
Indigenous Vibrio cholerae strains from a non-endemic region are pathogenic

1999, 181(24):7588. J. Bacteriol.
Shelley M. PayneElizabeth E. Wyckoff, Ana-Maria Valle, Stacey L. Smith and Transports Vibriobactin and Enterobactin
Vibrio choleraeTransporter System from A Multifunctional ATP-Binding Cassette
http://jb.asm.org/content/181/24/7588Updated information and services can be found at:
These include:
REFERENCEShttp://jb.asm.org/content/181/24/7588#ref-list-1at:
This article cites 52 articles, 31 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
on N
ovember 11, 2014 by guest
http://jb.asm.org/
Dow
nloaded from

JOURNAL OF BACTERIOLOGY,0021-9193/99/$04.0010
Dec. 1999, p. 7588–7596 Vol. 181, No. 24
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
A Multifunctional ATP-Binding Cassette Transporter Systemfrom Vibrio cholerae Transports Vibriobactin and Enterobactin
ELIZABETH E. WYCKOFF,* ANA-MARIA VALLE, STACEY L. SMITH, AND SHELLEY M. PAYNE
Department of Molecular Genetics and Microbiology, and Institute for Cellular and Molecular Biology,University of Texas, Austin, Texas 78712-1095
Received 14 July 1999/Accepted 5 October 1999
Vibrio cholerae uses the catechol siderophore vibriobactin for iron transport under iron-limiting conditions.We have identified genes for vibriobactin transport and mapped them within the vibriobactin biosynthetic genecluster. Within this genetic region we have identified four genes, viuP, viuD, viuG and viuC, whose proteinproducts have homology to the periplasmic binding protein, the two integral cytoplasmic membrane proteins,and the ATPase component, respectively, of other iron transport systems. The amino-terminal region of ViuPhas homology to a lipoprotein signal sequence, and ViuP could be labeled with [3H]palmitic acid. This suggeststhat ViuP is a membrane lipoprotein. The ViuPDGC system transports both vibriobactin and enterobactin inEscherichia coli. In the same assay, the E. coli enterobactin transport system, FepBDGC, allowed the utilizationof enterobactin but not vibriobactin. Although the entire viuPDGC system could complement mutations in fepB,fepD, fepG, or fepC, only viuC was able to independently complement the corresponding fep mutation. Thisindicates that these proteins usually function as a complex. V. cholerae strains carrying a mutation in viuP orin viuG were constructed by marker exchange. These mutations reduced, but did not completely eliminate,vibriobactin utilization. This suggests that V. cholerae contains genes in addition to viuPDGC that function inthe transport of catechol siderophores.
Pathogenic bacteria require iron for growth and survival. Inmost environments, however, iron is not readily available. Inthe mammalian host, the vast majority of iron is present inintracellular iron proteins, such as hemoproteins and ferritin.Most of the extracellular iron is complexed with proteins, suchas transferrin, lactoferrin, hemopexin, or haptoglobin. Thesupply of free iron also is limited in many environments out-side the human host. To obtain iron, bacteria have evolvedhigh-affinity iron acquisition systems, and many bacteria haveseveral of these systems (6). Some of these iron acquisitionsystems support direct utilization of iron from specific host ironcompounds. Others involve the production and transport ofsiderophores, which are small molecules that bind iron withhigh affinity and are then transported back into the cell. Theexpression of iron acquisition genes is repressed by iron, usu-ally by the negative regulatory transcription factor Fur (4).
The gram-negative pathogen Vibrio cholerae, the causativeagent of cholera (13, 25), has multiple iron transport systems.V. cholerae transports free heme and hemoglobin-associatedheme through the action of the HutABCD system (21, 22, 31).V. cholerae strains also transport siderophores, including thehydroxamate siderophore ferrichrome (17), although the pro-duction of hydroxamate siderophores has not been observed inV. cholerae (42). The siderophore produced and used by mostV. cholerae strains is the catechol vibriobactin (17) (Fig. 1).Vibriobactin contains three 2,3-dihydroxybenzoyl residueslinked to a backbone of norspermidine, an abundant poly-amine in most members of the Vibrionaceae (55, 56). Two ofthese dihydroxybenzoyl residues are joined to the backbone viaL-threonine, whereas the third is linked directly to the norsper-midine moiety (17) (Fig. 1). Dihydroxybenzoate is synthesizedfrom chorismate by VibABC (17, 53). The late steps in vibri-
obactin biosynthesis, in which dihydroxybenzoate, threonine,and norspermidine are joined to each other, are not well un-derstood, but at least four proteins, VibD, VibE, VibF, andVibH, are required for this process.
Less is known about the transport of vibriobactin. Becausevibriobactin is structurally similar to enterobactin, the catecholsiderophore produced by Escherichia coli and related species(Fig. 1), it appeared likely that the transport systems would besimilar. The E. coli Fep system, which transports enterobactin,is typical of high-affinity iron transport systems found in gram-negative bacteria. These consist of several components. Theouter membrane receptors, such as FepA, bind their ligandwith high affinity (6, 7, 11). Transport of the ligand through theouter membrane requires the activity of TonB, ExbB, andExbD, which are thought to transduce the energy required fortransport (30). Transport of the siderophore through theperiplasm and across the inner membrane requires a periplas-mic binding protein-dependent ATP-binding cassette (ABC)transport system (5, 6). In this system, the siderophore specif-ically binds its periplasmic binding protein, e.g., FepB, whichthen delivers the ligand to the corresponding inner membranepermease complex. The permease usually consists of two inte-gral membrane proteins, which form a tight complex with eachother within the cytoplasmic membrane. Each of the integralmembrane proteins is bound to an additional protein that hasATPase activity. The hydrolysis of ATP by the ATPase subunitis thought to generate the energy for transport of the ligandacross the inner membrane. In E. coli, FepD and FepG formthe integral membrane permease complex (10, 41), and FepCis the ATPase (41). Following transport of enterobactin, acytoplasmic protein, Fes, catalyzes the removal of iron fromthe ferri-siderophore complex (11).
In V. cholerae, the vibriobactin receptor, ViuA, has beenidentified and characterized (9, 45). Like enterobactin, vibri-obactin transport is dependent upon a functional TonB system(22, 31). Another vibriobactin utilization protein, ViuB, isanalogous to Fes and removes the iron from the iron-vibri-
* Corresponding author. Mailing address: Department of MolecularGenetics and Microbiology, University of Texas, Austin, TX 78712-1095. Phone: (512) 471-5204. Fax: (512) 471-7088. E-mail: [email protected].
7588
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

obactin complex in the cytoplasm (8). Prior to this work, how-ever, there was no information available about the transport ofvibriobactin through the periplasm and across the cytoplasmicmembrane. In this report we describe viu genes located withinthe vibriobactin biosynthetic gene cluster which encode anABC transporter system. The system encoded by these genes isdifferent from Fep in that it can transport both vibriobactinand enterobactin. Further, we show that the Viu and Feppermeases each function as a complex, and, with the exceptionof the ATPase homologues, the individual Viu and Fep pro-teins are not interchangeable. We also present evidence that V.cholerae has an additional system for the transport of catecholsiderophores.
MATERIALS AND METHODS
Bacterial strains, plasmids, and media. Bacterial strains and plasmids used inthis study are listed in Table 1. The iron chelator ethylenediamine di(ortho-hydroxyphenylacetic acid) (EDDA) was deferrated by the method of Rogers(36). When added, the antibiotic concentrations used were 250 mg of carbeni-cillin per ml, 50 mg of kanamycin per ml, and 50 mg of chloramphenicol per ml(for E. coli) or 5 mg of chloramphenicol per ml (for V. cholerae).
DNA sequencing. DNA was sequenced by using an Applied Biosystems Prism377 DNA sequencer (Perkin-Elmer Corp.). Routine DNA sequence analysis wasperformed by using the program DNA Strider (27). Amino acid sequence align-ments were performed by using the gap program of the Genetics ComputerGroup DNA sequence analysis package. The BLAST program (2) was used tosearch the National Center for Biotechnology Information protein database.
Detection of siderophore production and utilization. The colorimetric test forthe production of catechols was performed by the method of Arnow (3). Thebioassay for siderophore production and utilization was performed as previouslydescribed (53).
Construction of chromosomal mutations in V. cholerae. To construct a muta-tion in viuP, a SmaI fragment containing the chloramphenicol cassette frompMTLcam (54) was inserted into the MscI site in viuP in the plasmid pVIB119to give the plasmid pVIB148. The PstI fragment containing the disrupted viuPgene was then subcloned into the PstI site of pWSc1 (31), a plasmid carrying thesacB gene, which confers sensitivity to sucrose (15). This plasmid was introducedinto the V. cholerae wild-type strain Lou15 by electroporation, and the markerexchange mutant EWV103 was obtained by selecting colonies resistant to bothsucrose and chloramphenicol as previously described (53). To construct SSV121,containing a mutation in viuG, the chloramphenicol gene from pMA9 was in-serted into the StuI site within viuG in the plasmid pVIB121. The SalI-XbaIfragment of the resulting plasmid was cloned into SalI-XbaI-digested pHM5, togive pUNK143. Following transfer of pUNK143 by conjugation into Lou15,sucrose-resistant, chloramphenicol-resistant colonies were selected. The pres-ence of the cam cassette within the chromosomal viuP gene in strain EWV103and within viuG in SSV121 was confirmed by Southern hybridization (data notshown).
Construction of pViuAB. pViuAB was constructed by PCR amplification of theviuA and viuB genes from V. cholerae O395. The PCR was performed as follows:1 ml of an overnight culture of O395 was pelleted by centrifugation and wasresuspended in 100 ml of sterile water. One microliter of this cell suspension wasmixed with 100 pmol of each deoxynucleoside triphosphate, 50 pmol of eachprimer, Qiagen PCR buffer (13 final concentration), 1.25 U of Taq polymerase
(Qiagen), and 1.25 U of Pfu polymerase (Stratagene) in a total volume of 100 ml.The reaction was 95°C for 5 min, followed by 30 cycles of 94°C for 30 s, 45°C for60 s, and 72°C for 20 min in a GeneAmp PCR system 2400 (Perkin-Elmer). Theprimer sequences were 59-AAGCTTTGTAGGAAGGGAA and 59-TCTAGATAAGCAATGTGCTCATAAA. The 3.6-kbp product was made blunt with Kle-now and ligated in the SmaI site of pWSK29 (50).
Labeling of lipoproteins with [3H]palmitic acid. Overnight cultures werediluted into 1 ml of L broth containing 20 mCi of [3H]palmitic acid per ml and50 mg of carbenicillin per ml. Cultures were grown for 8 h with shaking at 37°C.Cells were harvested by centrifugation, resuspended in 50 ml of sodium dodecylsulfate (SDS) gel sample buffer and lysed by boiling for 15 min. Proteins wereseparated by SDS-polyacrylamide gel electrophoresis (PAGE). The gel wastreated with EN3HANCE (NEN Life Science Products, Inc., Boston, Mass.), and[3H]-labeled proteins were visualized by fluorography.
RESULTS AND DISCUSSION
Identification of genes encoding potential iron transportproteins. As a continuation of our studies of V. cholerae irontransport systems, we characterized genes for the transport ofvibriobactin across the cytoplasmic membrane. Because sid-erophore synthesis and transport genes are often clustered, wetested previously isolated cosmid clones containing vibriobac-tin biosynthetic genes (53) for the presence of iron transportfunctions. Preliminary experiments showed that one of thecosmid clones complemented a mutation in the E. coli enter-obactin transport gene, fepC (data not shown). Further sub-cloning and complementation experiments indicated that theV. cholerae iron transport protein genes reside in a 4,100-bpregion which is within the vibriobactin synthetic gene cluster(Fig. 2). The nucleotide sequence of this region was obtainedon both strands (GenBank accession no. U52150). This regioncontained four open reading frames (ORFs), which have beennamed viuPDGC. They are flanked by the vibriobactin biosyn-thetic genes vibH and vibD (Fig. 2 and unpublished data).
The first ORF, viuP, encodes a protein with a predictedmolecular weight of 36,100 and predicted pI of 5.97. It hasamino acid sequence homology with E. coli FepB (Table 2),the periplasmic binding protein for the transport of enterobac-tin (12). The amino-terminal region of ViuP is rich in hydro-phobic amino acids but lacks a good match to the standardsignal peptidase cleavage site (34) (Fig. 3A). The ViuP amino-terminal sequence is a better match to the consensus sequencefor the prolipoprotein signal peptidase (Fig. 3D) (52), suggest-ing that ViuP may be a lipoprotein. In gram-positive bacteria,the binding protein components of membrane transport sys-tems are membrane lipoproteins (46). In gram-negative bac-teria, a few of the periplasmic binding protein homologues alsoappear to be membrane lipoproteins. Examples include the
FIG. 1. Structure of the catechol siderophores enterobactin and vibriobactin (17).
VOL. 181, 1999 CATECHOL TRANSPORT IN V. CHOLERAE 7589
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

FatB protein for anguibactin transport in Vibrio anguillarum(1) and the CeuE protein for enterobactin transport in Campy-lobacter jejuni and Campylobacter coli (32, 35). In bacteriallipoproteins, the lipid is covalently attached to a cysteine res-idue present within a consensus sequence located near theamino-terminal end of the protein (34, 46, 52). The cysteineresidue in the amino-terminal region of ViuP aligns with thecysteines in FatB and in CeuE that are believed to be themodified residues (Fig. 3D).
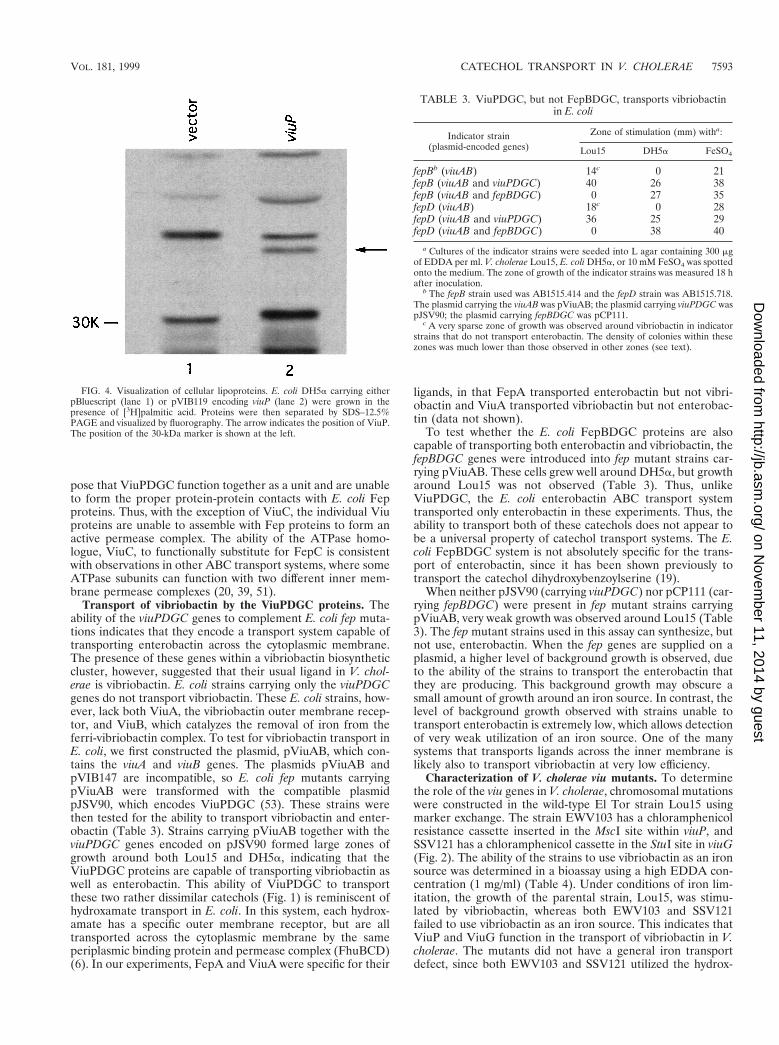
To determine whether ViuP is a membrane lipoprotein, E.coli carrying either pBluescript encoding viuP or the pBlue-script vector as a control were labeled with [3H]palmitic acid.SDS-PAGE of tritium-labeled proteins showed the presence ofa labeled protein with a molecular weight of approximately35,000, the predicted size of ViuP. This labeled protein waspresent in cells carrying viuP on a plasmid, but not in cellscarrying the pBluescript vector (Fig. 4), suggesting that ViuP isa lipoprotein. Interestingly, a cysteine in HutB, the putativeperiplasmic binding protein for heme transport in V. cholerae,also can be aligned with the cysteine in FatB and ViuP asshown by a ClustalW alignment of the amino-terminal regionsof these three proteins (Fig. 3D). These data raise the possi-bility that attachment of the binding protein component of irontransport systems to the inner membrane may be a commonfeature in Vibrio spp.
The second ORF, viuD, encodes a protein with homology toFepD and other integral membrane proteins of cytoplasmic
membrane permeases (Table 2). viuD has two possible trans-lation starts, one overlapping the viuP stop codon and a sec-ond, in-frame ATG four codons downstream of the first one.The tight linkage of viuP and viuD suggests that they arecotranscribed. The calculated molecular weight for the largerpossible ViuD protein is 37,033, and the calculated pI is 9.40.Highly basic pIs are typical of cytoplasmic membrane per-mease proteins. ViuD is extremely hydrophobic, as would beexpected for an integral membrane protein. Staudenmaier etal. (44) found that the inner membrane permease proteins ofiron transport systems share a high level of sequence conser-vation throughout their entire length and also identified sevenregions of these proteins with especially high amino acid se-quence conservation. Each of these seven sequence motifs ispresent within the ViuD sequence (data not shown).
The third ORF, viuG, encodes a protein with homology toFepG and other cytoplasmic membrane permease proteins(Table 2). The translational start site of viuG has not beenidentified, but we believe that the most likely start is an ATG20 nucleotides upstream of the viuD stop codon. The predictedprotein has a molecular weight of 37,470 and a pI of 11.36.Like ViuD, ViuG has the amino acid sequence features char-acteristic of cytoplasmic membrane permease proteins. As hasbeen reported for other inner membrane protein pairs, ViuDand ViuG have homology with each other (Table 2).
The fourth ORF in the gene cluster, viuC, encodes a proteinwith homology to FepC and other ATPase proteins of ABC
TABLE 1. Bacterial strains and plasmids used in this study
Strain or plasmid Relevant characteristic Source orreference
StrainsLou15 V. cholerae El Tor 42O395 V. cholerae classical S. CalderwoodEWV103 V. cholerae Lou15 viuP::cam This studySSV121 V. cholerae Lou15 viuG::cam This studyMBG14 V. cholerae O395 viuA::TnphoA 16DH5a E. coli cloning strain, Ent1 18SM10lpir E. coli host for pHM5 29AB1515.414 E. coli AB1515 fepB::Tn5 C. EarhartAB1515.718 E. coli AB1515 fepD::Tn5 C. Earhart (10)AB1515.764 E. coli AB1515 fepG::Tn5 C. Earhart (10)AB1515.199 E. coli AB1515 fepC::Tn5 C. Earhart (10)
PlasmidspBluescript SK2 Cloning vector StratagenepSU18 Cloning vector 28pWSK29 Cloning vector 50pWKS30 Cloning vector 50pMA9 cam resistance gene 24pMTLcam cam resistance gene in the BamHI site of pMTL24 54pWSc1 pWKS30 carrying sacB 31pHM5 pGP704 carrying sacB 37pCP111 pACYC184 with an 11-kbp EcoRI fragment carrying fepBDGC in the EcoRI site 33pJSV90 pACYC184 with a 9-kbp EcoRI fragment carrying viuPDGC and vibD and vibH in the
EcoRI site53
pVIB109 5.5-kbp PstI-SalI fragment of pJSV90 subcloned into the PstI-SalI sites of pSU18 This studypVIB119 2.9-kbp BamHI fragment containing vibH and viuP subcloned into the BamHI site of
pBluescript SK2This study
pVIB121 HindIII fragment of pVIB109 cloned into the HindIII site of pBluescript SK2 This studypVIB147 BglII fragment from pJSV90 subcloned in the BamHI site of pWKS30 This studypVIB148 pVIB119 with a cam cassette inserted in the MscI site in viuP This studypVIB154 pVIB147 with a cam cassette inserted in the MscI site in viuP This studypVIB159 pVIB147 from which sequences to the right of the EcoRV site were deleted This studypViuAB PCR fragment containing viuA and viuB subcloned in the SmaI site of pWSK29 This studypUNK143 pHM5 containing viuG::cam This study
7590 WYCKOFF ET AL. J. BACTERIOL.
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

transporters (Table 2). ViuC contains both Walker motif A,GPNGCGKS (amino acids 52 to 60), and motif B, YLLL-DEPT (amino acids 177 to 184) (49), indicating that it is likelyto be the ATPase component of the complex. It has a predictedmolecular weight of 30,914 and a predicted pI of 7.56. ViuC ismore hydrophilic than either ViuD or ViuG, consistent with itbeing more loosely associated with the cytoplasmic membrane.The vibriobactin biosynthetic gene vibD is located immediatelydownstream of viuC (Fig. 3C).
Upstream of the viuP translation start is a potential pro-moter sequence (Fig. 3A). This region contains a sequencewith homology to the E. coli Fur box, suggesting that theexpression of these genes is regulated by iron. Approximately200 bp upstream of viuP is the potential promoter region forthe vibriobactin biosynthetic gene vibH, which also contains apotential Fur binding site. The vibH promoter does not appearto overlap the promoter for viuP. In E. coli, fepB and thedivergently transcribed gene entC are separated by a complexregion that contains direct repeats, possible stem-loop struc-tures, and an ORF of unknown function (12). The function ofthese features is not well understood, but they appear to in-fluence fepB expression (40). No direct or inverted repeatsequences are located between the viuP and vibH genes, andno ORFs of significant size are present, indicating that theregulation of viuP is likely to be different from that of fepB.
viuG and viuC are separated by 116 nucleotides (Fig. 3B).This region contains a small, inverted repeat sequence, whichmight function as a rho-independent transcription terminator.The region also contains a sequence with homology to theconsensus Fur box. These data suggest that viuC is transcribedseparately from viuPDG. The presence of a promoter in thisregion is supported by the complementation data shown below.
Frequently, genes for inner membrane permease proteinsare linked to the gene for the outer membrane receptor for thesame ligand (11). Our analysis of this region did not reveal thepresence of the vibriobactin receptor gene, viuA, or any otherpotential outer membrane receptor protein gene. Data fromthe unfinished TIGR (The Institute for Genomic Research) V.cholerae database also failed to show a closely linked potential
receptor protein gene. Recent physical mapping studies indi-cate that the V. cholerae genome consists of two large repli-cons. viuA and the vibriobactin gene cluster described here areboth located on replicon I but are separated by about 106 bp(48).
Complementation of E. coli fep mutations with viu regionclones. Preliminary complementation data, together with thesequence data presented above, suggest that the ViuPDGCproteins function as a periplasmic binding protein-dependenttransport system. In initial experiments to determine the func-tions of these genes, segments of the viu gene region weresubcloned into low-copy-number vectors, and these plasmidswere tested for the ability to complement mutations in E. colifep genes (Fig. 2). E. coli strains with a Tn5 insertion in one ofeach of the fep genes were transformed with a plasmid con-taining the entire viu region (pVIB147). Each of these trans-formed strains was able to use the siderophore enterobactin as
FIG. 2. Organization of the vibriobactin transport genes and complementation of E. coli fep mutations. The horizontal arrows indicate the direction of transcriptionof the viu genes, and selected restriction sites are indicated below the line. The vertical arrowheads above the lines indicate the position of the cam cassette insertionin the marker exchange mutations in EWV103 and SSV121. The arrow in pVIB154 indicates the position of insertion of a cam cassette into the MscI site in viuP. Theability of plasmids to complement E. coli fep mutants in a bioassay is shown to the right. 1, growth on enterobactin; 2, no growth on enterobactin. Strains used forcomplementation studies were as follows: fepB, AB1515.414; fepD, AB1515.718; fepG, AB1515.764; fepC, AB1515.199.
TABLE 2. Selected homologies of vibriobactin transport proteins toother transport proteins
Transportprotein Compared protein Identity
(%)Similarity
(%)
ViuP E. coli FepB (12) 39 48
ViuD E. coli FepD (10, 41) 42 52Erwinia chrysanthemi CbrB (26) 39 47V. cholerae HutC (31) 38 45
ViuG Mycobacterium smegmatis FxuA (14) 49 57E. coli FepG (41) 45 56E. chrysanthemi CbrC (26) 35 45V. cholerae HutC (31) 37 46V. cholerae ViuD (this study) 35 42
ViuC M. smegmatis FxuB (14) 63 72E. coli FepC (41) 58 70E. chrysanthemi CbrD (26) 55 65V. cholerae HutD (31) 40 48
VOL. 181, 1999 CATECHOL TRANSPORT IN V. CHOLERAE 7591
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

an iron source, as measured in a bioassay (Fig. 2). This indi-cates that the four Viu proteins can functionally substitute forthe Fep proteins in the transport of enterobactin.
In the above experiments, all four of the viu genes werepresent on the plasmids. Additional plasmids were constructedto determine whether the Viu proteins must assemble togetheras a Viu complex, or whether the individual Viu protein ViuP,ViuD, ViuG, or ViuC can replace the homologous Fep pro-tein, FepB, FepD, FepG, or FepC, respectively, to form anactive transport complex. The plasmid pVIB154, in which apolar chloramphenicol cassette was inserted into the MscI sitein viuP (Fig. 2), complemented a mutation in fepC but not infepB, fepD, or fepG. The cassette inserted into viuP should bepolar on viuD and viuG but not on viuC, which appears to haveits own promoter. Thus, viuC is the only gene likely to beexpressed from this plasmid, suggesting that ViuC can substi-tute for FepC. This is supported by the observation that the
plasmid pVIB109, containing viuG and viuC also comple-mented the fepC mutation. pVIB109 did not complement thefepG mutation. The plasmid pVIB159, containing viuP, failedto complement a mutation in the viuP homologue, fepB. Al-though the E. coli fepB mutation is a Tn5 insertion and is polaron downstream genes, the fepDGC genes are transcribed froma separate promoter (11), and thus the transport defect isspecific for fepB. When the plasmids pVIB109 and pVIB159were carried together in the same cell, complementation ofeach of the fep mutations was observed (Fig. 2). This indicatesthat the lack of fepB or fepG complementation observed with asingle plasmid is not due to poor expression of the viuP andviuG genes from these plasmids.
The observation that pVIB109 and pVIB159 did not indi-vidually complement the fepBDG mutations, but did comple-ment these mutations when present together, suggests that theproteins encoded by these two plasmids may interact. We pro-
FIG. 3. Portions of the nucleotide sequence and the predicted amino acid sequences of ViuP and related proteins. (A) The nucleotide sequence and predictedtranslation of the 59 end of viuP and the sequence of the vibH-viuP intergenic region. Possible fur boxes for vibH and viuP are underlined. The potential cleavage sitefor lipoprotein modification is indicated by an arrowhead. (B) The viuG-viuC intergenic region. An inverted repeat that may function as a transcription terminator isindicated by arrows, and a possible fur box for the viuC transcript is underlined. (C) The viuC-vibD intergenic region. The end of viuC and predicted start of vibD areshown. Three potential ATG start codons are present at the 59 end of the vibD ORF. (D) The predicted amino terminal sequence of FatB (1), C. coli CeuE (35), HutB(31), and ViuP. The consensus lipoprotein cleavage site is also shown; the asterisk below the alignments indicates the conserved cysteine.
7592 WYCKOFF ET AL. J. BACTERIOL.
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
pose that ViuPDGC function together as a unit and are unableto form the proper protein-protein contacts with E. coli Fepproteins. Thus, with the exception of ViuC, the individual Viuproteins are unable to assemble with Fep proteins to form anactive permease complex. The ability of the ATPase homo-logue, ViuC, to functionally substitute for FepC is consistentwith observations in other ABC transport systems, where someATPase subunits can function with two different inner mem-brane permease complexes (20, 39, 51).
Transport of vibriobactin by the ViuPDGC proteins. Theability of the viuPDGC genes to complement E. coli fep muta-tions indicates that they encode a transport system capable oftransporting enterobactin across the cytoplasmic membrane.The presence of these genes within a vibriobactin biosyntheticcluster, however, suggested that their usual ligand in V. chol-erae is vibriobactin. E. coli strains carrying only the viuPDGCgenes do not transport vibriobactin. These E. coli strains, how-ever, lack both ViuA, the vibriobactin outer membrane recep-tor, and ViuB, which catalyzes the removal of iron from theferri-vibriobactin complex. To test for vibriobactin transport inE. coli, we first constructed the plasmid, pViuAB, which con-tains the viuA and viuB genes. The plasmids pViuAB andpVIB147 are incompatible, so E. coli fep mutants carryingpViuAB were transformed with the compatible plasmidpJSV90, which encodes ViuPDGC (53). These strains werethen tested for the ability to transport vibriobactin and enter-obactin (Table 3). Strains carrying pViuAB together with theviuPDGC genes encoded on pJSV90 formed large zones ofgrowth around both Lou15 and DH5a, indicating that theViuPDGC proteins are capable of transporting vibriobactin aswell as enterobactin. This ability of ViuPDGC to transportthese two rather dissimilar catechols (Fig. 1) is reminiscent ofhydroxamate transport in E. coli. In this system, each hydrox-amate has a specific outer membrane receptor, but are alltransported across the cytoplasmic membrane by the sameperiplasmic binding protein and permease complex (FhuBCD)(6). In our experiments, FepA and ViuA were specific for their
ligands, in that FepA transported enterobactin but not vibri-obactin and ViuA transported vibriobactin but not enterobac-tin (data not shown).
To test whether the E. coli FepBDGC proteins are alsocapable of transporting both enterobactin and vibriobactin, thefepBDGC genes were introduced into fep mutant strains car-rying pViuAB. These cells grew well around DH5a, but growtharound Lou15 was not observed (Table 3). Thus, unlikeViuPDGC, the E. coli enterobactin ABC transport systemtransported only enterobactin in these experiments. Thus, theability to transport both of these catechols does not appear tobe a universal property of catechol transport systems. The E.coli FepBDGC system is not absolutely specific for the trans-port of enterobactin, since it has been shown previously totransport the catechol dihydroxybenzoylserine (19).
When neither pJSV90 (carrying viuPDGC) nor pCP111 (car-rying fepBDGC) were present in fep mutant strains carryingpViuAB, very weak growth was observed around Lou15 (Table3). The fep mutant strains used in this assay can synthesize, butnot use, enterobactin. When the fep genes are supplied on aplasmid, a higher level of background growth is observed, dueto the ability of the strains to transport the enterobactin thatthey are producing. This background growth may obscure asmall amount of growth around an iron source. In contrast, thelevel of background growth observed with strains unable totransport enterobactin is extremely low, which allows detectionof very weak utilization of an iron source. One of the manysystems that transports ligands across the inner membrane islikely also to transport vibriobactin at very low efficiency.
Characterization of V. cholerae viu mutants. To determinethe role of the viu genes in V. cholerae, chromosomal mutationswere constructed in the wild-type El Tor strain Lou15 usingmarker exchange. The strain EWV103 has a chloramphenicolresistance cassette inserted in the MscI site within viuP, andSSV121 has a chloramphenicol cassette in the StuI site in viuG(Fig. 2). The ability of the strains to use vibriobactin as an ironsource was determined in a bioassay using a high EDDA con-centration (1 mg/ml) (Table 4). Under conditions of iron lim-itation, the growth of the parental strain, Lou15, was stimu-lated by vibriobactin, whereas both EWV103 and SSV121failed to use vibriobactin as an iron source. This indicates thatViuP and ViuG function in the transport of vibriobactin in V.cholerae. The mutants did not have a general iron transportdefect, since both EWV103 and SSV121 utilized the hydrox-
FIG. 4. Visualization of cellular lipoproteins. E. coli DH5a carrying eitherpBluescript (lane 1) or pVIB119 encoding viuP (lane 2) were grown in thepresence of [3H]palmitic acid. Proteins were then separated by SDS–12.5%PAGE and visualized by fluorography. The arrow indicates the position of ViuP.The position of the 30-kDa marker is shown at the left.
TABLE 3. ViuPDGC, but not FepBDGC, transports vibriobactinin E. coli
Indicator strain(plasmid-encoded genes)
Zone of stimulation (mm) witha:
Lou15 DH5a FeSO4
fepBb (viuAB) 14c 0 21fepB (viuAB and viuPDGC) 40 26 38fepB (viuAB and fepBDGC) 0 27 35fepD (viuAB) 18c 0 28fepD (viuAB and viuPDGC) 36 25 29fepD (viuAB and fepBDGC) 0 38 40
a Cultures of the indicator strains were seeded into L agar containing 300 mgof EDDA per ml. V. cholerae Lou15, E. coli DH5a, or 10 mM FeSO4 was spottedonto the medium. The zone of growth of the indicator strains was measured 18 hafter inoculation.
b The fepB strain used was AB1515.414 and the fepD strain was AB1515.718.The plasmid carrying the viuAB was pViuAB; the plasmid carrying viuPDGC waspJSV90; the plasmid carrying fepBDGC was pCP111.
c A very sparse zone of growth was observed around vibriobactin in indicatorstrains that do not transport enterobactin. The density of colonies within thesezones was much lower than those observed in other zones (see text).
VOL. 181, 1999 CATECHOL TRANSPORT IN V. CHOLERAE 7593
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

amate siderophore ferrichrome (Table 4). When the mutantstrains were transformed with plasmids carrying the viu genes,vibriobactin utilization was restored, indicating that the ob-served defects were due to the viu mutation. Both EWV103and SSV121 stimulated the growth of V. cholerae strains underconditions of iron limitation, indicating that neither strain wasdefective in vibriobactin production (data not shown).
Evidence for additional genes for the transport of catecholsiderophores in V. cholerae. The V. cholerae strains were alsotested for siderophore transport at a reduced EDDA concen-tration of 500 mg/ml (Table 5). Transport of enterobactin hasbeen previously observed in V. cholerae (38), and at this EDDAconcentration, the transport of enterobactin was observed inour assay. When assayed at this EDDA concentration, the viumutants used both vibriobactin and enterobactin, but the zoneswere smaller and less dense than those observed with thewild-type strain. In the mutant strains, the zones around vibri-obactin and enterobactin are of approximately equal size, incontrast to the wild-type strain, which had a larger zone aroundvibriobactin (Table 5). Although it was necessary to reduce theEDDA concentration to observe growth of the mutants onvibriobactin, the amount of EDDA used (500 mg/ml) is still arelatively high concentration of the chelator. It is expected thata high-affinity transport system would be required to observeiron transport under these conditions. The ability of the viumutants to use vibriobactin under these conditions suggests thepresence of an additional system for the transport of catecholsiderophores in V. cholerae.
To investigate this further, the transport phenotype of astrain carrying a mutation in the vibriobactin outer membranereceptor gene, viuA, was further examined. The strain MBG14(viuA::TnphoA) and its classical biotype parental strain, O395,were tested for transport of vibriobactin and enterobactin (Ta-
ble 4). MBG14 was completely defective in vibriobactin trans-port, consistent with previous characterization of this mutant(45). Transport of enterobactin, however, was unaffected in themutant, indicating that enterobactin enters the cell via a ViuA-independent transport system. This indicates that additionalgenes for the transport of catechols are present in V. cholerae.
In this paper, we have presented additional characterizationof the genes within a vibriobactin locus in V. cholerae. Four ofthe genes within this locus encode proteins with sequencehomology to periplasmic binding proteins and cytoplasmic per-mease proteins (Table 2). These proteins transport both vibri-obactin and enterobactin in E. coli. With the exception ofViuC, they appear to function together as a complex ratherthan assembling with the E. coli Fep proteins to form a mixedcomplex. In V. cholerae, the Viu proteins function in the trans-port of vibriobactin, but additional proteins that transport cat-echol siderophores are also present.
A model for the transport of catechols in V. cholerae ispresented in Fig. 5. In this model, vibriobactin crosses theouter membrane through ViuA, and enterobactin crossesthrough a separate outer membrane protein, which is specificfor enterobactin. This protein has been designated VctA forVibrio catechol transport. The ViuPDGC system can transportboth vibriobactin and enterobactin in E. coli, and we anticipatethat it can also transport both of these ligands in V. cholerae.Because the viuP and viuG mutations reduced, but did notcompletely abolish, catechol transport, we also propose that V.cholerae contains at least one additional system for the trans-port of catechol siderophores across the inner membrane (des-ignated VctPDGC). In Fig. 5, we show a single, additionalcytoplasmic membrane permease system that transports bothvibriobactin and enterobactin. This is the simplest model con-sistent with the data, but more complex models involving mul-tiple, additional permease systems are also possible. At thistime, there is no information about the organization of theseunidentified catechol transport protein genes. It is also notknown whether the Vct system transports intact vibriobactinand enterobactin, or whether it transports breakdown productsderived from these siderophores. As shown in Fig. 5, the finalstep in iron transport is the removal of iron from the iron-siderophore complex by ViuB. Previously published data sug-gest that ViuB can function in the utilization of both ferri-vibriobactin and ferri-enterobactin (8).
FIG. 5. Model for catechol siderophore transport in V. cholerae.
TABLE 4. Vibriobactin transport at high EDDA concentrationsrequires the Viu proteins in V. choleraea
Indicator strain Genotype orphenotype
Zone of stimulation(mm) with:
Lou15 Ferrichrome
Lou15 Wild type 14 15EWV103 viuP 0 12EWV103/pVIB147 ViuP1 9 15SSV121 viuG 0 11SSV121/pVIB147 ViuG1 15 16
a Cultures of the indicator strains were seeded into L agar containing 1 mg ofEDDA per ml. V. cholerae Lou15 or ferrichrome was spotted onto the medium.The zone of growth was measured 18 h after inoculation.
TABLE 5. Evidence for an additional catechol transport systemin V. choleraea
Indicator strain Genotype orphenotype
Zone of stimulation (mm) with:
Lou15 DH5a Ferrichrome
Lou15 Wild type 26 17 28EWV103 viuP 9 9 23EWV103/pVIB147 ViuP1 22 12 23SSV121 viuG 8 10 17SSV121/pVIB147 ViuG1 25 21 28O395 Wild type 21 12 21MBG14 viuA 0 13 17
a Cultures of the indicator strains were seeded into L agar containing 500 mgof EDDA per ml. V. cholerae Lou15, E. coli DH5a, or ferrichrome was spottedonto the medium. The zone of growth was measured 18 h after inoculation.
7594 WYCKOFF ET AL. J. BACTERIOL.
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

It has been difficult to determine the relative contribution ofthe different iron transport systems to the survival and growthof V. cholerae in mammalian hosts. Classical strains carrying amutation in the gene for vibriobactin synthesis (43), vibriobac-tin transport (23), or heme transport (23, 47) are only weaklyattenuated in animal models. The virulence of the viuA-hutAdouble mutant was more attenuated than any of the singlemutants, suggesting that both of these transport systems con-tribute to iron acquisition in vivo (23, 47). While it is possiblethat an unidentified system is responsible for most iron acqui-sition in vivo, it is more likely that multiple systems are present,each contributing to the acquisition of iron in different envi-ronments or at different times during the course of infection.The presence of multiple systems may reflect the overall im-portance of iron acquisition in this organism.
ACKNOWLEDGMENTS
This work was supported by the Foundation for Research and bygrant AI16935 from the National Institutes of Health.
We thank Charles Earhart for providing strains and for helpfuldiscussions. We also thank Charles Earhart, Douglas Henderson, andLaura Runyen-Janecky for comments on the manuscript and ChrisTinkle for technical assistance.
REFERENCES
1. Actis, L. A., M. Tolmasky, L. M. Crosa, and J. Crosa. 1995. Characterizationand regulation of the expression of FatB, an iron transport protein encodedby the pJM1 virulence plasmid. Mol. Microbiol. 17:197–204.
2. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990.Basic local alignment search tool. J. Mol. Biol. 215:403–410.
3. Arnow, L. E. 1937. Colorimetric determination of the components of 3,4-dihydroxyphenylalanine tyrosine mixtures. J. Biol. Chem. 118:531–537.
4. Bagg, A., and J. B. Neilands. 1987. Ferric uptake regulation protein acts asa repressor, employing iron (II) as a cofactor to bind the operator of an irontransport operon in Escherichia coli. Biochemistry 26:5471–5477.
5. Boos, W., and J. M. Lucht. 1996. Periplasmic binding protein-dependentABC transporters, p. 1175–1209. In F. C. Neidhardt, R. Curtiss III, J. L.Ingraham, E. C. C. Lin, K. Brooks Low, B. Magasanik, W. S. Reznikoff, M.Riley, M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Sal-monella: cellular and molecular biology, 2nd ed., vol. 1. ASM Press, Wash-ington, D.C.
6. Braun, V., K. Hantke, and W. Koster. 1998. Bacterial iron transport: mech-anisms, genetics, and regulation, p. 67–145. In A. Sigel and H. Sigel (ed.),Metal ions in biological systems, vol. 35. Marcel Dekker, New York, N.Y.
7. Buchanan, S. K., B. S. Smith, L. Venkatramani, D. Xia, L. Esser, M. Palnit-kar, R. Chakraborty, D. van der Helm, and J. Deisenhofer. 1999. Crystalstructure of the outer membrane active transporter FepA from Escherichiacoli. Nat. Struct. Biol. 6:56–63.
8. Butterton, J. R., and S. B. Calderwood. 1994. Identification, cloning, andsequencing of a gene required for ferric vibriobactin utilization by Vibriocholerae. J. Bacteriol. 176:5631–5638.
9. Butterton, J. R., J. A. Stoebner, S. M. Payne, and S. B. Calderwood. 1992.Cloning, sequencing, and transcriptional regulation of viuA, the gene encod-ing the ferric vibriobactin receptor of Vibrio cholerae. J. Bacteriol. 174:3729–3738.
10. Chenault, S. S., and C. F. Earhart. 1991. Organization of genes encodingmembrane proteins of the Escherichia coli ferrienterobactin permease. Mol.Microbiol. 5:1405–1413.
11. Earhart, C. F. 1996. Uptake and metabolism of iron and molybdenum, p.1075–1090. In F. C. Neidhardt, R. Curtiss III, J. L. Ingraham, E. C. C. Lin,K. Brooks Low, B. Magasanik, W. S. Reznikoff, M. Riley, M. Schaechter, andH. E. Umbarger (ed.), Escherichia coli and Salmonella: cellular and molec-ular biology, 2nd ed., vol. 1. ASM Press, Washington, D.C.
12. Elkins, M. F., and C. F. Earhart. 1989. Nucleotide sequence and regulationof the Escherichia coli gene for ferrienterobactin transport protein FepB. J.Bacteriol. 171:5443–5451.
13. Faruque, S. M., M. J. Albert, and J. J. Mekalanos. 1998. Epidemiology,genetics and ecology of toxigenic Vibrio cholerae. Microbiol. Mol. Biol. Rev.62:1301–1314.
14. Fiss, E. H., S. Yu, and W. R. Jacobs. 1994. Identification of genes involvedin the sequestration of iron in mycobacteria: the ferric exochelin biosyntheticand uptake pathways. Mol. Microbiol. 14:557–569.
15. Gay, P., D. LeCoq, M. Steinmetz, T. Berkelman, and C. I. Kado. 1985.Positive selection procedure for entrapment of insertion sequence elementsin gram-negative bacteria. J. Bacteriol. 164:918–921.
16. Goldberg, M. B., V. J. DiRita, and S. B. Calderwood. 1990. Identification of
an iron-regulated virulence determinant in Vibrio cholerae, using TnphoAmutagenesis. Infect. Immun. 58:55–60.
17. Griffiths, G. L., S. P. Sigel, S. M. Payne, and J. B. Neilands. 1984. Vibri-obactin, a siderophore from Vibrio cholerae. J. Biol. Chem. 259:383–385.
18. Hanahan, D. 1983. Studies on transformation of Escherichia coli with plas-mids. J. Mol. Biol. 166:557–580.
19. Hantke, K. 1990. Dihydroxybenzoylserine—a siderophore for E. coli. FEMSMicrobiol. Lett. 67:5–8.
20. Hekstra, D., and J. Tommassen. 1993. Functional exchangeability of theABC proteins of the periplasmic binding protein-dependent transport sys-tems Ugp and Mal of Escherichia coli. J. Bacteriol. 175:6546–6552.
21. Henderson, D. P., and S. M. Payne. 1994. Characterization of the Vibriocholerae outer membrane heme transport protein HutA: sequence of thegene, regulation of expression, and homology to the family of TonB-depen-dent proteins. J. Bacteriol. 176:3269–3277.
22. Henderson, D. P., and S. M. Payne. 1993. Cloning and characterization ofthe Vibrio cholerae genes encoding the utilization of iron from haemin andhaemoglobin. Mol. Microbiol. 7:461–469.
23. Henderson, D. P., and S. M. Payne. 1994. Vibrio cholerae iron transportsystems: role of heme and siderophore iron transport in virulence and iden-tification of a gene associated with multiple iron transport systems. Infect.Immun. 62:5120–5125.
24. Hong, M., and S. M. Payne. 1997. Effect of mutations in Shigella flexnerichromosomal and plasmid-encoded lipopolysaccharide genes on invasionand serum resistance. Mol. Microbiol. 24:779–791.
25. Kaper, J. B., J. G. Morris, and M. M. Levine. 1995. Cholera. Clin. Microbiol.Rev. 8:48–86.
26. Mahe, B., C. Masclaux, L. Rauscher, C. Enard, and D. Expert. 1995. Dif-ferential expression of two siderophore-dependent iron-acquisition pathwaysin Erwinia chrysanthemi 3937: characterization of a novel ferrisiderophorepermease of the ABC transporter family. Mol. Microbiol. 18:33–43.
27. Marck, C. 1988. “DNA strider”: a “C” program for the fast analysis of DNAand protein sequences on the Apple Macintosh family of computers. NucleicAcids Res. 16:1829–1836.
28. Martinez, E., B. Bartolome, and F. de la Cruz. 1988. pACYC184-derivedcloning vectors containing the multiple cloning site and lacZ a reporter geneof pUC8/9 and pUC18/19 plasmids. Gene 68:159–162.
29. Miller, V. L., and J. J. Mekalanos. 1988. A novel suicide vector and its usein construction of insertion mutations: osmoregulation of outer membraneproteins and virulence determinants in Vibrio cholerae required toxR. J.Bacteriol. 170:2575–2583.
30. Moeck, G. S., and J. W. Coulton. 1998. TonB-dependent iron acquisition:mechanisms of siderophore-mediated active transport. Mol. Microbiol. 28:675–681.
31. Occhino, D. A., E. E. Wyckoff, D. P. Henderson, T. J. Wrona, and S. M.Payne. 1998. Vibrio cholerae iron transport: haem transport genes are linkedto one of two sets of tonB, exbB, exbD genes. Mol. Microbiol. 29:1493–1507.
32. Park, S. F., and P. T. Richardson. 1995. Molecular characterization of aCampylobacter jejuni lipoprotein with homology to periplasmic siderophore-binding protein. J. Bacteriol. 177:2259–2264.
33. Pierce, J. R., and C. F. Earhart. 1986. Escherichia coli K-12 envelope pro-teins specifically required for ferrienterobactin uptake. J. Bacteriol. 166:930–936.
34. Pugsley, A. P. 1993. The complete general secretory pathway in gram-neg-ative bacteriae. Microbiol. Rev. 57:50–108.
35. Richardson, P. T., and S. F. Park. 1995. Enterochelin acquisition in Campy-lobacter coli: characterization of components of a binding-protein-dependenttransport system. Microbiology 141:3181–3191.
36. Rogers, H. J. 1973. Iron-binding catechols and virulence in Escherichia coli.Infect. Immun. 7:445–456.
37. Runyen-Janecky, L. J., M. Hong, and S. M. Payne. 1999. Virulence plasmid-encoded impCAB operon enhances survival and induced mutagenesis inShigella flexneri after exposure to UV radiation. Infect. Immun. 67:1415–1423.
38. Rutz, J. M., T. Abdullah, S. P. Singh, V. I. Kalve, and P. E. Klebba. 1991.Evolution of the ferric enterobactin receptor in gram-negative bacteria. J.Bacteriol. 173:5964–5974.
39. Schlosser, A., T. Kampers, and H. Schrempf. 1997. The Streptomyces ATP-binding component of MsiK assists in cellobiose and maltose transport. J.Bacteriol. 179:2092–2095.
40. Schmitt, M. P., and S. M. Payne. 1991. Genetic analysis of the enterobactingene cluster in Shigella flexneri. J. Bacteriol. 173:816–825.
41. Shea, C. M., and M. A. McIntosh. 1991. Nucleotide sequence and geneticorganization of the ferric enterobactin transport system: homology to otherperiplasmic binding protein-dependent systems in Escherichia coli. Mol. Mi-crobiol. 5:1415–1428.
42. Sigel, S. P., and S. M. Payne. 1982. Effect of iron limitation on growth,siderophore production and expression of outer membrane proteins ofVibrio cholerae. J. Bacteriol. 150:148–155.
43. Sigel, S. P., J. A. Stoebner, and S. M. Payne. 1985. Iron-vibriobactin trans-port system is not required for virulence of Vibrio cholerae. Infect. Immun.47:360–362.
VOL. 181, 1999 CATECHOL TRANSPORT IN V. CHOLERAE 7595
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

44. Staudenmaier, H., B. Van Hove, Z. Yaraghi, and V. Braun. 1989. Nucleotidesequences of the fecBCDE genes and locations of the proteins suggest aperiplasmic-binding-protein-dependent transport mechanism of iron(III) di-citrate in Escherichia coli. J. Bacteriol. 171:2626–2633.
45. Stoebner, J. A., J. R. Butterton, S. B. Calderwood, and S. M. Payne. 1992.Identification of the vibriobactin receptor of Vibrio cholerae. J. Bacteriol.174:3270–3274.
46. Sutcliffe, I. C., and R. R. B. Russell. 1995. Lipoproteins of gram-positivebacteria. J. Bacteriol. 177:1123–1128.
47. Tashima, K. T., P. A. Carroll, M. B. Rogers, and S. B. Calderwood. 1996.Relative importance of three iron-regulated outer membrane proteins for invivo growth of Vibrio cholerae. Infect. Immun. 64:1756–1761.
48. Trucksis, M., J. Michalski, Y. K. Deng, and J. B. Kaper. 1998. The Vibriocholerae genome contains two unique circular chromosomes. Proc. Natl.Acad. Sci. USA 95:14464–14469.
49. Walker, J. E., M. Saraste, M. J. Runswick, and N. J. Gay. 1982. Distantlyrelated sequences in the alpha- and beta-subunits of ATP synthase, myosin,kinases and other ATP-requiring enzymes and a common nucleotide bindingfold. EMBO J. 1:945–951.
50. Wang, R. F., and S. R. Kushner. 1991. Construction of versatile low-copy-number vectors for cloning, sequencing and gene expression in Escherichiacoli. Gene 100:195–199.
51. Wilken, S., G. Schmees, and E. Schneider. 1996. A putative helical domainin the MalK subunit of the ATP-binding-cassette transport system for mal-tose of Salmonella typhimurium (MalFGK2) is crucial for interaction withMalF and MalG. A study using the LacK protein of Agrobacterium ra-diobacter as a tool. Mol. Microbiol. 22:655–666.
52. Wu, H. C. 1996. Biosynthesis of lipoprotein, p. 1005–1014. In F. C. Neid-hardt, R. Curtiss III, J. L. Ingraham, E. C. C. Lin, K. Brooks Low, B.Magasanik, W. S. Reznikoff, M. Riley, M. Schaechter, and H. E. Umbarger(ed.), Escherichia coli and Salmonella: cellular and molecular biology, 2nded., vol. 1. ASM Press, Washington, D.C.
53. Wyckoff, E., J. A. Stoebner, K. E. Reed, and S. M. Payne. 1997. Cloning of aVibrio cholerae vibriobactin gene cluster: identification of genes required forearly steps in siderophore biosynthesis. J. Bacteriol. 179:7055–7062.
54. Wyckoff, E. E., D. Duncan, A. G. Torres, M. Mills, K. Maase, and S. M.Payne. 1998. Structure of the Shigella dysenteriae haem transport locus and itsphylogenetic distribution in enteric bacteria. Mol. Microbiol. 28:1139–1152.
55. Yamamoto, S., M. A. R. Chowdhury, M. Kuroda, T. Nakano, Y. Koumoto,and S. Shinoda. 1991. Further study on polyamine compositions in Vibrion-aceae. Can. J. Microbiol. 37:148–153.
56. Yamamoto, S., S. Shinoda, M. Kawaguchi, K. Wakamatsu, and M. Makita.1983. Polyamine distribution in Vibrionaceae: norspermidine as a generalconstituent of Vibrio species. Can. J. Microbiol. 29:724–728.
7596 WYCKOFF ET AL. J. BACTERIOL.
on Novem
ber 11, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
Copyright © 2022 FDOKUMEN



















