Adoptively Transferred Tumor-Specific T Cells Stimulated Ex vivo Using Herpes Simplex Virus...
12
Adoptively Transferred Tumor-Specific T Cells Stimulated Ex vivo Using Herpes Simplex Virus Amplicons Encoding 4-1BBL Persist in the Host and Show Antitumor Activity In vivo Kyung H. Yi, 1,3 Hovav Nechushtan, 4 William J. Bowers, 5,6 Gail R. Walker, 2 Yu Zhang, 3 Dien G. Pham, 3 Eckhard R. Podack, 1 Howard J. Federoff, 7 Khaled A. Tolba, 3 and Joseph D. Rosenblatt 1,3 1 Department of Microbiology and Immunology and 2 Biostatistics Department, University of Miami Miller School of Medicine, and 3 University of Miami Sylvester Comprehensive Cancer Center, Miami, Florida; 4 Department of Oncology, Hebrew University Hadassah School of Medicine, Jerusalem, Israel; 5 Department of Neurology and 6 Center for Aging and Developmental Biology, University of Rochester School of Medicine, Rochester, New York; and 7 Georgetown University Medical Center, Washington, District of Columbia Abstract 4-1BB is a T-cell costimulatory receptor which binds its ligand 4-1BBL, resulting in prolonged T cell survival. We studied the antitumor effects of adoptively transferred tumor-specific T cells expanded ex vivo using tumors transduced with herpes simplex virus (HSV) amplicons expressing 4-1BBL as a direct source of antigen and costimulation. We constructed HSV amplicons encoding either the 4-1BBL (HSV.4-1BBL) or B7.1 (HSV.B7.1) costimulatory ligands. Lewis lung carcinoma cells expressing ovalbumin (LLC/OVA) were transduced with HSV.4-1BBL, HSV.B7.1, or control HSV amplicons and used to stimulate GFP + OVA-specific CD8 + T cells (OT-1/GFP) ex vivo . Naive or ex vivo stimulated OT-1/GFP cells were adoptively transferred into LLC/OVA tumor-bearing mice. Higher percentages of OT-1/ GFP cells were seen in the peripheral blood, spleen, and tumor bed of the HSV.4-1BBL–stimulated OT-1/GFP group compared with all other experimental groups. OT-1 cells identified within the tumor bed and draining lymph nodes of the HSV.4-1BBL– stimulated OT-1 group showed enhanced bromodeoxyuridine (BrdUrd) incorporation, suggesting ongoing expansion in vivo . Mice receiving HSV.4-1BBL–stimulated OT-1/GFP had signifi- cantly decreased tumor volumes compared with untreated mice (P < 0.001) or to mice receiving naive OT-1/GFP (P < 0.001). Transfer of HSV.B7.1-stimulated OT-1/GFP did not protect mice from tumor. Mice that received HSV.4-1BBL–stimulated OT-1/ GFP exhibited increased cytolytic activity against LLC/OVA and higher percentages of Ly-6C + OT-1/GFP in the spleen and tumor bed compared with controls. Tumor-specific Tcells stimulated ex vivo using tumor transduced with HSV.4-1BBL expand in vivo following adoptive transfer, resulting in tumor eradication and the generation of tumor-specific CD44 + Ly-6C + CD62L effector memory T cells. [Cancer Res 2007;67(20):10027–37] Introduction Adoptive transfer of autologous tumor-specific T cells is a promising approach to cancer immunotherapy. T cells isolated from tumor-infiltrating lymphocytes, tumor-draining lymph nodes (TDLN), or peripheral blood contain tumor-specific cells, which can be expanded ex vivo and transferred into the host. Responses have been observed in metastatic melanoma (1), renal cell carcinoma (2), and glioma (3), among other cancers, when treated with ex vivo expanded CTLs. Immune ablation may further augment responses seen with adoptive T-cell transfer (1). Efficacy of T-cell adoptive transfer may be improved through optimization of in vitro expansion, improved characterization of effector populations, and/or by enhancing the function and survival of transferred CTLs to facilitate establishment of immunologic memory. 4-1BB (CD137, ILA, TNFRSF9) is a type I transmembrane glycoprotein belonging to the tumor necrosis factor receptor superfamily (4, 5). 4-1BB expression is observed in a range of myeloid and lymphoid cells, including CD4 + and CD8 + T cells, intraepithelial lymphocytes, natural killer cells, monocytes, and dendritic cells (5, 6). In contrast to CD28 expression on naive T cells, 4-1BB is induced on T cells following activation. CD28 seems to relay an initial costimulatory signal followed by 4-1BB signaling, which serves to further shape the T-cell response. 4-1BB ligation induces cytokine secretion, especially IFN-g, enhances proliferation and survival of T cells in vitro and in vivo (5, 7–12), and plays a crucial role in the generation and expansion of effector and memory CTLs (8, 13). Administration of agonistic anti– 4-1BB monoclonal antibody (mAb) enhanced antitumor responses in the poorly immunogenic Ag104A sarcoma model (14) and improved antitumor effects seen with adoptive transfer of CD8 + T cells in several tumor models (15–17). 4-1BB–mediated antitumor effects have been ascribed to the prevention of programmed cell death, leading to the accumulation of antitumor effector cells (15, 18). Previous experiments have also shown the effectiveness of 4-1BBL gene transduction for antitumor immunity. Melero et al. (19) first transduced 4-1BBL into P815 mastocytoma using a retroviral vector and inoculated mice, which then developed a strong CTL response and long-term immunity against wild- type tumor. In the A20 lymphoma (20) and NRS1 squamous cell carcinoma (21) models, 4-1BBL expression also reduced tumor growth. Combined adenoviral 4-1BBL and interleukin-12 (IL-12) gene transfer conferred protection against hepatic metas- tases induced by a poorly immunogenic MCA26 colon carcinoma line (22). We used herpes simplex virus I (HSV) amplicons, replication- defective viral particles, for gene transfer of 4-1BBL because of their broad cellular tropism, large transgene capacity, and ability to induce high levels of gene expression. By triggering an innate response, HSV amplicons may facilitate a more vigorous adaptive Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). Requests for reprints: Joseph D. Rosenblatt, Division of Hematology/Oncology, University of Miami Sylvester Comprehensive Cancer Center, 1475 NW 12th Avenue (D8-4), Suite 3300, Miami, FL 33136. Phone: 305-243-9528; Fax: 305-243-9161; E-mail: [email protected]. I2007 American Association for Cancer Research. doi:10.1158/0008-5472.CAN-06-2391 www.aacrjournals.org 10027 Cancer Res 2007; 67: (20). October 15, 2007 Research Article Research. on March 3, 2016. © 2007 American Association for Cancer cancerres.aacrjournals.org Downloaded from
Transcript of Adoptively Transferred Tumor-Specific T Cells Stimulated Ex vivo Using Herpes Simplex Virus...

Adoptively Transferred Tumor-Specific T Cells Stimulated Ex vivo
Using Herpes Simplex Virus Amplicons Encoding 4-1BBL Persist
in the Host and Show Antitumor Activity In vivo
Kyung H. Yi,1,3Hovav Nechushtan,
4William J. Bowers,
5,6Gail R. Walker,
2Yu Zhang,
3Dien G. Pham,
3
Eckhard R. Podack,1Howard J. Federoff,
7Khaled A. Tolba,
3and Joseph D. Rosenblatt
1,3
1Department of Microbiology and Immunology and 2Biostatistics Department, University of Miami Miller School of Medicine, and3University of Miami Sylvester Comprehensive Cancer Center, Miami, Florida; 4Department of Oncology, Hebrew UniversityHadassah School of Medicine, Jerusalem, Israel; 5Department of Neurology and 6Center for Aging and DevelopmentalBiology, University of Rochester School of Medicine, Rochester, New York; and 7Georgetown University MedicalCenter, Washington, District of Columbia
Abstract
4-1BB is a T-cell costimulatory receptor which binds its ligand4-1BBL, resulting in prolonged T cell survival. We studied theantitumor effects of adoptively transferred tumor-specific Tcellsexpanded ex vivo using tumors transduced with herpes simplexvirus (HSV) amplicons expressing 4-1BBL as a direct source ofantigen and costimulation. We constructed HSV ampliconsencoding either the 4-1BBL (HSV.4-1BBL) or B7.1 (HSV.B7.1)costimulatory ligands. Lewis lung carcinoma cells expressingovalbumin (LLC/OVA) were transduced with HSV.4-1BBL,HSV.B7.1, or control HSV amplicons and used to stimulateGFP+ OVA-specific CD8+ T cells (OT-1/GFP) ex vivo. Naive orex vivo stimulated OT-1/GFP cells were adoptively transferredinto LLC/OVA tumor-bearing mice. Higher percentages of OT-1/GFP cells were seen in the peripheral blood, spleen, and tumorbed of the HSV.4-1BBL–stimulated OT-1/GFP group comparedwith all other experimental groups. OT-1 cells identified withinthe tumor bed and draining lymph nodes of the HSV.4-1BBL–stimulated OT-1 group showed enhanced bromodeoxyuridine(BrdUrd) incorporation, suggesting ongoing expansion in vivo.Mice receiving HSV.4-1BBL–stimulated OT-1/GFP had signifi-cantly decreased tumor volumes compared with untreated mice(P < 0.001) or to mice receiving naive OT-1/GFP (P < 0.001).Transfer of HSV.B7.1-stimulated OT-1/GFP did not protect micefrom tumor. Mice that received HSV.4-1BBL–stimulated OT-1/GFP exhibited increased cytolytic activity against LLC/OVA andhigher percentages of Ly-6C+ OT-1/GFP in the spleen and tumorbed comparedwith controls. Tumor-specificTcells stimulated exvivo using tumor transduced with HSV.4-1BBL expand in vivofollowing adoptive transfer, resulting in tumor eradication andthe generation of tumor-specific CD44+Ly-6C+CD62L� effectormemory T cells. [Cancer Res 2007;67(20):10027–37]
Introduction
Adoptive transfer of autologous tumor-specific T cells is apromising approach to cancer immunotherapy. T cells isolatedfrom tumor-infiltrating lymphocytes, tumor-draining lymph nodes
(TDLN), or peripheral blood contain tumor-specific cells, whichcan be expanded ex vivo and transferred into the host. Responseshave been observed in metastatic melanoma (1), renal cellcarcinoma (2), and glioma (3), among other cancers, when treatedwith ex vivo expanded CTLs. Immune ablation may furtheraugment responses seen with adoptive T-cell transfer (1). Efficacyof T-cell adoptive transfer may be improved through optimizationof in vitro expansion, improved characterization of effectorpopulations, and/or by enhancing the function and survival oftransferred CTLs to facilitate establishment of immunologicmemory.4-1BB (CD137, ILA, TNFRSF9) is a type I transmembrane
glycoprotein belonging to the tumor necrosis factor receptorsuperfamily (4, 5). 4-1BB expression is observed in a range of myeloidand lymphoid cells, including CD4+ and CD8+ T cells, intraepitheliallymphocytes, natural killer cells, monocytes, and dendritic cells (5, 6).In contrast to CD28 expression on naive T cells, 4-1BB is induced onT cells following activation. CD28 seems to relay an initialcostimulatory signal followed by 4-1BB signaling, which serves tofurther shape the T-cell response.4-1BB ligation induces cytokine secretion, especially IFN-g,
enhances proliferation and survival of T cells in vitro and in vivo(5, 7–12), and plays a crucial role in the generation and expansion ofeffector and memory CTLs (8, 13). Administration of agonistic anti–4-1BB monoclonal antibody (mAb) enhanced antitumor responses inthe poorly immunogenic Ag104A sarcoma model (14) and improvedantitumor effects seenwith adoptive transfer of CD8+ Tcells in severaltumor models (15–17). 4-1BB–mediated antitumor effects have beenascribed to the prevention of programmed cell death, leading to theaccumulation of antitumor effector cells (15, 18).Previous experiments have also shown the effectiveness of
4-1BBL gene transduction for antitumor immunity. Melero et al.(19) first transduced 4-1BBL into P815 mastocytoma using aretroviral vector and inoculated mice, which then developeda strong CTL response and long-term immunity against wild-type tumor. In the A20 lymphoma (20) and NRS1 squamouscell carcinoma (21) models, 4-1BBL expression also reducedtumor growth. Combined adenoviral 4-1BBL and interleukin-12(IL-12) gene transfer conferred protection against hepatic metas-tases induced by a poorly immunogenic MCA26 colon carcinomaline (22).We used herpes simplex virus I (HSV) amplicons, replication-
defective viral particles, for gene transfer of 4-1BBL because of theirbroad cellular tropism, large transgene capacity, and ability toinduce high levels of gene expression. By triggering an innateresponse, HSV amplicons may facilitate a more vigorous adaptive
Note: Supplementary data for this article are available at Cancer Research Online(http://cancerres.aacrjournals.org/).
Requests for reprints: Joseph D. Rosenblatt, Division of Hematology/Oncology,University of Miami Sylvester Comprehensive Cancer Center, 1475 NW 12th Avenue(D8-4), Suite 3300, Miami, FL 33136. Phone: 305-243-9528; Fax: 305-243-9161; E-mail:[email protected].
I2007 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-06-2391
www.aacrjournals.org 10027 Cancer Res 2007; 67: (20). October 15, 2007
Research Article
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

response. We have noted the strong activation of several toll-likereceptors (TLR), induction of cytokines, and NKG2D-ligandexpression following transduction with HSV amplicons in macro-phage cell lines and human chronic lymphocytic leukemia (CLL;ref. 23).8 Because HSV amplicons can readily transduce primarytumor cells, we reasoned that HSV amplicons encoding 4-1BBLcould facilitate direct antigen presentation by tumor cells to expandtumor-specific effectors for adoptive transfer. We hypothesized thatHSV.4-1BBL–expanded CD8+ T cells would show desirable effectorproperties, including in vivo expansion and therapeutic efficacy,as well as potentially confer a memory response.We used HSV.4-1BBL amplicons to transduce tumor for
purposes of activating and expanding tumor-specific CD8+ OT-1cells in vitro and studied the behavior of adoptively transferredex vivo expanded cells in LLC/OVA tumor-bearing mice. Ourstudies show that HSV.4-1BBL has the potential to inducesignificant expansion of CTLs in vitro and in vivo , and that theadoptive transfer of expanded T cells may result in a reduction oftumor growth in vivo as well as the persistence of CD44hiLy-6ChiCD62Lneg tumor-specific T cells with memory characteristics.
Materials and Methods
Animals and cells. C57BL/6 and Thy1.1 (C57BL/6.PL-Thy1a/Cy) mice
were obtained from The Jackson Laboratory. Thy1.1 mice, a C57BL/6Jcongenic strain, carry the T cell–specific Thy1a (Thy1.1) allele. OT-1 mice
(obtained courtesy of M. Bevan, Howard Hughes Medical Institute,
University of Washington, Seattle, WA) express a transgenic TCR that is
specific for OVA257-264 (SIINFEKL) peptide bound to H-2Kb. GFP+ mice (24)were bred with OT-1 mice to generate OT-1/GFP T cells expressing green
fluorescent protein (GFP). Mice were maintained in pathogen-free facilities
at the University of Miami, and procedures were done in agreement with theInstitutional Animal Care and Use Committee per NIH guidelines.
Lewis lung carcinoma (LLC) cells stably transfected with ovalbumin
(LLC/OVA; ref. 25) were grown in Iscove’s modified Dulbecco’s medium
plus 10% fetal bovine serum (FBS), penicillin (50 units/mL), streptomycin(50 Ag/mL; I-10), and 1 mg/mL geneticin.
Antibodies. Anti-mouse CD3q-PE, anti-mouse CD8a-PE or -Cy-Chrome
(PE-Cy5), anti-mouse CD4-FITC, anti-mouse Va2-FITC, and anti-mouse
Vh5.1,5.2-PE (BD PharMingen) mAbs were used to stain for T cells. Beforestaining, splenocytes were treated with anti-mouse CD16/CD32 (Fc-gIII/IIreceptor, BD PharMingen) to block Fc-mediated binding. Antibodies used
to assess activation and/or differentiation states of T cells include anti–CD44-PE (eBioscience), anti–CD62L-PE (BD PharMingen), anti-human
granzyme B-PE (Caltag), anti-mouse CD25-biotin (7D4) or anti-mouse
Ly-6C-biotin (BD PharMingen) followed by streptavidin-PE (Sigma), anti-
mouse 4-1BB (BD PharMingen) followed by anti-rat immunoglobulin G1
(IgG1)–biotin (BD PharMingen) and streptavidin-PE, and anti-CD107a
(1D4B; BD PharMingen) followed by anti-rat IgG (H+L)-PE (Caltag). B7.1
expression was detected with anti-human CD80-FITC (BD PharMingen)
and 4-1BBL expression with anti-murine 4-1BBL (BD PharMingen), anti-ratIgG (H+L)-biotin (Caltag), and streptavidin-PE (Sigma).
Purification of CD8+ OT-1 cells. Splenocytes from OT-1 mice were
purified by positive column selection using MACS anti-CD8a (Ly-2)
MicroBeads (Miltenyi Biotec) as described (25). Purified cells were >97%CD8+Va2+Vh5.1,5.2+ as assessed by flow cytometry.
HSV-amplicon-vector construction and helper-virus–free packaging.The cDNA of murine 4-1BBL with a KpnI site 5¶ and NheI site 3¶ wasamplified by reverse transcription-PCR from RNA of C57BL/6 spleen
stimulated with lipopolysaccharide (15 Ag/mL) for 24 h using the 5¶primer: 5¶-GGTACCGCCATGGACCAGCACACACTTG-3¶ and the 3¶ primer:
5¶-GCTAGCTTCCCATGGGTTGTCGGGTTTCAC-3¶ based on its publishednucleotide sequence (26). The cDNA was inserted into pCR-Script Amp
SK(+) (Stratagene; 415.pCRScript) and again amplified by PCR, to introduce
a 5¶ BamHI site and stop codon plus EcoRI 3¶ using the 5¶ primer:
5¶-TCGGATCCGTAATGGACCAGCACACACTTG-3¶ and the 3¶ primer:5¶-GAGAATTCTCATTCCCATGGGTTGTCGGGTTTCAC-3¶. The complete
murine 4-1BBL cDNA was then cloned into the BamHI and EcoRI sites
in the polylinker region of the HSV-1 amplicon vector pHSVPrPUC (27).
The cDNA of human B7.1 was cloned into HSVPrPUC as previouslydescribed (28).
Helper-virus–free replication-defective packaging of HSV-1 viral ampli-
cons was done as described previously (29, 30). Amplicons containing the
gene for Escherichia coli h-galactosidase (HSV.LacZ) were prepared usingthe same vector system.
OT-1/GFP in vitro expansion for in vivo administration. LLC/OVA
cells were resuspended at 106 cells/100 AL I-10 media, transduced with
either HSV.4-1BBL (MOI = 1) or HSV.B7.1 (MOI = 1), and incubated at 37jCfor 1 h before being transferred to a six-well plate. One day later, transduced
LLC/OVA were resuspended at 5 � 106 cells/mL, treated with 0.4 mg/mL
mitomycin C for 20 min at 37jC in PBS, and washed thrice in RPMI 1640
plus 10% FBS, penicillin, streptomycin, and 50 Amol/L 2-ME (R-10). Freshly
isolated OT-1/GFP cells were then plated with mitomycin C–treated
LLC/OVA at a 3:2 ratio in 24-well plates. Each well contained 2.4 � 106 OT-
1/GFP cells plus 1.6 � 106 tumor cells in 2 mL R-10. R-10 media (0.5–1 mL)
was added to each well after 2 days. Cells were harvested on the third day
and separated from the tumor using anti-CD8a magnetic beads before
adoptive transfer.
Intracellular staining and flow cytometry. Cells were stained on the
surface with fluorochrome-conjugated anti-CD4 and anti-CD8a antibodies
in PBS at 4jC for 20 min. Cells were then washed with PBS, fixed usingCytofix/Cytoperm buffer (BD PharMingen) for 20 min at 4jC. A 0.1%
saponin/1% FBS in PBS solution was used to wash, stain with fluorochrome-
conjugated antibodies, and wash cells again. Cells were analyzed using an
LSR flow cytometer and CellQuest software (BD Biosciences).Blood collection and preparation for flow cytometry. Sodium
heparin from a 10-mL Vacutainer blood collection tube (Becton Dickinson)
was resuspended in PBS (5 mL) and aliquoted into microfuge tubes (100 ALper tube). Blood was collected from the tail and lysed with ACK bufferthrice. Cells were resuspended in PBS for antibody staining.
Detection of GFP+ cells in frozen tissue sections. Spleens were frozenin OCT compound (Sakura Finetek) with dry ice and stored at �80jC untilsectioning. Tissues were sectioned 6 Am thick and adhered onto Superfrost
plus glass slides (VWR). Slides were kept cold to prevent diffusion of GFP
and exposed in a closed-lid container to 37% formaldehyde vapor at �20jCfor 24 h, as first described by Jockusch et al. (31). Tissues were outlined withan ImmEdge pen (Vector Laboratories), washed with PBS, and counter-
stained with 1 Ag/mL Hoechst 33342 (Sigma) for 15 min at 37jC. Afterwashing, slides were mounted with Prolong Gold anti-fade reagent
(Molecular Probes/Invitrogen). Sections were viewed using a Leica DMIRBInverted Microscope, and images were captured with MetaMorph Imaging
System (Molecular Devices Corporation).
Tumor measurements and statistics. Statistical analyses were doneusing Microsoft Excel, StatView, and SAS 9.1. Average cell counts were
compared by Student’s t test. Tumor burden was calculated as the volume
of a sphere with radius based on the average of two diameters, D1 and D2,
measured by caliper [Volume = 4/3p((D1 + D2)/4)3]. Where possible, t tests
were used to compare the average tumor volume in treatment groups at
the end of the experiment. For experiments involving animal sacrifice,
tumor growth was compared across groups by fitting a log-linear regression
model, and differences in tumor-free mice across groups were compared byFisher’s exact test.
CTL assay. Splenocytes were incubated with mitomycin C–treated LLC/
OVA at a 10:1 ratio for 6 days with recombinant mIL-2 (10–20 units/mL) inR-10 media and then plated in 96-well round-bottom plates at the indicated
effector:target ratios. LLC/OVA and LLC targets were labeled with 51Cr
(150 AL/106 cells) and plated at 5 � 104 cells per well. Plates were incubated
at 37jC for 8 h. Supernatant was collected and added to Ready Safe Liquid8 Unpublished data.
Cancer Research
Cancer Res 2007; 67: (20). October 15, 2007 10028 www.aacrjournals.org
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
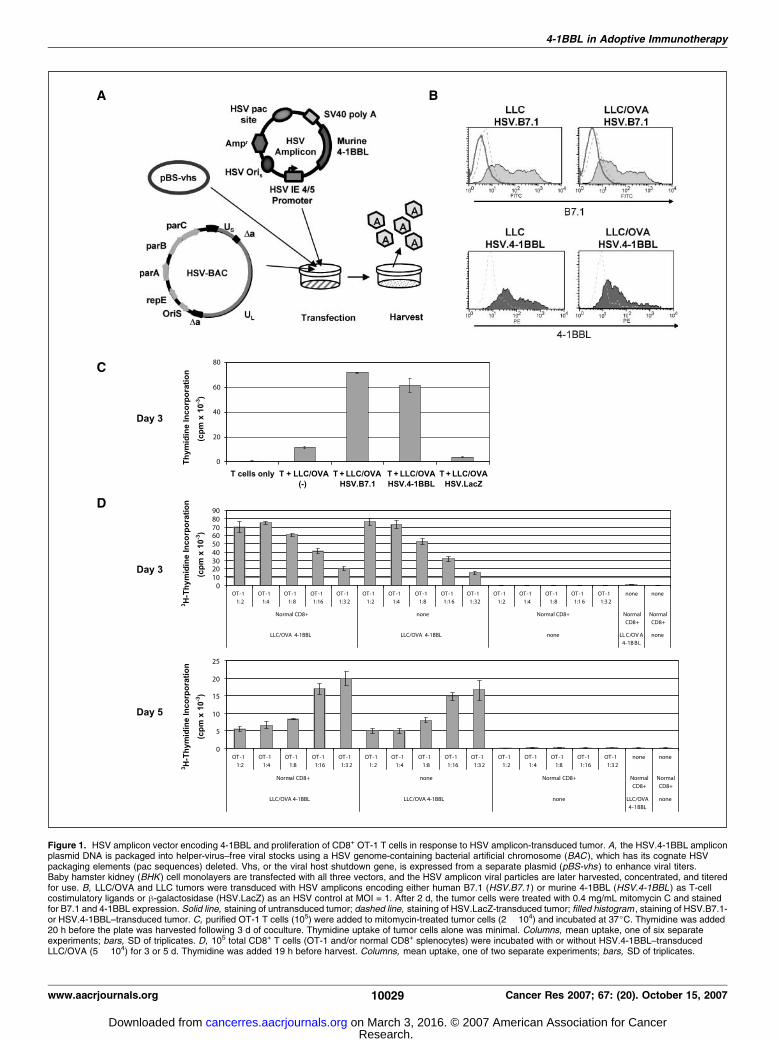
Figure 1. HSV amplicon vector encoding 4-1BBL and proliferation of CD8+ OT-1 T cells in response to HSV amplicon-transduced tumor. A, the HSV.4-1BBL ampliconplasmid DNA is packaged into helper-virus–free viral stocks using a HSV genome-containing bacterial artificial chromosome (BAC ), which has its cognate HSVpackaging elements (pac sequences) deleted. Vhs, or the viral host shutdown gene, is expressed from a separate plasmid (pBS-vhs ) to enhance viral titers.Baby hamster kidney (BHK ) cell monolayers are transfected with all three vectors, and the HSV amplicon viral particles are later harvested, concentrated, and titeredfor use. B, LLC/OVA and LLC tumors were transduced with HSV amplicons encoding either human B7.1 (HSV.B7.1 ) or murine 4-1BBL (HSV.4-1BBL ) as T-cellcostimulatory ligands or h-galactosidase (HSV.LacZ) as an HSV control at MOI = 1. After 2 d, the tumor cells were treated with 0.4 mg/mL mitomycin C and stainedfor B7.1 and 4-1BBL expression. Solid line, staining of untransduced tumor; dashed line, staining of HSV.LacZ-transduced tumor; filled histogram , staining of HSV.B7.1-or HSV.4-1BBL–transduced tumor. C, purified OT-1 T cells (105) were added to mitomycin-treated tumor cells (2 � 104) and incubated at 37jC. Thymidine was added20 h before the plate was harvested following 3 d of coculture. Thymidine uptake of tumor cells alone was minimal. Columns, mean uptake, one of six separateexperiments; bars, SD of triplicates. D, 105 total CD8+ T cells (OT-1 and/or normal CD8+ splenocytes) were incubated with or without HSV.4-1BBL–transducedLLC/OVA (5 � 104) for 3 or 5 d. Thymidine was added 19 h before harvest. Columns, mean uptake, one of two separate experiments; bars, SD of triplicates.
4-1BBL in Adoptive Immunotherapy
www.aacrjournals.org 10029 Cancer Res 2007; 67: (20). October 15, 2007
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

scintillation cocktail for aqueous samples (Beckman Coulter). Samples werecounted on a LS 6500 multipurpose scintillation counter (Beckman
Coulter). Percent lysis = (sample counts � spontaneous counts)/(maximum
counts � spontaneous counts) � 100.
In vivo bromodeoxyuridine labeling. Three and six days followingadoptive transfer of OT-1, Thy1.1+ mice were injected i.p. with 100 ALbromodeoxyuridine (BrdUrd; 1 mg; APC BrdU Flow kit, BD PharMingen).
On day 8, OT-1 numbers and incorporation of BrdUrd were assessed in the
spleen, tumor, and draining and nondraining lymph nodes by staining withanti–CD8-PE, anti–Thy1.2-FITC, and anti–BrdUrd-APC per manufacturer’s
instructions.
Results
Tumors transduced by HSV.4-1BBL express murine 4-1BBLand can induce proliferation of CD8+ OT-1 T cells. Murine4-1BBL cDNA was cloned into the HSV amplicon vectorpHSVPrPUC and packaged into amplicons (HSV.4-1BBL) using ahelper-virus–free packaging method (Fig. 1A ; refs. 29, 30). Packagedvirions contain only amplicon genomes, without the propagationof the helper virus (Fig. 1A). HSV.B7.1 and HSV.LacZ amplicons,which encode for human B7.1 and bacterial h-galactosidase,
respectively, were also packaged using the helper-free method.Both mouse and human B7.1 can stimulate T-cell CD28 receptorsof either species (24).We tested the ability of the amplicons to transduce LLC/OVA
or the parental LLC tumor cell line. LLC and LLC/OVA cellstransduced with HSV.B7.1 or HSV.4-1BBL showed high levels ofexpression of B7.1 or 4-1BBL, respectively, by day 2 as shownby flow cytometric analysis (Fig. 1B). HSV.LacZ transduction didnot induce either B7.1 or 4-1BBL expression (Fig. 1B). HSV.B7.1transduction did not induce 4-1BBL expression or vice versa (datanot shown). To more accurately follow the effects of 4-1BBL onantigen-specific T cells, we used the adoptive transfer of CD8+ OT-1T cells in our in vivo mouse experiments (32, 33). OT-1/GFP cellswere derived from TCR-transgenic OT-1 mice that were back-crossed into GFP+ mice to facilitate monitoring once adoptivelytransferred. HSV.B7.1- or HSV.4-1BBL–transduced LLC/OVA tumorcells were used to stimulate CD8+ OT-1 T cells for 3 to 5 days(Fig. 1C). At day 3, markedly increased proliferation was seen forOT-1 cells stimulated with either HSV.B7.1- or HSV.4-1BBL–transduced tumors, compared with untransduced tumor (Fig. 1C).HSV.LacZ-transduced LLC/OVA did not augment proliferation of
2–8
Figure 2. Phenotype of OT-1/GFP cellsbefore adoptive transfer and experimentaloutline. A, CD8+ OT-1/GFP T cells wereadded to mitomycin C–treated LLC/OVAtumor cells. Three days later, OT-1/GFPT cells were stained for surface activationmarkers and analyzed by flow cytometry.Filled histogram, OT-1/GFP + HSV.4-1BBL–transduced LLC/OVA; bold solid line,OT-1/GFP + HSV.B7.1-transduced LLC/OVA; dashed line, OT-1/GFP + HSV.LacZ–transduced LLC/OVA; dotted line, naiveOT-1/GFP. Data are representative of threeseparate experiments. B, LLC/OVA tumorcells were transduced with HSV.4-1BBL orHSV.B7.1 at an MOI of 1 and cultured for1 d. Tumor cells were then washed andtreated with mitomycin C. CD8+ OT-1/GFPcells were purified from spleen andcocultured with transduced tumor for3 d. Stimulated OT-1/GFP cells wereseparated from tumor cells using anti-CD8antibody-magnetic beads. OT-1/GFPcells were given i.v. into C57BL/6 micebearing LLC/OVA tumor. LLC/OVA tumorwas palpable following 2 to 4 d of s.c.injection. Peripheral blood, spleen, TDLN,and the tumor bed were examined for thepresence of OT-1/GFP cells. Spleen wasanalyzed for cytolytic activity, and thephenotype of OT-1/GFP cells wascharacterized in the spleen, TDLN, andtumor bed.
Cancer Research
Cancer Res 2007; 67: (20). October 15, 2007 10030 www.aacrjournals.org
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
OT-1 in vitro compared with untransduced LLC/OVA. OT-1stimulated with HSV.4-1BBL–transduced LLC/OVA continued toproliferate vigorously at day 5 (data not shown). OT-1 cells whichwere cultured with parental LLC, with or without costimulatoryligands, did not proliferate, signifying an absolute requirement for
signal one for T-cell activation (data not shown). OT-1 cellscultured with HSV.4-1BBL–transduced LLC/OVA did not showincreased proliferation on days 3 (Supplementary Fig. S1) or 5 (datanot shown), with the addition of soluble (2 Ag) or plate-bound(0.2 Ag) anti-CD28 antibody. Soluble anti-CD28 antibody instead
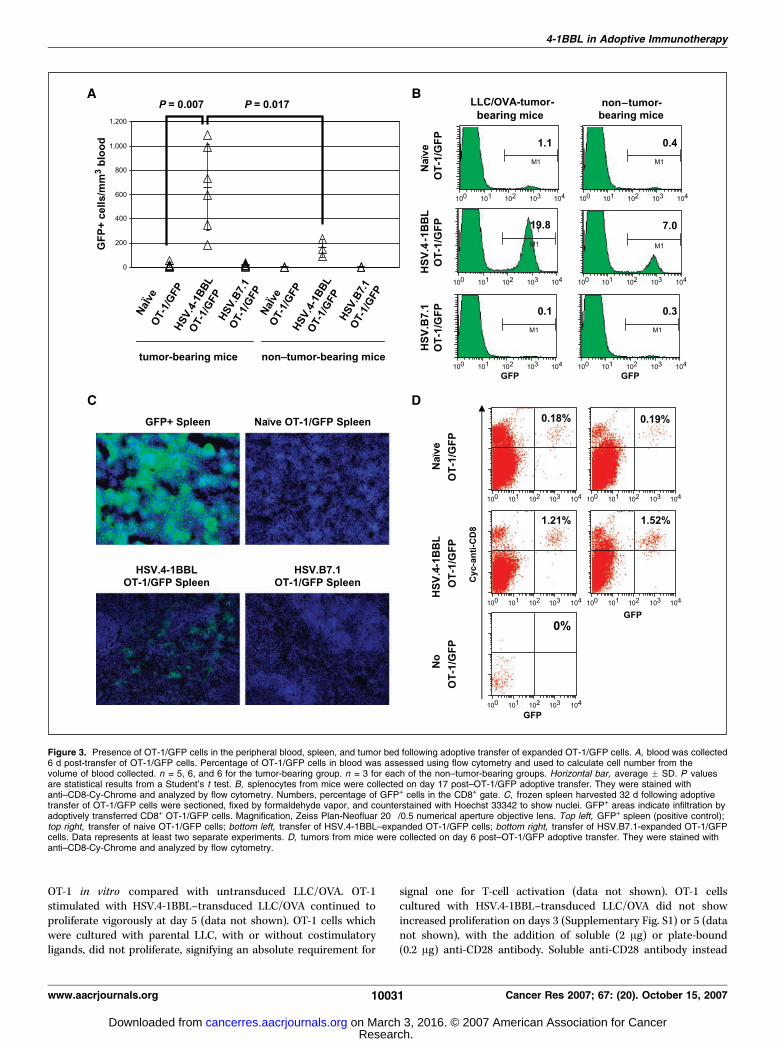
Figure 3. Presence of OT-1/GFP cells in the peripheral blood, spleen, and tumor bed following adoptive transfer of expanded OT-1/GFP cells. A, blood was collected6 d post-transfer of OT-1/GFP cells. Percentage of OT-1/GFP cells in blood was assessed using flow cytometry and used to calculate cell number from thevolume of blood collected. n = 5, 6, and 6 for the tumor-bearing group. n = 3 for each of the non–tumor-bearing groups. Horizontal bar, average F SD. P valuesare statistical results from a Student’s t test. B, splenocytes from mice were collected on day 17 post–OT-1/GFP adoptive transfer. They were stained withanti–CD8-Cy-Chrome and analyzed by flow cytometry. Numbers, percentage of GFP+ cells in the CD8+ gate. C, frozen spleen harvested 32 d following adoptivetransfer of OT-1/GFP cells were sectioned, fixed by formaldehyde vapor, and counterstained with Hoechst 33342 to show nuclei. GFP+ areas indicate infiltration byadoptively transferred CD8+ OT-1/GFP cells. Magnification, Zeiss Plan-Neofluar 20�/0.5 numerical aperture objective lens. Top left, GFP+ spleen (positive control);top right, transfer of naive OT-1/GFP cells; bottom left, transfer of HSV.4-1BBL–expanded OT-1/GFP cells; bottom right, transfer of HSV.B7.1-expanded OT-1/GFPcells. Data represents at least two separate experiments. D, tumors from mice were collected on day 6 post–OT-1/GFP adoptive transfer. They were stained withanti–CD8-Cy-Chrome and analyzed by flow cytometry.
4-1BBL in Adoptive Immunotherapy
www.aacrjournals.org 10031 Cancer Res 2007; 67: (20). October 15, 2007
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

markedly inhibited proliferation elicited by HSV.B7.1-transducedLLC/OVA, but not that elicited by HSV.4-1BBL–transduced tumor,indicating that soluble anti-CD28 antibody blocked the interactionbetween CD28 on T cells and B7.1 on the tumor cells (Supple-mentary Fig. S1). Addition of soluble anti–CTLA-4 antibody (12 Ag)to the cocultures of OT-1 cells with HSV.B7.1- or HSV.4-1BBL–transduced LLC/OVA did not further augment proliferation(Supplementary Fig. S1). Titration of OT-1 with HSV.4-1BBL–transduced LLC/OVA showed that tumor-specific OT-1 cells (105)could expand following serial dilution with normal CD8+
splenocytes (Fig. 1D). Dilutions of up to 1:32 (3,000 OT-1 cells perwell) are shown for days 3 and 5 (Fig. 1D). On day 5, OT-1 cellscontinue to proliferate at a dilution of 1:32 even when the wellswith a higher number of OT-1 have markedly decreased theirproliferation due possibly to limited nutrients or lack of intacttumor cells. Therefore, tumor-specific cells can be stimulated toproliferate using this system even when present at lower numbers.
OT-1 cells costimulated ex vivo with 4-1BBL display aneffector phenotype. Expanded OT-1/GFP cells were characterizedfollowing 3 days of coincubation with transduced LLC/OVA(Fig. 2A). CD44 is expressed on activated T cells and functions inlymphocyte homing and adhesion. CD25 (IL-2Ra) is a componentof the high-affinity IL-2 receptor up-regulated on effector T cells.Granzyme B is a serine protease stored in the granules ofCTLs along with perforin (34). CD107a (LAMP-1) is a widelyexpressed intracellular antigen that appears on CD8+ CTLsfollowing activation-induced degranulation (35). OT-1/GFP stimu-lated ex vivo with either HSV.4-1BBL– or HSV.B7.1-transducedLLC/OVA expressed high levels of CD44, intracellular granzyme B,and CD107a and modestly increased levels of CD25, indicating thatthey were activated and capable of cytotoxic activity (Fig. 2A).HSV.LacZ-stimulated OT-1/GFP expressed CD44 and CD107a atlower levels than those stimulated with HSV.4-1BBL or HSV.B7.1.4-1BB was expressed on OT-1 cells stimulated for 3 days withHSV.4-1BBL–transduced LLC/OVA, but was not detected on naiveOT-1 cells (Fig. 2A). Naive OT-1/GFP did not express any of theaforementioned activation markers.Ly-6C is a marker for previously activated Tcells andmemory CD8+
T cells (36). Expression of Ly-6C was highest in HSV.4-1BBL–
stimulated OT-1/GFP, compared with HSV.B7.1- and HSV.LacZ-stimulated OT-1/GFP and naive OT-1/GFP (Fig. 2A). Because Ly-6Ccan be up-regulated on Tcells by type I IFN secretion byHSVamplicon-transduced tumors (37), supernatant from cocultures of OT-1 andLLC/OVA transduced with HSV.4-1BBL or HSV.LacZ was collectedon days 1 to 3. IFN-a was not detected by ELISA at a detectionthreshold of 12.5 pg/mL (data not shown). More likely, IFN-g (38)produced by 4-1BBL–stimulated T cells induced Ly-6C expression.
T cells costimulated with 4-1BBL ex vivo expand in vivo inresponse to tumor. Ex vivo stimulated OT-1/GFP cells wereadoptively transferred into LLC/OVA tumor-bearing mice, andthe extent of expansion and antitumor response were measured(Fig. 2B). Mice were bled at several time points following transferof OT-1/GFP cells to detect expansion (Fig. 3A). Six days followingtransfer, the number of OT-1/GFP in the peripheral blood wassignificantly greater in the LLC/OVA tumor-bearing groupreceiving HSV.4-1BBL–stimulated OT-1/GFP cells compared withtumor-bearing groups receiving naive (P = 0.007) or HSV.B7.1-stimulated OT-1/GFP (Fig. 3A). Greater numbers of OT-1/GFP cellswere observed following transfer of 4-1BBL–stimulated OT-1/GFPcells into non–tumor-bearing mice than naive or B7.1-stimulatedOT-1/GFP cells, indicating continued proliferation of 4-1BBL–stimulated OT-1/GFP cells in vivo in the absence of tumor. Thenumber of 4-1BBL–stimulated OT-1/GFP cells was significantlygreater in the tumor-bearing mice compared with non–tumor-bearing mice (P = 0.017), suggesting that 4-1BBL-stimulated OT-1/GFP cells can respond in vivo to tumor-specific antigen (Fig. 3A).The total numbers and percentages of tumor-specific OT-1/GFP
cells in relation to the CD8+ population were determined in thespleen of tumor-bearing mice 17 days post-transfer. Representativemice are shown in Fig. 3B . The percentage of GFP+ cells in thespleen was the greatest in the tumor-bearing group given OT-1/GFP cells stimulated with HSV.4-1BBL–transduced tumor (19.8% ofCD8+ cells) versus HSV.B7.1-transduced tumor (0.1%) or naiveOT-1/GFP cells (1.1%; Fig. 3B). Substantial numbers of OT-1/GFPcells were also detected in the spleen of non–tumor-bearingmice 17 days after transfer of 4-1BBL-stimulated OT-1/GFP cells(7% of CD8+ cells), suggesting that tumor-specific 4-1BBL–stimulated T cells can also persist in vivo in the absence of antigen
Figure 4. BrdUrd incorporation byadoptively transferred OT-1. A, Thy1.1+mice were pulsed with BrdUrd on days 7 and10 and sacrificed on day 12 to determinein vivo proliferation of OT-1 CD8+ cells andtheir distribution in the spleen, TDLNs, andnon-TDLN. Numbers, percentage of BrdUrd+cells in a CD8+Thy1.2+ gate. B, the presenceof proliferating OT-1 cells in the tumor wasassessed by pulsing mice on days 3 and 6with 1 mg of BrdUrd and harvesting the tumoron day 8. Numbers indicate percentage inthe lymphocyte gate. Data are representativeof two experiments.
Cancer Research
Cancer Res 2007; 67: (20). October 15, 2007 10032 www.aacrjournals.org
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

(Fig. 3B). In addition, there were greater absolute numbers of OT-1/GFP cells present in the spleen for the 4-1BBL–stimulatedOT-1/GFP group (0.9 F 0.4 � 106 cells per spleen) versus B7.1-stimulated (0.9F 0.7� 105) or naive OT-1/GFP groups (1F 0.3� 105).Spleens were sectioned and examined for GFP+ cells 32 days
following adoptive transfer of OT-1/GFP. Spleens from mice givenHSV.4-1BBL–stimulated OT-1/GFP cells showed increased infiltrationwith GFP+ cells (Fig. 3C). Analysis by flow cytometry indicated that9.4% of the CD8+ splenocytes in the HSV.4-1BBL–stimulated OT-1/GFPgroupwereGFP+, comparedwith 0.1% and 0.2%GFP+ in the naiveOT-1/GFP and HSV.B7.1-stimulated OT-1/GFP groups, respectively.These results indicate that tumor-specific T cells stimulated ex vivowith HSV.4-1BBL–transduced tumor could expand and persist in vivo .The tumor bed was also analyzed for the presence of OT-1/GFP
cells. Six days post-transfer, flow cytometry analysis of dispersedtumor showed that the percentage of GFP+ cells was 7- to 8-foldgreater in mice that received the 4-1BBL–stimulated OT-1/GFPcells compared with those receiving naive cells (Fig. 3D). Later timepoints could not be examined due to tumor regression in theHSV.4-1BBL–stimulated OT-1/GFP group.
BrdUrd is incorporated by adoptively transferred cells. Todetermine whether adoptively transferred OT-1 divided in vivo ,we characterized the BrdUrd uptake of transferred cells in separateexperiments. Thy1.2+ OT-1 cells were transferred into Thy1.1+ mice,which were pulsed with BrdUrd on days 7 and 10. Mice weresacrificed on day 12 to determine in vivo proliferation of OT-1 CD8+
cells and their distribution in the spleen, TDLNs, and non-TDLNs(Fig. 4A). Significantly higher percentages of OT-1 cells that hadincorporated BrdUrd were present in the TDLNs in the 4-1BBLgroup compared with the naive OT-1 group (Fig. 4A).The presence of proliferating OT-1 cells in the tumor bed was
assessed by pulsing mice on days 3 and 6 with 1 mg of BrdUrd andharvesting the tumor on day 8. Overall, more Thy1.2+ OT-1 cellswere present in the 4-1BBL–stimulated group as compared with thenaive OT-1 or B7.1-stimulated group, and these cells had incorpo-rated BrdUrd (Fig. 4B).
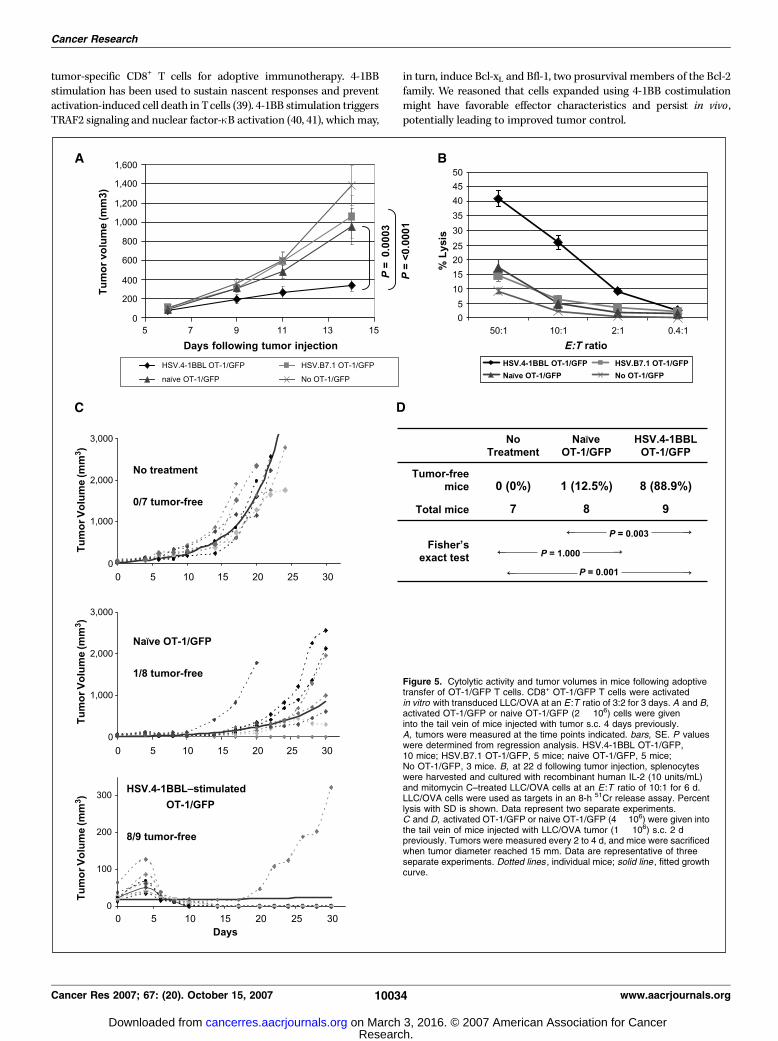
CD8+ T cells expanded in vitro with 4-1BBL possess cytolyticactivity and markedly decrease tumor growth. We measuredCTL activity from splenocytes of mice that received 2 � 106 OT-1/GFP cells by adoptive transfer. LLC/OVA tumor-bearing micetreated with HSV.4-1BBL–activated OT-1/GFP cells harbored lowertumor burden at day 14 than untreated mice (P < 0.0001) or micetreated with an identical number of HSV.B7.1-stimulated OT-1/GFP(P < 0.0001) or naive OT-1/GFP cells (P = 0.0003; Fig. 5A). NaiveOT-1/GFP treatment did not reduce tumor burden significantlywhen compared with the untreated group (P > 0.05). Administra-tion of HSV.B7.1-activated OT-1/GFP cells did not have majorinhibitory effects on tumor size when compared with no treatmentor to naive OT-1/GFP transfer (P > 0.05; Fig. 5A).Splenocytes from the HSV.4-1BBL–activated OT-1/GFP group
showed substantially higher CTL activity against LLC/OVAcompared with splenocytes from the HSV.B7.1-stimulated OT-1/GFP, naive OT-1/GFP, and untreated groups (Fig. 5B). These resultssuggest that T cells expanded with HSV.4-1BBL–transduced tumorcells may have more favorable effector characteristics than thoseobtained through HSV.B7.1-mediated stimulation.In a second experiment, 4 � 106 cells OT-1/GFP cells, either
naive or stimulated with HSV.4-1BBL–transduced tumor, weretransferred into LLC/OVA tumor-bearing mice to determine theeffects on tumor volume (Fig. 5C). Transfer of HSV.4-1BBL–stimulated OT-1/GFP resulted in a statistically significant decrease
in tumor growth compared with no treatment (P < 0.001) or tonaive OT-1/GFP transfer (P < 0.001; Fig. 5C). The estimated rate ofgrowth for the HSV.4-1BBL OT-1/GFP group from the regressionmodel was significantly lower than that for the naive OT-1/GFPgroup (0.013 versus 0.130; P < 0.001). By day 30, 88.9% (8/9) mice inthe HSV.4-1BBL OT-1/GFP group were tumor-free, compared with0% (0/7) tumor-free in the no-treatment group and 12.5% (1/8)tumor-free in the naive OT-1/GFP group. Analysis using Fisher’sexact test show that these differences between the HSV.4-1BBL OT-1/GFP group and the no-treatment group (P = 0.001) or the naive OT-1/GFP group (P = 0.003) were statistically significant (Fig. 5D).
4-1BBL–stimulated CD8+ OT-1 cells display memory pheno-typic characteristics. The phenotype of OT-1/GFP cells in eachgroup was characterized post-adoptive transfer (Fig. 6). On day 6,GFP+ cells in the spleen and tumor bed were analyzed for Ly-6C(Fig. 6A). The HSV.4-1BBL–stimulated OT-1/GFP group had agreater percentage of Ly-6C+GFP+ cells present in the spleen andtumor bed than the naive OT-1/GFP group. On day 17, OT-1/GFP+
cells in tumor-bearing and non–tumor-bearing mice were studiedfor levels of CD44, Ly-6C, and CD62L (L-selectin; Fig. 6B andSupplementary Fig. S2). Significantly higher levels of CD44+ OT-1/GFP were observed in the splenocytes of mice that had receivedHSV.4-1BBL–stimulated OT-1/GFP (12.7% in tumor-bearing mice,3.8% in non–tumor-bearing mice) than were seen for eitherHSV.B7.1-stimulated (0.1% in tumor-bearing and non–tumor-bearing mice) or naive OT-1/GFP transfer (0.4% in tumor-bearingmice, 0.1% in non–tumor-bearing mice; Fig. 6B and SupplementaryFig. S2). HSV.4-1BBL–stimulated OT-1/GFP cells continued to showhigher levels of Ly-6C expression on day 17 in comparison to naiveor HSV.B7.1-stimulated OT-1/GFP transferred cells (Fig. 6B andSupplementary Fig. S2). Further analysis showed that OT-1/GFPcells persisting in the spleen were CD62L negative (Fig. 6B andSupplementary Fig. S2), as well as CD25 negative and CD127(IL-7Ra) negative (data not shown). Similar characteristics,indicating the generation of effector memory cells, were observedin OT-1/GFP cells present in the spleen at day 32 (data not shown).This indicates that upon adoptive transfer of 4-1BBL–stimulatedT cells, the T cells persist at high levels and exhibit phenotypicattributes of effector memory cells.To determine whether this strategy could be used in a
nontransgenic setting, we inoculated GFP+ mice with LLC/OVAs.c. for 8 days and then harvested spleens and TDLNs. CD8+ T cellsin the spleen or lymphocytes from TDLNs were stimulated ex vivowith untransduced LLC/OVA or LLC/OVA transduced with HSV.4-1BBL. Increased Ly-6C expression was observed in vitro on theCD8+ splenocytes and, to a lesser degree, on the CD8+ TDLN cellsfollowing 5 days of cultivation with HSV.4-1BBL–transduced LLC/OVA compared with coculture with untransduced tumor (data notshown). Although 4-1BBL–transduced LLC/OVA induced prolifer-ation and modest expansion of GFP+ lymphocytes in vitro , levelswere f20-fold lower than found with OT-1, preventing adequateharvest for adoptive transfer. Adoptive transfer of 0.5 � 106
cocultured T cells showed modest effects on tumor growth. Thismay be due to low precursor frequency and reduced expansion.Therefore, further optimization of the ability to generate tumor-specific T cells and/or modification of transferred dose and doseschedules will be necessary to improve tumor control.
Discussion
The overall purpose of these studies was to understand thepotential utility of 4-1BB as a means of expanding and activating
4-1BBL in Adoptive Immunotherapy
www.aacrjournals.org 10033 Cancer Res 2007; 67: (20). October 15, 2007
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
tumor-specific CD8+ T cells for adoptive immunotherapy. 4-1BBstimulation has been used to sustain nascent responses and preventactivation-induced cell death in Tcells (39). 4-1BB stimulation triggersTRAF2 signaling and nuclear factor-nB activation (40, 41), whichmay,
in turn, induce Bcl-xL and Bfl-1, two prosurvival members of the Bcl-2family. We reasoned that cells expanded using 4-1BB costimulationmight have favorable effector characteristics and persist in vivo ,potentially leading to improved tumor control.
Figure 5. Cytolytic activity and tumor volumes in mice following adoptivetransfer of OT-1/GFP T cells. CD8+ OT-1/GFP T cells were activatedin vitro with transduced LLC/OVA at an E :T ratio of 3:2 for 3 days. A and B,activated OT-1/GFP or naive OT-1/GFP (2 � 106) cells were giveninto the tail vein of mice injected with tumor s.c. 4 days previously.A, tumors were measured at the time points indicated. bars, SE. P valueswere determined from regression analysis. HSV.4-1BBL OT-1/GFP,10 mice; HSV.B7.1 OT-1/GFP, 5 mice; naive OT-1/GFP, 5 mice;No OT-1/GFP, 3 mice. B, at 22 d following tumor injection, splenocyteswere harvested and cultured with recombinant human IL-2 (10 units/mL)and mitomycin C–treated LLC/OVA cells at an E :T ratio of 10:1 for 6 d.LLC/OVA cells were used as targets in an 8-h 51Cr release assay. Percentlysis with SD is shown. Data represent two separate experiments.C and D, activated OT-1/GFP or naive OT-1/GFP (4 � 106) were given intothe tail vein of mice injected with LLC/OVA tumor (1 � 106) s.c. 2 dpreviously. Tumors were measured every 2 to 4 d, and mice were sacrificedwhen tumor diameter reached 15 mm. Data are representative of threeseparate experiments. Dotted lines , individual mice; solid line , fitted growthcurve.
Cancer Research
Cancer Res 2007; 67: (20). October 15, 2007 10034 www.aacrjournals.org
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

HSV.4-1BBL amplicons were used to transduce LLC/OVA tumorsin vitro for purposes of ex vivo expansion of tumor-specific OT-1T cells. OT-1 cells responded to coculture with HSV.4-1BBL–transduced LLC/OVA by proliferating and expressing activationmarkers, namely, CD44, CD25, Ly-6C, CD107a, 4-1BB, and intra-cellular granzyme B, indicating priming and differentiation intocytolytic effectors.Adoptive transfer of HSV.4-1BBL–stimulated OT-1/GFP T cells
significantly conferred greater protection against LLC/OVA growthcompared with naive or HSV.B7.1-stimulated OT-1/GFP cells. Micetreated with HSV.4-1BBL–expanded OT-1/GFP cells exhibitedgreater CTL activity and showed higher percentages of tumor-specific OT-1/GFP+ cells in the spleen and tumor bed. FollowingHSV.4-1BBL stimulation, CD8+ OT-1 T cells expanded in vivo ,incorporated BrdUrd, and expressed high levels of CD44 and theCD8+ memory marker Ly-6C. Ly-6C has been shown to regulate
homing of CD8+ T cells to lymph nodes (42) and perhaps augmentsthe homing of transferred HSV.4-1BBL–stimulated OT-1/GFP cellsto secondary lymphoid organs, where they can be found on days 6,17, and 32 post-transfer. Most transferred OT-1/GFP cells wereCD62L negative even after 20 days post-tumor eradication in thehost, indicating, in combination with Ly-6C expression, an effectormemory phenotype. Perhaps, longer observations can show theconversion of tumor-specific cells to CD44hiLy-6ChiCD62Lhi centralmemory cells. In these studies, we show that T-cell costimulationwith 4-1BBL expressed on tumor cells may be useful in facilitatingthe expansion of tumor-specific T cells in vivo as well as in vitro .Other groups have examined the possibility of using 4-1BB
costimulation for the generation of tumor-reactive T cells foradoptive immunotherapy. Addition of an agonistic anti–4-1BBantibody to in vitro cultures of TDLN cells and anti-CD3/anti-CD28antibodies enhanced expansion, production of type 1 cytokines,
Figure 6. CD44, Ly-6C, and CD62Lexpression of OT-1/GFP in splenocytes andtumor. Splenocytes from mice were collectedon day 6 (A ) or day 17 (B) following OT-1/GFP adoptive transfer. Tumor was alsoanalyzed on day 6 (A). Cells were stainedwith anti–CD8-Cy-Chrome and biotinylatedanti-Ly-6C followed by streptavidin-PE(A and B ), anti-CD44-PE (B), or anti-CD62L-PE (B) and analyzed by flow cytometry.Numbers indicate percentages in thelymphocyte gate in A and the CD8+ gatein B . Data represent one of three experimentswith similar results.
4-1BBL in Adoptive Immunotherapy
www.aacrjournals.org 10035 Cancer Res 2007; 67: (20). October 15, 2007
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

and survival of T cells (43). When anti-CD3/anti-CD28/anti–4-1BB–expanded TDLN cells were adoptively transferred into MCA 205tumor-bearing mice, significantly fewer metastatic lesions andprolonged survival of mice were observed compared with TDLNcells stimulated without anti–4-1BB (43). Strome et al. (16) alsoobserved that the combined use of anti-CD3/anti-CD28/anti–4-1BBin activating T cells for adoptive immunotherapy resulted in thegeneration of T cells that were more effective than those activatedby anti-CD3 alone or anti-CD3/anti-CD28 in mediating antitumorreactivity.However, when groups used anti-CD3/anti–4-1BB antibodies,
without anti-CD28 antibody, to expand a polyclonal T-cellpopulation in culture, they were not successful in generatingtumor-reactive T cells due to nonspecific expansion (16, 17). Ourstudies indicate that it may be possible to specifically generatetumor-reactive clones, which can persist in vivo , using HSV.4-1BBL–transduced tumor cells in the absence of additional CD28costimulation. Whether the low levels of B7.1 present on LLC/OVAtumor are adequate for CD28 stimulation to work in combinationwith 4-1BBL costimulation is not known (44).Maus et al. (11) have shown ex vivo expansion of human
polyclonal and MHC tetramer-sorted antigen-specific CTL usingartificial antigen presenting cells, specifically K562 erythromyeloidcell lines stably transfected to express 4-1BBL and the Fcg receptorCD32 to bind anti-CD3 and anti-CD28 antibodies on the surface.In contrast to the work by Maus et al., the highly efficient HSVamplicon system provides a theoretical means by which to selec-tively expand tumor-specific effector populations using autologoustumor from patients without the need for presorting for tumor-reactive T cells.In addition to 4-1BB costimulation, HSV.4-1BBL amplicon
transduction of tumor may provide other stimuli that facilitatethe generation of effector cells. HSV amplicons can impart astrong innate response to transduced cells, including macrophagecell lines and human CLL cells, resulting in cytokine secretion andNKG2D-L expression by the transduced cells (23). HSV possessesat least three molecular components capable of activating theinnate immune system: (1) dsRNA generated through self-hybridization of viral genes transcribed from complementaryDNA strands (45); (2) envelope glycoproteins recognized by TLR2(46); and (3) unmethylated CpG motifs encoded in the viralgenome that activate TLR9 (47). Due to the fact that HSVamplicon DNA is concatamerized, CpG effects on TLR9 may bequite potent. The enhanced capacity of transduced tumors to
stimulate an innate immune response may lead to an improvedadaptive response.We did not test whether HSV.B7.1 will further augment effects
seen with HSV.4-1BBL stimulation. HSV.B7.1-stimulated OT-1failed to expand in vivo and inhibit tumor growth. Because B7.1serves as a ligand for both CD28 and CTLA-4, B7.1 may have alsobound to CTLA-4 expressed on activated OT-1 cells, inhibitingexpansion and survival. We, however, did not observe greaterproliferation in the presence of soluble anti–CTLA-4 antibody(Supplementary Fig. S1).In summary, our studies suggest that costimulation with 4-1BBL
may be employed to enhance expansion and cytolytic activity oftumor-specific CD8+ T cells for the generation of tumor-specificimmunity. We used the adoptive transfer of TCR-transgenic OT-1/GFP T cells to more accurately follow the effects of 4-1BBL onT cells that were specific for defined tumor-related antigens in ourin vivo mouse experiments, because antigen-specific CD8+ T cellsare normally present in very low numbers (32, 33) Although theOT-1 model was useful in demonstrating expansion of a tumor-specific response, results obtained with OT-1 may differ ascompared with what would be observed using a polyclonal T cellpopulation as a source of expanded T cells. Such a populationmight contain relatively fewer precursor T cells with antitumoractivity, and expansion of relevant effectors might prove moredifficult. Nevertheless, the vigorous cytolytic effector function aswell as the increased expansion and persistence seen using HSVamplicon-transduced tumor suggest that this method should beexplored further and may be potentially applicable in the humansetting. Because HSV vectors are theoretically safe and a highlyefficient means of gene transfer, the laboratory is pursuingpreclinical development of these vectors for potential human usein CLL. Further optimization may allow for efficient expansion ofrelatively rare precursor antitumor T cells.
Acknowledgments
Received 6/30/2006; revised 7/10/2007; accepted 8/1/2007.Grant support: NIH Grants RO1 CA87978, RO1 CA74273, and PO1 CA109094, the
Leukemia and Lymphoma Society of America, and the Rochester Nathan ShockCenter.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Ann Casey and Wade Narrow at the University of Rochester for HSVamplicon packaging and the Imaging Core supported by the University of Miami/Sylvester Cancer Center, and Dr. Seung-Uon Shin, Dr. Hyun-Mi Cho, and Dr. PiaChallita-Eid for their critiques and collaboration.
References1. Dudley ME, Wunderlich JR, Yang JC, et al. Adoptivecell transfer therapy following non-myeloablative butlymphodepleting chemotherapy for the treatment ofpatients with refractory metastatic melanoma. J ClinOncol 2005;23:2346–57.
2. Kawai K, Saijo K, Oikawa T, et al. Clinical course andimmune response of a renal cell carcinoma patient toadoptive transfer of autologous cytotoxic T lympho-cytes. Clin Exp Immunol 2003;134:264–9.
3. Tsuboi K, Saijo K, Ishikawa E, et al. Effects of localinjection of ex vivo expanded autologous tumor-specificT lymphocytes in cases with recurrent malignantgliomas. Clin Cancer Res 2003;9:3294–302.
4. Kwon BS, Kozak CA, Kim KK, Pickard RT. Genomicorganization and chromosomal localization of the T-cellantigen 4-1BB. J Immunol 1994;152:2256–62.
5. Pollok KE, Kim YJ, Zhou Z, et al. Inducible T cellantigen 4-1BB. Analysis of expression and function.J Immunol 1993;150:771–81.
6. Futagawa T, Akiba H, Kodama T, et al. Expression andfunction of 4-1BB and 4-1BB ligand on murine dendriticcells. Int Immunol 2002;14:275–86.
7. Hurtado JC, Kim SH, Pollok KE, Lee ZH, Kwon BS.Potential role of 4-1BB in T cell activation. Comparisonwith the costimulatory molecule CD28. J Immunol 1995;155:3360–7.
8. Cooper D, Bansal-Pakala P, Croft M. 4-1BB (CD137)controls the clonal expansion and survival of CD8 T cellsin vivo but does not contribute to the development ofcytotoxicity. Eur J Immunol 2002;32:521–9.
9. Shuford WW, Klussman K, Tritchler DD, et al.4-1BB costimulatory signals preferentially induce CD8+
T cell proliferation and lead to the amplification in vivoof cytotoxic T cell responses. J Exp Med 1997;186:47–55.
10. Wilcox RA, Flies DB, Wang H, et al. Impairedinfiltration of tumor-specific cytolytic T cells in theabsence of interferon-g despite their normal maturationin lymphoid organs during CD137 monoclonal antibodytherapy. Cancer Res 2002;62:4413–8.
11. Maus MV, Thomas AK, Leonard DG, et al. Ex vivoexpansion of polyclonal and antigen-specific cytotoxicT lymphocytes by artificial APCs expressing ligands forthe T-cell receptor, CD28 and 4-1BB. Nat Biotechnol2002;20:143–8.
12. Cannons JL, Lau P, Ghumman B, et al. 4-1BB ligandinduces cell division, sustains survival, and enhanceseffector function of CD4 and CD8 T cells with similarefficacy. J Immunol 2001;167:1313–24.
13. Bertram EM, Lau P, Watts TH. Temporal segregationof 4-1BB versus CD28-mediated costimulation: 4-1BBligand influences T cell numbers late in the primaryresponse and regulates the size of the T cell memory
Cancer Research
Cancer Res 2007; 67: (20). October 15, 2007 10036 www.aacrjournals.org
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

response following influenza infection. J Immunol 2002;168:3777–85.
14. Melero I, Shuford WW, Newby SA, et al. Monoclonalantibodies against the 4-1BB T-cell activation moleculeeradicate established tumors. Nat Med 1997;3:682–5.
15. May KF, Jr., Chen L, Zheng P, Liu Y. Anti–4-1BBmonoclonal antibody enhances rejection of large tumorburden by promoting survival but not clonal expansionof tumor-specific CD8+ T cells. Cancer Res 2002;62:3459–65.
16. Strome SE, Martin B, Flies D, et al. Enhancedtherapeutic potential of adoptive immunotherapy byin vitro CD28/4-1BB costimulation of tumor-reactiveT cells against a poorly immunogenic, major histocom-patibility complex class I–negative A9P melanoma.J Immunother 2000;23:430–7.
17. Kim JA, Averbook BJ, Chambers K, et al. Divergenteffects of 4-1BB antibodies on antitumor immunity andon tumor-reactive T-cell generation. Cancer Res 2001;61:2031–7.
18. Takahashi C, Mittler RS, Vella AT. Cutting edge: 4-1BB is a bona fide CD8 T cell survival signal. J Immunol1999;162:5037–40.
19. Melero I, Bach N, Hellstrom KE, Aruffo A, Mittler RS,Chen L. Amplification of tumor immunity by genetransfer of the co-stimulatory 4-1BB ligand: synergy withthe CD28 co-stimulatory pathway. Eur J Immunol 1998;28:1116–21.
20. Guinn BA, DeBenedette MA, Watts TH, BerinsteinNL. 4-1BBL cooperates with B7-1 and B7-2 in convertinga B cell lymphoma cell line into a long-lasting antitumorvaccine. J Immunol 1999;162:5003–10.
21. Mogi S, Sakurai J, Kohsaka T, et al. Tumour rejectionby gene transfer of 4-1BB ligand into a CD80(+) murinesquamous cell carcinoma and the requirements of co-stimulatory molecules on tumour and host cells.Immunology 2000;101:541–7.
22. Martinet O, Ermekova V, Qiao JQ, et al. Immuno-modulatory gene therapy with interleukin 12 and 4-1BBligand: long-term remission of liver metastases in amouse model. J Natl Cancer Inst 2000;92:931–6.
23. Tolba KA, Bowers WJ, Hilchey SP, et al. Develop-ment of herpes simplex virus-1 amplicon-basedimmunotherapy for chronic lymphocytic leukemia.Blood 2001;98:287–95.
24. Ikawa M, Yamada S, Nakanishi T, Okabe M. ‘‘Greenmice’’ and their potential usage in biological research.FEBS Lett 1998;430:83–7.
25. Strbo N, Oizumi S, Sotosek-Tokmadzic V, Podack ER.Perforin is required for innate and adaptive immunityinduced by heat shock protein gp96. Immunity 2003;18:381–90.
26. Goodwin RG, Din WS, Davis-Smith T, et al. Molecularcloning of a ligand for the inducible T cell gene 4-1BB: amember of an emerging family of cytokineswith homologyto tumor necrosis factor. Eur J Immunol 1993;23:2631–41.
27. Geller AI, Breakefield XO. A defective HSV-1 vectorexpresses Escherichia coli h-galactosidase in culturedperipheral neurons. Science 1988;241:1667–9.
28. Kutubuddin M, Federoff HJ, Challita-Eid PM, et al.Eradication of pre-established lymphoma using herpessimplex virus amplicon vectors. Blood 1999;93:643–54.
29. Stavropoulos TA, Strathdee CA. An enhanced pack-aging system for helper-dependent herpes simplex virusvectors. J Virol 1998;72:7137–43.
30. Bowers WJ, Howard DF, Brooks AI, Halterman MW,Federoff HJ. Expression of vhs and VP16 during HSV-1helper virus-free amplicon packaging enhances titers.Gene Ther 2001;8:111–20.
31. Jockusch H, Voigt S, Eberhard D. Localization ofGFP in frozen sections from unfixed mouse tissues:immobilization of a highly soluble marker protein byformaldehyde vapor. J HistochemCytochem 2003;51:401–4.
32. Kearney ER, Pape KA, Loh DY, Jenkins MK.Visualization of peptide-specific T cell immunity andperipheral tolerance induction in vivo . Immunity 1994;1:327–39.
33. Pape KA, Kearney ER, Khoruts A, et al. Use ofadoptive transfer of T-cell–antigen-receptor-transgenicT cell for the study of T-cell activation in vivo . ImmunolRev 1997;156:67–78.
34. MassonD, Tschopp J. A family of serine esterases in lyticgranules of cytolytic T lymphocytes. Cell 1987;49:679–85.
35. Betts MR, Brenchley JM, Price DA, et al. Sensitive andviable identification of antigen-specific CD8+ T cells bya flow cytometric assay for degranulation. J ImmunolMethods 2003;281:65–78.
36. Walunas TL, Bruce DS, Dustin L, Loh DY, BluestoneJA. Ly-6C is a marker of memory CD8+ T cells. JImmunol 1995;155:1873–83.
37. Schlueter AJ, Krieg AM, de Vries P, Li X. Type I interferonis the primary regulator of inducible Ly-6C expression onT cells. J Interferon Cytokine Res 2001;21:621–9.
38. LeClair KP, Bridgett MM, Dumont FJ, Palfree RG,Hammerling U, Bothwell AL. Kinetic analysis ofLy-6 gene induction in a T lymphoma by interfer-ons and interleukin 1, and demonstration of Ly-6inducibility in diverse cell types. Eur J Immunol 1989;19:1233–9.
39. Hurtado JC, Kim YJ, Kwon BS. Signals through 4-1BBare costimulatory to previously activated splenic T cellsand inhibit activation-induced cell death. J Immunol1997;158:2600–9.
40. Saoulli K, Lee SY, Cannons JL, et al. CD28-indepen-dent, TRAF2-dependent costimulation of resting T cellsby 4-1BB ligand. J Exp Med 1998;187:1849–62.
41. Lee HW, Park SJ, Choi BK, Kim HH, Nam KO, KwonBS. 4-1BB promotes the survival of CD8+ T lymphocytesby increasing expression of Bcl-xL and Bfl-1. J Immunol2002;169:4882–8.
42. Hanninen A, Jaakkola I, Salmi M, Simell O, JalkanenS. Ly-6C regulates endothelial adhesion and homingof CD8(+) T cells by activating integrin-dependentadhesion pathways. Proc Natl Acad Sci U S A 1997;94:6898–903.
43. Li Q, Carr A, Ito F, Teitz-Tennenbaum S, Chang AE.Polarization effects of 4-1BB during CD28 costimulationin generating tumor-reactive T cells for cancer immu-notherapy. Cancer Res 2003;63:2546–52.
44. Tirapu I, Huarte E, Guiducci C, et al. Low surfaceexpression of B7-1 (CD80) is an immunoescapemechanism of colon carcinoma. Cancer Res 2006;66:2442–50.
45. Jacquemont B, Roizman B. RNA synthesis in cellsinfected with herpes simplex virus. X. Properties of viralsymmetric transcripts and of double-stranded RNAprepared from them. J Virol 1975;15:707–13.
46. Aravalli RN, Hu S, Rowen TN, Palmquist JM,Lokensgard JR. Cutting edge: TLR2-mediated proin-flammatory cytokine and chemokine production bymicroglial cells in response to herpes simplex virus.J Immunol 2005;175:4189–93.
47. Lundberg P, Welander P, Han X, Cantin E. Herpessimplex virus type 1 DNA is immunostimulatory in vitroand in vivo . J Virol 2003;77:11158–69.
4-1BBL in Adoptive Immunotherapy
www.aacrjournals.org 10037 Cancer Res 2007; 67: (20). October 15, 2007
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

2007;67:10027-10037. Cancer Res Kyung H. Yi, Hovav Nechushtan, William J. Bowers, et al. vivo
In4-1BBL Persist in the Host and Show Antitumor Activity Using Herpes Simplex Virus Amplicons Encodingvivo
ExAdoptively Transferred Tumor-Specific T Cells Stimulated
Updated version
http://cancerres.aacrjournals.org/content/67/20/10027
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2007/10/03/67.20.10027.DC1.html
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/67/20/10027.full.html#ref-list-1
This article cites 47 articles, 30 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/67/20/10027.full.html#related-urls
This article has been cited by 3 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on March 3, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from