
A genetic approach to estimating natal dispersal distances and self-recruitment in resident...
10
33 A genetic approach to estimating natal dispersal distances and self-recruitment in resident rainforest birds Stefan Woltmann, Thomas W. Sherry and Brian R. Kreiser S. Woltmann ([email protected]) and T. W. Sherry, Dept of Ecology and Evolutionary Biology, Tulane Univ., New Orleans, LA 70118, USA. Present address for SW: Dept of Renewable Natural Resources, Louisiana State Univ., Baton Rouge, LA 70803, USA. – B. R. Kreiser, Dept of Biological Sciences, e Univ. of Southern Mississippi, Hattiesburg, MS 39406, USA. Natal dispersal is a fundamental component of the ecology and evolutionary history of birds, yet is often prohibitively difficult to study. We characterized natal dispersal for the first time in a bird using molecular genetic parentage analyses in a tropical rainforest understory species, the chestnut-backed antbird (amnophilidae: Myrmeciza exsul). Median natal dispersal distance was ∼800 m (mean 931 84 (SE) m, n 48), with ∼90% of all distances 1500 m. We found no evidence of sex-biased dispersal. An index of self-recruitment (i.e. individuals establishing a territory within the population of origin) was higher in sites largely or entirely surrounded by non-forest, suggesting birds are reluctant to disperse out of preferred forest habitat. Via simulations, we confirmed that the genetic data had sufficient resolution to correctly identify parent-offspring dyads, but lacked resolution to identify other relationships (full-sib and half-sib) with confidence. Chest- nut-backed antbirds have measurable self-recruitment rates caused by short natal dispersal distances, and self-recruitment may be amplified by reluctance to disperse out of sites bordered by non-forest. Some tropical forest understory birds have naturally short dispersal distances, and our results have implications for understanding how species will be affected by fragmented landscapes and for the design of reserves. Dispersal is fundamentally linked to species ecology and life- histories. At ecological timescales, dispersal influences local population growth rates, gene flow, and the persistence of populations in fragmented landscapes (Turchin 1998). At evolutionary timescales, dispersal ultimately influences the formation and distribution of species. Dispersal can be func- tionally categorized as either natal (‘dispersal from the site or group of birth to that of first reproduction or potential reproduction’, Greenwood and Harvey 1982) or breeding (movements between breeding sites) dispersal; the former is the predominant mechanism of gene flow in many spe- cies (Newton 2007). Characterizing and quantifying natal dispersal is important because of its implications for the evolution of inbreeding avoidance and sex-biased dispersal (Johnson and Gaines 1990, Bohonak 1999, Barton 2001). Knowledge of both natal and breeding dispersal patterns is also crucial for interpreting population genetic structure, understanding responses to habitat fragmentation, and the effective design of reserves (Brawn et al. 1998, Shanks et al. 2003, Chetkiewicz et al. 2006, Heinz and Strand 2006). Despite its importance, studying dispersal of individuals remains challenging in mobile taxa such as birds. Moreover, although it may be possible to track individuals via mark- recapture or telemetry or other methods, the consequences of dispersal are still often difficult to infer because success- ful movement does not guarantee subsequent reproductive success. Natal dispersal in small birds is particularly challeng- ing to study, often requiring tremendous effort in terms of finding nests, banding nestlings, and following or relocating fledglings once they mature and acquire a territory (Woolfen- den and Fitzpatrick 1989, Verhulst et al. 1997, Wheelwright and Mauck 1998, Robinson 2000, Winkler et al. 2005). Young birds are difficult to follow, and in migratory species, distinguishing death from emigration is especially problem- atic (Arcese et al. 1992, Wheelwright and Mauck 1998). In tropical forest systems, many of these difficulties are magni- fied by the fact that territories are often larger, nests are often more cryptic (many nests remain undescribed), and nest suc- cess is often far lower than in temperate counterparts (Rob- inson et al. 2000, Jetz et al. 2008), making it challenging to find enough fledglings to study even if one could follow them to the time they eventually acquire their own breeding territory. Molecular genetic parentage analysis has been used to study natal dispersal in many taxa, including plants (Austerlitz et al. 2004, Robledo-Arnuncio and Gil 2004, Sezen et al. 2005, Hanson et al. 2007), mammals (Peacock and Ray 2001, Telfer et al. 2003, Waser et al. 2006, Taylor et al. 2007), and fishes (Saenz-Agudelo et al. 2009, Christie et al. 2010). Peery et al. (2008) inferred source-sink popu- lation dynamics in an isolated California bird population using molecular parentage data, and their study supports Journal of Avian Biology 43: 33–42, 2012 doi: 10.1111/j.1600-048X.2011.05572.x © 2012 e Authors. Journal of Avian Biology © 2012 Nordic Society Oikos Subject Editor: Staffan Bensch. Accepted 29 November 2011
Transcript of A genetic approach to estimating natal dispersal distances and self-recruitment in resident...

33
A genetic approach to estimating natal dispersal distances and self-recruitment in resident rainforest birds
Stefan Woltmann, Thomas W. Sherry and Brian R. Kreiser
S. Woltmann ([email protected]) and T. W. Sherry, Dept of Ecology and Evolutionary Biology, Tulane Univ., New Orleans, LA 70118, USA. Present address for SW: Dept of Renewable Natural Resources, Louisiana State Univ., Baton Rouge, LA 70803, USA. – B. R. Kreiser, Dept of Biological Sciences, The Univ. of Southern Mississippi, Hattiesburg, MS 39406, USA.
Natal dispersal is a fundamental component of the ecology and evolutionary history of birds, yet is often prohibitively difficult to study. We characterized natal dispersal for the first time in a bird using molecular genetic parentage analyses in a tropical rainforest understory species, the chestnut-backed antbird (Thamnophilidae: Myrmeciza exsul). Median natal dispersal distance was ∼800 m (mean 931 84 (SE) m, n 48), with ∼90% of all distances 1500 m. We found no evidence of sex-biased dispersal. An index of self-recruitment (i.e. individuals establishing a territory within the population of origin) was higher in sites largely or entirely surrounded by non-forest, suggesting birds are reluctant to disperse out of preferred forest habitat. Via simulations, we confirmed that the genetic data had sufficient resolution to correctly identify parent-offspring dyads, but lacked resolution to identify other relationships (full-sib and half-sib) with confidence. Chest-nut-backed antbirds have measurable self-recruitment rates caused by short natal dispersal distances, and self-recruitment may be amplified by reluctance to disperse out of sites bordered by non-forest. Some tropical forest understory birds have naturally short dispersal distances, and our results have implications for understanding how species will be affected by fragmented landscapes and for the design of reserves.
Dispersal is fundamentally linked to species ecology and life-histories. At ecological timescales, dispersal influences local population growth rates, gene flow, and the persistence of populations in fragmented landscapes (Turchin 1998). At evolutionary timescales, dispersal ultimately influences the formation and distribution of species. Dispersal can be func-tionally categorized as either natal (‘dispersal from the site or group of birth to that of first reproduction or potential reproduction’, Greenwood and Harvey 1982) or breeding (movements between breeding sites) dispersal; the former is the predominant mechanism of gene flow in many spe-cies (Newton 2007). Characterizing and quantifying natal dispersal is important because of its implications for the evolution of inbreeding avoidance and sex-biased dispersal (Johnson and Gaines 1990, Bohonak 1999, Barton 2001). Knowledge of both natal and breeding dispersal patterns is also crucial for interpreting population genetic structure, understanding responses to habitat fragmentation, and the effective design of reserves (Brawn et al. 1998, Shanks et al. 2003, Chetkiewicz et al. 2006, Heinz and Strand 2006).
Despite its importance, studying dispersal of individuals remains challenging in mobile taxa such as birds. Moreover, although it may be possible to track individuals via mark-recapture or telemetry or other methods, the consequences of dispersal are still often difficult to infer because success-ful movement does not guarantee subsequent reproductive
success. Natal dispersal in small birds is particularly challeng-ing to study, often requiring tremendous effort in terms of finding nests, banding nestlings, and following or relocating fledglings once they mature and acquire a territory (Woolfen-den and Fitzpatrick 1989, Verhulst et al. 1997, Wheelwright and Mauck 1998, Robinson 2000, Winkler et al. 2005). Young birds are difficult to follow, and in migratory species, distinguishing death from emigration is especially problem-atic (Arcese et al. 1992, Wheelwright and Mauck 1998). In tropical forest systems, many of these difficulties are magni-fied by the fact that territories are often larger, nests are often more cryptic (many nests remain undescribed), and nest suc-cess is often far lower than in temperate counterparts (Rob-inson et al. 2000, Jetz et al. 2008), making it challenging to find enough fledglings to study even if one could follow them to the time they eventually acquire their own breeding territory.
Molecular genetic parentage analysis has been used to study natal dispersal in many taxa, including plants (Austerlitz et al. 2004, Robledo-Arnuncio and Gil 2004, Sezen et al. 2005, Hanson et al. 2007), mammals (Peacock and Ray 2001, Telfer et al. 2003, Waser et al. 2006, Taylor et al. 2007), and fishes (Saenz-Agudelo et al. 2009, Christie et al. 2010). Peery et al. (2008) inferred source-sink popu-lation dynamics in an isolated California bird population using molecular parentage data, and their study supports
Journal of Avian Biology 43: 33–42, 2012 doi: 10.1111/j.1600-048X.2011.05572.x
© 2012 The Authors. Journal of Avian Biology © 2012 Nordic Society Oikos Subject Editor: Staffan Bensch. Accepted 29 November 2011

34
the utility of parentage analyses in ecological studies (Peery et al. 2008, Pemberton 2008). To our knowledge, the use of parentage data to estimate natal dispersal distance has not been applied to birds. DeWoody (2005) provides a review of available genetic markers; Jones and Ardren (2003) and Jones et al. (2010) review analytical approaches and soft-ware for parentage analysis. The use of parentage analysis to understand natal dispersal specifically is best applied to spe-cies in which adults are sedentary or largely so, which helps ensure that parent-offspring distances are primarily the result of natal and not adult (breeding dispersal) movements. The ability to group individuals into pre- and post-reproductive classes is also useful.
Self-recruitment (i.e. an individual establishing a terri-tory in its population of origin) is widely discussed among benthic marine population ecologists. In these often patchy environments (e.g. coral reefs), large-scale population con-nectivity was long assumed because many organisms disperse via planktonic larvae, and self-recruitment was thought to be essentially zero (Levin 2006). However, self-recruitment has been found to be more widespread than expected in such communities, resulting from planktonic larvae having sophisticated means of settling in appropriate areas (Jones et al. 1999, Saenz-Agudelo et al. 2009, Christie et al. 2010). Because they can fly, birds are typically considered highly mobile and effective dispersers, with little self-recruitment. Although tropical ecologists have long suspected that some forest understory birds disperse poorly (especially across non-preferred habitats; Willis 1974, Mayr and Diamond 2001), evidence for this has been slow to come and is gen-erally based on behavioral observations, especially of adults (Develey and Stouffer 2001, Laurance 2004, Laurance and Gomez 2005, Boscolo et al. 2008, Lees and Peres 2009). However, such behavioral studies say little if anything about natal dispersal, which usually involves greater distances than breeding dispersal in birds (Newton 2007). In support of poor intrinsic dispersal capability in some tropical birds, recent evidence from Central America demonstrates poor flight capacity in several understory species (Moore et al. 2008, Ibarra-Macias et al. 2011). These different lines of evi-dence indicate that some – perhaps many – species of birds are physically or behaviorally incapable of being the ‘aerial plankton’ that dispersing birds have been assumed to be, and this has important implications for the persistence of popu-lations in anthropogenically fragmented landscapes.
We used genetic parentage analyses in the present study to measure natal dispersal characteristics and to estimate differences in self-recruitment among five populations of a non-migratory Neotropical forest bird, the chestnut-backed antbird Myrmeciza exsul. Previous research on this species has shown that breeding dispersal is minimal (Woltmann and Sherry 2011), indicating that geographical distances separating parent–offspring dyads within breeding popula-tions can be used to estimate natal dispersal distances. We predicted that 1) natal dispersal distances are relatively short (on the order of 1 km – Bates 2002), and 2) self-recruitment is higher in sites surrounded or mostly surrounded by non-forest habitat. Additionally, because female-biased dispersal is common in landbirds (Greenwood and Harvey 1982), and has important implications for overall gene flow, we looked for evidence of sex-biased dispersal in our dataset.
Methods
The chestnut-backed antbird is a medium-sized (28 g) under-story passerine that occurs in lowland forests from Honduras to Ecuador. Pairs hold year-round territories, and individuals only opportunistically forage at army ant (Eciton, Labidus spp.) swarms (Woltmann et al. 2010). Territory size is both small and consistent: mean territory size in forest in Costa Rica is 1.88 ha ( 0.62 SD; Losada-Prado 2009). Based on plumage characteristics, chestnut-backed antbirds can only be aged as either or one year old (Wolfe et al. 2009). This species is amenable to using parentage analysis to esti-mate natal dispersal because 1) individuals on territories are highly sedentary, and territory switches, if they occur, typically involve only 1–2 territory-widths (∼163 m), and 2) individuals 1 yr of age regularly acquire and hold ter-ritories (Woltmann and Sherry 2011). Thus, the geographic distance between a parent and a territorial offspring is a rea-sonable first estimate of natal dispersal distance.
We sampled five populations of chestnut-backed antbird in Heredía Province in northeastern Costa Rica from 2005 to 2010 (Fig. 1). This landscape was entirely forested until the late 1950s, after which it was rapidly deforested to its current level of 75% non-forest (Slud 1960, Sánchez-Azofeifa et al. 1999, Joyce 2006). At the La Selva Biological Reserve (hereafter ‘La Selva’ or ‘LS’) our sampling was mainly in a 300-ha study area, and in some cases we present results restricted to this plot to facilitate comparisons with other sites where we sampled similarly sized or smaller areas. We sampled LS from February–March in 2005–2009; 14 addi-tional samples from 2010 were taken in June and July. Seven
Figure 1. Map of the study area in northeastern Costa Rica showing the location of the five study sites and landscape configuration. Dark gray indicates remaining forest, and light shading indicates non-forest (mainly agriculture and residential areas). The area in black is above 1000 m a.s.l., and beyond the altitudinal range of chestnut-backed antbirds. TI Tirimbina Rainforest Preserve, LS La Selva Biological Reserve, RF Rio Frio, PL Plastico, QG Quebrada Gonzales. The latter two sites are in the Braullio Carrillo National Park.

35
individuals sampled at a site (‘Starke’) 1 km from LS were grouped with LS because there was no indication of genetic differentiation between these sites. We sampled four other sites during single years: Tirimbina (TI; 2006), Plastico (PL, 2007), Quebrada Gonzales (QG, 2009), and Rio Frio (RF, 2009). Both TI (300 ha) and RF (80 ha) are currently sur-rounded by matrix inhospitable to chestnut-backed antbirds (pasture, pineapple and banana plantations). La Selva (1600 ha) is also surrounded on three sides by inhospitable matrix, but is contiguous on its southern border with the Braullio Carrillo National Park (BCNP; 40 000 ha). Both PL and QG are embedded within contiguous forest in BCNP.
Birds were captured in mistnets and individually marked with a numbered aluminum band and three colored plastic leg bands. A blood sample (50–100 mL) was drawn from the brachial vein, stored in Queen’s lysis buffer (Seutin et al. 1991), and birds were released. Geographic coordinates (to within 10 m) of each capture were determined using existing GIS data (LS, TI, PL) or a handheld GPS unit (QG, RF).
Laboratory methods
Whole genomic DNA was extracted from blood with DNEasy kits (Qiagen) and stored at –20°C. We selected 14 microsatel-lite loci developed for chestnut-backed antbird by Barnett et al. (2007) and Feldheim et al. (2010). An additional locus devel-oped for spotted antbird Hylophylax naevioides cross-amplifies in chestnut-backed antbird (HyNa06: forward primer (5′-3′): TTCTTCTCGCTAACCCCGTCTGA, reverse: GCCAAT TT CTGCAGTTTTCGTATC; I. J. Lovette pers. comm.; Supplementary material Appendix 1, Table A1).
PCR conditions were as in Barnett et al. (2007) and Feldheim et al. (2010). PCR conditions for HyNa06 were as in Feldheim et al. (2010), but annealing temperature was 45°C, and we ran 32 cycles. PCR products were visualized with a Li-Cor 4300 DNA Analyzer. Alleles were scored by eye with the aid of sizing standards and GeneImager IR software (Li-Cor). We independently amplified and scored an aver-age of 25% (range 14–50% per locus) of individuals at least twice. Initial estimation of overall genotyping error was 0.03, and all these errors were resolved by amplifying and scoring individuals until a consistent genotype was recorded. Con-sidering individuals not run multiple times, an overall error rate of 0.02 is implied, but because most were rerun due to gel imperfections or loading error, the remaining genotyping error in our dataset is likely 0.02. We found no systematic evidence of linkage disequilibrium among loci (GenePop; Raymond and Rousset 2004). One locus (HyNa06) did not meet HWE in site LS (unadjusted p 0.03), but met HWE in all other sites, and we retained the locus in our analyses (GenePop). No evidence of null alleles was present in any of the 15 loci used (van Oosterhout et al. 2004).
Parentage analyses – empirical data
Our goal was to identify all parent–offspring dyads in our samples. Parentage analyses depend on accurate estimates of background allele frequencies (Marshall et al. 1998, Jones and Ardren 2003), so we first tested the five sampled popula-tions to see if they could be validly pooled for analysis (i.e. to confirm that allele frequencies did not differ significantly
between populations). We used an exact test of population differentiation as implemented in GenePop, using 1000 dememorizations, 500 batches, and 3000 iterations per batch to test significance.
We used ML-Relate (Kalinowski et al. 2006) to identify dyads consistent with various relationships within our data. ML-Relate employs a maximum likelihood approach to categorizing relationships based on a dyad’s occupation in ‘k-space’, where k is the probability of sharing alleles identi-cal by descent. The program assigns each dyad to one of four relationship categories: parent–offspring (PO), full-sib (FS), half-sib (HS), and unrelated (U), and gives the relative likeli-hoods of the other three relationships being the true rela-tionship. Although we were mainly interested in identifying PO dyads, we also examined the program’s ability to identify FS and HS relationships in our data, as these relationships also have implications for dispersal within and between sites (Supplementary material Appendix 1).
Wherever possible, we assigned directionality of a PO dyad (i.e. which individual had dispersed) based on consid-eration of the age of the individuals, or paired status of two putative parents of the same adult-plumaged individual. If two same-sex individuals are identified as PO (and neither individual’s mates were sampled), the sex of the disperser is still known, even if the directionality is not. We measured the geographic distance between individuals (approximate center of the territory) to the nearest 10 m in ArcView 8 (sites LS, PL, TI; ESRI, California) or GoogleEarth (sites QG, RF).
To compare self-recruitment among sites, we calculated a self-recruitment index as (NPO(NPO – 1)/2)/ND*100, where NPO the number of PO dyads detected within a site, and ND the number of dyads analyzed at each site (i.e. N(N–1)/2). When comparing sites, data from LS were limited to individuals sampled within a 300 ha focal plot in order to keep sampling area similar among sites. We tested whether the number of PO dyads detected dif-fered among sites with a G-test, and added a dummy PO dyad to site PL to avoid calculating a logarithm of zero in the observed cell.
Parentage analyses – Monte Carlo simulations
To assess the likelihood of our dataset containing false– positive relationships, we quantified the probability of any one dyad’s relationship arising by chance sharing of alleles (indicating the true parent or offspring was unsampled) using simulated datasets with the same allelic characteristics as our observed data (Supplementary material Appendix 1).
Results
We sampled 255 individuals across the 5 sites. Three nest-lings and two juveniles in the company of adults were removed from the dataset as undispersed young after con-firming that they were the offspring of the accompanying adults. We considered two additional offspring undispersed: a juvenile male sampled on the edge of his father’s territory, and a juvenile female captured in a contiguous neighbor pair’s territory.
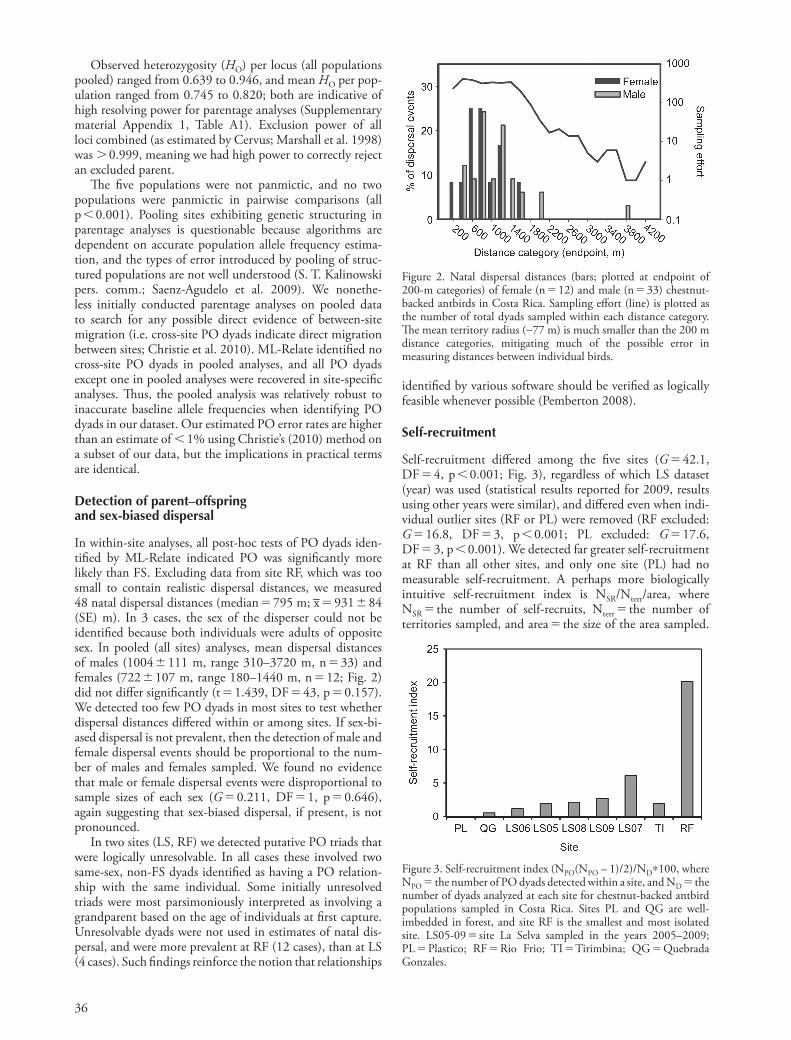
36
identified by various software should be verified as logically feasible whenever possible (Pemberton 2008).
Self-recruitment
Self-recruitment differed among the five sites (G 42.1, DF 4, p 0.001; Fig. 3), regardless of which LS dataset (year) was used (statistical results reported for 2009, results using other years were similar), and differed even when indi-vidual outlier sites (RF or PL) were removed (RF excluded: G 16.8, DF 3, p 0.001; PL excluded: G 17.6, DF 3, p 0.001). We detected far greater self-recruitment at RF than all other sites, and only one site (PL) had no measurable self-recruitment. A perhaps more biologically intuitive self-recruitment index is NSR/Nterr/area, where NSR the number of self-recruits, Nterr the number of territories sampled, and area the size of the area sampled.
Observed heterozygosity (HO) per locus (all populations pooled) ranged from 0.639 to 0.946, and mean HO per pop-ulation ranged from 0.745 to 0.820; both are indicative of high resolving power for parentage analyses (Supplementary material Appendix 1, Table A1). Exclusion power of all loci combined (as estimated by Cervus; Marshall et al. 1998) was 0.999, meaning we had high power to correctly reject an excluded parent.
The five populations were not panmictic, and no two populations were panmictic in pairwise comparisons (all p 0.001). Pooling sites exhibiting genetic structuring in parentage analyses is questionable because algorithms are dependent on accurate population allele frequency estima-tion, and the types of error introduced by pooling of struc-tured populations are not well understood (S. T. Kalinowski pers. comm.; Saenz-Agudelo et al. 2009). We nonethe-less initially conducted parentage analyses on pooled data to search for any possible direct evidence of between-site migration (i.e. cross-site PO dyads indicate direct migration between sites; Christie et al. 2010). ML-Relate identified no cross-site PO dyads in pooled analyses, and all PO dyads except one in pooled analyses were recovered in site-specific analyses. Thus, the pooled analysis was relatively robust to inaccurate baseline allele frequencies when identifying PO dyads in our dataset. Our estimated PO error rates are higher than an estimate of 1% using Christie’s (2010) method on a subset of our data, but the implications in practical terms are identical.
Detection of parent–offspring and sex-biased dispersal
In within-site analyses, all post-hoc tests of PO dyads iden-tified by ML-Relate indicated PO was significantly more likely than FS. Excluding data from site RF, which was too small to contain realistic dispersal distances, we measured 48 natal dispersal distances (median 795 m; x– 931 84 (SE) m). In 3 cases, the sex of the disperser could not be identified because both individuals were adults of opposite sex. In pooled (all sites) analyses, mean dispersal distances of males (1004 111 m, range 310–3720 m, n 33) and females (722 107 m, range 180–1440 m, n 12; Fig. 2) did not differ significantly (t 1.439, DF 43, p 0.157). We detected too few PO dyads in most sites to test whether dispersal distances differed within or among sites. If sex-bi-ased dispersal is not prevalent, then the detection of male and female dispersal events should be proportional to the num-ber of males and females sampled. We found no evidence that male or female dispersal events were disproportional to sample sizes of each sex (G 0.211, DF 1, p 0.646), again suggesting that sex-biased dispersal, if present, is not pronounced.
In two sites (LS, RF) we detected putative PO triads that were logically unresolvable. In all cases these involved two same-sex, non-FS dyads identified as having a PO relation-ship with the same individual. Some initially unresolved triads were most parsimoniously interpreted as involving a grandparent based on the age of individuals at first capture. Unresolvable dyads were not used in estimates of natal dis-persal, and were more prevalent at RF (12 cases), than at LS (4 cases). Such findings reinforce the notion that relationships
Figure 2. Natal dispersal distances (bars; plotted at endpoint of 200-m categories) of female (n 12) and male (n 33) chestnut-backed antbirds in Costa Rica. Sampling effort (line) is plotted as the number of total dyads sampled within each distance category. The mean territory radius (∼77 m) is much smaller than the 200 m distance categories, mitigating much of the possible error in measuring distances between individual birds.
Figure 3. Self-recruitment index (NPO(NPO – 1)/2)/ND*100, where NPO the number of PO dyads detected within a site, and ND the number of dyads analyzed at each site for chestnut-backed antbird populations sampled in Costa Rica. Sites PL and QG are well- imbedded in forest, and site RF is the smallest and most isolated site. LS05-09 site La Selva sampled in the years 2005–2009; PL Plastico; RF Rio Frio; TI Tirimbina; QG Quebrada Gonzales.

37
genetically effective dispersal much greater than what we detected is unlikely because even populations connected by forest and only 14–20 km apart (i.e. LS-PL and PL-QG) were not panmictic. In a North American migratory bird, by comparison, ∼90% of natal dispersal distances of tree swal-lows Tachycineta bicolor were 10 km, but distances of up to 200 km were recorded (Winkler et al. 2005). Moreover, continent-wide genetic structure of tree swallows is evident only at scales of 1000s of km (Stenzler et al. 2009), implying that there is enough long-distance dispersal (natal, breeding or both) in tree swallows to maintain panmixia at vastly larger spatial scales than in chestnut-backed antbird. A more apt temperate comparison is the Florida scrub jay Aphelocoma coerulescens, a highly endangered habitat specialist which has mean natal dispersal distances 10 km, and which dis-plays significant genetic population structure within its small geographical range (Breininger 1999, Coulon et al. 2008). Although too few studies document natal dispersal distances in landbirds to draw broad conclusions, year-round resi-dence may be more important in selecting for relatively short dispersal distances than latitude (Arguedas and Parker 2000, Dingemanse et al. 2003, Wilson et al. 2011). It is important to note, however, that short dispersal distances alone do not necessarily lead to genetic population structure. For example, median dispersal distance of resident house sparrows Passer domesticus in Finland is similar to that of chestnut-backed antbirds (i.e. 1 km), yet house sparrow populations are panmictic across a much larger geographic scale than in our study (Kekkonen et al. 2011).
We detected no PO dyads (self-recruitment 0) in one of the sites (PL) well-embedded in a large expanse of con-tinuous forest, suggesting that dispersal distances at this site are considerably greater than at our other sites. In contrast, the other well-embedded site (QG) did have measurable (but relatively low) self-recruitment (Fig. 3). QG is situated in a narrow, steep valley, and is bordered on one side by a large river (100 m wide). We suspect both features are dispersal barriers to chestnut-backed antbirds. In the Andes, significant genetic structure was found among valley popu-lations of rufous-collared sparrow Zonotrichia capensis, and isolation has led to reproductive asynchrony in populations only 25 km apart, demonstrating that barriers to dispersal can exist on a very local scale, even for widespread habitat generalists (Moore et al. 2005). Indeed, relatively small-scale barriers have had a large influence on tropical avian
The two indices were strongly correlated in our analyses (Pearson r 0.939, p 0.001), and in practice a territory-based index may often be correlated with our dyad-based self-recruitment index.
Parentage analyses – Monte Carlo simulations
Type I error rates (false–positives) in simulated datasets var-ied greatly among relationship types, but varied less within relationship type among the five sites (Table 1). PO dyads were rarely detected in simulations, occurring at rates of 1–2%. We are thus confident that PO dyads identified in our observed data are not the result of chance sharing of alleles.
Discussion
Natal dispersal distances and self-recruitment
Our study is the first we are aware of to explicitly estimate self-recruitment and natal dispersal distances in a bird using genetic parentage analysis. Consistent with our predictions, natal dispersal distances were short, and self-recruitment was measurable at all but one of our sites. In contrast to our expectations, however, we found no evidence of female-biased dispersal. Considering the relatively small sampling areas (generally 300 ha), we found a high degree of self-recruitment in several of our sites, which is surprising given the widespread perception of birds as highly vagile. Mean dispersal distances of chestnut-backed antbird were ∼1 km, and self-recruitment appeared to be higher in sites that were partly or entirely surrounded by non-forest habitats (Fig. 3), suggesting that individuals are hesitant to disperse out of these sites. Despite short dispersal distances, close relatives generally did not occur on adjacent territories, as is frequent in other ecologically similar species (Robinson 2000, Gill and Stutchbury 2010).
Most (∼90%) of the natal dispersal distances we measured were 1.5 km. We likely did not capture the full range of natal dispersal distances, and natal dispersal distances larger than the maximum 3.7 km detected seem probable (Van Houtan et al. 2007, Cooper et al. 2008). As is almost inevi-table in such studies, the potential to detect longer dispersal distances declines with distance (Fig. 2). However, frequent
Table 1. Comparison of observed vs randomly generated (simulated) relationships in chestnut-backed antbird in Costa Rica. Simulations were carried out to determine site-specific type I error rates (i.e. the number of ‘relationships’ in randomized datasets). Simulated numbers are scaled to match sample sizes (number of dyads) analyzed at each site, and are rounded to whole numbers. Error (%) was calculated from non-rounded numbers. See Supplementary material Appendix 1 for details of the simulations.
Observed/simulated Error (%)
Site Parent–offspring Full-sib Half-sib Parent–offspring Full-sib Half-sib
LS all 43/1 40/10 905/800 2 30 110LS (300 ha) 32/0 15/5 336/417 1 45 167PL 0/0 0/1 26/45 – – 181QG 2/0 1/0 9/21 2 31 211RF 16/0 9/1 55/75 2 18 132TI 5/0 3/1 37/60 1 43 157All pooled 66/1 99/33 2993/2916 2 34 97

38
an individual switches to another territory is low (0.04 for males, 0.09 for females; Woltmann and Sherry 2011), which minimizes error associated with measuring distances between territories as a proxy for natal dispersal distance. In situations where breeding dispersal is not well understood, the finding of PO dyads within a study area is still indicative of short overall dispersal distances, but more caution is warranted in drawing inferences about natal vs. breeding dispersal dis-tances. As in nearly all studies of dispersal, the challenge of adequately sampling large areas to detect rare large dispersal distances can be daunting (Koenig et al. 1996). Only in rare cases (e.g. islands) can it be claimed that the entire area of potential dispersal has been sampled, and that the true dis-persal curve has been described. However, our data on short dispersal distances combined with lack of panmixia among our populations indicates that effective genetic dispersal is 14 km, and probably less, for this species in this land-scape.
Mating systems will influence the applicability and inter-pretation of parentage analyses in the study of natal dispersal. For example, extra-pair fertilization (EPF) and egg-dumping will complicate interpretations of natal dispersal. If egg-dumping is prevalent (as in some ducks), measuring natal dispersal distances will be difficult, if not impossible, using these methods, but estimates of self-recruitment would still be generally valid. Socially monogamous species (like chestnut-backed antbird) with 2-egg clutches and extended parental care are predicted (and in some cases confirmed) to be genetically monogamous (Fleischer et al. 1997, Stutch-bury and Morton 2001, Taylor et al. 2008), and no egg-dumping has been reported in thamnophilids or ecologically similar species.
The confidence with which PO and other relationships can be assigned has strong implications regarding conclu-sions drawn from any particular study, yet quantification (or even simple description) of various sources of genotyping and assignment error remain too infrequent in the literature (Hoffman and Amos 2005, Jones et al. 2010). Genotyping (scoring) errors are relatively easy to quantify and correct, whereas other types of error can often only be estimated with simulations. FS is the relationship most likely to be con-founded with PO, and we addressed this by using likelihood ratio tests on all PO dyads to confirm initial assignments made by the software (Blouin et al. 1996, Berg et al. 2009). Given our poor ability to identify relationships beyond parent–offspring, despite a relatively large number of micro-satellite loci, caution in simply accepting the other relation-ship assignments produced by ML-Relate is warranted.
Confidence of parentage assignments must also be evalu-ated on a site-by-site basis if samples come from genetically differentiated populations, because the power of the genetic markers may vary by site. The highest proportion of ran-domly generated ‘PO’ dyads in our simulations were gen-erated with allele frequencies of those at the smallest and most isolated site (RF). This site also contained the largest number of logically unresolvable ‘PO’ dyads identified by ML-Relate. Because the marker set overall had considerable power to correctly identify PO relationships, we infer that the high number of relatives of varying degrees at RF, as well as the limited number of relationship types that ML-Relate can search for (and reliably identify) made disentangling
diversity (Haffer 1969, Capparella 1988), and understory birds are particularly sensitive to such barriers (Burney and Brumfield 2009).
We found no evidence of sex-biased dispersal in chestnut-backed antbird, either based on mean dispersal distances, or the proportion of dispersal events detected by sex. Although female-biased dispersal is predominant among birds (Green-wood and Harvey 1982, Pusey 1987), few resident tropi-cal species have been examined. It is possible that the small sample size of female dispersal events provided insufficient power to detect differences in mean dispersal distance, but we note that mean dispersal distances tended to be greater for males than females in chestnut-backed antbird (Fig. 2), although not significantly so. We also found no evidence of sex-biased dispersal in a landscape-scale analysis of popula-tion structure in chestnut-backed antbird (Woltmann 2010, Woltmann et al. in press), although the bias in dispersal dis-tances may need to be quite strong to be detected via micro-satellite analyses (Goudet et al. 2002).
Several consequences of high self-recruitment can be pre-dicted from our study. First, high local self-recruitment could lead to fine-scale genetic population structure, even in the absence of obvious barriers to dispersal (Coltman et al. 2003, Moore et al. 2005, Cabe et al. 2007). Second, elevated self-recruitment in isolated populations could decrease genetic diversity over time (Epps et al. 2005, Boessenkool et al. 2007; but see Tarr et al. 1998). In the long-term, such populations may eventually become inbred (Bensch et al. 1994), contrib-uting to a high risk of local extirpation, assuming such popu-lations have not already been extirpated for other reasons, such as demographic stochasticity (Frankham 1998, 2005). In contrast, short dispersal distances may increase the speed at which populations can become locally adapted to particu-lar ecological conditions, as has been demonstrated in Mexi-can jay Aphelocoma ultramarina (McCormack and Smith 2008). We note that the sometimes striking local genetic structure found in some cooperatively or lek breeding spe-cies (Shorey et al. 2000) is a consequence of self-recruitment, although some of this local differentiation should in theory be counteracted by dispersal of the non-self-recruiting sex (Coulon et al. 2008). Additionally, high self-recruitment and local population genetic structure does not necessarily lead to detection of inbreeding, especially when populations are large or when the genetic markers are chosen for their inher-ent variability (e.g. because of high allelic richness, micro-satellites are not well-suited for detecting subtle evidence of inbreeding).
The use of genetic parentage analysis to study natal dispersal
Given that the majority of bird species are year-round tropi-cal residents, the use of genetic parentage analysis to under-stand natal dispersal of tropical understory birds has great potential, but its utility must be carefully evaluated for each species. It is critically important to understand breeding dispersal to some extent. In chestnut-backed antbird, territo-ries are consistently small (2 ha or less; Losada-Prado 2009), movements of territory holders are infrequent, and largely limited to movements of 1–2 territory-widths (Woltmann and Sherry 2011). Additionally, the annual likelihood that

39
conservation of tropical forest bird populations in increas-ingly fragmented landscapes. To the extent that our method of quantifying natal dispersal can be applied in broad avian comparative studies, it also opens up further possibilities to link dispersal capability in birds with their ecology (Burney and Brumfield 2009), morphology (Desrochers 2010), and life-history traits.
Acknowledgements – We thank I. J. Lovette and J. R. Barnett (Cornell Univ.), and K. A. Feldheim (The Field Museum) for assis-tance with microsatellite development, D. McClearn, J. Hurtado, E. Castro and D. L. Roberts for logistical assistance, M. L. Brady, M. F. Cashner, R. S. Terrill and J. Wielfaert for assistance in the field, and E. Robakis for lab assistance. A. Boyle provided two sam-ples from Plastico, and D. M. Visco collected all samples from La Selva in 2010. We thank K. Pillar (Southeastern Louisiana Univ.) and H. L. Bart (Tulane Univ.) for use of their lab facilities. Funding for this study came from Sigma Xi, the Stone Center for Latin American Studies (Tulane), The Organization for Tropical Studies, the Cooper Ornithological Society, the American Ornithologists’ Union, the Gunning Fund (Tulane), the American Museum of Natural History, and the Louisiana Experimental Program to Stim-ulate Competitive Research (EPSCoR) funded by the National Sci-ence Foundation and the Board of Regents Support Fund. We thank MINAE for permission to conduct this work in Costa Rica, and thank the land managers at Tirimbina and Rio Frio for allow-ing us to work on their properties. The comments of L. Lens and W. Cresswell improved this manuscript. This study was conducted under Tulane Univ. IACUC protocol no. 0298R-UT-C.
References
Arcese, P., Smith, J. N. M., Hochachka, W. M., Rogers, C. M. and Ludwig, D. 1992. Stability, regulation, and the determination of abundance in an insular song sparrow population. – Ecology 73: 805–822.
Arguedas, N. and Parker, P. G. 2000. Seasonal migration and genetic population structure in house wrens. – Condor 102: 517–528.
Austerlitz, F., Dick, C. W., Dutech, C., Klein, E. K., Oddou- Muratorio, S., Smouse, P. E. and Sork, V. L. 2004. Using genetic markers to estimate the pollen dispersal curve. – Mol. Ecol. 13: 937–954.
Barnett, J. R., Woltmann, S., Bogdanowicz, S. M. and Lovette, I. J. 2007. Isolation and characterization of microsatellite markers from the chestnut-backed antbird, Myrmeciza exsul. – Mol. Ecol. Not. 7: 1070–1072.
Barton, N. H. 2001. The evolutionary consequences of gene flow and local adaptation: future approaches. – In: Clobert, J., Danchin, E., Dhondt, A. A. and Nichols, J. D. (eds), Disper-sal. Oxford Univ. Press, pp. 329–340.
Bates, J. M. 2002. The genetic effects of forest fragmentation on five species of Amazonian birds. – J. Avian Biol. 33: 276–294.
Bensch, S., Hasselquist, D. and von Schantz, T. 1994. Genetic similarity between parents predicts hatching failure: noninces-tuous inbreeding in the great reed warbler. – Evolution 48: 317–326.
Berg, E. C., Eadie, J. M., Langen, T. A. and Russell, A. F. 2009. Reverse sex-biased philopatry in a cooperative bird: genetic con-sequences and a social cause. – Mol. Ecol. 18: 3486–3499.
Birdlife International. 2000. Threatened birds of the world. – Lynx Edicions.
Blouin, M. S., Parsons, M., Lacaille, V. and Lotz, S. 1996. Use of microsatellite loci to classify individuals by relatedness. – Mol. Ecol. 5: 393–401.
relationships within RF more difficult than in the other sites. In the context of our study, the reduced resolving power of the data at RF further supports the idea that individuals of this species are reluctant to disperse out of sites surrounded by inhospitable matrix.
Conservation considerations
Understanding dispersal tendencies of organisms targeted for conservation is critical to the effective design of reserves (Palumbi 2003, Shanks et al. 2003), yet dispersal remains one of the most poorly known aspects of tropical bird biol-ogy. The fact that the majority of globally threatened bird species are year-round residents of lowland tropical forests (BirdLife International 2000) that share many life-history characteristics with chestnut-backed antbird (Stutchbury and Morton 2001) makes understanding tropical forest bird dispersal behavior even more critical. As short as natal dispersal distances of chestnut-backed antbirds were in our study, they are still generally greater than distances measured via mark-recapture in song wrens Cyphorinus phaeocepha-lus (Robinson 2000) and buff-throated wrens Cantorchilus leucotis (Gill 2003) in Panama, both in terms of absolute dis-tances and territory-widths, suggesting limited dispersal in chestnut-backed antbirds may not be unusual among tropi-cal forest understory insectivores.
Elevated self-recruitment of chestnut-backed antbirds at three of our sites is cause for concern from conservation and management perspectives, and the case of the chestnut-backed antbird illuminates potential dangers of relying on untested assumptions regarding population persistence. The species is unusual for a tropical forest-dependent species in its widespread occurrence in fragmented landscapes, and its apparent ability to persist in even small or very isolated forest fragments (Wetmore 1972, Roberts 2007, Woltmann et al. 2010). It was logical to presume in the past that chestnut-backed antbirds are better able to disperse than other antbirds, and thus persist in fragmented landscapes (i.e. as a classic metapopulation) where many ecologically similar species do not. However, chestnut-backed antbirds have poor physical dispersal abilities (Moore et al. 2008), and naturally short dispersal distances (Woltmann and Sherry 2011; this study), suggesting that many isolated popula-tions in fragmented landscapes may be (at least temporarily) persisting in the absence of significant immigration, i.e. per-sisting as non-equilibrium metapopulations. To the extent that populations of this, and other ecologically similar species are indeed isolated on small habitat fragments (e.g. the population of chestnut-backed antbird at the Rio Frio site), as our data suggest, their risk of local extirpation increases due to known risk factors associated with small populations (e.g. inbreeding, genetic drift, demographic stochasticity).
Additional research is needed to better understand mech-anisms that regulate fragmented populations in tropical landscapes. In particular, understanding dispersal behavior in large, unfragmented contexts is a challenging but critical need, and the method we have applied successfully to the chestnut-backed antbird could be applied to many tropical and other non-migratory landbirds. Understanding intrin-sically weak (local) dispersal ability and self- recruitment in particular birds will be necessary for the effective

40
Fleischer, R. C., Tarr, C. L., Morton, E. S., Sangmeister, A. and Derrickson, K. C. 1997. Mating system of the dusky antbird, a tropical passerine, as assessed by DNA fingerprinting. – Condor 99: 512–514.
Frankham, R. 1998. Inbreeding and extinction: island populations. – Conserv. Biol. 12: 665–675.
Frankham, R. 2005. Genetics and extinction. – Biol. Conserv. 126: 131–140.
Gill, S. A. 2003. Territory acquisition, mate fidelity, pair formation, and parental care in a Neotropical wren, Thryothorus leucotis. – PhD thesis, York Univ., Toronto.
Gill, S. A. and Stutchbury, B. J. M. 2010. Delayed dispersal and territory acquisition in Neotropical buff-breasted wrens (Thryothorus leucotis). – Auk 127: 372–378.
Goudet, J., Perrin, N. and Waser, P. M. 2002. Tests for sex-biased dispersal using bi-parentally inherited genetic markers. – Mol. Ecol. 11: 1103–1114.
Greenwood, P. J. and Harvey, P. H. 1982. The natal and breeding dispersal of birds. – Annu. Rev. Ecol. Evol. Syst. 13: 1–21.
Haffer, J. 1969. Speciation in Amazonian forest birds. – Science 165: 131–137.
Hanson, T., Brunsfeld, S., Finegan, B. and Waits, L. P. 2007. Con-ventional and genetic measures of seed dispersal for Dipteryx panamensis (Fabaceae) in continuous and fragmented Costa Rican rain forest. – J. Trop. Ecol. 23: 635–642.
Heinz, S. K. and Strand, E. 2006. Adaptive patch searching strategies in fragmented landscapes. – Evol. Ecol. 20: 113–130.
Hoffman, J. I. and Amos, W. 2005. Microsatellite genotyping errors: detection approaches, common sources and consequ-ences for paternal exclusion. – Mol. Ecol. 14: 599–612.
Ibarra-Macias, A., Robinson, W. D. and Gaines, M. S. 2011. Experimental evaluation of bird movements in a fragmented Neotropical landscape. – Biol. Conserv. 144: 703–712.
Jetz, W., Sekercioglu, C. H. and Bo, K. 2008. The worldwide variation in avian clutch size across species and space. – PLoS Biol. 6: 2650–2657.
Johnson, M. L. and Gaines, M. S. 1990. Evolution of dispersal: theoretical models and empirical tests using birds and mam-mals. – Annu. Rev. Ecol. Syst. 21: 449–480.
Jones, A. G. and Ardren, W. R. 2003. Methods of parentage anal-ysis in natural populations. – Mol. Ecol. 12: 2511–2523.
Jones, A. G., Small, C. M., Paczolt, K. A. and Ratterman, N. L. 2010. A practical guide to methods of parentage analysis. – Mol. Ecol. Res. 10: 6–30.
Jones, G. P., Milicich, M. J., Emslie, M. J. and Lunow, C. 1999. Self-recruitment in a coral reef fish population. – Nature 402: 802–804.
Joyce, A. T. 2006. Land use change in Costa Rica. – Litografía e Imprenta LIL, S.A., San José, Costa Rica.
Kalinowski, S. T., Wagner, A. P. and Taper, M. L. 2006. ML-Relate: a computer program for maximum likelihood estima-tion of relatedness and relationship. – Mol. Ecol. Not. 6: 576–579.
Kekkonen, J., Seppä, P., Hanski, I. K., Jensen, H., Väisänen, R. A. and Brommer, J. E. 2011. Low genetic differentiation in a sedentary bird: house sparrow population genetics in a con-tiguous landscape. – Heredity 106: 183–190.
Koenig, W. D., Van Vuren, D. and Hooge, P. N. 1996. Detectabil-ity, philopatry, and the distribution of dispersal distances in vertebrates. – Trends Ecol. Evol. 11: 514–517.
Laurance, S. G. W. 2004. Responses of understory rain forest birds to road edges in central Amazonia. – Ecol. Appl. 14: 1344–1357.
Laurance, S. G. W. and Gomez, M. S. 2005. Clearing width and movements of understory rainforest birds. – Biotropica 37: 149–152.
Boessenkool, S., Taylor, S. S., Tepolt, C. K., Komdeur, J. and Jamieson, I. G. 2007. Large mainland populations of South Island robins retain greater genetic diversity than offshore island refuges. – Conserv. Genet. 8: 705–714.
Bohonak, A. J. 1999. Dispersal, gene flow, and population struc-ture. – Q. Rev. Biol. 74: 21–45.
Boscolo, D., Candia-Gallardo, C., Awade, M. and Metzger, J. P. 2008. Importance of interhabitat gaps and stepping-stones for lesser woodcreepers (Xiphorhynchus fuscus) in the Atlantic Forest, Brazil. – Biotropica 40: 273–276.
Brawn, J. D., Robinson, S. K., Stotz, D. F. and Robinson, W. D. 1998. Research needs for the conservation of Neotropical birds. – In: Marzluff, J. M. and Sallabanks, R. (eds), Avian conserva-tion: research and management. Island Press, pp. 323–335.
Breininger, D. R. 1999. Florida scrub-jay demography and disper-sal in a fragmented landscape. – Auk 116: 520–527.
Burney, C. W. and Brumfield, R. T. 2009. Ecology predicts levels of genetic differentiation in Neotropical birds. – Am. Nat. 174: 358–368.
Cabe, P. R., Page, R. B., Hanlon, T. J., Aldrich, M. E., Connors, L. and Marsh, D. M. 2007. Fine-scale population differentiation and gene flow in a terrestrial salamander (Plethodon cinereus) living in continuous habitat. – Heredity 98: 53–60.
Capparella, A. P. 1988. Neotropical avian diversity and riverine barriers. – Acta XIX Congressus Internationalis Ornithologici, pp. 307–316.
Chetkiewicz, C.-L. B., St Clair, C. C. and Boyce, M. S. 2006. Corridors for conservation: integrating pattern and process. – Annu. Rev. Ecol. Evol. Syst. 37: 317–342.
Christie, M. R. 2010. Parentage in natural populations: novel methods to detect parent-offspring pairs in large data sets. – Mol. Ecol. Res. 10: 115–128.
Christie, M. R., Johnson, D. W., Stallings, C. D. and Hixon, M. A. 2010. Self-recruitment and sweepstakes reproduction amid extensive gene-flow in a coral reef fish. – Mol. Ecol. 19: 1042–1057.
Coltman, D. W., Pilkington, J. G. and Pemberton, J. M. 2003. Fine-scale genetic structure in a free-living ungulate popula-tion. – Mol. Ecol. 12: 733–742.
Cooper, C. B., Daniels, S. J. and Walters, J. R. 2008. Can we improve estimates of juvenile dispersal distance and survival? – Ecology 89: 3349–3361.
Coulon, A., Fitzpatrick, J. W., Bowman, R., Stith, B. M., Makare-wich, C. A., Stenzler, L. M. and Lovette, I. J. 2008. Congruent population structure inferred from dispersal behaviour and intensive genetic surveys of the threatened Florida scrub-jay (Aphelocoma coerulescens). – Mol. Ecol. 17: 1685–1701.
Desrochers, A. 2010. Morphological response of songbirds to 100 years of landscape change in North America. – Ecology 91: 1577–1582.
Develey, P. F. and Stouffer, P. C. 2001. Effects of roads on move-ments by understory birds in mixed-species flocks in central Amazonian Brazil. – Conserv. Biol. 15: 1416–1422.
DeWoody, J. A. 2005. Molecular approaches to the study of par-entage, relatedness, and fitness: practical applications for wild animals. – J. Wildl. Manage. 69: 1400–1418.
Dingemanse, N. J., Both, C., Van Noordwijk, A. J., Rutten, A. L. and Drent, P. J. 2003. Natal dispersal and personalities in great tits (Parus major). – Proc. R. Soc. B 270: 741–747.
Epps, C. W., Palsboll, P. J., Wehausen, J. D., Roderick, G. K., Ramey, R. R. and Mccullough, D. R. 2005. Highways block gene flow and cause a rapid decline in genetic diversity of desert bighorn sheep. – Ecol. Lett. 8: 1029–1038.
Feldheim, K. A., Woltmann, S. and Kreiser, B. R. 2010. Charac-terization of 20 additional microsatellite loci for Myrmeciza exsul (Aves: Thamnophilidae: chestnut-backed antbird). – Mol. Ecol. Res. 10: 404–408.

41
Sezen, U. U., Chazdon, R. L. and Holsinger, K. E. 2005. Genetic consequences of tropical second-growth forest regeneration. – Science 307: 891.
Shanks, A. L., Grantham, B. A. and Carr, M. H. 2003. Propagule dispersal distance and the size and spacing of marine reserves. – Ecol. Appl. 13: S159–S169.
Shorey, L., Piertney, S., Stone, J. and Höglund, J. 2000. Fine-scale genetic structuring on Manacus manacus leks. – Nature 408: 352–353.
Slud, P. 1960. The birds of finca ‘La Selva’, Costa Rica: a tropical wet forest locality. – Bull. Am. Mus. Nat. Hist. 121: 49–148.
Stenzler, L. M., Makarewich, C. A., Coulon, A., Ardia, D. R., Lovette, I. J. and Winkler, D. W. 2009. Subtle edge-of-range genetic structuring in transcontinentally distributed North American tree swallows. – Condor 111: 470–478.
Stutchbury, B. J. M. and Morton, E. S. 2001. Behavioral ecology of tropical birds. – Academic Press.
Tarr, C. L., Conant, S. and Fleischer, R. C. 1998. Founder events and variation at microsatellite loci in an insular passerine bird, the laysan finch (Telespiza cantans). – Mol. Ecol. 7: 719–731.
Taylor, A. C., Tyndale-Biscoe, H. and Lindenmayer, D. B. 2007. Unexpected persistence on habitat islands: genetic signatures reveal dispersal of a eucalypt-dependent marsupial through a hostile pine matrix. – Mol. Ecol. 16: 2655–2666.
Taylor, S. S., Boessenkool, S. and Jamieson, I. G. 2008. Genetic monogamy in two long-lived New Zealand passerines. – J. Avian Biol. 39: 579–583.
Telfer, S., Piertney, S. B., Dallas, J. F., Stewart, W. A., Marshall, F., Gow, J. L. and Lambin, X. 2003. Parentage assignment detects frequent and large-scale dispersal in water voles. – Mol. Ecol. 12: 1939–1949.
Turchin, P. 1998. Quantitative analysis of movement – Sinauer.Van Houtan, K. S., Pimm, S. L., Halley, J. M., Bierregaard Jr,
R. O. and Lovejoy, T. E. 2007. Dispersal of Amazonian birds in continuous and fragmented forest. – Ecol. Lett. 10: 219–229.
van Oosterhout, C., Hutchinson, W. F., Wills, D. P. M. and Shipley, P. 2004. MICRO-CHECKER: software for identi-fying and correcting genotyping errors in microsatellite data. – Mol. Ecol. Not. 4: 535–538.
Verhulst, S., Perrins, C. M. and Riddington, R. 1997. Natal dis-persal of great tits in a patchy environment. – Ecology 78: 864–872.
Waser, P. M., Busch, J. D., Mccormick, C. R. and Dewoody, J. A. 2006. Parentage analysis detects cryptic precapture dispersal in a philopatric rodent. – Mol. Ecol. 15: 1929–1937.
Wetmore, A. 1972. The birds of the Republic of Panama, part 3. – Smithsonian Inst. Press.
Wheelwright, N. T. and Mauck, R. A. 1998. Philopatry, natal dis-persal, and inbreeding avoidance in an island population of savannah sparrows. – Ecology 79: 755–767.
Willis, E. O. 1974. Populations and local extinctions of birds on Barro Colorado Island, Panama. – Ecol. Monogr. 44: 153–169.
Wilson, A. G., Arcese, P., Chan, Y. L. and Patten, M. A. 2011. Micro-spatial genetic structure in song sparrows (Melospiza melodia). – Conserv. Genet. 12: 213–222.
Winkler, D. W., Wrege, P. H., Allen, P. E., Kast, T. L., Senesac, P., Wasson, M. F. and Sullivan, P. J. 2005. The natal dispersal of tree swallows in a continuous mainland environment. – J. Anim. Ecol. 74: 1080–1090.
Wolfe, J. D., Pyle, P. and Ralph, C. J. 2009. Breeding seasons, molt patterns, and gender and age criteria for selected northeastern Costa Rican resident landbirds. – Wilson J. Ornithol. 121: 556–567.
Woltmann, S. 2010. Dispersal of a tropical rainforest understory bird, the chestnut-backed antbird (Myrmeciza exsul). – PhD thesis, Tulane Univ. New Orleans, LA, USA.
Woltmann, S. and Sherry, T. W. 2011. High apparent survival and stable territory dynamics of chestnut-backed antbird
Lees, A. C. and Peres, C. A. 2009. Gap-crossing movements predict species occupancy in Amazonian forest fragments. – Oikos 118: 280–290.
Levin, L. A. 2006. Recent progress in understanding larval disper-sal: new directions and digressions. – Integr. Comp. Biol. 46: 282–297.
Losada-Prado, S. 2009. Home range and movements of Myrmeciza exsul (Aves: Thamnophilidae) in two fragmented landscapes in Costa Rica: evaluating functional connectivity. – PhD thesis, CATIE, Turrialba, Costa Rica.
Marshall, T. C., Slate, J., Kruuk, L. E. B. and Pemberton, J. M. 1998. Statistical confidence for likelihood-based paternity inference in natural populations. – Mol. Ecol. 7: 639–655.
Mayr, E. and Diamond, J. 2001. The birds of northern Melanesia: speciation, ecology and biogeography. – Oxford Univ. Press.
Mccormack, J. E. and Smith, T. B. 2008. Niche expansion leads to small-scale adaptive divergence along an elevation gradient in a medium-sized passerine bird. – Proc. R. Soc. B 275: 2155–2164.
Moore, I. T., Bonier, F. and Wingfield, J. C. 2005. Reproductive asynchrony and population divergence between two tropical bird populations. – Behav. Ecol. 16: 755–762.
Moore, R. P., Robinson, W. D., Lovette, I. J. and Robinson, T. R. 2008. Experimental evidence for extreme dispersal limitation in tropical forest birds. – Ecol. Lett. 11: 960–968.
Newton, I. 2007. The migration ecology of birds. – Academic Press.Palumbi, S. R. 2003. Population genetics, demographic connectiv-
ity, and the design of marine reserves. – Ecol. Appl. 13: S146–S158.
Peacock, M. M. and Ray, C. 2001. Dispersal in pikas (Ochotona princeps): combining genetic and demographic approaches to reveal spatial and temporal patterns. – In: Clobert, J., Danchin, E., Dhondt, A. A. and Nichols, J. D. (eds), Dispersal. Oxford Univ. Press, pp. 43–56.
Peery, M. Z., Beissinger, S. R., House, R. F., Bérubé, M., Hall, L. A., Sellas, A. and Palsbøll, P. J. 2008. Characterizing source-sink dynamics with genetic parentage assignments. – Ecology 89: 2746–2759.
Pemberton, J. M. 2008. Wild pedigrees: the way forward. – Proc. R. Soc. B 275: 613–621.
Pusey, A. E. 1987. Sex-biased dispersal and inbreeding avoidance in birds and mammals. – Trends Ecol. Evol. 2: 295–299.
Raymond, M. and Rousset, F. 2004. GenePop on the Web ver. 3.4. – http://wbiomed.curtin.edu.au/genepop/ .
Roberts, D. L. 2007. Effects of tropical forest fragmentation on ecol-ogy and conservation of migrant and resident birds in lowland Costa Rica. – PhD thesis, Univ. of Idaho, Moscow, ID, USA.
Robinson, T. R. 2000. Factors affecting natal dispersal by song wrens (Cyphorhinus phaeocephalus): ecological constraints and demography. – PhD thesis, Univ. of Illinois at Urbana- Champagne, IL, USA.
Robinson, W. D., Robinson, T. R., Robinson, S. K. and Brawn, J. D. 2000. Nesting success of understory forest birds in central Panama. – J. Avian Biol. 31: 151–164.
Robledo-Arnuncio, J. J. and Gil, L. 2004. Patterns of pollen dis-persal in a small population of Pinus sylvestris L. revealed by total-exclusion paternity analysis. – Heredity 94: 13–22.
Saenz-Agudelo, P., Jones, G. P., Thorrold, S. R. and Planes, S. 2009. Estimating connectivity in marine populations: an empirical evaluation of assignment tests and parentage analysis under different gene flow scenarios. – Mol. Ecol. 18: 1765–1776.
Sánchez-Azofeifa, A., Quesada-Mateo, C., Gonzalez-Quesada, P., Dayanandan, S. and Bawa, K. S. 1999. Protected areas and conservation of biodiversity in the tropics. – Conserv. Biol. 13: 407–411.
Seutin, G., White, B. N. and Boag, P. T. 1991. Preservation of avian blood and tissue samples for DNA analysis. – Can. J. Zool. 69: 82–90.

42
Woltmann, S., Kreiser, B. R. and Sherry, T. W. in press. Fine-scale genetic population structure of an understory rainforest bird in Costa Rica. – Conserv. Genet.
Woolfenden, G. E. and Fitzpatrick, J. W. 1989. Sexual asymmetries in the life history of the Florida scrub jay. – In: Rubenstein, D. I. and Wrangham, R. W. (eds), Ecological aspects of social evolu-tion: birds and mammals. Princeton Univ. Press, pp. 87–107.
(Myrmeciza exsul) in a large rain forest preserve. – Wilson J. Ornithol. 123: 15–23.
Woltmann, S., Terrill, R. S., Miller, M. J. and Brady, M. L. 2010. Chestnut-backed antbird (Myrmeciza exsul). – In: Schulenberg, T. S. (ed.), Neotropical birds online. Cornell Univ., http://neotropical.birds.cornell.edu/portal/species/overview?p_p_spp 393811 .
Supplementary material (Appendix J5572 at www. oikosoffice.lu.se/appendix ). Appendix 1.




















