
Valuing multiple threatened species and ecological communities in ...
Upload
independentCategory
view
1download
0
1 23
Journal of InsectConservationAn international journal devotedto the conservation of insectsand related invertebrates ISSN 1366-638X J Insect ConservDOI 10.1007/s10841-011-9422-7
A preliminary assessment of changes inplant-dwelling insects when threatenedplants are translocated
Melinda L. Moir, Peter A. Vesk, KarlE. C. Brennan, Lesley Hughes, DavidA. Keith, Michael A. McCarthy, DavidJ. Coates & Sarah Barrett

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media B.V.. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

ORIGINAL PAPER
A preliminary assessment of changes in plant-dwelling insectswhen threatened plants are translocated
Melinda L. Moir • Peter A. Vesk • Karl E. C. Brennan •
Lesley Hughes • David A. Keith • Michael A. McCarthy •
David J. Coates • Sarah Barrett
Received: 28 March 2011 / Accepted: 7 July 2011
� Springer Science+Business Media B.V. 2011
Abstract Translocation of threatened species is a tool
used increasingly to conserve biodiversity, but the suite of
co-dependent species that use the threatened taxa as hosts
can be overlooked. We investigate the preliminary impact
of translocating three threatened plant species on insect
species and the integrity of insect assemblages that depend
on these plants as their hosts. We compare the insect
assemblages between natural populations of the threatened
species, related non-threatened plant species growing wild
near the threatened plants, and threatened plants translo-
cated to another site approximately 40 km away. We used
host breadth models and a coextinction risk protocol to
determine which insect species are potentially host-specific
on the threatened plants, and then assessed these insects’
potential presence at the translocation site. We found that
insect assemblages on naturally-occurring threatened plants
had more individuals, higher species density and higher
species richness than assemblages on translocated plants.
For one plant species, Leucopogon gnaphalioides, species
composition differed significantly between wild and
translocated populations (P \ 0.001). Furthermore, four
insect species that were host-specific to Banksia brownii
and B. montana were not detected on the translocated
plants. Instead, translocated plants supported insect assem-
blages more similar to those of related plant species from the
surrounding area. We conclude that threatened plant trans-
locations that involve seed collection and propagation may
have limited benefit for individual dependent species or the
supported insect assemblage. Additional conservation
actions will be required to maintain the diversity of insect
assemblages and host-dependent relationships.
Keywords Assisted colonization �Conservation planning � Coextinction risk �Host specificity � Plant-insect interactions � Translocation
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10841-011-9422-7) contains supplementarymaterial, which is available to authorized users.
M. L. Moir (&) � P. A. Vesk � M. A. McCarthy
School of Botany, University of Melbourne, Parkville,
VIC 3010, Australia
e-mail: [email protected]
K. E. C. Brennan
Western Australian Department of Environment &
Conservation, PO Box 10173, Kalgoorlie, WA 6430, Australia
L. Hughes
Department of Biological Sciences, Macquarie University,
North Ryde, NSW 2109, Australia
D. A. Keith
NSW National Parks & Wildlife Service, PO Box 1967,
Hurstville, NSW 2220, Australia
D. A. Keith
School of Biological, Earth and Environmental Sciences,
University of New South Wales, Sydney, NSW 2052, Australia
D. J. Coates
Western Australian Department of Environment &
Conservation, Locked Bag 104, Bentley DC,
WA 6983, Australia
S. Barrett
Western Australian Department of Environment &
Conservation, 120 Albany Hwy, Albany, WA 6330, Australia
123
J Insect Conserv
DOI 10.1007/s10841-011-9422-7
Author's personal copy

Introduction
Disturbances such as habitat fragmentation and incursion
by exotic species are occurring at a rapid rate, and threaten
many species with extinction. Rapid transformation in
climate over the next 60–90 years will add to existing
environmental stresses and may extinguish or severely
reduce the populations of many threatened species (e.g.,
McClean et al. 2005; Thuiller et al. 2005; Williams et al.
2007; Wilson and Maclean 2011). The potential for loss of
global biodiversity through co-extinction is high because
many of the organisms currently at risk of extinction act as
hosts to numerous dependent species (Koh et al. 2004;
Moir et al. 2010a, 2011).
As one of the 25 biodiversity hotspots identified by
Myers et al. (2000), the south-west of Australia is partic-
ularly susceptible to loss of species. The south-west is
home to an estimated 8,000 plant species (Hopper and
Gioia 2004), 1,909 of which are listed by the State Gov-
ernment as threatened or in need of conservation actions
(Western Australian Herbarium 2011). Translocation trials
of 16 threatened plant species in the south coast region by
the Western Australian Department of Environment and
Conservation have aimed to alleviate the immediate risk of
extinction primarily due to Phytophthora dieback caused
by Phytophthora cinnamomi and inappropriate fire regimes
(Barrett et al. 2008). The ultimate goal of these translo-
cations is to establish viable self-sustaining populations
and to produce seed to establish further populations in
Phytophthora free areas. For this study, we selected three
of the most threatened species found in the montane
regions of the Stirling Range, which also demonstrated
rapid growth when translocated. These are Leucopogon
gnaphalioides Stschegl. (Ericaceae), Banksia brownii R.Br.
(Proteaceae: Spicigerae), and Banksia montana (A.
S. George) (Proteaceae: Spathulatae), all of which are lis-
ted nationally as Endangered under the Environmental
Protection and Biodiversity Conservation Act 1999, and
are also listed as Critically Endangered under the Western
Australian Wildlife Conservation Act 1953. Global climate
change may detrimentally affect these predominantly
montane species through drying of their habitat (e.g.,
Bureau of Meteorology—Australia (BoM) 2011). We
examined the experimental translocation of these plant
species and assessed the preliminary changes in the insect
assemblages between their in situ and ex situ plant popu-
lations. This is a preliminary study because the transloca-
tion experiments have only been underway for a few years
with some plants only recently reaching reproductive
maturity. However, this investigation is warranted given
that insect species associated with the threatened plants are
potentially under immediate risk of coextinction. We
addressed the following questions:
• How distinctive are the assemblages of insects on these
threatened plants in the wild?
• How similar are the species compositions of insect
assemblages found on wild threatened plants and
translocated individuals of the same plant species?
• Are the insect species that use these threatened plants
also found elsewhere, and are the most host-specific
insects also found on translocated individuals?
• Which co-occurring plant species have insect assem-
blages that most resemble those of translocated host
plants?
Methods
Study area
The study areas of the Stirling Range National Park and
Torndirrup Peninsula (Albany city reserve), plus the
translocation site between the two native sites, are located
within the South-west Australian Floristic Region of
Western Australia (Fig. 1). The Stirling Range National
Park covers 115,920 hectares of remnant vegetation, with
sampling occurring in approximately half the area.
Torndirrup Peninsula is remnant vegetation which is joined
to the 3,868 ha Torndirrup National Park, although sam-
pling only occurred within 1 ha of the Peninsula itself.
Banksia montana and L. gnaphalioides are restricted to
areas of high elevation in the Stirling Range ([950 m
above sea level). Banksia brownii occurs in areas of high
elevation in the Stirling Range, but it also persists in sev-
eral disjunct populations within 80 km to the south and
south-east, including at Torndirrup Peninsula. Although
there is no historical record of B. brownii occurring within
the translocation site or the surrounding area, recent pop-
ulation genetic studies suggest that it was probably dis-
tributed throughout the region during at least the mid
Pleistocene (Coates and McArthur 2010). These studies
also suggest that the current disjunct distribution is the
result of subsequent climatic events (Hopper 2009) and
significantly predates any land clearing over the last
150 years (Coates and McArthur 2010). Individuals of
each of the threatened plant species, L. gnaphalioides,
B. brownii, and B. montana, were grown from seed or
cuttings, and transplanted as seedlings approximately
50 km from their nearest current natural populations
(herein termed ‘in situ’) in the Stirling Range National Park
(see Cochrane et al. 2011; Fig. 1). Developed in 2003, the
site to which the plants were translocated is in a 2.8 km2
remnant of woodland on lateritic soils surrounded by
farmland and other native remnants near Kamballup. This
contrasts with in situ sites which consist of thickets and
mallee-heath on rocky sand clay loam soils in the Stirling
J Insect Conserv
123
Author's personal copy

Range, and mallee heath and low woodland on sandy clay
soils over granite at the Torndirrup Penisula (Coates and
McArthur 2010). Climate of all three regions hasn’t been
recorded but 5 km north of Torndirrup, Albany has an
average annual rainfall of 929 mm, and 30 km west of
Kamballup, Mt Barker receives 729 mm annually (Bureau
of Meteorology—Australia (BoM) 2011). Approximately
50 km northeast of the Stirling Range, Ongerup receives
385 mm annually (Bureau of Meteorology—Australia
(BoM) 2011), which may be a close approximation of the
rainfall in the low-lying regions of the Stirling Range, but
montane conditions where the threatened plants occur are
wetter and cooler. The translocation site was selected for
this study due to its location between both in situ sites
(Fig. 1). The translocated plants (herein termed ‘ex situ’)
are being grown within this native vegetation in several
monospecific transects of two rows each, numbering
approximately 40 plants per transect.
Sampling
There were three study sites in total: the translocation site at
Kamballup, and two in situ (or natural/native) sites at
Torndirrup Peninsula and Stirling Range National Park
(Fig. 1). Sampling occurred during September 2007 for the
translocation site, October 2008 at one in situ site (Tornd-
irrup) and from September—December 2007, plus October
2008, within the second in situ site (Stirling Range). The
length of time taken was due to the number of plants sampled
(3,026 individuals, 104 species) for this, and a larger study
on the coextinction of insects on threatened plants. Sampling
was also stratified according to altitude, to account for
delayed responses of insect assemblages at higher altitudes
due to slower temperature increases in these areas. For
example, Fielding et al. (1999) found that for every 100 m in
elevation, the appearance of adults of a Cercopidae bug was
delayed by 5.6 days. Therefore, lowland sites (15–300 m
above sea level: Translocation, Torndirrup and some Stirling
Range areas) were sampled in early spring, while higher
mountain altitudes (350–650 m above sea level) were
sampled later in spring, and the highest altitudes
(700–1,039 m above sea level) sampled in very late spring.
At in situ sites (Stirling Range and Torndirrup), insects
were collected from both threatened and non-threatened,
co-occurring plant species by beating and vacuuming.
When used in combination, these techniques have been
shown to be efficient for sampling insects within this type
of vegetation (Moir et al. 2005b; McCarthy et al. 2010). All
plant individuals were sampled with similar sampling
intensity regardless of site, with all above-ground plant
structures vacuumed with the samples frozen and sorted
under a microscope (see Moir et al. 2005a), or plants were
beaten and insects fell into a modified butterfly net
(*70 cm in diameter) which had the netting replaced with
white calico material. Each plant that was beaten or vac-
uumed was also searched by hand to collect cryptic insect
fauna, such as galls and lerps, not readily captured by the
other sampling methods. Fifteen individual plants of
B. brownii and L. gnaphalioides were vacuumed and
another 15 beaten, while only seven individuals of
B. montana were vacuumed on Bluff Knoll due to the low
numbers of plants remaining here (\15 mature plants). An
additional 14 individuals of 15–22 confamilial species and
genera for each plant species were sampled (seven indi-
viduals vacuumed, seven beaten) to determine which
insects were restricted in host breadth to the focal plant
Fig. 1 Location of study sites
within the south-west of
Western Australia showing the
plant species and number of
individual plants sampled in
square brackets (adapted from
Google Earth, 2010)
J Insect Conserv
123
Author's personal copy

species (Appendix 1—see Electronic supplementary
material; Fig. 1). Confamilial taxa for each threatened
plant species were selected on the basis that they 1.)
occurred in the same area (within 1 km), or 2.) were sister
species and were sampled up to 20 km away.
At the translocation site, 15 individuals of B. montana,
B. brownii (originating from Stirling Range, not Torndir-
rup) and L. gnaphalioides (39, 3, and 28 months-old
respectively) were sampled by vacuuming. We were not
granted permission to beat translocated individuals due to
the low numbers of plants available and concerns that
beating might damage plants. To account for the potential
difference in assemblages captured using the different
sampling techniques, vacuum samples were analysed sep-
arately, as well as in combination with all other samples.
The heights of translocated plants were generally smaller
than those plants sampled at in situ sites due to the dif-
ferences in age (i.e. B. montana translocated 76.2 cm, in
situ *150 cm; B. brownii translocated *70 cm, in situ
*120–280 cm, L. gnaphalioides translocated 31.4 cm, in
situ *56 cm), however, translocated plants grew more
vigorously than the same aged plants at in situ sites (e.g.,
translocated B. montana were on average 76.2 cm and
same aged plants in situ, 6.8 cm; 2-year old translocated
L. gnaphalioides were on average 31.4 cm while 6-year old
plants in situ were 4.7 cm due to grazing: Barrett unpub-
lished data). Three to four confamilial species, and seven to
eight unrelated species, were also sampled at the translo-
cation site by all methods (Appendix 1—see Electronic
supplementary material; Fig. 1). These surrounding plant
species were within 10–100 m of the translocated plants.
Identification of specimens
We preserved all plant-dwelling insects, and identified the
beetles (Coleoptera) and bugs (Hemiptera) to species level,
with validation from taxonomic experts. The smaller orders
(representing 10% of total individuals collected), Orthop-
tera, Phasmatodea, Hymenoptera, Thysanoptera and larval
Lepidoptera, were identified to morpho-species. Introduced
insect species of Coleoptera and Hemiptera (e.g., aphids)
were removed from the analysis as the objective of the
study was to assess native insect assemblages.
Data analysis
Insect assemblages
Differences in insect species density (sensu Gotelli and
Colwell 2001) and insect abundance between sites were
compared for each threatened plant species using one-way
analysis of variance (ANOVA) using SPSS (SPSS Inc.
2001). Data for insect abundance on all plants, and species
density on the banksias, were square-root transformed to
improve normality of the data and homogeneity of variance;
F-ratios were considered significant at P \ 0.05. Smoothed
species accumulation curves (50,000 randomizations)
against observed number of insect individuals for each
threatened plant species at each site were constructed. The
number of species that remain undetected on each plant
species was estimated with the Abundance-based Coverage
Estimator (ACE). Curves were constructed using EstimateS
8.2 (Colwell 2006).
We used analysis-of-similarities (ANOSIM) (Clarke
1993) to assess differences in insect assemblages between
the focal threatened plant species and co-occurring species
(see Appendix 1—see Electronic supplementary material).
We also used ANOSIM to determine the differences in
composition between insect assemblages on in situ and
translocated populations of the three focal plant species.
ANOSIM R-statistics generally range between 0 and 1; a
value of 0 indicates that two assemblages are identical
whereas a value of 1 indicates that two assemblages are
entirely different (see Clarke 1993). Similarity matrices were
constructed using the Bray–Curtis measure for the abun-
dance of insect species on each plant individual. Square-root
transformation down-weights the importance of abundant
species, and increases the contribution from rarer species
(Clarke 1993), which was desirable in this study to prevent
common species from dominating analyses and overlooking
rarer but potentially host-specific insects on each plant spe-
cies. As a precautionary approach, we reduced the chance of
a Type II error (failing to detect dissimilarity when it exists)
by considering ANOSIM R-statistics significant at P \ 0.10.
Similarity percentages (SIMPER) identified the contribution
of each insect species to the dissimilarity between plant
species to help identify insect species that may be under- or
over-represented on translocated hosts relative to in situ
hosts. Non-metric multi-dimensional scaling (MDS) was
performed (1,000 restarts) on the Bray–Curtis matrix to
produce ordinations. These analyses were conducted in
PRIMER-E version 6.1.11 (PRIMER-E Ltd 2008).
Insect species
The taxonomy and/or ecology of the majority of insects
comprising the assemblages on the plants at the study sites
are unknown. To determine the insect species most likely
to be host-specific to the focal plant species we used a host
breadth model using hierarchical Bayesian Zero-Inflated
Poisson modelling. Details of this model are presented
elsewhere (Vesk et al. 2010). Briefly, the model includes
sub-models for host use, insect occupancy and abundance
and ‘tourist’ insect occupancy. The model that we use here
is a modification of those presented in Vesk et al. (2010)
J Insect Conserv
123
Author's personal copy

with the additional aim of estimating occupancy of insect
species at different sites—the in situ and translocated
populations of the focal plant species.
The data Yi are observed counts of insects on individual
plants. We model the data as having three components—
counts of zero, counts of one and other counts. Zeros arise
from either the plant species not being used by the insect
species or from the insect species using the plant species as a
host, but being absent from the sample. Absence from the
sample can be either because the insect truly did not occupy
the individual plant sampled or it occupied the plant but
evaded capture. We do not distinguish between these two
processes in terms of their relative contribution to detection.
Counts of one may arise either from true occupancy of
insects on individual plants that they use, or from ‘tourist’
occupancy on plants that the insects do not actually use.
Counts greater than one, we assumed, would only result from
detections of insects on plants that they truly use as hosts, i.e.
we explicitly assumed that tourist occupancy would not
result in two or more individuals of an insect species on a
plant from a species that they did not actually use.
In contrast with Vesk et al. (2010), here we model the
likelihood of individual observations. We have found that
this approach has better Markov Chain Monte Carlo sam-
pling properties, and while more computationally
demanding and thus slower, provides a more reliable route
for analysis (PA Vesk unpublished data).
The likelihoods are as follows.
LðYi ¼ yjhi;-i;ui;kiÞ
¼hið1�-iÞ þ ð1� hiÞð1�uiÞ; y¼ 0
ð1� hiÞuiþ hi-iki expð�kiÞ=ð1� expð�kiÞÞ; y¼ 1
hi-ikyi expð�kiÞ=ðy!ð1� expð�kiÞÞÞ; y[1
8><
>:
here, pi indicates whether the invertebrate species actually
uses the plant species on which it was recorded in obser-
vation i (hi = 1) or not (hi = 0). The parameter xi is the
probability that the individual plant represented by obser-
vation i is occupied by the insect species represented by
observation i conditional on the plant species being used by
that invertebrate. /i is the probability that the observation
is that of a tourist individual when the plant species is not
used, and ki is the mean abundance of the insect species on
the individual plant when present (but not as a tourist).
Being counts, abundance was assumed to be Poisson dis-
tributed, and truncated to values above 1 because absence
was modelled separately.
We used a hierarchical structure to write models for
each of the parameters hi, xi and ki. The actual use of an
individual plant by an invertebrate species hi was modelled
as a Bernoulli realization with the specified probability pi
for that plant family and invertebrate species combination,
to account for appropriate dependencies in the data.
Variation between plant species within the family was
unmodelled ‘noise’ (Vesk et al. 2010). Occupancy by an
invertebrate species was modelled at the level of species-
by-site combinations. This enabled occupancy of the par-
ticular invertebrate species to be compared between sites,
e.g. in situ and translocated populations of the three focal
threatened plant species. The average abundance ki was
modelled at the level of insect orders. This is quite coarse,
but our focus here is on occupancy. The logit of the
probabilities of use, occupancy and tourist occurrence were
assumed to have normal distributions with unknown means
and standard deviations that were estimated from the data.
We used Bayesian inference, employing vague priors
throughout. Parameters representing means were drawn
from normal distributions with means equal to 0 and stan-
dard deviations equal to H1000. Standard deviations were
specified as drawn from uniform distributions between 0
and 20. The prior distribution for the probability of a tourist
occurrence /i was specified as uniform between 0 and 1.
We used Markov chain Monte Carlo sampling to esti-
mate posterior probability distributions of parameters. The
model was implemented in the freeware OpenBUGS 3.11
(Thomas et al. 2006). The code is presented in Appendix
2—see Electronic supplementary material.
We routinely inspected chain traces and found that
convergence usually occurred within 2,000 iterations. We
discarded the first 5,000 iterations as a burn in and sampled
at least the next 10,000 samples from the Markov chain.
Using the host breadth model, we estimated: the prob-
ability of an insect using each of the three focal plant
species B. montana, B. brownii or L. gnaphalioides; the
host breadth of those insect species and; the probability of
occupancy of those insect species at each site. Insects
identified as having [40% probability of using each
threatened host plant species and a 25% chance that the
host breadth was three plant species or fewer were then
subjected to the decision protocol (Fig. 1 of Moir et al.
2011) to assess their potential as co-threatened taxa. We
used 25% as an example lower bound or threshold to
reduce the risk of failing to recognise a host-restricted
invertebrate (Type II error) below that which might be
expected from use of means or medians. Because host
breadth can only be positive and is akin to a count, there is
a mean–variance relationship that results in higher mean
host breadth with greater uncertainty. The actual threshold
used would be a matter for managers, and would depend
upon their attitude to risk.
Results
Our database of plants and their insect affiliates comprised
1,240 plants representing 44 plant species (11 genera from
J Insect Conserv
123
Author's personal copy
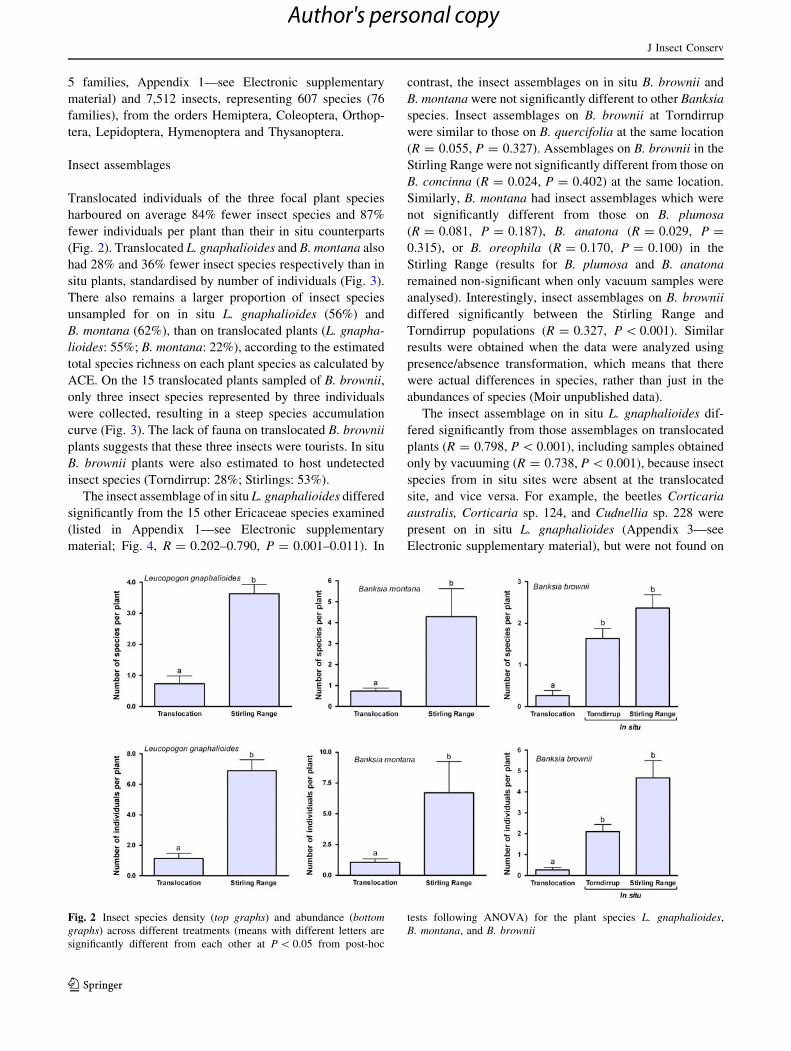
5 families, Appendix 1—see Electronic supplementary
material) and 7,512 insects, representing 607 species (76
families), from the orders Hemiptera, Coleoptera, Orthop-
tera, Lepidoptera, Hymenoptera and Thysanoptera.
Insect assemblages
Translocated individuals of the three focal plant species
harboured on average 84% fewer insect species and 87%
fewer individuals per plant than their in situ counterparts
(Fig. 2). Translocated L. gnaphalioides and B. montana also
had 28% and 36% fewer insect species respectively than in
situ plants, standardised by number of individuals (Fig. 3).
There also remains a larger proportion of insect species
unsampled for on in situ L. gnaphalioides (56%) and
B. montana (62%), than on translocated plants (L. gnapha-
lioides: 55%; B. montana: 22%), according to the estimated
total species richness on each plant species as calculated by
ACE. On the 15 translocated plants sampled of B. brownii,
only three insect species represented by three individuals
were collected, resulting in a steep species accumulation
curve (Fig. 3). The lack of fauna on translocated B. brownii
plants suggests that these three insects were tourists. In situ
B. brownii plants were also estimated to host undetected
insect species (Torndirrup: 28%; Stirlings: 53%).
The insect assemblage of in situ L. gnaphalioides differed
significantly from the 15 other Ericaceae species examined
(listed in Appendix 1—see Electronic supplementary
material; Fig. 4, R = 0.202–0.790, P = 0.001–0.011). In
contrast, the insect assemblages on in situ B. brownii and
B. montana were not significantly different to other Banksia
species. Insect assemblages on B. brownii at Torndirrup
were similar to those on B. quercifolia at the same location
(R = 0.055, P = 0.327). Assemblages on B. brownii in the
Stirling Range were not significantly different from those on
B. concinna (R = 0.024, P = 0.402) at the same location.
Similarly, B. montana had insect assemblages which were
not significantly different from those on B. plumosa
(R = 0.081, P = 0.187), B. anatona (R = 0.029, P =
0.315), or B. oreophila (R = 0.170, P = 0.100) in the
Stirling Range (results for B. plumosa and B. anatona
remained non-significant when only vacuum samples were
analysed). Interestingly, insect assemblages on B. brownii
differed significantly between the Stirling Range and
Torndirrup populations (R = 0.327, P \ 0.001). Similar
results were obtained when the data were analyzed using
presence/absence transformation, which means that there
were actual differences in species, rather than just in the
abundances of species (Moir unpublished data).
The insect assemblage on in situ L. gnaphalioides dif-
fered significantly from those assemblages on translocated
plants (R = 0.798, P \ 0.001), including samples obtained
only by vacuuming (R = 0.738, P \ 0.001), because insect
species from in situ sites were absent at the translocated
site, and vice versa. For example, the beetles Corticaria
australis, Corticaria sp. 124, and Cudnellia sp. 228 were
present on in situ L. gnaphalioides (Appendix 3—see
Electronic supplementary material), but were not found on
Fig. 2 Insect species density (top graphs) and abundance (bottomgraphs) across different treatments (means with different letters are
significantly different from each other at P \ 0.05 from post-hoc
tests following ANOVA) for the plant species L. gnaphalioides,
B. montana, and B. brownii
J Insect Conserv
123
Author's personal copy
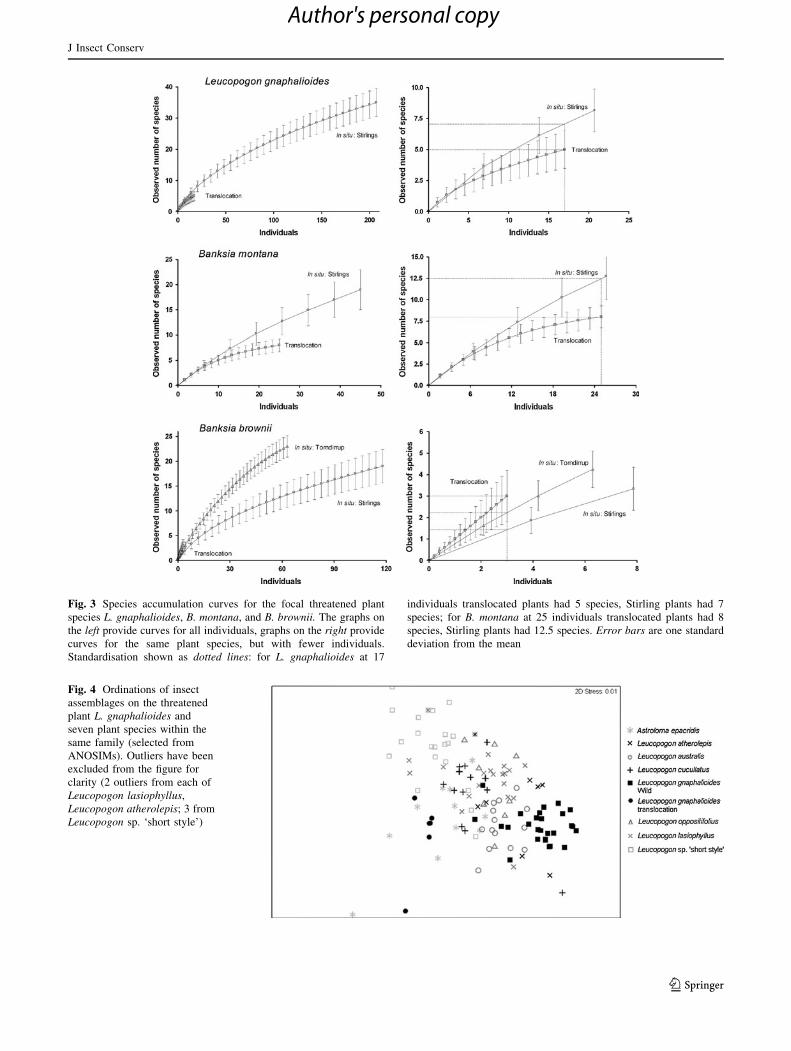
Fig. 3 Species accumulation curves for the focal threatened plant
species L. gnaphalioides, B. montana, and B. brownii. The graphs on
the left provide curves for all individuals, graphs on the right provide
curves for the same plant species, but with fewer individuals.
Standardisation shown as dotted lines: for L. gnaphalioides at 17
individuals translocated plants had 5 species, Stirling plants had 7
species; for B. montana at 25 individuals translocated plants had 8
species, Stirling plants had 12.5 species. Error bars are one standard
deviation from the mean
Fig. 4 Ordinations of insect
assemblages on the threatened
plant L. gnaphalioides and
seven plant species within the
same family (selected from
ANOSIMs). Outliers have been
excluded from the figure for
clarity (2 outliers from each of
Leucopogon lasiophyllus,
Leucopogon atherolepis; 3 from
Leucopogon sp. ‘short style’)
J Insect Conserv
123
Author's personal copy

translocated plants. Instead, translocated L. gnaphalioides
had an insect assemblage more similar to a closely related
plant species within the surrounding area, Astroloma
epacridis (R = 0.006, P = 0.376, Fig. 4). The dominant
insect in the assemblage on both Astroloma epacridis and
L. gnaphalioides at the translocation site was the Achilidae
bug Plectoderini sp. 12 (Appendix 3—see Electronic sup-
plementary material).
Insect species
After assessing the complete insect assemblage with host
breadth models and the decision protocol, no insect species
were identified as potentially host-specific to L. gnapha-
lioides in the wild (Table 1). Interestingly, some beetles
(Corticaria australis, Corticaria sp. 124, and Cudnellia sp.
228), highlighted by the community assessment above as
important components of the distinctive assemblage of in
situ L. gnaphalioides, were not host-specific to this plant
species, but were also found on other epacrids in the Stir-
ling Range. For Corticaria, this is most likely due to its
non-specific mycetophagous (fungal) feeding habitats
(Matthews 1992). In contrast, Cudnellia sp. 228 is a her-
bivore and potentially a specialist at family-level as it was
only found on epacrids. Moreover, it is flightless and was
restricted to the summit of Bluff Knoll, suggesting that it is
characteristic of that plant community. These results indi-
cate that, rather than consisting of multiple host-specific
insect species, the assemblage on in situ L. gnaphalioides
was an unusual combination of insect species, which were
not seen together on other epacrids (similar to assemblages
consisting of Threatened Ecological Communities which
may not feature a threatened species; Nicholson et al.
2009).
Host-breadth models and the decision protocol identified
one insect species on in situ B. montana and three insect
species from in situ B. brownii that appeared host-specific
to these plant species (Table 1). Of these four species, the
three hemipterans are highly likely to be host-specific as all
feed exclusively on the sap of the plants. Although the
fourth insect, beetle Bruchinae sp. 282, was not found on
any other plant species, its host-specificity on B. brownii is
uncertain as the Australian members of this subfamily of
insects are thought to be seed feeders as larvae and pollen
feeders as adults (Hangay and Zborowski 2010), and plants
were not in flower when sampled. In any case, the proba-
bility that any of these four insects were present at the
translocated site was very low (\0.6%, Table 2). These
insects were not found on translocated individuals of
B. montana or B. brownii, despite one, the plant-louse
Trioza sp. 03, occurring on in situ populations of
B. brownii at both Torndirrup and Stirling Range.
Discussion
This study suggests that plant translocations that involve
seed collection and propagation, may have limited benefit
for individual host-specific dependent species or the sup-
ported insect assemblage within the first few years, unless
the insects are also assisted in colonising the plants at the
new location. The accumulation of new insect species on
translocated plants was limited in terms of number of
species per individuals collected, but was similar to the
insect colonisation of plant species introduced to countries
or continents outside their natural range (Brandle et al.
2008). Furthermore, insect assemblages on translocated
plants were most similar to those from surrounding, related
plant species at the translocated site, a finding consistent
with what is known about colonisation patterns of intro-
duced exotic species (e.g., Strong et al. 1977; Brandle et al.
2008). Guild structure of the insect assemblages between in
Table 1 Summary of insects collected from targeted threatened plant species, with the insect species potentially restricted to each plant
identified, following analysis by host breadth models (Vesk et al. 2010) and the decision protocol (Moir et al. 2011)
Plant species Location Total insect
species
Total insect
abundance
Insect species restricted to each plant species
Leucopogon gnaphalioides In situ (Stirling) 35 207 0
Translocated 5 17 NA
Banksia montana In situ (Stirling) 19 45 Pseudococcus sp. 15 (Pseudococcidae)
Translocated 8 25 NA
Banksia brownii In situ (Torndirrup) 23 63 Bruchinae sp. 282 (Chrysomelidae)
Ceronema sp. 18 (Coccidae)
Trioza sp. 03 (Triozidae)
In situ (Stirling) 19 118 Trioza sp. 03 (Triozidae)
Translocated 3 3 NA
J Insect Conserv
123
Author's personal copy

situ and translocated plants remain to be investigated, and
may give different results as to the efficacy of host trans-
location on insect assemblages (e.g., Andrew and Hughes
2007). Similarly, as we only examined predominantly one
trophic level (herbivores) present on threatened plants, the
impact of translocation on other trophic levels, as well as
on the cascade of interactions between trophic levels,
remains unknown.
Natural reassembly of insect assemblages on individual
plants that have been reintroduced or translocated into an
area is possible, but depends on several factors including
dispersal capabilities of the insects (i.e. Moir et al. 2010b),
competition from insects already established on the plants
(e.g., Gonzalez-Megıas and Gomez 2003), parasitism and
predation (e.g., Denno et al. 2003), and the physiological
requirements of the insect species themselves (e.g., New
2008). Time lags are expected in the dispersal and estab-
lishment of insect populations on translocated host popu-
lations (Moir et al. 2010b), although reassembly can occur
quickly when source insect populations are located nearby
(e.g., Gratton and Denno 2005). Insect species with poor
dispersal abilities, such as those without wings, may not be
capable of reaching new sites and thus require assistance to
establish (Moir et al. 2005b; Watts et al. 2008). Even
winged insects can be poor dispersers, such as Mitchell’s
satyr butterfly, which does not travel further than
420 m year-1 (Szymanski et al. 2004). In the case pre-
sented here, the translocation site was [40 km away,
located within a landscape matrix fragmented by agricul-
tural development and occurred within different vegetation
types to that of the in situ sites (montane and coastal heath,
versus jarrah/marri woodland), perhaps making dispersal
more difficult. Competition and predation may not inhibit
insect species establishment in this study as most of the
translocated plants were host to very few other insects,
particularly B. brownii. Finally, the physiological require-
ments of the insects examined here may possibly contribute
to their absence from translocated plants as the majority
occupy cooler, wetter areas than the habitats at the trans-
location site. Elucidating the mechanisms as to why insect
species or assemblages are not present on translocated
plants requires further study. In any case, to maintain the
diversity of insect assemblages and host-dependent rela-
tionships that occur on the threatened plant species in the
wild, the next step should involve further conservation
actions, and in particular, overcoming taxonomic impedi-
ments by species descriptions of all host-specific insects
(note that all host-specific species identified here are
undescribed).
Our study has significant implications for assisted
migration or other movements of threatened host species,
which often occur over much greater distances than those
described within this study. Namely, that dependent spe-
cies and associated assemblages are highly unlikely to
re-establish naturally. Rather than a single species
approach when translocating for conservation purposes,
land managers should consider moving suites of species
(Seddon 2010), including dependent herbivores and other
dependent invertebrates with the host taxa (see Moir et al.
in review). This broader method of translocation may be
moving towards the novel ecosystem approach (see Hobbs
et al. 2006), whereby assemblages of species under threat
of extinction through climate change are translocated to an
area where they do not currently occur, thereby creating a
novel ecosystem in the new location (Lindenmayer et al.
2010). Such an approach may carry risks if the translocated
species spread and displace indigenous species in the
vicinity of the translocation site, a question likely to require
significant experimentation to resolve. According to Hunter
(2007), our translocation site would provide a good initial
test of insect translocation as it ‘‘was well connected his-
torically, but is currently surrounded by human-dominated
landscapes that might be a barrier if the translocated spe-
cies had unacceptable effects’’.
Table 2 Estimated probability
of occurrence at in situ (Stirling
Range [Stirl] or Torndirrup
[Torn]) and translocation sites
of insect species identified as
host specific to the targeted
threatened plant species in the
wild (see Table 1)
Plant host Insect species (family) Location Estimated probability
of occurrence (%)
Banksia montana Pseudococcus sp. 15 (Pseudococcidae) Translocation 0.4
In situ (Stirl) 6.8
Banksia brownii Bruchinae sp. 282 (Chrysomelidae) Translocation 0.5
In situ (Stirl) 0.3
In situ (Torn) 5.6
Banksia brownii Ceronema sp. 18 (Coccidae) Translocation 0.3
In situ (Stirl) 0.3
In situ (Torn) 5.2
Banksia brownii Trioza sp. 03 (Triozidae) Translocation 0.5
In situ (Stirl) 10.7
In situ (Torn) 2.8
J Insect Conserv
123
Author's personal copy

Limitations and management needs
This preliminary study has two limitations; lack of site
replication and the relatively limited time since transloca-
tion. Both factors were imposed by logistic constraints on
the availability of both in situ and translocated sites for all
three threatened plant species, but suggest important
directions for future adaptive management. The lack of
replication for both native and translocation sites means
that we were unable to assess the influence of site variation
on the results. While some sources of site variation cannot
be accounted for, differences in plant growth rates, local
richness of invertebrate species, and sizes of plant popu-
lations appear not to explain the observed differences in
invertebrate assemblages between the translocated site and
the in situ sites. Plants in the translocated site grew faster
compared to similar aged plants within in situ sites and, in
the case of B. montana, reaching sexual maturity and
flowering far earlier than in the source population (Barrett
pers. obs.). The translocation site also had as many indi-
viduals of the focal plant species as the in situ sites. There
was a rich diversity of insects in the translocation site (144
insect species in total, excluding introduced species), and
when a plant species occurring naturally both within in situ
and ex situ sites was examined (Banksia sessilis), plants in
the translocated site had more insects than those situated
within in situ sites (translocated site: 110 individual insects,
32 insect species; Stirling Range ‘northeast’ population: 42
individuals, 20 species; Stirling Range ‘southeast’ popu-
lation: 21 individuals, 7 species). Despite these findings, it
is likely that some ex situ sites will produce better trans-
location outcomes than others. Establishment and sampling
of additional translocation sites would improve under-
standing about factors that influence success for both plants
and dependent invertebrates, providing a more informed
basis for planning future translocation projects. Increased
replication of in situ locations is greatly constrained by the
fact that some of the plant species such as B. montana only
remain in one in situ location.
The temporal dynamics of invertebrate occupation of
translocated host plants is unknown (e.g., Moir et al.
2005b, 2010b). Our sampling began relatively soon after
plants were translocated (3–39 months later), and pre-
sumably recorded the early stages of a trajectory in which
increasing numbers of invertebrate species find and colo-
nise newly available host plants. Whether this trajectory
continues to increase in subsequent years requires further
work to resolve. Thus, an important adaptive action is the
continued monitoring of invertebrate assemblages on
translocated plants, relative to in situ populations of the
hosts, as well as the surrounding ecosystem, to determine
the needs for supplementary translocations of invertebrate
species that fail to colonise their translocated hosts. The
Stirling Range has been identified as a hotspot for inver-
tebrate endemism in the south-west (e.g., Department of
Environment & Conservation 2008; Moir et al. 2009), with
invertebrate fauna restricted to this area due to the onset of
aridity in the late Miocene-Pliocene (Main 1999; Moir
et al. 2009; Cooper et al. 2011). Given this, and also that
the majority of insects identified here are wingless and/or
small, together with the long distance of the translocation
site from native areas, we expect that most species will not
recolonise for many decades, if at all.
Given the immediate threats to these host plants in the
wild (see Barrett et al. 2008), and the fact that populations
of host-specific insects may decline faster than that of their
hosts (e.g., Taylor and Moir 2009; Moir et al. 2010a),
conservation actions to assist in the insects survival are
required urgently. This study provides the first step in the
development of an adaptive management strategy that
guides a structured process of learning by doing (e.g., Keith
et al. 2011) to improve translocation outcomes for depen-
dent invertebrate fauna as well as their threatened plant
hosts.
Acknowledgments Grants from the Australian Research Council
(DP0772057) and Australia & Pacific Science Foundation (APSF
07/3) supported this work. We thank P. Luscombe for his enthusiasm
in the translocation experiment conducted on his property, and
assistance with plant identifications. We gratefully thank B. Hanich,
R. Oberprieler, G.S. Taylor, A. Szito, P.J. Gullan, S. Barker and
C.A.M. Reid for taxonomic assistance. Finally, two anonymous
reviewers and Editor T. Shreeve are thanked for their time and useful
suggestions on this paper.
References
Andrew NR, Hughes L (2007) Potential host colonization by insect
herbivores in a warmer climate: a transplant experiment. Glob
Change Biol 13:1539–1549
Barrett S, Shearer BL, Crane CE, Cochrane A (2008) An extinction-
risk assessment tool for flora threatened by Phytophthoracinnamomi. Aust J Bot 56:477–486
Brandle M, Kuhn I, Klotz S, Belle C, Brandl R (2008) Species
richness of herbivores on exotic host plants increases with time
since introduction of the host. Divers Distribut 14:905–912
Bureau of Meteorology—Australia (BoM) (2011) Trend in Annual
Total Rainfall 1960–2009 (mm/10 years). Australian climate
variability & change—Trend maps. BoM, Canberra, Australia.
http://www.bom.gov.au/cgi-bin/climate/change/trendmaps.cgi?
map=rain&area=wa&season=0112&period=1960 Accessed 10
March 2011
Clarke KR (1993) Non-parametric multivariate analysis of changes in
community structure. Aust J Ecol 18:117–143
Coates DJ, McArthur S (2010) Assessing the taxonomic status of
Banksia brownii and patterns of genetic diversity in extinct and
extant populations. In: Bankwest landscope project. Western
Australian Department of Environment and Conservation, Perth,
p 8
Cochrane A, Barrett S, Monks L, Dillon R (2011) Partnering
conservation actions. Inter situ solutions to recover threatened
species in South West Western Australia. Kew Bulletin 65:1–6
J Insect Conserv
123
Author's personal copy

Colwell RK (2006) EstimateS. http://viceroy.eeb.uconn.edu/estimates
6.0b1 edn. University of Connecticut
Cooper SJB, Harvey MS, Saint KM, Main BY (2011) Deep
phylogeographic structuring of populations of the trapdoor
spider Moggridgea tingle (Migidae) from southwestern Austra-
lia: evidence for long-term refugia within refugia. Molecular
Ecology doi:10.1111/j.1365-294X.2011.05160.x
Denno RF, Gratton C, Debel H, Finke DL (2003) Predation risk
affects relative strength of top-down and bottom-up impacts on
insect herbivores. Ecology 84:1032–1044
Department of Environment & Conservation (2008) Wildlife man-
agement plan for the threatened and relictual invertebrates of the
Stirling range. 2008–2013. Wildlife management plan. Depart-
ment of Environment & Conservation, Perth
Fielding CA, Whittaker JB, Butterfield JEL, Coulson JC (1999)
Predicting responses to climate change: the effect of altitude and
latitude on the phenology of the Spittlebug Neophilaenuslineatus. Funct Ecol 13:65–73
Gonzalez-Megıas A, Gomez JM (2003) Consequences of removing a
keystone herbivore for the abundance and diversity of arthropods
associated with a cruciferous shrub. Ecol Entomol 28:299–308
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures
and pitfalls in the measurement and comparison of species
richness. Ecol Lett 4:379–391
Gratton C, Denno RF (2005) Restoration of arthropod assemblages in
a Spartina salt marsh following removal of the invasive plant
Phragmites australis. Rest Ecol 13:358–372
Hangay G, Zborowski P (2010) A guide to the beetles of Australia.
CSIRO publishing, Collingwood
Hobbs RJ et al (2006) Novel ecosystems: theoretical and management
aspects of the new ecological world order. Glob Ecol Biogeog
15:1–7
Hopper S (2009) OCBIL theory: towards an integrated understanding
of the evolution, ecology and conservation of biodiversity on
old, climatically buffered, infertile landscapes. Plant Soil
322:49–86
Hopper SD, Gioia P (2004) The southwest Australian floristic region:
evolution and conservation of a global hotspot of diversity. Ann
Rev Ecol Evol Syst 35:623–650
Hunter ML (2007) Climate change and moving species: furthering the
debate on assisted colonization. Conserv Biol 21:1356–1358
Keith DA, Martin TG, McDonald-Madden E, Walters C (2011)
Uncertainty and adaptive management for biodiversity conser-
vation. Biol Conserv 144:1175–1178
Koh LP, Dunn RR, Sodhi NS, Colwell RK, Proctor HC, Smith VS
(2004) Species coextinctions and the biodiversity crisis. Science
305:1632–1634
Lindenmayer DB et al (2010) Conservation strategies in response to
rapid climate change: Australia as a case study. Biol Conserv
143:1587–1593
Main BY (1999) Biological anachronisms among trapdoor spiders
reflect Australia’s environmental changes since the Mesozoic.
In: Ponder W, Lunney D (eds) The other 99%: the conservation
and biodiversity of invertebrates. The Royal Zoological Society
of New South Wales, Sydney, pp 236–245
Matthews EG (1992) A guide to the beetles of South Australia, part 6,
Polyphaga: Lymexyloidea, Cleroidea and Cucujoidea. South
Australian Museum, Adelaide
McCarthy MA et al (2010) Resource allocation for efficient
environmental management. Ecol Lett 13:1280–1289
McClean CJ et al (2005) African plant diversity and climate change.
Ann Missouri Bot Garden 92:139–152
Moir ML, Brennan KEC, Majer JD, Koch JM, Fletcher MJ (2005a)
Toward an optimal sampling protocol for Hemiptera on under-
storey plants. J Insect Conserv 9:3–20
Moir ML, Brennan KEC, Koch JM, Majer JD, Fletcher MJ (2005b)
Restoration of a forest ecosystem: the effects of vegetation and
dispersal capabilities on the reassembly of plant-dwelling
arthropods. Forest Ecol Manage 217:294–306
Moir ML, Brennan KEC, Harvey MS (2009) Diversity, endemism and
species turnover of millipedes within the southwest Australia
global biodiversity hotspot. J Biogeography 36:1958–1971
Moir ML, Vesk PA, Brennan KEC, Keith DA, Hughes L, McCarthy
MA (2010a) Current constraints and future directions in
estimating coextinction. Conserv Biol 24:682–690
Moir ML, Brennan KEC, Majer JD, Koch JM, Fletcher MJ (2010b)
Plant species redundancy and the restoration of fauna habitat:
lessons from plant-dwelling bugs. Rest Ecol 18:136–147
Moir ML, Vesk PA, Brennan KEC, Keith DA, McCarthy MA,
Hughes L (2011) Identifying and managing cothreatened inver-
tebrates through assessment of coextinction risk. Conserv Biol
doi:10.1111/j.1523-1739.2011.01663.x
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J
(2000) Biodiversity hotspots for conservation priorities. Nature
403:853–858
Moir ML et al. (in review) Avoiding coextinction of dependent
species during ex situ conservation and assisted migrations of
threatened hosts. Conserv Biol. manuscript available
New TR (2008) Conserving narrow range endemic insects in the face
of climate change: options for some Australian butterflies.
J Insect Conserv 12:585–589
Nicholson E, Keith DA, Wilcove DS (2009) Assessing the threat
status of ecological communities. Conserv Biol 23:259–274
PRIMER-E Ltd (2008) Primer 6 for windows, 6.1.11 edn. PRIMER-E
Ltd, Plymouth
Seddon PJ (2010) From reintroduction to assisted colonization:
moving along the conservation translocation spectrum. Rest Ecol
18:796–802
SPSS Incorporated (2001) SPSS 11.0 for Windows. Chicago, IL
Strong DR, McCoy ED, Rey JR (1977) Time and the number of
herbivore species: the pests of sugarcane. Ecology 58:167–175
Szymanski J, Shuey JA, Oberhauser K (2004) Population structure of
the endangered Mitchell’s Satyr, Neonympha mitchellii mitchel-lii (French): implications for conservation. Am Midl Nat
152:304–322
Taylor GS, Moir ML (2009) In threat of co-extinction: new species of
Acizzia Heslop-Harrison (Hemiptera: Psyllidae) from vulnerable
species of Acacia and Pultenaea. Zootaxa 2249:20–32
Thomas A, O’Hara R, Ligges U, Sturtz S (2006) Making BUGS
Open. RNews 6:12–17
Thuiller W, Lavorel S, Araujo MB, Sykes MT, Prentice IC, Mooney
HA (2005) Climate change threats to plant diversity in Europe.
Proc Natl Acad Sci USA 102:8245–8250
Vesk PA, Moir ML, McCarthy MA (2010) How many hosts?
Modelling host breadth from field samples. Meth Ecol Evolut
1:292–299
Watts C, Stringer I, Sherley G, Gibbs G, Green C (2008) History of
weta (Orthoptera: Anostostomatidae) translocation in New
Zealand: lessons learned, islands as sanctuaries and the future.
J Insect Conserv 12:359–370
Western Australian Herbarium (2011) Florabase. Department of
Environment and Conservation. http://florabase.dec.wa.gov.au/
Accessed 27 Feb 2011
Williams JW, Jackson ST, Kutzbach JE (2007) Projected distributions
of novel and disappearing climates by 2100 AD. Proc Natl Acad
Sci USA 104:5738–5742
Wilson R, Maclean I (2011) Recent evidence for the climate change
threat to Lepidoptera and other insects. J Insect Conserv
15:259–268
J Insect Conserv
123
Author's personal copy
Copyright © 2022 FDOKUMEN




















