
A new method using neuromagnetic stimulation to measure conduction time within the cauda equina
14
ELSEVIER Electroencephalographyand clinical Neurophysiology 101 (1996) 153-166 Q A new method using neuromagnetic stimulation to measure conduction time within the cauda equina P.J. Maccabee a,*, M.E. Lipitz a, T. Desudchit a R.W. Golub c, V.W. Nitti c, j.p. Bania a, J.A. Wilier a, R.Q. Cracco a, j. Cadwell a, G.C. Hotson d, L.P. Eberle b, V.E. Amassian a,b Departments of a Neurology, t, Physiology, c Surgery and a Radiology, State University of New York, Health Science Center at Brooklyn, Box 35, 450 Clarkson Avenue, Brooklyn, NY 11203, USA Accepted for publication: 11 December 1995 Abstract Using principles derived from electric field measurements and studies of phrenic nerve in vitro, neuromagnetic stimuli in humans were predicted to excite selective low threshold sites in proximal and distal cauda equina. Physical models, in which induced electric fields were recorded in a segment of human lumbosacral spine immersed in a saline filled tank, supported this prediction. Conclusions from the model were tested and confirmed in normal human subjects. Ipsilateral motor evoked potentials were elicited in lower limb muscles and striated sphincters by magnetic coil (MC) stimulation of both proximal and distal cauda equina. Over proximal cauda equina a vertically oriented MC junction and cranially directed induced current elicited a newly identified compound muscle action potential (CMAP). The F response latency and lack of attenuation when the target muscle was vibrated suggest that the proximal response is a directly elicited M response arising near or at the rootlet exit zone of the conus medullaris. Over distal cauda equina, lumbar roots were optimally excited by a horizontally oriented MC junction, and sacral roots by an approximately vertically oriented MC junction, eliciting CMAPs with similar appearance but shorter latency consistent with the known intrathecal lengths of the lower lumbar and sacral nerve roots. The induced current was usually most effective when directed towards the spinal fluid filled thecal sac. Normal subjects showed stable CMAP onset latencies elicited at proximal and distal cauda equina despite wide variation in amplitude. Thus, cauda equina conduction time can be directly calculated. This new method may improve the detection and classification of peripheral neuropathies affecting lower limbs and striated sphincters. Keywords: Neuromagneticstimulation;Cauda equina; Conductiontime; Lumbosacral spine; Compoundmuscle action potential(CMAP) 1. Introduction Neuromagnetic stimulation of lumbosacral roots has conventionally been performed using a centrally placed round magnetic coil (MC) which is bisected by the long axis of the vertebral column and sacrum (Chokroverty et al., 1990). By moving the coil in a cranial or caudal direction, the windings may be positioned to parallel and directly overlay targeted nerve roots and/or their neuro- foramina. Despite movement of the coil, the elicited re- * Corresponding author. Tel.: 718-270 2430; Fax: 718-270 1946. sponses appear to arise at fixed low threshold sites ("hot spots"), giving stable onset latencies despite widespread variations in amplitude (Ugawa et al., 1989; Britton et al., 1990). As previously demonstrated in the cervical spine, "hot spots" may be created when induced current chan- neled through neuroforamina results in excitation of nerve roots transiting or emerging there (Maccabee et al., 1991). For lumbar and sacral roots, the available evidence is also consistent with this neuroforaminal hypothesis, implying a site of excitation at the distal end of the cauda equina (Ugawa et al., 1989; Britton et al., 1990). In the human, low threshold sites to magnetic coil stimuli could also exist at the proximal in addition to the distal cauda equina. This suggestion partly evolves from 0924-980X/96/$15.00 © 1996 Elsevier Science Ireland Ltd. All rights reserved SSDI 0013-4694(95)00264-2 EEM 95078
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of A new method using neuromagnetic stimulation to measure conduction time within the cauda equina

ELSEVIER Electroencephalography and clinical Neurophysiology 101 (1996) 153-166
Q A new method using neuromagnetic stimulation to measure conduction
time within the cauda equina
P.J. Maccabee a,*, M.E. Lipitz a, T. Desudchit a R.W. Golub c, V.W. Nitti c, j.p. Bania a, J.A. Wilier a, R.Q. Cracco a, j. Cadwell a,
G.C. Hotson d, L.P. Eberle b, V.E. Amassian a,b Departments o f a Neurology, t, Physiology, c Surgery and a Radiology, State University o f New York, Health Science Center at Brooklyn, Box 35,
450 Clarkson Avenue, Brooklyn, NY 11203, USA
Accepted for publication: 11 December 1995
Abstract
Using principles derived from electric field measurements and studies of phrenic nerve in vitro, neuromagnetic stimuli in humans were predicted to excite selective low threshold sites in proximal and distal cauda equina. Physical models, in which induced electric fields were recorded in a segment of human lumbosacral spine immersed in a saline filled tank, supported this prediction. Conclusions from the model were tested and confirmed in normal human subjects. Ipsilateral motor evoked potentials were elicited in lower limb muscles and striated sphincters by magnetic coil (MC) stimulation of both proximal and distal cauda equina.
Over proximal cauda equina a vertically oriented MC junction and cranially directed induced current elicited a newly identified compound muscle action potential (CMAP). The F response latency and lack of attenuation when the target muscle was vibrated suggest that the proximal response is a directly elicited M response arising near or at the rootlet exit zone of the conus medullaris. Over distal cauda equina, lumbar roots were optimally excited by a horizontally oriented MC junction, and sacral roots by an approximately vertically oriented MC junction, eliciting CMAPs with similar appearance but shorter latency consistent with the known intrathecal lengths of the lower lumbar and sacral nerve roots. The induced current was usually most effective when directed towards the spinal fluid filled thecal s a c .
Normal subjects showed stable CMAP onset latencies elicited at proximal and distal cauda equina despite wide variation in amplitude. Thus, cauda equina conduction time can be directly calculated. This new method may improve the detection and classification of peripheral neuropathies affecting lower limbs and striated sphincters.
Keywords: Neuromagnetic stimulation; Cauda equina; Conduction time; Lumbosacral spine; Compound muscle action potential (CMAP)
1. Introduction
Neuromagnetic stimulation of lumbosacral roots has conventionally been performed using a centrally placed round magnetic coil (MC) which is bisected by the long axis of the vertebral column and sacrum (Chokroverty et al., 1990). By moving the coil in a cranial or caudal direction, the windings may be positioned to parallel and directly overlay targeted nerve roots a n d / o r their neuro- foramina. Despite movement of the coil, the elicited re-
* Corresponding author. Tel.: 718-270 2430; Fax: 718-270 1946.
sponses appear to arise at fixed low threshold sites ( "ho t spots"), giving stable onset latencies despite widespread variations in amplitude (Ugawa et al., 1989; Britton et al., 1990). As previously demonstrated in the cervical spine, "hot spots" may be created when induced current chan- neled through neuroforamina results in excitation of nerve roots transiting or emerging there (Maccabee et al., 1991). For lumbar and sacral roots, the available evidence is also consistent with this neuroforaminal hypothesis, implying a site of excitation at the distal end of the cauda equina (Ugawa et al., 1989; Britton et al., 1990).
In the human, low threshold sites to magnetic coil stimuli could also exist at the proximal in addition to the distal cauda equina. This suggestion partly evolves from
0924-980X/96/$15.00 © 1996 Elsevier Science Ireland Ltd. All rights reserved SSDI 0013-4694(95)00264-2 EEM 95078

154 P.J. Maccabee et al. / Electroencephalography and clinical Neurophysiology 101 (1996) 153-166
the fundamental principles elaborated in vitro, where a small twin coil was used to excite mammalian phrenic nerve immersed in a container filled with Ringer solution (Maccabee et al., 1993a). These studies demonstrate that a straight nerve in a homogeneous volume conductor is optimally excited by the peak negative-going first spatial derivative of the induced electric field parallel to the long axis of the nerve (Durand et al., 1989; Reilly, 1989; Roth and Basser, 1990). By contrast, "hot spots" created by a bend in the nerve or current channeling by high resistance barriers are optimally excited at the peak of the induced field (Maccabee et al., 1993a). Possible anatomical corre- lates of the in vitro findings are: (1) for the distal "hot spot," transit of nerve rootlets through or near interverte- bral foramina, and (2) for the proximal hot spot, the entry and exit of cauda equina rootlets into the spinal cord. In the following report, we first test the hypothesis using physical models of human lumbosacral spine immersed in saline. We then determine the optimal twin coil orientation suggested by the model in human subjects. These experi- ments, in part, were previously presented as abstracts (Maccabee et al., 1994a,b).
2. Methods
2.1. Stimulator, current reversing with or without phase damping, magnetic coils
Many details concerning the principles of magnetic stimulation have previously been described (Cadwell, 1990; Barker, 1991). Briefly, the time-varying magnetic field was generated by a coil energized by an MES-10 stimula- tor (Cadwell Laboratories, Kennewick, WA). The output of the stimulator was fed into a specially built phase damping device (designed by J. Cadwell) whose output consisted of either an unmodified, conventional MES-10 polyphasic pulse or a predominantly monophasic pulse. The induced major phase of the monophasic pulse exactly superimposed upon the first phase of the induced polypha- sic pulse; the amplitude of the second phase was 20% that of the first phase (Maccabee et al., 1993a). In addition, the device also provided selective reversal of current polarity. Thus, current direction could be reversed and current profile could be modified without moving the MC.
The output of this device was fed into 1 of 3 large twin coils wrapped in electrical insulation tape. It was antici- pated that if excitation of cauda equina lying deep below the surface was to be secured, wide diameter coils were required. However, the appropriate coil dimensions for subjects of different size had to be empirically derived and could not be theoretically predicted. In these coils the transverse, longitudinal outer diameters of each half coil were: 7.25 by 9.25 cm; 12.3 by 12.3 cm; 15.8 by 18.6 crn. The length of the junction of each of the 3 coils was 7.5, 6.0 and 11.5 cm. The smallest (and mid-size) coils are
illustrated in Figs. 1, 2, 4, 6, and 7). The largest coils were used to elicit normal values.
2.2. Electric field measurements in the physical model
As in earlier experiments on the cervical spine (Mac- cabee et al., 1991), a segment of human lumbosacral spine was immersed eccentrically within a large saline filled tank (30 cm in diameter, 12.4 cm height, 0.3 cm wall thickness) and near its inner wall. The electric fields were induced in the tank by the same twin coil. The electric field was predominantly monophasic. The descriptive terms "horizontal" and "ver t ica l" MC junction denote the long axis of the twin coil junction (and its continuation in the handle) in reference to the long axis of the vertebral column (Figs. 1 and 2); these terms are also used to describe MC orientations in the human. The electric field was measured across intervertebral neuroforamina and lon- gitudinally within the central sacral canal with a single coaxial cable (RG 174U, 2.57 mm OD) threaded through a 1 ml plastic pipette (see Fig. 1, in Maccabee et al., 1990). For measurements across neuroforamina, the distal 30 mm of the cable was bent at a right angle. The distalmost 10 mm of the outside insulation and shield of the cable was stripped and when submerged in isotonic saline, the volt- age drop was recorded across the 10 mm between cable shield and bared distal tip. The shield of the cable was grounded to the oscilloscope for single-ended recording. The electric fields recorded by the single co-axial cable were fed into a digital recording oscilloscope (Hitachi VC-6020).
2.3. Human subjects
Investigations were approved by the Institutional Re- view Board. The neuromagnetic stimulation was confined to the lower spine and the responses elicited in peripheral nerve rootlets or roots were recorded from lower limb muscles and /or the urinary and anal striated sphincters. Data were obtained in 7 male and 1 female volunteers who gave informed consent (age range 26-41 years).
2.4. Recording electrodes and stimulation technique in the human
Stimulation with one or more of 3 large twin MCs was tested on the lumbosacral roots. The orientation of the coils over the spine is described in Results. Compound muscle action potentials (CMAPs) were recorded with an Excel Electromyography System (Cadwell Laboratories). Standard 10 mm Ag-AgCI disk electrodes (Medelec) were placed over the following muscles in both lower limbs: vastus medialis (G1 on muscle belly, G2 over medial patella); tibialis anterior (G1 on the muscle belly at mid- shin, G2 3-5 cm distal); abductor hallucis (G1 1 cm posterior and 0.5-1.0 cm inferior to the navicular promi-

P.J. Maccabee et al./Electroencephalography and clinical Neurophysiology 101 (1996) 153-166 155
nence, G2 over the medial extensor hailucis longus). In 1 subject, G1 was also placed over the flexor hallucis brevis instead of the abductor hallucis. When recording the H soleus reflex, electrodes were conventionally placed over the soleus muscle. The ground electrode (Medelec) was round (32 mm in diameter) and was placed over the posterior thigh. Limb recordings were performed with the subject prone. Pillows were placed beneath the lower abdomen so as to slightly flex the lower spine.
Recordings from the striated sphincters utilized com- mercially available electrode assemblies. Initially, the ex- ternal anal sphincter (EAS) recordings were obtained by inserting a cylindrical soft sponge platform (length 40 mm; diameter 25 mm; Dantec 13L81), with thin disk recording electrodes positioned approximately mid-cylinder, 180 ° apart. Recordings were obtained simultaneously from left and right EAS (Fig. 3). Subsequently, we confirmed that reliable motor evoked responses from left or right rectal musculature could be obtained with the St. Mark 's gloved finger pudendal electrode (Dantec 13L40) when G1 was at the tip of the index finger and G2 was located over the middle phalanx (Swash and Snooks, 1986). By changing the depth of the finger, ipsilateral responses from both EAS and puborectalis (PR) could be individually obtained to stimulation of proximal and distal cauda equina (Swash and Snooks, 1986, 1992). Recordings from the urinary striated sphincter (USS) were obtained with a cylindrical
recording electrode mounted on the surface of a 10 French Foley catheter (Dantec 21L10), which was inserted by a urologist. Striated sphincter recordings were performed with the subject in a lateral decubitus position with the lower spine slightly flexed.
All recordings were fed into the preamplifier input of the EMG system. Negativity of G1 was displayed up. Filter settings were 5 or 10-3000 Hz. In the text, induced current directed in the long axis of the spine in a caudal to cranial direction is sometimes described as "cranially in- duced current"; that induced in a cranial to caudal direc- tion as "caudal ly induced current."
3. Results
3.1. Physical model
3.1.1. Sacral canal The physical model was studied to test the prediction
that the largest induced electric fields would he generated in the long axis of the sacral canal with the MC junction oriented longitudinally. By contrast, a horizontally oriented MC junction should induce the smallest field owing to a reduced width of the sacral canal compared to its length. Also, a junction axis 45 ° off midline should induce a substantial but not maximal field, large enough to dep01ar-
Ventral Aspect Record: Vertical probe in Sacral canal
Tip inferior $1 level Positive up
Mid- Junction rotation at $1 MC 2O%
-:.--. / / fi '~ "\\
I I I
I
%%% ~\ ~" /I/I
. , ,~.
,.." ,.. ~. . t I \ I
~ x % L I
20mV/mm
0.1 ms
Fig. 1. View of the ventral surface of a human sacrum. Longitudinal electric fields (mV/mm; positive up) are induced by the (small) twin MC and recorded approximately in the center of the long axis of the sacral canal. The tip (positive up) of the coaxial cable extends to the inferior S1 level and the center of the MC junction pivots about this point at 0 ° (horizontal), 45 ° (oblique), and 90 ° (vertical).
156 PJ. Maccabee et al. / Electroencephalography and clinical Neurophysiology lO1 (1996) 153-166
ize ipsilateral nerve fibers as suggested by the findings of Fig. 1.
In Fig. 1, a vertically oriented coaxial cable probe is fixed in place centrally within the sacral canal, i.e., approx- imately paralleling the vertical trajectory of the sacral rootlets before they angle ipsilaterally towards their ventral neuroforamina. In Fig. 1, the results confirm the predic- tions: the horizontal junction orientation induces no re- sponse: a 45 ° orientation induces a substantial response; a vertical orientation induces a maximal response. Thus, the findings in the model confirm the predictions.
3.1.2. Lumbar and sacral neuroforamina A further prediction was that the foramina containing
lumbar roots (e.g. L4-5 for the L4 root, L5-S1 for the L5 root) would be best excited with a roughly horizontal MC junction. This is because the plane of the electric field
Ventral Aspect Record: Neuroforamina Junction horizontal at L5-$1 MC 20% ~
Right , ~ , ~ . . . ~ ~ . . . -~.LS...~ Left
S2 - - ~ ~ , ~ s - - $2
$4 ~ - - S4
~--C-f ' ~ t 20mV/mm 01 ms
Ventral Aspect Record: Neuroforarnina Junction horizontal at $1 MC 2O%
!..3-4 - - ~ - - - - ~ 1.3-4
L4-5 L4-5
L5-S1 ~ ' ~ L5-$1
$1 ~ ' - - ~ Sl
$2 . . . ~ ~ . ; ~ ' ~ ~ . . - - ~ ' $2
$3 ~ ' ~ - - ~ , - $3
$4 S4
20mV/mm
01 ms
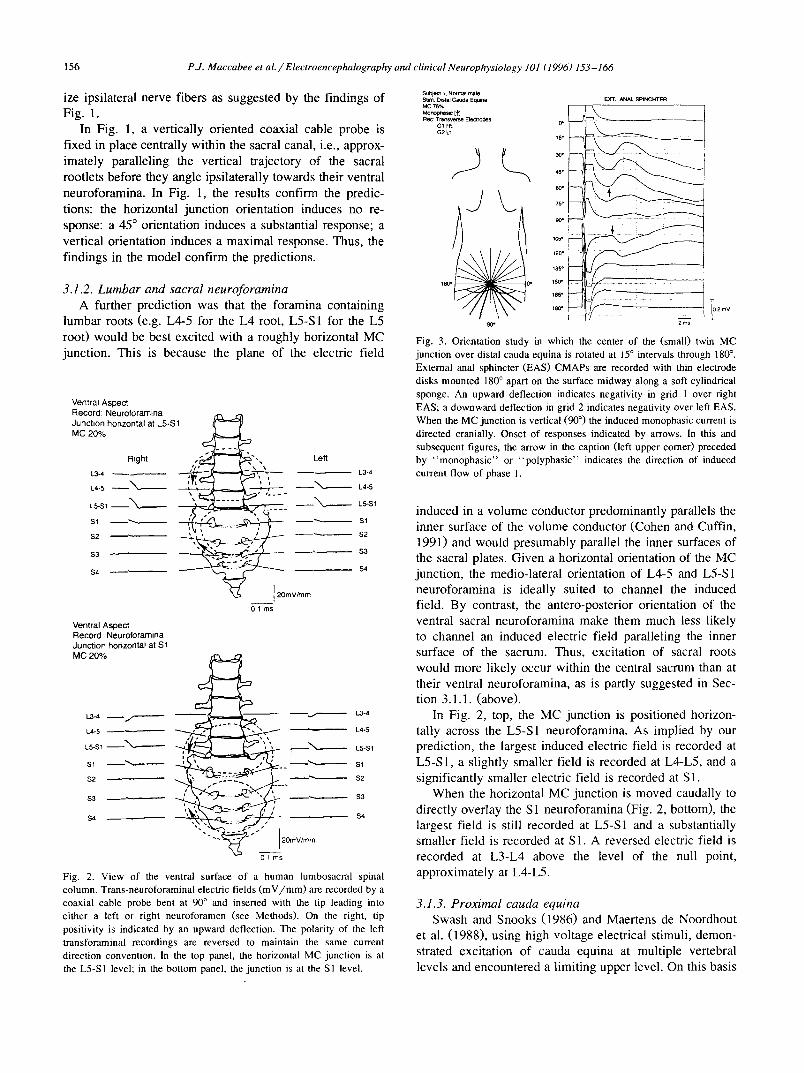
Fig. 2. View of the ventral surface of a human lumbosacral spinal column. Trans-neuroforaminal electric fields (mV/mm) are recorded by a coaxial cable probe bent at 90 ° and inserted with the tip leading into either a left or right neuroforamen (see Methods). On the right, tip positivity is indicated by an upward deflection. The polarity of the left transforaminal recordings are reversed to maintain the same current direction convention, in the top panel, the horizontal MC junction is at the L5-S1 level; in the bottom panel, the junction is at the S1 level.
SubjeCt 1. Normal male Stim: Dtstal Cau~a Eq~ina EXT, ANAL SPINCHTER MC 76%
Rec Transverse Electrodes ' ~ G1 Rt (~P
) \ '
135"
18CJ 150"
165" T
18~° [0 2 mV
90* 2 ms
Fig. 3. Orientation study in which the center of the (small) twin MC junction over distal cauda equina is rotated at 15 ° intervals through 180 °. External anal sphincter (EAS) CMAPs are recorded with thin electrode disks mounted 180 ° apart on the surface midway along a soft cylindrical sponge. An upward deflection indicates negativity in grid 1 over right EAS; a downward deflection in grid 2 indicates negativity over left EAS. When the MC junction is vertical (90 °) the induced monopbasic current is directed cranially. Onset of responses indicated by arrows. In this and subsequent figures, the arrow in the caption (left upper comer) preceded by "monophasic" or "polyphasic" indicates the direction of induced current flow of phase 1.
induced in a volume conductor predominantly parallels the inner surface of the volume conductor (Cohen and Cuffin, 1991) and would presumably parallel the inner surfaces of the sacral plates. Given a horizontal orientation of the MC junction, the medio-lateral orientation of L4-5 and L5-S 1 neuroforamina is ideally suited to channel the induced field. By contrast, the antero-posterior orientation of the ventral sacral neuroforamina make them much less likely to channel an induced electric field paralleling the inner surface of the sacrum. Thus, excitation of sacral roots would more likely occur within the central sacrum than at their ventral neuroforamina, as is partly suggested in Sec- tion 3.1.1. (above).
In Fig. 2, top, the MC junction is positioned horizon- tally across the L5-S1 neuroforamina. As implied by our prediction, the largest induced electric field is recorded at L5-S1, a slightly smaller field is recorded at L4-L5, and a significantly smaller electric field is recorded at S1.
When the horizontal MC junction is moved caudally to directly overlay the S 1 neuroforamina (Fig. 2, bottom), the largest field is still recorded at L5-S1 and a substantially smaller field is recorded at S1. A reversed electric field is recorded at L3-L4 above the level of the null point, approximately at L4-L5.
3.1.3. Proximal cauda equina Swash and Snooks (1986) and Maertens de Noordhout
et al. (1988), using high voltage electrical stimuli, demon- strated excitation of cauda equina at multiple vertebral levels and encountered a limiting upper level. On this basis

P.I. Maccabee et al. / Electroencephalography and clinical Neurophysiology l O1 (1996) 153-166 157
we speculated that an inhomogeneity would possibly arise at the interface between nerve rootlet and conus owing to the sudden change in conductance between the spinal fluid filled thecal sac and the conus. This was suggested by earlier studies in which an induced electric field in a saline conductor is abruptly displaced and deformed by a cylin- drical non-conducting inhomogeneity (Maccabee et al., 1991, Fig. 3, middle and right upper two traces). The maximal fields are then displaced to either side, at the outer boundary of the obstruction. Therefore, a low thresh- old excitation site might be expected at the junction of the proximal cauda equina and conus medullaris. This inhomo- geneity may be added to by the acute caudal angle of the nerve roots with the bend of the fibers as they enter each segment. I f the obstruction-nerve bend hypothesis were correct, then the direction of induced current would be critical for excitation; an induced current directed cranially would excite preferentially because of membrane current exiting at the rootlet exit zone.
3.2. Human studies: direct motor responses
3.2.1. Bilateral external anal sphincter responses ($2-$3- S4)
Initial studies simultaneously recorded right and left EAS responses from G1 and G2 recording electrodes (Fig. 3). The smallest MC was optimally positioned over the distal spine and its junction was rotated about its center at 15 ° intervals through 180 °. The small coil was used be- cause the two larger twin coils created unacceptable stimu- lus artifact. The largest ipsilateral CMAP was typically obtained when the axis of the MC junction was between 0 °
and 45 ° from the midline in different subjects a n d / o r sides. In Fig. 3, large responses are obtained at 30 -60 ° on the right (i.e., 30 -60 ° from the vertical; G1 negative up) and 105-120 ° on the left (i.e., 15-30 ° from the vertical; G2 negative down). When using monophasic current, re- sponse thresholds were usually a little lower when the induced current was directed cranially. However, in this subject, the threshold was slightly lower with caudally induced current (not shown). In Fig. 3, despite a constant MC output intensity, the responses are significantly attenu- ated as the junction axis approaches the vertical. It was deduced that the disappearance of the response with a vertical junction axis reflected simultaneous activation of G1 and G2 electrodes, whereas the ipsilateral responses were obtained when the junction axis was off the midline, i.e., when efficiently exciting ipsilateral but not contralat- eral sacral nerve roots.
3.2.2. Distal versus proximal foot muscle responses (S/, S2)
The hypotheses derived from the model were further tested in the flexor hallucis brevis muscle. A mid-size, vertically oriented, twin coil was positioned over the lower spine (Fig. 4). The coil was moved cranially in 2 cm steps from the coccyx (i.e., location 0). Coil position is refer- enced to the distal divergence of the MC junction. Two experiments are shown at MC output intensity of 50% (left) and 90% (right). On the left (MC 50% traces), at location 2, over the distal sacrum, the largest response is obtained at an approximate onset latency of 24.7 msec. (Note that at position 2, the center of the 6 cm MC junction, where the induced field should be maximal, is at
Su#ject 2, Normal male Stim: Distal and Proximal Cauda Equina Monophasic [~'] Re<: Rt FHB
F I
MC 50%
24
22
2O
IS
16 g
12
,o ~
6
4
2
0
--1
MC 90%
5 m s
Fig. 4. CMAPs from right FHB elicited at two different MC output intensities by the mid-size twin MC oriented vertically. The coil is moved in 2 cm intervals from position 0 at the coccyx to position 24, referenced to the distal divergence of the MC junction. At the lower MC output intensity (50%, left traces) the maximal amplitude CMAP over distal cauda equina was elicited at position 2 which corresponds to position 5 at the mid-point of the MC junction. At the higher MC output intensity (90%, right traces) the maximal amplitude CMAP over proximal cauda equina is elicited at positions 14 and 16 which corresponds to positions 17 and 19 (interpolated at position 18) at the mid-point of the MC junction. Therefore, the sites of optimal excitation are estimated to be (18 - 5) = 13 cm apart.
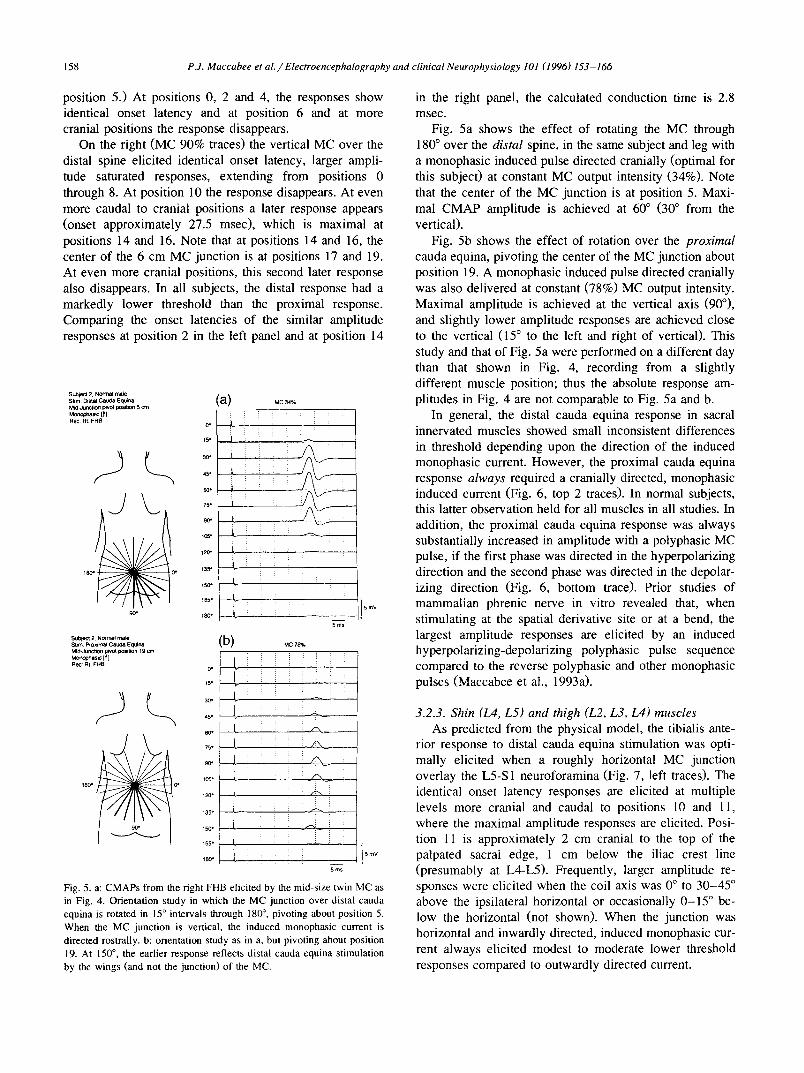
158 PJ. Maccabee et al./Electroencephalography and clinical Neurophysiology 101 (1996) 153-166
position 5.) At positions 0, 2 and 4, the responses show identical onset latency and at position 6 and at more cranial positions the response disappears.
On the right (MC 90% traces) the vertical MC over the distal spine elicited identical onset latency, larger ampli- tude saturated responses, extending from positions 0 through 8. At position 10 the response disappears. At even more caudal to cranial positions a later response appears (onset approximately 27.5 msec), which is maximal at positions 14 and 16. Note that at positions 14 and 16, the center of the 6 cm MC junction is at positions 17 and 19. At even more cranial positions, this second later response also disappears. In all subjects, the distal response had a markedly lower threshold than the proximal response. Comparing the onset latencies of the similar amplitude responses at position 2 in the left panel and at position 14
Su~)ect 2, N~n~t male StJm. Distal Cauda Equ~na (a) MK~-Junctlon pivo~ pos~on 5 cm MorK~ohasic It] RBC Rt FHB O ° ~ .
15"
45°
lC~°
180o Do 135"
t50*
155"
90* 180"
Sublect 2, Nocmal makB Stirn Ptoxin'~al CauOa Eq~in= Mid.unction pwot po~itio n 19 cm Mon~orlas0c [~] Rec Rt FHB
O"
15,
45° ~°
~ 75" 911" ~05 •
180 ~ G"
120"
135" 150 °
165"
180"
..r3 • • - -
i
.---L
5mY 5ms
(b ) , , : ,~
_J. .~__
5 m s
Fig. 5. a: CMAPs from the right FHB elicited by the mid-size twin MC as in Fig. 4. Orientation study in which the MC junction over distal cauda equina is rotated in 15 ° intervals through 180 °, pivoting about position 5. When the MC junction is vertical, the induced monophasic current is directed rostrally, b: orientation study as in a, but pivoting about position 19. At 150 °, the earlier response reflects distal cauda equina stimulation by the wings (and not the junction) of the MC.
in the right panel, the calculated conduction time is 2.8 msec.
Fig. 5a shows the effect of rotating the MC through 180 ° over the distal spine, in the same subject and leg with a monophasic induced pulse directed cranially (optimal for this subject) at constant MC output intensity (34%). Note that the center of the MC junction is at position 5. Maxi- mal CMAP amplitude is achieved at 60 ° (30 ° from the vertical).
Fig. 5b shows the effect of rotation over the proximal cauda equina, pivoting the center of the MC junction about position 19. A monophasic induced pulse directed cranially was also delivered at constant (78%) MC output intensity. Maximal amplitude is achieved at the vertical axis (90°), and slightly lower amplitude responses are achieved close to the vertical (15 ° to the left and right of vertical). This study and that of Fig. 5a were performed on a different day than that shown in Fig. 4, recording from a slightly different muscle position; thus the absolute response am- plitudes in Fig. 4 are not comparable to Fig. 5a and b.
In general, the distal cauda equina response in sacral innervated muscles showed small inconsistent differences in threshold depending upon the direction of the induced monophasic current. However, the proximal cauda equina response always required a cranially directed, monophasic induced current (Fig. 6, top 2 traces). In normal subjects, this latter observation held for all muscles in all studies. In addition, the proximal cauda equina response was always substantially increased in amplitude with a polyphasic MC pulse, if the first phase was directed in the hyperpolarizing direction and the second phase was directed in the depolar- izing direction (Fig. 6, bottom trace). Prior studies of mammalian phrenic nerve in vitro revealed that, when stimulating at the spatial derivative site or at a bend, the largest amplitude responses are elicited by an induced hyperpolarizing-depolarizing polyphasic pulse sequence compared to the reverse polyphasic and other monophasic pulses (Maccabee et al., 1993a).
3.2.3. Shin (L4, L5) and thigh (L2, L3, L4) muscles As predicted from the physical model, the tibialis ante-
rior response to distal cauda equina stimulation was opti- mally elicited when a roughly horizontal MC junction overlay the L5-S 1 neuroforamina (Fig. 7, left traces). The identical onset latency responses are elicited at multiple levels more cranial and caudal to positions 10 and 11, where the maximal amplitude responses are elicited. Posi- tion 11 is approximately 2 cm cranial to the top of the palpated sacral edge, 1 cm below the iliac crest line (presumably at L4-L5). Frequently, larger amplitude re- sponses were elicited when the coil axis was 0 ° to 30-45 ° above the ipsilateral horizontal or occasionally 0-15 ° be- low the horizontal (not shown). When the junction was horizontal and inwardly directed, induced monophasic cur- rent always elicited modest to moderate lower threshold responses compared to outwardly directed current.

P.J. Maccabee et al . / Electroencephalography and clinical Neurophysiology 101 (1996) 153-166 159
In Fig. 7, the onset latency of the maximal distal response at positions 10 and 11 is 15.4 msec. When the MC was subsequently positioned vertically over the spine (handle pointing cranially) and moved in 2 cm steps at 78% output, a proximal response (onset 17.8 msec) ap- peared at positions 12 through 22, which was maximal at positions 16-18 and which attenuated and disappeared at more caudal to cranial positions (note that at interpolated position 17, the center of the 6 cm MC junction is at position 20). In all 5 subjects studied, the proximal re- sponse to tibialis anterior was always optimally elicited when the MC junction was vertical. Comparing the onset latencies of the maximal amplitude responses in the left panel at positions 10, 11 and at positions 16, 18 in the right panel, the calculated conduction time is 2.4 msec.
When recording from vastus medialis (not shown), identical observations were generally found, although the horizontal MC junction was, as expected, optimally lo- cated more cranially, approximately over the L4-5 neuro- foramina. Also, the optimal position of the vertical MC junction over proximal cauda equina was displaced cra- nially 0-1 cm more than that required for tibialis anterior. In some normal subjects the distal vastus medialis re- sponse is biphasic, consisting of an early component (onset 8 -10 msec) and a late component (onset 15 msec). The early component was enhanced when the MC junction was angled 30-45 ° above the ipsilateral horizontal, giving a predominantly fused CMAP. By contrast, stimulation prox- imally in these subjects elicited the typical monophasic response. In one normal subject, the late component was
Subject 2, Normal male Stim Proximal Caucla Equina D~stal Divergence F)Silion 16 cm RBC Lt FHB
,;;--:,,~ if-- , i . i I
%'~ a;~ /t15
- - 10
- 2
Induced Current
Mono 1
Mono l
Po~y ~ .~
M C t ~ %
5ms
S m V
Fig. 6. Proximal cauda equina CMAPs in left FHB elicited by the mid-size twin MC held vertically at the optimal level (distal divergence at position 16). In the top trace, monophasic current is induced in a caudal to cranial direction. Second trace from top: reversal of monophasic current in this typical, thin, normal subject elicited no response, despite the same 100% MC output. Third trace from top: a slightly larger amplitude CMAP is elicited by a polyphasic pulse (phase 1 directed cranially, i.e., depolarizing at proximal cauda equina; phase 2 directed caudally, i.e., hyperpolarizing at proximal cauda equina). Bottom: a substantially larger amplitude response is elicited by the reverse polypha- sic pulse yielding a hyperpolarizing-depolarizing sequence at proximal cauda equina.
SU~)lect 2. N o r m a l male Stim: Distal Cauda Equlna (Lt)
Proximal Caucia Equina ( Rt ) Monophasic
Horizontal [in] Vertical [~]
Rec: Lt T ib Ant
MC 88*/. Me 78%
7 4 24
5 Io le
12
10 U
I ° ' ¢ 4
Z
.. 5 mv
5 ms
Fig. 7. CMAPs elicited by the large twin MC in the left tibialis anterior. Left traces: the distal cauda equina responses are elicited maximally by a horizontal MC junction (inwardly directed current) at positions l 0 and 1 l (interpolated to position 10.5). Right traces: when the MC junction is vertical and the monophasic induced current is directed cranially, the maximal proximal cauda equina response is elicited at positions 16 and 18 (interpolated to position 17 which corresponds to position 20 at the mid-point of the MC junction). Therefore, the sites of optimal excitation are estimated to be ( 2 0 - 0 . 5 ) = 9.5 cm apart (cf., 13 cm for FHB in Fig. 4).
attenuated by vibrating the vastus medialis, suggesting that it may be an H reflex (Zhu et al., 1992). In those subjects without a late component, a slightly lower threshold re- sponse was elicited by inwardly directed, induced current, as is always the case for tibialis anterior. Typically, the early response is thus elicited by both directions of current. However, in those normals showing both an early and late response in vastus medialis, and holding the MC junction horizontal, outwardly directed induced current elicited the early response and inwardly directed induced current elicited the late response.
3.3. Intensity series
An intensity series at proximal and distal cauda equina stimulation sites was performed in 5 normal subjects. The MC output intensities were chosen to elicit a range of CMAP amplitudes up to the maximum. Therefore, the MC output intensities were not increased in fixed incremental steps. All studies, as illustrated in Fig. 8, reveal that widespread fluctuations in CMAP amplitude are accompa-

160 P.J. Maccabee et al. / Electroencephalography and clinical Neurophysiology 101 (1996) 153-166
nied by stable onset latencies when the elicited amplitude exceeds 40-50% maximal. Below 40-50% maximal am- plitude, the elicited onset latencies were inconsistently delayed by up to 0 . 5 - ! . 5 msec.
3.4. Effect o f direction of induced current on latency
When stimulating lumbar roots adjusted to at least 50% of maximal amplitude, distal cauda equina responses did not reveal consistent CMAP onset latency shifts with reversal of current direction. Occasionally, when stimulat- ing sacral roots, inconsistent latency decreases up to 0.5 msec were observed with caudally directed induced current compared to cranially directed current.
3.5. F wave latency
It was considered important to measure F wave laten- cies to validate conclusions as to the sites of distal and especially proximal M wave excitation. Early in this part of the study it became apparent that careful measurement of F wave latency was required at an amplification that was appropriate for MC elicited M responses from cauda equina. Thus, in Fig. 9, at identical high gains (middle and bottom traces): (F + M - 1 ) / 2 = (43.9 + 2.5 - 1 ) / 2 = 22.7 msec. Thus the estimated conduction times from
Subject 3, Normal male MC: Output
Low to saturation Nonqinear
Monophasic Horizontal [in] Distal Vertical [q]
LT VASTUS MEDIALIS
Proximal )L_
i _ _
Distal
CE
LT TISIALIS ANTERIOR
Proxima E.
INTENSITY SERIES
Subject 4, Norma l male Rec: Rt. AH Polyphasic [~,.~' ]
Distal C E.
MC Stim. ( 100% |
~ P r o x i m a l C E . - -
--Med. Mal leo lus-
Electric f S t i m
- - Med. M a l l e o l u s - -
['-- Distal C. E. MC Stim. ]
100% L-Proximal C. E ~ - -
..... i ............. i + T1 T2 t8.8 22.8
m ........ -L
9
T1 2.50
'! i
10 mV +
- - q i
i
T1 18,0
i 0.2 m V
+
T2 43.9
_ _ +
T2 1 0 ms 21.7
Fig. 9. Comparison of latencies of F wave and cauda equina (CE) responses in AH. Top and bottom records: the motor responses to MC stimulation of proximal and distal cauda equina are illustrated at low gain and at high gain, respectively. Middle records illustrate motor responses to electrical stimulation at the medial malleolus at low and also at high gain to reveal F responses in 7 superimposed sequential sweeps,
distal and proximal excitation sites to the anterior horn cell are (22.7 - 18.0) = 4.7 msec and (22.7 - 21.7) = 1.0 msec, respectively at the same high amplification. At unequal gains (Fig. 9, top), different erroneous values would result. When recording from right AH in 4 normal subjects the corresponding latencies from anterior horn to distal cauda equina were 2.86, 4.36, 5.44 and 4.70 msec (mean 4.34 msec) and the latencies from anterior horn to the presumed rootlet exit zone were 0.86, 1.96, 2.14 and 1.0 msec (mean 1.48 msec).
Distal
LT. ABDUCTOR C E I CE
HALLUCIS
Proxim
5 ms
5 m Y
Fig. 8. Response to stimulus intensity increase. Distal and proximal cauda equina CMAPs are elicited in left vastus medialis, tibialis anterior, and abductor hallucis by a large twin MC.
3.6. Soleus muscle H and M responses
When recording from soleus in 5 normal subjects, motor responses were elicited by stimulation at 3 levels and H soleus reflexes were elicited simultaneously at two of these levels (Fig. 10): (1) at the popliteal fossa the conventional electrical stimulus elicited an M response followed by an H reflex (bottom trace); (2) at distal cauda equina the MC stimulus elicited an M response followed by a later wave which has been shown to be an H reflex (Britton et al., 1990; Chokroverty et al., 1990; Zhu et al.,
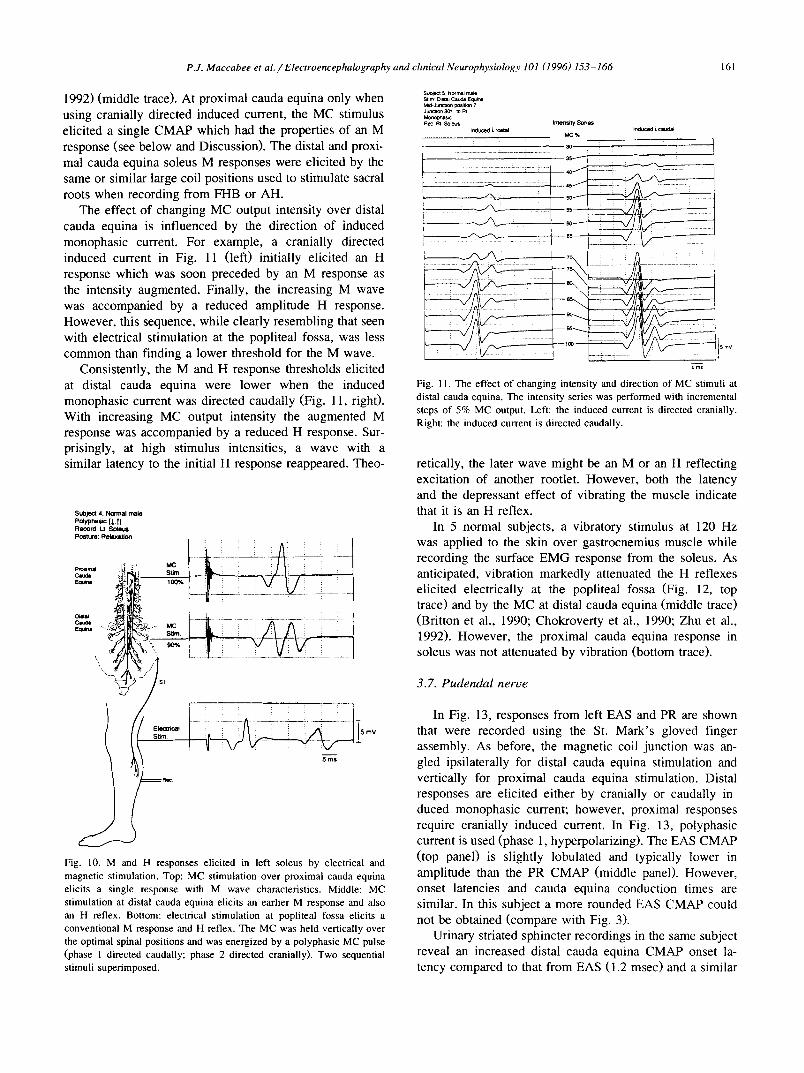
P J. Maccabee et al. / Electroencephalography and clinical Neurophysiology 101 (1996) 153-166 161
1992) (middle trace). At proximal cauda equina only when using cranially directed induced current, the MC stimulus elicited a single CMAP which bad the properties of an M response (see below and Discussion). The distal and proxi- mal cauda equina soleus M responses were elicited by the same or similar large coil positions used to stimulate sacral roots when recording from FHB or AH.
The effect of changing MC output intensity over distal cauda equina is influenced by the direction of induced monophasic current. For example, a cranially directed induced current in Fig. 11 (left) initially elicited an H response which was soon preceded by an M response as the intensity augmented. Finally, the increasing M wave was accompanied by a reduced amplitude H response. However, this sequence, while clearly resembling that seen with electrical stimulation at the popliteal fossa, was less common than finding a lower threshold for the M wave.
Consistently, the M and H response thresholds elicited at distal cauda equina were lower when the induced monophasic current was directed caudally (Fig. 11, right). With increasing MC output intensity the augmented M response was accompanied by a reduced H response. Sur- prisingly, at high stimulus intensities, a wave with a similar latency to the initial H response reappeared. Theo-
Subject 4, Normal male Paypna~e [~,t] Record: I t . So le t~ Post'~ce: Relaxat ion
Proxlrn~
Earn=
:: i i V i v i ::
5, Nor m~tl rna~ mira Dlstab C ~ Equ~na M ~ J u n t ~ positron 7 Junc~n 3o* ~ I
Rec ~ S~e~ Indoced t, rolltal
Irmm~a7 Series i n a ~ ~ c~da l
7 5 _ - ...... ...... :
5ms
Fig. 11. The effect of changing intensity and direction of MC stimuli at distal cauda equina. The intensity series was performed with incremental steps of 5% MC output. Left: the induced current is directed cranially. Right: the induced current is directed caudally.
retically, the later wave might be an M or an H reflecting excitation of another rootlet. However, both the latency and the depressant effect of vibrating the muscle indicate that it is an H reflex.
In 5 normal subjects, a vibratory stimulus at 120 Hz was applied to the skin over gastrocnemius muscle while recording the surface EMG response from the soleus. As anticipated, vibration markedly attenuated the H reflexes elicited electrically at the popliteal fossa (Fig. 12, top trace) and by the MC at distal cauda equina (middle trace) (Britton et al., 1990; Chokroverty et al., 1990; Zhu et al., 1992). However, the proximal cauda equina response in soleus was not attenuated by vibration (bottom trace).
3.7. Pudendal nerve
: : i : : i i i
- f f i --+-- ? -::- - - - - -
5 ms
5 mV
Fig. 10. M and H responses elicited in left soleus by electrical and magnetic stimulation. Top: MC stimulation over proximal cauda equina elicits a single response with M wave characteristics. Middle: MC stimulation at distal cauda equina elicits an earlier M response and also an H reflex. Bottom: electrical stimulation at popliteal fossa elicits a conventional M response and H reflex. The MC was held vertically over the optimal spinal positions and was energized by a polyphasic MC pulse (phase 1 directed caudally; phase 2 directed cranially). Two sequential stimuli superimposed.
In Fig. 13, responses from left EAS and PR are shown that were recorded using the St. Mark's gloved finger assembly. As before, the magnetic coil junction was an- gled ipsilaterally for distal cauda equina stimulation and vertically for proximal cauda equina stimulation. Distal responses are elicited either by cranially or caudally in- duced monophasic current; however, proximal responses require cranially induced current. In Fig. 13, polyphasic current is used (phase t, hyperpolarizing). The EAS CMAP (top panel) is slightly lobulated and typically lower in amplitude than the PR CMAP (middle panel). However, onset latencies and cauda equina conduction times are similar. In this subject a more rounded EAS CMAP could not be obtained (compare with Fig. 3).
Urinary striated sphincter recordings in the same subject reveal an increased distal cauda equina CMAP onset la- tency compared to that from EAS (1.2 msec) and a similar

162 P.J. Maccabee et al. / Electroencephalography and clinical Neurophysiology 101 (1996) 153-166
Sut)leCt 5, Normal male Record: Rt. Soleus Posture: Relaxation Vibration [120 Hz] to Soleus Sequential stimuli x4
Control Stim: Post. Tib. n, POD. Fossa Cathodal
Control
Stim: L Distal C E MC 80% Polyphasic [J,'T]
Stlm ProximaIMC 100%CE / Polyphasi¢ [,~, ~']
5 ms
5 mV
Fig. 12. Effect of vibrating soleus on responses to stimulation of distal and proximal cauda equina and at the popliteal fossa. After obtaining control responses with the vibrator plate on the skin overlying gastrocne- mius soleus, the vibrator was energized at 120 Hz for 10 sec and the responses were recorded. For each paradigm, 4 sequential stimuli were superimposed.
increase (1.1 msec) in cauda equina conduction time. Responses were maximally elicited with a vertically ori- ented junction over distal (Eardley et al., 1990) as well as proximal cauda equina.
3.8. Procedures in normal subjects
On the basis of the above experiments, in 5 normal subjects cauda equina conduction times were obtained recording from right vastus medialis, tibialis anterior, ab- ductor hallucis, and soleus (Table l). When recording from abductor hallucis and soleus, responses were elicited using the optimal MC orientations and positions generally de- rived from Figs. 4, 5a and 5b. When recording from vastus
Subject 6, Normal male PoJyphasic [l,t]
Distal Left I C E External Anal SphinCter Proximal
C E &=26ms
CE.
Urinary Striated Sphincter
!05mY
i i &=29ms ........ +
i i
3 mS
Fig. 13. CMAP responses from left external anal sphincter, left puborec- talis, and the urinary striated sphincter recorded in a normal subject by MC stimulation of proximal and distal cauda equina. Four sequential stimuli were superimposed.
medialis and tibialis anterior, responses were elicited using optimal MC orientations and positions illustrated in Fig. 7. Although similar, the optimal MC positions and orienta- tions differed slightly in each subject. To maximize CMAP amplitudes, motor responses were elicited at proximal and distal cauda equina by a polyphasic MC pulse with a hyperpolarizing-depolarizing sequence (Fig. 6, bottom; and Fig. 1E in Maccabee et al., 1993a). Compared to the monophasic pulse, the hyperpolarizing-depolarizing polyphasic pulse yields a slightly longer response latency (by approximately 0.125-0.150 msec), presumably reflect- ing the duration of the hyperpolarizing phase. Onset laten- cies to vastus medialis, tibialis anterior and abductor hallu-
Table 1 Cauda equina conduction times of motor fibers innervating the indicated right lower limb muscles of 5 normal males
Subject Age Height (years) (cm)
M proximal CE to M distal CE H distal CE to M proximal CE M proximal CE to M distal CE
Vastus medialis Tibialis anterior Abductor hallucis Soleus Soleus (msec) (msec) (msec) (msec) (msec)
1 40 178 1.8 1.8 4.0 3.7 3.3 2 26 180 1.9 2.2 3.1 3.7 2.4 3 32 183 2.0 2.4 4.0 3.6 3.8 4 33 168 1.9 2.5 3.0 3.1 3.0 5 28 165 1.8 2.7 3.3 3.0 3.1
Mean 32 175 1.9 2.3 3.5 3.4 3.1
In vastus medialis, tibialis anterior and abductor hallucis, cauda conduction times were calculated from M response onset latency. In soleus, conduction times were calculated from the negative peaks of M responses and H reflexes (see text). Latency in msec. Abbreviation: CE = cauda equina.

P.J. Maccabee et a l . / Electroencephalography and clinical Neurophysiology 101 (1996) 153-166 163
cis were measured to the first inflection point from base- line, visualized from the superposition of responses ob- tained at maximal and submaximal output intensities (as in Fig. 8, CMAP onset latencies in all subjects remained stable despite widespread variation in amplitude). How- ever, when recording from soleus, maximal M and H responses were measured from their respective negative peak latencies. This is because the onset of the H reflex elicited at distal cauda equina is obscured by the tail of the preceding M response (Zhu et al., 1992; and Fig. 10, middle traces).
4. Discussion
4.1. The utility of the lumbosacral model in predicting human responses
The conclusions derived from the physical model and from earlier in vitro experiments (Amassian et al., 1992; Maccabee et al., 1993a) initially guided the method of stimulation in humans and the predictions were substan- tially confirmed. The predictions and confirmatory data include:
(1) Given that the L4-5 and L5-S1 neuroforamina are anatomically oriented medio-laterally the induced current should be channeled with the same orientation. In the model, maximal fields were elicited when the MC junction was roughly horizontal and was placed over the L4-5 and L5-S1 neuroforamina (Fig. 2). Similarly, in the normal human (Fig. 7), distal excitation of the L4 and L5 segmen- tal roots to tibialis anterior occurred at lowest threshold when the MC junction was horizontal and located approxi- mately midway between the top edge of the sacrum and the L4-L5 interspace at the level of the superior iliac crests. The possibility that proximal excitation occurred and was masked by collision with distal excitation can be excluded by the intensity series in Fig. 8. Frequently, the elicited lumbar root response to either tibialis anterior or vastus medialis was optimal when the junction was tilted above the ipsilateral horizontal. This latter result is similar to previous studies of cervical root stimulation (Epstein et al., 1991; Mills et al., 1993). Mills et al. (1993) provided anatomical evidence that cervical neuroforamina are also angled above the ipsilateral horizontal. Thus, it is inferred that the MC junction is optimally positioned when parallel to and directly overlaying a cervical neuroforamen. Possi- bly, the caudal bend of the cervical roots at their exit from the neuroforamina represents an additional low threshold site. A similar mechanism may possibly operate when stimulating lumbar roots.
(2) Large electric fields were also recorded within the longitudinal central sacral axis when the MC junction was either vertical or 45 ° from the vertical. In part, this reflects the relatively greater length of the vertical sacral axis
compared to the horizontal sacral axis (Fig. 1). Also, as anticipated, much smaller fields were recorded across ven- tral sacral neuroforamina because their antero-posterior orientation severely limits the channeling of induced cur- rent there (Figs. 1 and 2). In the human, sacral roots were optimally excited by a vertical or near vertical MC junc- tion (Fig. 5a). It also appears likely that the distal cauda equina site of excitation for lumbar and sacral roots is identical to that obtained with a round coil by others (Chokroverty et al., 1990; MacDonell et al., 1992). Experi- ments comparing round coil and twin coil excitation in the same subject reveal virtually identical CMAP onset laten- cies (unpublished observations). It is not obvious from either the model or the known anatomy why, over distal cauda equina, the most effective direction of induced monophasic current is always inward for lumbar roots and usually rostral for sacral roots. Although the more effective direction of induced current is towards the spinal fluid filled thecal sac and away from the more distal root segments covered by epidural fat, this should not be a critical factor; the magnitude of the induced current should be the same for both directions of current.
(3) A vertically oriented MC junction positioned over the proximal cauda equina should excite the region of the rootlet exit zone because the induced current flowing in the axis of the middle or lower cauda equina is rostrally bounded by the higher resistance conus medullaris with displacement of current to the sides of the conus. In addition, the acute caudal angle of the nerve rootlets with respect to the conus tip positions them ideally for excita- tion by cranially directed current which passes near or at their interface with the conus. In the normal human, proximal cauda equina responses are elicited exclusively by cranially directed, induced monophasic current. Fur- thermore, if at the entry zone the rootlets were angled cranially (as occurs with tethered cord syndrome) instead of caudally, the threshold for excitation could be substan- tially increased. In one patient with tethered cord syn- drome, distal, but not proximal, responses were elicited (unpublished observation).
(4) When exciting a straight nerve in a homogeneous media volume conductor, the stimulating function has been proven to be the negative-going spatial derivative of the induced electric field (Maccabee et al., 1993a). In those experiments, moving the small twin MC junction along the nerve also moved the derivative and the site of excitation but this was always limited by the relatively short length of the nerve. It would therefore be anticipated that moving the large twin coils along the spine might also excite different portions of the mainly straight cauda equina rootlets. However, responses with intermediate or incre- menting latency between proximal and distal low threshold sites were never observed. This supports the conclusion that excitation occurs at sites other than those along straight nerve fibers, such as at bends at the rootlet exit zone and at the intervertebral neuroforamina.

164 P.J. Maccabee et al . / Electroencephalography and clinical Neurophysiology 101 (1996) 153-166
4.2. Direct excitation of motor fibers in eauda equina
The motor responses elicited by MC excitation over proximal and distal cauda equina most likely reflect direct excitation of peripheral motor fibers. The available evi- dence indicating direct excitation of peripheral motor fibers includes:
(1) The responses elicited over proximal cauda equina are similar in appearance to those elicited distally, but with longer onset latencies, typically 1.9 msec for vastus medi- alis, 2.3 msec for tibialis anterior, and 3.5 msec for abductor hallucis (Table 1). For a conduction velocity of 55 m / s e c the above latency differences correspond to conduction distances of 10.5 cm, 12.7 cm, and 19.3 cm. These calculated distances are roughly similar to the lengths of the L3, L5 and S1 intrathecal segments measured in cadavers (Sunderland, 1976). However, an accurate com- parison cannot be made in the absence of data on the heights of Sunderland's subjects which could presumably vary with the lengths of the cauda equina.
(2) Vibrating a muscle has previously been shown to markedly reduce the amplitude of the H reflex. However, vibration of the vastus medialis, tibialis anterior and foot muscles did not significantly reduce the CMAPs described above (but did markedly reduce a later CMAP, see below).
(3) Comparison of the latencies elicited over proximal and distal cauda equina (measured at high gain) with the calculated one-way F response latencies (measured at the same gain) revealed that the one-way F response latencies exceeded those of the proximal response.
(4) Prior in vitro experiments using relatively simple homogeneous volume conductors reveal that low threshold sites are maximally activated at the peak of the induced electric field under the center of the MC junction (Mac- cabee et al., 1993a). In the subject whose responses are illustrated in Figs. 4 and 7, the peak electric field along the 6 cm MC junction is located 3 cm proximal to the distal divergence. For the flexor hallucis brevis (Fig. 4), the MC junction displacement from optimal caudal to cranial posi- tions is approximately 13 cm (i.e., central vertical junction positions 5-18). For tibialis anterior (Fig. 7) the corre- sponding MC junction displacement is approximately 9.5 cm (interpolated horizontal junction position 10.5 to inter- polated vertical junction position 20). These MC displace- ment distances are roughly supported by measurements taken from an MRI scan in the same subject: 14 cm from conus tip to the right S1 ventral foramen and 12 cm from conus tip to the left L5-S1 neuroforamen (anatomically another 2 cm could be added to the S1 and 3 cm added to the L5 measurements (Haymaker and Woodhall, 1953) owing to the more caudal to cranial locations of the SI and L5 rootlet exit zones; this yields 16 cm for S1 and 15 cm for L5). Moreover, proximal to distal latency shifts are 2.8 msec for the right AHB and 2.4 msec for the left tibialis anterior (corresponding, at approximately 55 m/sec , to 15.4 and 13.2 cm, respectively).
For the distal response to abductor hallucis (Fig. 4), the optimal placement of the central junction at position 5 overlays upper sacrum. Theoretically, excitation of sacral rootlets could take place either at the abrupt change in conductivity where the rootlets emerge from the sacral sac (usually at the level of the $2 foramina) or where the roughly vertical rootlets within the central sacrum bend anteriorly to enter the ventral sacral foramina. The distance between these two postulated low threshold sites is not more than a few centimeters. For the distal cauda equina response recorded from tibialis anterior, excitation of the L5 nerve root appears to occur at the level of the L5-S 1 neuroforamen (Fig. 7).
Occasionally, using the twin coil, sacral root CMAP onset latencies to abductor hallucis decreased when the induced current was directed caudally. This may possibly reflect excitation jumping between the sacral sac and near foraminal site or to an intermediate site such as the spinal fluid surrounding the rootlets for short distances beyond their emergence from the thecal sac.
(5) Patients with S I radiculopathy and/or severe sen- sory neuropathy have shown absent popliteal and distal cauda equina H reflexes but preserved proximal cauda equina responses from soleus and from abductor hallucis, tibialis anterior and vastus medialis (Maccabee et al., 1995). This indicates that the proximal cauda equina re- sponses do not reflect indirect excitation of sensory affer- ent pathways.
Proof of the exact sites at which distal and proximal excitation of cauda equina motor fiber occurs awaits mea- surement of CMAP response latencies before and during surgical exposure.
4.3. Indirect (transsynaptic) activation of motor fibers in cauda equina
With distal stimulation of the cauda equina and record- ing from soleus, a later wave (Fig. 10, middle trace) is evident following the M response. This later wave differs fundamentally from the wave elicited by proximal stimula- tion in being markedly reduced by vibrating the muscle (Fig. 12, middle trace). In agreement with others (Britton et al., 1990; Chokroverty et al., 1990; Zhu et al., 1992), the later wave depends upon activation of presynaptic fibers with subsequent transsynaptic activation of motor neurons, i.e., it is physiologically equivalent to the H response elicited at the popliteal fossa (Fig. 12, top trace). An additional possibility exists that the distal cauda equina H reflex is added to by excitation in spinal cord elements including the corticospinal tract. This seems unlikely when considering that the H reflex is elicited only when the stimulating MC is held over the sacrum. Furthermore, in a patient with sensory greater than motor neuropathy, the distal M response was preserved but the H reflex was absent when elicited at distal cauda equina and the popliteal fossa (Maccabee et al., 1995).

P.J. Maccabee et al. / Electroencephalography and clini(~d Neurophysiology 101 (1996) 153-166 165
The soleus motor response elicited by proximal cauda equina stimulation (Fig. 12, bottom trace) is not signifi- cantly reduced by vibration and appears to be the same response elicited in abductor hallucis, tibialis anterior, and vastus medialis; however, it might also include compo- nents derived from activation of presynaptic fibers. No- tably, in the patient referred to above, the motor responses to proximal cauda equina, distal cauda equina and popliteal fossa were preserved despite the loss of H reflexes. There- fore, we conclude that the proximal cauda equina response in soleus is a direct M and not a transsynaptic reflex.
4.4. Direct activation of motor fiber to striated sphincters
are prolonged to as much as 10-12 msec (compare with Table 1) and are roughly equivalent to conduction veloci- ties of 15 m / s e c or less. In the future, it is anticipated that this method will find widespread application in the detec- tion and classification of peripheral neuropathies affecting lower limbs and striated sphincters.
Acknowledgement
The authors wish to thank Ms. Helen R. Watson for her secretarial assistance.
Swash and Snooks (1992) have recently emphasized that PR is innervated by a nerve branch separate from, but with length similar to that innervating EAS. In addition, the nerve to PR bends slightly ventral from the inner sacral surface. Possibly, excitation could take place at this bend analogous to experiments with mammalian peripheral nerve in vitro (Maccabee et al., 1993a). However, in the normal subject depicted in Fig. 13, terminal latencies from distal cauda equina to PR and EAS are similar, which might suggest a common site of excitation. The longer terminal latency to USS is consistent with the longer length of its innervating nerve (Snooks and Swash, 1984; Swash and Snooks, 1992). The longer cauda equina conduction time in USS compared to EAS may further suggest that either slower conducting fibers innervate USS or that the sites of low threshold MC excitation in cauda equina are not identical.
4.5. Clinical aspects
Previously, direct detection of conduction abnormalities within the cauda equina required cortical stimulation and calculation of conventional central motor conduction time (CMCT), a distance that extends from motor cortex to the site of peripheral stimulation over distal cauda equina. To the lower limbs, CMCT reflects the sum of conduction times from motor cortex to anterior horn, and from anterior horn to distal cauda equina. Thus, any increase in conduc- tion time in the cauda equina is added to a much longer conduction time from motor cortex to the rootlet exit zone. By contrast, our method can directly measure motor (L4, L5, S1-$4) and sensory (S1) conduction in a cauda equina segment, which may be entirely intrathecal. This is the same locus that is often most severely affected in acquired and experimental demyelinating polyneuropathies (Van der Meche and Van Doom, 1995). Also, CMCT to lower limbs may now be calculated much more accurately, from motor cortex to rootlet exit zone, excluding the cauda segment.
Preliminary studies in some patients with demyelinating neuropathy reveal markedly slowed conduction times re- stricted to or predominantly within the cauda equina (Mac- cabee et al., 1995). These cauda segment conduction times
References
Amassian, V.E., Eberle, L.P., Maccabee, P.J. and Cracco, R.Q. Mod- elling magnetic coil excitation in human cerebral cortex with a peripheral nerve immersed in a brain-shaped volume conductor: the significance of fiber bending in excitation. Electroenceph. clin. Neu- rophysiol., 1992, 85: 291-301.
Barker, A.T. An introduction to the basic principles of magnetic nerve stimulation. J. Clin. Neurophysiol., 1991, 8: 26-37.
Britton, T.C., Meyer, B.U., Herdman, J. and Benecke, R. Clinical use of the magnetic stimulator in the investigation of peripheral conduction time. Muscle Nerve, 1990, 13: 396-406.
Cadwell, J. Principles of magnetoelectric stimulation. In: S. Chokroverty (Ed.), Magnetic Stimulation in Clinical Neurophysiology. Butter- worth, Boston, MA, 1990: 13-32.
Chokroverty, S., Spire, J.P., DiLullo, J., Moody, E. and Maselli, R. Magnetic stimulation of the human peripheral nervous system. In: S. Chokroverty (Ed.), Magnetic Stimulation in Clinical Neurophysiol- ogy. Butterworth, Boston, MA, 1990: 249-273.
Cohen, D. and Cuffin, B.N. Developing a more focal magnetic simulator. 1. Some basic principles. J. Clin. Neurophysiol., 1991, 8: 102-111.
Durand, D., Ferguson, A.S. and Dalbasti, T. Induced electric fields by magnetic stimulation in non-homogeneous conducting media. IEEE Eng. Med. Biol., 1989, 5: 1251-1253.
Eardley, I., Nagendran, K., Kirby, R.S. and Fowler, C.J. A new technique for assessing the efferent innervation of the human striated urethral sphincter. J. Urol., 1990, 144: 948-951.
Epstein, C.M., Fernandez-Beer, E., Weissman, J.D. and Matsuura, S. Cervical magnetic stimulation: the role of the neural foramen. Neurol- ogy, 1991, 41: 677-680.
Haymaker, W. and Woodhall, B. Peripheral Nerve Injuries. Saunders, Philadelphia, PA, 1953:32 (Fig. 25).
Maccabee, P.J., Eberle, L.P., Amassian, V.E., Cracco, R.Q., Rudell, A.P. and Jayachandra, M. Spatial distribution of the electrical field induced in volume by round and figure "8' magnetic coils: relevance to activation of sensory nerve fibers. Electroenceph. clin. Neurophysiol., 1990, 76: 131-141.
Maccabee, P.J., Amassian, V.E., Eberle, L.P., Rudell, A.P., Cracco, R.Q., Lai, K.S. and Somasundaram, M. Measurement of the electric field induced into inhomogeneous volume conductors by magnetic coils. Application to human spinal neurogeometry. Electroenceph. clin. Neurophysiol., 1991, 81: 224-237.
Maccabee, P.J., Amassian, V.E., Eberle, L.P. and Cracco, R.Q. Magnetic coil stimulation of straight and bent amphibian and mammalian peripheral nerve in vitro: locus of excitation. J. Physiol. (Lond.), 1993a, 460: 201-219.
Maccabee, P.J., Eberle, L.P., Amassian, V.E., Anselmi, G.D., Tatarian, G.T., Lipitz, M.E., Rosen, T. and Cracco, R.Q. Optimal polarity sequence for exciting peripheral nerve and spinal roots. Neurology, 1993b, 43 (Suppl. 2): A288 (abstract).

166 P.J. Maccabee et aL / Electroencephalography and clinical Neurophysiology 101 (1996) 153-166
Maccabee, P.J., Lipitz, M.E., Golub, R.W., Eberle, L.P., Cracco, R.Q., Amassian, V.E., Desudchit, T.P. and Bania, J.P. Mechanism and method of selective lumbosacral nerve root stimulation using the magnetic coil. Neurology, 1994a, 44 (Suppl. 2): A349 (abstract).
Maccabee, P.J., Lipitz, M.E., Desudchit, T., Golub, R.W., Amassian, V.E., Bania, J.P., Eberle, L.P., Nitti, V.W. and Cracco, R.Q. Motor evoked responses elicited over proximal and distal cauda equina by a large twin magnetic coil. Soc. Neurosci. Abst., 1994b, 20: 787.
Maccabee, P.J., Lipitz, M.E., Desudchit, T., Golub, R.W., Amassian, V.E., Nitti, V.W., Cracco, R.Q., Hotson, G.L., Eberle, L.P., Jalal, S., Wilier, J.A. and Bania, J.P. Detection of proximal demyelinating neuropathy in cauda equina by neuromagnetic stimulation. Neurology, 1995, 45 (Suppl. 4): A170 (abstract).
MacDonell, R.A.L., Cros, D. and Shahani, B.T. Lumbosacral nerve root stimulation comparing electrical with surface magnetic coil tech- niques. Muscle Nerve, 1992, 15: 885-890.
Maertens de Noordhout, A., Rothwell, J.C., Thompson, P.D., Day, B.L. and Marsden, C.D. Percutaneous electric stimulation of lumbosacral roots in man. J. Neurol. Neurosurg. Psychiat., 1988, 51: 174-181.
Mills, K.R., McLeod, C., Sheffy, J. and Loh, L. The optimal current direction for excitation of human cervical motor roots with a double coil magnetic stimulator. Electroenceph. clin. Neurophysiol., 1993, 89: 138-144.
Reilly, J.P. Peripheral nerve stimulation by induced electric currents: exposure to time-varying magnetic fields. Med. Biol. Eng. Comput., 1989, 27: 101-110,
Roth, B.J. and Basser, P. Model of the stimulation of a nerve fiber by electromagnetic induction. IEEE Trans. Biomed. Eng., 1990, 37: 588-597.
Snooks, S.J. and Swash, M. Peroneal nerve and transcutaneous spinal stimulation: new methods for investigation of the urethral striated sphincter musculature. Br. J. Urol., 1984, 56: 406-409.
Sunderland, S. Avulsion of nerve roots. In: P.J. Vinken and G.W. Bruyn (Eds.), Handbook of Clinical Neurology, Vol. 11. Elsevier, Amster- dam, 1976: 393-435.
Swash, M. and Snooks, S.J. Slowed motor conduction in lumbosacral nerve roots in cauda equina lesions: a new diagnostic technique. J. Neurol. Neurosurg. Psychiat., 1986, 49: 808-816.
Swash, M. and Snooks, S.J. Motor nerve conduction studies of the pelvic floor innervation. In: K.M. Henry and M. Swash (Eds.), Colonproctol- ogy and the Pelvic Floor. Butterworth-Heinemarm, Boston, MA, 1992: 196-206.
Ugawa, Y., Rothwell, J.C., Day, B.L., Thompson, P.D. and Marsden, C.D. Magnetic stimulation over the spinal enlargements. J. Neurol. Neurosurg. Psychiat., 1989, 52: 1025-1032.
Van der Meche, F.G.A. and Van Doom, P.A. Guillain-Barr6 syndrome and chronic inflammatory demyelinating polyneuropathy: immune mechanisms and update on current therapies. Ann. Neurol., 1995, 37 (Suppl. 1): 514-531.
Zhu, Y., Starr, A., Su, S.H., Woodward, G. and Haldeman, S. The H-reflex to magnetic stimulation of lower-limb nerves. Arch. Neurol., 1992, 49: 66-71.




















