
Genome turnover in great millet and related millets ( Poaceae
11
P1. Syst. Evol. 179: 155-165 (1992) Plant Systematics and Evolution © Springer-Verlag 1992 Printed in Austria Genome turnover in great millet and related millets (Poaceae) LALITHA SUNIL KUMAR, LAKSHMI SIVARAMAN, a n d PRABHAKAR K. RANJEKAR Received August 3, 1990; in revised version September 18, 1991 Key words: Angiosperms, Poaceae, millets.- Homogeneous/heterogeneous repeat families, fossil repeats, genome turnover. Abstract: Based on optical reassociation studies of total nuclear DNAs at 55 °C, 62 °C, 69 °C, and 75 °C, it is concluded that repeat families in great millet, little millet, barn yard millet and finger millet are heterogeneous while those of fox tail millet are homogeneous. In great millet, almost one third of the sequences that behave as single copy at standard conditions are actually fossil repeats. Such "fossil" repeats are not a prominent feature of the genomes of the other four millets. The ratios of sequence complexities of repeats isolated at 75°C to those isolated at 55°C are 2.2, 3.5, 81, and 0.3 in case of little millet, finger millet, fox tail millet, and great millet respectively. On the basis of the above three obser- vations, it is suggested that among these millets, the rate of turnover of the genome of fox tail millet is the slowest while that of great millet is the fastest. Such comparative estimates of differences in the turnover rates of genomes of related species are expected to generate useful data about the evolution of genomes. It is now well known that repeated DNA sequences are not highly conserved during evolution (FLAVELL 1986). The various changes that take place in the repetitive component of any genome have been described by FLAVELL (1982, 1986), FLAVELL & al. (1980), DOVER (1980, 1982), DOVER & FLAVELL (1984), and TnoMvSON & MURRAY (1980) and are depicted in Fig. 1. In this model of genome evolution, amplification processes are responsible for the increase in the DNA content. The newly created repeat sequences are then subjected to deletions, mutations, trans- positions, and compound amplification events (FLAVELL 1982, 1986) leading to turnover of DNA sequences during evolution (FLAVELL 1980, THOMPSON & MUR- RAY 1980). The turnover process also includes events such as crossing over and gene conversion (DOVER 1982). All these processes resulting in the turnover of repeated DNA sequences in a genome are involved along with natural selection and genetic drift in the fixation of repeated DNA in populations (DovER 1982). Families of repeated DNA sequences would vary in the extent of base sequence divergence among members of the same family as well as between members of different families and this divergence in turn may reflect the time elapsed since the family was first formed as well as the turnover rate of the genome.
Transcript of Genome turnover in great millet and related millets ( Poaceae

P1. Syst. Evol. 179: 155-165 (1992) Plant
Systematics and
Evolution © Springer-Verlag 1992 Printed in Austria
Genome turnover in great millet and related millets (Poaceae)
LALITHA SUNIL KUMAR, LAKSHMI SIVARAMAN, and PRABHAKAR K. RANJEKAR
Received August 3, 1990; in revised version September 18, 1991
Key words: Angiosperms, Poaceae, millets.- Homogeneous/heterogeneous repeat families, fossil repeats, genome turnover.
Abstract: Based on optical reassociation studies of total nuclear DNAs at 55 °C, 62 °C, 69 °C, and 75 °C, it is concluded that repeat families in great millet, little millet, barn yard millet and finger millet are heterogeneous while those of fox tail millet are homogeneous. In great millet, almost one third of the sequences that behave as single copy at standard conditions are actually fossil repeats. Such "fossil" repeats are not a prominent feature of the genomes of the other four millets. The ratios of sequence complexities of repeats isolated at 75°C to those isolated at 55°C are 2.2, 3.5, 81, and 0.3 in case of little millet, finger millet, fox tail millet, and great millet respectively. On the basis of the above three obser- vations, it is suggested that among these millets, the rate of turnover of the genome of fox tail millet is the slowest while that of great millet is the fastest. Such comparative estimates of differences in the turnover rates of genomes of related species are expected to generate useful data about the evolution of genomes.
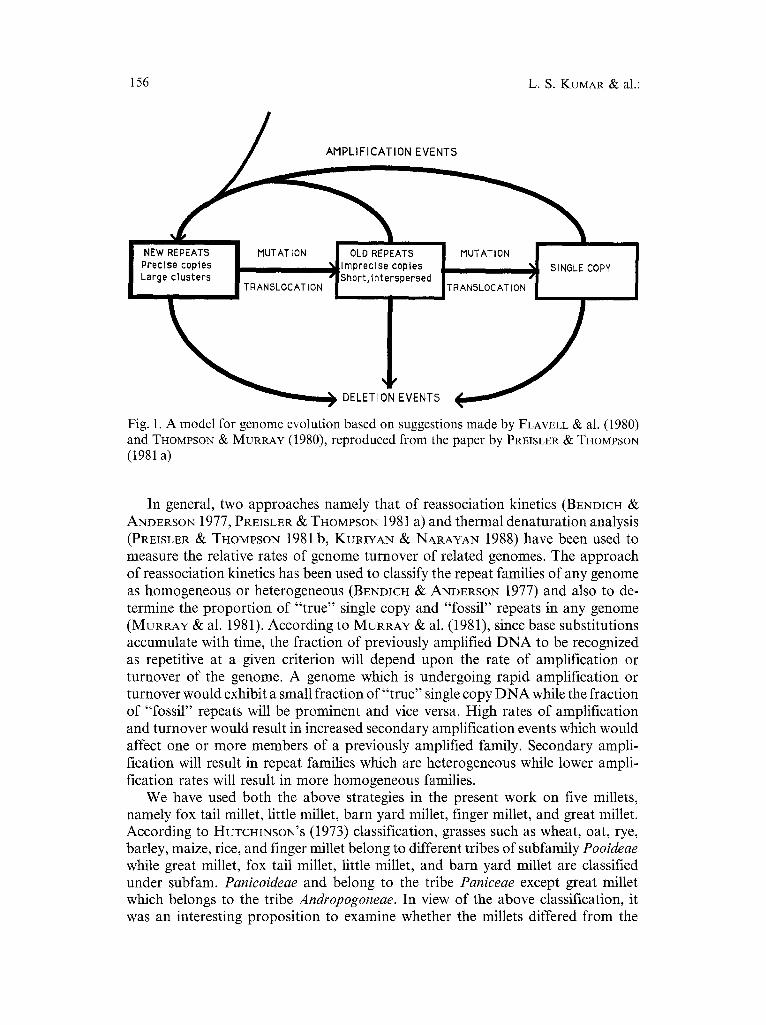
It is now well known that repeated D N A sequences are not highly conserved during evolution (FLAVELL 1986). The various changes that take place in the repetitive component of any genome have been described by FLAVELL (1982, 1986), FLAVELL & al. (1980), DOVER (1980, 1982), DOVER & FLAVELL (1984), and TnoMvSON & MURRAY (1980) and are depicted in Fig. 1. In this model of genome evolution, amplification processes are responsible for the increase in the D N A content. The newly created repeat sequences are then subjected to deletions, mutations, trans- positions, and compound amplification events (FLAVELL 1982, 1986) leading to turnover of D N A sequences during evolution (FLAVELL 1980, THOMPSON & MUR- RAY 1980). The turnover process also includes events such as crossing over and gene conversion (DOVER 1982). All these processes resulting in the turnover of repeated D N A sequences in a genome are involved along with natural selection and genetic drift in the fixation of repeated D N A in populations (DovER 1982). Families of repeated D N A sequences would vary in the extent of base sequence divergence among members of the same family as well as between members of different families and this divergence in turn may reflect the time elapsed since the family was first formed as well as the turnover rate of the genome.
156 L.S. KUMAR 8¢ al.:
AMPLIFICATION EVENTS
NEW REPEATS MUTATION OLD REPEATS MUTATION Precise copies ~ l m p r e c l s e copies Large clusters Short,Interspersed
TRANSLOCATION TRANSLOCATION
SINGLE COPY
DELETION EVENTS
Fig. 1. A model for genome evolution based on suggestions made by FLAVELL 8~; al. (1980) and THOMPSON & MURRAY (1980), reproduced from the paper by PREISLER ~ THOMPSON (1981 a)
In general, two approaches namely that of reassociation kinetics (BENDICH 8¢ ANDERSON 1977, PREISLER & THOMPSON 1981 a) and thermal denaturation analysis (PREISLER 8¢ THOMPSON 1981 b, KURIYAN & NARAYAN 1988) have been used to measure the relative rates of genome turnover of related genomes. The approach of reassociation kinetics has been used to classify the repeat families of any genome as homogeneous or heterogeneous (BENDICH & ANDERSON 1977) and also to de- termine the proportion of "true" single copy and "fossil" repeats in any genome (MURRAY & al. 1981). According to MURRAY 8¢ al. (1981), since base substitutions accumulate with time, the fraction of previously amplified DNA to be recognized as repetitive at a given criterion will depend upon the rate of amplification or turnover of the genome. A genome which is undergoing rapid amplification or turnover would exhibit a small fraction of"true" single copy DNA while the fraction of "fossil" repeats will be prominent and vice versa. High rates of amplification and turnover would result in increased secondary amplification events which would affect one or more members of a previously amplified family. Secondary ampli- fication will result in repeat families which are heterogeneous while lower ampli- fication rates will result in more homogeneous families.
We have used both the above strategies in the present work on five millets, namely fox tail millet, little millet, barn yard millet, finger millet, and great millet. According to HUTCHINSON'S (1973) classification, grasses such as wheat, oat, rye, barley, maize, rice, and finger millet belong to different tribes of subfamily Pooideae while great millet, fox tail millet, little millet, and barn yard millet are classified under subfam. Panicoideae and belong to the tribe Paniceae except great millet which belongs to the tribe Andropogoneae. In view of the above classification, it was an interesting proposition to examine whether the millets differed from the

Genome turnover in millets 157
other grasses at the molecular level and if so, did these changes offer a selective advantage to them. Selective advantages may be in the form of reassortment of genes and other sequences, high genetic flexibility and/or extensive specialization. In the present work, an attempt has been made to interpret, in terms of an evo- lutionary perspective, the significance of the differences observed between the DNAs of millets and cereals as well as the differences observed among the millet DNAs.
Material and methods
Seed material. Seeds of barn yard millet, EchinochIoa frumentacea (RoxB.) W. F. WIGHT CV. 'Madira', were obtained from Vivekananda Parvatiya Krishi Anusandhan Shala, ICAR, Almora, Uttar Pradesh. Italian fox tail millet, Setaria italica (L.) P. BEAUVO~S, were received from IDRC Project, Cotton Research Station, Nandyal, Andhra Pradesh. Little millet, Panicum miliare L., seeds were procured from Regional Research Station, OVAT, Semi- liguda, Sunabeda, Orissa State. Seeds of great millet, Sorghum vulgare (L.) MOENCH CV. 'Maldandi', and finger millet, Eleusine coracana (L.) GaERXN CV. 'Madwa VL 101', were purchased locally.
Methods. The extraction procedure of plant DNA as well as the shearing and sizing of DNA were done as described by SIVARAMAN • al. (1984). Experiments of DNA reasso- ciation kinetics and the isolation of S 1-resistant repeat DNA fractions at 55 °C and 75 °C were done as described by SIVARAMAN & RANJEKAR (1984).
Results
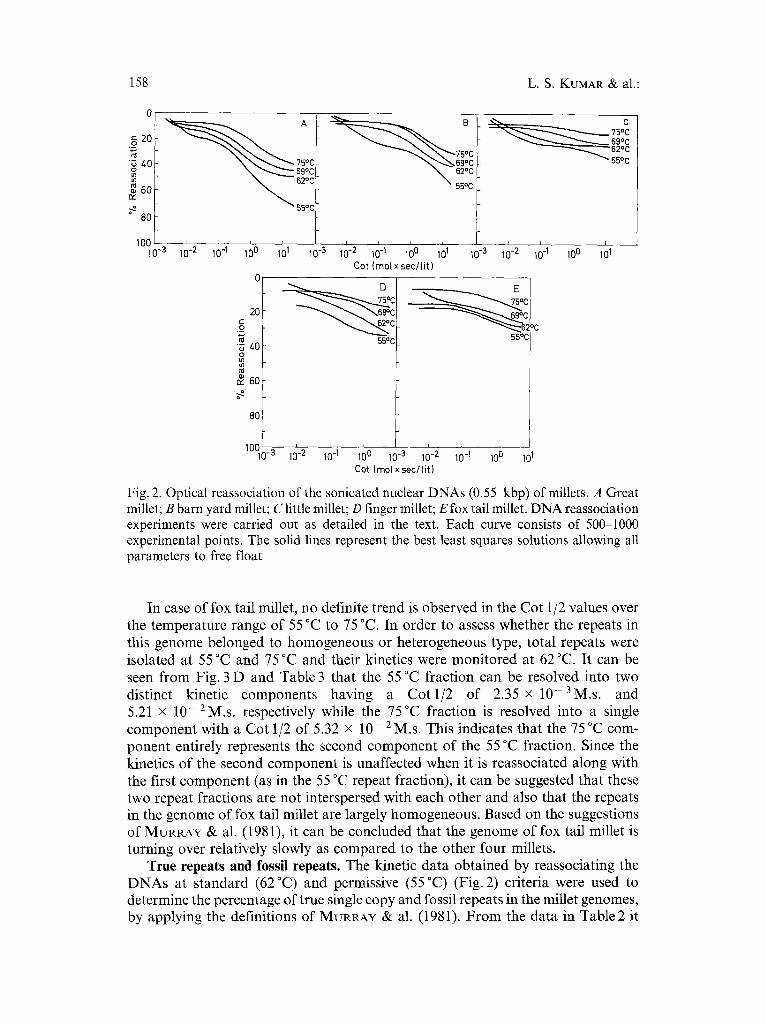
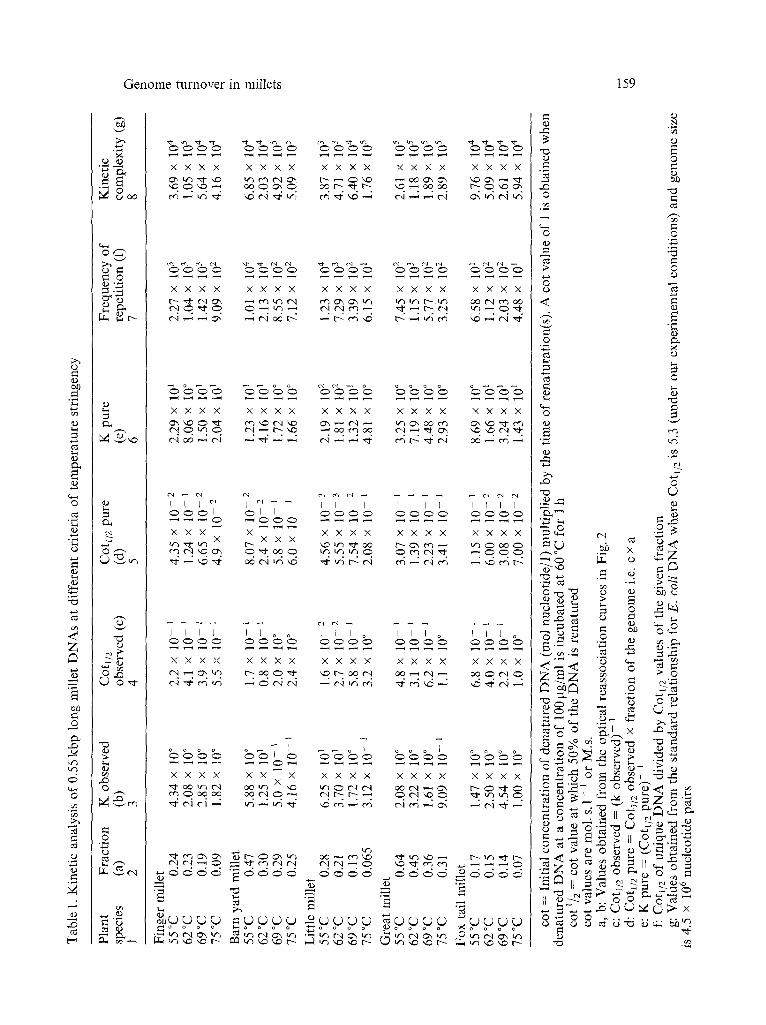
Optical reassociation studies of total nuclear DNAs at 55 °C, 62 °C, 69 °C, and 75 °C. To determine the nature of repeat families, reassociation of 550 bp long DNAs were studied optically up to a Cot value of 10 M.s. at four different temperatures namely 55 °C, 62 °C, 69 °C, and 75 °C (Fig. 2). All the reassociation curves were evaluated using a computer aided non-linear regression method (Table 1). Though in a few cases, the curves could be modelled into two components, all the data were fitted as a single component to facilitate a better comparison of their rate constants. The Cot 1/2 of single copy DNA in all the reassociation experiments was considered to be the same as that at 62 °C.
From Fig. 2 it can be seen that all the millet DNAs contain rapidly reassociating DNA sequences which is typical of higher eukaryotes. For determining the nature of repeat families, only those repetitive fractions which followed second order kinetics were analyzed. As the conditions of reassociation are made more stringent, differences in the proportions of repetitive DNA as well as their kinetic complexity, copy number and rate of reaction are evident (Table 1). In all the five millet DNAs, the fraction scored as repetitive decreases as the temperature of incubation is raised. This observation is consistent with the presence of either the homogeneous or heterogeneous model of repeat families proposed by BENDrCI4 & ANDERSON (1977). According to these authors, young and old families of repeats have the same copy number and hence an increase in the observed Cot 1/2 values with an increase in temperature would be expected if the repeat families in the genome were predom- inantly of the heterogeneous type and no significant change would be observed in the Cot 1/2 values if the genome contained predominantly homogeneous repeat families. In case of great millet, little millet, barn yard millet, and finger millet, an increase in the Cot 1/2 values is observed suggesting that the repeat sequences in these plant genomes are predominantly heterogeneous in nature.
158 L. S, KUMAR & al.:
._o= 20
~ 40 o
60 tY
8O
A
75oc \ ~ 6 9 o c
~ 62oc
~ 5 5 o c
100 . . . . 0-3 10-2 10-1 100 101 10 -3
0
2O
'5 40 o
~. 60
80
B
75oc 69°C
~ 62oc ~ 5 5 o c
, i i i
10-2 10-1 100 101 10 -3 Cot(molx sec/lit)
ct 55oC
~ vs c 69oC 62oc 55oc
10-2 10-1 10 0 101
55oc
i i i i i
10 0- 3 10_2 10.1 100 10_3 10~2 10_1 100 101 Cot (mo[ x sec/lit)
Fig. 2. Optical reassociation of the sonicated nuclear DNAs (0.55 kbp) of millets. A Great millet; B barn yard millet; C little millet; D finger millet; E fox tail millet. DNA reassociation experiments were carried out as detailed in the text. Each curve consists of 500-1000 experimental points. The solid lines represent the best least squares solutions allowing all parameters to free float
In case of fox tail millet, no definite trend is observed in the Cot 1/2 values over the temperature range of 55 °C to 75 °C. In order to assess whether the repeats in this genome belonged to homogeneous or heterogeneous type, total repeats were isolated at 55 °C and 75 °C and their kinetics were monitored at 62 °C. It can be seen from Fig. 3 D and Table 3 that the 55 °C fraction can be resolved into two distinct kinetic components having a Cot 1/2 of 2.35 x 10-3M.s . and 5.21 x 10-2M.s . respectively while the 75°C fraction is resolved into a single component with a Cot 1/2 of 5.32 x 10- 2 M.s. This indicates that the 75 °C com- ponent entirely represents the second component of the 55 °C fraction. Since the kinetics of the second component is unaffected when it is reassociated along with the first component (as in the 55 °C repeat fraction), it can be suggested that these two repeat fractions are not interspersed with each other and also that the repeats in the genome of fox tail millet are largely homogeneous. Based on the suggestions of MURRAY & al. (1981), it can be concluded that the genome of fox tail millet is turning over relatively slowly as compared to the other four millets.
True repeats and fossil repeats. The kinetic data obtained by reassociating the DNAs at standard (62 °C) and permissive (55 °C) (Fig. 2) criteria were used to determine the percentage of true single copy and fossil repeats in the millet genomes, by applying the definitions of MURRAY & al. (1981). F rom the data in Table2 it
Genome turnover in millets 159
>-. O
,.-t
O
"r--,
"T--, O
"4
Z n Z
_.9.,o
,_O
, 5
O
r.9
O
[,..
~0
..~ 0 0 0 0 O 0 O 0 0 0 0 0 ~ 0 O 0 0 C~ 0 C~
~ , - " X X X X X X X X X X X X X X X X X X X X
'.-.1~ O '.D ~*','~ oo C~, C~ ~ 00 I>- "~ r-- ,.@ ,,,--~ oo oo t..- C~ '..o O',
~ o o o o o o o o o o o o o o o o
~:~ x x x x x x x x x x x x x x x x
---- O 0 0 0 0
X X X X X X X X X X X X X X X X
6) ;::1 Ira..,
O . . - ~ :O~, ,
q , . o r,.p o ,~
>
O
O
~ ' ~
i l l ~ ~ - I I I I
X X X X X X X X X X X X
I l l l I I I I I o o o o o o b b o o o b
X X X X X X X X X X X X
~ ~ o l o o o o o
X X X X X X ~ X X X X X
~ d d d d - " " " " ~ o o o o d o o d
X X X X
X X X X
, 7 7 ,
7777 r l l F
X X X X
X X X X X X X X
o o o 7 0 o o 0
X X X X X X X X
Zdddd ~ dddd..
eppp " o PPPP
(D ,xZ
(D
O
©
N]
O
¢,)
©
" a
> © O
<
,.,_/
© ,._..,
¢)
¢.) r.m
O ,"u ro
~ ,..,2 ° ~ ~ "~ E~rO ~ x ~ <
. . . . , . b Z
. ~ .~ d ~.~
o = ~ o Nrd m ~ - . ~ = '.-' ~,-N_ m
< - - < o > o Z ~ Z ~ = ~.~
m' x "°41
o ~ ~ .,:_,:s
o ~ ~ 0 ~ ~.-~. ~-~ :~ o . ' < ~ 4..~.,._ ~ , ~ 4..a ~ II 0 0 " ~ 4'-~ 0 ~ ~ ~ , - 0 ,C " r ~ . , - . , - ~ 0
II ~ - - - > . ._o~ o ~ x 0 +.~ 0 0 ~ . . . . . . . . ~ c) ~ r..;, o ~ o " ~ ¢,;,,4.q e.0.~

160 L.S. KUMAR & al.:
20
t0
60
8O
¢J o 100
2o
Z,0:
60
80
100 10-3
A
(Sl-55)
( s = - 5 5 ) ~
1:2; 2', (st_75) ~sl-55) Component 1
~ _~,~5) Component 2 \
I0-2 I0-I 10 0 I0 -3 I0-2 i0-I 10 0 101 Cot (tool x secl l i t )
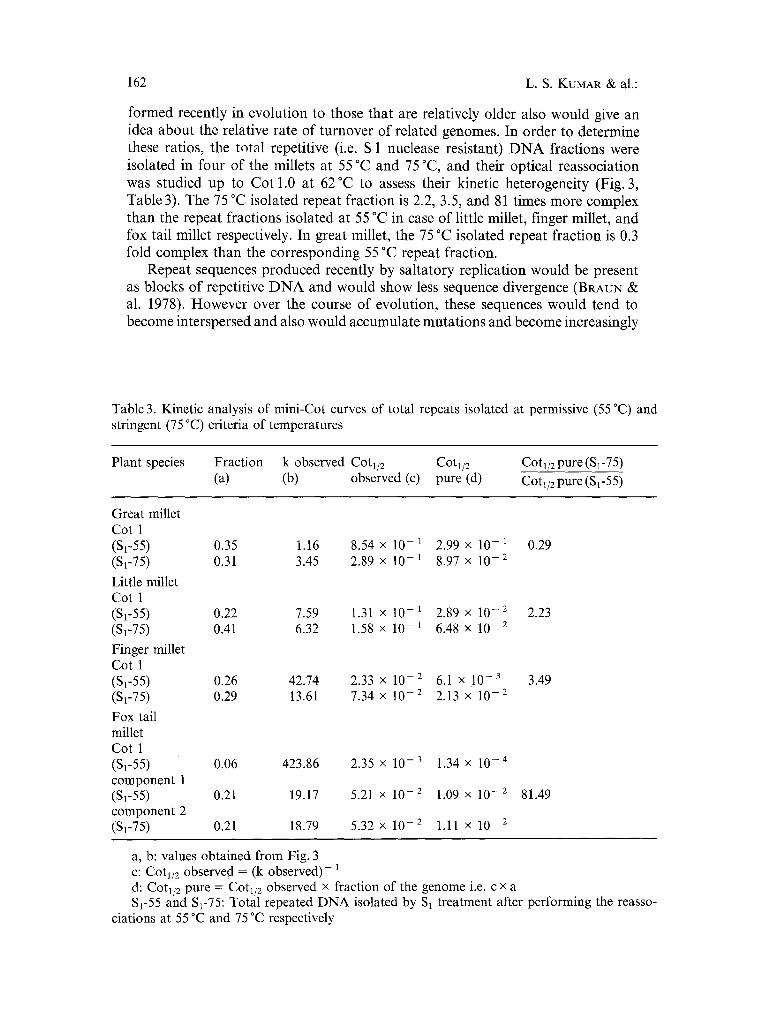
Fig. 3. Mini Cot curves of the total repeats isolated at permissive (55 °C) and stringent (75 °C) temperatures, monitored at 62 °C. A Great millet; B little millet; C finger millet; D fox tail millet. Enriched total repeat fractions, isolated by S 1 nuclease treatment, were melted and their optical reassociation was followed in 0.12 M sodium phosphate buffer, pH 6.8 up to a Cot value of 1.0 M.s. at 62 °C. Each curve consists of 500-1000 experimental points. The solid lines represent the best least squares fit, allowing all parameters to free float. S 1-55 and S 1-75: Total repeats isolated by S 1 treatment after performing the reas- sociations at 55 °C and 75 °C respectively
can be seen that finger millet, fox tail millet, and barn yard millet have the m a x i m u m percent of true single copy D N A of 98.48%, 96.42%, and 95.43% respectively and this value decreases to 87.44% in little millet and 63.07% in great millet. Thus as compared to the other four millets, a greater amoun t of single copy D N A in great millet behaves as fossil repeats at lower temperatures. A smaller fraction of fossil repeats and a larger fraction of true single copy indicates a relatively low rate of turnover of the genome while a larger fraction of fossil repeats and a lower fraction of true single copy indicates a relatively higher rate of turnover (MURRAY • al. 1981). Our data thus suggests that the genome of great millet is turning over at a rate faster than that of the other four millets.
Self-reassociation at 62 °C of the total repeat fractions isolated at permissive (55 °C) and stringent (75 °C) temperatures. The ratios of kinetic complexities of repeats

Gen0me turnover in millets 161
©
~D
o
© o
o
©
e-i o
,x3
c~
• ~ ~ . ~ 0
o ~ ' ~ ,
;2 ~
~ . ~ o o
I
~ ~
~ ~
X X
I'~ ¢¢)
¢¢3 t'~
c q ' ~ - e¢~ t"- c 5 c 5
o
X X X
o o ~ oo
@ ~ e q
e z ~
u ' 3 0 tg3
e,i c5 m
t 'q q') 1-~ tt3 ,4~ ,,D
X X ~ ~ X ~
ID
~D
(D
©
~D
e-I
©
< Z
©
~D
~× tt'3
~'~
o
[I . o
~
~ , s z ~ -~
O
O
X
, . a l o
o i
O
X
~5
© o o
©
©
0 .~, o
X (LI N
(D
O
(D
162 L.S. KUMAR & al.:
formed recently in evolution to those that are relatively older also would give an idea about the relative rate of turnover of related genomes. In order to determine these ratios, the total repetitive (i.e. S 1 nuclease resistant) D N A fractions were isolated in four of the millets at 55 °C and 75 °C, and their optical reassociation was studied up to Cot 1.0 at 62°C to assess their kinetic heterogeneity (Fig. 3, Table 3). The 75 °C isolated repeat fraction is 2.2, 3.5, and 81 times more complex than the repeat fractions isolated at 55 °C in case of little millet, finger millet, and fox tail millet respectively. In great millet, the 75 °C isolated repeat fraction is 0.3 fold complex than the corresponding 55 °C repeat fraction.
Repeat sequences produced recently by saltatory replication would be present as blocks of repetitive D N A and would show less sequence divergence (BRAUN & al. 1978). However over the course of evolution, these sequences would tend to become interspersed and also would accumulate mutat ions and become increasingly
Table 3. Kinetic analysis of mini-Cot curves of total repeats isolated at permissive (55 °C) and stringent (75 °C) criteria of temperatures
Plant species Fraction k observed Cotl/2 COtl/2 (a) (b) observed (c) pure (d)
COtl/2 pure ($1-75)
Cotl/2 pure ($1-55)
Great millet Cot 1 (81-55) 0.35 1.16 8.54 x 10- ~ 2.99 x 10- 1 0.29 (SF75) 0.31 3.45 2.89 x 10- i 8.97 x 10-2
Little millet Cot 1 (S1-55) 0.22 7.59 1.31 x 10-1 2.89 x 10-2 2.23 (81-75) 0.41 6.32 1.58 x 10-1 6.48 x 10-2
Finger millet Cot 1 (S1-55) 0.26 42.74 2.33 x 10-2 6.1 x 10-3 3.49 ($1-75) 0.29 13.61 7.34 x 10-2 2.13 x 10-2
Fox tail millet Cot 1 (81-55) 0.06 423.86 2.35 x 10- 3 1.34 x 10- 4 component 1 ($1-55) 0.21 19.17 5.21 x 10-2 1.09 x 10-2 81.49 component 2 ($1-75) 0.21 18.79 5.32 x 10-2 1.11 x 10-2
a, b: values obtained from Fig. 3 c: Cotl/2 observed = (k observed)- 1 d: Cotl/a pure = Cotl/e observed x fraction of the genome i.e. c x a $1-55 and $1-75: Total repeated DNA isolated by $1 treatment after performing the reasso-
ciations at 55 °C and 75 °C respectively

Genome turnover in millets 163
diverged with time. The rate at which these processes occur would determine the turnover rate of the genome. Repeat sequences having functional significance would remain conserved. Thus, the sequences reassociating at 75 °C would include some of these conserved sequences as well as recently formed sequences. In a genome exhibiting a high turnover rate, the ratio of sequence complexities of the repeats isolated at 75 °C to those isolated at 55 °C would be less than 1 as seen in great millet while higher ratios would be expected for a genome which is turning over at a slower rate as observed in the other three millets (Table 3). Our data thus suggest that the genome of great millet is turning over at a faster rate than that of the other three millets.
Discussion
The present work provides some experimental evidence to the predictions of MUR- RAY & al. (1981) where attempts have been made to correlate rate of genome turnover with DNA content, nature of repeat families, and interspersion patterns. Among the five millets under consideration, the turnover rate of the great millet genome is the fastest while that of fox tail millet is the slowest. This conclusion has been arrived at from the present analysis of the reassociation kinetics data. The data (Table 1) obtained from such studies indicate the presence of homogeneous families in fox tail millet and of heterogeneous families in the other four millets. Analysis of the 55 °C and 62 °C optical reassociation curves indicate that in great millet, of the total DNA reassociating as single copy at standard criterion, almost one third behaves as fossil repeats at permissive criterion (Table 2). In case of the other four millets, most of the single copy sequences are true single copy. The self- reassociation of the repeats isolated at 55 °C and 75 °C at 62 °C has shown that the fraction isolated at 75 °C is 81, 3.5, and 2.2 times more complex than the 55 °C fraction in case of fox tail millet, finger millet, and little millet respectively, while it is only 0.3 fold complex in case of great millet. Thus it is suggested that among these millets, the rate of turnover of the genome of fox tail millet is the slowest. The presence of a relatively higher proportion of fossil repeats in great millet than in the other four millets indicates that the rate of turnover of the great millet genome is comparatively the fastest. The higher genome turnover rate of great millet is also reflected by its higher DNA content and higher repetitive DNA content as compared to the other four millets (SIvARAMAN • al. 1984).
A comparison of the important features of the genomes of millets under con- sideration and those of cereals reveals that the genomes of the millets are turning over at a relatively slower rate than those of the cereals. This conclusion follows from the predictions of FLAVELL (1986) and PREISLER ~; THOMPSON (1981 a) that the genome turnover rate would be expected to be greater in large genomes since more DNA has to be replicated. According to MURRAY ~; al. (1981) such high turnover or rapid amplifications would produce highly repetitive genomes with extensive short period interspersion, while slower turnover should result in a smaller fraction of repetitive DNA and an increase in the length of interspersed single copy and repeat DNA. The cereal genomes show predominantly a short period inter- spersion pattern and have a high proportion of repeated DNA that exhibits almost 13-15% base mismatch (SMITH ~; FLAVELL 1975; FLAVELL & SMITH 1976; RAN- JEKAR & al. 1974, 1976; FLAVELL & al. 1981). On the contrary, the millets have a

164 L.S. KVMAR & al.:
lower D N A content (1 C < 5 pg) and in general a lower repetitive D N A content and show a base sequence divergence of 6-8% (finger millet shows a base sequence divergence of 10-13%) and exhibit either a long or mixed period interspersion pattern (DESHPANDE • RANJEKAR 1980; SIVARAMAN & RANJEKAR 1984; GUPTA & RANJEKAR 1981 ; SIVARAMAN & al. 1984, 1986). Though finger millet is classified under the same subfamily as the cereals, it shows a base sequence divergence that is intermediate between the cereals and millets and a low D N A content and in- terspersion pattern like the millets (DESHPANDE & RANJEKAR 1980, GUPa'A & RANJEKAR 1981).
After having assessed the relative rate of genome turnover in the millets and cereals, a question can be raised about its probable significance. We postulate that specific functional constraints are imposed on the millet genomes to maintain a low rate of turnover. We suggest that environmental factors are responsible for the maintenance of low turnover rates. Turnover rates in turn reflect on the genome size. Presence of long tandem arrays of clustered repeats is assumed to bring about a folding of chromosomes and thus a reduction in genome size. Genome size in turn is very useful in predicting plant responses to climate. A selective force which determines the relationship between genome size and growth under different eco- logical conditions arises from the differential effect of temperature on cell division and cell expansion (PRICE 1988 a, b). Species growing at low temperatures (tem- perate climates) have long mitotic and meiotic cycles resulting in cell enlargement. This enlargement is not observed in plants growing in warm and arid environment and they usually show smaller genomes. All the five millets taken up in the present study have low D N A content and are of tropical habitat. Thus, temperature appears to control the turnover rates in these millets which in turn controls the genome size.
R e f e r e n c e s
BENOICH, A. J., ANDERSON, R. S., 1977: Characterization of families of repeated DNA sequences from four vascular plants. - Biochemistry 16: 4655-4663.
BRAUN, B. A., SCHANKE, K. E., GRAHAM, D. E., 1978: Isolation of discrete repetitive sequence classes from Xenopus DNA by high temperature reassociation. - Nucleic Acids Res. 5: 4283-4303.
DESHPANDE, V. G., RANJEKAR, P. K., 1980: Repetitive DNA in three Gramineae species with low DNA content. - Hoppe-Seyler's Z. Physiol. Chem. 361: 1223-1233.
DOVER, G. A., 1980: Ignorant DNA? - Nature 285: 618-620. - 1982: Molecular drive: a cohesive mode of species evolution. - Nature 299:111-117. -- FLAVELL, R. B., 1984: Molecular coevolution. DNA divergence and the maintenance
of function. - Cell 38: 622-623. FLAVELL, R. B., 1980: The molecular characterization and organisation of plant chro-
mosomal DNA sequences. - Ann. Rev. P1. Physiol. 31: 569-596. - 1982: Amplification, deletion, and rearrangement: major sources of variation during
species divergence. - In DOVER, G. A., FLAVELL, R. B., (Eds.): Genome evolution, pp. 301-324. - New York, London: Academic Press.
- 1986: Repetitive DNA and chromosome evolution in plants. - Phil. Trans. Roy. Soc. London B 312: 227-242.
-- SMITH, D. B., 1976: Nucleotide sequence organization in the wheat genome. - Heredity 37:231 252.
- BEDBROOK, J. R., JONES, J., O'DELL, M., GERLACH, W. L., DYER, T. A., THOMPSON,

Genome turnover in millets 165
R. D., 1980: Molecular events in cereal genome evolution. - In DAVIES, D. R., HoP- WOOD, D. A., (Eds.): Proceedings of the fourth John Innes Symposium. The Plant Genome and second International Haploid Conference, pp. 15-30. - John Innes Inst. Norwich, U.K.
- O'DELL, M., HUTCHINSON, J., 1981: Nucleotide sequence organization in plant chro- mosomes and evidence for sequence translocation during evolution. - Cold Spring Harbor Symposium Quant. Biol. 45: 501-508.
GUPTA, V. S., RANJEKAR, P. K., 1981 : DNA sequence organization in finger millet (EIeusine coracana). - J. Biosci. 3: 417-430.
HUTCHINSON, J., 1973: In: The families of flowering plants. 3rd edn. - Oxford: Oxford University Press.
KURIYAN, P. N., NARAYAN, e. K. J., 1988: The distribution and divergence during evolution of families of repetitive DNA sequences in Lathyrus species. - J. Mol. Evol. 27: 303- 310.
MURRAY, M. G., PETERS, D. L., THOMPSON, W. F., 1981: Ancient repeated sequences in the pea and mung bean genomes and implications for genome evolution. - J. Mol. Evol. 17: 31-42.
PREISLER, R. S., THOMPSON, W. F., 1981 a: Evolutionary sequence divergence within re- peated DNA families of higher plant genomes. I. Analysis of reassociation kinetics. - J. Mol. Evol. 17: 78-84.
1981 b: Evolutionary sequence divergence within repeated DNA families of higher plant genomes. II. Analysis of thermal denaturation. - J. Mol. Evol. 17: 85-93.
PRICE, H. J., 1988 a: DNA content variation among higher plants. - Ann. Missouri Bot. Gard. 75: 1248-1257.
- 1988 b: Nuclear DNA content variation within angiosperm species. - Evol. Trends Plants 2: 53-60.
RANJEKAR, P. K., LAFONTAINE, J. a . , PALLOTTA, D., 1974: Characterization of repetitive DNA in rye (Secale cereale). - Chromosoma 48: 427-440.
-- PALLOTTA, D., LAFONTAINE, J. G., 1976: Analysis of the genome of plants. II. Char- acterization of repetitive DNA in barley (Hordeum vulgare) and wheat (Triticum aestivum). - Biochim. Biophys. Acta 425: 30-40.
SIVARAMAN, L., RANJEKAR, P. K., 1984: Novel molecular features of millet genomes. - Ind. J. Biochem. Biophys. 21: 299-303.
- GUPTA, V. S., RANJEKAR, P. K., 1984: Molecular organization of great millet (Sorghum vulgare) DNA. - J. Biosci. 6: 795-809.
- 1986: DNA sequence organization in the genomes of three related millet plant species. - P1. Mol. Biol. 6: 375-388.
SMITH, D. B., FLAVELL, R. B., 1975: Characterization of the wheat genome by renaturation kinetics. - Chromosoma 50: 223-242.
THOMPSON, W. F., MURRAY, M. G., 1980: Sequence organization in pea and mung bean DNA and a model for genome evolution. - In DAVIES, J. R., HOPWOOD, D. A., (Eds.): Proceedings of the fourth John Innes Symposium. The Plant Genome and second International Haploid Conference, pp. 31-45. - John Innes Inst., Norwich, U.K.
Address of the authors: LALITHA SUNIL KUMAR, LAKSHMI SIVARAMAN, and PRABHAKAR K. RANJEKAR, Division of Biochemical Sciences, National Chemical Laboratory, Poona 411008, India.
Accepted October 3, 1991 by F. EHRENDOFER




















