
Hot spots of retroviral integration in human CD34+ hematopoietic cells
Upload
independentCategory
view
1download
0
All-trans-retinoic acid up-regulates CD38but not c-Kit antigens on human marrow CD34þ cellswithout recruitment into cell cycle
OLIVIER HERAULT,1,2 JORGE DOMENECH,1,2 MICHEL DEGENNE,2 JEAN-LO UIS BREMO ND,2 LUC SENS EBE,4
MARIE-CHRISTINE BERNARD,1 CHRISTIAN BINET1,2
AND PHILI PPE COLOMBAT1,3
1JE 1993, Faculty of Medicine, Tours, 2Laboratory of Haematology, and 3Department of Haematology/Oncology,University Hospital, Tours, and 4Regional Blood Bank, Tours, France
Received 7 May 1998; accepted for publication 28 July 1998
Summary. Retinoids, especially all-trans-retinoic acid(ATRA), are well known for their differentiating activity onHL-60 cells. Moreover ATRA induces CD38 antigen over-expression on these cells. In this study we examined theeffects of ATRA on purified normal CD34þ cells from adulthuman marrows incubated with ATRA (1 mM) or stem cellfactor (SCF) after 7 d liquid cultures in serum-deprivedmedium. Before and after the incubation, CD34þ cells werestudied by flow cytometry to evaluate the cell-surfaceexpression of CD38 and c-Kit antigens and the cycle statusof these cells using high-resolution analysis (DNA content vKi-67 antigen expression) to clarify the functional meaningof antigenic variations. When compared with controlcultures, ATRA-treated cells displayed changes in theirimmunophenotypic profile. Particularly relevant was theup-regulation of CD38 antigen with a mean (6SEM) fold
increase of 2·1 6 0·1 (P ¼ 0·028) for geometric meanfluorescence intensity (GMFI), without modulation of c-Kitexpression. SCF only down-regulated expression of c-Kit witha fold decrease of 4·6 6 0·9 for GMFI (P ¼ 0·043). UnlikeSCF, ATRA did not induce CD34þ cells to entry into cell cycledespite increased levels of surface CD38 antigen. Moreovermorphological and functional assays did not argue for anATRA-induced maturation process. Contrary to steady-statecells, CD34þ cells treated with pharmacological doses ofATRA alone displayed CD38 over-expression without changein c-Kit levels and cycle status, suggesting an absence ofmaturation pressure.
Keywords: all-trans-retinoic acid, CD34þ cells, CD38antigen, c-Kit, cell cycle.
Human haemopoietic stem cells and primitive progenitorcells are characterized by surface CD34 expression andabsence of CD38 expression. During differentiation andcommitment of CD34þ cells the expression of CD38 antigenincreases with the expression of the c-kit proto-oncogeneproduct, the SCF receptor (Gore et al, 1995). Most terminallydifferentiated cells which have lost CD34 antigen no longerexpress CD38 and c-Kit antigens (Terstappen et al, 1991;Kawashima et al, 1996).
Retinoids, especially all-trans-retinoic acid (ATRA), havebeen shown to exert in vitro a wide range of biologicaleffects on malignant myeloid cells, including a strongdifferentiating activity on cells such as HL-60 cells, a line
derived from a human myeloblastic leukaemia (Breitmanet al, 1980, 1981). This differentiating activity is also wellknown on fresh acute promyelocytic leukaemia cells in vitro(Dan Dauer & Koeffler, 1983) as well as in vivo (Daenen et al,1986; Huang et al, 1988). On the other hand, ATRA is apotent and highly specific inducer of CD38 antigen on HL-60cells (Drach et al, 1993; Kontani et al, 1993), mediatedthrough one of its nuclear receptors (RAR-a) (Drach et al,1994; Mehta et al, 1997). However, little information isavailable on the effects of ATRA on the membranephenotype of normal CD34þ cells in relation to theirphysiological state. A recent study suggests an up-regulationof CD38 expression by ATRA (Drach et al, 1994). Oneapproach to a better understanding of these effects is thestudy of membrane antigen expression on CD34þ cellscoupled with cell cycle analysis to clarify the functionalmeaning of these variations.
British Journal of Haematology, 1998, 103, 343–350
343q 1998 Blackwell Science Ltd
Correspondence: Dr O. Herault, Laboratoire d’Hematologie, JE 1993,Faculte de Medecine, 2 bis boulevard Tonnelle, 37032 Tours Cedex,France.
The purpose of our study was to evaluate CD38 and c-Kitexpression together with cell cycle status on normalmarrow CD34þ cells treated in vitro with ATRA. CD34þ
cells were studied by flow cytometry immediately after theirisolation and after exposure to ATRA in serum-free medium.We analysed concomitantly the pattern of differentiation, themorphology and the clonogenic efficiency of these cells.
Our study identified ATRA as a regulator of the CD38expression on human marrow progenitor cells. ATRAinduced the overexpression of CD38 antigen without
increase of c-Kit expression on CD34þ cells and no clearevidence for entry into cell cycle, contrary toCD34þCD38high cells in steady state, suggesting an absenceof maturation pressure.
MATERIALS AND METHODS
Isolation of CD34þ cells. CD34þ bone marrow cells wereobtained from bone marrow aspirates of 12 healthy donors,who had given informed consent. Mononuclear cells were
q 1998 Blackwell Science Ltd, British Journal of Haematology 103: 343–350
344 Olivier Herault et al
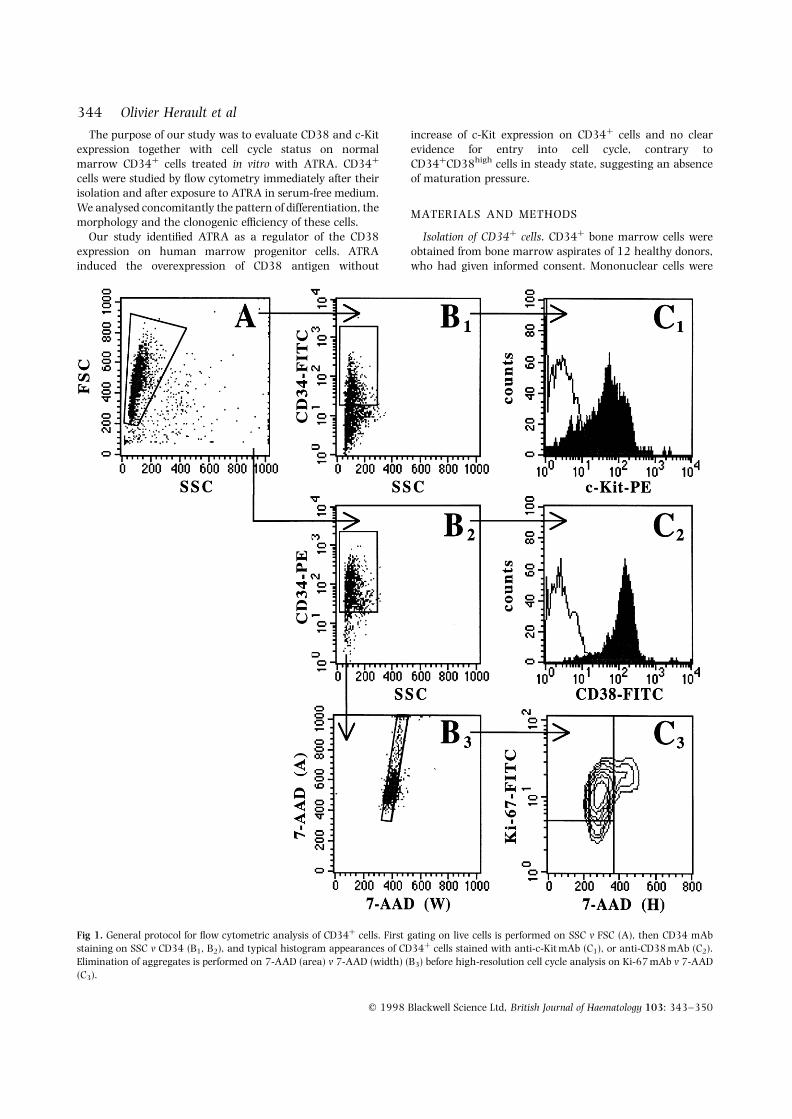
Fig 1. General protocol for flow cytometric analysis of CD34þ cells. First gating on live cells is performed on SSC v FSC (A), then CD34 mAbstaining on SSC v CD34 (B1, B2), and typical histogram appearances of CD34þ cells stained with anti-c-Kit mAb (C1), or anti-CD38 mAb (C2).Elimination of aggregates is performed on 7-AAD (area) v 7-AAD (width) (B3) before high-resolution cell cycle analysis on Ki-67 mAb v 7-AAD(C3).

345Effects of ATRA on CD38 and c-Kit Expression by CD34þ Cells
q 1998 Blackwell Science Ltd, British Journal of Haematology 103: 343–350
separated by centrifugation over a Ficoll-Hypaque gradientand enriched for CD34þ cells by direct immuno-separationusing magnetic beads (Dynal CD34 Progenitor Cell SelectionSystem , Dynal A.S., Oslo, Norway) according to themanufacturer’s instructions. To determine the purity of theselected population, cells were stained with HPCA-2 mono-clonal antibody (mAb) (anti-CD34 R-phycoerythrin (PE)directly conjugated; Becton Dickinson, Mountain View,Calif., U.S.A.), and analysed on a FACScalibur flowcytometer (Becton Dickinson).
CD34þ cell cultures in liquid medium. CD34þ cells wereincubated at a concentration of 2 × 104 cells/ml for 7 d at378C, 5% CO2 in the presence of ATRA (1 mM; Sigma, St Louis,Mo., U.S.A.) or SCF (10 ng/ml, kindly provided by Amgen,Thousand Oaks, Calif., U.S.A.), in a serum-free mediumconsisting of Iscove’s medium (Life Technologies, Cergy-Pontoise, France) supplemented with 1% deionized bovineserum albumin (Sigma), 0·3 mg/ml human transferrin(Boerhinger-Mannheim, Mannheim, Germany), 7·8 mg/mlcholesterol (Sigma), 30 mg/ml L-a-phosphatidyl cholin(Sigma), 0·25 U/ml human insulin (Orgasuline , Organon,St Denis, France) and 0·1 mM 2-mercaptoethanol (Sigma).To prepare stock solution, ATRA was dissolved at 1 mmol/lin dimethyl sulphoxide (Sigma) to produce a finalconcentration of DMSO not exceeding 0·01% in culturemedium.
Clonogenic assays and morphology. Cells were cultured ina semisolid system for CFU-GM and BFU-E assays.Culture medium consisted of 1% methylcellulose (Sigma)with Iscove’s medium supplemented with 30% fetalcalf serum (Life Technologies), 2% deionized bovineserum albumin, 0·1 mM 2-b-mercaptoethanol, 0·1 mM
pyruvate (Boerhinger-Mannheim), 2 mM L-alanyl-L-glutamine(Life Technologies), 1% penicillin-streptomycin (Bristol-Myers-Squibb, Paris La Defense, France) and 0·25 mg/mlamphotericin B. Clonogenic assays were performed usingSCF, granulocyte-macrophage colony-stimulating factor(GM-CSF) (Schering-Plough, Madison, N.J., U.S.A.), granulocyte
colony-stimulating factor (G-CSF) (Amgen), interleukin-3(IL-3) (Life Technologies) at 10 ng/ml and erythropoietin(Epo) (Boehringer-Mannheim) at 3 IU/ml. Cells were platedin triplicate 1 ml cultures at 103 cells/ml in Petri dishes(Nunc, Denmark) and incubated at 378C in a humidified 5%CO2 atmosphere. The culture plates were scored in situ on aninverted microscope on day 14 for CFU-GM (aggregates >40nonhaemoglobinized cells) and for BFU-E (aggregates>200 haemoglobinized cells). Morphological study requiredcytocentrifugation at 6000 rpm, using a cytocentrifuge(CytospinTM, Shandon-Southern, Eragny, France). The cellswere fixed in 100% methanol, stained with May-Grunwald-Giemsa, and morphologic features were examined by lightmicroscopy. Cell viability was evaluated after propidiumiodide and acridine orange staining and studied byepifluorescence microscopy.
Flow cytometric analysis: surface staining. All acquisitionswere performed using the CELLquest software (BectonDickinson) on a FACScalibur flow cytometer equipped with488 nm argon laser. Expression of CD38 protein wasdetermined using mAb in two-colour flow cytometricexperiments, where fluorescein isothiocyanate (FITC)-con-jugated Leu-17 (anti-CD38; Caltag, Burlingame, Calif.,U.S.A.) was used together with PE-conjugated HPCA-2. Asregards c-Kit antigen, PE-conjugated 95C3 (anti-c-Kit;Immunotech, Marseille, France) was used together withFITC-conjugated 581 (anti-CD34; Immunotech). All anti-bodies were used at saturating concentrations according tothe manufacturer’s recommendations and cells were incu-bated for 30 min on ice, followed by two washes withphosphate-buffered saline (PBS). Isotypic control antibodies(PE-conjugated IgG1 and FITC-conjugated IgG1; Dako,Glostrup, Denmark) were used to set the compensationsand to determine background fluorescences (PE and FITCfluorescences were detected on a logarithmic scale). CD38and c-Kit expression on CD34þ subpopulations was mea-sured after setting a live-gate on cells positive for the CD34antigen and collecting at least 5000 events. Intensity of
Table I. Characteristics of fresh and liquid cultured cells.
Day 7
Day 0 Control ATRA SCF
Cell number 100·0 6 0·0 42·4 6 4·0 51·8 6 4·5 77·5 6 5·5% of CD34þ cells 80·1 6 5·0 46·2 6 4·5 47·9 6 3·7 44·9 6 5·7% of viable cells 95·3 6 0·9 76·5 6 2·6 75·1 6 2·6 79·2 6 2·9
% of CD34þ cells expressing lineage markersCD13 16·1 6 3·1 19·5 6 4·5 26·5 6 5·4 9·6 6 2·9CD36 33·0 6 3·8 10·7 6 5·2 15·9 6 2·9 10·8 6 2·9CD19 19·9 6 5·9 0 0 0CD7 2·1 6 0·0 0 0 0
Cells were treated with ATRA (10¹6M) or SCF (10 ng/ml) for 7 d in a serum-free medium (in comparison with
untreated cells). Phenotypic analysis was performed at day 0 and day 7 using HPCA2-PE (anti-CD34 mAb), SJ1D1-FITC (anti-CD13 mAb), CLB-IVC7-FITC (anti-CD36 mAb), HD37-FITC (anti-CD19 mAb), DK24-FITC (anti-CD7mAb).
CD38 or c-Kit expression was shown as signal/noise (S/N)ratios defined as the geometric mean fluorescence of CD38 orc-Kit-expressing cells divided by the geometric meanfluorescence of cells stained with the isotypic controlantibody. Moreover, the pattern of differentiation of CD34þ
cells, including expression of myeloid (CD13), erythroid(CD36) and lymphoid (CD7, CD19) antigens was studiedusing FITC-conjugated mAbs against CD13 (SJ1D1,Immunotech), CD7 (DK24) and CD19 (HD37) obtainedfrom Dako, and mAb against CD36 (CLB-IVC7) obtainedfrom CLB (Amsterdam, The Netherlands).
Flow cytometric analysis: high-resolution cell cycle analysis.The cell cycle status (G0, G1 and SþG2/M) of CD34þ cellswas determined by simultaneous two-parameter analysisusing DNA content versus antibody staining for the Ki-67antigen (Jordan et al, 1996). After the culture, cells wereharvested, washed once, and resuspended in PBS plus 1%fetal calf serum (FCS). All subsequent steps were performedon ice. First, cell surface staining was done using PE-conjugated HPCA-2 for 30 min. Isotype control wasperformed in parallel for each sample. After surface stainingwas complete, cells were resuspended in PBS, and para-formaldehyde (CellFix ; Becton Dickinson) was added to afinal concentration of 1%. Cells were then incubated for30 min, an equal volume of PBS plus 0·2% Triton X-100(Sigma) was added and samples were left overnight at 48C.The fixed and permeabilized cells were washed, resuspendedin PBS plus 1% FCS and stained with FITC-conjugated MIB-1(anti-Ki-67; Immunotech). Finally, the cells were washedand resuspended in PBS plus 1% FCS containing 0·5 mg/ml7-aminoactinomycin D (7-AAD) (Sigma). Samples wereincubated in 7-AAD overnight before analysis. Sampleswere run at low differential pressure, in order to minimizeCV, and at least 3 × 104 events were collected. FL1 (FITC)and FL2 (PE) fluorescences were collected in a logscale, and FL3 (7-AAD) fluorescence in a linear scale.Samples were analysed by first gating on forward scatter(FSC) v side scatter (SSC) to select live cells. Next, SSC vCD34 fluorescence was visualized and gates wereestablished to define the subpopulations of interest. Finally,after a gating on FL3-area v FL3-width to eliminateaggregates, 7-AAD v Ki-67 was visualized, and the propor-tion of events corresponding to each cycle stage wasdetermined.
Statistical analysis. Significance of immunophenotypicchanges (geometric means) during liquid culture andbetween different culture conditions were evaluated usingthe Wilcoxon’s test for paired samples.
RESULTS
Fig 1 shows the protocol and typical scattergramand histogram appearances of CD34þ cells stained withanti-Ki-67 mAb and 7-AAD, anti-c-Kit mAb, or anti-CD38 mAb.
Functional, morphological and phenotypic characteristicsof cellsThe characteristics of selected and day 7 cells are shown inTable I (n ¼ 12). The drastic conditions of serum-free
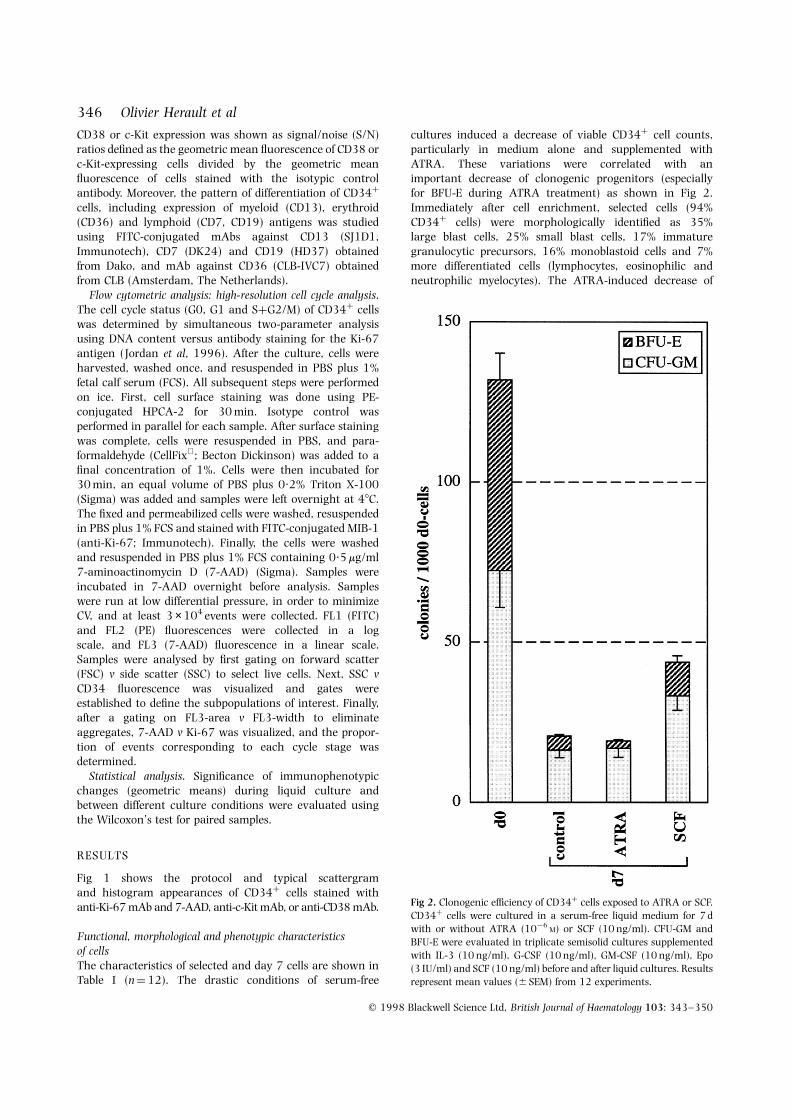
cultures induced a decrease of viable CD34þ cell counts,particularly in medium alone and supplemented withATRA. These variations were correlated with animportant decrease of clonogenic progenitors (especiallyfor BFU-E during ATRA treatment) as shown in Fig 2.Immediately after cell enrichment, selected cells (94%CD34þ cells) were morphologically identified as 35%large blast cells, 25% small blast cells, 17% immaturegranulocytic precursors, 16% monoblastoid cells and 7%more differentiated cells (lymphocytes, eosinophilic andneutrophilic myelocytes). The ATRA-induced decrease of
q 1998 Blackwell Science Ltd, British Journal of Haematology 103: 343–350
346 Olivier Herault et al
Fig 2. Clonogenic efficiency of CD34þ cells exposed to ATRA or SCF.CD34þ cells were cultured in a serum-free liquid medium for 7 dwith or without ATRA (10¹6
M) or SCF (10 ng/ml). CFU-GM andBFU-E were evaluated in triplicate semisolid cultures supplementedwith IL-3 (10 ng/ml), G-CSF (10 ng/ml), GM-CSF (10 ng/ml), Epo(3 IU/ml) and SCF (10 ng/ml) before and after liquid cultures. Resultsrepresent mean values (6 SEM) from 12 experiments.
347Effects of ATRA on CD38 and c-Kit Expression by CD34þ Cells
q 1998 Blackwell Science Ltd, British Journal of Haematology 103: 343–350
CD34þ cell population was not associated with aclear differentiation: morphological analysis demonstratedthat ATRA was unable to promote myeloid maturation,and cytofluorometric analysis of CD13 expression showeda very moderate commitment in myeloid compartment(Table I).
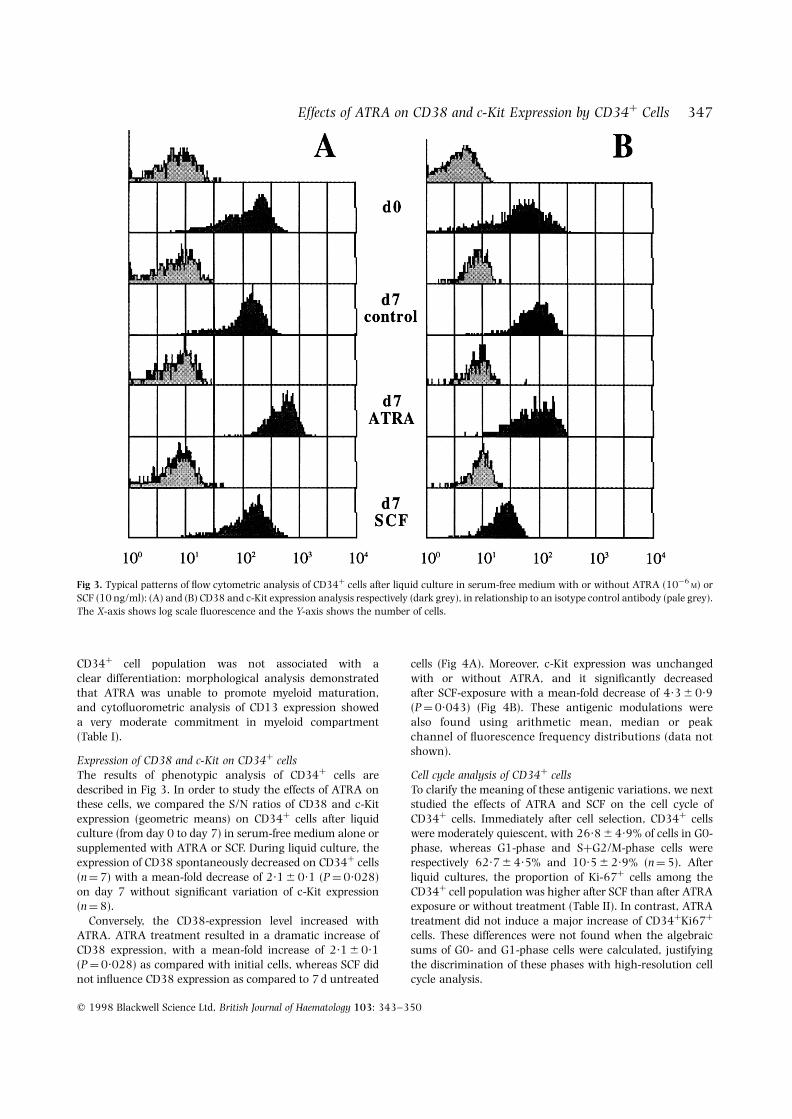
Expression of CD38 and c-Kit on CD34þ cellsThe results of phenotypic analysis of CD34þ cells aredescribed in Fig 3. In order to study the effects of ATRA onthese cells, we compared the S/N ratios of CD38 and c-Kitexpression (geometric means) on CD34þ cells after liquidculture (from day 0 to day 7) in serum-free medium alone orsupplemented with ATRA or SCF. During liquid culture, theexpression of CD38 spontaneously decreased on CD34þ cells(n ¼ 7) with a mean-fold decrease of 2·1 6 0·1 (P ¼ 0·028)on day 7 without significant variation of c-Kit expression(n ¼ 8).
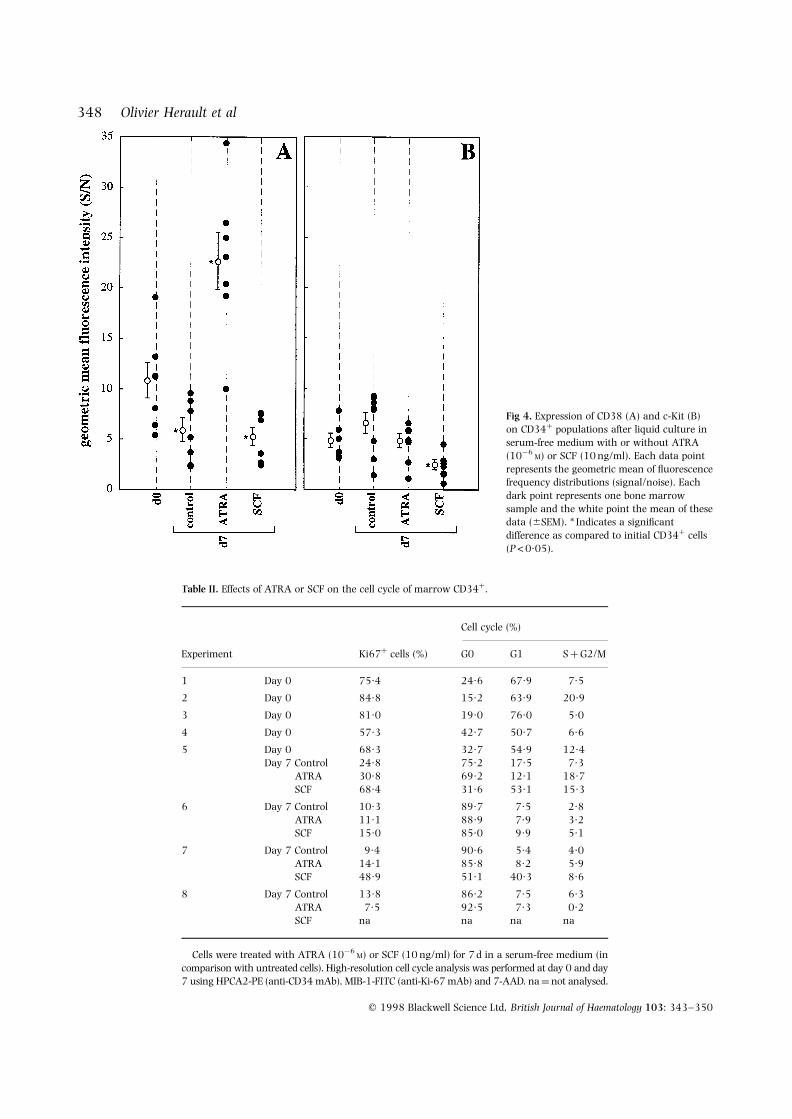
Conversely, the CD38-expression level increased withATRA. ATRA treatment resulted in a dramatic increase ofCD38 expression, with a mean-fold increase of 2·1 6 0·1(P ¼ 0·028) as compared with initial cells, whereas SCF didnot influence CD38 expression as compared to 7 d untreated
cells (Fig 4A). Moreover, c-Kit expression was unchangedwith or without ATRA, and it significantly decreasedafter SCF-exposure with a mean-fold decrease of 4·3 6 0·9(P ¼ 0·043) (Fig 4B). These antigenic modulations werealso found using arithmetic mean, median or peakchannel of fluorescence frequency distributions (data notshown).
Cell cycle analysis of CD34þ cellsTo clarify the meaning of these antigenic variations, we nextstudied the effects of ATRA and SCF on the cell cycle ofCD34þ cells. Immediately after cell selection, CD34þ cellswere moderately quiescent, with 26·8 6 4·9% of cells in G0-phase, whereas G1-phase and SþG2/M-phase cells wererespectively 62·7 6 4·5% and 10·5 6 2·9% (n ¼ 5). Afterliquid cultures, the proportion of Ki-67þ cells among theCD34þ cell population was higher after SCF than after ATRAexposure or without treatment (Table II). In contrast, ATRAtreatment did not induce a major increase of CD34þKi67þ
cells. These differences were not found when the algebraicsums of G0- and G1-phase cells were calculated, justifyingthe discrimination of these phases with high-resolution cellcycle analysis.
Fig 3. Typical patterns of flow cytometric analysis of CD34þ cells after liquid culture in serum-free medium with or without ATRA (10¹6M) or
SCF (10 ng/ml): (A) and (B) CD38 and c-Kit expression analysis respectively (dark grey), in relationship to an isotype control antibody (pale grey).The X-axis shows log scale fluorescence and the Y-axis shows the number of cells.
q 1998 Blackwell Science Ltd, British Journal of Haematology 103: 343–350
348 Olivier Herault et al
Fig 4. Expression of CD38 (A) and c-Kit (B)on CD34þ populations after liquid culture inserum-free medium with or without ATRA(10¹6
M) or SCF (10 ng/ml). Each data pointrepresents the geometric mean of fluorescencefrequency distributions (signal/noise). Eachdark point represents one bone marrowsample and the white point the mean of thesedata (6SEM). * Indicates a significantdifference as compared to initial CD34þ cells(P<0·05).
Table II. Effects of ATRA or SCF on the cell cycle of marrow CD34þ.
Cell cycle (%)
Experiment Ki67þ cells (%) G0 G1 S þ G2/M
1 Day 0 75·4 24·6 67·9 7·5
2 Day 0 84·8 15·2 63·9 20·9
3 Day 0 81·0 19·0 76·0 5·0
4 Day 0 57·3 42·7 50·7 6·6
5 Day 0 68·3 32·7 54·9 12·4Day 7 Control 24·8 75·2 17·5 7·3
ATRA 30·8 69·2 12·1 18·7SCF 68·4 31·6 53·1 15·3
6 Day 7 Control 10·3 89·7 7·5 2·8ATRA 11·1 88·9 7·9 3·2SCF 15·0 85·0 9·9 5·1
7 Day 7 Control 9·4 90·6 5·4 4·0ATRA 14·1 85·8 8·2 5·9SCF 48·9 51·1 40·3 8·6
8 Day 7 Control 13·8 86·2 7·5 6·3ATRA 7·5 92·5 7·3 0·2SCF na na na na
Cells were treated with ATRA (10¹6M) or SCF (10 ng/ml) for 7 d in a serum-free medium (in
comparison with untreated cells). High-resolution cell cycle analysis was performed at day 0 and day7 using HPCA2-PE (anti-CD34 mAb), MIB-1-FITC (anti-Ki-67 mAb) and 7-AAD. na ¼ not analysed.

349Effects of ATRA on CD38 and c-Kit Expression by CD34þ Cells
q 1998 Blackwell Science Ltd, British Journal of Haematology 103: 343–350
DISCUSSION
Previous studies have indicated that CD34þCD38¹ cells arehighly enriched for blast colony-forming cells, whereasCD34þCD38þ cells are associated with lineage commitment(Terstappen et al, 1991). Moreover, there is a goodcorrelation between CD38 expression and cycle status:CD34þCD38¹ cells represent a noncycling population,whereas cells that express high levels of CD38 are veryactively cycling (Hao et al, 1995; Jordan et al, 1996; Reems &Torok-Storb, 1995). Moreover, primitive haemopoietic pro-genitors in cell cycle dormancy are Kitlow whereas activelycycling, more mature, progenitors are Kithigh (Katayama et al,1993). The expression of Kit declines during furthermaturation (Kawashima et al, 1996), except in the mastcell lineage.
In order to evaluate the direct effect of ATRA on humanmarrow progenitor cells, we chose to use CD34þ cellscultured in vitro in a serum-free medium. Activity of ATRAadded at pharmacological doses (1 mM) was compared to thatof SCF at 10 ng/ml (a low concentration sufficient to inducecommitment) and to control cultures. To understand themeaning of the antigenic modulations observed, we analysedthe cell cycle. Using monoparametric analysis of cell cycle(propidium iodide alone), it is impossible to distinguish cellsin G0 v G1. As there was no difference between ATRA- andSCF-treated cells (Table I) considering the algebraicsums of G0 and G1-phase cells (G0/G1 peak), we used ahigh-resolution cell cycle analysis (Ki-67 v 7-AAD).
Most of initial CD34þ cells were predominantly in activecycle before culture and were unable to remain sponta-neously in this cycle state after culture in medium aloneprobably because of the deprivation of cytokine. Theseculture conditions could explain the spontaneous decrease ofCD38 expression observed in the CD34þ population.
After exposure to SCF, as compared to initial levels and 7 dlevels, we observed a significant down-regulation of c-Kit, aspreviously described (Blume-Jensen et al, 1991; Gore et al,1995), without modulation of CD38 expression. However,we concomitantly found a clear commitment of CD34þ cellsinto cell cycle showing that our system could detect such anactivation even in serum-free culture.
Conversely, ATRA was able to significantly increase CD38expression on normal CD34þ marrow cells, as previouslyshown on HL-60 cells and has been suggested for CD34þ
cells (Drach et al, 1994). CD38 up-regulation occurredwithout concomitant over-expression of c-Kit antigen asexpected during physiological commitment of CD34þ cells(Katayama et al, 1993; Kawashima et al, 1996). The highlevel of CD38 expression is not correlated with a maturationprocess, as demonstrated by the weak increase of CD13expression, which appears theoretically very early in myeloidmaturation (Tjonnfjord et al, 1996), without morphologicalargument for myeloid differentiation. Morever, this ATRA-induced up-regulation was not associated with increasedlevels of cycle activity, contrary to marrow CD34þCD38high
cells in steady state (Jordan et al, 1996). These data areconcordant with the decrease of clonogenic progenitors, asthe haemopoietic growth factors used for clonogenic assays
were only effective on G1 cells. The higher decrease of BFU-Ecan be explained by inhibition of the erythroid differentiationprogramme induced by ATRA (Labbaye et al, 1994). Theseantigenic variations cannot be explained by a decrease of theimmature progenitor cell compartment, considering the verylow number of these cells in the initial CD34þ cellpopulation.
In conclusion, ATRA is capable of directly inducing CD38cell expression uncoupled with c-Kit modulation and entryinto cell cycle on cultured CD34þ cells. Therefore, after invitro treatment with pharmacological doses of ATRA, CD38antigen expression cannot attest a maturation of marrowCD34þ cells. To clarify the effects of ATRA on marrow long-term culture-initiating cells (LTC-IC), further functionalstudies (long-term cultures with limiting dilution assay)are needed to determine whether these phenotypic changesare associated with a decrease of the early progenitor cellpopulation.
ACKNOWLEDGMENTS
We are grateful to Professor Michel Marchand for providingsamples from patients undergoing cardiac surgery. We thankNathalie Heraud and Marie-Therese Georget for theirtechnical assistance. This work was supported by grantsfrom the Conseil Regional de la Region Centre (grant number99500 R3 0005), the Comites du Cher et de l’Indre-et-Loirede la Ligue Nationale contre le Cancer and the Ministere del’Enseignement Superieur et de la Recherche.
REFERENCES
Blume-Jensen, P., Cleasson-Welsh, L., Siegbahn, A., Zsebo, K.M.,Westermark, B. & Heldin, C.H. (1991) Activation of the c-kitproduct by ligand-induced dimerization mediates circular actinreorganization and chemotaxis. EMBO Journal, 10, 4121–4128.
Breitman, T.R., Collins, S.J. & Keene, B.R. (1981) Terminaldifferentiation of human promyelocytic leukemic cells in primaryculture in response to retinoic acid. Blood, 57, 1000–1004.
Breitman, T.R., Selonick, S.E. & Collins, S.J. (1980) Induction ofdifferentiation of the human promyelocytic leukemia cell line (HL-60) by retinoic acid. Proceedings of the National Academy of Sciencesof the United States of America, 86, 2936–2940.
Daenen, S., Vellenga, E., van Dobbenburgh, O.A. & Halie, M.R.(1986) Retinoic acid as antileukemic therapy in a patient withacute promyelocytic leukemia and Aspergillus pneumonia. Blood,67, 559–661.
Dan Dauer, H. & Koeffler, P. (1983) Effects of retinoids on normal andabnormal hematopoiesis. Modulation and Mediation of Cancer byVitamins (ed. by F. L. Meyskens and K. N. Prasad), pp. 236–243.Karger, Basel.
Drach, J., McQueen, T., Engel, H., Andreef, M., Robertson, K.A.,Collins, S.J., Malavasi, F. & Mehta, K. (1994) Retinoic acid-inducedexpression of CD38 antigen in myeloid cells is mediated throughretinoic acid receptor-a. Cancer Research, 54, 1746–1752.
Drach, J., Zhao, S., Malavasi, F. & Mehta, K. (1993) Rapid inductionof CD38 antigen on myeloid leukemia cells by all trans-retinoicacid. Biochemical and Biophysical Research Communications, 195,545–550.
Gore, S.D., Amin, S., Weng, L.J. & Civin, C.I. (1995) Steel factorsupports the cycling of isolated human CD34þ cells in the absenceof other growth factors. Experimental Hematology, 23, 413–421.

Hao, Q.L., Shah, A.J., Thiemann, F.T., Smogorzewska, E.M. & Crooks,G.M. (1995) A functional comparison of CD34þCD38¹ cells incord blood and bone marrow. Blood, 86, 3745–3753.
Huang, M., Ye, Y., Chen, S., Chai, J.R., Lu, J.X., Zhoa, L., Gu, L.J. &Wang, Z.Y. (1988) Use of all-trans retinoic acid in the treatment ofpromyelocytic leukemia. Blood, 72, 567–572.
Jordan, C.T., Yamasaki, G. & Minamoto, D. (1996) High-resolutioncell cycle analysis of defined phenotypic subsets within primitivehuman hematopoietic cell populations. Experimental Hematology,24, 1347–1355.
Katayama, N., Shih, J.P., Nishikawa, S.I., Kina, T., Clark, S.C. &Ogawa, M. (1993) Stage-specific expression of c-Kit protein bymurine hematopoietic progenitors. Blood, 82, 2353–2360.
Kawashima, I., Zanjani, E.D., Almaida-Porada, G., Flake, A.W.,Zeng, H.Q. & Ogawa, M. (1996) CD34þ human marrow cells thatexpress low levels of Kit protein are enriched for long-termmarrow-engrafting cells. Blood, 87, 4136–4142.
Kontani, K., Nishina, H., Ohoka, Y., Takahashi, K. & Katada, T.(1993) NAD glycohydrolase specifically induced by retinoic acid inhuman leukemic HL-60 cells. Journal of Biological Chemistry, 268,16895–16898.
Labbaye, C., Valtieri, M., Testa, U., Giampaolo, A., Meccia, E.,Sterpetti, P., Parolini, I., Pelosi, E., Bulgarini, D., Cayre, Y.E. &Peschle, C. (1994) Retinoic acid downmodulates erythroiddifferentiation and GATA1 expression in purified adult-progenitorculture. Blood, 83, 651–656.
Mehta, K., McQueen, T., Manshouri, T., Andreeff, M., Collins, S. &Albitar, M. (1997) Involvement of retinoic acid receptor-a-mediated signaling pathway in induction of CD38 cell-surfaceantigen. Blood, 89, 3607–3614.
Reems, A. & Torok-Storb, B. (1995) Cell cycle and functionaldifferences between CD34þCD38high and CD34þCD38low
human marrow cells after in vitro cytokine exposure. Blood, 6,1480–1487.
Terstappen, L.W.M.M., Huang, S., Safford, M., Lansdorp, P.M. &Coken, M.R. (1991) Sequential generations of hematopoieticcolonies derived from single nonlineage-committed CD34þCD38¹
progenitor cells. Blood, 77, 1218–1227.Tjonnfjord, G.E., Steen, R., Veiby, O.P. & Egeland, T. (1996) Lineage
commitment of CD34þ human hematopoietic progenitor cells.Experimental Hematology, 24, 875–882.
q 1998 Blackwell Science Ltd, British Journal of Haematology 103: 343–350
350 Olivier Herault et al
Copyright © 2022 FDOKUMEN




















