
Mechanistic aspects of biosynthesis of silver nanoparticles by several Fusarium oxysporum strains
Upload
independentCategory
view
4download
0
ARTICLE IN PRESS
0885-5765/$ - se
doi:10.1016/j.pm
�CorrespondE-mail addr
Physiological and Molecular Plant Pathology 69 (2006) 73–85
www.elsevier.com/locate/pmpp
A role for the mycotoxin deoxynivalenol in stem colonisation duringcrown rot disease of wheat caused by Fusarium graminearum and
Fusarium pseudograminearum
Agnieszka M. Mudgea, Ruth Dill-Mackyb, Yanhong Dongb, Donald M. Gardinera,Rosemary G. Whitec, John M. Mannersa,�
aCSIRO Plant Industry, Queensland Bioscience Precinct, 306 Carmody Road, St. Lucia, Brisbane Qld. 4067, AustraliabDepartment of Plant Pathology, University of Minnesota, 495 Borlaug Hall, 1991 Upper Buford Circle, St. Paul, MN 55108, USA
cCSIRO Plant Industry, PO Box 1600, Canberra ACT 2601, Australia
Accepted 25 January 2007
Abstract
Fusarium graminearum (Fg) can cause head blight and crown rot (CR) diseases of wheat but fungal colonisation and mycotoxin
production by Fg during CR are little understood. Studies of an Australian strain of Fg demonstrated that expression of the Tri5 gene of
Fg and deoxynivalenol (DON) production were induced during infection of the stem base and to levels equivalent to those observed in
inoculated heads. To study fungal colonisation and DON production in CR disease, we inoculated stem bases, 14 days after sowing, with
macroconidia of Australian and USA strains of Fg and of an Australian strain of Fusarium pseudograminearum, a related pathogen
frequently associated with CR. At maturity, the fungal pathogen was subsequently detected by isolation on growth media in high
percentages of plants at the stem base and the lowest node adjacent to the inoculation site (50–100%), in the non-inoculated flag-leaf
node (FLN) (35–95%) and in asymptomatic non-inoculated mature heads (50–60%) and kernels (20%). Microscopic analysis suggested
that colonisation of upper nodes occurred primarily via the pith parenchyma and lumen. Significant concentrations of DON (up to
35 ppm) were detected in the FLN and the head/rachis following inoculation of the stem base. To test the role of DON in CR a transgenic
strain of Fg from the USA with the Tri5 gene deleted was compared with its original wild-type and these produced similar levels of CR
lesion development and necrosis indicating that DON production was not necessary to cause CR disease symptoms. However, the DON-
minus mutant was less frequently recovered from the FLN than the wild type suggesting that DON has a role in stem colonisation by the
fungus.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Head blight; Head scab; Gibberella zeae; Fungal gene expression; Trichothecene; Fungal infection
1. Introduction
Fusarium head blight (FHB) is a re-emerging disease ofwheat that is now globally important [1–3]. Severalpathogens in the genus Fusarium can cause FHB but inmany regions of the world members of the Fusarium
graminearum Schwabe species complex (Fg) (teleomorphGibberella zeae [Schw.] Petch), are the principle pathogensresponsible for this disease. It is well known that infection
e front matter r 2007 Elsevier Ltd. All rights reserved.
pp.2007.01.003
ing author. Tel.: +617 32142304; fax: +61 7 3214 2920.
ess: [email protected] (J.M. Manners).
of wheat heads by Fg, and other Fusarium patho-gens, reduces grain quality through the production ofmycotoxins such as the trichothecene deoxynivalenol(DON) making grain unsafe for human and livestockconsumption [1,2].Epidemics of FHB caused by Fg have been reported in
Australia but these are not frequent and are usuallylocalised to regions that have a higher incidence of rainfallduring anthesis and grain filling and where maize (Zea
mays L.) or sorghum (Sorghum bicolour L. Moench) havebeen grown in rotation with wheat [4–6]. However, Fg andother Fusarium pathogens can infect other parts of the

ARTICLE IN PRESSA.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–8574
wheat plant and are the principle cause of crown rot (CR)that is a chronic problem in wheat production in Australia[4,7–9] and other parts of the world [10,11] causingsignificant yield losses each year. The primary fungalspecies causing CR of wheat in Australia is Fusarium
pseudograminearum T. Aoki & O’Donnell (Fp) and is onlyrarely observed as its teleomorph, Gibberella coronicola T.Aoki and O’Donnell [4,7,9,12]. The species Fp and Fg werepreviously termed F. graminearum Group 1 and Group 2,respectively [13], but were subsequently separated into Fp
and Fg following molecular phylogenetic analysis andmating studies [14,15]. The Fg species complex is stillundergoing phylogenetic examination and it has recentlybeen proposed, on the basis of a sequence analysis of eightloci, that nine phylogenetic species may be recognisedwithin Fg [16]. Although surveys of Fusarium pathogens innorthern wheat growing regions of Australia have shownthat Fg isolates are primarily obtained from infected headswhile Fp isolates are more frequently obtained frominfected crowns [4], there is strong evidence from thesefield surveys and associated glasshouse infection tests thatFg and Fp can both cause FHB and CR [4,5]. There is littleinformation on host resistance mechanisms to CR [17] andthere is a dearth of information on the host–pathogenbiology of CR disease. Because of the global research focuson Fg and a knowledge base of its biology which is growingrapidly due to the availability of the genome sequence andan expanding collection of mutants for specific genes [18],this pathogen species represents a good model for studiesof CR development by Fusarium pathogens.
The symptoms associated with CR are necrosis of thecrown and stem base and in severe cases, the formation of‘whiteheads’, which are heads that die prematurely duringgrain development, when infected plants are stressed bywater deficit [7]. Primary infection is thought to occur atthe emerging shoot, crown and stem base from aninoculum of spores and mycelium on residual stubble butthere is little information on the histopathology of theinfection process [7]. Once infection is established in thestem base and crown, reports so far suggest that onlylimited fungal colonisation can occur in the stem,occasionally reaching the flag-leaf node (FLN), and thatfungal colonisation of the plant is stimulated by waterdeficit stress [7,19,20]. There are no reports of fungalcolonisation of wheat heads following inoculation of thecrown or stem base [7]. It is therefore thought that plantdeath and the formation of whiteheads results fromnecrosis in the crown and stem interfering with thetransport of water and nutrients to the developing head [7].
Both Fg and Fp are known to produce the trichothecenemycotoxin DON in culture and in planta when incitingFHB disease [5,21,22]. The biosynthesis of trichothecenemycotoxins is determined by multiple enzymes many ofwhich are encoded on a gene cluster in Fg [22]. One of themost intensively studied of these genes is Tri5 that encodesa trichodiene synthase, the enzyme responsible for the firstcommitted step in the synthesis of trichothecenes [22,23].
For Fg there is compelling evidence from gene knock outexperiments in the fungus and from the manipulation ofDON metabolism in transgenic wheat plants, that DON,and thus the Tri5 gene, contribute to the aggressiveness ofFg in FHB [24–27]. However, the production and potentialrole of DON in CR during the colonisation of stem tissueby either Fg or Fp have not been reported [7].We report here on an analysis of fungal colonisation,
Tri5 gene expression and DON production, in wheatfollowing inoculation of the stem base of wheat seedlingsby Fusarium pathogens. The experiments described in thispaper were conducted both in Australia and in the USAwith isolates from each of these respective countries, butare described here together because consistent results wereobtained in both studies. These experiments suggest thatthe Fusarium pathogen and trichothecene mycotoxins maybe more extensively distributed throughout the tissues ofwheat plants following infection via the stem base than haspreviously been recognised and these toxins have animportant role in colonisation of the stem.
2. Materials and methods
2.1. Fungal strains
The Australian strains used in these experiments wereCS3002 (Fp), isolated from infected crowns (Queensland,Australia) and CS3040 and CS3005 (Fg), isolated frominfected wheat heads (New South Wales, Australia). Therespective identities of CS3002 as Fp, and CS3040 andCS3005 as Fg as recently defined by O’Donnell et al. [16],were confirmed by DNA sequence and phylogeneticanalysis of the translation elongation factor-1a (TEF)and phosphate permease (PHO) 2 gene regions [28]. TheUSA strains tested included Butte86ADA-11 (B86A11), anisolate of Fg collected from a commercial wheat field inMinnesota that is highly aggressive in causing head blight[29]. Studies also were conducted on the wild-type strainGz3639 of Fg and its near-isogenic strain GzT40 which is atransgenic mutant derived from Gz3639 by deletion of theTri5 gene [27]. It should be noted that because ofquarantine regulations, experiments with fungal isolateswere conducted in the country of origin of each isolate.
2.2. Fungal culture conditions
Fungi were cultured in Australia on SNA (SpeziellerNahrstoffarmer Agar) plates [4,30] and kept at roomtemperature. Spores for inoculations were produced in100ml Mung Bean Broth [31] inoculated with 10 SNA agarplugs (6mm diameter) colonised with either strain andincubated with shaking (150 rpm) at room temperature for5–7 days. Spores were collected by filtering the broththrough 100 mm mesh nylon cloth (Millipore) and rinsed atleast twice in sterile water. For the production of vegetativemycelium for mycotoxin and gene expression analysis,100ml of potato dextrose broth (PDB) from Bacto

ARTICLE IN PRESSA.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–85 75
Laboratories, Liverpool, NSW, Australia, were inoculatedwith a spore suspension from one SNA plate and the liquidcultures incubated at room temperature for 48 h. In theUSA macroconidia of the three Fg isolates were producedby culture on mung bean agar as described [29]. Briefly,isolates were cultured on mung bean agar under fluorescentlight (3:1 cool white:black light, 12:12 light:dark, 36mol/m2 s)at room temperature (20–22 1C) for 14 days. Spores werethen washed from the agar surface with 10–20ml ofdistilled water per petri dish. The suspension of spores andmycelial fragments were filtered through one layer ofcheesecloth and spore concentrations determined using ahemacytometer. Spore suspensions were adjusted to 5� 105
spores per ml in distilled water and stored at �80 1C untilneeded.
2.3. Plant growth and inoculation technique
Seeds of the susceptible bread wheat cultivar Kennedywhich is known to be susceptible to both CR and FHB,were grown in Brisbane, Australia in controlled environ-ment rooms with photoperiod length of 18 h, 22 1C and60% RH at 500 mmol/m2 s. Night-time temperature was setat 18 1C, with 80% RH. Five to six seeds were sown per pot(14 cm diameter, 17 cm height) containing steamed pottingmix (50% peat and 50% sand by volume) and thinned tosingle plants after germination. Inoculations of the stembase were performed as described [4]. Briefly, at 14 daysafter sowing a small piece (30� 15mm) of sterilisedabsorbent cotton as wrapped around the base of the stemimmediately above the soil layer and held in place with asleeve of plastic wrap. Plants were inoculated by applying700 ml of a suspension of 5� 105 spores per ml of water.Control plants were mock-inoculated with water only.Inoculation of the stem base in this manner results in abrown lesion that spread up the stem and down throughthe crown producing symptoms identical to that of CRdisease observed in the field [4]. For FHB disease, wheat cv.Kennedy was sown as described above but grown in a glasshouse at 25 1C day and 15 1C night temperatures (73 1C)under natural light conditions. Inoculations were per-formed when heads were at mid-anthesis, as described byAkinsanmi et al. [4]. A 1.5ml suspension of 5� 105 sporesper ml of water was atomised onto each head and headswere then covered with a plastic bag for 48 h andsubsequently with a paper bag until disease assessment.For experiments conducted in the USA the hard red springwheat cultivar Wheaton (FHB susceptible) was used.Plants were grown in pots (6.4 cm diameter� 25 cm depth;D40 Deepots, Stuewe & Sons, Inc., Corvallis, OR)containing commercial potting medium (Metromix 200,Scotts Co., Marysville, OH) and held in trays (D40T,Stuewe & Sons, Inc., Corvallis, OR) in a growth chamber(Conviron E15; Controlled Environments Ltd., Winnipeg,Man., Canada) with photoperiod length of 16 h at825 mmol/m2 s. Day-time settings were 22 1C and 60%RH and night-time settings 18 1C and 80% RH. Three
seeds were sown per pot and thinned to one plant per pot 7days after planting. Inoculations of the stem base wereperformed on 14-day-old seedlings as described above.
2.4. DNA Extractions and PCR
DNA for PCR analysis was extracted following amodified method of Irelan et al. [32]. Fungal myceliumwas grown in 0.5ml PDB for 2–5 days, collected bycentrifugation at 13,000 rpm for 10min. Mycelium wasresuspended in 0.5ml isolation buffer [33] by briefvortexing. Cells were disrupted by boiling for 3min.Proteins were precipitated on ice for 10min after additionof 0.3ml of 7.5M ammonium acetate. After centrifugationat 13,000 rpm for 5min, 0.7ml of the supernatant wasadded to 0.5ml of isopropanol and incubated on ice10min. The precipitated DNA was collected by centrifuga-tion at 13,000 rpm for 10min, washed with 70% EtOH, airdried, and resuspended in 50 ml water.PCR reactions were performed in a total volume of 25 ml
and included 1 ml template DNA, 240 nM of each forwardand reverse primer, 2.0mM MgCl2, 0.2mM dNTPs and0.5U Taq DNA polymerase in a 1� reaction buffer (NewEngland BioLabs, Beverly, MA, USA). The DNA ampli-fication program was as follows: an initial 95 1C denatura-tion for 2min was followed by 30 cycles of 95 1C for 30 s,55 1C for 30 s, 72 1C for 50 s, followed by a final extensionof 72 1C for 4min. PCR fragments were separated on a1.8% 0.5�TBE gel stained with EtBr and photographed.The primers used to identify Fp were Fp1-1 and Fp1-2,
published previously by Aoki and O’Donnell [14]. Theprimers used to identify Fg were Fg1-1 (50-CMCTCA-CACGACGACTCGATAC-30) and Fg1-2 (50-GAGA-ATGTGATGACAGCAGTG-30), amplifying a 363 bpfragment from EF-1a (translation elongation factor),designed after Aoki and O’Donnell [14]. The primers usedto amplify the GFP gene present on vector pTG present infungal transformants were APgfp50 (50-CAAGGGCGAG-GAGCTGTTC-30) and APgfp30 (50-CAGCTTGTGCCC-CAGGATG-30), generating a 417 bp amplicon.
2.5. Real-time quantitative PCR analysis of Tri5 expression
Total RNA was extracted from samples using the CsClmethod described in Masel et al. [34]. For real-timequantitative PCR (qRT-PCR) 5 mg total RNA was treatedwith 1U amplification grade DNaseI (Invitrogen, Carls-bad, CA, USA) according to manufacturer’s instructions,then reverse transcribed using SuperScript III (Invitrogen,Carlsbad, CA, USA) according to manufacturer’s instruc-tions, and the resulting cDNA diluted 1:10 with water.PCR was conducted in a total volume of 12 ml with a finalconcentration of 1� SYBR GREEN PCR master mix(Applied Biosystems, Scoresby, Vic., Australia), 1 ml cDNAand 500 nM of each forward and reverse primer. Cyclingconditions consisted of a 15 s denaturation step at 95 1Cfollowed by anneal/extension step of 1min at 60 1C,

ARTICLE IN PRESSA.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–8576
repeated a total of 40 times. A final denaturing step wasdone to generate the dissociation curve. Primers used toamplify Tri5 were APT5newF (50-CACTTTGCTCA-GCCTCGCC-30) and APT5newR (50-CGATTGTTTG-GAGGGAAGCC-30), producing an amplicon of 76 bp.Amplification rate of Tri5 was compared to Fg 18S rRNAamplified using primers 18sF50 (50-GTCCGGCCG-GGCCTTTCC-30) and 18sF30 (50-AAGTCCTGTTTCC-CCGCCACGC-30), producing an amplicon of 68 bp.Primers used to differentiate wheat template from fungaltemplate were 18sP50 (50-CGACCTACTCGACCCTTC-GGCCGG-30) and 18sP30 (50-CGATGCCGGAAACAC-GACCCGG-30). Data was analysed using the programLinReg PCR, version 7.5 to calculate internal PCRamplification efficiencies and absolute gene expressionvalues were determined using average efficiencies raisedto the negative crossing threshold [35].
2.6. Assessment of fungal colonisation following inoculation
of the stem base
For the experiments conducted in Australia, transgenichygromycin-resistant strains of both Fg and Fp were used.Firstly, this ensured that fungal isolates recovered fromnon-inoculated regions of inoculated plants could beunambiguously identified as the same strain used forinoculation, and secondly it meant that hygromycin couldbe included in isolation plates to recover fungal isolatesfrom plants thereby minimising contamination with otherpotential saprophytes and endophytes. Therefore, the wild-type strains CS3040 and CS3002 were genetically trans-formed with the vector pTG (A. Mudge, unpublished)derived from pAN7.1 [36] and contains the bacterial hph
gene conferring resistance to the antibiotic hygromycinunder the regulatory sequences of the trpC gene fromAspergillus nidulans (Eidam). The transformation protocolused was essentially as described previously [37] except thatabout 2 g wet weight mycelium was incubated in 10mlMSP (1.2M MgSO4 in 10mM phosphate buffer pH 5.8)and 10ml SSO (0.6M sorbitol, 0.6M sucrose pH 5.6) inwhich were dissolved 1 g glucanex (NovoNordisk, Gen-thofte, Denmark) and 0.5 g Driselase (Sigma, St. Louis,MO, USA), and incubated at 30 1C. Transformation of Fg
and Fp yielded transgenic strains FgT21 and FpT2,respectively. Transformants were maintained at roomtemperature on SNA plates supplemented with hygromycinat 100 ppm for FgT21 and 50 ppm for FpT2.
Two inoculation experiments were conducted withFgT21 (labelled FgA and FgB in Fig. 3), each with 20inoculated and 20 control plants and one experiment wasconducted with the FpT2 strain, using 10 inoculated and 10control plants. Plants were inoculated as described aboveand plants and mock-inoculated control plants were thenallowed to mature and senesce. Plant tissue samples wereharvested prior to the plants completely drying off. Toreduce the potential of any accidental splash dispersal offungal spores from inoculated plants onto other plants the
pots were placed in trays of water, no deeper than 2 cm,which were topped up as needed to water the plants. Headswere covered in glassine bags (Clarke Packaging, Brisbane,Australia) as they emerged. At maturity the length of thelesion at the inoculation site was measured and rated fordegree of discoloration using a three-point scale where 0—non-visible, 1—moderate browning and 2—intense brown-ing. To minimise cross-contamination during furthersampling, first the control plants and then the inoculatedplants were collected and dissected for analysis with tissuesexcised in the following order: heads, FLNs and crowns.Plant tissues were surface sterilised with 70% EtOH forat least 1min, air dried, then cut in half and one half ofthe dissected explant placed cut side down onto SNAsupplemented with 100 ppm or 50 ppm hygromycin forFgT21 or FpT2 inoculated plants, respectively. The otherhalf of each tissue sample was reserved for DON analysis.Plated tissues were incubated for up to 4 weeks and thefrequency of recovery of fungal isolates recorded. Fungalisolates were retained for analysis by the polymerase chainreaction (PCR). In the USA an experiment, similar to thatdescribed above, was subsequently conducted using theUSA strains of Fg with 39–41 plants inoculated with eachstrain. At maturity plants were sampled in a similarmanner except that fungal isolations were conducted usingKomada medium [38] to select for Fusarium isolates andeach plant was scored for either the presence or absence offungus as well as for disease symptoms. The meanfrequencies of recovery of Fusarium from each pool ofreplicate plants inoculated with specific USA strains andthat of controls were analysed statistically using the Fisherexact test.
2.7. Microscopic analysis
Wheat plants (cv. Kennedy) were inoculated with Fg
(CS3005) at the stem base using the method of Mitter et al.[39], and sampled at 8 weeks post-inoculation and handsections prepared from the site of inoculation (internode 1)and the next upper non-inoculated internode and sectionsstained with 0.05% toluidine blue in sodium benzoate pH4.4 and rinsed with water. Standard light microscopy wasused and in some instances final micrographs werecompiled by combining images obtained from multiplefocal planes through a section using the Auto-Montagesoftware package of Syncroscopy (Cambridge, UK).
2.8. Mycotoxin measurements
DON assays were performed as described by Mirocha etal. [40], except that the samples were ground with a pestlein a mortar chilled with liquid nitrogen. Acid washed sandwas used in the Australian experiments to aid the grindingprocess. Briefly, DON was extracted from the sampledtissues by placing the ground tissue into a glass centifugetube and adding 1.5ml of acetonitrile:water (84:16 v:v)solution to vials which were then placed on a rotary shaker

ARTICLE IN PRESS
0
500
1000
1500
2000
2500
3000
3500
Rela
tive T
ri5 e
xpre
ssio
n
0.000
0.200
0.400
0.600
0.800
1.000
1.200
Fu
ng
al /
wheat
18S
rR
NA
0
100
200
300
400
500
600
FH1 FH7 FH14 CR3 CR14 CR28
DO
N p
pm
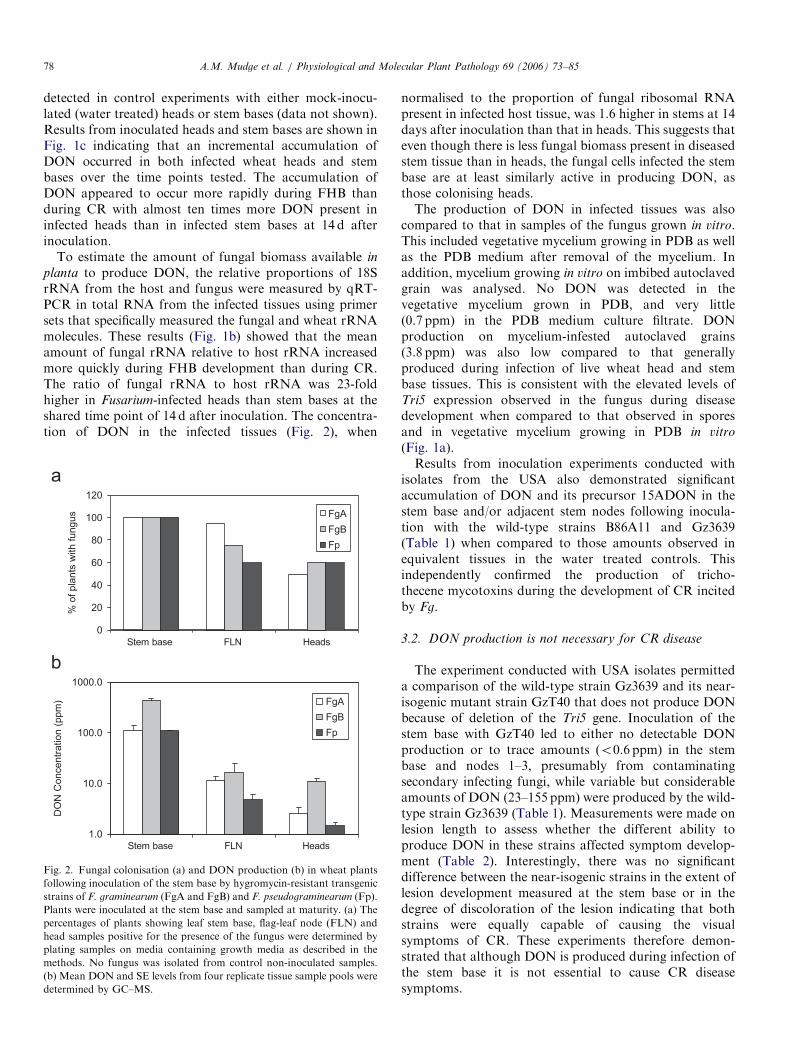
Fig. 1. A comparison of Tri5 gene expression (a), fungal biomass
production (b) and DON production (c) during FHB and CR disease
development caused by Fg. (a) Tri5 gene expression was determined
relative to fungal 18S rRNA using qRT-PCR and values normalised to
that of mycelium cultured in PDB. (b) Fungal and wheat 18S ribosomal
RNA species were measured by qRT-PCR to estimate relative fungal
biomass. (c) DON was assayed using GC–MS with no DON found in
mock inoculated controls. Means and SE values for FHB diseased head
tissue at 1, 7 and 14 days after inoculation of the heads (FH1, FH7 and
FH14, respectively) and CR diseased stem base tissue at 3, 14 and 28 days
after inoculation of the stem base (CR3, CR14 and CR28, respectively) in
three independent experiments are shown.
A.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–85 77
for 2 h. The extraction fluid was then passed through aspecially designed column (one part C18 packing materialto three parts aluminum oxide in a 1mm tuberculin syringe[41] and 1ml was removed and placed in a 2-mm screw topvial and evaporated to dryness with nitrogen. DON in theextract was derivatised to its trimethylsilyl ether derivativefor GC/MS analysis [40]. A 1 ml sample was injected into acombination gas chromatograph and mass spectrometer(Model QP2010; Shimadzu Ltd., Kyoto, Japan). Thepresence of DON was verified based on retention timesand ion monitoring (m/z). DON concentrations weredetermined by comparing peak areas to known standardsincluded at regular intervals and adjusted according tosample weights. In some assays measurements were alsomade for 3-acetyl-deoxynivalenol (3ADON) and 15-acetyl-deoxynivalenol (15ADON). DON production was alsoexamined on wheat grain culture. Twenty grams of wheatseed (cv. Kennedy) was place in a 250ml flask, soaked inwater overnight, drained, autoclaved, and inoculated withone 6mm plug of mycelium growing on SNA. Theinoculated grain culture was incubated at room tempera-ture for 10 days, air dried and processed for DON asdescribed above. Statistical analysis of DON and 15ADONlevels used the non-parametric methods of Kruskal–WallisANOVA and Mann–Whitney pairwise contrasts.
3. Results
3.1. Tri5 gene expression and DON production during CR
disease
Quantitative real-time PCR (qRT-PCR) was used tomeasure the abundance of transcripts of the Tri5 genefollowing inoculation of the stem base of cv. Kennedy bythe Australian wild-type Fg strain CS3040 and during thesubsequent development of CR (Fig. 1a). The qRT-PCRassay was performed on RNA extracted from stem tissuessampled at 3, 14 and 28 d after inoculation. To compareTri5 expression with that occurring during FHB develop-ment, RNA from head tissues were also sampled at 1, 7 and14 d after inoculation of heads, also with wild-type Fg.These infection stages were tested because they representedearly, mid and late time points of symptom developmentfor the respective diseases. For comparative purposes, theexpression of Tri5 was determined in spores (the inoculumsource) and vegetative mycelium-cultured axenically in arich media (PDB). Transcript levels of the Tri5 gene in eachsample were normalised to those of fungal 18S rRNAmeasured using primers that distinguished fungal fromplant 18S rRNA. From these results (Fig. 1a), it can beseen that Tri5 was expressed during the progression of bothFHB and CR. The mean abundance of Tri5 transcripts washigher in Fg throughout the progress of CR than at any ofthe points measured for FHB and was significantlydifferent at the shared time point of 14 days followinginoculation where direct comparisons can be made(P ¼o0.05, t-test). Interestingly, the expression of Tri5
peaked at the mid time point of FHB development and wasat its lowest in heads 14 d after inoculation. In contrast,Tri5 expression remained fairly constant throughout CRdisease development. It was also evident from this analysisthat Tri5 expression was much higher in the fungus duringboth CR and FHB development than in either vegetativemycelium growing in rich media which had a relative valueof 1 compared to that of the infected plant samples shownin Fig. 1a and in spores which had a relative value of 8.To compare trichothecene mycotoxin levels produced
during the development of FHB and CR, DON wasassayed in samples that were used for the qRT-PCRanalysis of Tri5 expression described above. No DON was
ARTICLE IN PRESSA.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–8578
detected in control experiments with either mock-inocu-lated (water treated) heads or stem bases (data not shown).Results from inoculated heads and stem bases are shown inFig. 1c indicating that an incremental accumulation ofDON occurred in both infected wheat heads and stembases over the time points tested. The accumulation ofDON appeared to occur more rapidly during FHB thanduring CR with almost ten times more DON present ininfected heads than in infected stem bases at 14 d afterinoculation.
To estimate the amount of fungal biomass available in
planta to produce DON, the relative proportions of 18SrRNA from the host and fungus were measured by qRT-PCR in total RNA from the infected tissues using primersets that specifically measured the fungal and wheat rRNAmolecules. These results (Fig. 1b) showed that the meanamount of fungal rRNA relative to host rRNA increasedmore quickly during FHB development than during CR.The ratio of fungal rRNA to host rRNA was 23-foldhigher in Fusarium-infected heads than stem bases at theshared time point of 14 d after inoculation. The concentra-tion of DON in the infected tissues (Fig. 2), when
1.0
10.0
100.0
1000.0
DO
N C
oncentr
ation (
pp
m) FgA
FgB
Fp
0
20
40
60
80
100
120
Stem base FLN Heads
Stem base FLN Heads
% o
f p
lan
ts w
ith
fu
ng
us FgA
FgB
Fp
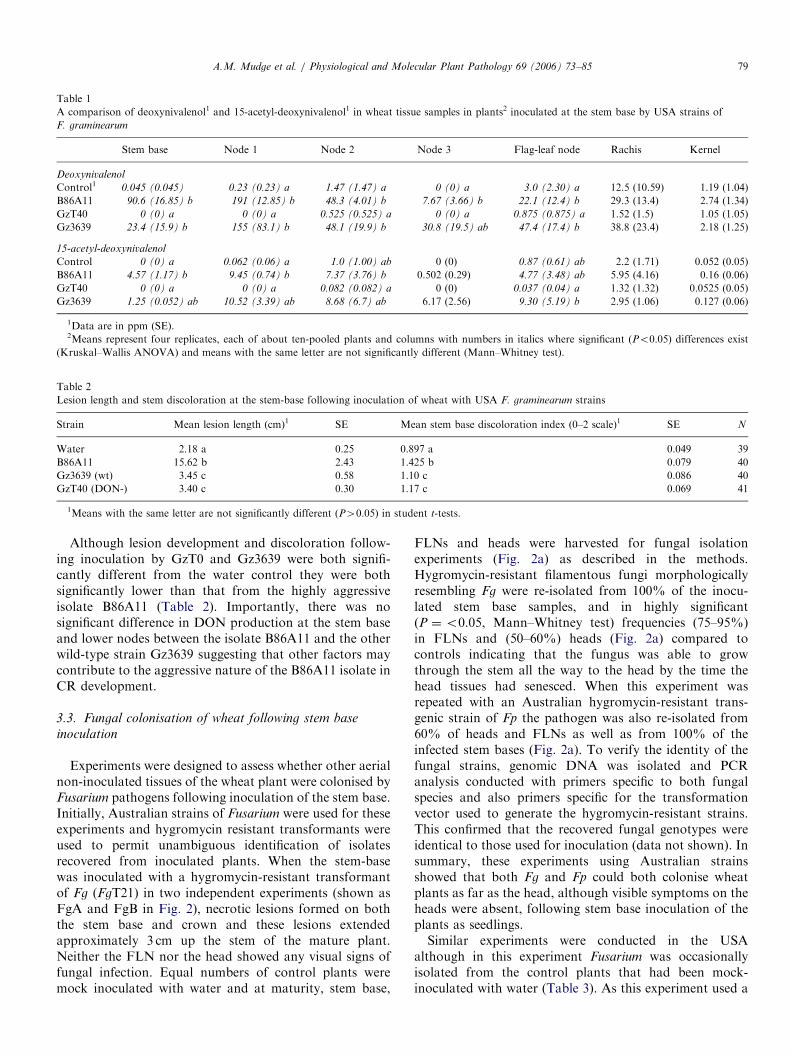
Fig. 2. Fungal colonisation (a) and DON production (b) in wheat plants
following inoculation of the stem base by hygromycin-resistant transgenic
strains of F. graminearum (FgA and FgB) and F. pseudograminearum (Fp).
Plants were inoculated at the stem base and sampled at maturity. (a) The
percentages of plants showing leaf stem base, flag-leaf node (FLN) and
head samples positive for the presence of the fungus were determined by
plating samples on media containing growth media as described in the
methods. No fungus was isolated from control non-inoculated samples.
(b) Mean DON and SE levels from four replicate tissue sample pools were
determined by GC–MS.
normalised to the proportion of fungal ribosomal RNApresent in infected host tissue, was 1.6 higher in stems at 14days after inoculation than that in heads. This suggests thateven though there is less fungal biomass present in diseasedstem tissue than in heads, the fungal cells infected the stembase are at least similarly active in producing DON, asthose colonising heads.The production of DON in infected tissues was also
compared to that in samples of the fungus grown in vitro.This included vegetative mycelium growing in PDB as wellas the PDB medium after removal of the mycelium. Inaddition, mycelium growing in vitro on imbibed autoclavedgrain was analysed. No DON was detected in thevegetative mycelium grown in PDB, and very little(0.7 ppm) in the PDB medium culture filtrate. DONproduction on mycelium-infested autoclaved grains(3.8 ppm) was also low compared to that generallyproduced during infection of live wheat head and stembase tissues. This is consistent with the elevated levels ofTri5 expression observed in the fungus during diseasedevelopment when compared to that observed in sporesand in vegetative mycelium growing in PDB in vitro
(Fig. 1a).Results from inoculation experiments conducted with
isolates from the USA also demonstrated significantaccumulation of DON and its precursor 15ADON in thestem base and/or adjacent stem nodes following inocula-tion with the wild-type strains B86A11 and Gz3639(Table 1) when compared to those amounts observed inequivalent tissues in the water treated controls. Thisindependently confirmed the production of tricho-thecene mycotoxins during the development of CR incitedby Fg.
3.2. DON production is not necessary for CR disease
The experiment conducted with USA isolates permitteda comparison of the wild-type strain Gz3639 and its near-isogenic mutant strain GzT40 that does not produce DONbecause of deletion of the Tri5 gene. Inoculation of thestem base with GzT40 led to either no detectable DONproduction or to trace amounts (o0.6 ppm) in the stembase and nodes 1–3, presumably from contaminatingsecondary infecting fungi, while variable but considerableamounts of DON (23–155 ppm) were produced by the wild-type strain Gz3639 (Table 1). Measurements were made onlesion length to assess whether the different ability toproduce DON in these strains affected symptom develop-ment (Table 2). Interestingly, there was no significantdifference between the near-isogenic strains in the extent oflesion development measured at the stem base or in thedegree of discoloration of the lesion indicating that bothstrains were equally capable of causing the visualsymptoms of CR. These experiments therefore demon-strated that although DON is produced during infection ofthe stem base it is not essential to cause CR diseasesymptoms.
ARTICLE IN PRESS
Table 1
A comparison of deoxynivalenol1 and 15-acetyl-deoxynivalenol1 in wheat tissue samples in plants2 inoculated at the stem base by USA strains of
F. graminearum
Stem base Node 1 Node 2 Node 3 Flag-leaf node Rachis Kernel
Deoxynivalenol
Control1 0.045 (0.045) 0.23 (0.23) a 1.47 (1.47) a 0 (0) a 3.0 (2.30) a 12.5 (10.59) 1.19 (1.04)
B86A11 90.6 (16.85) b 191 (12.85) b 48.3 (4.01) b 7.67 (3.66) b 22.1 (12.4) b 29.3 (13.4) 2.74 (1.34)
GzT40 0 (0) a 0 (0) a 0.525 (0.525) a 0 (0) a 0.875 (0.875) a 1.52 (1.5) 1.05 (1.05)
Gz3639 23.4 (15.9) b 155 (83.1) b 48.1 (19.9) b 30.8 (19.5) ab 47.4 (17.4) b 38.8 (23.4) 2.18 (1.25)
15-acetyl-deoxynivalenol
Control 0 (0) a 0.062 (0.06) a 1.0 (1.00) ab 0 (0) 0.87 (0.61) ab 2.2 (1.71) 0.052 (0.05)
B86A11 4.57 (1.17) b 9.45 (0.74) b 7.37 (3.76) b 0.502 (0.29) 4.77 (3.48) ab 5.95 (4.16) 0.16 (0.06)
GzT40 0 (0) a 0 (0) a 0.082 (0.082) a 0 (0) 0.037 (0.04) a 1.32 (1.32) 0.0525 (0.05)
Gz3639 1.25 (0.052) ab 10.52 (3.39) ab 8.68 (6.7) ab 6.17 (2.56) 9.30 (5.19) b 2.95 (1.06) 0.127 (0.06)
1Data are in ppm (SE).2Means represent four replicates, each of about ten-pooled plants and columns with numbers in italics where significant (Po0.05) differences exist
(Kruskal–Wallis ANOVA) and means with the same letter are not significantly different (Mann–Whitney test).
Table 2
Lesion length and stem discoloration at the stem-base following inoculation of wheat with USA F. graminearum strains
Strain Mean lesion length (cm)1 SE Mean stem base discoloration index (0–2 scale)1 SE N
Water 2.18 a 0.25 0.897 a 0.049 39
B86A11 15.62 b 2.43 1.425 b 0.079 40
Gz3639 (wt) 3.45 c 0.58 1.10 c 0.086 40
GzT40 (DON-) 3.40 c 0.30 1.17 c 0.069 41
1Means with the same letter are not significantly different (P40.05) in student t-tests.
A.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–85 79
Although lesion development and discoloration follow-ing inoculation by GzT0 and Gz3639 were both signifi-cantly different from the water control they were bothsignificantly lower than that from the highly aggressiveisolate B86A11 (Table 2). Importantly, there was nosignificant difference in DON production at the stem baseand lower nodes between the isolate B86A11 and the otherwild-type strain Gz3639 suggesting that other factors maycontribute to the aggressive nature of the B86A11 isolate inCR development.
3.3. Fungal colonisation of wheat following stem base
inoculation
Experiments were designed to assess whether other aerialnon-inoculated tissues of the wheat plant were colonised byFusarium pathogens following inoculation of the stem base.Initially, Australian strains of Fusarium were used for theseexperiments and hygromycin resistant transformants wereused to permit unambiguous identification of isolatesrecovered from inoculated plants. When the stem-basewas inoculated with a hygromycin-resistant transformantof Fg (FgT21) in two independent experiments (shown asFgA and FgB in Fig. 2), necrotic lesions formed on boththe stem base and crown and these lesions extendedapproximately 3 cm up the stem of the mature plant.Neither the FLN nor the head showed any visual signs offungal infection. Equal numbers of control plants weremock inoculated with water and at maturity, stem base,
FLNs and heads were harvested for fungal isolationexperiments (Fig. 2a) as described in the methods.Hygromycin-resistant filamentous fungi morphologicallyresembling Fg were re-isolated from 100% of the inocu-lated stem base samples, and in highly significant(P ¼o0.05, Mann–Whitney test) frequencies (75–95%)in FLNs and (50–60%) heads (Fig. 2a) compared tocontrols indicating that the fungus was able to growthrough the stem all the way to the head by the time thehead tissues had senesced. When this experiment wasrepeated with an Australian hygromycin-resistant trans-genic strain of Fp the pathogen was also re-isolated from60% of heads and FLNs as well as from 100% of theinfected stem bases (Fig. 2a). To verify the identity of thefungal strains, genomic DNA was isolated and PCRanalysis conducted with primers specific to both fungalspecies and also primers specific for the transformationvector used to generate the hygromycin-resistant strains.This confirmed that the recovered fungal genotypes wereidentical to those used for inoculation (data not shown). Insummary, these experiments using Australian strainsshowed that both Fg and Fp could both colonise wheatplants as far as the head, although visible symptoms on theheads were absent, following stem base inoculation of theplants as seedlings.Similar experiments were conducted in the USA
although in this experiment Fusarium was occasionallyisolated from the control plants that had been mock-inoculated with water (Table 3). As this experiment used a

ARTICLE IN PRESS
Table 3
Systemic colonisation of wheat plants demonstrated by recovery of Fusarium isolates from tissue samples of wheat following inoculation at the stem base
with USA strains of F. graminearum
Sample N Stem Base Node 1 Node 2 Flag-leaf node Rachis Kernel
% Plants where Fusarium isolates were obtained from each explant1
Control 39 25.6 12.8 17.9 10.3 7.7 2.6
B86A11 40 95 97.5 95 35 20 20.5
GzT40 40 78 90.2 87.8 17.1 10 4.9
Gz3639 41 52.5 62.5 77.5 37.5 17.5 17.5
Pairwise comparison P-values1
B86A11 vs. control 6.14E�11 7.3E–16 5.08E–13 0.014 0.029
Gz3639 vs. control 0.021 2.3E–06 1.11E–07 0.0075 0.057
GzT40 vs. control 4.7E�06 1.04E–12 2.32E–10 0.52 1
GzT40 vs. Gz3639 0.020 7.77E–03 0.253 0.048 0.088
1Columns where significant differences (Po0.05) between treatments were detected using the Fisher exact test for two-way tables are in italics and the
P-values shown for selected pairwise comparisons.
A.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–8580
smaller growth chamber some cross-plant contaminationwas not unexpected. Statistical analysis of the recoveryrates from inoculated plants in comparison to controlsrevealed significantly higher frequencies of recovery of thewild-type fungal strains B86A11 and Gz3639 from theFLN and kernels of plants inoculated at the stem basetherefore confirming the findings of the experiments withthe Australian strains (Table 3).
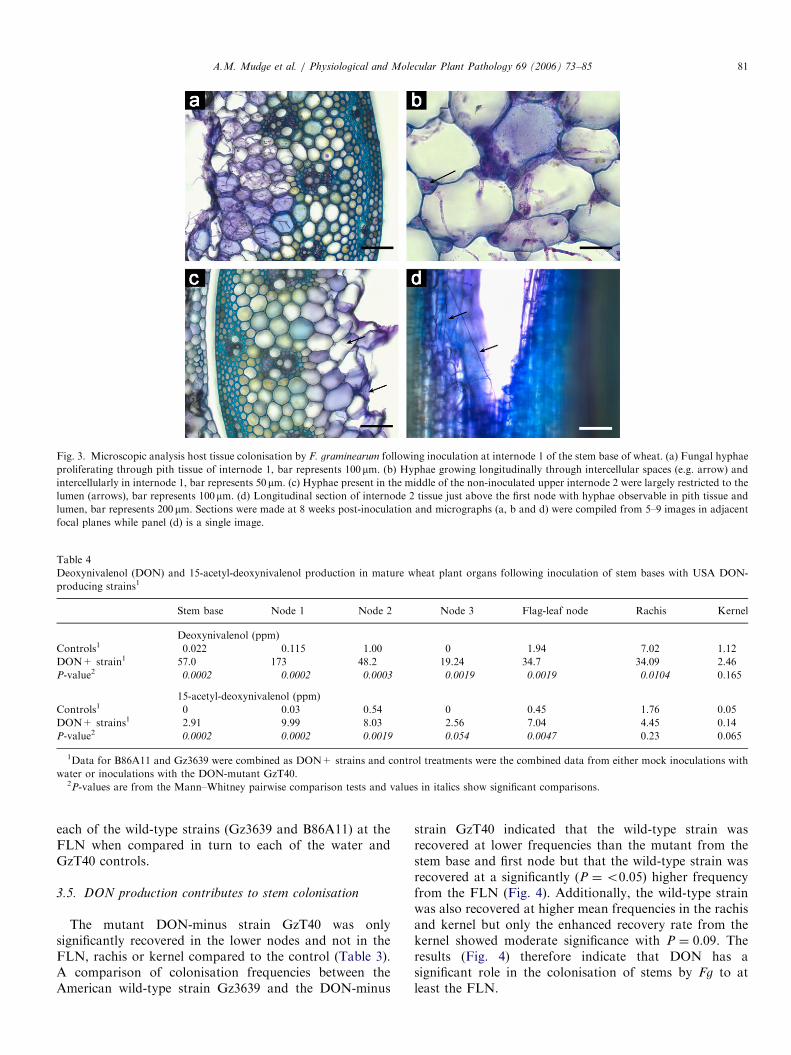
To assess which tissues in the host were colonised ininoculated and upper non-inoculated parts of the stem,sections were prepared from the site of inoculation oninternode 1 and at the next upper internode at 8 weeks afterinoculation with an Australian Fg isolate (Fig. 3). Theupper non-inoculated internode was symptomless whensampled. At the site of infection the fungus was primarilylocated in the pith parenchyma with intra and intercellularhyphae being observed (Figs. 3a and b) In the upper non-inoculated internodes the hyphal density was greatlyreduced and the fungus was mainly observed as inter-cellular hyphae in the central stem region in associationwith the pith parenchyma (Fig. 3c). In longitudinal sections(Fig. 3d) long hyphal strands were observed extendingvertically within the lumen. There was very little colonisa-tion of the vascular tissues at the infection site and nohyphae were observed in the vascular bundles in the non-inoculated upper internode.
3.4. DON production in distal non-inoculated tissues
following stem base infection
Because Fusarium was recovered from distal parts of theplant following inoculation of the stem base, DON assayswere performed on the plant samples described above. Inthe experiment conducted using the Australian isolates(Fig. 2b), large amounts of DON, in the range of111–438 ppm, were detected in Fusarium-infected crowns(Fig. 2b). Furthermore, the Australian Fp strain used inthis study appeared to be as competent in DON productionas the Fg strain tested and produced substantial amounts
(111 ppm) of DON at the infection site. This is also thefirst time that the production of DON by Fp in infectedstem bases has been reported. Of most interest was thatDON was also readily detected in the distal non-inoculatedFLNs and the heads of wheat plants inoculated at thestem base by both Australian Fg and Fp strains. Thelevels of DON in the heads of inoculated plants rangedfrom 1 to 11 ppm (Fig. 2b) which were significantly(P ¼o0.05) greater than controls, but were sub-stantially lower than the levels generally detected in FHBfollowing direct inoculation of the heads in this study(Fig. 1c). Plants inoculated at the stem base with Fp
appeared to have lower levels of DON production in theheads of these tillers than those inoculated similarly withFg. No DON was detected in any of the tissue samplesfrom the control mock-inoculated plants in the study inAustralia.In the experiment studying American isolates, just as
Fusarium was recovered from some of the control mock-inoculated plants, these plants also showed low butvariable recoveries of DON and 15ADON. Because onlyfour replicate samples were used for the DON and15ADON analysis from each organ sample, a pooledstatistical approach was taken to increase the N value forcomparisons assessing DON accumulation systemicallyfollowing infection (Table 4). Firstly, to test the premisethat DON and 15ADON are produced in upper non-inoculated organs we compared the combined data fromthe wild-type DON-producing strains B86A11 and Gz3639with the combined control treatments of mock inoculatedand those inoculated with the DON-minus mutant GzT40(Table 4). Both controls act as a measure of cross-plantcontamination by DON producing strains in the experi-ment. This comparison showed that significant (P ¼o0.05) DON was produced in distal plant parts up tothe rachis and significant 15ADON up to the FLN in theplants inoculated with the wild-type strains when com-pared to controls. At the level of individual treatments(Table 1) significant levels of DON were observed from
ARTICLE IN PRESS
Fig. 3. Microscopic analysis host tissue colonisation by F. graminearum following inoculation at internode 1 of the stem base of wheat. (a) Fungal hyphae
proliferating through pith tissue of internode 1, bar represents 100mm. (b) Hyphae growing longitudinally through intercellular spaces (e.g. arrow) and
intercellularly in internode 1, bar represents 50mm. (c) Hyphae present in the middle of the non-inoculated upper internode 2 were largely restricted to the
lumen (arrows), bar represents 100mm. (d) Longitudinal section of internode 2 tissue just above the first node with hyphae observable in pith tissue and
lumen, bar represents 200mm. Sections were made at 8 weeks post-inoculation and micrographs (a, b and d) were compiled from 5–9 images in adjacent
focal planes while panel (d) is a single image.
Table 4
Deoxynivalenol (DON) and 15-acetyl-deoxynivalenol production in mature wheat plant organs following inoculation of stem bases with USA DON-
producing strains1
Stem base Node 1 Node 2 Node 3 Flag-leaf node Rachis Kernel
Deoxynivalenol (ppm)
Controls1 0.022 0.115 1.00 0 1.94 7.02 1.12
DON+ strain1 57.0 173 48.2 19.24 34.7 34.09 2.46
P-value2 0.0002 0.0002 0.0003 0.0019 0.0019 0.0104 0.165
15-acetyl-deoxynivalenol (ppm)
Controls1 0 0.03 0.54 0 0.45 1.76 0.05
DON+ strains1 2.91 9.99 8.03 2.56 7.04 4.45 0.14
P-value2 0.0002 0.0002 0.0019 0.054 0.0047 0.23 0.065
1Data for B86A11 and Gz3639 were combined as DON+ strains and control treatments were the combined data from either mock inoculations with
water or inoculations with the DON-mutant GzT40.2P-values are from the Mann–Whitney pairwise comparison tests and values in italics show significant comparisons.
A.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–85 81
each of the wild-type strains (Gz3639 and B86A11) at theFLN when compared in turn to each of the water andGzT40 controls.
3.5. DON production contributes to stem colonisation
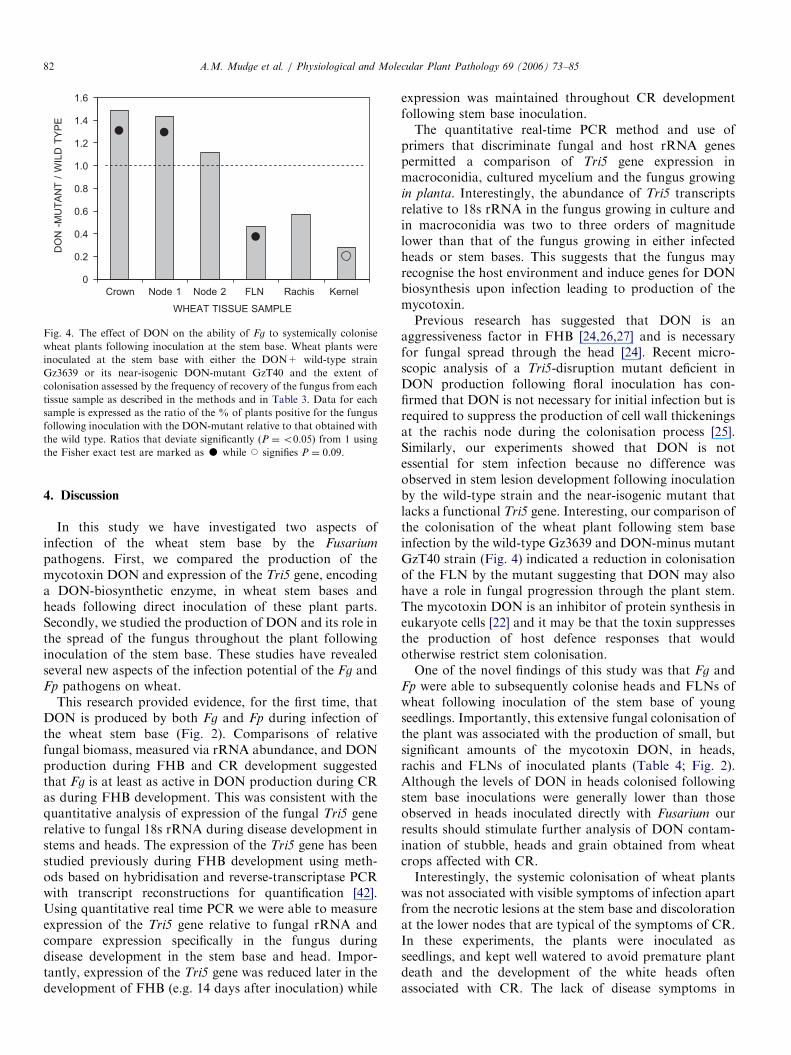
The mutant DON-minus strain GzT40 was onlysignificantly recovered in the lower nodes and not in theFLN, rachis or kernel compared to the control (Table 3).A comparison of colonisation frequencies between theAmerican wild-type strain Gz3639 and the DON-minus
strain GzT40 indicated that the wild-type strain wasrecovered at lower frequencies than the mutant from thestem base and first node but that the wild-type strain wasrecovered at a significantly (P ¼o0.05) higher frequencyfrom the FLN (Fig. 4). Additionally, the wild-type strainwas also recovered at higher mean frequencies in the rachisand kernel but only the enhanced recovery rate from thekernel showed moderate significance with P ¼ 0.09. Theresults (Fig. 4) therefore indicate that DON has asignificant role in the colonisation of stems by Fg to atleast the FLN.
ARTICLE IN PRESS
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Crown Node 1 Node 2 FLN Rachis Kernel
WHEAT TISSUE SAMPLE
DO
N -
MU
TA
NT
/ W
ILD
TY
PE
Fig. 4. The effect of DON on the ability of Fg to systemically colonise
wheat plants following inoculation at the stem base. Wheat plants were
inoculated at the stem base with either the DON+ wild-type strain
Gz3639 or its near-isogenic DON-mutant GzT40 and the extent of
colonisation assessed by the frequency of recovery of the fungus from each
tissue sample as described in the methods and in Table 3. Data for each
sample is expressed as the ratio of the % of plants positive for the fungus
following inoculation with the DON-mutant relative to that obtained with
the wild type. Ratios that deviate significantly (P ¼o0.05) from 1 using
the Fisher exact test are marked as K while J signifies P ¼ 0.09.
A.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–8582
4. Discussion
In this study we have investigated two aspects ofinfection of the wheat stem base by the Fusarium
pathogens. First, we compared the production of themycotoxin DON and expression of the Tri5 gene, encodinga DON-biosynthetic enzyme, in wheat stem bases andheads following direct inoculation of these plant parts.Secondly, we studied the production of DON and its role inthe spread of the fungus throughout the plant followinginoculation of the stem base. These studies have revealedseveral new aspects of the infection potential of the Fg andFp pathogens on wheat.
This research provided evidence, for the first time, thatDON is produced by both Fg and Fp during infection ofthe wheat stem base (Fig. 2). Comparisons of relativefungal biomass, measured via rRNA abundance, and DONproduction during FHB and CR development suggestedthat Fg is at least as active in DON production during CRas during FHB development. This was consistent with thequantitative analysis of expression of the fungal Tri5 generelative to fungal 18s rRNA during disease development instems and heads. The expression of the Tri5 gene has beenstudied previously during FHB development using meth-ods based on hybridisation and reverse-transcriptase PCRwith transcript reconstructions for quantification [42].Using quantitative real time PCR we were able to measureexpression of the Tri5 gene relative to fungal rRNA andcompare expression specifically in the fungus duringdisease development in the stem base and head. Impor-tantly, expression of the Tri5 gene was reduced later in thedevelopment of FHB (e.g. 14 days after inoculation) while
expression was maintained throughout CR developmentfollowing stem base inoculation.The quantitative real-time PCR method and use of
primers that discriminate fungal and host rRNA genespermitted a comparison of Tri5 gene expression inmacroconidia, cultured mycelium and the fungus growingin planta. Interestingly, the abundance of Tri5 transcriptsrelative to 18s rRNA in the fungus growing in culture andin macroconidia was two to three orders of magnitudelower than that of the fungus growing in either infectedheads or stem bases. This suggests that the fungus mayrecognise the host environment and induce genes for DONbiosynthesis upon infection leading to production of themycotoxin.Previous research has suggested that DON is an
aggressiveness factor in FHB [24,26,27] and is necessaryfor fungal spread through the head [24]. Recent micro-scopic analysis of a Tri5-disruption mutant deficient inDON production following floral inoculation has con-firmed that DON is not necessary for initial infection but isrequired to suppress the production of cell wall thickeningsat the rachis node during the colonisation process [25].Similarly, our experiments showed that DON is notessential for stem infection because no difference wasobserved in stem lesion development following inoculationby the wild-type strain and the near-isogenic mutant thatlacks a functional Tri5 gene. Interesting, our comparison ofthe colonisation of the wheat plant following stem baseinfection by the wild-type Gz3639 and DON-minus mutantGzT40 strain (Fig. 4) indicated a reduction in colonisationof the FLN by the mutant suggesting that DON may alsohave a role in fungal progression through the plant stem.The mycotoxin DON is an inhibitor of protein synthesis ineukaryote cells [22] and it may be that the toxin suppressesthe production of host defence responses that wouldotherwise restrict stem colonisation.One of the novel findings of this study was that Fg and
Fp were able to subsequently colonise heads and FLNs ofwheat following inoculation of the stem base of youngseedlings. Importantly, this extensive fungal colonisation ofthe plant was associated with the production of small, butsignificant amounts of the mycotoxin DON, in heads,rachis and FLNs of inoculated plants (Table 4; Fig. 2).Although the levels of DON in heads colonised followingstem base inoculations were generally lower than thoseobserved in heads inoculated directly with Fusarium ourresults should stimulate further analysis of DON contam-ination of stubble, heads and grain obtained from wheatcrops affected with CR.Interestingly, the systemic colonisation of wheat plants
was not associated with visible symptoms of infection apartfrom the necrotic lesions at the stem base and discolorationat the lower nodes that are typical of the symptoms of CR.In these experiments, the plants were inoculated asseedlings, and kept well watered to avoid premature plantdeath and the development of the white heads oftenassociated with CR. The lack of disease symptoms in

ARTICLE IN PRESSA.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–85 83
colonised FLNs and heads and in other aerial plant organsabove the lesion at the crown and stem base suggested thatthe pathogen had colonised the host plant as might anendophyte or biotroph rather than as a necrotroph asobserved at the infection site on the stem base wherenecrosis is evident. Previous studies have indicated that Fp
occasionally reaches the FLN in plants with CR [7], but thepresence of the pathogen in the heads following stem baseinoculation has not been reported. Similarly F. culmorum
has been reported to systemically colonise the aboveground tissues of wheat plants [43,44]. There are somesignificant differences between these previous studies andour study. Firstly, in earlier studies wheat seedlings wereinoculated indirectly with the inoculum applied to the soil[19,43,45] or by wound inoculation of upper nodes ofmaturing plants while we inoculated the surface of stembase of emerged seedlings directly. However, we believethat one important difference was that in the study ofAustralian isolates we used hygromycin-resistant Fg andFp strains. This was done primarily so that an unambig-uous identification of recovered isolates could be made. Asmost fungi and bacteria are very sensitive to hygromycinthis antibiotic was included in the isolation plates on whichplant explants were incubated to track the presence of thehygromycin-resistant fungal strains. The use of hygromy-cin also allowed for longer incubation periods than mightusually be employed in such experiments by suppressinggrowth of other saprophytes or endophytes. This appearedto be important because in many instances it took severaldays for the Fusarium to appear in culture from head andFLN tissue excised from inoculated plants.
This paper describes similar findings from experimentsusing Australian and USA isolates of Fg studying fungalcolonisation and DON production following stem baseinoculation. The two experiments strengthen the conclu-sions which indicate that Fg has a flexible pathogeniclifestyle and can produce DON at high levels in multipletissues of wheat. It is evident from the results of experimentconducted in St. Paul with the USA isolates that higherbackground levels of fungal contamination and DONproduction were observed in the mock-inoculated controlplants than were observed during the Australian studywhere no isolations were made or DON detected in thecontrols. This may be because in the USA study wild-typestrains were used and therefore hygromycin could not beused in the selection media to limit the recovery ofcontaminating fungi; secondly the experiments conductedin the USA used smaller growth chambers resulting incrowded plant growth conditions which may have facili-tated cross-plant fungal transfer. It is also possible thatseed-borne Fg may have resulted in a background level ofinfection. Nonetheless the replication in these experimentswas sufficient to demonstrate significant recoveries of thepathogen in the inoculated plants compared to controls,from upper plant organs, including kernels (Table 3), andalso significant DON levels in FLN and rachis (Tables 1and 4) compared to controls for DON production.
Exactly how Fg and Fp colonise the FLN and headfollowing inoculation of the stem base is not completelyclear from our research. We can rule out the possibility thataerial ascospore dispersal from the infected stem baseoccurred. While ascospores are aerially dispersed theAustralian isolates of both Fg and Fp used in theseexperiments produce few perithecia [4] and no perithecia orascospores were observed on the infected stem tissue in allthe experiments conducted herein. It is possible thatmacroconidia were produced and dispersed, but it isunlikely that the environmental conditions favoured thegermination of these spores or the infection of aerial planttissues. Fusarium isolates were not recovered from control(non-inoculated) plants that were co-located amongst theinoculated plants in the growth chamber in Australia andwere recovered at significantly lower frequencies in theUSA experiment. Care was taken to water plants via thetrays in which pots were placed to avoid splash dispersal ofmacroconidia. Our initial microscopic analysis of non-inoculated upper internodes has provided some evidenceon how the fungus colonised the stem and suggests that thefungus grows primarily in the pith parenchyma at itsinterface with the lumen of the culm (Fig. 3). The lumenappears to provide a pathway for vertical growth that isfree of host structural barriers while the surroundingparenchyma cells provide a potential nutrient source andhumid environment. When Fg colonises the stem down-wards from an inoculated head during FHB disease [46] itappears also to spread via the pith parenchyma and lumen,but invades the vascular system as well. Microscopicstudies of F. culmorum colonising wheat plants havedemonstrated the presence of fungal hyphae in the vascularsystem at nodes distant from the point of inoculation [43].In contrast to these reports we found no evidence ofhyphae of Fg in vascular cells in the non-inoculated uppernodes and infected vascular cells were also infrequentcompared to parenchyma cells in the lesion at the site ofinoculation (Fig. 3). The vascular bundles of the lower stembase of wheat are surrounded by a layer of lignifiedparenchyma cells and these may be difficult for the Fg
pathogen to penetrate from the adjacent pith parenchyma.In summary, this report demonstrates that under
controlled conditions Fg and Fp can extensively colonisewheat plants following stem-based inoculation and pro-duce DON in distal, non-inoculated tissues where itappears to assist colonisation. This suggests that thesepathogens have broader infection strategies than pre-viously recognised. From an industry perspective it wouldbe interesting to test asymptomatic heads and stubbleproduced in CR-affected wheat fields to determine thepresence and extent of mycotoxin contamination.
Acknowledgements
We are grateful for the support of a postdoctoralfellowship for A.M. Mudge from the Ken and YasukoMyer Plant Science Foundation, a Visiting Fellowship for

ARTICLE IN PRESSA.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–8584
R. Dill-Macky from the Grains Research and Develop-ment Corporation and support for D. Gardiner from theCooperative Research Centre for Tropical Plant Protec-tion. Research in Ruth Dill-Macky’s laboratory at theUniversity of Minnesota was funded by the US Wheat andBarley Scab Initiative and the Minnesota Small GrainsInitiative. We are grateful for the help and advice of Drs.Vivek Mitter, Kemal Kazan and Sukumar Chakraborty,for technical assistance from Amar Elakkad and KarenWennberg. Dr. Alec Zwart of CSIRO Mathematics andInformation Sciences assisted with the statistical analyses.
References
[1] Champei A, Dore T, Fourbet JF. Fusarium head blight: epidemio-
logical origin of the effects of cultural practices on head blight attacks
and the production of mycotoxins by Fusarium in wheat grains. Plant
Sci 2004;166:1389–415.
[2] Goswami RS, Kistler HC. Heading for disaster: Fusarium graminear-
um on cereal crops. Mol Plant Pathol 2004;5:515–25.
[3] McMullen M, Jones R, Gallenberg D. Scab of wheat and barley: a re-
emerging disease of devastating impact. Plant Dis 1997;81:1340–8.
[4] Akinsanmi OA, Mitter V, Simpfendorfer S, Backhouse D, Chakra-
borty S. Identity and pathogenicity of Fusarium spp. isolated from
wheat fields in Queensland and northern New South Wales. Aust
J Agric Res 2004;55:97–107.
[5] Burgess LW, Klein TA, Bryden WL, Tobi NF. Head blight of wheat
caused by Fusarium graminearum Group 1 in New South Wales in
1983. Australas Plant Pathol 1987;16:72–8.
[6] Southwell RJ, Moore KJ, Manning W, Hayman PT. An outbreak of
Fusarium head blight of durum wheat on the Liverpool Plains in
northern New South Wales in 1999. Australas Plant Pathol 2003;
32:465–71.
[7] Burgess LW, Backhouse D, Summerell BA, Swan LJ. Crown rot of
wheat. In: Summerell BA, Leslie JF, Backhouse D, Bryden WL,
Burgess LW, editors. Fusarium. St. Paul, MN: APS Press; 2001.
p. 271–94.
[8] Burgess LW, Wearing AH, Toussoun TA. Surveys of Fusaria
associated with crown rot of wheat in eastern Australia. Aust
J Agric Res 1975;26:791–9.
[9] Williams KJ, Dennis JI, Smyl C, Wallwork H. The application of
species-specific assays based on the polymerase chain reaction to
analyse Fusarium crown rot of durum wheat. Australas Plant Pathol
2002;31:119–29.
[10] Braun HJ, Mergoum M, Morgounov A, Nicol J. Adaptation of
Winter Wheat to Central and West Asia. CIMMYT research results,
2004, /http://www.cimmyt.org/research/wheat/map/research_results/
reshighlights/pdfs/reshigh_adaptwinter.pdfS.
[11] Smiley RW, Patterson L-M. Pathogenic fungi associated with
Fusarium foot rot of winter wheat in semiarid Pacific Northwest
USA. Plant Dis 1996;80:944–9.
[12] Backhouse D, Burgess LW. Climatic analysis of the distribution of
Fusarium graminearum, F. pseudograminearum and F. culmorum on
cereals in Australia. Australas Plant Pathol 2002;31:321–7.
[13] Francis RG, Burgess LW. Characteristics of two populations of
Fusarium roseum ‘Graminearum’ in eastern Australia. Trans Br
Mycol Soc 1977;68:421–7.
[14] Aoki T, O’Donnell K. Morphological and molecular characterisation
of Fusarium pseudograminearum sp. nov., formerly recognised as
Group 1 population of F. graminearum. Mycologia 1999;91:597–609.
[15] Aoki T, O’Donnell K. Morphological characterisation of Gibberella
coronicola sp. nov., obtained through mating experiments of
Fusarium pseudograminearum. Mycoscience 1999;40:443–53.
[16] O’Donnell K, Ward TJ, Geiser DM, Kistler HC, Aoki T.
Genealogical concordance between the mating type locus and seven
other genes supports formal recognition of nine phylogenetically
distinct species within the Fusarium graminearum clade. Fungal Genet
Biol 2004;41:600–23.
[17] Desmond OJ, Edgar CI, Manners JM, Maclean DJ, Schenk PM,
Kazan K. Methyl jasmonate induced gene expression in wheat delays
symptom development by the crown rot pathogen Fusarium
pseudograminearum. Physiol Mol Plant Pathol 2005;67:171–9.
[18] Trail F, Gaffoor I, Guenther JC, Hallen HE. Using genomics to
understand the disease cycle of the fusarium head blight fungus,
Gibberella zeae (anamorph Fusarium graminearum). Can J Plant
Pathol 2005;27:486–98.
[19] Beddis AL, Burgess LW. The influence of plant water stress on
infection and colonisation of wheat seedlings by Fusarium graminear-
um Group 1. Phytopathology 1992;82:78–83.
[20] Burgess LW, Dodman RL, Pont W, Mayers P. Fusarium diseases of
wheat, maize and grain sorghum in eastern Australia. In: Nelson PA,
Toussoun TA, Cook RJ, editors. Fusarium: Diseases, Biology and
Taxonomy. University Park: Pennsylvania State University Press;
1981. p. 64–76.
[21] Blaney BJ, Dodman RL. Production of the mycotoxins zearalenone,
4-deoxynivalenol and nivalenol by isolates of Fusarium graminearum
Groups 1 and 2 from cereals in Queensland. Aust J Agric Res
1988;39:21–9.
[22] McCormick S. The role of DON in pathogenicity. In: Leonard KJ,
Bushnell WR, editors. Fusarium head blight of wheat and barley. St.
Paul, MN: APS Press; 2003. p. 165–83.
[23] Hohn TM, Desjardins AE. Isolation and gene disruption of the Tox5
gene encoding trichodiene synthase in Gibberella pulicaris. Mol Plant-
Microbe Interact 1992;5:249–56.
[24] Bai GH, Desjardins AE, Plattner RD. Deoxynivalenol non-produ-
cing Fusarium graminearum causes initial infection, but does not
cause disease spread in wheat spikes. Mycopathology 2001;153:91–8.
[25] Jansen C, von Wettstein D, Schafer W, Kogel K-H, Felk A, Maier
FJ. Infection patterns in barley and wheat spikes inoculated with
wild-type and trichodiene synthase gene disrupted Fusarium grami-
nearum. Proc Natl Acad Sci USA 2005;102:16892–7.
[26] Okubara P, Blechl AE, McCormick SP, Alexander NJ, Dill-Macky
R, Hohn TM. Engineering deoxynivalenol metabolism in wheat
through the expression of a fungal trichothecene acetyltransferase
gene. Theor Appl Genet 2002;106:74–83.
[27] Proctor RH, Hohn TM, McCormick SP. Reduced virulence of
Gibberella zeae caused by disruption of a trichothecene toxin
biosynthetic gene. Mol Plant-Microbe Interact 1995;8:593–601.
[28] Scott JB, Chakraborty S. Multilocus sequence analysis of Fusarium
pseudograminearum reveals a single phylogenetic species. Mycol Res
2006, in press.
[29] Evans CK, Xie W, Dill-Macky R, Mirocha CJ. Biosynthesis of
deoxynivalenol in spiklelets of barley inoculated with macroconidia
of Fusarium graminearum. Plant Dis 2000;84:654–60.
[30] Nirenberg H. Untersuchungen uber die morphologische und biolo-
gische Differenzierung in der Fusarium-Sektion Liseola. Mitteilungen
aus der Biologischen Bundesansstalt fur Land und Forestwirtschaft.
Berlin: Dahlem; 1976. p. 169.
[31] Bai GH, Shaner G. Variation in Fusarium graminearum and cultivar
resistance to wheat scab. Plant Dis 1996;80:975–9.
[32] Irelan J, Miao V. Selker EUSmall scale DNA preps for Neurospora
crassa. Fungal Genet Newslett 1993;40:24.
[33] Yoder OC. Genetics of Cochliobolus heterostrophus. Adv Plant Pathol
1988;6:93–122.
[34] Masel AM, Struijk N, McIntyre CLM, Irwin JAG, Manners JM. A
strain specific cyclin homolog in the fungal phytopathogen Colleto-
trichum gloeosporioides. Gene 1993;133:141–5.
[35] Ramakers C, Ruijter JM, Lekanne Deprez RH, Moormann AFM.
Assumption-free analysis of quantitative real-time polymerase chain
reaction (PCR) data. Neurosci Lett 2003;339:62–6.
[36] Punt PJ, Oliver RP, Dingemanse MA, Pouwels PH, van den Hondel
CAMJJ. Transformation of Aspergillus based on the hygromycin B
resistance marker from Escherichia coli. Gene 1987;56:117–24.

ARTICLE IN PRESSA.M. Mudge et al. / Physiological and Molecular Plant Pathology 69 (2006) 73–85 85
[37] Poplawski AM, He C, Irwin JAG, Manners JM. Transfer of an
autonomously replicating vector between vegetatively incompatible
biotypes of Colletotrichum gloeosporioides. Curr Genet 1997;32:66–72.
[38] Komada H. Development of a selective medium for quantitative
isolation of Fusarium oxysporum, from natural soil. Rev Plant Prot
Res 1975;8:114–25.
[39] Mitter V, Zhang MC, Liu CJ, Ghosh R, Ghosh M, Chakraborty S. A
high-throughput glasshouse bioassay to detect crown rot resistance in
wheat germplasm. Plant Pathol 2006;55:433–41.
[40] Mirocha CJ, Kolaczkowski E, Xie W, Yu H, Jelen H. analysis of
deoxynevalenol and its derivatives (batch and single kernel) using gas
chromatography/mass spectrometry. J Agric Food Chem 1998;46:
1414–8.
[41] Tacke BK, Casper HH. Determination of deoxynivalenol in wheat,
barley and malt by column cleanup and gas chromatography with
electron capture detection. J AOAC Int 1996;79:472–5.
[42] Doohan FM, Weston G, Rezanoor HN, Parry DW, Nicholson P.
Development and use of a reverse transcription-PCR assay to study
expression of Tri5 by Fusarium species in vitro and in planta. Appl
Environ Microbiol 1999;65:3850–4.
[43] Clement JA, Parry DW. Stem-base disease and fungal colonisation of
winter wheat grown in compost inoculated with Fusarium culmorum,
F. graminearum and Microdochium nivale. Eur J Plant Pathol 1998;
104:323–30.
[44] Snijders CHA. Systemic fungal growth of Fusarium culmorum in
stems of winter wheat. J Phytopathol 1990;129:133–40.
[45] Liddell CM, Burgess LW, Taylor PWJ. Reproduction of crown rot of
wheat caused by Fusarium graminearum group 1 in the glasshouse.
Plant Dis 1986;70:632–5.
[46] Guenther JC, Trail F. The development and differentiation of
Gibberella zeae (anamorph: Fusarium graminearum) during colonisa-
tion of wheat. Mycologia 2005;97:229–37.
Copyright © 2022 FDOKUMEN




















