
Accelerated evolution of CES7, a gene encoding a novel major urinary protein in the cat family
22
and Evolution. All rights reserved. For permissions, please e-mail: [email protected] The Author 2010. Published by Oxford University Press on behalf of the Society for Molecular Biology Accelerated Evolution of CES7, a Gene Encoding a Novel Major Urinary Protein in the Cat Family Li Gang, Jan E. Janecka, and William J. Murphy * Department of Veterinary Integrative Biosciences, College of Veterinary Medicine & Biomedical Sciences, Texas A&M University, College Station, TX 77843-4458 *Corresponding author: William Murphy, VIBS, Texas A&M University, 4458 TAMU, College Station, TX 77843-4458; EMAIL: [email protected] ; FAX: 979-845-9972. Key words: Evolution, Positive Selection, MUP, Cat Family Runing head: Rapid Evolution of CES7 Gene in Felidae MBE Advance Access published October 21, 2010 by guest on June 2, 2016 http://mbe.oxfordjournals.org/ Downloaded from
Transcript of Accelerated evolution of CES7, a gene encoding a novel major urinary protein in the cat family

and Evolution. All rights reserved. For permissions, please e-mail: [email protected] The Author 2010. Published by Oxford University Press on behalf of the Society for Molecular Biology
Accelerated Evolution of CES7, a Gene Encoding a Novel Major
Urinary Protein in the Cat Family
Li Gang, Jan E. Janecka, and William J. Murphy*
Department of Veterinary Integrative Biosciences, College of Veterinary Medicine & Biomedical Sciences, Texas A&M University, College Station, TX 77843-4458 *Corresponding author: William Murphy, VIBS, Texas A&M University, 4458 TAMU, College Station, TX 77843-4458; EMAIL: [email protected]; FAX: 979-845-9972. Key words: Evolution, Positive Selection, MUP, Cat Family
Runing head: Rapid Evolution of CES7 Gene in Felidae
MBE Advance Access published October 21, 2010 by guest on June 2, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

Abstract Cauxin is a novel urinary protein recently identified in the domestic cat that
regulates the excretion of felinine, a pheromone precursor involved in sociochemical communication and territorial marking of domestic and wild felids. Understanding the evolutionary history of cauxin may therefore illuminate molecular adaptations involved in the evolution of pheromone-based communication, recognition and mate selection in wild animals. We sequenced the gene encoding cauxin, CES7, in 22 species representing all major felid lineages, and multiple outgroups and show that it has undergone rapid evolutionary change preceding and during the diversification of the cat family. A comparison between feline cauxin and orthologous carboxylesterases from other mammalian lineages revealed evidence of strong positive Darwinian selection within and between several cat lineages, enriched at functionally important sites of the protein. The higher rate of radical amino acid replacements in small felids, coupled with the lack of felinine and extremely low levels of cauxin in the urine of the great cats (Panthera), correlates with functional divergence of this gene in Panthera, and its putative loss in the snow leopard. Expression studies found evidence for several alternatively spliced transcripts in testis and brain, suggesting additional roles in male reproductive fitness and behavior. Our work presents the first report of strong positive natural selection acting on a major urinary protein of non-rodent mammals, providing evidence for parallel selection pressure on the regulation of pheromones in different mammalian lineages, despite the use of different metabolic pathways. Our results imply that natural selection may drive rapid changes in the regulation of pheromones in urine among the different cat species, which in turn may influence social behavior, such as territorial marking, and conspecific recognition, therefore serving as an important mechanism for the radiation of this group of mammals.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Introduction Major urinary proteins (MUPs) play a critical role in transferring information
during urine-derived scent marking (Beynon and Hurst 2003). MUPs are well characterized in laboratory rodent species, and are known to be important for pheromone communication and individual recognition (Novotny et al. 1999; Hurst et al. 2001). Even though there is no direct evidence indicating MUPs play a role in reproductive isolation, the use of urine for scent-marking during the mating season, and social communication throughout the year, suggests MUPs may be excellent candidate genes. Molecular evolutionary characterization of MUPs may provide insight into the diversity and evolutionary history of chemical signaling compounds and the genetic basis of communication among wild animals (Emes et al. 2004).
Rodent MUPs belong to a multi-gene family produced by a series of duplication events, and are characterized by extremely high levels of polymorphism within and between different species. In wild mice, the diversity of expression of MUPs can exceed MHC (major histocompatibility complex) variation (Hurst et al. 2001). Comparative genomic analyses provide evidence that significant adaptive selection has driven the evolution of rat and mouse MUPS (Emes et al. 2004). The expansion of this gene family is unique to rodents, suggesting different proteomic pathways may serve to regulate pheromone-based communication across different mammalian lineages. Outside of the murid rodent lineage, however, very little research has sought to identify and characterize MUPs in other mammals.
With the exception of lions, domestic cats, and male cheetahs, most members of the cat family, Felidae, do not exhibit complex social behaviors, and direct interactions are often limited to mating and territorial disputes (Bekoff, Daniels, and Gittleman 1984; Kitchener 1999). Scent marking is therefore one of the most common and reliable signals to indirectly convey information regarding sex, reproductive status and territory (Rich and Hurst 1998; Kitchener 1999). Urine spraying/marking is one of the most important mechanisms for sociochemical communication used by cats (Verberne and Leyhausen 1976; Macdonald 1980; Mellen 1993) and suggests a tight linkage with reproductive behavior (Molteno, Sliwa, and Richardson 1998). Felinine is a low molecular mass, volatile compound identified in the urine of both sexes of different felids (Datta and Harris 1951; Datta and Harris 1953; Westall 1953). Felinine, which is known to be involved in scent marking, is regulated by sex hormones and slowly released at natural temperatures for more than 30 days (Hendriks et al. 1995; Rutherfurd et al. 2007; Hendriks et al. 2008).
While most mammals excrete low levels of urinary proteins, felines exhibit non-pathological proteinuria. Recently, Miyazaki et al. (2003) characterized a 70-kilodalton protein in the carboxylesterase (CES) superfamily, named cauxin, that comprises >90% of the total urinary protein content in domestic cats. It has not been detected in the urine of non-felid mammals. Cauxin is responsible for hydrolyzing the felinine precursor, 3-methylbutanolcysteinylglycine, to felinine and glycine (Miyazaki et
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

al. 2006b). The highest urinary concentration of cauxin is found in mature intact male domestic cats, relative to females and immature males. Many factors affect the excretion of cauxin across different species, including age and gender, which are closely correlated with levels of felinine (Miyazaki et al. 2006a). Cauxin is also present in species of the genus Panthera (i.e., lions, jaguars, tigers, & leopards), the sister lineage of all other living cat species, though at markedly lower (fivefold) concentrations (McLean et al. 2007). However, members of Panthera lack felinine production. This pattern suggests that urinary excretion of cauxin evolved once in the felid common ancestor, and that the large difference in excretion levels between the smaller felids and the big cats may be due to functional divergence of the protein following their diversification (McLean et al. 2007).
Given the role cauxin plays in regulating felinine production, as well as possibly serving as a carrier for other urinary compounds, we hypothesized that natural selection may be driving its evolution. We therefore sequenced all CES7 exons, and flanking intron regions, in 22 felid species. We found evidence for strong positive selection acting on CES7 in small cats, which excrete high levels of cauxin in their urine and produce the pheromone felinine. In contrast, big cats with low levels of urinary cauxin, and lacking felinine, had lower rates of nonsynonymous substitutions within CES7. This suggests a novel functional change within felids associated with pheromone production. Our results imply that adaptive evolution of CES7 confers rapid changes in the regulation of pheromone production and sociochemical communication, which may be important in the development of premating selection barriers.
Materials and Methods: Samples and sequencing To test for a signature of natural selection on CES7, we amplified and sequenced the coding gene sequences from multiple felids. We used the 2X genome sequence assembly of the domestic cat (felCat3) to design 12 PCR primer pairs (Table S1) flanking each annotated exon, which allowed for amplification and subsequent sequencing of the complete CES7 coding sequences, partial introns, and 3-UTR segments from the genomic DNA of 22 felines which represent all major lineages of Felidae (Supporting Information), Other primers were designed using domestic dog and cat genome sequence alignments in order to amplify and sequence the CES7 sequences of three carnivoran outgroup taxa (Table S1). Phylogenetic reconstruction and divergence time estimation Amino acid sequence alignments were performed with the software CLUSTAL-W1.81 (Thompson et al. 1997) and Bioedit (http://www.mbio.ncsu.edu/BioEdit/bioedit.html) with manual checking of alignment gaps and indels by eye. Nucleotide coding sequence alignments were aided by referencing the amino acid sequence alignment. Nucleotide substitution models and
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

parameters were estimated using Modeltest 3.7 using default settings (Posada and Crandall 1998). Phylogenetic tree reconstruction was performed with both maximum likelihood (ML) and Bayesian approaches. RAxML 7.0.0 (Stamatakis 2006) was used to search for the best ML tree and bootstrap analysis (500 replicates) and Mrbayes3.1 (Ronquist and Huelsenbeck 2003) was used to obtain Bayesian posterior probabilities (BPP) for each node with the best nucleotide substitution model suggested by Modeltest. For MrBayes analyses, one million generations were used for 6 simultaneous Markov chains, with the burnin set to 500,000 generations. We used Consel (Shimodaira and Hasegawa 2001) to perform approximately unbiased (AU), Kishino-Hasegawa (KH), and Shimodaira-Hasegawa (SH) tests to compare the CES7 gene tree topology with published species trees (Johnson et al. 2006), using default parameters to generate P-values. PAML4.4 (Yang 2007) was used to produce the log-likelihoods of site-patterns of the different trees, as input for the Consel analysis. We estimated divergence times for feline CES7 gene orthologues using the software estbranches (F84 model) and multidivtime (Thorne, Kishino, and Painter 1998; Kishino, Thorne, and Bruno 2001; Thorne and Kishino 2002) using published fossil constraints (Johnson et al. 2006). Chains were run for one million generations with a burnin of 100,000 generations.
Estimation of dN/dS ratios Coding sequences of 27 predicted orthologs of the CES7 gene were obtained from non-felid mammals available in the Ensembl database (Supporting Information). The software MEGA4 (Tamura et al. 2007) was used to estimate the average ratio of synonymous nucleotide substitution per synonymous site (dS) and nonsynonymous nucleotide substitutions per synonymous site (dN) among the different mammal lineages using the embedded Pamilo-Bianchi-Li method. To characterize discrete dN/dS (ω) variation among the different amino acid sites of the CES7 gene, we performed a sliding window analysis in SWAAP 1.0.2 (http://www.bacteriamuseum.org/SWAAP/ SwaapPage.htm), comparing felids with other mammals. The Nei and Gojobori (Nei and Gojobori 1986) method was used for the estimation of dN and dS with a window size of 180 nucleotides and 15 nucleotide sliding increments. To test for variation of the dN/dS ratio among different branches in the felid CES7 gene tree, the PAML4.4 package was used to analyze the 23 feline CES7 coding sequences using a series of models comparisons to explore patterns observed in the dN/dS ratios. Different models tested included the branch-specific model (the dN/dS ratio and the numbers of nonsynonymous/synonymous substitutions for each branch) and the site-specific model (the dN/dS ratio of amino acid sites to identify residues under positive selection). Branch-specific models involved the ‘free-ratio’, ‘one ratio’ and ‘two ratio’ models to represent the different presumed hypothesis. The ‘Free-ratio’ model assumes an independent ω value for each branch in the phylogenetic tree. By contrast the ‘one-ratio’ model assumes a constant ω value across all branches. Rejection of the ‘one-ratio’ model
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

indicates variation in the ω value among branches of the CES7 gene tree. The ‘two-ratio’ model was used to confirm positive selection pressure on different branches of the CES7 tree by setting different ω values to different branches of the phylogeny: branches with an estimated ω lower than one and branches with an estimated ω value higher than one (indicated by the results of the free-ratio test). To examine whether natural selection pressure acting on different parts of CES7 correlates with specific functional regions, we used two pairs of comparisons to identify positively selected amino acid sites: (1) the near neutral model (model 1a, null hypothesis) versus a positive selection model (model 2a, alternative hypothesis) to detect whether a signature of positive selection was present among the different amino acid positions. (2) Model 7 (beta) versus model 8 (beta & ω) was used to identify specific amino acid sites which are potentially under positive selection. Clade model (Bielawski and Yang 2004) tests were further used to assess variation in CES7 rates between big cats of the genus Panthera and the small felines, where cauxin expression and felinine production are markedly different (Hendriks et al. 1995; McLean et al. 2007). In these tests, rejection of the null hypothesis indicates a proportion of sites evolving under different selective constraints between the small and big cat clades. Estimation of amino acid properties
We evaluated the effects of amino acid substitution in each branch of the CES7 gene tree using the software TreeSAAP (Woolley et al. 2003). This program categorizes amino acids based on 31 physicochemical properties, and can be used to infer whether positive or negative selective pressures likely govern specific substitutions. The numbers of changes classified in categories 7 and 8 with high significance (p<0.001) were used as an index for the degree of radical amino acid substitution, suggesting structural changes to the protein.
RT-PCR and RACE Analysis We determined expression profiles of cauxin of adult male domestic cat tissues using RT-PCR, as described elsewhere (Murphy et al. 2006). Primer pairs were designed using the published domestic cat mRNA sequence (Suppl. Table 1). To examine variation in cauxin expression we assayed kidney, brain, testis, lymph node, spleen, thymus and muscle. We performed 5’ and 3’ RACE using nested internal primers as described previously to detect splicing variants (Murphy et al. 2006). We purified the RT-PCR and RACE products to obtain complete 5’ and 3’ transcripts and used cloning to isolate the independent transcripts. The positive clones were sequenced with traditional Sanger methodology on an ABI3730. Domestic cat brain and testis tissues were used in RACE experiments.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Results Sequence data The final concatenated sequence alignment was 5.6-kb including complete CES7 coding sequences, partial introns, and 3-UTR segments of 23 species of Felidae (GenBank accession numbers from HQ379709 to HQ379734). Felid CES7 coding sequences are shorter than other mammalian orthologs, due to a GT insertion that introduces a stop codon prior to the ancestral mammalian C-terminus (Holmes, Cox, and Vandeberg 2008), ablating 30 residues from the end. We identified numerous fixed differences between the 23 studied felids and other mammals. All of the felid CES7 genes that were sequenced showed intact coding sequences, with the exception of an 11-bp deletion in the 7th exon of the snow leopard (Panthera uncia). This deletion was verified in multiple unrelated individuals, and while we infer the snow leopard cauxin product is non-functional because it removes a large fraction of the second carbesterase domain (Fig. 1b), we were unable to examine its expression profile, and snow leopard urine was not included in previous biochemical studies (McLean et al. 2007).
Phylogenetic reconstruction Modeltest suggested GTR+G as the best fitting nucleotide substitution model for the CES7 sequence alignment. Phylogenetic tree reconstruction was performed using ML (RaxML) and Bayesian (Mr,Bayess) methods with the GTR+G model. Both analyses yielded identical topologies with proportionately similar statistical support levels (Fig. S1). The initial split in the CES7 gene tree separates the Panthera lineage (the “big cats”, subfamily Pantherinae), and other feline species (“small cats”, subfamily Felinae) (Fig. 2). Within the small cat clade, 7 monophyletic lineages supported by previous phylogenetic studies (Johnson et al. 2006) were also supported in the CES7 gene tree with high statistical support: domestic cat lineage (Felis), lynx lineage (Lynx), leopard cat lineage (Prionailurus), puma lineage (Puma and Acinonyx), bay cat lineage (Pardofelis), caracal lineage (Profelis) and ocelot lineage (Leopardus). Of these, five lineages (domestic cat, lynx, leopard cat, bay cat, and puma) clustered with high support (>99 to ML bootstrap value and 1.00 for Bayesian posterior probability [BPP]), a result consistent with previous species trees (Johnson et al. 2006). The main differences between the CES7 gene tree and the Johnson et al. (2006) species tree were the relationships between the small cat lineages. For example, in the CES7 gene tree the first and second divergences in the subfamily Felinae lead to the ocelot lineage, followed by the caracal lineage, and are well supported with moderate to high bootstrap/BPP values. In contrast, the bay cat lineage represents the most basal divergence within Felinae in the species tree (Johnson et al. 2006). The alternative CES7 gene tree topology was significantly better than the species tree, in likelihood ratio tests (p<0.001 in approximately unbiased [AU], as well as Shimodaira-Hasegawa [SH] tests), and were also supported by analyses of only intronic sequences (data not shown).
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Sliding window test results A comparison of average dN/dS ratio between felid CES7 genes and 50 additional mammalian orthologues showed very large differences between the two groups. The average dN/dS ratio based on a 50-taxon alignment of mammalian CES7 orthologs was 0.43. Independent calculations for different lineages produced dN/dS values ranging from 0.36 (rodents) to 0.45 (artiodactyls and hominids), 0.4 for non-felid carnivores, and 0.2 for canids respectively. By contrast, the average pairwise dN/dS value across 23 felid species was 1.07. This ratio fluctuates across the sequence (Fig. 1), with 4 peaks significantly greater that 1. These peaks were absent in all other mammals examined. Branch-specific models
The CES7 gene tree was used as the guide tree to run several tests of natural selection. The free ratio model produced dN/dS values >1 for nearly 50% of the branches in the felid CES7 tree (Fig. 2, Table S2). A further test for two ratio categories among the branches, those with ω values greater than one and those with ω values less than one, resulted in rejection of the one-ratio model in favor of the unrestricted two-ratio model (2∆ℓ = 15.9; df = 1; p < 0.001, Table 1). Moreover, the LRT between the unconstrained two-ratio model (alternative hypothesis, the estimated ω ratio of the suggested positively selected branches) and the constrained two-ratio model (null hypothesis, the ω ratio of the suggested positively selected branches fixed to 1) distinguished the former as significantly better than the latter (2∆ℓ = 5.8; df = 1; p < 0.05, Table 1).
Site-specific models The LRT test for site-specific models rejected model 1a and model 7 (Table 1) and identified 57 (of 545) amino acids that are under positive selection with dN/dS ratios approaching 5.09; 12 of these residues with high probability (Table S2). Five positively selected sites occur within conserved functional domains of carboxylesterase proteins, including both carbesterase family domains, and other domains known to be involved in disulfide bond formation (Fig. 1, Table S2).
Clade models All three clade model tests (Table S3) rejected the simpler models in favor of the complex model (Table 1) showing accelerated evolution of amino acid sites in the small cat clade following its divergence from the ancestor of Panthera in the late Miocene. Fourteen amino acids were identified as being under divergent selective pressures in the small cat clade with high probability (Table S3). Mapping these amino acids onto the predicted protein structure revealed three positively selected residues located in the first disulfide bond region, three in the helix region, and one site (410V) which is one of the three ‘side door’ residues (Fleming et al. 2005; Bencharit et al. 2006) that play an important role as part of the entrance to the active site of this enzyme.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Sequence divergence comparison Others factors aside from positive selection can contribute to dN/dS values that exceed 1. These include intense fluctuation of population size over a short period of time, and relaxation of functional constraint on some branches (Wyckoff, Wang, and Wu 2000; Zhang, Nielsen, and Yang 2005). Therefore, we compared the evolution of the felid CES7 sequences to 30 random coding and non-coding nucleotide gene segments (Johnson et al. 2006). These segments were randomly distributed among 13 autosomes and the sex chromosomes of the domestic cat genome, and sequenced from the same 23 Felidae species (Fig. 3). The average sequence divergence calculated for synonymous sites and adjacent non-coding sequence was comparable for both CES7 and other random gene segments. However, CES7 exons had three times as many substitutions compared to coding sequences of the other genes. Further, the rate of sequence divergence at nonsynonymous sites in those exons was four times higher in CES7 than in the other gene segments (Fig. 3), while divergence at intronic sequences between CES7 and random genes did not show this same tendency. The highest sequence divergence rate is found at nonsynonymous sites, but the intronic sequence rate differences between CES7 and random nuclear segments were the lowest (CES7intronic /Random intronic =1.08), which is expected as intronic substitutions are generally regarded to be neutral across most parts of the genome. The difference in synonymous substitution rates between CES7 and random segments is much lower (CES7synonymous/Random synonymous =1.6) than that of the non-synonymous substitutions (CES7non-synonymous/Random non-synonymous=4.52). These results show non-synonymous substitutions are contributing more to CES7 sequence divergence than random nuclear gene loci. Divergence time estimation and changes of amino acid properties Analyses using TreeSAAP indentified several felid lineages that had radical functional changes; more than 10 substitutions classified in categories 7 and 8, which are the most radical property change categories, indicating a highly significant (P<0.001) level of physicochemical divergence that may be caused by positive selection. These include the ancestral lineage leading to lynx, ocelot and the branches of domestic cat and jaguarundi (Fig. S2). Our estimates of divergence times for CES7 using a Bayesian relaxed clock framework revealed no significant correlation between the time interval and the levels of the radical physicochemical changes of CES7 gene on specific branches (Spearman’s rank correlation test, p> 0.05, Fig. S2). CES7 gene expression patterns in domestic cat. We used RT-PCR and rapid amplification of cDNA ends (RACE) (Frohman, Dush, and Martin 1988), to examine CES7 mRNA expression and potential splicing variation among different domestic cat tissues (see Fig. 4). The longest published transcript was found in the kidney, confirming previously published results (Miyazaki et al. 2003). However we found evidence of expression of the putative full-length transcript in testis,
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

which had not been reported. Additional RT-PCR reactions using internal sets of primers indicated transcription in a broader range of tissues, though expression levels in testis and kidney were always strongest, with brain being higher than most tissues as well (Fig. 4). Therefore we further examined transcript diversity using RACE, and found several novel transcripts in the brain and testis (Fig. 4). Two brain transcripts lack the first and second exons, with a novel amino-terminus. We also found two novel exons in the testis transcripts, denoted exon 1a and exon 9a. Exon1a is located ~37kb upstream from exon1 and encodes a novel 5’ UTR. The other novel transcript in testis is short, consisting of only exons 10 through 13 but with a novel exon 9a located in the intronic region between common exon 9 and 10. These exons were not present in EST and cDNA databases for other eutherian mammals and are therefore either unique to the domestic cat, or have yet to be characterized in other mammals.
Discussion
Major urinary proteins are important for individual recognition mechanisms in rodents (Hurst et al. 2001), and previous comparisons of mouse and rat MUP repertoires detected strong signals of adaptive evolution (Emes et al. 2004). However, the evolution of proteins present in the urine of non-rodent mammals remains largely undescribed. In felids, scent marking through urine spraying serves important social functions. Felinine is the major felid-specific chemical element of urine that produces an olfactory response in domestic cats (Hendriks et al. 1995). Physiological research has confirmed the feline major urinary protein cauxin, encoded by the CES7 gene, is critical for the catalysis of 3-methylbutanol-glutathione (3-MBG), the precursor of felinine. The 3-MBG compound only occurs in blood, and is transformed into felinine in the kidney (Miyazaki et al. 2006b). Felinine has been indentified in the urine of members of multiple small cat lineages, including the domestic cat, leopard cat, lynx and ocelot (Datta and Harris 1951; Datta and Harris 1953; Hendriks et al. 1995).
We observed evidence of strong positive natural selection in CES7 sequences across multiple clades of cats. The comparison of felid CES7 coding sequences with other mammalian orthologs emphasizes the remarkably higher dN/dS ratio in felids. The highest dN/dS ratio peaks correspond to two important functional sites in carboxylesterase proteins, which include disulfide bond formation and a CES family protein motif region. This pattern was also supported by detection of numerous amino acid sites under positive selection. Based on the predicted structure information (Holmes, Cox, and Vandeberg 2008), 3 sites were located in the signal peptide, 15 sites located in the helix region, 4 sites in disulfide bond region, and 2 sites in the CES family protein motif region. Two residues that were under particularly strong selection, 410V and 411L, are in the ‘side door’ region of carboxylesterase orthologs, which is an area critical for the interaction of this enzyme with its substrates (Redinbo and Potter 2005; Bencharit et al. 2006; Holmes, Cox, and Vandeberg 2008).
In 6 of 7 small cat lineages (Johnson et al. 2006), nonsynonymous substitutions
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

were fixed faster than synonymous substitutions. Several evolutionary mechanisms may explain varying dN/dS ratios across different lineages, including bottlenecks, population expansion, and fluctuations in effective population size and generation time (Nielsen 2005). However, our comparison of the sequence evolution patterns in CES7 with 30 gene segments from different autosomes and both sex chromosomes indicated a nearly threefold higher substitution rate in the coding sequence of CES7 than in the other genes. Relaxed selection cannot explain the higher DNA mutation rates of nonsynonymous sites when compared to presumably neutral synonymous and intronic sites. Our results are therefore more consistent with positive selection driving the evolution of CES7, rather than relaxation of functional constraint. In addition, the published functional and behavioral data demonstrate urinary cauxin excretion is unique to the cat family (Miyazaki et al. 2008) and that it correlates with production of a hormone known to elicit behavioral responses in cats. This functional importance of cauxin is consistent with the positive selection patterns we observed and also argues against relaxed constraint as a mechanism for the accumulated fixed differences in the protein.
The biochemical context of several mutations that were under positive selection is consistent with a scenario involving ongoing adaptive evolution of CES7 prior to and during the felid radiation. Crystal structure analysis of human orthologs, carboxylesterase 1 (CES1), suggests three critical regions largely responsible for the precise function of this protein (Bencharit et al. 2003; Bencharit et al. 2006). Sites in these regions are conserved in other members of the CES family, including cauxin. These include the active site, the Z-site, and the side door. The predicted ‘Z-site’ of cauxin (residue 347) is believed to play a crucial role in the binding of substrates to the active site (Fleming et al. 2005; Redinbo and Potter 2005; Bencharit et al. 2006). Multispecies alignments show this residue to be highly conserved across mammals. In felids, however, a second codon position G to C mutation substitutes alanine for glycine. The known function and strong conservation at this site in non-felid mammals suggests its importance for enzymatic activity of cholesterol ester metabolism was altered in the felid ancestor.
Further evidence for adaptive functional divergence of cauxin is based on the markedly higher levels of urinary cauxin expression in members of small cat lineages relative to large pantherine cats, which do not excrete felinine as a pheromone (McLean et al. 2007). Accordingly, CES7 dN/dS ratios are much higher in the small cat lineages, with the distribution of positively selected sites concentrated in the disulfide bond and helix regions that are involved in determining protein structure. These results imply that different selection patterns acted on CES7 in the two major clades of cats, following the split from the ancestral modern felid, perhaps as a consequence of sub-functional divergence of cauxin between small and big cats and its role in regulating pheromones in different species.
Although the functional role of cauxin in big cats is unknown, the low levels of cauxin excreted in the urine of the four big cats studied (tiger, lion, jaguar, and leopard) coupled with lack of felinine production, may imply a change in function for members of
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

this lineage. Such a change is supported by the frame shift mutation we found in the snow leopard. The most recent studies assaying urinary cauxin excretion did not include samples from a snow leopard, so it is unknown whether the identification here of a putative loss of function mutation (11-bp deletion) translates to a reduction in, or lack thereof, cauxin excretion in snow leopard urine. The snow leopard CES7 protein may exist as an alternatively spliced form that excludes this exon, though to test this hypothesis would require appropriate tissues samples. Alternatively, CES7 may have retained an ancestral function in reproductive tissues, or acquired a new function in the big cats.
Relevant to this discussion is the recent finding of additional MUPs expressed in the urine of tiger, some of which are not reported as being present in the domestic cat or other small cat species (McLean et al. 2007). One of these novel MUPs is transthyretin, a carrier protein for retinol in human blood (Hamilton and Benson 2001). Recent data from our lab has shown that the transthyretin gene (TTR) has accumulated an inordinate number of substitutions per unit time in the early branches leading to the two major lineages of Panthera (Davis, Li, and Murphy 2010), while no substitutions distinguish lion/leopard/jaguar or snow leopard/tiger. Hence the phylogenetic signal indicates TTR is strongly tracking the early phylogeny of pantherids (Davis, Li, and Murphy 2010), suggesting this MUP has been shaped by adaptive evolutionary divergence, perhaps where it played a role in premating selection by regulating pheromone levels in big cats that lack felinine. Evaluating these hypotheses will require further biochemical and proteomic characterization of MUPs and pheromones in the big cats.
While most members of the CES gene superfamily are expressed and excreted in liver and intestine with the function of detoxification and metabolism of carboxylesters, aromatic esters and thioesters (Hosokawa et al. 1995), CES7 is unique in that it is expressed in a variety of different tissues across mammals: cerebrospinal fluid of the human brain (Ota et al. 2004), testis (Holmes, Cox, and Vandeberg 2008) and male reproductive fluids of several species (e.g. sheep, pig, mouse and domestic cat (Ecroyd et al. 2006)) playing a transfer role for lipid and cholesterol. Recent studies found extremely low mRNA levels in the kidney of ram, but with no cauxin protein detected in this organ. By contrast, domestic cat CES7 expression levels are highest in kidney (Miyazaki et al. 2003), which has not been reported in other mammal species and likely represents a lineage-specific innovation to regulate production of felinine that is unique among mammals. While our RT-PCR results confirm highest expression in the kidney, we did identify several novel transcripts in the brain and testis. These results suggest multiple functions of CES7 in different tissues are regulated by alternative splicing, some of which may relate to male reproductive fitness.
Feline urine carries information such as gender, age, health, and reproductive status (Molteno, Sliwa, and Richardson 1998). A comparative study of 20 species from 7 lineages of the small cat clade supports the remarkable uniformity of urine marking and other social behaviors (Mellen 1993). Moreover, European lynxes have been shown to
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

discriminate individuals based on species, sex and age, but also can detect different conspecific individuals through urinary odors (Sokolov, Naidenko, and Serbenyuk 1996). These behaviors are especially critical during the mating season. Felinine is thought to be an important territorial maker and may function to attract members of the opposite sex (Hendriks et al. 1995; Tarttelin, Hendriks, and Moughan 1998). Indeed, behavioral tests show that cats express excessive interest when 3-mercapto-3-methyl-1-butanol, a derivative of felinine, is presented (Miyazaki et al. 2008). In vitro enzyme assays also reveal a significant correlation between the concentration of cauxin and felinine in the urine of intact (non-neutered/spayed) cats of different gender and age, while the concentration of the felinine precursor does not vary by gender (Miyazaki et al. 2006b). Taken together, these studies suggest cauxin regulates, via interaction with sex hormones, the urinary concentration of felinine in different sexes, and during different phases of sexual maturity (Miyazaki et al. 2006a; Miyazaki et al. 2006b).
Accelerated evolution of reproductive genes, particularly those involved in sperm competition and gamete recognition, has provided strong evidence for positive selection shaping postmating isolation mechanisms (Kelley and Swanson 2008). Whole genome comparisons of mammals reveal reproductive gene repertoires vary considerably between species, including closely related species pairs such as chimpanzee and human (Swanson and Vacquier 2002). Research on Drosophila provides evidence for natural selection acting on genes responsible for premating selection (e.g. sensory systems, behavior, habitat preference), both at the sequence level (Vieira, Sanchez-Gracia, and Rozas 2007) and expression level (Shirangi et al. 2009). By contrast, research into the molecular evolution of premating selection in vertebrates has been very limited. Based on our observations of significant positive Darwinian selection driving the evolution of CES7 in felids we suggest that the novel amino acid changes enriched at functional sites within cauxin may alter the regulation of felinine production across cat species, possibly influencing species recognition and mating partner choice. Although we currently have no direct evidence that cauxin, or other uncharacterized MUPs, have a role in reproductive isolation, we hypothesize that the rapid evolution of CES7 suggests that it may be involved in the development of premating barriers among recently diverged cat species. The function and rapid evolution of CES7, when contrasted with other mammals, indicates that urine based pheromone communication systems evolve rapidly. Further genomic and proteomic studies of mammalian MUPs will provide an excellent opportunity to better understand the molecular mechanisms of urinary communication, recognition and mate selection in free ranging species.
Acknowledgments We thank Emma Teeling and Brian Davis for helpful discussion and two anonymous reviewers for their helpful comments. This work was funded in part by NSF grant EF0629849 to WJM.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

References
Bekoff M, Daniels TJ, Gittleman JL. 1984. Life history patterns and the comparative social ecology of carnivores. Annu Rev Ecol Syst.15:191-232.
Bencharit S, Edwards CC, Morton CL, Howard-Williams EL, Kuhn P, Potter PM, Redinbo MR. 2006. Multisite promiscuity in the processing of endogenous substrates by human carboxylesterase 1. J Mol Biol. 363:201-214.
Bencharit S, Morton CL, Xue Y, Potter PM, Redinbo MR. 2003. Structural basis of heroin and cocaine metabolism by a promiscuous human drug-processing enzyme. Nature Struct Biol. 10:349-356.
Beynon RJ, Hurst JL. 2003. Multiple roles of major urinary proteins in the house mouse, Mus domesticus. Biochem Soc Trans. 31:142-146.
Bielawski JP, Yang ZH. 2004. A maximum likelihood method for detecting functional divergence at individual codon sites, with application to gene family evolution. J Mol Evol. 59:121-132.
Datta SP, Harris H. 1951. A convenient apparatus for paper chromatography - results of a survey of the urinary amino-acid patterns of some animals. J Physiol (Lond). 114:P39-P41.
Datta SP, Harris H. 1953. Urinary amino acid patterns of some mammals. Ann Eugen. 18:107-116.
Davis BW, Li G, Murphy WJ. 2010. Supermatrix and species tree methods resolve phylogenetic relationships within the big cats, Panthera (Carnivora: Felidae). Mol Phylogenet Evol. 56:64-76.
Ecroyd H, Belghazi M, Dacheux JL, Miyazaki M, Yamashita T, Gatti JL. 2006. An epididymal form of cauxin, a carboxylesterase-like enzyme, is present and active in mammalian male reproductive fluids. Biol Reprod. 74:439-447.
Emes RD, Beatson SA, Ponting CP, Goodstadt L. 2004. Evolution and comparative genomics of odorant- and pheromone-associated genes in rodents. Genome Res. 14:591-602.
Fleming CD, Bencharit S, Edwards CC, Hyatt JL, Tsurkan L, Bai F, Fraga C, Morton CL, Howard-Williams EL, Potter PM, Redinbo MR. 2005. Structural insights into drug processing by human carboxylesterase 1: tamoxifen, mevastatin, and inhibition by benzil. J Mol Biol. 352:165-177.
Frohman MA, Dush MK, Martin GR. 1988. Rapid production of full-length cDNAs from rare transcripts: amplification using a single gene-specific oligonucleotide primer. Proc Natl Acad Sci U S A. 85:8998-9002.
Hamilton JA, Benson MD. 2001. Transthyretin: a review from a structural perspective. Cell Mol Life Sci. 58:1491-1521.
Hendriks WH, Moughan PJ, Tarttelin MF, Woolhouse AD. 1995. Felinine: a urinary amino acid of Felidae. Comp Biochem Physiol B. 112:581-588.
Hendriks WH, Rutherfurd-Markwick KJ, Weidgraaf K, Ugarte C, Rogers QR. 2008.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Testosterone increases urinary free felinine, N-acetylfelinine and methylbutanolglutathione excretion in cats (Felis catus). J Anim Physiol Anim Nutr (Berl). 92:53-62.
Holmes RS, Cox LA, Vandeberg JL. 2008. Mammalian carboxylesterase 5: Comparative biochemistry and genomics. Comp Biochem Physiol Part D. 3:195-204.
Hosokawa M, Endo T, Fujisawa M, Hara S, Iwata N, Sato Y, Satoh T. 1995. Interindividual variation in carboxylesterase levels in human liver microsomes. Drug Metab Dispos. 23:1022-1027.
Hurst JL, Payne CE, Nevison CM, Marie AD, Humphries RE, Robertson DH, Cavaggioni A, Beynon RJ. 2001. Individual recognition in mice mediated by major urinary proteins. Nature 414:631-634.
Johnson WE, Eizirik E, Pecon-Slattery J, Murphy WJ, Antunes A, Teeling E, O'Brien SJ. 2006. The late Miocene radiation of modern Felidae: a genetic assessment. Science 311:73-77.
Kelley JL, Swanson WJ. 2008. Positive selection in the human genome: from genome scans to biological significance. Annu Rev Genomics Hum Genet. 9:143-160.
Kishino H, Thorne JL, Bruno WJ. 2001. Performance of a divergence time estimation method under a probabilistic model of rate evolution. Mol Biol Evol. 18:352-361.
Kitchener AC. 1999. Watch with mother: A review of social learning in the Felidae. In: Box HO, Gibson KR, editors. Mammalian social learning: Comparative and ecological perspectives. Cambridge: Cambridge University Press. p. 236-258.
Macdonald DW. 1980. Patterns of scent marking with urine and faeces amongst carnivore communities. Symp Zool Soc Lond. 45: 106–121.
McLean L, Hurst JL, Gaskell CJ, Lewis JC, Beynon RJ. 2007. Characterization of cauxin in the urine of domestic and big cats. J Chem Ecol. 33:1997-2009.
Mellen JD. 1993. A comparative analysis of scent-marking, social and reproductive behavior in 20 species of small cats (Felis). Amer Zool. 33:151-166.
Miyazaki M, Kamiie K, Soeta S, Taira H, Yamashita T. 2003. Molecular cloning and characterization of a novel carboxylesterase-like protein that is physiologically present at high concentrations in the urine of domestic cats (Felis catus). Biochem J. 370:101-110.
Miyazaki M, Yamashita T, Hosokawa M, Taira H, Suzuki A. 2006a. Species-, sex-, and age-dependent urinary excretion of cauxin, a mammalian carboxylesterase. Comp Biochem Physiol B. 145:270-277.
Miyazaki M, Yamashita T, Suzuki Y, Saito Y, Soeta S, Taira H, Suzuki A. 2006b. A major urinary protein of the domestic cat regulates the production of felinine, a putative pheromone precursor. Chem Biol. 13:1071-1079.
Miyazaki M, Yamashita T, Taira H, Suzuki A. 2008. The biological function of cauxin, a major urinary protein of the domestic cat (Felis catus). In: Hurst JL, Beynon RJ, Roberts, SC, Wyatt T, editors. Chemical Signals in Vertebrates 11. Springer. p. 51–60.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Molteno AJ, Sliwa A, Richardson PRK. 1998. The role of scent marking in a free-ranging, female black-footed cat (Felis nigripes). J Zool. 245:35-41.
Murphy WJ, Wilkerson AJP, Raudsepp T, Agarwala R, Schaffer AA, Stanyon R, Chowdhary BP. 2006. Novel gene acquisition on carnivore Y chromosomes. PLoS Genet. 2:353-363.
Nei M, Gojobori T. 1986. Simple methods for estimating the numbers of synonymous and nonsynonymous nucleotide substitutions. Mol Biol Evol. 3:418-426.
Nielsen R. 2005. Molecular signatures of natural selection. Annu Rev Genet. 39:197-218. Novotny MV, Ma WD, Wiesler D, Zidek L. 1999. Positive identification of the
puberty-accelerating pheromone of the house mouse: the volatile ligands associating with the major urinary protein. Proc R Soc Lond (Biol). 266:2017-2022.
Ota T, Suzuki Y, Nishikawa T, et al. (154 co-authors). 2004. Complete sequencing and characterization of 21,243 full-length human cDNAs. Nat Genet. 36:40-45. (Please note that 154 includes all authors of the article.)
Posada D, Crandall KA. 1998. MODELTEST: testing the model of DNA substitution. Bioinformatics 14:817-818.
Redinbo MR, Potter PM. 2005. Mammalian carboxylesterases: from drug targets to protein therapeutics. Drug Discov Today. 10:313-325.
Rich TJ, Hurst JL. 1998. Scent marks as reliable signals of the competitive ability of mates. Anim Behav. 56:727-735.
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19:1572-1574.
Rutherfurd SM, Kitson TM, Woolhouse AD, McGrath MC, Hendriks WH. 2007. Felinine stability in the presence of selected urine compounds. Amino Acids 32:235-242.
Shimodaira H, Hasegawa M. 2001. CONSEL: for assessing the confidence of phylogenetic tree selection. Bioinformatics 17:1246-1247.
Shirangi TR, Dufour HD, Williams TM, Carroll SB. 2009. Rapid evolution of sex pheromone-producing enzyme expression in Drosophila. Plos Biol. 7: e1000168.
Sokolov VE, Naidenko SV, Serbenyuk MA. 1996. Recognition by the European lynx (Lynx lynx, Felidae, Carnivora) of the species and sex and age of conspecific, familiar, and unfamiliar individuals according to urinary odors. Izv Akad Nauk Ser Biol. 5:571-577.
Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22:2688-2690.
Swanson WJ, Vacquier VD. 2002. The rapid evolution of reproductive proteins. Nat Rev Genet. 3:137-144.
Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol. 24:1596-1599.
Tarttelin MF, Hendriks WH, Moughan PJ. 1998. Relationship between plasma testosterone and urinary felinine in the growing kitten. Physiol Behav. 65:83-87.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. 1997. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25:4876-4882.
Thorne JL, Kishino H. 2002. Divergence time and evolutionary rate estimation with multilocus data. Syst Biol. 51:689-702.
Thorne JL, Kishino H, Painter IS. 1998. Estimating the rate of evolution of the rate of molecular evolution. Mol Biol Evol. 15:1647-1657.
Verberne G, Leyhausen P. 1976. Marking behaviour of some Viverridae and Felidae: time-interval analysis of the marking pattern. Behaviour 58:192-253.
Vieira FG, Sanchez-Gracia A, Rozas J. 2007. Comparative genomic analysis of the odorant-binding protein family in 12 Drosophila genomes: purifying selection and birth-and-death evolution. Genome Biol. 8:R235.
Westall RG. 1953. The amino acids and other ampholytes of urine. 2. The isolation of a new sulphur-containing amino acid from cat urine. Biochem J. 55:244-248.
Woolley S, Johnson J, Smith MJ, Crandall KA, McClellan DA. 2003. TreeSAAP: Selection on amino acid properties using phylogenetic trees. Bioinformatics 19:671-672.
Wyckoff GJ, Wang W, Wu CI. 2000. Rapid evolution of male reproductive genes in the descent of man. Nature 403:304-309.
Yang Z. 2007. PAML 4: phylogenetic analysis by maximum likelihood. Mol Biol Evol. 24:1586-1591.
Zhang J, Nielsen R, Yang Z. 2005. Evaluation of an improved branch-site likelihood method for detecting positive selection at the molecular level. Mol Biol Evol. 22:2472-2479.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from
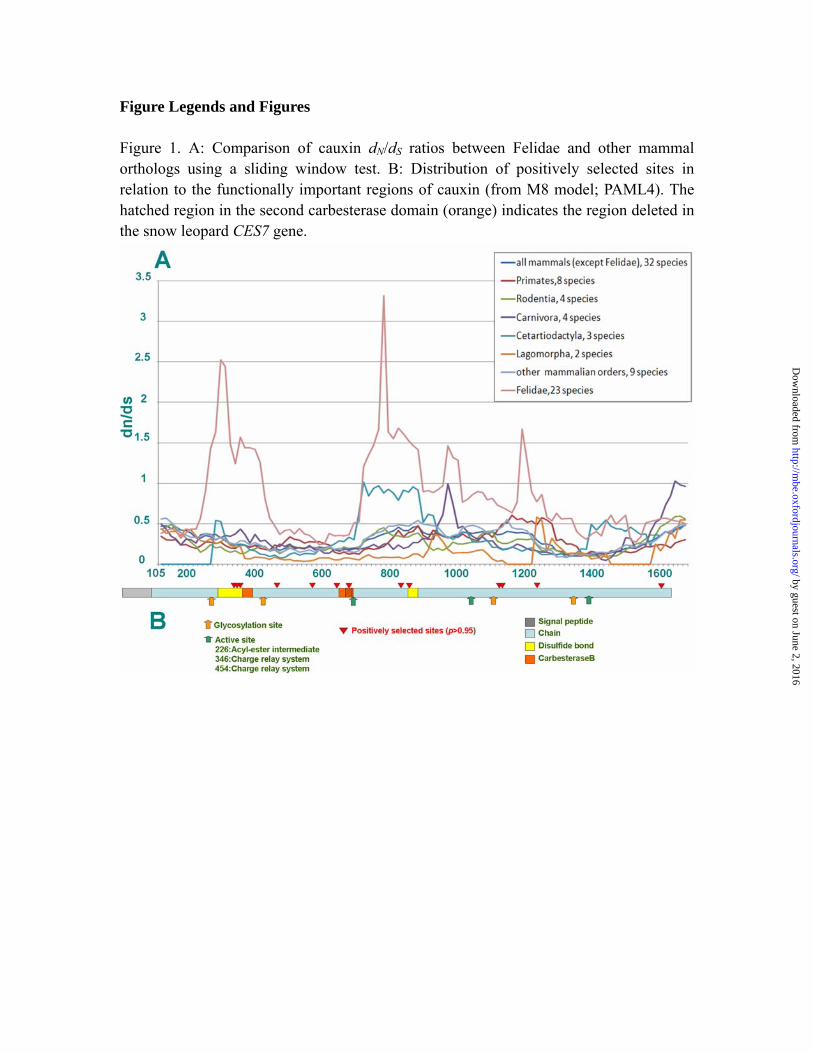
Figure Legends and Figures Figure 1. A: Comparison of cauxin dN/dS ratios between Felidae and other mammal orthologs using a sliding window test. B: Distribution of positively selected sites in relation to the functionally important regions of cauxin (from M8 model; PAML4). The hatched region in the second carbesterase domain (orange) indicates the region deleted in the snow leopard CES7 gene.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Figure 2. Free ratio results of CES7 gene in select members of Felidae. The topology shown is based on the ML gene tree. The number shown above each branch is the estimated dN/dS ratio (branches with a dN/dS ratio greater than one are shown in orange color), while numbers below the branches are the estimated numbers of nonsynonymous substitutions (before the diagonal) and estimated numbers of synonymous substitutions (after the diagonal).
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from
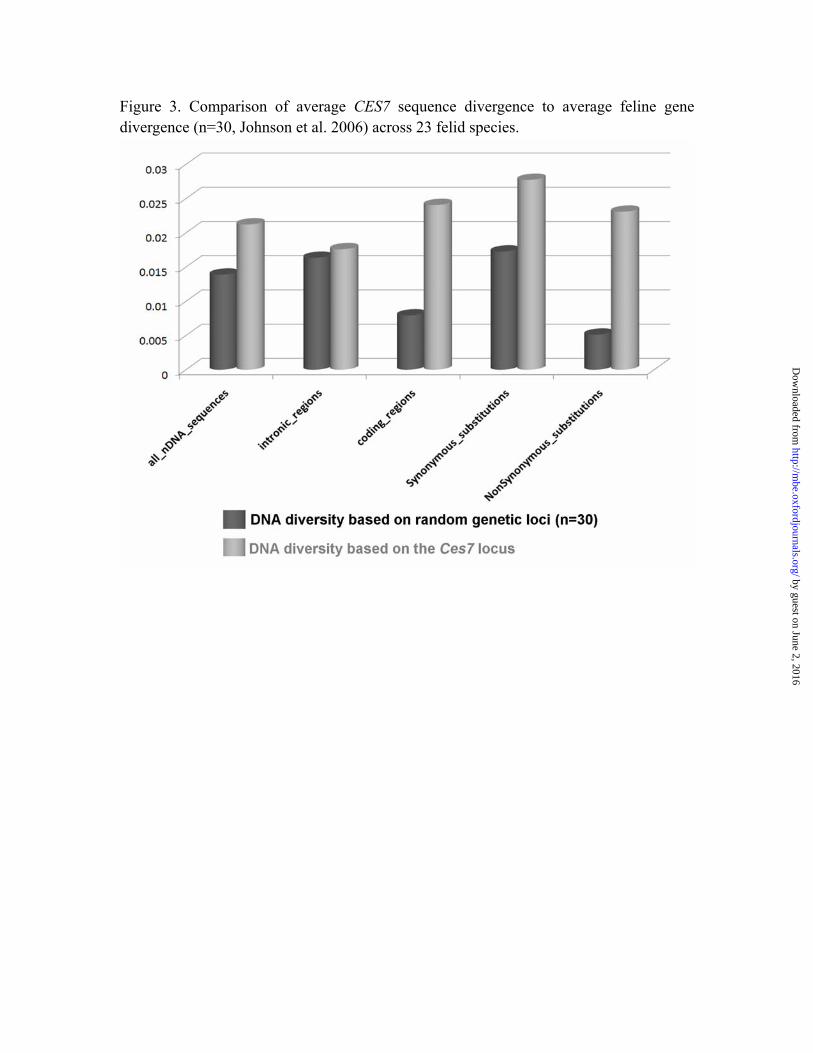
Figure 3. Comparison of average CES7 sequence divergence to average feline gene divergence (n=30, Johnson et al. 2006) across 23 felid species.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from

Figure 4. Results from RT-PCR assays of CES7 expression in 7 domestic cat tissues using different primer combinations (TH: Thymus; SP: Spleen; LN: Lymph node; TS: Testes; KD: Kidney; BN: Brain; MU: Muscle). The position of the primers are shown with reference to the CES7 gene structure map (bottom), and different splicing forms detected by RACE. Gray boxes are coding exons, white boxes are non-coding exons.
by guest on June 2, 2016http://m
be.oxfordjournals.org/D
ownloaded from
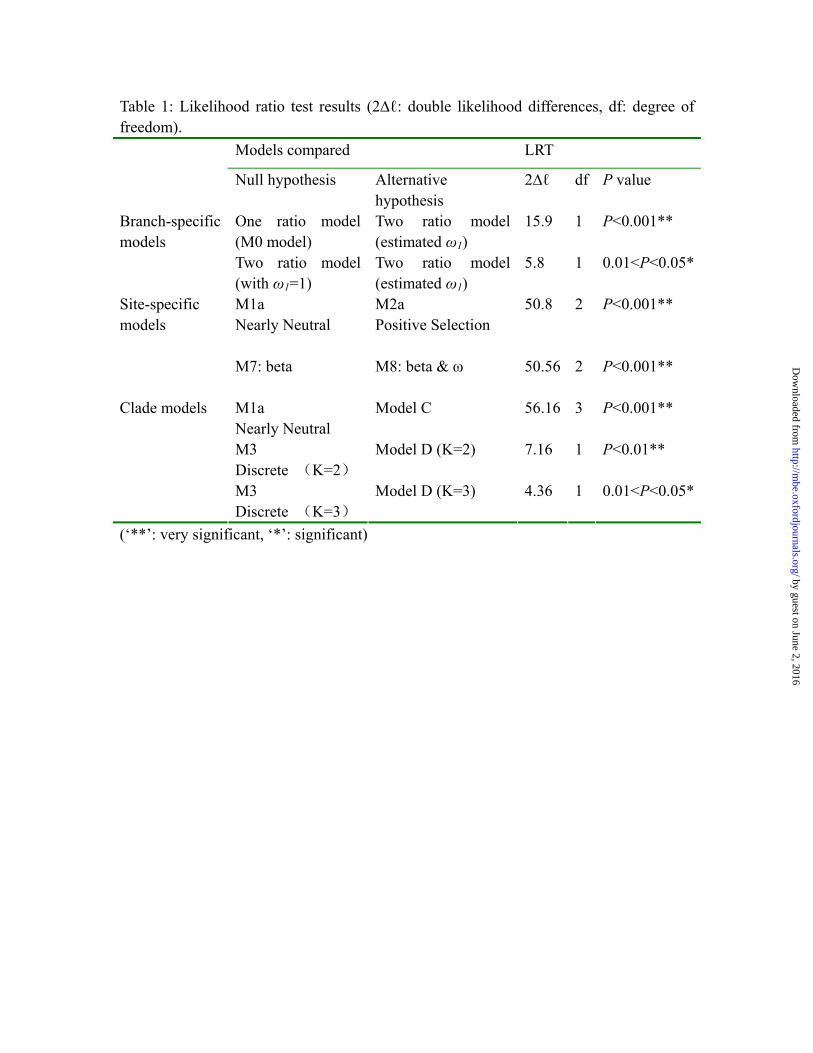
Table 1: Likelihood ratio test results (2∆ℓ: double likelihood differences, df: degree of freedom).
(‘**’: very significant, ‘*’: significant)
Models compared LRT
Null hypothesis Alternative hypothesis
2∆ℓ df P value
One ratio model (M0 model)
Two ratio model (estimated ω1)
15.9 1 P<0.001** Branch-specific models
Two ratio model (with ω1=1)
Two ratio model (estimated ω1)
5.8 1 0.01<P<0.05*
M1a Nearly Neutral
M2a Positive Selection
50.8 2 P<0.001** Site-specific models
M7: beta
M8: beta & ω
50.56 2 P<0.001**
M1a Nearly Neutral
Model C 56.16 3 P<0.001**
M3 Discrete (K=2)
Model D (K=2) 7.16 1 P<0.01**
Clade models
M3 Discrete (K=3)
Model D (K=3) 4.36 1 0.01<P<0.05*
by guest on June 2, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from




















