
A First Quantitative Census of Vascular Epiphytes in Rain forests of Colombian Amazonia
20
-1 A first quantitative census of vascular epiphytes in rain forests of Colombian Amazonia ANA MARI ´ A BENAVIDES D. 1,3 , ALVARO J. DUQUE M. 2,3,* , JOOST F. DUIVENVOORDEN 2 , G. ALEJANDRA VASCO 1,3 and RICARDO CALLEJAS 1 1 Instituto de Biologı´a, Universidad de Antioquia; 2 Institute for Biodiversity and Ecosystem Dynamics (IBED), Centre for Geo-Ecological Research (ICG), Universiteit van Amsterdam, P.O. Box 94062, 1090 GB Amsterdam, The Netherlands; 3 Tropenbos-Colombia Foundation; *Author for correspon- dence (e-mail: [email protected]) Received 20 May 2003; accepted in revised form 11 November 2003 Key words: Detrended correspondence analysis, Hemi-epiphytes, Holo-epiphytes, Landscape, Mantel test, Phorophyte Abstract. Epiphytism in Colombian Amazonia was described by counting vascular epiphytes in thirty 0.025-ha (5 · 50 m) plots, well-distributed over the main landscape units in the middle Caqueta´ area of Colombian Amazonia. Each plot was directly adjacent to a 0.1-ha plot at which the species composition of trees and lianas (diameter at breast height (DBH) ‡ 2.5 cm) had been recorded 3 years earlier. The purpose of the study was to explore abundance, diversity, and dis- tribution of epiphytes between the principal landscape units. A total of 6129 individual vascular epiphytes were recorded belonging to 27 families, 73 genera, and 213 species (which included 59 morpho-species). Araceae, Orchidaceae, and Bromeliaceae were the most speciose and abundant families. A total of 2763 phorophytes were registered, 1701 (62%) of which with DBH ‡ 2.5 cm. About 40–60% of the woody plants with DBH ‡ 2.5 cm carried epiphytes, which points at low phorophyte limitation throughout all landscapes. Epiphytism was concentrated on stem bases. Just as trees, epiphyte species assemblages were well associated with the main landscapes. Contrary to trees, however, epiphyte abundance and diversity (species richness, Fisher’s alpha index) hardly differed between the landscapes. This calls for caution when explanations for distribution and dynamics of tree species are extrapolated to growth forms with a totally different ecology. Introduction Northwestern Amazonia has been recognized as a region with high tree diversity (Valencia et al. 1994), but also where the epiphyte communities exhibit high abundance and diversity (Gentry and Dodson 1987b; Nieder et al. 2001). In the past decades, most studies carried out on vascular plants have focused on the tree component, despite the fact that the non-tree vegetation is responsible for a high percentage of the total diversity in the tropical forests (Gentry and Dodson 1987a; Galeano et al. 1998; Schnitzer and Carson 2000). Epiphytes are plants that inhabit a discontinuous and three-dimensional landscape, directly in contact with the forest soil or not (Bennett 1986). Pat- terns of distribution and floristic composition of epiphytic plants have been related to factors of dispersal (Benzing 1986; Wolf 1993), humidity and soils Biodiversity and Conservation 14: 739–758, 2005. Ó Springer 2005 DOI 10.1007/s10531-004-3920-9
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of A First Quantitative Census of Vascular Epiphytes in Rain forests of Colombian Amazonia

-1
A first quantitative census of vascular epiphytes
in rain forests of Colombian Amazonia
ANA MARIA BENAVIDES D.1,3, ALVARO J. DUQUE M.2,3,*,JOOST F. DUIVENVOORDEN2, G. ALEJANDRA VASCO1,3 andRICARDO CALLEJAS11Instituto de Biologıa, Universidad de Antioquia; 2Institute for Biodiversity and Ecosystem Dynamics
(IBED), Centre for Geo-Ecological Research (ICG), Universiteit van Amsterdam, P.O. Box 94062,
1090 GB Amsterdam, The Netherlands; 3Tropenbos-Colombia Foundation; *Author for correspon-
dence (e-mail: [email protected])
Received 20 May 2003; accepted in revised form 11 November 2003
Key words: Detrended correspondence analysis, Hemi-epiphytes, Holo-epiphytes, Landscape,
Mantel test, Phorophyte
Abstract. Epiphytism in Colombian Amazonia was described by counting vascular epiphytes in
thirty 0.025-ha (5 · 50 m) plots, well-distributed over the main landscape units in the middle
Caqueta area of Colombian Amazonia. Each plot was directly adjacent to a 0.1-ha plot at which
the species composition of trees and lianas (diameter at breast height (DBH) ‡ 2.5 cm) had been
recorded 3 years earlier. The purpose of the study was to explore abundance, diversity, and dis-
tribution of epiphytes between the principal landscape units. A total of 6129 individual vascular
epiphytes were recorded belonging to 27 families, 73 genera, and 213 species (which included 59
morpho-species). Araceae, Orchidaceae, and Bromeliaceae were the most speciose and abundant
families. A total of 2763 phorophytes were registered, 1701 (62%) of which with DBH ‡ 2.5 cm.
About 40–60% of the woody plants with DBH ‡ 2.5 cm carried epiphytes, which points at low
phorophyte limitation throughout all landscapes. Epiphytism was concentrated on stem bases. Just
as trees, epiphyte species assemblages were well associated with the main landscapes. Contrary to
trees, however, epiphyte abundance and diversity (species richness, Fisher’s alpha index) hardly
differed between the landscapes. This calls for caution when explanations for distribution and
dynamics of tree species are extrapolated to growth forms with a totally different ecology.
Introduction
Northwestern Amazonia has been recognized as a region with high treediversity (Valencia et al. 1994), but also where the epiphyte communitiesexhibit high abundance and diversity (Gentry and Dodson 1987b; Nieder et al.2001). In the past decades, most studies carried out on vascular plants havefocused on the tree component, despite the fact that the non-tree vegetation isresponsible for a high percentage of the total diversity in the tropical forests(Gentry and Dodson 1987a; Galeano et al. 1998; Schnitzer and Carson 2000).
Epiphytes are plants that inhabit a discontinuous and three-dimensionallandscape, directly in contact with the forest soil or not (Bennett 1986). Pat-terns of distribution and floristic composition of epiphytic plants have beenrelated to factors of dispersal (Benzing 1986; Wolf 1993), humidity and soils
Biodiversity and Conservation 14: 739–758, 2005. � Springer 2005
DOI 10.1007/s10531-004-3920-9

(Gentry and Dodson 1987b; Leimbeck and Balslev 2001), and variability ofstructure, superficial area and inclination and size of branches of host trees(phorophytes) (Nieder et al. 1999; Freiberg 1996, 2001). Recently, in nearbyrain forests of the Yasunı area, Leimbeck and Balslev (2001) reported sub-stantial differences in aroid epiphytism between floodplains of the Tiputiniriver and surrounding uplands, suggesting a strong role of phorophytelimitation in floodplain forests.
Here we make the first attempt to quantitatively describe vascular epiphyt-ism in Colombian Amazonia. We counted vascular epiphytes in thirty 0.025-haplots, well-distributed over the main landscape units in a part of the basin ofthe middle Caqueta river (Figure 1). Each plot was directly adjacent to a 0.1-haplot at which the species composition of trees and lianas (diameter at breast
Figure 1. Location of the Meta area in Colombian Amazonia.
740

height (DBH) ‡ 2.5 cm) had been recorded 3 years earlier (Duque et al. 2001).The purpose of this paper is to present these species data, while focusing on thequestion whether or not there existed any difference in abundance, diversity, ordistribution of epiphytes between the principal landscape units in the Metaarea.
Study site
The study area comprised about 1000 km2 and was situated along the middlestretch of the Caqueta River in Colombian Amazonia near the mouth of theMeta river, roughly between 1�–2� S and 70�–73� W (Figure 1). The principallandscape units found here were well-drained floodplains, swampy areas(including permanently inundated back swamps and basins in floodplains),areas covered with white-sand soils (found on high terraces of the Caquetariver and in less dissected parts of the Tertiary sedimentary plain), and well-drained uplands or terra firme (never flooded by river water and including lowand high fluvial terraces of the Caqueta river and a Tertiary sedimentary plain)(Duivenvoorden and Lips 1993; Lips and Duivenvoorden 2001). Soils werecalled well-drained when they showed a FAO drainage class of 2 or higher, andpoorly drained when this class was below 2 (FAO 1977). The height of thestudied forests varied between 10–15 m (white-sand areas), 15–25 m (well-drained floodplains and swamps), and 25–35 m (terra firme). Extensive foreststructural information is given in Duque et al. (2001). The area received a meanannual precipitation of about 3060 mm (1979–1990) with a mean monthlyrainfall always above 100 mm (Duivenvoorden and Lips 1993). Mean annualtemperature was 25.7 �C (1980–1989) (Duivenvoorden and Lips 1993).
Methods
Rectangular plots of 5 · 50 m were established directly contiguous to the longside of previously established 20 · 50 m plots. These latter plots were installedin each one of the above-mentioned landscape units, which had been recog-nized on aerial photographs (Duivenvoorden 2001). During walks through theforests, soils and terrain forms were rapidly described, and the forest wasvisually examined. In this way, forest stands with more or less homogeneoussoils were identified. In these stands, plots were located without bias withrespect to floristic composition. Recent gaps due to fallen canopy trees wereavoided. All plots were established in mature forests that did not show signs ofrecent human intervention, at a minimum distance of 500 m between plots(Figure 1). Plots were mapped with GPS. In 1997 and 1998, the density andspecies composition of lianas and trees with DBH ‡ 2.5 cm were recorded inthese 0.1-ha plots (Duque et al. 2001, 2002). During a new fieldwork fromMarch to June 2000, the adjacent 0.025-ha plots were censused for epiphytism.
741

The 5 · 50 m plots were subdivided into subplots of 5 · 10 m, in which allvascular epiphytes occurring on trees and lianas with a stem basis inside theplot area were recorded.
Field collection of epiphytes was done with the help of indigenous climbers.Binoculars were used to examine epiphyte individuals occurring in distantcrowns. With the help of poles, crowns were surveyed and all observed indi-vidual epiphyte plants were collected. For each epiphyte plant, the positionabove ground (in the case of hemi-epiphytes the maximum height was con-sidered), and position on the phorophyte (main stem or branches) were re-corded. Three plant positions were considered: (1) base: individuals found at orbelow 3 m above ground level; (2) stem: individuals found above 3 m andbelow the first branch; (3) branches or crowns: individuals found on stems orbranches in crowns.
For each phorophyte, the following variables were recorded: (1) DBH (fromphorophytes with height lower than 1.3 m the stem diameter was recorded athalf of the total height). (2) Total height and height of first branch, measuredor estimated by means of poles of 8 m length. For trees, we calculated theconical superficial area of the phorophyte stems as 3.14 · the product of thestem radius and the height of the first branch (if there were no branches,the total height was employed).
All species in each plot were collected applying vouchers numbered AMB100–1300. Species identification took place at the Herbario Universidad deAntioquia (HUA), Herbario Amazonico Colombiano (COAH), and HerbarioNacional Colombiano (COL), by means of taxonomic keys, comparison withherbarium collections, and consultations of specialists. The nomenclature offamilies follows Cronquist (1988) for angiosperms and Tryon and Tryon (1982)for pteridophytes. Within families or groups of closely allied families, speci-mens that could not be identified as species because of a lack of sufficientdiagnostic characteristics, were clustered into morpho-species on the basis ofsimultaneous morphological comparisons with all other specimens.
In this study, the term epiphyte is used, in a broad sense, for plants thatspend most of their life cycle attached to other plants (Benzing 1987). Onlythose epiphyte individuals that were in contact with the forest soil were re-corded as hemi-epiphyte. All other epiphyte individuals were recorded as holo-epiphyte. Clones from rhizomatous plants were considered as one individual.
Numerical analysis
To calculate the diversity, Fisher’s alpha index was employed (Fisher et al.1943; Condit et al. 1996). Differences of diversity, species richness, epiphyteabundance, and superficial area of the phorophytes between the landscapeswere analyzed by ANOVA and subsequent Tukey–Kramer tests. The conditionof normal distribution of residuals was checked by means of Shapiro–Wilktests. The analyses were developed using JMP 3.2.2 (SAS 1994).
742

Patterns of epiphyte species composition were explored by DetrendedCorrespondence Analysis (DCA, Hill 1979) in CANOCO version 4 (terBraak and Smilauer 1998), applying plot data of abundance and presence–absence. Correlations between epiphyte species, trees and liana species in theadjacent plots, and the spatial position of the plots, were analyzed byMantel and partial Mantel tests (Legendre and Legendre 1998), applyingR-package for Macintosh (Casgrain and Legendre 2002). The floristic sim-ilarity matrices were constructed on the basis of the abundance data usingthe Steinhaus index. A Euclidean distance matrix was calculated using thegeographical coordinates of the plots (Legendre and Legendre 1998). Thesignificance of the Mantel r coefficient was tested by means of 10,000permutations.
Results
A total of 6129 individual vascular epiphytes were recorded in the 30 plots of0.025 ha each. Precisely 1200 botanical collections were made pertaining to 27families, 74 genera, and 213 species (which included 59 morpho-species). Atotal of 141 species (66%) were found in more than one plot and just 17 species(8%) represented 50% of the total number of individuals registered. Manyspecies (78) were found both as hemi-epiphyte and holo-epiphyte. Most species(107), however, were strictly holo-epiphytic, while 28 species were alwayshemi-epiphytic.
Araceae, Orchidaceae, and Bromeliaceae were the most speciose andabundant families (see Appendix and Figure 2a). Of these, Araceae was themost diverse family in all landscape units. Two genera of Araceae, Philoden-dron and Anthurium, had the highest species richness (Figure 2b). There were117 monocotyledonous species (5 families, 36 genera), 45 species of pterido-phytes (12 families, 20 genera), and 49 dicotyledonous species (10 families, 18genera). Five species were found in all landscape units: Aechmea nivea (Bro-meliaceae), Asplenium serratum (Aspleniaceae), Codonanthe crassifolia (Ges-neriaceae), Anthurium ernestii (Araceae), and Philodendron linnaei (Araceae).Trichomanes ankersii (Hymenophyllaceae) was the most abundant species,being present mainly in upland forests.
A total number of 2763 phorophytes were registered, 1701 (62%) of whichwith DBH ‡ 2.5 cm. On average, one phorophyte carried 2.2 (standard devi-ation = 1.9) epiphyte individuals and 1.8 (SD = 1.2) epiphyte species. Basedon the density of trees and lianas in the adjacent 0.1-ha plots (Duque et al.2001) about 40–60% of the woody plants with DBH ‡ 2.5 cm carriedepiphytes, and about 50–85% in case of DBH ‡ 5 cm (Table 1).
Many (44–60%) epiphyte individuals were found 0–3 m above the ground,and far less (4–12%) were in the crowns or on the branches, throughout alllandscape units (Table 2). Stem bases also carried the highest number of epi-phyte species, but differences with the upper parts of the phorophytes were less
743
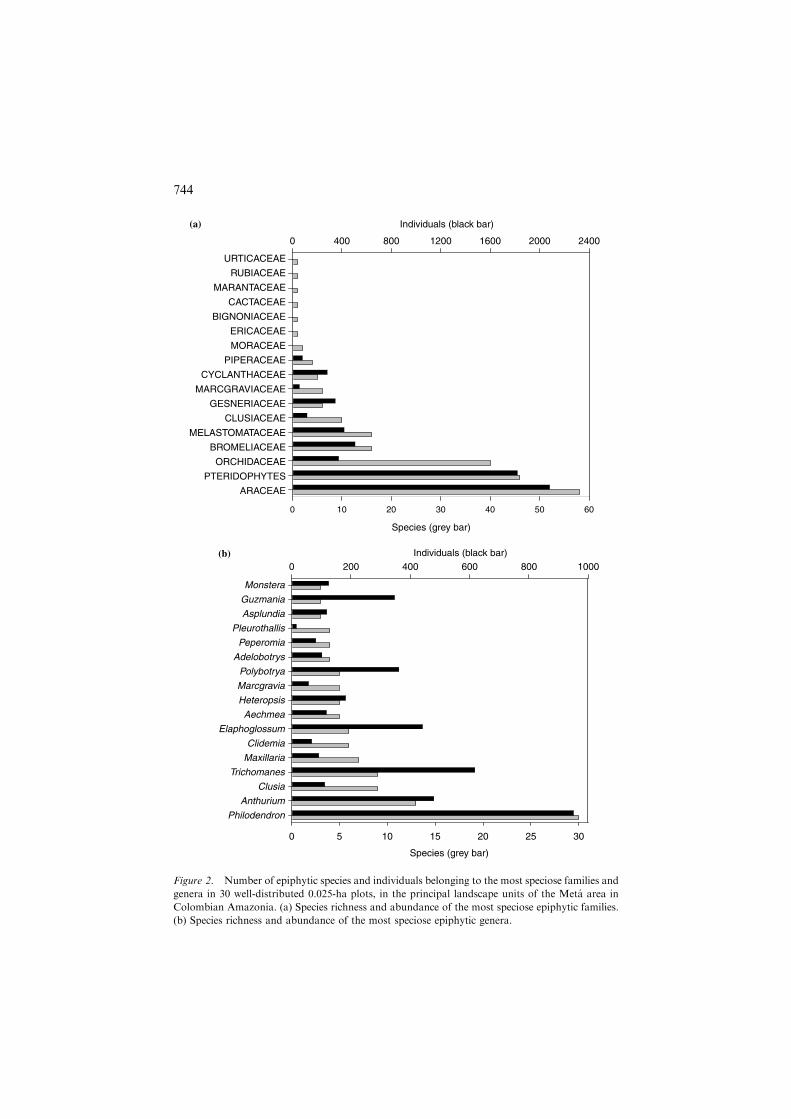
ARACEAE
PTERIDOPHYTES
ORCHIDACEAE
BROMELIACEAE
CLUSIACEAE
MARCGRAVIACEAE
MORACEAE
CACTACEAE
MELASTOMATACEAE
CYCLANTHACEAE
GESNERIACEAE
PIPERACEAE
ERICACEAE
MARANTACEAE
RUBIACEAE
URTICACEAE
BIGNONIACEAE
Individuals (black bar)
Species (grey bar)
0 10 20 30 40 50 60
0 400 800 1200 1600 2000 2400
Species (grey bar)
0 5 10 15 20 25 30
0 200 400 600 800 1000
Anthurium
Philodendron
Aechmea
Clusia
Trichomanes
Heteropsis
Marcgravia
Elaphoglossum
Clidemia
Maxillaria
Guzmania
Polybotrya
Asplundia
Adelobotrys
Individuals (black bar)
Peperomia
Pleurothallis
Monstera
(a)
(b)
Figure 2. Number of epiphytic species and individuals belonging to the most speciose families and
genera in 30 well-distributed 0.025-ha plots, in the principal landscape units of the Meta area in
Colombian Amazonia. (a) Species richness and abundance of the most speciose epiphytic families.
(b) Species richness and abundance of the most speciose epiphytic genera.
744
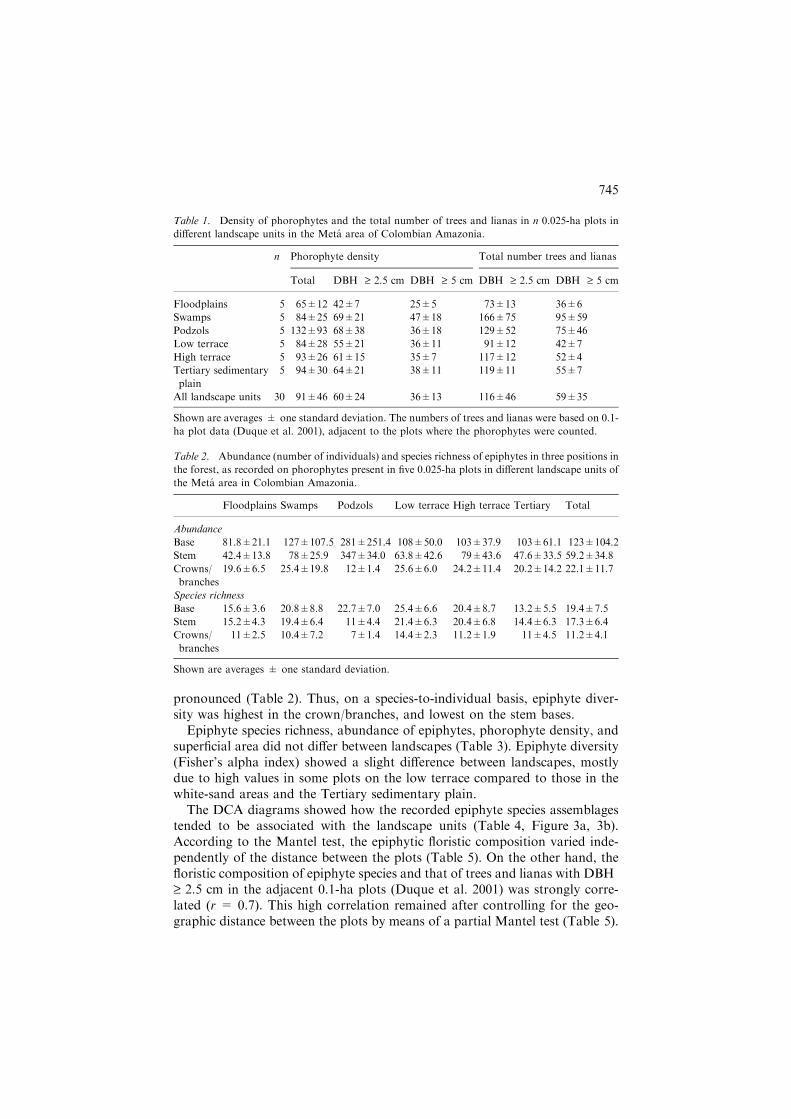
pronounced (Table 2). Thus, on a species-to-individual basis, epiphyte diver-sity was highest in the crown/branches, and lowest on the stem bases.
Epiphyte species richness, abundance of epiphytes, phorophyte density, andsuperficial area did not differ between landscapes (Table 3). Epiphyte diversity(Fisher’s alpha index) showed a slight difference between landscapes, mostlydue to high values in some plots on the low terrace compared to those in thewhite-sand areas and the Tertiary sedimentary plain.
The DCA diagrams showed how the recorded epiphyte species assemblagestended to be associated with the landscape units (Table 4, Figure 3a, 3b).According to the Mantel test, the epiphytic floristic composition varied inde-pendently of the distance between the plots (Table 5). On the other hand, thefloristic composition of epiphyte species and that of trees and lianas with DBH‡ 2.5 cm in the adjacent 0.1-ha plots (Duque et al. 2001) was strongly corre-lated (r = 0.7). This high correlation remained after controlling for the geo-graphic distance between the plots by means of a partial Mantel test (Table 5).
Table 1. Density of phorophytes and the total number of trees and lianas in n 0.025-ha plots in
different landscape units in the Meta area of Colombian Amazonia.
n Phorophyte density Total number trees and lianas
Total DBH ‡ 2.5 cm DBH ‡ 5 cm DBH ‡ 2.5 cm DBH ‡ 5 cm
Floodplains 5 65±12 42±7 25±5 73±13 36±6
Swamps 5 84±25 69±21 47±18 166±75 95±59
Podzols 5 132±93 68±38 36±18 129±52 75±46
Low terrace 5 84±28 55±21 36±11 91±12 42±7
High terrace 5 93±26 61±15 35±7 117±12 52±4
Tertiary sedimentary
plain
5 94±30 64±21 38±11 119±11 55±7
All landscape units 30 91±46 60±24 36±13 116±46 59±35
Shown are averages ± one standard deviation. The numbers of trees and lianas were based on 0.1-
ha plot data (Duque et al. 2001), adjacent to the plots where the phorophytes were counted.
Table 2. Abundance (number of individuals) and species richness of epiphytes in three positions in
the forest, as recorded on phorophytes present in five 0.025-ha plots in different landscape units of
the Meta area in Colombian Amazonia.
Floodplains Swamps Podzols Low terrace High terrace Tertiary Total
Abundance
Base 81.8±21.1 127±107.5 281±251.4 108±50.0 103±37.9 103±61.1 123±104.2
Stem 42.4±13.8 78±25.9 347±34.0 63.8±42.6 79±43.6 47.6±33.5 59.2±34.8
Crowns/
branches
19.6±6.5 25.4±19.8 12±1.4 25.6±6.0 24.2±11.4 20.2±14.2 22.1±11.7
Species richness
Base 15.6±3.6 20.8±8.8 22.7±7.0 25.4±6.6 20.4±8.7 13.2±5.5 19.4±7.5
Stem 15.2±4.3 19.4±6.4 11±4.4 21.4±6.3 20.4±6.8 14.4±6.3 17.3±6.4
Crowns/
branches
11±2.5 10.4±7.2 7±1.4 14.4±2.3 11.2±1.9 11±4.5 11.2±4.1
Shown are averages ± one standard deviation.
745

Table3.
Speciesrichness,abundance
(number
ofindividuals),anddiversity
(Fisher’sAlphaindex)ofepiphytesfoundonphorophytesin
n0.025-haplotsin
differentlandscapeunitsoftheMetaareain
ColombianAmazonia.
Floodplains
(n=
5)
Swamps
(n=
5)
Podzols
(n=
5)
Low
terrace
(n=
5)
Highterrace
(n=
5)
Tertiary
sedim
entary
plain
(n=
5)
Alllandscapes
(n=
30)
ANOVA
F
Speciesrichness
25±7
32±
10
29±
736±7
32±10
23±7
29±
92.1
ns
Number
ofindividuals
143±33
230±
107
278±
214
197±96
206±81
170±
92
204±
115
0.8
ns
Fisher’sAlphaindex
9.3±3.1
(ab)
16.1±13.8
(ab)
9.6±2.7
(a)
13.2±1.1
(b)
10.6±4.3
(ab)
7.6±2.3
(a)
11±
6.4
3.4*
Number
ofphorophytes
65±13
84±
28
132±
93
84±32
93±
29
94±
33
92±
46
1.2
ns
Superficialarea(m
2)
59.7±
19
71.2±29.3
57±
26
68.6±26
76.2±22
89.3±23
70.3±24.1
1.2
ns
Alsoshownare
thenumber
andthesuperficialareaofthephorophytesin
theseplots.Figuresrepresentaverages
±onestandard
deviation.Therightcolumn
gives
theFvalues
oftheANOVA
betweenlandscapeunits(ns=
nonsignificant;*0.05<
p<
0.01).Theletter
codes
(a),(ab),and(b)indicate
theresultof
theTukey–Kramer
post
hoctest
ofdifference
betweenlandscapeunits.
746

Table 4. Summary information of Detrended Correspondence Analyses (DCA), based on vascular
epiphyte species composition on phorophytes in thirty 0.025-ha plots.
Axis 1 Axis 2 Axis 3 Axis 4 Total inertia
A: Presence–absence data
Eigenvalues 0.45 0.28 0.17 0.12 4.23
Length of gradient (sd units) 4.1 3.3 2.8 2.2
B: Abundance data
Eigenvalues 0.54 0.27 0.16 0.12 4.78
Length of gradient (sd units) 4.7 3.2 2.3 1.9
Axi
s 2
0
1
2
3
0 1 2 3 4 5
Axi
s 2
0
1
2
3
FloodplainsSwampsPodzols
Tertiary plainHigh terracesLow terraces
Axis 1
0 1 2 3 4 5
(a)
(b)
Figure 3. Detrended Correspondence Analysis of vascular epiphytes in the Meta area of
Colombian Amazonia. (a) Based on the presence–absence of epiphyte species. (b) Based on the
abundance (number of individuals) of epiphyte species.
747
Discussion
The species belonging to the most speciose families in this study weremore similar to those reported for wet and moist forests in lowlands (Gentryand Dodson 1987b; Foster 1990; Balslev et al. 1998), than those located indrier forests where the aroid component decreased, and Orchidaceae andPteridophytes increased (Wolf and Flamenco-S. 2003). Three of the mostspeciose families (Araceae, Orchidaceae, and Bromeliaceae) have been reportedwithin the most abundant and diverse families in other studies that includedepiphytes as well (Gentry and Dodson 1987b; Balslev et al. 1998; Galeano et al.1998).
The recorded number of epiphyte species is within the range of other reportsfrom Neotropical forests (Gentry and Dodson 1987b) and among the highestfor the Amazonian region (Gentry and Dodson 1987b; Prance 1990; Balslevet al. 1998; Carlsen 2000; Nieder et al. 2000). Our total of 213 vascular epiphytespecies comprised 14% of the species of trees and lianas (DBH ‡ 2.5 cm)found in the adjacent plots. In the same area, Duivenvoorden (1994) found thathemi-epiphytes represented about 5% of the vascular plant species, but hereported undersampling of the upper stems and crowns of high trees. All thesefigures remain well below the estimates of studies in western Ecuador andCosta Rica where between 25 and 35% of vascular species in small plotspertained to epiphytes (Whitmore et al. 1985; Gentry and Dodson 1987a, b).
Recording epiphytes in forest canopies with binoculars is common practice(e.g., Leimbeck and Balslev 2001). However, even though much care has beentaken to observe and sample the epiphytes by climbing into tree crowns, itremains possible that small epiphyte plants have been missed in our study,especially in high trees of floodplains, swamps and terra firme, accounting
Table 5. Mantel and partial Mantel test results of vascular epiphyte species against species of trees
and lianas, and geographic distance (space) in the Meta area of Colombian Amazonia.
Mantel r Partial Mantel r Probability
Matrix A = All vascular epiphytic species
Matrix B
Trees 0.7 0.0001
Space �0.05 0.18
Matrix B Matrix C
Trees Space 0.7 0.0001
Space Trees �0.02 0.33
Matrix A is composed of Steinhaus similarity coefficients between epiphytic species data from thirty
0.025-ha plots. Trees is the matrix composed of Steinhaus similarity coefficients between species
data of trees and lianas (DBH ‡ 2.5 cm) from thirty 0.1-ha plots, each directly adjacent to the
0.025-ha plots where epiphytes were recorded. Space is the matrix composed of Euclidean distances
between plots. Mantel r is the Mantel correlation coefficient between matrix A and matrix B. Partial
Mantel r is the Mantel correlation between matrix A and matrix B when the effect of matrix C is
removed.
748

partially for the high density and species richness of epiphytes at the stem basis.Only by more intensive sampling, for example including careful destructivefelling of all branches, an exhaustive census of epiphyte diversity in tree crownscan be made. To test if the branches and crowns might have been undersam-pled, we cut down 30 trees with a DBH between 20 and 30 cm well outside theplot areas but close to each plot. Each of these trees had a visually defined largeepiphyte load along the stem and in the crown. Contrary to our expectations,the analyses of these data, which are still in a preliminary stage of speciesidentification and therefore not shown here, did not reveal significant differ-ences in the number of epiphyte individuals and epiphyte species in branchesand crowns compared to the phorophytes in similar diameter-class sampled inthe plots.
About 4–6 out of every 10 woody plants (DBH ‡ 2.5 cm) and 5–8 out ofevery 10 woody plant with DBH ‡ 5 cm carried epiphytes, suggesting thatepiphytes fail to effectively colonize a substantial number of potential phoro-phytes in the Meta area. Leimbeck and Balslev (2001), in floodplains of nearbyYasunı, found that 98% of the trees with DBH ‡5 cm carried aroid epiphytes.These authors hypothesized that aroid epiphytes experienced limitation forphorophytes in floodplains. Their floodplain saturation percentage of 98%corresponded to about 25 phorophytes with aroid epiphytes per 0.025 ha whenbased on the tree density (DBH ‡ 5 cm) of 1012/ha reported by these authors.In the five floodplain plots of the Meta area, the average number of phoro-phytes with aroid epiphytes was 21/0.025 ha, corresponding to 58% of the treesand lianas with DBH ‡ 5 cm. So, on a plot area basis, the forests of thefloodplain of the Caqueta river contained 16% less phorophytes covered witharoid epiphytes, and their phorophyte saturation level for aroids was about40% lower than in Yasunı. It seems unlikely, in this light, that the aroidepiphytes in the Meta experience phorophyte limitation to the same degree asmight take place in Yasunı floodplains. For the transition and upland areas inYasunı, about 31 and 32 phorophytes with aroids were found in sample areasof 0.025 ha, which corresponded to 82–86% of the total tree density (DBH‡ 5 cm). In the three terra firme units this average number ranged between 14/0.025 and 29/0.025 ha, corresponding to 26–70% of the tree and liana density(DBH ‡ 5 cm). This comparison suggests that a lower number of trees andlianas are covered by aroid epiphytes in upland forests of the Meta areacompared to Yasunı, and that the saturation level and phorophyte limitation iscomparatively low too, just as in the floodplains. Overall climate and humiditylevels of the Yasunı area and Meta areas hardly differ (Lips and Duiven-voorden 2001). Yasunı forests might be subjected to a greater immigration ofaroid epiphytes from the surrounding forests, especially from the nearby An-des, compared to the Caqueta area. The Andes have been mentioned as a richcentre of diversity for aroid epiphytes (Gentry 1982).
In the Meta area, epiphytes showed a more or less similar abundance andspecies diversity in all landscapes. This is remarkably different from trees,which show a well-documented gradient in species diversity from swamps and
749

podzols to well-drained floodplains and well-drained uplands (Duivenvoorden1996; Duque et al. 2001). Why might landscape factors not affect epiphytediversity in the same way as they do for trees? Epiphytes in upper canopies inall lowland forests are generally subjected to high temperatures and low levelsof air humidity (ter Steege and Cornelissen 1989), leading to energetic losses bytissue respiration and water balance stress (Andrade and Nobel 1997; Zotz andAndrade 1997). In forest understories stress factors differ between forest types.In the understory of tall forests, air humidity tends to be higher and moreconstant but light availability and associated rates of carbon fixation lower(Kessler 2002). In the understory of low forests, light penetration in understoryis higher, but temperature and drought are also higher leading to less favorablegrowth conditions for epiphytes. Therefore, the epiphytes in both high and lowforests in the various landscape units might experience a more or less similarnet degree of stress. Second, epiphytes are claimed to have a high dispersalability (Benzing 1987; Nieder et al. 1999), which would allow a more rapidcolonization reducing possible effects of forest development on epiphyte speciesdiversity. This explanation, however, seems only valid for epiphytes occuring inupper canopy crowns, but not for understory environments where dispersal bywind is less effective. A high epiphyte dispersal ability should lead to a widedistribution of many epiphyte species in all landscapes, which is not in corre-spondence to the high epiphyte–landscape association recorded in the Metaarea.
Epiphyte species compositional patterns were well related to the principallandscape units (Figure 3a, b and Table 4). In view of the dominance of epi-phytes in the understory this is hardly a surprise. The floodplain and swampplots are subjected to an annual inundation by the Caqueta river, during whichwater levels may rise several meters above the forest soil. This, plus the closerproximity of river and swamp water during periods of low river water levelslikely produce a higher humidity (including mist in early mornings), at annualand daily time-scale, compared to upland conditions. Yearly sedimentation ofsilty deposits, which are partially of Andean origin, makes the rooting envi-ronment at the trunk bases more fertile than in upland forests. Leimbeck andBalslev (2001) further mentioned enhanced vegetation reproduction due tomechanical damage or separation of plant parts into ramets when submerged.The lower stand height and simpler structure of white-sand forests might in-duce less habitat diversity, as well as better light penetration and wider dailyamplitude in temperature and humidity in the understory environment,compared to the generally taller forests in the other landscape units.
Contrary to trees, landscape patterns of species diversity and species com-position for epiphytes are uncoupled. In conclusion, we hypothesize that someepiphyte species are more favoured by high humidity (floodplains andswamps), or are better adapted to withstand drought (in low podzol forests)than others without leading to competitive exclusion as this latter process iseffectively counterbalanced by immigration from regional pools in situations oflow phorophyte limitation. We need more explorative studies, and additional
750

studies on the dispersal ability and autobiology of epiphytic taxa and thedynamics of epiphyte populations (Benzing 1995; Nieder and Zotz 1998). Ourresults suggest that caution is needed when knowledge of tree species distri-bution and dynamics are extrapolated to growth forms with a totally differentecology and vice versa.
Acknowledgements
The authors are thankful to members of the Mirana community and to theherbaria of the Missouri Botanical Garden, Herbario Universidad de Antio-quia (HUA), Herbario Amazonico Colombiano (COAH), and the HerbarioNacional Colombiano (COL). Comments on the manuscript of Jan Wolf andtwo anonymous reviewers were gratefully included. This study was partiallyfinanced by the European Commission (ERB IC18 CT960038), Tropenbos-Colombia, Colciencias, the Schure-Beijerink-Popping Fonds (KNAW) and theNetherlands Foundation for the Advancement of Tropical Research – WO-TRO (WB85-335).
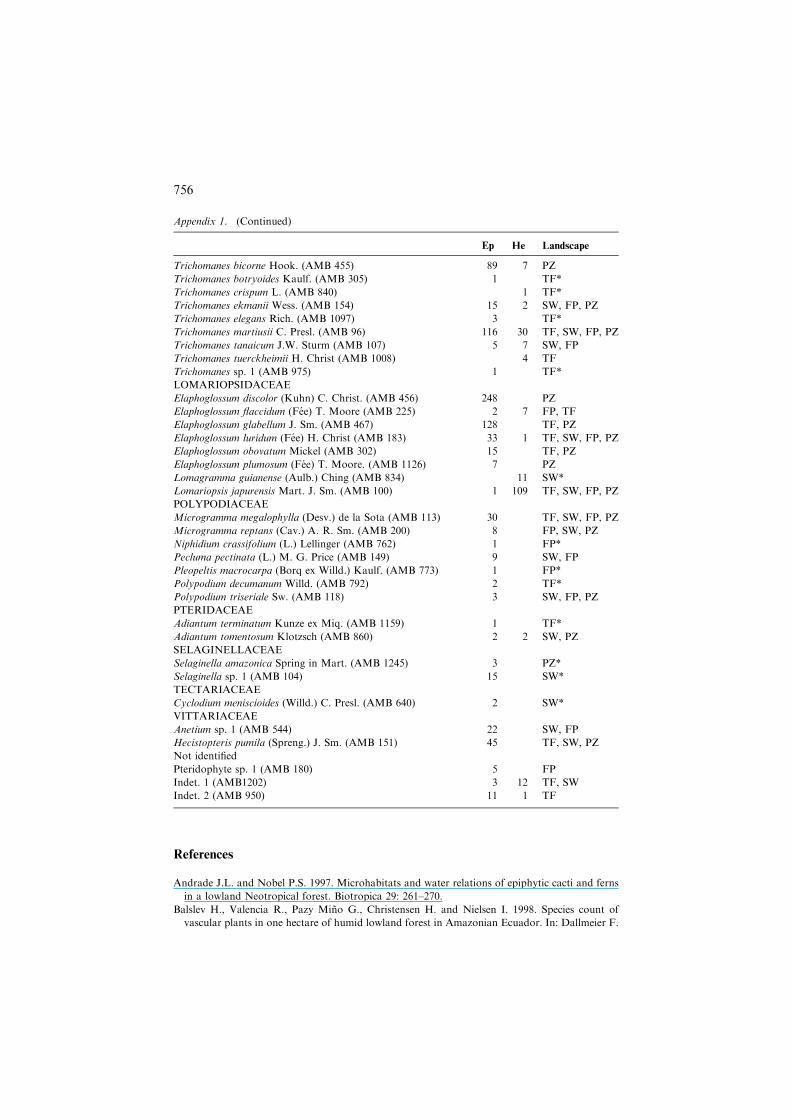
Appendix 1
Species found in 30 widely distributed 0.025-ha plots in the Meta area of Colombian Amazonia.
Voucher codes are added between parentheses. Also, for each species, the number of plant indi-
viduals per habit is given, as well as the main landscape units where the species were recorded.
Habit codes: Ep, Holo-epiphyte; He, Hemi-epiphyte. Landscape codes: TF, Terra firme; FP, Flood
plains; Sw, Swamps; PZ, Podzol. *Species only found in one plot.
Ep He Landscape
Angiosperms
ARACEAE
Anthurium acrobates Sodiro (AMB 821) 1 TF*
Anthurium atropurpureum Schult.
and Maguire (AMB 429)
53 5 TF, SW, PZ
Anthurium clavigerum Poepp. (AMB 177) 1 1 FP*
Anthurium eminens Schott (AMB 142) 10 2 TF, SW, FP
Anthurium ernestii Engl. (AMB 621) 202 15 TF, SW, FP, PZ
Anthurium galactospadix Croat (AMB 245) 6 FP*
Anthurium gracile (Rudge) Schott (AMB 120) 51 4 TF, SW, FP
Anthurium obtusum (Engl.) Grayum (AMB 148) 17 1 TF, SW, FP, PZ
Anthurium pentaphyllum (Aubl.) G. Don (AMB 308) 13 29 TF, FP
Anthurium polydactylum Madison (AMB 141) 2 1 TF, SW
Anthurium sinuatum Benth. ex Schott (AMB 111) 5 24 TF, SW
Anthurium sp. 2 (AMB 175) 21 1 FP, PZ
Anthurium uleanum Engl. (AMB 642) 9 6 FP*
Heteropsis flexuosa (Kunth) Bunting (AMB 208) 58 TF, SW, FP
Heteropsis spruceana Schott (AMB 741) 62 TF, SW, FP
Heteropsis steyermarkii Bunting (AMB 306) 2 49 TF, SW, PZ
Heteropsis sp. 1 (AMB 1173) 2 TF*
751
Appendix 1. (Continued)
Ep He Landscape
Heteropsis sp. 3 (AMB 803) 9 TF*
Monstera gracilis Engl. (AMB 808) 6 TF*
Monstera obliqua Miq. (AMB 770) 70 TF, SW, FP
Monstera spruceana (Schott) Engl. (AMB 342) 1 49 TF
Philodendron acutatum Schott (AMB 315) 7 7 TF, FP
Philodendron applanatum G.M. Barroso (AMB 597) 23 14 TF, SW, FP, PZ
Philodendron asplundii Croat and Soares (AMB 868) 4 4 TF, SW, PZ
Philodendron barrosoanum G.S. Bunting (AMB 339) 6 9 TF, FP
Philodendron buntingianum Croat (AMB 364) 8 13 TF
Philodendron chinchamayense Engl. (AMB 764) 36 TF, SW, FP
Philodendron elaphoglossoides Schott (AMB 583) 1 SW*
Philodendron fragantissimum Kunth (AMB 196) 35 119 TF, SW, FP, PZ
Philodendron guttiferum Kunth (AMB 215) 8 49 TF, FP
Philodendron hederaceum (Jacq.) Schott (AMB 545) 1 30 SW, FP
Philodendron herthae K. Krause (AMB 549) 7 16 TF, SW, FP, PZ
Philodendron holtonianum Schott (AMB 768) 1 FP*
Philodendron hylaeae Bunting (AMB 122) 17 1 TF, SW, PZ
Philodendron insigne Schott (AMB 358) 6 39 TF
Philodendron linnaei Kunth (AMB 121) 184 20 TF, SW, FP, PZ
Philodendron megalophyllum Schott (AMB 99) 56 10 TF, SW, FP, PZ
Philodendron melinonii Brongn. ex Regel (AMB909) 1 4 TF
Philodendron panduriforme (Kunth) Kunth (AMB 1145) 1 TF*
Philodendron pteropus Mart. ex Schott (AMB 173) 7 64 TF, SW, FP
Philodendron pulchrum Barroso (AMB 430) 14 4 TF, SW, PZ
Philodendron tripartitum (Jacq.) Schott (AMB 264) 15 18 TF, SW, FP
Philodendron venustum Bunting (AMB 489) 7 5 TF, SW, PZ
Philodendron sp. 1 (AVG 201) 1 SW*
Philodendron sp. 2 (AMB 785) 8 1 TF, PZ
Philodendron sp. 3 (AMB 851) 6 11 TF, SW, FP
Philodendron sp. 4 (AMB 816 3 TF*
Philodendron sp. 10 (AMB 1203) 1 4 TF
Philodendron sp. 11 (AMB 817) 2 23 TF
Philodendron sp. 12 (AMB 653) 11 TF
Philodendron sp. 13 (AMB 178 6 2 SW, FP
Rhodospatha venosa Gleason (AMB 805) 6 4 TF
Rhodospatha sp. 3 (AMB 739) 197 98 TF, SW, PZ
Stenospermation amomifolium Schott (AMB486) 14 TF, SW, PZ
Stenospermation sp. 1 (AMB 1247) 2 FP, PZ
Syngonium podophyllum Schott (AMB 270) 2 20 FP
BIGNONIACEAE
Schlegelia sp. 1 (AMB 1201) 1 TF*
BROMELIACEAE
Aechmea contracta (Mart. ex Schult.f.)
Mez (AMB 252)
40 TF, SW, FP, PZ
Aechmea corymbosa (Mart. ex Schult.
and Schult. f.) Mez (AMB 135)
15 TF, FP, PZ
Aechmea nivea L.B. Sm. (AMB 368) 41 TF, SW, FP, PZ
Aechmea tillandsioides (Mart. ex Schult.
and Schult. f.) Baker (AMB 318)
19 TF, SW, PZ
752

Appendix 1. (Continued)
Ep He Landscape
Aechmea sp. 1 (AMB 382) 2 TF, PZ
Brocchinia cf. paniculata Schult. f. (AMB 416) 3 TF*
Guzmania brasiliensis Ule (AMB 340) 50 TF, PZ
Guzmania lingulata (L.) Mez (AMB 428) 283 TF, SW, FP
Guzmania vittata (Mart. ex Schult. f.)
Mez (AMB 877)
14 TF, SW
Neoregelia stolonifera L.B. Sm. (AMB 732) 1 SW*
Neoregelia sp. 1 (AMB 492) 2 PZ*
Pepinia sprucei (Baker) Varad. and
Gilmartin (AMB 171)
11 TF, FP
Pepinia uaupensis (Baker) Varad. and
Gilmartin (AMB 363)
5 TF, SW, PZ
Streptocalyx colombianus L.B. Sm. (AMB 303) 5 TF*
Streptocalyx poeppigii Beer (AMB 199) 15 TF, SW, FP
Tillandsia paraensis Mez (AMB 1076) 1 TF*
CACTACEAE
Disocactus amazonicus (K. Schum.)
D.R. Hunt (AMB 1199)
1 TF*
CLUSIACEAE
Clusia cf . amazonica Planch. and
Triana (AMB 490)
8 TF, SW, PZ
Clusia caudata (Planch. and Triana)
Pipoly (AMB 1073)
1 TF*
Clusia flavida (Benth.) Pipoly (AMB 423) 27 TF, SW, PZ
Clusia grandiflora Splitg. (AMB 892) 6 1 TF*
Clusia hammeliana Pipoly (AMB 898) 4 1 TF
Clusia sp. 1 (AVG 374) 21 5 TF
Clusia sp. 2 (AVG 329) 17 1 TF
Clusia sp. 3 (AMB 624) 17 TF, FP, PZ
Clusia sp. 5 (AMB 152) 2 1 SW*
Clusiaceae sp. 1 (AMB 850) 7 SW*
CYCLANTHACEAE
Asplundia vaupesiana Harling (AMB 292) 21 67 TF
Asplundia xiphophylla Harling (AMB 436) 7 24 TF, SW, FP, PZ
Evodianthus funifer (Poit.) Lindm. (AMB 123) 19 35 TF, SW, PZ
Ludovia lancifolia Brongn. (AMB 709) 28 6 TF, FP
Ludovia sp. 1 (AMB 885) 73 4 TF, PZ
ERICACEAE
Psammisia sp. 1 (AMB 443) 11 TF, PZ
Satyria cf . panurensis (Benth. ex Meisn.) Benth.
and Hook. f. ex Nied. (AMB 1097)
1 TF*
GESNERIACEAE
Alloplectus sp.1 (AMB 457) 4 6 PZ*
Codonanthe calcarata (Miq.) Hanst (AMB 427) 90 TF, PZ
Codonanthe crassifolia (H. Focke)
C.V. Morton (AMB 158)
175 TF, SW, FP, PZ
Codonanthopsis dissimulata (H.E. Moore)
Wiehler (AMB 185)
20 TF, SW, FP
Paradrymonia ciliosa (Mart.) Wiehler (AMB 194) 36 16 TF, FP, PZ
753

Appendix 1. (Continued)
Ep He Landscape
Gesneriaceae sp. 1 (AMB 160) 1 SW*
MARANTACEAE
Monotagma laxum (Poepp. and Endl.)
Schum. (AMB 304)
1 TF*
MARCGRAVIACEAE
Marcgravia cf. strenua J.F. Macbr. (AMB 581) 8 13 TF, SW, PZ
Marcgravia sp. 1 (AVG 200) 1 6 TF, SW, FP
Marcgravia sp. 2 (AMB 1209) 1 TF*
Marcgravia sp. 3 (AVG 219) 5 TF
Marcgravia sp. 4 (AMB 184) 12 11 TF, FP, PZ
Marcgraviastrum sp. 1 (AMB 999) 1 TF*
MELASTOMATACEAE
Adelobotrys linearifolia Uribe (AMB 738) 1 46 TF, SW
Adelobotrys marginata Brade (AMB 321) 1 39 TF
Adelobotrys praetexta Pilg. (AMB 902) 9 TF
Adelobotrys spruceana Cogn. (AMB 134) 4 2 SW, FP
Clidemia alternifolia Wurdack (AMB 1152) 2 TF
Clidemia epibaterium DC. (AMB 137) 2 17 TF, SW, PZ
Clidemia sp. 1 (AMB 1196) 1 2 TF
Clidemia sp. 2 (AMB 1061) 1 TF*
Clidemia sp. 3 (AMB 105) 2 SW*
Clidemia sp. 4 (AMB 917) 7 34 TF, PZ
Leandra candelabrum (J.F. Macbr.) Wurdack (AMB 341) 153 TF
Leandra sp. 1 (AMB 165) 1 2 SW*
Tococa lancifolia Spruce ex Triana (AMB 136) 1 SW*
Tococa cf . ulei Pilg. (AMB 1148) 1 TF*
Tococa sp. 1 (AMB 1127) 1 TF*
Melastomataceae sp. 2 (AMB 1115) 89 TF*
MORACEAE
Ficus paraensis (Miq.) Miq. (AMB 1195) 1 TF*
Ficus sp. 1 (AMB 163) 1 TF*
ORCHIDACEAE
Adipe longicornis (Lindl.) M. Wolfe (AMB 316) 4 TF, PZ
Braemia vittata (Lindl.) Jenny (AMB 110) 23 2 TF, SW, FP
Campylocentrum poeppigii (Rchb. f.) Rolfe (AMB 484) 4 FP*
Catacetum sp. 1 (AVG 288) 10 SW, FP
Dichaea hookeri Garay and Sweet (AMB 613) 9 SW
Dichaea rendlei Gleason (AMB 1092) 10 TF, PZ
Epidendrum cf. nocturnum Jacq. (AMB 1256) 1 PZ*
Epidendrum longicolle Lindl. (AMB 139) 83 SW, PZ
Epidendrum microphyllum Lindl. (AMB 523) 10 SW, PZ
Gongora quinquenervis Ruiz and Pav. (AMB 505) 6 PZ*
Masdevallia aff. trigonopetala Kraenzl. (AMB 223) 3 FP*
Maxillaria cf. parkeri Hook. (AMB 521) 53 TF, PZ
Maxillaria cf. triloris E. Morren (AMB 1056) 18 TF, PZ
Maxillaria sp. 1 (AMB 596) 1 PZ*
Maxillaria sp. 3 (AMB 1232) 1 SW*
Maxillaria sp. 4 (AMB 206) 1 FP*
Maxillaria superflua Rchb. f. (AMB 359) 17 TF, PZ
754

Appendix 1. (Continued)
Ep He Landscape
Maxillaria uncata Lindl. (AMB 716) 1 TF*
Notylia sp. 1 (AMB 465 5 PZ*
Octomeria brevifolia Cogn. (AMB 371) 5 TF
Octomeria erosilabia C. Schweinf. (AMB 421) 7 TF
Octomeria sp. 1 (AMB 1219) 31 TF
Ornithocephalus cf. cochleariformis C. Schweinf. (AMB 262) 1 FP*
Paphinia cf. seegeri Gerlach (AMB 470) 9 TF, PZ
Pleurothallis aff. aurea Lindl. (AMB 500) 7 PZ
Pleurothallis cf. flexuosa (Poepp. and Endl.) Lindl. (AMB 517) 3 SW, FP, PZ
Pleurothallis grobyi Bateman ex Lindl. (AMB 717) 1 TF*
Pleurothallis miqueliana (H. Focke) Lindl. (AMB 609) 4 SW
Polyotidium huebneri (Mansf.) Garay (AMB 463) 10 4 TF, SW, PZ
Polystachya sp. 1 (AMB 774) 1 PZ*
Sobralia macrophylla Rchb. f. (AMB 182) 5 FP, PZ
Sobralia sp. 1 (AMB 1074) 1 PZ*
Vanilla cf. columbiana Rolfe (AMB 777) 1 FP*
Vanilla penicillata Garay and Dunst. (AMB 618) 2 SW, FP
Vanilla sp. 1 (AMB 140) 2 SW*
Orchidaceae sp. 1 (AMB 532) 6 PZ
Orchidaceae sp. 2 (AVG 360) 4 TF
Orchidaceae sp. 3 (AMB 758) 2 FP*
Orchidaceae sp. 4 (AMB 1294) 1 FP*
PIPERACEAE
Peperomia cardenasii Trel. (AMB 240) 45 TF, SW, FP, PZ
Peperomia macrostachya (Vahl) A. Dietr. (AMB 181) 12 FP
Peperomia pseudopereskiaefolia C.DC (AMB 560) 8 TF, FP
Peperomia serpens Loud. (AMB 202) 17 FP
URTICACEAE
Pilea sp. 1 (AMB 757) 1
Pteridophytes
ASPLENIACEAE
Asplenium serratum L. (AMB 191) 41 9 TF, SW, FP, PZ
BLECHNACEAE
Salpichlaena hookeriana (Kuntze) Alston (AMB 854) 3 26 SW*
DENNSTAEDTIACEAE
Lindsaea klotzschiana Moritz (AMB 462) 18 PZ
Lindsaea lancea (L.) Bedd. (AMB 114) 8 TF, SW
DRYOPTERIDACEAE
Polybotrya caudata Kunze (AMB 257) 107 TF, FP
Polybotrya polybotryoides (Baker) H. Christ (AMB 115) 16 TF, SW
Polybotrya pubens Mart. (AMB 350) 1 235 TF, SW
Polybotrya sessilisora R. C. Moran (AMB 986) 1 TF*
GRAMMITIDACEAE
Cochlidium furcatum (Hook. and Grev.) C. Chr. (AMB 982) 11 TF, PZ
HYMENOPHYLLACEAE
Hymenophyllum hirsutum (L.) Sw. (AMB 916) 2 TF*
Hymenophyllum sp. 1 (AMB 1254) 5 PZ*
Trichomanes ankersii C. Parker ex Hook. and Grev. (AMB 288) 9 325 TF, SW
Trichomanes arbuscula Desv. (AMB 616) 2 3 SW*
755
References
Andrade J.L. and Nobel P.S. 1997. Microhabitats and water relations of epiphytic cacti and ferns
in a lowland Neotropical forest. Biotropica 29: 261–270.
Balslev H., Valencia R., Pazy Mino G., Christensen H. and Nielsen I. 1998. Species count of
vascular plants in one hectare of humid lowland forest in Amazonian Ecuador. In: Dallmeier F.
Appendix 1. (Continued)
Ep He Landscape
Trichomanes bicorne Hook. (AMB 455) 89 7 PZ
Trichomanes botryoides Kaulf. (AMB 305) 1 TF*
Trichomanes crispum L. (AMB 840) 1 TF*
Trichomanes ekmanii Wess. (AMB 154) 15 2 SW, FP, PZ
Trichomanes elegans Rich. (AMB 1097) 3 TF*
Trichomanes martiusii C. Presl. (AMB 96) 116 30 TF, SW, FP, PZ
Trichomanes tanaicum J.W. Sturm (AMB 107) 5 7 SW, FP
Trichomanes tuerckheimii H. Christ (AMB 1008) 4 TF
Trichomanes sp. 1 (AMB 975) 1 TF*
LOMARIOPSIDACEAE
Elaphoglossum discolor (Kuhn) C. Christ. (AMB 456) 248 PZ
Elaphoglossum flaccidum (Fee) T. Moore (AMB 225) 2 7 FP, TF
Elaphoglossum glabellum J. Sm. (AMB 467) 128 TF, PZ
Elaphoglossum luridum (Fee) H. Christ (AMB 183) 33 1 TF, SW, FP, PZ
Elaphoglossum obovatum Mickel (AMB 302) 15 TF, PZ
Elaphoglossum plumosum (Fee) T. Moore. (AMB 1126) 7 PZ
Lomagramma guianense (Aulb.) Ching (AMB 834) 11 SW*
Lomariopsis japurensis Mart. J. Sm. (AMB 100) 1 109 TF, SW, FP, PZ
POLYPODIACEAE
Microgramma megalophylla (Desv.) de la Sota (AMB 113) 30 TF, SW, FP, PZ
Microgramma reptans (Cav.) A. R. Sm. (AMB 200) 8 FP, SW, PZ
Niphidium crassifolium (L.) Lellinger (AMB 762) 1 FP*
Pecluma pectinata (L.) M. G. Price (AMB 149) 9 SW, FP
Pleopeltis macrocarpa (Borq ex Willd.) Kaulf. (AMB 773) 1 FP*
Polypodium decumanum Willd. (AMB 792) 2 TF*
Polypodium triseriale Sw. (AMB 118) 3 SW, FP, PZ
PTERIDACEAE
Adiantum terminatum Kunze ex Miq. (AMB 1159) 1 TF*
Adiantum tomentosum Klotzsch (AMB 860) 2 2 SW, PZ
SELAGINELLACEAE
Selaginella amazonica Spring in Mart. (AMB 1245) 3 PZ*
Selaginella sp. 1 (AMB 104) 15 SW*
TECTARIACEAE
Cyclodium meniscioides (Willd.) C. Presl. (AMB 640) 2 SW*
VITTARIACEAE
Anetium sp. 1 (AMB 544) 22 SW, FP
Hecistopteris pumila (Spreng.) J. Sm. (AMB 151) 45 TF, SW, PZ
Not identified
Pteridophyte sp. 1 (AMB 180) 5 FP
Indet. 1 (AMB1202) 3 12 TF, SW
Indet. 2 (AMB 950) 11 1 TF
756

and Comiskey J.A. (eds), Forest Biodiversity in North, Central and South America, and the
Caribbean. UNESCO, Paris, France, pp. 585–594.
Bennett B. 1986. Patchiness, diversity, and abundance relationships of vascular epiphytes. Selbyana
9: 70–75.
Benzing D.H. 1986. The vegetative basis of vascular epiphytism. Selbyana 9: 23–43.
Benzing D.H. 1987. Vascular epiphytism: Taxonomical participation and adaptative diversity.
Annals of the Missouri Botanical Garden 74: 183–204.
Benzing D.H. 1995. Vascular epiphytes. In: Lowman M. and Nadkarni N. (eds), Forest Canopies.
Academic Press, San Diego, California, pp. 225–253.
Carlsen M. 2000. Structure and diversity of the vascular epiphyte community in the overstory of a
tropical rain forest in Surumoni, Amazonas state, Venezuela. Selbyana 21: 7–10.
Casgrain P. and Legendre P. 2002. The R Package for Multivariate and Spatial Analysis, Version
4.0d6. Universite de Montreal, Montreal, Canada.
Condit R., Hubbel S.P., Lafrankie J.V., Sukumar R., Manokaran N., Foster R.B. and Ashton P.S.
1996. Species–area and species–individual relationships for tropical trees: a comparison of three
50-ha plots. Journal of Ecology 84: 549–562.
Cronquist A. 1988. The Evolution and Classification of Flowering Plants. The New York Botanical
Garden, New York.
Duivenvoorden J.F. 1994. Vascular plant species counts in the rain forests of the middle Caqueta
area, Colombian Amazonia. Biodiversity and Conservation 3: 685–715.
Duivenvoorden J.F. 1996. Patterns of tree species richness in rain forests of the middle Caqueta
area, Colombia, NW Amazonia. Biotropica 28: 142–158.
Duivenvoorden J.F. 2001. Mapa de la ecologıa del paisaje del medio Caqueta: Plancha Meta. In:
Duivenvoorden J.F., Balslev H., Cavelier J., Grandez C., Tuomisto H. and Valencia R. (eds),
Evaluacion de recursos vegetales no maderables en la Amazonıa noroccidental. Universiteit van
Amsterdam, Amsterdam, The Netherlands.
Duivenvoorden J.F. and Lips J.M. 1993. Ecologıa del paisaje del medio Caqueta: memoria ex-
plicativa de los mapas. Tropenbos-Colombia, Santafe de Bogota, Colombia.
Duque A., Sanchez M., Cavelier J., Duivenvoorden J.F., Mirana P., Mirana J. and Matapı A 2001.
Relacion bosque-ambiente en el Medio Caqueta, Amazonıa colombiana. In: Duivenvoorden
J.F., Balslev H., Cavelier J., Grandez C., Tuomisto H. and Valencia R. (eds), Evaluacion de
recursos vegetales no maderables en la Amazonıa noroccidental. Universiteit van Amsterdam,
Amsterdam, The Netherlands, pp. 99–130.
Duque A., Sanchez M., Cavelier J. and Duivenvoorden J.F. 2002. Different floristic patterns of
woody understory and canopy plants in Colombian Amazonia. Journal of Tropical Ecology 18:
499–525.
FAO 1977. Guidelines for Soil Profile Description. FAO, Rome, Italy.
Fisher R.A., Corbet A.S. and Williams C.B. 1943. The relation between the number of species and
the number of individuals in a random sample of animal population. Journal of Animal Ecology
7: 42–57.
Foster R. 1990. The floristic composition of Rio Manu floodplain forest. In: Gentry A. (ed.), Four
Neotropical Rainforests. Yale University Press, New Haven, Connecticut, pp. 99–111.
Freiberg M. 2001. The influence of epiphyte cover on branch temperature in a tropical tree. Plant
Ecology 153: 241–250.
Freiberg M. 1996. Spatial distribution of vascular epiphytes on three emergent canopy trees in
French Guiana. Biotropica 283: 345–355.
Galeano G., Suarez S. and Balslev H. 1998. Vascular species count in wet forest in the Choco area
on the Pacific coast of Colombia. Biodiversity and Conservation 7: 1563–1575.
Gentry A.H. 1982. Neotropical floristic diversity: phytogeographical connections between Central
and South America. Pleistocene climatic fluctuations, or an accident of the Andean orogeny?
Annals of the Missouri Botanical Garden 69: 557–593.
Gentry A.H. and Dodson C.H. 1987a. Contribution of non-trees to species richness of a tropical
rain forest. Biotropica 19: 149–156.
757

Gentry A.H. and Dodson C.H. 1987b. Diversity and biogeography of Neotropical vascular epi-
phytes. Annals of the Missouri Botanical Garden 74: 205–233.
Hill M.O. 1979. DECORANA–a FORTRAN Program for Detrended Correspondence Analysis
and Reciprocal Averaging. Cornell University, Ithaca, New York.
Kessler M. 2002. Species richness and ecophysiological types among Bolivian bromeliad commu-
nities. Biodiversity and Conservation 11: 987–1010.
Legendre P. and Legendre L. 1998. Numerical Ecology. Elsevier, Amsterdam, The Netherlands.
Leimbeck R.M. and Balslev H. 2001. Species richness and abundance of epiphytic Araceae on
adjacent floodplain and upland forest in Amazonian Ecuador. Biodiversity and Conservation 10:
1579–1593.
Lips J.M. and Duivenvoorden J.F. 2001. Caracterizacion ambiental. In: Duivenvoorden J.F.,
Balslev H., Cavelier J., Grandez C., Tuomisto H. and Valencia R. (eds), Evaluacion de recursos
vegetales no maderables en la Amazonıa noroccidental. Universiteit van Amsterdam, Amster-
dam, The Netherlands, pp. 19–46.
Nieder J. and Zotz G. 1998. Methods of analyzing the structure and dynamics of vascular epiphyte
communities. Ecotropica 6: 33–39.
Nieder J., Engwald S. and Barthlott W. 1999. Patterns of neotropical epiphyte diversity. Selbyana
201: 66–75.
Nieder J., Engwald S., Klawun M. and Barthlott W. 2000. Spatial distribution of vascular epi-
phytes including hemi-epiphytes in a lowland Amazonian forest Surumoni Crane Plot of
Southern Venezuela. Biotropica 323: 385–396.
Nieder J., Prosperi J. and Michaloud G. 2001. Epiphytes and their contribution to canopy
diversity. Plant Ecology 153: 51–63.
Prance G.T. 1990. The floristic composition of the forest of Central Amazonian Brazil. In: Gentry
A. (ed), Four Neotropical Rainforests. Yale University Press, New Haven, Connecticut, pp. 112–
140.
SAS Institute 1994. JMP Statistic and Graphics Guide. SAS Institute Inc., Cary, North Carolina.
Schnitzer S. and Carson W. 2000. Have we forgotten the forest because of the trees? Trends in
Ecology and Evolution 15: 375–376.
ter Braak C.J.K. and Smilauer P. 1998. CANOCO Reference Manual and User’s Guide to CA-
NOCO for Windows: Software for Canonical Community Ordination Version 4. Microcomputer
Power, Ithaca, New York.
ter Steege H. and Cornelissen J.H.C. 1989. Distribution and ecology of vascular epiphytes in
lowland rain forest of Guyana. Biotropica 214: 331–339.
Tryon R.M. and Tryon A.F. 1982. Ferns and Allied Plants, with Special Reference to Tropical
America. Springer, New York.
Valencia R., Balslev H. and Pazy Mino G. 1994. High alpha-diversity in Amazonian Ecuador.
Biodiversity and Conservation 3: 21–28.
Whitmore T.C., Peralta R. and Brown K. 1985. Total species count in a Costa Rican tropical rain
forest. Journal of Tropical Ecology 1: 375–378.
Wolf J.H.D. 1993. Ecology of Epiphytes and Epiphyte Communities in Montane Rain Forest,
Colombia. Ph.D. Thesis, University of Amsterdam, The Netherlands.
Wolf J.H.D. and Flamenco-S A. 2003. Patterns in species richness and distribution of epiphytes in
tropical America. Journal of Biogeography 30: 1689–1707.
Zotz G. and Andrade J.L. 1997. Water relations of two co-occurring epiphytic bromeliads. Journal
of Plant Physiology 152: 545–554.
758




















