
Continuous Positive Airway Pressure and Noninvasive Ventilation
Upload
independentCategory
view
1download
0
0041-1337/01/7107-966/0TRANSPLANTATION Vol. 71, 966–976, No. 7, April 15, 2001Copyright © 2001 by Lippincott Williams & Wilkins, Inc. Printed in U.S.A.
ACTIVATION OF HUMAN AIRWAY EPITHELIAL CELLS BY NON-HLA ANTIBODIES DEVELOPED AFTER LUNG
TRANSPLANTATION: A POTENTIAL ETIOLOGICAL FACTOR FORBRONCHIOLITIS OBLITERANS SYNDROME1
ANDRES JARAMILLO,2 BASHOO NAZIRUDDIN,2 LEIYING ZHANG,2 SCOTT I. REZNIK,2 MICHAEL A. SMITH,2
AVIVA A. ALOUSH,2 ELBERT P. TRULOCK,3 G. ALEXANDER PATTERSON, 2 AND T. MOHANAKUMAR2,4,5
Departments of Surgery, Medicine, and Pathology and Immunology, Washington University School of Medicine,St. Louis, MO 63110-1093
Background. The main cause of morbidity and mor-tality after lung transplantation (LT) is bronchiolitisobliterans syndrome (BOS). Anti-HLA antibodies de-velopment after LT has been shown to play an impor-tant role in BOS pathogenesis. However, the nature ofnon-HLA antibodies developed after LT and their rolein BOS pathogenesis have not been determined.
Methods. Sera from 16 BOS1 patients and 11 BOS2patients were collected at 12, 24, 36, and 48 monthsafter LT. Anti-HLA class I and class II antibodies wereabsorbed with pooled human platelets and pooled hu-man lymphoblastoid cell lines, respectively. Then, thepresence of non-HLA antibodies against several celllines from different origin was determined by flowcytometric analysis. Antibody-positive samples weretested for induction of proliferation and growth factorproduction in two selected airway epithelial cell(AEC) lines.
Results. Five of 16 BOS1 patients (31.2%) and 0 of 11BOS- patients (0%) developed anti-AEC antibodies af-ter LT (P50.05). No reactivity against endothelialcells, lymphocytes, monocytes, or granulocytes wasdetected. Further analysis of two selected sera demon-strated the development of reactivity against a 60-kDaantigen expressed by 60% of AEC lines and only 12% ofcell lines from other tissues. Antibody binding to thisantigen induced intracellular Ca11 influx, tyrosinephosphorylation, proliferation, and up-regulation oftransforming growth factor-b and heparin-bindingepidermal growth factor mRNA transcription in AECs.
Conclusions. These results indicate that anti-AECantibodies may play a role in the immunopathogenesisof BOS in the absence of anti-HLA antibodies.
INTRODUCTION
The main cause of long-term morbidity and mortality afterlung transplantation (LT) is the development of bronchiolitisobliterans syndrome (BOS), which is considered to representchronic lung allograft rejection (1). BOS is a fibroproliferativeprocess identified histologically by inflammation and fibrosisof the lamina propria and lumen and clinically by progressiveairflow obstruction (2). The specific etiology and pathogene-sis of BOS are still unclear. However, a growing body ofevidence suggests that BOS is caused by the immune re-sponse developed against antigenic determinants expressedby airway epithelial cells (AEC) and endothelial cells (EC) ofthe lung allograft (3). In fact, it has been shown that en-hanced immunosuppression slows the progression of the dis-ease (4, 5).
Previous studies have shown that AECs are immunologicaltargets during lung allograft rejection (6–8). Interestingly,multiple studies have shown that, upon activation, epithelialcells from different tissues have the ability to produce highquantities of several growth factors including epidermalgrowth factor (EGF) (9), heparin-binding EGF (HB-EGF)(10), endothelin (ET)-1 (11), basic fibroblast growth factor(bFGF) (10, 12), granulocyte-monocyte colony-stimulatingfactor (GM-CSF) (13), insulin-like growth factor (IGF)-1 (14),platelet-derived growth factor (PDGF) (10, 12), and trans-forming growth factor (TGF)-b (10, 12). All these growthfactors are involved in the activation of proliferation of fibro-blasts and smooth muscle cells (14–22).
Growth factor-mediated proliferation of fibrous tissue, pre-sumably in response to acute and chronic lung allograft in-jury, leads to airway constriction and luminal obliteration asBOS develops. The specific growth factors and their levels ofexpression involved in the regulation of the tissue remodel-ing observed during the course of BOS are still unknown.Heterotopic allogeneic transplantation of mouse or rat tra-cheas reveal fibroproliferation with pathology identical toBOS although similar isograft developed the same lesion onlywhen exposed to exogenous PDGF or bFGF (23, 24). OnlyPDGF and TGF-b have been found to be increased during theprocess of BOS development after LT in humans (25–27).ET-1 has been found to be steadily increased in serial bron-choalveolar lavage samples collected from LT patients (28).However, there is no information regarding the levels of thisgrowth factor before or at the time of BOS development.
1 This work was supported by the National Institute of HealthGrant No. HL56643. AJ is a recipient of the National ResearchService Award AI07163; SIR is a recipient of the National ResearchService Award HL07776; MAS is a recipient of the National Re-search Service Award HL09557; and TM is a recipient of the Jac-queline G. and William E. Maritz endowed Professorship in Immu-nology and Oncology.
2 Department of Surgery.3 Department of Medicine.4 Department of Pathology and Immunology.5 Address correspondence to: T. Mohanakumar, PhD, Department
of Surgery, Washington University School of Medicine, Box 8109-3328.CSRB, 660 South Euclid Avenue, St. Louis, MO 63110-1093.
966

Development of anti-HLA antibodies after LT has beenidentified as one of the major predisposing factors for BOS(29, 30). A possible role for anti-HLA class I antibodies in thedevelopment of chronic allograft rejection has been suggested(31, 32). These studies showed that binding of anti-HLA classI antibodies stimulated the proliferation of ECs and smoothmuscle cells. Similar studies from our laboratory have alsoshown that binding of anti-HLA antibodies induce prolifera-tion of AECs (33). Although most attention has focused on therole of anti-HLA antibodies in graft rejection, it has becomeclear that antibodies directed against other non-HLA anti-gens may also be of clinical importance. The presence ofnon-HLA antibodies has been shown to be significant inchronic rejection of cardiac, renal, and liver allografts (34–37). However, the nature of non-HLA antibodies developedafter LT has not been determined and their importance inchronic lung allograft rejection has not been defined. Thepresent study was conducted to determine the role of anti-AEC antibodies in BOS pathogenesis, to determine theirmechanism of action, and to characterize their antigenicspecificity.
MATERIALS AND METHODS
Patients. Sixteen LT patients who developed BOS (BOS1) and 11LT patients who did not develop BOS (BOS2) were included in thisstudy. Mean follow-up times for the BOS1 and the BOS2 groupswere 71.3612.2 and 69.5612.0 months after LT, respectively. Meantime for BOS onset was 35.7620.8 months after LT. Sera wereobtained immediately before LT and collected again at 12, 24, 36,and 48 months after LT. Sera from two selected patients (patients 13and 14) with high reactivity to AECs and no reactivity to HLAantigens were also collected at 60 and 72 months after LT. None ofthe patients showed anti-HLA or anti-AEC antibodies before LT. Thecomplement activity in all sera was abrogated by means of incuba-tion at 56°C for 30 min. All sera were kept frozen at 270°C until use.
Patients were included in the BOS2 group if their forced expira-tory volume in 1 second remained at or above 80% of their previouslyestablished baseline value and there was no histological diagnosis ofobliterative bronchiolitis. Patients were included in the BOS1 groupif their forced expiratory volume in 1 second persistently fell below80% of their previously established baseline value or presented apositive histological diagnosis of obliterative bronchiolitis. Otherspecific etiologies for decreased spirometry, such as infection andbronchial anastomotic stricture, were also ruled out before makingthe diagnosis of BOS (38). Two BOS1 patients were diagnosed bymeans of histological observation of obliterative bronchiolitis. Theremaining BOS1 patients were diagnosed by means of physiologicalobservation of decreased spirometry.
Cells. AEC lines were developed from lung biopsies, immortalizedby transfection with the pRSV-Tag plasmid, and cultured in LHC-9medium (39). Primary ECs were obtained from Clonetics (San Diego,CA) and also developed in our laboratory from donor aortas andcultured in M-199 medium. Primary bronchial smooth muscle cells(BSMC) were obtained from Clonetics and cultured in SmGM-2medium (Clonetics) supplemented following the manufacturer’s in-structions. Kidney cell lines (KCL) were developed from the renalcortex of cadaveric kidneys, immortalized by transfection with thepRSV-Tag plasmid, and cultured in RPMI-1640 medium (Sigma, St.Louis, MO) supplemented with 15% fetal bovine serum, 2 mM L-glutamine, 25 mM HEPES buffer, 1 mM sodium pyruvate, 100 U/mlpenicillin, and 0.1 mg/ml streptomycin. Lymphoblastoid cell lines(LCL) were developed by EBV-induced transformation and culturedin RPMI-1640 medium supplemented as described above. A549 (lungcarcinoma), BEP-1 (lung carcinoma), NCI-H69 (lung small cell car-cinoma), MRC-5 (lung fibroblast), HBL-100 (breast carcinoma),
BT-20 (breast carcinoma), and U937 (monocyte) cell lines were ob-tained from the American Type Culture Collection (Manassas, VA)and cultured in RPMI-1640 medium supplemented as describedabove. BEAS-2B (transformed AEC) cell line was obtained from theAmerican Type Culture Collection and cultured in LHC-9 medium(40).
Detection of anti-HLA antibodies. Anti-HLA reactivity was de-tected by means of the PRA-STAT ELISA system (SangStat MedicalCorporation, Menlo Park, CA) that was performed according to themanufacturer’s instructions. Briefly, 100 ml of serum (diluted 1/101)were dispensed into HLA-coated microtiter plates and incubated for120 min at 22°C. After washing (33), horseradish peroxidase-conju-gated goat anti-human IgG was added at appropriate concentrationand the plates were incubated for 60 min at 22°C. The plates werewashed again (33 to remove any unbound conjugate, and 100 ml ofO-phenylenediamine dihydrochloride (0.5 mg/ml) was added. After a15-min incubation, the color development was stopped by the addi-tion of 100 ml HCl (1.0 M), and the optical density was measured at492 nm with a reference wavelength of 630 nm. An assay wasconsidered valid if the positive and negative control values fell withinthe range specified by the manufacturer. A delta value for each testwas determined as the optical density of the well minus the opticaldensity of the corresponding no-antigen well. A test was consideredpositive when the delta value for the well was greater than 0.35 themean positive control value. The pattern of optical density detectedwith an ELISA plate reader was analyzed with the SOFT-STATcomputer program (SangStat Medical Corporation). Patients wereconsidered positive for anti-HLA antibodies if there was positivereactivity against 2% or more of the panel wells with different HLAantigens in any of the sera collected during the follow-up time.
Detection of non-HLA antibodies. The presence of non-HLA anti-bodies directed against the different cell types tested in the presentstudy was determined by means of flow cytometric analysis. Briefly,the cells (53105) were incubated for 30 min at 4°C with AB1 normalhuman serum (NHS) (C-six Diagnostics, Germantown, WI) diluted1:2 in phosphate-buffered saline supplemented with 5% fetal bovineserum, 25 mM HEPES buffer, and 0.02% sodium azide. The cellswere then incubated for 60 min at 25°C with the patient serum (1:2),NHS (negative control) (1:2), or a broadly allo-reactive anti-HLAhuman serum (internal positive control) (1:2). The cells were thenincubated for 30 min at 4°C with fluorescein-conjugated F(ab')2 goatanti-human IgG (1:100) (Jackson ImmunoResearch Laboratories,West Grove, PA). The cells were then fixed in 1% paraformaldehydeand single-color flow cytometric analysis was performed using aFACScan flow cytometer (Becton Dickinson, Mountain View, CA). Atest sample was considered positive if its mean fluorescent channel(MFC) shift was more than 0.5 SD above the NHS (negative control)MFC value.
Anti-HLA antibody absorption. Regardless of their anti-HLA an-tibody status (positive or negative), all samples were processedequally for anti-HLA class I and class II antibody absorption. Toabsorb anti-HLA class I antibodies, the serum was incubated with apooled human platelet concentrate (BCA, West Chester, PA) follow-ing the manufacturer’s instructions. After 60 min at 25°C, the mix-ture was centrifuged and the absorption was repeated twice. Thisprotocol has been shown to reduce the anti-HLA class I reactivity by90.2610.9% in sera from three of the BOS1 patients included in thisstudy (30). To absorb anti-HLA class II antibodies, the serum wasincubated with a pool of five LCLs with different HLA haplotypes(503106 cells/ml of serum). After 60 min. at 25°C, the mixture wascentrifuged and the absorption was repeated twice. After anti-HLAantibody absorption, all sera were tested for the presence of residualanti-HLA antibodies by means of flow cytometric analysis as de-scribed above using a pool of two LCLs with different HLA haplo-types. A test sample was considered positive if its MFC shift wasmore than 0.5 SD above the NHS (negative control) MFC. Afterabsorption of anti-HLA antibodies, all samples were negative for the
JARAMILLO ET AL.April 15, 2001 967

presence of reactive antibodies against the pool of LCLs (data notshown).
Anti-AEC antibody absorption. Ten million AECs were lysed in500 ml of 1% NP-40 diluted in Tris-buffered saline supplementedwith 4 mM EDTA, 0.5 mM zinc chloride, 1 mM sodium orthovana-date, 2 mM PMSF, 5 mM aprotinin, 10 mM leupeptin, and 10 mMpepstatin (stop buffer). After 30 min at 4°C, the samples were cen-trifuged for 10 min. at 14,0003g. To absorb anti-AEC antibodies, 500ml of sera diluted 1:250 in “stop buffer” were mixed with the AEClysate. After 60 min at 4°C, the absorption was repeated twice.
Intracellular Ca11 ([Ca11]i) concentration measurement.Changes in [Ca11]i induced by anti-AEC antibodies binding weremeasured in cultured AECs loaded with the Ca11-specific fluores-cent probe indo 1-AM (Molecular Probes, Eugene, OR). Briefly, AECswere cultured in glass-bottom petri dishes to 80% confluency inLHC-9 medium. Then, the LHC-9 medium was replaced with Ca11
and Mg11-free HBSS supplemented with 2% BSA and 10 mM indo1-AM dissolved in 0.02% DMSO and pluronic acid as a dispersingagent. After 30 min, the AECs were washed and patient serum (1:4)or NHS (negative control) (1:4) were added to the AECs and thefluorescent changes were measured using a laser scanning confocalmicroscope equipped with argon laser and a water immersion flourobjective (403) (RCM 8000; Nikon Inc, Melville, NY). Indo 1-AM is adual emission dye and when excited at 354 nM two wavelengths areemitted at 405 nm corresponding to Ca11-bound indo 1-AM and at480 nm corresponding to free indo 1-AM. Two photo multiplierssimultaneously detecting both emission wavelengths and the analogsignals are then digitized, and displayed in color as high-resolutionspatial concentration images. For calibration, the intensities of testsolutions of Ca11 varying from 0 to 1 mM were determined with 5mM penta-potassium indo-1. The ratiometric calculations were doneusing the method described by Grynkiewicz et al. (40).
Western blotting for phosphotyrosine. AECs were grown to 80%confluency in 6-well plates in LHC-9 medium. Then, the LHC-9medium was replaced with RPMI-1640 medium supplemented with2% BSA, L-glutamine, HEPES buffer, sodium pyruvate, penicillin,and streptomycin (starvation medium). After 24 hr, the AECs wereharvested and 53105 cells were treated with 100 ml of patient serum(1:4) or NHS (negative control) (1:4) in eppendorf tubes at 37°C for 5min. The phosphorylation reactions were stopped by the addition of“stop buffer”. After washing (23) with stop buffer, the AECs werethen lysed in stop buffer containing 1% NP-40. After 30 min at 4°C,the samples were centrifuged for 10 min at 14,0003g. Laemmlisample buffer was then added and the cell lysates were boiled for 5min. The proteins were then separated under reducing conditions(with 1% 2-mercaptoethanol) by means of 4–20% gradient SDS-PAGE. The proteins were transferred to a nitrocellulose membrane,blocked for 60 min in Tris-buffered saline containing 5% BSA and0.5% Tween-20, and immunoblotted for 60 min. with the PY-20anti-phosphotyrosine mouse IgG monoclonal antibody (1:1000)(Transduction Laboratories, Lexington, KY) or the MOPC monoclo-nal antibody (1:1000) (ICN Pharmaceuticals, Costa Mesa, CA) asisotype control. The membranes were then incubated for 60 min withperoxidase-conjugated goat anti-mouse IgG (Organon Teknika,Durham, NC) diluted 1:3000 in Tris-buffered saline containing 5%non-fat dry milk and 0.5% Tween-20. The reaction was then visual-ized by means of the ECL Western blot detection system (AmershamInc, Buckinhamshire, England).
Western blotting for cellular antigens. The cells were lysed in“stop buffer” containing 1% NP-40. After 30 min at 4°C, the celllysates were centrifuged for 10 min. at 14,0003g. Laemmli samplebuffer was then added and the cell lysates were boiled for 5 min. Theproteins were then separated under reducing conditions (with 2-ME)or nonreducing conditions (without 2-ME) by means of 4–20% gra-dient sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Theproteins were transferred to a nitrocellule membrane, blocked for 60min. in Tris-buffered saline containing 5% non-fat dry milk and 0.5%Tween-20, and immunoblotted for 60 min. with patient serum (1:
1500) or NHS (negative control) (1:1500). The membranes were thenincubated for 60 min with peroxidase-conjugated goat anti-humanIgG (1:5000) (Jackson ImmunoResearch Laboratories). The reactionwas then visualized by means of the ECL western blot detectionsystem (Amersham Inc.).
Proliferation assay. AECs (2.53103 cells/well) were incubated for24 hr in flat-bottom 96-well plates in LHC-9 medium. Then, theLHC-9 medium was replaced with “starvation medium” as describedabove. After a starvation period of 24 hr, the AECs were incubatedwith patient serum (1:20) or NHS (negative control) (1:20) for 24, 48,and 72 hr. AECs incubated in LHC-9 medium were used as positivecontrol. [3H]-thymidine (1 mCi/well) was added 24 hr before termi-nation of the cultures and the amount of [3H]-thymidine incorpo-rated into DNA was measured by b-scintillation counting. The re-sults expressed as stimulation indices corresponding to the ratio ofthe mean counts per minute of the test cultures to the mean countsper minute of the cultures in “starvation medium”.
Reverse transcription–polymerase chain reaction. AECs weregrown to 80% confluency in 6-well plates in LHC-9 medium. Then,the LHC-9 medium was replaced with “starvation medium” as de-scribed above. After a starvation period of 24 hr, the AECs wereincubated with patient serum (1:20) or NHS (negative control) (1:20)for 24 hr. The cells were then homogenized with 1 ml TRIzol reagent(Life Technologies, Gaithersburg, MD) and total RNA was isolatedaccording to the manufacturer’s instructions. cDNA was synthesizedby extension of primers using Superscript reverse transcriptase (LifeTechnologies) according to manufacturer’s instructions. PCR wasthen performed using the HotSartTaq master mix (Qiagen, Valencia,CA) in the presence of previously designed and validated oligonucle-otide primers for EGF, HB-EGF, ET-1, bFGF, IGF-1, IGF-2, PDGF,TGF-a, TGF-b, and GM-CSF (10, 16, 41–43). Previously designedand validated oligonucleotide primers for b-actin were used as inter-nal standard control (10). Primers were designed to cross-intron-exon boundaries and were verified to be unique for the genes studied,based on the Genbank database. Primers were tested with high cycleamplification (40 cycles) and all the reactions produced single bandsat the appropriate molecular weight on agarose gel electrophoresis.
Statistical analysis. Statistical analysis of the data was carriedout by means of the Fisher exact test or Student’s paired-sample ttest with the alpha set a priori at P#0.05.
RESULTS
Correlation between development of specific anti-AEC an-tibodies and development of BOS after LT. To determinewhether there was a correlation between the development ofanti-AEC antibodies and the development of BOS after LT,we performed anti-HLA class I and class II antibody absorp-tions as described above. Then, by means of flow cytometricanalysis, we measured the binding of antibodies to AECs(n55, pooled) in serum samples collected serially from all theLT patients included in the study. As shown in Table 1, therewas a slightly significant correlation between the develop-ment of anti-AEC antibodies and the development of BOSafter LT. Five of 16 BOS1 patients (31.2%) and 0 of 11 BOS2patients (0%) developed anti-AEC antibodies after LT
TABLE 1. Correlation between development of anti-AECantibodies and development of BOS after LTa
Anti-AEC1b Anti-AEC2 Total
BOS1 5 11 16BOS2 0 11 11Total 5 22 27
a P50.05; Fisher exact test.b A test sample was considered positive if its MFC shift was more
than 0.5 SD above the NHS (negative control) MFC value.
TRANSPLANTATION968 Vol. 71, No. 7

(P50.05, Fisher exact test). Subsequently, antibody reactiv-ity was tested against ECs (n55, pooled) and peripheralblood monocytes, lymphocytes, or granulocytes (n55, pooled)by flow cytometric analysis as described above. The samesamples did not show any reactivity against any of thesetypes of cells (data not shown). These data indicate the spe-cific nature of the antibody reactivity against AECs and thetotal absence of residual anti-HLA antibodies in the samples.
As shown in Table 2, there was no correlation between thedevelopment of cytomegalovirus infection and the develop-ment of anti-AEC antibodies after LT. Two of 5 anti-AEC1,BOS1 patients (40.0%) and 6 of 11 anti-AEC2, BOS1 pa-tients (54.5%) developed at least one active cytomegalovirusinfection after LT (P.0.99, Fisher exact test). There was nocorrelation between the development of acute rejection epi-sodes and the development of anti-AEC antibodies after LT.Five of five anti-AEC1, BOS1 patients (100%) and 10 of 11anti-AEC2, BOS1 patients (90.9%) developed at least oneacute rejection episode after LT (P.0.99, Fisher exact test).There was no correlation between the development of cyto-megalovirus infection and the development of BOS after LT.Eight of 16 BOS1 patients (50.0%) and 5 of 11 BOS2 pa-tients (45.4%) developed at least one active cytomegalovirusinfection after LT (P.0.99, Fisher exact test). There was nocorrelation between the development of acute rejection epi-sodes and the development of BOS after LT. Fifteen of 16BOS1 patients (93.7%) and 10 of 11 BOS2 patients (90.9%)developed at least one acute rejection episode after LT(P.0.99, Fisher exact test). There was a significant correla-tion between the development of anti-HLA antibodies and
the development of BOS after LT. Ten of 16 BOS1 patientsdeveloped anti-HLA antibodies (62.5%) while 0 of 11 BOS2patients (0%) developed anti-HLA antibodies (P50.001,Fisher exact test). There was no correlation between thedevelopment of anti-HLA antibodies and the development ofanti-AEC antibodies after LT. One of 5 anti-AEC1 patients(20.0%) also developed anti-HLA antibodies and 9 of 11 anti-AEC2 patients (81.8%) also developed anti-HLA antibodies(P50.35, Fisher exact test). Therefore, most of the BOS1patients show an exclusive development of either anti-HLAantibodies or anti-AEC antibodies.
Sequential measurements of the anti-AEC antibodies ob-served in two sera with the highest anti-AEC reactivityshowed different kinetics of antibody development. As shownin Figure 1A, patient 13 developed anti-AEC antibodies after36 months after LT. The highest titer of anti-AEC antibodieswas observed at 48 months after LT, time of BOS onset inthis patient. Subsequently, the anti-AEC reactivity in thispatient was reduced significantly at 72 months after LT. Incontrast, anti-AEC antibodies were observed as early as 12months after LT in patient 14 who developed BOS 32 monthsafter LT (Fig. 1B). In this patient, the level of anti-AECreactivity remained stable during the length of the study.Therefore, serum samples from this patient were used forfurther biochemical and functional analysis in our study.
Differential AEC recognition by anti-AEC antibodies devel-oped after LT. To further characterize the antibody reactiv-ity developed in patients 13 and 14, the binding of anti-AECantibodies was assessed against a panel of AEC lines. Theanti-AEC reactivity developed by patient 13 was limited to 6
TABLE 2. Clinical and laboratory evaluation of LT patients
Patient BOS Diagnosis Anti-HLA Anti-AEC CMV infection Acute rejection
1 1 Ha 1 2 2 12 1 H 1 2 2 13 1 Pb 1 2 1 14 1 P 1 2 1 25 1 P 1 2 2 16 1 P 1 2 1 17 1 P 1 2 1 18 1 P 1 2 2 19 1 P 1 2 1 1
10 1 P 1 1 2 111 1 P 2 1 2 112 1 P 2 1 2 113 1 P 2 1 1 114 1 P 2 1 1 115 1 P 2 2 2 116 1 P 2 2 1 117 2 NAc 2 2 2 118 2 NA 2 2 1 119 2 NA 2 2 2 120 2 NA 2 2 2 121 2 NA 2 2 2 122 2 NA 2 2 1 123 2 NA 2 2 1 124 2 NA 2 2 1 125 2 NA 2 2 2 126 2 NA 2 2 2 227 2 NA 2 2 1 1
a Histological observation of obliterative bronchiolitis.b Physiological observation of decreased spirometry.c NA, Not applicable.
JARAMILLO ET AL.April 15, 2001 969

of 9 AEC lines (66.6%) (Fig. 2A) and the anti-AEC reactivitydeveloped by patient 14 was limited to 6 of 10 AEC lines(60%) (Fig. 2B). Interestingly, both serum samples recog-nized the same AEC lines in the panel tested. These datasuggested that both sera are recognizing the same allele of apolymorphic antigenic system. To rule out the possibility thatthe anti-AEC reactivity observed in these serum samples wasnot against a surface antigen up-regulated by the pRSV-Tag-induced transformation, we tested the reactivity of serumfrom patient 14 against a panel of 8 primary AEC lines, 6 ofwhich were also pRSV-Tag-transformed and used in thescreening experiments showed in Figures 2A and B. As ob-served in Figure 2C, a similar reactivity pattern was ob-served with the primary AECs. The anti-AEC reactivity de-veloped by patient 14 after LT was limited to five of eightprimary AEC lines (62.5%) as was the case with the pRSV-Tag-transformed AEC lines. Only one of the primary AEClines showed a significant up-regulation of surface antigenic-ity recognized by serum 14 after transformation with thepRSV-Tag plasmid (KIK-511 AEC line) (Figs. 2B and C).
Lack of recognition of cell lines from different tissue originby anti-AEC antibodies developed after LT. To determinewhether the reactivity developed in patient 14 was specificfor AECs, we tested the reactivity of this serum sampleagainst several cell lines from different tissues, i.e., theMRC-5 lung fibroblast cell line, the HBL-100 and BT-20breast carcinoma cell lines, the U937 monocytic cell line,KCLs, LCLs, BSMCs, and ECs. As shown in Figure 3, onlythe BT-20 breast cancer cell line was recognized by serum
from patient 14. Overall, these data indicate that the surfaceantigen recognized by patient 14 is expressed by the majorityof AECs and a limited number of epithelial cells from othertissues.
Recognition in whole cell extracts of a 60-kDa antigen byanti-AEC antibodies developed after LT. To characterize theantigen(s) recognized by sera from patients 13 and 14, thereactivity of these samples was determined by western blot-ting with whole cell extracts from the KCC-266 AEC line,which was recognized by the anti-AEC antibodies by flowcytometry (surface antigen expression-positive, Fig. 2), andthe KJE-206 AEC line, which was not recognized by theanti-AEC antibodies by flow cytometry (surface antigen ex-pression-negative Fig. 2). As shown in Figure 4, an antigen ofapproximately 60 kDa was detected in both AEC lines byserum from patient 14. This result indicates a differentialsurface expression of this antigen in AECs rather than rec-ognition of a polymorphic antigenic system. The same 60-kDaantigen was detected by serum from patient 14 both underreducing and nonreducing conditions indicating the mono-meric nature of the antigen (data not shown). Similar reac-tivity was observed in serum samples from patient 14 col-
FIGURE 2. Differential AEC recognition by the anti-AEC anti-bodies developed after LT. Anti-AEC reactivity was assessedin patients 13 (A) and 14 (B) by means of FACS analysis usingSV-40-transformed AEC lines. Anti-AEC reactivity was alsoassessed in patient No. 14 (C) by means of FACS analysisusing primary AEC lines. A broadly reactive (100%) anti-HLAalloserum was used as internal positive control. Data arepresented as MFC shift over the NHS (negative control) MFCvalue. Number of cells analyzed: 33103.
FIGURE 1. Sequential measurements of the anti-AEC antibod-ies developed after LT. Serum samples were serially collectedevery 12 months until 72 months after LT. Anti-AEC antibod-ies were detected in patient 13 (A) and 14 (B) by means ofFACS analysis using a pool of five AEC lines. Data are pre-sented as MFC shift over the NHS (negative control) MFCvalue. Number of cells analyzed: 33103.
TRANSPLANTATION970 Vol. 71, No. 7

lected at 36, 48, and 60 months after LT (data not shown). Todemonstrate that serum from other patients who tested pos-itive by FACS analysis also recognized this 60-kDa antigen,we tested serum from patient 13 collected at 48 months afterLT. As shown in Figure 4, serum from patient 13 also recog-nized this 60-kDa antigen as well as several other antigens inboth the KCC-266 and the KJE-206 AEC lines. To demon-strate that the reactivity against this 60-kDa antigen wasdeveloped after LT, we tested serum samples from patients13 and 14 collected before LT (0 months, Fig. 1A and B,respectively). As shown in Figure 4, no antibody reactivitywas observed against both AEC lines in both serum samplescollected before LT. In addition, no reactivity was observed inNHS (used as negative control in all experiments).
To demonstrate that the anti-AEC antibodies developedafter LT recognize the same 60-kDa antigen in both theKCC-266 (surface antigen-positive) and the KJE-206 (surface
antigen-negative) AEC lines, we absorbed the serum frompatient 14 with whole cell extract from the KCC-266 or theKJE-206 AEC lines. Then, we determine by Western blottingwhether the anti-AEC reactivity against whole cell extractsfrom both cell lines could be absorbed by this procedure. Asshown in Figure 5, incubation of this serum sample with theKCC-266 AEC line completely absorbed the reactivityagainst a 60-kDa antigen present in the KCC-266 AEC ly-sates as well as in the KJE-206 AEC lysates. Similar resultswere obtained when the same serum sample was absorbedwith a whole cell extract from the KJE-206 AEC line (Fig. 5).These results further indicate the specific recognition in bothsurface antigen-positive and surface antigen-negative AEClines of the same 60-kDa antigen by the anti-AEC antibodiesdeveloped after LT. The fact that high levels of anti-AECreactivity was detected against 60% of primary AEC lineswith the serum sample from patient 14 (Fig. 2C) suggest thatthis 60-kDa antigen is expressed on the surface of AECs in asignificant proportion of lung allografts.
Further analysis by western blotting with whole cell ex-tracts from all the AEC lines tested showed that comparablelevels of the 60-kDa antigen are detected in surface antigen-positive AEC lines (BEP-1, KBU-866, KHE-472, KIK-511,KDW-568, and KCC-266) (Fig. 6A) as well as surface antigen-negative AEC lines (A-549, BEAS-2B, NCI-H69, and KJE-206) (Fig. 6B). All the AEC lines tested expressed similarintracellular levels of the antigen. Also, all the positive AEClines demonstrated significant expression of this antigencomparable to the HLA expression, which was used as posi-tive control in the FACS analysis. Figure 6B also demon-strates that similar levels of the 60-kDa antigen are detectedin epithelial cell lines from different tissues (BT-20, HBL-100, KCLs), lung fibroblasts (MRC-5), BSMCs, and ECs.Very low intracellular levels of this antigen were detected inLCLs suggesting a differential expression of this protein indifferent tissues at the intracellular level.
FIGURE 3. Lack of recognition of cell lines from different tis-sue origin by the anti-AEC antibodies developed after LT.Antibody reactivity was assessed in patient 14 by means ofFACS analysis using cell lines from different origin. Abroadly reactive (100%) anti-HLA alloserum was used as in-ternal positive control. Data are presented as MFC shift overthe NHS (negative control) MFC value. Number of cells ana-lyzed: 33103.
FIGURE 4. Recognition of a 60-kDa antigen in both surfaceantigen-positive and antigen-negative AEC lines by the anti-AEC antibodies developed after LT. Proteins from whole cellextracts from the KCC-266 (surface antigen-positive) and theKJE-206 (surface antigen-negative) AEC lines were separatedunder reducing (with 2-ME) by SDS-PAGE. The AEC sampleswere then analyzed by western blotting using sera from pa-tients 13 and No. 14 collected before and after LT as well asNHS as negative control. Results shown are representative oftwo different experiments.
FIGURE 5. The same 60-kDa antigen is recognized in bothsurface antigen-positive and antigen-negative AEC lines bythe anti-AEC antibodies developed after LT. Proteins fromwhole cell extracts from the KCC-266 (surface antigen-posi-tive) and the KJE-206 (surface antigen-negative) AEC lineswere separated under reducing conditions (with 2-ME) bySDS-PAGE. The AEC samples were then analyzed by westernblotting using serum from patient 14 preabsorbed with theKCC-266 or the KJE-206 AEC lines. Nonabsorbed serum frompatient 14 and NHS were used as positive and negative con-trols, respectively. Results shown are representative of twodifferent experiments.
JARAMILLO ET AL.April 15, 2001 971
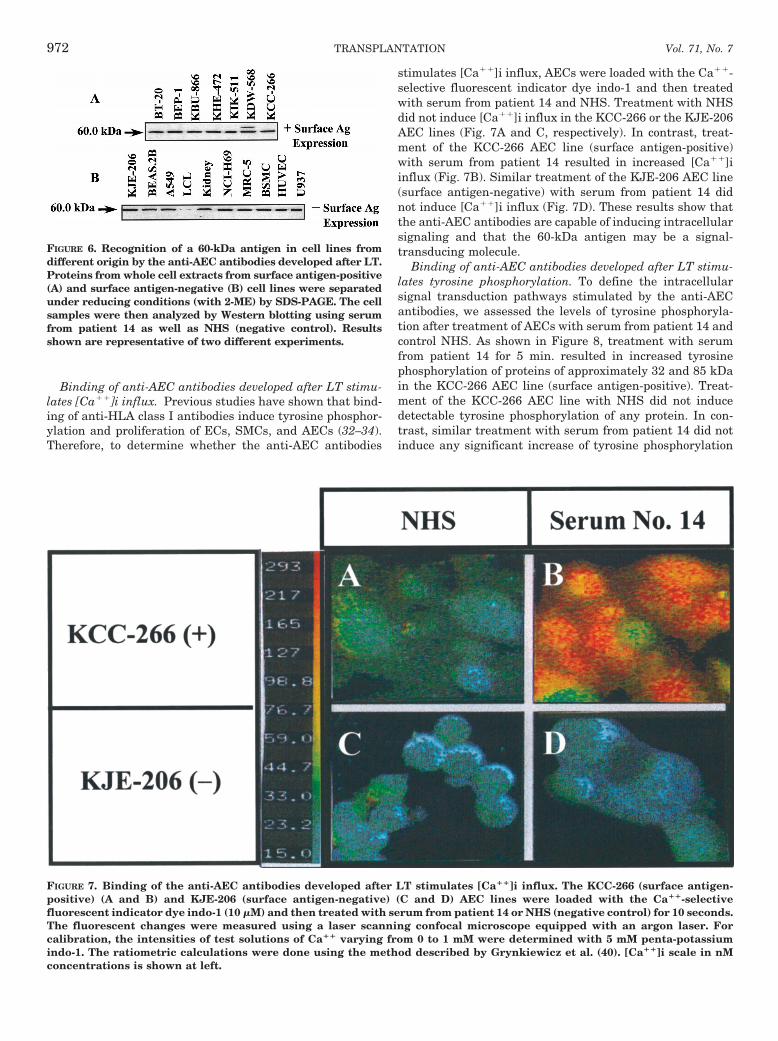
Binding of anti-AEC antibodies developed after LT stimu-lates [Ca11]i influx. Previous studies have shown that bind-ing of anti-HLA class I antibodies induce tyrosine phosphor-ylation and proliferation of ECs, SMCs, and AECs (32–34).Therefore, to determine whether the anti-AEC antibodies
stimulates [Ca11]i influx, AECs were loaded with the Ca11-selective fluorescent indicator dye indo-1 and then treatedwith serum from patient 14 and NHS. Treatment with NHSdid not induce [Ca11]i influx in the KCC-266 or the KJE-206AEC lines (Fig. 7A and C, respectively). In contrast, treat-ment of the KCC-266 AEC line (surface antigen-positive)with serum from patient 14 resulted in increased [Ca11]iinflux (Fig. 7B). Similar treatment of the KJE-206 AEC line(surface antigen-negative) with serum from patient 14 didnot induce [Ca11]i influx (Fig. 7D). These results show thatthe anti-AEC antibodies are capable of inducing intracellularsignaling and that the 60-kDa antigen may be a signal-transducing molecule.
Binding of anti-AEC antibodies developed after LT stimu-lates tyrosine phosphorylation. To define the intracellularsignal transduction pathways stimulated by the anti-AECantibodies, we assessed the levels of tyrosine phosphoryla-tion after treatment of AECs with serum from patient 14 andcontrol NHS. As shown in Figure 8, treatment with serumfrom patient 14 for 5 min. resulted in increased tyrosinephosphorylation of proteins of approximately 32 and 85 kDain the KCC-266 AEC line (surface antigen-positive). Treat-ment of the KCC-266 AEC line with NHS did not inducedetectable tyrosine phosphorylation of any protein. In con-trast, similar treatment with serum from patient 14 did notinduce any significant increase of tyrosine phosphorylation
FIGURE 6. Recognition of a 60-kDa antigen in cell lines fromdifferent origin by the anti-AEC antibodies developed after LT.Proteins from whole cell extracts from surface antigen-positive(A) and surface antigen-negative (B) cell lines were separatedunder reducing conditions (with 2-ME) by SDS-PAGE. The cellsamples were then analyzed by Western blotting using serumfrom patient 14 as well as NHS (negative control). Resultsshown are representative of two different experiments.
FIGURE 7. Binding of the anti-AEC antibodies developed after LT stimulates [Ca11]i influx. The KCC-266 (surface antigen-positive) (A and B) and KJE-206 (surface antigen-negative) (C and D) AEC lines were loaded with the Ca11-selectivefluorescent indicator dye indo-1 (10 mM) and then treated with serum from patient 14 or NHS (negative control) for 10 seconds.The fluorescent changes were measured using a laser scanning confocal microscope equipped with an argon laser. Forcalibration, the intensities of test solutions of Ca11 varying from 0 to 1 mM were determined with 5 mM penta-potassiumindo-1. The ratiometric calculations were done using the method described by Grynkiewicz et al. (40). [Ca11]i scale in nMconcentrations is shown at left.
TRANSPLANTATION972 Vol. 71, No. 7
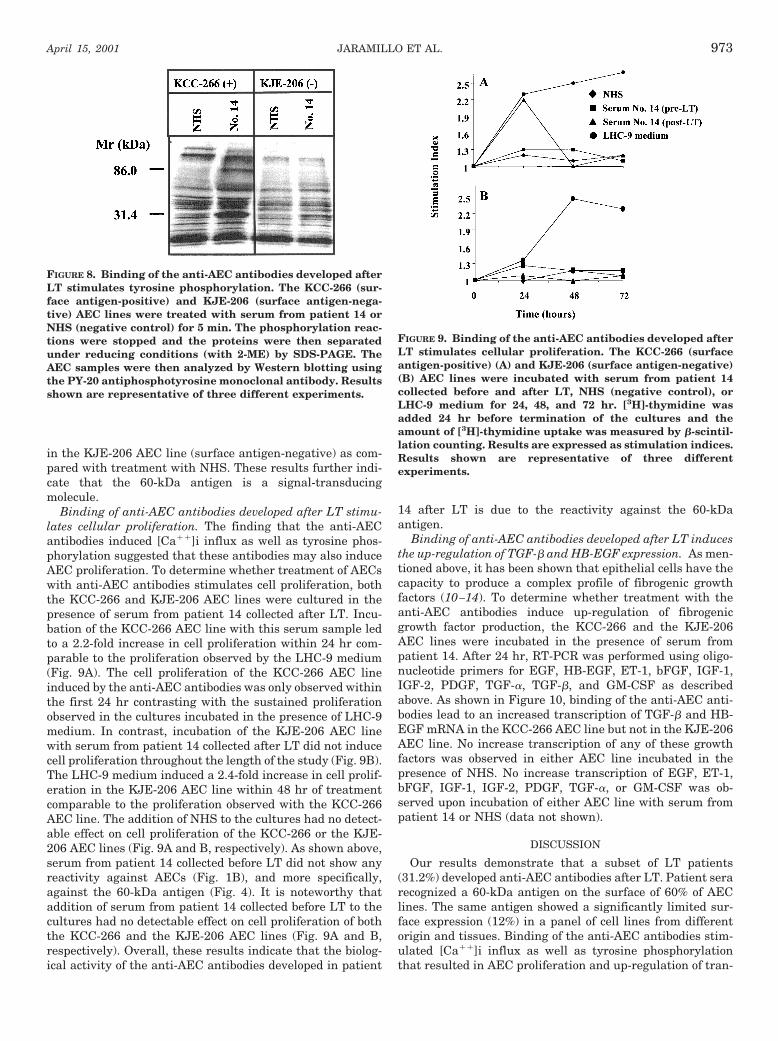
in the KJE-206 AEC line (surface antigen-negative) as com-pared with treatment with NHS. These results further indi-cate that the 60-kDa antigen is a signal-transducingmolecule.
Binding of anti-AEC antibodies developed after LT stimu-lates cellular proliferation. The finding that the anti-AECantibodies induced [Ca11]i influx as well as tyrosine phos-phorylation suggested that these antibodies may also induceAEC proliferation. To determine whether treatment of AECswith anti-AEC antibodies stimulates cell proliferation, boththe KCC-266 and KJE-206 AEC lines were cultured in thepresence of serum from patient 14 collected after LT. Incu-bation of the KCC-266 AEC line with this serum sample ledto a 2.2-fold increase in cell proliferation within 24 hr com-parable to the proliferation observed by the LHC-9 medium(Fig. 9A). The cell proliferation of the KCC-266 AEC lineinduced by the anti-AEC antibodies was only observed withinthe first 24 hr contrasting with the sustained proliferationobserved in the cultures incubated in the presence of LHC-9medium. In contrast, incubation of the KJE-206 AEC linewith serum from patient 14 collected after LT did not inducecell proliferation throughout the length of the study (Fig. 9B).The LHC-9 medium induced a 2.4-fold increase in cell prolif-eration in the KJE-206 AEC line within 48 hr of treatmentcomparable to the proliferation observed with the KCC-266AEC line. The addition of NHS to the cultures had no detect-able effect on cell proliferation of the KCC-266 or the KJE-206 AEC lines (Fig. 9A and B, respectively). As shown above,serum from patient 14 collected before LT did not show anyreactivity against AECs (Fig. 1B), and more specifically,against the 60-kDa antigen (Fig. 4). It is noteworthy thataddition of serum from patient 14 collected before LT to thecultures had no detectable effect on cell proliferation of boththe KCC-266 and the KJE-206 AEC lines (Fig. 9A and B,respectively). Overall, these results indicate that the biolog-ical activity of the anti-AEC antibodies developed in patient
14 after LT is due to the reactivity against the 60-kDaantigen.
Binding of anti-AEC antibodies developed after LT inducesthe up-regulation of TGF-b and HB-EGF expression. As men-tioned above, it has been shown that epithelial cells have thecapacity to produce a complex profile of fibrogenic growthfactors (10–14). To determine whether treatment with theanti-AEC antibodies induce up-regulation of fibrogenicgrowth factor production, the KCC-266 and the KJE-206AEC lines were incubated in the presence of serum frompatient 14. After 24 hr, RT-PCR was performed using oligo-nucleotide primers for EGF, HB-EGF, ET-1, bFGF, IGF-1,IGF-2, PDGF, TGF-a, TGF-b, and GM-CSF as describedabove. As shown in Figure 10, binding of the anti-AEC anti-bodies lead to an increased transcription of TGF-b and HB-EGF mRNA in the KCC-266 AEC line but not in the KJE-206AEC line. No increase transcription of any of these growthfactors was observed in either AEC line incubated in thepresence of NHS. No increase transcription of EGF, ET-1,bFGF, IGF-1, IGF-2, PDGF, TGF-a, or GM-CSF was ob-served upon incubation of either AEC line with serum frompatient 14 or NHS (data not shown).
DISCUSSION
Our results demonstrate that a subset of LT patients(31.2%) developed anti-AEC antibodies after LT. Patient serarecognized a 60-kDa antigen on the surface of 60% of AEClines. The same antigen showed a significantly limited sur-face expression (12%) in a panel of cell lines from differentorigin and tissues. Binding of the anti-AEC antibodies stim-ulated [Ca11]i influx as well as tyrosine phosphorylationthat resulted in AEC proliferation and up-regulation of tran-
FIGURE 8. Binding of the anti-AEC antibodies developed afterLT stimulates tyrosine phosphorylation. The KCC-266 (sur-face antigen-positive) and KJE-206 (surface antigen-nega-tive) AEC lines were treated with serum from patient 14 orNHS (negative control) for 5 min. The phosphorylation reac-tions were stopped and the proteins were then separatedunder reducing conditions (with 2-ME) by SDS-PAGE. TheAEC samples were then analyzed by Western blotting usingthe PY-20 antiphosphotyrosine monoclonal antibody. Resultsshown are representative of three different experiments.
FIGURE 9. Binding of the anti-AEC antibodies developed afterLT stimulates cellular proliferation. The KCC-266 (surfaceantigen-positive) (A) and KJE-206 (surface antigen-negative)(B) AEC lines were incubated with serum from patient 14collected before and after LT, NHS (negative control), orLHC-9 medium for 24, 48, and 72 hr. [3H]-thymidine wasadded 24 hr before termination of the cultures and theamount of [3H]-thymidine uptake was measured by b-scintil-lation counting. Results are expressed as stimulation indices.Results shown are representative of three differentexperiments.
JARAMILLO ET AL.April 15, 2001 973

scription of two fibrogenic growth factors: TGF-b andHB-EGF.
Previous studies have shown that the development of anti-HLA antibodies after transplantation is associated withchronic rejection in heart, kidney, and lung allografts (29, 30,44–47). Although most attention has focused on the role ofanti-HLA antibodies in allograft rejection, it is becomingincreasingly clear that antibodies against non-HLA antigensexpressed by different cell types in the allograft may be alsoof clinical significance. It has been shown that the develop-ment of anti-EC and anti-kidney epithelial cell antibodies isa significant risk factor for acute and chronic allograft rejec-tion as well as early graft loss (34, 35). In addition, previousstudies from our laboratory have shown that non-HLA anti-bodies directed against a 90– to 100–kDa epithelial cellantigen could be eluted from rejected kidneys (48). Moreover,Fredrich et al. (37) have recently found a significant correla-tion between the development of anti-EC antibodies duringcardiac allograft rejection and a lower 2-year allograftsurvival.
The nature of non-HLA antibodies developed after LT hasnot been determined and their role in chronic lung allograftrejection has not been defined. In our study we observed aslightly significant correlation between the development ofanti-AEC antibodies and the development of BOS after LT(P50.05, Fisher exact test). The time of BOS onset wascomparable between the anti-HLA1 patients (34621 monthsafter LT) and the anti-AEC1 patients (3769 months afterLT). Therefore, the detection of the anti-AEC antibodies pre-ceded the onset of BOS with a significant mean difference of22.5 months (P50.01, Student’s paired-sample t test). Inter-estingly, we have previously observed that anti-HLA anti-bodies developed after LT preceded the onset of BOS by 20.1
months (30). These results suggest that the anti-AEC anti-bodies may play a role in the pathogenesis of BOS in theabsence of anti-HLA antibodies.
BOS is characterized histologically by small airway inflam-mation and dense fibrosis of the lamina propria and lumenwith some degree of interstitial fibrosis (2). Up to this date,only two fibrogenic growth factors, PDGF and TGF-b, havebeen shown to be increased during the process of BOS devel-opment after LT in humans (25, 26). Increased concentra-tions of PDGF from mononuclear cells were observed in bron-choalveolar lavage samples from BOS1 patients before theonset of BOS (25). It is interesting that immunohistochemicalstaining demonstrated the presence of PDGF at the apicalsurface of AECs in some regions of the lesion. Another studydemonstrated by immunohistochemistry, an increase concen-tration of infiltrating macrophages with a concomitant up-regulation of PDGF and TGF-b in chronically rejected lungallografts (26). Previous studies have shown that anti-HLAclass I antibodies induced activation and proliferation of ECs,SMCs, and AECs (31–33). Our study shows that binding ofanti-AEC antibodies directed against a 60-kDa surface anti-gen induces AEC activation and up-regulation of TGF-b andHB-EGF production by these cells. These data suggest thatanti-AEC antibodies directed against non-HLA antigens mayalso play a role in the pathogenesis of BOS by up-regulatingthe production of fibrogenic growth factors by AECs in situ.
In our study, the antibodies developed after LT did notshow any reactivity to ECs, lymphocytes, monocytes, or gran-ulocytes tested by means of flow cytometric analysis (data notshown). These results further indicate the total absence ofresidual anti-HLA antibodies in these samples and the AEC-specificity of these antibodies. These results are also inagreement with previous studies that suggest the importanceof AECs as the main target for allogeneic immune responseduring BOS development after LT (6–8). Romaniuk et al. (6)showed that MHC class II antigens were expressed by AECsin rejecting rat pulmonary allografts but not in normal lungepithelium. Kubota et al. (7) also demonstrated the up-regu-lation of expression of MHC class I and class II antigens byAECs in rejecting rat pulmonary allografts. Despite theirability to express both MHC class I and class II antigens,very little data are available on the antigen-presenting abil-ity of AECs to CD41 T cells. In a related study, Mauck andHosenpud (10) demonstrated the ability of a human AEC lineto up-regulate intercellular ICAM-1 and both MHC class Iand class II antigens after treatment with IFN-g. This treat-ment with IFN-g rendered this AEC line capable of inducingstrong T cell proliferative response in vitro not observed withuntreated cells. Overall these data indicate that during thedevelopment of BOS, AECs are capable of antigen presenta-tion and of inducing activation of CD41 T cells.
It is noteworthy that a significant proportion of CD41 Th2cells has been shown in the mononuclear cell infiltrate oflung allografts during BOS development (49, 50). In addition,B cells have been shown to accumulate in intraluminal fi-brous tissue and to form follicles in perivascular and peri-bronchiolar spaces during the BOS development (50). Over-all, these data indicate that infiltrating CD41 Th2 cellsactivated by AECs, infiltrating dendritic cells (51), or both,may be able to induce a humoral immune response againstthe lung allograft epithelium during the course of BOSdevelopment.
FIGURE 10. Binding of the anti-AEC antibodies developed af-ter LT induces the up-regulation of TGF-b and HB-EGF ex-pression. The KCC-266 (surface antigen-positive) and KJE-206 AEC lines (surface antigen-negative) were incubatedwith serum from patient 14 or NHS (negative control) for 24hr. Total RNA was isolated, reversed transcribed into cDNA,PCR amplified, and electrophoresed in 2% agarose gel. Pre-viously designed and validated oligonucleotide primers forseveral fibrogenic growth factors and b-actin (internal stan-dard control) were used. The amplified products of b-actin,TGF-b, and HB-EGF are shown. Results shown are represen-tative of two different experiments.
TRANSPLANTATION974 Vol. 71, No. 7

Our results show that the 60-kDa antigen recognized bynon-HLA antibodies developed after LT is preferentially ex-pressed on the surface of AECs although it remains in asubcellular compartment in all other cell types. Also, none ofthe positive cell lines expressed intermediate surface expres-sion of this antigen as compared to the surface expression ofHLA Ags. These data suggest a tissue-specific regulation ofexpression of this non-polymorphic 60-kDa antigen. Furthercharacterization of the 60-kDa antigen expressed on AECsmay provide important information about the regulation ofexpression of this protein.
Our results indicate that specific anti-AEC antibodies maybe contributing to the development of BOS by binding tonon-HLA surface antigens and directly inducing the activa-tion of AECs that results in proliferation as well as increasedgrowth factor production by these cells. These may lead tothe activation of lung fibroblasts and BSMCs resulting in thetissue remodeling and proliferation of fibrous tissue observedduring the development of BOS. Our findings provide a basisfor the involvement of humoral immune responses againstspecific AEC antigens as one of the mechanisms for BOSpathogenesis after LT.
Acknowledgments. The authors thank Ms. Billie Glasscock forsecretarial assistance and Mr. Brian Duffy for flow cytometricanalysis.
REFERENCES
1. Reinsmoen NL, Bolman RM, Savik K, Butters K, Hertz MI. Aremultiple immunopathogenic events occurring during the de-velopment of obliterative bronchiolitis and acute rejection?Transplantation 1993; 55: 1040.
2. Kelly K, Hertz MI. Obliterative bronchiolitis. Clin Chest Med1997; 18: 319.
3. Tullius SG, Tilney NL. Both alloantigen-dependent and indepen-dent factors influence chronic allograft rejection. Transplanta-tion 1995; 59: 313.
4. Allen MD, Burke CM, McGregor CGA, Baldwin JC, JamiesonSW, Theodore J. Steroid-responsive bronchiolitis after humanheart-lung transplantation. J Thorac Cardiovasc Surg 1986;92: 449.
5. Glanville AR, Baldwin JC, Burke CM, Robin ED. Obliterativebronchiolitis after heart-lung transplantation: Apparent arrestby augmented immunosuppression. Ann Intern Med 1987;107: 300.
6. Romaniuk A, Prop J, Petersen AH, Wildevuur CRH, Nieuwen-huis P. Expression of class II major histocompatibility complexantigens by bronchial epithelium in rat lung allografts. Trans-plantation 1987; 44: 209
7. Kubota H, Yagyu K, Takeshita OM, Furuse A. Importance ofbronchus-associated lymphoid tissue and major histocompati-bility complex class I and II antigen expression on bronchialepithelium in acute lung allograft rejection and lung infectionin rats. Transplant Proc 1994; 26: 1856.
8. Spurzem JR, Sacco O, Rossi GA, Beckmann JD, Rennard SI.Regulation of major histocompatibility complex class II geneexpression on bovine bronchial epithelial cells. J Lab Clin Med1992; 120: 94.
9. Zhao J, Sime PJ, Bringas P, Gauldie J, Warburton D. Epitheli-um-specific adenoviral transfer of a dominant-negative mutantTGF-b type II receptor stimulates embryonic lung branchingmorphogenesis in culture and potentiates EGF and PDGF-AA.Mech Dev 1992; 72: 89.
10. Mauck KA, Hosenpud JD. The bronchial epithelium: A potentialallogenic target for chronic rejection after lung transplanta-
tion. J Heart Lung Transplant 1996; 15: 709.11. Markewitz BA, Kohan DE, Michael JR. Endothelin-1 synthesis,
receptors, and signal transduction in alveolar epithelium: ev-idence for an autocrine role. Am J Physiol 1995; 268: L192.
12. Wagner CR, Morris TE, Shipley GD, Hosenpud JD. Regulation ofhuman aortic endothelial cell-derived mesenchymal growthfactors by allogeneic lymphocytes in vitro. A potential mecha-nism for cardiac allograft vasculopathy. J Clin Invest 1993; 92:1269.
13. O’Brien AD, Standiford TJ, Christensen PJ, Wilcoxen SE, PaineR. Chemotaxis of alveolar macrophages in response to signalsderived from alveolar epithelial cells. J Lab Clin Med 1998;131: 417.
14. Cambrey AD, Kwon OJ, Gray AJ, et al. Insulin-like growthfactor I is a major fibroblast mitogen produced by primarycultures of human airway epithelial cells. Clin Sci 1995; 89:611.
15. Luo D, Lin Y, Liu X, et al. Effect of prostatic growth factor, basicfibroblast growth factor, epidermal growth factor, and steroidson the proliferation of human fetal prostatic fibroblasts. Pros-tate 1996; 28: 352.
16. Michael JR, Markewitz BA. Endothelins and the lung. Am JRespir Crit Care Med 1996; 154: 555.
17. Ohgi S, Johnson PW. Glucose modulates growth of gingivalfibroblasts and periodontal ligament cells: correlation withexpression of basic fibroblast growth factor. J Periodont Res1996; 31: 579.
18. Cerutis DR, Nogami M, Anderson JL, et al. Lysophosphatidicacid and EGF stimulate mitogenesis in human airway smoothmuscle cells. Am J Physiol 1997; 273: L10.
19. Clark RA, McCoy GA, Folkvord JM, McPherson JM. TGF-b1stimulates cultured human fibroblasts to proliferate and pro-duce tissue-like fibroplasia: a fibronectin matrix-dependentevent. J Cell Physiol 1997; 170: 69.
20. Postiglione L, Montagnani LS, Riccio A, Ladogana P, Salzano S,Callefuoco LRG. Expression of GM-CSF receptor and “in vitro”effects of GM-CSF on human fibroblasts. Life Sci 1998; 63: 327.
21. Pukac L, Huangpu J, Karnovsky MJ. Platelet-derived growthfactor-BB, insulin-like growth factor-I, and phorbol ester acti-vate different signaling pathways for stimulation of vascularsmooth muscle cell migration. Exp Cell Res 1998; 242: 548.
22. Stepien O, Gogusev J, Zhu DL, et al. Amlodipine inhibition ofserum-, thrombin-, or fibroblast growth factor-induced vascu-lar smooth-muscle cell proliferation. J Cardiovasc Pharmacol1998; 31: 786.
23. Hertz MI, Jessurun J, King MB, Murray JJ. Reproduction of theobliterative bronchiolitis lesion after heterotopic transplanta-tion of mouse airways. Am J Pathol 1993 142; 1945.
24. Al-Dossari GA, Jessurun J, Bolman RM, et al. Pathogenesis ofobliterative bronchiolitis. Possible roles of platelet-derivedgrowth factor and basic fibroblast growth factor. Transplanta-tion 1995; 59: 143.
25. Hertz MI, Henke CA, Nakhleh RE, et al. Obliterative bronchi-olitis after lung transplantation: A fibroproliferative disorderassociated with platelet-derived growth factor. Proc Natl AcadSci USA 1992; 89: 10385.
26. Hirabayashi T, Demertzis S, Schafers J, Hoshino K, Nashan B.Chronic rejection in lung allografts: Immunohistological anal-ysis of fibrogenesis. Transplant Int 1996; 9: S293.
27. El-Gamel A, Awad M, Sim E, et al. Transforming growth fac-tor-b1 and lung allograft fibrosis. Eur J Cardiothorac Surgery1998; 13: 424.
28. Schersten H, Hedner T, McGregor CGA, et al. Increased levels ofendothelin-1 in bronchoalveolar lavage fluid of patients withlung allografts. J Thorac Cardiovasc Surg 1996; 111: 253.
29. Sundaresan S, Mohanakumar T, Smith MA, et al. HLA-A locusmismatches and development of antibodies to HLA after lungtransplantation correlate with the development of bronchioli-
JARAMILLO ET AL.April 15, 2001 975

tis obliterans syndromw. Transplantation 1998; 65: 648.30. Jaramillo A, Smith MA, Phelan D, et al. Development of ELISA-
detected anti-HLA antibodies precedes the development ofbronchiolitis obliterans syndrome and correlates with progres-sive decline in pulmonary function after lung transplantation.Transplantation 1999; 67: 1155.
31. Bian H, Harris PE, Mulder A, Reed EF. Anti-HLA antibodyligation to HLA class I molecules expressed by endothelial cellsstimulates tyrosine phosphorylation, inositol phosphate gener-ation, and proliferation. Hum Immunol 1997; 53: 90.
32. Harris PE, Bian H, Reed EF. Induction of high affinity fibroblastgrowth factor receptor expression and proliferation in humanendothelial cells by anti-HLA antibodies: a possible mecha-nism for transplant atherosclerosis. J Immunol 1997; 159:5697.
33. Reznik SI, Jaramillo A, Zhang L, Patterson GA, Cooper JD,Mohanakumar T. Anti-HLA antibody binding to HLA class Imolecules induces proliferation of airway epithelial cells: Apotential mechanism for bronchiolitis obliterans syndrome.J Thorac Cardiovasc Surg 2000; 119: 39
34. Harmer AW, Haskard D, Koffman CG, Welsh KI. Novel antibod-ies associated with unexplained loss of renal allografts. Trans-pland Int 1990; 3: 66.
35. Al-Hussein KA, Talbot D, Proud G, Taylor RMR, Shenton BK.The clinical significance of post-transplantation non-HLA an-tibodies in renal transplantation. Transplant Int 1995; 8: 214.
36. Dubel L, Farges O, Johanet C, Sebagh M, Bismuth H. Highincidence of anti-tissue antibodies in patients experiencingchronic liver allograft rejection. Transplantation 1998; 65:1072.
37. Fredrich R, Toyoda M, Czer LSC, et al. The clinical significanceof antibodies to human vascular endothelial cells after cardiactransplantation. Transplantation 1999; 67: 385.
38. Cooper JD, Billingham M, Egan T. A working formulation for thestandardization of nomenclature and for clinical staging ofchronic dysfunction in lung allografts. J Heart Lung Trans-plant 1993; 12: 713.
39. Lechner JF, LaVeck MA. A serum-free method for culturingnormal human bronchial epithelial cells at clonal density. JTiss Cult Methods 1985; 9: 43.
40. Gryniewicz G, Poenie M, Tsien RY. A new generation of Ca21
indicators with greatly improved fluorescein properties. J BiolChem 1985; 260: 3440.
41. Itoh Y, Yanagisawa M, Ohkubo S, et al. Cloning and sequenceanalysis of cDNA encoding the precursor of a human endothe-lium-derived vasoconstrictor peptide, endothelin: identity of
human and porcine endothelin. FEBS Lett 1988; 231: 440.42. Dahiya R, Lee C, Haughney PC, Chui R, Ho R, Deng G. Differ-
ential gene expression of transforming growth factors a and b,epidermal growth factor, keratinocyte growth factor, and theirreceptors in fetal and adult human prostatic tissues and cancercell lines. Urology 1996; 48: 963.
43. Trutmann M, Terracciano L, Noppen C, et al. GM-CSF geneexpression and protein production in human colorectal cancercell lines and clinical tumor specimens. Int J Cancer 1998; 77:378.
44. Duijvestijn AM, Van Breda Vriesman PJC. Chronic renal allo-graft rejection: Selective involvement of the glomerular endo-thelium in humoral immune reactivity and intravascular co-agulation. Transplantation 1991; 52: 195.
45. Cherry R, Nielsen H, Reed E, Reemtsma K, Suciu-Foca N, Mar-boe CC. Vascular (humoral) rejection in human cardiac allo-graft biopsies: Relation to circulating anti-HLA antibodies.J Heart Lung Transplant 1992; 11: 24.
46. Davenport A, Younie ME, Parson JEM, Klouda PT. Developmentof cytotoxic antibodies following renal allograft transplanta-tion is associated with reduced graft survival due to chronicvascular rejection. Nephrol Dial Transplant 1994; 9: 1315.
47. Reed EF, Hong B, Ho E, Harris PE, Weinberger J, Suciu-Foca N.Monitoring of soluble HLA alloantigens and anti-HLA antibod-ies identifies heart allograft recipients at risk of transplant-associated coronary artery disease. Transplantation 1996; 61:566.
48. Joyce S, Flye MW, Mohanakumar T. Characterization of kidneycell-specific, non-MHC alloantigen using antibodies elutedfrom rejected human renal allografts. Transplantation 1988;46: 362.
49. Winter JB, Clelland C, Gouw A, Prop J. Distinct phenotypes ofinfiltrating cells during acute and chronic lung rejection inhuman heart-lung transplants. Transplantation 1995; 59: 63.
50. Hasegawa S, Ockner DM, Ritter JH, et al. Expression of class IImajor histocompatibility complex antigens (HLA-DR) and lym-phocyte subset immunotyping in chronic pulmonary trans-plant rejection. Arch Pathol Lab Med 1995; 119: 432.
51. Yousem SA, Ray L, Paradis IL, Dauber JA, Griffith BP. Potentialrole of dendritic cells in bronchiolitis obliterans in heart-lungtransplantation. Ann Thorac Surg 1990; 49: 424.
Received 24 August 2000.Provisionally accepted 3 November 2000.Accepted 10 November 2000.
TRANSPLANTATION976 Vol. 71, No. 7
Copyright © 2022 FDOKUMEN




















