
A comparison of living and dead molluscs on coral reef associated hard substrata in the northern Red...
24
Palaeogeography, Palaeoclimatology, Palaeoecology 159 ( 2000) 167–190 www.elsevier.nl/ locate/palaeo A comparison of living and dead molluscs on coral reef associated hard substrata in the northern Red Sea — implications for the fossil record Martin Zuschin a, * , Johann Hohenegger a, Fritz F. Steininger b a Institut fu ¨r Pala ¨ontologie, Universita ¨t Wien, Althanstraße 14, A-1090 Vienna, Austria b Forschungsinstitut und Naturmuseum Senckenberg, Senckenberganlage 25, D-60325 Frankfurt am Main, Germany Received 4 May 1999; received in revised form 7 December 1999; accepted for publication 22 December 1999 Abstract Fidelity of death assemblages to live shelly faunas is one of the major palaeontological questions, but quantitative data are scarce and most case studies on this topic have been performed in non-reef sediments. Therefore we studied di erent types of subtidal reef-associated hard substrata (reef flats, reef slopes, coral carpets, coral patches, rock grounds), each with di erent coral associations, in order to determine the agreement of assemblages of living and dead shell-bearing molluscs. A total area of 340.5 m2 was investigated and 2846 individuals were counted at 68 sample localities ranging from shallow subtidal to 40 m water depth. Most taxa found dead in the study area were also found live and vice versa; di erences in this respect can be related to quantitatively unimportant taxa. However, strong di erences exist in the proportion of living and dead fauna, dominant taxa, and molluscan distribution patterns. The ratio of live to dead molluscs is high. Living molluscs are strongly dominated by taxa with distinct relations to corals, mainly Pedum, Coralliophila and Tridacna, and the encrusting gastropod Dendropoma. Five distinct groups of living molluscs can be di erentiated and related to specific hard substrata, which are characterized by distinct molluscan life habits. In contrast, the death assemblages are always strongly dominated by encrusting bivalves, mainly Chamoidea and Spondylidae, and cerithiid gastropods in varying dominances. Correspondingly there is no significant correlation of the total abundance of living and dead molluscs and their overall similarity is only 6%. Similarity between living and dead faunas is above 50% at only 12 of the 68 sample locations, and at 17 sample locations significant correlations of living and dead molluscs were recognized. These correlations are mainly based on Chamoidea, which dominate both the living and dead fauna, on rock grounds. Therefore rock grounds are the only bottom type with consistent correlations and similarities of living and dead molluscs. The observed bias is due to the close relationship of molluscan life habits and post-mortem history of shells. Molluscs that live permanently attached to or within living corals (mostly bivalves and encrusting Dendropoma) can easily be overgrown after death by the large amounts of living substrata available. Rapid transport of dead shells into surrounding sediments or into crevices within corals is typical of gastropods that feed on corals. Molluscs that colonize dead surfaces preferentially accumulate on rock grounds. The ecologic information that can be derived from the shells depends on the di erent post-mortem histories of the faunas. © 2000 Elsevier Science B.V. All rights reserved. Keywords: mollusca; taphonomy; palaeoecology; coral reef; recent; Red Sea * Corresponding author. Fax: +43-1-31336-784. E-mail address: [email protected] ( M. Zuschin) 0031-0182/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved. PII: S0031-0182(00)00045-6
Transcript of A comparison of living and dead molluscs on coral reef associated hard substrata in the northern Red...

Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190www.elsevier.nl/locate/palaeo
A comparison of living and dead molluscs on coral reefassociated hard substrata in the northern Red Sea —
implications for the fossil record
Martin Zuschin a,*, Johann Hohenegger a, Fritz F. Steininger ba Institut fur Palaontologie, Universitat Wien, Althanstraße 14, A-1090 Vienna, Austria
b Forschungsinstitut und Naturmuseum Senckenberg, Senckenberganlage 25, D-60325 Frankfurt am Main, Germany
Received 4 May 1999; received in revised form 7 December 1999; accepted for publication 22 December 1999
Abstract
Fidelity of death assemblages to live shelly faunas is one of the major palaeontological questions, but quantitativedata are scarce and most case studies on this topic have been performed in non-reef sediments. Therefore we studieddifferent types of subtidal reef-associated hard substrata (reef flats, reef slopes, coral carpets, coral patches, rockgrounds), each with different coral associations, in order to determine the agreement of assemblages of living anddead shell-bearing molluscs. A total area of 340.5 m2 was investigated and 2846 individuals were counted at 68 samplelocalities ranging from shallow subtidal to 40 m water depth. Most taxa found dead in the study area were also foundlive and vice versa; differences in this respect can be related to quantitatively unimportant taxa. However, strongdifferences exist in the proportion of living and dead fauna, dominant taxa, and molluscan distribution patterns. Theratio of live to dead molluscs is high. Living molluscs are strongly dominated by taxa with distinct relations to corals,mainly Pedum, Coralliophila and Tridacna, and the encrusting gastropod Dendropoma. Five distinct groups of livingmolluscs can be differentiated and related to specific hard substrata, which are characterized by distinct molluscanlife habits. In contrast, the death assemblages are always strongly dominated by encrusting bivalves, mainly Chamoideaand Spondylidae, and cerithiid gastropods in varying dominances. Correspondingly there is no significant correlationof the total abundance of living and dead molluscs and their overall similarity is only 6%. Similarity between livingand dead faunas is above 50% at only 12 of the 68 sample locations, and at 17 sample locations significant correlationsof living and dead molluscs were recognized. These correlations are mainly based on Chamoidea, which dominateboth the living and dead fauna, on rock grounds. Therefore rock grounds are the only bottom type with consistentcorrelations and similarities of living and dead molluscs. The observed bias is due to the close relationship ofmolluscan life habits and post-mortem history of shells. Molluscs that live permanently attached to or within livingcorals (mostly bivalves and encrusting Dendropoma) can easily be overgrown after death by the large amounts ofliving substrata available. Rapid transport of dead shells into surrounding sediments or into crevices within corals istypical of gastropods that feed on corals. Molluscs that colonize dead surfaces preferentially accumulate on rockgrounds. The ecologic information that can be derived from the shells depends on the different post-mortem historiesof the faunas. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: mollusca; taphonomy; palaeoecology; coral reef; recent; Red Sea
* Corresponding author. Fax: +43-1-31336-784.E-mail address: [email protected] (M. Zuschin)
0031-0182/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved.PII: S0031-0182 ( 00 ) 00045-6

168 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
1. Introduction (sensu Bromley, 1975), rock grounds (sensuGoldring, 1995), cobbles (e.g. Surlyk andChristensen, 1974; Wilson, 1987) and a wide rangeOne of the major questions in considering the
quality of the fossil record is how accurately shelly of skeletons (e.g. Nebelsick et al., 1997; Zuschinand Piller, 1997a), most of which might be referreddeath assemblages preserve the composition and
structure of the shelled portion of the original to as shell grounds (after Dodd and Stanton,1990). Coral colonies, in contrast, are not only thecommunity (see review by Kidwell and Bosence,
1991). In spite of its importance, only few tapho- primary frame builders of coral reefs (for a reviewsee Scoffin, 1992), but are also the major type ofnomic investigations in modern environments have
provided data to answer this question, and com- hard substrata characterized by a living surface.Large parts of coral reefs provide such a livingparisons between living and dead faunas in marine
environments are mainly restricted to molluscan surface (e.g. Liddell and Ohlhorst, 1988) and,especially among molluscs, a wide range of taxaassemblages in soft sediments (e.g. Cadee, 1968;
Warme, 1969, 1971; Staff et al., 1986; Carthew and and different life habits are closely linked to suchsubstrata (e.g. Hadfield, 1976; Morton, 1983a;Bosence, 1986; Fursich and Flessa, 1987; Flessa
and Fursich, 1991). However, after removing sam- Kleemann, 1990, 1992; Schuhmacher, 1993).The coral-dominated shallow marine sub-pling artifacts, the agreement between live:dead
assemblages in these studies is generally considered tropical Northern Bay of Safaga (Fig. 1) is com-posed of various sedimentary facies that are closelyto be high (for reviews see Kidwell and Flessa,
1995; Kidwell, in press). Only a few studies com- associated with different types of hard substrata,including reef flats, reef slopes, coral carpets, coralpare the living and dead faunas of hard substrata
(e.g. Pandolfi and Minchin, 1995; Pandolfi and patches and rock bottoms (Piller and Pervesler,1989). These types of hard substrata are charac-Greenstein, 1997a; Greenstein and Pandolfi, 1997),
and to our knowledge there is no study dealing terized by different coral associations and bydifferent degrees of surface coverage (Riegl andquantitatively with the distribution of living and
dead molluscs in coral reef environments, which Velimirov, 1994; Riegl and Piller, 1997). The distri-butions of living bivalves are correlated withusually provide a close interfingering of living and
dead hard substrata. environmental gradients and the molluscan assem-blages characteristic of the different facies haveThe relation between live and dead assemblages
of hard and soft substrata probably differs signifi- different life/death ratios (Zuschin and Piller,1997b,c).cantly, because of some important differences
regarding composition of living communities and The objectives of the present study are to: (1)evaluate the degree of coincidence between livingaccumulation of their hard parts. Soft sediments
are mainly colonized by sessile or vagile infauna and dead molluscan faunas in a setting of mixedliving and dead hard substrata, and (2) determineand epifauna, and their shelly death assemblages
often map communities better than any single the influence of substratum type and molluscanlife habits on live:dead agreement.census of the living shelled community (e.g.
Carthew and Bosence, 1986). This is due to timeaveraging and low aggradation rates in mostmarine sediments, which smooth out the patchy 2. Materials and methodsdistribution of most communities (for a review seeKidwell and Bosence, 1991). In contrast, most Different subtidal hard substrata were sampled
for shelled molluscs at 68 localities (Fig. 2, Table 1)types of hard substrata provide a dead surfacethat is mainly populated by encrusting and endo- with a 0.25 m2 aluminium square frame. At each
locality the location of the first frame was selectedlithic colonizers. These usually become entombedin their life sites and thus allow the detailed study randomly, the following frames were positioned
along a line extending from the point. In mixedof faunal succession (Brett, 1988). Among suchhard substrata are, for example, hard grounds hard substrata/loose ground and/or soft-bottom

169M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Fig. 1. Location map and general bathymetry of the study area (after Piller and Pervesler, 1989). Dense stippled fields in the rightmap are intertidal areas. AM=aerial mast, H=‘Safaga Hotel’.
habitats, frames were only placed on hard substra- (Table 2). The present study is based on quantita-tive samples from the seafloor and from easilytum. A mean of 5.0 m2 (±1.7) of seafloor was
investigated per locality, with a range from 3 to accessible cavities. Due to methodological limita-tions, deeper cavities within coral reefs have not11 m2 (Table 2). The number of investigated
frames depended on mollusc density ( low density been considered (e.g. fig. 6 in Scoffin, 1992).Samples were taken during daylight (usuallyrequired a higher number of frames and vice versa)
and on water depth (greater water depths required between 10 am and 5 pm). The molluscs wereidentified either immediately in their environmentshorter dive times). The mean water depth was
14.3 m (±9.9) with a range from 1 to 40 m (most bivalves and sessile gastropods) or were

170 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
collected (many vagile gastropods) for moredetailed investigation. The fauna was separatedinto live and dead individuals and hermit crab-occupied shells. The information on live and deadindividuals was used for the statistical analysis;shells occupied by hermit crabs were treated as animportant descriptive feature affecting the distribu-tion of dead gastropod shells.
Due to a highly questionable taxonomy,Chamoidea, Spondylidae and Ostreoidea, exceptfor Lopha cristagalli, were not differentiated tospecies level; most of the taxonomic studies in factmainly describe ecophenotypes (Oliver, 1992,1995). Some other taxa were identified only abovethe species level due to problems with identificationin the field or poor preservation (e.g. Conus spp.).
It is difficult to recognize smaller molluscs bySCUBA diving. Therefore, to provide a consistentdatabase, molluscs<2 cm were excluded from thequantitative treatment. Most boring bivalves (e.g.gastrochaenids, lithophagins) have been excludedfrom the quantitative collection because they aredifficult to identify from the surficial appearanceof their small bore holes. A qualitative study ofboring bivalves has been presented by Kleemann(1992).
The complete database contains 54 taxa, butstatistical analysis was confined to the 18 taxa thateach contribute more than 1% to either the live orFig. 2. Sample locations. AM=aerial mast, H=‘Safaga Hotel’.dead mollusc content of all samples (Fig. 3,
Table 1Classification of the investigated hard substrata and corresponding assignment of sample sites. Classification of hard substratamodified after data provided in Piller and Pervesler (1989), Riegl and Velimirov (1994) and Riegl and Piller (1997, 1999)
Hard substrata Coral associations Living coral coverage (%) Sample locations
Reef flats Stylophora association 30 1, 2, 3Reef slopes Acropora association 40–80 15, 16, 17
Porites association 50–100 18, 19, 20Acropora–Millepora association 50 25, 26, 27, 38Porites–Millepora association 80–100 14Stylophora association <20 64
Coral carpets Porites association 60–90 4, 21, 59, 62, 63, 66, 67, 68faviid association 20–30 5, 7, 9, 11, 12, 13, 29, 30, 32, 33, 34,
35, 41, 43, 44, 47, 48, 51, 53, 58, 60, 61depauperate faviid association <20 6, 36, 37, 50
Rock grounds Sarcophyton association 10–20 10, 22, 31, 46, 55, 56, 57platy scleractinian association 30–50 8, 23, 42, 45, 49, 52, 54, 65
Coral patches Acropora patch association 60–80 24, 28, 40Stylophora–Acropora patch association 60–80 39
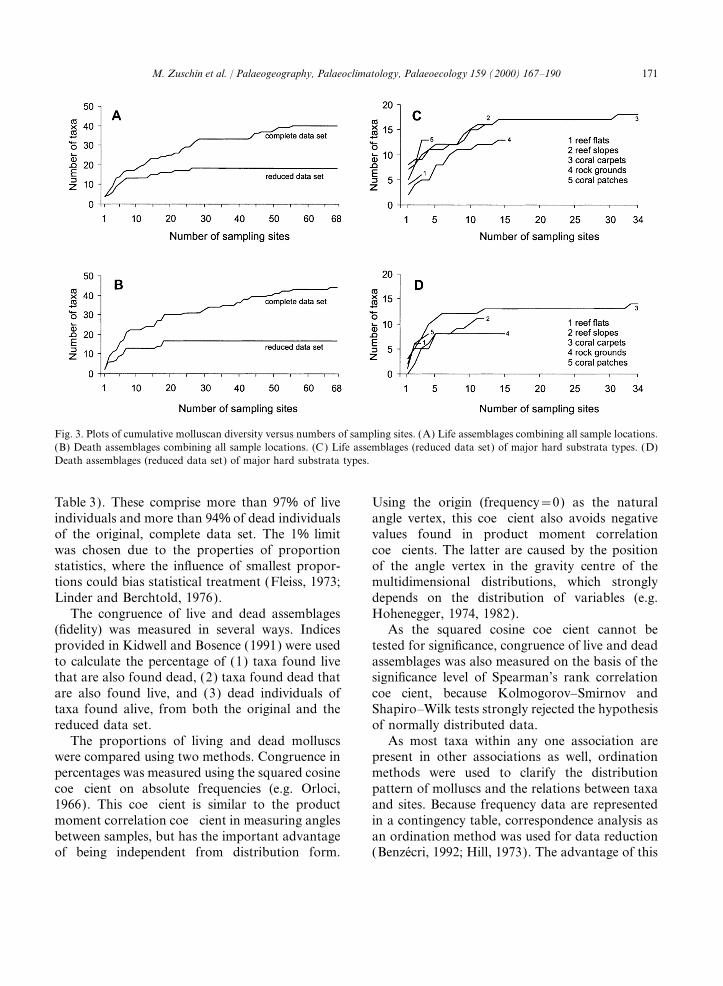
171M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Fig. 3. Plots of cumulative molluscan diversity versus numbers of sampling sites. (A) Life assemblages combining all sample locations.(B) Death assemblages combining all sample locations. (C) Life assemblages (reduced data set) of major hard substrata types. (D)Death assemblages (reduced data set) of major hard substrata types.
Table 3). These comprise more than 97% of live Using the origin (frequency=0) as the naturalangle vertex, this coefficient also avoids negativeindividuals and more than 94% of dead individuals
of the original, complete data set. The 1% limit values found in product moment correlationcoefficients. The latter are caused by the positionwas chosen due to the properties of proportion
statistics, where the influence of smallest propor- of the angle vertex in the gravity centre of themultidimensional distributions, which stronglytions could bias statistical treatment (Fleiss, 1973;
Linder and Berchtold, 1976). depends on the distribution of variables (e.g.Hohenegger, 1974, 1982).The congruence of live and dead assemblages
(fidelity) was measured in several ways. Indices As the squared cosine coefficient cannot betested for significance, congruence of live and deadprovided in Kidwell and Bosence (1991) were used
to calculate the percentage of (1) taxa found live assemblages was also measured on the basis of thesignificance level of Spearman’s rank correlationthat are also found dead, (2) taxa found dead that
are also found live, and (3) dead individuals of coefficient, because Kolmogorov–Smirnov andShapiro–Wilk tests strongly rejected the hypothesistaxa found alive, from both the original and the
reduced data set. of normally distributed data.As most taxa within any one association areThe proportions of living and dead molluscs
were compared using two methods. Congruence in present in other associations as well, ordinationmethods were used to clarify the distributionpercentages was measured using the squared cosine
coefficient on absolute frequencies (e.g. Orloci, pattern of molluscs and the relations between taxaand sites. Because frequency data are represented1966). This coefficient is similar to the product
moment correlation coefficient in measuring angles in a contingency table, correspondence analysis asan ordination method was used for data reductionbetween samples, but has the important advantage
of being independent from distribution form. (Benzecri, 1992; Hill, 1973). The advantage of this

172 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Table 2Basic data on sample locations (water depth, investigated area, molluscan density) and results of correlation and similarity analysison living and dead molluscs. Bold type=1% significance level, underlined type=5% significance level
Site Water Investigated Mollusc density (per 0.25 m2) % Similarity Significancedepth (m) area (m2) level
Living fauna Dead fauna
Mean St. dev. Min. Max. Mean St. dev. Min. Max.
1 1 4.5 11.8 8.2 0 30 0.2 0.5 0 2 0.0 0.7932 1 8 2.3 3.0 0 13 0.2 0.4 0 1 22.7 0.1053 1 5.75 5.0 6.7 0 25 0.1 0.5 0 2 0.0 0.7764 6 3 9.3 6.1 2 21 0.1 0.3 0 1 32.0 0.0245 10 3 2.8 2.6 0 10 1.3 1.1 0 3 67.6 0.0006 6 4 1.8 1.4 0 5 1.0 0.7 0 2 61.8 0.0007 15 4 0.8 1.2 0 4 0.6 0.7 0 2 63.9 0.0008 23 3 1.1 1.2 0 4 1.7 1.6 0 5 98.7 0.0009 18 3 1.8 3.1 0 11 0.4 0.7 0 2 26.3 0.08610 24 3 0.4 0.8 0 2 1.5 1.2 0 4 49.8 0.00211 17 4 1.6 3.8 0 13 0.3 0.6 0 2 0.0 0.74712 17 4 0.9 1.5 0 5 0.6 0.6 0 2 29.0 0.10313 12 4 2.6 3.4 0 13 1.0 1.3 0 4 12.6 0.51214 1 6 1.3 1.8 0 6 0.0 0.0 0 0 0.0 –15 13 6 0.6 1.1 0 4 0.1 0.3 0 1 0.6 0.95416 8 6 0.1 0.3 0 1 0.3 0.9 0 4 16.7 0.26517 5 6 0.7 0.9 0 2 0.1 0.4 0 2 4.0 0.91318 6 3 9.5 5.9 1 20 0.3 0.7 0 2 24.8 0.08019 4 3 3.5 2.8 0 8 0.0 0.0 0 0 0.0 –20 2 3 6.4 2.5 4 13 0.0 0.0 0 0 0.0 –21 10 6 2.3 1.5 0 5 0.2 0.4 0 1 69.9 0.00022 28 3 0.3 0.5 0 1 0.8 1.1 0 4 78.8 0.00023 30 6 0.0 0.0 0 0 0.0 0.2 0 1 0.0 –24 4 3 1.5 1.8 0 6 0.6 0.7 0 2 11.0 0.41425 9 6 0.7 1.1 0 3 0.3 0.6 0 2 0.0 0.53126 5 6 1.1 2.0 0 9 0.0 0.2 0 1 0.0 0.55327 5 8 1.4 2.4 0 12 0.4 0.9 0 3 6.2 0.64628 11 6 1.6 3.1 0 14 0.4 0.5 0 1 0.9 0.98129 12 6 0.9 1.3 0 4 0.3 0.6 0 2 1.7 0.79630 18 11 0.6 0.8 0 3 0.1 0.3 0 1 21.0 0.13631 6 5.25 0.9 1.0 0 3 1.1 1.5 0 7 78.6 0.00032 11 8 2.5 3.0 0 15 0.1 0.4 0 2 7.5 0.57733 20 6 1.0 1.1 0 4 0.1 0.3 0 1 0.1 0.61234 9 6 2.4 2.0 0 10 0.4 1.0 0 4 8.1 0.56335 15 4 1.9 1.9 0 6 0.4 0.6 0 2 5.0 0.61736 6 6 1.6 1.8 0 6 0.4 1.0 0 4 19.2 0.19837 10 6 1.1 1.3 0 4 0.3 0.6 0 2 22.4 0.22238 16 6 0.8 1.6 0 7 0.1 0.3 0 1 1.9 0.97139 7 7 1.3 1.5 0 6 0.1 0.4 0 1 2.9 0.77240 4 6 0.4 0.8 0 3 0.6 0.8 0 3 35.9 0.01941 15 8 0.5 0.9 0 4 0.4 0.7 0 3 4.8 0.67342 33 7 0.1 0.4 0 2 0.4 0.5 0 1 97.0 0.00043 22 7 0.6 0.8 0 3 0.3 0.5 0 2 1.5 0.74244 19 8 0.7 1.4 0 7 0.2 0.5 0 2 0.1 0.61045 30 4 0.2 0.4 0 1 0.3 0.4 0 1 8.3 0.63746 17 3.5 0.9 0.9 0 3 2.0 1.7 0 5 95.1 0.00047 25 7 0.7 1.1 0 4 0.3 0.5 0 2 1.5 0.91548 34 5 0.4 0.9 0 3 0.4 0.7 0 3 1.0 0.790
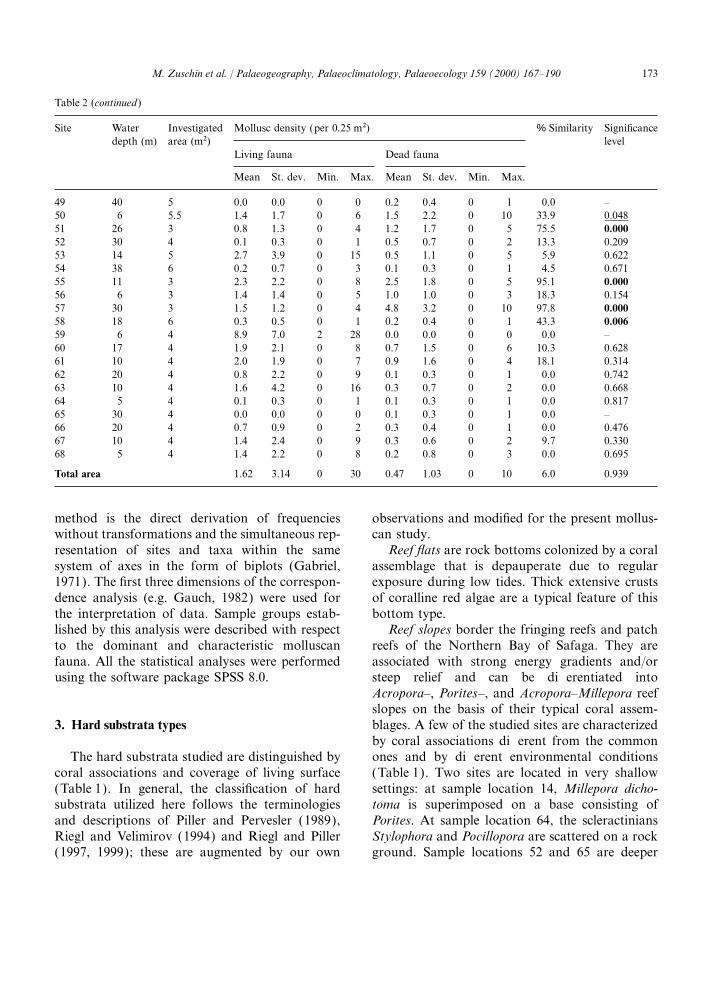
173M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Table 2 (continued )
Site Water Investigated Mollusc density (per 0.25 m2) % Similarity Significancedepth (m) area (m2) level
Living fauna Dead fauna
Mean St. dev. Min. Max. Mean St. dev. Min. Max.
49 40 5 0.0 0.0 0 0 0.2 0.4 0 1 0.0 –50 6 5.5 1.4 1.7 0 6 1.5 2.2 0 10 33.9 0.04851 26 3 0.8 1.3 0 4 1.2 1.7 0 5 75.5 0.00052 30 4 0.1 0.3 0 1 0.5 0.7 0 2 13.3 0.20953 14 5 2.7 3.9 0 15 0.5 1.1 0 5 5.9 0.62254 38 6 0.2 0.7 0 3 0.1 0.3 0 1 4.5 0.67155 11 3 2.3 2.2 0 8 2.5 1.8 0 5 95.1 0.00056 6 3 1.4 1.4 0 5 1.0 1.0 0 3 18.3 0.15457 30 3 1.5 1.2 0 4 4.8 3.2 0 10 97.8 0.00058 18 6 0.3 0.5 0 1 0.2 0.4 0 1 43.3 0.00659 6 4 8.9 7.0 2 28 0.0 0.0 0 0 0.0 –60 17 4 1.9 2.1 0 8 0.7 1.5 0 6 10.3 0.62861 10 4 2.0 1.9 0 7 0.9 1.6 0 4 18.1 0.31462 20 4 0.8 2.2 0 9 0.1 0.3 0 1 0.0 0.74263 10 4 1.6 4.2 0 16 0.3 0.7 0 2 0.0 0.66864 5 4 0.1 0.3 0 1 0.1 0.3 0 1 0.0 0.81765 30 4 0.0 0.0 0 0 0.1 0.3 0 1 0.0 –66 20 4 0.7 0.9 0 2 0.3 0.4 0 1 0.0 0.47667 10 4 1.4 2.4 0 9 0.3 0.6 0 2 9.7 0.33068 5 4 1.4 2.2 0 8 0.2 0.8 0 3 0.0 0.695
Total area 1.62 3.14 0 30 0.47 1.03 0 10 6.0 0.939
method is the direct derivation of frequencies observations and modified for the present mollus-can study.without transformations and the simultaneous rep-
resentation of sites and taxa within the same Reef flats are rock bottoms colonized by a coralassemblage that is depauperate due to regularsystem of axes in the form of biplots (Gabriel,
1971). The first three dimensions of the correspon- exposure during low tides. Thick extensive crustsof coralline red algae are a typical feature of thisdence analysis (e.g. Gauch, 1982) were used for
the interpretation of data. Sample groups estab- bottom type.Reef slopes border the fringing reefs and patchlished by this analysis were described with respect
to the dominant and characteristic molluscan reefs of the Northern Bay of Safaga. They areassociated with strong energy gradients and/orfauna. All the statistical analyses were performed
using the software package SPSS 8.0. steep relief and can be differentiated intoAcropora–, Porites–, and Acropora–Millepora reefslopes on the basis of their typical coral assem-blages. A few of the studied sites are characterized3. Hard substrata typesby coral associations different from the commonones and by different environmental conditionsThe hard substrata studied are distinguished by
coral associations and coverage of living surface (Table 1). Two sites are located in very shallowsettings: at sample location 14, Millepora dicho-(Table 1). In general, the classification of hard
substrata utilized here follows the terminologies toma is superimposed on a base consisting ofPorites. At sample location 64, the scleractiniansand descriptions of Piller and Pervesler (1989),
Riegl and Velimirov (1994) and Riegl and Piller Stylophora and Pocillopora are scattered on a rockground. Sample locations 52 and 65 are deeper(1997, 1999); these are augmented by our own

174 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Table 3Ecological properties of the 18 most abundant taxa and their corresponding classification into six life habit groups using integratedtaxonomic and life habit information. Group 1: bivalves associated with living corals. Group 2: bivalves encrusting dead hardsubstrata. Group 3: bivalve crevice dwellers in dead hard substrata. Group 4: encrusting gastropods. Group 5: gastropods feedingon living corals. Group 6: gastropods that avoid living corals
Taxon Substrate relations Feeding strategy Classification
Barbatia foliata (Forsskal, 1775) byssally attached in crevices of living Porites spp. suspension feeder 1Barbatia setigera (Reeve, 1844) byssally attached in crevices of dead hard substrate suspension feeder 3Chamoidea encrusted to dead hard substrates suspension feeder 2Ctenoides annulata (Lamarck, 1819) byssally attached in crevices of dead hard substrate suspension feeder 3Isognomon legumen (Gmelin, 1791) byssally attached in crevices of dead hard substrate suspension feeder 3Lima lima (Linnaeus, 1758) byssally attached in crevices of dead hard substrate suspension feeder 3Lopha cristagalli (Linnaeus, 1758) encrusted to dead hard substrates suspension feeder 2Ostreoidea encrusted to dead hard substrates suspension feeder 2Pedum spondyloideum (Gmelin, 1791) boring and byssally attached in living coral suspension feeder 1Pteriidae byssally attached to living corals suspension feeder 1Spondylidae encrusted to dead hard substrates suspension feeder 2Streptopinna saccata (Linnaeus, 1758) byssally attached in crevices of living and dead suspension feeder 1
hard substrateTridacna maxima (Roding, 1798) byssally attached among corals or within corals various food sources 1Cerithium spp. vagile on hard substrates algal detritus feeder 6Conus spp. vagile on hard substrates predator 6Coralliophila neritoidea (Lamarck, 1816) vagile on living Porites spp. parasitic 5Dendropoma maxima (Sowerby, 1825) encrusted to living corals/dead hard substrates suspension feeder 4Drupella cornus (Roding, 1798) vagile on living corals (acroporans) predator 5
reef slope settings and consist of rock grounds Coral patches (sensu Piller and Pervesler, 1989)are a special reef type, which is widespread in thewith a patchy platy scleractinian cover.
Coral carpets (sensu Reiss and Hottinger, 1984) Northern Bay of Safaga, commonly occurring inshallow water on sandy substrata. Coral patchesoccur over wide areas of the Northern Bay of
Safaga, to a depth of 40 m; they are up to 3 m are only a few metres in diameter and do not showa clear ecological zonation (Riegl and Piller, 1999).thick in shallow settings. Coral carpets build a
framework, but lack a distinct zonation since they They can be differentiated into two major groupsbased on their coral composition: Acropora– andonly grow in areas without distinct gradients (Riegl
and Piller, 1999). On the basis of coral assemblages Stylophora–Acropora coral patches; typically coralpatches grade into faviid carpets.they can be differentiated into Porites and
faviid carpets, which grade into one another.Depauperate faviid carpets are characterized byless than 20% of living coral coverage (Table 1). 4. Life habits of molluscsOn coral carpets, the highly variable heights ofindividual coral colonies create diverse molluscan The 18 taxa considered in the statistical analysis
consist of 13 bivalves and five gastropods, whichhabitats.Rock grounds (sensu Goldring, 1995) consist of are classified using integrated taxonomic and life
habit information provided in Fankboner and Reidnon-framework building Sarcophyton carpets(Riegl and Piller, 1999) on bare rocky surfaces (1990), Goreau et al. (1973), Hadfield (1976),
Hughes and Lewis (1974), Kleemann (1992),and platy scleractinian carpets, where distinct mol-luscan habitats are lacking except for the high Kohn (1983), Kohn and Leviten (1976), Leviten
and Kohn (1980), Morton (1983a,b), Reicheltamount of exposed coral limestone between thefaviid corals with platy growth forms (Table 1). (1982), Robertson (1970), Taylor (1971), Taylor

175M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
and Reid (1984), Schuhmacher (1993), Zuschin assemblage. The percentage of dead individualsthat are from taxa found alive is greater than 96%and Piller (1997b,c) and from our own field obser-
vations (Table 3). for the complete data set and is 100% for thereduced data set (Table 5).Group 1 consists of bivalves that have a close
substrate relation to living corals: these taxa bore The dominant taxa in the live and dead assem-blages are strikingly different, however. Livingin living corals (Pedum), settle in crevices of living
corals (B. foliata and Streptopinna), are epizoic on molluscs are strongly dominated by bivalves asso-ciated with living corals, especially Pedum andliving corals and the hydrozoan Millepora
(Pteriidae), or live in close association with living Tridacna, the encrusting gastropod Dendropoma,and gastropods feeding on corals, mainly repre-corals (Tridacna). Group 2 consists of bivalves that
are encrusters on dead hard substrata (Chamoidea, sented by the parasitic Coralliophila. Dead mol-luscs, in contrast, are strongly dominated byOstreoidea and Spondylidae). Group 3 consists of
bivalves that are crevice dwellers of dead hard encrusting bivalves, especially Chamoidea andSpondylidae. Overall, in the reduced data set gas-substrata (Barbatia setigera, Ctenoides annulata,
Isognomon legumen, Lima lima). Group 4 consists tropods make up 36% of the living molluscs butonly 8% of dead molluscs; more than 73% of theof the encrusting gastropod Dendropoma, which
shows a rather indifferent relation to living or dead shells of dead gastropods were inhabited by hermitcrabs (Table 4) (the corresponding values for thehard substrata because it occurs on both types.
Group 5 consists of vagile gastropods that have a complete data set are 37%, 11% and 66%).In accordance with the strong differencesclose feeding relation to living corals (Coralliophila
and Drupella). Group 6 consists of vagile gastro- regarding the dominating taxa, no statisticallysignificant correlation between the total abundancepods that try to avoid living corals (Conus spp.
and Cerithium spp). of living and dead molluscs exists, the overallsimilarity being only 6% (Table 2). 17 samplelocations display a significant or highly significantcorrelation of live and dead assemblages. They5. Comparison of live and dead molluscsbelong to rock grounds (8, 10, 22, 31, 42, 46, 55,57), coral carpets with Porites, faviid and depau-5.1. Congruence of live and dead assemblages
(fidelity) perate faviid associations (4, 5, 6, 7, 21, 50, 51and 58) and Acropora patch associations (40).Only at 12 of these sample locations is the sim-At 68 sample locations a total area of
340.5 m2 was investigated (Fig. 2, Table 2) and ilarity between living and dead fauna above 50%(Table 2). However, at most of these sample loca-2846 individuals were counted. Most of these
individuals were alive (2211; 77.7%) and only a tions major similarities are evident between livingand dead Chamoidea. These results are supportedminor part was dead (635; 22.3%). Generally,
molluscan densities are characterized by high by a correspondence analysis, which shows strongdifferences in the composition of living and deadstandard deviations (Tables 2 and 4), indicating
uneven distribution within and between sites. molluscs at most sample locations (Fig. 4).Corresponding to the much higher number ofliving molluscs, the overall density of living mol- 5.2. Distribution patterns of living and dead
molluscsluscs (1.62 indiv./0.25 m2) is much higher than thatof the dead fauna (0.47 indiv./0.25 m2).
The percentages of live taxa found dead and of According to the results of a correspondenceanalysis, the distribution pattern of living molluscsdead taxa found live on hard substrata in the
study area are above 60% for the complete data shows five distinct groups (Fig. 5), whereas forthe dead fauna no distinct groups can be recog-set. They are up to 100% for the reduced data set,
in which only those taxa are considered that con- nized (Fig. 6). At least four of the groups of livingmolluscs can be related to specific hard substratatribute at least 1% to either the live or dead

176 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Table 4Abundance, relative proportion and density (per 0.25 m2) of the 18 most abundant taxa. Life habit groups as in Table 3
Taxon ( life habit group) Living Dead Hermit crab inhabited
No. % Mean St. dev. No. % Mean St. dev. No. %
Pedum spondyloideum (1) 548 25.4 0.40 1.42 5 0.8 0.00 0.06 – –Dendropoma maxima (4) 387 17.9 0.28 1.99 1 0.2 0.00 0.03 – –Coralliophila neritoidea (5) 338 15.7 0.25 1.57 2 0.3 0.00 0.04 1 50.0Tridacna maxima (1) 257 11.9 0.19 0.53 33 5.5 0.02 0.15 – –Chamoidea (2) 142 6.6 0.10 0.39 318 53.2 0.23 0.79 – –Barbatia setigera (3) 87 4.0 0.06 0.33 20 3.3 0.01 0.14 – –Ctenoides annulata (3) 74 3.4 0.05 0.30 7 1.2 0.01 0.08 – –Spondylidae (2) 50 2.3 0.04 0.22 89 14.9 0.07 0.27 – –Streptopinna saccata (1) 40 1.9 0.03 0.17 1 0.2 0.00 0.03 – –Lopha cristagalli (2) 36 1.7 0.03 0.26 43 7.2 0.03 0.37 – –Barbatia foliata (1) 35 1.6 0.03 0.20 1 0.2 0.00 0.03 – –Ostreoidea (2) 34 1.6 0.02 0.20 23 3.8 0.02 0.16 – –Cerithium spp. (6) 30 1.4 0.02 0.19 30 5.0 0.02 0.17 22 73.3Pteriidae (1) 27 1.3 0.02 0.23 0 0.0 0.00 0.00 – –Isognomon legumen (3) 25 1.2 0.02 0.15 2 0.3 0.00 0.04 – –Drupella cornus (5) 20 0.9 0.01 0.31 7 1.2 0.01 0.07 6 85.7Lima lima (3) 18 0.8 0.01 0.13 7 1.2 0.01 0.07 – –Conus spp. (6) 11 0.5 0.01 0.10 9 1.5 0.01 0.08 7 77.8
Table 5Fidelity of death assemblage to living molluscs with respect to taxonomic composition after indices provided in Kidwell and Bosence(1991) and in comparison with data from soft substrata molluscs and corals. Sample groups I–V after results of correspondenceanalysis (compare Fig. 5)
% Live taxa % Dead taxa % Dead individualsfound dead found live from taxa found alive
Complete Reduced Complete Reduced Complete Reduceddata set data set data set data set data set data set
MolluscsSafaga (this study)Group I (on reef flats) 66.7 83.3 54.5 83.3 61.1 90.9Group II (on Porites associations) 50.0 60.0 60.0 75.0 45.8 50.0Group III (on rock grounds) 53.3 60.0 53.3 75.0 96.3 98.9Group IV (mainly on faviid carpets) 58.8 82.4 62.5 100.0 94.2 100.0Group V (on Acropora–Millepora reef slopes) 30.0 30.0 60.0 60.0 77.8 77.8
Study area 68.4 94.4 61.9 100.0 96.7 100.0
Non-reef mean ( Kidwell and Bosence, 1991) 87 44 83
CoralsShallow Florida Keys mean (Greenstein and Pandolfi, 1997) 67 57 69Deep Florida Keys mean (Pandolfi and Greenstein, 1997a) 77 54 56Madang Lagoon (Pandolfi and Minchin, 1995) 54 90 94

177M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Fig. 4. Ordination of samples and taxa and relationships between taxa and sites using the first two dimensions of the correspondenceanalysis on living and dead molluscs. (A) Points close to one another represent samples that are more similar in taxonomic compositionthan points farther away from one another. Note the displacement of the life assemblages from the death assemblages, suggestingdifferences in taxonomic composition between them, which can be inferred from the ordination of taxa (B). Patterns in life assemblagezonation are poorly reproduced in the death assemblages (compare also Figs. 5 and 6).
types and will be described regarding their live and within this group. Moreover, the wide dispersionof group IV samples between samples of groupsdead fauna.
The density of living molluscs is higher than II and III indicates the transitional character ofthis group (Fig. 5). Correspondingly, most of thethat of dead molluscs in four of the five differenti-
ated sample groups. Only in sample group III is recognized hard substrata are characterized by adistinct association of living molluscs. However,the dead fauna more abundant than the living
fauna (Table 6). at least two important bottom types, the Acroporaassociation on reef slopes and the coral patchThe living fauna of four sample groups are
distinct, with a few taxa and life habits characteriz- association, are not reflected by a distinct faunaof living molluscs, but are also included withining the composition (Figs. 5 and 7); only sample
group IV has a broad variety of taxa, reflecting sample group IV.The death assemblages in all five groups ofthe more heterogeneous character of samples

178 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190

179M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
samples are dominated by the same taxa (mainly 1997a; Greenstein and Pandolfi, 1997). For thisreason it could well be that a future fossil mollus-Chamoidea, Spondylidae and Cerithium spp., in
different rankings) and the same life habits (mainly can assemblage is more similar to the original lifeassemblage than the present death assemblage isencrusters to dead hard substrata) (Fig. 7,
Table 6). Consequently, the ordination of dead to the present life assemblage (see Greenstein et al.,1998b for such an example among reef corals).molluscs does not reveal groupings that can be
related to specific hard substrata types (Fig. 6):here the results of the correspondence analysis 6.2. Taxonomic composition of live and dead
assemblagesexplain a few samples only, that are characterizedby exceptional faunal compositions (samples 4and 18). More than 68% of the live taxa are also found
dead on the hard substrata in our study area andA distinct distribution pattern of sample loca-tions with significant correlations of living and this percentage is greater than 94% if only those
taxa are considered that contribute at least 1% todead molluscs is recognized: seven sample loca-tions of sample group III are characterized by either the live or dead assemblage (reduced data
set). These values are similar to the mean valuesignificant correlation coefficients, but only one ofsample group II and none from sample groups I (87%) for non-reef molluscs of various sedimentary
environments ( Kidwell and Bosence, 1991). Theand V. From sample group IV, which contains thehighest number of sample locations (44), nine nearly 62% of dead taxa found alive in the study
area, which is distinctly above the mean valuesample locations show significant correlations ofliving and dead fauna. Of those, however, at least (44%) for non-reef molluscs of various sedimentary
environments ( Kidwell and Bosence, 1991),four mark a transition from sample group IV tosample group III, which is reflected in the distribu- increase to 100% when only the reduced data set
is considered.tion pattern of samples in Fig. 5.Including also data from molluscan death
assemblages in sediments from the same area(Zuschin, 1997) increases the proportion of live6. Discussiontaxa found dead to 79% for the complete data set,indicating that many individuals of hard substrata6.1. Sampling biastaxa are preserved in the adjoining sediment orwithin crevices of the coral framework. TheThe logistics of our sampling method provided
a strong bias against dead molluscs, that lived in remaining differences from non-reef molluscs ofvarious sedimentary environments can beclose association with living corals or coralline red
algae (bivalves of life habit group 1 in Table 3 and explained by preferential overgrowth of many mol-luscan taxa living in close association with a livingencrusting Dendropoma). Such molluscs are easily
overgrown after death and therefore we did not substrata. The comparatively high value of deadtaxa found alive can be related to the high percen-pick them with our sampling regime, but in fact
they have a very good chance of being incorporated tage of living individuals, which is in markedcontrast to results from studies dealing with mol-into the fossil record. Similar sampling biases are
also reported against massive corals in modern luscan assemblages in sediments ( Kidwell andBosence, 1991) and again points to preferentialdeath assemblages (Pandolfi and Greenstein,
Fig. 5. Ordination of samples and taxa and relationships between taxa and sites using the first three dimensions of the correspondenceanalysis on living molluscs. Points close to one another represent samples that are more similar in taxonomic composition than pointsfarther away from one another. Using dimensions 1 and 2 (A) shows sample groups (SG) I, II, III and IV established by (B)distribution pattern of living molluscs. Using dimensions 2 and 3 (C) shows sample groups (SG) II, III, IV and V established by(D) distribution pattern of living molluscs. #=Sample locations and taxa, $=sample locations with significant correlation coeffi-cients of living and dead fauna.

180 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Fig. 6. Ordination of samples and taxa and relationships between taxa and sites using the first three dimensions of the correspondenceanalysis on dead molluscs. Points close to one another represent samples that are more similar in taxonomic composition than pointsfarther away from one another. Using dimensions 1 and 2 (A) no sample groups were gained because (B) the distribution patternof dead molluscs is indistinct. Using dimensions 2 and 3 (C) again no sample groups were gained because (D) the distribution patternof dead molluscs is indistinct. Results of the correspondence analysis explain a few samples only, that are characterized by exceptionalfaunal compositions (samples 4 and 18). #=Sample locations and taxa, $=sample locations with significant correlation coefficientsof living and dead fauna.

181M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Table 6Molluscan composition of sample groups I–V, recognized by the correspondence analysis. Bold numbers are for dominant andcharacteristic taxa in sample groups

182 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Fig. 7. Assignment of molluscan life habits to sample groups shown by correspondence analysis.
overgrowth and transport into adjoining sediments 6.3. Abundances of taxa in live and deadassemblagesof the dead individuals.
The considerable increase of percentages whenconsidering only the reduced data set strongly In contrast to the good agreement between
living and dead molluscs regarding taxonomicsuggests that living taxa not found in the deathassemblage and dead taxa not found alive are composition, nearly all taxa that were abundant
as living individuals were rare as dead individualsquantitatively unimportant. This is supported bythe high percentage of dead individuals from taxa and vice versa: Pedum, Dendropoma, Coralliophila
and Tridacna are dominant in the living assem-found alive for the complete data set (Table 5).Comparisons with fidelity indices from reef blage, whereas Chamoidea, Spondylidae and ceri-
thiid gastropods are dominant in the deathcorals are difficult, because strong differences existbetween values from the Indo-Pacific and the assemblage. Generally, gastropods, which contri-
bute considerably to the total of living molluscs,Caribbean, which are related to diversity differ-ences, and between values from shallow and deep are very rare among dead molluscs on hard sub-
strata. This is in marked contrast to soft substrata,water studies, which are related to changes incommunity structure and/or differences in time where molluscan death assemblages consistently
show strong fidelity to relative abundances in theaveraging (Table 5) (Pandolfi and Greenstein,1997a). However, reef corals also have life histories live community ( Kidwell and Flessa, 1995;
Kidwell, in press).different from molluscs and are characterized byselective preservation of their taxa; these features Rapid post-mortem overgrowth of taxa that
live in close association with a living substratumare potentially responsible for observed differencesbetween fidelity indices of molluscs and reef corals (corals or coralline red algae) and post-mortem
transport of vagile fauna into surrounding sedi-in general (Pandolfi and Minchin, 1995).

183M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
ments are considered to be the best explanations in the studied sediments, whereas contributions ofbivalves living in crevices of living or dead coralsfor the observed biased record of living and dead
molluscs. and boring into living corals are minor (Zuschinand Hohenegger, 1998).Additionally, hermit crab occupancy of empty
shells may be of some importance, but dead shellsof Coralliophila, which is the quantitatively most 6.4. Interactions of substrata and molluscan life
habitsimportant live vagile gastropod, are only found insediments around coral-dominated hard substrata(Zuschin and Hohenegger, 1998). Cerithium spp., Reef flats (sample group I ) are strongly domi-
nated by the encrusting gastropod Dendropomathe quantitatively most important dead gastropod,is found in the habitat of the living snails, despite maxima which lives cemented to dead hard sub-
strata or embedded into the tissue of living corals.frequent hermit crab occupancy. Therefore wewould rather exclude a major habitat anomaly due Obviously, these gastropods do not have a special
substrate preference, but require the agitated waterto hermit crab transport of shells, as for exampledescribed by Shimoyama (1979). Nevertheless, conditions on reef flats in order to spread their
mucous net for feeding (Hughes and Lewis, 1974).preferential hermit crab occupation of some gas-tropod taxa (compare also Frey, 1987) keeps their Such very high densities of these sessile gastropods,
however, are a phenomenon largely restricted todead shells in the original habitat for longer thanothers, producing an abundance anomaly (Walker, the Red Sea (compare Mastaller, 1978; Taylor and
Reid, 1984), with similar abundances only being1989) and being responsible for similarities/correlations of living and dead Cerithium spp. on reported from the SE Pacific (e.g. Salvat, 1971).
Despite the low proportion of living coral cover-rock grounds. However, distinct pagurization fea-tures like massive encrustations, alterations of the age, the very low abundance of dead individuals
can be explained by rapid overgrowth after death,aperture or pagurid facets (reviewed in Walker,1992 and Taylor, 1994) as found on a rocky especially as coralline red algae are very abundant.
Even with living Dendropoma usually only theintertidal in the Northern Bay of Safaga (Zuschinand Piller, 1997a) were never observed on the aperture of the animal can be recognized, as the
rest of the shell is already completely covered byinvestigated subtidal hard substrata. Only subtleencrustations of coralline red algae were obligatory coralline red algae or corals. Therefore on this
bottom type the similarity and correlation of livingon hermit crab inhabited shells of these environ-ments. Therefore recognition of hermit crab occu- and dead fauna is poor.
Sample group II, which represents most of thepancy of these shells in the fossil record would berather difficult. Porites-dominated reef slopes and coral carpets
(except sample location 21), is strongly dominatedAlternative possibilities for the observed biasare predatory loss of those taxa that settle within by molluscs with a close substrate relation to living
corals. Coralliophila and B. foliata are very typicalor in close association with living scleractinians ortheir taphonomic loss (see Murray and Alve, 1999 faunal elements, because they are nearly restricted
to this facies (Fig. 5). B. foliata has always beenfor an example of selective taphonomic loss amongforaminifera). However, no extensive predation on observed to live in crevices of living Porites spp.,
usually with direct contact between the bivalvethese taxa has been observed and the loss ofexactly those taxa that live in close association and the coral tissue. This is in contrast to Taylor
(1971), who reported this species from dead partswith a living substratum seems to be very unlikely.Our taphonomic interpretation is also supported of coral branches or massive corals. Coralliophila
violacea is a parasitic gastropod, that feeds onby a comparison with a detailed study on mollus-can death assemblages in sediments in the Porites and Porites (Synerea) (Robertson, 1970;
Schuhmacher, 1993). The low proportion of deadNorthern Bay of Safaga: the upper valves ofbivalves encrusting dead hard substrata and the individuals in this sample group again strongly
suggests rapid overgrowth of bivalves, which isshells of gastropods feeding on corals are abundant

184 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
supported by the high surface coverage by living can life habits to be developed on these bottomtypes (Fig. 7). Due to its intermediate positionscleractinians. Coralliophila is interpreted to fall
off after death and correspondingly will be found between sample groups II and III, two majortrends in the molluscan composition within thisin the surrounding sediments or in crevices between
coral heads. A comparison with a quantitative group can be recognized. Sample locations near tosample group II in the correspondence analysisstudy on molluscan death assemblages in sediments
confirms this view, as coralliophilids are an impor- are mainly characterized by molluscs with a dis-tinct relation to living corals. Streptopinna in mosttant faunal element in coral associated sedimentary
facies (Zuschin and Hohenegger, 1998). cases is embedded in living massive coral colonies,but sometimes is also found in crevices of deadAdditionally, those individuals that mainly con-
tribute to the dead fauna belong to encrusters of coral heads. The host-specific Pedum is associatedwith a variety of scleractinians (preferentiallysmall-sized dead hard substrata within the overall
living surface. These individuals take longer to be Montipora spp.) in the northern Red Sea( Kleemann, 1990). It occurs in living corals (e.g.overgrown than those that live in direct contact
with living corals. Therefore on this bottom type Yonge, 1967; Waller, 1972), where it lives as aborer and attaches byssally ( Kleemann, 1990).only one sample location (4) shows significant
correlations of living and dead fauna. Tridacna maxima is a coral reef-associated bivalve(e.g. Yonge, 1974), which is usually found byssallySample group III, which summarizes rock
grounds, contains many of those sample locations attached within or between living coral colonies.Sometimes penetration into the coral substratumwhich have a very high proportion of dead surface
and correspondingly a low coverage by living was obvious (compare Yonge, 1980). In a fewcases large, unattached specimens were found lyingscleractinians. Five samples from Sarcophyton car-
pets belong here and three sample locations from on the surface of reef slopes and coral carpets. Allof the mentioned taxa can also be found as abun-platy scleractinian carpets. Due to their high pro-
portion of dead surface, these bottoms are domi- dant constituents of sample group II, from whichthis assemblage mainly differs by the absence ofnated by Chamoidea, which have been observed
to colonize mainly bare rocky surfaces (Zuschin B. foliata and Coralliophila. Generally, these fea-tures suggest relatively high amounts of livingand Piller, 1997b,c). The second typical faunal
element is Cerithium spp. (C. echinatum, C. nodulo- surface, which increase towards sample group II.The somewhat strange coral composition of samplesum, C. ruppelli) which are well-known algal–
detritus feeders (Houbrick, 1992). Typically, after location 14 is reflected in the molluscan fauna andcorrespondingly a rather isolated position withindeath all this fauna remains at the surface of the
bottom for very long, as it will not easily be sample group IV.Sample locations near to sample group III areovergrown by corals or coralline red algae. In
accordance with this scenario, sample group III is characterized by bivalves that live as crevice dwell-ers within dead hard substrata, where they arethe only sample group strongly dominated by dead
individuals and characterized by consistent signifi- byssally attached (Barbatia setigera, Ctenoidesannulata, Isognomon legumen, Lima lima) (comparecant correlations and high similarities of living and
dead fauna. Hadfield, 1976; Morton, 1983a; Taylor, 1971). Inmost cases these hard substrata are dead coralSample group IV is the only sample group that
shows a very heterogeneous composition of bottom colonies or dead parts of otherwise living coralcolonies. A second important feature aretypes, molluscan taxa and molluscan life habits.
This is due to its intermediate position between Spondylidae and Ostreoidea, which are usuallyfound as encrusters on dead coral heads. Lophasample groups II, III and V (Fig. 5). More than
half of the sample locations, however, belong to cristagalli often produces striking clusters on uppervalves of spondylids, which themselves arecoral carpets with faviid or depauperate faviid
associations. The very high coral diversity (Riegl cemented to dead coral heads (Zuschin and Piller,1997b). Additionally Conus spp., which lives as aand Piller, 1997) enables a broad variety of mollus-

185M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
predator on a variety of invertebrates (e.g. Taylorand Reid, 1984) and whose favourable portions ofreefs have <20% cover of living coral (Kohn,1983), shows a relation to this facies. This faunareflects the low amount of living surface, whichdecreases towards sample group III in the corre-spondence analysis. Corresponding to the ratherlow coverage by living corals, the proportion ofdead individuals is relatively high on these bottomtypes. The interpretation of the importance ofovergrowth after death is supported on dead coralheads by the large proportion of dead crevicedwellers, which do not live in direct contact witha living surface. In correspondence to these obser-vations, four of eight sample locations with sig-nificant correlations of live and dead fauna on thisbottom type mark a transition to sample groupIII (rock grounds), but none is transitional tosample group II (Porites associations).
Sample group V represents most sample loca-tions of the Acropora–Millepora-dominated reefslopes (except sample location 27) and is domi-nated by molluscs with a close relation to livingcoelenterates: the typical Pteriidae (Electroma ala-corvi and Pteria spp.) are epizoic bivalves and notmuch is known about their habitat preferences(Oliver, 1992); we observed them byssally attached
A
Bto live branching corals (mostly Acropora) and the
Fig. 8. Recent and fossil Dendropoma: (A) living Dendropomahydrozoan Millepora. Drupella is a predator onin the Northern Bay of Safaga, scale bar=10 cm; (B) fossil
corals (Robertson, 1970; Schuhmacher, 1993) and Dendropoma from Pleistocene outcrops along the Red Seawas only encountered on Acroporans. Both taxa coast, scale bar=10 cm.are interpreted to fall off after death and thereforewill be found in the surrounding sediments. A the study area in general, and a dominance of live
fauna on all bottom types with a high proportioncomparison with quantitative bulk samples fromreef slopes confirms this view (Zuschin, unpub- of living surface (corals or coralline red algae). In
contrast, the only bottom type with a dominancelished material ). Therefore, on this bottom typeno significant correlations or high similarities of of dead individuals are rock grounds. We can
prove the fossil occurrence of molluscan taxaliving and dead fauna exist.within corals at least for Dendropoma maxima,which is a typical constituent of Pleistocene coralreefs along the Red Sea coast (Fig. 8).7. Palaeontological implications
Our interpretation also fits previous results,which show that coral reefs have a high temporalOur data indicate that most of the fauna living
in close association with a living hard substratum resolution and are well suited to preserve short-term fluctuations of the fauna due to the rapid(coral colonies and coralline red algae) should be
found embedded in the framework of fossil coral rates of reef growth compared with the aggrada-tion of soft sediments (Jackson, 1983; Crame,reefs. Our interpretation is supported by the high
proportion of live individuals on hard substrata in 1980, 1981).

186 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Therefore, the fauna rapidly overgrown by the high number of inner-reef specialists amongmolluscs, which are strongly affected by sea-levelliving substrata should provide considerable eco-
logical and temporal information. In contrast, the fluctuations (Paulay, 1990, 1996; Taviani, 1998).Finally, the fossil occurrence of the studiedfauna that colonizes dead hard substrata remains
much longer on the surface, indicated by the strong fauna depends on the preservation potential of therespective substrata, which may vary considerablydominance of dead individuals on rock grounds,
which mainly provide a dead surface. Con- due to the prevailing disintegrative processes anddiagenetic environments (for a review see Scoffin,sequently this fauna should be taphonomically
altered and considerably time averaged. 1992). For example, corals in higher energy envi-ronments may be less degraded than those fromTaphonomic alteration is obvious, as most dead
encrusting bivalves are disarticulated; time averag- lower energy, and fragile corals may be betterpreserved than robust coral growth forms due toing was not quantified. However, if this fauna
becomes entombed at their life sites, it may at least different residence times in the taphonomicallyactive zone (Pandolfi and Greenstein, 1997b).allow the study of faunal succession (Brett, 1988).
Fauna preferentially transported into surrounding The diagenetic history of uplifted Pleistoceneterraces along the northern Red Sea coast reflectssoft substrata will be affected by extensive time
averaging and taphonomic disintegration typically complex inter-relationships of climatic changes,sea-level variations of different orders, and tecton-occurring in soft sediments and the associated loss
of much ecological information (Flessa et al., ics and their diagenetic features are characterizedby a highly variable spatial distribution (Strasser1993; Flessa and Kowalewski, 1994; Fursich, 1978;
Fursich and Aberhan, 1990; Greenstein, 1989; et al., 1992; Strasser and Strohmenger, 1997). ThePleistocene coral reef faunas tend to be diageneti-Greenstein et al., 1995; Kidwell, 1998; Kidwell and
Bosence, 1991; Kowalewski, 1996; Kowalewski cally reduced in numbers compared with modernsediments from the same area. The reduction inet al., 1998; Meldahl et al., 1997; Olszewski, 1999;
Powell et al., 1989). diversity is due to widespread vadose diagenesis inthe Pleistocene sediments and is especially true ofHowever, the fossil occurrence of the studied
fauna will ultimately depend on the long-term aragonitic molluscs (Dullo, 1983, 1984, 1990). Thefaunal reduction increases with age of thestability of the studied assemblages as well as on
the preservation potential of the respective sub- Pleistocene terraces (Strasser et al., 1992; Strasserand Strohmenger, 1997) and only thick-shelledstrata. For reef corals, community structure is
stable in the long term (103–105 years) (Jackson, Tridacna and Strombus shells are well preserved,whereas smaller shells are not recorded (Dullo,1992; Jackson et al., 1996; Pandolfi, 1996, 1999).1984). The diagenetic environment seems toStudies in the Caribbean show that Pleistocenegovern shell preservation as the latter is very goodreefs exhibit excellent coral preservation and sig-(even primary coloration of molluscs is preserved)nificant similarities regarding taxonomy, zonationin uplifted terraces with slightly different climaticand diversity to living shallow water coral associa-factors and geological settings along the Red Seations (Mesolella, 1967; Greenstein and Curran,coast (Dullo, 1984).1997; Greenstein and Pandolfi, 1997; Pandolfi and
However, at least for the total species list, littleGreenstein, 1997a; Pandolfi and Jackson, 1997;differences exist between Pleistocene and modernGreenstein et al., 1998a). These Pleistocene reefsmolluscan assemblages from coral reefs of thecan even be used to calibrate an ecological baselineKenya coast (Crame, 1986; compare alsowith which to compare modern reef assemblagesValentine, 1989).(Greenstein et al., 1998a). In contrast, comparisons
of Pleistocene and modern molluscan assemblagesshow that significant faunal differences exist, which
8. Conclusionscan be related to significant habitat loss due toQuaternary sea-level fluctuations (Taylor, 1978;Crame, 1986; Paulay, 1996). These differences 1. Most taxa found dead in the study area were
also found live and vice versa; differences frombetween corals and molluscs might be related to

187M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
results on molluscan assemblages in sediments sediments from the same study area, observa-tions in Pleistocene deposits along the Red Seacan be related to the special accumulation con-
ditions of dead faunas on hard substrata. Living coast, and previous results, which show thatrapid reef growth is potentially suited to pre-taxa not found in the death assemblage and
dead taxa not found alive are proven to be serve ecological information.6. The approach of our investigation was to com-quantitatively unimportant.
2. Strong differences regarding dominating taxa bine the life habits of organisms with their post-mortem history. The fossil occurrence of theof living and dead molluscs are the result of
distinct biases in the death assemblage. Dead studied faunas, however, finally depends on thelong-term stability of the studied assemblagesmolluscs, that lived in close association with
living corals or coralline red algae, are strongly and on the preservation potential of the respec-tive substrata, which may vary considerablyunder-represented in the death assemblage.
Such molluscs are easily overgrown after death due to the prevailing disintegrative processesand diagenetic environments.and therefore were not recognized by our sam-
pling regime. These molluscs, however, have avery good chance of being incorporated intothe fossil record. Dead gastropods, that feed on
Acknowledgementscorals, are also strongly under-represented inthe studied death assemblage, because of rapid
Thanks are due to Abbas M. Mansour, Wernerpost-mortem transport into surrounding sedi-E. Piller, Michael Rasser and Bernhard Riegl forments or into crevices within corals.help with field work. Ronald Janssen, Eike3. Major similarities and significant correlationsNeubert and P. Graham Oliver helped with taxo-of living and dead faunas are mainly observednomic problems. Karl Kleemann, Werner E. Pilleron rock grounds, where the amount of scle-and Bernhard Riegl provided stimulating discus-ractinians can be neglected and encrustingsions on coral–mollusc interactions and the classi-Chamoidea dominate live and death assem-fication of hard substrata.blages. Similarities between living and dead
The authors are especially grateful to RobertCerithium spp. are influenced by hermit crabs,J. Stanton Jr., Susan M. Kidwell and reviewerswhich preferentially keep these shells in theirFranz T. Fursich and John M. Pandolfi, whosehabitat after death.comments improved the manuscript greatly.4. From a palaeoecological point of view the hard
Financial support was provided by Projectsubstrata fauna studied can be differentiatedP10715-Geo to FFS of the Austrian ‘Fonds zurinto three groups: fauna preferentially over-Forderung der wissenschaftlichen Forschung’grown by a living substratum should provide(FWF). The manuscript was finished during aconsiderable temporal and ecological informa-Max Kade fellowship to MZ at Texas A&Mtion as it will be preserved within a rapidlyUniversity.growing reef framework. Fauna encrusting dead
hard substrata remains much longer at thesurface and if it becomes entombed at their lifesites, it may finally allow the detailed study of Referencesfaunal succession. Fauna preferentially trans-ported into surrounding soft substrata will be Benzecri, J.P., 1992. Correspondence Analysis Handbook.
Marcel Dekker, New York. II+665 pp.affected by time averaging and taphonomicBrett, C.E., 1988. Paleoecology and evolution of marine harddisintegration typically occurring in sediments
substrate communities: an overview. Palaios 3, 374–378.and the associated loss of much ecologicalBromley, R.G., 1975. Trace fossils at omission surfaces. In:
information. Frey, R.W. (Ed.), The Study of Trace Fossils. Springer, New5. Our interpretation is supported by a compari- York, pp. 399–428.
Cadee, G.C., 1968. Molluscan biocoenoses and thanato-son with molluscan death assemblages in soft

188 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
coenoses in the Ria de Arosa, Galicia, Spain. Zool. Verh. with application to principal component analysis. Biomet-95, 1–121. rika 58, 435–467.
Carthew, R., Bosence, D., 1986. Community preservation in Gauch, H.G., 1982. Multivariate Analysis in Community Ecol-recent shell-gravels, English Channel. Palaeontology 29 ogy. Cambridge University Press, Cambridge. 298 pp.(2), 243–268. Goldring, R., 1995. Organisms and the substrate: response and
Crame, J.A., 1980. Succession and diversity in the Pleistocene effect. In: Bosence, D.W.J., Allison, P.A. (Eds.), Marinecoral reefs of the Kenya coast. Palaeontology 23 (1), 1–37. Palaeoenvironmental Analysis from Fossils, Geol. Soc.
Crame, J.A., 1981. Ecological stratification in the Pleistocene Spec. Publ. 83, 151–180.coral reefs of the Kenya coast. Palaeontology 24 (3), Goreau, T.F., Goreau, N.I., Yonge, C.M., 1973. On the utiliza-609–646. tion of photosynthetic products from zooxanthellae and of
Crame, J.A., 1986. Late Pleistocene molluscan assemblages a dissolved amino acid in Tridacna maxima f. elongata (Mol-from the coral reefs of the Kenya coast. Coral Reefs 4, lusca: Bivalvia). J. Zool. 169, 417–454.183–196. Greenstein, B.J., 1989. Mass mortality of the West-Indian echi-
Dodd, J.R., Stanton Jr., R.J., 1990. Palaeoecology: Concepts noid Diadema antillarum (Echinodermata: Echinoidea): aand Applications. Wiley, New York. xvii+502 pp. natural experiment in taphonomy. Palaios 4, 487–492.
Dullo, W.C., 1983. Diagenesis of fossils of the Miocene Leitha Greenstein, B.J., Curran, H.A., 1997. How much ecologicalLimestone of the Paratethys, Austria: an example for faunal information is preserved in fossil reefs and how reliable ismodifications due to the changing diagenetic environment. it? In: Proc. 8th Int. Coral Reef Symp., Panama City,Facies 8, 1–112. Panama Vol. 1., 417–422.
Dullo, W.C., 1984. Progressive diagenetic sequence of aragonite Greenstein, B.J., Pandolfi, J.M., 1997. Preservation of com-structures: Pleistocene coral reefs and their modern counter- munity structure in modern reef coral life and death assem-parts on the eastern Red Sea coast, Saudi Arabia. Palaeo- blages of the Florida Keys: implications for the Quaternaryntogr. Am. 54, 254–260. fossil record of coral reefs. Bull. Marine Sci. 61, 431–452.
Dullo, W.C., 1990. Facies, fossil record and age of Pleistocene Greenstein, B.J., Curran, H.A., Pandolfi, J.M., 1998a. Shiftingreefs from the Red Sea. Facies 22, 1–46. ecological baselines and the demise of Acropora cervicornis
Fankboner, P.V., Reid, R.G.B., 1990. Nutrition in giant clams in the western North Atlantic and Caribbean Province: a(Tridacnidae). In: Morton, B. (Ed.), The Bivalvia — Proc.
Pleistocene perspective. Coral Reefs 17, 249–261.Memorial Symp. in Honour of Sir Charles Maurice Yonge,
Greenstein, B.J., Pandolfi, J.M., Curran, H.A., 1998b. The com-Edinburgh, 1986. Hong Kong University Press, Hong Kong,
pleteness of the Pleistocene fossil record: implications forpp. 195–209.
stratigraphic adequacy. In: Donovan, S.K. (Ed.), The Ade-Fleiss, J.L., 1973. Statistical Methods for Rates and Propor-
quacy of the Fossil Record. Wiley, London, pp. 75–109.tions. Wiley, New York. 223 pp.Greenstein, B.J., Pandolfi, J.M., Moran, P.J., 1995. TaphonomyFlessa, K.W., Fursich, F.T., 1991. Quantitative analyses of mol-
of crown-of-thorns starfish: implications for the recognitionluscan communities and taphocoenoses of Bahia la Choyaof ancient population explosions. Coral Reefs 14, 91–97.(Gulf of California, Sonora, Mexico). In: Fursich, F.T.,
Hadfield, M.G., 1976. Molluscs associated with living corals.Flessa, K.W. (Eds.), Ecology, taphonomy and paleoecologyMicronesica 12, 133–148.of Recent and Pleistocene molluscan faunas of Bahia la
Hill, M.O., 1973. Reciprocal averaging: an eigenvector methodChoya, northern Gulf of California, Zitteliana 18, 79–88.of ordination. J. Ecol. 61, 237–249.Flessa, K.W., Kowalewski, M., 1994. Shell survival and time-
Hohenegger, J., 1974. Uber einfache Gruppierungsmethodenaveraging in nearshore and shelf environments: estimatesvon Fossil-Vergesellschaftungen am Beispiel obertriadischerfrom the radiocarbon literature. Lethaia 27, 153–165.Foraminiferen. Neues Jh. Geol. Palaontol. Abh. 146 (3),Flessa, K.W., Cutler, A.H., Meldahl, K.H., 1993. Time and263–297.taphonomy: quantitative estimates of time-averaging and
Hohenegger, J., 1982. Numerische Klassifikation von Indivi-stratigraphic disorder in a shallow marine habitat. Paleo-duen und Merkmalsnormierung. Sitz. Osterreich. Akad.biology 19, 266–286.Wissenschaften, Mathematisch-naturwissensch. Klasse,Frey, R.W., 1987. Hermit crabs: neglected factors in taphonomyAbt. I 191 1–4, 15–72.and paleoecology. Palaios 2, 313–322.
Houbrick, R.S., 1992. Monograph of the Genus Cerithium Bru-Fursich, F.T., 1978. The influence of faunal condensation andguiere in the Indo-Pacific (Cerithiidae: Prosobranchia).mixing on the preservation of fossil benthic communities.Smithsonian Contrib. Zool. 510, 211 pp.Lethaia 11, 243–250.
Hughes, R.N., Lewis, A.H., 1974. On the spatial distribution,Fursich, F.T., Aberhan, M., 1990. Significance of time-averag-feeding and reproduction of the vermetid gastropod Dendro-ing for palaeocommunity analysis. Lethaia 23, 143–152.poma maxima. J. Zool., London 172, 531–547.Fursich, F.T., Flessa, K.W., 1987. Taphonomy of tidal flat mol-
Jackson, J.B.C., 1983. Biological determinants of present andluscs in the Northern Gulf of California: paleoenviron-past sessile animal distributions. In: Tevesz, M.J.S., McCall,mental analysis despite the perils of preservation. PalaiosP.L. (Eds.), Biotic Interactions in Recent and Fossil Benthic2, 543–559.
Gabriel, K.R., 1971. The biplot — graphic display of matrices Communities. Plenum Press, New York, pp. 39–120.

189M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Jackson, J.B.C., 1992. Pleistocene perspectives on coral reef Saleuddin, A.S.M., Wilbur, K.M. (Eds.), Physiology, Part2. The Mollusca Vol. 5. Academic Press, New York,community structure. Am. Zool. 32, 719–731.
Jackson, J.B.C., Budd, A.F., Pandolfi, J.M., 1996. The shifting pp. 65–147.Murray, J.W., Alve, E., 1999. Natural dissolution of modernbalance of natural communities? In: Jablonski, D., Erwin,
D.H., Lipps, J.H. (Eds.), Evolutionary Paleobiology. The shallow water benthic foraminifera: taphonomic effects onthe palaeoecological record. Palaeogeogr. Palaeoclimatol.University of Chicago Press, Chicago, pp. 89–122.
Kidwell, S.M., 1998. Time-averaging in the marine fossil record: Palaeoecol. 146, 195–209.Nebelsick, J.H., Schmid, B., Stachowitsch, M., 1997. Theoverview of strategies and uncertainties. Geobios 30 (7),
977–995. encrustation of fossil and recent sea-urchin tests: ecologicaland taphonomic significance. Lethaia 30, 271–284.Kidwell, S.M., in press. Ecological fidelity of molluscan death
assemblages. In: Aller, R.C. (Ed.), Organism–Sediment Oliver, P.G., 1992. Bivalved Seashells of the Red Sea. Hemmen,Wiesbaden. 330 pp.Interactions: Papers in Honour of Donald C. Rhoads (Belle
W. Baruch Symp. Vol.). University of South Carolina Press. Oliver, P.G., 1995. Bivalvia. In: Dance, S.P. (Ed.), Seashells ofEastern Arabia. Motivate Publishing, pp. 194–281.Kidwell, S.M., Bosence, D.W.J., 1991. Taphonomy and time-
averaging of marine shelly faunas. In: Allison, P.A., Briggs, Olszewski, T., 1999. Taking advantage of time-averaging. Paleo-biology 25, 226–238.D.E.G. (Eds.), Taphonomy: Releasing the Data Locked in
the Fossil Record. Plenum Press, New York, pp. 115–209. Orloci, L., 1966. Geometric models in ecology. I. The theoryand application of some ordination methods. J. Ecol. 54,Kidwell, S.M., Flessa, K.W., 1995. The quality of the fossil
record: populations, species, and communities. Annu. Rev. 193–215.Pandolfi, J.M., 1996. Limited membership in Pleistocene coralEcol. Syst. 26, 269–299.
Kleemann, K., 1990. Coral associations, biocorrosion and space reef assemblages from the Huon Peninsula, Papua NewGuinea: constancy during global change. Paleobiology 22,competition in Pedum spondyloideum (Gmelin) (Pectinacea,
Bivalvia). PSZNI Marine Ecol. 11, 77–94. 152–176.Pandolfi, J.M., 1999. Response of Pleistocene coral reefs toKleemann, K., 1992. Coral communities and coral–bivalve
associations in the northern Red Sea at Safaga, Egypt. environmental change over long temporal scales. Am. Zool.39, 113–130.Facies 26, 125–134.
Kohn, A.J., 1983. Microhabitat factors affecting abundance and Pandolfi, J.M., Greenstein, B.J., 1997a. Preservation ofcommunity structure in death assemblages of deep waterdiversity of Conus on coral reefs. Oecologia 60, 293–301.
Kohn, A.J., Leviten, P.A., 1976. Effect of habitat complexity Caribbean reef corals. Limnol. Oceanogr. 42, 1505–1516.Pandolfi, J.M., Greenstein, B.J., 1997b. Taphonomic alterationon population density and species richness in tropical inter-
tidal predatory gastropod assemblages. Oecologia 25, of reef corals: effects of reef environment and coral growthform. I. The Great Barrier Reef. Palaios 12, 27–42.199–210.
Kowalewski, M., 1996. Time-averaging, overcompleteness and Pandolfi, J.M., Jackson, J.B.C., 1997. The maintenance ofdiversity on coral reefs: examples from the fossil record. In:the geological record. J. Geol. 104, 317–326.
Kowalewski, M., Goodfriend, G.A., Flessa, K.W., 1998. High- Proc. 8th Int. Coral Reef Symp., Panama City, PanamaVol. 1., 397–404.resolution estimates of temporal mixing within shell beds:
the evils and virtues of time-averaging. Paleobiology 24, Pandolfi, J.M., Minchin, P.R., 1995. A comparison of taxo-nomic composition and diversity between reef coral life and287–304.
Leviten, P.J., Kohn, A.J., 1980. Microhabitat resource use, death assemblages in Madang Lagoon, Papua, New Guinea.Palaeogeogr. Palaeoclimatol. Palaeoecol. 119, 321–341.activity patterns, and episodic catastrophe: Conus on tropi-
cal intertidal reef rock benches. Ecol. Monogr. 50, 55–75. Paulay, G., 1990. Effects of late Cenozoic sea-level fluctuationson the bivalve faunas of tropical oceanic islands. Paleobiol-Liddell, W.D., Ohlhorst, S.L., 1988. Hard substrata community
patterns, 1–120 m, North Jamaica. Palaios 3, 413–423. ogy 16, 415–434.Paulay, G., 1996. Dynamic clams: changes in the bivalve faunaLinder, A., Berchtold, W., 1976. Statistische Auswertung von
Prozentzahlen. Birkhauser, Basel. 232 pp. of Pacific islands as a result of sea-level fluctuations. Am.Malacol. Bull. 12, 45–57.Mastaller, M., 1978. The marine molluscan assemblages of Port
Sudan, Red Sea. Zool. Mededel. 53, 117–144. Piller, W.E., Pervesler, P., 1989. The Northern Bay of Safaga(Red Sea, Egypt): an actuopalaeontological approach. I.Meldahl, K.H., Flessa, K.W., Cutler, A.H., 1997. Time-averag-
ing and postmortem skeletal survival in benthic fossil assem- Topography and bottom facies. Beitr. Palaontol. Osterreich15, 103–147.blages: quantitative comparisons among Holocene
environments. Paleobiology 23, 207–229. Powell, E.N., Staff, G.M., Davies, D.J., Callender, W.R., 1989.Macrobenthic death assemblages in modern marine environ-Mesolella, K.J., 1967. Zonation of uplifted Pleistocene coral
reefs on Barbados, West Indies. Science 156, 638–640. ments: formation, interpretation, and application. Rev.Aquat. Sci. 1 (4), 555–589.Morton, B., 1983a. Coral-associated bivalves of the Indo-
Pacific. In: Russel-Hunter, W.D. (Ed.), Ecology. The Mol- Reichelt, R.E., 1982. Space: a non-limiting resource in theniches of some abundant coral reef gastropods. Coral Reefslusca Vol. 6. Academic Press, New York, pp. 139–224.
Morton, B., 1983b. Feeding and digestion in Bivalvia. In: 1, 3–11.

190 M. Zuschin et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 159 (2000) 167–190
Reiss, Z., Hottinger, L., 1984. The Gulf of Aqaba. Ecological Taylor, P.D., 1994. Evolutionary palaeoecology of symbiosisbetween bryozoans and hermit crabs. Hist. Biol. 9, 157–205.Micropaleontology. Springer, Berlin. VIII+354 pp.
Taylor, J.D., Reid, D.G., 1984. The abundance and trophicRiegl, B., Piller, W.E., 1997. Distribution of coral associationsclassification of molluscs upon coral reefs in the Sudanesein Northern Safaga Bay (Red Sea, Egypt). Facies 36,Red Sea. J. Nat. Hist. 18, 175–209.141–162.
Valentine, J.W., 1989. How good was the fossil record? CluesRiegl, B., Piller, W.E., 1999. Coral frameworks revisited — reefsfrom the Californian Pleistocene. Paleobiology 15 (2),and coral carpets in the northern Red Sea. Coral Reefs83–94.18, 241–253.
Walker, S.E., 1989. Hermit crabs as taphonomic agents. PalaiosRiegl, B., Velimirov, B., 1994. The structure of coral communi-4, 439–452.ties at Hurghada in the Northern Red Sea. PSZNI Marine
Walker, S.E., 1992. Criteria for recognizing marine hermit crabsEcol. 15, 213–231.in the fossil record using gastropod shells. J. Paleontol.Robertson, R., 1970. Review of the predators and parasites of66, 535–558.stony corals, with special reference to symbiotic proso-
Waller, T.R., 1972. The Pectinidae (Mollusca: Bivalvia) of Eni-branch gastropods. Pacific Sci. 24, 43–54.wetok Atoll, Marshall Islands. Veliger 14, 221–264.Salvat, B., 1971. Mollusques lagunaires et recifaux de l’ile de
Warme, J.E., 1969. Live and dead molluscs in a coastal lagoon.Raevavae (Australes, Polynesie). Malacol. Rev. 4, 1–15.J. Paleontol. 43, 141–150.Scoffin, T.P., 1992. Taphonomy of coral reefs: a review. Coral
Warme, J.E., 1971. Paleoecological aspects of a modern coastalReefs 11, 57–77.lagoon. Univ. Calif. Publ. Geol. Sci. 87, 131 pp.Schuhmacher, H., 1993. Impact of some corallivorous snails on
Wilson, M.A., 1987. Ecological dynamics on pebbles, cobblesstony corals in the Red Sea. In: Proc. Seventh Int. Coraland boulders. Palaios 2, 594–599.Reef Symp., Guam, 1992., 840–846.
Yonge, C.M., 1967. Observations on Pedum spondyloideumShimoyama, S., 1979. Modification of shell distribution pat-(Chemnitz) Gmelin, a scallop associated with reef-buildingterns by hermit crabs in a protected shore. Marine Sci. 11,corals. Proc. Malacol. Soc. London 37, 311–323.527–535.
Yonge, C.M., 1974. Coral reefs and molluscs. Trans. Roy. Soc.Staff, G.M., Stanton Jr., R.J., Powell, E.N., Cummins, H., 1986.
Edinb. 69, 147–166.Time-averaging, taphomomy, and their impact on paleo-
Yonge, C.M., 1980. Functional morphology and evolution incommunity reconstruction: death assemblages in Texas
the Tridacnidae (Mollusca: Bivalvia: Cardiacea). Rec.bays. Geol. Soc. Am. Bull. 97, 428–443. Austral. Mus. 33, 735–777.
Strasser, A., Strohmenger, C., 1997. Early diagenesis in Pleisto- Zuschin, M., 1997. Actuopalaeontological investigations oncene coral reefs, southern Sinai, Egypt: response to tectonics, shallow water mollusc associations in the Northern Bay ofsea-level and climate. Sedimentology 44, 537–558. Safaga (Red Sea, Egypt). Unpublished Ph.D. Thesis, Uni-
Strasser, A., Strohmenger, C., Davaud, E., Bach, A., 1992. versity of Vienna, 151 pp.Sequential evolution and diagenesis of Pleistocene coral Zuschin, M., Hohenegger, J., 1998. Subtropical coral reef asso-reefs (South Sinai, Egypt). Sediment. Geol. 78, 59–79. ciated sedimentary facies characterized by molluscs (North-
Surlyk, F., Christensen, W.K., 1974. Epifaunal zonation on an ern Bay of Safaga, Red Sea, Egypt). Facies 38, 229–254.Upper Cretaceous rocky coast. Geology 2, 529–534. Zuschin, M., Piller, W.E., 1997a. Gastropod shells recycled —
Taviani, M., 1998. Stable tropics not so stable: climatically an example from a rocky tidal flat in the northern Red Sea.driven extinctions of reef-associated molluscan assemblages Lethaia 30, 127–134.(Red Sea and western Indian Ocean, last interglaciation to Zuschin, M., Piller, W.E., 1997b. Bivalve distribution on coralpresent). Spec. Publ. Int. Assoc. Sediment. 25, 69–76. carpets in the northern Bay of Safaga (Red Sea, Egypt) and
Taylor, J.D., 1971. Intertidal zonation of Aldabra Atoll. Philos. its relation to environmental parameters. Facies 37,Trans. Roy. Soc. London, Ser. B 260, 173–213. 183–194.
Taylor, J.D., 1978. Faunal response to the instability of reef Zuschin, M., Piller, W.E., 1997c. Molluscan hard-substratehabitats: Pleistocene molluscan assemblages of Aldabra associations in the Northern Red Sea. PSZNI Marine Ecol.
18 (4), 361–378.Atoll. Palaeontology 21 (1), 1–30.




















