved in antigen processing and presentation, apoptosisand the Toll-like receptor pathway were significantly enriched. In addition, confirmation of local and
systemic modulation of subsets of biomarkers was achieved using Real-Time PCR and Cytometric BeadAssays. Time course and magnitude of cellular infiltration (F4/80+ and CD11b+ cells) to the injection sitewas changed in response to formulation of hCMVgB pDNA with Vaxfectin®. Since the expression level ofthe pDNA-encoded transgene in the muscle was not affected by formulation Vaxfectin® mechanism ofaction is expected to rely primarily on modulation of immune pathways and not on an increase in trans-fection of the antigen-encoding pDNA. Taken together, these data help explain the Vaxfectin®-dependentrobust enhancement of immune responses.© 2009 Elsevier Ltd. All rights reserved.
. Introduction
Expression profiling is a promising new approach to under-tanding the underlying mechanisms of action of vaccines anddjuvants. A growing body of work [1–9] indicates that the usef DNA arrays can help dissect the influences of antigen, adju-ant and route of administration on vaccine performance. Recentublished work by Mosca et al. [9] identified biomarkers asso-iated with adjuvants MF59, CpG and Alum in the absence of ao-administered antigen. To achieve this, analysis of expressionrofiles from mouse muscles was conducted after intramuscular
njection using whole-mouse genome DNA microarray analysis.he results identified unique profiles of cytokine, cytokine recep-or as well as other biomarkers consistent with the adjuvant’siological activity. Work by Purcell et al. [5] provides a compre-ensive example of the molecular response to vaccination withon-adjuvanted plasmid DNA (pDNA) in a commercial pDNA vac-ine against infectious hematopoietic necrosis virus (IHNV). Theesults showed that intramuscular administration of anti-IHNV
∗ Corresponding author. Tel.: +1 858 646 1108; fax: +1 858 646 1150.E-mail address: [email protected] (A. Vilalta).
vaccine in fish results in strong changes in gene expression rel-ative to administration of pDNA with no antigen encoded. Theresults also showed that the IHNV pDNA vaccine increased typeI IFN system across multiple tissues, which is the basis for effectiveanti-viral immunity. Microarray results from Byon et al. [10] com-pared immune responses for pDNA vs. recombinant protein-basedanti-viral hemorrhagic septicemia virus (VHSV) vaccine. The resultsshowed that differences in the nature and magnitude of B- andT-cell responses from vaccination of Japanese flounder fish witheither recombinant glycoprotein or pDNA VHSV vaccine correlatedwith the up-regulation of different sets of stress and immune-related genes.
The adjuvant activity of cationic lipid systems has been doc-umented extensively [11–18]. In spite of the large amount ofimmunological data reported, the mechanism of action of thesedelivery systems has only been partially elucidated.
The experimental results presented here sought to identifyexpression markers that could provide insight into the biologicalactivity of the cationic lipid system Vaxfectin®. Characterizationof the early, local biological response to intramuscular vaccina-tion with a Vaxfectin®-formulated pDNA vaccine in mice wasachieved using several techniques including DNA microarrays,Real-Time PCR, Cytometric Bead Assays and immunohistochem-
264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2009.08.075

7410 A. Vilalta et al. / Vaccine 27 (2009) 7409–7417
istry. Taken together, these data provide insight into Vaxfectin®
adjuvant activity and creates a knowledge base that can beused to the development of more potent and well-balancedvaccines.
2. Materials and methods
2.1. Plasmid DNA and formulation
Plasmid VR6365 has been described extensively elsewhere [19].Briefly, the pDNA encodes a secreted form of the human CMV(hCMV) gB (aminoacids 1–713, GenBank accession AY646879) thatlacks the transmembrane and cytoplasmic domains of the full-length glycoprotein to facilitate secretion, but retains the nativesignal sequence. Sequence encoding hCMV gB was introduced intoexpression plasmid VR10551; gene expression is under the con-trol of the constitutive hCMV immediate early promoter-enhancer.pDNA was manufactured under cGMP using standard bacterialfermentation followed by alkaline lysis; pDNA was further puri-fied using a two-step chromatographic process. pDNA used in thisstudy contained less than 0.01 EU endotoxin/�g pDNA and less than0.05 �g RNA/�g pDNA.
Vaxfectin® consists of an equimolar mixture of VC1052 ((±)-N-(3-aminopropyl)-N,N-dimethyl-2,3-bis (myristoleyloxy)-1-propa-naminium bromide) and DPyPE (diphytanoylphosphatidyl-ethanolamine). Preparation of Vaxfectin® and its formulationswere first described by Hartikka et al. [11]. Briefly, both VC1052and DPyPE were resuspended in chloroform, mixed in 1:1 molarratio, aliquoted into vials, and dried to create Vaxfectin® reagentdry lipid film. On the day of injection, the lipid film vials wereresuspended in 1 mL 0.9% saline and diluted if necessary. pDNAwas prepared in 0.9% saline, 20 mM sodium phosphate, pH 7.2.pDNA was formulated with Vaxfectin® by gently streaming thelipid into pDNA of equal volume. All the required doses wereprepared by formulating at 0.2–0.5 mg/mL range and dilutingdown to lower concentration as required. The final DNA: cationiclipid molar ratio for formulations was 4:1.
2.2. Animal procedures
All animal procedures were reviewed and approved by Vical’sInstitutional Animal Care and Use Committee (IACUC), and com-plied with the standards set forth in the Guide for the Care andUse of Laboratory Animals (U.S. Department of Health Services,National Institutes of Health; Bethesda, MD). Female BALB/c mice6–8 weeks old (Jackson Laboratories; Bar Harbor, MN) were used.VR6365 plasmid was administered either unformulated (PBS) orformulated with Vaxfectin®. Mice received a single administra-tion of 10 �g of the pDNA (±Vaxfectin®) as bilateral, intramuscular(rectus femoris) injections; injected muscles were collected at 1,2 and 7 days post-injection. Collected tissues were transferredto plastic tubes and either snap-frozen in liquid nitrogen (DNAmicroarray analysis) or preserved in RNALater (Applied Biosys-tems/Ambion; Austin, TX) according to manufacturer’s directions.Tissues collected for immunohistochemistry analysis were pre-served using a 2-methyl butane/OCT embedding method (seeSection 2.7).
2.3. DNA expression arrays
Microarray data were collected at Expression Analysis (Durham,NC) using the GeneChip® Mouse Genome 430 2.0 Array(Affymetrix; Santa Clara, CA). These arrays contain probes forapproximately 39,000 transcripts and variants. Before target pro-duction, the quality and quantity of each RNA sample was assessedusing a 2100 BioAnalyzer (Agilent; Santa Clara, CA). Target was
prepared and hybridized according to the “Affymetrix Techni-cal Manual”. Total RNA (10 �g) was converted into cDNA usingReverse Transcriptase (Invitrogen; Carlsbad, CA) and a modi-fied oligo(dT)24 primer that contains T7 promoter sequences(GenSet; Paris, France). After first strand synthesis, residual RNAwas degraded by the addition of RNaseH and a double-strandedcDNA molecule was generated using DNA Polymerase I and DNALigase. The cDNA was then purified and concentrated using aphenol:chloroform extraction followed by ethanol precipitation.The cDNA products were incubated with T7 RNA Polymerase andbiotinylated ribonucleotides using an in vitro Transcription kit(Enzo Diagnostics). One-half of the cRNA product was purifiedusing an RNeasy column (Qiagen; Valencia, CA) and quantifiedwith a spectrophotometer. The cRNA target (20 �g) was incu-bated at 94 ◦C for 35 min in fragmentation buffer (Tris, MgOAc,KOAc). The fragmented cRNA was diluted in hybridization buffer(MES, NaCl, EDTA, Tween 20, Herring Sperm DNA, Acetylated BSA)containing biotin-labeled OligoB2 and Eukaryotic HybridizationControls (Affymetrix). The hybridization cocktail was denaturedat 99 ◦C for 5 min, incubated at 45 ◦C for 5 min and then injectedinto a GeneChip® cartridge. The GeneChip® array was incubatedat 42 ◦C for at least 16 h in a rotating oven at 60 rpm. GeneChip®
was washed with a series of non-stringent (25 ◦C) and strin-gent (50 ◦C) solutions containing variable amounts of MES, Tween20 and SSPE. The microarrays were then stained with strep-tavidin phycoerythrin and the fluorescent signal was amplifiedusing a biotinylated antibody solution. Fluorescent images weredetected in a GeneChip® Scanner 3000 and expression data wereextracted using the MicroArray Suite 5.0 software (Affymetrix).All GeneChip®s were scaled to a median intensity setting of500.
2.4. Expression array data analysis
Muscles were collected 1 and 2 days (n = 3 for each treatmentand time point) after vaccination. GeneChip® arrays were used todetermine differences in transcript levels between unformulated(PBS) and Vaxfectin®-formulated VR6365 pDNA in 24 h and 48 htissue samples. In an attempt to focus on the most promisingof the modulated transcripts, we deemed a change significantif p ≤ 0.05 and the fold change ≥2. Modulated features wereedited to remove redundancy. GeneChip® data were analyzed byExpression Analysis (Durham, NC) using the Permutation Analysisfor Differential Expression (PADE) method. Briefly, the two-groupcomparison analysis is conducted on normalized expression valuesthat are individually transformed using the base 2 logarithm of therelevant expression index. A floor of 1.0 is used to avoid negativetransformed values (log2(max(1.0,Signal))). After computing themean for each group using the log transformed signal values, themean is then converted back to the original probeset signal units.Averaging on the logarithm scale and then inverse transforming(also known as the geometric averaging) provides a more robustestimate of overall expression that is less impacted by outliers orskewed expression levels relative to a simple arithmetic averageof the raw expression values from each array. Raw fold change iscalculated as the simple ratio of overall expression values from thetwo groups. The higher overall expression is divided by the loweroverall expression. If the baseline group expression is higher, thenfold change is designated to have a negative value. PADE is a specificimplementation of permutation testing dealing with the very largedimensional aspect (transcripts) to each individual sample (array).For more details consult Expression Analysis web site (http://www.expressionanalysis.com/pdf/PADE TechNote 2005.pdf). ProteinCluster analysis was performed using bioinformatics tools atthe DAVID 2.0 website (http://david.abcc.ncifcrf.gov/; NIAID;Bethesda, MD) [20].

A. Vilalta et al. / Vaccine 27
2.5. Real-Time PCR analysis of muscle RNA samples
2.5.1. RNA isolationTotal RNA was isolated from muscle samples collected (n = 5) 1,
2 and 7 days after vaccination using QIAGEN RNeasy Midi Kit (QIA-GEN, Valencia, CA) following manufacturer’s recommendations.
Single strand cDNA synthesis. Reverse transcription was doneusing total RNA (5 �g) as template and High-Capacity cDNA ArchiveKit (Applied Biosystems; Foster City, CA) reagents.
Biomarker transcript levels. Relative transcript levels of a sub-set of immune-related up-regulated genes (CD274, Cxcl9, Igtp,IFN-�, IL-4, IL-6, Irgm, MyD88, Tgtp, TRIF, TLR2, TLR3 and TLR9)were determined using commercially available TaqMan® GeneExpression Assays (Applied Biosystems). Analysis was performedusing the ABI Prism® 7900HT Sequence Detection System (AppliedBiosystems) and cycling conditions suggested by the manufac-turer. A comparative method was used to determine relativequantitation as outlined in Applied Biosystems User Bulletin# 2(http://www3.appliedbiosystems.com/cms/groups/mcb support/documents/generaldocuments/cms 040980.pdf). Briefly, Ct valuesfor target gene amplification were normalized by subtracting theCt values for the house keeping gene GAPDH using equation: Ct(target gene) − Ct (GAPDH) = �Ct. No biases were detected in thelevels of GAPDH. The target gene stimulated �Ct was subtractedfrom the naïve tissue �Ct to calculate the fold change in geneexpression: �Ct (stimulated) − �Ct (naïve) = ��Ct. Fold increasesin gene expression were calculated using the following equation:2−��Ct is fold change in expression.
2.5.2. Plasmid-encoded antigen expressionRNA samples from muscle were assayed for hCMV gB transcript-
specific sequences using a Real-Time PCR assay as described [21]with normalization to GAPDH. Data are presented as a comparisonto transcript levels in naïve tissue. This allows for an estimate of thecontribution of tissue damage caused by injection to the changesin gene expression.
2.6. Systemic cytokine levels
Systemic levels of CCL2, CCL3, Cxcl9, IL-2, IL-4, IL-5, IL-6, IL-10, IL12p70, IFN-� and TNF-� in serum were evaluated usingCytometric Bead Array (CBA) flex sets (BD Pharmingen; La Jolla,CA). Flow cytometric analysis was performed at Flow CytometryResearch Core facility (VA Hospital; San Diego, CA) using FAC-SCanto flow cytometer (BD Biosciences; Mountain View, CA). Datawere acquired and analyzed using BD FACSArray software (BD Bio-sciences).
2.7. Histological examination of injected muscle tissue
Injected muscles were immersed in OCT embedding (VWRScientific; San Francisco, CA) compound and frozen using dryice/2-methyl butane slurry. Samples were kept at −70 ◦C until sec-tioned on a cryostat. All immuno-staining experiments includedboth negative and positive controls. Sections were acetone fixedand washed in PBS; endogenous peroxidases were removed byimmersing in 0.03% H2O2. Endogenous biotin was removed byincubating first with 0.1% avidin in PBS and then with 0.01%biotin in PBS following by blocking with 1% bovine serum albu-min/PBS. Slides were incubated for 30 min with the followingantibodies diluted in 1% BSA: biotinylated F4/80, biotinylatedCD45/B220, biotinylated CD11b/Mac-1 and biotinylated CD3e(BD Pharmingen). Streptavidin-labeled HRP (Jackson ImmunoRe-search; West Grove, PA) was added followed by substratedevelopment with the AEC (amino ethyl carbozole) kit (VectorLaboratories; Burlingame, CA) and a hematoxylin nuclear counter-
(2009) 7409–7417 7411
stain. All incubations were followed by three wash steps with PBS(pH 7.1).
2.8. Statistics
Statistical analysis of DNA microarray data was done as outlinedin Section 2.4. Analysis of Real-Time PCR and CBA data was carriedout using Minitab 15 (State College, PA); statistical relevance wasdetermined using un-paired Student’s t-test.
3. Results
3.1. Modulation of pDNA-encoded hCMV gB antigen
hCMV gB expression results as determined by Real-Time PCR aresummarized in Table 1. Normalized Ct values indicate that hCMV gBwas expressed at about the same level in VR6365-PBS and VR6365-Vaxfectin® muscle samples 1, 2 and 7 days after injection.
3.2. Modulation of gene expression by VR6365-Vaxfectin®
Analysis of the 24 h VR6365-Vaxfectin® muscle samples (n = 3per treatment) indicates that the abundance of 510 transcripts wassignificantly increased (≥2×; p ≤ 0.05) compared to the VR6365-PBS samples; 115 transcripts were up-regulated at least 5-fold.Approximately 522 genes were significantly (p ≤ 0.05) down-regulated 2-fold or more and seventeen 5-fold or more. The meanfold change ranged from 1.1 to 27.6. Analysis of samples collected48 h after injection showed 541 transcripts up-regulated at least 2-fold (p ≤ 0.05); about 136 were overexpressed at least 5-fold. About337 transcripts were down-regulated at least 2-fold while 22 tran-scripts showed a 5-fold or more reduction in expression (p ≤ 0.05).The mean fold change at 48 h ranged from 1.1 to 47.6. Fig. 1 summa-rizes expression and associated gene function data for transcriptswith the highest level of modulation (at least 10-fold increase ordecrease) after treatment with VR6365-Vaxfectin® 24 h and 48 hafter injection.
3.3. Vaxfectin® modulates inflammatory and innate immunitypathways at the site of injection
Functional analysis of modulated genes (1032 genes; mod-ulation by at least 2-fold; p ≤ 0.05) identified three groupsof genes significantly enriched. These were antigen processing
and presentation [Kyoto Encyclopedia of Genes and Genomes(KEGG) mm04612, p = 7.9 × 10−6], Toll-like receptor pathway(KEGG mm04620, p = 9.6 × 10−3) and apoptosis (KEGG mm04210,p = 1.2 × 10−3); the results are summarized in Fig. 2. Furtheranalysis identified an overlapping group of 87 genes associatedTable 1Effect of formulation on pDNA-encoded gene expression after vaccineadministration.
gB Ct GAPDH Ct Fold
Day 1PBS 28.6 ± 0.2 18.1 ± 0.1 1.0Vaxfectin® 28.1 ± 0.2 18.7 ± 0.1 2.1 ± 1.1
Day 2PBS 30.8 ± 1.6 17.6 ± 0.1 1.0Vaxfectin® 31.1 ± 0.5 18.4 ± 0.1 1.4 ± 1.4
Day 7PBS 31.4 ± 1.2 17.6 ± 0.2 1.0Vaxfectin® 31.0 ± 0.7 17.9 ± 0.2 1.6 ± 1.7
Ct values obtained using hCMV gB Real-Time PCR assay described in Section 2. RNAsamples from five injected muscles were used for the analysis. Fold changes overpDNA-PBS were calculated using method described in Section 2.5.

7412 A. Vilalta et al. / Vaccine 27 (2009) 7409–7417
Fig. 1. Gene modulation by hCMV gB pDNA-Vaxfectin® . Microarray data of genesmodulated by pDNA-Vaxfectin® was analyzed for fold change ratios (VR6365-Vaxfectin®/VR6365-PBS) as described in Section 2. mRNA transcripts with thehap
w(6wtr
Fig. 2. Functional analysis of microarray transcription profiles in mouse muscle.Functional analysis of hCMV gB pDNA-Vaxfectin®-modulated genes (1032 genes;fold change ratio ≥2 and p ≤ 0.05 in at least one time point) was done as describedin Section 2. Cluster analyses for antigen presentation and processing, apoptosis andTLR pathways are presented for 24 h and 48 h as indicated. Gene accession numbers
ighest modulation (fold change ratio ≥10 and p ≤ 0.05 in at least one time point)re presented. Gene accession numbers, functions and log2 ratios color scale arerovided.
ith the Gene Ontology (GO) “immune system process” clusterGO:0002376, p = 7.6 × 10−15). Transcript levels for Cxcl9, IL-4, IL-
, IFN-�, CD274 (PD-L1), Igtp, Irgm, MyD88, Tgtp, TLR2 andTLR3ere confirmed using Real-Time PCR analysis. These specificranscripts were chosen based on their relevance to immuneesponses as determined by the functional analysis described
and statistical significance of each category of genes are presented. Fold changes(VR6365-Vaxfectin®/VR6365-PBS) are expressed as log2 ratios using the indicatedcolor scale.
above and the high level of modulation after injection of VR6365pDNA formulated with Vaxfectin®. No significant change in TRIFor TLR9 mRNA was observed by microarray analysis. However,due to their potential relevance in adjuvant activated immuneresponse transcript levels for TRIF and TLR9 were assayed byReal-Time PCR. Real-Time PCR results showed that there wasno significant difference in TRIF expression between treatmentswhereas TLR9 transcripts were up-regulated about 3-fold byVaxfectin®-formulated VR6365 compared to VR6365-PBS. Tran-script level results for selected biomarkers as determined byReal-Time PCR are summarized in Figs. 3 and 4. Real-Time PCRdata confirmed DNA chip results of the selected biomarkers 24 hand 48 h after vaccine administration. Day 7 Real-Time PCR wasadded to determine the duration of the Vaxfectin®-dependentstimulation.
3.4. Systemic response to pDNA-Vaxfectin®
Serum was obtained from the same mice used for Real-TimePCR tissue collection to evaluate the systemic effects caused byvaccination. Levels of CCL2, CCL3, Cxcl9, IL-2, IL-4, IL-5, IL-6,IL-10, IL12p70, IFN-� and TNF-� in serum (Fig. 5) were evalu-ated by multiplex Cytometric Bead Arrays (CBA). The cytokineswith a significant increase in systemic levels compared to pre-bleed levels for any of the treatments are shown (Cxcl9, CCL2,IL-6 and IFN-�). Serum levels of Cxcl9, CCL2 and IFN-� in theVR6365-Vaxfectin® groups were statistically higher than theVR6365-PBS groups 24 h after vaccine administration. No signif-icant difference in IL-6 levels for VR6365-Vaxfectin® comparedto the VR6365-PBS groups was observed. However, a signifi-
cant difference in IL-6 levels in the serum was observed 48 hrsafter VR6365-Vaxfectin® administration compared to VR6365-PBS. The levels of all four cytokines returned to baseline byday 7.
A. Vilalta et al. / Vaccine 27 (2009) 7409–7417 7413
Fig. 3. Real-Time PCR confirmation of increased expression of TLR-pathway molecules. Transcript levels of key elements of the TLR-pathway were evaluated in injectedmuscle as described in Section 2. Panel (A) shows the level of adaptor molecule MyD88 was increased in response to Vaxfectin® while the level of TRIF was unaffected byformulation; panel (B) shows the mRNA for Toll-like receptors 2, 3 and 9 increased after vaccination with hCMV gB pDNA-Vaxfectin® . Symbols represent the arithmetic meanof five independent samples; error bars represent the corresponding S.E.M.
3.5. Expression modulation of Th1/Th2 and inflammation markers
Microarray data were interrogated for relative expression lev-els for Th1/Th2 markers (CD80, CD86, IL-2, IL-4, IL-5, IL-6, IL-10,IL-12p70, IL-13, and IFN-�). These data showed that IL-6 wasover expressed in the muscle in response to VR6365-Vaxfectin®
(4.6-fold, p = 0.023) 24 h after administration and remained high(3.2-fold, p = 0.004) 48 h after injection; Real-Time PCR data con-firmed overexpression at these two time points and furtherindicated that IL-6 mRNA levels in the injected muscle returned tobaseline by day 7 (Fig. 3). IL-6 levels in the serum (Fig. 5) were alsoelevated in response to pDNA-Vaxfectin®. In addition, Real-TimePCR data showed that IFN-� mRNA levels were increased in theVR6365-Vaxfectin® samples (10-fold over VR6365-PBS) while IL-4mRNA increased by approximately 2-fold in response to Vaxfectin®
compared to the VR6365-PBS control. Systemic levels of IFN-�were also elevated (5-fold over VR6365-PBS) in the Vaxfectin®
groups while systemic levels of IL-4 were unchanged relative to theVR6365-PBS controls. Other Th1/Th2 and inflammation markerswere unaffected by formulation as determined by DNA microarrayand Cytometric Bead Assays (data not shown).
3.6. Vaxfectin® enhances recruitment of CD11b+ and F4/80+ cellsto the injection site
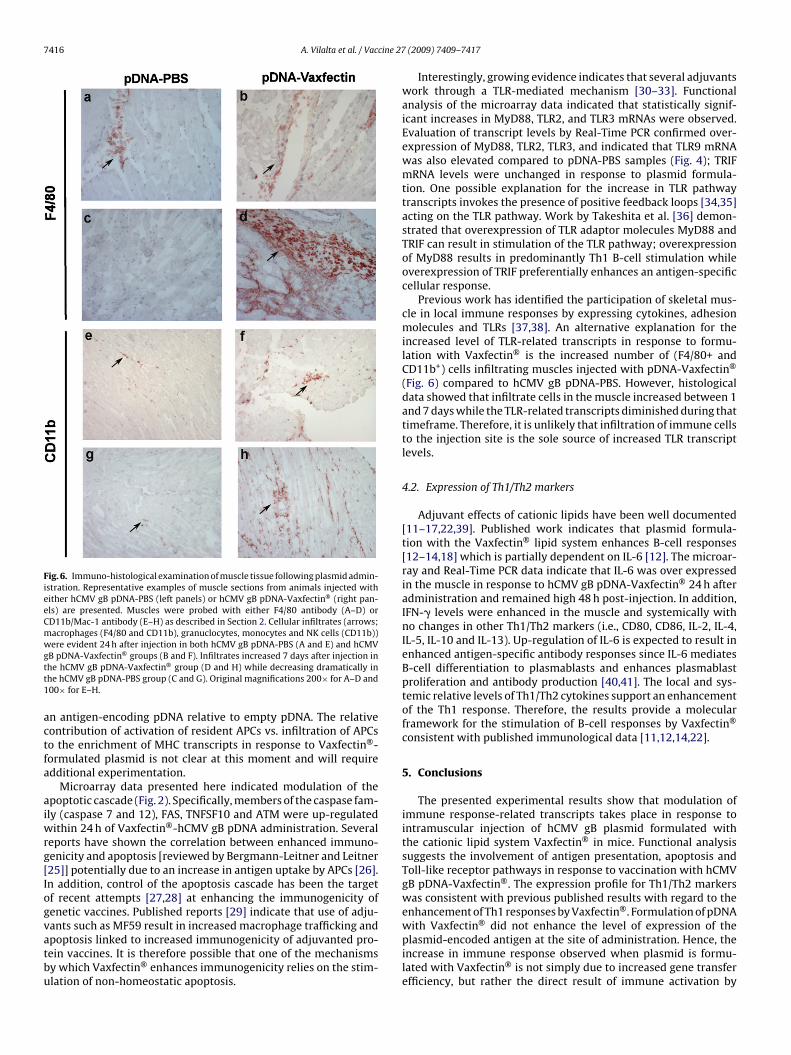
Immunohistochemistry of injected muscles showed increasedinfiltration of CD11b+ (macrophages, granulocytes, monocytes andNK cells) and F4/80+ (macrophages) cells in response to Vaxfectin®
formulation (Fig. 6). Cellular infiltrates in the VR6365-Vaxfectin®
samples increased between days 1 and 7 while in VR6365-PBS sam-ples infiltrates seemed to decrease between days 1 and 7. A smallincrease in B220+ (B-) cells was observed in VR6365-Vaxfectin®-injected muscle tissue (data not shown); no CD3e+ (T-) cells weredetected in injected muscle tissue.

7414 A. Vilalta et al. / Vaccine 27 (2009) 7409–7417
Fig. 4. Real-Time PCR confirmation of overexpression of a subset of biomarkers. Transcript levels for four cytokines (A) and four other biomarkers (B) identified through theDNA microarray were evaluated using quantitative PCR as described in Section 2. Symbols represent the arithmetic mean of five independent samples; error bars representthe corresponding S.E.M.

A. Vilalta et al. / Vaccine 27 (2009) 7409–7417 7415
Fig. 5. Vaxfectin® formulation results in transient systemic elevation of pro-inflammatory cytokines. Cytokines levels in the serum were investigated using Cytometric BeadAssays as described in Section 2. Cytokines found to change in response to Vaxfectin® are presented here. Symbols represent the geometric mean; error bars represent thecorresponding S.E.M.
4. Discussion
4.1. Experimental approach
Several publications have demonstrated that immunogenicity ofpDNA vaccines can be significantly enhanced through formulationwith Vaxfectin® [11,12,14,22]; this increase in immunogenicitywas confirmed for the hCMV gB pDNA vaccine (VR6365) used inthis study (data not shown). Conceptually, Vaxfectin® can enhanceimmunogenicity of pDNA vaccines by enhancing pDNA uptakeand/or modulating immune responses through an adjuvant activ-ity. Work presented here explored the contributions of these twofactors by evaluating the level of expression of a plasmid-encodedantigen in the injected tissue and the patterns of endogenous geneexpression at the site of vaccine administration. To address theeffect on pDNA transfection, pDNA-encoded hCMV gB transcriptlevels in the muscle were determined using a Real-Time PCR assaythat discriminates between hCMV gB pDNA and mRNA. Transgeneexpression data indicated that formulation of hCMV gB pDNA withVaxfectin® did not increase the level of hCMV gB transcript inthe injected muscle 1, 2 or 7 days after administration relativeto the hCMV gB pDNA-PBS control. These findings are in agree-ment with previous published data [11] indicating that no changesin the expression of the reporter �-galactosidase could be foundbetween PBS and Vaxfectin®-formulated �-galactosidase pDNAone day post-administration. The muscle gene transfer resultstherefore suggest that the reported increase in immune responsesto hCMV gB pDNA formulated with Vaxfectin® is not likely dueto an increase in expression of the plasmid-encoded antigen(s) at
the site of administration. Effect of adjuvants on the expressionpatterns of immune cells after vaccination has been explored byseveral groups [1,2,23,6–8,24]. Also, the effect of adjuvants with-out a co-administered antigen [9] has been explored recently.However, there are limited comprehensive data of the effect ofgenetic vaccination on the expression profile, with or withoutadjuvants, at the site of administration [5,6]. In the present work,transcriptome-wide DNA arrays and Real-Time PCR assays wereused to explore the immuno-modulatory activity of Vaxfectin®
on early gene expression at the injection site. Transcript levels inmuscle injected with Vaxfectin®-formulated hCMV gB pDNA werecompared to those observed in muscle injected with hCMV gBpDNA in PBS. This approach was expected to generate formulation-specific expression data since transcription changes due solely toadministration of plasmid DNA and injection-induced tissue dam-age would be controlled.
The microarray data showed that vaccination with Vaxfectin®-formulated plasmid resulted in the up-regulation of both MHCclass I (H2-D1, H2-Q1, H2-K1 and H2-T9, 10, 22, 23, 24) and classII (H2-Ab1, H2-DMa, H2-DMb1 and H2-Eb1) genes 24–48 h afterinjection (Fig. 2). In addition, other genes involved in antigen pro-cessing and presentation such as B2M, TAP and TAPBP were alsoenriched in the Vaxfectin®-hCMV gB pDNA groups. Mosca et al. [9]observed a similar pattern of gene activation by several adjuvantsin the absence of co-administered antigen. Their data also indicatethat the enrichment of MHC class II transcripts tracked with thecellular infiltration of MHC class II+ cells to the injection site. Inter-estingly, Purcell et al. [5] observed a more substantial enrichmentin antigen presentation transcripts in response to vaccination with
7416 A. Vilalta et al. / Vaccine 27 (2009) 7409–7417
Fig. 6. Immuno-histological examination of muscle tissue following plasmid admin-istration. Representative examples of muscle sections from animals injected witheither hCMV gB pDNA-PBS (left panels) or hCMV gB pDNA-Vaxfectin® (right pan-els) are presented. Muscles were probed with either F4/80 antibody (A–D) orCD11b/Mac-1 antibody (E–H) as described in Section 2. Cellular infiltrates (arrows;macrophages (F4/80 and CD11b), granuclocytes, monocytes and NK cells (CD11b))were evident 24 h after injection in both hCMV gB pDNA-PBS (A and E) and hCMVgtt1
actfa
aiwrg[Iogvatbu
Interestingly, growing evidence indicates that several adjuvantswork through a TLR-mediated mechanism [30–33]. Functionalanalysis of the microarray data indicated that statistically signif-icant increases in MyD88, TLR2, and TLR3 mRNAs were observed.Evaluation of transcript levels by Real-Time PCR confirmed over-expression of MyD88, TLR2, TLR3, and indicated that TLR9 mRNAwas also elevated compared to pDNA-PBS samples (Fig. 4); TRIFmRNA levels were unchanged in response to plasmid formula-tion. One possible explanation for the increase in TLR pathwaytranscripts invokes the presence of positive feedback loops [34,35]acting on the TLR pathway. Work by Takeshita et al. [36] demon-strated that overexpression of TLR adaptor molecules MyD88 andTRIF can result in stimulation of the TLR pathway; overexpressionof MyD88 results in predominantly Th1 B-cell stimulation whileoverexpression of TRIF preferentially enhances an antigen-specificcellular response.
Previous work has identified the participation of skeletal mus-cle in local immune responses by expressing cytokines, adhesionmolecules and TLRs [37,38]. An alternative explanation for theincreased level of TLR-related transcripts in response to formu-lation with Vaxfectin® is the increased number of (F4/80+ andCD11b+) cells infiltrating muscles injected with pDNA-Vaxfectin®
(Fig. 6) compared to hCMV gB pDNA-PBS. However, histologicaldata showed that infiltrate cells in the muscle increased between 1and 7 days while the TLR-related transcripts diminished during thattimeframe. Therefore, it is unlikely that infiltration of immune cellsto the injection site is the sole source of increased TLR transcriptlevels.
4.2. Expression of Th1/Th2 markers
Adjuvant effects of cationic lipids have been well documented[11–17,22,39]. Published work indicates that plasmid formula-tion with the Vaxfectin® lipid system enhances B-cell responses[12–14,18] which is partially dependent on IL-6 [12]. The microar-ray and Real-Time PCR data indicate that IL-6 was over expressedin the muscle in response to hCMV gB pDNA-Vaxfectin® 24 h afteradministration and remained high 48 h post-injection. In addition,IFN-� levels were enhanced in the muscle and systemically withno changes in other Th1/Th2 markers (i.e., CD80, CD86, IL-2, IL-4,IL-5, IL-10 and IL-13). Up-regulation of IL-6 is expected to result inenhanced antigen-specific antibody responses since IL-6 mediatesB-cell differentiation to plasmablasts and enhances plasmablastproliferation and antibody production [40,41]. The local and sys-temic relative levels of Th1/Th2 cytokines support an enhancementof the Th1 response. Therefore, the results provide a molecularframework for the stimulation of B-cell responses by Vaxfectin®
consistent with published immunological data [11,12,14,22].
5. Conclusions
The presented experimental results show that modulation ofimmune response-related transcripts takes place in response tointramuscular injection of hCMV gB plasmid formulated withthe cationic lipid system Vaxfectin® in mice. Functional analysissuggests the involvement of antigen presentation, apoptosis andToll-like receptor pathways in response to vaccination with hCMVgB pDNA-Vaxfectin®. The expression profile for Th1/Th2 markerswas consistent with previous published results with regard to theenhancement of Th1 responses by Vaxfectin®. Formulation of pDNAwith Vaxfectin® did not enhance the level of expression of the
B pDNA-Vaxfectin® groups (B and F). Infiltrates increased 7 days after injection inhe hCMV gB pDNA-Vaxfectin® group (D and H) while decreasing dramatically inhe hCMV gB pDNA-PBS group (C and G). Original magnifications 200× for A–D and00× for E–H.
n antigen-encoding pDNA relative to empty pDNA. The relativeontribution of activation of resident APCs vs. infiltration of APCso the enrichment of MHC transcripts in response to Vaxfectin®-ormulated plasmid is not clear at this moment and will requiredditional experimentation.
Microarray data presented here indicated modulation of thepoptotic cascade (Fig. 2). Specifically, members of the caspase fam-ly (caspase 7 and 12), FAS, TNFSF10 and ATM were up-regulated
ithin 24 h of Vaxfectin®-hCMV gB pDNA administration. Severaleports have shown the correlation between enhanced immuno-enicity and apoptosis [reviewed by Bergmann-Leitner and Leitner25]] potentially due to an increase in antigen uptake by APCs [26].n addition, control of the apoptosis cascade has been the targetf recent attempts [27,28] at enhancing the immunogenicity ofenetic vaccines. Published reports [29] indicate that use of adju-ants such as MF59 result in increased macrophage trafficking and
poptosis linked to increased immunogenicity of adjuvanted pro-ein vaccines. It is therefore possible that one of the mechanismsy which Vaxfectin® enhances immunogenicity relies on the stim-lation of non-homeostatic apoptosis.plasmid-encoded antigen at the site of administration. Hence, theincrease in immune response observed when plasmid is formu-lated with Vaxfectin® is not simply due to increased gene transferefficiency, but rather the direct result of immune activation by

A. Vilalta et al. / Vaccine 27 (2009) 7409–7417 7417
Vaxfectin®. These results provide further insight into the activa-tion pathways in which Vaxfectin® is able to increase the immuneresponse to pDNA following intramuscular administration.
Acknowledgments
The authors would like to acknowledge Jane Morrow and VicalVivarium staff for performing all of the vaccinations and tissueharvests. Michael Komai provided all of the pDNA formulationsfor this study. The authors are grateful to Neal Sekiya for pro-viding assistance with the FACS analysis (VA Hospital San Diego,flow cytometry research core facility) and to Professor Nissi M.Varki (UCSD) for advice and support with histology and immuno-histochemistry. The authors appreciate the critical review of thismanuscript by Dr. Jukka Hartikka.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.vaccine.2009.08.075.
References
[1] Regnstrom K, Ragnarsson EG, Rydell N, Sjoholm I, Artursson P. Tetanus antigenmodulates the gene expression profile of aluminum phosphate adjuvant inspleen lymphocytes in vivo. Pharmacogenomics J 2002;2(1):57–64.
[2] Regnstrom K, Ragnarsson E, Artursson P. Gene expression after vaccinationof mice with formulations of diphtheria toxoid or tetanus toxoid and differ-ent adjuvants: identification of shared and vaccine-specific genes in spleenlymphocytes. Vaccine 2003;21(19–20):2307–17.
[3] Regnstrom K, Burgess DJ. Pharmacogenomics and its potential impacton drug and formulation development. Crit Rev Ther Drug Carrier Syst2005;22(5):465–92.
[4] Halonen SK, Woods T, McInnerney K, Weiss LM. Microarray analysis of IFN-gamma response genes in astrocytes. J Neuroimmunol 2006;175(1–2):19–30.
[5] Purcell MK, Nichols KM, Winton JR, Kurath G, Thorgaard GH, Wheeler P, etal. Comprehensive gene expression profiling following DNA vaccination ofrainbow trout against infectious hematopoietic necrosis virus. Mol Immunol2006;43(13):2089–106.
[6] Regnstrom K, Ragnarsson EG, Fryknas M, Koping-Hoggard M, Artursson P. Geneexpression profiles in mouse lung tissue after administration of two cationicpolymers used for nonviral gene delivery. Pharm Res 2006;23(3):475–82.
[7] Rogers JV, Casbohm SL, Giannunzio LF, Sinnott LT, Rust SW, Brys AM, et al.Transcriptional analysis of protective antigen-stimulated PBMC from non-human primates vaccinated with the anthrax vaccine absorbed. Vaccine2006;24(17):3609–17.
[8] Royaee AR, Jong L, Mendis C, Das R, Jett M, Yang DC. Cholera toxininduced novel genes in human lymphocytes and monocytes. Mol Immunol2006;43(8):1267–74.
[9] Mosca F, Tritto E, Muzzi A, Monaci E, Bagnoli F, Iavarone C, et al. Molecularand cellular signatures of human vaccine adjuvants. Proc Natl Acad Sci USA2008;105(30):10501–6.
[10] Byon JY, Ohira T, Hirono I, Aoki T. Comparative immune responses in Japaneseflounder, Paralichthys olivaceus after vaccination with viral hemorrhagic sep-ticemia virus (VHSV) recombinant glycoprotein and DNA vaccine using amicroarray analysis. Vaccine 2006;24(7):921–30.
[11] Hartikka J, Bozoukova V, Ferrari M, Sukhu L, Enas J, Sawdey M, et al. Vaxfectinenhances the humoral immune response to plasmid DNA-encoded antigens.Vaccine 2001;19(15–16):1911–23.
[12] Reyes L, Hartikka J, Bozoukova V, Sukhu L, Nishioka W, Singh G, et al. Vaxfectinenhances antigen specific antibody titers and maintains Th1 type immuneresponses to plasmid DNA immunization. Vaccine 2001;19(27):3778–86.
[13] Fischer L, Minke J, Dufay N, Baudu P, Audonnet JC. Rabies DNA vac-cine in the horse: strategies to improve serological responses. Vaccine2003;21(31):4593–6.
[14] Hermanson G, Whitlow V, Parker S, Tonsky K, Rusalov D, Ferrari M, et al. Acationic lipid-formulated plasmid DNA vaccine confers sustained antibody-mediated protection against aerosolized anthrax spores. Proc Natl Acad SciUSA 2004;101(37):13601–6.
[15] Sedegah M, Rogers WO, Belmonte A, Belmonte M, Banania G, Patterson N,et al. Vaxfectin enhances immunogenicity and protective efficacy of P. yoeliicircumsporozoite DNA vaccines. Vaccine 2006;24(11):1921–7.
[16] Jimenez GS, Planchon R, Wei Q, Rusalov D, Geall A, Enas J, et al. Vaxfectin-formulated influenza DNA vaccines encoding NP and M2 viral proteins protectmice against lethal viral challenge. Hum Vaccines 2007;3(5):157–64.
[17] Lalor PA, Webby RJ, Morrow J, Rusalov D, Kaslow DC, Rolland A, et al. Plas-mid DNA-based vaccines protect mice and ferrets against lethal challenge withA/Vietnam/1203/04 (H5N1) influenza virus. J Infect Dis 2008;197(12):1643–52.
[18] Pan CH, Jimenez GS, Nair N, Wei Q, Adams RJ, Polack FP, et al. Vaxfectin®-formulated measles DNA vaccine encoding the hemagglutinin and fusionproteins protects juvenile and infant rhesus macaques from measles. Clin Vac-cine Immunol 2008;15(8):1214–21.
[19] Selinsky C, Luke C, Wloch M, Geall A, Hermanson G, Kaslow D, et al. A DNA-basedvaccine for the prevention of human cytomegalovirus-associated diseases.Hum Vaccines 2005;1(1):16–23.
[20] Dennis Jr G, Sherman BT, Hosack DA, Yang J, Gao W, Lane HC, et al. DAVID:database for annotation, visualization, and integrated discovery. Genome Biol2003;4(5):P3.
[21] Mahajan R, Feher B, Jones B, Jones D, Marjerison L, Sam M, et al. A TaqMan®
Reverse Transcription Polymerase Chain Reaction (RT-PCR) in vitro potencyassay for plasmid-based vaccine products. Mol Biotechnol 2008.
[22] Margalith M, Vilalta A. Sustained protective rabies neutralizing antibody titersafter administration of cationic lipid-formulated pDNA vaccine. Genet VaccinesTher 2006;4:2.
[23] Rosenkrands I, Agger EM, Olsen AW, Korsholm KS, Andersen CS, Jensen KT, etal. Cationic liposomes containing mycobacterial lipids: a new powerful Th1adjuvant system. Infect Immun 2005;73(9):5817–26.
[24] Yan W, Chen W, Huang L. Mechanism of adjuvant activity of cationic lipo-some: phosphorylation of a MAP kinase, ERK and induction of chemokines.Mol Immunol 2007;44(15):3672–81.
[25] Bergmann-Leitner ES, Leitner WW. Danger, death and DNA vaccines. MicrobesInfect 2004;6(3):319–27.
[26] Albert ML, Sauter B, Bhardwaj N. Dendritic cells acquire antigen from apoptoticcells and induce class I-restricted CTLs. Nature 1998;392(6671):86–9.
[27] Li AF, Hough J, Henderson D, Escher A. Co-delivery of pro-apoptotic BAX witha DNA vaccine recruits dendritic cells and promotes efficacy of autoimmunediabetes prevention in mice. Vaccine 2004;22(13–14):1751–63.
[28] Chattergoon MA, Muthumani K, Tamura Y, Ramanathan M, Shames JP, SaulinoV, et al. DR5 activation of caspase-8 induces DC maturation and immuneenhancement in vivo. Mol Ther 2008;16(2):419–26.
[29] Dupuis M, Denis-Mize K, LaBarbara A, Peters W, Charo IF, McDonald DM, etal. Immunization with the adjuvant MF59 induces macrophage trafficking andapoptosis. Eur J Immunol 2001;31(10):2910–8.
[30] Sieling PA, Chung W, Duong BT, Godowski PJ, Modlin RL. Toll-like receptor 2 lig-ands as adjuvants for human Th1 responses. J Immunol 2003;170(1):194–200.
[31] Schellack C, Prinz K, Egyed A, Fritz JH, Wittmann B, Ginzler M, et al. IC31, anovel adjuvant signaling via TLR9, induces potent cellular and humoral immuneresponses. Vaccine 2006;24(26):5461–72.
[32] Seya T, Akazawa T, Tsujita T, Matsumoto M. Role of Toll-like receptors inadjuvant-augmented immune therapies. Evid Based Complement Altern Med2006;3(1):31–8 [discussion 133–7].
[33] Johnston D, Zaidi B, Bystryn JC. TLR7 imidazoquinoline ligand 3M-019 isa potent adjuvant for pure protein prototype vaccines. Cancer ImmunolImmunother 2007;56(8):1133–41.
[34] Plavec I, Sirenko O, Privat S, Wang Y, Dajee M, Melrose J, et al. Method foranalyzing signaling networks in complex cellular systems. Proc Natl Acad SciUSA 2004;101(5):1223–8.
[35] Oda K, Kitano H. A comprehensive map of the toll-like receptor signaling net-work. Mol Syst Biol 2006;2:0015.
[36] Takeshita F, Tanaka T, Matsuda T, Tozuka M, Kobiyama K, Saha S, et al. Toll-likereceptor adaptor molecules enhance DNA-raised adaptive immune responsesagainst influenza and tumors through activation of innate immunity. J Virol2006;80(13):6218–24.
[37] Wiendl H, Hohlfeld R, Kieseier BC. Immunobiology of muscle: advancesin understanding an immunological microenvironment. Trends Immunol2005;26(7):373–80.
[38] Wiendl H, Hohlfeld R, Kieseier BC. Muscle-derived positive and negative regu-lators of the immune response. Curr Opin Rheumatol 2005;17(6):714–9.
[39] Nukuzuma C, Ajiro N, Wheeler CJ, Konishi E. Enhancing effect of Vaxfectin on theability of a Japanese encephalitis DNA vaccine to induce neutralizing antibodyin mice. Viral Immunol 2003;16(2):183–9.
[40] Van Snick J, Cayphas S, Szikora JP, Renauld JC, Van Roost E, Boon T, et al. cDNAcloning of murine interleukin-HP1: homology with human interleukin 6. Eur JImmunol 1988;18(2):193–7.
[41] Matsuda T, Yamasaki K, Taga T, Hirano T, Kishimoto T. Current concepts of Bcell modulation. Int Rev Immunol 1989;5(2):97–109.




















