
Accelerated Diversification of Nonhuman Primate Malarias in Southeast Asia: Adaptive Radiation or...
38
Article Accelerated Diversification of Nonhuman Primate Malarias in Southeast Asia: Adaptive Radiation or Geographic Speciation? Michael P. Muehlenbein,* , y,1 M. Andre ına Pacheco, y,2 Jesse E. Taylor, 2 Sean P. Prall, 1 Laurentius Ambu, 3 Senthilvel Nathan, 3 Sylvia Alsisto, 3 Diana Ramirez, 3 and Ananias A. Escalante* ,2,4 1 Department of Anthropology, Indiana University, Bloomington 2 Center for Evolutionary Medicine and Informatics, The Biodesign Institute, Arizona State University, Tempe 3 Sabah Wildlife Department, Kota Kinabalu, Sabah, Malaysia 4 School of Life Sciences, Arizona State University, Tempe y These authors contributed equally to this work. *Corresponding author: E-mail: [email protected]; [email protected]. Associate editor: Claudia Russo Abstract Although parasitic organisms are found worldwide, the relative importance of host specificity and geographic isolation for parasite speciation has been explored in only a few systems. Here, we study Plasmodium parasites known to infect Asian nonhuman primates, a monophyletic group that includes the lineage leading to the human parasite Plasmodium vivax and several species used as laboratory models in malaria research. We analyze the available data together with new samples from three sympatric primate species from Borneo: The Bornean orangutan and the long-tailed and the pig- tailed macaques. We find several species of malaria parasites, including three putatively new species in this biodiversity hotspot. Among those newly discovered lineages, we report two sympatric parasites in orangutans. We find no differences in the sets of malaria species infecting each macaque species indicating that these species show no host specificity. Finally, phylogenetic analysis of these data suggests that the malaria parasites infecting Southeast Asian macaques and their relatives are speciating three to four times more rapidly than those with other mammalian hosts such as lemurs and African apes. We estimate that these events took place in approximately a 3–4-Ma period. Based on the genetic and phenotypic diversity of the macaque malarias, we hypothesize that the diversification of this group of parasites has been facilitated by the diversity, geographic distributions, and demographic histories of their primate hosts. Key words: host range, macaques, malaria, orangutan, parasite speciation, phylogeny Plasmodium, population structure. Introduction Parasitism is one of the most common multispecies interac- tions observed in nature (Poulin 2005). Most research pro- grams in evolutionary parasitology focus on long-term coevolutionary processes where the adaptation of parasites to their hosts is assumed to have an essential role in parasite speciation (Page 2003; Poulin et al. 2011). A complementary approach considers that, in addition to adaptation to a host or hosts, geographic factors come into play when explaining the diversity of parasite species (Huyse et al. 2005; Brooks et al. 2006; Hoberg et al. 2008; Ricklefs 2010). This second perspec- tive is particularly valuable wherever there are not obvious phylogenetic concordances between host species and their parasites. Plasmodium (phylum Apicomplexa, family Plasmodiidae) is a group of vector-borne parasitic protozoa that are better known by the handful of species that cause malaria in humans. The global health importance of human malarias, however, overshadows the extraordinary diversity in this genus represented by more than 200 described species para- sitizing many vertebrate hosts including rodents and nonhuman primates (Garnham 1966; Coatney et al. 1971; Valki unas 2005; Telford 2009). Of particular interest are those Plasmodium found in nonhuman primates and their relationships to the extant human malarias (Ayala et al. 1998; Escalante et al. 1998; Singh et al. 2004; Escalante et al. 2005; Krief et al. 2010; Liu et al. 2010; Prugnolle et al. 2013; Pacheco et al. 2013). Less attention, however, has been given to the evolutionary processes leading to the staggering diversity ob- served in primate malarial parasites (Hayakawa et al. 2008; Pacheco et al. 2011; Pacheco et al. 2012b). Based on a rate of speciation that was considered rapid for Plasmodium, it was proposed that such a “big-bang” pattern of parasite diversity was evidence for an adaptive radiation of the malarias infecting mammalian hosts facilitated by multi- ple host switches (Hayakawa et al. 2008). Although the hy- pothesis of adaptive radiation is appealing for parasites, such processes are not easy to study in any group of organisms, parasitic, or free-living. Specifically, showing rapid diversifica- tion as result of putative adaptations to a newly available ecological opportunity is complex, especially in a diverse and geographically widespread group of organisms (Hunter ß The Author 2014. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution. This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited. For commercial re-use, please contact [email protected] Open Access 422 Mol. Biol. Evol. 32(2):422–439 doi:10.1093/molbev/msu310 Advance Access publication November 10, 2014 by guest on January 20, 2015 http://mbe.oxfordjournals.org/ Downloaded from
Transcript of Accelerated Diversification of Nonhuman Primate Malarias in Southeast Asia: Adaptive Radiation or...

Article
Accelerated Diversification of Nonhuman Primate Malarias inSoutheast Asia Adaptive Radiation or Geographic SpeciationMichael P Muehlenbeiny1 M Andreına Pachecoy2 Jesse E Taylor2 Sean P Prall1 Laurentius Ambu3
Senthilvel Nathan3 Sylvia Alsisto3 Diana Ramirez3 and Ananias A Escalante24
1Department of Anthropology Indiana University Bloomington2Center for Evolutionary Medicine and Informatics The Biodesign Institute Arizona State University Tempe3Sabah Wildlife Department Kota Kinabalu Sabah Malaysia4School of Life Sciences Arizona State University TempeyThese authors contributed equally to this work
Corresponding author E-mail ananiasescalanteasuedu mpm1indianaedu
Associate editor Claudia Russo
Abstract
Although parasitic organisms are found worldwide the relative importance of host specificity and geographic isolationfor parasite speciation has been explored in only a few systems Here we study Plasmodium parasites known to infectAsian nonhuman primates a monophyletic group that includes the lineage leading to the human parasite Plasmodiumvivax and several species used as laboratory models in malaria research We analyze the available data together with newsamples from three sympatric primate species from Borneo The Bornean orangutan and the long-tailed and the pig-tailed macaques We find several species of malaria parasites including three putatively new species in this biodiversityhotspot Among those newly discovered lineages we report two sympatric parasites in orangutans We find no differencesin the sets of malaria species infecting each macaque species indicating that these species show no host specificity Finallyphylogenetic analysis of these data suggests that the malaria parasites infecting Southeast Asian macaques and theirrelatives are speciating three to four times more rapidly than those with other mammalian hosts such as lemurs andAfrican apes We estimate that these events took place in approximately a 3ndash4-Ma period Based on the genetic andphenotypic diversity of the macaque malarias we hypothesize that the diversification of this group of parasites has beenfacilitated by the diversity geographic distributions and demographic histories of their primate hosts
Key words host range macaques malaria orangutan parasite speciation phylogeny Plasmodium population structure
IntroductionParasitism is one of the most common multispecies interac-tions observed in nature (Poulin 2005) Most research pro-grams in evolutionary parasitology focus on long-termcoevolutionary processes where the adaptation of parasitesto their hosts is assumed to have an essential role in parasitespeciation (Page 2003 Poulin et al 2011) A complementaryapproach considers that in addition to adaptation to a hostor hosts geographic factors come into play when explainingthe diversity of parasite species (Huyse et al 2005 Brooks et al2006 Hoberg et al 2008 Ricklefs 2010) This second perspec-tive is particularly valuable wherever there are not obviousphylogenetic concordances between host species and theirparasites
Plasmodium (phylum Apicomplexa family Plasmodiidae)is a group of vector-borne parasitic protozoa that are betterknown by the handful of species that cause malaria inhumans The global health importance of human malariashowever overshadows the extraordinary diversity in thisgenus represented by more than 200 described species para-sitizing many vertebrate hosts including rodents and
nonhuman primates (Garnham 1966 Coatney et al 1971Valkiunas 2005 Telford 2009) Of particular interest arethose Plasmodium found in nonhuman primates and theirrelationships to the extant human malarias (Ayala et al 1998Escalante et al 1998 Singh et al 2004 Escalante et al 2005Krief et al 2010 Liu et al 2010 Prugnolle et al 2013 Pachecoet al 2013) Less attention however has been given to theevolutionary processes leading to the staggering diversity ob-served in primate malarial parasites (Hayakawa et al 2008Pacheco et al 2011 Pacheco et al 2012b)
Based on a rate of speciation that was considered rapid forPlasmodium it was proposed that such a ldquobig-bangrdquo patternof parasite diversity was evidence for an adaptive radiation ofthe malarias infecting mammalian hosts facilitated by multi-ple host switches (Hayakawa et al 2008) Although the hy-pothesis of adaptive radiation is appealing for parasites suchprocesses are not easy to study in any group of organismsparasitic or free-living Specifically showing rapid diversifica-tion as result of putative adaptations to a newly availableecological opportunity is complex especially in a diverseand geographically widespread group of organisms (Hunter
The Author 2014 Published by Oxford University Press on behalf of the Society for Molecular Biology and EvolutionThis is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License(httpcreativecommonsorglicensesby-nc40) which permits non-commercial re-use distribution and reproduction in anymedium provided the original work is properly cited For commercial re-use please contact journalspermissionsoupcom Open Access422 Mol Biol Evol 32(2)422ndash439 doi101093molbevmsu310 Advance Access publication November 10 2014
by guest on January 20 2015httpm
beoxfordjournalsorgD
ownloaded from
1998 Glor 2010 Aguilee et al 2013) Furthermore apparentpatterns of accelerated evolution could be consistent withmany other processes if they are observed at broad timeand geographic scales (Glor 2010)
One group of malarial parasites that is especially well suitedfor exploring processes such as adaptive radiation andorgeographic speciation is composed of the parasites thatinfect nonhuman primates in South East Asia These speciesare part of a monophyletic group that exhibits extraordinaryphenotypic diversity and its radiation is connected with theorigin of the human parasite Plasmodium vivax (Escalanteet al 1998 2005 Pacheco et al 2011 Prugnolle et al 2013)This group includes species that are zoonotic (Singh et al2004 Ta et al 2014) and others that are considered modelorganisms in malaria research (Coatney et al 1971 Cox-Singhand Singh 2008) In addition this clade is found in a definedgeographic region that shares a common geologic historywith many parasite species exhibiting overlapping hostranges (the host species utilized by a parasite species) andspatial distributions (Coatney et al 1971 Fooden 1994)
Here we report new data on the diversity of Plasmodiumspecies infecting macaques and orangutans at two study sitesin Borneo and analyze them together with previously re-ported data (Hayakawa et al 2008 Pacheco et al 2011Pacheco et al 2012b) We find several lineages that suggestnew parasite species in this biodiversity hotspot Amongthose we report divergent sympatric lineages in orangutansas well as evidence of parasite population genetic structuresdetermined by geographic isolation We also present evidencesuggesting that the diversification rate of the Southeast Asianparasites is accelerated relative to other Plasmodium clades(eg the lemur parasites or the Laverania clade that infectsAfrican apes and humans)
Thus taking into consideration the apparent acceleratedrate of evolution in this monophyletic group and that manyspecies share multiple hosts we hypothesize that Plasmodiumspecies diversity in Southeast Asia was driven at least in partby the history of the host species utilized by these parasites
However determining whether such rapid diversification wasadaptive in nature will require additional evidence beyond theobservation of accelerated evolution
ResultsIn this study we characterized the diversity of nonhumanprimate malarias found in blood sampled from one popula-tion of Pongo pygmaeus morio and two populations ofMacaca nemestrina and Macaca fascicularis from SabahMalaysian Borneo
Plasmodium Species Diversity and PhylogeneticRelationship
Table 1 describes the diversity of nonhuman primate malariasfound in one population of orangutans (Po p morio) and twopopulations from different macaque species (M nemestrinaand M fascicularis) present in Sabah Malaysian Borneo Bloodsmears were collected and some of the samples were positiveby microscopy however the quality of the smears did notallow us to identify the parasites beyond the genus level Thuswe relied on molecular diagnostics for species identification
All blood samples from 15 pig-tailed macaques (M nemes-trina) and 26 long-tailed macaques (M fascicularis) were pos-itive for malarial parasites by polymerase chain reaction (PCR)using cytochrome b (cytb) and some of them had mixedinfections We were able to generate 34 parasite mitochon-drial genomes (mtDNA) for different Plasmodium parasites16 from M nemestrina and 18 from M fascicularis Howeverwe were unable to obtain complete mtDNA sequences forseveral positive M fascicularis samples probably because oflow parasitemia Plasmodium inui was the most commonparasite in these two macaque species (915 for M nemestrinaand 826 for M fascicularis table 1) Regardless of the rela-tively small sample size we found the same six Plasmodiumspecies (P knowlesi P coatneyi Plasmodium sp P fieldi Pinui and P cynomolgi) in both species of Macaca (table 1)
Table 1 Diversity of Plasmodium Parasites in Primates from Sabah Borneo (complete mtDNA genomes)
Host Species Parasite Species No Positive Samples
Macaca nemestrina (Sundaland pig-tailed macaque) Plasmodium knowlesi 215P coatneyi 115Plasmodium sp 115P fieldi 315a
P inui 915a
P cynomolgi 115
Macaca fascicularis (long-tailed macaque) P knowlesi 426a
P coatneyi 126Plasmodium sp 126P fieldi 126P inui 826a
P cynomolgi 326a
Hepatocystis sp 226a
Pongo pygmaeus (Bornean orangutan) Plasmodium sp (clade A)b 738a
Plasmodium sp (clade B)b 1338a
Plasmodium sp (clade C)b 1038a
aMixed infectionbFor clade information see figure 1
423
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Figure 1 shows a Bayesian phylogenetic analysis of a com-bined set of nearly complete mitochondrial genome se-quences including those generated in this and severalprevious studies (supplementary table S1 Supplementary
Material online) We used the African parasite P gonderiand one found in drills in order to root the diversity of speciesobserved in this study (the six Plasmodium species listed intable 1) together with other parasites already reported in
FIG 1 Phylogenetic tree of Plasmodium species based on complete mitochondrial genomes Bayesian and ML methods yielded identical topologies andso only the Bayesian tree is shown Both phylogenetic methods used a general time reversible model with gamma-distributed substitution rates and aproportion of invariant sites (GTR +G+ I) The values above branches are posterior probabilities together with bootstrap values (in bold) as apercentage obtained for the ML tree (see Materials and Methods) Macaque parasites from Borneo are labeled in blue and Plasmodium speciesfrom orangutans are labeled in red Numbers of individuals with each parasite are shown Samples with indicate mixed infection The outgroup isindicated by the gray branches
424
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
nonhuman primates from southeast Asia (Escalante et al2005 Pacheco et al 2013 Pacheco et al 2012b) Overallthis phylogeny is similar to those obtained in previous studies(Pacheco et al 2011 2013 Pacheco et al 2012b) but includesseveral lineages that may correspond to new Plasmodiumspecies Of particular interest is a lineage that was presentin both macaque species which shares a recent commonancestor with P coatneyi This Plasmodium species is diver-gent from the other species in that clade and does not appearto belong to either P knowlesi or P coatneyi (table 2)Specifically the divergence between Plasmodium sp and Pcoatneyi (0023 0002 table 2) was comparable to that ob-served between other pairs of Plasmodium species includingthe human parasite P vivax and the macaque parasite Pcynomolgi (0012 0002 table 2) and the chimpanzee para-site P reichenowi and the human parasite P falciparum(0025 0002 table 2) This result may indicate that bothspecies of macaques harbor a new Plasmodium species Inaddition we found Hepatocystis sp in two individuals of Mfascicularis but not in M nemestrina (table 1 sequences notreported)
Of the 38 orangutans sampled for this study 23 were pos-itive (605) by PCR and 10 harbored mixed infections(table 1) Three samples had low parasitemia and the PCRdid not yield bands that would allow us to clone themUnfortunately although positive blood smears were available
these were not of sufficient quality to determine whetherthese parasites coincide with any of the Plasmodium speciespreviously described in orangutans by Garnham (1966) andPeters et al (1976) We generated 30 parasite mitochondrialgenomes from the remaining 20 samples these haplotypesbelonged to three different lineages (AndashC fig 1) The phylo-genetic analysis showed that two of these lineages (A and B)form a monophyletic group and share a common ancestorwith P inui a species of macaque parasite with broad geo-graphic distribution (Coatney et al 1971 Fooden 1994)
Lineage C is at the base of a monophyletic group thatincludes P hylobati from gibbons P inui from macaques aswell as lineages A and B from orangutans This parasite phy-logeny is not congruent with any primate phylogenetic hy-potheses estimated using nuclear andor mitochondrial genes(eg Perelman et al 2011) such discrepancy indicates thathost switches likely took place during the evolution of thisPlasmodium clade Specifically P inui seems to derive fromparasites infecting Asian apes (Pacheco et al 2012b)The average distance between the haplotypes belonging tolineages A and B in orangutan malarias (0009 0001 for thecomplete mtDNA table 2) is similar to the divergence be-tween P vivax and P cynomolgi (0012 0002 table 2) Onthe other hand the average distances between clades A and C(0023 0002) and between clades B and C (0024 0002)are somewhat higher than those between clades A and B
Table 2 Genetic Divergences (substitutions per site) among Different Plasmodium Species Using the Kimura 2-Parameter Model As Implementedin MEGA v522 (Tamura et al 2011)
Species Genetic distance (d SE)
n COX1 COX3 CYTB COX1 + CYTB Complete mtDNA
Plasmodium sp (orangutan-clade A) 12 0005 0001 0006 0002 0003 0001 0004 0001 0004 00003
Plasmodium sp (orangutan-clade B) 13 0003 0001 0004 0001 0004 0001 0003 0001 0003 00003
Plasmodium sp (orangutan-clade C) 10 0002 0001 0003 0001 0004 0001 0003 0001 0003 00003
Orangutan clade A orangutan clade B 12 vs 13 0011 0002 0019 0005 0008 0002 0010 0001 0009 00010
Orangutan clade A orangutan clade C 12 vs 10 0034 0004 0043 0007 0028 0004 0031 0003 0023 00017
Orangutan clade B orangutan clade C 13 vs 10 0034 0004 0049 0008 0028 0005 0031 0003 0024 00018
Orangutan clade A + orangutan clade B 25 0008 0001 0012 0003 0006 0001 0007 0001 0006 00006
P cynomolgi (all strains) 14 0005 0001 0006 0001 0005 0001 0004 0001 0004 00006
P cynomolgi (clade A-Berok) 4 0007 0002 0007 0002 0006 0002 0006 0001 0006 00008
P cynomolgi (clade B-B) 10 0001 0001 0001 0001 0001 0001 0001 0000 0001 00002
P cynomolgi (clade A vs clade B) 4 vs10 0010 0002 0010 0003 0009 0002 0009 0002 0009 00011
P vivax 109 0001 0000 0001 0001 0 0001 0000 0001 00002
P vivaxndashP cynomolgi 109 vs 14 0014 0003 0017 0004 0013 0003 0014 0002 0012 00015
P inui (all) 31 0013 0002 0023 0004 0015 0002 0014 0001 0013 00010
P inui (Macaca nemestrina)-(M fascicularis) 9 vs 8 0004 0001 0005 0001 0003 0001 0003 0000 0004 00000
P inui (all)ndashP sp from orang clade A 31 vs 12 0023 0003 0045 0007 0018 0002 0024 0003 0023 00010
P inui (all)ndashP sp from orang clade B 31 vs 13 0025 0004 0054 0008 0027 0004 0026 0002 0025 00020
P inui (all)ndashP sp from orang clade A + B 31 vs 25 0024 0003 0050 0007 0026 0004 0025 0002 0024 00020
P knowlesi (all) 65 0001 0000 0002 0001 0001 0000 0001 0000 0001 00000
P knowlesi (NCBI)ndashSepilok 60 vs 5 0002 0000 0002 0001 0001 0000 0001 0000 0001 00000
P knowlesindashP coatneyi 65 vs 2 0043 0005 0034 0007 0025 0005 0035 0004 0032 00020
Plasmodium sp (MnendashMfa) 1 vs 1 0 0 0003 0001 0001 0001 0003 00010
P knowlesindashPlasmodium sp 65 vs 2 0037 0005 0037 0007 0020 0004 0030 0003 0029 00020
P coatneyindashPlasmodium sp 2 vs 2 0032 0005 0037 0007 0019 0004 0026 0003 0023 00020
P falciparum 101 0 0000 0001 0001 0001 0001 0000 0000 00001
P reichenowindashP falciparum 2 vs 101 0032 0005 0055 0009 0025 0005 0029 0003 0025 00018
425
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
between P vivax and P cynomolgi and between P reichenowiand P falciparum These results suggest that the haplotypesidentified in figure 1 as AndashC may belong to three differentspecies Clade A includes haplotypes described previously in adifferent population of orangutans (Pacheco et al 2012b)
In the case of P cynomolgi we found two closely relatedlineages in both species of macaques These lineages belong toone of the two clades (fig 1) that have been described for Pcynomolgi using antigen-encoding genes (Pacheco et al 20072010 Pacheco et al 2012a Pacheco et al 2012b) which con-tains the Berok PT2 and Gombak strains Indeed the averagegenetic distance among these four P cynomolgi strains(0006 00008) is relatively small if we consider its broadgeographic range (see table 2) It is also worth noting thatthe divergence of these two clades of P cynomolgi is compa-rable to that observed between P vivax and P cynomolgi(table 2) an observation that has been corroborated by ge-nomic data (Tachibana et al 2012)
Malaria Antigens Apical Membrane Antigen-1 andMerozoite Surface Protein 1 42 kDa
In addition to mtDNA genomes phylogenetic analyses werealso performed separately on data from several macaque andorangutan parasites using nuclear genes encoding two majorantigens Apical membrane antigen-1 (AMA-1 fig 2 and sup-plementary table S1 Supplementary Material online) andmerozoite surface protein 1 42 kDa (MSP-142 fig 3 and sup-plementary table S1 Supplementary Material online) Despitethe smaller number of species for which data is available thephylogenies estimated from these two loci are largely com-patible with previous studies using genes encoding merozoiteantigens (MSP-1 Pacheco et al 2007 Pacheco et al 2012bRAP-1 Pacheco et al 2010 MSP-8MSP-10 Pacheco et al2012a) In particular in both phylogenies we observe thatseveral of the orangutan malaria lineages form a monophy-letic group sharing a common ancestor with the macaqueparasite P inui One difference between the phylogenies gen-erated using either mtDNA or these two antigen-encodinggenes is in the relative position of P hylobati This is notsurprising considering that the position of P hylobati hasmodest support in the mtDNA (085 posterior probabilityand 63 bootstrap support see fig 1) Although in the mito-chondrial tree this lineage is located within the clade thatincludes the three orangutan lineages and P inui in theama-1 and msp-142 phylogenies it is located at the base ofthe clade containing P inui and the three orangutan parasiteclades (A-C fig 1) This discrepancy could have three differentexplanations First because the mitochondrial and nuclearloci are unlinked there could be random differences in thegenealogies of these loci Second this could be due to diver-sifying- or frequency-dependent selection acting on the anti-gen loci Finally this could be an artifact of our inability togenerate antigen sequences for all of the taxa included in themitochondrial tree Indeed because we were unable to gen-erate any sequences of ama-1 that can be unambiguouslyassigned to mtDNA clade C the position of P hylobati relativeto the orangutan clades is unresolved (fig 2)
In the case of ama-1 we were able to obtain sequences forP knowlesi Plasmodium sp P cynomolgi P inui from Mnemestrina and Plasmodium sp from orangutan The geneticpolymorphism of ama-1 per species is reported in the sup-plementary table S2 Supplementary Material online Theama-1 allele found in this study for P cynomolgi was mostclosely related to that contained in the Berok strain this resultis consistent with the mtDNA phylogeny Again the unde-scribed Plasmodium sp isolated from macaques forms a cladealong with P coatneyi and the average genetic distance esti-mated between the ama-1 allele for this parasite and P coat-neyi is very high (0076 0073) supporting the hypothesisthat the former organism is a new species On the other handwe only were able to generate msp-142 sequences for P inuiand the Plasmodium species detected in orangutans The ge-netic polymorphism per species for msp-142 is reported insupplementary table S3 Supplementary Material onlineOverall using these additional msp-142 sequences we recov-ered the same topology that was previously reported for thisantigen (Pacheco et al 2007 Pacheco et al 2012b)
Phylogenetic-based analyses provide some insight into therole of selection at these two loci (Pond et al 2011) In thecase of ama-1 several lineages contain codons that appear tobe under positive selection ( 41 Plt 005 corrected formultiple testing using HolmndashBonferroni methodSupplementary table S4 and fig S1 Supplementary Materialonline) Furthermore many of the nodes that may corre-spond to host switches also have significant uncorrected se-quential likelihood ratio tests (LRTs) A similar patternemerges from msp-142 where this method identifies four lin-eages that appear to be under episodic diversifying selection(supplementary table S4 and fig S1 Supplementary Materialonline)
Haplotype Networks
A median joining network of 41 Plasmodium mtDNA haplo-types isolated from orangutans from Borneo is depicted infigure 4 This network includes both the 30 PlasmodiummtDNA haplotypes from Sabah and the 11 mtDNA haplo-types from the Kalimantan orangutans As expected thethree clades identified in the mtDNA tree were clearly sepa-rated in the haplotype network Furthermore clade A itself(colored in green) contains two genetically differentiated pop-ulations (light green Sabah dark green Kalimantan) andindeed the fixation index (FST) between these two popula-tions is high (056) In another context the Venn diagramdepicted in figure 4 shows the number of orangutans withmixed infections of Plasmodium At least 10 of the 20 orang-utan samples harbored mixed infections with parasites fromtwo distinct clades but none was found to be simultaneouslyinfected by all three species
Figure 5 shows a median joining network for the 16 P inuimtDNA haplotypes from both species of Macaca (eight hap-lotypes from M nemestrina and eight from M fascicularis)No evidence of population structure between hosts wasfound in this analysis in fact the fixation index (FST) is effec-tively 0 (0024) In addition we also identified a divergent P
426
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 2 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding AMA-1 Bayesian and ML methods yielded identical topologiesand so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) The values abovebranches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials and Methods)Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shown Cladenumbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section Samples with indicate mixed infectionThe outgroup is indicated by the gray branch
427
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 3 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding MSP-142 kDa Bayesian and ML methods yielded identicaltopologies and so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) Thevalues above branches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials andMethods) Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shownClade numbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section The outgroup is indicated by thegray branch
428
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
inui mtDNA haplotype that was carried by a single individualof M nemestrina (see also fig 1)
The Ages and Branching Rates of the Asian PrimateMalarias
The mitochondrial genome phylogeny was used to estimatethe ages of the major clades of South Asian primate malariasunder three different calibration scenarios (fig 6 and supple-mentary fig S2 Supplementary Material online) These timesalong with the 95 credibility intervals (Crls) were calculatedusing BEAST v175 (Drummond et al 2012) and are shown intable 3 (See supplementary fig S2 Supplementary Materialonline for the node numbers) As expected there is extensiveoverlap among the Crls obtained under the three differentscenarios used for calibration When the phylogeny is cali-brated by assuming that the Asian and African primate par-asites split in the same time frame that their hosts becamegeographically isolated (6ndash142 Ma) and that lemur parasitesoriginated as a monophyletic group at least 20 Ma (seeMaterials and Methods) the Asian primate malarias are esti-mated to have originated between 538 and 852 Ma The splitbetween P inui and the orangutan Plasmodium lineages in
clades A and B was around 292 and 480 Ma which is inagreement with an earlier study of clade A (246ndash438 MaPacheco et al 2012b) Furthermore clades A and B divergedapproximately 10ndash23 Ma (table 3) this is clearly older thantime estimates for the split of the Bornean and Sumatranorangutans using nuclear single nucleotide polymorphisms(SNPs) but earlier than estimates for the same two speciesusing the mitochondrial genome (Ma et al 2013) On theother hand the common ancestor of the orangutanPlasmodium lineages in clade C appears to have existed be-tween 371 and 598 Ma well after the time of the split be-tween orangutan and gibbons or the most recent commonancestor of Asian apes with macaques
Previous phylogenetic studies have indicated that P coat-neyi and P knowlesi are sister taxa (eg Escalante et al 2005)and indeed these two species are usually referred as ldquocloselyrelatedrdquo in the malaria literature In this study the age of themost recent common ancestor of P knowlesi and P coatneyiwas estimated to be 554 Ma (CrI 417ndash710 table 3) whichlies within the time interval proposed for the radiation ofmacaques and other nonhuman primates in the Malaysianarchipelago (Delson 1980 Perelman et al 2011 Springer et al2012)
FIG 4 Venn diagram of mixed infections and median joining network of Plasmodium sp mtDNA haplotypes from orangutans from BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to differentclades green = clade A red = clade B and blue = clade C
429
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

1998 Glor 2010 Aguilee et al 2013) Furthermore apparentpatterns of accelerated evolution could be consistent withmany other processes if they are observed at broad timeand geographic scales (Glor 2010)
One group of malarial parasites that is especially well suitedfor exploring processes such as adaptive radiation andorgeographic speciation is composed of the parasites thatinfect nonhuman primates in South East Asia These speciesare part of a monophyletic group that exhibits extraordinaryphenotypic diversity and its radiation is connected with theorigin of the human parasite Plasmodium vivax (Escalanteet al 1998 2005 Pacheco et al 2011 Prugnolle et al 2013)This group includes species that are zoonotic (Singh et al2004 Ta et al 2014) and others that are considered modelorganisms in malaria research (Coatney et al 1971 Cox-Singhand Singh 2008) In addition this clade is found in a definedgeographic region that shares a common geologic historywith many parasite species exhibiting overlapping hostranges (the host species utilized by a parasite species) andspatial distributions (Coatney et al 1971 Fooden 1994)
Here we report new data on the diversity of Plasmodiumspecies infecting macaques and orangutans at two study sitesin Borneo and analyze them together with previously re-ported data (Hayakawa et al 2008 Pacheco et al 2011Pacheco et al 2012b) We find several lineages that suggestnew parasite species in this biodiversity hotspot Amongthose we report divergent sympatric lineages in orangutansas well as evidence of parasite population genetic structuresdetermined by geographic isolation We also present evidencesuggesting that the diversification rate of the Southeast Asianparasites is accelerated relative to other Plasmodium clades(eg the lemur parasites or the Laverania clade that infectsAfrican apes and humans)
Thus taking into consideration the apparent acceleratedrate of evolution in this monophyletic group and that manyspecies share multiple hosts we hypothesize that Plasmodiumspecies diversity in Southeast Asia was driven at least in partby the history of the host species utilized by these parasites
However determining whether such rapid diversification wasadaptive in nature will require additional evidence beyond theobservation of accelerated evolution
ResultsIn this study we characterized the diversity of nonhumanprimate malarias found in blood sampled from one popula-tion of Pongo pygmaeus morio and two populations ofMacaca nemestrina and Macaca fascicularis from SabahMalaysian Borneo
Plasmodium Species Diversity and PhylogeneticRelationship
Table 1 describes the diversity of nonhuman primate malariasfound in one population of orangutans (Po p morio) and twopopulations from different macaque species (M nemestrinaand M fascicularis) present in Sabah Malaysian Borneo Bloodsmears were collected and some of the samples were positiveby microscopy however the quality of the smears did notallow us to identify the parasites beyond the genus level Thuswe relied on molecular diagnostics for species identification
All blood samples from 15 pig-tailed macaques (M nemes-trina) and 26 long-tailed macaques (M fascicularis) were pos-itive for malarial parasites by polymerase chain reaction (PCR)using cytochrome b (cytb) and some of them had mixedinfections We were able to generate 34 parasite mitochon-drial genomes (mtDNA) for different Plasmodium parasites16 from M nemestrina and 18 from M fascicularis Howeverwe were unable to obtain complete mtDNA sequences forseveral positive M fascicularis samples probably because oflow parasitemia Plasmodium inui was the most commonparasite in these two macaque species (915 for M nemestrinaand 826 for M fascicularis table 1) Regardless of the rela-tively small sample size we found the same six Plasmodiumspecies (P knowlesi P coatneyi Plasmodium sp P fieldi Pinui and P cynomolgi) in both species of Macaca (table 1)
Table 1 Diversity of Plasmodium Parasites in Primates from Sabah Borneo (complete mtDNA genomes)
Host Species Parasite Species No Positive Samples
Macaca nemestrina (Sundaland pig-tailed macaque) Plasmodium knowlesi 215P coatneyi 115Plasmodium sp 115P fieldi 315a
P inui 915a
P cynomolgi 115
Macaca fascicularis (long-tailed macaque) P knowlesi 426a
P coatneyi 126Plasmodium sp 126P fieldi 126P inui 826a
P cynomolgi 326a
Hepatocystis sp 226a
Pongo pygmaeus (Bornean orangutan) Plasmodium sp (clade A)b 738a
Plasmodium sp (clade B)b 1338a
Plasmodium sp (clade C)b 1038a
aMixed infectionbFor clade information see figure 1
423
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Figure 1 shows a Bayesian phylogenetic analysis of a com-bined set of nearly complete mitochondrial genome se-quences including those generated in this and severalprevious studies (supplementary table S1 Supplementary
Material online) We used the African parasite P gonderiand one found in drills in order to root the diversity of speciesobserved in this study (the six Plasmodium species listed intable 1) together with other parasites already reported in
FIG 1 Phylogenetic tree of Plasmodium species based on complete mitochondrial genomes Bayesian and ML methods yielded identical topologies andso only the Bayesian tree is shown Both phylogenetic methods used a general time reversible model with gamma-distributed substitution rates and aproportion of invariant sites (GTR +G+ I) The values above branches are posterior probabilities together with bootstrap values (in bold) as apercentage obtained for the ML tree (see Materials and Methods) Macaque parasites from Borneo are labeled in blue and Plasmodium speciesfrom orangutans are labeled in red Numbers of individuals with each parasite are shown Samples with indicate mixed infection The outgroup isindicated by the gray branches
424
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
nonhuman primates from southeast Asia (Escalante et al2005 Pacheco et al 2013 Pacheco et al 2012b) Overallthis phylogeny is similar to those obtained in previous studies(Pacheco et al 2011 2013 Pacheco et al 2012b) but includesseveral lineages that may correspond to new Plasmodiumspecies Of particular interest is a lineage that was presentin both macaque species which shares a recent commonancestor with P coatneyi This Plasmodium species is diver-gent from the other species in that clade and does not appearto belong to either P knowlesi or P coatneyi (table 2)Specifically the divergence between Plasmodium sp and Pcoatneyi (0023 0002 table 2) was comparable to that ob-served between other pairs of Plasmodium species includingthe human parasite P vivax and the macaque parasite Pcynomolgi (0012 0002 table 2) and the chimpanzee para-site P reichenowi and the human parasite P falciparum(0025 0002 table 2) This result may indicate that bothspecies of macaques harbor a new Plasmodium species Inaddition we found Hepatocystis sp in two individuals of Mfascicularis but not in M nemestrina (table 1 sequences notreported)
Of the 38 orangutans sampled for this study 23 were pos-itive (605) by PCR and 10 harbored mixed infections(table 1) Three samples had low parasitemia and the PCRdid not yield bands that would allow us to clone themUnfortunately although positive blood smears were available
these were not of sufficient quality to determine whetherthese parasites coincide with any of the Plasmodium speciespreviously described in orangutans by Garnham (1966) andPeters et al (1976) We generated 30 parasite mitochondrialgenomes from the remaining 20 samples these haplotypesbelonged to three different lineages (AndashC fig 1) The phylo-genetic analysis showed that two of these lineages (A and B)form a monophyletic group and share a common ancestorwith P inui a species of macaque parasite with broad geo-graphic distribution (Coatney et al 1971 Fooden 1994)
Lineage C is at the base of a monophyletic group thatincludes P hylobati from gibbons P inui from macaques aswell as lineages A and B from orangutans This parasite phy-logeny is not congruent with any primate phylogenetic hy-potheses estimated using nuclear andor mitochondrial genes(eg Perelman et al 2011) such discrepancy indicates thathost switches likely took place during the evolution of thisPlasmodium clade Specifically P inui seems to derive fromparasites infecting Asian apes (Pacheco et al 2012b)The average distance between the haplotypes belonging tolineages A and B in orangutan malarias (0009 0001 for thecomplete mtDNA table 2) is similar to the divergence be-tween P vivax and P cynomolgi (0012 0002 table 2) Onthe other hand the average distances between clades A and C(0023 0002) and between clades B and C (0024 0002)are somewhat higher than those between clades A and B
Table 2 Genetic Divergences (substitutions per site) among Different Plasmodium Species Using the Kimura 2-Parameter Model As Implementedin MEGA v522 (Tamura et al 2011)
Species Genetic distance (d SE)
n COX1 COX3 CYTB COX1 + CYTB Complete mtDNA
Plasmodium sp (orangutan-clade A) 12 0005 0001 0006 0002 0003 0001 0004 0001 0004 00003
Plasmodium sp (orangutan-clade B) 13 0003 0001 0004 0001 0004 0001 0003 0001 0003 00003
Plasmodium sp (orangutan-clade C) 10 0002 0001 0003 0001 0004 0001 0003 0001 0003 00003
Orangutan clade A orangutan clade B 12 vs 13 0011 0002 0019 0005 0008 0002 0010 0001 0009 00010
Orangutan clade A orangutan clade C 12 vs 10 0034 0004 0043 0007 0028 0004 0031 0003 0023 00017
Orangutan clade B orangutan clade C 13 vs 10 0034 0004 0049 0008 0028 0005 0031 0003 0024 00018
Orangutan clade A + orangutan clade B 25 0008 0001 0012 0003 0006 0001 0007 0001 0006 00006
P cynomolgi (all strains) 14 0005 0001 0006 0001 0005 0001 0004 0001 0004 00006
P cynomolgi (clade A-Berok) 4 0007 0002 0007 0002 0006 0002 0006 0001 0006 00008
P cynomolgi (clade B-B) 10 0001 0001 0001 0001 0001 0001 0001 0000 0001 00002
P cynomolgi (clade A vs clade B) 4 vs10 0010 0002 0010 0003 0009 0002 0009 0002 0009 00011
P vivax 109 0001 0000 0001 0001 0 0001 0000 0001 00002
P vivaxndashP cynomolgi 109 vs 14 0014 0003 0017 0004 0013 0003 0014 0002 0012 00015
P inui (all) 31 0013 0002 0023 0004 0015 0002 0014 0001 0013 00010
P inui (Macaca nemestrina)-(M fascicularis) 9 vs 8 0004 0001 0005 0001 0003 0001 0003 0000 0004 00000
P inui (all)ndashP sp from orang clade A 31 vs 12 0023 0003 0045 0007 0018 0002 0024 0003 0023 00010
P inui (all)ndashP sp from orang clade B 31 vs 13 0025 0004 0054 0008 0027 0004 0026 0002 0025 00020
P inui (all)ndashP sp from orang clade A + B 31 vs 25 0024 0003 0050 0007 0026 0004 0025 0002 0024 00020
P knowlesi (all) 65 0001 0000 0002 0001 0001 0000 0001 0000 0001 00000
P knowlesi (NCBI)ndashSepilok 60 vs 5 0002 0000 0002 0001 0001 0000 0001 0000 0001 00000
P knowlesindashP coatneyi 65 vs 2 0043 0005 0034 0007 0025 0005 0035 0004 0032 00020
Plasmodium sp (MnendashMfa) 1 vs 1 0 0 0003 0001 0001 0001 0003 00010
P knowlesindashPlasmodium sp 65 vs 2 0037 0005 0037 0007 0020 0004 0030 0003 0029 00020
P coatneyindashPlasmodium sp 2 vs 2 0032 0005 0037 0007 0019 0004 0026 0003 0023 00020
P falciparum 101 0 0000 0001 0001 0001 0001 0000 0000 00001
P reichenowindashP falciparum 2 vs 101 0032 0005 0055 0009 0025 0005 0029 0003 0025 00018
425
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
between P vivax and P cynomolgi and between P reichenowiand P falciparum These results suggest that the haplotypesidentified in figure 1 as AndashC may belong to three differentspecies Clade A includes haplotypes described previously in adifferent population of orangutans (Pacheco et al 2012b)
In the case of P cynomolgi we found two closely relatedlineages in both species of macaques These lineages belong toone of the two clades (fig 1) that have been described for Pcynomolgi using antigen-encoding genes (Pacheco et al 20072010 Pacheco et al 2012a Pacheco et al 2012b) which con-tains the Berok PT2 and Gombak strains Indeed the averagegenetic distance among these four P cynomolgi strains(0006 00008) is relatively small if we consider its broadgeographic range (see table 2) It is also worth noting thatthe divergence of these two clades of P cynomolgi is compa-rable to that observed between P vivax and P cynomolgi(table 2) an observation that has been corroborated by ge-nomic data (Tachibana et al 2012)
Malaria Antigens Apical Membrane Antigen-1 andMerozoite Surface Protein 1 42 kDa
In addition to mtDNA genomes phylogenetic analyses werealso performed separately on data from several macaque andorangutan parasites using nuclear genes encoding two majorantigens Apical membrane antigen-1 (AMA-1 fig 2 and sup-plementary table S1 Supplementary Material online) andmerozoite surface protein 1 42 kDa (MSP-142 fig 3 and sup-plementary table S1 Supplementary Material online) Despitethe smaller number of species for which data is available thephylogenies estimated from these two loci are largely com-patible with previous studies using genes encoding merozoiteantigens (MSP-1 Pacheco et al 2007 Pacheco et al 2012bRAP-1 Pacheco et al 2010 MSP-8MSP-10 Pacheco et al2012a) In particular in both phylogenies we observe thatseveral of the orangutan malaria lineages form a monophy-letic group sharing a common ancestor with the macaqueparasite P inui One difference between the phylogenies gen-erated using either mtDNA or these two antigen-encodinggenes is in the relative position of P hylobati This is notsurprising considering that the position of P hylobati hasmodest support in the mtDNA (085 posterior probabilityand 63 bootstrap support see fig 1) Although in the mito-chondrial tree this lineage is located within the clade thatincludes the three orangutan lineages and P inui in theama-1 and msp-142 phylogenies it is located at the base ofthe clade containing P inui and the three orangutan parasiteclades (A-C fig 1) This discrepancy could have three differentexplanations First because the mitochondrial and nuclearloci are unlinked there could be random differences in thegenealogies of these loci Second this could be due to diver-sifying- or frequency-dependent selection acting on the anti-gen loci Finally this could be an artifact of our inability togenerate antigen sequences for all of the taxa included in themitochondrial tree Indeed because we were unable to gen-erate any sequences of ama-1 that can be unambiguouslyassigned to mtDNA clade C the position of P hylobati relativeto the orangutan clades is unresolved (fig 2)
In the case of ama-1 we were able to obtain sequences forP knowlesi Plasmodium sp P cynomolgi P inui from Mnemestrina and Plasmodium sp from orangutan The geneticpolymorphism of ama-1 per species is reported in the sup-plementary table S2 Supplementary Material online Theama-1 allele found in this study for P cynomolgi was mostclosely related to that contained in the Berok strain this resultis consistent with the mtDNA phylogeny Again the unde-scribed Plasmodium sp isolated from macaques forms a cladealong with P coatneyi and the average genetic distance esti-mated between the ama-1 allele for this parasite and P coat-neyi is very high (0076 0073) supporting the hypothesisthat the former organism is a new species On the other handwe only were able to generate msp-142 sequences for P inuiand the Plasmodium species detected in orangutans The ge-netic polymorphism per species for msp-142 is reported insupplementary table S3 Supplementary Material onlineOverall using these additional msp-142 sequences we recov-ered the same topology that was previously reported for thisantigen (Pacheco et al 2007 Pacheco et al 2012b)
Phylogenetic-based analyses provide some insight into therole of selection at these two loci (Pond et al 2011) In thecase of ama-1 several lineages contain codons that appear tobe under positive selection ( 41 Plt 005 corrected formultiple testing using HolmndashBonferroni methodSupplementary table S4 and fig S1 Supplementary Materialonline) Furthermore many of the nodes that may corre-spond to host switches also have significant uncorrected se-quential likelihood ratio tests (LRTs) A similar patternemerges from msp-142 where this method identifies four lin-eages that appear to be under episodic diversifying selection(supplementary table S4 and fig S1 Supplementary Materialonline)
Haplotype Networks
A median joining network of 41 Plasmodium mtDNA haplo-types isolated from orangutans from Borneo is depicted infigure 4 This network includes both the 30 PlasmodiummtDNA haplotypes from Sabah and the 11 mtDNA haplo-types from the Kalimantan orangutans As expected thethree clades identified in the mtDNA tree were clearly sepa-rated in the haplotype network Furthermore clade A itself(colored in green) contains two genetically differentiated pop-ulations (light green Sabah dark green Kalimantan) andindeed the fixation index (FST) between these two popula-tions is high (056) In another context the Venn diagramdepicted in figure 4 shows the number of orangutans withmixed infections of Plasmodium At least 10 of the 20 orang-utan samples harbored mixed infections with parasites fromtwo distinct clades but none was found to be simultaneouslyinfected by all three species
Figure 5 shows a median joining network for the 16 P inuimtDNA haplotypes from both species of Macaca (eight hap-lotypes from M nemestrina and eight from M fascicularis)No evidence of population structure between hosts wasfound in this analysis in fact the fixation index (FST) is effec-tively 0 (0024) In addition we also identified a divergent P
426
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 2 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding AMA-1 Bayesian and ML methods yielded identical topologiesand so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) The values abovebranches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials and Methods)Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shown Cladenumbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section Samples with indicate mixed infectionThe outgroup is indicated by the gray branch
427
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 3 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding MSP-142 kDa Bayesian and ML methods yielded identicaltopologies and so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) Thevalues above branches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials andMethods) Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shownClade numbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section The outgroup is indicated by thegray branch
428
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
inui mtDNA haplotype that was carried by a single individualof M nemestrina (see also fig 1)
The Ages and Branching Rates of the Asian PrimateMalarias
The mitochondrial genome phylogeny was used to estimatethe ages of the major clades of South Asian primate malariasunder three different calibration scenarios (fig 6 and supple-mentary fig S2 Supplementary Material online) These timesalong with the 95 credibility intervals (Crls) were calculatedusing BEAST v175 (Drummond et al 2012) and are shown intable 3 (See supplementary fig S2 Supplementary Materialonline for the node numbers) As expected there is extensiveoverlap among the Crls obtained under the three differentscenarios used for calibration When the phylogeny is cali-brated by assuming that the Asian and African primate par-asites split in the same time frame that their hosts becamegeographically isolated (6ndash142 Ma) and that lemur parasitesoriginated as a monophyletic group at least 20 Ma (seeMaterials and Methods) the Asian primate malarias are esti-mated to have originated between 538 and 852 Ma The splitbetween P inui and the orangutan Plasmodium lineages in
clades A and B was around 292 and 480 Ma which is inagreement with an earlier study of clade A (246ndash438 MaPacheco et al 2012b) Furthermore clades A and B divergedapproximately 10ndash23 Ma (table 3) this is clearly older thantime estimates for the split of the Bornean and Sumatranorangutans using nuclear single nucleotide polymorphisms(SNPs) but earlier than estimates for the same two speciesusing the mitochondrial genome (Ma et al 2013) On theother hand the common ancestor of the orangutanPlasmodium lineages in clade C appears to have existed be-tween 371 and 598 Ma well after the time of the split be-tween orangutan and gibbons or the most recent commonancestor of Asian apes with macaques
Previous phylogenetic studies have indicated that P coat-neyi and P knowlesi are sister taxa (eg Escalante et al 2005)and indeed these two species are usually referred as ldquocloselyrelatedrdquo in the malaria literature In this study the age of themost recent common ancestor of P knowlesi and P coatneyiwas estimated to be 554 Ma (CrI 417ndash710 table 3) whichlies within the time interval proposed for the radiation ofmacaques and other nonhuman primates in the Malaysianarchipelago (Delson 1980 Perelman et al 2011 Springer et al2012)
FIG 4 Venn diagram of mixed infections and median joining network of Plasmodium sp mtDNA haplotypes from orangutans from BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to differentclades green = clade A red = clade B and blue = clade C
429
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
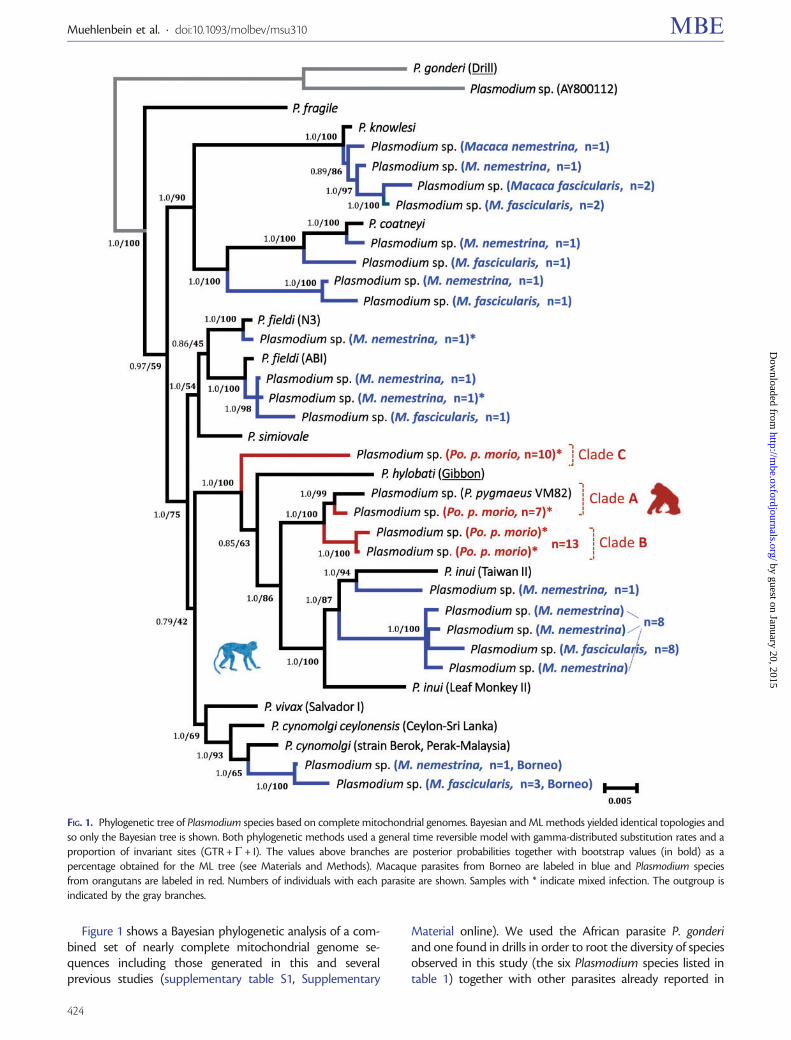
Figure 1 shows a Bayesian phylogenetic analysis of a com-bined set of nearly complete mitochondrial genome se-quences including those generated in this and severalprevious studies (supplementary table S1 Supplementary
Material online) We used the African parasite P gonderiand one found in drills in order to root the diversity of speciesobserved in this study (the six Plasmodium species listed intable 1) together with other parasites already reported in
FIG 1 Phylogenetic tree of Plasmodium species based on complete mitochondrial genomes Bayesian and ML methods yielded identical topologies andso only the Bayesian tree is shown Both phylogenetic methods used a general time reversible model with gamma-distributed substitution rates and aproportion of invariant sites (GTR +G+ I) The values above branches are posterior probabilities together with bootstrap values (in bold) as apercentage obtained for the ML tree (see Materials and Methods) Macaque parasites from Borneo are labeled in blue and Plasmodium speciesfrom orangutans are labeled in red Numbers of individuals with each parasite are shown Samples with indicate mixed infection The outgroup isindicated by the gray branches
424
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
nonhuman primates from southeast Asia (Escalante et al2005 Pacheco et al 2013 Pacheco et al 2012b) Overallthis phylogeny is similar to those obtained in previous studies(Pacheco et al 2011 2013 Pacheco et al 2012b) but includesseveral lineages that may correspond to new Plasmodiumspecies Of particular interest is a lineage that was presentin both macaque species which shares a recent commonancestor with P coatneyi This Plasmodium species is diver-gent from the other species in that clade and does not appearto belong to either P knowlesi or P coatneyi (table 2)Specifically the divergence between Plasmodium sp and Pcoatneyi (0023 0002 table 2) was comparable to that ob-served between other pairs of Plasmodium species includingthe human parasite P vivax and the macaque parasite Pcynomolgi (0012 0002 table 2) and the chimpanzee para-site P reichenowi and the human parasite P falciparum(0025 0002 table 2) This result may indicate that bothspecies of macaques harbor a new Plasmodium species Inaddition we found Hepatocystis sp in two individuals of Mfascicularis but not in M nemestrina (table 1 sequences notreported)
Of the 38 orangutans sampled for this study 23 were pos-itive (605) by PCR and 10 harbored mixed infections(table 1) Three samples had low parasitemia and the PCRdid not yield bands that would allow us to clone themUnfortunately although positive blood smears were available
these were not of sufficient quality to determine whetherthese parasites coincide with any of the Plasmodium speciespreviously described in orangutans by Garnham (1966) andPeters et al (1976) We generated 30 parasite mitochondrialgenomes from the remaining 20 samples these haplotypesbelonged to three different lineages (AndashC fig 1) The phylo-genetic analysis showed that two of these lineages (A and B)form a monophyletic group and share a common ancestorwith P inui a species of macaque parasite with broad geo-graphic distribution (Coatney et al 1971 Fooden 1994)
Lineage C is at the base of a monophyletic group thatincludes P hylobati from gibbons P inui from macaques aswell as lineages A and B from orangutans This parasite phy-logeny is not congruent with any primate phylogenetic hy-potheses estimated using nuclear andor mitochondrial genes(eg Perelman et al 2011) such discrepancy indicates thathost switches likely took place during the evolution of thisPlasmodium clade Specifically P inui seems to derive fromparasites infecting Asian apes (Pacheco et al 2012b)The average distance between the haplotypes belonging tolineages A and B in orangutan malarias (0009 0001 for thecomplete mtDNA table 2) is similar to the divergence be-tween P vivax and P cynomolgi (0012 0002 table 2) Onthe other hand the average distances between clades A and C(0023 0002) and between clades B and C (0024 0002)are somewhat higher than those between clades A and B
Table 2 Genetic Divergences (substitutions per site) among Different Plasmodium Species Using the Kimura 2-Parameter Model As Implementedin MEGA v522 (Tamura et al 2011)
Species Genetic distance (d SE)
n COX1 COX3 CYTB COX1 + CYTB Complete mtDNA
Plasmodium sp (orangutan-clade A) 12 0005 0001 0006 0002 0003 0001 0004 0001 0004 00003
Plasmodium sp (orangutan-clade B) 13 0003 0001 0004 0001 0004 0001 0003 0001 0003 00003
Plasmodium sp (orangutan-clade C) 10 0002 0001 0003 0001 0004 0001 0003 0001 0003 00003
Orangutan clade A orangutan clade B 12 vs 13 0011 0002 0019 0005 0008 0002 0010 0001 0009 00010
Orangutan clade A orangutan clade C 12 vs 10 0034 0004 0043 0007 0028 0004 0031 0003 0023 00017
Orangutan clade B orangutan clade C 13 vs 10 0034 0004 0049 0008 0028 0005 0031 0003 0024 00018
Orangutan clade A + orangutan clade B 25 0008 0001 0012 0003 0006 0001 0007 0001 0006 00006
P cynomolgi (all strains) 14 0005 0001 0006 0001 0005 0001 0004 0001 0004 00006
P cynomolgi (clade A-Berok) 4 0007 0002 0007 0002 0006 0002 0006 0001 0006 00008
P cynomolgi (clade B-B) 10 0001 0001 0001 0001 0001 0001 0001 0000 0001 00002
P cynomolgi (clade A vs clade B) 4 vs10 0010 0002 0010 0003 0009 0002 0009 0002 0009 00011
P vivax 109 0001 0000 0001 0001 0 0001 0000 0001 00002
P vivaxndashP cynomolgi 109 vs 14 0014 0003 0017 0004 0013 0003 0014 0002 0012 00015
P inui (all) 31 0013 0002 0023 0004 0015 0002 0014 0001 0013 00010
P inui (Macaca nemestrina)-(M fascicularis) 9 vs 8 0004 0001 0005 0001 0003 0001 0003 0000 0004 00000
P inui (all)ndashP sp from orang clade A 31 vs 12 0023 0003 0045 0007 0018 0002 0024 0003 0023 00010
P inui (all)ndashP sp from orang clade B 31 vs 13 0025 0004 0054 0008 0027 0004 0026 0002 0025 00020
P inui (all)ndashP sp from orang clade A + B 31 vs 25 0024 0003 0050 0007 0026 0004 0025 0002 0024 00020
P knowlesi (all) 65 0001 0000 0002 0001 0001 0000 0001 0000 0001 00000
P knowlesi (NCBI)ndashSepilok 60 vs 5 0002 0000 0002 0001 0001 0000 0001 0000 0001 00000
P knowlesindashP coatneyi 65 vs 2 0043 0005 0034 0007 0025 0005 0035 0004 0032 00020
Plasmodium sp (MnendashMfa) 1 vs 1 0 0 0003 0001 0001 0001 0003 00010
P knowlesindashPlasmodium sp 65 vs 2 0037 0005 0037 0007 0020 0004 0030 0003 0029 00020
P coatneyindashPlasmodium sp 2 vs 2 0032 0005 0037 0007 0019 0004 0026 0003 0023 00020
P falciparum 101 0 0000 0001 0001 0001 0001 0000 0000 00001
P reichenowindashP falciparum 2 vs 101 0032 0005 0055 0009 0025 0005 0029 0003 0025 00018
425
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
between P vivax and P cynomolgi and between P reichenowiand P falciparum These results suggest that the haplotypesidentified in figure 1 as AndashC may belong to three differentspecies Clade A includes haplotypes described previously in adifferent population of orangutans (Pacheco et al 2012b)
In the case of P cynomolgi we found two closely relatedlineages in both species of macaques These lineages belong toone of the two clades (fig 1) that have been described for Pcynomolgi using antigen-encoding genes (Pacheco et al 20072010 Pacheco et al 2012a Pacheco et al 2012b) which con-tains the Berok PT2 and Gombak strains Indeed the averagegenetic distance among these four P cynomolgi strains(0006 00008) is relatively small if we consider its broadgeographic range (see table 2) It is also worth noting thatthe divergence of these two clades of P cynomolgi is compa-rable to that observed between P vivax and P cynomolgi(table 2) an observation that has been corroborated by ge-nomic data (Tachibana et al 2012)
Malaria Antigens Apical Membrane Antigen-1 andMerozoite Surface Protein 1 42 kDa
In addition to mtDNA genomes phylogenetic analyses werealso performed separately on data from several macaque andorangutan parasites using nuclear genes encoding two majorantigens Apical membrane antigen-1 (AMA-1 fig 2 and sup-plementary table S1 Supplementary Material online) andmerozoite surface protein 1 42 kDa (MSP-142 fig 3 and sup-plementary table S1 Supplementary Material online) Despitethe smaller number of species for which data is available thephylogenies estimated from these two loci are largely com-patible with previous studies using genes encoding merozoiteantigens (MSP-1 Pacheco et al 2007 Pacheco et al 2012bRAP-1 Pacheco et al 2010 MSP-8MSP-10 Pacheco et al2012a) In particular in both phylogenies we observe thatseveral of the orangutan malaria lineages form a monophy-letic group sharing a common ancestor with the macaqueparasite P inui One difference between the phylogenies gen-erated using either mtDNA or these two antigen-encodinggenes is in the relative position of P hylobati This is notsurprising considering that the position of P hylobati hasmodest support in the mtDNA (085 posterior probabilityand 63 bootstrap support see fig 1) Although in the mito-chondrial tree this lineage is located within the clade thatincludes the three orangutan lineages and P inui in theama-1 and msp-142 phylogenies it is located at the base ofthe clade containing P inui and the three orangutan parasiteclades (A-C fig 1) This discrepancy could have three differentexplanations First because the mitochondrial and nuclearloci are unlinked there could be random differences in thegenealogies of these loci Second this could be due to diver-sifying- or frequency-dependent selection acting on the anti-gen loci Finally this could be an artifact of our inability togenerate antigen sequences for all of the taxa included in themitochondrial tree Indeed because we were unable to gen-erate any sequences of ama-1 that can be unambiguouslyassigned to mtDNA clade C the position of P hylobati relativeto the orangutan clades is unresolved (fig 2)
In the case of ama-1 we were able to obtain sequences forP knowlesi Plasmodium sp P cynomolgi P inui from Mnemestrina and Plasmodium sp from orangutan The geneticpolymorphism of ama-1 per species is reported in the sup-plementary table S2 Supplementary Material online Theama-1 allele found in this study for P cynomolgi was mostclosely related to that contained in the Berok strain this resultis consistent with the mtDNA phylogeny Again the unde-scribed Plasmodium sp isolated from macaques forms a cladealong with P coatneyi and the average genetic distance esti-mated between the ama-1 allele for this parasite and P coat-neyi is very high (0076 0073) supporting the hypothesisthat the former organism is a new species On the other handwe only were able to generate msp-142 sequences for P inuiand the Plasmodium species detected in orangutans The ge-netic polymorphism per species for msp-142 is reported insupplementary table S3 Supplementary Material onlineOverall using these additional msp-142 sequences we recov-ered the same topology that was previously reported for thisantigen (Pacheco et al 2007 Pacheco et al 2012b)
Phylogenetic-based analyses provide some insight into therole of selection at these two loci (Pond et al 2011) In thecase of ama-1 several lineages contain codons that appear tobe under positive selection ( 41 Plt 005 corrected formultiple testing using HolmndashBonferroni methodSupplementary table S4 and fig S1 Supplementary Materialonline) Furthermore many of the nodes that may corre-spond to host switches also have significant uncorrected se-quential likelihood ratio tests (LRTs) A similar patternemerges from msp-142 where this method identifies four lin-eages that appear to be under episodic diversifying selection(supplementary table S4 and fig S1 Supplementary Materialonline)
Haplotype Networks
A median joining network of 41 Plasmodium mtDNA haplo-types isolated from orangutans from Borneo is depicted infigure 4 This network includes both the 30 PlasmodiummtDNA haplotypes from Sabah and the 11 mtDNA haplo-types from the Kalimantan orangutans As expected thethree clades identified in the mtDNA tree were clearly sepa-rated in the haplotype network Furthermore clade A itself(colored in green) contains two genetically differentiated pop-ulations (light green Sabah dark green Kalimantan) andindeed the fixation index (FST) between these two popula-tions is high (056) In another context the Venn diagramdepicted in figure 4 shows the number of orangutans withmixed infections of Plasmodium At least 10 of the 20 orang-utan samples harbored mixed infections with parasites fromtwo distinct clades but none was found to be simultaneouslyinfected by all three species
Figure 5 shows a median joining network for the 16 P inuimtDNA haplotypes from both species of Macaca (eight hap-lotypes from M nemestrina and eight from M fascicularis)No evidence of population structure between hosts wasfound in this analysis in fact the fixation index (FST) is effec-tively 0 (0024) In addition we also identified a divergent P
426
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 2 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding AMA-1 Bayesian and ML methods yielded identical topologiesand so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) The values abovebranches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials and Methods)Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shown Cladenumbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section Samples with indicate mixed infectionThe outgroup is indicated by the gray branch
427
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 3 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding MSP-142 kDa Bayesian and ML methods yielded identicaltopologies and so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) Thevalues above branches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials andMethods) Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shownClade numbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section The outgroup is indicated by thegray branch
428
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
inui mtDNA haplotype that was carried by a single individualof M nemestrina (see also fig 1)
The Ages and Branching Rates of the Asian PrimateMalarias
The mitochondrial genome phylogeny was used to estimatethe ages of the major clades of South Asian primate malariasunder three different calibration scenarios (fig 6 and supple-mentary fig S2 Supplementary Material online) These timesalong with the 95 credibility intervals (Crls) were calculatedusing BEAST v175 (Drummond et al 2012) and are shown intable 3 (See supplementary fig S2 Supplementary Materialonline for the node numbers) As expected there is extensiveoverlap among the Crls obtained under the three differentscenarios used for calibration When the phylogeny is cali-brated by assuming that the Asian and African primate par-asites split in the same time frame that their hosts becamegeographically isolated (6ndash142 Ma) and that lemur parasitesoriginated as a monophyletic group at least 20 Ma (seeMaterials and Methods) the Asian primate malarias are esti-mated to have originated between 538 and 852 Ma The splitbetween P inui and the orangutan Plasmodium lineages in
clades A and B was around 292 and 480 Ma which is inagreement with an earlier study of clade A (246ndash438 MaPacheco et al 2012b) Furthermore clades A and B divergedapproximately 10ndash23 Ma (table 3) this is clearly older thantime estimates for the split of the Bornean and Sumatranorangutans using nuclear single nucleotide polymorphisms(SNPs) but earlier than estimates for the same two speciesusing the mitochondrial genome (Ma et al 2013) On theother hand the common ancestor of the orangutanPlasmodium lineages in clade C appears to have existed be-tween 371 and 598 Ma well after the time of the split be-tween orangutan and gibbons or the most recent commonancestor of Asian apes with macaques
Previous phylogenetic studies have indicated that P coat-neyi and P knowlesi are sister taxa (eg Escalante et al 2005)and indeed these two species are usually referred as ldquocloselyrelatedrdquo in the malaria literature In this study the age of themost recent common ancestor of P knowlesi and P coatneyiwas estimated to be 554 Ma (CrI 417ndash710 table 3) whichlies within the time interval proposed for the radiation ofmacaques and other nonhuman primates in the Malaysianarchipelago (Delson 1980 Perelman et al 2011 Springer et al2012)
FIG 4 Venn diagram of mixed infections and median joining network of Plasmodium sp mtDNA haplotypes from orangutans from BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to differentclades green = clade A red = clade B and blue = clade C
429
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
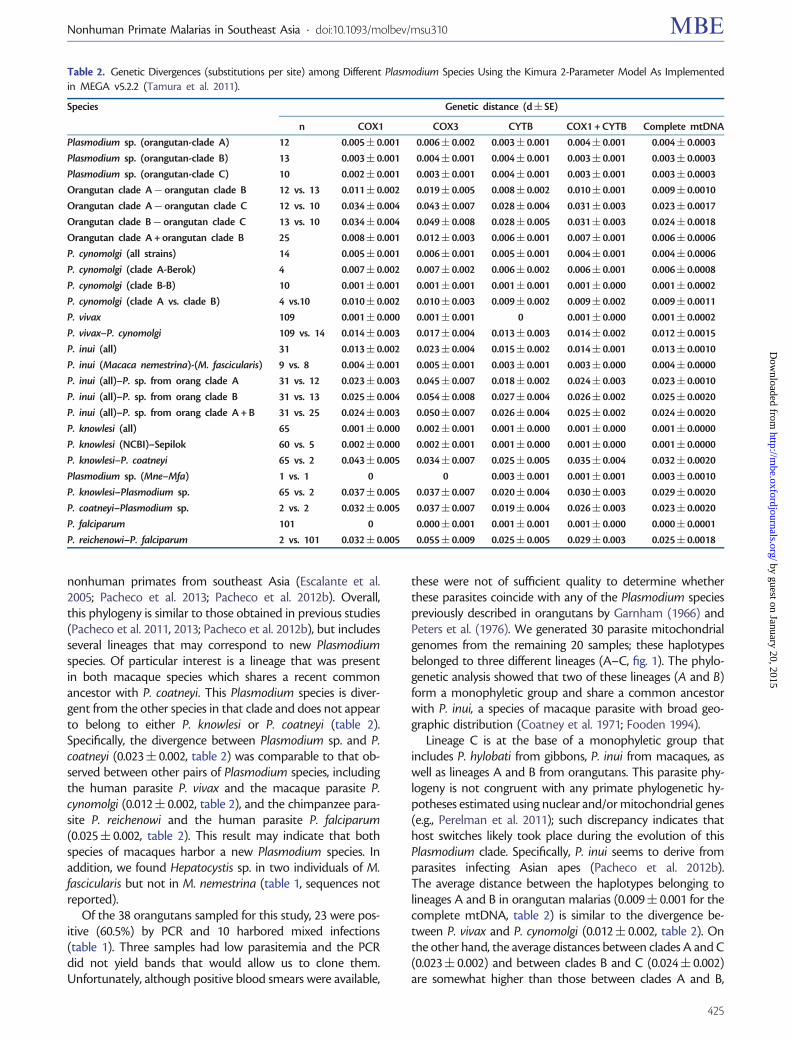
nonhuman primates from southeast Asia (Escalante et al2005 Pacheco et al 2013 Pacheco et al 2012b) Overallthis phylogeny is similar to those obtained in previous studies(Pacheco et al 2011 2013 Pacheco et al 2012b) but includesseveral lineages that may correspond to new Plasmodiumspecies Of particular interest is a lineage that was presentin both macaque species which shares a recent commonancestor with P coatneyi This Plasmodium species is diver-gent from the other species in that clade and does not appearto belong to either P knowlesi or P coatneyi (table 2)Specifically the divergence between Plasmodium sp and Pcoatneyi (0023 0002 table 2) was comparable to that ob-served between other pairs of Plasmodium species includingthe human parasite P vivax and the macaque parasite Pcynomolgi (0012 0002 table 2) and the chimpanzee para-site P reichenowi and the human parasite P falciparum(0025 0002 table 2) This result may indicate that bothspecies of macaques harbor a new Plasmodium species Inaddition we found Hepatocystis sp in two individuals of Mfascicularis but not in M nemestrina (table 1 sequences notreported)
Of the 38 orangutans sampled for this study 23 were pos-itive (605) by PCR and 10 harbored mixed infections(table 1) Three samples had low parasitemia and the PCRdid not yield bands that would allow us to clone themUnfortunately although positive blood smears were available
these were not of sufficient quality to determine whetherthese parasites coincide with any of the Plasmodium speciespreviously described in orangutans by Garnham (1966) andPeters et al (1976) We generated 30 parasite mitochondrialgenomes from the remaining 20 samples these haplotypesbelonged to three different lineages (AndashC fig 1) The phylo-genetic analysis showed that two of these lineages (A and B)form a monophyletic group and share a common ancestorwith P inui a species of macaque parasite with broad geo-graphic distribution (Coatney et al 1971 Fooden 1994)
Lineage C is at the base of a monophyletic group thatincludes P hylobati from gibbons P inui from macaques aswell as lineages A and B from orangutans This parasite phy-logeny is not congruent with any primate phylogenetic hy-potheses estimated using nuclear andor mitochondrial genes(eg Perelman et al 2011) such discrepancy indicates thathost switches likely took place during the evolution of thisPlasmodium clade Specifically P inui seems to derive fromparasites infecting Asian apes (Pacheco et al 2012b)The average distance between the haplotypes belonging tolineages A and B in orangutan malarias (0009 0001 for thecomplete mtDNA table 2) is similar to the divergence be-tween P vivax and P cynomolgi (0012 0002 table 2) Onthe other hand the average distances between clades A and C(0023 0002) and between clades B and C (0024 0002)are somewhat higher than those between clades A and B
Table 2 Genetic Divergences (substitutions per site) among Different Plasmodium Species Using the Kimura 2-Parameter Model As Implementedin MEGA v522 (Tamura et al 2011)
Species Genetic distance (d SE)
n COX1 COX3 CYTB COX1 + CYTB Complete mtDNA
Plasmodium sp (orangutan-clade A) 12 0005 0001 0006 0002 0003 0001 0004 0001 0004 00003
Plasmodium sp (orangutan-clade B) 13 0003 0001 0004 0001 0004 0001 0003 0001 0003 00003
Plasmodium sp (orangutan-clade C) 10 0002 0001 0003 0001 0004 0001 0003 0001 0003 00003
Orangutan clade A orangutan clade B 12 vs 13 0011 0002 0019 0005 0008 0002 0010 0001 0009 00010
Orangutan clade A orangutan clade C 12 vs 10 0034 0004 0043 0007 0028 0004 0031 0003 0023 00017
Orangutan clade B orangutan clade C 13 vs 10 0034 0004 0049 0008 0028 0005 0031 0003 0024 00018
Orangutan clade A + orangutan clade B 25 0008 0001 0012 0003 0006 0001 0007 0001 0006 00006
P cynomolgi (all strains) 14 0005 0001 0006 0001 0005 0001 0004 0001 0004 00006
P cynomolgi (clade A-Berok) 4 0007 0002 0007 0002 0006 0002 0006 0001 0006 00008
P cynomolgi (clade B-B) 10 0001 0001 0001 0001 0001 0001 0001 0000 0001 00002
P cynomolgi (clade A vs clade B) 4 vs10 0010 0002 0010 0003 0009 0002 0009 0002 0009 00011
P vivax 109 0001 0000 0001 0001 0 0001 0000 0001 00002
P vivaxndashP cynomolgi 109 vs 14 0014 0003 0017 0004 0013 0003 0014 0002 0012 00015
P inui (all) 31 0013 0002 0023 0004 0015 0002 0014 0001 0013 00010
P inui (Macaca nemestrina)-(M fascicularis) 9 vs 8 0004 0001 0005 0001 0003 0001 0003 0000 0004 00000
P inui (all)ndashP sp from orang clade A 31 vs 12 0023 0003 0045 0007 0018 0002 0024 0003 0023 00010
P inui (all)ndashP sp from orang clade B 31 vs 13 0025 0004 0054 0008 0027 0004 0026 0002 0025 00020
P inui (all)ndashP sp from orang clade A + B 31 vs 25 0024 0003 0050 0007 0026 0004 0025 0002 0024 00020
P knowlesi (all) 65 0001 0000 0002 0001 0001 0000 0001 0000 0001 00000
P knowlesi (NCBI)ndashSepilok 60 vs 5 0002 0000 0002 0001 0001 0000 0001 0000 0001 00000
P knowlesindashP coatneyi 65 vs 2 0043 0005 0034 0007 0025 0005 0035 0004 0032 00020
Plasmodium sp (MnendashMfa) 1 vs 1 0 0 0003 0001 0001 0001 0003 00010
P knowlesindashPlasmodium sp 65 vs 2 0037 0005 0037 0007 0020 0004 0030 0003 0029 00020
P coatneyindashPlasmodium sp 2 vs 2 0032 0005 0037 0007 0019 0004 0026 0003 0023 00020
P falciparum 101 0 0000 0001 0001 0001 0001 0000 0000 00001
P reichenowindashP falciparum 2 vs 101 0032 0005 0055 0009 0025 0005 0029 0003 0025 00018
425
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
between P vivax and P cynomolgi and between P reichenowiand P falciparum These results suggest that the haplotypesidentified in figure 1 as AndashC may belong to three differentspecies Clade A includes haplotypes described previously in adifferent population of orangutans (Pacheco et al 2012b)
In the case of P cynomolgi we found two closely relatedlineages in both species of macaques These lineages belong toone of the two clades (fig 1) that have been described for Pcynomolgi using antigen-encoding genes (Pacheco et al 20072010 Pacheco et al 2012a Pacheco et al 2012b) which con-tains the Berok PT2 and Gombak strains Indeed the averagegenetic distance among these four P cynomolgi strains(0006 00008) is relatively small if we consider its broadgeographic range (see table 2) It is also worth noting thatthe divergence of these two clades of P cynomolgi is compa-rable to that observed between P vivax and P cynomolgi(table 2) an observation that has been corroborated by ge-nomic data (Tachibana et al 2012)
Malaria Antigens Apical Membrane Antigen-1 andMerozoite Surface Protein 1 42 kDa
In addition to mtDNA genomes phylogenetic analyses werealso performed separately on data from several macaque andorangutan parasites using nuclear genes encoding two majorantigens Apical membrane antigen-1 (AMA-1 fig 2 and sup-plementary table S1 Supplementary Material online) andmerozoite surface protein 1 42 kDa (MSP-142 fig 3 and sup-plementary table S1 Supplementary Material online) Despitethe smaller number of species for which data is available thephylogenies estimated from these two loci are largely com-patible with previous studies using genes encoding merozoiteantigens (MSP-1 Pacheco et al 2007 Pacheco et al 2012bRAP-1 Pacheco et al 2010 MSP-8MSP-10 Pacheco et al2012a) In particular in both phylogenies we observe thatseveral of the orangutan malaria lineages form a monophy-letic group sharing a common ancestor with the macaqueparasite P inui One difference between the phylogenies gen-erated using either mtDNA or these two antigen-encodinggenes is in the relative position of P hylobati This is notsurprising considering that the position of P hylobati hasmodest support in the mtDNA (085 posterior probabilityand 63 bootstrap support see fig 1) Although in the mito-chondrial tree this lineage is located within the clade thatincludes the three orangutan lineages and P inui in theama-1 and msp-142 phylogenies it is located at the base ofthe clade containing P inui and the three orangutan parasiteclades (A-C fig 1) This discrepancy could have three differentexplanations First because the mitochondrial and nuclearloci are unlinked there could be random differences in thegenealogies of these loci Second this could be due to diver-sifying- or frequency-dependent selection acting on the anti-gen loci Finally this could be an artifact of our inability togenerate antigen sequences for all of the taxa included in themitochondrial tree Indeed because we were unable to gen-erate any sequences of ama-1 that can be unambiguouslyassigned to mtDNA clade C the position of P hylobati relativeto the orangutan clades is unresolved (fig 2)
In the case of ama-1 we were able to obtain sequences forP knowlesi Plasmodium sp P cynomolgi P inui from Mnemestrina and Plasmodium sp from orangutan The geneticpolymorphism of ama-1 per species is reported in the sup-plementary table S2 Supplementary Material online Theama-1 allele found in this study for P cynomolgi was mostclosely related to that contained in the Berok strain this resultis consistent with the mtDNA phylogeny Again the unde-scribed Plasmodium sp isolated from macaques forms a cladealong with P coatneyi and the average genetic distance esti-mated between the ama-1 allele for this parasite and P coat-neyi is very high (0076 0073) supporting the hypothesisthat the former organism is a new species On the other handwe only were able to generate msp-142 sequences for P inuiand the Plasmodium species detected in orangutans The ge-netic polymorphism per species for msp-142 is reported insupplementary table S3 Supplementary Material onlineOverall using these additional msp-142 sequences we recov-ered the same topology that was previously reported for thisantigen (Pacheco et al 2007 Pacheco et al 2012b)
Phylogenetic-based analyses provide some insight into therole of selection at these two loci (Pond et al 2011) In thecase of ama-1 several lineages contain codons that appear tobe under positive selection ( 41 Plt 005 corrected formultiple testing using HolmndashBonferroni methodSupplementary table S4 and fig S1 Supplementary Materialonline) Furthermore many of the nodes that may corre-spond to host switches also have significant uncorrected se-quential likelihood ratio tests (LRTs) A similar patternemerges from msp-142 where this method identifies four lin-eages that appear to be under episodic diversifying selection(supplementary table S4 and fig S1 Supplementary Materialonline)
Haplotype Networks
A median joining network of 41 Plasmodium mtDNA haplo-types isolated from orangutans from Borneo is depicted infigure 4 This network includes both the 30 PlasmodiummtDNA haplotypes from Sabah and the 11 mtDNA haplo-types from the Kalimantan orangutans As expected thethree clades identified in the mtDNA tree were clearly sepa-rated in the haplotype network Furthermore clade A itself(colored in green) contains two genetically differentiated pop-ulations (light green Sabah dark green Kalimantan) andindeed the fixation index (FST) between these two popula-tions is high (056) In another context the Venn diagramdepicted in figure 4 shows the number of orangutans withmixed infections of Plasmodium At least 10 of the 20 orang-utan samples harbored mixed infections with parasites fromtwo distinct clades but none was found to be simultaneouslyinfected by all three species
Figure 5 shows a median joining network for the 16 P inuimtDNA haplotypes from both species of Macaca (eight hap-lotypes from M nemestrina and eight from M fascicularis)No evidence of population structure between hosts wasfound in this analysis in fact the fixation index (FST) is effec-tively 0 (0024) In addition we also identified a divergent P
426
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 2 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding AMA-1 Bayesian and ML methods yielded identical topologiesand so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) The values abovebranches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials and Methods)Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shown Cladenumbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section Samples with indicate mixed infectionThe outgroup is indicated by the gray branch
427
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 3 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding MSP-142 kDa Bayesian and ML methods yielded identicaltopologies and so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) Thevalues above branches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials andMethods) Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shownClade numbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section The outgroup is indicated by thegray branch
428
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
inui mtDNA haplotype that was carried by a single individualof M nemestrina (see also fig 1)
The Ages and Branching Rates of the Asian PrimateMalarias
The mitochondrial genome phylogeny was used to estimatethe ages of the major clades of South Asian primate malariasunder three different calibration scenarios (fig 6 and supple-mentary fig S2 Supplementary Material online) These timesalong with the 95 credibility intervals (Crls) were calculatedusing BEAST v175 (Drummond et al 2012) and are shown intable 3 (See supplementary fig S2 Supplementary Materialonline for the node numbers) As expected there is extensiveoverlap among the Crls obtained under the three differentscenarios used for calibration When the phylogeny is cali-brated by assuming that the Asian and African primate par-asites split in the same time frame that their hosts becamegeographically isolated (6ndash142 Ma) and that lemur parasitesoriginated as a monophyletic group at least 20 Ma (seeMaterials and Methods) the Asian primate malarias are esti-mated to have originated between 538 and 852 Ma The splitbetween P inui and the orangutan Plasmodium lineages in
clades A and B was around 292 and 480 Ma which is inagreement with an earlier study of clade A (246ndash438 MaPacheco et al 2012b) Furthermore clades A and B divergedapproximately 10ndash23 Ma (table 3) this is clearly older thantime estimates for the split of the Bornean and Sumatranorangutans using nuclear single nucleotide polymorphisms(SNPs) but earlier than estimates for the same two speciesusing the mitochondrial genome (Ma et al 2013) On theother hand the common ancestor of the orangutanPlasmodium lineages in clade C appears to have existed be-tween 371 and 598 Ma well after the time of the split be-tween orangutan and gibbons or the most recent commonancestor of Asian apes with macaques
Previous phylogenetic studies have indicated that P coat-neyi and P knowlesi are sister taxa (eg Escalante et al 2005)and indeed these two species are usually referred as ldquocloselyrelatedrdquo in the malaria literature In this study the age of themost recent common ancestor of P knowlesi and P coatneyiwas estimated to be 554 Ma (CrI 417ndash710 table 3) whichlies within the time interval proposed for the radiation ofmacaques and other nonhuman primates in the Malaysianarchipelago (Delson 1980 Perelman et al 2011 Springer et al2012)
FIG 4 Venn diagram of mixed infections and median joining network of Plasmodium sp mtDNA haplotypes from orangutans from BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to differentclades green = clade A red = clade B and blue = clade C
429
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

between P vivax and P cynomolgi and between P reichenowiand P falciparum These results suggest that the haplotypesidentified in figure 1 as AndashC may belong to three differentspecies Clade A includes haplotypes described previously in adifferent population of orangutans (Pacheco et al 2012b)
In the case of P cynomolgi we found two closely relatedlineages in both species of macaques These lineages belong toone of the two clades (fig 1) that have been described for Pcynomolgi using antigen-encoding genes (Pacheco et al 20072010 Pacheco et al 2012a Pacheco et al 2012b) which con-tains the Berok PT2 and Gombak strains Indeed the averagegenetic distance among these four P cynomolgi strains(0006 00008) is relatively small if we consider its broadgeographic range (see table 2) It is also worth noting thatthe divergence of these two clades of P cynomolgi is compa-rable to that observed between P vivax and P cynomolgi(table 2) an observation that has been corroborated by ge-nomic data (Tachibana et al 2012)
Malaria Antigens Apical Membrane Antigen-1 andMerozoite Surface Protein 1 42 kDa
In addition to mtDNA genomes phylogenetic analyses werealso performed separately on data from several macaque andorangutan parasites using nuclear genes encoding two majorantigens Apical membrane antigen-1 (AMA-1 fig 2 and sup-plementary table S1 Supplementary Material online) andmerozoite surface protein 1 42 kDa (MSP-142 fig 3 and sup-plementary table S1 Supplementary Material online) Despitethe smaller number of species for which data is available thephylogenies estimated from these two loci are largely com-patible with previous studies using genes encoding merozoiteantigens (MSP-1 Pacheco et al 2007 Pacheco et al 2012bRAP-1 Pacheco et al 2010 MSP-8MSP-10 Pacheco et al2012a) In particular in both phylogenies we observe thatseveral of the orangutan malaria lineages form a monophy-letic group sharing a common ancestor with the macaqueparasite P inui One difference between the phylogenies gen-erated using either mtDNA or these two antigen-encodinggenes is in the relative position of P hylobati This is notsurprising considering that the position of P hylobati hasmodest support in the mtDNA (085 posterior probabilityand 63 bootstrap support see fig 1) Although in the mito-chondrial tree this lineage is located within the clade thatincludes the three orangutan lineages and P inui in theama-1 and msp-142 phylogenies it is located at the base ofthe clade containing P inui and the three orangutan parasiteclades (A-C fig 1) This discrepancy could have three differentexplanations First because the mitochondrial and nuclearloci are unlinked there could be random differences in thegenealogies of these loci Second this could be due to diver-sifying- or frequency-dependent selection acting on the anti-gen loci Finally this could be an artifact of our inability togenerate antigen sequences for all of the taxa included in themitochondrial tree Indeed because we were unable to gen-erate any sequences of ama-1 that can be unambiguouslyassigned to mtDNA clade C the position of P hylobati relativeto the orangutan clades is unresolved (fig 2)
In the case of ama-1 we were able to obtain sequences forP knowlesi Plasmodium sp P cynomolgi P inui from Mnemestrina and Plasmodium sp from orangutan The geneticpolymorphism of ama-1 per species is reported in the sup-plementary table S2 Supplementary Material online Theama-1 allele found in this study for P cynomolgi was mostclosely related to that contained in the Berok strain this resultis consistent with the mtDNA phylogeny Again the unde-scribed Plasmodium sp isolated from macaques forms a cladealong with P coatneyi and the average genetic distance esti-mated between the ama-1 allele for this parasite and P coat-neyi is very high (0076 0073) supporting the hypothesisthat the former organism is a new species On the other handwe only were able to generate msp-142 sequences for P inuiand the Plasmodium species detected in orangutans The ge-netic polymorphism per species for msp-142 is reported insupplementary table S3 Supplementary Material onlineOverall using these additional msp-142 sequences we recov-ered the same topology that was previously reported for thisantigen (Pacheco et al 2007 Pacheco et al 2012b)
Phylogenetic-based analyses provide some insight into therole of selection at these two loci (Pond et al 2011) In thecase of ama-1 several lineages contain codons that appear tobe under positive selection ( 41 Plt 005 corrected formultiple testing using HolmndashBonferroni methodSupplementary table S4 and fig S1 Supplementary Materialonline) Furthermore many of the nodes that may corre-spond to host switches also have significant uncorrected se-quential likelihood ratio tests (LRTs) A similar patternemerges from msp-142 where this method identifies four lin-eages that appear to be under episodic diversifying selection(supplementary table S4 and fig S1 Supplementary Materialonline)
Haplotype Networks
A median joining network of 41 Plasmodium mtDNA haplo-types isolated from orangutans from Borneo is depicted infigure 4 This network includes both the 30 PlasmodiummtDNA haplotypes from Sabah and the 11 mtDNA haplo-types from the Kalimantan orangutans As expected thethree clades identified in the mtDNA tree were clearly sepa-rated in the haplotype network Furthermore clade A itself(colored in green) contains two genetically differentiated pop-ulations (light green Sabah dark green Kalimantan) andindeed the fixation index (FST) between these two popula-tions is high (056) In another context the Venn diagramdepicted in figure 4 shows the number of orangutans withmixed infections of Plasmodium At least 10 of the 20 orang-utan samples harbored mixed infections with parasites fromtwo distinct clades but none was found to be simultaneouslyinfected by all three species
Figure 5 shows a median joining network for the 16 P inuimtDNA haplotypes from both species of Macaca (eight hap-lotypes from M nemestrina and eight from M fascicularis)No evidence of population structure between hosts wasfound in this analysis in fact the fixation index (FST) is effec-tively 0 (0024) In addition we also identified a divergent P
426
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 2 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding AMA-1 Bayesian and ML methods yielded identical topologiesand so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) The values abovebranches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials and Methods)Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shown Cladenumbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section Samples with indicate mixed infectionThe outgroup is indicated by the gray branch
427
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 3 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding MSP-142 kDa Bayesian and ML methods yielded identicaltopologies and so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) Thevalues above branches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials andMethods) Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shownClade numbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section The outgroup is indicated by thegray branch
428
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
inui mtDNA haplotype that was carried by a single individualof M nemestrina (see also fig 1)
The Ages and Branching Rates of the Asian PrimateMalarias
The mitochondrial genome phylogeny was used to estimatethe ages of the major clades of South Asian primate malariasunder three different calibration scenarios (fig 6 and supple-mentary fig S2 Supplementary Material online) These timesalong with the 95 credibility intervals (Crls) were calculatedusing BEAST v175 (Drummond et al 2012) and are shown intable 3 (See supplementary fig S2 Supplementary Materialonline for the node numbers) As expected there is extensiveoverlap among the Crls obtained under the three differentscenarios used for calibration When the phylogeny is cali-brated by assuming that the Asian and African primate par-asites split in the same time frame that their hosts becamegeographically isolated (6ndash142 Ma) and that lemur parasitesoriginated as a monophyletic group at least 20 Ma (seeMaterials and Methods) the Asian primate malarias are esti-mated to have originated between 538 and 852 Ma The splitbetween P inui and the orangutan Plasmodium lineages in
clades A and B was around 292 and 480 Ma which is inagreement with an earlier study of clade A (246ndash438 MaPacheco et al 2012b) Furthermore clades A and B divergedapproximately 10ndash23 Ma (table 3) this is clearly older thantime estimates for the split of the Bornean and Sumatranorangutans using nuclear single nucleotide polymorphisms(SNPs) but earlier than estimates for the same two speciesusing the mitochondrial genome (Ma et al 2013) On theother hand the common ancestor of the orangutanPlasmodium lineages in clade C appears to have existed be-tween 371 and 598 Ma well after the time of the split be-tween orangutan and gibbons or the most recent commonancestor of Asian apes with macaques
Previous phylogenetic studies have indicated that P coat-neyi and P knowlesi are sister taxa (eg Escalante et al 2005)and indeed these two species are usually referred as ldquocloselyrelatedrdquo in the malaria literature In this study the age of themost recent common ancestor of P knowlesi and P coatneyiwas estimated to be 554 Ma (CrI 417ndash710 table 3) whichlies within the time interval proposed for the radiation ofmacaques and other nonhuman primates in the Malaysianarchipelago (Delson 1980 Perelman et al 2011 Springer et al2012)
FIG 4 Venn diagram of mixed infections and median joining network of Plasmodium sp mtDNA haplotypes from orangutans from BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to differentclades green = clade A red = clade B and blue = clade C
429
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
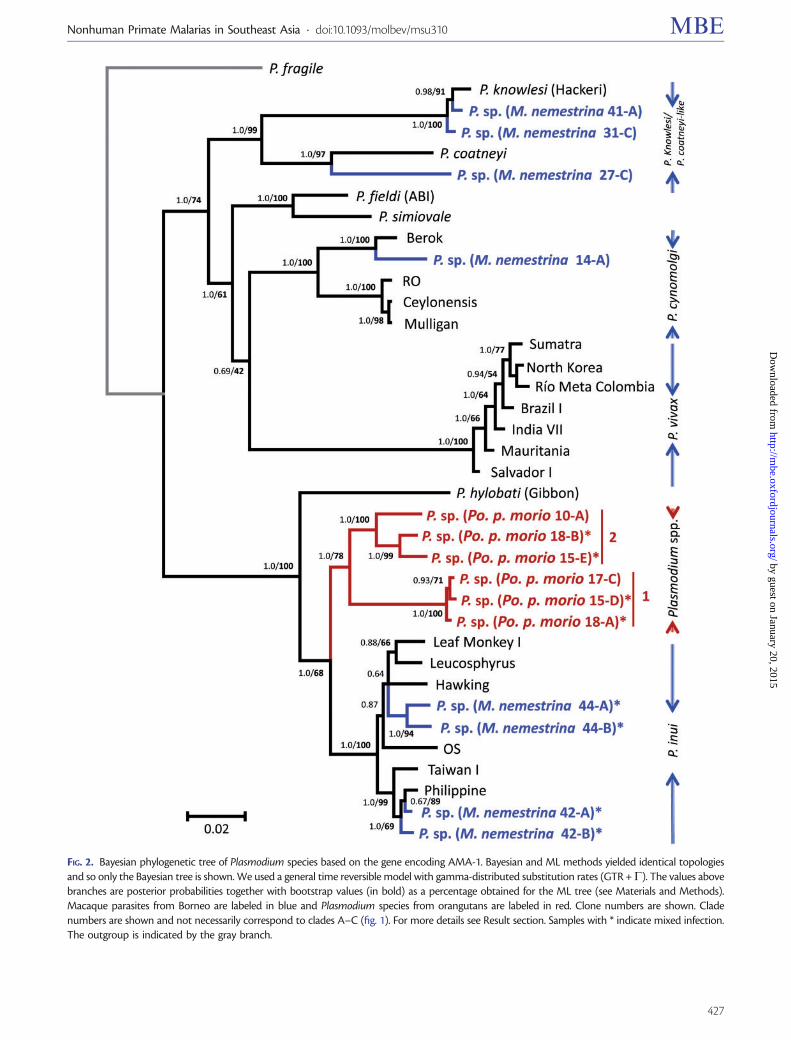
FIG 2 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding AMA-1 Bayesian and ML methods yielded identical topologiesand so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) The values abovebranches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials and Methods)Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shown Cladenumbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section Samples with indicate mixed infectionThe outgroup is indicated by the gray branch
427
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
FIG 3 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding MSP-142 kDa Bayesian and ML methods yielded identicaltopologies and so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) Thevalues above branches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials andMethods) Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shownClade numbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section The outgroup is indicated by thegray branch
428
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
inui mtDNA haplotype that was carried by a single individualof M nemestrina (see also fig 1)
The Ages and Branching Rates of the Asian PrimateMalarias
The mitochondrial genome phylogeny was used to estimatethe ages of the major clades of South Asian primate malariasunder three different calibration scenarios (fig 6 and supple-mentary fig S2 Supplementary Material online) These timesalong with the 95 credibility intervals (Crls) were calculatedusing BEAST v175 (Drummond et al 2012) and are shown intable 3 (See supplementary fig S2 Supplementary Materialonline for the node numbers) As expected there is extensiveoverlap among the Crls obtained under the three differentscenarios used for calibration When the phylogeny is cali-brated by assuming that the Asian and African primate par-asites split in the same time frame that their hosts becamegeographically isolated (6ndash142 Ma) and that lemur parasitesoriginated as a monophyletic group at least 20 Ma (seeMaterials and Methods) the Asian primate malarias are esti-mated to have originated between 538 and 852 Ma The splitbetween P inui and the orangutan Plasmodium lineages in
clades A and B was around 292 and 480 Ma which is inagreement with an earlier study of clade A (246ndash438 MaPacheco et al 2012b) Furthermore clades A and B divergedapproximately 10ndash23 Ma (table 3) this is clearly older thantime estimates for the split of the Bornean and Sumatranorangutans using nuclear single nucleotide polymorphisms(SNPs) but earlier than estimates for the same two speciesusing the mitochondrial genome (Ma et al 2013) On theother hand the common ancestor of the orangutanPlasmodium lineages in clade C appears to have existed be-tween 371 and 598 Ma well after the time of the split be-tween orangutan and gibbons or the most recent commonancestor of Asian apes with macaques
Previous phylogenetic studies have indicated that P coat-neyi and P knowlesi are sister taxa (eg Escalante et al 2005)and indeed these two species are usually referred as ldquocloselyrelatedrdquo in the malaria literature In this study the age of themost recent common ancestor of P knowlesi and P coatneyiwas estimated to be 554 Ma (CrI 417ndash710 table 3) whichlies within the time interval proposed for the radiation ofmacaques and other nonhuman primates in the Malaysianarchipelago (Delson 1980 Perelman et al 2011 Springer et al2012)
FIG 4 Venn diagram of mixed infections and median joining network of Plasmodium sp mtDNA haplotypes from orangutans from BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to differentclades green = clade A red = clade B and blue = clade C
429
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
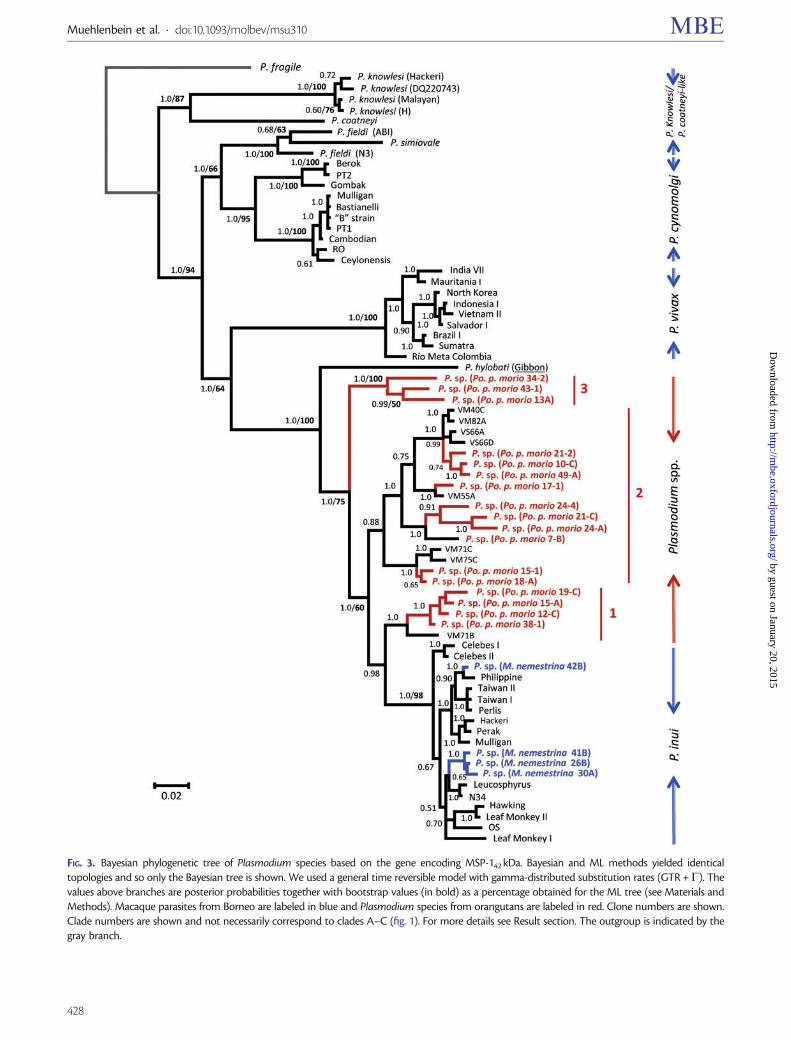
FIG 3 Bayesian phylogenetic tree of Plasmodium species based on the gene encoding MSP-142 kDa Bayesian and ML methods yielded identicaltopologies and so only the Bayesian tree is shown We used a general time reversible model with gamma-distributed substitution rates (GTR +G) Thevalues above branches are posterior probabilities together with bootstrap values (in bold) as a percentage obtained for the ML tree (see Materials andMethods) Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red Clone numbers are shownClade numbers are shown and not necessarily correspond to clades AndashC (fig 1) For more details see Result section The outgroup is indicated by thegray branch
428
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
inui mtDNA haplotype that was carried by a single individualof M nemestrina (see also fig 1)
The Ages and Branching Rates of the Asian PrimateMalarias
The mitochondrial genome phylogeny was used to estimatethe ages of the major clades of South Asian primate malariasunder three different calibration scenarios (fig 6 and supple-mentary fig S2 Supplementary Material online) These timesalong with the 95 credibility intervals (Crls) were calculatedusing BEAST v175 (Drummond et al 2012) and are shown intable 3 (See supplementary fig S2 Supplementary Materialonline for the node numbers) As expected there is extensiveoverlap among the Crls obtained under the three differentscenarios used for calibration When the phylogeny is cali-brated by assuming that the Asian and African primate par-asites split in the same time frame that their hosts becamegeographically isolated (6ndash142 Ma) and that lemur parasitesoriginated as a monophyletic group at least 20 Ma (seeMaterials and Methods) the Asian primate malarias are esti-mated to have originated between 538 and 852 Ma The splitbetween P inui and the orangutan Plasmodium lineages in
clades A and B was around 292 and 480 Ma which is inagreement with an earlier study of clade A (246ndash438 MaPacheco et al 2012b) Furthermore clades A and B divergedapproximately 10ndash23 Ma (table 3) this is clearly older thantime estimates for the split of the Bornean and Sumatranorangutans using nuclear single nucleotide polymorphisms(SNPs) but earlier than estimates for the same two speciesusing the mitochondrial genome (Ma et al 2013) On theother hand the common ancestor of the orangutanPlasmodium lineages in clade C appears to have existed be-tween 371 and 598 Ma well after the time of the split be-tween orangutan and gibbons or the most recent commonancestor of Asian apes with macaques
Previous phylogenetic studies have indicated that P coat-neyi and P knowlesi are sister taxa (eg Escalante et al 2005)and indeed these two species are usually referred as ldquocloselyrelatedrdquo in the malaria literature In this study the age of themost recent common ancestor of P knowlesi and P coatneyiwas estimated to be 554 Ma (CrI 417ndash710 table 3) whichlies within the time interval proposed for the radiation ofmacaques and other nonhuman primates in the Malaysianarchipelago (Delson 1980 Perelman et al 2011 Springer et al2012)
FIG 4 Venn diagram of mixed infections and median joining network of Plasmodium sp mtDNA haplotypes from orangutans from BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to differentclades green = clade A red = clade B and blue = clade C
429
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

inui mtDNA haplotype that was carried by a single individualof M nemestrina (see also fig 1)
The Ages and Branching Rates of the Asian PrimateMalarias
The mitochondrial genome phylogeny was used to estimatethe ages of the major clades of South Asian primate malariasunder three different calibration scenarios (fig 6 and supple-mentary fig S2 Supplementary Material online) These timesalong with the 95 credibility intervals (Crls) were calculatedusing BEAST v175 (Drummond et al 2012) and are shown intable 3 (See supplementary fig S2 Supplementary Materialonline for the node numbers) As expected there is extensiveoverlap among the Crls obtained under the three differentscenarios used for calibration When the phylogeny is cali-brated by assuming that the Asian and African primate par-asites split in the same time frame that their hosts becamegeographically isolated (6ndash142 Ma) and that lemur parasitesoriginated as a monophyletic group at least 20 Ma (seeMaterials and Methods) the Asian primate malarias are esti-mated to have originated between 538 and 852 Ma The splitbetween P inui and the orangutan Plasmodium lineages in
clades A and B was around 292 and 480 Ma which is inagreement with an earlier study of clade A (246ndash438 MaPacheco et al 2012b) Furthermore clades A and B divergedapproximately 10ndash23 Ma (table 3) this is clearly older thantime estimates for the split of the Bornean and Sumatranorangutans using nuclear single nucleotide polymorphisms(SNPs) but earlier than estimates for the same two speciesusing the mitochondrial genome (Ma et al 2013) On theother hand the common ancestor of the orangutanPlasmodium lineages in clade C appears to have existed be-tween 371 and 598 Ma well after the time of the split be-tween orangutan and gibbons or the most recent commonancestor of Asian apes with macaques
Previous phylogenetic studies have indicated that P coat-neyi and P knowlesi are sister taxa (eg Escalante et al 2005)and indeed these two species are usually referred as ldquocloselyrelatedrdquo in the malaria literature In this study the age of themost recent common ancestor of P knowlesi and P coatneyiwas estimated to be 554 Ma (CrI 417ndash710 table 3) whichlies within the time interval proposed for the radiation ofmacaques and other nonhuman primates in the Malaysianarchipelago (Delson 1980 Perelman et al 2011 Springer et al2012)
FIG 4 Venn diagram of mixed infections and median joining network of Plasmodium sp mtDNA haplotypes from orangutans from BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to differentclades green = clade A red = clade B and blue = clade C
429
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

Visual inspection of the Plasmodium phylogeny (fig 6)suggests that the speciation rate may have been elevatedwithin the clade of malaria parasites infecting Asian ma-caques In fact this hypothesis is consistent with the resultsobtained when we used the program MEDUSA to comparedifferent birthndashdeath models on a sample of 50 trees describ-ing the relationships between the Plasmodium species knownto infect mammalian hosts (table 4 and supplementary fig S3Supplementary Material online) In every case the correctedAkaike information criterion (AICc) calculated by MEDUSAwas minimized by a two component birthndashdeath model withaccelerated branching within the clade of Asian macaque-infecting parasites Although in five cases the reduction inAICc was too small to reject a simpler model with homoge-neous birth and death rates across the entire tree in theremaining 45 cases the two-component model was identifiedas the best-fitting model by the stepwise selection procedureimplemented in MEDUSA (table 4) Maximum likelihood(ML) estimates of the branching and extinction rates withinthe two-component models suggest that the parasites infect-ing Asian macaques have speciated approximately three tofour times more rapidly than the other lineages contained inthe tree Across the 50 trees analyzed the diversification rates(r = bndashd eventsmillion years) range from 01245 to 03257(mean 02055) within the Asian clade and 00394 to 00728
(mean 00580) within the rest of the tree Furthermore thesedifferences appear to be almost entirely due to changes in thebranching rate rather than the extinction rate Indeed inevery case the extinction rate was estimated to be less than02 of the branching rate
DiscussionAlthough there has been increasing awareness of the impor-tance of parasitic organisms for biodiversity (Poulin 2005)most studies continue to focus on the harm caused by par-asites to their hosts (Nunn and Altizer 2006) and pay relativelylittle attention to the role of parasites as intrinsic elements ofan ecosystem Nonetheless (Brooks and McLennan 1993 Page2003 Poulin 2005 Brooks et al 2006 Poulin et al 2011) it isnotable that research programs in evolutionary parasitologyaddressing host specialization (host range) and parasite spe-ciation have followed intertwined paths Although host evo-lutionary history is considered a major driver in parasitespeciation (Poulin et al 2011) the discussion has been re-cently enriched by highlighting the importance of geographicpatterns in the host or hosts populations (Nieberding et al2008 Ricklefs 2010) However a common framework thatencompasses host-range and different forms of geographicspeciation is still a work in progress (Huyse et al 2005)Malarial parasites and other Apicomplexa allow such basic
FIG 5 Median joining network of Plasmodium inui mtDNA haplotypes from macaques (Macaca nemestrina and M fascicularis) from Sabah BorneoBranch lengths are proportional to divergence and node sizes are proportional to total haplotype frequencies Colors correspond to different species ofMacaca blue = M nemestrina and maroon = M fascicularis
430
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
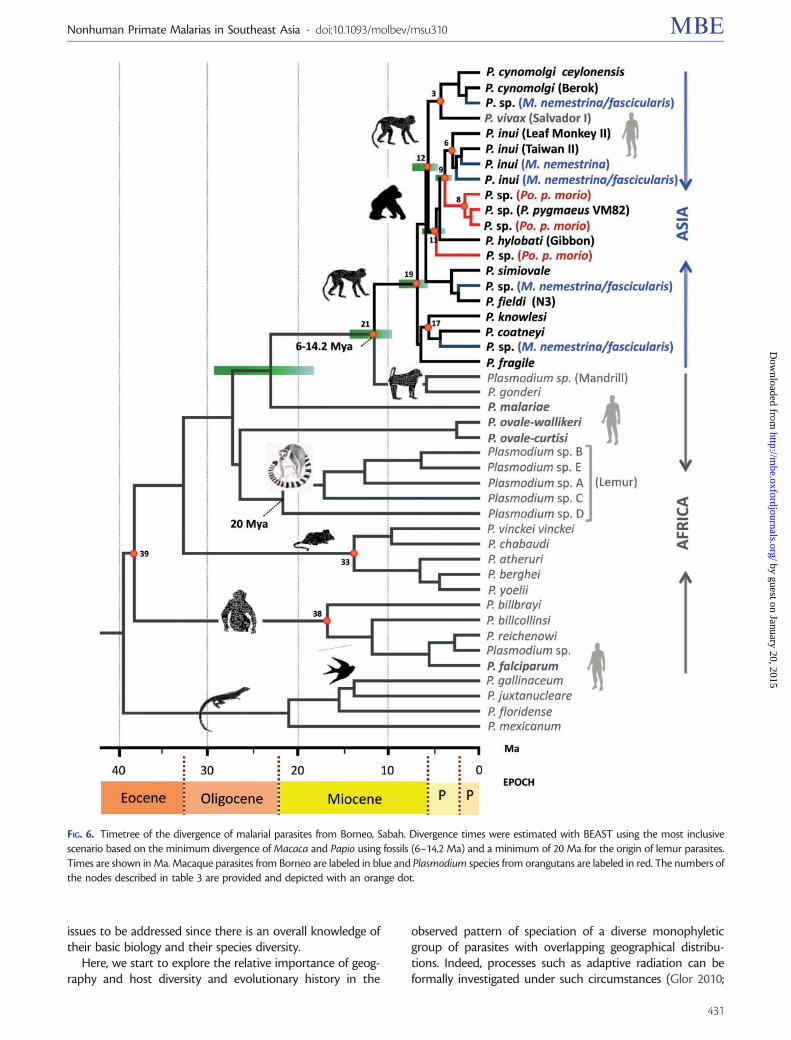
issues to be addressed since there is an overall knowledge oftheir basic biology and their species diversity
Here we start to explore the relative importance of geog-raphy and host diversity and evolutionary history in the
observed pattern of speciation of a diverse monophyleticgroup of parasites with overlapping geographical distribu-tions Indeed processes such as adaptive radiation can beformally investigated under such circumstances (Glor 2010
FIG 6 Timetree of the divergence of malarial parasites from Borneo Sabah Divergence times were estimated with BEAST using the most inclusivescenario based on the minimum divergence of Macaca and Papio using fossils (6ndash142 Ma) and a minimum of 20 Ma for the origin of lemur parasitesTimes are shown in Ma Macaque parasites from Borneo are labeled in blue and Plasmodium species from orangutans are labeled in red The numbers ofthe nodes described in table 3 are provided and depicted with an orange dot
431
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
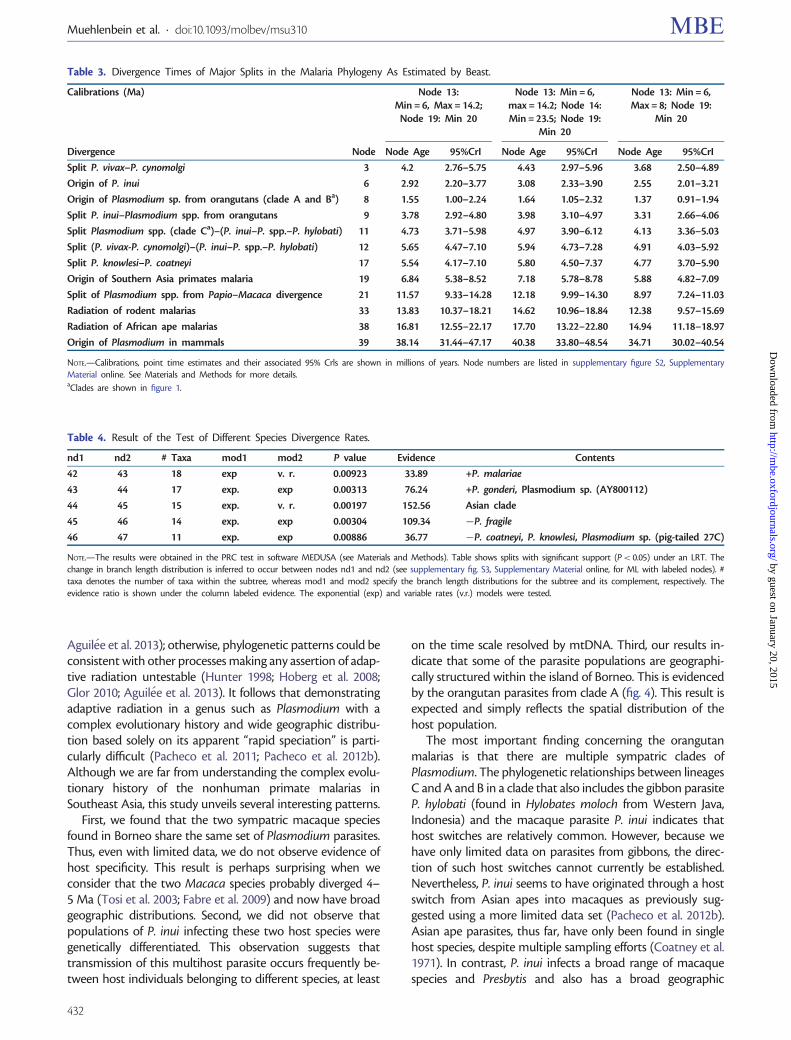
Aguilee et al 2013) otherwise phylogenetic patterns could beconsistent with other processes making any assertion of adap-tive radiation untestable (Hunter 1998 Hoberg et al 2008Glor 2010 Aguilee et al 2013) It follows that demonstratingadaptive radiation in a genus such as Plasmodium with acomplex evolutionary history and wide geographic distribu-tion based solely on its apparent ldquorapid speciationrdquo is parti-cularly difficult (Pacheco et al 2011 Pacheco et al 2012b)Although we are far from understanding the complex evolu-tionary history of the nonhuman primate malarias inSoutheast Asia this study unveils several interesting patterns
First we found that the two sympatric macaque speciesfound in Borneo share the same set of Plasmodium parasitesThus even with limited data we do not observe evidence ofhost specificity This result is perhaps surprising when weconsider that the two Macaca species probably diverged 4ndash5 Ma (Tosi et al 2003 Fabre et al 2009) and now have broadgeographic distributions Second we did not observe thatpopulations of P inui infecting these two host species weregenetically differentiated This observation suggests thattransmission of this multihost parasite occurs frequently be-tween host individuals belonging to different species at least
on the time scale resolved by mtDNA Third our results in-dicate that some of the parasite populations are geographi-cally structured within the island of Borneo This is evidencedby the orangutan parasites from clade A (fig 4) This result isexpected and simply reflects the spatial distribution of thehost population
The most important finding concerning the orangutanmalarias is that there are multiple sympatric clades ofPlasmodium The phylogenetic relationships between lineagesC and A and B in a clade that also includes the gibbon parasiteP hylobati (found in Hylobates moloch from Western JavaIndonesia) and the macaque parasite P inui indicates thathost switches are relatively common However because wehave only limited data on parasites from gibbons the direc-tion of such host switches cannot currently be establishedNevertheless P inui seems to have originated through a hostswitch from Asian apes into macaques as previously sug-gested using a more limited data set (Pacheco et al 2012b)Asian ape parasites thus far have only been found in singlehost species despite multiple sampling efforts (Coatney et al1971) In contrast P inui infects a broad range of macaquespecies and Presbytis and also has a broad geographic
Table 3 Divergence Times of Major Splits in the Malaria Phylogeny As Estimated by Beast
Calibrations (Ma) Node 13Min = 6 Max = 142
Node 19 Min 20
Node 13 Min = 6max = 142 Node 14Min = 235 Node 19
Min 20
Node 13 Min = 6Max = 8 Node 19
Min 20
Divergence Node Node Age 95CrI Node Age 95CrI Node Age 95CrI
Split P vivaxndashP cynomolgi 3 42 276ndash575 443 297ndash596 368 250ndash489
Origin of P inui 6 292 220ndash377 308 233ndash390 255 201ndash321
Origin of Plasmodium sp from orangutans (clade A and Ba) 8 155 100ndash224 164 105ndash232 137 091ndash194
Split P inuindashPlasmodium spp from orangutans 9 378 292ndash480 398 310ndash497 331 266ndash406
Split Plasmodium spp (clade Ca)ndash(P inuindashP sppndashP hylobati) 11 473 371ndash598 497 390ndash612 413 336ndash503
Split (P vivax-P cynomolgi)ndash(P inuindashP sppndashP hylobati) 12 565 447ndash710 594 473ndash728 491 403ndash592
Split P knowlesindashP coatneyi 17 554 417ndash710 580 450ndash737 477 370ndash590
Origin of Southern Asia primates malaria 19 684 538ndash852 718 578ndash878 588 482ndash709
Split of Plasmodium spp from PapiondashMacaca divergence 21 1157 933ndash1428 1218 999ndash1430 897 724ndash1103
Radiation of rodent malarias 33 1383 1037ndash1821 1462 1096ndash1884 1238 957ndash1569
Radiation of African ape malarias 38 1681 1255ndash2217 1770 1322ndash2280 1494 1118ndash1897
Origin of Plasmodium in mammals 39 3814 3144ndash4717 4038 3380ndash4854 3471 3002ndash4054
NOTEmdashCalibrations point time estimates and their associated 95 Crls are shown in millions of years Node numbers are listed in supplementary figure S2 SupplementaryMaterial online See Materials and Methods for more detailsaClades are shown in figure 1
Table 4 Result of the Test of Different Species Divergence Rates
nd1 nd2 Taxa mod1 mod2 P value Evidence Contents
42 43 18 exp v r 000923 3389 +P malariae
43 44 17 exp exp 000313 7624 +P gonderi Plasmodium sp (AY800112)
44 45 15 exp v r 000197 15256 Asian clade
45 46 14 exp exp 000304 10934 P fragile
46 47 11 exp exp 000886 3677 P coatneyi P knowlesi Plasmodium sp (pig-tailed 27C)
NOTEmdashThe results were obtained in the PRC test in software MEDUSA (see Materials and Methods) Table shows splits with significant support (Plt 005) under an LRT Thechange in branch length distribution is inferred to occur between nodes nd1 and nd2 (see supplementary fig S3 Supplementary Material online for ML with labeled nodes) taxa denotes the number of taxa within the subtree whereas mod1 and mod2 specify the branch length distributions for the subtree and its complement respectively Theevidence ratio is shown under the column labeled evidence The exponential (exp) and variable rates (vr) models were tested
432
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

distribution Thus it seems interesting that a parasite cladewith many species with apparently high host specificity inAsian apes has given rise to a generalist parasite with abroad host range
In the case of orangutan malarias the occurrence of suchsympatric lineages within a host could be the result of sym-patric speciation secondary contacts between originally iso-lated host populations with divergent parasite lineages orsecondary acquisition via host switches as has been proposedin avian parasites (Ricklefs 2010) Although we cannot ruleout the possibility of sympatric speciation the life cycle ofmalarial parasites involving sexual reproduction in the mos-quito vector makes it likely that different parasite lineagesinfecting the same host species in the same area will fre-quently mate Under these conditions sympatric speciationis expected to occur only if there is very strong selectionpromoting divergence and reproductive isolation (Gavrilets2004 Giraud et al 2006) For this reason we believe that ourdata are best explained by scenarios involving secondary con-tacts The finding of sympatric divergent lineages in orangu-tans (A and B) is the third case of ldquocanonicallyrdquo sympatricparasite species or subspecies (same host and sympatric in ageographic sense) described in primate malarias where thehost demographic history seems to provide a suitable expla-nation for this pattern (Sutherland et al 2010 Pacheco et al2013) Indeed the two lineages A and B shared their mostrecent common ancestor 1ndash2 Ma a time frame that lieswithin the demographic history of the extant orangutans agenus that evolved under complex climatic and geographicevents that changed the connectivity of its populations(Nieberding et al 2008 Locke et al 2011)
Finally our data suggest that speciation has occurred at anelevated rate within the group of malaria parasites that infectSoutheast Asian nonhuman primates (table 4) This rateseems to be 3ndash4 time faster than in the other groups includedin this study (eg lemur malarias or the Leverania lineagesobserved in African Apes) in a time period of approximately3ndash4 Ma from the origin of the southeast Asia parasite clade tothe split of P inui and the orangutan parasites those eventsalso include the divergence of the lineage leading to thehuman parasite P vivax from its closest relative in macaquesP cynomolgi (table 4) Furthermore we found no evidence ofa significant extinction rate in the sampled species of mam-malian malarias Although this inference is contingent on theassumption that we have sampled all extant Plasmodiumspecies infecting mammalian hosts we obtained qualitativelysimilar results when we repeated the MEDUSA analyses butspecified that the lemur parasite clade contained an addi-tional three species that have not yet been sequencedIndeed there are partial mtDNA sequences that suggest theexistence of three such species (unpublished data) It is alsoimportant to highlight that additional sampling of nonhumanprimates in Southeast Asia may increase the number of spe-cies contained within this clade (eg parasites in gibbons seeCoatney et al 1971) in which case the evidence for acceler-ated branching would be even stronger
In addition to accelerated cladogenesis there are otherlines of evidence needed to determine whether this pattern
is the result of adaptations to new hosts andor environ-ments This aspect is far more complex to address Here wehave found some evidence for episodic selection in two pro-teins involved in the invasion of the red blood cellConsidering the number of proteins that participate in theinvasion of the red blood cell these two antigens only providea first glimpse of the many molecular adaptations that havepresumably emerged in this group of parasites This is alreadyevident when we consider that some parasites like P knowlesiand P coatneyi exhibit antigenic variation while other speciesin this clade do not (Lapp et al 2013) In addition there is anextraordinary diversity of life history traits in this group(Coatney et al 1971) including the preferred type of redblood cell (eg reticulocytes for P cynomolgi whereas P know-lesi invades all including mature erythrocytes) and their pe-riodicity (parasite replication cycle) including a 24 h cycle forP knowlesi (the only one in mammals with such a short cycle)and a 72 h for P inui (a convergent trait with an unrelatedparasite P malariae) among other traits At this stage we canonly speculate that such phenotypic diversity may allow thecoexistence of multiple species by reducing competitionIndeed we observed a relatively high number of mixed infec-tions in this study (table 1) Nevertheless all this phenotypicdiversity and variation at antigen-encoding genes reportedhere (supplementary table S2 and S3 SupplementaryMaterial online) and elsewhere emerged in a relatively shortperiod of 3ndash4 Ma (Pacheco et al 2011 Pacheco et al 2012bRice et al 2014 and this investigation) these facts are consis-tent with but of course do not prove that this relativelyspeciose clade of parasites is the product of an adaptiveradiation
In summary contrary to previous studies the evidencedoes not suggest that host switches have played a centralrole in the diversification of this group of parasites On theone hand we find no evidence of host specialization in themacaque parasites In particular we did not observe differen-tiation of P inui lineages between the two host species nor didwe find parasite species restricted to a single macaque speciesOn the other hand although the orangutan parasites doappear to be host specific phylogenetic analyses suggestthat at least two of these lineages have diversified withinorangutans The two lineages that do appear to have origi-nated via host switches are P inui which emerged from line-ages found in Asian apes (Pacheco et al 2012b) and thelineage leading to P vivax which now infects humans andAfrican apes (Prugnolle et al 2013) We hypothesize that thehost ranges of these parasite species underwent several his-torical changes as result of each host species demographichistory that affected their population densities and spatialdistributions Importantly this notion of a host range evolu-tionary dynamic does not assume that parasites affected theirhostsrsquo speciation or distribution (eg via differential virulenceor other coevolutionary processes) However hostndashparasiteantagonisms could be part of the parasite evolutionarydynamics as suggested by the observation of episodic selec-tion in the antigens included in this investigationAlthough hypothesizing adaptive radiation is tempting inlight of the rapid diversification of host species we are
433
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

unable to draw such a conclusion in this case due to theconfounding effects of biogeographical processes Overall elu-cidating the relative importance of adaptive radiation andbiogeographical processes is a matter of great importanceUnderstanding the evolutionary dynamic of the host rangesin parasite speciation will provide valuable information aboutthe origin of the observed biodiversity of parasites
Materials and Methods
Study Sites
The island of Borneo is considered a biodiversity hotspot(Meijaard and Nijman 2003) Just in the state of Sabah(73618 km2 in the northern portion of Borneo) there aremore than ten species of nonhuman primatesApproximately 11000 Eastern Bornean orangutans (Po pmorio) are found throughout the major lowland forests andfloodplains (Ancrenaz et al 2004) An unknown number ofSundaland pig-tailed macaque (M nemestrina) and commonlong-tailed macaque (M fasicularis) are found almost ubiqui-tously throughout the state The samples were collected intwo locations separated by approximately 300 km in SabahThe Sepilok Orangutan Rehabilitation Centre (SORC) and asatellite facility operated by the Sabah Wildlife Department
The SORC is located 22 km outside of the city of Sandakan(the second largest city in Sabah with a human population of~500000) on the northern edge of the 5529 hectare Kabili-Sepilok Virgin Jungle Forest Reserve bordered by oil palmplantations poultry farms and fruit orchards with at leastsix hotels located within 1 km of the Centre (supplementaryfig S4 Supplementary Material online) Established in 1964 asa sanctuary for the rehabilitation of orphaned injured andorconfiscated orangutans (Po p morio) and other endangeredspecies SORC is operated by the Sabah Wildlife DepartmentFollowing a 6-month quarantine period orangutans aretaught how to transverse the forest and forage for foodFollowing extensive health inspections these animals areeventually relocated or released into the surrounding forestTo facilitate public education and generate operational fundsthe public is allowed to view two daily feedings of the free-ranging animals (Ambu 2007)
The satellite facility operated by the Sabah WildlifeDepartment included in this study is located at theShangri-La Rasa Ria Resort on Pantai Dalit Beach in TuaranIt is approximately 30 km north of Kota Kinabalu surroundedby 400 hectares of tropical forest and contains an OrangutanEducation Centre part of the Rasa Ria Nature Reserve andhome to four orangutans at the time of sampling These an-imals were originally housed at SORC but brought to Rasa Riafor further rehabilitation Like SORC the public is allowed toview two daily feedings of the animals
Sample Collection
As part of routine husbandry practices by the Sabah WildlifeDepartment veterinary staff at the SORC and Rasa Riarsquos med-ical clinics inspected and sampled animals for health statusThis included 34 orangutans (Po p morio 17 female 17 male)ages 16ndash186 years at the SORC Eleven of these animals were
collected from the clinicrsquos indooroutdoor nursery whereasthe remainders were free-ranging animals in the surroundingforest An additional four orangutans (two female and twomale) ages 2ndash4 years were collected and sampled at the RasaRia indooroutdoor nursery For sampling purposes all butone orangutan (the largest male) were manually restrainedfor clinical inspection and sample collection None of theseanimals was injured or febrile at the time
At SORC samples were collected from 15 pig-tailed ma-caques (M nemestrina seven females and eight males alladults) and 26 long-tailed macaques (M fascicularis 10 fe-males and 16 males all adults) These animals were inspectedand sampled after approximately 2 mgkg of Zoletil 100 (zola-zepam-tiletamine Virbac South Africa) was delivered intra-muscularly via syringe dart through a Vario CO2 rifle orblowpipe from Telinject (Saugus California) Respirationand heart rate were monitored while anesthetized For themacaques and orangutans alike blood was collected from thefemoral vein using standard venipuncture technique withsterile single-use Vacutainer productions (Beckton-Dickson) Multiple aliquots of whole blood were stored incryovials frozen at 0 C for 2 weeks before shipment to theUnited States where they were stored at 80 C Aliquots ofwhole blood were also stored in cryovials using RNAlater(500ml of blood mixed with 13 ml RNAlater LifeTechnologies Corporation) frozen at 0 for 2 weeks beforeshipment to the United States where they were stored at80 C All the samples were collected in July 2010 andNovember 2011 No animal was sampled more than once
Samples were imported from Sabah to Indiana Universityusing CDC Public Health Service import permit numbers2011-03-048 and 2013-06-111 and CITES permit numbers12US77006A9 and 9062 Permission to conduct all researchin Sabah was granted by the Sabah Wildlife Department andall animal handling was done by trained staff of the SabahWildlife Department Ethical permission was granted by theBloomington Institutional Animal Care and Use Committeeat Indiana University
DNA Extraction and Malaria Molecular Diagnostics
Genomic DNA was extracted from whole blood usingQIAamp DNA Blood Mini kit (Qiagen GmbH HildenGermany) and each sample was screened for Plasmodiumparasites by nested PCR using primers for a 1200 bp fragmentof the Cytochrome b (cytb) gene that have been used inprevious studies (Pacheco et al 2011 2013 Pacheco et al2012b) The cytb external primers were Forward 50-TGTAAT GCC TAG ACG TAT TCCReverse 50-GT CAA WCAAAC ATG AAT ATA GAC and the internal primers wereForward 50-T CTA TTA ATT TAG YWA AAG CACReverse50-G CTT GGG AGC TGT AAT CAT AAT The primary PCRamplifications were carried out in a 50ml volume reactionusing 20 ng of total genomic DNA 3 mM MgCl2 1 PCRbuffer 125 mM of each deoxynucleoside triphosphate04 mM of each primer and 003 Uml AmpliTaq polymerase(Applied Biosystems Roche-USA) The primary PCR condi-tions were A partial denaturation at 94 C for 4 min and 36
434
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

cycles with 1 min at 94 C 1 min at 53 C and 2 min extensionat 72 C and a final extension of 10 min at 72 C was added inthe last cycle
The nested PCRs were also made in a 50ml volume reac-tion using only 1ml of the primary PCRs 15 mM MgCl21 PCR buffer 125 mM of each deoxynucleoside triphos-phate 04 mM of each primer and 003 Uml AmpliTaq po-lymerase The nested PCR conditions were A partialdenaturation at 94 C for 4 min and 25 cycles with 1 min at94 C 1 min at 56 C and 2 min extension at 72 C and a finalextension of 10 min at 72 C was added in the last cycle Bothstrands for all the cytb fragments were direct sequenced usingan Applied Biosystems 3730 capillary sequencer and identi-fied as Plasmodium using BLAST (Altschul et al 1997)
Molecular Data
Three loci were amplified from malaria positive samples Firstwe amplified approximately 5800 bp of the parasites mito-chondrial genomes (mtDNA) The mtDNA is a linear genomewith three genes encoding Cytochrome B Cox I and Cox III(Pacheco et al 2011) We use the mtDNA genome because nopattern consistent with positive selection has been identifiedit has a consistent A+T content across Plasmodium species(in contrast with the nuclear genome) and there is a relativelyrich database of Plasmodium mitochondrial data that facili-tates phylogenetic studies (Escalante et al 1998 Pacheco et al2011 Pacheco et al 2012b) In addition we sequenced twonuclear genes encoding major antigens AMA-1 and MSP-142These two malarial antigens are essential in the invasion of thehost red blood cell and have been widely studied (Chesne-Seck et al 2005 Pacheco et al 2007) As in the case of themtDNA there is a rich data set on these antigens that facil-itates species identification
The mtDNA was amplified using the oligos Forward 50-GAGGA TTC TCT CCA CAC TTC AAT TCG TAC TTCReverse50-CAG GAA AAT WAT AGA CCG AAC CTT GGA CTC withTaKaRa LA TaqTM Polymerase (TaKaRa Mirus Bio Inc) ThePCR conditions were A partial denaturation at 94 C for1 min and 30 cycles with 30 s at 94 C and 7 min at 68 C afinal extension of 10 min at 72 C was added To detect mixedinfections in all cases at least two independent PCR productswere purified using QIAquick Gel extraction kit (QiagenGmbH) cloned in the pGEM-T Easy Vector systems(Promega USA) and a minimum of four clones and bothstrands were sequenced from each individual
The AMA-1 was amplified by using degenerated primersForward 50-AT GAA TAA AAT ATA CTR CAT AMTReverse50-TC AGT AGT AWG GCT TCT CCA PCR was carried out ina 50ml volume reaction using 20 ng of total genomic DNA25 mM MgCl2 1 PCR buffer 125 mM of each deoxynu-cleoside triphosphate 04 mM of each primer and 003 UmlAmpliTaq Gold polymerase and the PCR conditions were Apartial denaturation at 94 C for 4 min and 36 cycles with1 min at 94 C 1 min at 52 C and 2 min extension at72 C and a final extension of 10 min at 72 C was addedin the last cycle For each sample at least two independentPCR products were also purified from the agarose gel using
QIAquick gel extraction kit (Qiagen GmbH) cloned andboth strands for a minimum of five clones were sequencedfrom each individual
Finally the MSP-142 was amplified by using the primersForward 50-GAC CAA GTA ACA ACG GGA GReverse 50-CAAA GAG TGG CTC AGA ACC in 50ml volume with 20 ngmlof total genomic DNA 25 mM MgCl2 1 PCR buffer125 mM of each deoxynucleoside triphosphate 04 mM ofeach primer and 003 UmM of AmpliTaq Gold DNA poly-merase (Applied Biosystems Roche-USA) The PCR condi-tions were a partial denaturation at 94 C for 4 min and 35cycles of 1 min at 94 C 1 min at 55 C and 2 min extensionat 72 C and a final extension of 10 min was added in the lastcycle Like in the case of the mtDNA at least two independentPCR products per sample were purified from the agarose gelusing QIAquick gel extraction kit (Qiagen GmbH) clonedand both strands were sequenced for a minimum of fiveclones for each sample
Genetic diversity within different Plasmodium specieslin-eages was quantified by calculating the parameter (nucle-otide diversity) for the mtDNA and the two antigens (AMA-1and MSP-142) This statistic estimates the average number ofsubstitutions between any two sequences as implemented inMEGA v522 (Tamura et al 2011) Standard errors of esti-mates were calculated with 1000 bootstrap replicates
The sequences reported in this investigation were submit-ted to GenBank under the accession numbers KJ569800 toKJ569917 (see supplementary table S1 SupplementaryMaterial online for details)
Phylogenetic Analyses
Independent alignments for nucleotide sequences of themtDNA (5717 bp excluding gaps) ama-1 (1663 bp excludinggaps) and msp-142 (1057 bp excluding gaps) genes weremade using ClustalX v2012 and Muscle as implemented inSeaView v435 with manual editing First phylogenetic rela-tionships were inferred for the mtDNA using both the MLmethod implemented in PhyML v30 (Guindon and Gascuel2003) and Bayesian methods using MrBayes v312 with thedefault priors (Ronquist and Huelsenbeck 2003) The reliabil-ity of the nodes in the ML tree was assessed by the bootstrapmethod with 200 pseudoreplications In the Bayesian analysiseach of the three mitochondrial genes plus the noncodingregions were used as separate partitions (Pacheco et al 2011)Bayesian support for the nodes was inferred in MrBayes bysampling every 100 generations from two independent chainslasting 8 106 Markov Chain Monte Carlo (MCMC) stepsThe chains were assumed to have converged once the averagestandard deviation of the posterior probability was below001 and the value of the potential scale reduction factorwas between 100 and 102 (Ronquist and Huelsenbeck2003) Fifty percent of the sample was then discarded as aburn-in once convergence was reached Both phylogeneticmethods used a general time reversible model withgamma-distributed substitution rates and a proportion ofinvariant sites (GTR +G+ I) and indeed this model had thelowest Bayesian Information Criterion scores as estimated by
435
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

MEGA v522 (Nei and Kumar 2000 Tamura et al 2011) Inorder to compare the different Plasmodium species found inthis study we estimated the genetic divergences of themtDNA genomes among and within different species usingthe Kimura 2-parameter model as implemented in MEGAv522 (Tamura et al 2011)
Phylogenetic relationships were also inferred for the ama-1and msp-142 kDa genes using both the ML method imple-mented in PhyML v30 and Bayesian methods with the defaultpriors (Ronquist and 2003) In the case of the Bayesian infer-ence two independent chains were sampled every 100 gen-erations in runs lasting 6 106 MCMC steps for ama-1 geneand 3 106 for msp-142 As before 50 of the sample wasdiscarded as a burn-in period once the chain had achievedconvergence We used a general time reversible model withgamma-distributed substitution rates (GTR +G) for bothgenes because it best fit the data as estimated by MEGAv522 (Nei and Kumar 2000 Tamura et al 2011)
Supplementary table S1 Supplementary Material onlineprovides a complete list of the species and sequences ob-tained in this study it also includes those previously publishedthat were used as part of our phylogenetic analyses Becausethe same Plasmodium species (almost identical mtDNA withthe exception of few SNPs) were found in both species ofMacaca we only amplified the genes encoding the two majorantigens (ama-1 and msp-142) from the 15 positive samples ofpig-tailed macaques (M nemestrina) Information about basicbiology geographic distribution and host-range of these spe-cies can be found elsewhere (Coatney et al 1971)
Haplotype Networks
The median joining network for a complete set of 41Plasmodium sp mtDNA haplotypes from orangutans wasestimated using Network v4610 (Fluxus Technologies2011) with transversions weighted twice as large as transi-tions and the epsilon parameter set equal to 0 In this analysiswe included all 30 Plasmodium sp mtDNA haplotypes ob-tained from orangutan in this study (Sabah) and also 11mtDNA haplotypes from Kalimantan orangutans that havebeen reported in somewhere else (Pacheco et al 2012b) Alsowe estimated a median joining network for a complete set of16 P inui mtDNA haplotypes from both species of Macaca(eight haplotypes from M nemestrina and eight from Mfascicularis) as was described before In addition we alsoused DnaSP v5 (Librado and Rozas 2009) to estimate thefixation index (FST) between the two populations (Sabahand Kalimantan Borneo) of Plasmodium sp from orangu-tan-clade A (for more details about the clades see theResults section) and between the two populations of P inuifrom M nemestrina and M fascicularis (Sabah)
Signatures Consistent with Natural Selection
Although we cannot attest that msp-1 or ama-1 genes areparticularly important in parasite speciation their role in theinvasion of the red blood cell could indicate adaptation to anew host We use a branch-site method or REL (Pond et al2011) as implemented in HyPhy (Pond et al 2005) This
approach aims to identify lineages where a proportion ofcodons evolve with 41 by using an LRT This methoddoes not make assumptions about which lineages areunder selection so it is ideal for this type of exploratory anal-yses Because these methods assume the true phylogeny weused the consensus phylogeny derived from a Bayesian anal-ysis that only included lineages that we consider ldquogood spe-ciesrdquo or clearly supported lineages (10 posterior probabilityon both antigens) Although many of these lineages are ac-cepted species others have just been proposed in thisinvestigation
Estimation of Divergence Times
BEAST v175 (Drummond et al 2012) was used to estimatetime trees for the mtDNA sequences (supplementary tableS1 Supplementary Material online) using four partitions ofthe data each gene (cox1 cox3 cytb) plus the noncodingregions (Pacheco et al 2011 2013 Pacheco et al 2012b)Relaxed clock methods were applied with a general time re-versible model with gamma-distributed substitution ratesand a proportion of invariant sites (GTR +G+ I) with het-erogeneity both among sites and across the four partitionsTwo independent chains were run for each analysis untilconvergence appeared to have been achieved Three scenar-ios were explored each of them incorporating different cali-bration points two based on host fossils and the third basedon a biogeographical landmark for the host
The first calibration point assumes that African parasitesfound in Mandrillus spp and Cercocebus spp diverged fromthose Plasmodium spp found in Southeast Asia macaqueswhen Macaca branched from Papio (Mu et al 2005Hayakawa et al 2008 Pacheco et al 2011) Fossils identifiedas Macaca spp indicate that such an event took place 6ndash8 Maas minimum and maximum boundaries respectively (Delson1980) These time points are used as calibration with an ex-ponential prior assuming that the divergence of the Africanand Asian parasites took place with 95 of probability be-tween 6 and 8 Ma (Mu et al 2005) However these fossils areused as minimum times when studying primates (Perelmanet al 2011) A more inclusive alternative calibration was con-structed around this event assuming that the divergence atthe same node took place at any time between 6 and 142 Mausing a uniform prior this incorporates molecular estimatesfor the same PapiondashMacaca divergence event (Pacheco et al2011) The use of 142 Ma as a maximum is consistent witholder fossils reported for Macaca spp (see PaleobiologyDatabase at httpwwwpaleodborg last accessedNovember 17 2014) and considers also the fact that P gonderiis a parasite of Chlorocebus (a Cercopithecini) In either casethese calibrations assume that the parasite lineages did notdiverge before their host taxa had diverged giving an upperbound of either 142 or 8 Ma They also assume that theparasite lineages did not diverge after the host taxa hadbecome isolated on different continents that is parasite di-versification did not involve intercontinental dispersal sepa-rate from that of the host taxa giving a lower bound of 6 Ma
436
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

A second fossil-based calibration is the minimum of 235Ma for the humanMacaca split (Benton and Donoghue2007) which is assumed to be the time when the monophy-letic group that includes P malariae (a parasite found inhumans) and all the Asian parasites originated This event isused as calibration with an exponential prior set conserva-tively with 975 of the prior probability below to 65 Ma so itallows for the clade to be as old as the origin of primates oreven older if the data support that (Perelman et al 2011Springer et al 2012) In addition to these calibrations thatincorporate information from fossils an additional calibrationpoint considers the origin of lemur malarias (a biogeograph-ical event) a monophyletic group that radiated inMadagascar (Pacheco et al 2011) The colonization ofMadagascar by terrestrial mammals including lemurs hap-pened during a relatively short time frame in the Cenozoic(~6520 Ma) during which ocean currents allowed for theeastward transport of vegetation rafts from Africa toMadagascar (Poux et al 2005 Ali and Huber 2010) Thustimes younger than approximately 20 Ma are unlikely to bebiologically realistic for the origin of lemur malarias (Pachecoet al 2011) This event was used as calibration with an expo-nential prior set conservatively with 975 of the prior prob-ability below to 65 Ma (Perelman et al 2011 Springer et al2012)
The three scenarios considered in this investigation utilizethe calibrations described above as follow 1) a combinationof the relaxed 6ndash142 Ma calibration for PapiondashMacaca diver-gence (Delson 1980 Mu et al 2005 Hayakawa et al 2008)with a minimum of 20 Ma for the origin of lemur parasites(Poux et al 2005 Ali and Huber 2010) 2) a combination of the6ndash142 Ma calibration with a minimum of 235 Ma for thehumanMacaca split and 20 Ma for the origin of the lemurparasites and 3) the 6ndash8 Ma narrowly defined around thefossils of the PapiondashMacaca divergence (Delson 1980) with aminimum of 20 Ma for the origin of the lemur lineage (seePacheco et al 2011 2013 Pacheco et al 2012b) Although thetime estimates under these scenarios are expected to overlaptheir comparison is important since the calibrations rely ondifferent assumptions (Pacheco et al 2011 Ramiro et al2012) An extensive discussion on different timing methodsand calibrations was reported elsewhere (Pacheco et al 2011)
Species Diversification Rates
Two methods were used to test whether the branching rate(cladogenesis) has changed across the Plasmodium mtDNAphylogeny In both cases we retained only those 38 lineagesthought to correspond to good species while excluding clo-sely related haplotypes that appear to have been sampledfrom within species We first carried out the nonparametriccladogenesis test (Nater et al 1992) which looks for evidenceof accelerated branching within a phylogeny This was appliedto a ML tree inferred for these 38 taxa We then used theprogram MEDUSA (Alfaro et al 2009) to analyze a sample of50 trees from a Bayesian phylogenetic analysis using an inho-mogeneous birthndashdeath model Given a rooted treeMEDUSA fits a sequence of increasingly complicated birthndash
death models to that tree and uses a stepwise procedureusing a corrected AICc to select the best-fitting model Themodels differ in the number of components into which thetree is partitioned with each component being assigned itsown birth and death rates In our analyses we consideredmodels with up to ten components The trees used in thisanalysis were generated by two independent runs of BEASTv175 using the same prior distributions and calibrationpoints as previously described with one tree being sampledevery million generations from generations 20ndash50 million inthe first chain and from generations 32ndash50 million in thesecond chain Monophyly of the parasites infecting mamma-lian hosts was enforced in these analyses and the clade con-taining four species with avian or reptilian hosts was prunedfrom each tree prior to analysis with MEDUSA
Supplementary MaterialSupplementary tables S1ndashS4 and figures S1ndashS4 are available atMolecular Biology and Evolution online (httpwwwmbeoxfordjournalsorg)
Acknowledgments
Permission to conduct this research in Malaysia was grantedby the Sabah Wildlife Department (SWD) Marc AncrenazBenoit Goossens Sail Jamaludin Timothy Wright the SabahWildlife Rescue Team and the Sabah Wildlife Advisory Panelprovided valuable assistance The authors thank all the mem-bers of the Sabah Wildlife Department the SORC and theNature Reserve at Shangri-La Rasa Ria The authors also thankthe DNA laboratory at the School of Life Sciences for theirtechnical support and Scott Hetrick for producing the mapsof Sabah This work was supported by the US NationalInstitutes of Health (R01 GM080586 to AAE) The fieldwork was supported by Indiana University (FacultyResearch Support Program to MPM) and is part of thelarger Sabah Ecosystem Health Project began by MPMand colleagues in 2005 The content is solely the responsibilityof the authors and does not represent the official views of theNIH
ReferencesAguilee R Claessen D Lambert A 2013 Adaptive radiation driven by the
interplay of eco-evolutionary and landscape dynamics Evolution 671291ndash1306
Ali JR Huber M 2010 Mammalian biodiversity on Madagascar con-trolled by ocean currents Nature 463653ndash656
Alfaro ME Santini F Brock C Alamillo H Dornburg A Rabosky DLCarnevale G Harmon LJ 2009 Nine exceptional radiations plus highturnover explain species diversity in jawed vertebrates Proc NatlAcad Sci U S A 10613410ndash13414
Altschul SF Madden TL Scheuroaffer AA Zhang J Zhang Z Miller WLipman DJ 1997 Gapped BLAST and PSI-BLAST a new generationof protein database search programs Nucleic Acids Res 1253389ndash3402
Ambu L 2007 Strategy of the Sabah Wildlife Department for wildlifeconservation in Sabah First International Conservation Conferencein Sabah the Quest for Gold Standards Kota Kinabalu MalaysiaSabah Wildlife Department
Ancrenaz M Gimenez O Ambu L Andau P Goossens B Payne JSawang A Tuuga A Lackman-Ancrenaz I 2004 Aerial surveysgive new estimates for orangutans in Sabah Malaysia PLoS Biol 3e3
437
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

Ayala FJ Escalante AA Lal AA Rich SM 1998 Evolutionary relationshipsof human malaria parasites In Sherman IW editor Malaria biologypathogenesis and protection Washington (DC) ASM Pressp 285ndash300
Benton MJ Donoghue PC 2007 Paleontological evidence to date thetree of life Mol Biol Evol 2426ndash53
Brooks DR Leon-Regagnon V McLennan DA Zelmer D 2006 Ecologicalfitting as a determinant of the community structure of platyhel-minth parasites of anurans Ecology 87S76ndashS85
Brooks DR McLennan DA 1993 Comparative study of adaptive radia-tions with an example using parasitic flatworms (platyhelminthescercomeria) Am Nat 142755ndash778
Chesne-Seck ML Pizarro JC Vulliez-Le Normand B Collins CR BlackmanMJ Faber BW Remarque EJ Kocken CH Thomas AW Bentley GA2005 Structural comparison of apical membrane antigen 1 ortho-logues and paralogues in apicomplexan parasites Mol BiochemParasitol 14455ndash67
Coatney RG Collins WE Warren M Contacos PG 1971 The primatemalarias Washington (DC) US Government Printing Office
Cox-Singh J Singh B 2008 Knowlesi malaria newly emergent and ofpublic health importance Trends Parasitol 24406ndash410
Delson E 1980 Fossil macaques phyletic relationships and a scenario ofdeployment In Lindburg DG editor The macaquesmdashstudies inecology behavior and evolution New York Van NostrandReinhold Co p 10ndash29
Drummond AJ Suchard MA Xie D Rambaut A 2012 Bayesian phylo-genetics with BEAUti and the BEAST 17 Mol Biol Evol 291969ndash1973
Escalante AA Cornejo OE Freeland DE Poe AC Durrego E Collins WELal AA 2005 A monkeyrsquos tale the origin of Plasmodium vivax as ahuman malaria parasite Proc Natl Acad Sci U S A 1021980ndash1985
Escalante AA Lal AA Ayala FJ 1998 Genetic polymorphism and naturalselection in the malaria parasite Plasmodium falciparum Genetics149189ndash202
Fabre PH Rodrigues A Douzery EJ 2009 Patterns of macroevolutionamong Primates inferred from a supermatrix of mitochondrial andnuclear DNA Mol Phylogenet Evol 53808ndash825
Fooden J 1994 Malaria in macaques Int J Primatol 15573ndash596Garnham PCC 1966 Malaria parasites and other haemosporidia
Oxford Blackwell Scientific PublicationsGavrilets S 2004 Fitness landscapes and the origin of species
Monographs in population biology Princeton (NJ) PrincetonUniversity Press
Giraud T Villareal LMMA Austerlitz F Le Gac M Lavigne C 2006Importance of the life cycle in sympatric host race formation andspeciation of pathogens Phytopathology 96280ndash287
Glor RE 2010 Phylogenetic insights on adaptive radiation Annu Rev EcolEvol Syst 41251ndash270
Guindon S Gascuel O 2003 A simple fast and accurate algorithm toestimate large phylogenies by maximum likelihood Syst Biol 52696ndash704
Hayakawa T Culleton R Otani H Horii T Tanabe K 2008 Big bang in theevolution of extant malaria parasites Mol Biol Evol 252233ndash2239
Hoberg Eric P Brooks Daniel R 2008 A macroevolutionarymosaic episodic host-switching geographical colonization and di-versification in complex host-parasite systems J Biogeograph 351533ndash1550
Hunter JP 1998 Key innovations and the ecology of macroevolutionTrends Ecol Evol 1331ndash36
Huyse T Poulin R Theron A 2005 Speciation in parasites a populationgenetics approach Trends Parasitol 21469ndash475
Krief S Escalante AA Pacheco MA Mugisha L Andre C Halbwax MFischer A Krief JM Kasenene JM Crandfield M et al 2010 On thediversity of malaria parasites in African apes and the origin ofPlasmodium falciparum from Bonobos PLoS Pathog 6e1000765
Lapp SA Korir-Morrison C Jiang J Bai Y Corredor V Galinski MR 2013Spleen-dependent regulation of antigenic variation in malaria para-sites Plasmodium knowlesi SICAvar expression profiles in splenicand asplenic hosts PLoS One 8(10)e78014
Librado P Rozas J 2009 DnaSP v5 a software for comprehensive analysisof DNA polymorphism data Bioinformatics 251451ndash1452
Liu W Li Y Learn GH Rudicell RS Robertson JD Keele BF Ndjango JBSanz CM Morgan DB Locatelli S et al 2010 Origin of the humanmalaria parasite Plasmodium falciparum in gorillas Nature 467420ndash425
Locke DP Hillier LW Warren WC Worley KC Nazareth LV Muzny DMYang SP Wang Z Chinwalla AT Minx P et al 2011 Comparativeand demographic analysis of orang-utan genomes Nature 469529ndash533
Ma X Kelley JL Eilertson K Musharoff S Degenhardt JD Martins ALVinar T Kosiol C Siepel A Gutenkunst RN et al 2013 Populationgenomic analysis reveals a rich speciation and demographic historyof orang-utans (Pongo pygmaeus and Pongo abelii) PLoS One8(10)e77175
Meijaard E Nijman V 2003 Primate hotspots on borneo predictivevalue for general biodiversity and the effects of taxonomy ConservBiol 17725ndash732
Mu J Joy DA Duan J Huang Y Carlton J Walker J Barnwell J Beerli PCharleston MA Pybus OG et al 2005 Host switch leads to emer-gence of Plasmodium vivax malaria in humans Mol Biol Evol 221686ndash1693
Nater A Nietlisbach P Arora N van Schaik CP van Noordwijk MAWillems EP Singleton I Wich SA Goossens B Warren KS et al 1992Tempo and mode of evolution revealed from molecular phyloge-nies Proc Natl Acad Sci U S A 898322ndash8326
Nei M Kumar S 2000 Molecular evolution and phylogenetics NewYork Oxford University Press
Nieberding CM Durette-Desset MC Vanderpoorten A Casanova JCRibas A Deffontaine V Feliu C Morand S Libois R Michaux JR2008 Geography and host biogeography matter for understand-ing the phylogeography of a parasite Mol Phylogenet Evol 47538ndash554
Nunn CL Altizer S 2006 Infectious diseases in primates behaviorecology and evolution New York Oxford University Press
Pacheco MA Battistuzzi FU Junge RE Cornejo OE Williams CV LandauI Rabetafika L Snounou G Jones-Engel L Escalante AA 2011Timing the origin of human malarias the lemur puzzle BMC EvolBiol 11299
Pacheco MA Cranfield M Cameron K Escalante AA 2013 Malarialparasite diversity in chimpanzees the value of comparativeapproaches to ascertain the evolution of Plasmodium falciparumantigens Malar J 12(1)328
Pacheco MA Elango AP Rahman AA Fisher D Collins WE Barnwell JWEscalante AA 2012a Evidence of purifying selection on merozoitesurface protein 8 (MSP8) and 10 (MSP10) in Plasmodium spp InfectGenet Evol 12978ndash986
Pacheco MA Poe AC Collins WE Lal AA Tanabe K Kariuki SKUdhayakumar V Escalante AA 2007 A comparative study of thegenetic diversity of the 42kDa fragment of the merozoite surfaceprotein 1 in Plasmodium falciparum and P vivax Infect Genet Evol 7180ndash187
Pacheco MA Reid MJC Schillaci MA Lowenberger CA Galdikas BMFJones-Engel L Escalante AA 2012b The origin of malarial parasites inorangutans PLoS One 7e34990
Pacheco MA Ryan EM Poe AC Basco L Udhayakumar V Collins WEEscalante AA 2010 Evidence for negative selection on the geneencoding rhoptry-associated protein 1 (RAP-1) in Plasmodiumspp Infect Genet Evol 10655ndash661
Page RDM 2003 Tangled trees phylogeny cospeciation and coevolu-tion Chicago The University of Chicago Press
Perelman P Johnson WE Roos C Seuanez HN Horvath JE Moreira MAKessing B Pontius J Roelke M Rumpler Y et al 2011 A molecularphylogeny of living primates PLoS Genet 7e1001342
Peters W Garnham PCC Killick-Kendrick R Rajapaksa N Cheong WHCadigan FC 1976 Malaria of the orang-utan (Pongo pygmaeus) inBorneo Philos Trans R Soc Lond B Biol Sci 275439ndash482
Pond SL Frost SD Muse SV 2005 HyPhy hypothesis testing usingphylogenies Bioinformatics 21676ndash679
438
Muehlenbein et al doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

Pond SL Murrell B Fourment M Frost SD Delport W Scheffler K 2011A random effects branch-site model for detecting episodic diversi-fying selection Mol Biol Evol 283033ndash3043
Poulin R 2005 Detection of interspecific competition in parasite com-munities J Parasitol 911232ndash1235
Poulin R Krasnov BR Mouillot D Thieltges DW 2011 The comparativeecology and biogeography of parasites Philos Trans R Soc Lond B BiolSci 3662379ndash2390
Poux C Madsen O Marquard E Vieites DR de Jong WW Vences M2005 Asynchronous colonization of Madagascar by the four en-demic clades of primates tenrecs carnivores and rodents as inferredfrom nuclear genes Syst Biol 54719ndash730
Prugnolle F Rougeron V Becquart P Berry A Makanga B Rahola NArnathau C Ngoubangoye B Menard S Willaume E et al 2013Diversity host switching and evolution of Plasmodium vivaxinfecting African great apes Proc Natl Acad Sci U S A 1108123ndash8128
Ramiro RS Reece SE Obbard DJ 2012 Molecular evolution and phylo-genetics of rodent malaria parasites BMC Evol Biol 12219
Rice BL Acosta MM Pacheco MA Carlton JM Barnwell JW EscalanteAA 2014 The origin and diversification of the merozoite surfaceprotein 3 (msp3) multi-gene family in Plasmodium vivax and relatedparasites Mol Phylogenet Evol 78C172ndash184
Ricklefs RE 2010 Host-pathogen coevolution secondary sympatry andspecies diversification Philos Trans R Soc Lond B Biol Sci 3651139ndash1147
Ronquist F Huelsenbeck JP 2003 MrBayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 191572ndash1574
Singh B Kim Sung L Matusop A Radhakrishnan A Shamsul SS Cox-Singh J Thomas A Conway DJ 2004 A large focus of naturally
acquired Plasmodium knowlesi infections in human beings Lancet3631017ndash1024
Springer MS Meredith RW Gatesy J Emerling CA Park J Rabosky DLStadler T Steiner C Ryder OA Janecka JE et al 2012Macroevolutionary dynamics and historical biogeography of pri-mate diversification inferred from a species supermatrix PLoS One7e49521
Sutherland CJ Tanomsing N Nolder D Oguike M Jennison CPukrittayakamee S Dolecek C Hien TT do Rosario VE Arez APet al 2010 Two nonrecombining sympatric forms of the humanmalaria parasite Plasmodium ovale occur globally J Infect Dis 2011544ndash1550
Ta TH Hisam S Lanza M Jiram AI Ismail N Rubio JM1 2014 First caseof a naturally acquired human infection with Plasmodium cyno-molgi Malar J 13(1)68
Tachibana S Sullivan SA Kawai S Nakamura S Kim HR Goto N ArisueN Palacpac NM Honma H Yagi M et al 2012 Plasmodium cyno-molgi genome sequences provide insight into Plasmodium vivax andthe monkey malaria clade Nat Genet 441051ndash1055
Tamura K Peterson D Peterson N Stecher G Nei M Kumar S 2011MEGA5 molecular evolutionary genetics analysis using maximumlikelihood evolutionary distance and maximum parsimony meth-ods Mol Biol Evol 282731ndash2739
Telford SR Jr 2009 Hemoparasites of the reptilian Boca Raton (FL) CRCPress
Tosi AJ Disotell TR Morales JC Melnick DJ 2003 Cercopithecine Y-chromosome data provide a test of competing morphological evo-lutionary hypotheses Mol Phylogenet Evol 27510ndash521
Valkiunas G 2005 Avian malaria parasites and other haemosporidiaBoca Raton (FL) CRC
439
Nonhuman Primate Malarias in Southeast Asia doi101093molbevmsu310 MBE by guest on January 20 2015
httpmbeoxfordjournalsorg
Dow
nloaded from
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
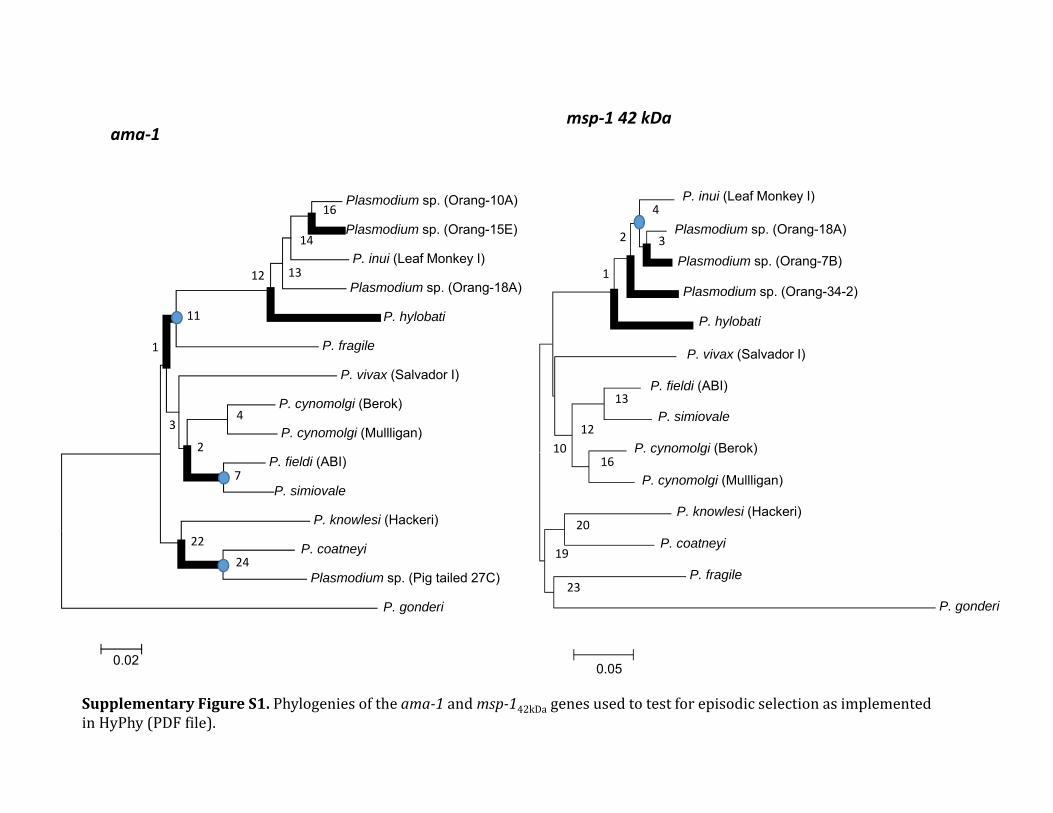
Plasmodium sp (Orang-10A)
Plasmodium sp (Orang-15E)
P inui (Leaf Monkey I)
Plasmodium sp (Orang-18A)
P hylobati
P fragile
P vivax (Salvador I)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P knowlesi (Hackeri)
P coatneyi
Plasmodium sp (Pig tailed 27C)
P gonderi
002
43
7
2
1
11
12
16
14
24
22
13
ama‐1
P coatneyi
P fragile
P gonderi
005
P knowlesi (Hackeri)
P cynomolgi (Berok)
P cynomolgi (Mullligan)
P fieldi (ABI)
P simiovale
P vivax (Salvador I)
P hylobati
Plasmodium sp (Orang-34-2)
Plasmodium sp (Orang-7B)
Plasmodium sp (Orang-18A)
P inui (Leaf Monkey I)4
32
1
13
12
1610
20
23
19
msp‐1 42 kDa
SupplementaryFigureS1Phylogeniesoftheama‐1 andmsp‐142kDa genesusedtotestforepisodicselectionasimplementedinHyPhy (PDFfile)
P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-

P vinckei vinckei
P cynomolgi (Berok)P cynomolgi ceylonensis
P vivax (Salvador I)
P inui (Taiwan II)P inui (Leaf Monkey II)
P hylobati (Gibbon)
P simiovale
P fieldi (N3)
P coatneyiP knowlesi
P fragile
P gonderi
P ovale‐wallikeriP ovale‐curtisi
P bergheiP yoelii
Plasmodium sp (Mandrill)
P malariae
Plasmodium sp D (Lemur)Plasmodium sp C (Lemur)Plasmodium sp A (Lemur)
Plasmodium sp B (Lemur)Plasmodium sp E (Lemur)
P atheruriP chabaudi
P billbrayiP billcollinsi
Plasmodium spP reichenowi
P falciparum
P mexicanumP floridense
P gallinaceumP juxtanucleare
P sp (M nemestrinafascicularis)
P sp (M nemestrina)P sp (M nemestrinafascicularis)P sp (P pygmaeus)P sp (P pygmaeus VM82)
P sp (P pygmaeus)
P sp (M nemestrinafascicularis)
P sp (M nemestrinafascicularis)
6‐142 Mya
20 Mya
235 Mya
SupplementaryFigureS2BEASTnodenumbersforthePlasmodium phylogenyasusedinTable6
12
3
45
6
78
9
10
11
12
1314
15
1617
18
19
20
21
22
23
2425
2627
28
29
3031
3233
34
3536
3738
39
4041
42
43
P sp (P pygmaeus)
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
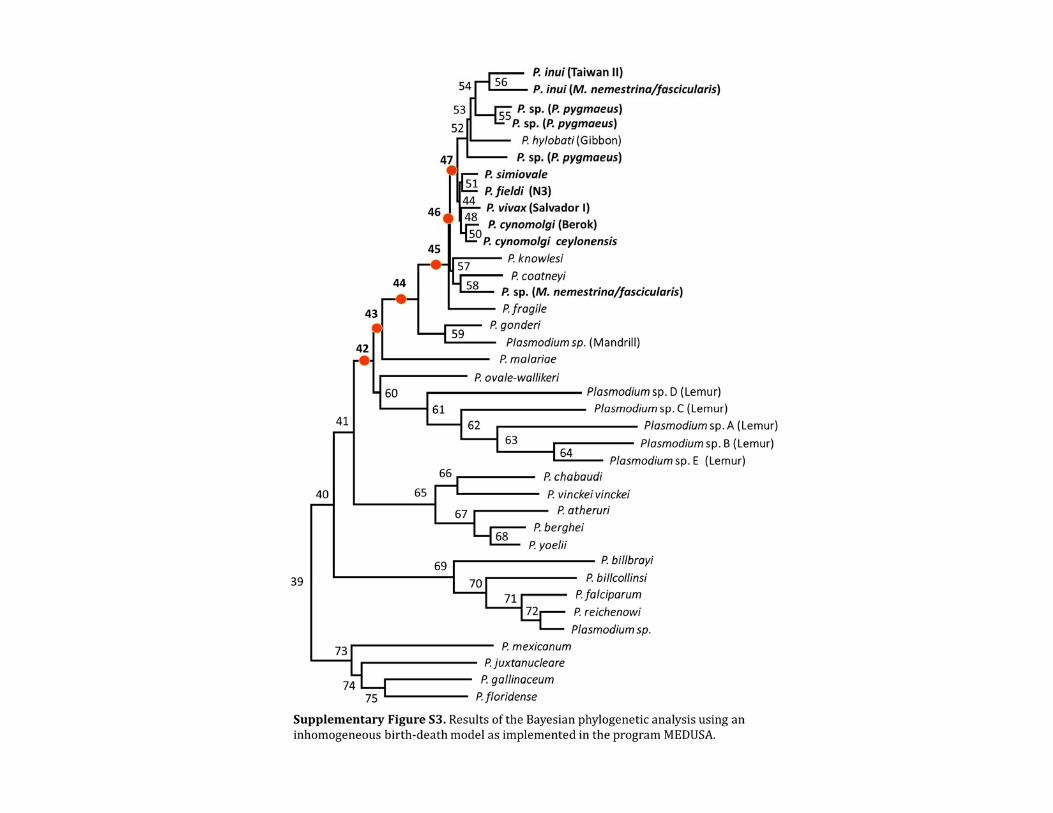
Supplementary Table S2 Genetic polymorphism observed in the ama1 gene in different Plasmodium species
ama1 π (SE)
P inui (n=10) CDS 00220 (00022) Domain I 00192 (00026) Domain II+III 00250 (00030) Plasmodium spp (Orangutan) Clade 1 (n=3) CDS 00020 (00009) Domain I 00016 (00011) Domain II+III 00025 (00015) Clade 2 (n=3) CDS 00231 (00029) Domain I 00269 (00045) Domain II+III 00190 (00035) Clade 1 + 2 (n=6) CDS 00370 (00034) Domain I 00412 (00049) Domain II+III 00327 (00044) P cynomolgi (n=5) CDS 00338 (00033) Domain I 00262 (00039) Domain II+III 00421 (00047)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
Supplementary Table S3 Genetic polymorphism observed in the msp‐142 gene in different Plasmodium species
Msp‐142 kDa π (SE)
P inui (n=19) 42 kDa 00228 (00021) 33 kDa 00274 (00028) 19 kDa 00076 (00028) Plasmodium spp (Orangutan) Clade 1 (n=5) 42 kDa 00212 (00028) 33 kDa 00241 (00034) 19 kDa 00111 (00043) Clade 2 (n=17) 42 kDa 00345 (00038) 33 kDa 00412 (00043) 19 kDa 00112 (00039) Clade 3 (n=3) 42 kDa 00420 (00046) 33 kDa 00503 (00060) 19 kDa 00132 (00060) Clade 1 + 2 (n=22) 42 kDa 00388 (00036) 33 kDa 00462 (00045) 19 kDa 00112 (00040) P cynomolgi (n=10) 42 kDa 00373 (00037) 33 kDa 00408 (00043) 19 kDa 00251 (00063)
Clade numbers are shown in Figure 3 and do not necessarily correspond to clades A B and C (Fig 1) π is the average nucleotide diversity and SE is Standard Error For more details see Result section
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
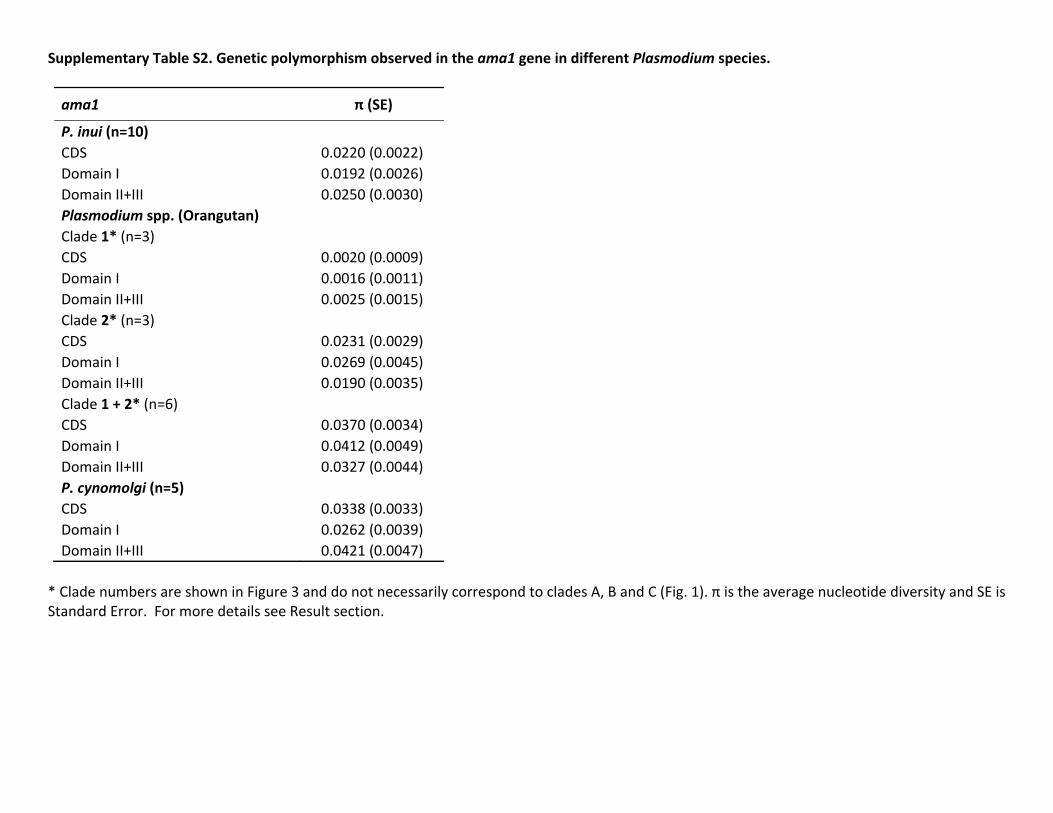
Supplementary Table S4 Inferred branch‐specific distributions of site‐wise ω and the Likelihood Ratio Test (LRT) test results for evidence of
episodic diversifying selection (EDS) Branches are sorted in the order of decreasing level of support for EDS See Supplementary Figure S1 for branch
names and nodes Mean ω the ω ratio inferred under the MG94xREV model which permits lineage‐to‐lineage but no site‐to‐site ω variation ω‐ the
maximum likelihood estimate (MLE) of the first rate class with ω le 1 Pr[ω=ω‐] the MLE of the proportion of sites evolving at ω‐ ωN the MLE of the
second rate class with ω le 1 Pr[ω=ωN] the MLE of the proportion of sites evolving at ωN ω+ the MLE of the rate class with unconstrained ω ge 1 an
indicates that the estimate (gt10000) is not reliable due to high variance (eg no synonymous substitutions at those sites) Pr[ω=ω+] the MLE of
the proportion of sites evolving at ω+ LRT Likelihood Ratio Test statistic for ω+ = 1 (null) versus ω+ unrestricted (alternative) p‐value the uncorrected
p‐value for the LRT test and Corrected p‐value the p‐value corrected for multiple testing using the Holm‐Bonferroni method
Branch Mean ω ω‐ Pr[ω=ω‐] ωN Pr[ω=ωN] ω+ Pr[ω=ω+] LRT p‐value Corrected
p‐value
ama‐1
Node 24 054 032 096 1 002 6463 002 1617 lt00001 0 Node 7 106 032 095 042 0 2434 005 1245 0 001 P hylobati 065 044 097 041 0 2338 003 1156 0 001 P sp (Orang‐15E) 019 011 098 011 001 gt10000 001 1155 0 001 Node 11 10 0 078 lt00001 0 54645 022 858 0 004
msp‐1
Node 4 432 1 083 1 015 gt10000 002 184 lt00001 0 P sp (Orang‐7B) 052 0 092 0 001 159 007 1479 lt00001 0 P hylobati 05 043 098 043 0 gt10000 002 1046 0 001 P sp (Orang‐34‐2) 045 0 087 0 002 649 011 1033 0 001
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
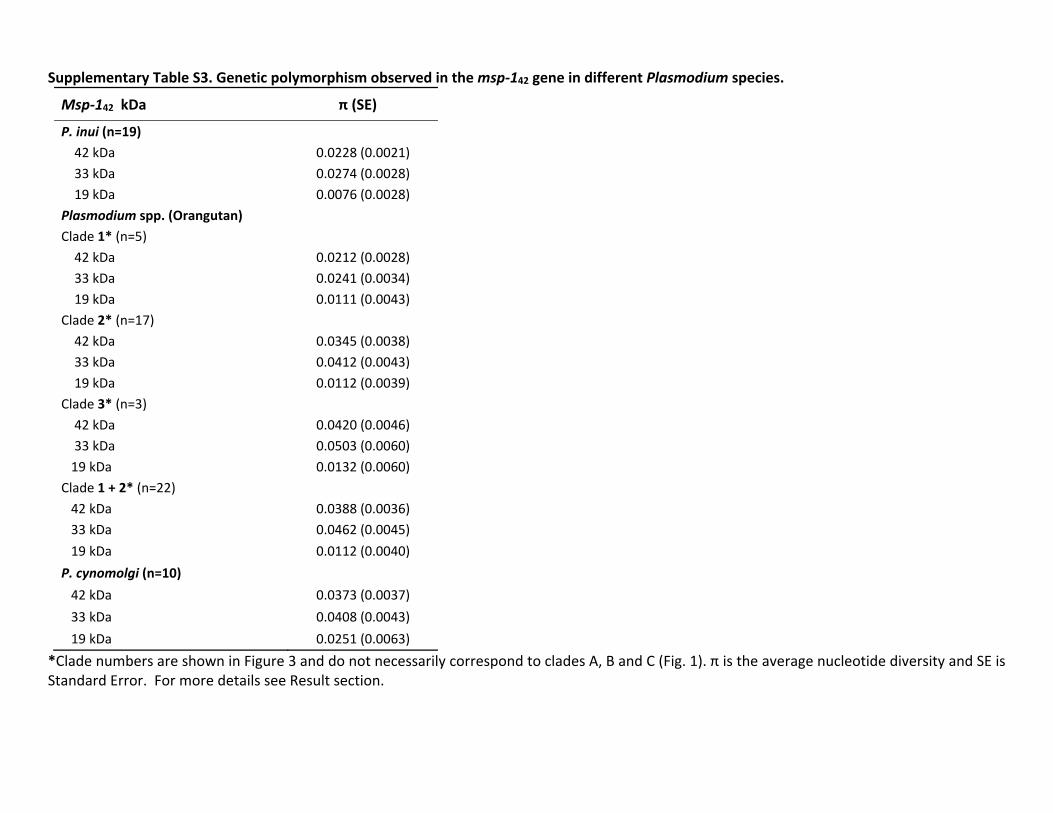
Supplementary13 Table13 S113 Plasmodium13 species13 included13
Species13 -shy‐13 Strain NCBI13 No Natural13 Host Geogrphic13 range Refrences
P13 vivax13 -shy‐13 Salvador13 I AY598140 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 cynomolgi13 -shy‐13 Berok AB444129 Macaca13 nemestrina
Kuala13 Lumpur13 Perak13 Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 cynomolgi13 (clone13 47A) KJ569865 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 22A) KJ569866 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 E2) KJ569867 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 (clone13 3AB) KJ569868 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
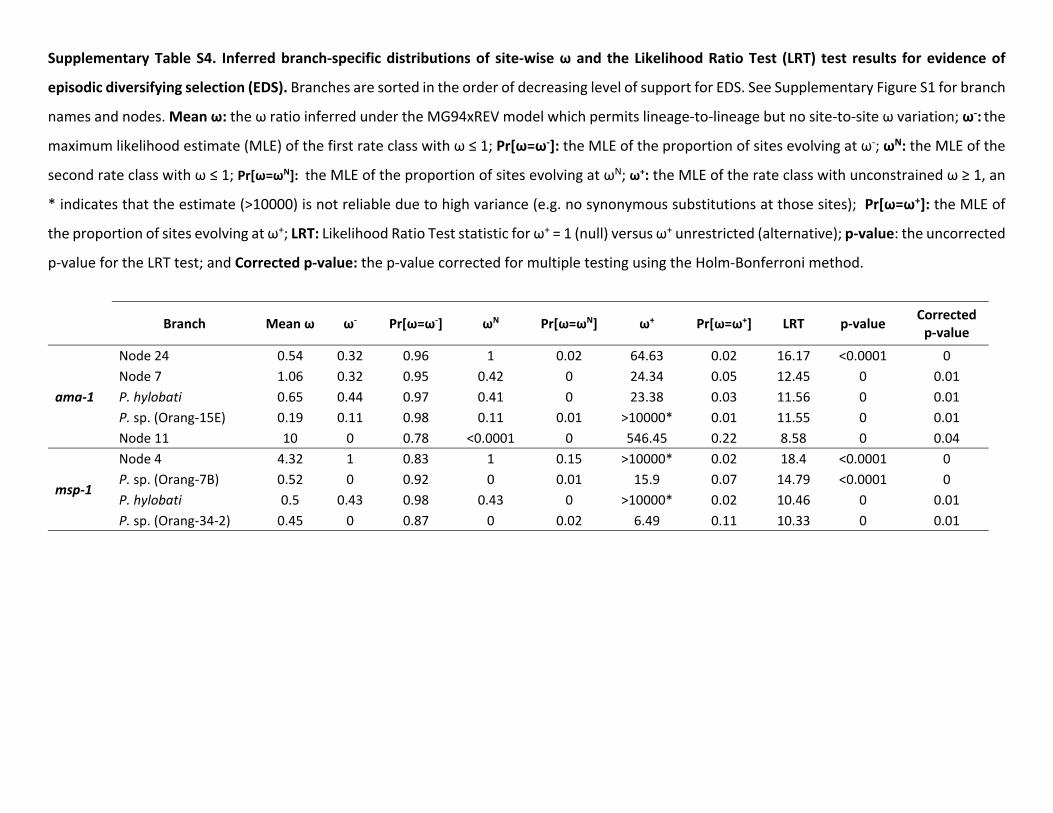
P13 cynomolgi13 ceylonensis AB44412513 Macaca13 sinica Ceylon13 (Sri13 Lanka)
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
Macaca13 sinica13 M13 nemestrina13 M13 fascicularis13 M13 mulatta13
P13 inui-shy‐13 Leaf13 Monkey13 II13 GQ355482 M13 radiata13 M13 cyclopis
Perlis13 state13 West13 Malaysia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 inui13 (clone13 42C) KJ569834 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 26A) KJ569835 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 28C) KJ569836 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui-shy‐13 Taiwan13 II13 GQ355483 South13 and13 East13 Asia
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
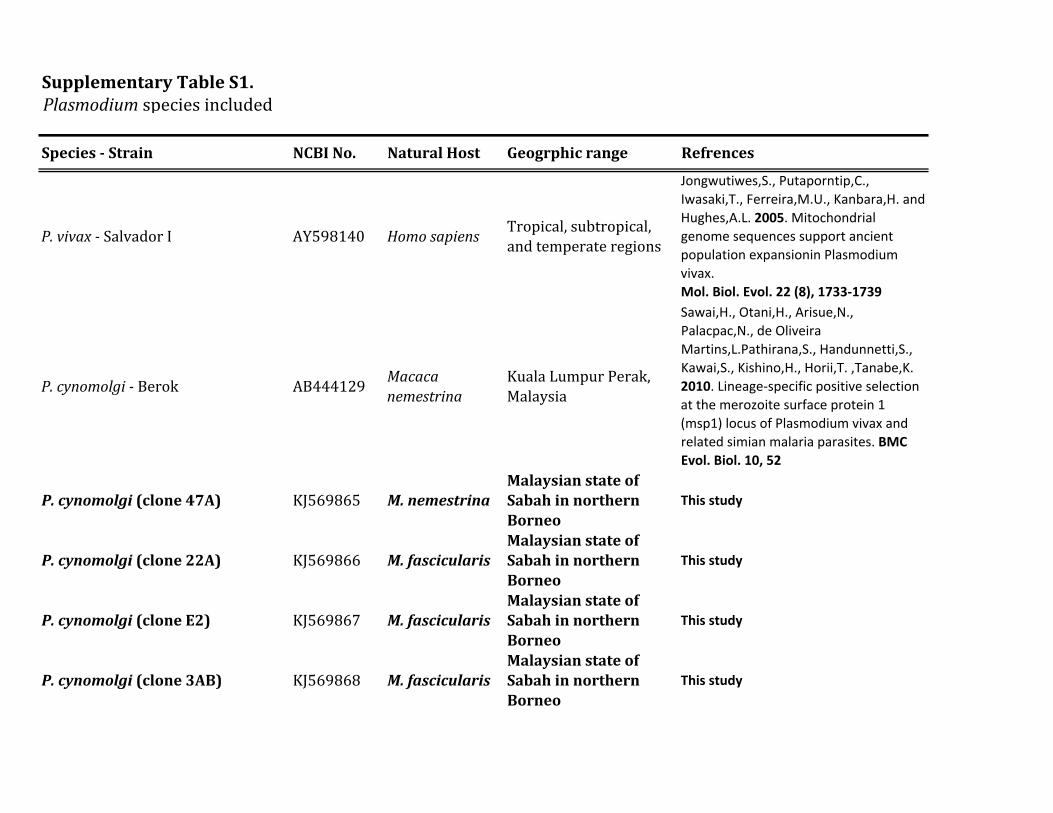
P13 inui13 (clone13 29G) KJ569837 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 30F) KJ569838 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 44E) KJ569839 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 45C) KJ569840 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 46C) KJ569841 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 48-shy‐2) KJ569842 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1AB) KJ569843 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 1A) KJ569845 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 3AA) KJ569846 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 14C) KJ569849 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 15A) KJ569844 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
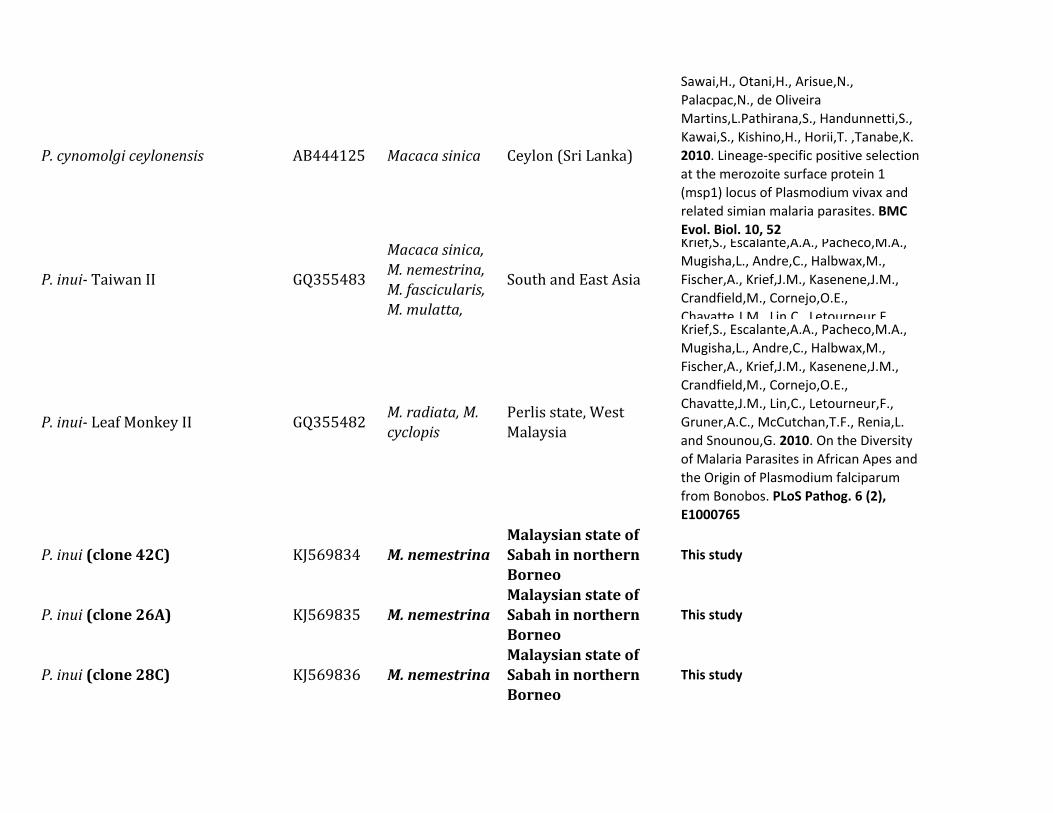
P13 inui13 (clone13 16C) KJ569847 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 17A) KJ569848 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 inui13 (clone13 20B) KJ569850 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 hylobati13 AB354573 Hylobati13 moloch Indonesia13 Malaysia13 (Borneo)
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
Plasmodium13 sp13 (VM82) JQ308531 Pongo13 pygmaeus Kalimantan13 Indonesia
PachecoMA13 ReidMJ13 SchillaciMA13 LowenbergerCA13 GaldikasBM13 Jones-shy‐EngelL13 and13 Escalante13 AA13 201213 The13 origin13 of13 malarial13 parasites13 in13 orangutans13 PLoS13 ONE13 713 (4)13 E3499013
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Yoda_7B) KJ569827
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Matamiss_19C) KJ569828
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Lumid_49C) KJ569829
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Anne_38B) KJ569830
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Salamat_24-shy‐2) KJ569831
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
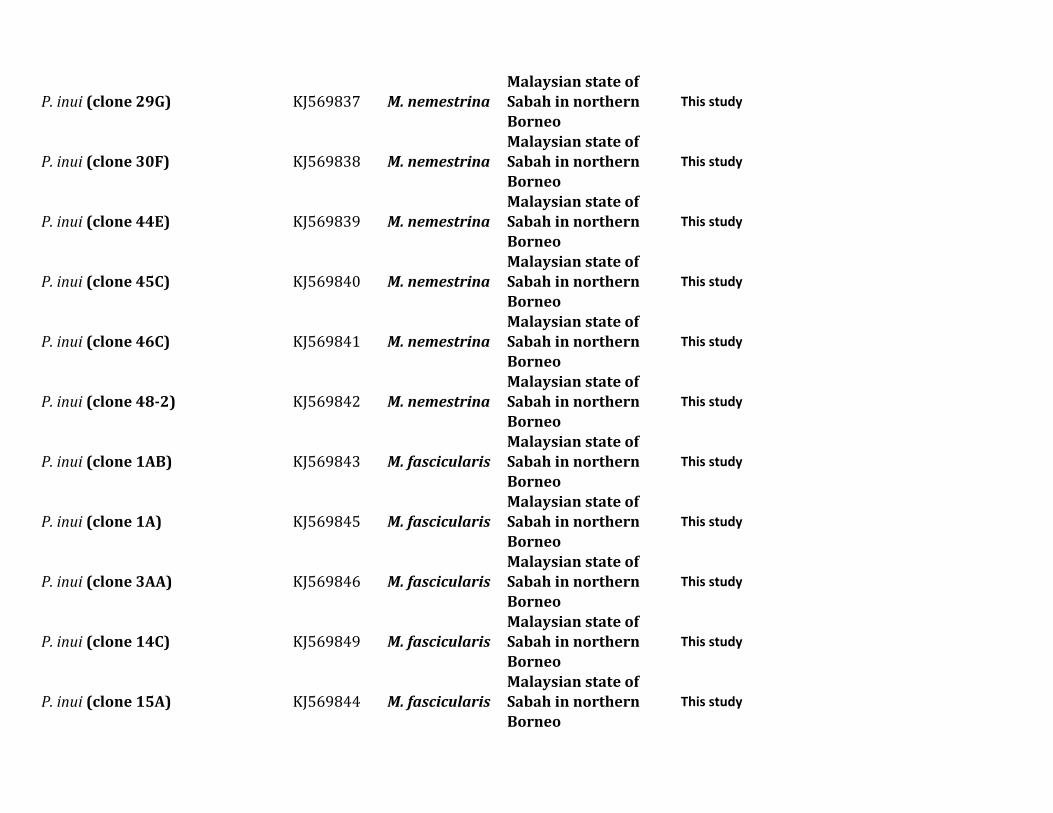
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Tambi_15E) KJ569832
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 A13 (clone13 Ganang_43A) KJ569833
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Gamai_10C) KJ569814
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Michelle_18C) KJ569815
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Poogel_9G) KJ569816
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Ceria_21B) KJ569817
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anne_38A) KJ569818
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Yoda_7C) KJ569819
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Salamat_24E) KJ569820
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rubin_17B) KJ569821
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 OTan_35B) KJ569822
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
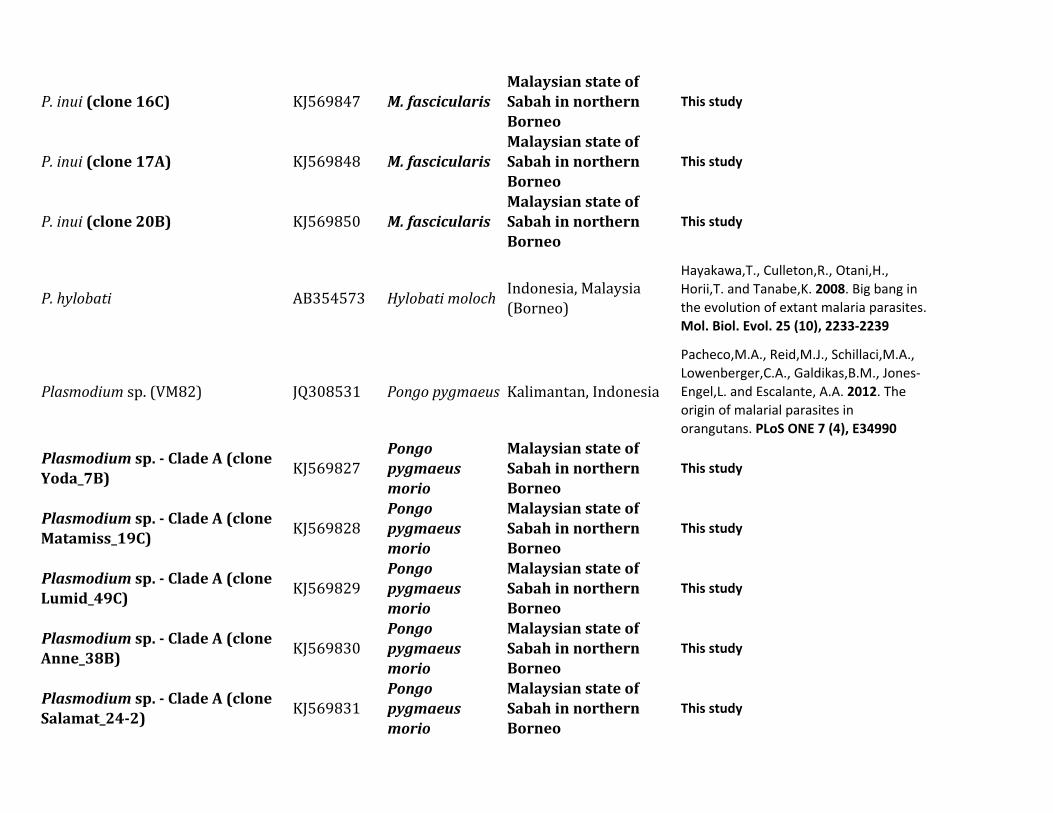
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Rosalita_36A) KJ569823
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Anakara_37A) KJ569824
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Hope_39B) KJ569825
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 B13 (clone13 Lumid_49B) KJ569826
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Poogel_9A) KJ569804
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Rosa_11C) KJ569805
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tobby_13A) KJ569806
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tambi_15B) KJ569807
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Angkung_34C) KJ569808
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Otan_35A) KJ569809
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Anakara_37C) KJ569810
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
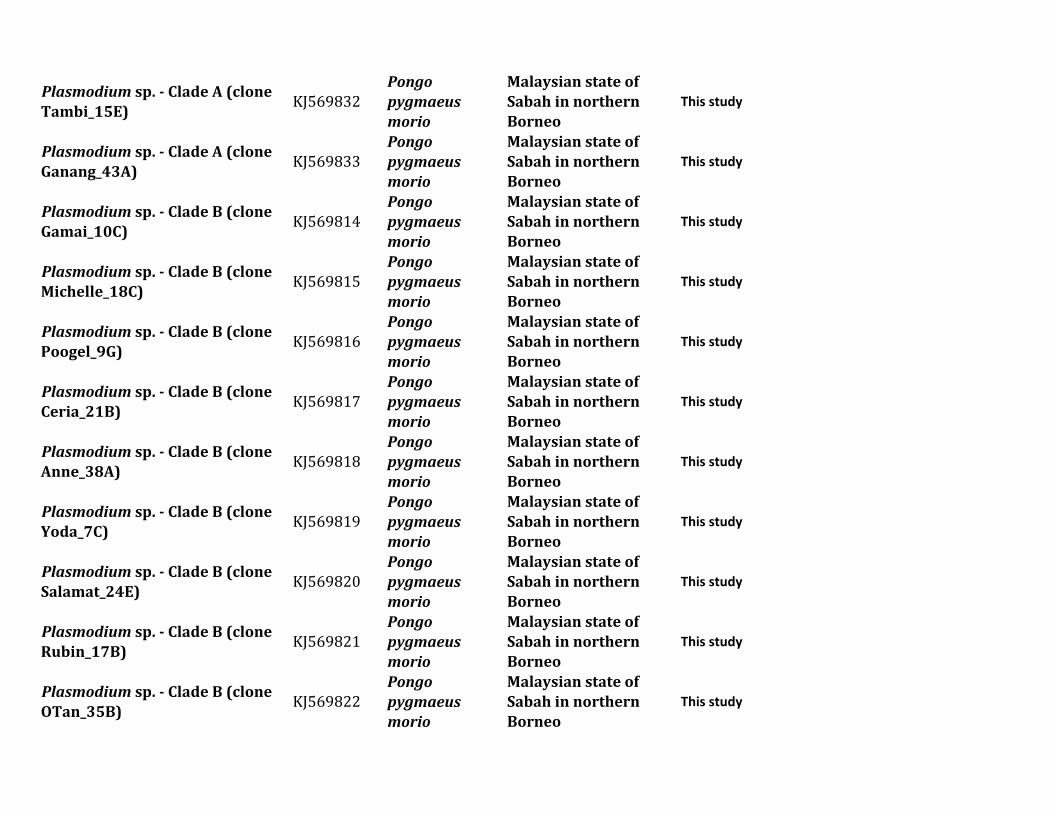
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Hope_39A) KJ569811
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Tiger_40C) KJ569812
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 -shy‐13 Clade13 C13 (clone13 Ganang_43D) KJ569813
Pongo13 pygmaeus13 morio
Malaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 simiovale AB434920 M13 sinica Sri13 Lanka
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi-shy‐13 N-shy‐313 AB354574 M13 nemestrina13 M13 fascicularis Malaysia
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 fieldi13 (clone13 48-shy‐4) KJ569863 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi-shy‐13 ABI13 AB444132 M13 nemestrina13 M13 fascicularis Malaysia
SawaiH13 OtaniH13 ArisueN13 PalacpacN13 de13 Oliveira13 MartinsLPathiranaS13 HandunnettiS13 KawaiS13 KishinoH13 HoriiT13 TanabeK13 201013 Lineage-shy‐specific13 positive13 selection13 at13 the13 merozoite13 surface13 protein13 113 (msp1)13 locus13 of13 Plasmodium13 vivax13 and13 related13 simian13 malaria13 parasites13 BMC13 Evol13 Biol13 1013 5213
P13 fieldi13 (clone13 32A) KJ569861 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
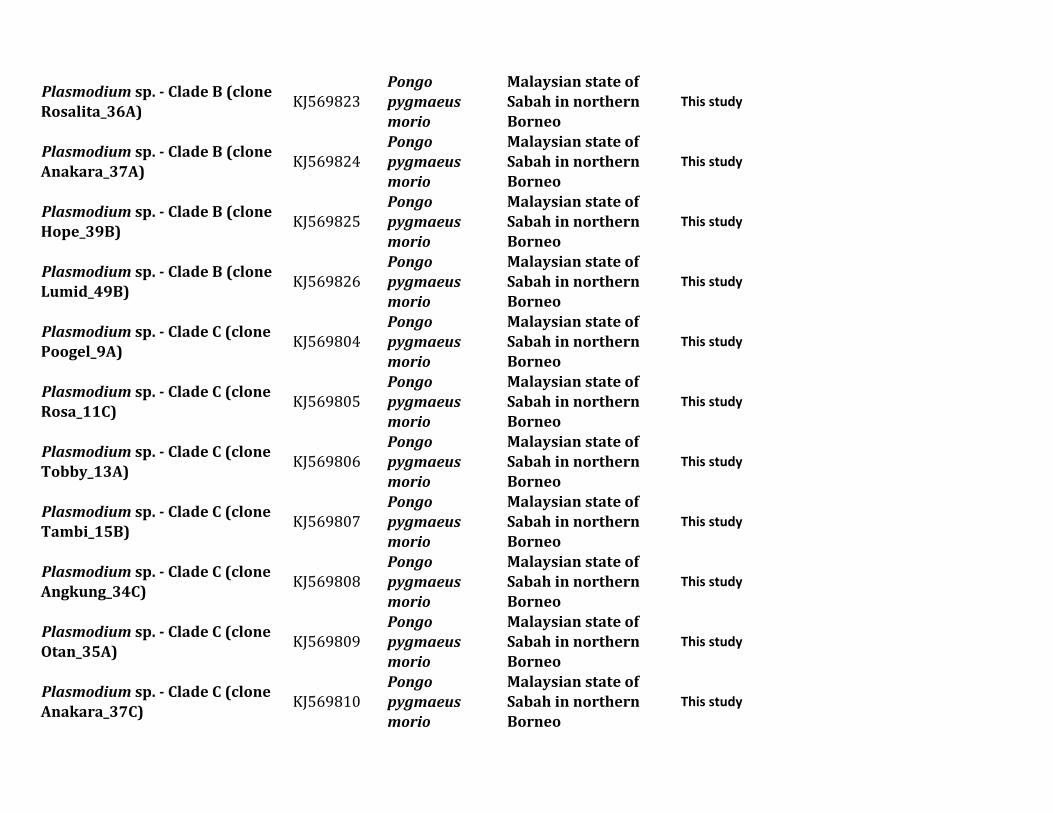
P13 fieldi13 (clone13 48B) KJ569862 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fieldi13 (clone13 2AA) KJ569864 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 coatneyi13 AB354575 M13 fascicularis Malaysia13 Philippines
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 knowlesi13 -shy‐13 Malayan NC_007232M13 nemestrina13 M13 fascicularis13 13 13 13 13 13 13 13 13 13 13 13 13 M13 nigra
Southeast13 Asia
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
Plasmodium13 sp13 (clone13 14F) KJ569851 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 31A) KJ569852 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 47-shy‐2) KJ569853 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 27C) KJ569854 M13 nemestrinaMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 SA1) KJ569855 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
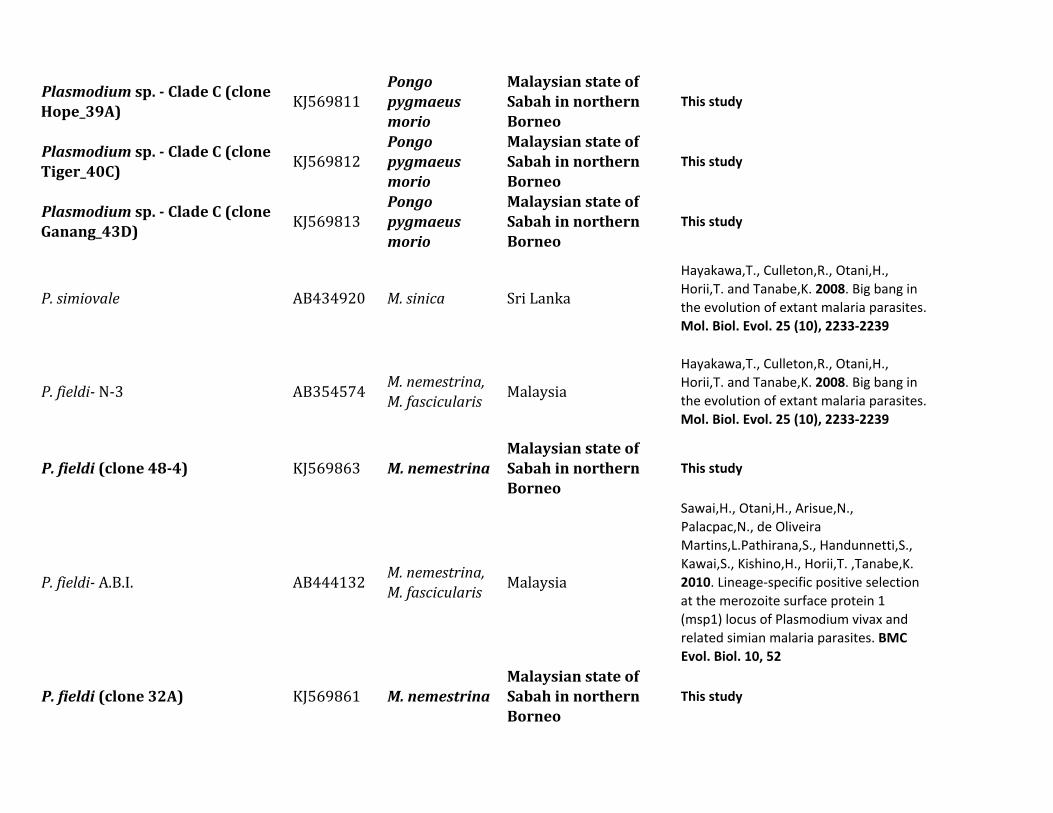
Plasmodium13 sp13 (clone13 1AA) KJ569856 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 23A) KJ569857 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 8A) KJ569858 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 4AB) KJ569859 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
Plasmodium13 sp13 (clone13 3-shy‐1) KJ569860 M13 fascicularisMalaysian13 state13 of13 Sabah13 in13 northern13 Borneo
This13 study
P13 fragile13 AY722799M13 radiata13 M13 mulatta13 Prebytis13 spp
Southern13 India13 Sri13 Lanka
JongwutiwesS13 PutaporntipC13 IwasakiT13 FerreiraMU13 KanbaraH13 and13 HughesAL13 200513 Mitochondrial13 genome13 sequences13 support13 ancient13 population13 expansionin13 Plasmodium13 vivax13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 13 Mol13 Biol13 Evol13 2213 (8)13 1733-shy‐173913
P13 gonderi13 AY800111Cercocebus13 atys13 Cercopithecus13 spp
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
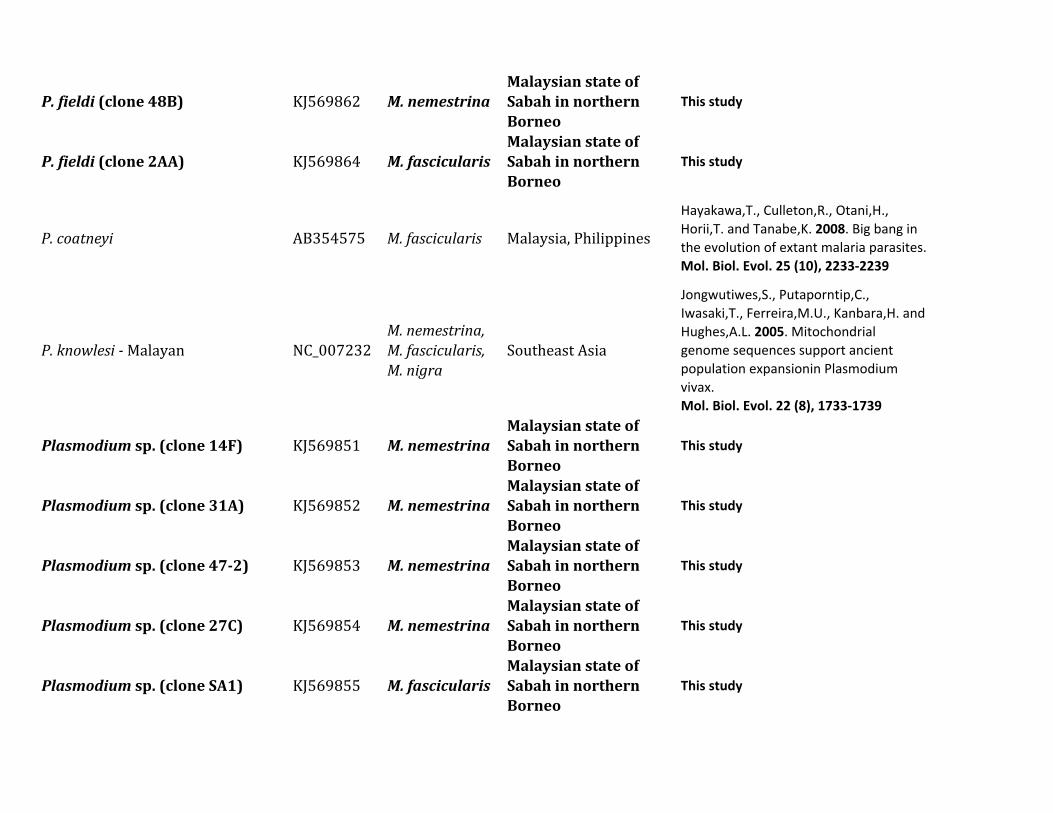
Plasmodium13 sp13 13 13 AY800112
Mandrillus13 sphinx13 (Cercopithecidae)
Central13 Africa
MuJ13 JoyDA13 DuanJ13 HuangY13 CarltonJ13 WalkerJ13 BarnwellJ13 BeerliP13 CharlestonM13 PybusOG13 and13 SuXZ13 200513 Host13 switch13 leads13 to13 emergence13 of13 Plasmodium13 vivax13 malaria13 in13 humans13 Mol13 Biol13 Evol13 2213 (8)13 1686-shy‐169313
P13 malariae AB354570 Homo13 sapiens Tropical13 subtropical13 and13 temperate13 regions
HayakawaT13 CulletonR13 OtaniH13 HoriiT13 and13 TanabeK13 200813 Big13 bang13 in13 the13 evolution13 of13 extant13 malaria13 parasites13 Mol13 Biol13 Evol13 2513 (10)13 2233-shy‐223913
P13 ovale-shy‐wallikeri HQ712053 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 ovale-shy‐curtisi13 HQ712052 Homo13 sapiens Tropical13 and13 subtropical13 regions
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Plasmodium13 sp13 (A)13 HQ712054 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (B)13 HQ712055 Varecia13 variegata
Plasmodium13 sp13 (C)13 HQ712056 Hapalemur13 griseus13 griseus
Plasmodium13 sp13 (D)13 HQ712057 Indri13 indri
Plasmodium13 sp13 (E)13 JN131536 Eulemur13 macaco13 macaco13
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
Madagascar13 (Eastern13 rainforest)13
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
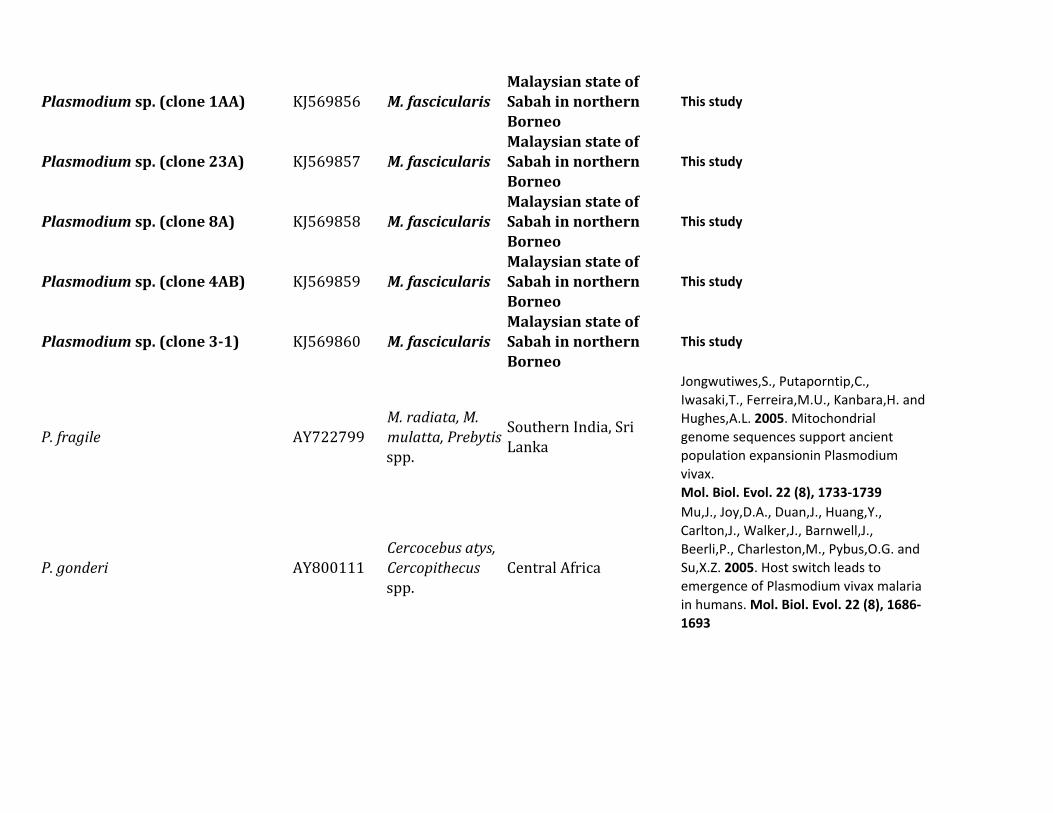
P13 berghei AF014115 Grammomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 yoelii13 M29000 Thamnomys13 sp Africa
VaidyaAB13 AkellaR13 and13 SuplickK13 198913 Sequences13 similar13 to13 genes13 for13 two13 mitochondrial13 proteins13 and13 portions13 of13 ribosomal13 RNA13 in13 tandemly13 arrayed13 6-shy‐kilobase-shy‐pair13 DNA13 of13 a13 malarial13 parasite13 Mol13 Biochem13 Parasitol13 3513 (2)13 97-shy‐10713
P13 atheruri13 HQ712051Atherurus13 africanus13 (Porcupine)
Central13 Africa
PachecoMA13 BattistuzziFU13 JungeRE13 CornejoOE13 WilliamsCV13 LandauI13 RabetafikaL13 SnounouG13 Jones-shy‐EngelL13 EscalanteAA13 201113 Timing13 the13 origin13 of13 human13 malarias13 the13 lemur13 puzzle13 BMC13 Evol13 Biol13 1113 29913
P13 chabaudi13 AF014116 Thamnomys13 sp Central13 AfricaTanTMC13 NoviyantiR13 Syafruddin13 MarzukiS13 and13 TingRCY13 200013 NCBI13 Direct13 Submission
P13 vinckei13 vinckei AB599931Thamnomys13 spGrammomys13 sp
Central13 Africa
Hikosaka13 K13 Watanabe13 Y13 Kobayashi13 F13 Waki13 S13 Kita13 K13 Tanabe13 K13 201113 Highly13 conserved13 gene13 arrangement13 of13 the13 mitochondrial13 genomes13 of13 2313 Plasmodium13 species13 Parasitol13 Int13 201113 Jun60(2)175-shy‐80
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
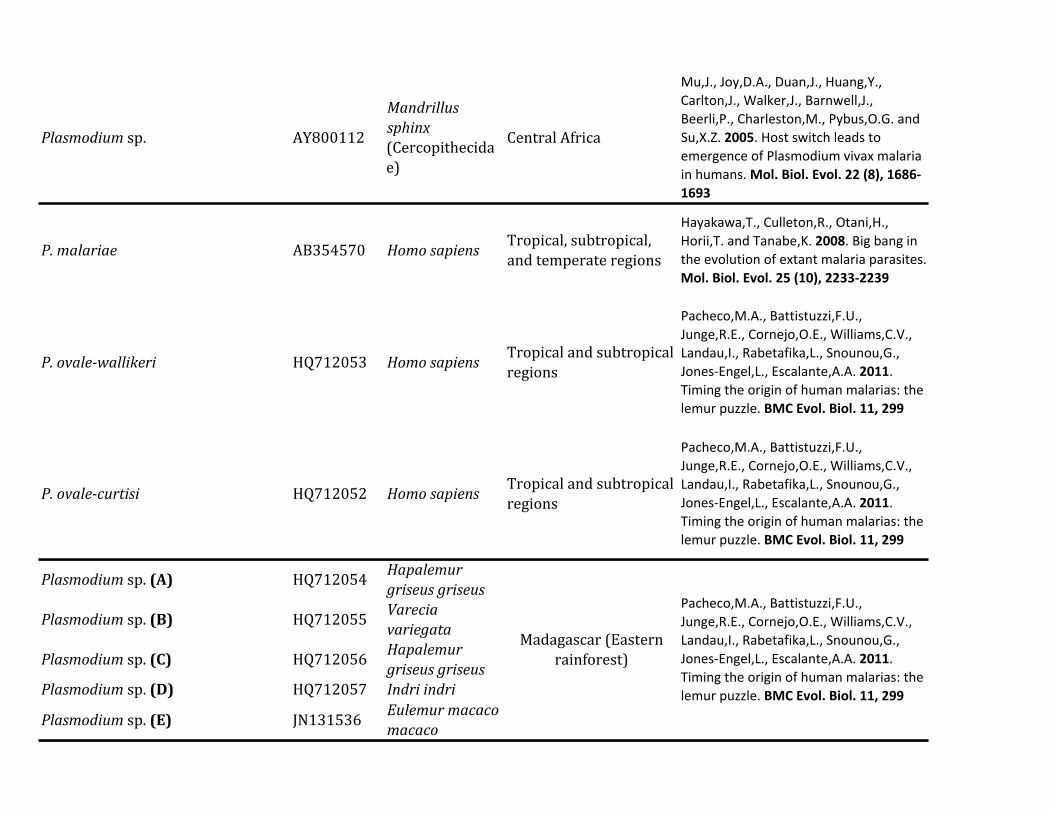
Plasmodium13 billbrayi GQ355468 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
Plasmodium13 billcollinsi GQ355479 Pan13 troglodytes Uganda13 Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 reichenowi13 NC-shy‐002235 Pan13 troglodytes Africa
ConwayDJ13 FanelloC13 LloydJM13 Al-shy‐JouboriBM13 BalochAH13 SomanathSD13 RoperC13 OduolaAM13 MulderB13 PovoaMM13 SinghB13 and13 ThomasAW13 200013 Origin13 of13 Plasmodium13 falciparum13 malaria13 is13 traced13 by13 mitochondrial13 DNA13 Mol13 Biochem13 Parasitol13 11113 (1)13 163-shy‐17113
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-
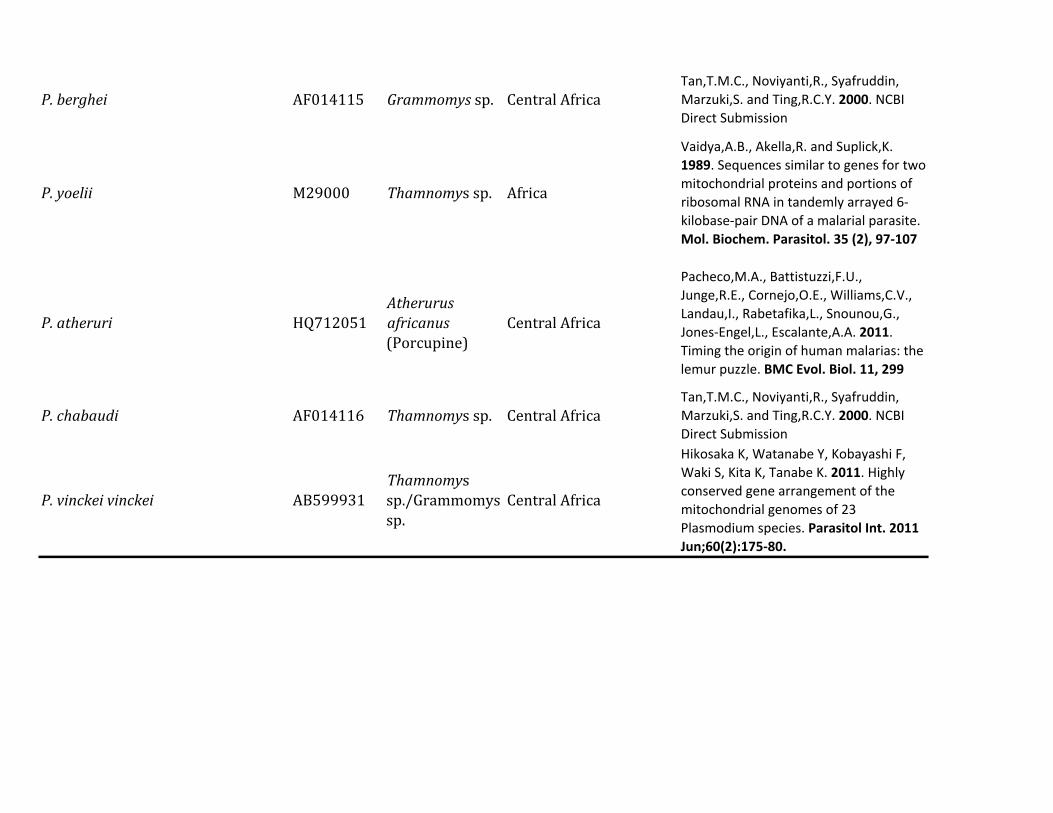
Plasmodium13 sp GQ355476 Pan13 troglodytes Republic13 of13 the13 Congo
KriefS13 EscalanteAA13 PachecoMA13 MugishaL13 AndreC13 HalbwaxM13 FischerA13 KriefJM13 KaseneneJM13 CrandfieldM13 CornejoOE13 ChavatteJM13 LinC13 LetourneurF13 GrunerAC13 McCutchanTF13 ReniaL13 and13 SnounouG13 201013 On13 the13 Diversity13 of13 Malaria13 Parasites13 in13 African13 Apes13 and13 the13 Origin13 of13 Plasmodium13 falciparum13 from13 Bonobos13 PLoS13 Pathog13 613 (2)13 E100076513
P13 falciparum13 AY282930 Homo13 sapiens Worldwide13 Tropical13 regions
JoyDA13 FengX13 MuJ13 FuruyaT13 ChotivanichK13 KrettliAU13 HoM13 WangA13 WhiteNJ13 SuhE13 BeerliP13 and13 SuXZ13 200313 Early13 origin13 and13 recent13 expansion13 of13 Plasmodium13 falciparum13 Science13 30013 (5617)13 318-shy‐32113
P13 juxtanucleare NC_008279 Galliformes13 and13 domestic13 birds
Tropical13 and13 subtropical13 regions
P13 gallinaceum13 NC_008288 Galliformes13 and13 Sphenisciformes
P13 floridense13 NC_009961 Anolis13 sagrei Caribbean
P13 mexicanum13 NC_00996013 Sceloporus13 occidentalis California13 USA
PerkinsSL13 200613 Whole13 genome13 amplification13 and13 sequencing13 of13 mitochondrial13 DNA13 of13 saurian13 malaria13 parasites13 Unpublished13 NCBI13 Direct13 Submission
OmoriS13 SatoY13 IsobeT13 YukawaM13 and13 MurataK13 200713 Complete13 nucleotide13 sequences13 of13 the13 mitochondrial13 genomes13 of13 two13 avian13 malaria13 protozoa13 Plasmodium13 gallinaceum13 and13 Plasmodium13 juxtanucleare13 Parasitol13 Res13 10013 (3)13 661-shy‐66413
- Muehlenbein et al 2015
-
- Muehlenbein et al 2015
- Supp-FigS1-3
- Supp-Fig-S4
- Supplementary_Table_S2-S4-OCT31
- Supp-Table-1
-
- Supp-Table-1
-















![Serotonin transporter binding and genotype in the nonhuman primate brain using [C-11]DASB PET](https://static.fdokumen.com/doc/165x107/633625ec02a8c1a4ec02115b/serotonin-transporter-binding-and-genotype-in-the-nonhuman-primate-brain-using-c-11dasb.jpg)




