
An emended description of the genus Fogedia (Bacillariophyceae) with reports of four species new to...
10
An emended description of the genus Fogedia (Bacillariophyceae) with reports of four species new to science from a Korean sand flat JINSOON PARK 1 ,JONG SEONG KHIM 1 *, JONGSEONG RYU 2 ,CHUL-HWAN KOH 1 AND ANDRZEJ WITKOWSKI 3 1 School of Earth and Environmental Sciences and Research Institute of Oceanography, Seoul National University, Seoul, South Korea 2 Department of Marine Biotechnology, Anyang University, Ganghwagun, Incheon, South Korea 3 Institute of Marine Sciences, University of Szczecin, Mickiewicza 18, PL-70-383 Szczecin, Poland PARK J., KHIM J.S., RYU J., KOH C.-H. AND WITKOWSKI A. 2013. An emended description of the genus Fogedia (Bacillariophyceae) with reports of four species new to science from a Korean sand flat. Phycologia 52: 437–446. DOI: 10. 2216/12–120.1 A taxonomic investigation of the microphytobenthos from the lower intertidal zone of a Korean sandflat revealed unprecedented dominance and diversity of the marine diatom genus Fogedia. Four new species are described: Fogedia coreana sp. nov., F. densa sp. nov., F. elliptica sp. nov., and F. lyra sp. nov. A great abundance of Fogedia species in the study area provided a unique opportunity to observe the frustule ultrastructure and cell contents. The first reported observation of the presence of two girdle-appressed plastids each with a single rod-like pyrenoid was made in species of Fogedia, indicating its close relationship to Navicula sensu stricto and Hippodonta. Ultrastructural characteristics (e.g. hymenate areola occlusions) of Fogedia also support such a relationship. Meanwhile the genus Fogedia differs from Navicula sensu stricto with respect to the raphe system and differs from Hippodonta with respect to the girdle bands and apical striae. The present study increased the overall worldwide number of the taxa of Fogedia to 13. An emended generic description of Fogedia is provided along with the biogeographic information of the species belonging to the genus. KEY WORDS: Distribution, Diversity, Fogedia, Korea, New species, Plastid, Pyrenoid, Saemangeum, Sand flat, Ultrastructure INTRODUCTION Round et al. (1990) stated that the genus Navicula was a heterogenous entity even after certain genera (e.g. Neidium and Anomoeoneis) were removed. Since then, many new genera have been established from Navicula sensu lato, especially species found in freshwater and terrestrial environments [e.g. Geissleria (Lange-Bertalot & Metzeltin 1996), Hippodonta (Lange-Bertalot et al. 1996), Mayamaea and Fistulifera (Lange-Bertalot 1997), Adlafla (Moser et al. 1998), Microcostatus (Johansen & Sray 1998), Neidiopsis (Lange-Bertalot & Genkal 1999), Decussata (Lange-Bertalot 2000), Envekadea (Gligora et al. 2009), and Pseudofallacia (Liu et al. 2012)]. The genus Muelleria should also be mentioned in that it was once combined into Navicula sensu lato (Hustedt 1961–1966) and then later reclassified as a separate genus (Spaulding & Stoermer 1997). During the same period, only a few marine genera were separated from Navicula sensu lato [e.g. Amicula, Austariella, and Kurpiszia (Witkowski et al. 2000)]. This may result from freshwater habitats having a wider range of environmental parameters (nutrients, temperature, altitude, alkalinities, pH, etc.) and from local isolation that should facilitate evolution. In addition, there is generally less scientific expertise and research conducted on marine naviculoid diatoms compared with corresponding studies in freshwater. The genus Fogedia, based on Navicula giffeniana Foged (Witkowski et al. 1997), is a marine diatom separated from Navicula sensu lato. Originally Fogedia was differentiated based on the placement of the internal raphe fissure and the presence of a lateral area. In addition, the apical raphe endings of Fogedia species were short and only slightly bent, terminating at a certain distance from the apices (Witkowski et al. 1997). These differ from the apical raphe endings of Navicula sensu stricto, which are clearly geniculate (i.e. strongly bent towards the valve secondary side). The authors mostly worked on processed material, hence observations on the shape and number of plastids in Fogedia have not been published. Three more species were recorded (Witkowski et al. 2000, 2010b), and presently eight Fogedia species are known (Table 1). While the species have been reported to inhabit many different places of the world, a majority of the recorded taxa were the only member of the genus found in one location studied in the above-mentioned literature. Likewise, a predominance of Fogedia species in diatom assemblages has not been previously reported. We conducted a taxonomic investigation in the lower tidal zone of the estuarine Gyehwa sand flat that is located in the Saemangeum area on the west coast of Korea (35830 0 N to 35850 0 N and 125840 0 E to 126800 0 E). Before the construction of a dike, the Saemangeum area was governed by a macro tidal regime (tidal range ¼ 1.2–7.2 m), and it was the largest estuary in South Korea, with a breadth of 20 km at its widest location (Oh & Koh 1995). Two large rivers, Mangyeong and Dongjin, flow into the Saemangeum area supplying a large amount of nutrient-rich freshwater. The grain size of the Saemangeum tidal flat was mud-dominated in the upper intertidal areas and sand-dominated in the lower intertidal * Corresponding author ([email protected]). Phycologia (2013) Volume 52 (5), 437–446 Published 4 September 2013 437
Transcript of An emended description of the genus Fogedia (Bacillariophyceae) with reports of four species new to...

An emended description of the genus Fogedia (Bacillariophyceae)
with reports of four species new to science from a Korean sand flat
JINSOON PARK1, JONG SEONG KHIM
1*, JONGSEONG RYU2, CHUL-HWAN KOH
1AND ANDRZEJ WITKOWSKI
3
1School of Earth and Environmental Sciences and Research Institute of Oceanography, Seoul National University, Seoul, SouthKorea
2Department of Marine Biotechnology, Anyang University, Ganghwagun, Incheon, South Korea3Institute of Marine Sciences, University of Szczecin, Mickiewicza 18, PL-70-383 Szczecin, Poland
PARK J., KHIM J.S., RYU J., KOH C.-H. AND WITKOWSKI A. 2013. An emended description of the genus Fogedia(Bacillariophyceae) with reports of four species new to science from a Korean sand flat. Phycologia 52: 437–446. DOI: 10.2216/12–120.1
A taxonomic investigation of the microphytobenthos from the lower intertidal zone of a Korean sandflat revealedunprecedented dominance and diversity of the marine diatom genus Fogedia. Four new species are described: Fogediacoreana sp. nov., F. densa sp. nov., F. elliptica sp. nov., and F. lyra sp. nov. A great abundance of Fogedia species in thestudy area provided a unique opportunity to observe the frustule ultrastructure and cell contents. The first reportedobservation of the presence of two girdle-appressed plastids each with a single rod-like pyrenoid was made in species ofFogedia, indicating its close relationship to Navicula sensu stricto and Hippodonta. Ultrastructural characteristics (e.g.hymenate areola occlusions) of Fogedia also support such a relationship. Meanwhile the genus Fogedia differs fromNavicula sensu stricto with respect to the raphe system and differs from Hippodonta with respect to the girdle bands andapical striae. The present study increased the overall worldwide number of the taxa of Fogedia to 13. An emended genericdescription of Fogedia is provided along with the biogeographic information of the species belonging to the genus.
KEY WORDS: Distribution, Diversity, Fogedia, Korea, New species, Plastid, Pyrenoid, Saemangeum, Sand flat,Ultrastructure
INTRODUCTION
Round et al. (1990) stated that the genus Navicula was aheterogenous entity even after certain genera (e.g. Neidiumand Anomoeoneis) were removed. Since then, many newgenera have been established from Navicula sensu lato,especially species found in freshwater and terrestrialenvironments [e.g. Geissleria (Lange-Bertalot & Metzeltin1996), Hippodonta (Lange-Bertalot et al. 1996), Mayamaeaand Fistulifera (Lange-Bertalot 1997), Adlafla (Moser et al.1998), Microcostatus (Johansen & Sray 1998), Neidiopsis(Lange-Bertalot & Genkal 1999), Decussata (Lange-Bertalot2000), Envekadea (Gligora et al. 2009), and Pseudofallacia(Liu et al. 2012)]. The genus Muelleria should also bementioned in that it was once combined into Navicula sensulato (Hustedt 1961–1966) and then later reclassified as aseparate genus (Spaulding & Stoermer 1997). During thesame period, only a few marine genera were separated fromNavicula sensu lato [e.g. Amicula, Austariella, and Kurpiszia(Witkowski et al. 2000)]. This may result from freshwaterhabitats having a wider range of environmental parameters(nutrients, temperature, altitude, alkalinities, pH, etc.) andfrom local isolation that should facilitate evolution. Inaddition, there is generally less scientific expertise andresearch conducted on marine naviculoid diatoms comparedwith corresponding studies in freshwater.
The genus Fogedia, based on Navicula giffeniana Foged(Witkowski et al. 1997), is a marine diatom separated from
Navicula sensu lato. Originally Fogedia was differentiatedbased on the placement of the internal raphe fissure and thepresence of a lateral area. In addition, the apical rapheendings of Fogedia species were short and only slightly bent,terminating at a certain distance from the apices (Witkowskiet al. 1997). These differ from the apical raphe endings ofNavicula sensu stricto, which are clearly geniculate (i.e.strongly bent towards the valve secondary side). The authorsmostly worked on processed material, hence observations onthe shape and number of plastids in Fogedia have not beenpublished. Three more species were recorded (Witkowski etal. 2000, 2010b), and presently eight Fogedia species areknown (Table 1). While the species have been reported toinhabit many different places of the world, a majority of therecorded taxa were the only member of the genus found inone location studied in the above-mentioned literature.Likewise, a predominance of Fogedia species in diatomassemblages has not been previously reported.
We conducted a taxonomic investigation in the lower tidalzone of the estuarine Gyehwa sand flat that is located in theSaemangeum area on the west coast of Korea (358300N to358500N and 1258400E to 1268000E). Before the constructionof a dike, the Saemangeum area was governed by a macrotidal regime (tidal range¼ 1.2–7.2 m), and it was the largestestuary in South Korea, with a breadth of 20 km at its widestlocation (Oh & Koh 1995). Two large rivers, Mangyeongand Dongjin, flow into the Saemangeum area supplying alarge amount of nutrient-rich freshwater. The grain size ofthe Saemangeum tidal flat was mud-dominated in the upperintertidal areas and sand-dominated in the lower intertidal* Corresponding author ([email protected]).
Phycologia (2013) Volume 52 (5), 437–446 Published 4 September 2013
437

areas. The Saemangeum reclamation project was initiated inthe study area beginning in the early 1990s, with theobjective to build a 33-km-long dike and convert ~400km2 of tidal flats within the dike into farmlands. Dikeconstruction was completed in April 2006, blocking all tidalcurrents (Koh et al. 2010; Ryu et al. 2011). Our studyrevealed the dominance of Fogedia species, providing aunique opportunity to observe the frustule ultrastructure andcell contents (e.g. plastids and pyrenoids). We providedescriptions of the four new species, an emended descriptionof Fogedia, and information on the autecology anddistribution of species.
MATERIAL AND METHODS
Sediment samples of the Saemangeum sand flat surface (0–0.5 cm depth) were collected from a low tidal area (358460N,1268350E) during the ebb tide in August 2003. Samples weretransferred to the laboratory, and live material was observedfor chloroplast information using the cover slip method(Round 1953). Sample material was treated with concen-trated HCl and H2O2 to remove calcium carbonate particlesand organic substances, respectively. Permanent slides weremade using Mountmedia (Wako Pure Chemical Industries,Osaka, Japan), which is a commercial equivalent of Pleurax.Photomicrographs were obtained using an Olympus BX60light microscope (LM; Olympus, Tokyo, Japan) with a 360Plan-Apochromatic objective lens or a Zeiss Axioscop LM(Carl Zeiss, Oberkochen, Germany) with a 3100 Plan-Apochromat objective lens. For scanning electron micros-copy (SEM), a few drops of cleaned material were air-driedon aluminium stubs, coated with gold and examined at 5 kVin a Hitachi S4500 SEM (Hitachi Corp., Tokyo, Japan). TheSEM stubs were deposited in the H. Lange-BertalotCollection, Botanical Institute, J.-W. Goethe University,Frankfurt amMain, Germany. Additional SEM informationwas also obtained for the same material using a JEOL 6301FSEM (JEOL Ltd, Tokyo, Japan) at 5 kV (SupplementaryMaterials).
RESULTS
Fogedia Witkowski, Lange-Bertalot & Metzeltin emended by
Witkowski & Park
[Witkowski A., Metzeltin D., Lange-Bertalot H. & Bafana G.1997. Nova Hedwigia 65: 81.]
DESCRIPTION: Cells solitary, naviculoid in shape, with two girdle-appressed plastids located along the valve margin (as typical inNavicula sensu stricto), and with two pyrenoids. Plates not smooth (asin typical Navicula sensu stricto) but somewhat thickened andundulated in the middle, often accompanied by four conspicuousstorage granules near the isthma and plastid juncture. Valves broadlylanceolate with slightly protracted apices. Valve face flat, abruptlybending into a shallow mantle. Transapical striae uniseriate,composed of apically elongate, simple slit-like areolae occluded byhymenes (similar to Navicula sensu stricto). Areolae around thecentral area often differed from the others with altered shape andorientation. Striae in some species interrupted by hyaline lateral areasslightly depressed below the surface of the valve face. Raphe medial,straight, not internally on side of the raphe costa. External centralraphe endings straight, externally rather expanded, internally simple.Apical raphe endings very short, turned towards the same side of thevalve; however, may be rather prolonged but never strongly geniculate(as in Navicula sensu stricto). Girdle narrow, composed of twounperforated bands. Species found in marine, and rarely in brackish,water habitats. Members of the epipelon.
TYPE SPECIES: Fogedia giffeniana (N. Foged) A. Witkowski, H.Lange-Bertalot, D. Metzeltin, & G. Bafana in A. Witkowski, D.Metzeltin, H. Lange-Bertalot, & G. Bafana (1997) Nova Hedwigia 65:81.
Fogedia coreana Park, Khim, Koh & Witkowski sp. nov.(Figs 1–5, S1–S5, S20–S23)
DESCRIPTION: Valves moderately variable from elliptic or broadlyelliptical-lanceolate to linear-elliptic, ends cuneate and finally shortlyprotracted, subrostrate, 17–46 lm in length, 9–13 lm in width. Raphecentral external endings punctiform, while external apical rapheendings slightly bent in the same direction. Axial area very narrow,linear throughout. Central area small, formed by a few shortenedstriae, rhomboid in shape. Lateral area (typical of some Fogedia spp.)absent. Transapical striae 12–14 in 10 lm. The striae composed ofareolae with differentiated density.
Table 1. List of Fogedia species and their geographic distributions.
Species Locality Reference
F. acuta (Salah) Witkowski, Lange-Bertalot & Metzeltin
England, Oregon (USA) Salah 1955; Witkowski et al. 2000
F. christensenii Witkowski, Metzeltin &Lange-Bertalot
Fiji Witkowski et al. 1997
F. finmarchica (Cleve & Grunow)Witkowski, Metzeltin & Lange-Bertalot
Norway, England, Greenland, AegeanSea, Fiji, Baltic Sea, Chile
Cleve & Grunow 1880; Salah 1955; vanden Heuvel & Prud’homme van Reine1985; Foged 1973, 1985, 1987;Witkowski et al. 1997
F. geisslerae Witkowski, Metzeltin &Lange-Bertalot
Baltic Sea, Danish-German North Sea Witkowski 1994; Witkowski et al. 1997
F. giffeniana (Foged) Witkowski,Metzeltin & Lange-Bertalot
Tanzania, Fiji, Oman Foged 1975, 1987; Witkowski et al. 1997
F. giffeniana var. yementica Bafana Yemen Witkowski et al. 1997F. heterovalvata (Simonsen) Witkowski,Metzeltin & Lange-Bertalot
Baltic Sea, Carribean Sea Simonsen 1959; Witkowski et al. 2000
F. krammeri Witkowski, Lange-Bertalot,Kociolek & Kulikovskiy
California (USA) Witkowski et al. 2010b
438 Phycologia, Vol. 52 (5), 2013
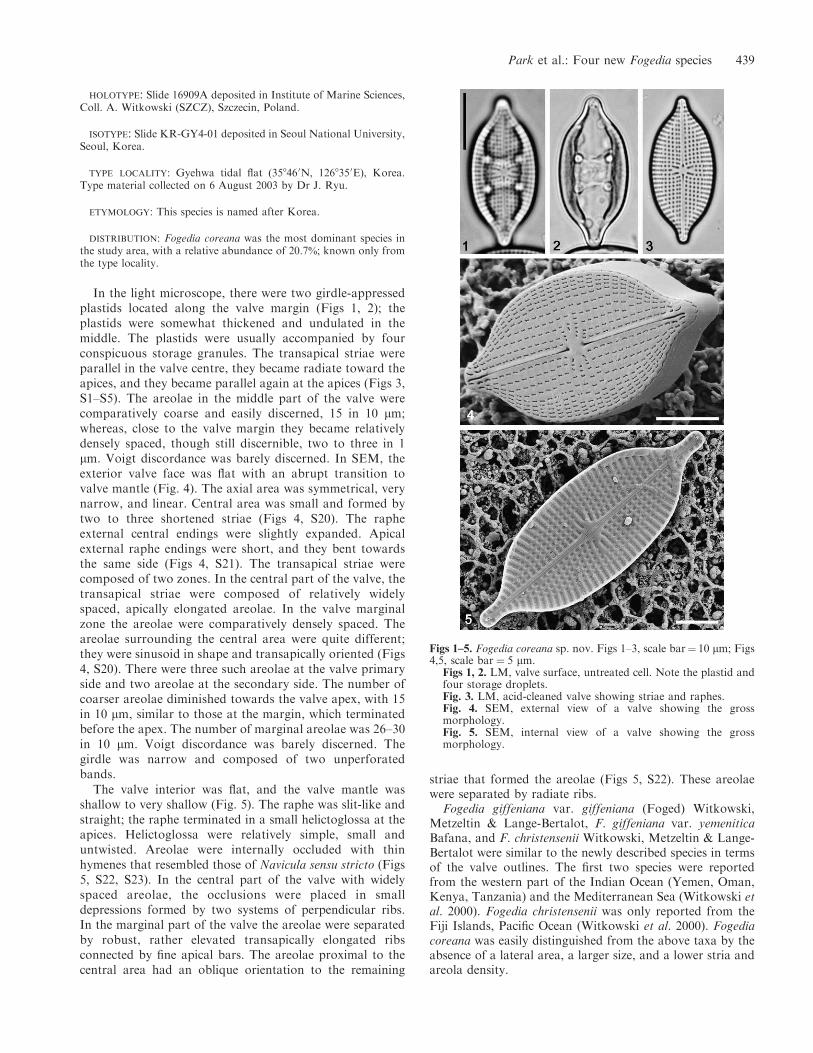
HOLOTYPE: Slide 16909A deposited in Institute of Marine Sciences,Coll. A. Witkowski (SZCZ), Szczecin, Poland.
ISOTYPE: Slide KR-GY4-01 deposited in Seoul National University,Seoul, Korea.
TYPE LOCALITY: Gyehwa tidal flat (358460N, 1268350E), Korea.Type material collected on 6 August 2003 by Dr J. Ryu.
ETYMOLOGY: This species is named after Korea.
DISTRIBUTION: Fogedia coreana was the most dominant species inthe study area, with a relative abundance of 20.7%; known only fromthe type locality.
In the light microscope, there were two girdle-appressedplastids located along the valve margin (Figs 1, 2); theplastids were somewhat thickened and undulated in themiddle. The plastids were usually accompanied by fourconspicuous storage granules. The transapical striae wereparallel in the valve centre, they became radiate toward theapices, and they became parallel again at the apices (Figs 3,S1–S5). The areolae in the middle part of the valve werecomparatively coarse and easily discerned, 15 in 10 lm;whereas, close to the valve margin they became relativelydensely spaced, though still discernible, two to three in 1lm. Voigt discordance was barely discerned. In SEM, theexterior valve face was flat with an abrupt transition tovalve mantle (Fig. 4). The axial area was symmetrical, verynarrow, and linear. Central area was small and formed bytwo to three shortened striae (Figs 4, S20). The rapheexternal central endings were slightly expanded. Apicalexternal raphe endings were short, and they bent towardsthe same side (Figs 4, S21). The transapical striae werecomposed of two zones. In the central part of the valve, thetransapical striae were composed of relatively widelyspaced, apically elongated areolae. In the valve marginalzone the areolae were comparatively densely spaced. Theareolae surrounding the central area were quite different;they were sinusoid in shape and transapically oriented (Figs4, S20). There were three such areolae at the valve primaryside and two areolae at the secondary side. The number ofcoarser areolae diminished towards the valve apex, with 15in 10 lm, similar to those at the margin, which terminatedbefore the apex. The number of marginal areolae was 26–30in 10 lm. Voigt discordance was barely discerned. Thegirdle was narrow and composed of two unperforatedbands.
The valve interior was flat, and the valve mantle wasshallow to very shallow (Fig. 5). The raphe was slit-like andstraight; the raphe terminated in a small helictoglossa at theapices. Helictoglossa were relatively simple, small anduntwisted. Areolae were internally occluded with thinhymenes that resembled those of Navicula sensu stricto (Figs5, S22, S23). In the central part of the valve with widelyspaced areolae, the occlusions were placed in smalldepressions formed by two systems of perpendicular ribs.In the marginal part of the valve the areolae were separatedby robust, rather elevated transapically elongated ribsconnected by fine apical bars. The areolae proximal to thecentral area had an oblique orientation to the remaining
striae that formed the areolae (Figs 5, S22). These areolaewere separated by radiate ribs.
Fogedia giffeniana var. giffeniana (Foged) Witkowski,Metzeltin & Lange-Bertalot, F. giffeniana var. yemeniticaBafana, and F. christensenii Witkowski, Metzeltin & Lange-Bertalot were similar to the newly described species in termsof the valve outlines. The first two species were reportedfrom the western part of the Indian Ocean (Yemen, Oman,Kenya, Tanzania) and the Mediterranean Sea (Witkowski etal. 2000). Fogedia christensenii was only reported from theFiji Islands, Pacific Ocean (Witkowski et al. 2000). Fogediacoreana was easily distinguished from the above taxa by theabsence of a lateral area, a larger size, and a lower stria andareola density.
Figs 1–5. Fogedia coreana sp. nov. Figs 1–3, scale bar¼ 10 lm; Figs4,5, scale bar ¼ 5 lm.
Figs 1, 2. LM, valve surface, untreated cell. Note the plastid andfour storage droplets.Fig. 3. LM, acid-cleaned valve showing striae and raphes.Fig. 4. SEM, external view of a valve showing the grossmorphology.Fig. 5. SEM, internal view of a valve showing the grossmorphology.
Park et al.: Four new Fogedia species 439

Fogedia densa Park, Khim, Koh & Witkowski sp. nov.
(Figs 6–10, S6–S11, S24–S27)
DESCRIPTION: Valves elliptic-lanceolate to linear-elliptic, with
short-protracted and acutely rounded apices, 13–21 lm in length,5.5–6 lm in width. Raphe central external endings punctiform;external apical raphe endings slightly bent in the same direction. Axialarea very narrow, linear throughout. Central area small, formed by afew shortened striae, rhomboid in shape. Lateral area typical of someFogedia spp. absent. Transapical striae 18–21 in 10 lm, Areolae 21–24in 10 lm.
HOLOTYPE: Slide 16909B deposited in the Institute of Marine
Sciences, Coll. A. Witkowski (SZCZ), Szczecin, Poland.
ISOTYPE: Slide KR-GY4-02 in deposited at Seoul National
University, Seoul, Korea.
TYPE LOCALITY: Gyehwa tidal flat (358460N, 1268350E), Korea; type
material was sampled on 6 August 2003 by Dr J. Ryu.
ETYMOLOGY: Latin densa¼ dense; refers to densely spaced areolaeand striae.
DISTRIBUTION: F. densa had a relative abundance of 5.6% at thetype locality. The species was also observed at a sand flat at GyeonggiBay (378350N, 1268220E), Yellow Sea, Korea, and at a sand flat inTianjin (398500N, 1178390E), Yellow Sea, China.
The chloroplasts of Fogedia densa were the same as in F.coreana when viewed in the light microscope (Figs 6, 7). Thetransapical striae radiated in the middle of the valve andwere parallel at the apices (Figs 8, S6–S11). The areolae weredensely and evenly spaced. Voigt discordance was barelydiscerned. In the SEM, the exterior valve face was flat withan abrupt transition to the very shallow valve mantle (Fig.9). The axial area was symmetrical, very narrow, and linear.The central area was small, formed by two or threeshortened striae. The external central endings of the raphewere slightly expanded (Figs 9, S24). Apical external rapheendings were slightly bent toward the same side (Figs 9, S25).
Figs 6–10. Fogedia densa sp. nov. Figs 6–9, scale bar ¼ 10 lm; Fig. 10, scale bar¼ 1 lm.Figs 6, 7. LM, valve surface, untreated cell. Note the plastid and storage droplets.Fig. 8. LM, acid-cleaned valve, showing striae and raphes.Fig. 9. SEM, external view of a valve showing the gross morphology.Fig. 10. SEM, external valve apex showing the simple raphe terminal ending.
440 Phycologia, Vol. 52 (5), 2013

The areolae and transapical striae were evenly spaced. Theareolae that surrounded the central area were rather differentfrom those with a transapical orientation and sometimes hada sinuous shape (Figs 9, S24).
The valve interior was flat, and the valve mantle was veryshallow (Figs S26, S27). The raphe slit ran quite straightfrom the central nodule to the helictoglossa. Helictoglossawere relatively simple, small, and untwisted (Fig. S27). Theareolae were internally occluded with thin hymens, whichresembled those of Navicula sensu stricto. The areolae andtransapical striae were evenly spaced. A few areolaeproximal to the central area had an oblique orientation tothe remaining striae forming areolae (Fig. S26). Fogediadensa was easily distinguished from the other members of thegenus by the absence of a lateral area, a smaller size, and ahigh stria and areola density.
Fogedia elliptica Park, Khim, Koh & Witkowski sp. nov.(Figs 11–16, S12–S15, S28–S31)
DESCRIPTION: Valves elliptic to elliptic-lanceolate, 16–41 lm inlength, 8–11.5 lm in width. Raphe central external endingspunctiform, slightly expanded. Raphe external central endings slightlyexpanded. Axial area narrow, linear throughout. Central area small,formed by a few slightly shortened striae, rhomboid in shape. Typicallateral area of Fogedia absent. Transapical striae 9–12.5 in 10 lm,areolae 21 in 10 lm.
HOLOTYPE: Slide 20703, deposited in the Institute of MarineSciences, Coll. A. Witkowski (SZCZ), Szczecin, Poland.
ISOTYPE: Slide KR-GY4-03, deposited in Seoul National Univer-sity, Seoul, Korea.
TYPE LOCALITY: Gyehwa tidal flat (358460N, 1268350E), Korea; typematerial was sampled on 6 August 2003 by Dr J. Ryu.
ETYMOLOGY: Latin elliptica ¼ elliptic; refers to its elliptic valveoutline.
DISTRIBUTION: Fogedia elliptica had a relative abundance of 5.1% atthe type locality. The species was also observed at a sand flat atTianjin (398500N, 1178390E), northern coast, Yellow Sea, China, and amud flat at Yancheng (338150N, 1208450E), southern Yellow Sea,China.
The chloroplasts of Fogedia elliptica were same as in F.coreana when viewed in the light microscope (Figs 11, 12).The apices were broadly rostrate in large specimens, but theywere not protracted in small specimens (Figs S12–S15).External apical raphe endings were difficult to discern. Thetransapical striae were parallel in the middle of the valve, andthey were slightly radiate at the apices (Figs 13, 14). Theareolae were densely and evenly spaced. Voigt discordancewas barely discerned. In the SEM, the exterior valve face wasflat with an abrupt transition to the valve mantle (Fig. 15).The axial area was symmetrical and narrowly linear. Centralarea was small, formed by a few slightly shortened striae; theshortened striae were due to deformation and packing ratherthan omission of certain areolae. Raphe external centralendings were slightly expanded. Apical external rapheendings were unlike those in other Fogedia except F.giffeniana; the raphe endings were abruptly deflected towardsthe secondary sides (Figs 15, S28). They were somewhat
similar to those of Navicula sensu stricto, yet they were notstrongly geniculate but diagonally straight, and thereforedifferentiated from Navicula spp. Areolae and transapicalstriae were evenly spaced. A few areolae proximal to thecentral area were round in shape and oblique in orientationto the remaining striae that formed areolae (Figs 15, S29).The girdle was narrow and composed of two unperforatedbands.
Figs 11–16. Fogedia elliptica sp. nov. Scale bars ¼ 10 lm.Figs 11, 12. LM, valve surface, untreated cell. Note the plastidand four storage droplets.Figs 13, 14. LM, acid-cleaned valves, showing striae and raphes.Fig. 15. SEM, external view of a valve showing the grossmorphology.Fig. 16. SEM, internal view of a valve showing the grossmorphology.
Park et al.: Four new Fogedia species 441

Valve interior was flat, and the valve mantle was very
shallow (Figs 16, S30). The raphe slit ran relatively straight
from the central nodule to the helictoglossa. Helictoglossa
were relatively simple, small, and untwisted (Figs 16, S31).
The areolae were internally occluded with thin hymenes thatresembled those in Navicula sensu stricto. Areolae andtransapical striae were evenly spaced. A few areolaeproximal to the central area were round in shape andoblique in orientation to the remaining striae that formedareolae (Figs 16, S30). Fogedia elliptica was easily distin-guished from the other members of the genus by its roundedvalve outline and the absence of a lateral area.
Fogedia lyra Park, Khim, Koh & Witkowski sp. nov.(Figs 17–21, S16–S19, S32–S35)
DESCRIPTION: Valves broadly elliptic or broadly elliptical-lanceo-late to linear-elliptic, with cuneate and finally short protracted,subrostrate apices, 29–40 lm in length, 11–12.5 lm in width. Raphecentral external endings punctiform, slightly expanded. Axial areavery narrow, linear throughout. Lateral area present. Transapicalstriae 12–14 in 10 lm, areolae 26–29 in 10 lm.
HOLOTYPE: Slide 20704, deposited in the Institute of MarineSciences, Coll. A. Witkowski (SZCZ), Szczecin, Poland.
ISOTYPE: Slide KR-GY4-04, deposited in Seoul National Univer-sity, Seoul, Korea.
TYPE LOCALITY: Gyehwa tidal flat (358460N, 1268350E), Korea; typematerial was sampled on 6 August 2003 by Dr J. Ryu.
ETYMOLOGY: Latin lyra¼ lyre; refers to the shape of its lateral area.
DISTRIBUTION: F. lyra had a relative abundance of 3.0% at the typelocality. The species was also observed in samples from sand flats atMinami-Arao (328580N, 1308260E), Ariake Sea, Kyushu, Japan, andHakata Bay (338410N, 1308250E), Kyushu, Japan.
The chloroplasts of Fogedia lyra were the same as in F.coreana (Figs 17, 18). The external apical raphe endings weredifficult to discern. Axial area expanded in the middle andformed very small central area that was lanceolate in shape.Approximately half-way between the valve margin and theraphe sternum, a typical Fogedia lateral area was well-developed (Figs 19, S16–S19). The transapical striae wereparallel in the middle of the valve, they became radiatetowards apices, and finally they became parallel again at theapices. The areolae were easily discerned. In the SEM, theexterior valve face was flat with an abrupt transition to thevalve mantle (Fig. 20). Axial area was symmetrical, verynarrow, and linear; it widened towards the valve centre intoa small lanceolate central area. The raphe external centralendings were slightly expanded. The apical external rapheendings were unlike other Fogedia except F. giffeniana and F.elliptica, and they were abruptly deflected towards thesecondary sides (Figs 20, S20). They were somehow similarto those of Navicula sensu stricto, yet they were not stronglygeniculate but diagonally straight, and therefore differenti-ated from Navicula spp. The transapical striae werecomposed of apically elongate S-shaped areolae, which wereevenly distributed. The areolae located close to the centralarea were rather different; they changed orientation from theapical plane to the transapical plane (Figs 20, S21). Therewere two such areolae at the valve primary side and three atthe valve secondary side. The number of areolae were 26–29in 10 lm. Approximately half-way between the valve marginand the raphe sternum there was a distinct lyra-shaped
Figs 17–21. Fogedia lyra sp. nov. Scale bars ¼ 10 lm.Figs 17, 18. LM, valve surface, untreated cell. Note the plastidand storage droplets.Fig 19. LM, acid-cleaned valve, showing striae and raphes.Fig. 20. SEM, external view of a valve showing the grossmorphology.Fig. 21. SEM, internal view of a valve showing the grossmorphology.
442 Phycologia, Vol. 52 (5), 2013

lateral area. This area was generally depressed below thevalve face surface.
The valve interior was flat with a very shallow mantle (Fig.21). The raphe slit was markedly curved near the centralpores; however, the internal slit ran relatively straight fromthe central nodule to the helictoglossa, not twisted (unlike inNavicula sensu stricto; Figs 21, S34, S35). The striae formingareolae were situated internally in distinct yet shallow grovesformed by transapical ribs. The striae were crossed by adistinct lyra-shaped elevation, which corresponded to adepression at the valve face exterior.
Fogedia giffeniana var. giffeniana, F. giffeniana var.yemenitica, and F. christensenii had similar valve outlines.Of these, F. giffeniana var. giffeniana occurred in the westernpart of the Indian Ocean (Oman, Kenya, Tanzania, SouthAfrica) and in the Mediterranean (Witkowski et al. 2000); F.giffeniana var. yemenitica was only reported from thewestern part of the Indian Ocean (Gulf of Aden); F.christensenii was reported only from Fiji, Pacific Ocean.These three species were easily distinguished from F. lyrabecause the latter had smaller sized cells as well as higherstria and areola densities.
DISCUSSION
All eight species of Fogedia have a linear-lanceolate, rarelyelliptic, valve shape as well as a straight and simple slit-likeraphe on the valve interior. Other common features includethe development of the lateral area and simple distal rapheends on the valve exterior. However, F. heterovalvata(Simonsen) Witkowski, Metzeltin & Lange-Bertalot wasdescribed as lacking a lateral area (Simonsen 1959,Witkowski et al. 2000), and F. giffeniana was described ashaving deflected external raphe endings. The eight Fogediaspecies have a wide geographic distribution in the oceans;however, a majority of these species have been reported asthe only member of the genus at the locations where theyoccur. Furthermore, there have been no records of theFogedia predominance in diatom assemblages. The onlyFogedia species previously observed at relatively highabundance was F. geisslerae, which was found in the NorthSea at Rm Island, Denmark (Witkowski, unpublishedresults). Even so, the abundance of F. geisslerae in theNorth Sea does not parallel the dominance of Fogediaspecies in our Korean study area.
In addition to four new species, this study provides thefirst report of Fogedia in the Northwest Pacific Ocean, and itis the first report of Fogedia dominance in the marine littoralenvironment. It is especially noteworthy that the number ofFogedia species discovered in one locality during the presentstudy (n ¼ 4) is equal to the half of the total number of theFogedia species previously reported from the entire world (n¼ 8).
We also observed a degree of variation in the morphologyof the Korean species (and Yellow Sea material, in general).As illustrated by SEM, these differences include variation inthe external apical raphe endings as well as the presence orabsence of lateral area. While the majority of the newlydescribed Fogedia species have simple external apical raphe
endings typical for the genus, F. elliptica and F. lyra haveexternal terminal raphe endings which are bent to thesecondary sides as in F. giffeniana. Additionally, terminalfissures are not strongly geniculate but diagonally straightwith abrupt deflections; thus, they are different from those oftypical Navicula sensu stricto. It was also shown that, asearlier implicated in the case of F. heterovalvata, Fogediaspecies do not necessarily possess a lateral area on the valveexterior; three out of the four new Fogedia species describedhere lack the lateral area. It is also noteworthy that F.coreana has a zoned distribution of areolae on valve facewith a sparse density in the middle of the valve; this gives animpression that Fogedia has a continuum of the lateral areadevelopment from the absence of the lateral area to a zoneddistribution of the areolae to finally the presence of a lateralarea. Interestingly, the structure of the lateral area itself alsovaries among species in terms of SEM morphologicalcharacteristics (cf. lateral area, table 1 in Witkowski et al.1997). Furthermore, certain species of Navicula sensu strictohave been reported to have lateral areas (Mann 1999;Kulikovsky et al. 2012). Thus the presence or absence of alateral area is not a single decisive criterion, and other majorcharacteristics of Fogedia are needed to separate the twogenera: viz. ‘simple external raphe endings’ is emended hereto ‘simple or deflected straight raphe endings’. Othermorphological characteristics, such as a flat valve face anda shallow valve mantle, may be valuable in the description offurther new Fogedia species.
We reported the first extensive observations on thechloroplast structure, which was possible because of thehigh abundance of Fogedia in our study area. Thechloroplasts are consistently positioned in the cell; the twogirdle-appressed plastids are located along the valve margin.The chloroplasts are slightly thickened and undulated in themiddle, and typically there are four conspicuous granulesthat accompany the plastids. Similar chloroplast structureswere observed for F. geisslerae from Bremerhaven, Germany(personal observation). Each chloroplast contains a singlerod-like pyrenoid, which was reported here for the first time.As shown by Cox (1987) and Cox & Williams (2006),chloroplasts provide very strong evidence when determiningphylogenetic relationships, and a chloroplast comparisonsuggests a close phylogenetic relationship for Fogedia,Hippodonta, and Navicula sensu stricto. Based on thechloroplast information, Fogedia may be included in thefamily of Naviculaceae, together with Hippodonta and theexamples of Haslea Simonsen (Simonsen 1974), Pseudogom-phonema Medlin (Medlin & Round 1986), Seminavis D.G.Mann (Round et al. 1990; Danielidis & Mann 2002), andTrachyneis Cleve (e.g. Round et al. 1990; Cox & Williams2006). The separation of Hippodonta from Navicula wassubject to debate (Cox 1999), but recent molecular datapublished by Bruder & Medlin (2008) show that Hippodontaand Navicula are sister taxa. The relationship betweenFogedia and Hippodonta is also supported by the raphesystem; Hippodonta has a simple slit similar to Fogedia(Lange-Bertalot et al. 1996; Witkowski et al. 1997; Cox 1999;Lange-Bertalot 2001). However, Hippodonta differs mor-phologically from Fogedia with respect to the mantle and theapices of the valves. Electron microscopic examinationshows that Hippodonta has a high girdle that is composed
Park et al.: Four new Fogedia species 443
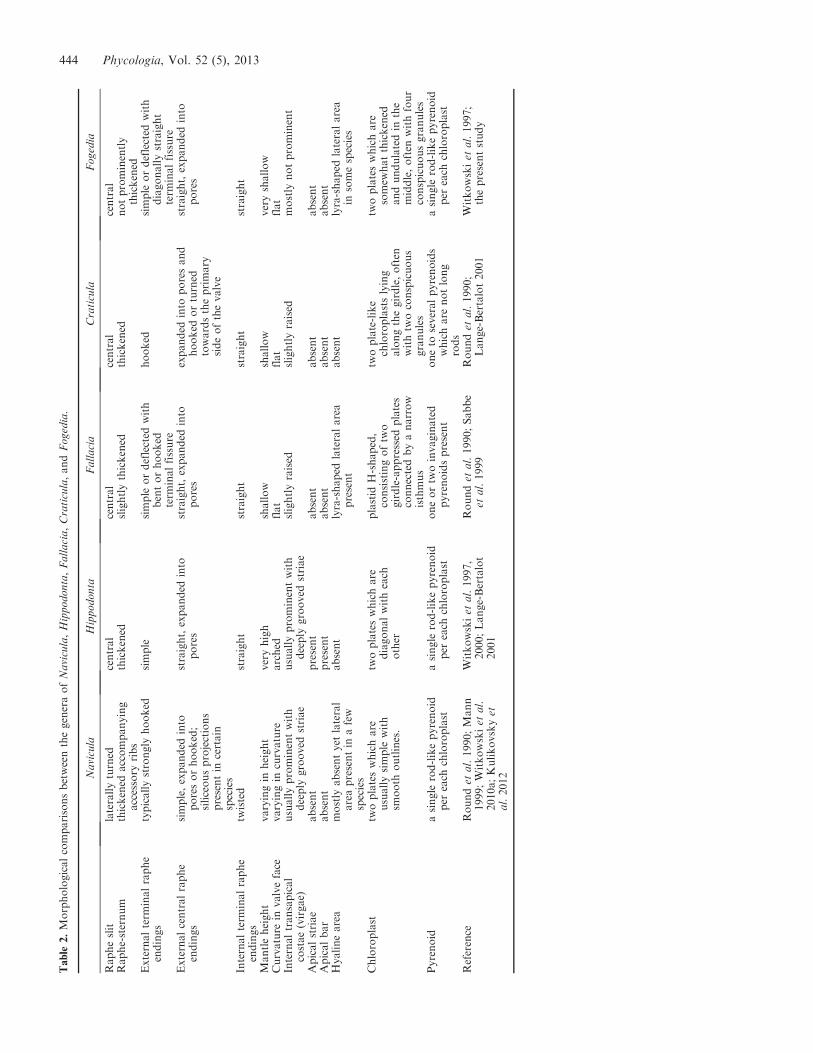
Table
2.MorphologicalcomparisonsbetweenthegeneraofNavicula,Hippodonta,Fallacia,Craticula,andFogedia.
Navicula
Hippodonta
Fallacia
Craticula
Fogedia
Rapheslit
laterallyturned
central
central
central
central
Raphe-sternum
thickened
accompanying
accessory
ribs
thickened
slightlythickened
thickened
notprominently
thickened
Externalterm
inalraphe
endings
typicallystrongly
hooked
simple
simple
ordeflected
with
bentorhooked
term
inalfissure
hooked
simple
ordeflected
with
diagonallystraight
term
inalfissure
Externalcentralraphe
endings
simple,expanded
into
poresorhooked;
siliceousprojections
presentin
certain
species
straight,expanded
into
pores
straight,expanded
into
pores
expanded
into
poresand
hooked
orturned
towardstheprimary
sideofthevalve
straight,expanded
into
pores
Internalterm
inalraphe
endings
twisted
straight
straight
straight
straight
Mantleheight
varyingin
height
veryhigh
shallow
shallow
veryshallow
Curvature
invalveface
varyingin
curvature
arched
flat
flat
flat
Internaltransapical
costae(virgae)
usuallyprominentwith
deeply
grooved
striae
usuallyprominentwith
deeply
grooved
striae
slightlyraised
slightlyraised
mostly
notprominent
Apicalstriae
absent
present
absent
absent
absent
Apicalbar
absent
present
absent
absent
absent
Hyalinearea
mostly
absentyet
lateral
areapresentin
afew
species
absent
lyra-shaped
lateralarea
present
absent
lyra-shaped
lateralarea
insomespecies
Chloroplast
twoplateswhichare
usuallysimple
with
smooth
outlines.
twoplateswhichare
diagonalwitheach
other
plastid
H-shaped,
consistingoftw
ogirdle-appressed
plates
connectedbyanarrow
isthmus
twoplate-like
chloroplastslying
alongthegirdle,often
withtw
oconspicuous
granules
twoplateswhichare
somew
hatthickened
andundulatedin
the
middle,often
withfour
conspicuousgranules
Pyrenoid
asingle
rod-likepyrenoid
per
each
chloroplast
asingle
rod-likepyrenoid
per
each
chloroplast
oneortw
oinvaginated
pyrenoidspresent
oneto
severalpyrenoids
whichare
notlong
rods
asingle
rod-likepyrenoid
per
each
chloroplast
Reference
Roundet
al.1990;Mann
1999;Witkowskiet
al.
2010a;Kulikovskyet
al.2012
Witkowskiet
al.1997,
2000;Lange-Bertalot
2001
Roundet
al.1990;Sabbe
etal.1999
Roundet
al.1990;
Lange-Bertalot2001
Witkowskiet
al.1997;
thepresentstudy
444 Phycologia, Vol. 52 (5), 2013

of very broad bands; whereas, Fogedia has a low girdle thatis composed of narrow open bands. In addition Hippodontahas apical striae at both apices, which are missing in Fogedia(Lange-Bertalot et al. 1996; Witkowski et al. 1997; Lange-Bertalot 2001). Unique for Hippodonta is the presence of asiliceous apical bar at both valve ends, which can be seen atthe valve interior in SEM (cf. Lange-Bertalot et al. 1996;Lange-Bertalot 2001). Therefore, Fogedia is as an indepen-dent genus that is closely related to Navicula sensu stricto andHippodonta.
There are other naviculoid genera that are comparable toFogedia, e.g. Fallacia has similar valve outlines and lateralareas, particularly when using LM. However, the chloroplastof Fallacia is H-shaped and consists of two girdle-appressedplates that are connected by a narrow isthmus. The pyrenoidmorphology also differs; Fallacia has one or two invaginatedpyrenoids. Craticula has two plate-like chloroplasts that areoften accompanied by two conspicuous droplets (illustratedin Round et al. 1990); however, Craticula has one to severalpyrenoids that are not long rods as occurs in Fogedia andNavicula. The overall morphology of Navicula, Hippodonta,Fallacia, Craticula, and Fogedia is summarized in Table 2.
Our study increased the number of Fogedia species from 8to 12 taxa, and it expanded the worldwide distribution toinclude the northwest Pacific Ocean (Fig. 22). Only F.finmarchica (Cleve & Grunow) Witkowski, Metzeltin &Lange-Bertalot appears to be cosmopolitan; the otherknown species have a wider geographical distribution thanour study area (Fig. 22). Our newly described species mayplay important ecological roles as indicated by the domi-nance of several of these species in diatom assemblages.
ACKNOWLEDGEMENTS
This work was supported by National Research Foundation
of Korea Grant (KRF-2006-612-C00040 & NRF-
2013R1A1A2A10011073) and by Polish Ministry of Science
and Higher Education (N306 468538). The authors are
grateful to Prof. David G. Mann for his advice regarding the
cytological characteristics of the genus Fogedia, to Dr
Ohtsuka for his kind help in taking SEM photos, to
Manfred Ruppel for SEM operation, and to anonymous
reviewers and editorial staffs for their valuable suggestions
and comments.
SUPPLEMENTARY DATA
Supplementary data associated with this article can be found
online at http://dx.doi.org/10.2216/12–120.1.s1.
REFERENCES
BRUDER K. & MEDLIN L.K. 2008. Morphological and molecularinvestigations of naviculoid diatoms. III. Hippodonta andNavicula s. s. Diatom Research 23: 331–347.
CLEVE P.T. & GRUNOW A. 1880. Beitrage zur kenntniss derarctischen Diatomeen. Kongliga Svenska vetenskaps-akademienshandlingar 17: 1–121.
COX E.J. 1987. Placoneis Mereschkowsky: the re-evaluation of adiatom genus originally characterized by its chloroplast type.Diatom Research 2: 145–157.
Fig. 22. Worldwide distribution of the Fogedia species.
Park et al.: Four new Fogedia species 445

COX E.J. 1999. Studies on the diatom genus Navicula Bory. VIII.Variation in valve morphology in relation to the generic diagnosisbased on Navicula tripunctata (O.F. Muller) Bory. DiatomResearch 14: 207–237.
COX E.J. & WILLIAMS D.M. 2006. Systematics of naviculoid diatoms(Bacillariophyta): a preliminary analysis of protoplast andfrustule characters for family and order level classification.Systematics and Biodiversity 4: 385–399.
DANIELIDIS D.B. & MANN D.G. 2002. The systematic of Seminavis(Bacillariophyta): the lost identities of Amphora angusta, A.ventricosa and A. macilenta. European Journal of Phycology 37:429–448.
FOGED N. 1973. Diatoms from southwest Greenland. Meddelelserom Grnland 194: 1–84.
FOGED N. 1975. Some littoral diatoms from the coast of Tanzania.Biblotheca Phycologia 16: 1–127.
FOGED N. 1985. Diatoms in Kos and Kalymnos, two Greek islandsin the Aegean. Biblotheca Diatomologica 10: 1–105.
FOGED N. 1987. Diatoms from Viti Levu, Fiji Islands. BiblothecaDiatomologica 14: 1–128.
GLIGORA M., KRALJ K., PLENKOVIC-MORAJ A., HINZ F., ACS E.,GRIGORSZKY I., COCQUYT C. & VAND DE VIJVER B. 2009.Observations on the diatom Navicula hedinii Hustedt (Bacillar-iophyceae) and its transfer to a new genus Envekadea Van deVijver et al. gen. nov. European Journal of Phycology 44: 123–138.
HUSTEDT F. 1961–1966. Die Kieselalgen Deutschlands, Osterreichsund der Schweiz unter Berucksichtigung der ubrigen LanderEuropas sowie der argrenzenden Meeresgebiete. In Kryptogamen-Flora von Deutschland, Osterreich und der Schweiz 7(3)(Raben-horst L., editor), 1–815. Akademische Verlagsgesellschasft,Leipzig.
JOHANSEN J.R. & SRAY J.C. 1998. Microcostatus gen. nov., a newaerophilic diatom genus based on Navicula krasskei Hustedt.Diatom Research 13: 93–101.
KOH C.H., RYU J. & KHIM J.S. 2010. The Saemangeum: history andcontroversy. Journal of the Korean Society for Marine Environ-mental Engineering 13: 327–334.
KULIKOVSKY M.S., LANGE-BERTALOT H., METZELTIN D., &WITKOWSKI
A. 2012. Lake Baikal: hotspot of endemic diatoms I. IconographiaDiatomologica 23: 7–608.
LANGE-BERTALOT H. 1997. Frankophila, Mayamaea und Fistulifera:drei neue Gattungen der Klasse Bacillariophyceae. Archiv furProtistenkunde 148: 65–76.
LANGE-BERTALOT H. 2000. Transfer to the generic rank of DecussataPatrick as a subgenus of Navicula Bory. In IconographiaDiatomologica. Annotated Diatom Micrographs. Vol. 9. Phytoge-ography-Diversity-Taxonomy (Ed. by H. Lange-Bertalot), pp.670–673. Koeltz Scientific Books, Konigstein.
LANGE-BERTALOT H. 2001. Navicula sensu stricto, 10 generaseparated from Navicula sensu lato, Frustulia. Diatoms of Europe2: 1–526.
LANGE-BERTALOT H. & GENKAL S.I. 1999. Diatomeen aus Siberien.I. Insel im Arktischen Ozean (Yugorsky-Shar Strait). InIconographia Diatomologica. Annotated Diatom Micrographs.Vol.6. Phytogeography-Diversity-Taxonomy (Ed. by H. Lange-Bertalot), pp. 1–303, 74 pl. Koeltz Scientific Books, Konigstein.
LANGE-BERTALOT H. & METZELTIN D. 1996. Indicators of oligo-trophy - 800 taxa representative of three ecologically distinct laketypes, carbonate buffered - Oligodystrophic - weakly buffered softwater. In Iconographia Diatomologica. Annotated Diatom Micro-graphs. Vol. 2. Ecology, Diversity, Taxonomy (Ed. by H. Lange-Bertalot), pp. 1–390. Koeltz Scientific Books, Konigstein.
LANGE-BERTALOT H., METZELTIN D. & WITKOWSKI A. 1996. Hippo-donta gen. nov. Umschreibung und Begrundung einer neuen
Gattung der Naviculaceae. Iconographia Diatomologica 4: 247–275.
LIU Y., KOCIOLEK J.P., FAN Y. &WANG Q. 2012. Pseudofallacia gen.nov., a new freshwater diatom (Bacillariophyceae) genus based onNavicula occulta Krasske. Phycologia 51: 620–626.
MANN D.G. 1999. The species concept in diatoms. Phycologia 38:437–495.
MEDLIN L.K. & ROUND F.E. 1986. Taxonomic studies of marinegomphonemoid diatoms. Diatom Research 1: 205–225.
MOSER G., LANGE-BERTALOT H. & METZELTIN D. 1998. Insel derEndemiten. Geobotanisches Phinomen Neukaledonien. Bibliothe-ca Diatomologica 38: 1–464.
OH S.H. & KOH C.H. 1995. Distribution of diatoms in the surficialsediments of the Mangyung-Dongjin tidal flat, west coast ofKorea (Eastern Yellow Sea). Marine Biology 122: 487–496.
RIVERA P.R. 1968. Sinopsis de las diatomeas de la Bahia DeComception. Gayana Botanica 18: 1–11.
ROUND F.E. 1953. An investigation of two benthic algal commu-nities in Malham Tan, Yorkshire. Journal of Ecology 41: 174–197.
ROUND F.E., CRAWFORD R.M. & MANN D.G. 1990. The diatoms,biology and morphology of the genera. Cambridge UniversityPress, Cambridge, 747 pp.
RYU J., KHIM J.S., CHOI J.W., SHIN H.C., AN S., PARK J., KANG D.,LEE C.H. & KOH C.H. 2011. Environmentally associated spatialchanges of a macrozoobenthic community in the Saemangeumtidal flat, Korea. Journal of Sea Research 65: 390–400.
SABBE K., VYVERMAN W. & MUYLAERT K. 1999. New and little-known Fallacia species (Bacillariophyta) from brackish andmarine intertidal sandy sediments in Northwest Europe andNorth America. Phycologia 38: 8–22.
SALAH M.M. 1955. Some new diatoms from Blakeney Point(Norfolk). Hydrobiologia 7: 88–102.
SIMONSEN R. 1959. Neue Diatomeen aus der Ostsee. I. KielerMeeresforschungen 15: 74–83.
SIMONSEN R. 1974. The diatom plankton of the Indian OceanExpedition of R/V ‘Meteor’ 1964–1965. ‘Meteor’ Forschungser-gebnisse, Rehe D 19: 1–107.
SPAULDING S.A. & STOERMER E.F. 1997. Taxonomy and distributionof the genus Muelleria Frenguelli. Diatom Research 12: 95–113.
VAN DEN HEUVEL H.M. & PRUD’HOMME VAN REINE W.F. 1985.Marine, mainly benthic diatoms of the west coast of the Island LaPalma (Canary Islands). Vieraea 14: 11–31.
WITKOWSKI A. 1994. Recent and fossil diatom flora of the Gulf ofGdansk, Southern Baltic Sea. Biblotheca Diatomologica 28: 1–313.
WITKOWSKI A., METZELTIN D., LANGE-BERTALOT H. & BAFANA G.1997. Fogedia gen. nov. (Bacillariophyceae), a new naviculoidgenus from the marine littoral. Nova Hedwigia 65: 79–98.
WITKOWSKI A., LANGE-BERTALOT H. & METZELTIN D. 2000. Diatomflora of Marine Coasts I. Iconographia Diatomologica 7: 1–925.
WITKOWSKI A., KULIKOVSKY M., NEVROVA E., LANGE-BERTALOT H.& GOGOREV R. 2010a. The genus Navicula in ancient basins. I.Two novelties from the Black Sea. Plant Ecology and Evolution143: 307–317.
WITKOWSKI A., LANGE-BERTALOT H., KOCIOLEK J.P., KULIKOWSKIY
M., BAK M. & RUPPEL M. 2010b. Diatom flora of San Franciscobay and vicinity. II. Fogedia krammeri sp. nov. Polish BotanicalJournal 55: 49–53.
Received 19 December 2012; accepted 19 June 2013Associate Editor: Andrew Alverson
446 Phycologia, Vol. 52 (5), 2013




















