
A dual role for MYB60 in stomatal regulation and root growth of Arabidopsis thaliana under drought...
13
A dual role for MYB60 in stomatal regulation and root growth of Arabidopsis thaliana under drought stress Jee Eun Oh • Yerim Kwon • Jun Hyeok Kim • Hana Noh • Suk-Whan Hong • Hojoung Lee Received: 7 October 2010 / Accepted: 16 May 2011 / Published online: 3 June 2011 Ó Springer Science+Business Media B.V. 2011 Abstract In response to environmental challenges, plant cells activate several signaling pathways that trigger the expression of transcription factors. Arabidopsis MYB60 was reported to be involved in stomatal regulation under drought conditions. Here, two splice variants of the MYB60 gene are shown to play a crucial role in stomatal move- ment. This role was demonstrated by over-expressing each variant, resulting in enhanced sensitivity to water deficit stress. The MYB60 splice variants, despite the fact that one of which lacks the first two exons encoding the first MYB DNA binding domain, both localize to the nucleus and promote guard cell deflation in response to water deficit. Moreover, MYB60 expression is increased in response to a low level of ABA and decreased in response to high level of ABA. At initial stage of drought stress, the plant system may modulate the root growth behavior by regulating MYB60 expression, thus promotes root growth for increased water uptake. In contrast, severe drought stress inhibits the expression of the MYB60 gene, resulting in stomatal closure and root growth inhibition. Taken together, these data indicate that MYB60 plays a dual role in abiotic stress responses in Arabidopsis through its involvement in stomatal regulation and root growth. Keywords Arabidopsis thaliana Auxin Drought stress Guard cell MYB60 Introduction The world population has been steadily growing since early 1400s. According to projections, the population will con- tinue to increase until approximately 2050, resulting in various environmental problems such as global warming. Temperature rise is expected to cause severe drought stresses in crops and thus, eventually results in food shortages. Hence, it is believed that elucidation on and manipulation of drought stress tolerance in plants may alleviate human suffering in the face of agricultural stres- ses. Various studies in Arabidopsis have implicated that a large proportion of the genome responds to drought (Shinozaki and Yamaguchi-Shinozaki 2000; Shinozaki et al. 2003) and high salinity stresses (Xiong et al. 2002; Zhu 2002). Guard cells, responsible for controlling the stomatal openings in the epidermis of leaves and other tissues, represent the primary plant defense against drought stress. However, several factors also play important roles in the regulation of stomatal movements including light, carbon dioxide concentration, phytohormones and water avail- ability. Among these, the role of abscisic acid (ABA) has been well documented in the control of guard cell move- ments. ABA inhibits guard cell K in channels while acti- vating Ca 2? -permeable channels and anion channels for anion efflux. These changes lead to guard cell deflation J. E. Oh and Y. Kwon authors contributed equally. Electronic supplementary material The online version of this article (doi:10.1007/s11103-011-9796-7) contains supplementary material, which is available to authorized users. J. E. Oh Y. Kwon J. H. Kim H. Lee (&) College of Life Sciences and Biotechnology, Korea University, 1, 5-ka Anam-dong, Sungbuk-ku, Seoul 136-713, Republic of Korea e-mail: [email protected] H. Noh S.-W. Hong Department of Molecular Biotechnology, College of Agriculture and Life Sciences, Bioenergy Research Institute, Chonnam National University, Gwangju, Korea 123 Plant Mol Biol (2011) 77:91–103 DOI 10.1007/s11103-011-9796-7
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of A dual role for MYB60 in stomatal regulation and root growth of Arabidopsis thaliana under drought...

A dual role for MYB60 in stomatal regulation and root growthof Arabidopsis thaliana under drought stress
Jee Eun Oh • Yerim Kwon • Jun Hyeok Kim •
Hana Noh • Suk-Whan Hong • Hojoung Lee
Received: 7 October 2010 / Accepted: 16 May 2011 / Published online: 3 June 2011
� Springer Science+Business Media B.V. 2011
Abstract In response to environmental challenges, plant
cells activate several signaling pathways that trigger the
expression of transcription factors. Arabidopsis MYB60
was reported to be involved in stomatal regulation under
drought conditions. Here, two splice variants of the MYB60
gene are shown to play a crucial role in stomatal move-
ment. This role was demonstrated by over-expressing each
variant, resulting in enhanced sensitivity to water deficit
stress. The MYB60 splice variants, despite the fact that one
of which lacks the first two exons encoding the first MYB
DNA binding domain, both localize to the nucleus and
promote guard cell deflation in response to water deficit.
Moreover, MYB60 expression is increased in response to a
low level of ABA and decreased in response to high level
of ABA. At initial stage of drought stress, the plant system
may modulate the root growth behavior by regulating
MYB60 expression, thus promotes root growth for
increased water uptake. In contrast, severe drought stress
inhibits the expression of the MYB60 gene, resulting in
stomatal closure and root growth inhibition. Taken
together, these data indicate that MYB60 plays a dual role
in abiotic stress responses in Arabidopsis through its
involvement in stomatal regulation and root growth.
Keywords Arabidopsis thaliana � Auxin �Drought stress � Guard cell �MYB60
Introduction
The world population has been steadily growing since early
1400s. According to projections, the population will con-
tinue to increase until approximately 2050, resulting in
various environmental problems such as global warming.
Temperature rise is expected to cause severe drought
stresses in crops and thus, eventually results in food
shortages. Hence, it is believed that elucidation on and
manipulation of drought stress tolerance in plants may
alleviate human suffering in the face of agricultural stres-
ses. Various studies in Arabidopsis have implicated that a
large proportion of the genome responds to drought
(Shinozaki and Yamaguchi-Shinozaki 2000; Shinozaki
et al. 2003) and high salinity stresses (Xiong et al. 2002;
Zhu 2002).
Guard cells, responsible for controlling the stomatal
openings in the epidermis of leaves and other tissues,
represent the primary plant defense against drought stress.
However, several factors also play important roles in the
regulation of stomatal movements including light, carbon
dioxide concentration, phytohormones and water avail-
ability. Among these, the role of abscisic acid (ABA) has
been well documented in the control of guard cell move-
ments. ABA inhibits guard cell Kin channels while acti-
vating Ca2?-permeable channels and anion channels for
anion efflux. These changes lead to guard cell deflation
J. E. Oh and Y. Kwon authors contributed equally.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11103-011-9796-7) contains supplementarymaterial, which is available to authorized users.
J. E. Oh � Y. Kwon � J. H. Kim � H. Lee (&)
College of Life Sciences and Biotechnology, Korea University,
1, 5-ka Anam-dong, Sungbuk-ku, Seoul 136-713,
Republic of Korea
e-mail: [email protected]
H. Noh � S.-W. Hong
Department of Molecular Biotechnology, College of Agriculture
and Life Sciences, Bioenergy Research Institute, Chonnam
National University, Gwangju, Korea
123
Plant Mol Biol (2011) 77:91–103
DOI 10.1007/s11103-011-9796-7

(Blatt 2000; Schroeder et al. 2001; Hetherington 2001;
Pandey et al. 2007). This inhibits stomatal opening, pro-
motes stomatal closure, and reduces plant water loss
(Sirichandra et al. 2009). In contrast, the transcriptional
control of stomatal movement has not been studied, in part
due to the belief that alterations in gene expression are
temporally incompatible with a prompt drought stress
response by guard cells. Therefore, it was considered more
plausible that cell volume changes are more rapidly
achieved by altering membrane transport via channels or
pumps. However, the level of PP2CA transcripts, encoding
a type 2C protein phosphatase, were found to increase in
Arabidopsis guard cell protoplasts upon treatment with
ABA (Leonhardt et al. 2004). The idea that stomatal
movement could be regulated by changes in gene expres-
sion was further supported by the observation that several
transcription factors can modulate stomatal movements
when plants are exposed to different environmental stimuli.
To date, a few MYB transcription factors have been
implicated in the control of stomatal movement. The MYB
factors, comprising about 196 family members in Arabid-
opsis thaliana, are classified into four different subgroups
(1R-, R2R3-, 3R- and 4R-MYB) according to their
arrangement of MYB domains (Chen et al. 2006; Dubos et al.
2010). This domain may consist of one to four imperfect
repeats and each of which includes about 50–53 amino acid
sequences forming three a-helices thus, building a helix-
turn-helix DNA-binding motifs (Rosinsky and Atchley
1998; Jia et al. 2004). The characterization of MYB60, an
Arabidopsis R2R3-MYB gene, revealed a guard cell-specific
expression pattern that is reduced in response to drought
stress (Cominelli et al. 2005). Plants containing a MYB60
null mutation were shown to exhibit a constitutive reduction
in stomatal opening, resulting in a decreased wilting
response under water stress conditions. Additionally, over-
expression of another guard cell-specific MYB family
member, MYB44, leads to enhanced ABA sensitivity and a
more rapid stomatal closure response in comparison to wild-
type plants (Jung et al. 2008). Thus, the reduced expression
of genes encoding type 2C protein phosphatases, known to
be negative regulators, partly explains the increased toler-
ance of MYB44 over-expressing plants to abiotic stresses.
Another MYB superfamily member, MYB61, also belongs to
the R2R3-MYB family and mediates responses leading to
stomatal closure (Liang et al. 2005). More recently, the
MYB96-mediated ABA signals were shown to be integrated
into an auxin signaling pathway (Seo et al. 2009). MYB96 is
induced by ABA while MYB60 is repressed by ABA. This
indicates that the expressions of different MYB genes are
carefully controlled in their unique and specific manner. This
notion was supported by the observation that MYB96-over-
expressing Arabidopsis plants exhibited enhanced drought
resistance with reduced lateral roots (Seo et al. 2009).
To understand the role of MYB60 in drought stress, two
splice variants were identified from the TAIR database.
One of these variants was initially suspected to have an
alternative role in stomatal movements due to the absence
of the first two exons, which encode a R2-type repeat of
R2R3-MYB domain found in full MYB60 sequence. The
results presented here demonstrate that both MYB60 splice
variants confer sensitivity to drought stress conditions
when overexpressed. Microarray analysis also detected
decreased levels of several aquaporin transcripts in plants
overexpressing the MYB60.2 splice variant under drought
stress, confirming the previous hypothesis reported by
Cominelli et al. (2005). Furthermore, it is clarified that
MYB60 transcripts are expressed in roots as well as in
guard cells. This result correlates with reduced expression
levels of the auxin-responsive DR5::GUS (beta-glucuroni-
dase) construct and an increased number of lateral roots in
MYB60-overexpressing plants. Taken together, these
results support that MYB60.1 and MYB60.2 are essential
transcriptional regulators involved in monitoring the status
of water availability and fine-tuning of the plant responses
to drought stress by regulating guard cell movements and
root growth.
Materials and methods
Plant materials and growth conditions
Arabidopsis thaliana, ecotype Columbia-0, was used for all
experiments. To examine soil growth phenotypes, seeds
were allowed to germinate and grow at 23�C with a 16 h
light/8 h dark cycle at a relative humidity of 70% following
3 days of stratification at 4�C. To examine plant growth
and developmental phenotypes on sterilized medium, seeds
were surface-sterilized and sown in Murashige and Skoog
(MS) agar supplemented with 2% sucrose. After 3 days of
stratification at 4�C, seeds were germinated at 23 ± 1�C
with a 16 h light/8 h dark cycle. Five-day-old seedlings
were transferred to test medium containing MS basal salts,
2% (w/v) sucrose and 1.2% phytoagar (Duchefa Biochemie
B.V., The Netherlands), and grown vertically.
Transgenic plants
Since the Arabidopsis Information Resource (TAIR,
www.arabidopsis.org) indicated that At1g08810 has two
transcripts, two sets of primers were designed as follows:
for the longer transcript, MYB60.1, forward 50-ATGGGTA
GGCCTCCATGC-30 and reverse 50-GAGCTCTCTAATA
TGCTTTAA-30, and for the shorter transcript, MYB60.2,
forward 50-ATGTGTCCTGATGTTTGTATCTTA-30 and
reverse 50-GAGCTCTCTAATATGCTTTAA-30. To amplify
92 Plant Mol Biol (2011) 77:91–103
123

the promoter region, the reverse primer MYB60proR
(50-CTCTCTGAGTCTTATGCTCTC-30) was used in com-
bination with MYB60proF1 (50-GCTTGTGACTCTTCT
TCCA-30), resulting in the synthesis of a *1.2 kb fragment,
with MYB60proF2 (50-CACATCCTTCACGTAGATGAC-
30), resulting in the synthesis of a *800 bp fragment, or
with MYB60proF3 (50-GTCTCTTCTTACGTTCCCTTC-30),resulting in the synthesis of a *400 bp fragment. For
overexpressing lines, DNA fragments were introduced into
the binary vector pBI121 (Promega Corp., WI, USA) under
the control of 35S CaMV promoter. Agrobacterium tum-
efaciens strain GV3101 was used to transform Arabidopsis
plants by the floral dipping method (Clough and Bent 1998).
Microscopy
Arabidopsis Col-0 wild-type and transgenic plants were
grown on moist soil until the rosettes were fully developed.
A month after germination, leaves were excised, allowed to
dry for 5 h, and monitored by inverted fluorescence
microscopy (Carl-Zeiss, Germany) to determine stomatal
opening. For confocal microscopy, green fluorescence
images were obtained with a Confocal Laser Scanning
Microscope (Bio-Rad, UK) using a 510–580 nm emission
setting. GFP was excited with a 568 nm krypton laser and
confocal images were collected using a 600–620 nm
emission setting. FM4-64 was excited using the 488 nm
line from an argon ion laser, and emitted fluorescence was
monitored using the 650LP filter.
RNA gel blot analysis
Wild-type and transgenic plants grown for 2 weeks on MS
medium were treated with hormones and chemicals as
indicated in the figures. Total RNA was extracted from
whole seedlings as previously described (Lee et al. 2002).
20 lg of total RNA was subjected to electrophoresis on
1.5% (w/v) agarose gels containing formaldehyde and then
transferred to a HybondTM-XL membrane (Amersham
Biosciences, NY, USA). The membranes were pre-
hybridized for 1 h and hybridized overnight at 65�C with
probes labeled with 32P-dCTP using the Random Primers
DNA Labeling System (Invitrogen Corp., CA, USA) fol-
lowing the manufacturer’s instruction.
qRT–PCR
Total RNA and the subsequent first-strand cDNA were pre-
pared from 2 week-old Col-0 and overexpressor seedlings.
RNA was extracted using Trizol reagent (Invitrogen, CA,
USA) and cDNA was synthesized using M-MLV Reverse
Transcriptase (iNtRON Biotechnology, Korea) according to
the manufacturer’s instructions. qRT-PCR reactions (total
volumes of 20 ll) were conducted using iCycler iQ
(Bio-Rad, CA, USA). Specific primer sets were designed as
follows. Primer sets to specifically detect MYB60.1 or
MYB60.2 transcripts, forward 50-ATGGGTAGGCCTCCA
TGC-30 and reverse 50-TTGTTACCCAATAAGGCTTGC-
30 or forward 50-ATGGTTGTGGTAAGATGGG-30 and
reverse 50- GCTTGAAGCATAGGTAGATC-30 were used
respectively. For qRT-PCR to quantify flavonoid biosyn-
thetic genes, primer pairs were as follows: PAP1(50-AT-
GGAGGGTTCGTCCAAAG-30 and 50-CAAGGTGCTCC
CCTTTTCTGTT-30) and F3H (50- ATGGCTCCAGGAA
CTTTGAC-30 and 50- AGCGAAGATTTGGTCGACAG-
30). Data represent the average of three independent
experiments.
Microarray analysis
The transcript levels of flavonoid-related genes in desic-
cation-treated Col-0 and MYB60.2-ox plants were obtained
by microarray analysis. four-week-old soil grown Col-0
and MYB60.2-ox plants were dehydrated for 3 days prior to
sampling for microarray analysis. Total RNA from the
rosette leaves of 1-month-old plants, grown on soil and
dried for 3 days, was purified using the RNeasy Plant Mini
kit (Qiagen, MD, USA). 5 lg of fibroblast total RNA was
used for probe-labeling and hybridization, detection and
scanning were completed according to the manufacturer’s
standard protocols, Affiymetrix, Inc. Using One-Cycle
cDNA Synthesis Kit (Affymetrix, CA, USA), single-
stranded cDNA was synthesized from total RNA and
double-stranded cDNA was obtained using DNA ligase,
DNA polymerase I, RNase H, and T4 DNA polymerase.
In vitro transcription was performed with ds-cDNA for in
vitro transcription using the GeneChip IVT Labeling Kit
(Affymetrix). After cleaning up with Sample Cleanup
Module (Affymetrix), 10–15 lg of labeled cRNA was
fragmented from 35 to 200 bp and hybridized to Human
Genome U133A 2.0 gene chips (Affymetrix)) at 45�C for
16 h. Then the arrays were washed in a GeneChip Fluidics
Station 450 with wash buffers according to the Affymetrix
standard protocol. The arrays were then stained and the
intensities were measured with GeneChip scanner 3000
(Affymetrix) by GCOS Affymetrix software.
Histochemical GUS assays
Two- or six-week-old Arabidopsis plants were subjected to
various abiotic stresses as indicated in the figures. The
GUS assay was performed by incubating whole plant tissue
in GUS assay solution at 37�C overnight. Chlorophyll was
then removed with 70% ethanol at 70�C prior to observa-
tion as described (Jefferson et al. 1987).
Plant Mol Biol (2011) 77:91–103 93
123

Statistical analysis
Data are expressed as means ± standard error (SE).
Comparisons were made between groups with the Stu-
dent’s t tests.
Results
The Arabidopsis MYB60 gene has two splice variants
Initially, the role of MYB60 as a negative regulator
against plant drought stress was investigated. Two splice
variants of MYB60, specifying cDNAs of 843 and 594 bp,
were identified from the TAIR database. These variants
were designated MYB60.1 and MYB60.2, respectively
(Fig. 1a). MYB60 is also classified as a member of the
R2R3-type MYB subfamily which contains 125 addi-
tional genes. However, the first R domain is absent in the
MYB60.2 transcript and its translation initiates with an
ATG in the latter part of the second intron (Fig. 1a). As
the MYB domain plays a crucial role in the recognition
of cis-acting elements in DNA, the individual role of the
MYB60.2 transcript was scrutinized and characterized to
determine whether it had a role distinct from that of
Fig. 1 Sequence analysis of Arabidopsis MYB60. a AtMYB60 is
alternative spliced during post-transcriptional processing, resulting in
two non-canonical transcripts. The longer transcript contains 843 bp
and the shorter transcript contains 594 bp. The shorter transcript
initiates translation with an ATG codon located within the second
intron of the gene. Probe locations for MYB60.1 or MYB60.2 are as
indicated. b Sequence alignment of MYB60.1 and MYB60.2 with
other R2R3-MYB family members of Arabidopsis. Among 126
R2R3-MYBs, AtMYB30, AtMYB94, and AtMYB96 share the
highest similarity. Interestingly, the shorter transcript lacks the first
R2 domain. c MYB60.1 and MYB60.2 transcripts were cloned into
Col-0 plants. The transcript expression levels from T3 generation of
each homozygous transgenic plant line were confirmed via northern
blot analysis and quantified
94 Plant Mol Biol (2011) 77:91–103
123
MYB60.1. Based on DNA sequence alignment, MYB60.1
and MYB60.2 were found to have high sequence
homology and to contain several conserved domains (or
blocks) also found in MYB30, MYB96 and MYB94.
Amino acid sequence homology between MYB60.1 and
MYB60.2 is 69.3% as determined by protein blast anal-
ysis while their sequences are 99% identical within the
shared region as determined by using Blastp Suit-2.
Moreover, the amino acid sequence of MYB60.1 shares
89, 78, and 76% homology with MYB30, MYB96 and
MYB94, respectively (Fig. 1b).
To examine potential hormone-responsive elements in
the MYB60 promoter, three fragments of the MYB60 pro-
moter were fused to a GUS-encoding sequence in the
vector pBI121. The resulting constructs, containing
*1.2 kb, *800 bp, and *400 bp of sequence upstream of
the start codon, were named MYB60proF1, MYB60proF2,
and MYB60proF3, respectively (Fig. S1). As shown in Fig.
S1, the 1.2 kb element resulted in the highest GUS
expression levels. GUS staining was observed in the sto-
mata of leaves, stems, and inflorescences.
Soil-grown MYB60.1-ox and MYB60.2-ox plants are
more susceptible to drought stress conditions
MYB60.1 and MYB60.2 transcripts were overexpressed to
further examine their roles in environmental stress
responses. The MYB60 transcript levels of MYB60.1-ox and
MYB60.2-ox seedlings were detected by northern analysis
(Fig. 1c). To specifically detect the MYB60.1 transcript, a
DNA fragment encompassing the first and second exons,
which are absent in AtMYB60.2, was used as a probe.
Substantial levels of MYB60 expression were detected in
the MYB60.1-ox lines #9 and 16, and in MYB60.2-ox lines
#3, 7 and 8 (Fig. 1c). As MYB60 is predicted to encode a
transcription factor with an R2R3-type MYB DNA-binding
domain, the protein was suspected to localize to the
nucleus. To investigate this hypothesis, MYB60.1::GFP or
MYB60.2::GFP construct was separately inserted into
onion epidermal cells by particle bombardment. As
expected, green fluorescence of GFP-fused each MYB
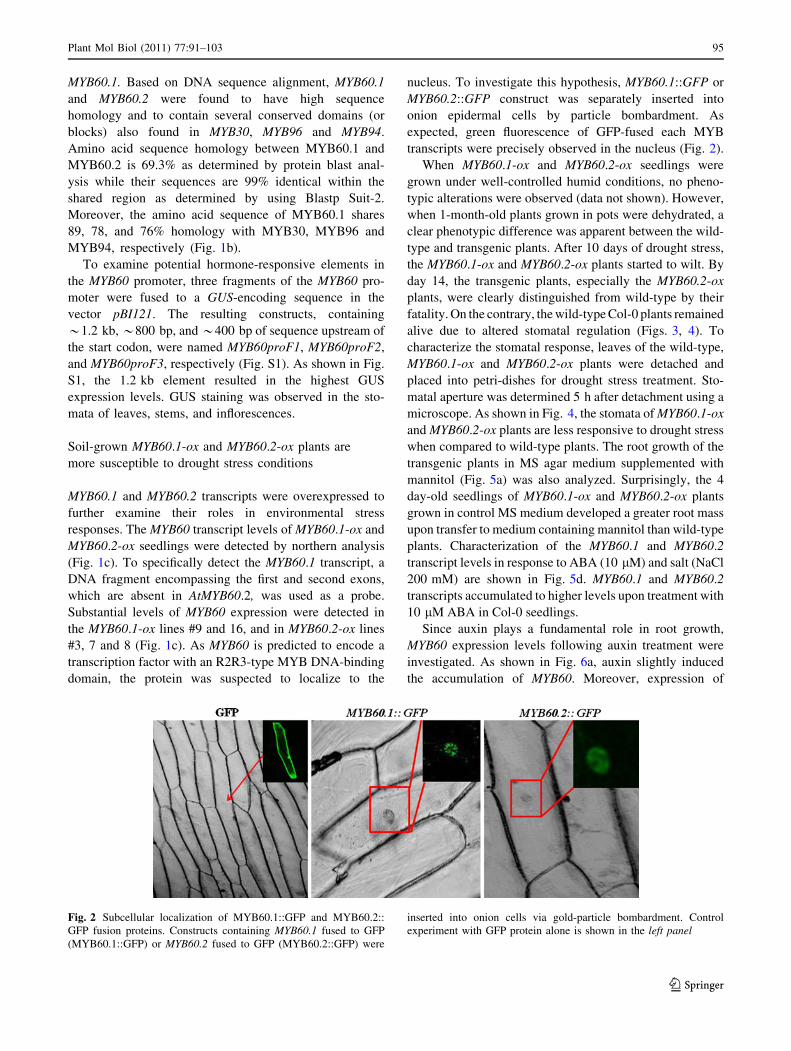
transcripts were precisely observed in the nucleus (Fig. 2).
When MYB60.1-ox and MYB60.2-ox seedlings were
grown under well-controlled humid conditions, no pheno-
typic alterations were observed (data not shown). However,
when 1-month-old plants grown in pots were dehydrated, a
clear phenotypic difference was apparent between the wild-
type and transgenic plants. After 10 days of drought stress,
the MYB60.1-ox and MYB60.2-ox plants started to wilt. By
day 14, the transgenic plants, especially the MYB60.2-ox
plants, were clearly distinguished from wild-type by their
fatality. On the contrary, the wild-type Col-0 plants remained
alive due to altered stomatal regulation (Figs. 3, 4). To
characterize the stomatal response, leaves of the wild-type,
MYB60.1-ox and MYB60.2-ox plants were detached and
placed into petri-dishes for drought stress treatment. Sto-
matal aperture was determined 5 h after detachment using a
microscope. As shown in Fig. 4, the stomata of MYB60.1-ox
and MYB60.2-ox plants are less responsive to drought stress
when compared to wild-type plants. The root growth of the
transgenic plants in MS agar medium supplemented with
mannitol (Fig. 5a) was also analyzed. Surprisingly, the 4
day-old seedlings of MYB60.1-ox and MYB60.2-ox plants
grown in control MS medium developed a greater root mass
upon transfer to medium containing mannitol than wild-type
plants. Characterization of the MYB60.1 and MYB60.2
transcript levels in response to ABA (10 lM) and salt (NaCl
200 mM) are shown in Fig. 5d. MYB60.1 and MYB60.2
transcripts accumulated to higher levels upon treatment with
10 lM ABA in Col-0 seedlings.
Since auxin plays a fundamental role in root growth,
MYB60 expression levels following auxin treatment were
investigated. As shown in Fig. 6a, auxin slightly induced
the accumulation of MYB60. Moreover, expression of
Fig. 2 Subcellular localization of MYB60.1::GFP and MYB60.2::
GFP fusion proteins. Constructs containing MYB60.1 fused to GFP
(MYB60.1::GFP) or MYB60.2 fused to GFP (MYB60.2::GFP) were
inserted into onion cells via gold-particle bombardment. Control
experiment with GFP protein alone is shown in the left panel
Plant Mol Biol (2011) 77:91–103 95
123

MYB60 was detected in the root tissues of wild-type plants
upon 10 lM of auxin treatment (Fig. 6a). Additionally,
GUS staining confirmed that the MYB60 promoter in the
roots and guard cells of the transgenic plants was sensitive
to IAA, although patch-type expression was detected in the
root tissue (Fig. 6b).
Possible target genes of MYB60 under drought stress
To comprehensively analyze the regulatory role of MYB60
and to gain more insights into the coordination of drought
stress responses with developmental and environmental
signaling systems, microarray analysis was conducted
using Col-0 and MYB60.2-ox plants (Tables 1, 2). Four-
week-old soil-grown Col-0 and AtMYB60.2-ox plants were
dehydrated for 3 days prior to sampling for microarray
analysis. Large numbers of drought-regulated genes,
including many novel genes, were detected to be altered.
The interactions between drought, rehydration, plant hor-
mones, and other environmental factors were investigated
using microarray analysis and in silico comparisons. Many
flavonoid biosynthetic genes were up-regulated in MYB60-
ox plants. As flavonoids are known to interfere with polar
auxin transport, plant phenotypes in response to auxin were
characterized. DR5::GUS plants were crossed with
MYB60.1-ox and MYB60.2-ox plants, and their progenies
were screened to identify double homozygous lines.
Fig. 3 Drought tolerance of MYB60.1-ox and MYB60.2-ox plants.
Col-0, MYB60.1-ox and MYB60.2-ox plants were seeded on soil.
1 month after germination, water was withheld for up to 2 weeks
prior to image collections
Fig. 4 Stomatal response of MYB60.1-ox and MYB60.2-ox plants
under drought stress. Col-0, MYB60.1-ox and MYB60.2-ox plants
were seeded on soil. To determine stomatal responses, the leaves of
wild-type, MYB60.1-ox and MYB60.2-ox plants at 1 month after
germination were detached and placed in a petri-dish (without the lid)
for drought stress treatment. 5 h after detachment, the stomatal
aperture was determined under the microscope (a) and pictures were
taken (b). The data represents the average of three independent
experiments with 100 stomata evaluated for each determination.
Lower case letters (a, b, c, d) indicate differences that are statistically
significant (t test P \ 0.05–0.01, comparison made between Col-0
and different overexpressing lines in the indicated treatment). Verticalbars indicate SE (n = 3)
96 Plant Mol Biol (2011) 77:91–103
123

Fig. 5 Drought-related
phenotypes of MYB60.1-ox and
AtMYB60.2-ox plants. a Root
architecture of Col-0, MYB60.1-
ox and MYB60.2-ox plants
grown on media containing
mannitol. Main root lengths
b and root mass c were
measured from each line. Seeds
were germinated on MS
medium and 5-day-old
seedlings were transferred to
medium supplemented with
mannitol and allowed to grow
for 2 additional weeks. The data
represents the average of three
independent experiments with
20 seedlings evaluated for each
determination. Lower caseletters (a, b, c, d) indicate
differences that are statistically
significant (t test
P \ 0.05–0.01, comparison
made between Col-0 and
different overexpressing lines in
the indicated treatment).
Vertical bars indicate SE
(n = 3). d For qRT-PCRanalysis, wild-type seedlings
were grown on growth medium
for 2 weeks and then transferred
to medium supplemented with
ABA (10 lM) or NaCl
(200 mM) for 3 or 6 h as
indicated prior to RNA
extraction. Transcript levels of
MYB60.1and MYB60.2 are
shown. Actin transcript levels
were used as an internal control
Plant Mol Biol (2011) 77:91–103 97
123

DR5::GUS x AtMYB60.1-ox F3 plants, DR5::GUS x At-
MYB60.2-ox F3 plants, and DR5::GUS plants were grown
in control MS-agar medium for 2 weeks and then trans-
ferred to media supplemented with 0, 0.1 or 10 lM IAA
for 6 h. As shown in Fig. 7, GUS expression was signifi-
cantly decreased in the roots and leaves of both DR5::GUS
x AtMYB60.1-ox F3 and DR5::GUS x AtMYB60.2-ox F3
plants.
Anthocyanin, a subgroup of plant secondary metabolite
flavonoid, was considered as a potential regulator of polar
auxin transport in plant tissues. Anthocyanin levels were
determined in the wild-type, MYB60.1-ox lines #9 and 16,
and MYB60.2-ox lines #7 and 8 (data not shown). However,
the differences in anthocyanin accumulation levels
between the control and overexpressor plants were
insignificant. Since some flavonoid biosynthetic genes,
such as PAP1 and PAP2, are reported to express in
response to sucrose (Teng et al. 2005), the transcript level
of MYB60 was examined by northern analysis after sucrose
treatments. As shown in Fig. 8, the MYB60 transcript level
was increased strongly after 3 or 6 h of incubation in
medium containing 4, 6, or 8% sucrose.
Interestingly enough that number of flavonoid biosyn-
thetic genes were determined to be highly expressed in
MYB60.2-ox plants after drought treatment as listed by
microarray analysis data (Table 2). The PAP1 and flava-
none-3ß-hydroxylase (F3H) expression levels were quan-
tified with quantitative RT-PCR to assess the relative
transcript levels of the flavonoid biosynthetic genes such as
PAP1 and F3H in MYB60-ox plants in Fig. 9. The tran-
scription level of PAP1and F3H were increased upon 8%
sucrose treatment.
Discussion
Drought and highly saline soils are among the most serious
challenges to crop production in the world today. Both
traditional breeding and genetic engineering of crop plants
have been utilized to improve drought and high salinity
tolerance or resistance with the goal of increasing agri-
cultural productivity in affected regions. An understanding
of the plant responses to abiotic stress at the genomic level
is expected to provide an essential foundation for future
breeding and genetic engineering strategies. Thus, to fulfill
the ultimate aim of crop-yield improvement, the function
of MYB60, which is involved in abiotic stress response, was
investigated in detail. MYB60 is a member of the R2R3-
type MYB subfamily. In the Arabidopsis Information
Resources (TAIR), MYB60 is listed as encoding two splice
variants designated MYB60.1 and MYB60.2. Interestingly,
MYB60.2 lacks the first R region and is apparently trans-
lated from an ATG located in the latter part of the second
intron (Fig. 1a). Each MYB repeat is known to play a
crucial role in the recognition of cis-acting elements in the
DNA. Hence, this study investigated whether these two
transcripts have roles that are distinct from or redundant to
one another. Full MYB60.1 protein was located in the
nucleus (Fig. 2) as predicted from its biomolecular func-
tion as a transcription factor. Moreover, the MYB60.2
variant, which is missing the first and second exons of
MYB60 (Fig. 1), also localizes to the nucleus (Fig. 2)
supporting that it may act as a transcriptional regulator
similar to MYB60.1.
The two splice variants of MYB60, MYB60.1 and
MYB60.2, alter plant responses to drought stress by con-
trolling stomatal movement (Figs. 3, 4). The regulatory
mechanism of guard cells as drought-adaptive plant
Fig. 6 MYB60 is induced by auxin. a The MYB60 transcript was
monitored in whole or root tissues isolated from three-week-old wild-
type, MYB60.1-ox and myb60-ko plants by northern analysis. Seeds
were germinated on MS medium and subsequent seedlings were
transferred to liquid medium supplemented with 10 lM IAA for the
indicated time period. The rRNA gene is shown as a loading control.
b To determine whether the MYB60 gene promoter is responsive to
IAA, a plasmid containing the promoter region (*1.2 kb) of MYB60fused to the coding region of the reporter gene GUS was constructed.
Histochemical assays were conducted to visualize the location of
b-glucurodinase (GUS) activity in transgenic plants at 0, 6 or 24 h
after incubation with 10 lM IAA. Two-week-old plants were stained
with GUS solution overnight, and chlorophyll was removed by
treatment with 70% ethanol
98 Plant Mol Biol (2011) 77:91–103
123

response was recently reviewed (Sirichandra et al. 2009).
As the epidermis is covered with an impermeable wax
layer, stomata play a crucial role in gas exchange between
plants and the atmosphere. Stomatal movement appears to
be controlled by an interaction between the activities of ion
channels and vacuolar and membrane transporters, and by
metabolite conversions (Roelfsema and Hedrich 2005;
Wasilewska et al. 2008). Additionally, the importance of
MYB60 in modulating stomatal movements has been
established (Cominelli et al. 2005). However, previous
reports did not realize the presence of a splice variant.
Recognizing its presence, this variant is the focus of this
study, as splice variants have been shown to sometimes
exhibit opposing functions. For instance, SULF1, a recently
discovered member of the sulfatase gene family from quail
embryo, has one splice variant (SULF1B) that appears to
oppose the functional activity of a second splice variant
(SULF1A) (Sahota and Dhoot 2009). Because ectopic
expression of either MYB60 variant confers greater sus-
ceptibility to drought stress, MYB60.1 and MYB60.2 appear
to play similar roles in terms of stomatal movement
(Figs. 3, 4). In contrast to myb60 knock-out plants (Fig.
S2), MYB60.1-ox and MYB60.2-ox plants developed
greater root mass than wild-type plants in sealed growth
medium (Fig. 5). This finding is surprising because previ-
ous results revealed that the MYB60.1-ox and MYB60.2-ox
plants withered earlier than wild-type plants when grown
under drought conditions in soil. The difference between
the two experimental results (Figs. 3, 5) may occur from a
higher level of humidity in the sealed plate shown in Fig. 5.
Apparently, ectopic expression of MYB60 was not able to
enhance evaporation through the stomata of transgenic
plants, causing faster mortality rate, although their guard
cells were constantly wide open in the sealed plate.
In addition to the fact that MYB60 expression was lim-
ited in guard cells in aerial parts, MYB60 was also
Table 1 Microarray analysis of flavonoid-related genes in drought-treated Col-0 and MYB60.2-ox plants
Probe sets Col-
0
60.2oX P value Log2
ratio
Target description MIPS Gene
symbol
Up-regulated genes
260140_at 20.2 239.8 4.34E-06 3.57 AtMYB90; production of anthocyanin pigment 2 Atlg66390 PAP2
254283_s_at 41.4 199.6 4.91E-05 2.27 Putative leucoanthocyanidin dioxygenase; putative
anthocyanidin synthase
At4g22870 –
250207_at 130.3 565.0 1.06E-05 2.12 Chalcone synthase (TT4) At5g13930 CHS
249215_at 19.0 79.1 4.09E-05 2.06 Dihydroflavonol 4-reductase (TT3) At5g42800 DFR
245628_at 21.4 88.9 3.68E-05 2.06 AtMYB75; production of anthocyanin pigment 1 Atlg56650 PAPI
252123_at 101.9 299.9 0.000348 1.57 Flavanone 3-hydroxylase (TT6) At3g51240 F3H
250533_at 79.6 226.3 3.67E-05 1.51 Flavonol synthase At5g08640 FLS1
261804_at 67.1 189.2 0.000244 1.50 Flavonoid 3-o-rhamnosyltransferase Atlg30530 UGT78D1
253222_at 19.4 53.9 0.00063 1.49 Chalcone and stilbene synthase family protein At4p34850 –
267147_at 40.4 101.2 7.48E-05 1.33 Putative anthocyanidin synthase; 2oG-Fe(II) oxygenase
family protein
At2g38240 –
255773_at 81.0 189.2 0.000275 1.22 Putative flavonol 40-sulfotransferase; sulfotransferase 17 Atlg18590 SOT17
261907_at 33.8 75.3 2.71E-05 1.15 4-coumarate:CoA lipase 3 Atlg65060 4CL3
251827_at 97.3 211.2 1.57E-05 1.12 Chalcone flavanone isomerase (TT5) At3g55120 CHI
265091_s_at 35.8 75.9 0.0008 1.03 Similar to anthocyanin 5-aromatic acyltransferase Atlg03495 –
245624_at 22.0 45.3 0.005175 1.02 Anthocyanidin 5-o-glucosyl transferase At4gl4090 –
Down-regulated genes
257288_at 155.6 76.6 0.001297 -1.02 Similar to malonyl-CoA; isoflavone 7-o-glucoside-6,-o-
malonyltransferase
At3g29670 –
267337_at 768.4 367.4 2.45E-06 -1.06 Putative anthocyanin 5-aromatic acyltransferase At2g39980 –
254835_s_at 220.7 105.4 0.000183 -1.07 Flavonoid 3,5-hydroxylase -like; member of CYP706A At4g12310 CYP706A5
252534_at 74.9 32.5 0.006755 -1.21 AtMYB 48; production of flavonol glycoside 3 At3g46130
249494_at 174.8 67.1 2.23E-05 -1.38 Anthocyanin 5-aromatic acyltransferase; transferase family
protein
At5g39050 –
249493_at 149.0 46.4 0.000188 -1.69 Anthocyanin 5-aromatic acyltransferase; transferase family
protein
At5g39080 –
Transcript levels of flavonoid-related genes were analyzed in desiccation-treated Col-0 and MYB60.2-ox plants by microarray analysis. Four-
week-old plants grown in soil were dehydrated for 3 days prior to sampling for microarray analysis. 60.2ox stands for MYB60.2 overexpressor
Plant Mol Biol (2011) 77:91–103 99
123

specifically expressed at a low level in roots upon auxin
exposure (Fig. 6). Microarray analysis revealed that several
genes involved in flavonoid biosynthesis and auxin sig-
naling were altered in MYB60.2-ox plants (Tables 1, 2). In
accordance with this molecular phenotype, overexpression
of the splice variants resulted in reduced DR5::GUS
expression in response to auxin in the roots (Fig. 7). Ini-
tially, it was suspected that MYB60 overexpression would
bring about an altered phenotype in transgenic plants.
However, no alterations were detected under normal
growth conditions. As a result, microarray analysis was
utilized to explore the possible targets of MYB60 under
drought conditions (Table 1). The main focus was placed
on the enhanced levels of several flavonoid biosynthetic
genes that are induced in MYB60.2-ox plants experiencing
drought stress (Table 1). From this analysis, it appeared
that flavonoid accumulation in MYB60.2-ox plants were
arising via independent manner from the action of MYB60
in guard cells. One flavonoid synthetic enzyme, chalcone
synthase (CHS), is detected in epidermal cells but not in
guard cells (Zobel and Hrazdina 1995). This indicates that
MYB60 may function in the root where its expression was
observed in response to auxin (Fig. 6). It is well established
that flavonoids function in the modulation of polar auxin
transport as well as in auxin-dependent tropic responses
(Peer and Murphy 2007). Reduced levels of auxin-
responsive DR5 activity and increased expression of flavo-
noid biosynthetic genes were observed in the MYB60.1-ox
and MYB60.2-ox plants (Fig. 7). However, there was no
drastic elevation in anthocyanin accumulation levels
among Col-0, MYB60.1-ox and MYB60.2-ox plants in
response to auxin. Key flavonoid biosynthetic genes, such
as PAP1 or PAP2, have higher levels of transcription when
the plants are exposed to sucrose (Teng et al. 2005). As
demonstrated in Fig. 8, the level of the MYB60 transcript
increased strongly when the plants were exposed to high
levels of sucrose, suggesting that this gene may be involved
in sucrose-induced flavonoid biosynthesis or that a high
concentration of sucrose might cause enhanced osmotic
stress in seedlings. Indeed, MYB60.1 and MYB60.2 tran-
scripts were accumulated in response to 10 lM ABA
(Fig. 5d) whereas MYB60 has been reported to be
decreased by ABA treatment (Cominelli et al. 2005) to the
contrary. It should be noted that these authors used 100 lM
ABA in their experiments while this study used a much
lower concentration (10 lM) of ABA for 3 or 6 h. In this
study, incubation for greater than 24 h with 10 lM ABA
also resulted in decreased levels of MYB60 transcripts (data
not shown). As a major flavonoid biosynthetic gene PAP1
is induced by sucrose and number of flavonoid biosynthetic
genes are up-regulated in MYB60-ox plants (Table 2), a
keen observation of MYB60 regulation in response to
Table 2 Microarray analysis of phytohormone-related genes in drought-treated Col-0 and MYB60.2-ox plants
Probe sets Col-0 60.2OX P value Log2 ratio Description MIPS code
Auxin
250294 at 15.13 64.61 3.98 E-05 2.098 Auxin-reponsive-like protein At5gl3380
245244_at 196.77 620.54 1.29E-06 1.658 IAA-amino acid hydrolase Atlg44350
256178_s_at 332.23 985.95 0.00126 1.572 IAA-amino acid conjugate hydrolase subfamily (ILL5) Atlg51780
234809_at 63.74 186.68 4.9E-05 1.552 Auxin-responsive family protein Artgl2410
251291_at 36.79 17.89 0.007468 -1.069 Auxin-responsive family protein At3g61900
257690_at 615.81 154.71 1.9E-05 -1.997 Auxin-responsive family protein At3gl2830
254323_at 168.31 35.33 3.95E-05 -2.261 Auxin-responsive family protein At4g22620
264014_at 59.25 10.80 6.3E-05 -2.447 Putative auxin-regulated protein At2g21210
GA
266613_at 122.14 403.53 1.14E-05 1.724 Gibberellin-regulated family protein At2gl4900
260141_at 148.72 398.23 4.82E-05 1.424 Negative regulator of GA responses (RGLl); GRAS family Atlg66350
261768_at 71.21 30.05 0.000649 -1.236 Gibberellin 3 beta-hydroxylase (GA3oX1) Atlgl5550
ABA
248227_at 11.02 31.30 0.001641 1.492 ABA-inducible protein-like; similar to pollen coat protein At5g53820
Ethylene
266821_at 319.39 998.82 5.95E-06 1.645 ERF (ethylene response factor) subfamily B-3 (ERF13) At2g44840
253259_at 1598.74 4300.10 1.86E-06 1.427 ERF (ethylene response factor) subfamily B-3 (RRTFl) At4g34410
Cytokinin
253696_at 221.09 80.40 1.35E-05 -1.460 Cytokinin oxidase (CKX4) At4g29740
Transcript levels of phytohormone-related genes were analyzed in Col-0 and MYB60.2-ox plants after drought treatment by microarray analysis.
Four-week-old Col-0 and MYB60.2-ox plants grown in soil were dehydrated for 3 days previous to sampling for microarray analysis
100 Plant Mol Biol (2011) 77:91–103
123

sucrose would provide interesting results. Thus, the tran-
scription level of PAP1 and F3H before and after 8%
sucrose treatment in Col-0 and MYB60 overexpressor
plants were quantified and determined to be elevated upon
stress treatment. However, the role of MYB60 in sucrose
signaling was not extensively investigated in this study. It
is hypothesized that the effect of sucrose may be similar to
osmotic stress.
Robust root growth is an important determinant of plant
proliferation during abiotic stress challenges due to the
total root mass affecting the efficiency of water uptake
(Peret et al. 2009). Many factors regulate root development
with auxin levels playing a critical role. Therefore, regu-
lating auxin distribution to ensure that appropriate levels
are reached in the root tissues is a central aspect of auxin
homeostasis control (Petersson et al. 2009; Ikeda et al.
2009). Based on the observations reported here, a
hypothesis is proposed for the role of MYB60 (Fig. S3).
Under normal conditions, MYB60 is expressed at sub-
stantial levels and mediates stomatal opening. However,
under severe drought conditions, MYB60 is down-regu-
lated in the guard cells. In this case, stomatal opening is no
longer activated, and stomatal closure is triggered by
increased ABA levels (Cominelli et al. 2005). During mild
drought stress, a low level of ABA leads to an increase in
Fig. 7 Histochemical GUS
assays of DR5::GUS x
AtMYB60-ox F3 plants. a GUS
activity in DR5::GUS x
AtMYB60.1-ox and b DR5::GUSx AtMYB60.2-ox seedlings
treated with IAA. DR5::GUS is
included as a control, and GUS
staining was detected in leaves
and roots. Histochemical assays
were conducted to visualize
expression of b-glucurodinase
(GUS) activity in transgenic
plants 24 h after incubation with
0, 0.1, 1, 5 or 10 lM IAA.
Three week-old plants were
stained with GUS solution
overnight, and chlorophyll was
removed by treatment with 70%
ethanol
Fig. 8 MYB60 is responsive to sucrose. For northern analysis, wild-
type seedlings were grown on MS medium for 2 weeks and then
transferred to medium supplemented with 0, 4, 6 or 8% sucrose as
indicated prior to RNA extraction. Transcript levels of MYB60 are
shown. The rRNA is shown as a loading control
Plant Mol Biol (2011) 77:91–103 101
123

MYB60 transcript levels. This alters local flavonoid levels,
leading to the modulation of polar auxin transport, thus
affecting overall root growth (Fig. S3). In this study, an
increase in the flavonoid level of MYB60.1-ox and
MYB60.2-ox plants was not detected. This finding may be
due to the restriction of flavonoid biosynthesis to specific
tissues, or due to low levels of local flavonoid accumula-
tion. Additionally, another type of flavonoid may accu-
mulate in MYB60.1-ox and MYB60.2-ox plants. This
possibility was not investigated in the current study.
Therefore, during the initial stages of stress, the plant
system may modulate root growth behavior by regulating
MYB60 expression, which would result in the growth of
roots with an increased capacity for water uptake. In con-
trast, severe drought stress blocks MYB60 expression,
resulting in stomatal closure and root growth inhibition.
The dual role of MYB60 in stomatal function and root
growth reflects the possibility that MYB60 may target and
regulate different downstream genes in different tissues, as
evidenced in this study. However, the potential alternative
roles of the MYB60 splice variants remain to be fully
explored. Realizing that MYB60 overexpression results in
enhanced leaf withering in response to water deficit, the
engineering of MYB60 to enhance plant stress tolerance
may be a challenge. This is despite the knowledge that
MYB60 improves root growth under the same abiotic
stresses in the presence of high humidity. Analysis of the
fusion of MYB60 to a root-specific promoter will provide
insights into its role in water uptake and potentially
contribute to strategies for responding to unfavorable
environmental stresses.
Acknowledgments This work was supported by a grant from the
Korea Research Foundation to Hojoung Lee (2009; Grant #2011-
0003726) in part by a grant from the Korea Research Foundation to
Suk-Whan Hong (grant #2011-0003259 and #2011-0018393). The
authors express their gratitude to Dr. Cominelli for providing seeds of
the myb60 knock-out line.
References
Blatt MR (2000) Cellular signaling and volume control in stomatal
movements in plants. Annu Rev Cell Dev Biol 16:21–241
Chen Y, Yang X, He K, Liu M, Li J, Gao Z, Lin Z, Zhang Y, Wang X,
Qiu X, Shen Y, Zhang L, Deng X, Luo J, Deng XW, Chen Z, Gu
H, Qu LJ (2006) The MYB transcription factor superfamily of
Arabidopsis: Expression analysis and phylogenetic comparison
with the rice MYB family. Plant Mol Biol 60:09–126
Clough SJ, Bent AF (1998) Floral dip: a simplified method for
Agrobacterium-mediated transformation of Arabidopsis thali-ana. Plant J 16:35–43
Cominelli E, Galbiati M, Vavasseur A, Conti L, Sala T, Vuylsteke M,
Leonhardt N, Dellaporta S, Tonelli C (2005) A guard-cell
specific MYB transcription factor regulates stomatal movements
and plant drought tolerance. Curr Biol 15:196–1200
Dubos C, Stracke R, Grotewold E, Weisshaar B, Martin C, Lepiniec L
(2010) MYB transcription factors in Arabidopsis. Trends Plant
Sci 15:73–581
Hetherington AM (2001) Guard cell signaling. Cell 107:711–714
Ikeda Y, Men S, Fischer U, Stepanova AN, Alonso JM, Ljung K,
Grebe M (2009) Local auxin biosynthesis modulates gradient-
Fig. 9 Expression levels of flavonoid biosynthetic genes in MYB60-
ox seedlings after sucrose treatment. qRT-PCRs were done to assess
the relative mRNA levels of the major flavonoid biosynthetic genes,
such as a PAP1, and b F3H, determined to be highly expressed in
MYB60-ox plants according to the microarray analysis in Table 2.
Analysis of upregulated flavonoid biosynthetic genes upon 8%
sucrose treatment was done and actin was used as internal control.
Col-0, MYB60.1-ox#16 and MYB60.2-ox#7 seeds were germinated on
MS-agar medium and 2 week-old seedlings were treated with
sucrose-containing MS liquid media for 5 h. Error bars represent
the standard deviation
102 Plant Mol Biol (2011) 77:91–103
123

directed planar polarity in Arabidopsis. Nat Cell Biol
11:731–738
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: beta-
glucuronidase as a sensitive and versatile gene fusion marker in
higher plants. EMBO J 20:901–3907
Jia L, Clegg MT, Jiang T (2004) Evolutionary dynamics of the DNA-
binding domains in putative R2R3-MYB genes identified from
rice subspecies indica and japonica genomes. Plant Physiol
134:75–585
Jung C, Seo JS, Han SW, Koo YJ, Kim CH, Song SI, Nahm BH, Choi
YD, Cheong JJ (2008) Overexpression of AtMYB44 enhances
stomatal closure to confer abiotic stress tolerance in transgenic
Arabidopsis. Plant Physiol 146:23–635
Lee H, Guo Y, Ohta M, Xiong L, Stevenson B, Zhu JK (2002) LOS2,
a genetic locus required for cold-responsive gene transcription
encodes a bifunctional enolase. EMBO J 21:692–2702
Leonhardt N, Kwak JM, Robert N, Waner D, Leonhardt G, Schroeder
JI (2004) Microarray expression analyses of Arabidopsis guard
cells and isolation of a recessive abscisic acid hypersensitive
protein phosphatase 2C mutant. Plant Cell 16:596–615
Liang YK, Dubois C, Dodd IC, Holroyd GH, Hetherington AM,
Campbell MM (2005) AtMYB61, an R2R3-MYB transcription
factor controlling stomatal aperture in Arabidopsis thaliana.
Curr Biol 15:201–1206
Pandey S, Zhang W, Assmann SM (2007) Roles of ion channels and
transporters in guard cell signal transduction. FEBS Lett
581:325–2336
Peer WA, Murphy AS (2007) Flavonoids and auxin transport:
modulators or regulators? Trends Plant Sci 12:56–563
Peret B, Rybel BD, Casimiro I, Benkova E, Swarup R, Laplaze L,
Beeckman T, Bennett MJ (2009) Arabidopsis lateral root
development: an emerging story. Trends Plant Sci 14:399–408
Petersson SV, Johansson AI, Kowalczyk M, Makoveychuk A, Wang
JY, Moritz T, Grebe M, Benfey PN, Sandberg G, Ljung K (2009)
An auxin gradient and maximum in the Arabidopsis root apex
shown by high-resolution cell-specific analysis of IAA distribu-
tion and synthesis. Plant Cell 21:1659–1668
Roelfsema MRG, Hedrich R (2005) In the light of stomatal opening:
new insights into ‘the Watergate’. New Phytol 165:65–691
Rosinsky JA, Atchley WR (1998) Molecular evolution of the MYB
family of transcription factors: evidence for polyphyletic origin.
J Mol Evol 46:4–83
Sahota AP, Dhoot GK (2009) A novel SULF1 splice variant inhibits
Wnt signalling but enhances angiogenesis by opposing SULF1
activity. Exp Cell Res 315:752–2764
Schroeder JI, Kwak JM, Allen GJ (2001) Guard cell abscisic acid
signalling and engineering of drought hardiness in plants. Nature
410:27–330
Seo PJ, Xiang F, Qiao M, Park JY, Lee YN, Kim SG, Lee YH, Park
WJ, Park CM (2009) The MYB96 transcription factor mediates
abscisic acid signaling during drought stress response in
Arabidopsis. Plant Physiol 151:275–289
Shinozaki K, Yamaguchi-Shinozaki K (2000) Molecular responses to
dehydration and low temperature: differences and cross-talk
between two stress signaling pathways. Curr Opin Plant Biol
3:17–223
Shinozaki K, Yamaguchi-Shinozaki K, Seki M (2003) Regulatory
network of gene expression in the drought and cold stress
responses. Curr Opin Plant Biol 6:410–417
Sirichandra C, Wasilewska A, Vla F, Valon C, Leung J (2009) The
guard cell as a single-cell model towards understanding drought
tolerance and abscisic acid action. J Exp Bot 60:1439–1463
Teng S, Keurentjes J, Bentsink L, Koornneef M, Smeekens S (2005)
Sucrose-specific induction of anthocyanin biosynthesis in Ara-
bidopsis requires the MYB75/PAP1 gene. Plant Physiol
139:1840–1852
Wasilewska A, Vlad F, Sirichandra C, Redko Y, Jammes F, Valon C,
Frey N, Leung J (2008) An update on abscisic acid signaling in
plants and more. Mol Plant 1:198–217
Xiong L, Lee H, Ishitani M, Zhu JK (2002) Regulation of osmotic
stress-responsive gene expression by the LOS6/ABA1 locus in
Arabidopsis. J Biol Chem 277:8569–8588
Zhu JK (2002) Salt and drought stress signal transduction in plants.
Ann Rev Plant Biol 53:247–273
Zobel AM, Hrazdina G (1995) Chalcone synthase localization in early
stages of plant development. I. Immunohistochemical use of
plasmolysis for localizing the enzyme in epidermal cell
cytoplasm of illuminated buckwheat hypocotyls. Biotech Histo-
chem 70:1–6
Plant Mol Biol (2011) 77:91–103 103
123




















