
Chimeric Rat/Human HER2 Efficiently Circumvents HER2 Tolerance In Cancer Patients
Upload
independentCategory
view
2download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Vaccine 27 (2009) 5152–5160
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
Immunogenicity of candidate chimeric DNA vaccine against tuberculosisand leishmaniasis�
Ayan Deya, Umesh Kumara, Pawan Sharmab, Sarman Singha,∗
a Department of Laboratory Medicine, All India Institute of Medical Sciences, New Delhi 110029, Indiab Immunology Group, International Centre for Genetic Engineering and Biotechnology, New Delhi 110067, India
a r t i c l e i n f o
Article history:Received 5 July 2008Received in revised form10 November 2008Accepted 31 May 2009Available online 24 June 2009
Keywords:Visceral leishmaniasisChimeraMultivalent VaccineKinesin motor domainesat-6IFN-�
a b s t r a c t
Mycobacterium tuberculosis and Leishmania donovani are important intracellular pathogens, especially inIndian context. In India and other South East Asian countries, both these infections are highly endemicand in about 20% cases co-infection of these pathogens is reported. For both these pathogens cell medi-ated immunity plays most important role. The available treatment of these infections is either prolongedor cumbersome or it is ineffective in controlling the outbreaks and spread. Therefore, potentiation of acommon host defense mechanism can be used to prevent both the infections simultaneously. In this studywe have developed a novel chimeric DNA vaccine candidate comprising the esat-6 gene of M. tuberculosisand kinesin motor domain gene of L. donovani. After developing this novel chimera, its immunogenicitywas studied in mouse model. The immune response was compared with individual constructs of esat-6and kinesin motor domain. The results showed that immunization with chimeric DNA vaccine constructresulted in stronger IFN-� and IL-2 response against kinesin (3012 ± 102 and 367.5 ± 8.92 pg/ml) andESAT-6 (1334 ± 46.5 and 245.1 ± 7.72 pg/ml) in comparison to the individual vaccine constructs. The recip-rocal immune response (IFN-� and IL-2) against individual construct was lower (kinesin motor domain:1788 ± 36.48 and 341.8 ± 9.801 pg/ml and ESAT-6: 867.0 ± 47.23 and 170.8 ± 4.578 pg/ml, respectively).The results also suggest that using the chimeric construct both proteins yielded a reciprocal adjuvant affectover each other as the IFN-� production against chimera vaccination is statistically significant (p < 0.0001)than individual construct vaccination. From this pilot study we could envisage that the chimeric DNA vac-cine construct may offer an attractive strategy in controlling co-infection of leishmaniasis and tuberculosisand have important implication in future vaccine design.
© 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Mycobacterium tuberculosis (MTB) and Leishmania donovani (LD)both are important human intracellular pathogens. Even thoughboth are distinct in aetiology and transmission dynamics. Yet cellmediated immunity plays a pivotal role in effective containmentof these diseases [1,2]. In the mouse model, infection with viru-lent MTB results in the activation and subsequent increase in thepopulation of CD4+ and CD8+ T cells in the lung draining lymphnodes [3] and migrates to lungs as the infection progress. Thisindicates that activated T cells migrate to the site of infection andinteract with antigen presenting cells on site to prevent reactiva-
� The partial data was presented at the Vaccine Congress 2007, held at Amsterdam,The Netherlands from December 9–11, 2007 (abstract no. O-11).
∗ Corresponding author at: Division of Clinical Microbiology, All India Institute ofMedical Sciences, PO Box # 4938, Ansari Nagar, New Delhi 110029, India.Tel.: +91 11 2658 8484; fax: +91 11 2658 8663/41.
E-mail address: sarman [email protected] (S. Singh).URL: http://www.drsarmansingh.com (S. Singh).
tion [2,3]. Similarly, in visceral leishmaniasis (VL) also helper andcytotoxic T cells are known to play an integral role in the immuneresponse to this infection, connecting the innate immune responseto the development of efficient adaptive cellular immunity, mainlythrough IL-2 and IFN-� production [4,5]. Moreover cytokine anal-ysis of serum from cured cases of visceral leishmaniasis revealeda characteristic type 1 response with increased IFN-�+, IL-2+ neu-trophils, eosinophils and NK cells [4,5].
Both these pathogens are also reported to act as co-factors inaggravating the disease. It is reported that Tuberculosis (TB) cancause immunosuppression by blocking macrophage responses toIFN-� by inhibiting the transcription of IFN-�-responsive genes,which results in the progression of latent leishmanial infectionto clinical manifestation [6,7]. Moreover, M. tuberculosis also con-tributes to the inability of an adaptive immune response to providecomplete protection by secreting an inhibitory factor(s), one ofwhich is IL-6. IL-6 inhibits IFN-� signaling in macrophages [7].Reciprocally, L. donovani can also reactivate latent Mycobacterialinfection [8]. Leishmania is known to down modulate nitric oxideproduction, correlating with reduction in inducible nitric oxidesynthase (iNOS) activity. Moreover, it has also been observed that
0264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.doi:10.1016/j.vaccine.2009.05.100

Author's personal copy
A. Dey et al. / Vaccine 27 (2009) 5152–5160 5153
Leishmania infected macrophages are less efficient in promotingthe sustained TCR (T-Cell Receptor) signaling necessary for activa-tion of T cell and for IFN-� production [9,10].
In the last few years rapid spread of tuberculosis in Indiaresulted in overlapping of tuberculosis and kala-azar endemiczones thus contributing to increased number of case with vis-ceral leishmaniasis–tuberculosis (VL–TB) co-infection [11]. It hasbeen estimated that about 65 new cases of M. tuberculosis occurper 100,000 population in Kala-Azar endemic areas of Bihar, India.Apart from India, in Sudan also approximately 20% cases of co-infection of VL–TB are reported [12]. Moreover, some cases of tripleinfection of HIV-TB-Kala-azar have also been reported [13,14]. Thusit has become imperative to control and prevent this co-infectionparticularly in areas where L. donovani and M. tuberculosis areendemic and occur concomitantly. Further, development of a duallyprotective vaccine may provide a common tool for control of theseco-infections. A common vaccine will also reduce the number ofvaccine doses.
For generation of chimeric vaccine construct the candidate genesselected were esat-6 gene of M. tuberculosis and kinesin motordomain of L. donovani. The ESAT-6 antigen coded by esat-6 geneis one of the major secreted proteins found in M. tuberculosisculture filtrates and is a dominant target for cell-mediated immu-nity in the early phases of infection in TB patients and in variousanimal models [15,16]. The esat-6 belongs to esat-6 multigene fam-ily and has five copies of cluster of esat-6 loci. These clusterscontain members of the CFP-10 (lhp) and esat-6 (esat-6) gene fam-ilies (encoding early secreted antigenic target) as well as genesencoding secreted, cell-wall-associated subtilisin-like serine pro-teases, putative ABC transporters, ATP-binding proteins and othermembrane-associated proteins. This region is conserved in all viru-lent strains of M. tuberculosis and M. bovis but deleted in all M. bovisBCG strains [17]. Studies have proven that esat-6 is a suitable andpromising vaccine candidate. For this various strategies have beenapplied by various workers which include recombinant BCG withesat-6 gene [18], esat-6 subunit vaccines [19], and conjugate DNAvaccine with mpt64, Ag85B, and esat-6 [20].
The kinesin motor domain region of L. donovani selected isalready known candidate for inducing Th-1 specific response inmouse model [21]. The gene coding for kinesin motor domain pro-tein is a member of kinesin protein superfamily, playing importantrole in cell division and intracellular transport of various car-gos, including vesicles, organelles, large protein complexes, andcytoskeletal filaments. It is highly conserved in nature both atnucleotide sequence and amino acid sequence level, demonstrating80–90% identity with other Leishmania species and 35–45% iden-tity in other living beings such as trypanosomes, mice and humans[22–26]. The kinesin gene of several Leishmania species had alreadybeen characterized completely and the immunodominant repeatregion is reported to have its potential application in diagnosingvisceral leishmaniasis [22,25,27]. Further in silico analysis and invivo studies in BALB/c model reveals that kinesin motor domain isa good vaccine candidate [21].
In the present study by co-expressing both these vaccine candi-dates together in mammalian expression vector we evaluated theimmunogenecity of the candidate chimeric DNA vaccine in BALB/cmouse model.
2. Materials and methods
2.1. Selection and isolation of kinesin motor domain and esat-6gene
The first step toward developing any vaccine involves a selec-tion of specific candidate. The previous studies and in silico analysis
of the amino acid sequences using MHC-I-prediction software [28]predicted epitopes for various HLA types. Hence, with this infor-mation, we selected kinesin motor domain of L. donovani andesat-6 domain of M. tuberculosis as candidates for chimeric DNAvaccine.
The kinesin motor domain was isolated by using the primerscapable of amplifying the open reading frame of kinesin motordomain (361–1350 bp) of L. donovani. The primers were designedwith following sequences: forward, 5′-CCA ATG CAT TGG ATG CACCCT TCC ACT GTG CGG-3′ and reverse, 5′-GGG CCC GTC GAT CACGCG CCC GAG CGT CGT-3′. Similarly primers for amplification ofRv-3875 region of esat-6 gene (14–296 bp) were designed withfollowing sequences: forward primer, 5′-CCC AAG CTT ATG ACAGAG CAG CAG TGG A-3′ and reverse primer, 5′-GCG GAT CCT GCGAAC ATC CCA GTG ACG-3′. These primers were designed withspecific restriction sites for cloning in suitable vector. The lin-ear map of the sequences along with restriction sites is shown inFig. 1A.
Using these primers, kinesin motor domain region was ampli-fied from MHOM/IN/KE16/1998 strain of L. donovani (GenBankAccession No. AY615886) and esat-6 gene was amplified from H37Rv strain M. tuberculosis (GenBank Accession No. AF420491). Theamplified products were isolated, purified and cloned in pGEM-T®
vector and later sequenced (Microsynth®, Switzerland) to verify theinsertion.
2.2. Generation of chimeric and individual vaccine constructs
For generation of chimeric vaccine construct directional cloningstrategy was adopted as shown in Fig. 1B. The plasmid express-ing the kinesin motor domain and ESAT-6 domain was preparedby cloning the respective genes on either side of self-cleaving pep-tide termed 2A, of the foot-and-mouth-disease virus (FMDV). Thisallows individual expression of the both gene from same vector[29].
Following sequence confirmation, the respective fragmentswere cleaved out from the pGEM-T® vector using specific restrictionenzyme. Gene sequence coding for FMDV 2A was also got synthe-sized artificially from Microsynth®, Switzerland and was cleavedout and inserted between kinesin motor domain and esat-6 regionsand ligated using T4 DNA ligase (MBI®). The chimeric construct wasfinally inserted into mammalian expression vector pVAX-1® understrong CMV promoter to drive the expression of the genes. After lig-ation recombination deficient and endonuclease deficient DH5-� E.coli strain was transformed using pVAX1-chimeric construct. All therecombinant molecules generated were screened using antibioticpressure selection. Finally the plasmid DNA was purified from therecombinant colonies.
Individual vaccine constructs were also generated to comparethe immunogenic efficacy of chimeric construct. For generation ofindividual vaccine construct, kinesin motor domain gene or esat-6gene was digested out and ligated directly into pVAX-1® vector. Theconstructs were transformed and isolated similarly as mentionedabove.
2.3. In vitro expression analysis of the constructs
The ability of the recombinant construct to express kinesinmotor domain and esat-6 domain proteins was studied in vitrousing TNT (Transcription and Translation) coupled transcrip-tion/translation system (Promega®) according to the manufac-turer’s instructions. TNT uses a coupled transcription/translationreaction for in vitro protein synthesis. Transcription and trans-lation take place simultaneously in the reaction: while theRNA polymerase transcribes the template gene, the ribosomeprovided by the TNT translates the 5′-end of nascent mRNA.

Author's personal copy
5154 A. Dey et al. / Vaccine 27 (2009) 5152–5160
Fig. 1. (A) Linear map of esat-6, FMDV 2A, and kinesin motor domain gene inserts with their respective restriction sites. (B) Directional cloning strategies for developingchimeric vaccine construct.
Further Western blot analysis confirmed the expression of theproteins
2.4. Western blot analysis of chimeric and individual construct
In order to check the expression of kinesin motor domain andESAT-6 domain, Western blot analysis was carried out accordingto protocol described earlier [22]. The in vitro reaction mixture ofthe proteins were resolved on 16% Tricine SDS–PAGE and trans-ferred onto nitrocellulose membranes (Millipore®, USA) using asemi-dry blotting apparatus (Bio-Rad®, USA) following the instruc-tions of the manufacturer. The membranes were blocked with 5%skimmed milk; washed thrice for 10 min each with PBS, pH 7.0,0.1% Tween-20, then incubated for 1 h at room temperature with1:50 dilution (in PBS, pH 7.0, 1% BSA) of patient sera. The mem-branes were washed thrice with PBS, pH 7.0, 0.1% Tween-20, andthen incubated for 1 h at room temperature with 1:1000 dilutionof biotin-conjugated anti-human IgG (in PBS, pH 7.0, 1% BSA). Fol-lowed by incubation with secondary antibody the membranes werewashed and then incubated with primary substrate avidine conju-gated to horseradish peroxidase. After washing thrice for 10 mineach with PBS, pH 7.0, 0.1% Tween-20, the blots were developed
using a DAB (Amresco®, USA) and 0.1% H2O2 as substrate and bandswere visualized directly on membrane.
2.5. Mouse immunization
The immunogenicity study for the chimeric vaccine was carriedout in inbred BALB/c mouse model after having ethical approvalfrom institute animal ethics committee. Six- to eight-week-oldBALB/c mouse were used in this study. Two days before DNA inoc-ulation, the quadriceps muscles were injected with 100 �l of asolution containing 0.25% bupivacaine hydrochloride to enhancesubsequent DNA absorption [30]. Four groups of six mice each wereused for vaccination studies, cell proliferation, cytokine productionand antibody response. The first group of mice was injected withchimeric construct. The second and third groups were injected withindividual kinesin motor domain and esat-6 constructs, respec-tively. Mouse injected with empty plasmid serves as control. ForDNA inoculation, 100 �g of DNA construct in PBS was injected intothe same region of the muscle as the bupivacaine injection. Twoweeks after immunization, animals were given booster dose. Micewere sacrificed 2 weeks following the final injections and spleensfrom the mice in each group were dissected out, aseptically.

Author's personal copy
A. Dey et al. / Vaccine 27 (2009) 5152–5160 5155
2.6. Splenocyte proliferation and cytokine assay
Spleens were removed from mouse under aseptic conditionson a sterile dish containing RPMI 1640 media. Single cell suspen-sions were prepared by grinding the spleen with disk bottom ofthe plunger of 10 ml syringe. The 5–10 ml of RPMI-1640 mediumwas added to it and the contents were mixed to homogeneity. Thedish was kept undisturbed for 2 min and the clear supernatant waspipetted out slowly. Cells were pelleted by centrifugation at 4 ◦Cfor 10 min at 250 × g in Sigma Centrifuge. The pellet containingred blood cells (RBCs) and splenocytes were collected. The pel-let was washed with 0.9% ammonium chloride which lyses theRBCs. The isolated splenocytes were than resuspended in RPMI1640 medium containing 10% FCS and 0.05 �M 2-mercaptoethanolabd adjusted to a density of 2.5 × 106 cells/ml. From this suspen-sion 5 × 105 cells were added to 200 �l of RPMI-1640 media with10% FCS in 96-well plates. The cells were stimulated with 1 �g/wellof mitogen concavalin A (control), Kinesin and ESAT-6 antigen. Thecells were than incubated for 48 h at 37 ◦C in atmosphere con-taining 5% CO2 and 95% humidity. Proliferation was measured byMTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro-mide (Sigma Aldrich, St. Louis, MO, USA)] assay as describedelsewhere, with minor modifications [31]. In brief, MTT solution(l0 �l, 5 mg/ml) was added to 200 �l of medium and incubated for3 h at 37 ◦C. After incubation the culture medium was removed anddark blue formazan crystals were dissolved in acidic isopropanol(100 �l of 0.04 N HCl in isopropanol). Color formation was mea-sured at A570 nm wavelength, using ELISA plate reader (BiotekInstruments, USA). Wells with Concavalin A served as positive con-trol while wells without any stimulant served as negative control.
The level of IFN-�, IL-2 and IL-4 were determined in culturesupernatant using Endogen mouse IFN-�, IL-2 and IL-4 ELISA kit(Pierce Biotechnology, Inc., USA) according to the manufacturerprotocol.
2.7. Delayed Type Hypersensitivity (DTH) response
For DTH responses, footpad swelling was measured in naiveBALB/c mouse injected with chimeric vaccine construct. Twomonths after boosting, each animal was injected in a hind footpadwith 20 �g of total soluble protein of L. donovani and M. tubercu-losis in 20 �l of PBS. Footpad swelling was measured with a dial
Fig. 2. Restriction analysis of the chimeric pVAX-1® clones having kinesin motordomain region and esat-6 region cloned together on two sides of self-cleaving FMDV-2A peptide. Lanes 1, 2 and 5: chimera with kinesin (BstX1 and Apa 1); lane 3: chimerawith kinesin and FMDV (BamH1 and Apa1); lanes 4, 6 and 7: chimera with esat-6(HindIII and BamH1); lane 8: 100 bp Marker (MBI®).
caliper at various times, and the results were expressed as the dif-ference between the thicknesses of the footpads inoculated withthe antigen and the thickness of the footpads inoculated with 20 �lof PBS.
2.8. Statistical analysis
Statistical analysis of the results and graph creation were donewith the Graph Pad Prism (version 3.0) and Microsoft Excel (version7.0) for general statistical calculations, such as arithmetic mean andstandard deviation. p values of <0.05 were considered significant.
3. Results
3.1. Generation of chimeric and individual vaccine constructs
Ligation of kinesin motor domain gene and esat-6 gene on twosides of self cleaving peptide and further cloning it into pVAX-1®
vector results in generation of chimeric vaccine construct. Restric-tion digestion analysis was used to confirm the insertion of chimericsequence into the vector as shown in the Fig. 2. Sequence analysis(from Microsynth® Switzerland) further confirmed the insertionof appropriate sequence in the desired orientation. Individual con-structs were also generated and their sequences were verified byrestriction digestion and sequence analysis (Fig. 3A and B).
Fig. 3. (A and B) Restriction analysis of the individual pVAX-1® clones having kinesin motor domain region and esat-6 region cloned. (A) Lane 1: 100 bp Marker (MBI®); lanes2 and 3: pVAX1 with kinesin; lanes 4 and 5: empty plasmid; (B) lane 1: 100 bp Marker (MBI®); lanes 2 and 3: empty plasmid; lanes 4 and 5: pVAX1 with esat-6.

Author's personal copy
5156 A. Dey et al. / Vaccine 27 (2009) 5152–5160
3.2. In vitro expression of kinesin motor domain and ESAT-6domain
In vitro expression of chimeric DNA vaccine constructs usingTNT® in vitro translation system resulted in co-expression of both35 kDa kinesin motor domain and 6 kDaESAT-6 domain from samevector. Further Western blot analysis confirmed the expression ofboth kinesin motor domain and ESAT-6 domain (Fig. 4A and B). Invitro expression and Western blot analysis of individual constructsare shown in Fig. 4A and B. In vitro expression of empty plasmid didnot result in expression of kinesin and ESAT-6 domain, as revealedby Western blot analysis (Fig. 4C).
3.3. Cellular immune response against leishmanial andtuberculosis epitopes
3.3.1. Proliferative response against kinesin motor domainSplenocytes proliferation assay of mouse group injected with
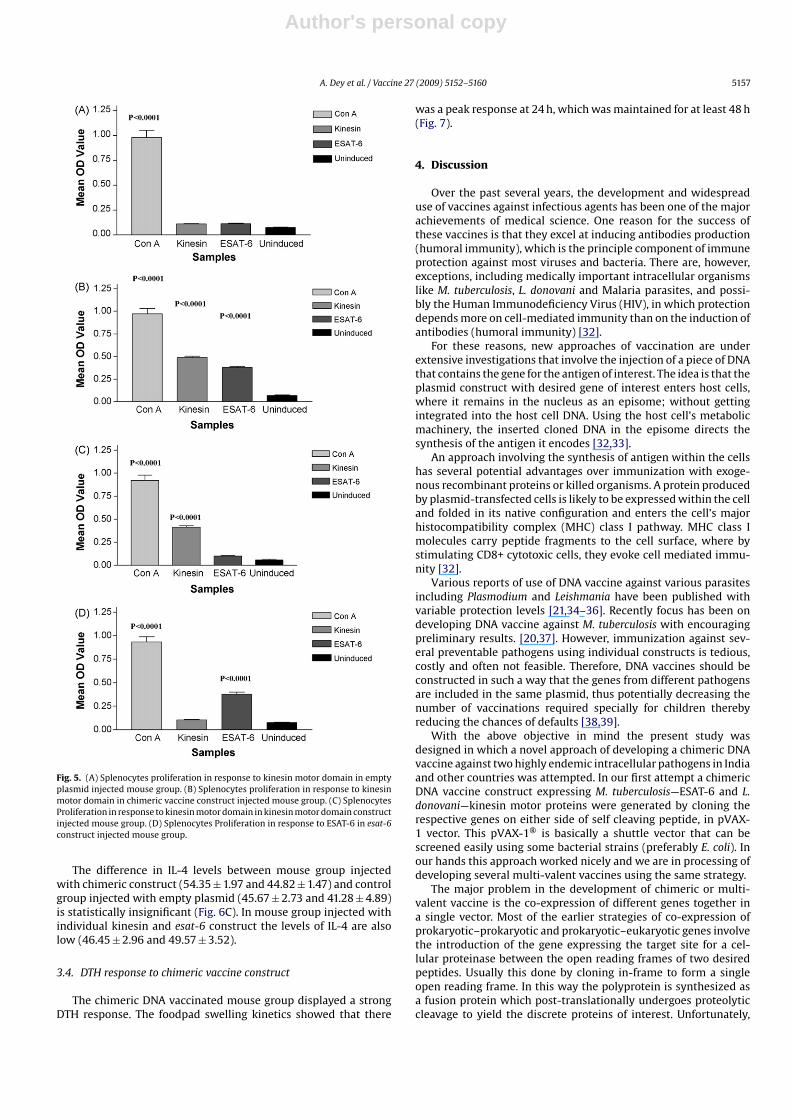
empty plasmid shows negligible proliferative responses againstexpressed kinesin motor domain (0.110 ± 0.004) (Fig. 5A). However,mouse group injected with Chimeric DNA vaccine construct prolif-erated vigorously against the kinesin motor domain (0.488 ± 0.015)which was statistically significant (p < 0.0001) than mouse groupinjected with empty plasmid (Fig. 5B). The mouse group injectedwith individual kinesin motor domain vaccine construct also gen-erated significant (p < 0.0001) proliferative response against kinesinmotor domain (0.415 ± 0.015) as shown in Fig. 5C. Concavalin A con-trols also showed good proliferative response both in vaccinatedand empty plasmid group.
3.3.2. Proliferative response against ESAT-6 domainSplenocytes isolated from mouse immunized with chimeric
DNA vaccine construct proliferated significantly (p < 0.0001)against stimulation with ESAT-6 (0.378 ± 0.013) as compared tothose obtained from mouse immunized with empty plasmid(0.069 ± 0.006) (Fig. 5A and B). However, the proliferative responseagainst ESAT-6 in comparison to kinesin motor domain was weak.This may be due to the fewer number of epitopes in ESAT-6 peptidesequence as compared to kinesin motor domain. The mouse groupinjected with individual esat-6 DNA construct also generated signif-icant (p < 0.0001) proliferative response (0.376 ± 0.022) as shown inFig. 5D.
3.3.3. IFN-� , IL-2 and IL-4 cytokine production against theconstructs
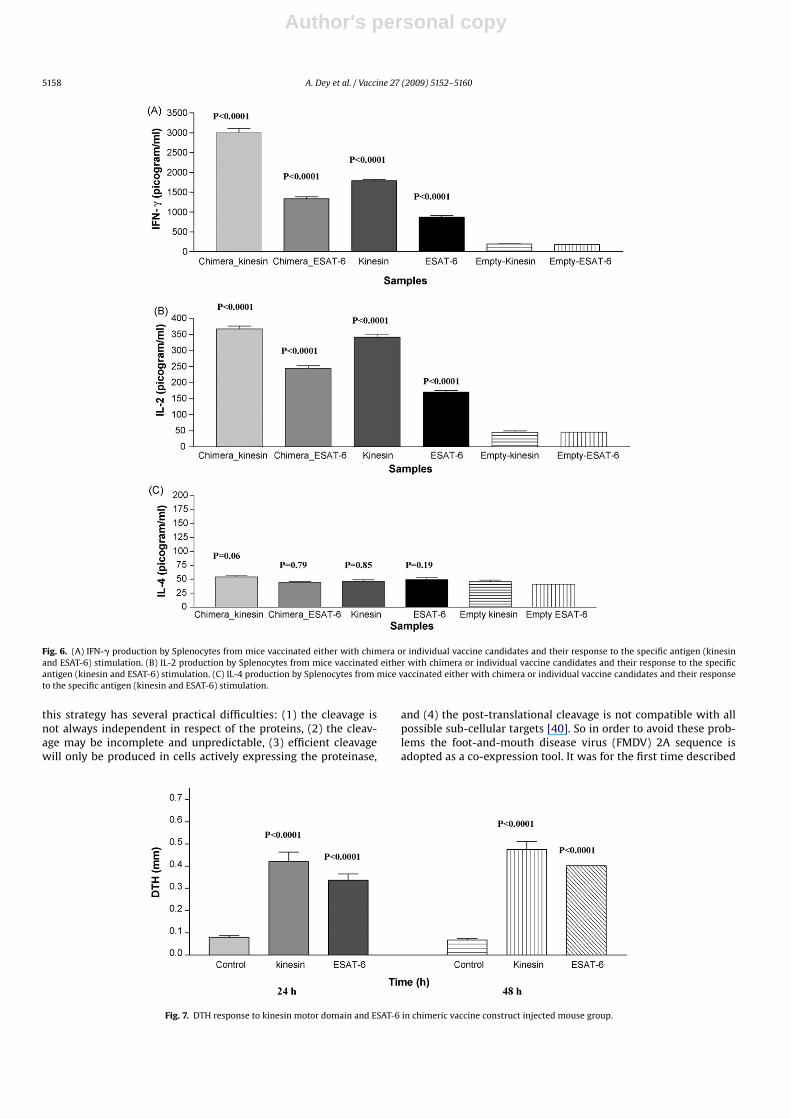
Cytokine analysis using ELISA revealed that Chimeric DNAvaccination generated strong cellular immune response result-ing in the production of higher levels of IFN-� (3012 ± 102and 1334 ± 46.5 pg/ml) against kinesin motor domain and ESAT-6 domain (Fig. 6A). The difference was statistically significant(p < 0.0001) as compared to mouse group injected with emptyplasmid (Fig. 6A). The mouse group injected with individ-ual vaccine constructs also generated higher levels of IFN-�against kinesin motor domain (1788 ± 36.48 pg/ml) and ESAT-6 (867.0 ± 47.23 pg/ml). However, mouse groups injected withchimeric construct generated stronger cellular response in compareto mouse groups injected individual vaccine construct as evidencedby higher IFN-� levels (p < 0.0001).
Mouse groups injected with chimeric vaccine construct showsIL-2 production against kinesin motor domain and ESAT-6(367.5 ± 8.92 and 245.1 ± 7.72 pg/ml) (Fig. 6B). Vaccination withindividual kinesin motor domain and esat-6 construct also resultedin production of IL-2 cytokine (341.8 ± 9.801 and 170.8 ± 4.578).Although vaccination with chimeric construct resulted in higherlevels of IL-2 production but the difference in IL-2 levels in chimeraand individual construct group was not statistically significant.
Fig. 4. A Western blot analysis of in vitro expression of kinesin motor domain. LaneM: prestained molecular weight marker (MBI®); lanes 1 and 2: in vitro expressionof kinesin from chimeric construct; lane 3: in vitro expression of kinesin from indi-vidual kinesin-pVAX1 construct; lane 4: purified rLvacc probed with VL positivepatient serum. (B) Western blot analysis in vitro expression of ESAT-6 domain. LaneM: prestained molecular weight marker (NEBpVAX1construct); lanes 2 and 3: invitro expression of ESAT-6 from chimeric construct; lane 4: in vitro expression ofESAT-6 from individual ESAT-6-pVAX1 construct; lane 5: purified ESAT-6 probedwith TB positive patient serum. (C) Western blot analysis of in vitro expression fromempty plasmid. Lane M: prestained molecular weight marker (MBI®); lanes 1–4:in vitro expression from empty plasmid using TNT translation system gives somenonspecific bands due to use of patient serum with polyclonal antibody.
Author's personal copy
A. Dey et al. / Vaccine 27 (2009) 5152–5160 5157
Fig. 5. (A) Splenocytes proliferation in response to kinesin motor domain in emptyplasmid injected mouse group. (B) Splenocytes proliferation in response to kinesinmotor domain in chimeric vaccine construct injected mouse group. (C) SplenocytesProliferation in response to kinesin motor domain in kinesin motor domain constructinjected mouse group. (D) Splenocytes Proliferation in response to ESAT-6 in esat-6construct injected mouse group.
The difference in IL-4 levels between mouse group injectedwith chimeric construct (54.35 ± 1.97 and 44.82 ± 1.47) and controlgroup injected with empty plasmid (45.67 ± 2.73 and 41.28 ± 4.89)is statistically insignificant (Fig. 6C). In mouse group injected withindividual kinesin and esat-6 construct the levels of IL-4 are alsolow (46.45 ± 2.96 and 49.57 ± 3.52).
3.4. DTH response to chimeric vaccine construct
The chimeric DNA vaccinated mouse group displayed a strongDTH response. The foodpad swelling kinetics showed that there
was a peak response at 24 h, which was maintained for at least 48 h(Fig. 7).
4. Discussion
Over the past several years, the development and widespreaduse of vaccines against infectious agents has been one of the majorachievements of medical science. One reason for the success ofthese vaccines is that they excel at inducing antibodies production(humoral immunity), which is the principle component of immuneprotection against most viruses and bacteria. There are, however,exceptions, including medically important intracellular organismslike M. tuberculosis, L. donovani and Malaria parasites, and possi-bly the Human Immunodeficiency Virus (HIV), in which protectiondepends more on cell-mediated immunity than on the induction ofantibodies (humoral immunity) [32].
For these reasons, new approaches of vaccination are underextensive investigations that involve the injection of a piece of DNAthat contains the gene for the antigen of interest. The idea is that theplasmid construct with desired gene of interest enters host cells,where it remains in the nucleus as an episome; without gettingintegrated into the host cell DNA. Using the host cell’s metabolicmachinery, the inserted cloned DNA in the episome directs thesynthesis of the antigen it encodes [32,33].
An approach involving the synthesis of antigen within the cellshas several potential advantages over immunization with exoge-nous recombinant proteins or killed organisms. A protein producedby plasmid-transfected cells is likely to be expressed within the celland folded in its native configuration and enters the cell’s majorhistocompatibility complex (MHC) class I pathway. MHC class Imolecules carry peptide fragments to the cell surface, where bystimulating CD8+ cytotoxic cells, they evoke cell mediated immu-nity [32].
Various reports of use of DNA vaccine against various parasitesincluding Plasmodium and Leishmania have been published withvariable protection levels [21,34–36]. Recently focus has been ondeveloping DNA vaccine against M. tuberculosis with encouragingpreliminary results. [20,37]. However, immunization against sev-eral preventable pathogens using individual constructs is tedious,costly and often not feasible. Therefore, DNA vaccines should beconstructed in such a way that the genes from different pathogensare included in the same plasmid, thus potentially decreasing thenumber of vaccinations required specially for children therebyreducing the chances of defaults [38,39].
With the above objective in mind the present study wasdesigned in which a novel approach of developing a chimeric DNAvaccine against two highly endemic intracellular pathogens in Indiaand other countries was attempted. In our first attempt a chimericDNA vaccine construct expressing M. tuberculosis—ESAT-6 and L.donovani—kinesin motor proteins were generated by cloning therespective genes on either side of self cleaving peptide, in pVAX-1 vector. This pVAX-1® is basically a shuttle vector that can bescreened easily using some bacterial strains (preferably E. coli). Inour hands this approach worked nicely and we are in processing ofdeveloping several multi-valent vaccines using the same strategy.
The major problem in the development of chimeric or multi-valent vaccine is the co-expression of different genes together ina single vector. Most of the earlier strategies of co-expression ofprokaryotic–prokaryotic and prokaryotic–eukaryotic genes involvethe introduction of the gene expressing the target site for a cel-lular proteinase between the open reading frames of two desiredpeptides. Usually this done by cloning in-frame to form a singleopen reading frame. In this way the polyprotein is synthesized asa fusion protein which post-translationally undergoes proteolyticcleavage to yield the discrete proteins of interest. Unfortunately,
Author's personal copy
5158 A. Dey et al. / Vaccine 27 (2009) 5152–5160
Fig. 6. (A) IFN-� production by Splenocytes from mice vaccinated either with chimera or individual vaccine candidates and their response to the specific antigen (kinesinand ESAT-6) stimulation. (B) IL-2 production by Splenocytes from mice vaccinated either with chimera or individual vaccine candidates and their response to the specificantigen (kinesin and ESAT-6) stimulation. (C) IL-4 production by Splenocytes from mice vaccinated either with chimera or individual vaccine candidates and their responseto the specific antigen (kinesin and ESAT-6) stimulation.
this strategy has several practical difficulties: (1) the cleavage isnot always independent in respect of the proteins, (2) the cleav-age may be incomplete and unpredictable, (3) efficient cleavagewill only be produced in cells actively expressing the proteinase,
and (4) the post-translational cleavage is not compatible with allpossible sub-cellular targets [40]. So in order to avoid these prob-lems the foot-and-mouth disease virus (FMDV) 2A sequence isadopted as a co-expression tool. It was for the first time described
Fig. 7. DTH response to kinesin motor domain and ESAT-6 in chimeric vaccine construct injected mouse group.

Author's personal copy
A. Dey et al. / Vaccine 27 (2009) 5152–5160 5159
in the 1980s that 18aa peptide (2A) of some picornaviruses (suchas FMDV) underwent a rapid co-translational self-processing anddirects its own separation from the growing polyprotein. During thelast decade or so, this mechanism has been sexploited to it fullestuse, leading to the development of a simple model in which a small2A peptide interacts with the exit tunnel of the ribosome to inducethe “skipping” of the last peptide bond at the C-terminus of 2A.All this happens during its translation stage while the ribosomecontinues to translat the downstream gene, after releasing the pre-vious protein which is simultaneously fused in its C-terminus to 2A[40]. This type of sequence is known as CHYSEL (cisacting hydrolaseelement).
We used the above co-translation element in present study,we demonstrated for the first time that a bi-valent DNA vac-cine expressing L. donovani and M. tuberculosis epitopes can begenerated. Our strategy worked satisfactorily as is evident fromsplenocytes proliferation assay which reveals induction of Th-1 spe-cific immune responses against the respective epitopes in mousemodel. ESAT-6 and Kinesin motor domain also produced IFN-� inresponse to stimulation with recombinant kinesin and esat-6 anti-gen in statistically significant amount (Fig. 6A). The splenocytescell lines were also assayed for IL-2 and IL-4 production. The IL-2 production was significantly more by these cells in response tostimulation with either kinesin or esat-6, but the levels of IL-4 werenot significantly higher than empty plasmid control group (Fig. 6Band C). This finding was also on the expected lines. As shown inFig. 7, control mice immunized with empty pVAX-1 plasmid didnot present any significant DTH reaction, whereas mice immunizedwith chimeric construct reacted significantly to L. donovani kinesinantigen and M. tuberculosis esat-6 antigens 24 h after injection. TheDTH reaction was maintained for up to 48 h after injection of theantigen. The measurement of DTH response is a parameter of signif-icance in identifying the presence or absence of a cellular immuneresponse [41,42]. Taken together our results clearly indicate that ourchimeric vaccine construct worked very satisfactorily and inducedcellular immune response against both the target pathogens. More-over, the immune response generated by chimeric construct wasfound to be superior to that of individual vaccine constructs, sug-gesting an adjuvant affect of these antigens over each other [43].Hence, our data shows that it is possible for two or more potentialTh-cell epitopes present in chimeric construct to act co-dominantly[43].
Ours is the first report in which the immunogenic epitopes fromthe genetically diverse causative agents the L. donovani, an eukary-ote and M. tuberculosis, a prokaryote were successfully clonedand expressed in a single DNA vaccine construct. These findingscould pave the way for chimeric multivalent DNA vaccines againstseveral diseases of man and animals and are endemic in givenrehions/countries.
Acknowledgements
This study was partially supported by an extramural grant fromAll India Institute of Medical Sciences, New Delhi and Departmentof Biotechnology, Government of India to SS. Fellowship to AD fromDepartment of Biotechnology, and Council of Scientific and Indus-trial Research, Government of India is acknowledged. Fellowship toUK from Council of Scientific and Industrial Research, Governmentof India is also acknowledged.
References
[1] Herwaldt BL. Leishmaniasis. Lancet 1999;354:1191–9.[2] Raja A. Immunology of tuberculosis. Indian J Med Res 2004;120:213–32.[3] Feng CG, Bean AGD, Hooi H, Briscoe H, Britton WJ. Increase in gamma interferon-
secreting CD8+, as well as CD4+ T cells in lungs following aerosol infection withMycobacterium tuberculosis. Infect Immun 1999;67:3242–7.
[4] Peruhype-Magalhães V, Martins-Filho OA, Prata A, Silva Lde A, Rabello A,Teixeira-Carvalho A, et al. Immune response in human visceral leishmania-sis: analysis of the correlation between innate immunity cytokine profile anddisease outcome. Scand J Immunol 2005;62(5):487–95.
[5] Kemp M, Kurtzhals JA, Bendtzen K, Poulsen LK, Hansen MB, Koech DK,et al. Leishmania donovani-reactive Th1- and Th2-like T-cell clones fromindividuals who have recovered from visceral leishmaniasis. Infect Immun1993;61:1069–73.
[6] Montalban C, Callejia JL. Leishmaniasis masking tuberculosis. Lancet1990;335(8684):299–300.
[7] Ting LM, Kim AC, Cattamanchi A, Ernst JD. Mycobacterium tuberculosis inhibitsIFN-� transcriptional responses without inhibiting activation of STAT1. JImmunol 1999;163:3898.
[8] Chaudhuri A. Leishmaniasis masking tuberculosis. Lancet 1989;2(8676):1396–7.
[9] Amprey JL, Spath GF, Porcelli SA. Inhibition of CD1 expression in humandendritic cells during intracellular infection with Leishmania donovani. InfectImmun 2004;72(1):589–92.
[10] Meier CL, Svensson M, Kaye PM. Leishmania-Induced inhibition ofmacrophage antigen presentation analyzed at the single-cell level. J Immunol2003;171:6706–13.
[11] Chadha VK, Kumar P, Gupta J, Jagannatha PS, Lakshminarayana, Magesh V, etal. The annual risk of tuberculous infection in the eastern zone of India. Int JTuberc Lung Dis 2004;8(5):537–44.
[12] el-Safi1 SH, Hamid N, Omer A, Abdel-Haleem A, Hammad A, Kareem HG, et al.Infection rates with Leishmania donovani and Mycobacterium tuberculosis in avillage in eastern Sudan. Trop Med Int Health 2004;9(12):1305–11.
[13] Pandey K, Sinha PK, Ravidas VN, Kumar N, Verma N, Lal CS, et al. Nexus ofinfection with human immunodeficiency virus, pulmonary tuberculosis andvisceral leishmaniasis: a case report from Bihar, India. Am J Trop Med Hyg2005;72(1):30–2.
[14] Wang J, Vanley C, Miyamoto E, Turner JA, Peng SK. Coinfection of visceral leish-maniasis and Mycobacterium in a patient with acquired immunodeficiencysyndrome. Arch Pathol Lab Med 1999;123(9):835–7.
[15] Brandt L, Oettinger T, Holm A, Anderson AB, Anderson P. Key epitopes on theESAT-6 antigen recognized in mice during the recall of protective immunity toMycobacterium tuberculosis. J Immunol 1996;157:3527–33.
[16] van Pinxteren LA, Ravn P, Agger EM, Pollock J, Anderson P. Diagnosis of tuber-culosis based on the two specific antigens ESAT-6 and CFP10. Clin Diagn LabImmunol 2000;7:155–60.
[17] Pittus NCGV, Gamieldien J, Hide W, Brown GD, Siezen RJ, Beyers AD. The esat-6gene cluster of Mycobacterium tuberculosis and other high G + C Gram-positivebacteria. Genome Biol 2001;2(10):1–18.
[18] Pym AS. Recombinant BCG exporting ESAT-6 confers enhanced protectionagainst tuberculosis. Nat Med 2003;9 [advance online publication].
[19] Brandt L, Elhay M, Rosenkrands I, Lindblad EB, Andersen P. esat-6 subunit vac-cination against Mycobacterium tuberculosis. Infect Immun 2000;68:791–5.
[20] Kamath AT, Feng CG, Macdonald M, Briscoe H, Warwick JB. Differential pro-tective efficacy of DNA vaccines expressing secreted proteins of Mycobacteriumtuberculosis. Infec Immun 1999;67:1702–7.
[21] Dey A, Sharma P, Redhu NS, Singh S. Kinesin Motor Domain of LeishmaniaDonovani as future vaccine candidate. Clin Vacc Immunol 2008;15(5):836–42.
[22] Sivakumar R, Sharma P, Chang KP, Singh S. Cloning, expression and purifica-tion of a novel recombinant antigen Leishmania donovani. Protein Expr Purif2006;46:156–65.
[23] Burns Jr JM, Shreffler WG, Benson DR, Ghalib HW, Badaro R, Reed SG. Molec-ular characterization of a kinesin-related antigen of Leishmania chagasi thatdetects specific antibody in African and American visceral Leishmaniasis. ProcNatl Acad Sci USA 1993;90:775–9.
[24] Sivakumar R, Dey A, Sharma P, Singh S. Expression and Characterization ofa recombinant kinesin antigen from an old Indian strain (DD8) of Leishma-nia donovani and comparing it with a commercially available antigen froma newly isolated (KE-16) strain of L. donovani. Infect Genet Evol 2008;8:313–22.
[25] Singh S, Gilman-Sachs A, Chang KP, Reed SG. Diagnostic and prognostic value ofK39 recombinant antigen in Indian leishmaniasis. J Parasitol 1995;81:1000–3.
[26] Iwai S, Ishiji A, Mabuchi I, Sutoh KA. Novel actin-bundling kinesin-related pro-tein from Dictyostelium discoideum. J Biol Chem 2004;6:4696–704.
[27] Takagi H, Islam MZ, Itoh M, Islam AU, Saifuddin Ekram AR, Hussain SM, et al.Short report: production of recombinant kinesin-related protein of Leishmaniadonovani and its application in the serodiagnosis of visceral leishmaniasis. AmJ Trop Med Hyg 2007;76(5):902–5.
[28] Singh H, Raghava GPS. ProPred: prediction of HLA-DR binding sites. Bioinfor-matics 2001;17(12):1236–7.
[29] Donnelly ML, Hughes LE, Luke G, Mendoza H, ten Dam E, Gani D, et al. The‘cleavage’ activities of foot-and-mouth disease virus 2A site-directed mutantsand naturally occurring ‘2A-like’ sequences. J Gen Virol 2001;82(5):1027–41.
[30] Zhu M, Xu X, Liu H, Liu X, Wang S, Dong F, et al. Enhancement of DNAvaccine potency against herpes simplex virus 1 by co-administration of aninterleukin-18 expression plasmid as a genetic adjuvant. J Med Microbiol2003;52(3):223–8.
[31] Mosmann T. Rapid colorimetric assay for cellular growth and survival:application to proliferation and cytotoxicity assays. J Immunol Methods1983;65:55–63.
[32] Seder RA, Gurunathan S. DNA vaccines—designer vaccines for the 21st century.N Engl J Med 1999;341(4):277–8.

Author's personal copy
5160 A. Dey et al. / Vaccine 27 (2009) 5152–5160
[33] Abdelnoor AM. Plasmid DNA vaccines. Curr Drug Targets Immune EndocrMetabol Disord 2001;1(1):79–92.
[34] Wang R, Doolan DL, Le TP, Hedstrom RC, Coonan KM, Charoenvit Y, et al. Induc-tion of antigen-specific cytotoxic T lymphocytes in humans by a malaria DNAvaccine. Science 1998;282(5388):476–80.
[35] Sukumaran B, Tewary P, Saxena S, Madhubala R. Vaccination with DNA encodingORFF antigen confers protective immunity in mice infected with Leishmaniadonovani. Vaccine 2003;21:1292–9.
[36] Basu R, Bhaumik S, Basu JM, Naskar K, De T, Roy S. Kinetoplastid membraneprotein-11 DNA vaccination induces complete protection against both pentava-lent antimonial-sensitive and -resistant strains of Leishmania donovani thatcorrelates with inducible nitric oxide synthase activity and IL-4 generation:evidence for mixed Th1- and Th2-like responses in visceral leishmaniasis. JImmunol 2005;174:7160–71.
[37] Khera A, Singh R, Shakila H, Rao V, Dhar N, Narayanan PR, et al. Elicitation of effi-cient, protective immune responses by using DNA vaccines against tuberculosis.Vaccine 2005;23:5655–65.
[38] Hasan UA, Abai AM, Harper DR, Wren BW, Morrow WJ. Nucleic acid immu-nization: concepts and techniques associated with third generation vaccines. JImmunol Methods 1999;229:1–22.
[39] McDonell WM, Askari FK. DNA vaccines. N Engl J Med 1996;334(1):42–5.[40] de Felipe P. Skipping the co-expression problem: the new 2A “CHYSEL” tech-
nology. Genet Vaccines Ther 2004;2(13), doi:10.1186/1479-0556-2-13.[41] Lyadova IV, Vordermeier HM, Eruslanov EB, Khaidukov SV, Apt AS, Hewinson
RG. Intranasal BCG vaccination protects BALB/c mice against virulent Mycobac-terium bovis and accelerates production of IFN-� in their lungs. Clin ExpImmunol 2001;126:274–9.
[42] Aguilar-Be I, da Silva Zardo R, Paraguai de Souza E, Borja-Cabrera GP, Rosado-Vallado M, Mut-Martin M, et al. Cross-protective efficacy of a prophylacticLeishmania donovani DNA vaccine against visceral and cutaneous murine leish-maniasis. Infect Immun 2005;73(2):812–9.
[43] Sharma P, Kumar A, Batni S, Chauhan VS. Co-dominant and reciprocal T-helpercell activity of epitopic sequences and formation of junctional B-cell determi-nants in synthetic T:B chimeric immunogens. Vaccine 1993;11(13):1321–6.
Copyright © 2022 FDOKUMEN




















