Acetyl salicylic acid and 24-epibrassinolide enhance root activity and improve root morphological...
9
This article was downloaded by: [University of Arid Agriculture] On: 22 April 2014, At: 22:29 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Acta Agriculturae Scandinavica, Section B - Soil & Plant Science Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/sagb20 Acetyl salicylic acid and 24-epibrassinolide enhance root activity and improve root morphological features in tomato plants under heat stress Abdul Rehman Khan a , Zhihui Cheng a , Bushra Ghazanfar a , Muhammad Azam Khan b & Zhu Yongxing a a College of Horticulture, Northwest Agriculture and Forestry University, Yangling, Shaanxi 712100, P.R China b PMAS Agriculture University, Rawalpindi, Pakistan Published online: 16 Apr 2014. To cite this article: Abdul Rehman Khan, Zhihui Cheng, Bushra Ghazanfar, Muhammad Azam Khan & Zhu Yongxing (2014): Acetyl salicylic acid and 24-epibrassinolide enhance root activity and improve root morphological features in tomato plants under heat stress, Acta Agriculturae Scandinavica, Section B - Soil & Plant Science, DOI: 10.1080/09064710.2014.906645 To link to this article: http://dx.doi.org/10.1080/09064710.2014.906645 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Acetyl salicylic acid and 24-epibrassinolide enhance root activity and improve root morphological...

This article was downloaded by: [University of Arid Agriculture]On: 22 April 2014, At: 22:29Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Acta Agriculturae Scandinavica, Section B - Soil &Plant SciencePublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/sagb20
Acetyl salicylic acid and 24-epibrassinolide enhanceroot activity and improve root morphologicalfeatures in tomato plants under heat stressAbdul Rehman Khana, Zhihui Chenga, Bushra Ghazanfara, Muhammad Azam Khanb & ZhuYongxinga
a College of Horticulture, Northwest Agriculture and Forestry University, Yangling,Shaanxi 712100, P.R Chinab PMAS Agriculture University, Rawalpindi, PakistanPublished online: 16 Apr 2014.
To cite this article: Abdul Rehman Khan, Zhihui Cheng, Bushra Ghazanfar, Muhammad Azam Khan & Zhu Yongxing(2014): Acetyl salicylic acid and 24-epibrassinolide enhance root activity and improve root morphological featuresin tomato plants under heat stress, Acta Agriculturae Scandinavica, Section B - Soil & Plant Science, DOI:10.1080/09064710.2014.906645
To link to this article: http://dx.doi.org/10.1080/09064710.2014.906645
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose ofthe Content. Any opinions and views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be reliedupon and should be independently verified with primary sources of information. Taylor and Francis shallnot be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and otherliabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

ORIGINAL ARTICLE
Acetyl salicylic acid and 24-epibrassinolide enhance root activity andimprove root morphological features in tomato plants under heat stress
Abdul Rehman Khana, Zhihui Chenga*, Bushra Ghazanfara, Muhammad Azam Khanb andZhu Yongxinga
aCollege of Horticulture, Northwest Agriculture and Forestry University, Yangling, Shaanxi 712100, P.R China;bPMAS Agriculture University, Rawalpindi, Pakistan
(Received 20 January 2014; accepted 14 March 2014)
High temperature has deleterious impacts on tomato growth and development and limits its production. Acetylsalicylic acid (ASA) and 24-epibrassinolide (EBL) have been widely reported as stress-ameliorating agents. Theeffect of exogenous application of varying levels of EBL (0.75, 1.5, and 3 µM) and ASA (0.25, 0.75, and 1.25mM) on root activity (RA) in terms of 2,3,5 triphenyl tetrazolium chloride (TTC) reduction and rootmorphological features was evaluated in four-week-old tomato seedlings (cultivar: Mei Jie Lo) grown underhigh-temperature stress (46°C/4 h per day) for 21 days. The daily heat stress treatment almost ceased the rootgrowth of chemically untreated seedlings. However, both EBL and ASA significantly attenuated the deleteriousimpacts of heat stress to different extents regarding root activity, total root length, surface area, volume, andnumber of nodes and connections. Different concentrations demonstrated signature effects. EBL (3 µM) wasover all the best treatment to improve root activity whereas ASA (0.25 mM) best enhanced root architecture(net length, volume, and area) as compared to the untreated heat-stressed controls. However, EBL (3 µM) andASA (1.25 mM) slightly inhibited mean root diameter. It is concluded that under high-temperature conditions,the exogenous EBL and ASA in studied doses improve root morphological features and root activity, henceenhance heat stress tolerance. Both chemical agents can be potential candidates in practical agriculture forextension of tomato growth period in summer by virtue of their heat stress amelioration ability.
Keywords: acetyl salicylic acid; heat tolerance; root architecture; tomato; 24-epibrassinolide
Introduction
Tomato (Lycopersicon esculentum Mill.) is an import-ant crop all over the world, consumed in cooked,processed, and fresh forms. It is heat-sensitive andextreme temperatures are a serious threat to tomatoproduction like many other crops. The high ambienttemperature in a significant proportion of the to-mato-growing areas of the world is one of the mostcrucial problems in tomato production (Peet et al.1997). Heat stress leads to inhibition in both veget-ative and reproductive plant growth (Hussain et al.2006; Singh et al. 2007) and causes decline in rootgrowth (Rodríguez et al. 2005).The application of plant growth regulators is recog-
nized to play an important role in plant response to
stress (Chakrabarti & Mukherjee 2003). Thus, induc-tion of thermotolerance in existing high-yielding culti-vars by foliar application with low concentrations ofgrowth hormones can be a fruitful effort (Wahid et al.2007). Steroidal hormones are a new class of planthormones named brassinosteroids. Their applicationimproved the basic thermotolerance in Brassica napusand tomato seedlings (Dhaubhadel et al. 1999; Singh& Shono 2005). Many workers demonstrated an up-regulatory role of brassionosteroids in root develop-ment in barley (Kartal et al. 2009), onions (Howellet al. 2007), Arabidopsis (Bao et al. 2004), maize(Romani et al. 1983), radish, and tomato (Takatsutoet al. 1983).Salicylic acid (SA) has been reported to induce
high-temperature stress tolerance (Dat et al. 1998;
*Corresponding author. Email: [email protected]
Acta Agriculturae Scandinavica, Section B – Soil & Plant Science, 2014http://dx.doi.org/10.1080/09064710.2014.906645
© 2014 Taylor & Francis
Dow
nloa
ded
by [
Uni
vers
ity o
f A
rid
Agr
icul
ture
] at
22:
29 2
2 A
pril
2014

Hayat et al. 2009; Senaratna et al. 2000). Acetylsalicylic acid (ASA) is a derivative of SA and itsapplication enhanced thermotolerance in potatomicroplants that facilitated the production of virus-free potato plants through tissue culture (Lopez-Delgado et al. 1998). Tomato seedlings grown fromseeds imbibed in aqueous solutions (0.1–0.5 mM) ofSA or ASA displayed enhanced tolerance and signi-ficant plant survival percentage against heat, chilling,and drought stresses (Senaratna et al. 2000).Root growth is sensitive to drought and heat
stresses and has a narrow optimum temperature rangewhen compared with other growth processes (Porter& Gawith, 1999). Roots play a critical role in planttolerance to heat stress, and maintaining viable andactive root growth is essential for plant growth andproductivity during periods of elevated air and soiltemperatures (Huang et al. 2012). An extensive rootsystem can contribute toward increased heat toler-ance by better moisture uptake and cooling effect byvirtue of increased transpiration (Kolb & Robberecht,1996; Xu & Huang, 2000a,b) and their involvementin hormone synthesis affecting shoot growth anddevelopment (McMichael & Burke 1994, 2002).Root viability and vitality can be assessed by 2,3,5triphenyl tetrazolium chloride (TTC) test (Ruf &Brunner 2003). The colorless TTC is reduced by themitochondrial dehydrogenases to red-colored triphe-nyl formazan. The efficiency of the red formazanformation depends on the activity of cytochromeoxidase (Block & Brouwer, 2002; Clemensson-Lindell 1994; Comas et al. 2000) and provides anestimate of dehydrogenase activity and rate of tissuerespiration (Richter et al. 2006; Steponkus 1971).Though the ameliorative role of 24-epibrassinolide
(EBL) and SA under heat stress has been extensivelydocumented in different crops, however effect of ASAand EBL in different concentrations on tomato rootgrowth and performance under heat stress conditionsyet remains an under-investigated area. Most of theworkers used a short-time exposure of seedlings toheat stress and then studied various indexes. But infield conditions, plants experience frequent heatstress. Opting a practical application approach, thepresent study was planned for manipulation of con-centration-specific plant responses under frequentheat stress and assessment of best suitable doses ofEBL and ASA as foliar application to determine theirrole in root development.
Materials and methods
Growing seedlings
Seeds of tomato cultivar Mei Jie Lo (MJL) weresterilized in 0.1% HgCl2 for two minutes, rinsed with
dH2O, and germinated in wet filter paper for 3 days ingrowth chamber at 26°C and 70% humidity in dark.The peat-based plant growth media was commerciallyprocured and was autoclaved (125 Pa/121°C) for 21minutes. The media was filled in plastic pots (length ×width × depth: 8 cm × 8 cm × 10 cm). Germinatedseeds were shifted in pots followed by shower irriga-tion and transferred to the growth chamber with 25°C/20°C day/night temperature, ~70% humidity level,and 12-h light period. Each seedling was supplied with25-ml commercial nutrition supplement on two leavesstage, in two splits.
Chemical treatments
24-epibrassinolide (EBL) and ASA were procuredfrom Sigma-Aldrich, Beijing, P.R. China. EBL wasdissolved in ethanol to make a stock solution andstored at –20°C for future preparation of concentra-tions as per treatment. However, ASA doses weredirectly prepared in distilled water using minimalquantity of ethanol as solvent. Both groups of tomatoplants were foliarly sprayed with respective doses ofEBL, ASA, or dH2O (controls) added with equalquantity of ethanol. The treated plants were thenkept at normal temperature (25°C/20°C: day/night,70% humidity, 4K lux light with 12-h day length) for3 days for satisfactory absorption. All the dosesincluding control were added with 0.1% tween-20as surfactant to ensure chemical absorption. Theeight treatments were: CLHT, control at high tem-perature; heat-stressed controls; CLNT, control atnormal temperature; EBL: E1, E2, E3 (0.75, 1.5,and 3.0 µM); ASA: A1, A2, A3 (0.25, 0.75, and 1.25mM), respectively.
Heat treatment
Two growth chambers were used with pre-adjustedinternal environmental conditions; 24±1°C day and20±1°C night temperatures, 12 h illumination periodand 70% humidity. In pre-experiments (data notshown), LT50, lethal temperature at which ~50%population of four-week-old tomato seedlings (MJLcultivar) would permanently wilt on 5-h exposure/daily for one week, was found to be 46°C at 70%humidity. Hence 46°C temperature was selected fordaily heat stress treatment for 21 days from 10.00 to14.00 h (4 h); keeping in view that it is the peak heatstress time in field conditions. After heat stresstreatment, the growth chamber would return tonormal temperature conditions daily. In order toavoid sudden heat shock, 46°C was gradually estab-lished: ~1°C increase for every three minutes. Pre-viously, Singh and Shono (2005) opted a heat stress of
2 A.R. Khan et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f A
rid
Agr
icul
ture
] at
22:
29 2
2 A
pril
2014

45°C for a brief exposure of 3 h for tomato (cultivar:Ailsa craig) seedlings. The CLNT plants were kept inother growth chamber keeping all conditions samebut with no heat treatment. All the plants wereirrigated with 20 ml of tap water on alternate daysand 70% humidity level was maintained by built-inauto-humidifier.
Root architecture
Plant functions are linked to the root system mor-phology and root activity (RA; Zhang et al. 2002).For root architecture studies, the roots were cleanedin running tap water by gentle agitation and thenscanned using Microtek Scan Maker i800plus scan-ner. Microtec Scan wizard EZ-2.3 software wasutilized to scan and analyze data regarding differentparameters of root morphological features. Theexperiment was conducted thrice and at least threeuniform roots were sampled from each treatment ineach replication.
Root activity
Root activity was estimated following the methodadapted by Onanuga et al. (2012) with slight mod-ifications. Briefly, at final harvesting stage on 25th dayof treatments (21 days of heat stress and 3 days ofrecovery time), at least three fresh root subsampleswere collected from uniform plants from each treat-ment, washed with dH2O thrice, blotted on filterpaper, and then stored at 4°C to be used same day.A standard curve was drawn based upon spectro-photometric absorption (λ = 485 nm) with varyingamounts of TTC (1g/0.1 l) solution with Na2S2O4
and ethyl acetate. The resulting linear equation; Y =0.355X, where R2 = 0.999 was used to calculate theroot activity. Roots were then sliced into one cen-timeter pieces, put into a test tube with 5ml of 1g/0.1 l(m/v) TTC, and 5 ml of 0.1 mol/l Na2HPO4–NaH2PO4 solution and incubated for 1h at 37°C inpreheated hot water bath. Then, 2 ml of 1 mol/lH2SO4 was added and the roots were taken out andground using mortar and pestle with 10 ml ethylacetate. The red colored extract was collected inseparate 10-ml test tubes and absorbance was spec-trophotometrically (spectrophotometer: UV-3802,UNIC, Shanghai, China) evaluated at λ = 485 nm.Root activity was calculated (Li 2000) as per linearequation obtained from standardization andexpressed as TTC reduction intensity: mg g–1 h–1:
TTC reduction intensity¼½TTC reductionmass ðmgÞ�/½root freshmass ðgÞ� time ðhÞ�
Experimental design and data analysis
Completely randomized design was followed and theexperiment was replicated thrice. The data wereanalyzed with one-way ANOVA and the differencesamong treatment means were determined by LSD atp < 0.05 using the analytical software Statistix-8.
Results
Root architecture
Root length
A significant reduction (72.2%) in total root lengthin the seedlings growing under heat stress in CLHT(386.7 cm) was recorded as compared to CLNT(1390.1 cm). However, all the chemical treatmentsattenuated the root length reduction impact andsignificantly enhanced total root length under heatstress conditions. The maximum root length amongthe heat-stressed seedlings was noted in A1 (1051.7cm) followed by E3 (910.8 cm; Figure 1a).
Surface area
Root surface area is an important parameter directlyattributed to nutrient absorption. The heat treatmentreduced the root surface area up to 82.7% in CLHTas compared with CLNT seedlings. Almost allchemical treatments enhanced the root surface areato varying extents but A1 (ASA: 0.25 mM) had bestresults (235.7 cm2; Figure 1b).
Root volume and diameter
The heat stress had severe deleterious impact on thetotal root volume and diameter in CLHT up to92.3% and 37.7% reduction, respectively, as com-pared with CLNT. Treatment A1 (0.25 mM) helpedseedlings to reduce this loss up to 76.7% and 32.2%for root volume and diameter, respectively. The restof concentrations A2, A3, E1, E2, and E3 alsosignificantly increased the total root volume tovarying extents as compared with the non-treatedheat-stressed seedlings (Figure 1c), whereas exceptA1, no treatment had any additive effect regardingmean root diameter, rather A3 (1.25 mM) and E3(3 µM) demonstrated a suppressive role (Figure 1d).
Number of root connections and nodes
Under heat stress conditions, number of root nodesand connections were significantly reduced (70.6%)and (74.9%), respectively, as compared to seedlingsat normal temperature. By virtue of chemicallyinduced amelioration, the total number of nodesand connections per root was significantly improved
Acta Agriculturae Scandinavica, Section B – Soil & Plant Science 3
Dow
nloa
ded
by [
Uni
vers
ity o
f A
rid
Agr
icul
ture
] at
22:
29 2
2 A
pril
2014

in all the chemical treatments, however, A1 (0.25 mM)produced highest number of both root parametersreducing the losses to 1.15% and 15.6%, respectively(Figure 1e).
Root activity
The heat stress reduced the root activity by 27.3% inCLHT seedlings as compared to CLNT. All the
ASA and EBL treatments significantly improvedroot respiratory activity as compared to CLHT interms of TTC reduction. E3 and E2 being the bestidentified doses, in this regard, enhanced rootactivity up to 70.7% and 67.3%, respectively, ascompared to CLHT seedlings. However, amongASA the best treatment was the middle level A2(0.75 mM) which improved root activity up to46.8% over CLHT. All three levels of EBL (0.75,
0
200
400
600
800
1000
1200
1400
1600
1800(a)
(b)
(c)
CLNT CLHT A1 A2 A3 E1 E2 E3
Tot
al r
oot l
engt
h (c
m)
a
f
b
cdecd
e de
bc
0
100
200
300
400
500
600
700
800
900
CLNT CLHT A1 A2 A3 E1 E2 E3
Roo
t sur
face
are
a (c
m2 )
a
d
b
cd cdd cd
c
0
5
10
15
20
25
30
35
40
45
Roo
t vol
ume
(cm
3 )
a
d
bbc cd c c c
CLNT CLHT A1 A2 A3 E1 E2 E3
Figure 1. Effects of exogenously applied EBL (E1, E2, E3: 0.25, 0.75, and 3 µM) and ASA (A1, A2, and A3: 0.25, 0.75,and 1.25 mM) concentrations on total length (a), total surface area (b), total volume (c), mean diameter (d), number of rootconnections and nodes (e), and root activity (f) of four weeks old seedlings of tomato (MJL cultivar) subjected to dailysevere heat stress (46°C/4 h) for three weeks. CLNT, control at normal temperature; CLHT, heat-stressed seedlings. Dataare the means of three replicates. Vertical bars refer to ±SE (n = 3). Means with different alphabets indicate significantdifference among treatments at p < 0.05. LSD test.
4 A.R. Khan et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f A
rid
Agr
icul
ture
] at
22:
29 2
2 A
pril
2014

1.5, and 3 µM) and A2 (0.75 mM) even supersededthe CLNT by 8.65%, 21.68%, 24.2%, and 6.8%,respectively (Figure 1f).
Discussion
Plant quality and survival capacity under hightemperatures depend upon root and shoot growth.Root morphological features can directly influencethe plant heat tolerance. Higher root number andincreased root length contributed to differences inheat tolerance among cultivars of Kentucky blue-grass (Lehman & Engelke, 1993).
In the present study, we observed a sharp declinein root architecture and root activity of tomatoseedlings grown under heat stress which confirmsthe findings of Huang and Fry (1998) who observedrestricted root growth and root metabolic activity inturf grass growing at 35°C. Root growth is moresensitive than shoot growth to temperature stressbecause of their lower optimal growth temperature.(Huang & Gao 2000; Xu & Huang 2000a,b).We observed different effects of different EBL
concentrations for certain studied parameters. Underheat stress conditions, EBL (1.5 and 3 µM) had asignificant promotional effect on the root growth
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Mea
n ro
ot d
iam
ater
(m
m)
a
bc b bcc bc bc
c
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000 No. of connections No. of nodesa
f
A
b
decd
e
c
e
A
BCB
C C
B
D
0
0.5
1
1.5
2
2.5
CLNT CLHT A1 A2 A3 E1 E2 E3
CLNT CLHT A1 A2 A3 E1 E2 E3
CLNT CLHT A1 A2 A3 E1 E2 E3
T
TC
red
uctio
n (
mg
g–1 h
–1)
b
dcd
b
c
ba a
(d)
(e)
(f)
Figure 1. (Continued)
Acta Agriculturae Scandinavica, Section B – Soil & Plant Science 5
Dow
nloa
ded
by [
Uni
vers
ity o
f A
rid
Agr
icul
ture
] at
22:
29 2
2 A
pril
2014
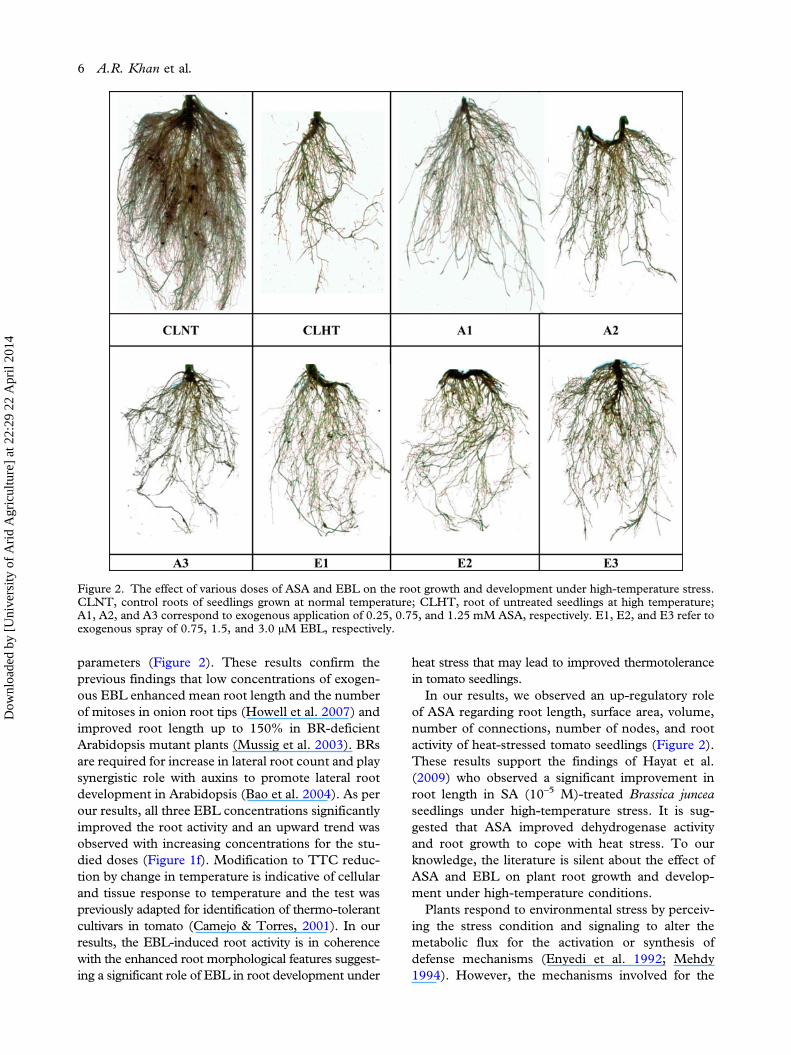
parameters (Figure 2). These results confirm theprevious findings that low concentrations of exogen-ous EBL enhanced mean root length and the numberof mitoses in onion root tips (Howell et al. 2007) andimproved root length up to 150% in BR-deficientArabidopsis mutant plants (Mussig et al. 2003). BRsare required for increase in lateral root count and playsynergistic role with auxins to promote lateral rootdevelopment in Arabidopsis (Bao et al. 2004). As perour results, all three EBL concentrations significantlyimproved the root activity and an upward trend wasobserved with increasing concentrations for the stu-died doses (Figure 1f). Modification to TTC reduc-tion by change in temperature is indicative of cellularand tissue response to temperature and the test waspreviously adapted for identification of thermo-tolerantcultivars in tomato (Camejo & Torres, 2001). In ourresults, the EBL-induced root activity is in coherencewith the enhanced root morphological features suggest-ing a significant role of EBL in root development under
heat stress that may lead to improved thermotolerancein tomato seedlings.In our results, we observed an up-regulatory role
of ASA regarding root length, surface area, volume,number of connections, number of nodes, and rootactivity of heat-stressed tomato seedlings (Figure 2).These results support the findings of Hayat et al.(2009) who observed a significant improvement inroot length in SA (10–5 M)-treated Brassica junceaseedlings under high-temperature stress. It is sug-gested that ASA improved dehydrogenase activityand root growth to cope with heat stress. To ourknowledge, the literature is silent about the effect ofASA and EBL on plant root growth and develop-ment under high-temperature conditions.Plants respond to environmental stress by perceiv-
ing the stress condition and signaling to alter themetabolic flux for the activation or synthesis ofdefense mechanisms (Enyedi et al. 1992; Mehdy1994). However, the mechanisms involved for the
Figure 2. The effect of various doses of ASA and EBL on the root growth and development under high-temperature stress.CLNT, control roots of seedlings grown at normal temperature; CLHT, root of untreated seedlings at high temperature;A1, A2, and A3 correspond to exogenous application of 0.25, 0.75, and 1.25 mM ASA, respectively. E1, E2, and E3 refer toexogenous spray of 0.75, 1.5, and 3.0 µM EBL, respectively.
6 A.R. Khan et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f A
rid
Agr
icul
ture
] at
22:
29 2
2 A
pril
2014

attenuation of high-temperature induced root declineby exogenous EBL and ASA in tomato seedlingsdeserve further investigations. It was observed thatboth chemicals offered protective effect, however,EBL contributed more toward biochemical activityand ASA better contributed toward structural mod-ifications in roots. We found that pretreatment oftomato seedlings (MJL cultivar) with lower concen-tration of ASA (0.25 mM) had more positive effect onthe studied parameters, whereas the middle andupper doses of EBL (1.5 and 3 µM) were moreeffective. This leads us to suggest study synergisticeffect of both EBL and ASA in varying concentrationsand combinations that could modify the plant defensesystem in field conditions.
FundingThis research work was supported by the project No.30370977 of National Natural Science Foundation, P.R.China.
References
Bao F, Shen J, Brady SR, Muday GK, Asami T, Yang Z.2004. Brassionosteroids interact with auxin to pro-mote lateral root development in Arabidopsis. PlantPhysiol. 1344:1624–1631.
BlockMD, BrouwerDD. 2002. A simple and robust in vitroassay to quantify the vigour of oilseed rape lines andhybrids. Plant Physiol Biochem. 40:845–852.
Camejo D, Torres W. 2001. High temperature effect ontomato pigment, protein content and cellular viability.Cultivos Tropicales. 223:13–17.
Chakrabarti N, Mukherjee S. 2003. Effect of phytohor-mones pretreatment on nitrogen metabolism in Vignaradiate under salt stress. Biol Plant. 36:63–66.
Clemensson-Lindell A. 1994. Triphenyl tetrazolium chlor-ide as an indicator of fine-root vitality and environ-mental stress in coniferous forest stands: applicationsand limitations. Plant Soil. 159:297–300.
Comas L, Eissenstat DM, Lakso AN. 2000. Assessing rootdeath and root system dynamics in a study of grapecanopy pruning. New Phytol. 147:171–178.
Dat JF, Foyer CH, Scott IM. 1998. Changes in salicylic acidand antioxidants during induced thermotolerance inmustard seedlings. Plant Physiol. 118:1455–1461.
Dhaubhadel S, Chaudhary S, Dobinson K, Krishna P. 1999.Treatment with 24-epibrassinolide, a brassinosteroidincreases the basic thermotolerance of Brassica napusand tomato seedlings. Plant Mol. Biol. 40:333–342.
Enyedi AJ, Yalpani N, Silverman P, Raskin I. 1992. Signalmolecules in systemic plant resistance to pathogensand pests. Cell. 70:879–886.
Hayat S, Masood A, Yusuf M, Fariduddin Q, Aqil A.2009. Growth of Indian mustard (Brassica juncea L.)in response to salicylic acid under high-temperaturestress. Braz. J. Plant Physiol. 21:187–195.
Howell WM, Keller GE, Kirkpatrick JD, Jenkins RL,Hunsinger RN, McLaughlin EW. 2007. Effect ofplant steroidal hormone, 24-EBL on the mitotic indexand growth of onion root tips. Gen Mol Res. 6:50–58.
Huang B, Fry JD. 1998. Root anatomical, physiologicaland morphological responses to drought stress for tallfescue cultivars. Crop Sci. 38:1017–1022.
Huang B, Gao, H. 2000. Growth and carbohydratemetabolism of creeping bentgrass in response toincreasing temperatures. Crop Sci. 40:1115–1120.
Huang B, Shimon R, Jichen X. 2012. Root carbon andprotein metabolism associated with heat tolerance. J.of Exp. Bot. 63:3455–3465.
Hussain T, Khan IA, Malik MA, Ali Z. 2006. BreedingPotential for high temperature tolerance in corn (Zeamays L.). Pakistan J. Bot. 38:1185–1195.
Kartal G, Temei A, Arican E, Gozukirmizi N. 2009.Effects of brassinosteroids on barley root growth,antioxidant system and cell division. Plant GrowthRegul. 58:261–267.
Kolb PF, Robberecht R. 1996. High temperature anddrought stress effects on survival of Pinus ponderosaseedlings. Tree Physiol. 16:665–672.
Lehman VG, Engelke MC. 1993. Heat resistance androoting potential of Kentucky bluegrass cultivars. IntTurf grass Soc Res J. 7:775–779.
Li HS. 2000. The experimental principle and technologyof plant physiology and biochemistry. Beijing: HigherEdu Press; p. 119–120.
Lopez-Delgado H, Dat JF, Foyer CH, Scott IM. 1998.Induction of thermo-tolerance in potatomicro-plants byacetyl salicylic acid and H2O2. J Exp Bot. 49:713–720.
McMichael BL, Burke JJ. 1994. Metabolic activity ofcotton roots in response to temperature. EnvironExp Bot. 34:201–206.
McMichael BL, Burke JJ. 2002. Temperature effects onroot growth. In: Waisel Y, Eshel Y, Kafkafi U, editors.Plant roots: the hidden half, 3rd ed. New York:Marcel Dekker; p. 717–728.
Mehdy MC. 1994. Active oxygen species in plant defenseagainst pathogens. Plant Physiol. 105:467–472.
Mussig C, Shine GH, Altman T. 2003. Brassinosteroidspromote root growth in Arabidopsis. Plant Physiol.133:1261–1271.
Onanuga AO, Pingan J, Sina A. 2012. Effect of phytohor-mones, phosphorus and potassium on cotton varieties(Gossypium hirsutum) root growth and root activitygrown in hydroponic nutrient solution. J Agri Sci.4:93–110.
Peet MM, Willits DH, Gardner R. 1997. Response ofovule development and post-pollen production pro-cesses in male-sterile tomatoes to chronic, sub-acutehigh temperature stress. J Exp Bot. 48:101–111.
Porter JR, Gawith M. 1999. Temperature and the growthand development of wheat: a review. Eur J Agron.10:23–36.
Richter AK, Emmanuel F, Ivano B. 2006. Polyphenols inthe woody roots of Norway spruce and Europeanbeech reduce TTC. Tree Physiol. 27:155–160.
Rodríguez M, Canales E, Borrás-Hidalgo O. 2005.Molecular aspects of abiotic stress in plants. Biotech-nol Appl. 22:1–10.
Romani G, Marre MT, Bonetti A, Cerana R, Lado P,Marre E. 1983. Effect of brassinosteroid on growthand electrogenic proton extrusion in maize rootsegments. Physiol Plant. 59:528–532.
Ruf M, Brunner I. 2003. Vitality of tree fine roots: reevalua-tion of the tetrazolium test. Tree Physiol. 23:257–263.
Senaratna T, Touchell D, Bunn E, Dixon K. 2000. Acetylsalicylic acid (Aspirin) and salicylic acid induce
Acta Agriculturae Scandinavica, Section B – Soil & Plant Science 7
Dow
nloa
ded
by [
Uni
vers
ity o
f A
rid
Agr
icul
ture
] at
22:
29 2
2 A
pril
2014

multiple stress tolerance in bean and tomato plants.Plant Growth Reg. 30:157–161.
Singh I, ShonoM. 2005. Physiological and molecular effectsof 24-epibrassinolide, a brassinosteroid on thermotoler-ance of tomato. Plant Growth Reg. 47:111–119.
Singh RP, Vara Prasad PV, Sunita K, Giri SN, Reddy KR.2007. Influence of high temperature and breeding forheat tolerance in cotton. Adv Agron. 93:313–385.
Steponkus PL. 1971. Effect of freezing on dehydrogenaseactivity and reduction of triphenyl tetrazolium chlor-ide. Cryobiology. 8:570–573.
Takatsuto S, Yazawa N, Ikekawa T, Takematsu T,Takeuchi Y, Koguchi M 1983. Structure, activityand relationship of brassionsteroids. Phytochemistry.22:2437–2441.
Wahid A, Gelani S, Ashraf M, Foolad MR. 2007. Heattolerance in plants: an overview. Environ Exp Bot.61:199–223.
Xu Q, Huang B. 2000a. Effects of air and soil temperatureon carbohydrate metabolism in creeping bentgrass.Crop Sci. 40:1368–1374.
Xu Q, Huang B. 2000b. Growth and physiologicalresponses of creeping bentgrass to differential shootand root temperatures. Crop Sci. 40:1363–1368.
Zhang L, Li JM, Wang HX. 2002. Physiological andecological responses of wheat (Triticum aestivm L.)root to cadmium stress. Chin J Soil Sci. 33: 61–65.
8 A.R. Khan et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f A
rid
Agr
icul
ture
] at
22:
29 2
2 A
pril
2014