
An fMRI investigation of working memory and sadness in females with bipolar disorder: a brief report
15
Brief Report An fMRI investigation of working memory and sadness in females with bipolar disorder: a brief report Neuropsychological studies have documented cog- nitive impairments in individuals with bipolar disorder (1–3), including difficulties with attention, memory, and executive functioning (4–15). One of the most consistently reported cognitive impair- ments in bipolar disorder is memory disturbance (e.g., 9, 12–14, 16). Previous studies by our group found that individuals with bipolar disorder have difficulties organizing information in working memory during encoding (the process of establish- ing a new memory representation) (17–20). This was associated with attenuated regional cerebral blood flow (rCBF) in the dorsolateral prefrontal cortex (DLPFC) (20). Functional magnetic reso- nance imaging (fMRI) studies of working memory Deckersbach T, Rauch SL, Buhlmann U, Ostacher MJ, Beucke J-C, Nierenberg AA, Sachs G, Dougherty DD. An fMRI investigation of working memory and sadness in females with bipolar disorder: a brief report. Bipolar Disord 2008: 10: 928–942. ª 2008 The Authors Journal compilation ª 2008 Blackwell Munksgaard Objective: Functional magnetic resonance imaging (fMRI) studies have documented abnormalities in the dorsolateral prefrontal cortex (DLPFC) and anterior cingulate cortex in bipolar disorder in the context of working memory tasks. It is increasingly recognized that DLPFC regions play a role in mood regulation and the integration of emotion and cognition. The purpose of the present study was to investigate with fMRI the interaction between acute sadness and working memory functioning in individuals with bipolar disorder. Methods: Nine depressed individuals with DSM-IV bipolar I disorder (BP-I) and 17 healthy control participants matched for age, gender, education, and IQ completed a 2-back working memory paradigm under no mood induction, neutral state, or acute sadness conditions while undergoing fMRI scanning. Functional MRI data were analyzed with SPM2 using a random-effects model. Results: Behaviorally, BP-I subjects performed equally well as control participants on the 2-back working memory paradigm. Compared to control participants, individuals with BP-I were characterized by more sadness-specific activation increases in the left DLPFC (BA 9 ⁄ 46) and left dorsal anterior cingulate (dACC). Conclusions: Our study documents sadness-specific abnormalities in the left DLPFC and dACC in bipolar disorder that suggest difficulties in the integration of emotion (sadness) and cognition. These preliminary findings require further corroboration with larger sample sizes of medication-free subjects. Thilo Deckersbach a , Scott L Rauch a,b , Ulrike Buhlmann a , Michael J Ostacher a , Jan-Carl Beucke a,c , Andrew A Nierenberg a , Gary Sachs a and Darin D Dougherty a a Department of Psychiatry, Massachusetts General Hospital and Harvard Medical School, Boston, b McLean Hospital, and Harvard Medical School, Belmont, MA, USA, c Department of Psychology, Humboldt University, Berlin, Germany Key words: bipolar disorder – fMRI – functional magnetic resonance imaging – sadness – working memory Received 12 September 2007, revised and accepted for publication 18 May 2008 Corresponding author: Thilo Deckersbach, PhD, Department of Psychiatry, 149-2628, Massachusetts General Hospital, Building 149, 13th Street, 2nd Floor, Charlestown, MA 02129, USA. Fax: 617 726 4078; e-mail: [email protected] Part of this research was presented at the annual meeting of the Seventh International Conference on Bipolar Disorder, June 7–9, 2007, Pittsburgh, PA, USA. The authors of this paper do not have any commercial associations that might pose a conflict of interest in connection with this manu- script. Bipolar Disorders 2008: 10: 928–942 ª 2008 The Authors Journal compilation ª 2008 Blackwell Munksgaard BIPOLAR DISORDERS 928
-
Upload
hms-harvard -
Category
Documents
-
view
0 -
download
0
Transcript of An fMRI investigation of working memory and sadness in females with bipolar disorder: a brief report

Brief Report
An fMRI investigation of working memoryand sadness in females with bipolar disorder:a brief report
Neuropsychological studies have documented cog-nitive impairments in individuals with bipolardisorder (1–3), including difficulties with attention,memory, and executive functioning (4–15). One ofthe most consistently reported cognitive impair-
ments in bipolar disorder is memory disturbance(e.g., 9, 12–14, 16). Previous studies by our groupfound that individuals with bipolar disorder havedifficulties organizing information in workingmemory during encoding (the process of establish-ing a new memory representation) (17–20). Thiswas associated with attenuated regional cerebralblood flow (rCBF) in the dorsolateral prefrontalcortex (DLPFC) (20). Functional magnetic reso-nance imaging (fMRI) studies of working memory
Deckersbach T, Rauch SL, Buhlmann U, Ostacher MJ, Beucke J-C,Nierenberg AA, Sachs G, Dougherty DD. An fMRI investigation ofworking memory and sadness in females with bipolar disorder: a briefreport.Bipolar Disord 2008: 10: 928–942. ª 2008 The Authors Journalcompilation ª 2008 Blackwell Munksgaard
Objective: Functional magnetic resonance imaging (fMRI) studieshave documented abnormalities in the dorsolateral prefrontal cortex(DLPFC) and anterior cingulate cortex in bipolar disorder in the contextof working memory tasks. It is increasingly recognized that DLPFCregions play a role in mood regulation and the integration of emotionand cognition. The purpose of the present study was to investigate withfMRI the interaction between acute sadness and working memoryfunctioning in individuals with bipolar disorder.
Methods: Nine depressed individuals with DSM-IV bipolar I disorder(BP-I) and 17 healthy control participants matched for age, gender,education, and IQ completed a 2-back working memory paradigm underno mood induction, neutral state, or acute sadness conditions whileundergoing fMRI scanning. Functional MRI data were analyzed withSPM2 using a random-effects model.
Results: Behaviorally, BP-I subjects performed equally well as controlparticipants on the 2-back working memory paradigm. Compared tocontrol participants, individuals with BP-I were characterized by moresadness-specific activation increases in the left DLPFC (BA 9 ⁄ 46) andleft dorsal anterior cingulate (dACC).
Conclusions: Our study documents sadness-specific abnormalities inthe left DLPFC and dACC in bipolar disorder that suggest difficulties inthe integration of emotion (sadness) and cognition. These preliminaryfindings require further corroboration with larger sample sizes ofmedication-free subjects.
Thilo Deckersbacha, Scott LRaucha,b, Ulrike Buhlmanna,Michael J Ostacher a, Jan-CarlBeuckea,c, Andrew A Nierenberga,Gary Sachsa and Darin DDoughertya
aDepartment of Psychiatry, Massachusetts General
Hospital and Harvard Medical School, Boston,bMcLean Hospital, and Harvard Medical School,
Belmont, MA, USA, cDepartment of Psychology,
Humboldt University, Berlin, Germany
Key words: bipolar disorder – fMRI – functional
magnetic resonance imaging – sadness – working
memory
Received 12 September 2007, revised and
accepted for publication 18 May 2008
Corresponding author: Thilo Deckersbach, PhD,
Department of Psychiatry, 149-2628,
Massachusetts General Hospital, Building 149,
13th Street, 2nd Floor, Charlestown, MA 02129, USA.
Fax: 617 726 4078;
e-mail: [email protected]
Part of this research was presented at the annual
meeting of the Seventh International Conference on
Bipolar Disorder, June 7–9, 2007, Pittsburgh, PA,
USA.
The authors of this paper do not have any commercial associations
that might pose a conflict of interest in connection with this manu-
script.
Bipolar Disorders 2008: 10: 928–942ª 2008 The Authors
Journal compilation ª 2008 Blackwell Munksgaard
BIPOLAR DISORDERS
928

in bipolar disorder have also found abnormalitiesin DLPFC, although the directionality of theabnormalities (i.e., hyper- or hypoactivation) var-ies across studies (21–24). For example, bothMonks et al. (21) and Lagopoulos et al. (22), usingn-back and Sternberg working memory tasks,reported reduced activations in the middle frontalgyrus [including Brodmann area (BA) 9], theinferior frontal gyrus, and the dorsal anteriorcingulate (dACC) in bipolar disorder comparedto normal control participants. Chang et al. (23),on the other hand, found increased activation inthe left DLPFC (BA 9 ⁄46) and dACC in a 2-backworking memory paradigm in a cohort of pediatricpatients with bipolar disorder. Overall, however,findings of abnormal dorsal prefrontal corticalactivation in bipolar disorder in the context offMRI working memory paradigms converge withresults from studies using other imaging techniques(e.g., 25, 26) and postmortem studies (27–30)implicating dorsal prefrontal cortical (in additionto ventral prefrontal) cortical areas in the patho-physiology of bipolar disorder (31).In the context of working memory, the DLPFC
has been described as being a part of a centralexecutive system that regulates the function ofsubordinate working memory systems (i.e., aphonological subsystem for storage and subvocalrehearsal of verbally coded information and avisuospatial sketchpad) (32, 33). More specifically,the DLPFC (BA 9 ⁄46) appears to support work-ing memory operations such as updating andmanipulating contents held in short-term storage(34, 35). The role of the anterior cingulate cortexin cognitive tasks is multifaceted. Dorsal portions(dACC) are part of a distributed attentionalnetwork (36) supporting functions such as re-sponse selection (37–41), performance monitoring(42, 43), error detection, and competition moni-toring (44–46). Rostral (or pregenual) areas serveregulatory functions and are viewed as beinginvolved in the integration of emotion and cogni-tion (47–49). More recently, the DLPFC had beendescribed as having a role in the interaction ofcognition and emotion (50–52). Increased DLPFCactivation has been found during the voluntarysuppression of sadness (50), reappraisal of emo-tive scenes (51), and n-back task performancefollowing the viewing of unpleasant movies (52),which suggests a role for the DLPFC in themodulation of emotion in addition to subservingcognitive processes involved in working memory.This is also reflected in a recent neurobiologicalmodel of bipolar disorder that hypothesizesabnormalities in DLPFC and dACC (amongother dorsal and ventral prefrontal regions) as
mediating impaired regulation and increased labil-ity of mood (31).To our knowledge, the interaction between
cognition and emotion in the context of workingmemory paradigms has never been investigated inbipolar disorder. Concentration and memory dif-ficulties often appear most pronounced duringperiods of depression. A commonly employedstrategy for developing a better understanding ofthe observed dorsal and ventral prefrontal changesduring depressive mood has been the use of moodinduction paradigms (e.g., 48, 53–55). In theseparadigms, subjects undergo transient periods ofsadness by recalling sad or neutral autobiograph-ical events while brain activity is recorded usingpositron emission tomography, single-photonemission computerized tomography, or fMRI.Studies employing this paradigm have revealedincreases in rCBF during sadness (versus neutralstate) in ventral regions including the subgenualprefrontal cortex, rostral ACC, insula, and ventralprefrontal cortex (e.g., 48, 53, 54), whereasdecreased rCBF associated with sad mood hasbeen reported for the DLPFC and dorsal ACC(e.g., 48, 53, 54), among other regions. Fordepressed individuals, there is evidence that sad-ness alters concurrent cognitive or emotionalprocessing. For example, Keedwell et al. (56)recently reported increased DLPFC and decreasedventral prefrontal activation in depressed individ-uals (compared to normal control participants) toemotionally valenced stimuli following sad moodinduction, suggesting that cognitive processing isaltered by induced sadness in depressive states.The overall purpose of the present study was
to investigate the interaction between sadness andn-back working memory performance in de-pressed patients with bipolar disorder andhealthy individuals using fMRI. N-back workingmemory tasks require participants to monitoritems on a screen and decide whether a presenteditem is identical to an item presented one, two,or three trials back (e.g., 2-back paradigm: L, M,L: correct answer = �yes�; L, M, P: correctanswer = �no�; see Fig. 1). N-back workingmemory paradigms have reliably yielded robustactivations in the DLPFC and dACC (57–59).Specifically, previous studies have shown a lin-ear increase in activations in the dACC andDLPFC with increasing working memory loadpresumably reflecting increased demands onholding information online, monitoring, manipu-lating, and updating of information (e.g., 59). Wedesigned an n-back working memory task thatvaried the demand on these working memoryprocesses by including a no-mood induction
fMRI investigation of working memory and sadness
929

condition as well as two conditions where par-ticipants completed a 2-back task during neutralstate and sadness induced via prerecorded auto-biographical scripts. We aimed to identify regionsin the dACC and DLPFC that show a linearincrease across the three conditions. We pre-dicted that, in these regions, similar to previousn-back fMRI studies in bipolar disorder, patientswith bipolar disorder would show abnormallyincreased activation during no-mood induction orneutral state, but that these activation abnormal-ities would be particularly pronounced during thesadness condition.
Methods
Participants
Study participants were 17 healthy control partic-ipants (all female) and nine individuals with a
primary diagnosis of DSM-IV bipolar I disorder(BP-I, all female) who met criteria for a majordepressive episode. BP-I participants wererecruited through the Bipolar Clinic and ResearchProgram (BCRP) and the Division of PsychiatricNeuroscience Research and Neurotherapeutics atthe Massachusetts General Hospital, Boston, MA,USA. Healthy control participants were recruitedthrough bulletin board notices within the hospital.None of the control participants were related to theBP-I participants. Diagnostic assessment of allparticipants was performed using the StructuredClinical Interview for DSM-IV (SCID) (60). Allparticipants provided written informed consentprior to participation in accordance with theguidelines of the Subcommittee on Human Studiesof the Massachusetts General Hospital. All BP-Iparticipants were free of current comorbid diag-noses. Two BP-I participants had a history of panicdisorder. At the time of the MRI scan, all BP-I
Run 1
Run 2
L
M
L
P
T
A
B
Target (‘yes’)
Foil (‘no’)
Time
0 0:30 1:30 2:00 4:30 5:30 6:00 8:30 9:00 10:00 Min
0 0:30 1:30 2:00 4:30 5:30 6:00 8:30 9:00 10:00 Min
+
+ 2-back no mood induction
2-back no mood induction
2-back neutral state
induction
2-back neutral state
induction
2-back sadness induction
2-back sadness state
induction
Listening toautobiographicalscript (30 sec)
Listening toautobiographicalscript (30 sec)
Listening toautobiographicalscript (30 sec)
Listening toautobiographicalscript (30 sec)
2-back no mood induction
2-back no mood induction
2-back no mood induction
2-back no mood induction
+
+
Fig. 1. A. Schematic view of the 2-back working memory task. Letters were shown in the center of a computer screen (one letter at atime). Participants were asked to decide via key press whether a given letter is identical to the letter presented two letters back (e.g., L,M, L: correct answer = �yes� = Target; L, M, P: correct answer = �no� = Foil). B. Structure of each run. The order of the 2-back–neutral state and 2-back–sadness conditions were counterbalanced.
Deckersbach et al.
930

participants were taking mood-stabilizing medica-tion (lithium, n = 5; lamictal, n = 2; valproicacid, n = 2).After enrollment, the mood of all BP-I partici-
pants was prospectively monitored for one weekusing the Hamilton Rating Scale for Depression(HAM-D) (61) and the Young Mania RatingScale (YMRS) (62). The BP-I group exhibited amoderate level of depression, while mania symp-toms as assessed by the YMRS were low (seeTable 1). All participants were medically healthyand had no history of significant head injury,seizure, neurologic condition, or current majormedical condition by report. BP-I patients whohad endocrinological disorders were not includedin the study. All participants were right handed(63). BP-I and control participants were alsomatched with respect to their stage in the men-strual cycle at the time of scanning. The groups didnot differ significantly with regard to age(t = )0.90, df = 24, p = 0.38; see Table 1), yearsof education (t = )0.16, df = 24, p = 0.88; seeTable 1), or IQ as estimated by the Wechsler Testof Adult Reading (WTAR) (64) (t = 1.38,df = 24, p = 0.18).
Following the diagnostic evaluation, studyparticipants prepared descriptions about sadness-provoking and neutral life events. These autobio-graphical scripts were prepared according to apreviously published procedure (65). Briefly, eachsubject wrote two paragraphs regarding her mostand second-most sadness-producing life experi-ences (e.g., �the day my mother died�) and of twoautobiographical neutral events (e.g., going for awalk). Based on the information provided, second-person, present-tense scripts were written andrecorded in a neutral voice for playback in theMRI scanner as part of the 2-back workingmemory paradigm (see below). All autobiograph-ical scripts for inducing sadness or neutral statewere 30 seconds long.
Functional MRI paradigm
N-back working memory. Participants completed a2-back working memory task, modeled afterBraver et al. (59), during sad and neutral stateswhile undergoing fMRI scanning. In the 2-backtask, participants were shown letters in the centerof a computer screen (one letter at a time) and wereasked to decide via key press whether a given letterwas identical to the letter presented two lettersback (e.g., L, M, L: correct answer = �yes�; L, M,P: correct answer = �no�; see Fig. 1). A given letterwas presented for 1.5 sec, followed by a blankscreen for 1.5 sec before the next letter wouldappear on the screen.The 2-back working memory paradigm included
three conditions: (i) 2-back working memory–nomood induction, (ii) 2-back working memory–neutral state induction, and (iii) 2-back workingmemory–sadness induction (see Fig. 1B). In each ofthe three conditions, subjects completed the 2-backtask as described above. Each of the three 2-backworking memory conditions included 18 targets (agiven letter is the same as two letters back) and 42foils (a given letter is not the same as two lettersback). In the 2-back working memory–no moodinduction condition, subjects completed the 2-backparadigm described above for a total of threeminutes without any neutral or sadness induction.In the 2-back working memory–neutral stateinduction, subjects completed the 2-back taskduring neutral state induction for three minutes.In the 2-back working memory–sadness induction,subjects completed the 2-back task during sadnessinduction for three minutes. The induction ofneutral state or sadness was conducted as follows:at the beginning of the three-minute period,subjects listened to the prerecorded autobiograph-ical script for 30 sec and then continued in that
Table 1. Demographic and behavioral characteristics of bipolar I disorderand control subjects
ControlsMean (SD)
Bipolar I disorderMean (SD)
Demographics
Age 25.6 (5.9) 27.6 (2.8)Education 16.1 (0.6) 16.2 (2.7)IQ 114.4 (4.7) 111.8 (4.2)HAM-D 0.6 (1.5) 25.2 (4.8)YMRS 0.2 (0.5) 2.8 (1.1)Measures
N-back reaction timesa
No mood induction 752.89 (122.51) 823.17 (119.90)Neutral state induction 787.64 (134.26) 856.28 (124.36)Sadness induction 796.90 (142.94) 886.39 (123.97)
N-back response accuracyb
No mood induction 94.43 (3.27) 92.73 (3.39)Neutral state induction 91.09 (4.27) 90.57 (4.49)Sadness induction 87.25 (7.66) 86.65 (8.98)
Visual analog scalesc
Ability to recallNeutral events 7.77 (1.25) 8.17 (1.12)Sad events 8.19 (1.67) 8.41 (1.52)
Sadness ratingNeutral events 1.48 (1.63) 1.10 (1.07)Sad events 8.38 (0.77) 8.18 (1.23)
aReaction times in milliseconds.bResponse accuracy = (number of correct responses ⁄ numberof possible correct responses)*100.cVisual analog scale range of scores from �0� = absent to�10� = maximum.HAM-D = Hamilton Rating Scale for Depression; YMRS = YoungMania Rating Scale.
fMRI investigation of working memory and sadness
931

mood state (sad or neutral) for an additional2.5 min (i.e., a total of three minutes). Participantscompleted the 2-back task for the entire three-minute period while concurrently listening to theprerecorded scripts and remaining in a neutral orsad state. A run included all three 2-back condi-tions. Each three-minute mood induction blockwas bracketed by a one-minute block of the 2-backworking memory–no mood induction condition(see Fig. 1B). The sadness or neutral state induc-tion conditions were presented either in the middleor at the end of the run (see Fig. 1B). At thebeginning and end of each run, a 30-sec fixationcross low-level baseline condition was presented(fixation cross in the center of the computer screen;see Fig. 1B). Each subject completed two runs witha three-minute resting period between the two runs.The order of the 2-back working memory–neutralstate and the 2-back working memory–sadnesswere counterbalanced (see Fig. 1B: e.g., run 1:neutral followed by sad; run 2: sad followed byneutral). Behavioral measures included (i) reactiontimes and (ii) response accuracy; we computedmean reaction times and percent response accuracyfor each of the conditions (no mood induction,neutral state induction, sadness induction) acrossthe two runs. Response accuracy was calculated bydividing the number of correct responses by thenumber of total possible correct responses multi-plied by 100.
Visual analog scales. After the scan, for each run,for each of the 2-back working memory–neutralstate conditions and the 2-back working memory–sadness induction conditions, subjects rated thedegree to which they could recall the eventpresented during neutral state or sadness and thedegree of sadness they experienced on a 0-10analog scale; 0 indicated a complete absence ofsadness and 10 indicated the maximum possibleexperience of sadness; similarly, 0 indicated acomplete inability to recall the event and 10indicated the maximum possible ability to recallthe event.
MRI scanning
MRI data were acquired using a 3.0-T whole-bodyscanner (Allegra; Siemens Medical Solutions)equipped for echo planar imaging (Siemens Med-ical Systems, Iselin, NJ, USA) with a 3-axis gradienthead coil. Head movements were restricted usingfoam cushions. Images were projected using a rearprojection system and Presentation stimulus pres-entation software (Neurobehavioral Systems, Inc.;http://www.neurobs.com). Following automated
scout and shimming procedures, two high-resolu-tion 3D MPRAGE sequences [repetition time(TR) = 2.53 ms, echo time (TE) = 3.45 ms, flipangle = 7o, voxel size = 1.3 · 1.0 · 1.3 mm]were collected for positioning of subsequent scans.Functional MRI images [i.e., blood oxygenationlevel dependent (BOLD) signal] were acquiredusing T2*-weighted sequence (20 coronal slicesaligned perpendicular to the plane intersecting theanterior and posterior commissures, 5 mm thick-ness, skip 1 mm, TE = 30 ms, TR = 1.5, flipangle = 90o).
MRI data analysis
Prior to the analysis, the first four volumesfrom each run were discarded to allow for T1equilibrium effects. Functional data were processedusing SPM2 software (Wellcome Department ofCognitive Neurology, London, UK; http://www.fil.ion.ucl.ac.uk/spm). Functional MRI imageswere motion corrected, spatially normalized tothe standardized space established by the MontrealNeurologic Institute (MNI) (http://www.bic.mni.mcgill.ca), resampled to 2 mm3 voxels andsmoothed with a three-dimensional Gaussiankernel of 6-mm width (full width half maximum).All collected data had minimal head motion(<3 mm linear movement in the orthogonalplanes; <0.5 degrees radians of angular move-ment). The general linear model was applied to thetime series, convolved with the canonical hemody-namic response (66) function and a 128s high-passfilter. For each subject, condition effects weremodeled with box-car regressors representing theoccurrence of each block type (2-back–no moodinduction, 2-back–neutral state induction, 2-back–sadness induction). For each subject, conditioneffects were estimated at each voxel, and statisticalparametric maps (SPMs) were produced for thecontrast of interest. Specifically, we implemented acontrast that reflected a linear increase in betaweights from the fixation condition to the 2-back–no mood induction, 2-back–neutral state inductionto the 2-back–sadness induction. For group anal-ysis, each subject�s contrast image (SPM) wasentered into a second-level random-effects analysis.For the a-priori specified regions of interest (ROI)(anterior cingulate, DLPFC; for details regardingthe ROI definitions, see below), we first identifiedloci of activation for the linear contrast of interest(main effect condition). This was achieved byentering the SPM contrast images in a one-samplet-test collapsed over patient and control partici-pants. (Note: main effects are independent fromgroup-by-condition interactions.) This analysis was
Deckersbach et al.
932

performed using a stringent statistical threshold ofp < 0.05 family-wise error (FWE) corrected (67)with the added requirement that at least threecontiguous voxels exceeded this statistical level(i.e., k ‡ 3). This corresponded to a voxel-levelp-value for statistical significance of z = 4.42. Forclusters of activation that exceeded the threshold,we extracted parameter estimates (beta weights) asan index of blood oxygenation level-dependentsignal change using MarsBaR (http://marsbar.sourceforge.net) (68). BP-I and control partici-pants were then compared using two-tailed t-testsfor between-group comparisons with an uncor-rected threshold of p = 0.05. The a-priori specifiedROIs were defined using masks provided by theAnatomical Automatic Labeling (69) tool imple-mented in the Wake Forest University PickAtlashttp://www.ansir.wfubmc.edu) (70, 71). For theanterior cingulate this involved Cingulum_Ant (leftand right). For the dorsal prefrontal cortex thisinvolved combined Frontal_Sup, Frontal_Mid,and Frontal Inf_Tri (left and right). Followingthese analyses, in a post-hoc fashion, we exploredthe effects of 2-back working memory–moodinduction, and sadness in a series of independentcontrasts within and across groups. Specifically, wetested activations associated with the 2-back work-ing memory task (all 2-back working memoryconditions combined versus fixation cross), moodinduction (2-back working memory–neutral stateand sadness induction combined versus 2-backworking memory–no mood induction) and sadness(2-back working memory–sadness induction versus2-back working memory–neutral state induction).For this exploratory, whole brain, non-FWE-corrected analysis, we adopted a voxel-wise thresh-old of z = 3.09 (p < 0.001) with the addedrequirement that at least five contiguous voxelsexceeded this statistical level (i.e., k ‡ 5).
Behavioral data statistical analysis
N-back reaction times and response accuracy wereevaluated by mixed model analysis of variance(ANOVA) with condition (2-back–no mood induc-tion versus 2-back–neutral state induction versus2-back–sadness induction) as the within-subjectsand group (controls versus BP-I) as the between-subjects factor. Visual analog scale ratings(sadness, ability to recall) were evaluated usingmixed model analysis of variance with condition(sad versus neutral) as the within-subjects factorand group (controls versus BP-I) as the between-subjects factor. Significant interactions werefollowed up by linear contrasts and t-tests.Correlations between BOLD data and depressive
symptoms (HAM-D scores) were investigatedcomputing Spearman rank correlations.
Results
Behavioral data
N-back. Reaction times and response accuracy areshown in Table 1. For reaction times, the mixedmodel ANOVA indicated a main effect of condi-tion (F = 12.65, df = 2,48, p < 0.001) but nomain effect of group (F = 2.13, df = 1,24,p ¼ 0.16) and no group-by-condition interaction(F = 0.57, df = 2,48 p = 0.57). A follow-up lin-ear contrast indicated that there was a linearslowing in reaction times from the 2-back–nomood induction to the 2-back–neutral state induc-tion to the 2-back–sadness induction condition(F = 17.37, df = 1,24, p < 0.001). For responseaccuracy, there was a main effect of condition(F = 12.57, df = 2,48, p < 0.001), but no maineffect of group (F = 0.29, df = 1,24, p = 0.60)and no group-by-condition interaction (F = 0.12,df = 2,48, p = 0.89). Follow-up t-tests indicatedthat both groups exhibited lower response accu-racy during the 2-back–neutral state conditioncompared to the 2-back–no mood inductioncondition (t = 4.21, df = 25, p < 0.001) andexhibited lower response accuracy in the 2-back–sadness induction condition compared to the2-back–neutral state condition (t = 3.03, df = 25,p = 0.006).
Visual analog scales. For sadness, the mixed modelANOVA yielded a main effect of condition indicat-ing that participants experienced more sadnessduring the 2-back–sadness condition compared tothe 2-back–neutral state condition (F = 344.24,df = 1,24, p < 0.001). There was no main effectof group (F = 0.68, df = 1,24, p = 0.42) andno group-by-condition interaction (F = 0.06,df = 1,24, p = 0.82). For the ability to recall, therewas no main effect of group (F = 0.33, df = 1,24,p = 0.57), no main effect of condition (F = 1,75,df = 1,24, p = 0.20), and no group-by-conditioninteraction (F = 0.11, df = 1,24, p = 0.74).
Functional MRI results
Linear contrast. Regions of linear increases(fixation < no mood induction < neutral state< sadness) collapsed over control and BP-Iparticipants are shown in Table 2 and Fig. 2.Consistent with our a-priori hypothesis, there wasa significant increase in BOLD signal in the dorsalanterior cingulate (BA 24 and 32; see Fig. 2A) and
fMRI investigation of working memory and sadness
933
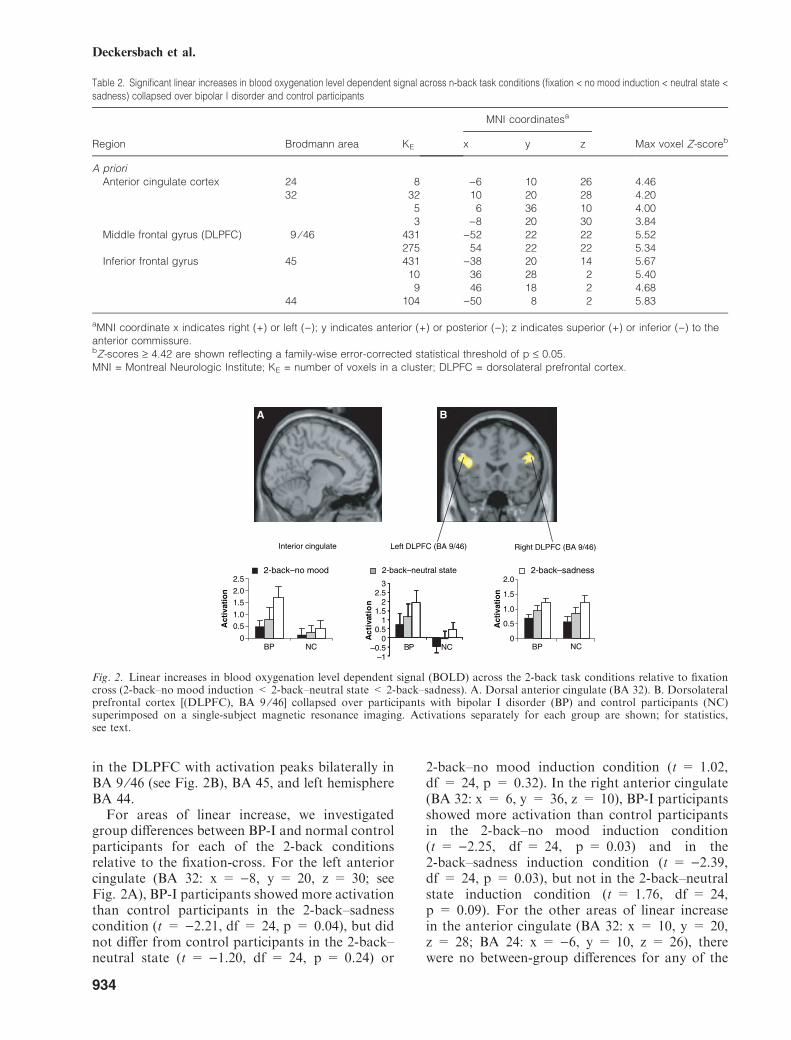
in the DLPFC with activation peaks bilaterally inBA 9 ⁄46 (see Fig. 2B), BA 45, and left hemisphereBA 44.For areas of linear increase, we investigated
group differences between BP-I and normal controlparticipants for each of the 2-back conditionsrelative to the fixation-cross. For the left anteriorcingulate (BA 32: x = )8, y = 20, z = 30; seeFig. 2A), BP-I participants showed more activationthan control participants in the 2-back–sadnesscondition (t = )2.21, df = 24, p = 0.04), but didnot differ from control participants in the 2-back–neutral state (t = )1.20, df = 24, p = 0.24) or
2-back–no mood induction condition (t = 1.02,df = 24, p = 0.32). In the right anterior cingulate(BA 32: x = 6, y = 36, z = 10), BP-I participantsshowed more activation than control participantsin the 2-back–no mood induction condition(t = )2.25, df = 24, p = 0.03) and in the2-back–sadness induction condition (t = )2.39,df = 24, p = 0.03), but not in the 2-back–neutralstate induction condition (t = 1.76, df = 24,p = 0.09). For the other areas of linear increasein the anterior cingulate (BA 32: x = 10, y = 20,z = 28; BA 24: x = )6, y = 10, z = 26), therewere no between-group differences for any of the
Table 2. Significant linear increases in blood oxygenation level dependent signal across n-back task conditions (fixation < no mood induction < neutral state <sadness) collapsed over bipolar I disorder and control participants
Region Brodmann area KE
MNI coordinatesa
Max voxel Z-scorebx y z
A prioriAnterior cingulate cortex 24 8 )6 10 26 4.46
32 32 10 20 28 4.205 6 36 10 4.003 )8 20 30 3.84
Middle frontal gyrus (DLPFC) 9 ⁄ 46 431 )52 22 22 5.52275 54 22 22 5.34
Inferior frontal gyrus 45 431 )38 20 14 5.6710 36 28 2 5.409 46 18 2 4.68
44 104 )50 8 2 5.83
aMNI coordinate x indicates right (+) or left ()); y indicates anterior (+) or posterior ()); z indicates superior (+) or inferior ()) to theanterior commissure.bZ-scores ‡ 4.42 are shown reflecting a family-wise error-corrected statistical threshold of p £ 0.05.MNI = Montreal Neurologic Institute; KE = number of voxels in a cluster; DLPFC = dorsolateral prefrontal cortex.
Left DLPFC (BA 9/46) Right DLPFC (BA 9/46)Interior cingulate
Act
ivat
ion
Act
ivat
ion
0
0.5
1.0
1.5
2.0
BP NC0
0.5
1.0
1.5
2.0
2.5
BP NC
2-back–neutral state2-back–no mood 2-back–sadness
A B
BP–1
–0.50
0.51
1.52
2.53
NC
Act
ivat
ion
Fig. 2. Linear increases in blood oxygenation level dependent signal (BOLD) across the 2-back task conditions relative to fixationcross (2-back–no mood induction < 2-back–neutral state < 2-back–sadness). A. Dorsal anterior cingulate (BA 32). B. Dorsolateralprefrontal cortex [(DLPFC), BA 9 ⁄ 46] collapsed over participants with bipolar I disorder (BP) and control participants (NC)superimposed on a single-subject magnetic resonance imaging. Activations separately for each group are shown; for statistics,see text.
Deckersbach et al.
934

conditions (all p ‡ 0.75). For the left DLPFC(BA 9 ⁄46: x = )52, y = 22, z = 22), BP-I par-ticipants showed a trend toward more activationthan control participants in the 2-back–nomood induction condition (t = 1.96, df = 24,p = 0.06). They showed significantly more activa-tion in the 2-back–sadness-condition (t = 2.59,df = 24, p = 0.02) but did not differ from controlparticipants in the 2-back–neutral state inductioncondition (t = 1.61, df = 24, p = 0.12). Therewere no between-group differences for the rightdorsal prefrontal cortex (BA 9 ⁄46: x = 54,y = 22, z = 22) for any of the conditions (allp ‡ 0.60).
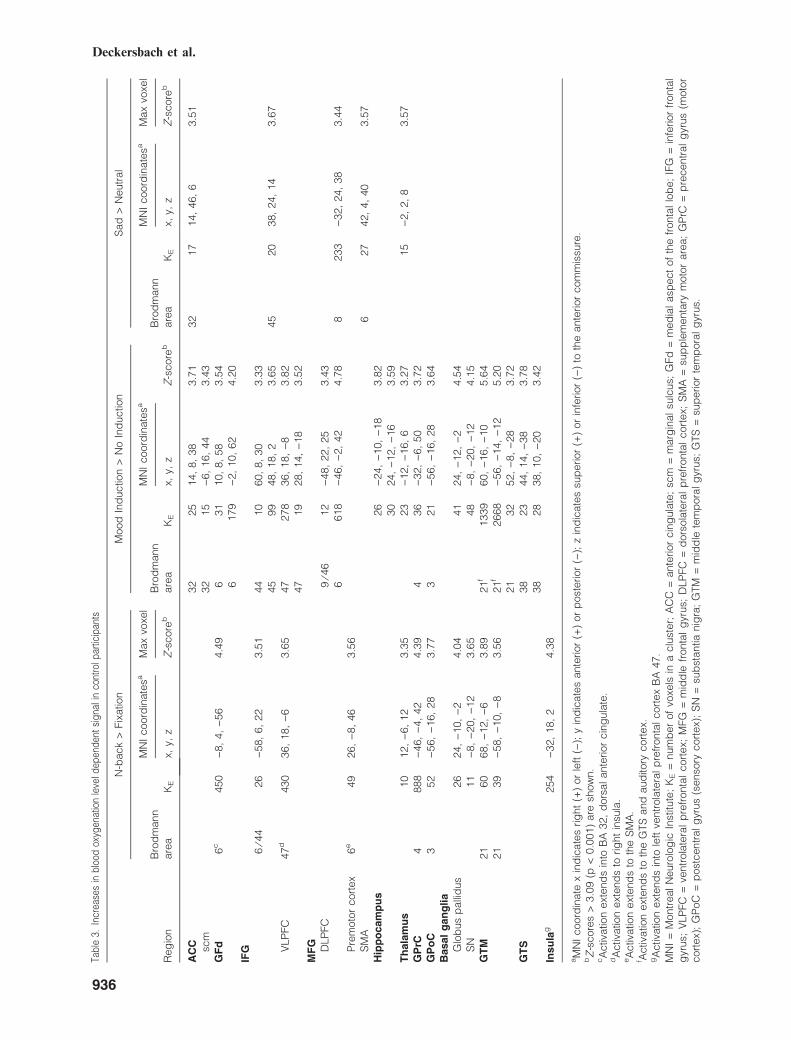
Exploratory analyses. Results of the within- andbetween-group contrasts for normal control andBP participants are shown in Tables 3–5.For the 2-back working memory versus fixation
cross contrast, compared to normal control par-ticipants, BP participants showed higher activa-tions in the right inferior frontal gyrus (BA 44),supplementary motor area (SMA), and in an areain the left precentral gyrus (BA 4 ⁄6; see Table 5).In addition, BP participants showed higher activa-tion in the right DLPFC in an area more anteriorand medial (x = 32, y = 34, z = 26; see Table 5)to the one for which there was a linear activationincrease across the three conditions (x = 54,y = 22, z = 22; see linear contrasts and Table 2).This right DLPFC area (x = 32, y = 34, z = 26)as well as the area in the SMA were also moreactivated in BP compared to control participantsfor the contrast 2-back working memory–moodinduction versus 2-back working memory–no mood induction (see Table 5). For the contrast2-back working memory–sad versus neutral stateinduction, there were no statistically significantdifferences between BP and control participants(see Table 5).
Correlations. For the a-priori hypothesized ROI,we investigated the association of depressionseverity (HAM-D scores) and linear increase inBOLD response across the conditions for the BP-I group. There were no significant correlationsbetween HAM-D scores and linear increase inany of the regions of interest (all p > 0.45). Theeffects of lithium and anticonvulsant medicationswere explored using multiple regression analysiswith lithium (yes = 1, no = 0) and anticonvul-sant (yes = 1, no = 0) as predictors of linearincrease in BOLD response across the conditionsfor the BP-I group. There were no significantcorrelations (all p > 0.10). There were also nosignificant correlations between HAM-D scores
in the BP group and BOLD signal in brainregions for which the exploratory analyses hadyielded significant group differences between BPand control participants (all p > 0.14). Themultiple regression analysis with lithium andanticonvulsants as predictors did not yield sig-nificant correlations for the exploratory analyses(all p > 0.09).
Discussion
We set out to investigate the functional neuroanat-omy of working memory during sadness in indi-viduals with bipolar disorder. We foundactivations in frontal and temporal cortical areasthat are typically observed in n-back workingmemory tasks that permit verbal coding (34, 57,58). These areas included the DLPFC (BA 9 ⁄46)and inferior prefrontal (BA 44, BA 45) cortex, aswell as the supplementary motor area. This patternof activation is commonly conceptualized asreflecting the activity of a central executive systemworking in conjunction with a phonological systemwhich jointly permit manipulating items �held�online in working memory.Our paradigm was specifically designed to
investigate working memory during sadness com-pared to neutral state or no mood induction. Wehypothesized that, for areas of linear activationincreases across the three conditions, duringsadness BP-I participants would be characterizedby abnormal activations in DLPFC and dACC,which support processes such as manipulatinginformation and performance monitoring �in�working memory. Consistent with our hypothe-sis, we found that patients with BP-I showedmore activation in the left DLPFC and rightdACC compared to control participants. For theleft and right DLPFC, these differences inactivation between BP and normal control par-ticipants tended to be present already during the2-back without any mood induction condition.However, the effect of increased left DLPFC anddACC activation in BP-I compared to controlparticipants was particularly pronounced duringsadness compared to the neutral or no moodstate induction conditions. This suggests that inindividuals with bipolar disorder, acute sadnessdisproportionately amplified functional abnor-malities in DLPFC and dACC that can alreadybe observed during a neutral state.Increased DLPFC and dACC activation in BP is
consistent with previous findings by Chang et al.(23), who also reported increased DLPFC anddACC activation in a sample of pediatric patientswith BP-I using a traditional 2-back working
fMRI investigation of working memory and sadness
935
Tab
le3.
Incr
ease
sin
blo
odox
ygen
atio
nle
veld
epen
den
tsi
gna
lin
cont
rolp
artic
ipan
ts
Reg
ion
N-b
ack
>Fix
atio
nM
ood
Ind
uctio
n>
No
Ind
uctio
nS
ad
>N
eutr
al
Bro
dm
ann
are
aK
E
MN
Icoord
inate
saM
ax
voxe
lB
rod
mann
are
aK
E
MN
Icoord
inate
sa
Z-s
core
bB
rod
mann
are
aK
E
MN
Icoord
inate
saM
ax
voxe
l
x,y,
zZ
-score
bx,
y,z
x,y,
zZ
-score
b
AC
C32
25
14,
8,
38
3.7
132
17
14,
46,
63.5
1sc
m32
15
)6,
16,
44
3.4
3G
Fd
6c
450
)8,
4,)
56
4.4
96
31
10,
8,
58
3.5
46
179
)2,
10,
62
4.2
0IF
G
6⁄4
426
)58,
6,
22
3.5
144
10
60,
8,
30
3.3
345
99
48,
18,
23.6
545
20
38,
24,
14
3.6
7V
LP
FC
47
d430
36,
18,)
63.6
547
278
36,
18,)
83.8
247
19
28,
14,)
18
3.5
2M
FG DLP
FC
9⁄4
612
)48,
22,
25
3.4
36
618
)46,)
2,
42
4.7
88
233
)32,
24,
38
3.4
4P
rem
oto
rcort
ex
6e
49
26,)
8,
46
3.5
6S
MA
627
42,
4,
40
3.5
7H
ipp
ocam
pu
s26
)24,)
10,)
18
3.8
230
24,)
12,)
16
3.5
9T
hala
mu
s10
12,)
6,
12
3.3
523
)12,)
16,
63.2
715
)2,
2,
83.5
7G
PrC
4888
)46,)
4,
42
4.3
94
36
)32,)
6,
50
3.7
2G
Po
C3
52
)56,)
16,
28
3.7
73
21
)56,)
16,
28
3.6
4B
asal
gan
glia
Glo
bus
palli
dus
26
24,)
10,)
24.0
441
24,)
12,)
24.5
4S
N11
)8,)
20,)
12
3.6
548
)8,)
20,)
12
4.1
5G
TM
21
60
68,)
12,)
63.8
921
f1339
60,)
16,)
10
5.6
421
39
)58,)
10,)
83.5
621
f2668
)56,)
14,)
12
5.2
021
32
52,)
8,)
28
3.7
2G
TS
38
23
44,
14,)
38
3.7
838
28
38,
10,)
20
3.4
2In
su
lag
254
)32,
18,
24.3
8
aM
NI
coord
inate
xin
dic
ate
srig
ht
(+)
or
left
());
yin
dic
ate
sante
rior
(+)
or
post
erior
());
zin
dic
ate
ssu
perior
(+)
or
infe
rior
())
toth
eante
rior
com
mis
sure
.bZ
-score
s>
3.0
9(p
<0.0
01)
are
show
n.
cA
ctiv
atio
next
end
sin
toB
A32,
dors
alante
rior
cin
gula
te.
dA
ctiv
atio
next
end
sto
rig
ht
insu
la.
eA
ctiv
atio
next
end
sto
the
SM
A.
f Activ
atio
next
end
sto
the
GTS
and
aud
itory
cort
ex.
gA
ctiv
atio
next
end
sin
tole
ftve
ntr
ola
tera
lp
refr
onta
lcort
ex
BA
47.
MN
I=
Montr
eal
Neuro
log
icIn
stitu
te;
KE
=num
ber
of
voxe
lsin
aclu
ster;
AC
C=
ante
rior
cin
gula
te;
scm
=m
arg
inal
sulc
us;
GFd
=m
ed
ial
asp
ect
of
the
fronta
llo
be;
IFG
=in
ferior
fronta
lg
yrus;
VLP
FC
=ve
ntr
ola
tera
lp
refr
onta
lcort
ex;
MFG
=m
idd
lefr
onta
lg
yrus;
DLP
FC
=d
ors
ola
tera
lp
refr
onta
lcort
ex;
SM
A=
sup
ple
menta
rym
oto
rare
a;
GP
rC=
pre
centr
al
gyr
us
(moto
rcort
ex)
;G
PoC
=p
ost
centr
alg
yrus
(senso
rycort
ex)
;S
N=
sub
stantia
nig
ra;
GTM
=m
idd
lete
mp
ora
lg
yrus;
GTS
=su
perior
tem
pora
lg
yrus.
Deckersbach et al.
936

Tab
le4.
Incr
ease
sin
blo
odox
ygen
atio
nle
veld
epen
den
tsi
gna
lin
par
ticip
ants
with
bip
olar
dis
ord
er
Reg
ion
N-b
ack
>Fix
atio
nM
ood
Ind
uctio
n>
No
Ind
uctio
nS
ad
>N
eutr
al
Bro
dm
ann
are
aK
E
MN
Icoord
inate
saM
ax
voxe
lB
rod
mann
are
aK
E
MN
Icoord
inate
sa
Z-s
core
bB
rod
mann
are
aK
E
MN
Icoord
inate
saM
ax
voxe
l
x,y,
zZ
-score
bx,
y,z
x,y,
zZ
-score
b
AC
C32
31
)12,
32,
22
4.1
8sc
m32
476
)10,
22,
40
4.6
332
108
)12,
20,
40
4.0
312,
18,
38
c4.4
632
37
6,
20,
42
3.6
1IF
G44
22
)58,
10,
63.8
844
14
)48,
10,
14
3.6
744
95
50,
6,
16
3.9
144
30
)56,
14,
18
3.9
245
253
)32,
26,
44.6
145
35
)30,
26,
63.7
345
⁄47
162
42,
20,
63.9
7V
LP
FC
36,
22,)
10
c3.8
8M
FG DLP
FC
9⁄4
623
36,
28,
24
4.1
69
⁄46
11
32,
36,
28
4.1
346
16
)52,
34,
16
3.2
132,
34,
28
c3.3
49
10
)50,
16,
28
3.1
7S
FG Pre
moto
rcort
ex
686
)40,)
4,
52
4.1
56
27
)40,)
4,
52
3.8
36
16
)26,)
2,
44
3.6
26
90
30,
2,
48
3.5
9A
myg
dala
37
)22,
0,)
14
3.9
442
)20,)
2,)
14
3.9
3G
PrC
616
)42,
4,
34
3.6
94
41
)56,)
16,
36
4.0
44
12
)56,)
16,
36
4.3
84
⁄612
)60,
0,
10
3.4
7G
Po
C43
15
)60,)
10,
20
4.5
2B
asal
gan
glia
Puta
men
22
20,
2,
14
4.7
020
20,
4,
14
4.1
4G
TM
21
⁄22
17
)66,)
14,)
24.1
721
⁄22
d79
60,)
12,)
24.0
421
14
60,)
16,)
23.6
762,)
22,
0c
3.5
4G
TS
22
18
)62,)
18,
43.5
222
d163
)62,)
20,
24.5
238
11
60,
12,)
12
4.0
938
27
60,
10,)
12
3.9
1In
su
la405
30,
14,
64.7
936
)38,
12,)
43.6
6
aM
NI
coord
inate
xin
dic
ate
srig
ht
(+)
or
left
());
yin
dic
ate
sante
rior
(+)
or
post
erior
());
zin
dic
ate
ssu
perior
(+)
or
infe
rior
())
toth
eante
rior
com
mis
sure
.bZ
-score
s>
3.0
9(p
<0.0
01)
are
show
n.
cS
econd
ary
activ
atio
np
eak
inth
esa
me
clu
ster.
dA
ctiv
atio
next
end
ing
into
aud
itory
cort
ex.
MN
I=
Montr
eal
Neuro
log
icIn
stitu
te;
KE=
num
ber
of
voxe
lsin
aclu
ster;
AC
C=
ante
rior
cin
gula
te;
scm
=m
arg
inal
sulc
us;
IFG
=in
ferior
fronta
lg
yrus;
VLP
FC
=ve
ntr
ola
tera
lp
refr
onta
lcort
ex;
MFG
=m
idd
lefr
onta
lgyr
us;
DLP
FC
=d
ors
ola
tera
lpre
fronta
lcort
ex;
SM
A=
sup
ple
menta
rym
oto
rare
a;G
PrC
=p
recentr
alg
yrus
(moto
rcort
ex)
;G
PoC
=p
ost
centr
alg
yrus
(senso
rycort
ex)
;G
TM
=m
idd
lete
mp
ora
lg
yrus;
GTS
=su
perior
tem
pora
lg
yrus.
fMRI investigation of working memory and sadness
937
memory paradigm. However, it is inconsistent withfindings from other fMRI working memory studies(21, 22, 24) that reported either decreased rightDLPFC or increased prefrontal activation that wasmore anterior (24) than that found in the presentstudy. These discrepancies across studies raise thepossibility that mood state is a critical source ofvariance in such experiments involving BP partic-ipants. Functional MRI working memory studiesby both Monks et al. (21) and Lagopoulos et al.(22) focused on euthymic individuals with BP withlow-level or residual depressive or manic symp-toms, while participants in the study by Changet al. (23) were characterized by more severeresidual depressive and manic symptoms. Thecurrent findings further suggest that the discrepan-cies across studies may be due to differences inmood state. However, the present study did notinclude a cohort of euthymic patients with bipolardisorder or retest depressed BP-I participantsduring a euthymic phase of their illness, whichwould permit a conclusion as to whether ourfindings reflect trait or state effects of BP. Inaddition, it should be emphasized that the sampleof individuals with bipolar disorder included in thisstudy was small. This limits the generalizability ofour results, such that caution is advised wheninterpreting the meaning and implication of ourfindings. Furthermore, whether sadness-specificfindings in this study are unique to sadness orcould also be observed with induction of otheremotions (happiness, anger) remains to be explored(for a discussion of disgust or fear-related findings,see 72). Likewise, it should be noted that in ourparadigm the 2-backonly conditionwasdivided intothree one-minute periods sandwiching the moodinduction conditions. It is possible that 2-back–no
mood induction blocks following the sadness (orneutral state) induction blocks partly reflect resid-ual activations from that condition, therebydecreasing the effect of activation differences acrossconditions. Auditory input (i.e., the presentation ofautobiographical scripts) occurred only during themood induction conditions, which may have pro-duced a potential confounding effect upon neuralactivity in the mood induction blocks compared tothe no-mood induction or fixation cross condition.Overall, participants in our study were well
matched with respect to age, education, and IQ,suggesting that these parameters do not accountfor our pattern of results. However, all BP-Iparticipants in the present study were takingmedications that can impair cognitive functioning(73–75). Mood-stabilizing medications such aslithium or anticonvulsants may reduce the effi-ciency of the processes supported by DLPFC anddACC to an extent that BP-I participants neededto mobilize additional resources in order tosuccessfully complete the 2-back working memorytask. This may be particularly pronounced in thecondition that involves sadness, thereby account-ing for the observed pattern of results. Althoughour regression analysis suggests that no singlemedication (lithium or anticonvulsant) accountsfor the pattern of results, it should be emphasizedthat statistical power for this analysis was limited.We should also note that our sample included
only female participants and that our findingscannot be unequivocally generalized to men. Inaddition, an earlier report of our group, whichincluded subjects from the cohorts presented here,initially found decreased activation in the anteriorcingulate during working memory during sadness(76) in an area in the anterior cingulate that was
Table 5. Increases in blood oxygenation level dependent signal in participants with bipolar disorder greater than control participants
Region
N-back > Fixation Mood Induction > No Induction
Brodmannarea KE
MNI coordinatesa
Max voxelZ-scoreb
Brodmannarea KE
MNI coordinatesa
Z-scorebx, y, z x, y, z
IFG 44 11 50, 8, 16 3.62MFG
DLPFC 9 14 32, 34, 26 3.46 9 11 32, 34, 26 3.32SMA 6 39 28, 6, 44 4.09 6 9 28, 6, 44 3.66
GPrC 4 ⁄ 6 16 )56, )12, 34 3.59
For the contrast 2-back working memory–sad versus neutral state induction, there were no statistically significant differences betweenbipolar disorder and control participants.aMNI coordinate x indicates right (+) or left ()); y indicates anterior (+) or posterior ()); z indicates superior (+) or inferior ()) to theanterior commissure.bZ-scores > 3.09 (p < 0.001) are shown.MNI = Montreal Neurologic Institute; KE= number of voxels in a cluster; IFG = inferior frontal gyrus; MFG = middle frontal gyrus;DLPFC = dorsolateral prefrontal cortex; SMA = supplementary motor area; GPrC = precentral gyrus (motor cortex).
Deckersbach et al.
938

anterior to the one reported here. BP-I and controlparticipants in this earlier report were not yet fullymatched with respect to the stage of their men-strual cycle and it is possible that this accounts forthe differences in findings. There is increasingawareness that for women, regional activationsmay vary with the stage of the menstrual cycle(e.g., follicular versus luteal phase, or early versuslate follicular phase) (77–83). Cycle-dependentvariations have been reported for the amygdala,hippocampus, ventral prefrontal cortex, anteriorcingulate, and DLPFC, among other regions (77–83), although findings about regions and directionsof activations (i.e., activation ⁄deactivation) varyacross paradigms and studies. Nonetheless, forfemale participants, this highlights the importanceof achieving a close match between female normalcontrol and BP participants to ensure that groupdifferences are not influenced by differences asso-ciated with the stage of the menstrual cycle.Within the limits of these methodological con-
siderations, the present findings raise the questionas to why abnormalities in the left DLPFC anddACC in bipolar disorder might be particularlypronounced during sadness. We hypothesized thatacute sadness (versus neutral states) would revealabnormalities in DLPFC and dACC by virtue oftaxing BP-I participants� ability to monitor andmanipulate information. Our task required partic-ipants to complete the 2-back working memoryparadigm under dual task conditions (i.e., workingmemory only or working memory during recall ofneutral and sad events). From the perspective of alimited capacity model, it is possible that for BP-Iparticipants, performing two tasks at once createsa disproportionate bottleneck occurring at a cen-tral, amodal stage of information processing thatimpairs response selection or decision-makingoperations from being concurrently executed (84).Previous investigations of dual-task paradigms inhealthy volunteers have highlighted brain regionsalso found in the present study, such as lateralprefrontal, dorsal premotor, and anterior cingu-late, as putative neural substrates of dual-taskinterference (84). However, while this interpreta-tion is consistent with an increase of activation inthese areas during both neutral and sadnessinduction, it does not explain why BP-I partici-pants showed disproportionately increased activa-tions specifically during sadness, given that theneutral state and sadness conditions were closelymatched other than for the emotional valence.Findings by Gray et al. (52) suggest that the leftDLPFC may play a vital role in the integration ofemotion and cognitive task demands (monitoring,manipulating, updating, etc.). They found selec-
tively increased left DLPFC activation during averbal n-back task (compared to a nonverbal)following exposure to unpleasant (versus pleasant)movies. Thus, it is possible that for depressed BP-Iparticipants, increased DLPFC and dACC activa-tion during sadness reflects difficulties integratingemotional and cognitive processes involved in ourworking memory paradigm.In summary, the present study replicated previ-
ous findings of abnormal DLPFC and dACCactivations in individuals with bipolar disorder.Our findings suggest that these abnormalities areamplified during periods of acute sadness whencompared to the neutral state, although our studydoes not permit any conclusions as to whether ornot this is associated with depression or medicationor represents a trait marker for bipolar disorder.Our findings support recent neurobiological mod-els of bipolar disorder (31) which view abnormal-ities in DLPFC and dACC (among other dorsaland ventral prefrontal regions) as being part ofthe neural substrate of impaired regulation andincreased lability of mood (31). In addition, thepresent study suggests that processes that integrateemotion (sadness) and cognition and that aresupported by DLPFC and dACC may be disturbedin bipolar disorder.
Acknowledgements
This work was supported by a Young Investigator Award fromthe National Alliance for Research in Schizophrenia andDepression to TD, and the Clinical Research TrainingProgram at Harvard Medical School (TD).
References
1. Martınez-Aran A, Vieta E, Colom F et al. Cognitivedysfunctions in bipolar disorder: evidence of neuropsycho-logical disturbances. Psychother Psychosom 2000; 69:2–18.
2. Bearden CE, Hoffman KM, Cannon TD. The neuropsy-chology and neuroanatomy of bipolar affective disorder: acritical review. Bipolar Disord 2001; 3: 106–150.
3. Murphy FC, Sahakian BJ. Neuropsychology of bipolardisorder. Br J Psychiatry, 2001; 178: 120–127.
4. Dupont RM, Jernigan TL, Butters N et al. Subcorticalabnormalities detected in bipolar affective disorder usingmagnetic resonance imaging. Arch Gen Psychiatry 1990;47: 55–59.
5. McKay AP, Tarbuck AF, Shapleske J, McKenna PJ.Neuropsychological function in manic-depressive psycho-sis. Evidence for persistent deficits in patients with chronic,severe illness. Br J Psychiatry 1995; 167: 51–57.
6. Tham A, Engelbrektson K, Mathe AA, Johnson L, OlssonM, Aberg-Wistedt A. Impaired neuropsychological per-formance in euthymic patients with recurring mooddisorders. J Clin Psychiatry 1997; 58: 26–29.
7. Atre-Vaidya N, Taylor MA, Seidenberg M, Reed R,Perrine A, Glick-Oberwise F. Cognitive deficits,
fMRI investigation of working memory and sadness
939

psychopathology, and psychosocial functioning in bipolarmood disorder. Neuropsychiatry Neuropsychol BehavNeurol 1998; 11: 120–126.
8. Ferrier IN, Stanton BR, Kelly TP, Scott J. Neuropsycho-logical function in euthymic patients with bipolar disorder.Br J Psychiatry 1999; 175: 246–251.
9. van Gorp WG, Altshuler L, Theberge DC, Wilkins J,Dixon W. Cognitive impairment in euthymic bipolarpatients with and without prior alcohol dependence. ArchGen Psychiatry 1998; 55: 41–46.
10. van Gorp WG, Altshuler L, Theberge DC, Mintz J.Declarative and procedural memory in bipolar disorder.Biol Psychiatry 1999; 46: 525–531.
11. Krabbendam L, Honig A, Wiersma J et al. Cognitivedysfunctions and white matter lesions in patients withbipolar disorder. Acta Psychiatr Scand 2000; 101: 274–280.
12. Clark L, Iversen SD, Goodwin GM. Sustained attentiondeficit in bipolar disorder. Br J Psychiatry 2002; 180: 313–319.
13. Cavanagh JTO, Van Beck M, Muir W, Blackwood DHR.Case-control study of neurocognitive function in euthymicpatients with bipolar disorder: an association with mania.Br J Psychiatry 2002; 180: 320–326.
14. Martınez-Aran A, Vieta E, Colom F et al. Cognitiveimpairment in euthymic bipolar patients. Bipolar Disord2002; 4 (Suppl. 1): 128.
15. Dickerson FB, Boronow JJ, Stallings CR, Origoni AE,Cole S, Yolken RH. Association between cognitive func-tioning and employment status of persons with bipolardisorder. Psychiatr Serv 2004; 55: 54–58.
16. Thompson JM, Gray JM, Hughes JH, Watson S, YoungAH, Ferrier IN. Impaired working memory monitoring ineuthymic bipolar patients. Bipolar Disord 2007; 9: 478–489.
17. Deckersbach T, McMurrich S, Ogutah J, Savage CR,Sachs G, Rauch SL. Characteristics of nonverbal memoryimpairment in bipolar disorder: the role of encodingstrategies. Psychol Med, 2004; 34: 823–832.
18. Deckersbach T, Savage CR, Reilly-Harrington N, Clark L,Sachs G, Rauch SL. Episodic memory impairment inbipolar disorder and obsessive-compulsive disorder: therole of memory strategies. Bipolar Disord 2004; 6: 233–244.
19. Deckersbach T, Savage CR, Dougherty DD et al. Spon-taneous and directed application of verbal learning strat-egies in bipolar disorder and obsessive-compulsivedisorder. Bipolar Disord 2005; 7: 166–175.
20. Deckersbach T, Dougherty DD, Savage CR et al. Imp-aired recruitment of the dorsolateral prefrontal cortex andhippocampus in bipolar disorder. Biol Psychiatry, 2005;59: 138–146.
21. Monks PJ, Thompson JM, Bullmore ET et al. A func-tional MRI study of working memory task in euthymicbipolar disorder: evidence for task-specific dysfunction.Bipolar Disord 2004; 6: 550–564.
22. Lagopoulos J, Ivanovski B, Malhi GS. An event-relatedfunctional MRI study of working memory in euthymicbipolar disorder. Rev Psychiatry Neurosci 2007; 32: 174–184.
23. Chang K, Adleman NE, Dienes K, Simenova DI, MenonV, Reiss A. Anomalous prefrontal-subcortical activation infamilial pediatric bipolar disorder. Arch Gen Psychiatry,2004; 61: 781–792.
24. Adler CM, Holland SK, Schmithorst V, Tuchfarber MJ,Strakowski SM. Changes in neuronal activation in patientswith bipolar disorder during performance of a workingmemory task. Bipolar Disord 2004; 6: 540–549.
25. Chang K, Adleman N, Dienes K, Barnea-Goraly N, ReissA, Ketter T. Decreased N-Acetylaspartate in children withfamilial bipolar disorder. Biol Psychiatry 2003; 53: 1059–1065.
26. Winsberg ME, Sachs N, Tate D, Adalsteinsson E, Spiel-man D, Ketter TA. Decreased dorsolateral prefrontalN-acetyl aspartate in bipolar disorder. Biol Psychiatry2000; 47: 475–481.
27. Rajkowska G, Halaris A, Selemon LD. Reductions inneuronal and glial density characterize prefrontal cortex inbipolar disorder. Biol Psychiatry 2001; 49: 741–752.
28. Cotter D, Mackay D, Chana G, Beasley C, Landau S,Everall IP. Reduced neuronal size in area 9 of thedorsolateral prefrontal cortex in subjects with majordepressive disorder. Cereb Cortex 2002; 12: 386–394.
29. Orlovskaya DD, Vostrikov VM, Rachmanova VI, Urano-va NA. Decreased numerical density of oligodendroglialcells in postmortem prefrontal cortex in schizophrenia,bipolar affective disorder and major depression. SchizophrRes 2000; 41: 105–106.
30. Todtenkopf MS, Vincent SL, Francine V, Benes FM. Across-study meta-analysis and three-dimensional compar-ison of cell counting in the anterior cingulate cortex ofschizophrenic and bipolar brain. Schizophr Res 2005; 73:79–89.
31. Phillips ML. The neural basis of mood dysregulation inbipolar disorder. Cognit Neuropsychiatry 2006; 11: 233–249.
32. Baddeley A. Human Memory. Theory and Practice. Hove:Earlbaum, 1990.
33. Baddeley A. Working memory: looking back and lookingforward. Nat Rev Neurosci 2003; 4: 829–839.
34. Smith EE, Jonides J. Storage and executive processes in thefrontal lobes. Science 1999; 283: 1657–1661.
35. Owen AM, Evans AC, Petrides M. Evidence for a two-stage model of spatial working memory processing withinthe lateral frontal cortex: a positron emission tomographystudy. Cereb Cortex 1996; 8: 353–364.
36. Mesulam M-M. Large-scale neurocognitive networks anddistributed processing for attention, language, and mem-ory. Ann Neurol 1990; 28: 597–613.
37. Posner MI, Petersen SE, Fox PT et al. Localization ofcognitive operations in the human brain. Science 1988; 240:1627–1631.
38. Frith CD, Friston K, Liddle PF, Frackowiak RS. Willedaction and the prefrontal cortex in man: a study with PET.Proc Biol Sci 1991; 244: 241–246.
39. Paus T, Petrides M, Evans AC et al. Role of the humananterior cingulate cortex in the control of oculomotor,manual, and speech responses: a positron emission tomog-raphy study. J Neurophysiol 1993; 70: 453–469.
40. Badgaiyan RD, Posner MI. Mapping the cingulate cortexin response selection and monitoring. Neuroimage 1998; 7:255–260.
41. Turken AU, Swick D. Response selection in the humananterior cingulate cortex. Nat Neurosci 1999; 2: 920–924.
42. Gehring WJ, Knight RT. Prefrontal-cingulate interactionsin action monitoring. Nat Neurosci 2000; 3: 516–520.
43. Luu P, Flaisch T, Tucker DM. Medial frontal cortex inaction monitoring. J Neurosci 2000; 20: 464–469.
44. Carter CS, Braver TS, Barch DM et al. Anterior cingulatecortex, error detection, and the online monitoring ofperformance. Science 1998; 280: 747–749.
45. Botvinic M, Nystrom LE, Fissell K et al. Conflict moni-toring versus selection- for-action in anterior cingulatecortex. Nature 1999; 402: 179–181.
Deckersbach et al.
940

46. Carter CS, Macdonald AM, Botvinick M et al. Parsingexecutive processes: strategic vs. evaluative functions of theanterior cingulate cortex. Proc Natl Acad Sci USA 2000;97: 1944–1948.
47. Mayberg HM. Limbic-cortical dysregulation: a proposedmodel of depression. J Neuropsych Clin N, 1997; 9: 471–481.
48. Mayberg HS, Liotti M, Brannan SK et al. Reciprocallimbic-cortical function and negative mood: convergingPET findings in depression and normal sadness. Am JPsychiatry 1999; 156: 675–682.
49. Seminowicz DA, Mayberg HS, McIntosh AR et al. Lim-bic-frontal circuitry in major depression: a path modelingmetanalysis. Neuroimage 2004; 22: 409–418.
50. Levesque J, Eugene F, Joanette Y et al. Neural circuitryunderlying voluntary suppression of sadness. Biol Psychi-atry 2003; 53: 502–510.
51. Ochsner KN, Ray RD, Cooper JC et al. For better or forworse: neural systems supporting the cognitive down- andup-regulation of negative emotion. Neuroimage 2004; 23:483–499.
52. Gray JR, Braver TS, Raichle ME. Integration of emotionand cognition in the lateral prefrontal cortex. 1. Proc NatlAcad Sci USA 2002; 99: 4115–4120.
53. Liotti M, Mayberg HS, Brannan SK, McGinnis S,Jerabek P, Fox PT. Differential limbic – cortical corre-lates of sadness and anxiety in healthy subjects: impli-cations for affective disorders. Biol Psychiatry 2000; 48:30–42.
54. Damasio AR, Grabowski TJ, Bechara A et al. Subcorticaland cortical brain activity during the feeling of self-generated emotions. Nat Neurosci 2000; 3: 1049–1056.
55. Malhi GS, Lagopoulos J, Ward PB et al. Cognitivegeneration of affect in bipolar depression: an fMRI study.Eur J Neurosci 2004; 19: 741–754.
56. Keedwell PA, Andrew C, Williams SC, Brammer MJ,Phillips ML. A double dissociation of ventromedialprefrontal cortical responses to sad and happy stimuli indepressed and healthy individuals. Biol Psychiatry 2005;58: 495–503.
57. Owen AM, Stern CE, Look RB, Tracey I, Rosen BR,Petrides M. Functional organization of spatial and non-spatial working memory processing within the humanlateral frontal cortex. Proc Natl Acad Sci USA 1998; 95:7721–7726.
58. Smith EE, Jonides J. Working memory: a view fromneuroimaging. Cognit Psychol 1997; 33: 5–42.
59. Braver TS, Cohen JD, Nystrom LE, Jonides J, Smith EE,Noll DC. A parametric study of prefrontal cortex involve-ment in human working memory. Neuroimage 1997; 5: 49–62.
60. First MB, Spitzer RL, Gibbon M, Williams JBW. Struc-tured Clinical Interview for DSM-IV Axis I Disorders-Patient Edition (SCID-I ⁄D, Version 2.0). New York:Biometrics Research Department, New York PsychiatricInstitute, 1995.
61. Hamilton M. A rating scale for depression. J NeurolNeurosurg Psychiatry 1960; 23: 56–62.
62. Young RC, Biggs JT, Ziegler VE, Meyer DA. A ratingscale for mania: reliability, validity and sensitivity. Br JPsychiatry 1978; 133: 429–435.
63. Oldfield RC. The assessment and analysis of handedness:The Edinburgh Inventory. Neuropsychologia 1971; 9: 97–113.
64. The Psychological Corporation. The Wechsler Test ofAdult Reading. San Antonio: The Psychological Corpora-tion, 2001.
65. Dougherty DD, Rauch SL, Deckersbach T et al. Ventro-medial prefrontal cortex and amygdala dysfunction duringan anger induction positron emission tomography study inpatients with major depressive disorder with anger attacks.Arch Gen Psychiatry 2004; 61: 795–804.
66. Friston KJ, Holmes AP, Worsley KJ, Poline JB, Frith CD,Frackowiak RS. Statistical parametric maps in functionalimaging: A general linear approach. Hum Brain Mapp1995; 2: 189–210.
67. Friston KJ (ed.). Statistical Parametric Mapping: TheAnalysis of Functional Brain Images. San Diego: Aca-demic Press, 2006.
68. Brett M, Anton J-L, Valabregue R, Poline J-B. Region ofInterest Analysis Using an SPM Toolbox [Abstract]. Pre-sented at the 8th International Conference of FunctionalMapping of the Human Brain, 2002 (June 2–6) Sendai,Japan.Available onCD-ROMinNeuroimage,Vol 16,No2,2002.
69. Tzourio-Mazoyer N, Landeau B, Papathanassiou D et al.Automated anatomical labeling of activations in SPMusing a macroscopic anatomical parcellation of the MNIMRI single-subject brain. Neuroimage 2002; 15: 273–289.
70. Maldjian JA, Laurienti PJ, Burdette JB, Kraft RA. Anautomated method for neuroanatomic and cytoarchitec-tonic atlas-based interrogation of fMRI data sets. Neuro-Image 2003; 19: 1233–1239.
71. Maldjian JA, Laurienti PJ, Burdette JH. Precentral gyrusdiscrepancy in electronic versions of the Talairach Atlas.Neuroimage 2004; 21: 450–455.
72. Malhi GS, Lagopoulos J, Sachdev PS, Ivanovski B, ShnierR, Ketter T. Is a lack of disgust something to fear? Afunctional magnetic resonance imaging facial emotionrecognition study in euthymic bipolar disorder patients.Bipolar Disord 2007; 9: 345–357.
73. Judd LL, Hubbard B, Janowsky DS, Huey LY, TakahashiKI. The effect of lithium carbonate on the cognitivefunctioning of normal subjects. Arch Gen Psychiatry 1977;34: 355–357.
74. Ananth J, Fhadirian AM, Engelsmann F. Lithium andmemory: a review. Can J Psychiatry 1987; 32: 312–316.
75. Goldberg JF, Burdick KE. Cognitive side effects ofanticonvulsants. J Clin Psychiatry 2001; 62 (Suppl. 14):27–33.
76. Deckersbach T, Nierenberg AA, Buhlmann U et al. Theinfluence of sadness on working memory in bipolardisorder: An fMRI investigation. Bipolar Disord 2007; 9(Suppl. 1): 31–32.
77. Amin Z, Epperson CN, Constable RT, Canli T. Effects ofestrogen variation on neural correlates of emotionalresponse inhibition. Neuroimage 2006; 32: 457–464.
78. Choi JC, Park SK, Kim YH et al. Different brainactivation patterns to pain and pain-related unpleasantnessduring the menstrual cycle. Anesthesiology 2006; 105: 120–127.
79. Dreher JC, Schmidt PJ, Kohn P, Furman D, Rubinow D,Berman KF. Menstrual cycle phase modulates reward-related neural function in women. Proc Natl Acad Sci USA2007; 104: 2465–2470.
80. Gizewski ER, Krause E, Karama S, Baars A, Senf W,Forsting M. There are differences in cerebral activationbetween females in distinct menstrual phases duringviewing of erotic stimuli: a fMRI study. Exp Brain Res2006; 174: 101–108.
81. Goldstein JM, Jerram M, Poldrack R et al. Hormonalcycle modulates arousal circuitry in women using func-tional magnetic resonance imaging. J Neurosci 2005; 25:9309–9316.
fMRI investigation of working memory and sadness
941

82. Protopopescu X, Pan H, Altemus M et al. Orbitofrontalcortex activity related to emotional processing changesacross the menstrual cycle. Proc Natl Acad Sci USA 2005;102: 16060–16065.
83. Schoning S, Engelien A, Kugel H et al. Functionalanatomy of visuo-spatial working memory during mental
rotation is influenced by sex, menstrual cycle, andsex steroid hormones. Neuropsychologia 2007; 45:3203–3214.
84. Dux PE, Ivanoff J, Asplund CL, Marois R. Isolation of acentral bottleneck of information processing with time-resolved fMRI. Neuron 2006; 52: 1109–1120.
Deckersbach et al.
942

![[CONFERENCE PAPER] Bipolar Bozuklukta BDT](https://static.fdokumen.com/doc/165x107/63328d1f4e0143040300b9b3/conference-paper-bipolar-bozuklukta-bdt.jpg)


















