
Occult Knowledge,Science and Gender on the Shakespearean stage
Upload
independentCategory
view
2download
0
At
Ea
D
Bttmswpctrwagjgninpwitpr(dlCtbntrd
Tcnlptmt
BA
SIC–
ALIM
ENTA
RY
TRA
CT
GASTROENTEROLOGY 2007;132:1912–1924
n Enteric Occult Reflex Underlies Accommodation and Slow Transit inhe Distal Large Bowel
AMONN J. DICKSON, NICK J. SPENCER, GRANT W. HENNIG, PETER O. BAYGUINOV, JIM REN, DANTE J. HEREDIA,nd TERENCE K. SMITH
epartment of Physiology & Cell Biology, University of Nevada School of Medicine, Reno, Nevada
tabeaasitgprto
siugstldq(ddttmrcwnf
leINd
ackground & Aims: Transit of fecal materialhrough the human colon takes >30 hours, whereasransit through the small intestine takes 24 hours. The
echanisms underlying colonic storage and slow tran-it have yet to be elucidated. Our aim was to determinehether an intrinsic neural mechanism underlies thesehenomena. Methods: Recordings were made from cir-ular muscle (CM) cells and myenteric neurons inhe isolated guinea pig distal colon using intracellularecordings and Ca2� imaging techniques. Video imagingas used to determine the effects of colonic fillingnd pellet transit. Results: Circumferential stretchenerated ongoing oral excitatory and anal inhibitoryunction potentials in the CM. The application of lon-itudinal stretch inhibited all junction potentials. N-�-itro-L-arginine (100 �mol/L) completely reversed the
nhibitory effects of longitudinal stretch suggesting thatitric oxide (NO) inhibited interneurons controllingeristaltic circuits. Ca2� imaging in preparations thatere stretched in both axes revealed ongoing firing
n nNOS �ve descending neurons, even when synap-ic transmission was blocked. Inhibitory postsynapticotentials were evoked in mechanosensitive interneu-ons that were blocked by N-�-nitro-L-arginine100 �mol/L). Pellet transit was inhibited by longitu-inal stretch. Filling the colon with fluid led to co-
onic elongation and an inhibition of motility.onclusions: Our data support the novel hypothesis
hat slow transit and accommodation are generatedy release of NO from descending (nNOS �ve) inter-eurons triggered by colonic elongation. We refer to
his powerful inhibitory reflex as the intrinsic occulteflex (hidden from observation) because it with-raws motor activity from the muscle.
he large bowel has evolved to serve several majorfunctions: to recover water and electrolytes from the
ontents of the intestinal lumen, to use bacteria to digestutrients (in instances in which appropriate enzymes are
acking), and to prevent being tracked by scent-orientatedredators and mark territories.1 To perform these func-ions, transit of intraluminal contents through the hu-
an colon is slow, taking �30 hours, whereas transit
hrough the small intestine takes 2– 4 hours.1– 4 As in-raluminal contents move down the large bowel, waternd electrolytes are absorbed, causing the contents toecome more viscous leading to stool formation. How-ver, the mechanism underlying colonic accommodationnd slow transit to promote absorption is unknown,1–5
lthough increased segmental contractions and extrinsicympathetic reflexes have been proposed.1– 4 Recently, anntrinsic neural mechanism involving an excess produc-ion of nitric oxide (NO) within the myenteric plexus, theanglionated network that controls motility,5,6 has beenroposed from immunohistochemical studies to have aole in the pathology underlying slow transit constipa-ion in the human large bowel.7,8 However, how an excessf NO might alter transit is unknown.
We have recently proposed that there are multiple sen-ory neurons in the myenteric plexus of the guinea pig largentestine.9 There appears to be 2 intrinsic sensory systemsnderlying peristaltic type motor patterns that are analo-ous to tension-sensitive Golgi tendon organs and length-ensitive muscle spindles in skeletal muscle. Activation ofhese systems by circumferential stretch applied to the co-on generates neurally mediated peristaltic waves that areependent on muscle tone (duration �40–60 seconds; fre-uency, �0.3/min)10 and ongoing peristaltic reflex activityduration, �3 seconds; frequency, �11/min), which is in-ependent of muscle tone but generated by increases in theiameter of the bowel.11–13 These 2 motor patterns appearo be generated by myenteric after-hyperpolarization (AH)-ype sensory neurons, which sense muscle tension,14 and
echanosensory S-type ascending and descending interneu-ons that detect changes in muscle length around the cir-umference of the bowel.11,13,15 Multipolar AH neurons,hich were until recently thought to be the only sensoryeuron in the gut, are defined for their long-lasting AH
ollowing an action potential. Unipolar synaptic (S) neu-
Abbreviations used in this paper: CM, circular muscle; CMMP, circu-ar muscle myenteric plexus; EJP, excitatory junction potential; FEPSP,xcitatory postsynaptic potential; IJP, inhibitory junction potential;PSP, inhibitory postsynaptic potential; LM, longitudinal muscle; L-NA,-�-nitro-L-arginine; PPADS, pyridoxal phosphate-6-azophenyl-2,=4=-isulfonic acid; TS, transmural stimulation.
© 2007 by the AGA Institute0016-5085/07/$32.00
doi:10.1053/j.gastro.2007.02.047

rie
cdbiisgrnewass
kacs((b1oncm
ptdtta1itlfnswrnat5rm
pLeadoeitejwt
rtsppsTers1wep
Fcs((sppt
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 ENTERIC OCCULT REFLEX AND LARGE BOWEL TRANSIT 1913
ons, on the other hand, receive fast excitatory synapticnput and comprise the mechanosensory interneurons andxcitatory and inhibitory motor neurons.11,15
Recently, low-threshold extrinsic afferent mechanore-eptors in the rectum have been shown not to exhibitirectional sensitivity because they respond equally tooth circular and longitudinal stretch.16 Given this find-
ng, we wanted to determine whether peristaltic activityn the colon is affected not only by circumferentialtretch but also by longitudinal stretch. We also investi-ated how intrinsic release of NO might affect theseeflexes. We demonstrate that a third myenteric sensoryeural system, which is inhibitory, is activated by coloniclongation. These mechanosensory neurons release NOithin the myenteric plexus resulting in a depression ofctivity of mechanosensitive interneurons driving peri-taltic nerve pathways activated by circumferentialtretch.
Materials and MethodsIntracellular Microelectrode Recordings FromSmooth MuscleMale and female guinea pigs (200 –350 g) were
illed in accordance with the Animal Ethics Committeet the University of Nevada-Reno, and a segment of distalolon (15–20 mm long) was removed. The mucosa andubmucosa were removed to reveal the circular muscleCM), and the segment was threaded through greasedHigh Vacuum Grease; Dow Corning, Midland, MI) rub-er diaphragms in 2 partitions (7 mm apart; see FigureA). Each chamber was separately perfused with warmedxygenated Krebs’ solution (36.0°C � 0.5°C) containingifedipine (1–2 �mol/L) to paralyze the smooth mus-le.9 –12 Simultaneous microelectrode recordings wereade from CM cells at both the oral and anal ends of the
igure 1. Effects of circumferential and longitudinal stretch. (A) Cir-umferential stretch applied to the distal colon activated ongoing peri-taltic reflex activity consisting of oral excitatory junction potentialsEJPs) that were coordinated with anal inhibitory junction potentialsIJPs) in the circular muscle. Transmural nerve stimulation (transmuraltimulating single pulse 0.5 ms, 30 V) applied to the middle of thereparation evoked a robust oral EJP and anal IJP. (B) When the samereparation was stretched only in the longitudinal axis, only low-ampli-
(ude neural activity was observed and gave only a small response to TS.
reparation. Preparations were stretched and pinned ei-her in the circumferential (12 mm, twice their restingiameter)11–13 or longitudinal directions or in both direc-ions simultaneously. Once control activity was recorded,he preparation was then stretched and repinned in bothxes, which took approximately 2–3 minutes. After a0-minute equilibration period, microelectrodes were re-
nserted into CM cells at either end of the tissue. Longi-udinal stretch was normalized to the slack segmentength. Simultaneous intracellular recordings were maderom CM cells using 2 independently mounted microma-ipulators (WPI; World Precision Instruments, Inc. Sara-ota, FL; model M3301R). Microelectrodes (ID, 0.5 mm)ere filled with 1.5 mol/L KCl solution and had tip
esistances of approximately 100 –150 M�. Electrical sig-als were amplified using a dual input Axoprobe 1Amplifier (Axon Instruments, Foster City, CA) and digi-ized at 2–5 kHz for neuronal recordings (see below) and00 Hz to 1 kHz for smooth muscle recordings on a PCunning Axoscope software (version 8.0; Axon Instru-
ents).
CM Myenteric Plexus PreparationsCM myenteric plexus (CMMP) preparations were
repared by removing the longitudinal muscle (LM). TheM was removed by teasing up muscle fibers at the oralnd, pinching these fibers with a pair of curved forcepsnd peeling the LM off the preparation, in an oral to analirection. This easily removed the LM, which often cameff as wide strips of muscle, without damaging the my-nteric plexus that remained intact upon the CM. Thentegrity of the enteric circuitry was assessed by the facthat the CM generated spontaneous coordinated oralxcitatory junction potentials (EJP) and anal inhibitoryunction potentials (IJP). This would not happen if thereas any damage to the myenteric plexus from removing
he LM.
Intracellular Microelectrode Recordings FromMyenteric NeuronsThe mucosa and submucosa and the LM were
emoved from a segment of distal colon 20 mm in lengtho create a CMMP preparation. The segment, which wastretched circumferentially to twice its slack width, wasinned in an organ bath with the myenteric plexus up-ermost. Retaining the CM ensures that neurons in peri-taltic nerve pathways are spontaneously active.13,15
ransmural stimulating wires were placed across the oralnd of the preparation approximately 10 mm from theecording site. Once a neuron was impaled, transmuraltimuli (TS) were applied (0.5-ms duration, 10 Hz for 5 to0 pulses). Standard intracellular recording techniquesere used to record from myenteric neurons at the anal
nd of the segment.13,15 The organ bath was separatelyerfused with warmed oxygenated Krebs’ solution
36.0°C � 0.5°C) containing nifedipine (1–2 �mol/L) to
p�
cpvNl4woFpattnw(svwG
ma
wbtsCahsh(m
FiaOolb
Fcafsftat�cc2s
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1914 DICKSON ET AL GASTROENTEROLOGY Vol. 132, No. 5
aralyze the smooth muscle9 –12 and guanethidine (0.5mol/L) to block norepinephine release.17
Calcium ImagingThe LM was peeled from a segment of colon to
reate the CMMP preparation to expose the myentericlexus.18 The preparation was stretched in both axes andiewed under an upright microscope (Eclipse E600FN;ikon Inc., Melville, NY) fitted with water immersion
enses (�10X, �20, �40: Nikon Plan Fluor).6,18 Fluo-AM (FluoroPure, AM, Molecular Probes, Eugene, OR)as loaded into neurons at room temperature as previ-usly described.18 The final loading concentration ofluo-4 in the bath was 50 nmol/L. The tissues were thenerfused with warm (36.0°C � 0.5°C) Krebs’ solutionnd illuminated using a Lambda DG-5 light source (Sut-er Instrument Company, Novato, CA). Appropriate fil-ers produced excitation of Fluo-4 between 460 and 490m and collected emissions �515 nm. Image sequencesere captured using a Cascade 512B digital camera
Roper Scientific Inc., Trenton, NJ) and MetaMorph 6.26oftware (Universal Imaging Corporation; Molecular De-ices Corporation, Downington, PA). Image sequencesere analyzed using custom-written software (Volumetry5; G.W.H.).18
ImmunohistochemistryFollowing calcium imaging experiments, whole
ount tissues were then (1) fixed for 15 minutes in
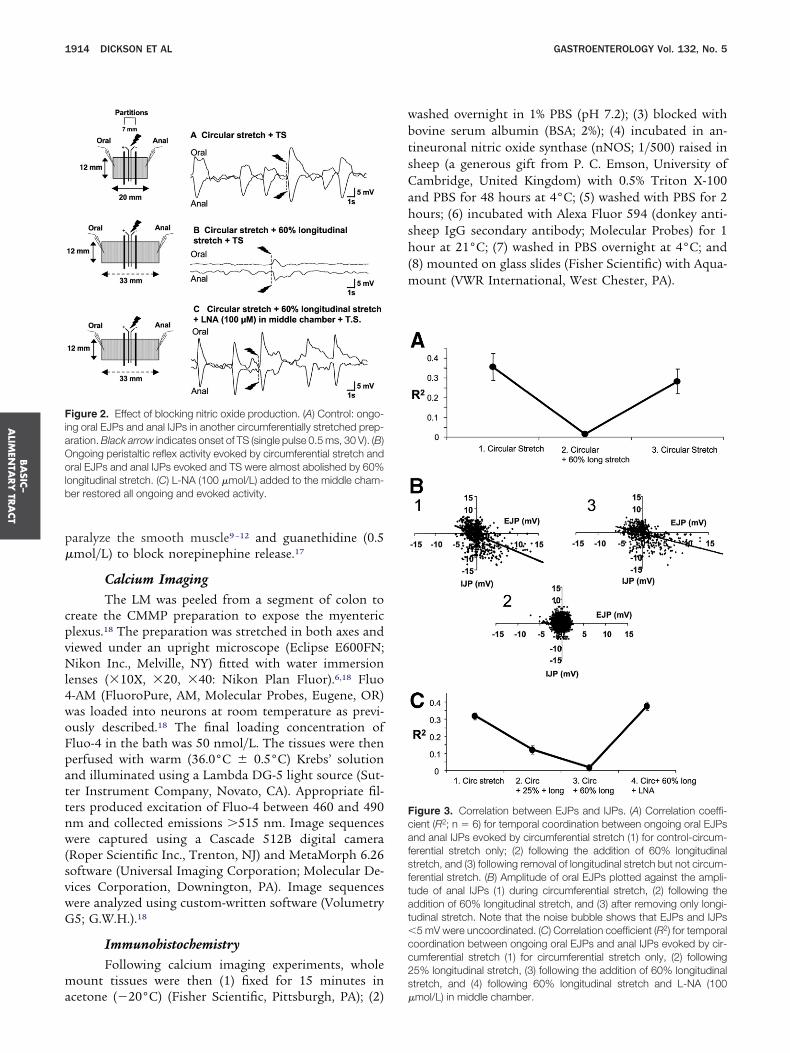
igure 2. Effect of blocking nitric oxide production. (A) Control: ongo-ng oral EJPs and anal IJPs in another circumferentially stretched prep-ration. Black arrow indicates onset of TS (single pulse 0.5 ms, 30 V). (B)ngoing peristaltic reflex activity evoked by circumferential stretch andral EJPs and anal IJPs evoked and TS were almost abolished by 60%
ongitudinal stretch. (C) L-NA (100 �mol/L) added to the middle cham-er restored all ongoing and evoked activity.
cetone (�20°C) (Fisher Scientific, Pittsburgh, PA); (2) �
ashed overnight in 1% PBS (pH 7.2); (3) blocked withovine serum albumin (BSA; 2%); (4) incubated in an-ineuronal nitric oxide synthase (nNOS; 1/500) raised inheep (a generous gift from P. C. Emson, University ofambridge, United Kingdom) with 0.5% Triton X-100nd PBS for 48 hours at 4°C; (5) washed with PBS for 2ours; (6) incubated with Alexa Fluor 594 (donkey anti-heep IgG secondary antibody; Molecular Probes) for 1our at 21°C; (7) washed in PBS overnight at 4°C; and
8) mounted on glass slides (Fisher Scientific) with Aqua-ount (VWR International, West Chester, PA).
igure 3. Correlation between EJPs and IJPs. (A) Correlation coeffi-ient (R2; n � 6) for temporal coordination between ongoing oral EJPsnd anal IJPs evoked by circumferential stretch (1) for control-circum-erential stretch only; (2) following the addition of 60% longitudinaltretch, and (3) following removal of longitudinal stretch but not circum-erential stretch. (B) Amplitude of oral EJPs plotted against the ampli-ude of anal IJPs (1) during circumferential stretch, (2) following theddition of 60% longitudinal stretch, and (3) after removing only longi-udinal stretch. Note that the noise bubble shows that EJPs and IJPs5 mV were uncoordinated. (C) Correlation coefficient (R2) for temporal
oordination between ongoing oral EJPs and anal IJPs evoked by cir-umferential stretch (1) for circumferential stretch only, (2) following5% longitudinal stretch, (3) following the addition of 60% longitudinaltretch, and (4) following 60% longitudinal stretch and L-NA (100
mol/L) in middle chamber.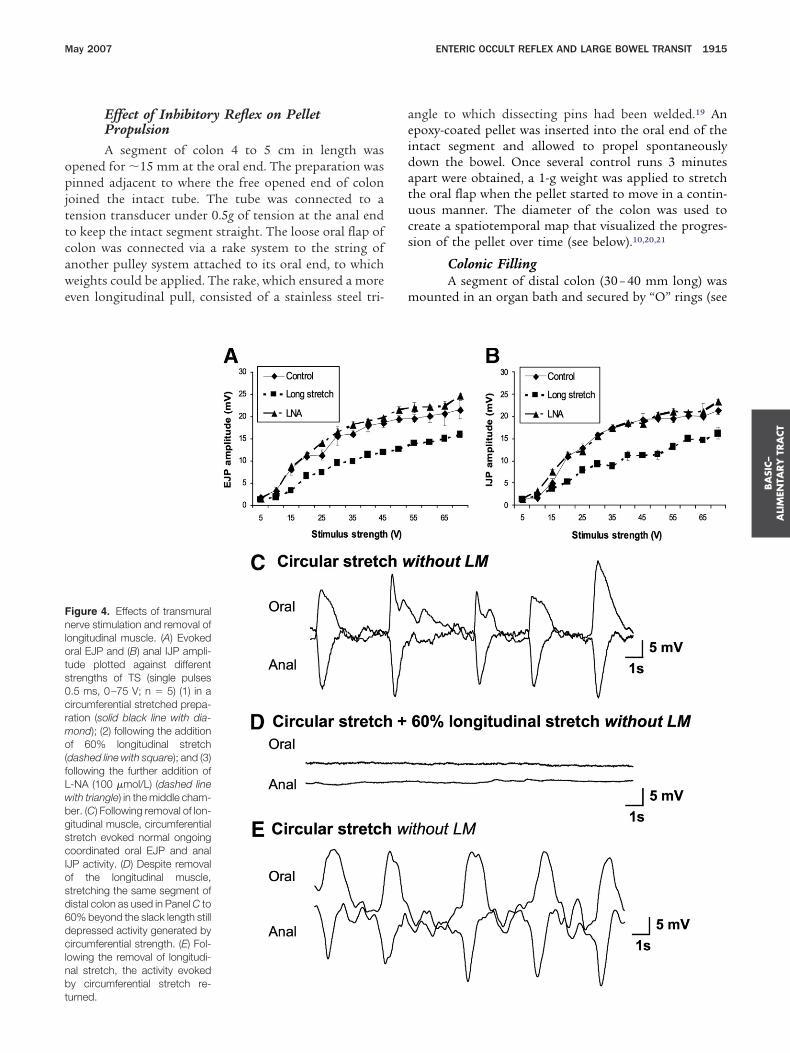
opjttcawe
aeidatucs
m
Fnlots0crmo(fLwbgscIosd6dclnbt
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 ENTERIC OCCULT REFLEX AND LARGE BOWEL TRANSIT 1915
Effect of Inhibitory Reflex on PelletPropulsionA segment of colon 4 to 5 cm in length was
pened for �15 mm at the oral end. The preparation wasinned adjacent to where the free opened end of colon
oined the intact tube. The tube was connected to aension transducer under 0.5g of tension at the anal endo keep the intact segment straight. The loose oral flap ofolon was connected via a rake system to the string ofnother pulley system attached to its oral end, to whicheights could be applied. The rake, which ensured a more
ven longitudinal pull, consisted of a stainless steel tri-
igure 4. Effects of transmuralerve stimulation and removal of
ongitudinal muscle. (A) Evokedral EJP and (B) anal IJP ampli-ude plotted against differenttrengths of TS (single pulses.5 ms, 0–75 V; n � 5) (1) in aircumferential stretched prepa-ation (solid black line with dia-ond); (2) following the additionf 60% longitudinal stretch
dashed line with square); and (3)ollowing the further addition of-NA (100 �mol/L) (dashed lineith triangle) in the middle cham-er. (C) Following removal of lon-itudinal muscle, circumferentialtretch evoked normal ongoingoordinated oral EJP and anal
JP activity. (D) Despite removalf the longitudinal muscle,tretching the same segment ofistal colon as used in Panel C to0% beyond the slack length stillepressed activity generated byircumferential strength. (E) Fol-
owing the removal of longitudi-al stretch, the activity evokedy circumferential stretch re-
urned.ngle to which dissecting pins had been welded.19 Anpoxy-coated pellet was inserted into the oral end of thentact segment and allowed to propel spontaneouslyown the bowel. Once several control runs 3 minutespart were obtained, a 1-g weight was applied to stretchhe oral flap when the pellet started to move in a contin-ous manner. The diameter of the colon was used toreate a spatiotemporal map that visualized the progres-ion of the pellet over time (see below).10,20,21
Colonic FillingA segment of distal colon (30 – 40 mm long) was
ounted in an organ bath and secured by “O” rings (see

FcsldweovCSoC
wacd
stnnts
Frss(aa
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1916 DICKSON ET AL GASTROENTEROLOGY Vol. 132, No. 5
igure 7A).10,20 The oral end was fixed and the anal endonnected to a moveable outflow tube, under 2g of ten-ion in an organ bath containing oxygenated Krebs’ so-ution at 36.0°C � 0.5°C (see Figure 7A). A force trans-ucer (TST125C; Biopac Systems Inc., Santa Barbara, CA)as glued (Vet bond) to the serosal surface at the oral
nd. Marker (eye glitter) arrays were glued along the wallf the colon to measure longitudinal movements andiewed with a video camera (WV-BP330; PanasonicCTV) and recorded on a VHS tape recorder (VWM-390;anyo). Warmed Krebs’ solution was injected into theral end with a syringe (Hamilton, Reno, NV; 0 – 800 �L).hanges in diameter and length of the intact segment
igure 5. Ca2 imaging of myenteric neurons following blockade of syemoved, was pinned in both axes. Hexamethonium (100 �mol/L), onolution to block fast synaptic transmission. (A) Average Ca2 fluoresuperimposed on averaged Ca2 fluorescence. (C) nNOS immunoreaD) Before the addition of L-NA, only 1 neuron in this ganglia exhibitednally projecting (see arrowhead 1 in A and B). (E) L-NA (100 �mol/L)
ctivity in other neurons (boxes 2–5; C and E) that were NOS �ve.ere followed over time by converting video images intosilhouette and then plotting the diameter along the
olon at each time point (33 ms) as a spatiotemporaliameter or D map.10,20,21
Drugs and SolutionsGuanethidine was used to block postganglionic
ympathetic nerve transmission. Nifedipine was usedo paralyze the muscle. Stock solutions of hexametho-ium bromide, ondansetron, and phosphate-6-ozophe-yl-2=-4=-disulfonic acid (PPADS) were made in dis-illed water and used to block all known forms of fastynoptic transmission. A combination of these drugs
c neurotransmission. A segment of colon, with the longitudinal muscleetron (1 �mol/L), and PPADs (30 �mol/L) were added to the Krebs’in myenteric neurons. (B) An overlay of nNOS immunoreactivy (red)
(red) overlaid with average Ca2 fluorescence in L-NA (100 �mol/L).ng activity (see arrowhead 1 in A–C). This neuron was nNOS ve andt effect the activity of the NOS ve neuron (black trace) but revealed
naptidans
cencectivyongoidid no

wo�(N17
wsdabrtFoldtmnrrrsOtmRsiviTistcattaOtpSa
t
pdscshbamtoWis
scpFtotp
Fimt(aseTp
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 ENTERIC OCCULT REFLEX AND LARGE BOWEL TRANSIT 1917
as diluted in Krebs’ solution at a final concentrationf hexamethonium (100 �m/L), ondansetron (1moL/L), and PPADS (30 �mol/L). Krebs’ solution
mmol/L): NaCl, 120.35; KCl, 5.9; NaHCO3, 15.5;aH2PO4, 1.2; MgSO4, 1.2; CaCl2, 2.5; and glucose,
1.5 (gassed with 3% CO2-97% O2 (vol/vol), pH 7.3–.4).
Statistical AnalysisAnalysis of spontaneous junction potentials (sJPs)
as performed using custom-written routines on shortamples of recordings of electrical activity (�2–3 minutes inuration) that were resampled to 100 Hz.22 A smoothinglgorithm (100-ms average, 5 iterations) and an averageaseline algorithm (10 seconds, 3 iterations) were applied toeduce high frequency noise and for calculation of ampli-udes of spontaneous junction potentials, respectively (seeigure 3). The first recording was used to identify the peaksf spontaneous EJPs (sEJPs) and spontaneous IJPs (sIJPs) by
ocating inflexion points. The amplitude, calculated as theifference between the peak of the junction potential andhe average baseline was calculated for each sJP. To deter-
ine whether sJPs in the second recording were coordi-ated in time and had similar amplitudes to sJPs in the firstecording, 2 different search routines were used. The firstoutine used the time at the peak of an sJP in the firstecording and looked for any inflexion (sIJP or sEJP) in theecond recording that was closest in time to that point.nce the peak of an sJP was located in the second trace,
he amplitude, maximum slope, and time to half-maxi-um amplitude of the second peak were calculated.esults from this analysis were used to compare the
imilarity of the amplitudes of sJPs between the 2 record-ngs. Regression analysis was used to summarize theariability in the amplitude of sJPs between the 2 record-ngs and was expressed as the correlation coefficient r.he second routine, instead of searching for any inflexion
n the second recording, looked only for sJPs that wereimilar to those in the first trace (eg, if there was an IJP inhe first recording, the routine would only return thelosest IJP in the second recording). Results from thisnalysis were used to compare differences in time be-ween similar sJPs and were expressed as standard devia-ions calculated at 3 different amplitude levels (1–5, 5–9,nd �9 mV). All results are expressed as means � SEM.ne-way and 2-way factorial analysis of variance statistical
ests were used to compare differences between groups. Arobability of less than 0.05 was considered significant.cheffé post hoc tests were used. N refers to the number ofnimals on which observations were made.
ResultsEffects of Circumferential vs LongitudinalStretch AloneWe recently reported that maintained circumferen-
ial stretch of the guinea pig distal colon generated ongoing (
eristaltic reflex activity. This activity consists of an ongoingischarge of oral EJPs and anal IJPs, which are temporallyynchronized, in the circular and longitudinal smooth mus-le layers (Figure 1A).11–13 This stretch-evoked ongoing peri-taltic reflex activity, which can be recorded for severalours, is stretch sensitive and independent of muscle toneecause it occurs in the presence of smooth muscle relax-nts such as nifedipine or papavarine (Figure 1B).13 Trans-ural nerve stimuli applied to the middle of the prepara-
ion evoked robust polarized responses, consisting of anral EJP and an anal IJP in the circular muscle (Figure 1A).e examined whether this ongoing peristaltic reflex activity
n the distal colon is affected not only by circumferentialtretch but also by longitudinal stretch.
We found that maintained longitudinal stretch by it-elf when applied to the same preparations to whichircumferential stretch was not applied failed to generaterominent neural activity in the circular muscle (n � 6,igure 1B). When transmural nerve stimuli were appliedo the middle of a segment of colon that was stretchednly in the longitudinally axis, only low-amplitude junc-ion potentials were evoked, which were similar in am-litude to those in Figure 2B as described below.
igure 6. NO-mediated synaptic events. In the presence of guaneth-dine (0.5 �mol/L) and nifedipine (1 �mol/L), recordings were made from
yenteric neurons in a circumferentially stretched CMMP preparationhat were situated approximately 10 mm anal of transmural stimulatingTS) electrodes. (A) IPSP evoked in S neuron exhibiting spontaneousction potentials (0.5-ms duration, 5 pulses at 10 Hz, 30 V). (B) A similartimulus evoked FEPSPs in a neuron with ongoing FEPSPs. (C) IPSPvoked in another neuron in the presence of quanethidine (0.5 �mol/L).his neuron exhibited spontaneous action potentials, FEPSPs, androximal process potentials. (D) Following the further addition of L-NA
100 �mol/L), the IPSP in the same neuron was blocked.

ofbbeclnwftnwb
wcoapas(aw
s0T�tetvptwe
stwdosmcliR(ce
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1918 DICKSON ET AL GASTROENTEROLOGY Vol. 132, No. 5
Effects of Longitudinal Stretch on OngoingPeristaltic Reflex ActivityRemarkably, longitudinal stretch (to 25% and 60%
f the relaxed length of colon) reduced in a gradedashion the amplitude (Figure 2) and coordination ofoth oral EJPs and anal IJPs (n � 6; Figure 3) generatedy circumferential stretch. Longitudinal stretch had noffect on the resting membrane potential of circular mus-le cells (circumferential stretch, 45.3 � 2.7 mV; plus 60%ongitudinal stretch 44.6 � 2.7 mV; n � 7; P � .01). Mostotably, the inhibitory effects of longitudinal stretchere reversible because activity returned to control levels
ollowing removal of longitudinal stretch (Figure 3). Fur-hermore, polarized responses activated by transmuralerve stimulation applied to the center of the preparation,hich evokes an oral EJP and an anal IJP, were also reducedy longitudinal stretch (Figures 2 and 3A and B).
Inhibitory Effects of Longitudinal Stretch AreMediated by NOWe were interested in the mechanism that could
ithdraw ongoing reflex activity from the smooth mus-le. Previously, we had shown that mucosal stimulationf the guinea pig distal colon that was stretched in bothxes activates myenteric interneurons in peristaltic reflexathways that can be inhibited by exogenously appliednd neurally released NO.23 However, as we have alsohown previously,12,22 the NO synthesis inhibitor L-NA100 �mol/L, n � 4) had no effect on the frequency,mplitude, and correlation of oral EJPs and anal IJPs
5
5
Oral StretchOral
Anal
5 mm
5 s
Oral
Anal
A
B
hen added to the middle chamber of a preparation m
tretched in only the circumferential axis (R2 control,.35 � 0.035; L-NA, 0.36 � 0.014; P � .01; n � 4).herefore, we were most surprised to find that L-NA (100mol/L; n � 6) applied to the middle chamber between
he 2 recording sites completely reversed the inhibitoryffects of longitudinal stretch on both the amplitude andhe coordination of ongoing junction potentials acti-ated by circumferential stretch (Figures 2C and 3C) andolarized responses evoked by transmural nerve stimula-ion (Figures 2C and 4A and B). This suggests that NOas being released by myenteric neurons to inhibit the
nteric neural circuitry.
Effect of Removing Longitudinal Muscle
Because colonic elongation inhibits ongoing peri-taltic activity, we were interested in determining whetherhe sensory receptors responsible for this inhibitory reflexere located in the LM. Removal of the LM by sharpissection to create a CMMP preparation had no effectn the junction potentials activated by circumferentialtretch, as we have recently reported (Figure 4C).13 Re-
arkably, however, in these CMMP preparations, theoordinated oral EJPs and anal IJPs were still inhibited byongitudinal stretch to 60% of the relaxed length follow-ng repinning of the same preparation (Figure 4D; n � 5).emoval of longitudinal stretch restored control activity
Figure 4E). This suggests that the mechanosensitive pro-esses (dendrites) of neurons underlying this reflex are inither the myenteric plexus or in the underlying circular
5 mm
OralStretchPre Post
Figure 7. Effects of colonicelongation on pellet propulsion.(A) As shown in the spatiotem-poral map, an epoxy-coated pel-let was allowed to propel spon-taneously down the distal colon.Once propulsion was initiated,the pellet propelled at a constantvelocity down the colon. (B) Inanother run, once the pelletstarted to move at a constant ve-locity, colonic elongation wasapplied to the oral flap. Elonga-tion of the flap to approximatelytwice its resting length almoststopped pellet propulsion. Fol-lowing removal of the stimulus,the flap gradually relaxed back toits resting length, and the pelletthen continued to move downthe colon at its prestretched ve-locity.
mm
s
uscle.

illtmawmptoPttavnogFtaieoap(g0sgnefsf
ulCmmtiphaflF
i1r(0pn6rmnsctm3a
toict0amflssFtirr2d�ppntpm8pi
stc
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 ENTERIC OCCULT REFLEX AND LARGE BOWEL TRANSIT 1919
Activity of Myenteric Neurons inPreparations Stretched in Both AxesClearly, the myenteric neurons releasing NO to
nhibit this ongoing reflex activity respond selectively toongitudinal stretch. We reasoned that such neurons areikely to be NOS ve descending interneurons.24 To testhis hypothesis, we spatially monitored the activity of
any myenteric neurons simultaneously using Ca2 im-ging18 in preparations devoid of LM. These preparationsere stretched in both axes so that the excitability ofyenteric motor neurons and interneurons in peristaltic
athways was inhibited. In these experiments, hexame-honium (100 �mol/L; nicotinic receptor antagonist),ndansetron (1 �mol/L; 5-HT3 receptor antagonist), andPADs (30 �mol/L; P2X receptor antagonist) were addedo block all known forms of fast synaptic transmission inhe myenteric plexus.25 Under these circumstances, anyctive neurons are likely to be mechanosensory and acti-ated by longitudinal stretch. We found that only 1 to 3eurons per ganglia (mean, 2.0 � 0.3, n � 6) exhibitedngoing firing of Ca2 transients (1.25 � 0.23 Hz), sug-esting that they are likely to be mechanosensory (seeigure 6D). Later immunohistochemistry of the prepara-ions revealed that these active neurons were nNOS vend anally projecting, suggesting that they are likely to benterneurons (Figure 5B). L-NA (100 �mol/L) had noffect on the frequency (L-NA, 1.42 � 0.19 Hz; P � .01)f Ca2 transients in these nNOS ve neurons (Figure 5Cnd E, black traces). In addition, in L-NA (100 �mol/L), 2opulations of NOS �ve neurons also became active
Figure 5C and E). In one group (2.8 � 0.30 neurons peranglia), Ca2 transients occurred at a frequency of.72 � 0.12 Hz, whereas, the other group of teardrop-haped unipolar neurons (5.33 � 0.33 neurons per gan-lia) elicited a Ca2 wave (duration, 7.8 � 0.7 seconds; 22eurons from 6 ganglia), which was initiated at the apicalnd of the neuron, in any given 30-second period. There-ore, these NOS �ve neurons are probably mechanosen-itive interneurons in peristaltic pathways13,15 becauseast synaptic transmission was blocked (Figure 5E).
NO-Mediated Synaptic EventsWe attempted to determine the synaptic events
nderlying NO-mediated inhibition of reflex activity byongitudinal stretch. In circumferentially stretchedMMP preparations from which the LM had been re-oved, we made intracellular electrical recordings from 5yenteric neurons that fired spontaneous action poten-
ials and from 5 myenteric neurons that exhibited ongo-ng spontaneous bursts of fast excitatory postsynapticotentials (FEPSPs) (Figure 6A and B). The neurons ex-ibiting spontaneous action potentials are likely mech-nosensory interneurons driving ongoing peristaltic re-ex activity, whereas the neurons showing bursts of
EPSPs are likely to be muscle motor neurons.15 Repet- ftive transmural nerve stimulation (duration, 0.5 seconds;0 Hz for 5–10 pulses) applied �10 mm oral of theecording site evoked inhibitory postsynaptic potentialsIPSPs; duration, 2.6 � 0.4 seconds; amplitude, 8.6 �.4 mV) in the 5 neurons exhibiting spontaneous actionotentials (Figure 6A) and bursts of FEPSPs in motoreurons showing ongoing bursts of FEPSPs (FigureB).15 IPSPs, although rare in myenteric neurons, can beeadily evoked in submucosal neurons and are largely
ediated by the release of norepinephrine from extrinsicerve fibers acting on � 2 receptors on the soma ofubmucosal neurons.17 Therefore, our experiments werearried out in the presence of guanethindine (1 �mol/L)o block norepinephrine release from extrinsic nerve ter-
inals. L-NA (100 �mol/L) completely blocked IPSPs inneurons exhibiting spontaneous action potentials afterperiod of 15 to 20 minutes (n � 3) (Figure 6C and D).
Inhibition of Transit of a PelletWe reasoned that, if descending interneurons ac-
ivated by elongation were involved in inhibiting transitf a pellet, then elongating the oral flap of colon should
nhibit pellet propulsion. In control segments, epoxy-oated pellets, after a brief delay, traveled smoothly downhe entire length of the intact colon at a velocity of 1.25 �.18 mm/s (n � 4; Figure 7A). When the pellet hadssumed a constant velocity at a distance of 10.4 � 2.2m (n � 4) from the entrance of the intact segment, we
ound that elongating the oral flap to twice its restingength significantly reduced the velocity of pellet propul-ion down the colon to 0.26 � 0.13 mm/s, which wasignificantly different from control (P � .01, n � 4;igure 7B). Pellet propulsion was inhibited throughouthe duration of the stimulus (12.3 � 1.7 seconds), mov-ng only 3.0 � 2.0 mm (n � 4) down the colon. Followingelease of oral longitudinal stretch, the oral flap graduallyelaxed back to its resting length after a period of 7.2 �.9 seconds, and then the pellet resumed its propulsionown the colon at 2.5 � 0.5 mm/s (n � 4). L-NA (100mol/L, n � 4) applied to the bath blocked pellet pro-ulsion in 3 experiments, as previously described.26 In theresence of LNA, longitudinal stretch of the oral flap hado effect on pellet propulsion. In 1 preparation, however,he application of L-NA (100 �mol/L) had no effect onellet propulsion down the colon (velocity in L-NA, 1.7m/s). However, in the presence of L-NA, oral stretch for
.7 seconds also had no detectable effect on pellet pro-ulsion in this preparation, suggesting that the descend-
ng inhibitory reflex was blocked.
Effects of Colonic FillingWe further investigated whether longitudinal
tretch might have physiologic significance by examininghe effects of filling an intact isolated segment of distalolon with fluid (Figure 8A). The colon distended circum-
erentially in response to increasing volumes of injected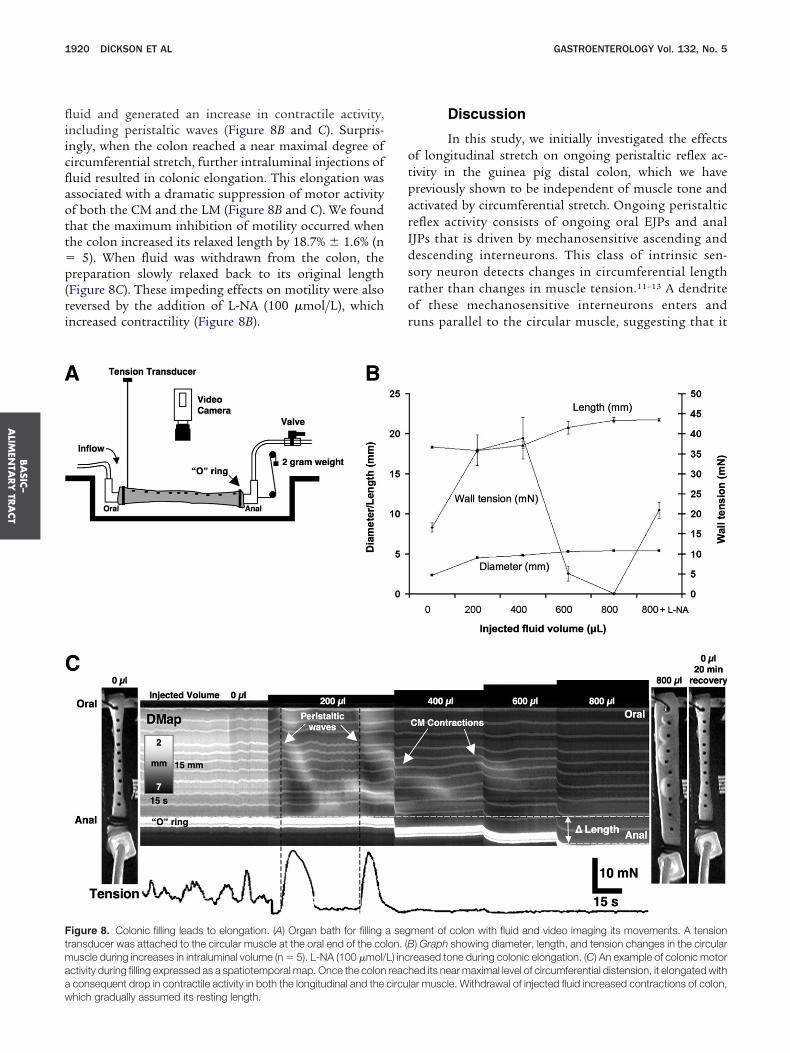
fliicflaott�p(ri
otparIdsror
Ftmaaw
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1920 DICKSON ET AL GASTROENTEROLOGY Vol. 132, No. 5
uid and generated an increase in contractile activity,ncluding peristaltic waves (Figure 8B and C). Surpris-ngly, when the colon reached a near maximal degree ofircumferential stretch, further intraluminal injections ofuid resulted in colonic elongation. This elongation wasssociated with a dramatic suppression of motor activityf both the CM and the LM (Figure 8B and C). We foundhat the maximum inhibition of motility occurred whenhe colon increased its relaxed length by 18.7% � 1.6% (n
5). When fluid was withdrawn from the colon, thereparation slowly relaxed back to its original length
Figure 8C). These impeding effects on motility were alsoeversed by the addition of L-NA (100 �mol/L), whichncreased contractility (Figure 8B).
igure 8. Colonic filling leads to elongation. (A) Organ bath for fillingransducer was attached to the circular muscle at the oral end of the couscle during increases in intraluminal volume (n � 5). L-NA (100 �mol/
ctivity during filling expressed as a spatiotemporal map. Once the colonconsequent drop in contractile activity in both the longitudinal and the
hich gradually assumed its resting length.Discussion
In this study, we initially investigated the effectsf longitudinal stretch on ongoing peristaltic reflex ac-ivity in the guinea pig distal colon, which we havereviously shown to be independent of muscle tone andctivated by circumferential stretch. Ongoing peristalticeflex activity consists of ongoing oral EJPs and analJPs that is driven by mechanosensitive ascending andescending interneurons. This class of intrinsic sen-ory neuron detects changes in circumferential lengthather than changes in muscle tension.11–13 A dendritef these mechanosensitive interneurons enters anduns parallel to the circular muscle, suggesting that it
ment of colon with fluid and video imaging its movements. A tension) Graph showing diameter, length, and tension changes in the circular
reased tone during colonic elongation. (C) An example of colonic motored its near maximal level of circumferential distension, it elongated with
lar muscle. Withdrawal of injected fluid increased contractions of colon,
a seglon. (BL) increachcircu

rttaa
stfgttbisradbwsalhh
saittitccpraAltgSn
Fctcmei(Aoma(tedpIjcastnlsrNSnci
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 ENTERIC OCCULT REFLEX AND LARGE BOWEL TRANSIT 1921
esponds to circumferential stretch.13,15 Ascending in-erneurons trigger excitatory motor neurons to bothhe CM and LM, whereas descending interneuronsctivate inhibitory motor neurons to the CM and LMt the same time.
Longitudinal Stretch and NOWe were most surprised to find that longitudinal
tretch decreased both the amplitude and the coordina-ion between oral EJPs and anal IJPs, activated by circum-erential stretch and transmural nerve stimulation, in araded manner at the same time. These effects of longi-udinal stretch were completely reversed by the NO syn-hesis inhibitor L-NA when applied to the middle cham-er between the 2 recording sites. This suggests that NO
s being released by myenteric neurons, which respondelectively to longitudinal stretch, to inhibit synaptic neu-otransmission within the myenteric plexus (Figure 9). Thislso suggests that the mechanosensitive interneuronsriving this reflex activated by circumferential stretch areeing inhibited by NO because they are the only neuronsithin the middle chamber that are likely to drive the
hort excitatory and inhibitory motor neurons at the oralnd anal ends of the preparation, respectively.13,15 In thearge intestine, motor neurons to the circular muscleave short projections (�1 mm), whereas interneurons
igure 9. Proposed neural cir-uit underlying colonic elonga-ion. (A) Schematic shows thatircumferential stretch activatesechanosensitive ascending
xcitatory (AEPs) and descend-ng inhibitory nerve pathwaysDIPs) underlying peristalsis.EPs and DIPs, which reinforcene another, activate excitatoryotor neurons (EMNs) orally
nd inhibitory motor neuronsIMNs) anally to both the longi-udinal and circular muscle lay-rs. Activation of EMNs pro-uces an excitatory junctionotential (EJP), and activation of
MNs produces an inhibitoryunction potential (IJP) in the cir-ular muscle, at the oral andnal ends of the segment, re-pectively. During circumferen-ial stretch, descending NOSeurons are quiescent. (B) Fol-
owing colonic elongation, de-cending NOS ve interneu-ons are activated to releaseO, which likely generates IP-Ps in mechanosensitive inter-eurons in AEPs and DIPs and aonsequent withdrawal of activ-
ty from both EMNs and IMNs.
ave projection lengths of �7 mm along the bowel.27,28 t
Interneurons in the ColonWe hypothesized that myenteric, NOS ve, de-
cending interneurons are likely to be mechanosensorynd selectively respond to longitudinal stretch by releas-ng NO. Furthermore, the neurons responding to longi-udinal stretch cannot be AH sensory neurons becausehey are not NOS ve. In the guinea pig distal colon,mmunohistochemical coding studies24 demonstrate thathere are 3 classes of ascending interneurons and 3lasses of descending interneurons that contain acetyl-holine (ACh), suggesting that they may be involved ineristaltic reflex pathways. A fourth descending interneu-on is immunoreactive only for NOS, whereas one of thebove descending interneurons contains in addition toCh, vasoactive intestinal polypeptide (VIP)/gastrin-re-
easing peptide (GRP)/�NOS/�Calbindin.24 The NOSve neuron responding to colonic elongation is likely to be
he class containing only NOS because ACh, calcitoninene-related peptide, and VIP excite myenteric neurons.25,29
imilar complex classes of ascending and descending inter-eurons have been observed in the human colon.30
Calcium Imaging of Activity in MyentericNeuronsBecause this inhibitory reflex triggered by longi-
udinal stretch is still present even when the LM has been

rtgCwabptpkcosftnmnrconr
iwteoieisnsstegbfbNstortbflbtt
tasiTwsl
sitditdpseppttsii
flpmgsTcsectodiflbbcm
omcp
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1922 DICKSON ET AL GASTROENTEROLOGY Vol. 132, No. 5
emoved suggests that the mechanosensory dendrites ofhese NOS-positive neurons must either be in myentericanglia or in the underlying circular muscle. We useda2 imaging of myenteric neurons in preparations thatere stretched in both axes so that peristaltic pathwaysctivated by circumferential stretch should be inhibitedy longitudinal stretch. Our rationale was that in thesereparations, the intrinsic sensory neurons that respondo longitudinal stretch should still be active. In thesereparations we used a combination of drugs to block allnown forms of fast synaptic transmission. Under theseircumstances, only 1 or 2 neurons per ganglia exhibitedngoing activity, suggesting that they are mechanosen-ory in response to longitudinal stretch. These wereound to be nNOS ve and anally projecting, suggestinghat they were likely to be descending interneurons. Theumber of ganglia in 1-cm length of colon is approxi-ately 1600 (40 � 40) ganglia,13 suggesting that the
umber of these mechanosensory nNOS ve interneu-ons (�2 per ganglia) is likely be approximately 3200 perm length of guinea pig distal colon. L-NA had no effectsn the frequency of Ca2 transients in these nNOS veeurons, which is not surprising because myenteric neu-ons that release NO are not themselves targets for NO.31
NO-Mediated Synaptic PotentialsClearly, NO released during longitudinal stretch
nhibits synaptic transmission in peristaltic nerve path-ays. Previous studies in the guinea pig ileum showed
hat sodium nitroprusside (NaNO; NO donor) had noffect on evoked FEPSPs, although it did produce “anccasional decrease in input resistance and hyperpolar-
zation” in some myenteric neurons.32 IPSPs can bevoked readily in submucous neurons,17 but they are raren myenteric neurons.33 IPSPs have, however, been ob-erved in some AH neurons and in some unidentified Seurons in the guinea pig distal colon.33 In this study, wehow for the first time that IPSPs can be generated inpontaneously firing mechanosensitive S interneuronshat underlie peristaltic nerve pathways but not appar-ntly in motor neurons.15 Presumably, the IPSPs wereenerated by the descending nNOS ve interneuronsecause our stimulus was applied oral to the gangliarom which recordings were made and the IPSPs werelocked by L-NA, suggesting that they are mediated byO. IPSPs would be expected to reduce the probability of
tretch-evoked action potentials in mechanosensory in-erneurons underlying the muscle tone independent ofngoing peristaltic reflex activity. Interestingly, in theeciprocal tendon reflex of the central nervous system,he � motor neurons to the extensor muscle are excitedy FEPSPs while, at the same time, � motor neurons toexor muscle are inhibited. This inhibition is producedy interneurons that release �-aminobutyric acid, ratherhan NO, which generates IPSPs to reduce action poten-
ial firing in � motor neurons.34 hPellet PropulsionWe showed that applying longitudinal stretch on
he oral segment of colon inhibited pellet propulsion fort least as long as the stimulus was maintained. Thisuggests that colonic elongation activates a descendingnhibitory reflex that can dramatically reduce transit.his reflex is an acute reflex because pellet propulsionas inhibited immediately following application of the
timulus. This descending reflex can be directly corre-ated with our electrophysiological findings.
Previously, it has been shown that the velocity of aingle pellet along the guinea pig distal colon is reducedn a dose-dependent manner by inhibiting NO produc-ion, suggesting that these effects are caused by blockingescending inhibition in front of a pellet.35 Interestingly,
t has also been demonstrated that the presence of mul-iple pellets in the guinea pig distal colon leads to aramatic slowing in the velocity and rate of emptying ofellets.21 In light of our current findings, this is noturprising because Gabella36 found that the colon waslongated over each pellet by approximately 29% com-ared with the region between pellets. In addition to theossible activation of descending inhibitory pathways byhe preceding pellet,21 we propose that the elongation ofhe colon by pellets also activates mechanosensitive de-cending (NOS ve) interneurons that release NO tonhibit enteric neurons in peristaltic circuits, thus slow-ng transit and promoting storage (Figure 9).
Colonic FillingWhen a segment of distal colon was filled with
uid, it first distended circumferentially and generatederistaltic waves, and then, as it reached its near maxi-um level of circumferential stretch, it began to elon-
ate. This elongation was associated with a dramaticuppression of motility that was shown to involve NO.his elongation is likely a natural consequence of theross-ply lattice arrangement of collagen fibers in theubmucosal layer that constrains the gut to distend inither the circumferential or longitudinal axis.36 Thisollagen matrix, which changes with the movements ofhe intestinal wall, appears to protect the bowel fromverstretching in both the circumferential and longitu-inal axes. It should be noted that accommodation in the
solated small intestine is considered to be the phase ofuid accumulation (filling phase), which is characterizedy radial distension accompanied by shortening of theowel, before the threshold for peristalsis is reached.37 Inontrast, our studies in the colon suggests that accom-odation is associated with elongation of the bowel.In summary, colonic filling by fluid leads to elongation
f the colon that is associated with a suppression ofotility. This is also supported by the observation that
olonic elongation applied oral to a pellet inhibits pelletropulsion. Therefore, our data are consistent with the
ypothesis that colonic elongation activates mechano-
stNintabeitfiaeccsocNtb
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
BA
SIC–
ALI
MEN
TARY
TRA
CT
May 2007 ENTERIC OCCULT REFLEX AND LARGE BOWEL TRANSIT 1923
ensitive NOS ve descending interneurons that respondo longitudinal stretch by releasing NO. The release ofO by these neurons is likely to generate IPSPs that
nhibit action potential firing in mechanosensitive inter-eurons and possibly other neurons underlying peristal-ic pathways (Figure 9). We refer to this elongation reflexs an intrinsic occult reflex (hidden from observation)ecause it withdraws all activity from the muscle. Ourvidence suggests that this intrinsic neural circuit is crit-cal for slow transit and accommodation of colonic con-ents. The occult reflex is probably deactivated followingeeding because this causes extrinsic vagal excitation ofntrinsic cholinergic motor neurons (gastrocolonic reflex)nd an increase in motility.1– 4 This excitation would bexpected to produce contraction of the longitudinal mus-le leading to a reduction in colonic elongation. Clini-ally, the intrinsic occult reflex provides a negative feedbackystem that would be expected to exacerbate the inhibitionf motility associated with constipation in an impactedolon. In support of this concept, an excess production ofO within the myenteric plexus has been recently proposed
o underlie slow transit constipation in the human largeowel.7,8
References
1. Christensen J. The motility of the colon. In: Johnson LR, ed.Physiology of the gastrointestinal tract, 4th ed. New York: Raven,1994;24:991–1024.
2. Smith TK, Sanders KM. Motility of the large intestine. In: YamadaT, Alpers DH, Owyang C, Powel DW, Silverstein FE, eds. Textbookof gastroenterology, vol 1. ch 10. Hagerstown, MD: Lippincott,Williams and Wilkins, 1995;234–260.
3. Camilleri M, Ford MJ. Colonic sensorimotor physiology in health,and its alteration in constipation and diarrhoeal disorders. Ali-ment Pharmacol Ther 1998;12:287–302.
4. Phillips SF. Functions of the large bowel: an overview. ScandJ Gastroenterol Suppl 1984;93:1–12.
5. Sanders K, Smith TK. Neural regulation of colonic motor function.Colonic Dis 2003;3:35–52.
6. Stevens RJ, Publicover NG, Smith TK. Induction and regulation ofCa2 waves by enteric neural reflexes. Nature 1999;399:62–66.
7. Cortesini C, Cianchi F, Infantino A, Lise M. Nitric oxide synthaseand VIP distribution in enteric nervous system in idiopathicchronic constipation. Dig Dis Sci 1995;40:2450–2455.
8. Tomita R, Fujisaki S, Ikeda T, Fukuzawa M. Role of nitric oxide inthe colon of patients with slow-transit constipation. Dis ColonRectum 2002;45:593–600.
9. Smith TK, Hennig GW, Spencer NJ. Sensory transduction in theENS. Physiol News 2005;58:1–3.
0. Smith TK, Oliver GR, Hennig GW, O’Shea DM, Vanden Berghe P,Kang SH, Spencer NJ. A smooth muscle tone-dependent stretch-activated migrating motor pattern in isolated guinea-pig distalcolon. J Physiol 2003;551:955–969.
1. Spencer NJ, Hennig GW, Smith TK. A rhythmic motor patternactivated by circumferential stretch in guinea-pig distal colon.J Physiol 2002;545:629–648.
2. Spencer NJ, Hennig GW, Smith TK. Stretch activated neuronalpathways in longitudinal and circular muscle of guinea-pig distal
colon. Am J Physiol 2003;284:G231–G241.3. Spencer NJ, Dickson EJ, Hennig GW, Smith TK. Sensory elementswithin the circular muscle are essential for mechanotransductionof ongoing peristaltic reflex activity in guinea-pig distal colon.J Physiol 2006;576:519–531.
4. Kunze WA, Furness JB, Bertrand PP, Bornstein JC. Intracellularrecording from myenteric neurons of the guinea-pig ileum thatrespond to stretch. J Physiol 1998;506:827–842.
5. Spencer NJ, Smith TK. Mechanosensory S-neurons rather thanAH neurons appear to generate a rhythmic motor pattern inguinea-pig distal colon. J Physiol 2004;558:577–596.
6. Lynn P, Zagorodnyuk V, Hennig GW, Costa M, Brookes S. Me-chanical activation of rectal intraganglionic laminar endings in theguinea pig distal gut. J Physiol 2005;564:589–601.
7. North RA, Surprenant A. Inhibitory synaptic potentials resultingfrom -adrenoceptor activation in guinea-pig submucous plexusneurones. J Physiol 1985;358:17–33.
8. Park KJ, Hennig GW, Lee HT, Spencer NJ, Ward SM, Smith TK,Sanders KM. Spatial and temporal mapping of pacemaker activ-ity in interstitial cells of Cajal in mouse ileum in situ. Am J PhysiolCell Physiol 2006;290:C1411–C1427.
9. Brookes SJH, Chen BN, Costa M, Humpheys CM. Initiation ofperistalsis by circumferential stretch of flat sheets of guinea-pigileum. J Physiol 1999;516:525–538.
0. Hennig GW, Costa M, Chen BN, Brookes SJ. Quantitative analysisof peristalsis in the guinea-pig small intestine using spatio-tem-poral maps. J Physiol 1999;517:575–590.
1. Hennig GW, Costa M, Humphreys CM, Brookes SJ. Analysis ofmotor patterns in the isolated guinea-pig large intestine by spatio-temporal maps. Neurogastroenterol Motil 2001;13:483–492.
2. Spencer NJ, Hennig GW, Smith TK. Spatial and temporal coordi-nation of junction potentials in circular muscle of guinea-pigdistal colon. J Physiol 2001;535:565–578.
3. Smith TK, McCarron S. Nitric oxide modulates cholinergic reflexpathways to the longitudinal and circular muscle in the isolatedguinea-pig distal colon. J Physiol 1998;512:893–906.
4. Lomax AE, Furness JB. Neurochemical classification of entericneurons in the guinea-pig distal colon. Cell Tissue Res 2000;302:59–72.
5. Galligan JJ. Pharmacology of synaptic transmission in the entericnervous system. Curr Opin Pharmacol 2002;2:623–629.
6. Foxx-Orenstein AE, Grider JR. Regulation of colonic propulsion byenteric excitatory and inhibitory neurotransmitters. Am J Physiol1996;271:G433–G437.
7. Neunlist M, Michel K, Aubé A-C, Galmiche J, Schemann M. Pro-jections of excitatory and inhibitory motor neurones to the circularand longitudinal muscle of the guinea pig colon. Cell Tissue Res2001;305:325–330.
8. Spencer NJ, Hennig GW, Dickson E, Smith TK. Synchronization ofenteric neuronal firing during the murine colonic MMC. J Physiol2005;564:829–847.
9. Wood JD. Physiology of the enteric nervous system. In: JohnsonLR, ed. Physiology of the gastrointestinal tract. 3rd ed. New York:Raven, 1994:423–482.
0. Wattchow DA, Porter AJ, Costa M. The polarity of neurochemicallydefined myenteric neurons in the human colon. Gastroenterology1997;113:497–506.
1. Shuttleworth CW, Xue C, Ward SM, de Vente J, Sanders KM.Immunohistochemical localization of 3’,5’-cyclic guanosinemonophosphate in the canine proximal colon: responses to nitricoxide and electrical stimulation of enteric inhibitory neurons.Neurosci 1993;56:513–522.
2. Tamura K, Schemann M, Wood JD. Actions of nitric oxide-gener-ating sodium nitroprusside in myenteric plexus of guinea pigsmall intestine. Am J Physiol 1993;265:G887–G893.
3. Lomax AE, Sharkey KA, Bertrand PP, Low AM, Bornstein JC,
Furness JB. Correlation of morphology, electrophysiology and
3
3
3
3
NS
Df
Gfc
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1924 DICKSON ET AL GASTROENTEROLOGY Vol. 132, No. 5
chemistry of neurons in the myenteric plexus of the guinea-pigdistal colon. J Auton Nerv Syst 1999;76:45–61.
4. Byrne JH. Postsynaptic potentials and synaptic integration. In:Byrne JH, Roberts JL, eds. From molecules to networks. NewYork: Elsevier Academic Press, 2003;459–478.
5. On the musculature of the gastro-intestinal tract. Anat Embryol1981;163:135–156.
6. Gabella G. The cross-ply arrangement of collagen fibres in thesubmucosa of the mammalian small intestine. Cell Tissue Res1987;248:491–497.
7. Waterman SA, Costa M, Tonini M. Accommodation mediated byenteric inhibitory reflexes in the isolated guinea-pig small intes-
tine. J Physiol 1994;474:539–546.Received November 14, 2006. Accepted February 15, 2007.Address requests for reprints to: Terence K. Smith, PhD, University of
evada School of Medicine, Physiology and Cell Biology, 1600 North Virginiatreet, Reno, Nevada 89557. e-mail: [email protected]; fax: (775) 784-6908.Supported by a grant from the National Institute of Diabetes and
igestive and Kidney Diseases (RO1 DK45713; to T.K.S. and N.J.S.) andor imaging performed in a Core laboratory by grant P20 RR-18751.
E.J.D., N.J.S., and G.W.H. contributed equally to this work.E.J.D., N.J.S., and J.R. performed muscle and neuron recordings;
.W.H. developed algorithms for data analysis; P.O.B. and E.J.D. per-ormed Ca2�� imaging; and D.J.H. performed pellet propulsion andolonic filling experiments.
There are no conflicts of interest to disclose.Copyright © 2022 FDOKUMEN




















