
A phytogeographical comparison between subalpine forests of Guatemala and Costa Rica
15
Feddes Repertorium 105 (1994) 1-2, 73-87 Berlin. Februar 1994 University of Amsterdam, Hugo de Vies Laboratory, The Netherlands G. A. ISLEBE; M. KAPPELLE A phytogeographical comparison between subalpine forests of Guatemala and Costa Rica With 4 Figures and 2 Tables Summary We conducted a comparative study on the phytogeo- graphical similarities between the vascular generic floras of subalpine forests in Guatemala and Costa Rica. In total 178 vascular plant genera in 80 families were recorded in Central American subalpine forests, with 103 in Guatemala and 127 in Costa Rica. Woody genera account for less than 30% in subalpine Guate- mala and over 50% in subalpine Costa Rica. The Guatemalan subalpine flora consists mainly of wide temperate, holarctic and neotropical herb genera, whereas the Costa Rican subalpine flora is principally made up of neotropical shrub and wide tropical tree genera, next to a small amount of wide temperate herb genera. Almost 30% of the I78 vascular genera are shared between both Central American subalpine floras, resulting in a Ssrensen Coefficient of Com- munity of 45%. The major part of the shared flora (57%) is temperate in distribution. the holarctic element being twice as important as the austral- antarctic element. In conclusion, we state that since the formation of the Panamanian Isthmus plant migration from the holarctic region into subalpine Costa Rica dominated over migration from the austral-antarctic region towards subalpine Guate- mala. Zusammenfassung Eine vergleichende Studie iiber die phytogeographi- schen Ahnlichkeiten auf Gattungsebene wurde zwi- schen der subalpinen Flora Guatemalas und Costa Ricas ausgefiihrt. Insgesamt wurden 178 Gattungen in 80 Familien fur die zentralamerikanischen sub- alpinen Walder aufgezeichnet. Fur Guatemala bzw. Costa Rica wurden 103 bzw. 127Gattungen fest- gestellt. Weniger aIs 30% sind holzige Gattungen im subalpinen Bereich von Guatemala, wlhrend es in Costa Rica mehr als 50% sind. Die guatemaltekische subalpine Flora besteht hauptsachlich aus gemaoig- ten, holarktischen und neotropischen Kriiutergattun- gen, wahrend in Costa Rica die subalpine Flora aus neotropischen Strauchgattungen und weitverbreite- ten tropischen Baumen besteht, ferner mit einer geringen Anzahl aus weitverbreiteten gemaBigten Krsutergattungen. Nahezu 30% der 178 Gattungen sind in beiden Gebieten vertreten. Der Serensen Gemeinschaftskoefizient ergab einen Wert von 45%. Der gr6Bte Teil der gemeinsamen Flora (57%) ist gemlBigt in seiner Verbreitung und das holarktische Element ist doppelt so stark vertreten im Vergleich zum austral-anatarktischen Florenelement. Zusam- menfassend kann gesagt werden, daD seit der Entste- hung des Isthmus von Panama die Pflanzenmigration a w der Holarktis in die subalpinen Zonen von Costa Rica ubenvog iiber die Migration aus der austral- antarktischen Region in die subalpine Region Guatemalas.
Transcript of A phytogeographical comparison between subalpine forests of Guatemala and Costa Rica

Feddes Repertorium 105 (1994) 1-2, 73-87 Berlin. Februar 1994
University of Amsterdam, Hugo de Vies Laboratory, The Netherlands
G. A. ISLEBE; M. KAPPELLE
A phytogeographical comparison between subalpine forests of Guatemala and Costa Rica
With 4 Figures and 2 Tables
S u m m a r y
We conducted a comparative study on the phytogeo- graphical similarities between the vascular generic floras of subalpine forests in Guatemala and Costa Rica. In total 178 vascular plant genera in 80 families were recorded in Central American subalpine forests, with 103 in Guatemala and 127 in Costa Rica. Woody genera account for less than 30% in subalpine Guate- mala and over 50% in subalpine Costa Rica. The Guatemalan subalpine flora consists mainly of wide temperate, holarctic and neotropical herb genera, whereas the Costa Rican subalpine flora is principally made up of neotropical shrub and wide tropical tree genera, next to a small amount of wide temperate herb genera. Almost 30% of the I78 vascular genera are shared between both Central American subalpine floras, resulting in a Ssrensen Coefficient of Com- munity of 45%. The major part of the shared flora (57%) is temperate in distribution. the holarctic element being twice as important as the austral- antarctic element. In conclusion, we state that since the formation of the Panamanian Isthmus plant migration from the holarctic region into subalpine Costa Rica dominated over migration from the austral-antarctic region towards subalpine Guate- mala.
Zusammenfassung
Eine vergleichende Studie iiber die phytogeographi- schen Ahnlichkeiten auf Gattungsebene wurde zwi- schen der subalpinen Flora Guatemalas und Costa Ricas ausgefiihrt. Insgesamt wurden 178 Gattungen in 80 Familien fur die zentralamerikanischen sub- alpinen Walder aufgezeichnet. Fur Guatemala bzw. Costa Rica wurden 103 bzw. 127Gattungen fest- gestellt. Weniger aIs 30% sind holzige Gattungen im subalpinen Bereich von Guatemala, wlhrend es in Costa Rica mehr als 50% sind. Die guatemaltekische subalpine Flora besteht hauptsachlich aus gemaoig- ten, holarktischen und neotropischen Kriiutergattun- gen, wahrend in Costa Rica die subalpine Flora aus neotropischen Strauchgattungen und weitverbreite- ten tropischen Baumen besteht, ferner mit einer geringen Anzahl aus weitverbreiteten gemaBigten Krsutergattungen. Nahezu 30% der 178 Gattungen sind in beiden Gebieten vertreten. Der Serensen Gemeinschaftskoefizient ergab einen Wert von 45%. Der gr6Bte Teil der gemeinsamen Flora (57%) ist gemlBigt in seiner Verbreitung und das holarktische Element ist doppelt so stark vertreten im Vergleich zum austral-anatarktischen Florenelement. Zusam- menfassend kann gesagt werden, daD seit der Entste- hung des Isthmus von Panama die Pflanzenmigration a w der Holarktis in die subalpinen Zonen von Costa Rica ubenvog iiber die Migration aus der austral- antarktischen Region in die subalpine Region Guatemalas.

74 Feddes Repert., Berlin 105 (1994) 1 - 2
Introduction In the second half of the 19th century botanical research started in the mountain regions. of Central America with the arrival of naturalists like WAGNER (1866) and SAPPER (1894) in Guatemala, and 0rsted, WERCKLE, TONDUZ and PITTIER in Costa Rica (GOMEZ & SAVAGE 1983). During the course of the present century they were followed by a series of outstanding botanists; who studied the diverse mountain floras and vegetation types occurring in this neotropical region. To mention just a few: RODR~GUEZ (1921), STANDLEY (1946- 1977), BARTRAM (1949) and HOLDRIDGE et al. (1950) in Guatemala, and STANDLEY (1934- 1937), HOLDRIDGE et al. (1971) and GOMEZ (1986) in Costa Rica.
Recently botanists, ecologists and foresters became even more interested in the vascular flora and plant communities, which occupy the mountain ranges of Central America. The main reason for this growing interest is the increasing human encroachment in this region, which causes a continuous forest depletion and land degradation (KAPPELLE 1993 ; ISLEBE, in press; KAPPELLE & JUAREZ, in prep.). However, most attention from botanists has been on either Central American montane forests or alpine vegetation (Lzacatonal’ in Guatemala, ‘paramo’ in Costa Rica). Examples of such studies are ISLEBE et al. (in press) for Guatemala and WEBER (1958), JIM~NEZ et al. (1988), KAPPELLE et al. (1989 and 1991) and CLEEF & CHAVERRI (1992) for Costa Rica. The subalpine belt, which is situated between the alpine and monta- ne belts, just below the upper tree line, has been ignored so far. Subalpine dwarf forests in Costa Rica are particularly fragile, since fires starting in (tropic-alpine) paramo habitats extend downwards, where they threaten the local, many times endemic subalpine flora (HORN 1989; KAPPELLE, in prep.). On this occasion we will give a first
approximation of the phytogeographical rela- tionships between the vascular generic floras of the only two subalpine forest areas presently found in Central America. The first of these floras is found on the slopes of the Guatemalan Sierra de Cuchumatanes, while the second occurs in the upwr region of the Costa Rican
will be made with earlier accounts on phyto- geographical spectra from alpine and montane vegetation belts available for the region.
Study Areas Guatemala Guatemalan subalpine forests are found be- tween 3150 and 3800 m alt. in the Sierra de 10s Cuchumatanes, a mountain chain in the north- western part of the country (Fig. 1). Here, subalpine forests are dominated by the conifer species Juniperus standleyi and Pinus hartwegii. Actually they are under a tremendous human pressure, and - if no inmediate action is undertaken - they will probably disappear during the forthcoming years (ISLEBE, 1993). At their lower limit they are bordered by different types of montane mixed forests cha- racterized by the conifers Abies guatemalensis, Pinus ayacahuite and P. montezumae, which are associated with broadleaved Alnus and Quercus species.
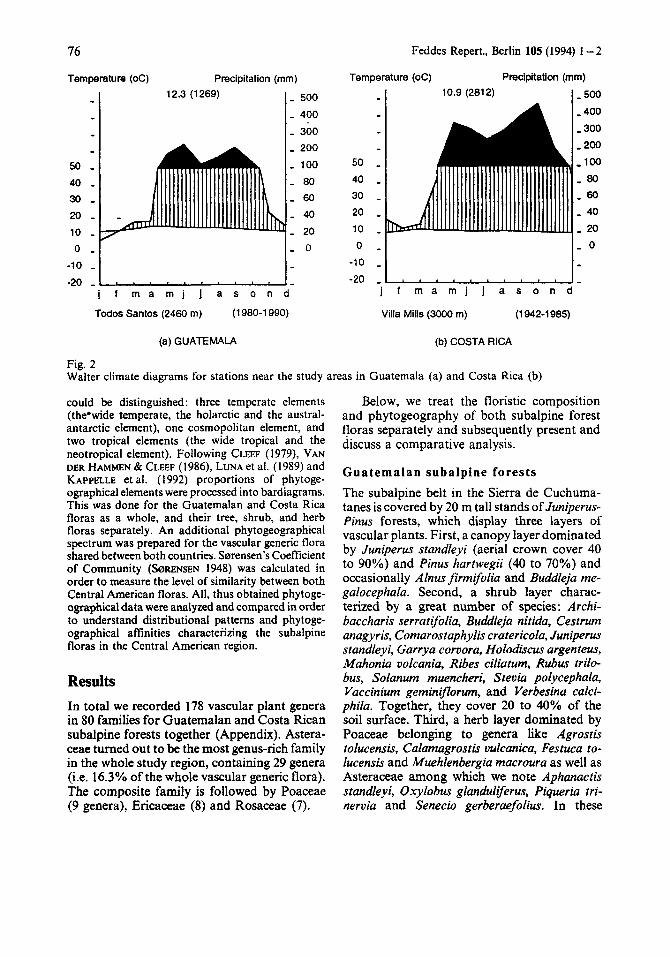
The climate in the subalpine Cuchumatanes region is cool and dry. The dry season lasts from November to April. Meteorological mea- surements at 3800m alt. in 1991 and 1992 by the first author showed, that the maximum temperature never exceeded 18.5 “C (April), while the minimum temperature could reach 3 “C below zero (December). Frosts are com- mon from November through March. The estimated rainfall fluctuates from 1200 to 1800mm per year (Perfil Ambiental, 1987). Fig. 2 presents a Walter climate diagram from Todos Santos a nearby town at 2460 m. Here, the average annual temperature is 12.3 “C and rainfall oscillates around 1269 mm.
During the Pliocene the central Guatemalan plateau was formed and the relief of the Sierra de Cuchumatanes increased due to material of plutonic origin (DENGO & BOHNENBERGER 1969; WEYL 1980). Quaternary glaciations helped shape the geomorphological landscape of the Sierra (WEYL 1980). This is evidenced by traces of glaciation near 3600 m alt., prob- ably of Wisconsin age, as observed and described by ANDERSON (1968) and HASTEN- RATH (1974). According to the F A 0 Soil Classification System subalpine soils in the
Cordillera de Talamaka. Brief comparisons Sierra de Cuchumatanes are of the Luvisol type

G. ISLEBE et al., Phytogeographical comparisons in neotropical (subalpine) forests 75
(Perfil Ambiental, 1987). Topsoil acidity at about 15 cm depth below surface ranges from 7 to 8.
Costa R i t a Subalpine forests in Costa Rica are mainly found between 3 100 and 3500 m in the Cordille- ra de Talamanca, in the south-central part of thecountry (Fig. 1). Here, 6 to 12 m tall (dwarf) forests are dominated by ericaceous species. Subalpine forests are bordered by montane Quercus-dominated forests at their lower limits, and alpine Chusquea-dominated bamboo para- mo vegetation at their upper limit, the upper forest line. Plant communities in the montane oakforest zone and (alpine) bamboo paramo belt have been treated in detail on other occasions (e.g. KAPPELLE et al. 1989, 1991; CLEF & CHAWRRI 1992).
The subalpine climate in the Cordillera de Talamanca is characterized by a dry season lasting from December to April. January is the coldest month with minimum temperatures near 3°C below zero (night frost). At about 3000 m alt. near the village of Villa Mills the average annual temperature is 10.9 "C, while the mean rainfall reaches about 2800 mm per year (IMN, 1988; Fig. 2).
Fig. 1 Location of the Central American study areas, marked with stars: the Sierra de Cuchurnatanes in Guate- mala and the Cordillera de Talamanca in Costa Ricd
The Cordillera de Talamanca, which had its final uplift during the Pliocene, is formed of marine sediments intercalated with plutonic rocks (WEYL 1980; CASTILLO 1984). The major peaks in the Cordillera, such as the Cerro Chirripi, (3819111 alt.) and the Cerro de la Muerte (3491 m alt.), have been affected by glacial periods, which left traces such as U- shaped valleys and glacial lakes (HASTENRATH 1973). At present the topography of the Cordil- lera de Talamanca is very rugged, with steep slopes covered by soils derived from volcanic ash deposits (VAs~mz 1983). The prevailing FAO-classified soil type is Andosol. In the topsoil at I5 cm depth pH values range from 3.5 to 5 (KAPPELLE et al. 1989).
Methods
On basis of recent botanical fieldwork in the subalpine forests of Guatemala (first author) and Costa Rica (second author) checklists of vascular plant species could be compiled for both areas (ISLEBE et al., in prep.; KAPPELLE etal. 1991 and unpubl.). Sub- sequently we determined the present geographical distribution of each vascular genus on basis of available literature (GOOD 1974; CLEEF 1979; WILLIS 1986; LUNA etal. 1989; CLEF & CHAVERRI 1992;
this way six different phytogeographical elements KAPPELLE et al. 1992; ULLOA & J0RGENSEN 1993). In
76
Temperature (oC) Precipitation (mm) 12.3 (1 269)
200
- 100
- 80
- 60 - 40
- 20
I - O j f m a m j j a s o n d
Todos Santos (2460 m) (1 980-1 990)
(a) GUATEMALA
Feddes Repert., Berlin 105 (1994) 1-2
Temperature (oC) Precipitation (mm) 10.9 (2812) I - 500
f m a m j j a s o n d
Villa Mills (3000 m) (1 942-1 985)
(b) COSTA RlCA
Fig. 2 Walter climate diagrams for stations near the study areas in Guatemala (a) and Costa Rica (b)
could be distinguished: three temperate elements (the'wide temperate, the holarctic and the austral- antarctic element), one cosmopolitan element, and two tropical elements (the wide tropical and the neotropical element). Following CLEEF (1979), VAN DER HAMMEN & CLEEF (1986), LUNA et al. (1989) and KAPPELLE et al. (1992) proportions of phytoge- ographical elements were processed into bardiagrams. This was done for the Guatemalan and Costa Rica floras as a whole, and their tree, shrub, and herb floras separately. An additional phytogeographical spectrum was prepared for the vascular generic flora shared between both countries. Sarensen's Coefficient of Community (SBRENSEN 1948) was calculated in order to measure the level of similarity between both Central American floras. All, thus obtained phytoge- ographical data were analyzed and compared in order to understand distributional patterns and phytoge- ographical affinities charactefizing the subalpine floras in the Central American region.
Results In total we recorded 178 vascular plant genera in 80 families for Guatemalan and Costa Rican subalpine forests together (Appendix). Astera- ceae turned out to be the most genus-rich family in the whole study region, containing 29 genera (i.e. 16.3% of the whole vascular generic flora). The composite family is followed by Poaceae (9 genera), Ericaceae (8) and Rosaceae (7).
Below, we treat the floristic composition and phytogeography of both subalpine forest floras separately and subsequently present and discuss a comparative analysis.
Guatemalan subalpine forests The subalpine belt in the Sierra de Cuchuma- tanes is covered by 20 m tall stands of Juniperus- Pinus forests, which display three layers of vascular plants. First, a canopy layer dominated by Juniperus standleyi (aerial crown cover 40 to 900/) and Pinus hartwegii (40 to 70%) and occasionally Alnus firmifalia and Buddleja me- galocephala. Second, a shrub layer charac- terized by a great number of species: Archi- baccharis serratifolia, Buddleja nitida, Cestrum anagyris, Comarostaphylis cratericola, Juniperus standleyi, Garrya corvora, Holodiscus argenteus, Mahonia volcania, Ribes ciliaturn, Rubw trilo- bus, Solanum muencheri, Stevia polycephala, Vaccinium geminiflorum, and Verbesina calci- phila. Together, they cover 20 to 40% of the soil surface. Third, a herb layer dominated by Poaceae belonging to genera like Agrostis tolucensis, Calamagrostis vulcanica, Festuca to- lucensis and Muehlenbergia macroura as well as Asteraceae among which we note Aphanactis standleyi, Oxylobus glanduliferus, Piqueria tri- nervia and Senecio gerberaefolius. In these

G. ISLEBE et al., Phytogeographical comparisons in neotropical (subalpine) forests 77
Juniperus-Pinus forests a total of 103 vascular genera were found, belonging to six trees, 22 shrubs, 70 herbs, one climber and four ferns. Most genus-rich families are Asteraceae (1 6 genera), Poaceae (8), Rosaceae (6) and Apia- ceae (5).
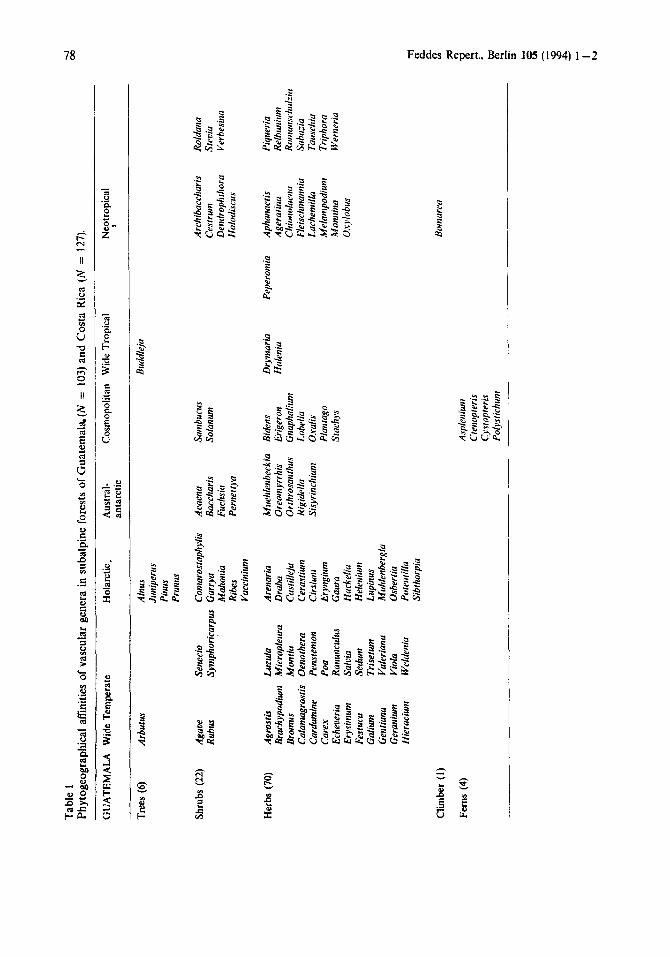
Looking at the phytogeographical spectrum for all Guatemalan vascular genera as presented in Tables 1 and 2 we note, that about 60% of them are temperate in their distribution with half ofthem wide temperate. A fifth of the total are truly holarctic, including common genera as Alnus, Juniperus, Pinus, Potentilla, Prunus, Ribes and Vaccinium. Only 12.6% of all genera are cosmopolitan, e.g. Bidens, Erigeron, Oxalis, Plantago, Sambucus and Solanum. The remai- ning quarter has a tropical distribution range, most of them being neotropical (22.4% of the total). Important neotropical taxa are Ar- chibaccharis, Cestrum, Holodiscus, Lachemilla, konnina, Relbunium, Roldana and the truly tropic-alpine Werneria. The case of Roman- schulzia as an endemic genus in the Mexican- Guatemalan-Costa Rican mountain region is noteworthy (CLEEF & CHAVERRI 1992). When we consider the phytogeographical spectrum for different growthforms some interesting trends can be revealed (Tables 1 and 2). Cer- tainly the most striking feature is the great number of herbs in these forests. They make up 68% of the total vascular flora and therefore dominate the floristic aspect. Most of these herbs are temperate in distribution. Over a quarter of the total vascular genera are wide temperate herbs, while holarctic herbs comprise only 10 to 15% of the vascular flora. Tropical herbs, on the other hand, are few in number and are mainly neotropical in distribution. Shrubs form the second most important growthform group, with over 20% of all vascular genera. These are more or less equally spread over temperate, cosmopolitan and tropical elements. Trees, climbers and ferns together account for a 10% of the vascular generic flora and do not influence the overall phytogeographical picture very much.
Costa Rican subalpine forests
Talamancan subalpine forests are dominated by the canopy species Comarostaphylis ar- butoides, Schefflera rodriguesiana, Vaccinium
consanguineum and Weinmannia trianae var. sulcata. Other important species in the tree layer are e.g. Buddleja nitida, Clethra gelida, Es- callonia myrtilloides, Myrica pubescens, Oreopa- nax xalapensis, Rhamnus oreodendron and Viburnum costaricanum. In the shrub layer numerous woody species dominate, of which some of the most conspicuous are Acaena elongata, Garrya laurifolia, Hesperomeles hetero- phylla, Holodiscus argenteus, Maytenus woo&oni, Pernettya coriacea and the hemi parasitic Gaia- dendron punctatum. The bamboo Chusguea replaces shrubby taxa in more open areas. The herb layer is characterized by a series of herbs and ferns. Some dominant genera are Castilleja, Centropogon, Gnaphalium, Lachemilla, Plantago and Valeriana. In these ericaceous subalpine forests a total of 127 vascular genera were found, belonging to 30 trees, 34 shrubs, 41 herbs, seven climbers, one bamboo and 14 ferns (including one fern-ally). Most genus-rich fami- lies are Asteraceae, Ericaceae and Rosaceae.
The phytogeographical spectrum of these subalpine forests shows, that the tropical elements make up half of the total vascular flora (Tables 1 and 2). Wide tropical and neotropical genera are present in more or less equal numbers, each comprising about a quarter of the whole flora. The temperate element is represented by almost 40% of all genera, and the remaining 12.6% consists of cosmopolitan genera. As important neotropical genera we mention Ageratina, Cestrum, Chusquea, Diplo- stephium, Disterigma, Monochaetum, Roldana and Verbesina. Myrrhidendron is considered a true neotropic-alpine (paramo) taxon (CLEEF & CHAWRRI 1992). Characteristic wide tropical genera are Abatia, Buddleja, Clethra, Crossope- talum, Maytenus, Phenax and Zanthoxylum. With regards to the temperate genera one finds for instance the holarctic Arenaria, Castilleja, Lupinus, Myricu, Prunus, Ribes and Vaccinium, the wide temperate Cardamine, Carex, Galium, Senecio, Valeriana and Viola, and the austral- antarctic Drymis, Escallonia, Gaultheria, Hy- drocotyle, Nertera and Weinmannia. Looking at the spectra for the different growthforms, one observes next to the many herb genera (32.3%) large numbers of shrub (26.8%) and tree (23.6%) genera. Thus, the woody component makes up over half of the vascular generic flora. Most important groups are the neotropical
Tabl
e I
Phyt
ogeo
grap
hica
l afin
ities
of
vasc
ular
gen
era
in s
ubal
pine
fore
sts
of G
uate
mal
a.(N
= 1
03) a
nd C
osta
Rica
(N
= 1
27).
GU
ATE
MA
LA
Wid
e Te
mpe
rate
H
olar
ctic
. A
ustr
al-
Cos
mop
olita
n W
ide
Trop
ical
N
eot
ropi
cal
anta
rctic
8
Tree
s (6)
Shru
bs (2
2)
Her
bs (7
0)
Clim
ber
Fern
s (4
Arb
ulus
Agu
w
Rub
us
Agr
ostis
B
ruch
ypod
ium
B
rom
us
Cul
umug
rost
is
Cur
dam
ine
Cur
ex
Ech
euer
iu
Ery
sim
um
Fesr
ucu
Gul
ium
G
enriu
nu
Ger
uniu
m
Hie
ruci
um
Sen
ecio
S
ymph
oric
urpu
s
Luzu
lu
Mic
ropl
eura
M
onlia
O
enot
heru
P
enst
emon
P
W
Runu
ncul
us
Sulu
ia
Sed
wn
Tris
etum
V
uler
iunu
V
iola
W
elde
niu
Aln
us
Juni
peru
s P
inus
P
riirr
tts
Com
uros
tuph
ylis
G
urry
u M
ahon
iu
Rib
es
Vuc
cini
um
Are
nuriu
D
ruh
C
usril
leju
C
eras
tium
C
irsiu
m
Ery
ngiu
m
Guu
ru
Huc
keliu
H
elen
ium
Lu
pinu
s M
ulile
nber
giu
Osb
vrliu
P
oten
tillu
S
ibtli
orpi
u
Aca
enu
Bac
chur
is
Fuch
sirr
Per
netty
a
Mue
hlen
beck
iu
Ore
oniy
rrhi
s 0r
thro
sunt
liu.s
R
igid
ella
S
isyr
incl
iitm
i
Sum
birc
us
Sol
anum
Bid
ens
Erig
rron
G
napl
uiliu
m
Lobe
lia
Oxu
lis
Pla
ntug
o St
uchy
s
Asp
leni
um
Cte
nopt
eris
Brtd
dlej
u
A rc
hibu
cchu
ris
Ces
trum
D
endr
ophf
horu
H
olod
iscr
rs
Dry
mur
ia
Pep
erom
iu
Aph
anuc
tis
Hul
eniu
A
geru
tinu
Ciii
onol
uenu
Fl
eisc
hmun
niu
L.uc
hem
illu
Mel
ampo
dium
M
onrr
inu
Oxy
lobu
s
Rot
tiure
a
Rol
duna
S
teui
u V
erhe
sinu
Piq
ueriu
R
elbr
miu
m
Rm
nuns
clnr
lziu
S
uha:
ia
Turr
schi
u T
ripho
ru
Wer
neriu
W
Pol
ystic
hron
0
2.
5’
Cy.
stop
teris
L

G . ISLEBE et al., Phytogeographical comparisons in neotropical (subalpine) forests 79
80 Feddes Repert., Berlin 105 (1994) 1-2
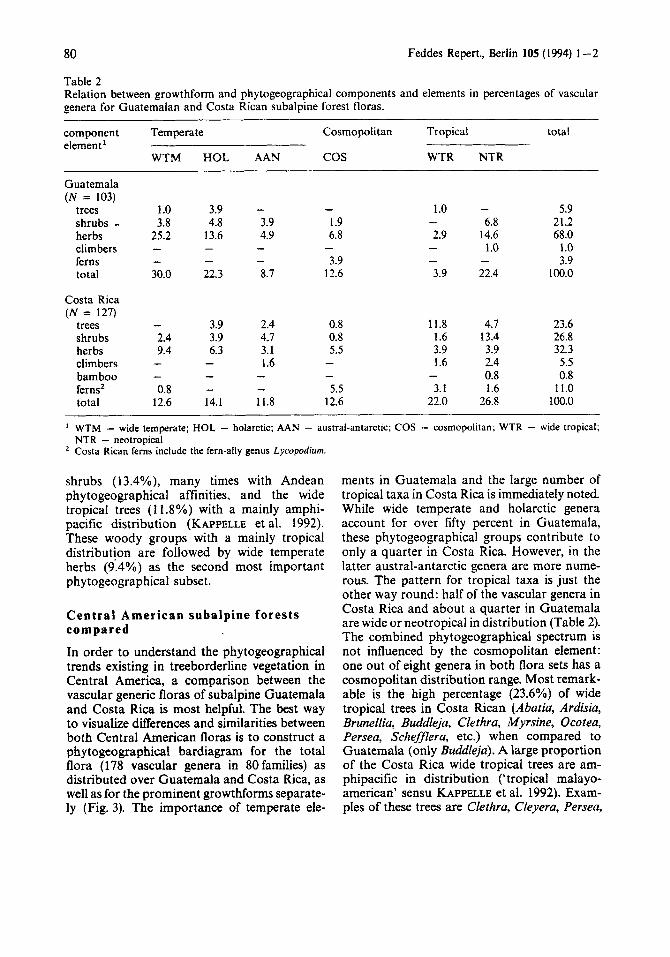
Table 2 Relation between growthform and phytogeographical components and elements in percentages of vascular genera for Guatemalan and Costa Rican subalpine forest floras.
component Temperate Cosmopolitan Tropical total element’
WTM HOL AAN cos WTR NTR
Guatemala ( N = 103)
trees shrubs - herbs climbers ferns total
Costa Rica ( N = 127)
trees shrubs herbs climbers bamboo ferns’ total
1 .o 3.9 - 3.8 4.8 3.9
25.2 13.6 4.9 - - - - - - 30.0 22.3 8.7
- 3.9 2.4 2.4 3.9 4.7 9.4 6.3 3.1 - - 1.6
0.8 - - -
- - 12.6 14.1 11.8
- 1.9 6.8
3.9 12.6
-
0.8 0.8 5.5 - - 5.5
12.6
1 .o - - 6.8 2.9 14.6 - 1 .o 3.9 22.4 - -
11.8 4.7 1.6 13.4 3.9 3.9 1.6 2.4 - 0.8 3.1 1.6
22.0 26.8
5.9 21.2 68.0
1 .o 3.9
100.0
23.6 26.8 32.3
5.5 0.8
11.0 100.0
WTM - wide temperate; HOL - holarctic; AAN - austral-antarctic; COS - cosmopolitan; WTR - wide tropical; NTR - neotropical Costa Rican ferns include the fern-ally genus Lycopodium.
shrubs (13.4%), many times with Andean phytogeographical affinities, and the wide tropical trees (11.8%) with a mainly amphi- pacific distribution (KAPPELLE et al. 1992). These woody groups with a mainly tropical distributjon are followed by wide temperate herbs (9.4%) as the second most important phytogeographical subset.
Central American subalpine forests compared
In order to understand the phytogeographical trends existing in treeborderline vegetation in Central America, a comparison between the vascular generic floras of subalpine Guatemala and Costa Rica is most helpful. The best way to visualize dflerences and similarities between both Central American floras is to construct a phytogeographical bardiagram for the total flora (178 vascular genera in 80 families) as distributed over Guatemala and Costa Rica, as well as for the prominent growthforms separate- ly (Fig. 3). The importance of temperate ele-
ments in Guatemala and the large number of tropical taxa in Costa Rica is immediately noted. While wide temperate and holarctic genera account for over fifty percent in Guatemala, these phytogeographical groups contribute to only a quarter in Costa Rica. However, in the latter austral-antarctic genera are more nume- rous. The pattern for tropical taxa is just the other way round: half of the vascular genera in Costa Rica and about a quarter in Guatemala are wide or neotropical in distribution (Table 2). The combined phytogeographical spectrum is not influenced by the cosmopolitan element: one out of eight genera in both flora sets has a cosmopolitan distribution range. Most remark- able is the high percentage (23.6%) of wide tropical trees in Costa Rican (Abatia, Ardisia, Brunellia, Buddleja, Clethra, Myrsine, Ocotea, Persea, Schefflera, etc.) when compared to Guatemala (only Buddleja). A large proportion of the Costa Rica wide tropical trees are am- phipacific in distribution (‘tropical malayo- american’ sensu KAPPELLE et al. 1992). Exam- ples of these trees are Clethra, Cleyera, Persea,

G. ISLEBE et al., Phytogeographical comparisons in neotropical (subalpine) forests
Nora proportions phflogeographical componenrs (2) cosmoporiran
COS = cosmopolitan
(3) tropicel Guatemala (I) temperate
WTM =wide temperate HOL = holarctic WTR =wide tropical
Costa Rica AAN = austral-antarctic NTR = neotropical
81
I
nr. of genera 30
25
20
15
10
5
0 WTM HOL AAN COS WTR NTR WIN HOL AAN COS WTR NTR
10
5
0 WTM HOL AAN COS WTR NTR
nr. of genera 30
25
20
15
10
5
0 WTM HOL AAN COS WTR NTR
Styrax and Symplocos. Instead of trees, herbs are the dominant growthform in subalpine Guatemala, many of which are wide temperate (among others Agrostis, Brornus, Echeueria, Festuca, Micropleura, Penstemon, Ranunculus and Trisetum) or holarctic (e.g. Draba, Cera- stium, Gaura, Helenium, Muhlenbergia and Po- tentilfa). In Costa Rica on the other hand there are proportionally less temperate herbs. Here, the number of wide tropical herbs is slightly larger than in the Guatemalan subalpine forests
due to the presence of genera such as Begonia, Phytolacca and Pitcairnia. Considering shrubs it can be noted, that neotropical shrub genera are twice as important in Costa Rica (with Cavendishia, Diplostephium, Disterigma, Ma- cleania, Monochaetum, etc.) as they are in Gua- temala. Austral-antarctic shrubs are also more frequent in Costa Rica (Desfontainea, Gaul- rheria, Ugni), while wide temperate shrubs are a more common feature in Guatemala (Agave, Symphoricarpus). At this occasion it is impor-
6 FddU Repert. 105 (1994) 1-2

82
temperate component
wide temperate element ( 1 1 )
holarctic element ( 1 2)
austral-antarctic element (6)
cosmopolitan component 0 cosmopolitan element (8)
tropical component
a wide tropical element (3)
E3 neotropical element (11)
Feddes Repert., Berlin 105 (1994) 1-2
Discussion and conclusions The Guatemalan subalpine flora is dominated by the temperate element (60%), whereas the Costa Rican subalpine zone shows a dominance of tropical taxa (48%). Due to the absence of other Central American subalpine vegetation areas we can conclude that a first approach on the generic level seems satisfactory to under- stand the present-day phytogeographical pat- tern governing the distribution of subalpine vascular floras in Central America. The generic approach earlier offered valuable accounts on the mountain floras of other neotropical regions (CLEEF 1980; G u m 1992; KAPPELLE etal. 1992). The present results exhibit a biogeo- graphical gradient in a north-south direction, with more temperate vascular genera in sub- alpine Guatemala and more tropical taxa in subalpine Costa Rica.
If we compare our vascular generic data sets with Mexican montane floras (VEL~ZQUEZ 1993), then we note a great phytogeographical affinity between the Guatemalan and Mexican subalpine vegetation (ISLEBE & VELAZQUEZ, in press). About 90% of all Guatemalan subalpine vascular genera occur also in pine forest com- munities of the Mexican Neovolcanic Range, thus supporting RZEDOWSKI’S concept of a ‘Megamexico’ (RZEDOWSKI 1991). However, a phytogeographical gradient analysis on the plant community level shows ecoclimatological differences, which advocate the division of ‘Megamexico’ into phytocoenologically more sound units (ISLEBE & VELAZQUEZ, in press). On basis of a phytogeographical comparison between montane oakforests in Costa Rica and Colombia (KAPPELLE et al. 1992) and own observations we might state that the Costa Rican subalpine flora also displays a close relationship with the subalpine northern Andes, at least concerning the shrubs. However, a more detailed analysis on this topic is needed to fully understand the phytogeographical position of subalpine Costa Rica, between both northern Central America (Guatemala) and the tropical Andes (Colombia, Ecuador).
Two major geographical barriers have influenced the floral composition of the entire Central American Region, namely (1) the Panamanian Isthmus and (2) the Nicaraguan Depression (KAPPELLE et al. 1992). The latter
Fig. 4 Proportional distribution of phytogeographical ele- ments on basis of 51 vascular genera occurring in both Guatemalan and Costa Rican subalpine forests Absolute numbers of genera are given between brackets. Note that over half o f the common genera are temperate in their distribution (57%)
tant to mention, that we must be aware of the fact, that many species recorded for either Guatemalan or Costa Rican subalpine forests can occur in one or the other country, in the montane forests below, or the alpine vegetation above the subalpine belt. An example is the genus Oenothera, which has been found in Guatemalan subalpine forests, but not in their equivalent in Costa Rica, although it is known from Costa Rican secondary plant communities in the montane belt (KAPPELLE et al., in press).
Let us now take a look at the vascular genera the Guatemalan and Costa Rican subalpine forest belts have in common. Fig. 4 presents the phytogeographical spectrum for the 51 vascular genera shared by both countries. Over half of these genera (56.9%) are temperate in distri- bution, most of them being wide temperate (21.6%) or holarctic (23.5%). OnIy a tenth of the shared genera (1 1.8%) have an austral- antarctic distribution pattern. This spectrum suggests that, in Central America, migration of holarctic genera (Pnmus, Ribes, Vaccinium, etc.) to the south has dominated over the migration of austral-antarctic genera (Acaena, Baccharis, Orthrosanrhus, etc.) to the north. Neotropical genera shared (e.g. Ageratina, Lachemilla, Mon- nina, Relbuniurn and Roldana) make up over a fifth (21.6%) and probably originated within the larger study area (Central America) or nearby neotropical mountain ranges (northern Andes). This implies a shorter migration route than austral-antarctic genera have. Wide tropi- cal genera found in both subalpine areas are few (Buddleja, Halenia and Peperomia).

G . ISLEBE et al., Phytogeographical comparisons in neotropical (subalpine) forests 83
has been a significant barrier for representatives of the Pinaceae. While this conifer family is present in the lowlands of Guatemala, Belize, Honduras and Nicaragua (i.e. Pinus caribea), it could not reach and colonize Costa Rica. If this circumstance is only due to a phytogeogra- phical barrier in a broad sense or if competition is another main cause, remains unknown.
During Pliocene times a connection was established through the isthmian region with the South American subcontinent (GRAHAM 1992). The uplift of the northern Andes took place during the last five million years (Hoo- GHIEMSTRA 1984; VAN DER HAMMEN 1989). This made an increased biotic interchange between North and South America possible (RAVEN & AXELROD 1974; GENTRY 1982).
Still little is known about the effect of the last glacial period on the natural environment. Paleo-ecological data are actually completely lacking for the high-elevated study area in Guatemala (ISLEBE, in prep.), while two decades ago HASTENRATH (1974) already found clear evidence of glaciation in the Sierra de 10s Cuchumatanes. LEYDEN (1984 and 1987) pro- vided indications of vegetation recovery in the Guatemalan Peten lowlands after Pleistocene aridity between 11,000- 10,000 BP. The latest results from paleo-ecological data available from Costa Rica show that the upper timber line was about 600 m displaced downslope during the last glacial maximum (MARTIN 1964; H~~GHIEMSTRA et al. 1992). Repeated lowering of the upper forest line in the Cordillera de Talamanca during the Plio-Pleistocene made speciation and present-day diversity possible (CLEEF & CHAVERRI 1992). Dry habitat species from Mexico and Guatemala acquired better possibilities for a southward migration during dry glacial conditions. At that time the pa- ram0 extention was at its maximum and ‘inter-neotropic-alpine’ distances were minimal (CLEEF & CHAVERRI 1992). However, as is mentioned above, the Nicaraguan Depression remained a major barrier during that period.
At present the Cordillera de Talamanca is the southernmost distribution centre of some holarctic elements like Comarostaphylis, Cir- sium, Garrya, Mahonia and Romanschulzia (CLEEF & CHAVERRI 1992). In contrast, some neotropical genera obtain their northernmost distribution in Guatemala, like Werneriu. Only
one true paramo element (Aphanactis) is present in the Guatemala subalpine forests and one (Myrrhidendron) in subalpine Costa Rica.
The high percentages of the temperate floral element in the subalpine zone is partly due to harsh environmental conditions prevailing in this vegetation belt. Local climatic fluctuations like day-night temperature oscillations, precipi- tation patterns with about five dry months and increased radiation are in favour of better- adapted temperate taxa. The rainfall regime in Costa Rica most probably favours a high diversity of trees and shrubs. Local speciation depending on factors like habitat .diversity may also result in generic differentiation as is the case with the formerly broad-sensed genera Eupatorium and Senecio in the Asteraceae occupying Mexican and Guatemalan highlands (e.g. KING & ROBINSON 1987). These tendencies are discussed by RZEDOWSKI (1991) in his attempt to explain the diversity and origin of the Mexican flora. He calculated the number of species per composite genus and found similar ratios for Guatemala (4.3 spp./genus), the adjacent Mexican region of Chiapas (4.2) and the Valley of Mexico (3.6).
Acknowledgements The first author was financially supported by the Karl Benz und Gottlieb Daimler-Stiftung at Ladenburg, Germany. During fieldwork he was assisted by Renate Hiibner, Cruz Jerbnimo CarilIo and Frank Gernaert. Karl-Uwe Krarner and Harold Robinson helped to identify Guatemalan ferns and Asteraceae, respectiv- ely. The second author was supported by grant nr. W 84-331 of the Netherlands Foundation for the Advancement of Tropical Research (WOTRO) to Antoine M. Cleef and by additional funds of the University of Amsterdam. Costa Rican plant speci- mens were mainly identified with help of Luis Poveda and Nelson Zamora. Antoine M.Cleef and Henri Hooghiemstra continuously provided logistic and scientific assistence.
References ANDERSON, T. H.; 1968, First evidence for glaciation
in the Sierra de 10s Cuchumatanes Range, Northwestern Guatemala. Geol. SOC. Amer. Special Paper, 121, 20 pp.
BARTRAM, E. B., 1949, Mosses of Guatemala. Field Mus. Nat. Hist., Bot. Ser., 26, 442 pp.
BEAMAN, J. H., 1965, A preliminary ecological study of the alpine flora of Popocattpetl and Iztac- cihuatl. Bol. Soc. Bot. Mex., 29, 63 - 75.

84 Feddes Repert., Berlin 105 (1994) 1-2
HOLDRIDGE, L. R.; W. C. GRENKE; W. H. HATHEWAY; T. LIANG; J. Tosijr., 1971, Forest environments in tropical life zones a, A pilot study. Oxford.
HOOGHIEMSTRA, H., 1984, Vegetational history of the high plane of Bogota, Colombia: a continous record of the last 3.5 million years. Diss. Bot., 79, Cramer, Vaduz, 42- 66.
HOOGHIEMSTRA, H.; A. M. CLEEF; G. W. NOLDUS; M. KAPPELLE, 1992, Upper Quaternary vegetation dynamics and paIeoclimatotogy of the La Chonta bog area (Cordillera de Talamanca, Costa Rica). J. Quat. Sci. 7(3), 205-225.
HORN, S. P., 1989, Postfire vegetation development in the Costa Rican paramos. Madroiio, 36(2),
IMN, 1988, Catastro de Ias series de precipitaciones medidas en Costa Rica. Instituto Meteorolbgico Nacional, MIRENEM, San Jose, Costa Rica.
ISLEBE, G., 1993, Will Guatemala's Junperus - Pinus forests survive? Environm. Conserv.
ISLEBE, G.; A. M. CLEEF; J. A. VEL~ZQUEZ, In press, Especies leiiosas de la Sierra de 10s Cuchumatanes y la Cadena Volcanica, Guatemala. Acta Bot. Mex.
ISLEBE, G.; J. A. VELAZQUEZ, In press, Affinity among mountain ranges in Megamexico.
JIMENEZ, W.; A. CHAVERRI; R. MIRANDA; I. ROJAS, 1988, Approximaciones silviculturales al manejo de un robledal (Quercus spp.) en San Gerardo de Dota, Costa Rica. Turrialba, 38(3), 208-214.
KAPPELLE, M., 1993, Recovery after clearing of an upper montane Quercirs forest, Costa Rjca. Rev. Biol. Trop., 41, 47-56.
KAPPELLE, M.; A. M. CLEEF; A. CHAVERRI, 1989, Phytosociology of montane Chusquea - Quercus forests, Cordillera de Talamanca, Costa Rica. Brenesia, 32, 73- 105.
KAPPELLE, M.; N. ZAMORA; T. FLORFS, 1991, Flora lefiosa de la zona aka (2000-3819111) de la Cordillera de Talamanca, Costa Rica. Brenesia, 34,121-144.
KAPPELLE, M.; A. M. CLEF; A. CHAVERRI, 1992, Phytogeography of Talamanca montane Quernrs forests, Costa Rica. I . Biogeogr., 19, 299-315.
KING, R. M.; H. ROBINSON, 1987, The genera of the Eupatorieae (Asteraceae). Monogr. Syst. Bot. Missouri Bot. Gard., 22, 1 - 576.
LEYDEN, B., 1984, Guatemalan forest synthesis after Pleistocene aridity. Proc. Nat. Acad. Sci. U.S.A.,
LEYDEN, B., 1987, Man and climate in the Maya lowlands. Quat. Research, 28, 407-414.
LUNA, I.; L. ALMEIDA; J. LLORENTE, 1989, Florktica y aspectos fitogeogrhficos del Bosque Mes6filo de Montaiia de las caiiadas de Ocuilan, Estados de Morelos y Mtxico. An. Inst. Biol. Univ. Aut6n. Mtxico, Ser. Bot., 59( I), 63 - 87.
747 pp.
93-114.
81, 4856-4859.
CASTILLO, R., 1984, Geologia de Costa Rica, Una sinopsis. Editorial de la Universidad de Costa Rica, San Pedro, Costa Rica. 182 pp.
CLEEF, A. M., 1979, The phytogeographical position of the Neotropical vascular paramo flora with special reference to the Colombian Cordillera Oriental. In: K. LARSEN; L. B. HOLM-NIELSEN (eds.), Tropical Botany. London. 175 - 184.
CLEEF, A. M., 1980, Posicibn fitogeograficade la flora vascular del paramo neotropical. Rev. Inst. Geogr. Augustin Codazzi, 7, 68-86.
CLEEF, A.M. ; A. CHAVERRI, 1992, Phytogeography of the paramo flora of the Cordillera de Tal- amanca, Costa R i a . In: H. BALSLEV; J. L. LUTEYN (eds.), Paramo: an Andean ecosystem under human influence. London. 45 -60.
DENGO, G., 1975, Paleozoic and Mksozoic tectonic belts in Mexico and Central America. In: A. E. M. NAIRN; F. G. STEHLI (eds.), The ocean basins and margins, The Gulf of Mexico and the Caribbean. Vol. 3. New York. 283-323.
DENGO, G.; D. BOHNENBERGER, 1969, Structural development of northern Central America. Amer. Assoc. Petrol. Geol., Mem., 11, 203-220.
GENTRY, A. H., 1978, Floristic knowledge and needs in pacific tropical America. Brittonia, 30 (2),
GENTRY, A. H., 1982, Neotropical floristic diversity: phytogeographical connections between Central and South America: Pleistocene climatic fluctua- tions or an accident of the Andean orogeny:! Ann. Missouri Bot. Gard., 69, 557-593.
GOMEZ, L. D., 1986, Vegetacion de Costa Rica. EUNED, San JOG., Costa Rica.
G&MEZ, L. D.; J. M. SAVAGE, 1983, Searchers on that rich coast: Costa Rican field biology, 1400- 1980. In: D. H. JANZEN (ed.), Costa Rican Natural Histoiy. Chicago, 111. 1 - 11.
GOOD, R., 1974, The geography of flowering plants. London. 557 pp.
GRAHAM, A., 1992, Utilization of the isthmian land-bridge during the Cenozoic : paleobotanical evidence for timing, and the selective influence of altitudes and climate. Rev. Paleobot. Palynol., 72,
GUTTE, P., 1992, Die Herkunft hwhandiner zentral- peruanischer Gattungen: Versuch einer Floren- analyse. Feddes Repert., 103,209-214.
HASTENRATH, S., 1973, On the Pleistocene glaciation of the Cordillera de Talamanca, Costa Rica. 2. Gletscherkd.~.Glazialgeol.,9 (1-2), 105- 121.
HASTENRATH, S., 1974, Spuren pleistoziiner Vereisung in den Altos de Cuchumatanes, Guatemala. Eiszeitalter u. Gegenw., 25, 25 - 34.
HOLDRIDGE, L. R.; B. LAMB; M. BERTJZLL, 1950, Los bosques de Guatemala. Vol. I-V. Instituto de Foment0 de la Produccibn de Guatemala, Guate- mala. 175 pp.
t34- 153.
119- 128.

G. ISLEBE et al., Phytogeographical comparisons in neotropical (subalpine) forests 85
MARTIN, P., 1964, Paleoclimatology and a tropical pollen profile. In: Proc. 6th INQUA Conf. Warsaw, 1961, 309-312.
Perfil Ambiental, 1987, Perfil ambiental: la Republica de Guatemala. Univ. Rafael Landivar, ICATA, Guatemala, 384 pp.
RAVEN, P.; D. AXELROD, 1974, Angiosperm bio- geography and past continental movements. Ann. Missouri Bot. Card., 61, 539-673.
RODRIGUEZ, M. L., 1921, Note sur le vegetation du Guatemala. Bull. Soc. Bot. France, 67,356-360.
ROSEN; D. E., 1985, Geological hierarchies and biogeographic congruence in the Caribbean. Ann. Missouri Bot. Card., 72, 636-659.
RZEDOWSKI, J., 1978, Vegetacion de Mkxico. Ed. Limusa, Mkxico D.F. 432 pp.
RZEDOWSKI, J. 1991. Diversidad y origenes de la flora fanerogamica de Mexico. Acta Bot. Mex., 14,
SAPPER, K., 1894, Grundziige der physischen Geo- graphie von Guatemala. Petermanns geogr. Mitt., Erganzungsh., 113.
SMITH, D. L., 1985, Caribbean plate relative motions. In. F. G. STEHLI; S. D. WEBB (eds.), The great American biotic interchange. New York. 17-48.
S~RENSEN, T., 1948, A method of establishing groups of equal amplitude in plant sociology based on similarity of species content. Det. Kong. Danske Vidensk. Selsk. Biol. Skr., 5(4), 1-34.
STANDLEY, P. C., 1934- 1937, Flora of Costa Rica. Field Mus. Nat. Hist., 18(1), Publ. 391,1116 pp.
STANDLEY, P. C., ed., 1946- 1977, Flora of Guate- mala. Field Mus. Nat. Hist., Bot. Ser., 24.
STEYERMARK, J., 1950, Flora of Guatemala. Ecology,
3-21.
31, 358-372. ULLOA, c . ; P. M. JBRGENSEN, 1993, Arboles y
arbustos de 10s Andes del Ecuador. AAU re-
Appendix list of 178 vascular genera found in Central American subalpine forests P t e r i d o p h y t a Adian thaceae
Adiantum L. Eriosorur F ~ E
Aspleniaceae Asplenium L.
Athyriaceae Cystopteris BERNH.
Blechneaceae Blechnum L.
Dryopteridaceae Dryopteris ADANS. Polystichum ROTH
Hymenophyllum SMITH H ymenophyllaceae
port 30. Aarhus University. Aarhus, Danmark.
VAN DER HAMMEN, T., 1989, History of the montane forests of the northern Andes. PI. Syst. Evol., 162,
VAN DER HAMMEN, T.; A. M. CLEEF, 1986, Develop- ment of the high Andean paramo flora and vegetation. In: F. VWLLEUMIER; M. MONASTERIO (eds.), High altitude tropical biogeography. New York. 153 - 200.
V~SQUEZ, A., 1983, Soils. In: D. H. JANZEN (ed.), Costa Rican Natural History. Chicago. 63-65.
VEL~ZQUEZ, J. A., 1993, Landscape ecology of Tlaloc and Pelado Volcanoes, Mexico, Ph. D. thesis. Hugo de Vries laboratory, University of Am- sterdam. 163 pp. ..
WAGNER, M., 1866, Uber den Character und Hohen- verhlltnisse der Vegetation in den Cordilleren von Veragua und Guatemala. Sitz.-Ber. Kgl. Bayr. Akad. Wiss., 1, Miinchen.
WEBER, H., 1958, Die Paramos von Costa Rica und ihre pflanzengeographische Verkettung mit den Hochanden Siidamerikas. Abh. Akad. Wiss. Lit. Mainz, math.-naturwiss. Kl., 3.
WEYL, R., 1980, Geology of Central America (Beitrage zur regionalen Geologie der Erde.) Berlin. 371 pp.
WILLIS, J. C., 1986, A dictionary of the flowering plants and ferns. Cambridge, UK.
264 PP.
109- 114.
Address of the authors: GERALD A. ISLEBE; MAARTEN KAPPELLE, University of Amsterdam, Hugo de Vries Laboratory, Kruis- laan 318, 1098 SM Amsterdam, Niederlande.
Manuskripteingang: 7. Juni 1993.
Lomariopsidaceae
L ycopodiaceae
Polypodiaceae
Elaphoglossum J. SMITH
Lycopodium L.
Campyloneurwn PRESL. Ctenopteris BLUME Grammitis Sw. Polypodium L.
Plagiogyria (KzE.) METT.
Vitraria SMITH
Plagiogyriaceae
Vittariaceae
G y m n o s p e r m a e
Cupressaceae Juniperus L.

86 Feddes Repert., Berlin 105 (1994) 1 -2
Chionolaena DC. Cirsium MILL. Diplostephium KUNTH Erigeron L. FIeischmannia SCH. BIP. Gnaphalium L. Helenium L. Hieracium L. Hypochoeris L. Melampodium L. Mikania WILLD. Neomirandea KING et ROBINSON Osbertia GREENE Oxylobus Moc. ex DC. Pentacalia CASS. Piqueria CAV. Pseudobaccharis CABRERA Roldana LA LLAVE Sabazia CASS. Senecio L. Sigesbeckia L. Stevia CAV. Verbesina L. Werneria KUNTH
Begoniaceae Begonia L.
Berberidaceae Berberis L. Mahonia N u n .
Bet ulaceae Alnus MILLER
Boraginaceae Hackelia OPIZ
Brassicaceae Cardamine L. Draba L. Erysimum L. Romanschulzia 0. E. ScnuLz
Brunelliaceae Brunellia RUIZ et P A V ~ N
Campanulaceae Centropogon PRESL. Lobelia L.
Caprifoliaceae Sambucus L. Symphoricarpos DUHAM. Virburnum L.
Cary ophyllaceae Arenaria L. Cerastium L. Drymaria WILLD. ex ROEM. et SCHULT. Stellaria L.
Crossopetalum P. BR. Maytanus MOLINA
Clethra GRONOV. ex L.
Celastraceae
Cletbraceae
Pinaceae
Monocotyledones Amar y llidaceae
Agave L. Bomarea MIRB.
Pitcairnia L'HBRIT.
Weldenia SCHULT.
Carex L. Rhynchospora VAHL Uncinia PERS.
Orthrosanthus SWEET RigidelIa LINDL. Sisyrinchium L.
Luzula DC.
Maianthemum WEBER
Malaxis SOLAND. ex SW. Triphora Nun.
Agrostis L. Brachypodium BEAUV. Bromus L. Calamagrostis ADANS. Chusquea KUNTH Festuca L. Muhlenbergia SCHREB. Poa L. Trisetum PERS.
Smilacaceae Smilax L.
Dicotyledones Apiaceae
Pinus L.
Bromeliaceae
Commelinaceae
Cyperaceae
Iridaceae
Juncaceae
Liliaceae
Orchidaceae
PoaCeae
Eryngium L. Hydrocotyle L. Micropleura LAG. Myrrhidendron COULT. et ROSE Oreomyrrhis ENDL. Tauschia SCHLECHD.
Aqui foliaceae Hex L.
Araliaceae Oreopanax DECNE. et PLANCH. Schefflera J. R. et G. FORST.
Agerutina 0. HOW. Aphanactis WEDD. Archibaccharis HEERING Baccharis L. Bidens L.
Asteraceae

G. ISLEBE et al., Phytogeographical comparisons in neotropical (subalpine) forests 87
Clusiaceae CIusia L.
Crassulaceae Echevaria DC. Sedum L.
Cunoniaceae Weinmannia L.
Ericaceae Arbutus L. Cavenchshia LINDL. Comarostaphylis ZUCC. Disterigma NIEDENZU Gaultheria KALM ex L. Macleania HOOK. Pernettya GAUDICH. Vaccinium L.
Abatia Rurz et PAV&N
Garrya DOUGL.
Gentiana L. Halenia BORCKH.
Geraniaceae Geranium L.
Grossulariaceae Escallonia M m s ex L. Ribes L.
Hypericum L.
SaIvia L. Stachys L.
Ocotea AUBL. Persea GAERTN.
Buddleja Housr. ex L. Desfontainea Rmz et PAV&N
Dendrophthora EICHL. Gaiadendron G. DON Phoradendron NUTT.
Miconia Rmz et PAV&N Monochaetum (DC.) NAUD.
Myrica L.
Ardisia SW. Myrsine L.
Myrcianthes BERG. Ugni TURCZ.
Fuchsia L. Oenothera L.
Oxalidaceae Oxalis L.
Flacourtiaceae
Garryaceae
Gen tianaceae
H ypericaceae
Lamiaceae
Lauraceae
Loganiaceae
Loranthaceae
Melastomataceae
M yricaceae
M yrsinaceae
Myrtaceae
Onagraceae
Papaveraceae Bocconia L.
Papilionaceae Lupinus L.
Phytolaccaceae Phytoiacca L.
Piperaceae Peperomia RUIZ et P A V ~ N
Plantaginaceae Plantago L.
Polygalaceae Monnina Rurz et PAV&N
Polygonaceae Muehlenbeckia MEISN. Rumex L.
Portulacacede Montia L.
Ranunculaceae Ranunculus L.
R hamnacede Rhamnus L.
Rosaceae Acaena M u m ex L. Hesperomeles LINDL. Holodiscus (C. KOCH) MAXIM. Lachemilla RYDB. Potentilla L. Prunus L. Rubus L.
Dydimaea HOOK. f . Galium L. Nertera B. et SOL. ex GAERTN. Relbuniwn BENTH. et HOOK. f.
Zanthoxylum L.
Castilleja MUTIS Hemichaena BENTH. Penstemon SCHMID. Sibthorpia L.
Cestrum L. Sobnum L.
St yracaceae Styrax L.
S ymplocaceae Symp[ocos L.
Theaceae Cleyera THUNB. emend. DC.
Urticaceae Phenax WEDD.
Valerianaceae Valeriana L.
Violaceae Viola L.
Winteraceae Drymis FORST.
Rubiaceae
Rutaceae
Scrophulariaceae
Solanaceae




















