
Altered lipid profile and changes in uroplakin properties of rat urothelial plasma membrane with...
8
TECHBOOKS Journal: MCBI MS Code: 106R1 PIPS No: DO00004505 DISK 25-11-2004 17:2 Pages: 7 Molecular and Cellular Biochemistry xxx: 1–7, 2004. c ❣ 2004 Kluwer Academic Publishers. Printed in the Netherlands. Altered lipid profile and changes in uroplakin properties of rat urothelial plasma membrane with diets of different lipid composition 1 2 3 Guillermina A. Bongiovanni, Aldo R. Eynard and Reyna O. Calder´ on 4 Instituto de Biolog´ ıa Celular, I a C´ atedra de Biolog´ ıa Celular, Histolog´ ıa y Embriolog´ ıa, Facultad de Ciencias M´ edicas, Universidad Nacional de C ´ ordoba, C´ ordoba, Argentina 5 6 Received; accepted 7 Abstract 8 Rigid plaques containing protein particles in plasma membrane build on the apical surface of the mammalian urothelium. We have previously shown that dietary fats modified the fatty acid profile as well as the fluorescence anisotropy of rat urothelial plasma membranes. In this study, we have further examined the proportion of phosphatidylcholine, phosphatidylethanolamine, cerebrosides, sulfatides and cholesterol in detergent resistant (DRM) and soluble (DSM) plasma membrane fractions as well as the properties of the particles. Four groups of weaned rats were fed for 12 weeks on a commercial diet (control), or on a formula containing 5% (w/w) of corn oil, fish oil or olein. The control DRM behaved as a distinctive domain since it was enriched in cholesterol and glycosphingolipids. DSM showed higher levels of phosphatidylcholine and phosphatidylethanolamine with respect to DRM. On the other hand, the lipid distributions were affected by the diets. Homogeneous lipid distributions between DSM and DRM were found in olein membranes, suggesting a decreased potential formation of lipid domains. In addition, properties of the uroplakins were altered by dietary treatments. Thus, uroplakins (UP) Ia, Ib, II and III observed by SDS-PAGE, were in lower proportions (mainly olein) than in controls. Moreover, a higher proportion of UPIII was cross-linked to UPIII and UPIb in olein treatment than in control. Meanwhile, only cross-linking to UPIII or UPIb was altered in corn and fish diets, respectively. These results suggest a role of the lipids in the establishment of the uroplakin interactions. Thus, specific dietary fats may have important functional implications. (Mol Cell Biochem xxx: 1–7, 2004) 9 10 11 12 13 14 15 16 17 18 19 20 21 22 Key words: asymmetric unit membrane, detergent-resistant membrane, diet, dietary fats, lipid domains, uroplakins, urothelial- particles, urothelium, permeability barrier 23 24 Introduction 25 The apical surface of luminal “umbrella” cells of the mam- 26 malian transitional epithelium (urothelium) elaborates a 27 thick, highly specialized plasma membrane called the asym- 28 metric unit membrane (AUM) [1]. The luminal plasma mem- 29 brane is built up of rigid appearing plaques that consist of 30 a two-dimensional crystalline conformation of hexagonally 31 arranged 16-nm glycoproteic particles [2]. Morphologically, 32 this unusual array gives rise to an asymmetric appearance 33 with a thick luminal leaflet and a thinner cytoplasmic leaflet 34 Address for offprints: Reyna Olga Calderon, Instituto de Biolog´ ıa Celular, I a C´ atedra de Biolog´ ıa Celular, Histolog´ ıa y Embriolog´ ıa, Facultad de Ciencias M´ edicas, Universidad Nacional de C´ ordoba, CP 5000, CC 220, Ciudad Universitaria, C´ ordoba, Rep ´ ublica Argentina (E-mail: [email protected])] [2, 3]. The two-dimensional particles are formed by four ma- Q1 35 jor proteins named uroplakins (UP) Ia, Ib, II and III [4]. Each 36 particle has a central 6 nm lipid-filled “hole” surrounded by 37 six inverted U-shaped subunits, each consisting of an inner 38 and an outer subdomain connected via a distal joint. The uro- 39 plakin pairs consisting of UPIa/II and UPIb/III are associated 40 with the inner and outer subdomains, respectively [5]. A fifth 41 variety, UPIIIb, very scarce, forms the UPIb-IIIb pair, and are 42 present at a much lower amount [6]. It has been suggested 43 that the urothelial AUM: (a) stabilizes the apical surface, thus 44 preventing urothelial rupture during bladder distention [7], 45 AUTHOR'S PROOFS
Transcript of Altered lipid profile and changes in uroplakin properties of rat urothelial plasma membrane with...

TECHBOOKS Journal: MCBI MS Code: 106R1 PIPS No: DO00004505 DISK 25-11-2004 17:2 Pages: 7
Molecular and Cellular Biochemistry xxx: 1–7, 2004.c�2004 Kluwer Academic Publishers. Printed in the Netherlands.
Altered lipid profile and changes in uroplakinproperties of rat urothelial plasma membranewith diets of different lipid composition
1
2
3
Guillermina A. Bongiovanni, Aldo R. Eynard and Reyna O. Calderon4
Instituto de Biologıa Celular, Ia Catedra de Biologıa Celular, Histologıa y Embriologıa, Facultad de Ciencias Medicas,Universidad Nacional de Cordoba, Cordoba, Argentina
56
Received; accepted7
Abstract8
Rigid plaques containing protein particles in plasma membrane build on the apical surface of the mammalian urothelium. Wehave previously shown that dietary fats modified the fatty acid profile as well as the fluorescence anisotropy of rat urothelialplasma membranes. In this study, we have further examined the proportion of phosphatidylcholine, phosphatidylethanolamine,cerebrosides, sulfatides and cholesterol in detergent resistant (DRM) and soluble (DSM) plasma membrane fractions as well asthe properties of the particles. Four groups of weaned rats were fed for 12 weeks on a commercial diet (control), or on a formulacontaining 5% (w/w) of corn oil, fish oil or olein. The control DRM behaved as a distinctive domain since it was enrichedin cholesterol and glycosphingolipids. DSM showed higher levels of phosphatidylcholine and phosphatidylethanolamine withrespect to DRM. On the other hand, the lipid distributions were affected by the diets. Homogeneous lipid distributions betweenDSM and DRM were found in olein membranes, suggesting a decreased potential formation of lipid domains. In addition,properties of the uroplakins were altered by dietary treatments. Thus, uroplakins (UP) Ia, Ib, II and III observed by SDS-PAGE,were in lower proportions (mainly olein) than in controls. Moreover, a higher proportion of UPIII was cross-linked to UPIIIand UPIb in olein treatment than in control. Meanwhile, only cross-linking to UPIII or UPIb was altered in corn and fish diets,respectively. These results suggest a role of the lipids in the establishment of the uroplakin interactions. Thus, specific dietaryfats may have important functional implications. (Mol Cell Biochem xxx: 1–7, 2004)
910111213141516171819202122
Key words: asymmetric unit membrane, detergent-resistant membrane, diet, dietary fats, lipid domains, uroplakins, urothelial-particles, urothelium, permeability barrier
2324
Introduction25
The apical surface of luminal “umbrella” cells of the mam-26malian transitional epithelium (urothelium) elaborates a27thick, highly specialized plasma membrane called the asym-28metric unit membrane (AUM) [1]. The luminal plasma mem-29brane is built up of rigid appearing plaques that consist of30a two-dimensional crystalline conformation of hexagonally31arranged 16-nm glycoproteic particles [2]. Morphologically,32this unusual array gives rise to an asymmetric appearance33with a thick luminal leaflet and a thinner cytoplasmic leaflet34
Address for offprints: Reyna Olga Calderon, Instituto de Biologıa Celular, Ia Catedra de Biologıa Celular, Histologıa y Embriologıa, Facultad de CienciasMedicas, Universidad Nacional de Cordoba, CP 5000, CC 220, Ciudad Universitaria, Cordoba, Republica Argentina (E-mail: [email protected])]
[2, 3]. The two-dimensional particles are formed by four ma-
Q1
35jor proteins named uroplakins (UP) Ia, Ib, II and III [4]. Each 36particle has a central 6 nm lipid-filled “hole” surrounded by 37six inverted U-shaped subunits, each consisting of an inner 38and an outer subdomain connected via a distal joint. The uro- 39plakin pairs consisting of UPIa/II and UPIb/III are associated 40with the inner and outer subdomains, respectively [5]. A fifth 41variety, UPIIIb, very scarce, forms the UPIb-IIIb pair, and are 42present at a much lower amount [6]. It has been suggested 43that the urothelial AUM: (a) stabilizes the apical surface, thus 44preventing urothelial rupture during bladder distention [7], 45
AUTHOR'S PROOFS

2
(b) regulates the apical surface area by their reversible re-46trieval from, and insertion into, the luminal plasma membrane47of the superficial urothelial cells [8, 9], and (c) is a major com-48ponent of the permeability barrier of mammals urothelium49[5, 10, 11]. Thus, it is tempting to speculate that uroplakins,50in conjunction with specific lipid components, can contribute51to the bladder barrier function, and be largely responsible52for the healthy on diseased conditions of urinary tracts. Fur-53thermore, the dynamic state of the lipid bilayer of cellular54membranes allows the manipulation of the lipid content of55the membrane by dietary means. These facts are worth not-56ing, since specific fat intake is related to an increased risk of57colorectal cancer and of urinary tract tumorigenesis and may58play a role in metastasis development [12, 13, 14, 15]. In this59regard, it has been shown that an inadequate intake of essen-60tial fatty acids (EFAs) for long periods favors the abnormal61proliferation and tumorigenesis of rat urothelium [16]. Addi-62tionally, we have shown that urothelial plasma membranes,63isolated from rats fed in olein, corn and fish oil rich diets,64significantly changed the fatty acid composition and its lipid65dynamic properties [17].66
As an approach to address the structural role of the lipids in67urothelial plasma membrane, we analyzed the effect of olein,68corn and fish oil rich diets on the lipid compositions of two69detergent-resistant and soluble rat urothelial plasma mem-70brane fractions and on uroplakins associations. It is worth71mentioning that the formulae used to feed the animals have72been characterized as protective (fish) or promoter (olein) di-73ets for the urothelial in situ carcinoma development [15] as74well as for the induction of morphological and biophysical75alterations [18, 19, 17].76
Materials and methods77
Animals and diets78
Following the same dietary protocol as published elsewhere79[17], Wistar rats were weaned at 25 days and then divided80into four groups of six males and six females. Three groups81were fed ad libitum for 12 weeks with semisynthetic formu-82lae containing (% w/w) 20.0 casein, 50.0 sucrose, 20.0 corn83starch, 3.5 salt mixture, 1.0 vitamins mixture, 0.3 methionine,840.1 choline and 5% of one of the following lipid sources: corn85oil, enriched in 18:2 n-6 (corn diet); fish oil, enriched in 18:386n-3 (fish diet); refined olein, enriched in 18:1 n-9 (olein diet)87[20]. One group was fed on commercial animal diet (Cargill88Co) (control diet).89
Animal handling and experimental procedures were car-90ried out in agreement with the Guidelines for the Care and91Use of Laboratory Animals approved by the Animal Care92Committee from National University of Cordoba, Argentina.93
Luminal plasma membrane purification 94
The urothelial cells were obtained using the procedure modi- 95fied by Calderon et al. [21] for exfoliating the luminal surface 96of bladders. The purification of luminal plasma membranes 97was carried out on a discontinuous sucrose gradient according 98to Wu et al. [4]. The 2 and 3 fractions (0.75/1.1 and 1.1/1.3 M 99sucrose, respectively) were added and called total membrane 100(TM) and were then treated with 2% N-lauroyl-sarcosine 101for 20 min. A pellet containing the detergent resistant mem- 102brane fraction (DRM) (enriched in uroplakins), and a super- 103natant enriched in the detergent soluble membrane fraction 104(DSM) were obtained by centrifugation at 12,000 × g per 10530 min. 106
Protein determination 107
Protein was determined by the method of Lowry et al. [22]. 108Samples were measured in the presence of 2% N-lauroyl- 109sarcosine and 2% SDS (sodium dodecyl sulfate). 110
Lipid determination 111
Cerebrosides (CB), Sulfatides (S), phosphatidylcholine (PC), 112phosphatidylethanolamine (PE) and cholesterol (CHOL) 113were quantified according to Macala et al. [23] by high per- 114formance thin layer chromatography (HPTLC) on aluminum- 115backed silica sheets (from Merck) and using a Shimadzu 116Chromato Scanner CS930. Lipid standards used were from 117Avanti Polar-Lipid, Inc. The molar ratios were calculated us- 118ing the molar weights of 811.27 (CB), 780.15 (PC), 743.06 119(PE), 909.34 (S), 386.6 and 649 (free and esterified choles- 120terol, respectively) (CHOL). 121
Production of polyclonal antibody to bovine uroplakins 122
Since the uroplakins are highly conserved [24], bovine uro- 123plakins were used to prepare rabbit antibody against uro- 124plakins. The luminal plasma membrane fraction was isolated 125from 25 bovine urinary bladders, and these were then treated 126with 2% N-lauroyl-sarcosine (see below). The pellet contain- 127ing the uroplakins (confirmed by SDS-PAGE) was solubilized 128using 2% octyl glucoside in phosphate buffer saline (PBS) for 12920 min at room temperature and dialyzed in PBS overnight 130at 4 ◦C to remove the detergent. The uroplakins were cross- 131linked to chicken egg albumin by incubation in 3% glutaralde- 132hyde and were then used to immunize four rabbits. Serums 133from two rabbits exhibited reactivity against UPIII (rat and 134bovine) after four boosters, as determined by Western blot- 135ting, and were called anti-UPIII antibody. A weak reactivity 136
AUTHOR'S PROOFS

3
against both rat UPIa and UPIb was observed in the other two137rabbits (not used).138
Cross-linking of uroplakins139
The uroplakin forming particles in the DRM (100 µg of pro-140teins/100 µl of HEPES buffer) were cross-linked by EGS141[ethylene glyco-bis(succinimidyl succinate)] following the142procedure described by Wu et al. [25].143
Electrophoretic analyses and immunoblotting144
The DRM containing the particles were solubilized at room145temperature in sample buffer and the proteins were separated146by 14% SDS-PAGE (sodium dodecyl sulfate-polyacrylamide147gel electrophoresis) according to the Laemmli method [26].148Proteins in the gels were stained with Silver stain kit from149BioRad or transferred to nitrocellulose sheet. Immunoblot150analysis was carried out using rabbit anti-UPIII antibody and151a goat anti-rabbit HR-peroxidase conjugated from BioRad.152The bands visualized by Silver nitrate in polyacrylamide gel153and those revealed by immunostaining in nitrocellulose were154quantified by scanning densitometry using a Shimadzu Chro-155mato Scanner CS-930. The range used meant that the areas156were proportional to the protein.157
Statistical analysis158
The Student’s t-test was used for statistical analysis. Values159of p < 0.05 were considered significant.160
Results161
After 12 weeks on the diets, all the animal were healthy and162only a minimum of alopecia in the ventral region was ob-163served for animals in olein diet from approximately the 3rd164to the 7th week of treatment. No macroscopic alterations were165registered in the mucosa of the urinary tract. Using the proce-166dure described above, we were able to purify uroplakins from167urothelium as detergent-resistant membrane fractions (DRM)168for all dietary conditions. Examination of control detergent-169resistant membrane fractions (DRM) by electron microscopy170revealed the presence of a typical lattice of particles forming171hexagonally packed crystalline patches (not shown).172
Effect of dietary fats on membrane lipid composition173
Based on previous data from our laboratory, showing that174olein, corn and fish oil diets induced a conspicuous modifi-175
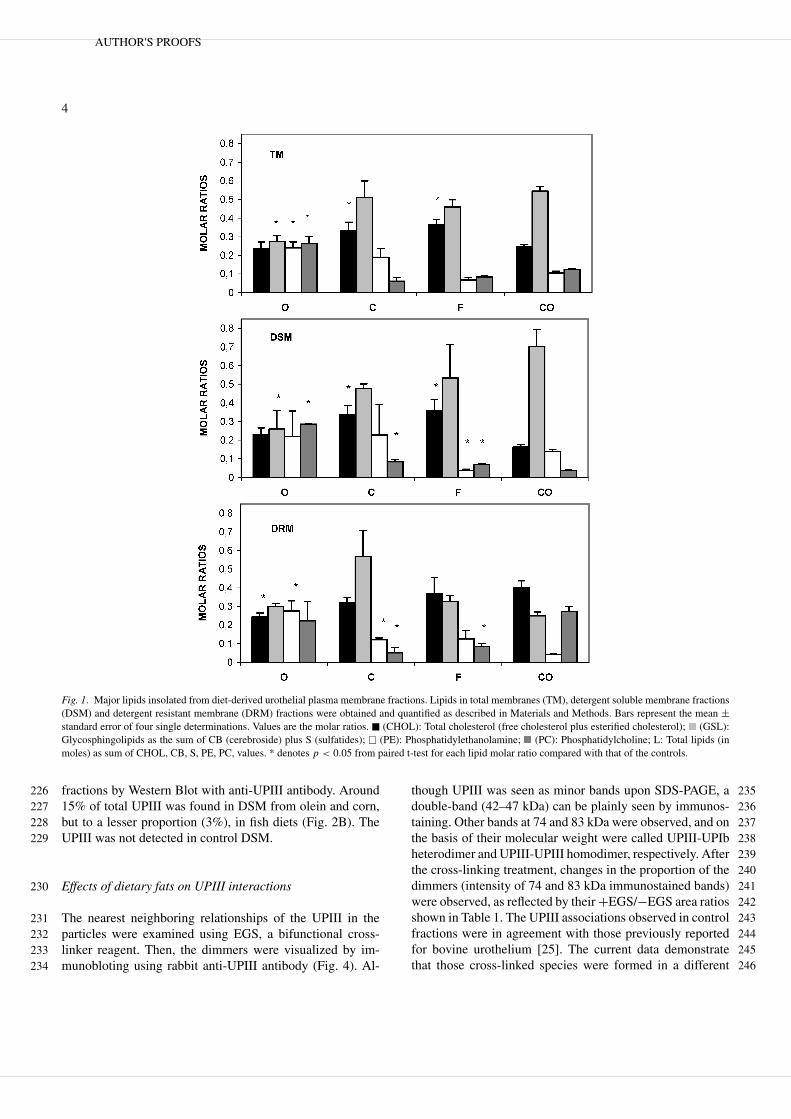
cation in fatty acid composition and also in the fluorescence 176anisotropy of rat luminal urothelial membrane [17], the ma- 177jor lipids in detergent soluble (DSM) and resistant (DRM) 178fractions of urothelial cells from animals fed on these di- 179ets were investigated. The lipid analysis on total membranes 180(TM) showed that all diets were able to alter the propor- 181tions of cholesterol (CHOL), Phosphatidylcholine (PC) phos- 182phatidylethanolamine (PE) and glycosphingolipids (GSL, as 183a sum of sulfatides plus cerebrosides) when compared to 184those of the control (Fig. 1: TM). The ratios of the lipids stud- 185ied were almost equal to unity in olein-derived membranes 186while those of the corn diet were similar to the controls. Inter- 187estingly, similar proportions of PC and CHOL were observed 188in all the fish-derived fractions. 189
The lipid distribution profile obtained after detergent treat- 190ment was not very different for DSM and DRM with respect 191to TM, although some quantitative differences were observed 192with respect to controls (Fig. 1: DSM, DRM). The increases 193of CB and S in DSM and PE in DRM in the olein diet com- 194pared with those in control fractions were the most significant 195changes observed (Fig. 1: DSM and DRM). 196
The DRM from rats fed with control diet had a relative 197enrichment of cholesterol CHOL and S when compared with 198the DSM (40 vs. 16% and 23 vs. 4%, respectively), while PC 199and PE were enriched in DSM with respect to DRM. 200
Effect of dietary fats on detergent solubility of uroplakins 201
The effects of dietary fats on the particle properties in rat 202asymmetrical unit membrane (AUM) were examined. Pro- 203teins from each diet-derived DRM were analyzed by SDS- 204PAGE (Fig. 2A). These fractions were enriched in 15, 27 and 20528 kDa proteins as well as in a double band at 42–47 kDa. 206They were called UPII, UPIa, UPIb and UPIII, respectively, 207according to Wu et al. [25]. Non-characterized bands of 18 208and 30 kDa and of high molecular weight appeared in the 209gel. Although equal amounts of proteins from DRM frac- 210tions, 40 µg, were loaded in the gel, a significant decrease 211in the amount of each uroplakins was found, mainly in olein, 212and to a lesser degree in corn and fish diets, with respect 213to the control (Fig. 2A). The area for each uroplakin related 214to total uroplakin area is show in Fig. 3. Uroplakin-particles 215in DRM from control fractions are composed of uroplakins 216Ia:Ib:II:III in the proportion of 58:34:6.6:1.4, respectively 217(Fig. 3). Although analogous ratios were observed under all 218dietary conditions, some uroplakins showed an altered pro- 219portion in comparison to the controls. Thus, an increase of 220UPIa proportion and a decrease of UPIb proportion were 221recorded for olein diet with respect to controls. On the other 222hand, an increase of UPIa proportion was found in corn diet 223whereas a decrease of UPII proportion was found in fish diet 224with respect to controls (Fig. 3). UPIII was assessed in DSM 225
AUTHOR'S PROOFS
4
Fig. 1. Major lipids insolated from diet-derived urothelial plasma membrane fractions. Lipids in total membranes (TM), detergent soluble membrane fractions(DSM) and detergent resistant membrane (DRM) fractions were obtained and quantified as described in Materials and Methods. Bars represent the mean ±standard error of four single determinations. Values are the molar ratios. � (CHOL): Total cholesterol (free cholesterol plus esterified cholesterol); (GSL):Glycosphingolipids as the sum of CB (cerebroside) plus S (sulfatides); � (PE): Phosphatidylethanolamine; (PC): Phosphatidylcholine; L: Total lipids (inmoles) as sum of CHOL, CB, S, PE, PC, values. * denotes p < 0.05 from paired t-test for each lipid molar ratio compared with that of the controls.
fractions by Western Blot with anti-UPIII antibody. Around22615% of total UPIII was found in DSM from olein and corn,227but to a lesser proportion (3%), in fish diets (Fig. 2B). The228UPIII was not detected in control DSM.229
Effects of dietary fats on UPIII interactions230
The nearest neighboring relationships of the UPIII in the231particles were examined using EGS, a bifunctional cross-232linker reagent. Then, the dimmers were visualized by im-233munobloting using rabbit anti-UPIII antibody (Fig. 4). Al-234
though UPIII was seen as minor bands upon SDS-PAGE, a 235double-band (42–47 kDa) can be plainly seen by immunos- 236taining. Other bands at 74 and 83 kDa were observed, and on 237the basis of their molecular weight were called UPIII-UPIb 238heterodimer and UPIII-UPIII homodimer, respectively. After 239the cross-linking treatment, changes in the proportion of the 240dimmers (intensity of 74 and 83 kDa immunostained bands) 241were observed, as reflected by their +EGS/−EGS area ratios 242shown in Table 1. The UPIII associations observed in control 243fractions were in agreement with those previously reported 244for bovine urothelium [25]. The current data demonstrate 245that those cross-linked species were formed in a different 246
AUTHOR'S PROOFS

5
Fig. 2. Effect of dietary fats on uroplakin profile of rat urothelium. Thedetergent resistant (DRM) and soluble (DSM) membranes were isolatedas described in Materials and Methods. (A) The SDS-solubilized proteinsfrom DRM (40 µg) were subjected to 14% SDS-PAGE and the separatedbands were stained with silver nitrate. O: olein; C: corn; F: fish; CO: control.The position of the Uroplakins II, Ia, Ib and III (15, 27, 28 and 42-47 kDa,respectively) is marked on the right. The position of the BioRad kaleidoscopeprestained molecular weight marker (18.1, 32.8, 41.8, 85.0 and 127 kDa) ismarked on the left; (B) UPIII from DSM (100 µg) was analyzed by WesternBlot with anti-UPIII. C: corn; Mr: prestained molecular weight markers (39and 57 kDa positions are marked on the left); F: fish; O: olein; CO: control.
proportion in olein, corn or fish diets. Thus, a higher pro-247portion of UPIII was cross-linked to UPIb in olein, and to a248lesser degree in corn diet-derived membranes, than in control249diet. The UPIII was cross-linked to UPIII in membranes from250all dietary conditions. However, it was in a smaller propor-251tion in olein and fish diet than controls (Fig. 4 and Table 1).252Consequently, while both homodimer and heterodimer were253markedly altered by olein diet, only heterodimer or homod-254imer was changed by corn or fish diets, respectively.255
Discussion256
Membrane domains in urothelial plasma membrane257
According to lipid segregation within the membranes de-258scribed by Schroeder et al. [27], the presence of at least259two lipid-protein domains in the AUM is supported by a dif-260ferent detergent solubility of the lipids associated with the261uroplakins within the aggregates (DRM), and with those cor-262responding to the bilayer regions far from the aggregates263(DSM).264
Table 1. Proportion of dimers formed by cross-linking in urothelialmembranes
Diet UPIb-UPIII UPIII-UPIII
OLEIN 7.35 ± 0.32a 2.41 ± 0.38a
CORN 2.69 ± 0.48a 6.83 ± 0.27
FISH 0.32 ± 0.04 2.51 ± 0.58a
CONTROL 0.41 ± 0.06 5.96 ± 0.97
Detergent resistant membranes from luminal plasma membranes were iso-lated as indicated in Materials and Methods and treated as in Fig. 4. Theareas of the bands were determined by scanning densitometry. The valuesrepresent the +EGS/−EGS area ratios to bands at 74 kDa (UPIb-UPIII) and83 kDa (UPIII-UPIII). Values are the average of three separate experimentsand were expressed as mean ± S.E.M. aDenotes p < 0.05 compared withcontrol.
Fig. 3. Effect of dietary fats on the relative amounts of uroplakins of raturothelium. The proteins (40 µg) were analyzed as shown in Fig. 2A anduroplakins were quantified by scanning densitometry. Bars represent the ratioarea of each uroplakin band (Ia, Ib, II, or III) compared to total uroplakinarea (UPs) and are the average of four separate experiments ± S.E.M. TheUPIII area was calculated adding the areas of 42 and 47 kDa bands, and UPsvalues were calculated by adding the area of all uroplakin bands. � : olein;
: corn; : fish; �: control. ∗ denotes p < 0.05 for each uroplakin arearatio versus the same ratio from controls using paired t-tests.
Fig. 4. Effect of dietary fats on the UPIII associations. The isolation ofdetergent resistant membrane was performed as indicated in Materials andMethods. Fractions were resuspended in buffer HEPES and then were incu-bated with (+) or without (−) EGS cross-linker. The reaction was stoppedwith sample buffer. The proteins (80 µg) were analyzed by Western blot withanti-UPIII. O: olein; C: corn; F: fish; CO: control. The molecular weights ofthe bands (42–47, 74, 83 and 100 kDa) are marked on the right. The positionof the Sigma prestained molecular weight markers (39, 57, 70 and 92 kDa)is marked on the left.
AUTHOR'S PROOFS

6
In searching for the structural role of the lipid, we modified265the lipid composition of the rat urothelial membranes by three266dietary treatments, and their membrane lipid composition was267determined. The membrane lipid molar fractions without de-268tergent treatment (Fig. 1: TM) indicate that the lipid distri-269butions were differently affected by each diet. Thus, the lipid270proportions from corn-derived membranes were not much271altered relative to those from the control membranes, with272all studied lipids being of a similar proportion in olein mem-273branes. On the other hand, one important feature regarding274the lipid profile of olein membranes was the increased con-275centration of PE, a phospholipid related to an increase of276transmembrane fluidity. Coincidently, the increased propor-277tion of PE, resulting in a decreased PC/PE and CHOL/(PC +278PE) ratios (1.35 vs. 4.89 and 0.43 vs. 0.66, respectively), is279in accordance with a decreased fluorescence anisotropy we280previously reported for the same olein-derived membranes281[17]. Taken together, this may indicate a decreased potential282formation of membrane domains in olein-derived urothelial283plasma membranes.284
The diets altered the lipid proportions in DSM and DRM285(Fig. 1). Thus, lipid distribution in both DSM and DRM frac-286tions from olein membranes repeated the same pattern with or287without detergent treatments. This indicates a homogeneous288lipid distribution between DSM and DRM, instead of definite289domains in olein membranes. In this regard, UPIII was also290distributed between DSM and DRM, revealing that at least291this uroplakin has lost its preferential locations within the292DRM shown in the control membranes (Fig. 2B). On the other293hand, the corn and fish diets affected the respective membrane294fractions in a manner that a preferential, although different295lipid distribution, between DSM and DRM, was observed.296
Uroplakins and different lipid environments297
Although the luminal plasma membrane is built up of rigid298appearing plaques that consist of uroplakin particles, the bulk299of the plaque surface (∼62%) is occupied by lipids [5]. Ad-300ditionally, because all uroplakin are integral membrane pro-301teins, we speculate that the different lipid environments may302regulate properties of the uroplakins embedded in its lat-303tice. In fact, a correlation between altered lipid membrane304profiles by dietary treatment and an altered uroplakin be-305havior was observed. Thus, olein diet induces conspicuous306changes on the hydrophobic membrane core properties, as307well as on the uroplakins when compared with the control308diet. In this regard, the total uroplakins was decreased in DRM309(Fig. 2A). Accordingly, the UPIII was increased on the DSM310(Fig. 2B). Beside, an altered neighboring relationship of the311UPIII (Table 1) was observed.312
Indeed, the amount of cross-linked species altered could313be indicative of changes in the interparticle distance as well314
as between uroplakins within the particles. As was shown 315in an earlier report, urothelium of rats suffering from essen- 316tial fatty acid deficiency, as may be induced by olein diet, 317have an abnormal distribution of the so-called “glycoprotein 318subunits”, a morphological term perhaps describing coarsely 319grouped uroplakins having Concanavalin binding sites [28]. 320
Possible functional implications of altered rat urothelial 321plasma membrane 322
Epidemiological studies pointed out that food and nutrition, 323in particular, dietary fat intake, are important factors for hu- 324man health. Taking into account that luminal plasma mem- 325brane of the superficial urothelial cells is a major compo- 326nent of the permeability barrier of mammals urothelium, it 327is tempting to speculate that the alterations induced by the 328diet, described in this paper, somehow perturb the perme- 329ability barrier function. The fact that major alterations were 330observed into olein-enriched diet increases the likelihood that 331diets enriched in non-essential fats may cause permeability 332barrier defects—a concept that may be associated to certain 333bladder diseases, as interstitial cystitis, cancer and lithiasis 334[29,14,15]. On the other hand, significant differences between 335olein and fish diets, would account for the protecting effect 336described to n-3 fatty acids [30]. However, the clinical impli- 337cations of the present results should be further clarified. 338
Acknowledgments 339
We thank Nelida Ramonda for skillful technical assistance 340and Paul Hobson, a native speaker, for critical reading of 341the manuscript. This investigation was partially supported 342by CONICET, Agencia Cordoba Ciencia, SeCyT-UNC and 343FONCyT, Argentina. 344
References 345
1. Vergara J, Longley W, Robertson JD: A hexagonal arrangement of sub- 346units in membrane of mouse urinary bladder. J Mol Biol 46: 593–596, 3471969 348
2. Hicks RM: The fine structure of the transitional epithelium of the rat 349ureter. J Cell Biol 26: 25–30, 1965 350
3. Caruthers JM, Bonneville MA: The asymmetric unit membrane (AUM) 351structure in the luminal plasma membrane of urothelium: The effect of 352trypsin on the integrity of the plaques. J Ultrastruct Res 71: 288–302, 3531980 354
4. Wu XR, Manabe M, Yu J, Sun TT: Large scale purification and im- 355munolocalization of bovine uroplakins I, II and III, molecular markers 356of urothelial differentiation. J Biol Chem 265: 19170–19179, 1990 357
5. Min G, Zhou G, Matthieu S, Sun TT, Kong XP: Structural basis of 358urothelial permeability barrier function as revealed by Cryo-EM studies 359of the 16 nm uroplakin particle. J Cell Sci 116: 4087–4094, 2003 360
AUTHOR'S PROOFS

7
6. Deng FM, Liang FX, Tu L, Resing KA, Hu P, Supino M, Hu CCA,361Zhou G, Ding M, Kreibich G, Sun TT: Uroplakin IIIb, a urothelial362differentiation marker, dimerizes with uroplakin Ib as an early step of363urothelial plaque assembly. J Cell Biol 159: 685–694, 2002364
7. Staehelin LA, Chlapowski FJ, Bonneville MA: Luminal plasma mem-365brane of the urinary bladder. I Three-dimensional reconstruction from366freeze-etch images. J Cell Biol 53: 73–91, 1972367
8. Minsky BD, Chlapowski FJ: Morphometrics analysis of the transloca-368tion of the luminal membrane between cytoplasm and cell surface of369transitional epithelial cells during the expansion-contraction cycles of370mammalian urinary bladder. J Cell Biol 77: 685–697, 1978371
9. Truschel ST, Ruiz WG, Shulman T, Pilewski J, Sun T-T, Apodaca G:372Primary Uroepithelial Cultures. A model system to analyze umbrella373cell barrier function. J Biol Chem 274: 15020–15029, 1999374
10. Hicks MR: The permeability of rat transitional epithelium. J Cell Biol37528: 21–23, 1966376
11. Hu P, Meyers S, Liang FX, Deng FM, Kachar B, Zeidel ML, Sun TT:377Role of membrane proteins in permeability barrier function: Uroplakin378ablation elevates urothelial permeability. Am J Physiol Renal Physiol379283: F1200–F1207, 2002380
12. Navarro A, Diaz MP, Munoz SE, Lantieri MJ, Eynard AR: Character-381ization of meat consumption and risk of colorectal cancer in Cordoba,382Argentina. Nutrition. 19: 7–10, 2003383
13. Pasqualini ME, Heyd VL, Manzo P, Eynard AR: Association between384E-cadherin expression by human colon, bladder and breast cancer cells385and the 13-HODE:15-HETE ratio. A possible role of their metastatic386potential. Prostaglandins Leukot Essent Fatty Acids. 68: 9–16, 2003387
14. Eynard AR: Is the risk of urinary tract tumorigenesis enhanced by a388marginal chronic essential fatty acid deficiency (EFAD)? Nutrition 14:389211–216, 1998390
15. Cremonezzi DC, Silva RA, Diaz P, Valentich MA, Eynard AR: Dietary391polyunsaturated fatty acids (PUFA) differentially modulate melamine-392induced preneoplastic urothelial proliferation and apoptosis in mice.393Protag Leuk Essent Fatty Acids 64: 151–159, 2001394
16. Monis B, Eynard AR: Abnormal cell proliferation, differentiation and395urothelial tumorigenesis in essential fatty acid deficient (EFAD) rats.396Prog Lip Res 20: 691–703, 1981397
17. Calderon RO, Eynard AR: Fatty acids specifically related to the398anisotropic properties of plasma membrane from rat urothelium.399Biochim Biophys Acta 1483: 174–184, 2000
18. Monis B, Eynard AR, Lis D, Kalinec F: Ultrastructural changes of 400the luminal plasma membrane of the transitional epithelium of the rat 401urinary tract in essential fatty acid deficiency. Experientia 32: 697–699, 4021976 403
19. Kalinec F, Calderon JH, Monis B: Ultrastructural variations of the 404urothelial membrane in essential fatty acid (EFA)-deficient rats. A 405digital densitometry study. J Histochem Cytochem 34: 1639–1644, 4061986 407
20. Eynard AR, Cejas V, Silva RA, Quiroga P, Munoz S: Histopathology of 408essential fatty acid deficient mice. Nutrition 8: 37–40, 1992 409
21. Calderon RO, Glocker M, Eynard AR: Lipids and fatty acids composi- 410tion of different fractions from rat urinary transitional epithelium. Lipids 41133: 1017–1022, 1998 412
22. Lowry OL, Rosebrough NJ, Farr AL, Randal RJ: Protein measurement 413with the folinphenol reagent. J Biol Chem 193: 265–275, 1951 414
23. Macala LJ, Yu RK, Ando S: Analysis of brain lipids by high performance 415thin-layer chromatography and densitometry. J Lip Res 24: 1243–1250, 4161983 417
24. Wu XR, Lin JH, Walz T, Hanr M, Yu J, Aebi U, Sun TT: Mammalian 418uroplakins. J Biol Chem 269: 13716–13724, 1994 419
25. Wu XR, Medina JJ, Sun TT: Selective interaction of UPIa and UPIb, 420two members of the transmembrane 4 superfamily, with distinct single 421transmembrane proteins in differentiated urothelial cells. J Biol Chem 422270: 29752–29759, 1995 423
26. Laemmli UK: Cleavage of structural proteins during the assembly of 424the head of the bacteriophge T4. Nature 227: 680–685, 1970 425
27. Schroeder RJ, Ahmed SN, Zhu Y, London E, Brown DA: Choles- 426terol and sphingolipid enhance the Triton X-100 insolubility of 427glycosylphosphatidylinositol-anchored proteins by promoting the for- 428mation of detergent-insoluble ordered membrane domains. J Biol Chem 429273: 1150–1157, 1998 430
28. Eynard AR, Monis B: Concanavalin A sites in urothelium and milk fat 431globules of essential fatty acid (EFA) deficient rats. J Submicros Cytol 432l5: 375–82, l983 433
29. Lavelle J, Meyers S, Ramage R, Bastacky S, Doty D, Apodaca G, Zeidel 434ML: Bladder permeability barrier: Recovery from selective injury of 435surface epithelial cells. Am J Physiol Renal Physiol 283: F242–F253, 4362002 437
30. Rose DP, Connolly JM: Omega-3 fatty acids as cancer chemopreventive 438agents. Pharmacol. Ther. 83: 217–244, 1999 439
AUTHOR'S PROOFS

Query
Q1. Publisher: Please provide Received and accepted dates.
AUTHOR'S PROOFS




















