
Serotherapy of B-Cell Neoplasms With Anti-B4Blocked Ricin: A Phase I Trial of Daily Bolus Infusion
Upload
universitalaquilaCategory
view
4download
0
A chimeric saporin-transferrin conjugate compared to
ricin toxin: role of the carrier in intracellular
transport and toxicityRODOLFO IPPOLITI,* EUGENIO LENDARO,* IRENE D’AGOSTINO,* MARIA LUISA FIANI,tDANTE GUIDARINI,t STEFANO VESTRI, PIER ALBERTO BENEDETTI, ANDMAURIZIO BRUNORI5”*Istituto Pasteur-Fondazione Cenci Bolognetti, Dipartimento di Scienze Biochimiche and C.N.R. Centre of
Molecular Biology, Universit#{224}di Roma “La Sapienza,” Roma, Italy; tI5tjtt0 Superiore di Sanit#{224},Roma, Italy; and1istituto di Biofisica del C.N.R., Pisa, Italy
RESEARCH COMMUNICATION
1220 0892.6638/95/0009.1 220/$01 .50. © FASEB
ABSTRACT Human transferrin (TI’) and saporin-6(Sap), a ribosome inactivating protein from Sapon-aria officinalis, were chemically conjugated: the re-action generated two chimeras (called Tf-Sap) thatproved to be cytotoxic to HepG2 cells. Electro-phoretic and chromatographic analysis revealed thatthe two conjugates contained saporin and Tf in a 2:1or 1:1 molar ratio (140 and 110 KDa, respectively).Free saporin is essentially nontoxic, whereas Tf-Sapefficiently kills HepG2 cells, although its 1D50 ( 6nM) is 1000-fold greater than that of ricin. Intracel-lular transport of these toxins was followed by in vivofluorescence video microscopy, preparing the conju-gates starting from rhodamine isothiocyanate -labeledsaporin. Image analysis of living HepG2 cells exposedto fluorescent Tf-Sap revealed that the endocytoticpathway involving passage through secondary en-dosomes is dictated by Tf and is different from thatof ricin (the dimeric toxin from Ricinus communis),which is delivered to the Golgi apparatus, the prob-able site of activation. We discuss whether differencesin toxicity between ricin and Tf-Sap can be attributedto the different mechanisms of transport and activa-tion.-Ippoliti, R., Lendaro, E., D’Agostino, I., Fiani,M. L., Guidarini, D., Vestri, S., P. A. Benedetti,Brunori, M. A chimeric saporin-transferrin conjugatecompared to ricin toxin: role of the carrier in intra-cellular transport and toxicity. FASEB J. 9,1220-1225 (1995)
Key Words: endocytosis - loxic conjugates
TRANSFERRIN (Tf,2 THE PROTEIN INVOLVED in iron uptake bycells, transports two metal ions via endocytosis after bind-ing to a specific membrane receptor (1). The intracellularcycle of Tf-receptor complexes, a well-characterized proc-
ess, involves transit through the acidic endosomal systemwhere iron ions are easily dissociated from the carrier pro-tein and released inside the cell (2, 3). The Tf receptor,widely distributed in different cell types, is usually overex-pressed in malignant cells (4, 5); this finding has inspired
the synthesis of artificial conjugates containing a toxinbound to Tf, an attempt to construct a “magic bullet” to kill
tumor cells with high efficiency (6).On the basis of their structure, ribosome inactivating pro-
teins (RIPs) from plants have been classified (7) as type I
monomeric proteins and type II dimeric molecules, the lat-ter being composed of an active A chain and a carrier Bchain. Once internalized, these toxins are translocated tothe cytoplasm where they inactivate ribosomal functionsand stop protein synthesis (8, 9). Their toxicity thereforedepends on finding their intracellular target after internali-zation following binding to the plasma membrane via anatural or an artificial carrier. The carrier may be either alectin (i.e., the B chain of the dimeric toxin ricin from
Ricinus communis) or another molecule (such as a hormoneor an immunoglobulin; see refs 10-12).
Previous studies have shown that complexes between hu-
man Tf and saporin 6 (SO-6 from Saponaria officinali.s; 13)or ricin A chain (14) have an inhibitory effect on the prolif-eration of erythroleukemic cells. However, these studies
provided little information on the intracellular routing orthe activation mechanism of these conjugates. We synthe-sized, isolated, and characterized two complexes between
saporin-6 (29 kDa) and human Tf (Tf-Sap) in 1:1 or 2:1
molar ratio, labeled these complexes with a fluorescentprobe, and followed with video microscopy their intracellu-
lar distribution dynamics in living human hepatoma cells(HepG2), a line expressing the Tf receptor (15). In thispaper we compare the toxic properties of Tf-Sap with those
of native ricin and ricin labeled with fluorescent com-pounds. It is known (16) that ricin toxicity depends on the
tTo whom correspondence and reprint requests should be addressed, at:
Dipartimento di Science Biochimiche “A. Rossi Fanelli,” Universit#{224} di
Roma “La Sapienza,” PIe. Aldo Mom 5, 00185 Roma, Italy.
2Abbreviations: Tf, transferrin; Tf-Sap, the conjugate between transfer-rin and saporin; B ricin-Sap, the conjugate between ricin B chain and
saporin; BFA, brefeldin A; C5-NBD-ceramide, 6-((N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)caproyl) sphingosine; RIPs, ribosome inactivatingproteins; D-MEM, Eagle modified Dulbecco medium; SPDP, N-succin-
imidyl 3-(2-pyridyldithio)proprionate; HBSA, Hank’s balanced salt solu-
tion containing 1% bovine serum albumin.

RESEARCH COMMUNICATION
CELLULAR PROPERTIES OF CHIMERIC SAPORIN 1221
reduction of the disulfide bridge between the A and B chainboth in ribosomes from cellular extracts and in intact cells,where probably only the active A chain is released into thecytoplasm. This process is likely to occur in the Golgi ap-paratus (17), where the toxin is rapidly delivered after itsbinding to the plasma membrane (18). Comparison of tox-icity and intracellular distribution between ricin and theTf-Sap complexes leads to interesting results, which arepresented and discussed.
EXPERIMENTAL PROCEDURES
Synthesis and purification of saporm TRITC
Isoform VI of saporin was purified from seeds of S. officinali.s as de-
scribed (19). The protein was modified with tetramethyl rhodamine B
isothiocyanate (Sigma, St. Louis, Mo.) in a phosphate buffer (10 mM pH
7.0 containing 0.15 M NaCI); the reaction mixture was purified by ion
exchange chromatography on a CM-Sepharose fast-flow (Pharmacia,Biotech, Uppsala, Sweden) column equilibrated with 10 mM phosphate
buffer, pH 6.0, and eluted with linear NaCI gradient from 0 to 0.3 M. The
purity of the labeled protein was tested by fast protein liquid chromatog-
raphy (Pharmacia).
Activation of human Tf with SPDP
Human Tf was activated by adding stoichiometric amounts of N-succin-imidyl 3-(2-pyridyldithio)propionate (SPDP) (20) dissolved in ethanol to
a protein solution (10 mg/mI) in 100 mM phosphate buffer, pH 7.0. The
reaction was carried out at mom temperature for 1 h. Excess of SPDP was
removed by gel filtration on a G25 column and the stoichiometry of thereaction was calculated spectrophotometrically by reduction of the 2-
pyridyl-disulfide group, which produces absorption at 343 nm (pyridine-2-thione at 343 nm = 8.08 i#{248}M1 - cm). The modified Tf contained
1 mol SPDP/mol.
Synthesis of the conjugates
Saporin or TRITC saporin were activated with 2-iminothiolane-HCI as
described (21). Excess SPDP was removed by chromatography on a
Sephadex G25 column and the activated toxin was immediately mixedwith SPDP-derivatized Tf (fivefold excess); after 72 h incubation at 4#{176}C,the reaction mixture was purified from free saporin by gel filtration on a
G200 Sephadex column. Fractions containing free Tf and the two mainconjugates (with one or two saporin molecules linked to Tf) were pooledand concentrated. Further purification to remove excess TI was achievedby using ion exchange chromatography on a CM-FastFlow column in 10mM phosphate buffer, pH 7.0, eluting the proteins with a NaCI gradient(0 to 0.3 M).
As a control for our hypothesis (see below) that intracellular transportof conjugates is dictated by the carrier, we synthesized a covalent com-plex (B ricin-Sap) between ricin B chain and saporin (rhodamine-la-beled), following a similar procedure.
Binding and uptake of Tf and Tf-Sap to HepG2 cells
Human TI and TI-Sap conjugate were labeled with 1251 by chloramine I
(22). Binding to membrane receptors was determined at 4#{176}Cby incuba-tion of HepG2 cells in 24-well plates (cell density 2 x 10#{176}/well)for 90mm with various concentrations of ‘25I-labeled Tf-Sap. For ligand uptake
studies, HepG2 (2 X 10#{176}/well)were incubated in 0.5 ml of Hank’sbalanced salt solution containing 1% bovine serum albumin (HBSA) withvarying amounts of t2#{176}I-labeledIf (specific activity 1.59 x i0 cpnilng)and ‘2#{176}I-labeledTI/Sap (specific activity 1.62 x 10 cpm/ng) for 60 mmat 37#{176}C.The cells were then washed three times with ice-cold PBS andlysed with 0.5 ml of 0.1 M NaOH. Cell lysates were assayed with agamma counter to determine the cell-associated ligand. Nonspecific up-
take was estimated in all cases by including 1 mg/mI holo-transferrin in
control wells.
Inhibition of protein synthesis
Toxicity of TI-Sap conjugate was tested on HepG2 cells grown in Eaglemodified Dulbecco medium (D-MEM, Gibco, Caithersburg, Md.) con:taming 10% fetal calf serum. Cells cultured for 2 days (1.5 x 10#{176}cells/well) were preincubated with Tf free medium for 1 h and then
treated with serial dilution of the chimeric proteins for 18 h at 37#{176}C.Themedium was then removed and 0.5 ml of leucine free medium containing3H-labeled leucine (1 ItCi/mI) was added to each well; cells were incu-bated for one additional hour at 37#{176}C.Cells were washed three times withice-cold HBSAmedium before adding 1 ml of 10% TCA to each well.
The protein precipitates were dissolved by adding 0.25 ml of 0.1 MNaOH to each well. Aliquots of cell lysates (100 RI) were mixed with 5ml of scintillation mixture and radioactivity was measured in a liquidscintillation counter. The results are expressed as a percentage of theradioactivity incorporated in control cells.
Video microscopy
A Zeiss Axiophot microscope equipped with a 100 W epi-illuminationlamp, an electric shutter, and the standard filter sets for fluorescence wasused throughout. A solid-state CCD camera (type 588, by 12S, France)was connected to the microscope outlet and an Apple LIGS PC was used
to control the camera and shutter of the microscope (23). Image analysis
was performed on an Apple MacIntosh IIvx using the program Image (byW. Rasband, NIH, Bethesda, Md., freely distributed) and Photoshop.
Cell labeling and images acquisition
HepG2 cells were grown in D-MEM (Gilico) at 37#{176}cunder 5% CO2 oncoverslides, up to an approximate density of 2 X 10#{176}cells/mI and incu-
bated with a 5 RM solution of 6-((N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)caproyl)sphingosine (C6-NBD-ceramide) as described (24).NBD-labeled cells were incubated in Hank’s medium with rhodamine-labeled If or Tf-Sap (600 nM); after 30 mm at 37#{176}Cthe coverslides werewashed, mounted on microscope slides, and transferred to the thermo-stated plate of the microscope (at 37#{176}C).A similar procedure was usedto perform the experiments in which cells were incubated either withfluorescein-labeled (on the A chain) ricin and rhodamine-labeled Tf-Sapor B ricin-Sap conjugate.
RESULTS
Purification
The reaction mixture yields two covalent conjugates con-taining saporin and Tf in a molar ratio of 2:1 and 1:1,respectively (see Fig. 1A). Further purification of the two
conjugates was not attempted, and we shall refer to the
chromatographic fraction (analyzed in Fig. 1B, lane 2) asTf-Sap. The reaction also yields high molecular weightcomplexes, which proved to be nontoxic to intact HepG2cells (data not shown).
In a previous paper (13), similar chimeras prepared by aslightly different procedure contained Tf and saporin invariable ratios; therefore, we think that the overall toxicityof the conjugates reported in ref 13 may have been under-estimated because of the presence of nontoxic high molecu-lar weight complexes.
Binding to membrane receptors and cellular uptake
Binding of ‘251-labeled Tf-Sap and ‘251-labeled Tf to

toxicity of Tf-Sap was scarcely affected, which suggests that
the pathway of intracellular activation of the chimeric con-jugate does not exclusively involve transport to the Golgi
apparatus. Thus, we directed observations of the intracellu-lar distribution of Tf-Sap by fluorescence video microscopy.
Fluorescence microscopy assays on living HepG2
cells
Fluorescent derivatives of Tf, Tf-Sap, and ricin were pre-pared (26) in order to monitor by optical microscopy theirinternalization and intracellular pathway in living cells.This approach has been used before to demonstrate the
rapid accumulation of ricin in the Golgi apparatus of living
CHO cells and to determine the kinetics of the process
(18).Figure 4.4 shows that after 30 mm of incubation, rho-
damine-labeled Tf-Sap (red emission) was distributed in
A
1 2 1 234
L_li . <-- If.-#{176}#{176}--- c-- Sep.
1#{149}A B
Figure 1. A) SDS-PAGE electrophoresis of the pool of the chroma-tographic fractionsfrom gel filtration on G200. Lane 1 shows the twocomplexes between TI and saporincontainingone ortwo saporinmolecules
(as estimated from the molecular weight),as wellas unreacted TI. Lane 2shows the same sample afterdenaturation in the presence of f3-mercap-toethanol. Arrows indicate the TI and saporin positions. B) SDS-PAGEelectrophoresis of TI-Sap after purification from freeTf as obtained by ionexchange chromatography on CM-Sepharose Fast Flow column. Lanes 1and 3: control TI under nonreducing and reducing conditions, respectively.Lanes 2 and 4: purified TI-Sap under nonreducing and reducing condi-tions, respectively.
HepG2 cells was assessed in order to quantitate the affinityfor the Tf receptor. Binding at 4#{176}Cof Tf and Tf-Sap to cellsshowed specificity and saturability with increasing ligandconcentrations. The total number of Tf receptors on the cellsurface was calculated to be 3 x 105/cell, with an affinityconstant (K5) of 4 x 1011 M-’ (see Fig. 2/1).
To assess whether both Tf-Sap and Tf are internalizedwith the same efficiency we performed cellular uptake ex-periments at 37#{176}C.Figure 2B shows the concentrationdependence of 1251-labeled toxin uptake via the Tf receptor.The results show that after incubation for 60 mm at 37#{176}C,uptake reaches apparent saturation at high ligand concen-
trations. This finding supports the hypothesis that Tf-Sapand Tf are internalized by the same mechanism.
Inhibition of protein synthesis
Cytotoxicity of Tf-Sap, ricin, and saporin on HepG2 cells isshown in Fig. 3A. It is evident that whereas saporin isnontoxic to intact HepG2 cells, Tf-Sap inhibits protein syn-thesis in a dose-dependent manner. The ID50 for Tf-Sap is
approximately 5-7 x i0 M, about three orders of magni-tude greater than that of ricin. Addition of free Tf (2.5 xiO M) to the incubation medium (filled and blank squaresin Fig. 3/1) increases the ID50 for Tf-Sap by 10- to 100-fold,showing that uptake of the conjugate occurs via the Tfreceptor, leading to transport at the site of processing.
It has been shown (25) that the fungal drug brefeldin A(BFA. from SIGMA) leads to disassembly of the Golgi ap-paratus, which is known (17) to be involved in the mecha-nism of activation of ricin. As expected, ricin toxicity was
almost completely abolished by preincubation of cells withBFA (Fig. 3B, blank and filled squares); on the contrary,
0 0.04 0.08 0.12
Bound (nM)
0.16 0.2
B
0.0O4-- - .
.0
- 0.003--- ----#{149}----#{149}--- --------- . - - - ----
- - ‘S
0.002t..-.-. - .-..‘ -
0.00+ I -.
IProteinconcentration4tg/mI)
Figure 2. A) Scatchard plot showing the binding of ‘251-labeled If-Sap tocellular membrane receptors of HepG2 cells at 4#{176}C.The slope yields Ka,and the intercept on the X axis shows the total number of binding sites/wellin the incubation plate. B) Uptake of ‘251-labeled TI (0) and Tf-Sap (#{149})on HepG2 cells, as determined after 60 mm incubation at 37#{176}C.The two
binding curves are largely superimposed, indicating that the affinity ofTI-Sap for the TI receptor is unchanged compared to Tf.
0 4 8 12 16 20
1222 Vol.9 September 1995 The FASEB journal IPPOLITI ETAL
RESEARCH COMMUNICATION

A#{149} --
80- #{149}-- -
--- --- -
4Q:
A
U-
-U
1
0
0
0
C’S
a
0
-“
.13 .12 .11 .10 -9 -8 -7 .6
Log. protein concentration (M)
B
-9.5 -9 -8.5 -8
Log. protein concentration (M)Figure 3. A) Inhibition of protein synthesis expressed as % of [3H]leucineincorporation in HepG2 cells. Symbols refer to: ricin (A), If-Sap (U),TI-Sap in the presence of constant amounts of If (500 tg per well) (0),sapormn (s).B) Effect of the drug brefeldin A (BFA) on the inhibition of
protein synthesis by If-Sap and ricin. Symbols refer to: ricin (0), ricin in
the presence of 5 Jig/mI BFA (U), If-Sap (0), If-Sap in the presence of5pg/m1BFA(S).
regularly shaped vesicular structures scattered throughout
the cytoplasm. In contrast, C6-NBD-ceramide (green emis-
sion), a phospholipid derivative that accumulates in theGolgi apparatus of living cells (24), showed a distinct per-
inuclear distribution characteristic of the Golgi complex.
These results suggest that Tf-Sap does not localize in theGolgi apparatus.
To compare the intracellular distribution of Tf-Sap (rho-
damine-labeled, red emission) witk that of ricin (fluo-rescein-labeled), we incubated cells with both toxinssimultaneously. As expected (see Fig. 4B), ricin is localizedin a very compact region of the cell scarcely overlappingthe vesicular structures containing Tf-Sap.
In a control experiment (Fig. 4C), cells were incubatedwith C6-NBD-ceramide and rhodamine-labeled Tf for 30mm. The pattern of intracellular distribution of Tf closelymatches that of Tf-Sap as shown in Fig. 4A, indicating thatTf-Sap follows the same intracellular pathway as Tf.
RESEARCH COMMUNICATION
CELLULAR PROPERTIES OF CHIMERIC SAPORIN 1223
Further experiments were performed with B ricin-Sap todemonstrate that the carrier molecule drives the intracellu-lar pathway of chimeras. As shown in Fig. 4D, B ricin-Sap
colocalizes with C6-NBD-ceramide; this behavior is similarto that of ricin (18) and suggests that ricin B chain linkedto saporin transports the toxin inside cells, imposing its own
pathway, which is different from that dictated by transferrin
as a vector.
DISCUSSION
In view of its highly efficient endocytotic pathway, Tf hasbeen used as a carrier to deliver various molecules insidecells. Among other examples, the conjugates between hu-
man Tf and RIPs are interesting for their potential as mod-
els for anti-cancer agents. Two chimenc proteins obtainedthrough reaction of Tf with ricin A chain or saporin have
been previously prepared (13, 14). These covalent conju-gates proved to be inhibitors of proliferation of humanerythroleukemic cell lines after long (> 48 h) incubation;under these conditions their ID50 (protein concentrationinhibiting 50% proliferation) was in the range of 10b0 to
10-11 M. To clarify the intracellular routing and the possibleactivation mechanism of these chimeric proteins, we syn-
thesized Tf-Sap, an artificial conjugate between human Tf
and either saporin or rhodamine-labeled saporin, and com-pared its toxicity and intracellular distribution with that ofricin (the toxic lectin from R. communis).
Inhibition of protein synthesis in human HepG2 cells
.7.5 .7 .6.5 .6 after treatment with Tf-Sap yields (Fig. 3A) an ID50 (about6 nM) approximately 1000-fold greater than that of ricinunder the same conditions. The conjugate between ricin A
chain and Tf was itself 100-fold less toxic than native ricinwhen tested on CEM cells (a T cell leukemic line; see ref
14). No comparative data were available for the Tf-Sapconjugates.
Differences in toxicity between ricin and artificial conju-
gates carrying a RIP may be tentatively correlated to differ-ences in the activation mechanism and the endocytoticpathway of these two molecules as dictated by the carrierprotein after binding to the plasma membrane (27). Ricin
binds to glycolipids and glycoproteins through its lectin-like B chain and is internalized by receptor-mediated endo-cytosis (see refs 16, 17 for recent reviews). The mechanismby which the two constituent polypeptide chains are sepa-rated and the toxic A chain released in an active form in thecytoplasm is still under investigation. We recently demon-
strated (18) that in living CHO cells ricin is rapidly deliv-
ered to the Golgi apparatus, the likely site of activation(17). On the other hand, artificial conjugates of RIPs syn-thesized in order to bind and be internalized via more spe-cific receptors may localize in unfavorable intracellular
compartments. Our results (Fig. 4) suggest that the distri-bution of rhodamine-labeled Tf-Sap conjugates, followed in
living HepG2 cells by fluorescence video microscopy, is
definitely different from that of ricin: in fact, the latter is
quickly (t1/2 30 mm) accumulated in the Golgi apparatus,
RESEARCH COMMUNICATION
1224 Vol.9 September 1995 The FASEB journal IPPOLITI ETAL
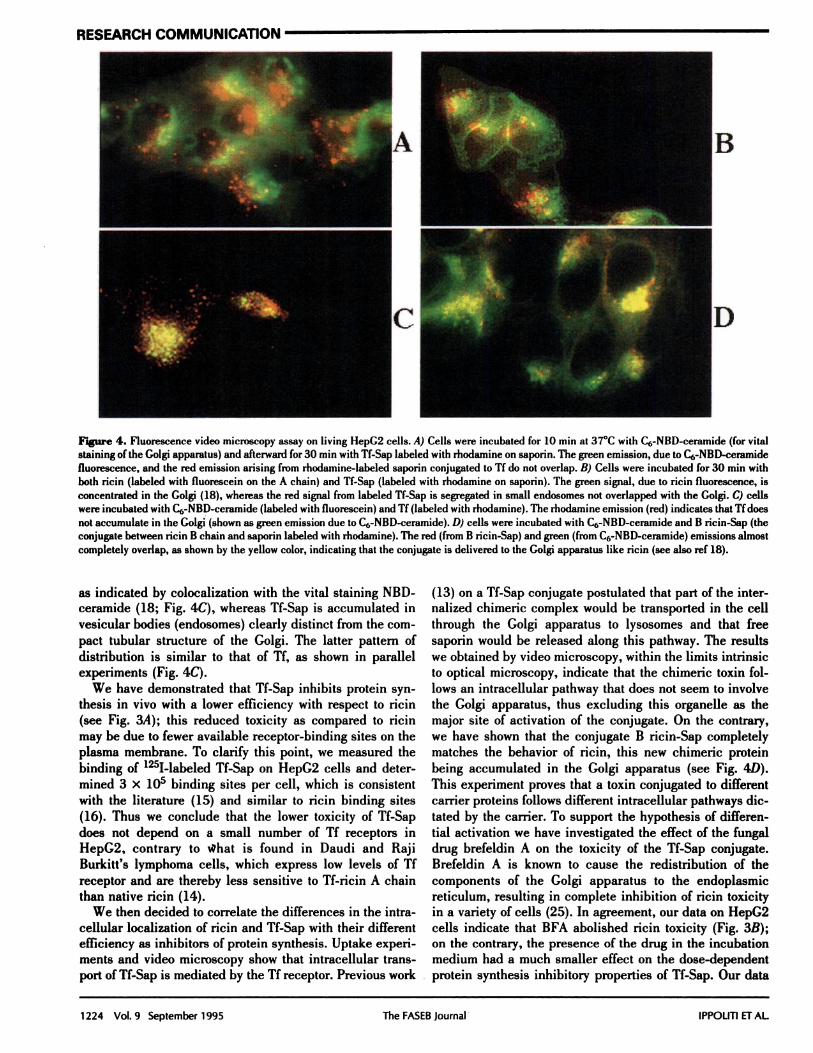
Figure 4. Fluorescence video microscopy assay on living HepC2 cells. A) Cells were incubated for 10 mm at 37#{176}Cwith C6-NBD-ceramide (for vital
staining of the Golgi apparatus) and afterward for 30 mm with If-Sap labeled with rhodamine on saporin. The green emission, due to C6-NBD-ceramidefluorescence, and the red emission arising from rhodamine-labeled saporin conjugated to If do not overlap. B) Cells were incubated for 30 mm with
both ricin (labeled with fluorescein on the A chain) and TI-Sap (labeled with rhodamnine on saporin). The green signal, due to ricin fluorescence, isconcentrated in the Golgi (18), whereas the red signal from labeled If-Sap is segregated in small endosomes not overlapped with the Golgi. C) cellswere incubated with C6-NBD-ceramide (labeled with fluorescein) and If (labeled with rhodamine). The rhodamine emission (red) indicates that II does
not accumulate in the Golgi (shown as green emission due to C6-NBD-ceramide). D) cells were incubated with C6-NBD-ceramide and B ncmn-Sap (theconjugate between ricin B chain and saporin labeled with rhodamine). The red (from B ricin-Sap) and green (from C6-NBD-ceramide) emissions almostcompletely overlap, as shown by the yellow color, indicating that the conjugate is delivered to the Golgi apparatus like ricin (see also ref 18).
as indicated by colocalization with the vital staining NBD-ceramide (18; Fig. 4C), whereas If-Sap is accumulated invesicular bodies (endosomes) clearly distinct from the com-
pact tubular structure of the Golgi. The latter pattern of
distribution is similar to that of Tf, as shown in parallelexperiments (Fig. 4C).
We have demonstrated that Tf-Sap inhibits protein syn-thesis in vivo with a lower efficiency with respect to ricin(see Fig. 3A); this reduced toxicity as compared to ricinmay be due to fewer available receptor-binding sites on theplasma membrane. To clarify this point, we measured thebinding of 1251-labeled Tf-Sap on HepG2 cells and deter-mined 3 x i0 binding sites per cell, which is consistentwith the literature (15) and similar to ricin binding sites(16). Thus we conclude that the lower toxicity of Tf-Sapdoes not depend on a small number of Tf receptors inHepG2, contrary to what is found in Daudi and RajiBurkitt’s lymphoma cells, which express low levels of Tfreceptor and are thereby less sensitive to Tf-ricin A chainthan native ricin (14).
We then decided to correlate the differences in the intra-cellular localization of ricin and Tf-Sap with their differentefficiency as inhibitors of protein synthesis. Uptake experi-ments and video microscopy show that intracellular trans-port of If-Sap is mediated by the Tf receptor. Previous work
(13) on a Tf-Sap conjugate postulated that part of the inter-nalized chimeric complex would be transported in the cell
through the Golgi apparatus to lysosomes and that freesaporin would be released along this pathway. The resultswe obtained by video microscopy, within the limits intrinsicto optical microscopy, indicate that the chimeric toxin fol-lows an intracellular pathway that does not seem to involvethe Golgi apparatus, thus excluding this organelle as themajor site of activation of the conjugate. On the contrary,we have shown that the conjugate B ricin-Sap completelymatches the behavior of ricin, this new chimeric proteinbeing accumulated in the Golgi apparatus (see Fig. 4D).
This experiment proves that a toxin conjugated to differentcarrier proteins follows different intracellular pathways dic-
tated by the carrier. To support the hypothesis of differen-tial activation we have investigated the effect of the fungaldrug brefeldin A on the toxicity of the Tf-Sap conjugate.Brefeldin A is known to cause the redistribution of the
components of the Golgi apparatus to the endoplasmicreticulum, resulting in complete inhibition of ricin toxicityin a variety of cells (25). In agreement, our data on HepG2cells indicate that BFA abolished ricin toxicity (Fig. 3B);
on the contrary, the presence of the drug in the incubationmedium had a much smaller effect on the dose-dependentprotein synthesis inhibitory properties of Tf-Sap. Our data

CELLULAR PROPERTIES OF CHIMERIC SAPORIN 1225
RESEARCH COMMUNICATION
therefore suggest that the lower toxicity of Tf-Sap comparedto ricin may be due to localization into a different intracel-lular compartment where activation presumably occurs withlower efficiency. However, the possibility that the chimericprotein complex may have an intrinsically lower catalytic
activity on eukaryotic ribosomes cannot be excluded.Artificial conjugates may be delivered to compartments
other than the site of activation, such as lysosomes whereproteolytic degradation is active. In the case of ricin A
chain toxicity on mouse macrophages (28), partial proteoly-sis has been involved as a mechanism of activation. Fur-thermore, in some cases eukaryotic cells were found to beresistant to the toxic action of RIPs (29), a behavior thatmay depend on some modification of the ribosomal proteins
or the RNA moiety involved in the mechanism of recogni-tion between the toxin and the ribosome. Some of thesephenomena may influence the toxicity of the conjugate.
In conclusion, we have obtained original data supportingthe view that natural and artificial toxins are both controlledin their endocytotic pathway by the carrier molecule,whether the B chain in the case of ricin or If in the case ofIf-Sap. The activation process, namely, separation of thetwo polypeptides and release of the active component into
the cytoplasm, is likely to occur as the toxin is deliveredeither to a highly specialized compartment (the Golgi) or toa vesicular structure (secondary endosomes). We proposethat the in vivo toxicity is related to differences in the
efficiency of the intracellular activation occurring in thecompartments where the toxin is (by and large) segregated,although it cannot be excluded that the whole chimerictoxin is still active on ribosomes, albeit with a differentcatalytic efficiency. Ihe latter possibility remains to beexcluded and will be crucial in order to decide on a soundstrategy for new chimeric toxins containing RIPs. Becausethese chimeras are potentially useful in cancer therapy,synthesis of conjugates that can be targeted to specific in-tracellular compartment where toxin activation is facilitatedmay be of medical interest.
Work was partially supported by the C.N.R. Target Project “Biotec-nologie e biostrumentazione.” We express our appreciation to the Uni-versity of Rome “La Sapienza” (Commissione di Ateneo) for a grant toacquire the videomicroscope. Very stimulating discussions with A.Bellelli (C.N.R., Rome) are gratefully acknowledged.
REFERENCES
1. Dautm-y-Varsat, A. (1986) Receptor-mediated endocytosis: the intracellularjoumney of transferrmn and its receptor. Biochimie 68, 375-381
2. Aisen, P., and Listowsky, L. (1980) Iron transport and storage proteins. Annu.
Rev. Biochern. 49, 359-3733. Killish, I., Steinlein, P., Romish, K., Hollinshead, R., Beng, H., and Gm-iftiths,
C. (1992) Characterization of early and late endocytic compartments of thetransfemn cycle.). Cell Sci. 103. 211-232
4. Fanlk, W. P., Hsi. B. L., and Stevens. P. L. (1980) Transferrin and transferrinreceptors in carcinoma of the breast. Lance: ii, 390-392
5. Sutherland, R., Delia, D., Schneider. C., Newman, R., Kenshead, J.. andcreaves, M. (1981) Ubiquitous cell-sum-face glycopromein on tumor cells is
proliferation-associated receptor for transfemn. Proc. Nat!. Acad. Sci. USA78,4515-4519
6. Wellhoner, H. H., Neville, D. M., Jr., Srinivasachar, K., and Erdmann, C.(1991) Uptake and concentration of bioactive macromolecules by K562 cellsvia the transferrin cycle utilizing an acid-labile transferrin conjugate. I. Rio!.Chem. 266, 4309-4314
7. Stirpe, F., and Barbieri, L. (1986) Ribosome inactivating proteins up to date.FEBS Lee. 195, 1-8
8. Endo, Y., and Tsurugi, K. (1987) RNA N-glycosidase activity of ricin A-chain.J. Biol. Chem. 262,8128-8130
9. Endo, Y., Mitsui, K., Motizuki, M., and Tsurugi, K. (1987) The mechanism ofaction of ricin and related toxic lectins on eukaryotic ribosomes.J. Biol. Chem.
262, 5908-591210. Singh, V., and Sairam, M. R. (1989) Hormonotoxins. 1. Strategy for synthesis
of ovine luteinizing hormone-gelonin conjugate bearing the toxin in the betasubunit. mt.J. Pept. Pro:. Res. 33, 22-28
11. Vitetta, E. S., and Uhr,J. W. (1985) Immunotoxins. Anne. Rev. Immuno!. 3,197-2 12
12. Ramakrishnan, S., Fryxel, D., Mahanraj, D., Olson, M., and Li, B. Y. (1992)Cytotoxic conjugates containing translational inhibitory proteins. Anne. Rev.Pharmacol. Toxicol. 32, 579-621
13. Bergamaschi, G., Cazzola, M., Dezza, L., Savino, E., Consonni, L., and Lappi,0. (1988) Killing of K562 cells with conjugates between human transferrinand a ribosome inactivating protein (SO-6). Br.). Hemato!. 68, 379-384
14. Raso, V., and Basala, M. (1984) A highly cytotoxic human Tf-ricin A chainconjugate used to select receptor-modified cells. J. Biol. Chem. 259,1143-1149
15. Stoorvogel, W., Genze, H. J., Griffith, J. M., and Strons, C. J. (1988) Thepathways of endocytosed transferrin and secretomy protein are connected inthe trans-Golgi reticulum.). Cell Biol. 106, 182 1-1829
16. Barbieri, L., Battelli, M. G., and Stirpe, F. (1993) Ribosome inactivatingproteins from plants. Biochim. Biophys. Acta 1154, 237-282
17. Sandvig, K., and van Deurs, B. (1994) Endocytosis and intracellular sortingof ricin and Shiga toxin. FEBS Leu. 346. 99-102
18. Lendaro, E., Ippoliti, R.. Bellelli, A.. Brunori, M., Evangelisma, V., Guidarini,D., and Benedetti, P. A. (1994) Intracellular dynamics of ricin followed by
fluorescence microscopy on living cells reveals a rapid accumulation of thedimeric toxin in the Golgi apparatus. FEBS Leu. 344,99-104
19. Barbieri, L., Stoppa, C., and Bolognesi, A. (1987) Large scale chroma-tographic purification of ribosome inactivating proteins. J. Chromatography408, 235-243
20. Carlson, J., Drevin, H., and Axen, R. (1978) Protein thiolation and reversibleprotein-protein conjugation. Biochem. J. 173. 723-737
21. Barbieri, L., Dinota, A., Gobbi, M., Tazzami, P. L., Rizzi, S., Bontadini, A.,Lemoli, R. M., Tura, S., and Stirpe, F. (1989) Immunotoxins containingsaporin 6 and monoclonal antibodies recognizing plasma cell associatedantigens: effects on target cells and on normal myeloid precursors. Eur. J.Haematol. 42, 238-245
22. Greenwood, F. C., Hunter, W. M., and Clover, J. S. (1963) The preparationof t3t I-labeled human growth hormone of high specific radioactivity. Biochem.J. 89, 114-123
23. Bellelli, A., Ippoliti, R., Lendaro, E., Brunori, M., Benedeui, P. A., Evangel-ista, V., Cuidarini, D., and Vestri, S. (1994) Analysis of biochemical processesin single living cells by quantitative microscopy. Biochem. Mo!. Biol. In:. 33,289-312
24. Sleight, R. C., and Pagano, R. E. (1984) Transport of a fluorescent phospha-tidylcholine analog from the plasma membrane to the Golgi apparatus.). CellBiol. 99, 742-751
25. Yoshida, T., Chen, C., Zhang, M., and Wu, H. C. (1981) Disruption of theGolgi apparatus by brefeldin A inhibits the cytotoxicity of ricin, modecin andPseudomonastoxin. Exp. Cell. Rev. 192, 389-395
26. Bellelli, A., Ippoliti, R., Brunori, M., Kam, Z., Benveniste, M., Emmanuel, F.,Turpin, E., Alfsen, A., and Fienoj, J. P. (1990) Binding and intemalization ofricin labelled with fluorescein isothiocyanate. Biochem. Bwphys. Rev. Corn-mun. 169, 602-609
27. Tonevitsky, A. C., Agapov, 1. 1., Mechetner, E. B., Ershow, C. V., Toptygin,A. Y., Sarma.T., Shamshiev, A. T.. and Pfueller, U. P. (1993) Comparison ofthe cytotoxic activity of the immunojoxins with different internalization rate.Biochem. Mol. Biol. In:. 31, 1059-1069
28. Blum, J. S., Fiani, M. L., and Stahl, P. D. (1991) Proteolytic cleavage of ricinA chain in endosomal vesicles. J. Biol. Chem. 266,22091-22095
29. Sallustio, S., and Stanley, P. (1990) Isolation of Chinese hamster ovaryribosomal mutants differentially resistant to ncin, abrin, and modeccin. J.Rio!. Chem. 265, 582-588
Received for publication March 20, 1995.Accepted for publication )une 22, 1995.
Copyright © 2022 FDOKUMEN




















