
A major terminal drought tolerance QTL of pearl millet is also associated with reduced salt uptake...
16
A major terminal drought tolerance QTL of pearl millet is also associated with reduced salt uptake and enhanced growth under salt stress Parbodh C. Sharma • Deepmala Sehgal • Dhananjay Singh • Gurbachan Singh • Rattan S. Yadav Received: 16 December 2009 / Accepted: 24 February 2010 / Published online: 13 March 2010 Ó Springer Science+Business Media B.V. 2010 Abstract The performance of a major quantitative trait locus (QTL) of terminal drought tolerance (DT) of pearl millet was assessed under salt stress. The test-cross hybrids of the QTL donor parent (drought tolerant, PRLT 2/89-33), QTL recipient parent (drought sensitive, H 77/833-2), and a set of six near isogenic lines introgressed with a terminal DT-QTL (QTL-NILs) were evaluated for germination and seedling emergence at 7 days after sowing (DAS) in Petri plates at four salinity levels, and at vegetative (24 DAS) and maturity stages at three salinity and alkalinity levels. Na ? and K ? accumulation, their compartmentation in different plant parts, and their effects on growth and yield parameters were evalu- ated. The DT-QTL donor parent and QTL-NILs accumulated less Na ? in shoot parts at seedling, vegetative and maturity stages, and also partitioned the accumulated Na ? more into nodes and internodes and less into leaves than the drought-sensitive recurrent parent. The pattern of reduced salt accu- mulation in the drought-tolerant parent and QTL- NILs was consistently associated with better growth and productivity in saline and alkaline treatments. It is concluded that the DT-QTL contributed by PRLT 2/89-33 exerted favourable effects on growth and productivity traits under salt stress by limiting Na ? accumulation in leaves. Keywords Pearl millet Á Drought and salt tolerance Á Terminal drought tolerance QTL Á QTL-NILs Á Growth under salt stress Á Ionic accumulation and compartmentation Abbreviations DAS Days after sowing DT-QTL Drought tolerance QTL DT-QTL-NILs DT-QTL-near isogenic lines ECiw Electrical conductivity of the irrigation water GCA General combining ability MABC Marker-assisted backcrossing Introduction Pearl millet (Pennisetum glaucum L. Br.) is an annual diploid crop belonging to the Poaceae. Globally, it is the seventh most important cereal crop and provides food security to 500 million of the poorest people living predominantly in parts of Asia and Africa. Among the cereals, it is considered the hardiest crop, which is grown in outlying areas peripheral to the major production areas where crops like sorghum and P. C. Sharma Á D. Singh Á G. Singh Central Soil Salinity Research Institute (CSSRI), Karnal, Haryana 132001, India D. Sehgal Á R. S. Yadav (&) Institute of Biological, Environmental & Rural Sciences (IBERS), Aberystwyth University, Gogerddan, Aberystwyth SY23 3EB, UK e-mail: [email protected] 123 Mol Breeding (2011) 27:207–222 DOI 10.1007/s11032-010-9423-3
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A major terminal drought tolerance QTL of pearl millet is also associated with reduced salt uptake...

A major terminal drought tolerance QTL of pearl milletis also associated with reduced salt uptake and enhancedgrowth under salt stress
Parbodh C. Sharma • Deepmala Sehgal •
Dhananjay Singh • Gurbachan Singh •
Rattan S. Yadav
Received: 16 December 2009 / Accepted: 24 February 2010 / Published online: 13 March 2010
� Springer Science+Business Media B.V. 2010
Abstract The performance of a major quantitative
trait locus (QTL) of terminal drought tolerance (DT)
of pearl millet was assessed under salt stress. The
test-cross hybrids of the QTL donor parent (drought
tolerant, PRLT 2/89-33), QTL recipient parent
(drought sensitive, H 77/833-2), and a set of six near
isogenic lines introgressed with a terminal DT-QTL
(QTL-NILs) were evaluated for germination and
seedling emergence at 7 days after sowing (DAS) in
Petri plates at four salinity levels, and at vegetative
(24 DAS) and maturity stages at three salinity and
alkalinity levels. Na? and K? accumulation, their
compartmentation in different plant parts, and their
effects on growth and yield parameters were evalu-
ated. The DT-QTL donor parent and QTL-NILs
accumulated less Na? in shoot parts at seedling,
vegetative and maturity stages, and also partitioned
the accumulated Na? more into nodes and internodes
and less into leaves than the drought-sensitive
recurrent parent. The pattern of reduced salt accu-
mulation in the drought-tolerant parent and QTL-
NILs was consistently associated with better growth
and productivity in saline and alkaline treatments. It
is concluded that the DT-QTL contributed by PRLT
2/89-33 exerted favourable effects on growth and
productivity traits under salt stress by limiting Na?
accumulation in leaves.
Keywords Pearl millet � Drought and salt
tolerance � Terminal drought tolerance QTL �QTL-NILs � Growth under salt stress �Ionic accumulation and compartmentation
Abbreviations
DAS Days after sowing
DT-QTL Drought tolerance QTL
DT-QTL-NILs DT-QTL-near isogenic lines
ECiw Electrical conductivity of the
irrigation water
GCA General combining ability
MABC Marker-assisted backcrossing
Introduction
Pearl millet (Pennisetum glaucum L. Br.) is an annual
diploid crop belonging to the Poaceae. Globally, it is
the seventh most important cereal crop and provides
food security to 500 million of the poorest people
living predominantly in parts of Asia and Africa.
Among the cereals, it is considered the hardiest crop,
which is grown in outlying areas peripheral to the
major production areas where crops like sorghum and
P. C. Sharma � D. Singh � G. Singh
Central Soil Salinity Research Institute (CSSRI),
Karnal, Haryana 132001, India
D. Sehgal � R. S. Yadav (&)
Institute of Biological, Environmental & Rural Sciences
(IBERS), Aberystwyth University, Gogerddan,
Aberystwyth SY23 3EB, UK
e-mail: [email protected]
123
Mol Breeding (2011) 27:207–222
DOI 10.1007/s11032-010-9423-3

maize fail to produce. In the hottest and driest parts of
arid and semi-arid tropics, it is mainly grown as a
rainfed crop where yearly variations in its productiv-
ity may be ascribed to climatic variations such as
severe droughts and untimely monsoonal rains during
the cropping season. Besides drought, the crop also
suffers from the problem of excessive salts that are
present in the soil or are added by poor-quality
irrigation water. Improved adaptation to dual stresses
of water and salt is thus a desirable trait to
incorporate in pearl millet cultivars.
Recent genetic mapping research in pearl millet has
defined a number of quantitative trait loci (QTL) for
components of grain and stover yield, and for yield
maintenance under terminal drought stress conditions.
Most importantly, a major QTL associated with grain
yield per se and for drought tolerance of grain yield
has been identified on linkage group (LG) 2 which
accounted up to 32% of the phenotypic variation of
grain yield in the mapping populations (Bidinger et al.
2007; Yadav et al. 2002, 2004). The effects of this
QTL have been validated in two independent marker-
assisted backcrossing programs in which the 30%
improvement in grain yield general combining ability
(GCA) expected of this QTL under terminal drought
stress conditions was recovered in introgression lines
based on the QTL (Serraj et al. 2005).
Exposure of crop plants to drought or salt stress
triggers many similar reactions. For example, both
stresses lead to cellular dehydration, which causes
osmotic stress and removal of water from the cyto-
plasm into the extracellular space, resulting in reduc-
tion of the cytosolic and vacuolar volumes (Munns
2002; Munns and Richards 2007; Zhu 2002). Recent
investigations in Arabidopsis and rice indicate that
responses to cold, drought and salt stress are controlled
by common and conserved regulatory pathways
(Kreps et al. 2002; Rabbani et al. 2003; Seki et al.
2002; Yamaguchi-Shinozaki and Shinozaki 2006)
and, therefore, provide the prospect that QTL/gene(s)
might have a pleiotropic effect on multiple stress
tolerance. The present study was, therefore, aimed at
examining whether a drought tolerance QTL in pearl
millet would have a pleiotropic effect on salt tolerance.
To test this hypothesis, the performance of a set of near
isogenic lines introgressed with a terminal drought
tolerance QTL (QTL-NILs) was tested under a range
of salinity and alkalinity treatments. The results of the
present study will inform deployment of this drought
tolerance QTL into pearl millet cultivars that are high-
yielding and grown in the areas where twin problem of
salinity and drought are common.
Materials and methods
Plant material
Test-cross hybrids of two parental genotypes [PRLT 2/
89-33 (drought-tolerant, donor parent for drought
tolerance QTL) and H 77/833-2 (drought-sensitive,
recipient parent for drought tolerance QTL)] and a set
of six genotypes (ICMR 01029, ICMR 01031, ICMR
01040, ICMR 01046, ICMR 02042 and ICMR 02044)
introgressed with a terminal drought tolerance QTL
(QTL-NILs) were selected for this study. The devel-
opment of QTL-NILs as well as details of F1 hybrid
seed produced on these lines, and on parental geno-
types, is described in Serraj et al. (2005). Briefly, the
genomic interval on LG 2 conferring drought tolerance
(Bidinger et al. 2007; Yadav et al. 2002, 2004) was
transferred from PRLT 2/89-33 (derived from the
Iniadi landrace material from Togo and Ghana and
known for its better grain-filling ability under terminal
drought stress conditions) to the genetic background of
H 77/833-2 (widely cultivated in the Thar desert
margins of north-western India and male parent of a
number of thermotolerant, extra-early, high-tillering
and high-yielding pearl millet hybrids) following four
generations of marker-assisted backcrossing (MABC).
At each backcross, the presence or absence of the
terminal drought tolerance QTL region was deter-
mined using molecular markers bracketing the drought
tolerance QTL interval on LG 2. After four generations
of MABC, two generations of selfing was done and
marker-assisted selection was performed to generate
QTL-NILs. These QTL-NILs were similar to the
recurrent parent (H 77/833-2) except for the drought
tolerance QTL interval on LG 2 transferred from the
donor parent PRLT 2/89-33. Test-cross hybrids were
produced for each of these six QTL-NILs (ICMR
01029 to ICMR 01044), as well as onto the two
parental lines (H 77/833-2 and PRLT 2/89-33), by
using each of these lines as a pollinator onto a common
male sterile line (female parent 843A). Performances
of the eight hybrids so produced were evaluated under
controlled saline and alkaline conditions. The test-
cross hybrids involving ICMR 01029, ICMR 01031
208 Mol Breeding (2011) 27:207–222
123

and ICMR 02041 were previously found to be superior
for their yield under terminal drought tolerance,
whereas ICMR 02042 and 02044 had a yield response
in the field similar to the test-cross hybrid of the
sensitive parent H 77/833-2 (Serraj et al. 2005).
Plant growth and exposure to salt stress
Test-cross hybrids of six QTL-NILs and two parental
genotypes were evaluated for their salt tolerance
under solution culture in Petri plates, and sand culture
in pots, as well as in soil in microplots. In Petri plates,
germination and seedling emergence at 7 days after
sowing (DAS) in saline solution was evaluated.
Under sand (salinity stress) and soil (alkali stress)
culture, the performance was evaluated at vegetative
(24 DAS) and maturity (at harvest) stages. The Petri
plate experiment was conducted in a growth chamber
while the sand and soil culture experiments were
conducted in a net house at the Central Soil Salinity
Research Institute (CSSRI), Karnal, Haryana, India,
from June to September, 2008, when natural growing
conditions (temperature 25–35�C, relative humidity
60–82%) are generally favourable for pearl millet.
Petri plate germination and seedling emergence
experiment
Germination and seedling emergence was studied in
Petri plates at four salinity levels (electrical conduc-
tivity of the irrigation water [Eciw] 15, 18, 22 and
26 dS m-1) and control (� strength Hoagland solu-
tion) in four replications at 25�C in a growth
chamber. Twenty-five uniform seeds were placed in
each Petri plate, lined with Whatman filter paper (no.
41), with the addition of 10 ml Hoagland saline
solution. Data on seed germination (measured as
protrusion of 2 mm radical) and seedling emergence
(measured as emergence of pair of first leaves)
percentage was recorded at 24 h intervals over 7
DAS. The saline water for irrigation was prepared by
adding NaCl, Na2SO4 and CaCl2, keeping the Na:Ca
and Cl:SO4 ratio of 4:1 in � strength Hoagland
nutrient solution using distilled water. Shoots of
7-day-old seedlings were separated from roots and
their Na? and K? contents determined by extraction
in 100 mM acetic acid for 2 h at 90�C as described in
Flowers and Yeo (1981).
Sand culture salinity experiment in pots
For studying plant responses at vegetative (24 DAS)
and maturity stages under saline conditions, hybrids
were evaluated with saline irrigation (ECiw 9 and
12 dS m-1) together with the control (� strength
Hoagland solution, ECiw 2 dS m-1) treatment in
ceramic pots of 20 kg capacity filled with sand. The
saline irrigation waters were prepared as described in
the previous section using normal tap water. The
salinity levels for each treatment were maintained in
each pot throughout the experiment by flushing
treatments at appropriate intervals (Sharma and Gill
1992). Fifteen seeds per pot were sown in four
replications. Each pot was thinned to five seedlings at
15 DAS. Two seedlings from each pot were harvested
at 24 DAS. Roots and shoots of these seedlings were
separated for recording fresh and dry weight and for
ionic analysis. The remaining three plants per pot
were grown until maturity. At maturity, these three
plants per pot were harvested and air dried to record
their biomass, stover and grain yield as detailed in
Yadav et al. (2002). Further, the main shoot of each
plant was separated into leaves, nodes and internodes
for ionic analysis using flame spectrophotometery.
Na? and K? contents were determined by wet
digestion method using flame spectrophotometery as
described in Sharma and Kumar (1999).
Soil culture alkalinity experiment in micro plots
In this experiment, the same set of eight pearl millet
hybrids was evaluated under alkali stress in micro
plots (6 9 4 m) filled with two levels of alkali soils
(pH 9.0 and 9.4) as well as reclaimed alkali soil (pH
8.5, used as control), in three replications. The
alkalinity levels were prepared by spraying required
amounts of Na2CO3 and NaHCO3 in different layers
of soil (15 cm each), which were then mixed
thoroughly before filling in micro plots (1 m deep).
Each replication comprised ten plants per row. At 24
DAS, two plants were sampled from each entry
across three replications and their shoot and root fresh
and dry weights and ionic accumulation determined
as described in the previous section. Eight plants per
row were harvested at maturity for recording dry
weight of stover, ear-heads, and grain. Extracts of the
plants harvested at 24 DAS, and at maturity, were
Mol Breeding (2011) 27:207–222 209
123

also subjected to ionic analysis using flame spectro-
photometery.
Statistical analysis
Statistical analysis and analysis of variance was con-
ducted using the statistical program package Indo-Stat
2007. Data were analyzed using salinity, alkalinity and
genotypes as main effects. Genotype 9 salinity and
genotype 9 alkalinity interactions were calculated sep-
arately for salinity and alkalinity experiments.
Results
Analysis of variance conducted on data recorded at
seedling and maturity stages is presented in Tables 1,
2, 3, 4, 5. Salinity and alkalinity stresses significantly
Table 1 Shoot and root dry weight (g per plant) at 24 DAS in hybrids of parental genotypes, and of QTL-NILs, at three salinity and
alkalinity levels
Genotypes Catalogue no. Salinity stress Alkalinity stress
Shoot dry
weight
Root dry
weight
Shoot dry
weight
Root dry
weight
ECiw 2 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 3.14f 0.38ef 1.00l 0.12i
Drought-tolerant parent 843A 9 PRLT 2/89-33 1.85cde 0.15ab 1.05l 0.08h
QTL-NILs 843A 9 ICMR 01029 2.88f 0.34cef 0.98l 0.11i
843A 9 ICMR 01031 2.71ef 0.43f 0.24cdefgh 0.05def
843A 9 ICMR 01040 2.53ef 0.40f 0.15a 0.03abcd
843A 9 ICMR 01046 3.22f 0.38ef 0.17a 0.02abc
843A 9 ICMR 02042 2.64ef 0.33cef 0.44jk 0.06efg
843A 9 ICMR 02044 2.33def 0.17abc 0.39gij 0.07gh
ECiw 9 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 1.21abc 0.12ab 0.08abc 0.01a
Drought-tolerant parent 843A 9 PRLT 2/89-33 0.95abc 0.17ab 0.58k 0.06efg
QTL-NILs 843A 9 ICMR 01029 1.26bc 0.12ab 0.19bcde 0.03abcd
843A 9 ICMR 01031 1.25bc 0.16ab 0.14abcd 0.01ab
843A 9 ICMR 01040 1.51bcd 0.16ab 0.22cde 0.03abcd
843A 9 ICMR 01046 1.11abc 0.15ab 0.24cdefg 0.04cde
843A 9 ICMR 02042 1.84cde 0.23bcde 0.39fghij 0.06gh
843A 9 ICMR 02044 1.54bcd 0.13ab 0.35efghij 0.07fgh
ECiw 12 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 0.29a 0.03a 0.05ab 0.01bcd
Drought-tolerant parent 843A 9 PRLT 2/89-33 1.03abc 0.12ab 0.15abcd 0.02ab
QTL-NILs 843A 9 ICMR 01029 1.48bcd 0.16ab 0.23cdefg 0.02abcd
843A 9 ICMR 01031 0.79ab 0.18abcd 0.15bcde 0.03abcd
843A 9 ICMR 01040 1.29bc 0.18abcd 0.21defghi 0.02abc
843A 9 ICMR 01046 0.78ab 0.10ab 0.15cdef 0.02ab
843A 9 ICMR 02042 0.82ab 0.08ab 0.08abcd 0.02abcd
843A 9 ICMR 02044 0.78ab 0.15ab 0.20bcde 0.02ab
LSD (P = 0.05)
Genotypes (G) 0.34* NS 0.07*** 0.01***
Salinity (S) 0.28*** 0.05*** 0.05*** 0.01***
G 9 S NS NS 0.14*** 0.02***
Means within a column that do not have a common letter are significantly different by LSD0.05 test
210 Mol Breeding (2011) 27:207–222
123

affected all the traits evaluated in this study. The two
parents and six QTL-NILs were not significantly
different for root dry weight at the seedling stage and
for shoot dry weight at maturity harvest under salinity
stress but were significantly different for the rest of
the traits evaluated under salinity and alkalinity stress
conditions. Genotypes 9 salinity treatments (G 9 S)
interactions were significant for all traits except for
shoot and root dry weight at 24 DAS, grain yield at
maturity, and concentration of Na? and K? ion in
roots, internodes and leaves at maturity. Geno-
types 9 alkalinity treatments (G 9 A) interactions
were significant for all traits but at a lower level of
significance (P = 0.05) for grain yield and biomass
yield. These data indicated that salinity and alkali
levels chosen for this study were appropriate for
expression of genotypic differences in parents and
QTL-NILs.
Table 2 Growth, and yield parameters measured at maturity stage in hybrids of parental genotypes and of QTL-NILs, at three
salinity levels
Genotypes Catalogue no. Biomass
yield (g per pot)
Stover
yield (g per pot)
Root dry
weight (g per pot)
Grain
yield (g per pot)
ECiw 2 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 351i 169i 105j 119d
Drought-tolerant parent 843A 9 PRLT 2/89-33 249efg 145gh 41ef 75abc
QTL-NILs 843A 9 ICMR 01029 316h 159hi 68i 107d
843A 9 ICMR 01031 320hi 157hi 52h 109d
843A 9 ICMR 01040 346i 171i 49gh 101cd
843A 9 ICMR 01046 271g 154h 43fg 71ab
843A 9 ICMR 02042 298h 146gh 48gh 101cd
843A 9 ICMR 02044 253fg 135fg 41ef 81bc
ECiw 9 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 210cd 100abcd 36cde 71ab
Drought-tolerant parent 843A 9 PRLT 2/89-33 225def 115de 33bcd 81bc
QTL-NILs 843A 9 ICMR 01029 249efg 128ef 48gh 78abc
843A 9 ICMR 01031 222de 111bcd 36de 81bc
843A 9 ICMR 01040 211cd 101abcd 34bcd 78abc
843A 9 ICMR 01046 176ab 100abcd 35bcd 53a
843A 9 ICMR 02042 212cd 113cd 36cde 79abc
843A 9 ICMR 02044 205bcd 115de 33bcd 64ab
ECiw 12 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 195abcd 95a 34bcd 72ab
Drought-tolerant parent 843A 9 PRLT 2/89-33 209cd 107abcd 32bcd 62ab
QTL-NILs 843A 9 ICMR 01029 194abcd 99abc 32bcd 67ab
843A 9 ICMR 01031 173a 94a 27a 57ab
843A 9 ICMR 01040 188abc 98abc 36cde 64ab
843A 9 ICMR 01046 176ab 95ab 31abcd 57ab
843A 9 ICMR 02042 184abc 93a 30abc 68ab
843A 9 ICMR 02044 195abcd 104abcd 30ab 66ab
LSD (P = 0.05)
Genotypes (G) 19*** NS 3*** 12***
Salinity (S) 9*** 5*** 2*** 8***
G 9 S 26*** 13*** 5*** NS
Means within a column that do not have a common letter are significantly different by LSD0.05 test
Mol Breeding (2011) 27:207–222 211
123
Growth and ionic accumulation at seedling
and vegetative stage
Seedling stage (7 DAS)
Germination as well as seedling emergence declined
with increase in salinity in all the eight test-cross hybrids
evaluated. At ECiw 26 dS m-1, seedling germination
declined by 60% in the drought-sensitive recurrent
parent H 77/833-2 and 40% in the hybrid of the drought-
tolerant parent PRLT 2/89-33 compared to control
treatment (data not shown). Mean seedling emergence
across hybrids declined by 19 and 66% at ECiw
15 dS m-1 and ECiw 26 dS m-1, respectively, in the
Petri plate experiment, compared to control (Fig. 1A).
At the highest salinity level of ECiw 26 dS m-1, an 80%
reduction in seedling emergence was recorded in the
hybrid of the drought-sensitive recurrent parent H 77/
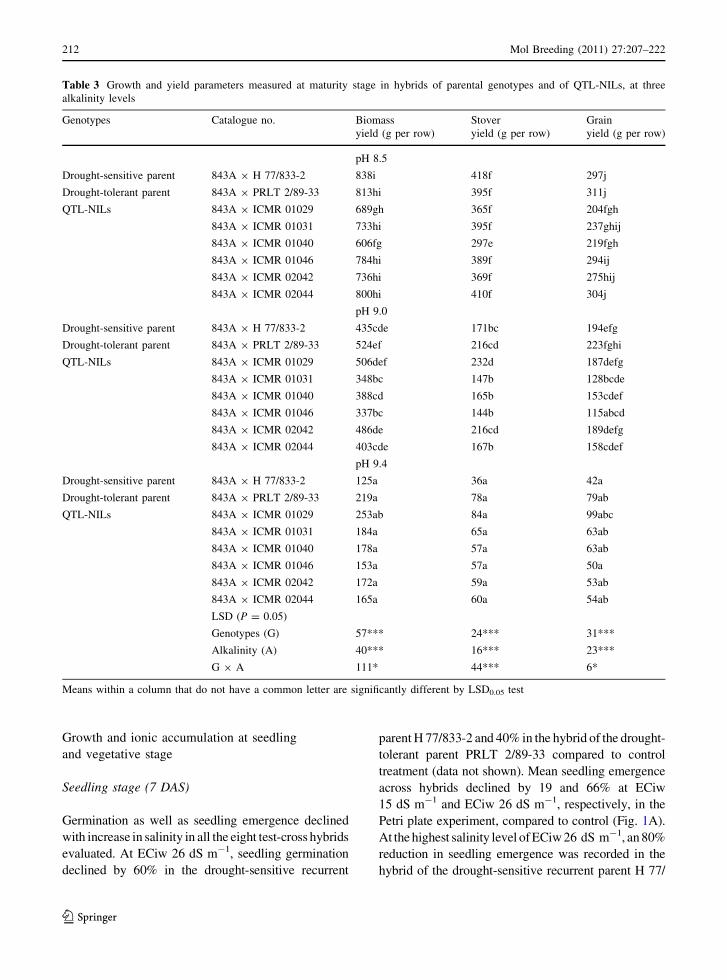
Table 3 Growth and yield parameters measured at maturity stage in hybrids of parental genotypes and of QTL-NILs, at three
alkalinity levels
Genotypes Catalogue no. Biomass
yield (g per row)
Stover
yield (g per row)
Grain
yield (g per row)
pH 8.5
Drought-sensitive parent 843A 9 H 77/833-2 838i 418f 297j
Drought-tolerant parent 843A 9 PRLT 2/89-33 813hi 395f 311j
QTL-NILs 843A 9 ICMR 01029 689gh 365f 204fgh
843A 9 ICMR 01031 733hi 395f 237ghij
843A 9 ICMR 01040 606fg 297e 219fgh
843A 9 ICMR 01046 784hi 389f 294ij
843A 9 ICMR 02042 736hi 369f 275hij
843A 9 ICMR 02044 800hi 410f 304j
pH 9.0
Drought-sensitive parent 843A 9 H 77/833-2 435cde 171bc 194efg
Drought-tolerant parent 843A 9 PRLT 2/89-33 524ef 216cd 223fghi
QTL-NILs 843A 9 ICMR 01029 506def 232d 187defg
843A 9 ICMR 01031 348bc 147b 128bcde
843A 9 ICMR 01040 388cd 165b 153cdef
843A 9 ICMR 01046 337bc 144b 115abcd
843A 9 ICMR 02042 486de 216cd 189defg
843A 9 ICMR 02044 403cde 167b 158cdef
pH 9.4
Drought-sensitive parent 843A 9 H 77/833-2 125a 36a 42a
Drought-tolerant parent 843A 9 PRLT 2/89-33 219a 78a 79ab
QTL-NILs 843A 9 ICMR 01029 253ab 84a 99abc
843A 9 ICMR 01031 184a 65a 63ab
843A 9 ICMR 01040 178a 57a 63ab
843A 9 ICMR 01046 153a 57a 50a
843A 9 ICMR 02042 172a 59a 53ab
843A 9 ICMR 02044 165a 60a 54ab
LSD (P = 0.05)
Genotypes (G) 57*** 24*** 31***
Alkalinity (A) 40*** 16*** 23***
G 9 A 111* 44*** 6*
Means within a column that do not have a common letter are significantly different by LSD0.05 test
212 Mol Breeding (2011) 27:207–222
123
833-2, compared to 50% in the drought-tolerant parent
PRLT 2/89-33. Similarly, most QTL-NILs (except for
ICMR 01046 and ICMR 02042) showed lower reduc-
tions in seedling emergence compared to the drought-
sensitive parent H 77/833-2, largely on the pattern of the
drought-tolerant parent PRLT 2/89-33.
Na?/K? ratios (on a molar basis) in shoots were
higher in the sensitive parent H 77/833-2 and lower in
the tolerant parent PRLT 2/89-33 and QTL-NILs
(Fig. 1B).
Vegetative stage (24 DAS)
Shoot and root dry weights of seedlings at 24 DAS
for the two parents and six QTL-NILs under different
salinity and alkalinity stress levels are given in
Table 1. In the control treatment of the sand-culture
experiment (ECiw 2.0 dS m-1), the drought-sensitive
parent (H 77/833-2) produced greater shoot dry
weight compared to the drought-tolerant parent
(PRLT 2/89-33). More specifically, the shoot dry
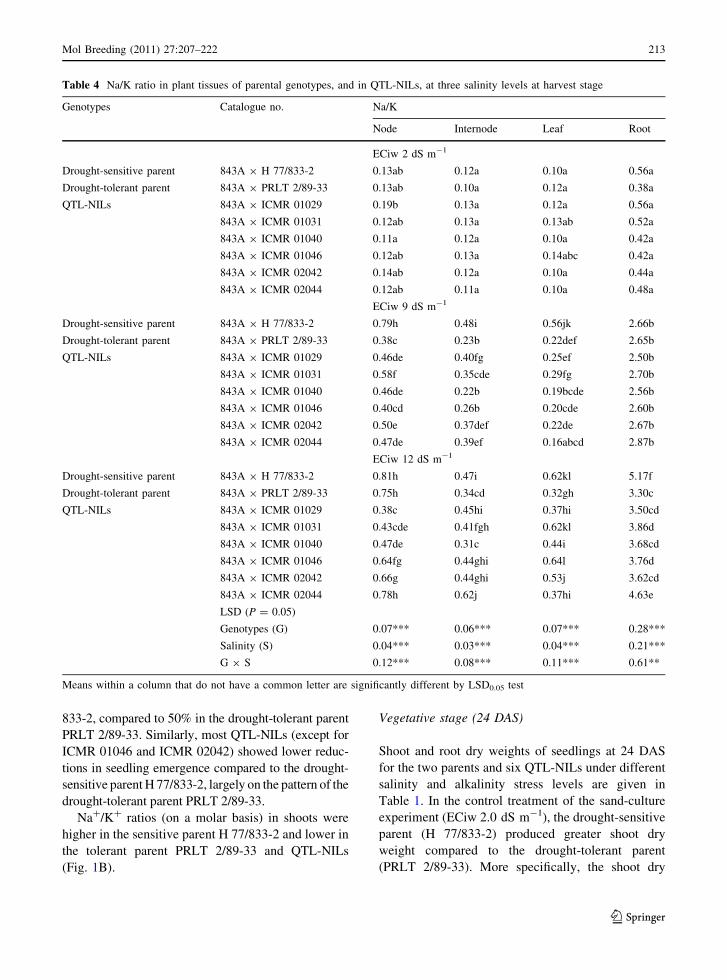
Table 4 Na/K ratio in plant tissues of parental genotypes, and in QTL-NILs, at three salinity levels at harvest stage
Genotypes Catalogue no. Na/K
Node Internode Leaf Root
ECiw 2 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 0.13ab 0.12a 0.10a 0.56a
Drought-tolerant parent 843A 9 PRLT 2/89-33 0.13ab 0.10a 0.12a 0.38a
QTL-NILs 843A 9 ICMR 01029 0.19b 0.13a 0.12a 0.56a
843A 9 ICMR 01031 0.12ab 0.13a 0.13ab 0.52a
843A 9 ICMR 01040 0.11a 0.12a 0.10a 0.42a
843A 9 ICMR 01046 0.12ab 0.13a 0.14abc 0.42a
843A 9 ICMR 02042 0.14ab 0.12a 0.10a 0.44a
843A 9 ICMR 02044 0.12ab 0.11a 0.10a 0.48a
ECiw 9 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 0.79h 0.48i 0.56jk 2.66b
Drought-tolerant parent 843A 9 PRLT 2/89-33 0.38c 0.23b 0.22def 2.65b
QTL-NILs 843A 9 ICMR 01029 0.46de 0.40fg 0.25ef 2.50b
843A 9 ICMR 01031 0.58f 0.35cde 0.29fg 2.70b
843A 9 ICMR 01040 0.46de 0.22b 0.19bcde 2.56b
843A 9 ICMR 01046 0.40cd 0.26b 0.20cde 2.60b
843A 9 ICMR 02042 0.50e 0.37def 0.22de 2.67b
843A 9 ICMR 02044 0.47de 0.39ef 0.16abcd 2.87b
ECiw 12 dS m-1
Drought-sensitive parent 843A 9 H 77/833-2 0.81h 0.47i 0.62kl 5.17f
Drought-tolerant parent 843A 9 PRLT 2/89-33 0.75h 0.34cd 0.32gh 3.30c
QTL-NILs 843A 9 ICMR 01029 0.38c 0.45hi 0.37hi 3.50cd
843A 9 ICMR 01031 0.43cde 0.41fgh 0.62kl 3.86d
843A 9 ICMR 01040 0.47de 0.31c 0.44i 3.68cd
843A 9 ICMR 01046 0.64fg 0.44ghi 0.64l 3.76d
843A 9 ICMR 02042 0.66g 0.44ghi 0.53j 3.62cd
843A 9 ICMR 02044 0.78h 0.62j 0.37hi 4.63e
LSD (P = 0.05)
Genotypes (G) 0.07*** 0.06*** 0.07*** 0.28***
Salinity (S) 0.04*** 0.03*** 0.04*** 0.21***
G 9 S 0.12*** 0.08*** 0.11*** 0.61**
Means within a column that do not have a common letter are significantly different by LSD0.05 test
Mol Breeding (2011) 27:207–222 213
123

weight of the drought-sensitive parent (H 77/833-2)
was 1.7 and 1.3 times higher than that of the drought-
tolerant parent (PRLT 2/89-33) in the control treat-
ment and salinity treatment of ECiw 9 dS m-1.
However, in the salinity treatment of ECiw
12 dS m-1, the trend was opposite. In this treatment,
the hybrid of the drought-tolerant parent (PRLT 2/89-
33) produced much greater shoot dry weight (3.5
times) than the sensitive-parent (H 77/833-2). Fur-
thermore, the QTL-NILs also produced much greater
shoot dry weight (3.3 times on average) compared to
the drought-sensitive parent at this salinity treatment
of ECiw 12 dS m-1. Among the QTL-NILs, ICMR
01029 and ICMR 01040 produced the highest shoot
dry weight at 24 DAS, even higher than the drought-
tolerant parent PRLT 2/89-33, at the highest salinity
level of ECiw 12 dS m-1. The maximum decline
(90% compared to controls) in shoot dry weight was
recorded in the drought-sensitive parent H 77/833-2
at this salinity treatment of ECiw 12 dS m-1. At
Table 5 Na/K ratio in plant tissues of parental genotypes, and in QTL-NILs, at three alkalinity levels at maturity stage
Genotypes Catalogue no. Na/K
Node Internode Leaf
pH 8.5
Drought-sensitive parent 843A 9 H 77/833-2 0.26a 0.25ab 0.47de
Drought-tolerant parent 843A 9 PRLT 2/89-33 0.31a 0.13a 0.26ab
QTL-NILs 843A 9 ICMR 01029 0.35a 0.26ab 0.25a
843A 9 ICMR 01031 0.26a 0.34ab 0.35abc
843A 9 ICMR 01040 0.32a 0.31ab 0.28ab
843A 9 ICMR 01046 0.28a 0.25ab 0.27ab
843A 9 ICMR 02042 0.40a 0.38b 0.24a
843A 9 ICMR 02044 0.35a 0.28ab 0.27ab
pH 9.0
Drought-sensitive parent 843A 9 H 77/833-2 2.62efg 1.34ef 0.96jk
Drought-tolerant parent 843A 9 PRLT 2/89-33 1.88c 0.44b 0.62fg
QTL-NILs 843A 9 ICMR 01029 2.21cde 1.14de 0.68gh
843A 9 ICMR 01031 2.71fgh 2.35h 0.67gh
843A 9 ICMR 01040 1.28b 0.75c 0.37bcd
843A 9 ICMR 01046 2.03cd 2.01g 0.60fg
843A 9 ICMR 02042 3.14hij 1.95g 0.37bcd
843A 9 ICMR 02044 2.38def 2.09g 0.40cd
pH 9.4
Drought-sensitive parent 843A 9 H 77/833-2 0.81m 3.73l 1.88l
Drought-tolerant parent 843A 9 PRLT 2/89-33 0.75kl 1.03d 0.85ij
QTL-NILs 843A 9 ICMR 01029 0.38kl 3.31k 1.03k
843A 9 ICMR 01031 0.43ijk 2.94j 0.77hi
843A 9 ICMR 01040 0.47ghi 1.40f 0.55ef
843A 9 ICMR 01046 0.64jk 2.61i 0.81i
843A 9 ICMR 02042 0.66l 2.43hi 0.66fgh
843A 9 ICMR 02044 0.78hij 2.49hi 0.88ij
LSD (P = 0.05)
Genotypes (G) 0.43*** 0.20*** 0.11***
Alkalinity (A) 0.25*** 0.13*** 0.06***
G 9 A 0.71* 0.37*** 0.18***
Means within a column that do not have a common letter are significantly different by LSD0.05 test
214 Mol Breeding (2011) 27:207–222
123

ECiw 12 dS m-1, the decline in shoot dry weight in
hybrids of the drought-tolerant parent PRLT 2/89-33
and QTL-NILs (e.g. ICMR 01029 and ICMR 01040)
was much less (44–48%) compared to the drought-
sensitive parent H 77/833-2. Furthermore, under
alkali stress, the mean shoot dry weight of the eight
hybrids declined by 47 and 65% at pH 9.0 and 9.4,
respectively, compared to their control plants. No
significant differences were recorded in root dry
weight of parents and QTL-NILs in salinity treat-
ments conducted under sand culture conditions at 24
DAS.
In the alkalinity experiment, among all the geno-
types evaluated, the highest shoot (1.0 g per plant)
and root (0.12 g per plant) dry weights were recorded
in the drought-sensitive parent (H 77/833-2) under
control conditions. However, shoot and root dry
weight reductions in the drought-sensitive parent
were much greater in alkali stress treatments com-
pared to the hybrids of the drought-tolerant parent
and QTL-NILs. Compared to control conditions, the
reduction in shoot dry weight in the drought-sensitive
parent (92%) was significantly higher than of the
drought-tolerant parent (45%) at pH 9.0. Moreover, at
pH 9.0, shoot dry weight in QTL-NIL ICMR 01029
was almost three times greater than the drought-
sensitive parent H 77/833-2. No significant differ-
ences were recorded in root dry weight of parents and
QTL-NILs in salinity treatments conducted under
sand culture conditions at 24 DAS.
Growth and ionic accumulation at maturity
Stover, grain and biomass yield
Total biomass (sum total of stover and grain yield)
accumulated by the parental hybrids and QTL-NILs
in salinity and alkalinity experiments are presented in
Tables 2 and 3. Under salinity stress, plant biomass
yield averaged across all hybrids declined by about
-20
0
20
40
60
80
100
120
843AxH 77/833-2 843A x ICMR 1029 843A x ICMR 1031 843A x ICMR 1040 843A x ICMR 1046 843A x ICMR 2042 843A x ICMR 2044 843A x PRLT 2/89-33
See
dlin
g e
mer
gen
ce (
%) 2 dS/m 15 dS/m 18 dS/m 22 dS/m 26 dS/m
A
0
1
2
3
4
5
2 15 18 22 26
Na/
K in
See
dlin
g S
ho
ot
Salinity levels (ECiw, dS/m)
843AxH 77/833-2843A x ICMR 1029843A x ICMR 1031843A x ICMR 1040843A x ICMR 1046843A x ICMR 2042843A x ICMR 2044843A x PRLT 2/89-33
B
Fig. 1 Seedling emergence percentage (A) and shoot Na/K
accumulation (B) recorded at 7 DAS in test-cross hybrids of
parental genotypes (H 77/833-2, PRLT 2/89-33), and of QTL-
NILs (ICMR 01029, ICMR 01031, ICMR 1040, ICMR 1046,
ICMR 02042 and ICMR 02044) differing by a terminal
drought tolerance QTL, at five salinity levels (ECiw 2, 15,
18, 22 and 26 dS m-1)
Mol Breeding (2011) 27:207–222 215
123

28 and 36% at salinity levels of ECiw 9 and
12 dS m-1, respectively, compared to plants growing
under the control treatment (ECiw 2 dS m-1). At
ECiw 2 dS m-1, the drought-sensitive parent (H 77/
833-2) produced the highest plant biomass (350 g per
pot) whereas the drought-tolerant parent (PRLT 2/89-
33) produced the least biomass (249 g per pot). As at
the vegetative stage, this trend was reversed in the
salinity treatments of ECiw 9.0 and 12.0 dS m-1. In
both these salinity treatments, the drought-tolerant
parent (PRLT 2/89-33) produced higher biomass than
the drought-sensitive parent (H 77/833-2) (Table 2).
Moreover, compared to control experiment (ECiw
2.0 dS m-1), the percentage reduction in plant bio-
mass was minimal in the drought-tolerant parent (10
and 16% at ECiw 9 and 12 dS m-1, respectively) and
maximal in the drought-sensitive parent (40 and 44%
at ECiw 9 and 12 dS m-1, respectively). Further-
more, hybrids of QTL-NILs also showed lower
percentage reductions (19–39% at ECiw 9 dS m-1
with an average of 28.9%, and 23–45% at ECiw
12 dS m-1 with an average of 37.6%) in plant
biomass compared to the sensitive parent. At ECiw
9 dS m-1, hybrids of genotypes ICMR 01029 and
ICMR 02044 showed least reduction in biomass (21
and 19% reduction, respectively) while hybrids of
genotypes ICMR 01040 and ICMR 01046 showed
biomass reductions (39 and 35%, respectively)
closer to hybrids of the drought-sensitive parent H
77/833-2.
In the control treatment of the alkalinity experi-
ment (reclaimed alkali soils, pH 8.5), again the hybrid
of the drought-sensitive parent (H 77/833-2) pro-
duced the highest biomass (838 g per row) among all
the eight (six QTL NILs and two parents) hybrids
tested. However, in alkali treatments (pH 9.0 and
9.4), it was the drought-tolerant parent (PRLT 2/89-
33) that produced the greatest biomass yield and the
least percentage reductions (73% in PRLT 2/89-33
compared to 85% in H 77/833-2 at pH 9.4) compared
to the control treatment (Table 3). Furthermore, the
QTL-NIL ICMR 01029 produced almost double the
biomass yield at pH 9.4 compared to the drought-
sensitive parent H 77/833-2. The hybrid of QTL-NIL
ICMR 01029 also produced around 15% higher
biomass yield over the hybrid of the drought-tolerant
parent PRLT 2/89-33.
Reductions of 21 and 26% in stover yield was
recorded in the drought-tolerant parent (PRLT 2/89-
33) compared to 41 and 44% in the drought-sensitive
parent (H 77/833-2) at ECiw 9 and 12 dS m-1,
respectively (Table 2). QTL-NILs also showed less
reduction in stover yield than the drought-sensitive
parent H 77/833-2 under higher salinity treatments.
Stover yield was also significantly affected by
alkalinity and a 52% mean reduction at pH 9.0 and
83% at pH 9.4 was recorded compared to the control
treatment (pH 8.5) (Table 3). Amongst the QTL-
NILs, the best performance was observed in QTL-
NIL ICMR 01029, which showed only 36% reduction
in stover yield at pH 9.0 and 76% at pH 9.4 compared
to 59 and 91%, respectively, in the drought-sensitive
parent H 77/833-2. QTL-NILs ICMR 01040 and
ICMR 02042 also showed lower percentage reduction
in stover yield (44 and 41% at pH 9.0, respectively)
compared to control plants.
At ECiw 9 dS m-1, QTL-NIL ICMR 01029
showed a grain yield reduction of 26% compared to
the drought-sensitive parent (H 77/833-2) which
showed a 40% reduction in grain yield. Under
alkalinity stress, similar results were recorded in
plant grain yield (Table 2). Amongst the QTL-NILs,
ICMR 01046 was recorded as having the maximum
reduction of 61% in plant grain yield at pH 9.0
compared to control plants.
Ionic accumulation at maturity
Data on Na? accumulation recorded in various plant
parts (root, leaf, node, internodes) of the hybrids of
parental genotypes, and of the QTL-NILs, are
presented in Figs. 2, 3, 4. In general, nodes retained
the highest Na? concentration in the shoot of the
eight hybrids followed by roots, internodes and
leaves (Fig. 2). In the control treatments (ECiw
2 dS m-1), Na? concentration in these parts was
similar in the hybrids of the two parental genotypes.
However, at ECiw 9.0 dS m-1, the drought-sensitive
parent (H 77/833-2) accumulated almost twice
(3.06 mmol g-1 dry weight) the concentration of
Na?, considering all parts together, compared to the
drought-tolerant parent PRLT 2/89-33 (1.72 mmol
g-1 dry weight). Notably, at the highest salinity level
of ECiw 12 dS m-1, the two parental genotypes (H
77/833-2 and PRLT 2/89-33) accumulated similar
Na? concentration in roots, nodes and internodes but
in leaves the drought-tolerant parent (PRLT 2/89-33)
accumulated less than half the concentration of Na?
216 Mol Breeding (2011) 27:207–222
123

(Fig. 2) compared to the hybrid of the drought-
sensitive parent (H 77/833-2).
Accumulation of Na? in the hybrids of QTL-NILs
was essentially similar to the hybrid of the drought-
tolerant parent PRLT 2/89-33 in both saline and
alkaline treatments. In salinity treatments, the amount
of Na? increased in all shoot parts of QTL-NILs but
this increase was greater in nodes and internodes than
in leaves. At the salinity level of ECiw 9 dS m-1, the
amount of Na? in leaves of QTL-NILs was, on
average, half that in the drought-sensitive parent H
77/833-2. Further, the leaf Na? in QTL-NILs was, on
average, 3–4 times lower compared to nodes and
internodes in the drought-sensitive parent. In alkali
stress treatments, the drought-sensitive parent
accumulated double the amount of Na? in internodes
(pH 9.0 and 9.4) and leaves (pH 9.4) in comparison to
the drought-tolerant parent (Fig. 3). The increment in
Na? accumulation in nodes was similar in the two
parental genotypes even at the highest alkali level of
pH 9.4. All QTL-NILs accumulated less Na? in
leaves, invariably 3–4 times less than nodes and
internodes. Furthermore, the mean of six QTL-NILs
hybrids accumulated Na? in leaves 2.3 times less
than the drought-sensitive parent (Fig. 3) at the
highest alkali stress level of pH 9.4.
K? accumulation generally declined with increase
in salinity and alkalinity stress in all plant parts. Total
K? content was similar in hybrids of both the parental
genotypes in control conditions (ECiw 2 dS m-1, pH
0.0
0.5
1.0
1.5
843AxH 77/833-2 843A x ICMR 1029 843A x ICMR 1031 843A x ICMR 1040 843A x ICMR 1046 843A x ICMR 2042 843A x ICMR 2044 843A x PRLT 2/89-33No
de
Na
(mm
ol/g
dry
wt)
Node A
0.0
0.5
1.0
1.5
843AxH 77/833-2 843A x ICMR 1029 843A x ICMR 1031 843A x ICMR 1040 843A x ICMR 1046 843A x ICMR 2042 843A x ICMR 2044 843A x PRLT 2/89-33
Inte
rno
de
Na
(mm
ol/g
d
ry w
t)
Internode B
0.0
0.5
1.0
1.5
843AxH 77/833-2 843A x ICMR 1029 843A x ICMR 1031 843A x ICMR 1040 843A x ICMR 1046 843A x ICMR 2042 843A x ICMR 2044 843A x PRLT 2/89-33Lea
f N
a (m
mo
l/g d
ry w
t)
LeavesC
0.0
0.5
1.0
1.5
843AxH 77/833-2 843A x ICMR 1029 843A x ICMR 1031 843A x ICMR 1040 843A x ICMR 1046 843A x ICMR 2042 843A x ICMR 2044 843A x PRLT 2/89-33
Ro
ot
Na
(mm
ol/g
d
ry w
t)
2 dS/m 9 dS/m 12 dS/m RootsD
Fig. 2 Na concentration (mmol g-1 dry weight) in nodes (A),
internodes (B), leaves (C) and roots (D) of pearl millet hybrids
on parental genotypes (H 77/833-2, PRLT 2/89-33), and of
QTL-NILs (ICMR 01029, ICMR 01031, ICMR 1040, ICMR
1046, ICMR 02042 and ICMR 02044) differing by a terminal
drought tolerance QTL, at three salinity levels (ECiw 2, 9 and
12 dS m-1)
Mol Breeding (2011) 27:207–222 217
123

8.5). However, it declined sharply in hybrid of the
drought-sensitive parent H 77/833-2, both with
increase in saline or alkaline stress compared to
hybrid of the drought-tolerant parent PRLT 2/89-33
(data not given). Amongst different plant parts,
nodes (1.48 mmol g-1 dry weight) and internodes
(1.51 mmol g-1 dry weight) accumulated higher
K? under salinity stress compared to leaves
(1.30 mmol g-1 dry weight), whereas the least accu-
mulation was recorded in roots (0.45 mmol g-1 dry
weight). Moreover, the differences in K? accumula-
tion in leaves were small and were not significantly
different among the eight test-cross hybrids of parents
and QTL-NILs in salinity treatments. Furthermore,
under alkalinity stress treatment of pH 9.0 of the
drought-tolerant parent hybrid, a higher K?
concentration was recorded in nodes (0.51 mmol g-1
dry weight) and internodes (0.77 mmol g-1 dry
weight) than in the drought-sensitive parent hybrid
(0.34 and 0.62 mmol g-1 dry weight in nodes and
internodes, respectively). A similar pattern was
evident for these plant parts at higher alkali stress
level of pH 9.4. Further, the K? content declined by
52% in the drought-sensitive parent hybrid H 77/833-
2 at pH 9.4 compared to 37% in the drought-tolerant
parent hybrid PRLT 2/89-33. A lower K? content
was recorded in different plant parts of the QTL-NILs
hybrids, ICMR 01029 and ICMR 01031, compared to
the other four QTL-NILs hybrids. Notably, in both
the salinity as well as in alkalinity treatments, K?
accumulation in QTL-NILs was not different from
either parent, unlike Na? accumulation.
Fig. 3 Na concentration (mmol g-1 dry weight) in nodes (A),
internodes (B) and leaves (C) of pearl millet hybrids on
parental genotypes (H 77/833-2, PRLT 2/89-33), and of QTL-
NILs (ICMR 01029, ICMR 01031, ICMR 1040, ICMR 1046,
ICMR 02042 and ICMR 02044) differing by a terminal
drought tolerance QTL, at three alkalinity levels (pH 8.5, 9.0
and 9.4)
218 Mol Breeding (2011) 27:207–222
123

Under salinity stress, on a mean basis, the highest
Na/K ratio was recorded in roots (2.35), and was
roughly five times higher than in nodes (0.42),
internodes (0.30) and leaves (0.29) (Table 4). How-
ever, under alkali stress (Table 5), the highest Na/K
ratio was recorded in nodes (2.06) compared to
internodes (1.42) and leaves (0.60). Averaged over
different plant parts, the drought-sensitive parent (H
77/833-2) hybrid had a higher Na/K ratio (1.76) at
ECiw 12 dS m-1 compared to drought-tolerant par-
ent hybrid (1.17). The mean Na/K ratio in six QTL-
NILs hybrids was 1.33 at this salinity level. Similarly,
under alkalinity (pH 9.4), the drought-sensitive parent
(H 77/833-2) showed a higher Na/K ratio (3.37)
compared to the drought-tolerant parent (PRLT 2/89-
33) hybrid (1.84), when averaged over different plant
parts. The mean Na/K ratio in the six QTL-NILs
hybrids was 2.24 at pH 9.4. Tissue Na/K ratios in
salinity treatments were higher in all plant parts of the
drought-sensitive parent (H 77/833-2) than the
drought-tolerant parent (PRLT 2/89-33) (Fig. 4).
Further, on averaging over alkali treatments, the
hybrid of the drought-sensitive parent (H 77/833-2)
recorded a higher Na/K ratio in nodes (2.46),
internodes (1.77) and leaves (1.10) compared to the
drought-tolerant parent (PRLT 2/89-33) as well as all
the QTL-NILs. At pH 9.4, nodes of the drought-
tolerant parent hybrid showed a lower Na/K ratio
(3.66) compared to the drought-sensitive parent
hybrid (4.51), ranging from 2.93 (ICMR 1040) to
3.92 (ICMR 2042) in QTL-NILs. Further, internodes
of the drought-sensitive parent hybrid recorded more
than three times the Na/K ratio (3.73) of the drought-
tolerant parent hybrid (PRLT 2/89-33) (1.03). Among
the QTL-NILs, the Na/K ratio ranged from 1.40
(ICMR 1040) to 3.31 (ICMR 1029).
Discussion
The aim of this study was to assess the performance
of a terminal drought tolerance QTL under salt stress.
To accomplish that, we evaluated hybrids of the QTL
donor and recipient parental genotypes and QTL-
NILs to which different proportions of the drought
tolerance QTL region were introgressed from PRLT
0.0
0.2
0.4
0.6
0.8
1.0
2 9 12 2 9 12 2 9 12
Na/
K
Salinity levels (dS/m)
843A x PRLT 2/89-33
843A x H 77/833-2
QTL-NILs
Node
InternodeLeaf
A
0.0
1.0
2.0
3.0
4.0
5.0
8.5 9.0 9.4 8.5 9.0 9.4 8.5 9.0 9.4
Na/
K
Alkalinity levels (pH)
843A x PRLT 2/89-33
843A x H 77/833-2
QTL-NILsNode
InternodeLeaf
B
Fig. 4 Na/K ratio in nodes, internodes and leaves of hybrids of parental genotypes and of QTL-NILs (values presented as average of
six NILs) at three salinity (ECiw 2, 9 and 12 dS m-1) (A) and alkalinity (pH 8.5, 9.0 and 9.4) levels (B)
Mol Breeding (2011) 27:207–222 219
123

2/89-33 parent into H 77/833-2, using markers
bracketing the QTL region (Bidinger et al. 2007;
Serraj et al. 2005; Yadav et al. 2002, 2004). These
genotypes were evaluated for growth and productiv-
ity traits under a range of salinity and alkalinity stress
treatments, as well as for ionic (Na? and K?)
accumulation and their compartmentalization in dif-
ferent plant parts. QTL-NILs were preferred for this
study because they offer a unique opportunity to
focus the analysis on the effects of a specific genomic
region containing the QTL in an otherwise uniform
genetic background (Giuliani et al. 2005). These
genotypes were evaluated as test-cross hybrids by
crossing individual QTL-NILs to a common male
sterile line tester (843-A) rather than inbred lines per
se, to restore heterotic vigor in parental and QTL-
NILs inbred lines that otherwise were too weak for
effective screening under salt stress conditions
(Yadav et al. 2002).
The differential behavior in growth and produc-
tivity parameters of hybrids of the drought-tolerant
parent, drought-sensitive parent, and QTL-NILs was
evident right from the germination and seedling
emergence stage through to maturity under both
salinity and alkalinity treatments. Differences in
seedling emergence in parental genotypes and in
QTL-NILs were insignificant until the salinity levels
reached 22 dS m-1. The differences were, however,
much more conspicuous at the salinity level of
26 dS m-1. In the drought-sensitive parent an 80%
reduction in seedling emergence was recorded at this
salinity level compared to 50% in the drought-
tolerant parent PRLT 2/89-33, and about 41% on
average in QTL-NILs (Fig. 1A). A similar trend was
observed for growth parameters at the vegetative
stage (24 DAS), where greater shoot growth was
recorded in the drought-tolerant parent and the QTL-
NILs than the drought-sensitive parent under both
salinity and alkalinity conditions. Similarly, at matu-
rity, under the highest salinity level (ECiw
12 dS m-1), stover yield and total biomass yield
were reduced by 26 and 16%, respectively, in the
drought-tolerant parent PRLT 2/89-33 compared
to 43 and 44% in the drought-sensitive parent H 77/
833-2.
Ionic accumulation and compartmentalization in
different plant parts at both seedling (Fig. 1B) and
maturity stages (Figs. 2, 3, 4) correlated with growth
and yield parameters in parents and QTL-NILs. The
drought-sensitive parent H 77/833-2 accumulated
significantly higher concentrations of toxic Na? in
shoot parts, irrespective of the growth stage, than the
drought-tolerant parent under both salt stress treat-
ments. It is worth noting that the difference in Na?
accumulation was evident only in leaves and not in
the rest of the shoot parts (i.e. nodes, internodes) of
the parental genotypes and QTL-NILs (Figs. 2, 3).
For example, at the highest salinity level (ECiw
12 dS m-1), the drought-sensitive parent accumu-
lated more than twice the concentration of Na? in
leaves compared to the drought-tolerant parent. As
for growth parameters, the QTL-NILs showed a
pattern of salt accumulation largely similar to that of
the drought-tolerant parent (PRLT 2/89-33) from
which this DT-QTL was introgressed. A lower
accumulation of Na? in leaves is a well-known
phenomenon associated with salt tolerance in pearl
millet (Krishnamurthy et al. 2007; Sharma and Gill
1992). The relationship between salt tolerance and
Na? concentration in leaves is also a well-known
mechanism reported in many plant species and has
shown promise as a selection tool for salinity
tolerance (Akita and Cabusaly 1990; Ashraf and
Khanum 1997; Ashraf and O’Leary 1996; Chippa and
Lal 1995; Lauchli 1984; Munns and James 2003;
Poustini and Siosemardeh 2004; Yeo and Flowers
1984; Yeo 1992; Zhu et al. 2004). In durum wheat,
for instance, genotypes with lower Na? concentration
in leaves yielded higher shoot biomass yield after
growing for 4 weeks in saline culture solution
(Munns and James 2003), and higher grain yield
after growing for 4 months in saline soil (Hussain
et al. 2003).
The other finding of the present study was that
Na? accumulation from root to shoot increased at a
higher rate in the drought-susceptible parent H 77/
833-2 (68.6%) compared to the drought-tolerant
parent PRLT 2/89-33 and the QTL-NILs (which on
average showed an increase by 37.3%) at the higher
salinity treatment. The well-documented differences
in salt tolerance between durum and bread wheat is
reported to be related to low rates of Na? transport
into shoot parts from roots (Gorhom et al. 1990;
Hussain et al. 2004; Munns et al. 2003). Interestingly,
we also found that Na? transport was independent of
K? transport in our study, as the hybrids of the two
parental genotypes had similar K? concentration in
their shoot in control as well as in the highest salinity
220 Mol Breeding (2011) 27:207–222
123

and alkalinity treatments. Furthermore, the pattern
and the concentration of K? in QTL-NILs was
dissimilar to that of either parent, suggesting that this
DT-QTL does not affect K? transport. Similar results
(that Na? accumulation is independent of K? accu-
mulation) have been reported in other plant species
(Flowers and Yeo 1981; Hussain et al. 2004; Munns
2002) including pearl millet (Krishnamurthy et al.
2007). From these data it can be concluded unam-
biguously that lower transport of Na? from roots to
shoots, together with preferential compartmentaliza-
tion of Na? in nodes and internodes leading to lower
Na? accumulation in leaves, is the mechanism
operating under salt stress, associated with this
drought tolerance QTL. This mechanism is in agree-
ment to what is known in pearl millet and other
species but different to what is reported in sorghum
where salt tolerance was largely dependent on
the capacity of roots to retain Na? (Netondo et al.
2004).
It is also worth highlighting that although the
performance of most of the QTL-NILs hybrids
analyzed in this study was largely similar to the
donor parent under salt stresses, not all QTL-NILs
behaved similarly. For example, ICMR 01046 and
ICMR 02042 recorded higher reductions in seed-
ling emergence (on the pattern of drought-sensitive
parent H 77/833-2) in the Petri plate experiment.
Similarly, higher reductions in biomass and shoot
weight under higher alkalinity stress (pH 9.4) were
recorded for ICMR 01046 and ICMR 02044. Also,
plant head weight showed maximal reductions in
ICMR 01031 and 01040, and ICMR 01046 and
02044, under highest salinity (ECiw 12 dS m-1)
and alkalinity (pH 9.4) treatments, respectively.
This differential response among QTL-NILs
hybrids could be due to the variable size of the
donor segment around the DT QTL on LG 2 that
has been introgressed into the QTL-NILs evaluated
in the study. Serraj et al. (2005) also reported
previously that all QTL-NILs did not behave
uniformly under terminal drought stress conditions.
ICMR 01029 and 01031 were found to be superior
in their study for yield maintenance while ICMR
02042 had a yield response more like H 77/833-2
and ICMR 02044 had an intermediate yield
response under terminal drought stress conditions.
The differential response of QTL-NILs emphasizes
the need for fine mapping of this important QTL
interval on LG 2. Efforts in this direction are
currently underway in our laboratory, in which a
high-resolution cross segregating only for the DT
QTL interval on LG 2 has been advanced using
QTL-NIL ICMR 01029 and H 77/833-2 as parental
genotypes. This highly specialized cross will not
only generate new markers within the QTL interval
and delimit the QTL region but also will serve as a
useful material to test association of putative
physiological processes associated with this impor-
tant QTL.
Conclusion
The results of the present study clearly establish that
the DT-QTL region on LG 2 contributed by PRLT 2/
89-33 also exerts favorable effects on growth and
productivity parameters of pearl millet under saline
and alkaline conditions. This DT-QTL does so by
limiting Na? transport to the photosynthesising
leaves. The exact mechanism by which the DT-
QTL limits Na? accumulation in leaves, however,
could not be established from the data generated in
this study. Preliminary physiological studies avail-
able on the drought-tolerant QTL donor parent and on
the QTL-NILs (Kholova et al. 2008) indicate that
they have lower transpiration rates and higher ABA
concentrations relative to the drought-sensitive recur-
rent parent H 77/833-2. Both low transpiration rate
(Munns and Richards 2007; Yadav et al. 1996) and
high ABA concentration (Voisin et al. 2006) can play
an important role in both Na? uptake and drought
tolerance and their possible roles in determining low
Na? transport observed in the QTL donor parent and
QTL introgressed NILs will be investigated and
reported in due course.
Acknowledgments The work reported in this study was
conducted under the auspices of the Collaborative Project with
Scientists & Technologists of Indian Origin Abroad Program
(CP-STIO) award to P.C.S. and R.S.Y. by the Department of
Science and Technology (DST), Government of India.
Financial support provided by the DST via grant number
DST/INT/CP-STIO/2006-07/60/2006 is gratefully acknow-
ledged. Plant materials used in the study was generated in a
separate project funded by the Biotechnology and Biological
Sciences Research Council (BBSRC) and Department for
International Development (DFID) to R.S.Y. via grant number
BB/F004133/1. The authors are grateful to Professor Tim
Flowers for critically reviewing the manuscript.
Mol Breeding (2011) 27:207–222 221
123

References
Akita S, Cabusaly GS (1990) Physiological basis of differential
response to salinity in rice cultivars. Plant Soil 123:277–294
Ashraf M, Khanum A (1997) Relationships between ion
accumulation and growth in two spring wheat lines dif-
fering in salt tolerance at different growth stages. J Agron
Crop Sci 178:39–51
Ashraf M, O’Leary JW (1996) Responses of some newly
developed salt-tolerant genotypes of spring wheat to salt
stress: 1. Yield components and ion distribution. J Agron
Crop Sci 176:91–101
Bidinger FR, Nepolean T, Hash CT et al (2007) Quantitative
trait loci for grain yield in pearl millet under variable
postflowering moisture conditions. Crop Sci 47:969–980
Chippa BR, Lal P (1995) Na/k ratios as the basis of salt tol-
erance in wheat. Aust J Agric Res 14:163–167
Flowers TJ, Yeo AR (1981) Variability in the resistance of
sodium chloride salinity within rice (Oryza sativa L)
varieties. New Phytol 88:363–373
Giuliani S, Sanguineti MC, Tuberosa R et al (2005) Root-ABA1a major constitutive QTL affects maize root architecture
and leaf ABA concentration at different water regimes.
J Exp Bot 56:3061–3070
Gorhom J, Wyn Jones RG, Bristol A (1990) Partial charac-
terization of the trait for enhanced K?-Na? discrimination
in the D genome of wheat. Planta 180:590–597
Hussain S, Munns R, Condon AG (2003) Effect of sodium
exclusion trait on chlorophyll retention and growth of
durum wheat in saline soil. Aust J Agric Res 54:589–597
Hussain S, Caemmerer S, Munns R (2004) Control of salt
transport from roots to shoots of wheat in saline soil.
Funct Plant Biol 31:1115–1126
Kholova J, Vadez V, Hash CT (2008) Mechanisms underlying
drought tolerance in pearlmillet (Pennisetum americanumL). In: 5th International crop science congress, 13–18
March 2008, Jeju South Korea Book of Abstracts, 188 pp
Kreps JA, Wu Y, Chang HS et al (2002) Transcriptome
changes for Arabidopsis in response to salt osmotic and
cold stress. Plant Physiol 130:2129–2141
Krishnamurthy L, Rachid S, Rai KN et al (2007) Identification
of pearl millet [Pennisetum glaucum (L) R Br] lines tol-
erant to soil salinity. Euphytica 158:179–188
Lauchli A (1984) Salt exclusion: an adaptation of legumes for
crops and pastures under saline conditions. In: Staples RC
(ed) Salinity tolerance in plants: strategies for crop
improvement. Wiley, New York, pp 171–187
Munns R (2002) Comparative physiology of salt and water
stress. Plant Cell Environ 25:239–250
Munns R, James RA (2003) Screening methods for salt tolerance:
a case study with tetraploid wheat. Plant Soil 253:201–218
Munns R, Richards RA (2007) Recent advances in breeding
wheat for drought and salt stresses In: Jenks MA (ed)
Advances in molecular breeding towards salinity and
drought tolerance, pp 565–585
Munns R, Rebetzke GJ, Hussain S et al (2003) Genetic control
of sodium exclusion in durum wheat. Aust J Agric Res
54:627–635
Netondo GW, Onyango JC, Beck E (2004) Sorghum and
salinity: I. Response of growth water relations and
ion accumulation to NaCl salinity. Crop Sci 44:797–
805
Poustini K, Siosemardeh A (2004) Ion distribution in wheat
cultivars in response to salinity stress. Field Crop Res
85:125–133
Rabbani MA, Maruyama K, Abe H et al (2003) Monitoring
expression profiles of rice genes under cold drought and
high-salinity stresses and abscisic acid application using
cDNA microarray and RNA gel-blot analyses. Plant
Physiol 133:1755–1767
Seki M, Narusaka M, Ishida J et al (2002) Monitoring the
expression profiles of 7000 Arabidopsis genes under
drought cold and high-salinity stresses using a full-length
cDNA microarray. Plant J 31:279–292
Serraj R, Hash CT, Rizvi SMH et al (2005) Recent advances in
marker-assisted selection for drought tolerance in pearl
millet. Plant Prod Sci 8:334–337
Sharma PC, Gill KS (1992) Effect of salinity on yield and ion
distribution in pearlmillet genotypes. Arid Soil Res
Rehabil 6:253–260
Sharma PC, Kumar P (1999) Alleviation of salinity stress
during germination in Brassica juncea by pre-sowing chill-
ing treatments to seeds. Biol Plantarum 42(3):451–455
Tester M, Davenport R (2003) Na? tolerance and Na? trans-
port in higher plants. Ann Bot 91:503–527
Voisin AS, Reidy B, Parent B et al (2006) Are ABA ethylene
or their interaction involved in the response of leaf growth
to soil water deficit? An analysis using naturally occurring
variation or genetic transformation of ABA production in
maize. Plant Cell Environ 291:829–1840
Yadav R, Flowers TJ, Yeo AR (1996) Involvement of tran-
spirational bypass flow in sodium uptake by high- and low
sodium-transporting lines of rice developed through in-
travarietal selection. Plant Cell Environ 19:329–336
Yadav RS, Hash CT, Bidinger FR et al (2002) Quantitative
trait loci associated with traits determining grain and
stover yield in pearl millet under terminal drought stress
conditions. Theor Appl Genet 104:67–83
Yadav RS, Hash CT, Bidinger FR et al (2004) Genomic
regions associated with grain yield and aspects post-
flowering drought tolerance in pearl millet across stress
environments and tester background. Euphytica 136:265–
277
Yamaguchi-Shinozaki K, Shinozaki K (2006) Transcriptional
regulatory network in cellular responses and tolerance to
dehydration and cold stress. Annu Rev Plant Biol 57:781–
803
Yeo AR (1992) Variation and inheritance of sodium transport
in rice. Plant Soil 146:104–116
Yeo AR, Flowers TJ (1984) Mechanisms of salinity resistance
in rice and their role as physiological criteria in plant
breeding. In: Staples RC, Toennissen GH (eds) Salinity
tolerance in plants: strategies for crop improvement.
Wiley, New York, pp 151–170
Zhu J-K (2002) Salt and drought signal transduction in plants.
Annu Rev Plant Biol 53:247–273
Zhu GY, Kinet J-M, Lutts S (2004) Characterisation of rice
(Oryza sativa) F3 populations selected for salt resistance
2. Relationship between yield-related parameters and
physiological properties. Aust J Exp Agric 44:333–342
222 Mol Breeding (2011) 27:207–222
123




















