
Emerging Green Techniques for the Extraction of Antioxidants ...
Upload
independentCategory
view
2download
0
A
ceowa2ca(irwse©
Ke
f
0
Toxicology 239 (2007) 213–223
An in vitro study on reproductive toxicity ofaluminium chloride on rabbit sperm: The
protective role of some antioxidants
Mokhtar I. Yousef a,∗, Kamel I. Kamel b,Marwa I. El-Guendi a, Fatma M. El-Demerdash a
a Department of Environmental Studies, Institute of Graduate Studies and Research, Alexandria University,163 Horreya Avenue, P.O. Box 832, El-Chatby, Alexandria 21526, Egypt
b Animal Production Research Institute, Dokki, Giza, Egypt
Received 11 April 2007; received in revised form 10 July 2007; accepted 11 July 2007Available online 17 July 2007
bstract
An in vitro study using rabbit sperm was designed to evaluate the cytotoxic effects of different concentrations of aluminiumhloride (AlCl3) at 0, 2 and 4 h of incubation on sperm motility and viability, oxidative status and the activities of some antioxidantnzymes (superoxide dismutase (SOD) and catalase (CAT)), transaminases and acid phosphatase. The role of vitamin C (1 mM)r vitamin E (2 mM) was also investigated in counteracting deterioration caused by AlCl3 on the tested parameters. Rabbit spermas incubated with different concentrations of AlCl3 (0, 1, 5, 10, 15 and 20 mM) with or without vitamin C or vitamin E for 2
nd 4 h. Results revealed that the percentage of motile and viable sperm decreased significantly after AlCl3 treatment at 10, 15 and0 mM and the response was both concentration and time dependent. Aluminium chloride at concentrations of 10, 15 and 20 mMaused significant induction of oxidative stress as evidenced by increased thiobarbituric acid reactive substances (TBARS) levelsnd inhibition in the activities of SOD and CAT. Increase in the activities of aspartate transaminase (AST) and alanine transaminaseALT) and decline in the activity of acid phosphatase (ACP) were also observed at AlCl3 concentrations of 15 and 20 mM. Co-ncubation with either vitamin C or vitamin E resulted in marked degrees of protection against AlCl3-induced cytotoxic effects,epresented in decreased TBARS levels and restoration of enzymes activities near control. On the other hand, no significant effect
as exerted from vitamin C or vitamin E on motility and viability. The present study demonstrated that AlCl3 caused deterioration inperm motility and viability, enhancement of free radicals and alterations in enzymes activities. The antioxidants revealed protectiveffects against the cytotoxicity of AlCl3.
2007 Elsevier Ireland Ltd. All rights reserved.
eywords: Reproductive toxicity; Aluminium chloride; Vitamin C; Vitamin E;nzymes
d
∗ Corresponding author. Tel.: +20 34 29 50 07;ax: +20 34 28 57 92.
E-mail address: [email protected] (M.I. Yousef).
300-483X/$ – see front matter © 2007 Elsevier Ireland Ltd. All rights reservoi:10.1016/j.tox.2007.07.011
Rabbit sperm; Sperm motility and viability; Free radicals; Antioxidant
1. Introduction
Taking into account that aluminium is ubiquitous,exposure to this element is in fact unavoidable dur-ing the entire life span. Humans consume an averageof 7600 �g/day of aluminium from drinking water and
ed.

icology
214 M.I. Yousef et al. / Toxfood (Yokel and McNamara, 2001). However, other alu-minium sources such as cosmetics, drugs or specialfoods increase the levels of exposure in some sub-groups of population. In relation to this, pregnant womenmay be potentially exposed to aluminium through thediet (including drinking water), dust and soil ingestionand some medications (Roig et al., 2006). Evidencehas shown that aluminium is linked to pathologiesas dialysis dementia, iron-adequate microcytic anemia,osteomalacia (Suwalsky et al., 2004; Domingo, 2006).Savory et al. (2006) reported that for four decades thecontroversial question concerning a possible role foraluminium neurotoxicity in contributing to the patho-genesis of Alzheimer’s disease has been debated, andstudies by different investigators have yielded con-tradictory results. Rabbits are particularly sensitive toaluminium neurotoxicity and they develop severe neu-rological changes that are dependent on dose, age androute of administration. The most prominent featureinduced by aluminium in rabbit brain is a neurofib-rillary degeneration that shares some similarity withthe neurofibrillary tangles found in Alzheimer’s diseasepatients. Domingo et al. (1987) reported that pregnantrats treated with aluminium nitrate on day 14 of gestationthrough day 21 of lactation, showed no overt fetotoxi-city, but growth of offspring was significantly delayed(body weight, body length and tail length) from birth toweaning.
Although the knowledge of Al toxicity has markedlyimproved in recent years; however, information con-cerning male reproductive toxicity in vitro of thiselement is still very limited. The study of Dawson etal. (1998) compared the semen levels of lead (Pb),cadmium (Cd) and aluminium (Al) in relation to livesperm in semen samples from 64 apparently healthymen. They reported that Al in human spermatozoaand seminal plasma were found to be correlated withdecreased sperm motility and viability. Testicular Alaccumulation, necrosis of spermatocytes/spermatids anda significant decrease in fertility were observed in malemice treated intraperitoneally with Al-nitrate at dosesof 0, 50, 100 and 200 mg/kg/day for 4 weeks (Llobet etal., 1995). In addition, the suppressive effects of long-term Al-chloride in drinking water at a concentrationof 1000 ppm for 12 weeks on both sexual and aggres-sive behavior, and fertility of male rats were also noted(Bataineh et al., 1998). Hovatta et al. (1998) studiedsemen quality and the concentrations of aluminium, cad-
mium and lead in spermatozoa and seminal plasma ina group of employees of a refinery and a polyolefinfactory and compared the results with data obtainedfrom sperm bank donor candidates over the same time239 (2007) 213–223
period. Relatively high concentrations of aluminiumin the spermatozoa were correlated with the motilityand morphology of the spermatozoa. Semen analysesof the highest aluminium concentrations in spermato-zoa showed asthenozoospermia. Guo et al. (2005) foundthat intraperitoneal injection with aluminium chloride at35 mg/kg BW/day for a period of 12 days significantlyincreased nitric oxide production and decreased bothtesticular adenosine 3,5-cyclic monophosphate (cAMP)and testosterone levels of male mice. They demonstratedthat excessive nitric oxide products, thus activatinginducible nitric oxide synthase, might be involved inreproductive toxicity of aluminium. Results obtainedfrom Yousef et al. (2005) revealed that rabbits orallyadministered Al-chloride at 34 mg/kg BW every otherday for 16 weeks showed significant decrease in ejac-ulate volume, sperm concentration, total sperm output,sperm motility, total motile sperm per ejaculate, packedsperm volume, total functional sperm fraction, normaland live sperm, while in dead and abnormal spermwere increased. Also, Chinoy et al. (2005a,b) foundthat aluminium induced toxicity in mice epididymis, vasdeferens, seminal vesicle, and ventral prostate of mice.They reported that vitamin C alleviated the toxicity ofaluminium.
Aluminium was shown to exert a pro-oxidant activ-ity and promotes biological oxidation both in vitro andin vivo (Exley, 2004). Also, Yousef et al. (2005) andYousef (2004) showed that AlCl3-induced free radicalsand inhibited antioxidant enzymes in blood and seminalplasma, and different organs (liver, testes, kidney, lungand brain) of rabbits.
To control the level of reactive oxygen species(ROS) and to protect cells under stress conditions,mammalian tissues contain several enzymes scavengingROS such as catalase, superoxide dismutase and glu-tathione peroxidase. Some vitamins also contribute tothe detoxification process from ROS such as ascorbicacid, which is considered the most water-soluble antiox-idant in extracellular fluids (Blokhina et al., 2003). Also,vitamin E, a major lipid-soluble antioxidant belong-ing to tocopherols, is the most effective chain-breakingantioxidant within cell membrane. It is able to repairoxidizing radicals directly, preventing the chain prop-agation step during lipid peroxidation (Zhang et al.,2001).
Therefore, our aim is to test the effect of differentconcentrations of AlCl on rabbit sperm motility, live
3and dead sperm, and oxidative status and the activitiesof antioxidant enzymes, and also, to investigate the roleof vitamin C or vitamin E in alleviating the cytotoxiceffect of AlCl3.
icology
2
2
upCfHloa2mA(si
2
iit1cwh3aod
taAc(mpswc
2
o5wotsm
M.I. Yousef et al. / Tox
. Materials and methods
.1. Chemicals and incubation medium
Aluminium chloride, vitamin C and vitamin E weresed in the present experiments. Aluminium chloride (itsurity was >99%) was purchased from Aldrich Chemicalompany (Milwaukee, WI, USA), vitamin C 20% from Neo-
arma (Italy) and vitamin E 50% from Lohmann Animalealth (Germany). All other chemicals used were of ana-
ytical reagent grade. Semen incubation medium consistedf tris-(hydroxymethyl)-aminomethane (37.85 g/l), citric acidnhydrous (21.15 g/l) and d(−)fructose (10 g/l) (Roca et al.,000). Aluminium chloride was dissolved in the incubationedium to produce stock solutions of 50, 250 and 500 mM.lso, stock solutions of vitamin C (50 mM) and vitamin E
10 mM) were prepared in the incubation medium. All stockolutions were prepared freshly at the beginning of each exper-ment.
.2. Animals
Male New Zealand white rabbits (age of 7 months andnitial weight of 2.985 ± 0.072 kg) were used. Animals werendividually housed in stainless steel cages at room tempera-ure (25 ± 2 ◦C) with a relative humidity of 50–60% and on a2 h light–dark cycle. The animals had free access to commer-ial pellet diet and water ad libitum. Rabbits were fed pelletshich consisted of 30% berseem (Trifolium alexandrinum)ay, 25% yellow corn, 26.2% wheat bran, 14% soybean meal,% molasses, 1% CaCl2, 0.4% NaCl, 0.3% mixture of miner-ls and vitamins and 0.1% methionine. Rabbits and diet werebtained from Animal Production Research Institute, Alexan-ria, Egypt.
Ejaculates were collected using an artificial vagina and aeaser doe. Aliquots from each ejaculate were initially evalu-ted under a light microscope for concentration and motility.
weak eosin solution was used for evaluation of sperm con-entration by the improved Neubauer haemocytometer slideGmbH + Co., Brandstwiete, Germany). The percentages ofotile sperm were estimated by visual examination under low-
ower magnification (10×) using light microscope with heatedtage. The ejaculates with the highest sperm motility (>70%)ere applied. Samples from rabbits with nearly the same per-
ent of motility were pooled.
.3. AlCl3 and vitamins treatments
There were three experimental groups; each group includedne control and five different concentrations of AlCl3 (1,, 10, 15 and 20 mM) without vitamins or in combination
ith either vitamin C (1 mM) (Verma and Kanwar, 1998)r vitamin E (2 mM) (Verma and Kanwar, 1999). One con-rol contained medium and sperm only, and two otherserved as positive controls, containing vitamin C or vita-in E. The pH was adjusted to 7.0 for all treatments at239 (2007) 213–223 215
the beginning of incubation. All groups were repeated 5times.
To evaluate the effect of AlCl3 on sperm motility and via-bility, semen was re-suspended in control and test incubationtubes at 1 × 105 sperm cell/ml incubation medium, and incu-bated with different concentrations of AlCl3 without vitaminsor in combination with either vitamin C or vitamin E in waterbath at 37 ◦C for 2 and 4 h. Vitamin C or vitamin E was addedprior to the addition of AlCl3. Time zero for the dosed samplesis approximately 1–2 min after the addition of AlCl3. Motilityand live sperm were estimated at 0, 2 and 4 h. The percent-age of motile sperm was estimated by visual examinationunder low-power magnification (10×) using light micro-scope. Live and dead sperm were estimated using 0.5% eosinstain.
2.4. Evaluation of TBARS and enzymes activities
To evaluate the effect of AlCl3 on enzyme activities, semenwas re-suspended in control and test incubation tubes at 8 × 106
sperm cell/ml incubation medium, and incubated as mentionedbefore. Samples were centrifuged after 2 or 4 h incubationperiods at 1000 × g for 20 min, the supernatant was col-lected and stored at −20 ◦C until analysis of thiobarbituricacid reactive substances level (TBARS), activities of superox-ide dismutase (SOD), catalase (CAT), asparate transaminase(AST), alanine transaminase (ALT) and acid phosphatase(ACP).
The levels of TBARS, as a marker for lipid peroxidation(LPO), were measured at 532 nm by using 2-thiobarbituric acid(2,6-dihydroxypyrimidine-2-thiol (TBA)). An extinction coef-ficient of 156 000 M−1 cm−1 was used for calculation (Okhawaet al., 1979). The activity of SOD (EC 1.15.1.1) activity wasmeasured according to Misra and Fridovich (1972). Cata-lase (EC 1.11.1.6) activity was determined using the Luckmethod involving the decomposition of hydrogen peroxide(Luck, 1974).
The activities of AST (EC 2.6.1.1) and ALT (EC 2.6.1.2)were determined with kits from Sentinel Ch. (via principleEugenio 5-20155 Milan, Italy). The principle reaction of thecolorimetric determination of AST or ALT activity is based onthe reaction of aspartate or alanine with alpha-ketoglutarate toform oxaloacetate or pyruvate, respectively. The oxaloacetateor pyruvate formed is measured by monitoring the concen-tration of oxaloacetate or pyruvate hydrazone formed with2,4-dinitrophenylhydrazine. The activity of ACP (EC 3.1.3.2)was determined by the hydrolysis of p-nitrophenyl under acidicconditions (Moss, 1984).
2.5. Statistical analysis
Data were analyzed as a completely randomized design(Steel and Torrie, 1981) using the General Linear Model pro-cedure of SAS (1986). Dunnett post hoc analysis was used tocompare means of different treatments against the control. Pvalues <0.05 were accepted as significant.

216 M.I. Yousef et al. / Toxicology 239 (2007) 213–223
, 2 andtment w
Fig. 1. Changes in motility (%) and viability (%) of rabbit sperm at 0(mean ± S.E.). Asterisk (*) indicates significant (p < 0.05) effect of treawith Vit. C compared to control with Vit. C.
3. Results
The effects of different concentrations of AlCl3 (0, 1,5, 10, 15 and 20 mM) without vitamins or in combina-tion with either vitamin C (1 mM) or vitamin E (2 mM)on rabbit sperm motility and live sperm after incuba-tion for 0, 2 and 4 h are presented in Figs. 1 and 2.Results showed significant decline (p < 0.05) in motility(%) and live sperm (%) in sperm cells exposed to AlCl3at concentrations of 10, 15 and 20 mM at 0, 2 and 4 h ofincubation, and this effect was concentration and timedependent. However, concentrations of 1 and 5 mM didaffect neither motility nor viability. Co-incubation withvitamin C and vitamin E did not alleviate the adverseeffects of AlCl3 on motility or viability at all incubationperiods.
Aluminium chloride significantly (p < 0.05) elevatedthiobarbituric acid reactive substances (TBARS) level,while the activities of superoxide dismutase (SOD)and catalase (CAT) were decreased in a concentration
4 h of incubation with AlCl3 alone or with vitamin C (Vit. C 1 mM)ith AlCl3 alone compared to control without Vit. C or in combination
and time-dependent manner. Both vitamins significantly(p < 0.05) reduced TBARS level and decreased theinhibitory effect of AlCl3 on SOD and CAT comparedto AlCl3 without vitamins (Figs. 3–6).
The activities of aspartate transaminase (AST) andalanine transaminase (ALT) were significantly (p < 0.05)increased, while acid phosphatase (ACP) activity wasdecreased in a concentration and time-dependent man-ner. Addition of either vitamin C or vitamin E to rabbitsperm incubated with AlCl3 showed significant decrease(p < 0.05) in AST and ALT leakage, and alleviated theinhibitory effect on ACP (Figs. 3–6).
4. Discussion
Results from this study showed nearly no alter-
ation in motility or viability at concentrations below10 mM AlCl3, in comparison with control. Our studywas performed at neutral pH, at which Al(OH)3 pre-cipitates in aqueous media. Moreover, the incubation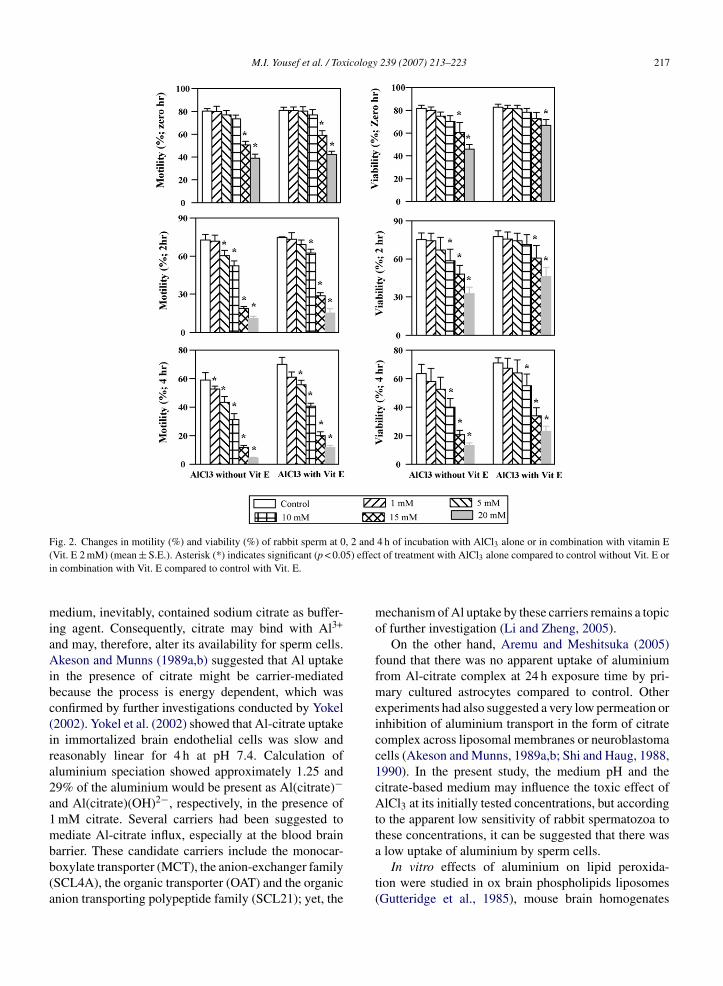
M.I. Yousef et al. / Toxicology 239 (2007) 213–223 217
F , 2 and( 5) effeci
miaAibc(ira2a1mbb(a
ig. 2. Changes in motility (%) and viability (%) of rabbit sperm at 0Vit. E 2 mM) (mean ± S.E.). Asterisk (*) indicates significant (p < 0.0n combination with Vit. E compared to control with Vit. E.
edium, inevitably, contained sodium citrate as buffer-ng agent. Consequently, citrate may bind with Al3+
nd may, therefore, alter its availability for sperm cells.keson and Munns (1989a,b) suggested that Al uptake
n the presence of citrate might be carrier-mediatedecause the process is energy dependent, which wasonfirmed by further investigations conducted by Yokel2002). Yokel et al. (2002) showed that Al-citrate uptaken immortalized brain endothelial cells was slow andeasonably linear for 4 h at pH 7.4. Calculation ofluminium speciation showed approximately 1.25 and9% of the aluminium would be present as Al(citrate)−nd Al(citrate)(OH)2−, respectively, in the presence ofmM citrate. Several carriers had been suggested toediate Al-citrate influx, especially at the blood brain
arrier. These candidate carriers include the monocar-oxylate transporter (MCT), the anion-exchanger familySCL4A), the organic transporter (OAT) and the organicnion transporting polypeptide family (SCL21); yet, the
4 h of incubation with AlCl3 alone or in combination with vitamin Et of treatment with AlCl3 alone compared to control without Vit. E or
mechanism of Al uptake by these carriers remains a topicof further investigation (Li and Zheng, 2005).
On the other hand, Aremu and Meshitsuka (2005)found that there was no apparent uptake of aluminiumfrom Al-citrate complex at 24 h exposure time by pri-mary cultured astrocytes compared to control. Otherexperiments had also suggested a very low permeation orinhibition of aluminium transport in the form of citratecomplex across liposomal membranes or neuroblastomacells (Akeson and Munns, 1989a,b; Shi and Haug, 1988,1990). In the present study, the medium pH and thecitrate-based medium may influence the toxic effect ofAlCl3 at its initially tested concentrations, but accordingto the apparent low sensitivity of rabbit spermatozoa tothese concentrations, it can be suggested that there was
a low uptake of aluminium by sperm cells.In vitro effects of aluminium on lipid peroxida-tion were studied in ox brain phospholipids liposomes(Gutteridge et al., 1985), mouse brain homogenates
218 M.I. Yousef et al. / Toxicology 239 (2007) 213–223
, AST aindicat
ontrol w
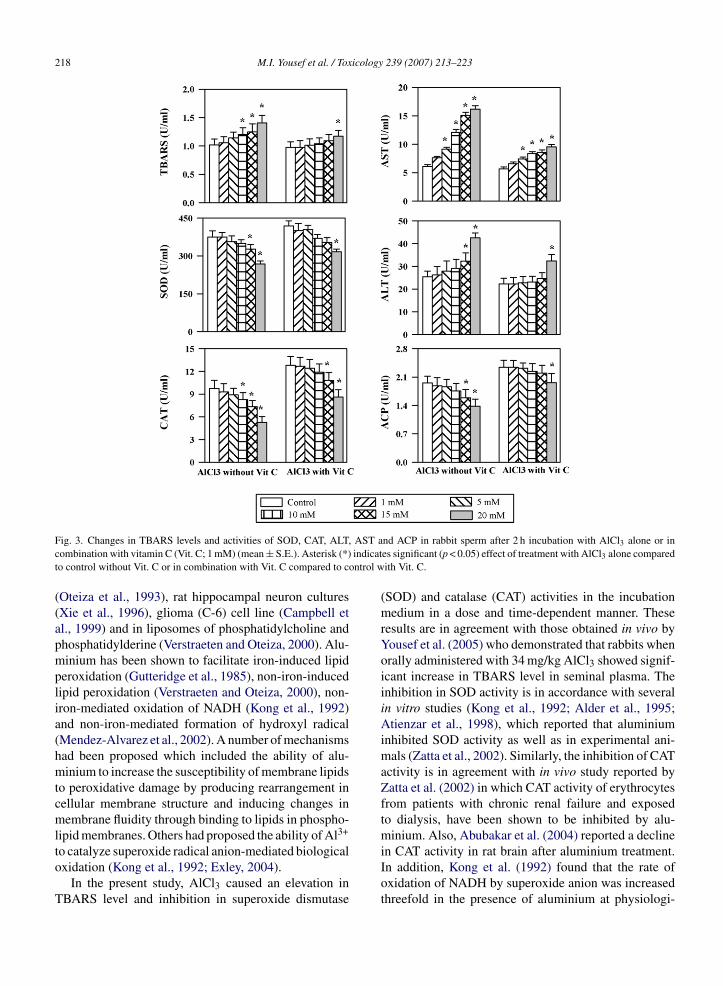
Fig. 3. Changes in TBARS levels and activities of SOD, CAT, ALTcombination with vitamin C (Vit. C; 1 mM) (mean ± S.E.). Asterisk (*)to control without Vit. C or in combination with Vit. C compared to c
(Oteiza et al., 1993), rat hippocampal neuron cultures(Xie et al., 1996), glioma (C-6) cell line (Campbell etal., 1999) and in liposomes of phosphatidylcholine andphosphatidylderine (Verstraeten and Oteiza, 2000). Alu-minium has been shown to facilitate iron-induced lipidperoxidation (Gutteridge et al., 1985), non-iron-inducedlipid peroxidation (Verstraeten and Oteiza, 2000), non-iron-mediated oxidation of NADH (Kong et al., 1992)and non-iron-mediated formation of hydroxyl radical(Mendez-Alvarez et al., 2002). A number of mechanismshad been proposed which included the ability of alu-minium to increase the susceptibility of membrane lipidsto peroxidative damage by producing rearrangement incellular membrane structure and inducing changes inmembrane fluidity through binding to lipids in phospho-lipid membranes. Others had proposed the ability of Al3+
to catalyze superoxide radical anion-mediated biologicaloxidation (Kong et al., 1992; Exley, 2004).
In the present study, AlCl3 caused an elevation inTBARS level and inhibition in superoxide dismutase
nd ACP in rabbit sperm after 2 h incubation with AlCl3 alone or ines significant (p < 0.05) effect of treatment with AlCl3 alone comparedith Vit. C.
(SOD) and catalase (CAT) activities in the incubationmedium in a dose and time-dependent manner. Theseresults are in agreement with those obtained in vivo byYousef et al. (2005) who demonstrated that rabbits whenorally administered with 34 mg/kg AlCl3 showed signif-icant increase in TBARS level in seminal plasma. Theinhibition in SOD activity is in accordance with severalin vitro studies (Kong et al., 1992; Alder et al., 1995;Atienzar et al., 1998), which reported that aluminiuminhibited SOD activity as well as in experimental ani-mals (Zatta et al., 2002). Similarly, the inhibition of CATactivity is in agreement with in vivo study reported byZatta et al. (2002) in which CAT activity of erythrocytesfrom patients with chronic renal failure and exposedto dialysis, have been shown to be inhibited by alu-minium. Also, Abubakar et al. (2004) reported a decline
in CAT activity in rat brain after aluminium treatment.In addition, Kong et al. (1992) found that the rate ofoxidation of NADH by superoxide anion was increasedthreefold in the presence of aluminium at physiologi-
M.I. Yousef et al. / Toxicology 239 (2007) 213–223 219
F d ACPw gnificanw Vit. E.
cpas
afaasaloeu
ipoo
ig. 4. Changes in TBARS levels, activities of SOD, CAT, AST, ALT anith vitamin E (Vit. E 2 mM) (mean ± S.E.). Asterisk (*) indicates siithout Vit. E or in combination with Vit. E compared to control with
al pH (media buffered at pH 6.0 and pH 7.0) in theresence of a significant excess of citrate, which may rel-tively coincide with the incubation environment in ourtudy.
The susceptibility of spermatozoa to oxidative dam-ge is attributed to the high concentration of unsaturatedatty acids in membrane phospholipids, the limitedntioxidant capacity of spermatozoa as well as thebility of spermatozoa to generate reactive oxygenpecies (Aitken, 1995). Induced oxidative stress byluminium may result in a fall in intracellular ATPevels which decrease sperm motility and lipid per-xidation associated with increased cell permeability,nzyme inactivation and production of spermicidal prod-cts (Verma and Kanwar, 1999).
Transminases and phosphatases in semen play an
mportant role in transamination and phosphorylationrocesses in sperm metabolism (Dhami et al., 1994). Inur study, aluminium treatments increased the activitiesf aspartate transaminase (AST) and alanine transami-in rabbit sperm after 2 h incubation with AlCl3 alone or in combinationt (p < 0.05) effect of treatment with AlCl3 alone compared to control
nase (ALT), which is in agreement with results obtainedin vitro with Wilhelm et al. (1996) in rat hepatic tis-sues, also with results obtained in vivo with Yousef et al.(2005) in rabbit seminal plasma. The increase in ASTand ALT activities may be attributed to some structuralalterations in the mitochondria in the mid-piece region,thereby causing leakage of these enzymes in the medium(Strzegek, 1988).
Our results showed also a decrease in acid phos-phatase (ACP) activity after incubation with AlCl3,which is supported by results obtained in vivo byYousef et al. (2005) and Domenech et al. (1992) whoobserved significant inhibition of ACP by aluminiumin Pseudomonas aeruginosa culture. This observationmay be explained considering that ACP is a magnesium-dependant enzyme, and a strong competition is expected
between Al3+ and Mg2+ ions for their close chemicalresemblance (Zatta et al., 2000).Regarding the oxidative damage caused by alu-minium in rabbit sperm, vitamin C and vitamin E were

220 M.I. Yousef et al. / Toxicology 239 (2007) 213–223
, ALT aindicat
ontrol w
Fig. 5. Changes in TBARS levels and activities of SOD, CAT, ASTcombination with vitamin C (Vit. C 1 mM) (mean ± S.E.). Asterisk (*)to control without Vit. C or in combination with Vit. C compared to c
chosen to evaluate their protective roles as antioxidants.Luck et al. (1995) reported that vitamin C should beconsidered an essential biochemical in the reproductiveprocess and a potentially significant factor in human fer-tility. In addition, vitamin E is believed to be the primarycomponents of the antioxidant system of the spermato-zoa (Surai et al., 1998). Yousef et al. (2003) showed thatthe formation of TBARS in rabbit sperm in vivo was sig-nificantly decreased by either vitamin alone or both incombination, which confirm our results in which vitaminC or vitamin E decreased TBARS level, alleviated theinhibitory effect of AlCl3 on SOD, CAT and ACP activi-ties, and reduced the leakage of AST and ALT. However,the observed decline in motility and viability was notimproved by either vitamin, in contrast with Yousef etal. (2005) who reported improvement in semen charac-
teristics in vivo after the administration of vitamin C withAlCl3. Zatta et al. (2000) demonstrated that aconitase, aprotein that binds citrate and catalyzes its isomerizationto isocitrate via the intermediate cis-aconitate in Krebsnd ACP in rabbit sperm after 4 h incubation with AlCl3 alone or ines significant (p < 0.05) effect of treatment with AlCl3 alone comparedith Vit. C.
cycle, showed decreased activity in the presence of alu-minium in brain homogenate in vitro, suggesting thataluminium may influence mitochondrial enzymes. Con-sequently, changes in mitochondrial functions may bereflected in sperm motility and viability. Therefore, itis suggested that minimal Al was taken up by the cellsand affecting mitochondrial functions through a mecha-nism other than causing oxidative damage, which mightexplain the inability of both vitamins of attenuating thenegative effect of AlCl3 on motility and viability.
In conclusion, the obtained results showed that alu-minium chloride exerted reproductive toxicity in adose and time-dependent manner on rabbit sperm invitro. Aluminium chloride increased TBARS level, andchanged the activities of antioxidant enzymes, transam-inases and acid phosphatase. It also caused a decline
in sperm motility and viability more likely throughother mechanisms than oxidative injury, which requiremore investigation. The use of vitamin C or vitamin Ein combination with aluminium chloride alleviated the
M.I. Yousef et al. / Toxicology 239 (2007) 213–223 221
F d ACPw gnificanw Vit. E.
da
R
A
A
A
A
A
A
ig. 6. Changes in TBARS levels, activities of SOD, CAT, AST, ALT anith vitamin E (Vit. E 2 mM) (mean ± S.E.). Asterisk (*) indicates siithout Vit. E or in combination with Vit. E compared to control with
isturbing effects on free radicals level and enzymaticctivity, but not on motility and viability.
eferences
bubakar, M.G., Taylor, A., Ferns, G.A.A., 2004. Regional accumu-lation of aluminium in the rat brain is affected by dietary vitaminE. J. Trace Elem. Med. Biol. 18, 53–59.
itken, R.J., 1995. Free radicals, lipid peroxidation and sperm func-tion. Reprod. Fertil. Dev. 7, 659–668.
keson, M.A., Munns, D.N., 1989a. Lipid bilayer permeation by neu-tral aluminum citrate and by three alpha-hydroxy carboxylic acids.Biochim. Biophys. Acta 984, 200–206.
keson, M.A., Munns, D.D., 1989b. Lipid bilayer permeation by neu-tral aluminum citrate and by three alpha-hydroxy carboxylic acids.Biochim. Biophys. Acta 984, 200–206.
lder, A.J., Caruso, C., Berlyne, G.M., 1995. The effect of alu-
minum on the vanadium-mediated oxidation of NADH. Nephron69, 34–40.remu, D.A., Meshitsuka, S., 2005. Accumulation of aluminum byprimary cultured astrocytes from aluminum amino acid complexand its apoptotic effect. Brain Res. 1031, 284–296.
in rabbit sperm after 4 h incubation with AlCl3 alone or in combinationt (p < 0.05) effect of treatment with AlCl3 alone compared to control
Atienzar, F., Desor, D., Burnel, D., Keller, J.M., Lehr, P., Vasseur, P.,1998. Effect of aluminum on superoxide dismutase activity in theadult rat brain. Biol. Trace Elem. Res. 65, 19–30.
Bataineh, H., Al-Hamood, M.H., Elbetieha, A.M., 1998. Assessmentof aggression, sexual behavior and fertility in adult male rat fol-lowing long-term ingestion of four industrial metals salts. Hum.Exp. Toxicol. 17, 570–579.
Blokhina, O., Virolainen, E., Fagerstedt, K.V., 2003. Antioxidants,oxidative damage and oxygen derivative stress: a review. Ann. Bot.91, 179–194.
Campbell, A., Prasad, K.N., Bondy, S.C., 1999. Aluminum-inducedoxidative events in cell lines: glioma are more responsive thanneuroblastoma. Free Radic. Biol. Med. 26, 1166–1171.
Chinoy, N.J., Momin, R., Jhala, D.D., 2005a. Fluoride and aluminiuminduced toxicity in mice epididymis and its mitigation by vitaminC. Fluoride 38, 115–121.
Chinoy, N.J., Momin, R., Sorathia, H.P., Jhala, D.D., 2005b. Recov-ery from fluoride plus aluminium toxicity in vas deferens, seminal
vesicle, and ventral prostate of mice by vitamin C. Fluoride 38,122–126.Dawson, E.B., Ritter, S., Harris, W.A., Evans, D.R., Powell, L.C., 1998.Comparison of sperm viability with seminal plasma metal levels.Biol. Trace Elem. Res. 64, 215–223.

icology
222 M.I. Yousef et al. / ToxDhami, A.J., Sahni, K.L., Mohan, G., Tripathi, R.P., 1994. Compara-tive evaluation of initially static and motile semen ejaculate fromFrieesian and Murrah buffalo bulls for physico-morphological, bio-chemical, enzymatic and mineral constituents of seminal plasma.Indian J. Anim. Sci. 64, 926–932.
Domenech, C.E., Lisa, T.A., Salvano, M.A., Garrido, M.N., 1992.Pseudomonas aeruginosa acid phosphatase activation by divalentcations and inhibition by aluminium ion. Fed. Eur. Biochem. Soc.299, 96–98.
Domingo, J.L., 2006. Aluminum and other metals in Alzheimer’sdisease: a review of potential therapy with chelating agents. J.Alzheimer’s Dis. 10, 331–341.
Domingo, J.L., Paternain, J.L., Llobet, J.M., Corbella, J., 1987. Effectsof oral aluminum administration on perinatal and postnatal devel-opment in rats. Res. Commun. Chem. Pathol. Pharmacol. 57,129–132.
Exley, C., 2004. The pro-oxidant activity of aluminum. Free Radic.Biol. Med. 36, 380–387.
Guo, C.H., Lin, C.Y., Yeh, M.S., Hsu, G.S.W., 2005. Aluminum-induced suppression of testosterone through nitric oxideproduction in male mice. Environ. Toxicol. Pharmacol. 19, 33–40.
Gutteridge, J.M.C., Quinlan, G.J., Clark, I., Halliwell, B., 1985.Aluminium salts accelerate peroxidation of membrane lipids stim-ulated by iron salts. Biochim. Biophys. Acta 835, 441–447.
Hovatta, O., Venalainen, E., Kuusimaki, L., Heikkila, J., Hirvi, T.,Reima, I., 1998. Aluminium, lead and cadmium concentrations inseminal plasma and spermatozoa, and semen quality in Finnishmen. Hum. Reprod. 13, 115–119.
Kong, S., Liochev, S., Fridovich, I., 1992. Aluminum(III) facilitatesthe oxidation of NADH by the superoxide anion. Free Radic. Biol.Med. 13, 79–81.
Li, J.G., Zheng, W., 2005. Regulation of neuroactive metals bythe choroid plexus. In: Zheng, W., Chodobski, A. (Eds.), TheBlood–Cerebrospinal Barrier. CRC Press, New York, pp. 211–239.
Llobet, J.M., Colomina, M.T., Sirvent, J.J., Domingo, J.L., Corbella,J., 1995. Reproductive toxicology of aluminum in male mice. Fun-dam. Appl. Toxicol. 25, 45–51.
Luck, H., 1974. Catalase. In: Bergmayer, M.V. (Ed.), Method of Enzy-matic Analysis. Verlag Chemic/Academic Press, New York, p.885.
Luck, M.R., Jeyaseelan, I., Scholes, R.A., 1995. Minireview: ascorbicacid and fertility. Biol. Reprod. 52, 262–266.
Mendez-Alvarez, E., Soto-Otero, R., Hermida-Ameijeiras, A., Lopez-Real, A.M., Labandeira-Garcia, J.L., 2002. Effects of aluminumand zinc on the oxidative stress caused by 6-hydroxydopamineautoxidation: relevance for the pathogenesis of Parkinson’s disease.Biochim. Biophys. Acta 1586, 155–168.
Misra, H., Fridovich, I., 1972. The role of superoxide anion in theautooxidation of epinephrine and a simple assay for superoxidedismutase. J. Biol. Chem. 247, 3170–3175.
Moss, D.W., 1984. In: Bergmeyer, H.U. (Ed.), Methods of EnzymaticAnalysis, vol. 4, 3rd ed. Verlag-Chemie, pp. 92–106.
Okhawa, H., Ohishi, W., Yagi, K., 1979. Assay formulation lipidperoxides in animal tissues by thiobarbituric acid reaction. Anal.Biochem. 95, 351.
Oteiza, P.I., Fraga, C.G., Keen, C.L., 1993. Aluminum has both oxidantand antioxidant effects in mouse brain membranes. Arch. Biochem.Biophys. 300, 517–521.
Roca, J., Martinez, S., Vazquez, J.M., Lucas, X., Parrilla, I., Martinez,E.A., 2000. Viability and fertility of rabbit spermatozoa diluted in
239 (2007) 213–223
Tris-buffer extenders and stored at 15 ◦C. Anim. Reprod. Sci. 64,103–112.
Roig, J.L., Fuentes, S., Colomina, M.T., Vicens, P., Domingo, J.L.,2006. Aluminum, restraint stress and aging: behavioral effects inrats after 1 and 2 years of aluminum exposure. Toxicology 218,112–124.
SAS, 1986. Statistical analysis system. In: SAS User’s Guide: Statis-tics, 5th ed. SAS Inst., Inc., Cary, NC, USA.
Savory, J., Herman, M.M., Ghribi, O., 2006. Mechanisms ofaluminum-induced neurodegeneration in animals: implications forAlzheimer’s disease. J. Alzheimer’s Dis. 10, 135–144.
Shi, B., Haug, A., 1988. Uptake of aluminum by lipid vesicles. Toxicol.Environ. Chem. 17, 337–349.
Shi, B., Haug, A., 1990. Aluminum uptake by neuroblastoma cells. J.Neurochem. 55, 551–558.
Steel, R.G.D., Torrie, J.H., 1981. Principles and Procedures of Statis-tics, 2nd ed. McGraw-Hill, Singapore.
Strzegek, J., 1988. Morphological structures of spermatozoa andtheir biological functions. Biul. Inf. ART (Akademia Rolniczo-Techniczna) Olsztyn 25, 67–72.
Surai, P., Kostjuk, I., Wishart, G., Macpherson, A., Speake, B., Noble,R., Ionov, I., Kutz, E., 1998. Effect of vitamin E and seleniumsupplementation of cockerel diets on glutathione peroxidase activ-ity and lipid peroxidation susceptibility in sperm, testes, and liver.Biol. Trace Elem. Res. 64, 119–132.
Suwalsky, M., Norris, B., Villena, F., Cuevas, F., Sotomayor, P., Zatta,P., 2004. Aluminum fluoride affects the structure and functions ofcell membranes. Food Chem. Toxicol. 42, 925–933.
Verma, A., Kanwar, K.C., 1998. Human sperm motility and lipid per-oxidation in different ascorbic acid concentrations; an in vitroanalysis. Andrology 30, 325–329.
Verma, A., Kanwar, K.C., 1999. Effect of vitamin E on humansperm motility and lipid peroxidation in vitro. Asian J. Androl.1, 151–154.
Verstraeten, S.V., Oteiza, P.I., 2000. Effects of Al(3+) and related met-als on membrane phase state and hydration: correlation with lipidoxidation. Arch. Biochem. Biophys. 375, 340–346.
Wilhelm, M., Jaeger, D.E., Schull-Cablitz, H., Hafner, D., Idel, H.,1996. Hepatic clearance and retention of aluminium: studies in theisolated perfused rat liver. Toxicol. Lett. 89, 257–263.
Xie, C.X., Mattson, M.P., Lovell, M.A., Yokel, R.A., 1996. Intra-neuronal aluminum potentiates iron-induced oxidative stress incultured rat hippocampal neurons. Brain Res. 743, 271–277.
Yokel, R.A., 2002. Brain uptake, retention, and efflux of aluminum andmanganese. Environ. Health Perspect. 110, 699–703.
Yokel, R.A., McNamara, P.J., 2001. Aluminium toxicokinetics: anupdated mini review. Pharmacol. Toxicol. 88, 159–167.
Yokel, R.A., Wilson, M., Harris, W.R., Halestrap, A.P., 2002. Alu-minum citrate uptake by immortalized brain endothelial cells:implications for its blood–brain barrier transport. Brain Res. 930,101–110.
Yousef, M.I., 2004. Aluminium-induced changes in hemato-biochemical parameters, lipid peroxidation and enzyme activitiesof male rabbits: protective role of ascorbic acid. Toxicology 199,47–57.
Yousef, M.I., Abdallah, G.A., Kamel, K.I., 2003. Effect of ascorbic acidand vitamin E supplementation on semen quality and biochemical
parameters of male rabbits. Anim. Reprod. Sci. 76, 99–111.Yousef, M.I., El-Morsy, A.M.A., Hassan, M.S., 2005. Aluminium-induced deterioration in reproductive performance and seminalplasma biochemistry of male rabbits: protective role of ascorbicacid. Toxicology 215, 97–107.

icology
Z
Z
Zhang, J.G., Nicholls-Grzemski, F.A., Tirmenstein, M.A., Fariss,
M.I. Yousef et al. / Tox
atta, P., Lain, E., Cagnolini, C., 2000. Effects of aluminum on activity
of Krebs cycle enzymes and glutamate dehydrogenase in rat brainhomogenate. Eur. J. Biochem. 267, 3049–3055.atta, P., Kiss, T., Suwalsky, M., Berthon, G., 2002. Aluminium(III)as a promoter of cellular oxidation. Coord. Chem. Rev. 228, 271–284.
239 (2007) 213–223 223
M.W., 2001. Vitamin E succinate protects hepatocytes against thetoxic effect of reactive oxygen species generated at mitochondrialcomplexes I and III by alkylating agents. Chem. Biol. Interact. 138,267–284.
Copyright © 2022 FDOKUMEN




















