
70-3-525.pdf - Oxford Academic
15
RESEARCH ARTICLE Vertical distribution of metabolically active eukaryotes in the water column and sediments of the Black Sea Marco J.L. Coolen 1 & Galina Shtereva 2 1 Marine Chemistry and Geochemistry Department, Woods Hole Oceanographic Institution, Woods Hole, MA, USA; and 2 Bulgarian Academy of Sciences, Institute of Oceanology (IO-BAS), Varna, Bulgaria Correspondence: Marco J.L. Coolen, Marine Chemistry and Geochemistry Department, Woods Hole Oceanographic Institution, 360 Woods Hole Road, Woods Hole, MA 02543, USA. Tel.: 11 508 289 2931; fax: 11 508 457 2164; e-mail: [email protected] Received 8 January 2009; revised 28 July 2009; accepted 29 July 2009. Final version published online 1 September 2009. DOI:10.1111/j.1574-6941.2009.00756.x Editor: Riks Laanbroek Keywords Black Sea; suboxic; sulfidic; 18S rRNA gene transcripts; DGGE; active eukaryotes. Abstract Recent DNA-based phylogenetic studies have reported high eukaryotal diversities in a wide range of settings including samples obtained from anoxic environments. However, parallel RNA-based surveys are required in order to verify whether the species detected are in fact metabolically active in such extreme environments. The Black Sea is the World’s largest anoxic basin but remains undersampled with respect to molecular eukaryotic diversity studies. Here, we report the distribution of active eukaryotes (18S rRNA-based survey) along a vertical nutrient and redox gradient in the water column and surface sediments of the Black Sea. A wide variety of eukaryotes were active in suboxic deep waters. Notably, certain species were active but escaped detection during a parallel 18S rDNA survey. The 18S rDNA survey from surface sediments yielded taxa of pelagic origin but none of these were identified from the water column at the time of sampling. Our data also indicate that gene transcripts do not always provide unequivocal proof that active microorganisms are indigenous to a specific position in an environ- mental gradient, because certain zoo- and phytoplankton species were still viable with detectable 18S rRNA in up to 300-year-old sulfidic sediments that underlie 830 m of sulfidic waters. Introduction Over the past several years, culture-independent molecular surveys using small subunit (SSU) rDNA sequences as phylogenetic markers have increasingly been used to study the environmental diversity of microbial eukaryotes. Pio- neering studies have revealed an unexpectedly high diversity of microbial eukaryotes in the deep sea (Lopez-Garcia et al., 2000) and in surface waters at different oceanic settings (D´ ıez et al., 2001; Moon-van der Staay et al., 2001). Some of the unique phylotypes from these studies represent novel lineages within known eukaryotic groups, such as alveolates and stramenopiles. In addition, 18S rDNA surveys have also been used to study microbial eukaryotic diversity in extreme environments, such as deep-sea hydrothermal vents (Edg- comb et al., 2002; Lopez-Garcia et al., 2003), acidic and iron-rich rivers (Zettler et al., 2002; Zettler et al., 2007), and various suboxic or anoxic marine settings, including marine and lacustrine surface sediments (Dawson & Pace, 2002), salt marsh environments (Stoeck et al., 2003), submarine caldera around fumaroles (Takishita et al., 2005), deep-sea methane cold-seep sediments (Takishita et al., 2007), strati- fied marine basins (Stoeck et al., 2003), and fjords (Behnke et al., 2006; Zuendorf et al., 2006). The latter studies revealed many novel phylotypes at high-taxonomic levels, although it has been argued that some of these phylotypes could represent artifacts such as chimeric sequences (e.g. Behnke et al., 2006). Another issue is that such DNA-based commu- nity analyses detect microorganisms irrespective of their viability or metabolic activity (Stoeck et al., 2007). The ability to identify living or active microbial eukaryotes is particularly important for settings where chemical environ- ments and microbial populations can change on a very small spatial scale (Stoeck et al., 2007), such as along the redox gradient in stratified, oxygen-depleted environments (Stoeck et al., 2003; Behnke et al., 2006). The identification of active microbial populations within environmental samples can be addressed by targeting SSU rRNA gene transcripts directly instead of DNA. This approach has mainly been used to identify active FEMS Microbiol Ecol 70 (2009) 525–539 c 2009 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved MICROBIOLOGY ECOLOGY Downloaded from https://academic.oup.com/femsec/article/70/3/525/533217 by guest on 14 July 2022
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of 70-3-525.pdf - Oxford Academic

R E S E A R C H A R T I C L E
Vertical distributionofmetabolicallyactive eukaryotes in thewatercolumnand sedimentsoftheBlackSeaMarco J.L. Coolen1 & Galina Shtereva2
1Marine Chemistry and Geochemistry Department, Woods Hole Oceanographic Institution, Woods Hole, MA, USA; and 2Bulgarian Academy of
Sciences, Institute of Oceanology (IO-BAS), Varna, Bulgaria
Correspondence: Marco J.L. Coolen, Marine
Chemistry and Geochemistry Department,
Woods Hole Oceanographic Institution, 360
Woods Hole Road, Woods Hole, MA 02543,
USA. Tel.: 11 508 289 2931; fax: 11 508 457
2164; e-mail: [email protected]
Received 8 January 2009; revised 28 July 2009;
accepted 29 July 2009.
Final version published online 1 September
2009.
DOI:10.1111/j.1574-6941.2009.00756.x
Editor: Riks Laanbroek
Keywords
Black Sea; suboxic; sulfidic; 18S rRNA gene
transcripts; DGGE; active eukaryotes.
Abstract
Recent DNA-based phylogenetic studies have reported high eukaryotal diversities
in a wide range of settings including samples obtained from anoxic environments.
However, parallel RNA-based surveys are required in order to verify whether the
species detected are in fact metabolically active in such extreme environments. The
Black Sea is the World’s largest anoxic basin but remains undersampled with
respect to molecular eukaryotic diversity studies. Here, we report the distribution
of active eukaryotes (18S rRNA-based survey) along a vertical nutrient and redox
gradient in the water column and surface sediments of the Black Sea. A wide
variety of eukaryotes were active in suboxic deep waters. Notably, certain species
were active but escaped detection during a parallel 18S rDNA survey. The 18S
rDNA survey from surface sediments yielded taxa of pelagic origin but none of
these were identified from the water column at the time of sampling. Our data
also indicate that gene transcripts do not always provide unequivocal proof
that active microorganisms are indigenous to a specific position in an environ-
mental gradient, because certain zoo- and phytoplankton species were still viable
with detectable 18S rRNA in up to 300-year-old sulfidic sediments that
underlie �830 m of sulfidic waters.
Introduction
Over the past several years, culture-independent molecular
surveys using small subunit (SSU) rDNA sequences as
phylogenetic markers have increasingly been used to study
the environmental diversity of microbial eukaryotes. Pio-
neering studies have revealed an unexpectedly high diversity
of microbial eukaryotes in the deep sea (Lopez-Garcia et al.,
2000) and in surface waters at different oceanic settings
(Dıez et al., 2001; Moon-van der Staay et al., 2001). Some of
the unique phylotypes from these studies represent novel
lineages within known eukaryotic groups, such as alveolates
and stramenopiles. In addition, 18S rDNA surveys have also
been used to study microbial eukaryotic diversity in extreme
environments, such as deep-sea hydrothermal vents (Edg-
comb et al., 2002; Lopez-Garcia et al., 2003), acidic and
iron-rich rivers (Zettler et al., 2002; Zettler et al., 2007), and
various suboxic or anoxic marine settings, including marine
and lacustrine surface sediments (Dawson & Pace, 2002),
salt marsh environments (Stoeck et al., 2003), submarine
caldera around fumaroles (Takishita et al., 2005), deep-sea
methane cold-seep sediments (Takishita et al., 2007), strati-
fied marine basins (Stoeck et al., 2003), and fjords (Behnke
et al., 2006; Zuendorf et al., 2006). The latter studies revealed
many novel phylotypes at high-taxonomic levels, although it
has been argued that some of these phylotypes could
represent artifacts such as chimeric sequences (e.g. Behnke
et al., 2006). Another issue is that such DNA-based commu-
nity analyses detect microorganisms irrespective of their
viability or metabolic activity (Stoeck et al., 2007). The
ability to identify living or active microbial eukaryotes is
particularly important for settings where chemical environ-
ments and microbial populations can change on a very small
spatial scale (Stoeck et al., 2007), such as along the redox
gradient in stratified, oxygen-depleted environments
(Stoeck et al., 2003; Behnke et al., 2006).
The identification of active microbial populations within
environmental samples can be addressed by targeting SSU
rRNA gene transcripts directly instead of DNA. This
approach has mainly been used to identify active
FEMS Microbiol Ecol 70 (2009) 525–539 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
MIC
ROBI
OLO
GY
EC
OLO
GY
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

prokaryotes in environmental samples (MacGregor et al.,
2001; Norris et al., 2002; Mills et al., 2004) and only recently
to detect active microbial eukaryotes (Coyne & Cary, 2005;
Stoeck et al., 2007; Alexander et al., 2009). rRNA is an
essential structural component of the ribosome, which is the
organelle responsible for protein synthesis in all prokaryotes
and eukaryotes. The number of cellular ribosomes, and thus
also the rRNA content, increases with growth rate and
decreases with starvation (Wagner, 1994; Muttray & Mohn,
1998; Buckley & Szmant, 2004; Chicharo & Chicharo, 2008;
Hawkins et al., 2008). Therefore, rRNA extracted from
environmental samples serves as a phylogenetic marker for
the identification and relative abundance of metabolically
active members within complex microbial communities.
Stoeck et al. (2007) constructed rRNA- and rDNA-based
clone libraries derived from an anoxic water sample from
the Framvaren Fjord (Norway) to compare the diversity and
relative abundance of active vs. inactive marine microeukar-
yotes. This study revealed 84 unique phylotypes of which
27% occurred in both libraries, 25% occurred exclusively in
the rRNA library and 48% occurred exclusively in the rDNA
library. Furthermore, phylotypes of phototrophic dinofla-
gellates, uncultured marine alveolates group I, and various
parasites were exclusively detected in the rDNA library and
represented nonindigenous members of the anoxic micro-
eukaryote community below the sulfidic chemocline
(Stoeck et al., 2007). Even though the analyzed clone library
was extensive (600 clones) and revealed many unique
phylotypes, only one depth was analyzed in the above study.
In addition, although the community structure below the
chemocline in stratified waters is likely to remain stable as
long as the stratification is not disturbed, pelagic samples
provide only a snapshot of photic zone-derived cells in
transit to the sediment (Coolen et al., 2007). Benthic rRNA
libraries in these settings would provide a more complete
picture of the capacity of organisms stemming from the
oxygenated part of the photic zone to remain active or viable
in suboxic or fully anoxic environments for substantial
periods of time. For example, another recent study has
shown that viable dinoflagellate cysts in anoxic Delaware’s
Inland Bays surface sediments could be identified using
PCR-based approaches from 18S rRNA libraries and even
from much more labile mRNA libraries (Coyne & Cary,
2005). This suggests that such gene transcripts could remain
detectable in resting stages of planktonic species not indi-
genous to anoxic water layers for substantial periods of time.
The Black Sea, the world’s largest permanently stratified
basin, is devoid of oxygen and contains abundant sulfide
from a depth of about 100 m to the seafloor at 2200 m. A
20–30-m-deep suboxic layer, depleted in both O2 and
sulfide, overlies the sulfide zone (Jørgensen et al., 1991).
This stable, vertically expanded redox gradient provides an
opportunity to study the distribution of microbial commu-
nities within different chemical environments, but thus far
remains undersampled with respect to eukaryotic diversity.
In the present study, we performed a high spatial resolution
analysis of the eukaryotic diversity (18S rDNA survey) and
sought to determine whether these organisms were metabo-
lically active (18S rRNA-based library) along a vertical
gradient of O2, H2S, and nutrients in the western Black Sea.
In addition, we performed a parallel survey on underlying
surface sediments to identify active benthic eukaryotes and,
more importantly, to establish whether eukaryotes derived
from the photic zones remain viable in this highly sulfidic
depositional setting.
Materials and methods
Water column chemistry and physical conditions
A Conductivity-Temperature-Depth profiler (CTD) was
deployed to measure conductivity, temperature, salinity,
fluorescence, dissolved oxygen (SBE-43 oxygen sensor; Sea-
Bird Electronics, Bellevue, WA) and to collect water to
measure N-species (nitrite, nitrate, ammonia), phosphate,
and sulfide. This effort revealed the exact depths of the
chlorophyll maximum, the upper and lower nitrite peaks,
the suboxic zone with o 0.2 mL L�1 of oxygen, as well as the
sulfidic chemocline. This information was used to select the
depths for the collection of particulate organic matter
(POM) for the molecular survey.
Nutrients (nitrite, nitrate, ammonia, and phosphate)
were measured spectrophotometrically according to Grassh-
off et al. (1983). Hydrogen sulfide was measured photome-
trically according to Cline (1969).
Sampling
Water samples were collected from 19 depths in the stratified
Black Sea (42146.56900N : 28140.64700E) aboard the R/VAkade-
mik (Institute of Oceanology, Bulgarian Academy of Sciences;
IO-BAS) using a SBE911 plus CTD (Sea-Bird Electronics)
equipped with twelve 5-L Niskin bottles. These depths in-
cluded the chlorophyll maximum at 22, 40, 60–115 m (5-m
intervals along the oxygenated mixolimnion), 120–135 m
(5-m intervals within the suboxic zone with o 0.2 mL L�1
oxygen), and within the sulfidic chemocline (150 and 160 m).
We took precautions to prevent contact of the oxygen-
depleted and sulfidic waters with the atmosphere by keeping
the water inside the Niskin bottle until cells in 3–5 L of water
were collected onto 142-mm, 0.2-mm pore size, polycarbonate
filters (Millipore, Billerica, MA). The water was directly
filtered from the Niskin bottles via a closed in-line tubing
system connected to the lower outlet of each Niskin
bottle under gentle pressure (o 50 mL min�1) provided by a
peristaltic pump (Masterflex, Cole and Palmer, Vernon Hills,
IL). The total water depth at this location was 971 m.
FEMS Microbiol Ecol 70 (2009) 525–539c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
526 M.J.L. Coolen & G. Shtereva
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

Fifty-centimeter-long undisturbed cores of laminated
sediments (Fig. 1b) were obtained at the same location
using a MC800 multicorer (Ocean Instruments, San Diego,
CA). One subcore (Core MC06) was subsectioned in 2-cm
intervals aboard ship and sections from the upper 8 cm of
undisturbed laminated sediments, including the top fluffy
layer, were used for subsequent extraction of nucleic acids as
described below.
Simultaneous DNA and RNA extraction andcDNA synthesis
POM
Directly after filtration, the filters were placed in a 15-mL
sterile conical centrifuge tube with a mixture of 4-mL
denaturing buffer (Hurt et al., 2001), 50mL 2-mercapto-
ethanol, and 2 mL of nucleases-free low-binding zirconium
beads with a diameter of 200mm (OPS Diagnostics, Leba-
non, NJ). Cells on the filter were mechanically disrupted to
release the nucleic acids by bead beating for 5 min at
maximum speed using the Vortex Genie 2s Vortex and
adapter (MoBio, Carlsbad, CA). The presence of the dena-
turing buffer and 2-mercaptoethanol ensured instant inacti-
vation of the nucleases present. Then the mixtures were
immediately stored at � 80 1C or colder until further
analysis. At the Woods Hole Oceanographic Institution
(WHOI), all steps involved in the extraction and purifica-
tion of nucleic acids were performed inside a HEPA-filtered
fume hood, which was UV-sterilized and cleaned with
Eliminase (Decon Laboratories, King of Prussia, PA) to
destroy nucleases before use. Additional precautions, such
as using filter tips during pipetting, using only RNAse-free
reagents and disposable plastic ware were also taken. Vinyl
gloves were worn at all times and cleaned in between steps
with Eliminase. Then, each deeply frozen filter/buffer/bead
mixture was transferred to precombusted (225 1C for 8 h)
and deeply frozen (� 80 1C) mortars and ground with a cold
pestle upon the thawing of the mixture. Circular movements
during grinding were omitted to minimize shearing of the
nucleic acids. The mixture was transferred into 50-mL-
sterile conical centrifuge tubes and an equal volume
(�4 mL) of phenol : chloroform : isoamylalcohol (PCI),
25 : 24 : 1, pH 8 (Ambion, Austin, TX) was added. After
vigorously vortexing, the mixture was placed on ice for
15 min to ensure the complete disassociation of nucleopro-
tein complexes, and centrifuged at 4 1C for 15 min at
10 000 g. The aqueous phase was transferred to a new sterile
tube and the nucleic acids were precipitated with
0.1 volume of 5 M NaCL (Ambion) and 1 volume of
isopropanol. After centrifugation at 10 000 g for 20 min at
room temperature, the air-dried pellet was dissolved in RNA
Storage Solution (Ambion).
Surface sediments
Total nucleic acids were extracted at 2-cm intervals from the
top 8 cm of sediment. Directly after sectioning, 1.5 mL of
sediment was transferred to 5-mL cyrovials and mixed with
equal volume of denaturing buffer (Hurt et al., 2001), 20 mL
of 2-mercaptoethanol, and 0.75 mL of zirconium beads.
Cells in the sediment were then immediately mechanically
disrupted to release the nucleic acids and to inactivate the
nucleases by bead beating as described above. The cyrovials
were stored in liquid nitrogen on board the R/V Akademik,
Fig. 1. (a) Sampling station in the western Black Sea off the coast of Varna, Bulgaria during cruise BS/AK06 (August 28–September 4, 2006) on the R/V
Akademik. All data presented in this report were obtained from filtered POM and multicore MC06 at Station 5 (Sta 5, 971 m). (b) MC06 with
undisturbed laminated anoxic top sediments.
FEMS Microbiol Ecol 70 (2009) 525–539 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
527Viable and active eukaryotes in the Black Sea
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

transferred on dry ice to WHOI, and stored at � 80 1C until
further analysis. At WHOI, the frozen mixture underwent
the same grinding procedure in sterile and deeply frozen
mortars as described above, but followed an additional
extraction step with 8 mL of extraction buffer as described
by Hurt et al. (2001). Following centrifugation (10 min,
10 000 g), total nucleic acids were extracted from the super-
natant using PCI and the additional steps described above.
The quality of the RNA and DNA extracts was verified by
agarose gel electrophoresis. The total nucleic acid extracts
from the POM and sediments were used for the 18S rDNA
survey.
Preparation of DNA-free RNA andcDNA synthesis
For the 18S rRNA survey, 0.1 volume of the extracted total
nucleic acids was treated for 30 min at 37 1C with 1 U DNAse
and 1� buffer using the DNA-freeTM kit (Ambion) followed
by inactivation of the DNAse using 0.1 volume of the
provided DNAse inactivation reagent. Before PCR, DNA-
free RNA samples were randomly transcribed into cDNA
using the iScript cDNA synthesis kit (Bio-Rad, Hercules,
CA). The 20 mL reaction mixtures contained RNAse-free
water, 1 U iScript reverse transcriptase, 1� iScript Reaction
Mix, 50 mg bovine serum albumin (BSA; Ambion), and
0.1 volume of DNA-free RNA sample. The reverse transcrip-
tase (RT) reactions were performed in a Mastercycler
(Eppendorf, Westbury, NY) and included incubation for
5 min at 25 1C, 30 min at 42 1C, and 5 min at 85 1C. The
latter cDNA served as template for PCR to study the diversity
of the metabolically active eukaryotes in the Black Sea.
PCR amplification of eukaryote 18S rDNA andreverse transcribed 18S rRNA
Each amplification reaction contained 0.25 mM of each
deoxynucleotide (dNTP), 4.5 mM of MgCl2, 0.5� (from
10 000� concentrated stock) SYBRsGreen I (Invitrogen,
Carlsbad, CA), 50 mg of BSA (Ambion), 5mL of
10� PicoMaxxTM reaction buffer (Stratagene, LaJolla, CA),
2.5 U of DNA polymerase (PicoMaxxTM high fidelity PCR
system; Stratagene), 0.2mM of primers (Thermo-Fisher,
Ulm, Germany), and 10 ng of DNA or cDNA template.
Partial (560-bp long) eukaryote 18S rDNA (or reverse
transcribed 18S rRNA) was PCR amplified using primer set
Euk1A and Euk516r-GC (Set A) as described previously
(Dıez et al., 2001) The reaction mixtures were adjusted to a
final volume of 50 mL with nucleic acids and nuclease-free
water (Ambion). Each PCR amplification series included a
reaction without a DNA template, which served as a control
for contaminations during the pipetting of the reaction
mixture components. For each cDNA sample, a parallel
reaction with 0.2 vol% of digested RNA was subjected to
PCR as a control for the presence of traces of incomplete
digested DNA in the various RNA templates used for cDNA
synthesis.
All reactions were performed in a Realplex quantitative
PCR cycler (Eppendorf) and involved initial denaturing
(4 min at 95 1C), followed by 38 cycles including denaturing
(30 s at 94 1C), primer annealing (40 s at 64 1C), primer
extension (60 s at 72 1C), and imaging of newly formed
fluorescent (SYBRsGreen I labeled) double-stranded
DNA (80 1C for 20 s). All DNA and cDNA samples
were then subjected again to real-time quantitative PCR
but this time all reactions were stopped at the end of the
exponential phase (i.e. after 25–32 cycles). This prevented
overamplification of individual 18S rDNA fragments
to allow semi-quantitative analysis by denaturing gradient
gel electrophoresis (DGGE; Schafer & Muyzer, 2001) as
outlined below.
Phylogenetic analysis of sequenced DGGE bands
For the majority of the samples, 100 ng of PCR-amplified
partial eukaryote 18S rRNA and RT-PCR amplified 18S
rRNA (600 bp including the 40-bp-long GC clamp) was
separated by DGGE (Schafer & Muyzer, 2001). The poly-
acrylamide gels (6%, w/v) contained a denaturing gradient
of 20–50% (with 100% denaturant equaling 7 M urea and
40% formamide). Gels were run for 15 h at 5 V cm�1 and
60 1C using a PhorU2 system (Ingeny, Leiden, the Nether-
lands). Afterwards, the gels were stained for 20 min by
covering the gels twice with 10 mL of 1�TAE buffer
(pH 8.3), containing 2 mL undiluted SYBRsGold (Invitro-
gen) followed by destaining for 60 min in 1�TAE buffer
(pH 8.3). In order to prevent DNA damage by UV, we used a
Dark Reader (Clare Chemicals Research Inc., Dolores, CO),
which uses visible light instead of UV in order to visualize
the SYBRsGold-stained DNA. Digital gel images were made
using a Foto/Analysts Express System (Fotodyne, Hartland,
WI) and IMAGEJ software. TOTALLAB TL100 v2006 1D-gel
analysis software (Nonlinear Dynamics, Durham, NC) was
used to determine the pixel density and vertical position of
each band. This information was used to determine the
relative abundance of each band within a given sample and
to identify the exact vertical position of each band in order
to characterize unique vs. identical bands between samples
(e.g. Dıez et al., 2001). DGGE bands were sliced from the gel
with a sterile scalpel and the DNA of each sliced gel fragment
was eluted in 75 mL sterile 10 mM Tris-HCl (pH 8.0) by
incubation for 48 h at 2 1C. One microliter of the eluted 18S
rDNAs (approximately 107 copies) was reamplified using
25 cycles and the primer combinations listed above, but this
time without the GC clamp. These amplicons served as DNA
for the subsequent cycle sequencing reactions. PCR reagents
were as described above but without the addition of
FEMS Microbiol Ecol 70 (2009) 525–539c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
528 M.J.L. Coolen & G. Shtereva
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

SYBRsGreen I or extra MgCl2. All reamplification reactions
were performed in a Master cycler (Eppendorf) and in-
volved initial denaturing (4 min at 95 1C), followed by
25 cycles including denaturing (30 s at 94 1C), primer
annealing (40 s at 58 1C), and primer extension (12 s at
72 1C). A final extension step was set at 72 1C for 30 min.
The quality of each sequencing template was checked by
agarose gel electrophoresis. The DNA concentration of each
amplicon was measured fluorometrically using picogreen
staining (e.g. Coolen et al., 2006). Thirty nanograms of
reamplified DGGE bands were submitted to Agencourt
(Beverly, MA) for subsequent bidirectional cycle sequencing
with primers EukA and Euk516r.
Sequences have been analyzed using the ARB software
package (version December 2007) (Ludwig et al., 2004) and
the corresponding SILVA SSURef 96 database (Pruesse et al.,
2007). After importing, all sequences were automatically
aligned according to the SILVA SSU reference alignment.
Manual refinement of the alignment was carried out taking
into account the secondary structure information of the
rRNA. Then, the phylogenetic bootstrap trees (1000 replica-
tions) were first reconstructed based on 1200-bp-long avail-
able sequences of the closest relatives using the neighbor-
joining method (ARB). The shorter aligned environmental
18S rDNA sequences from this study were inserted after-
wards using the parsimony interactive tool implemented in
the ARB software package without changing overall tree
topology. Sequences obtained in this study have been
deposited in the NCBI sequence database under accession
numbers GQ402464–GQ402489.
Results and discussion
Physical parameters and location of the suboxiczone, nutrients, and onset of sulfide
The upper 160 m of the Black Sea are brackish with salinities
between 17.2 psu at the surface and 20.6 psu toward the
sulfidic chemocline (Fig. 2). The surface water temperature
was 22.5 1C at the time of sampling, with a steep drop to
15.3 1C at the thermocline at 25 m, followed by relatively
constant temperatures (8.1� 0.6 1C) between 31 and 160 m
(Fig. 2). Fluorescence data show that a chlorophyll max-
imum was present just above the thermocline at 22 m,
whereas chlorophyll concentrations dropped close to zero
values o 80 m (Fig. 2). A small increase in fluorescence was
observed at the top of the suboxic zone at 120 m.
The suboxic zone with o 0.2 mL L�1 (i.e. o 10mM)
oxygen and undetectable sulfide concentrations (o 0.2mM)
(Murray et al., 1989; Codispoti et al., 1991) at Station 5 was
found between 120 m (st = 15.62) and 139 m (st = 15.90). At
140 m sulfide became detectable, showing 0.28mM, and
increased to 6mM at 150 m (st = 16.00) and 12.5mM at
160 m (st = 16.10) (Fig. 2).
Ammonia concentrations dropped from 8.5mM at
�160 m to undetectable levels (o 0.2mM) at the base of
Fig. 2. Water-depth profiles with physical parameters [temperature, salinity, st values, and fluorescence (chlorophyll a concentration)] as well as
chemical parameters (oxygen and sulfide) (left panel), nutrients (N-species and phosphate) (second panel from the left), and relative quantitative
distributions of 18S rDNA (closed circles) vs. 18S rRNA (open circles) of ‘total’ vs. active pelagic copepod, tunicate, haptophyte, dinoflagellate, ciliate,
and unclassified species. The suboxic zone with o 0.2 mL L�1 of dissolved oxygen and undetectable sulfide is marked in dark gray. The upper nitrite
maximum in the oxygenated waters and the lower nitrite maximum in the suboxic zone are marked in light gray.
FEMS Microbiol Ecol 70 (2009) 525–539 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
529Viable and active eukaryotes in the Black Sea
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

the suboxic zone. A small nitrite maximum (0.15mM) was
found in the suboxic zone at 130 m (Fig. 2). At depths
between �100 and 120 m where oxygen concentrations
reached up to 5 mL L�1, a second (upper) nitrite maximum
occurred, which agrees well with previous findings (Ward &
Kilpatrick, 1990; Murray & Yala-del-Rio, 2006). A nitrate
peak with up to 4mM of nitrate was observed between �80
and 120 m. Phosphate concentrations were highest at the
bottom of the suboxic zone and in the sulfidic waters down
to 160 m (�4 mM) (Fig. 2).
Distribution of metabolically active eukaryotesalong the environmental gradients
Metabolically active eukaryotes were identified within
19 POM samples collected along a vertical gradient (salinity,
oxygen, hydrogen sulfide, and nutrients), and within the
upper 8 cm of sulfidic sediments that underlie 830 m of
sulfidic water at the sampling location in the Black Sea. The
active population was studied from reverse transcribed and
PCR-amplified (two-step RT-PCR) 18S rRNA whereas com-
bined molecular signatures of both metabolically active and
inactive populations were studied from PCR-amplified
(genomic) 18S rDNA (i.e. 18S rRNA- vs. 18S rDNA-based
libraries). All amplicons were subjected to DGGE (Fig. 3a).
Unfortunately, we lost the DNA extract from 80 m during
the purification procedure and, as a result, no 18S rDNA
amplicons could be generated from that sample (Fig. 3a).
The corresponding DNAse-treated 18S rRNA extract from
the 80 m sample could subsequently be analyzed by RT-PCR
(Fig. 3a). Total DNA and RNA was successfully extracted
from 115, 75, 135, and 160 m, but the samples from these
depths contained fewer 18S rDNA (i.e. 115a) or 18S rRNA
(75b, 135b, 160b) template for PCR and yielded less
amplicons for subsequent DGGE analysis as compared with
other samples within the environmental gradient (Fig. 3a).
Fifty-six numbered DGGE bands were excised from the
DGGE gels and 53 of the excised bands were successfully
sequenced. These sequences represented 18 unique phylo-
types (based on a 99% sequence similarity cutoff) (Fig. 4).
Fig. 3. DGGE with PCR-amplified 18S rDNA and reverse transcribed and PCR-amplified 18S rRNA (from cDNA pool) from (a) water column POM
samples and (b) sulfidic sediments from MC06. Fifty-six bands were excised from the gel and 53 were successfully sequenced and revealed 18 unique
phylotypes, denoted 1–18 and with different colors for each taxonomic group. Bands from different samples that melted at identical heights in the gel
and appeared to represent the same phylotype carry the same number (1–18). Unsuccessfully sequenced bands are indicated with ‘X’ and chimeric
sequences with ‘C’. Sampling depths (in meter for POM samples and in centimeter for the sulfidic sediments) are located above panels (a) and (b),
respectively. Lanes that contained 18S rRNA amplicons are denoted with ‘a’ and lanes that contain reverse-transcribed and PCR-amplified 18S rRNA are
denoted with ‘b’. PM, position marker in the gel, not a sample; gDNA, genomic DNA that contains the 18S rRNA genes; cDNA, cDNA from total pool of
reverse transcribed RNA. The total DNA from 80 m was lost during extraction and, for that reason, did not result in 18S rDNA amplicons (indicated with
red ‘x’ in 80 m lane ‘a’).
FEMS Microbiol Ecol 70 (2009) 525–539c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
530 M.J.L. Coolen & G. Shtereva
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

Fig. 4. Phylogenetic tree showing the
relationship of eukaryote 18S rDNA and
18S rRNA sequences retrieved from Black
Sea water column and surface sediments
to reference sequences obtained from the
NCBI database (http://www.ncbi.nlm.nih.
gov). The eukaryote sequences from POM
and MC06 samples were determined
from the DGGE represented in Fig. 3.
Phylotypes are indicated as, for example,
‘3_ BSA6S5Euk’, which stands for DGGE
band representing the unique eukaryotic
phylotype # 3 obtained from the Black Sea
with the RV Akademik in 2006, Station 5.
Bar indicates 0.1 fixed point mutations per
nucleotide. Numbers at nodes indicate
bootstrap values out of 1000 replications
for phylogenetic trees calculated by
Felsenstein correction and Parsimony
methods.
FEMS Microbiol Ecol 70 (2009) 525–539 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
531Viable and active eukaryotes in the Black Sea
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

The DGGE bands that represented unique phylotypes are
numbered 1–18 in the DGGE (Fig. 3a). These phylotypes
comprised copepods (four phylotypes; 12_BSA6S5Euk,
16–18), tunicates (phylotype 11), haptophytes (phylotype
1), dinoflagellates (eight phylotypes; 3, 6, 7, 8, 9, 10, 13, 14),
and ciliates (phylotypes; 2 and 4) as well as a sequence
(phylotype 15), which did not fall into established taxo-
nomic groups but which showed up to 96% sequence
similarity to 18S rDNA sequences recovered from the
sulfidic waters of Framvaren Fjord (Fig. 4, Table 1) (Behnke
et al., 2006).
Copepods
A copepod (phylotype 12) with 100% sequence similarity to
partial 18S rDNA of Pseudocalanus elongatus (Fig. 4, Table 1)
was identified from both 18S rDNA/rRNA libraries in the
oxygenated part of the mixolimnion (between 40 and 95 m)
(Fig. 3a) and represented up to 40% of the total reverse-
transcribed and PCR-amplified 18S rRNA pool (Fig. 2).
Pseudocalanus elongatus is known to be highly abundant
throughout the Black Sea (Niermann & Greve, 1997) and
makes up �40% of the zooplankton in autumn within the
offshore regions (Unal et al., 2006). It is a cold-water species
with an upper temperature limit of occurrence of 13 1C,
which would explain its absence in the chlorophyll max-
imum (Figs 2 and 3a) that was 22.2 1C at the time of
sampling.
The sequence of phylotype 16 was identical to that of both
Calanus pacificus as well as Calanus helgolandicus (Fig. 4,
Table 1) and was present at depth intervals throughout
the oxygenated part of the water column (Figs 2 and 3a).
Most likely, phylotype 16 represents the endemic Calanus
euxinus (Hulsemann) (Besiktepe et al., 1998) instead, for
which no 18S rDNA sequence is available. Both C. helgolan-
dicus and C. euxinus are morphologically similar and show
o 0.5% sequence differentiation in the faster evolving
mitochondrial cytochrome oxidase subunit I gene (mtCOI).
This recently raised the question of whether C. euxinus
is a different species (Unal et al., 2006). Calanus helgolandi-
cus is an epipelagic copepod and typically inhabits the saline
waters of world’s oceans (Fleminger & Hulsemann, 1977)
with salinity values of 32–39, whereas C. euxinus is a key
component of the Black Sea pelagic ecosystem (Besiktepe
et al., 1998) and has adapted to the lower salinity of the
Black Sea (�18 psu). Furthermore, the predominance of
phylotype 16 in both 18S rDNA/rRNA libraries within the
deeper waters, notably the suboxic zone, corroborates with
the lifestyle of C. euxinus in the Black Sea. Previous studies
have shown that C. euxinus descends during the day to the
cold suboxic zone thereby significantly decreasing its total
metabolism as well as enabling it to utilize the energy of
consumed food for growth and lipid accumulation (Sve-
tlichny et al., 2000; Svetlichny et al., 2006).
The copepod represented by phylotype 17 was metaboli-
cally active at most depths between 40 and 80 m, whereas the
closely related phylotype 18 was identified only from the
narrow interval within the oxygenated upper nitrite max-
imum between 105 and 110 m (Figs 2, 3a, and 4, Table 1).
The closest relative of both phylotypes was an unassigned
Fig. 4b. Continued.
FEMS Microbiol Ecol 70 (2009) 525–539c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
532 M.J.L. Coolen & G. Shtereva
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

Table 1. General information about the eukaryotic phylotypes recovered from the Black Sea
Phyl. no. Tax. grp Closest (named) relative(s)
Sequence
similarity (%) General observations
12 Cop Pseudocalanus elongatus (AY 446903) 100 Present and active between 40 and 95 m
16 Cop Calanus helgolandicus (AY446908) 100 Present throughout most analyzed depths but mainly
active between 105 and 150 m
Calanus pacificus (L81939) 100
17 Cop Clone of Baffin Bay (EU182744) 100 Present and active at most depths between 40 and
80 m
18 Cop Clone of Baffin Bay (EU182744) 98.5 Active at 105 m, just present at 110 m
11 Tun Oikopleura dioica (AY116613) 99 Active in the chlorophyll maximum and in the upper
nitrite maximum (between 110 and 115 m)
1 Hap Cruciplacolithus neohelis (AB058348) 96 Active at most analyzed depths between 40 and
115 m
9 Din Gyrodinium rubrum (AB120003) and clones
from the Sargasso Sea deep chlorophyll
maximum (AY664955/72)
96–97 Present and active between 40 and 95 m, absent
from chlorophyll-depleted deeper waters
6 Din Clone SL163B11 (AY664961) Ross Sea 99 Found only at 60 m (active)
3 Din Gyrodinium fusiforme (AB120002) 98.5 Present between 90 and 100 m (80 m DNA sample
missing); active between 80 and 95 m
7 Din Gyrodinium fusiforme (AB120002) 99 Present at 100 m; activity not confirmed
14 Din Clone BB01_39 (AY885030) Mid-Atlantic
Estuary (Barnegat Bay, New Jersey)
98 Active at the oxic/suboxic zone transition (120 m);
present but inactive at 125 m
10 Din Clone SCM37C45 (AY664929) 99 Present at the oxic/suboxic zone transition (120 m);
activity not confirmed
Sargasso Sea deep chlorophyll maximum
13 Din Clones from sulfidic waters Mariager Fjord
(DQ103845/46/60/71)
98–99.8 Present in suboxic zone at 125 m; activity not
confirmed
8 Din Clone SCM28C60 (AY664961) Sargasso Sea
deep chlorophyll maximum
98.5 Present and active in suboxic zone (120–130 m) and
sulfidic chemocline at 150 m in the presence of 6 mM
of sulfide
4 Cil Clones from sulfidic waters of Framvaren Fjord
(DQ310304/14)
99.9 Active in fully oxygenated part of the photic zone at
40 m; not detected from sulfidic Black Sea waters
2 Cil Strombidium styliferum (DQ631805) 98 Active at the oxic/suboxic zone transition (120 m);
present but inactive at 125 m
15 Unc Clone FV23_2E8 (DQ310250) 83 Present at 150 and 160 m; active at 150 m in the
presence of 6 mM of sulfide
27 Cop Tortanus sp. New Caledonia (AY626995) 90 Present and active in up to 200-year-old (upper 4 cm)
sulfidic sediments
24 Rot Mytilina mucronata (DQ297708) 98 Low 18S rDNA content but activity confirmed in up to
300-year-old (upper 6 cm) sulfidic sediments
22 Hap Emiliania huxleyi (AB183618) 98 Low 18S rDNA content but activity confirmed in
upper 2 cm of sulfidic sediments
23 Din Scrippsiella sp. MBIC11164 (AB183674) 100 Present in upper 8 cm; active in upper 6 cm
21 Din Pentapharsodinium sp. CCMP771 (AF274270) 99.8 Present between 2 and 6 cm; activity not confirmed
25 Cil Clone M3_18E07 (DQ103803) 93 Present between 2 and 4 cm; activity not confirmed
Sulfidic waters Mariager Fjord
20 Cil Clone BS_DGGE_Euk-6 (DQ234286); Black Sea
sulfidic Unit I sediment
100 Present between 2 and 8 cm; activity not confirmed
26 Cil Clone M3_18E07 (DQ103803) 96 Present between 4 and 8 cm; activity not confirmed
Sulfidic waters Mariager Fjord
19 Cil Clone BS_DGGE_Euk-1 (DQ234281); Black Sea
sulfidic sapropel Unit II
97 Present between 6 and 8 cm; activity not confirmed
Phyl. no., phylotype number depicted from Fig. 4; Tax. grp, taxonomic group; Cop, copepods; Tun, tunicates; Hap, haptophytes; Din, dinoflagellates;
Cil, ciliates; Unc, unclassified; Rot, rotifers. Light- and dark-gray shaded part of the table represent phylotypes recovered from, respectively, the water
column and the sulfidic sediments of the Black Sea.
FEMS Microbiol Ecol 70 (2009) 525–539 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
533Viable and active eukaryotes in the Black Sea
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

copepod found at a depth of 180 m in the Arctic Baffin Bay
(Fig. 4, Table 1) with low in situ temperatures (1 1C) and low
chlorophyll concentrations (Hamilton et al., 2008). A paral-
lel identification of these unassigned copepod species based
on morphological characteristics was no longer possible
because the filters were completely used up by the nucleic
acids extraction.
Tunicata
Phylotype 11 represented Oikopleura dioica, the only re-
corded gelatinous tunicate (appendicularian) in the Black
Sea (Shiganova, 2005) (Fig. 4, Table 1). Its 18S rRNA (DGGE
band 11, Fig. 3a) was mainly found in the chlorophyll
maximum at 22 m but a faint DGGE band 11 was also
present in the lower part of the upper nitrite maximum
(between 110 and 115 m) (Figs 2 and 3a). The predomi-
nance of its 18S rDNA at 22 m is in accordance with the
knowledge that most individuals of this euryhaline and
eurythermic tunicate are present in the thermocline,
whereas fewer individuals appear to be present in the deeper
oxygenated waters of the Black Sea (Shiganova, 2005 and
references therein). The coinciding presence of Oikopleura
18S rRNA implies that viable Oikopleura individuals are
present in the chlorophyll maximum as well as in the deeper
oxygenated waters of the Black Sea (Figs 2 and 3a).
Oikopleura dioica populations grow exceptionally fast (Seo
et al., 2001) and this species ingests phytoplankton (5%) but
its main food is bacteria and detritus (95%) (Shiganova,
2005). The specialized filter apparatus of O. dioica retains
particles no larger than 15 mm and this feature allows feeding
in deep, detritus-based ecosystems. This utilization of small
size classes decreases competition with other zooplankton
species (Tiselius et al., 2003). The presence of viable
O. dioica in the nitrite maximum just above the suboxic
zone with low chlorophyll levels (Figs 2 and 3a) suggests that
O. dioica could be actively involved in filtering mainly
bacteria and/or detritus at this depth.
Haptophytes
Our molecular survey revealed a previously overlooked
haptophyte (i.e. phylotype 1), which is affiliated with
calcifying haptophytes and Cruciplacolithus neohelis as its
closest named relative (Fig. 4, Table 1). Whereas 18S rRNA
of this active haptophyte represented up to 30% of the total
RT-PCR-amplified 18S rRNA pool (Fig. 2), the correspond-
ing genomic 18S rDNA of this species was often below
detection on DGGE (Fig. 3a) and most likely also escaped
prior identification from an 18S rDNA library constructed
for additional locations in the Black Sea (Coolen et al.,
2006). This discrepancy can be caused by the fact that DGGE
is insensitive to less dominant species (e.g. Coolen et al.,
2007), or those with low genomic rRNA gene copy numbers.
The presence of active cells of this haptophyte down to
115 m could point to a capacity for heterotrophic growth,
given the extremely low light intensity and narrow light
spectrum supporting photosynthetic activity at this depth
(Overmann et al., 1992). Many haptophytes from early
diverging, noncalcifying lineages (Prymnesiales, Phaeocys-
tales) have retained heterotrophic behavior such as preda-
tion on bacteria and small algae (i.e. phagotrophy) (Kawachi
et al., 1991; Tillmann, 1998). Phagotrophy is restricted to
motile haptophyte cells with an emerging prey-catching
haptonema (Kawachi et al., 1991; Jones et al., 1994;
Tillmann, 1998). Also the motile haploid flagellate phase of
certain calcifying haptophytes were found to be phago-
trophic (Houdan et al., 2006). A phagotrophic life style for
the haptophyte related to calcifying species is thus plausible
and could be verified as soon as a culture from the Black
Sea’s deep suboxic zone would become available.
The widespread marine bloom-forming calcareous hap-
tophyte Emiliania huxleyi, which is also common in the
Black Sea (Eker-Develi & Kideys, 2003), was not identified in
our molecular surveys. A test with parallel tag-encoded FLX
amplicon pyrosequencing (e.g. Sogin et al., 2006) of a
130-bp-long fragment of the V9 region revealed that 18S
rDNA of haptophytes of the order Isochrysidales to which E.
huxleyi belongs, represented only 2% of the total eukaryotic
18S rDNA in the chlorophyll maximum at 22 m (M.J.L.
Coolen & C. Davis, unpublished data). As outlined earlier,
DGGE is biased to identify only phylotypes that represent
4�2% of the total pool of PCR-amplified DNA (e.g. Coolen
et al., 2007), and would explain why E. huxleyi was missed in
our general eukaryotic 18S rDNA PCR/DGGE approach.
Dinoflagellates
Our sampling set revealed eight unique dinoflagellate phy-
lotypes, which were found to exhibit high vertical stratifica-
tion, and their presence often coincided with the presence of
specific nitrogen species or suboxic/anoxic conditions. For
example, the dinoflagellates represented by phylotypes 9 and
3 were present and active at specific depths between 40 and
95 m (Figs 2 and 3a), and were closely affiliated with
uncultivated dinoflagellates from the deep chlorophyll max-
imum in the Sargasso Sea and/or phagotrophic Gyrodinium
spp. (Fig. 4, Table 1).
In addition, nucleic acids of some unassigned dinoflagel-
lates were found only in the suboxic or anoxic waters of the
Black Sea. The dinoflagellates represented by phylotypes 10
and 13 (Fig. 4, Table 1) were rare in the water column and
only their 18S rDNA was identified from the upper part of
the suboxic zone (Figs 2 and 3a). Therefore, their activity or
their indigeneity to suboxic waters could not be confirmed.
FEMS Microbiol Ecol 70 (2009) 525–539c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
534 M.J.L. Coolen & G. Shtereva
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022
On the other hand, phylotype 14 (Fig. 4, Table 1) was clearly
active at the oxic/suboxic transition at 120 in the presence of
3 mM of oxygen (Figs 2 and 3a). The DNA of this dino-
flagellate was most abundant at 125 m, whereas its RNA was
below the detection limit at this depth, underlying the value
of comparative analysis of gene transcripts for verification of
metabolic activity of microbiota within environmental gra-
dients. Phylotype 8, related to a clone from the Sargasso
Sea deep chlorophyll maximum, represented the only dino-
flagellate to be active in the deeper part of the suboxic zone
with undetectable levels of oxygen, as well as at 150 m in the
presence of 6mM of hydrogen sulfide (Fig. 2).
The sulfidic chemocline furthermore contained tran-
scripts of the 18S rRNA gene of an unclassified eukaryote
(i.e. phylotype 15) with up to 96% sequence similarity to
18S rDNA sequences recovered from sulfidic waters of
Framvaren Fjord (Fig. 4, Table 1) (Behnke et al., 2006). This
phylotype was not detected from the oxygenated waters
above, but a more sensitive PCR approach using species-
specific primers would be required to verify whether the
presence of this eukaryote was restricted to the slightly
sulfidic part of the stratified water column of the Black Sea.
Ciliates
Information about the species composition of ciliates in the
Black Sea is sparse and mostly relates to coastal areas
(Kovaleva & Golemanski, 1979; Bouvier et al., 1998). Our
survey revealed only two unique phylotypes of ciliates
throughout the upper 160 m of the water column. Phylotype
4 (Fig. 4, Table 1) is closely related to ciliates of the class
Oligohymenophorea and was found in the RNA-based (me-
tabolically active) library from fully oxygenated waters at a
depth of 40 m (Figs 2 and 3a). Environmental sequences
from DNA-based libraries of highly sulfidic Framvaren
Fjord waters (Behnke et al., 2006) were the closest relatives
of phylotype 4 (Fig. 4).
A ciliate with Strombidium styliferum as its closest relative
(phylotype 2) (Fig. 4, Table 1) was only found to be
metabolically active at the oxic/suboxic zone transition at
120 m and just present at 125 m. However, more precise
genotyping would be required to identify this ciliate because
a total of 63 morphotypes have been described from this
genus (Agatha, 2004) and the estimates of genetic diversity is
much greater (Song & Packroff, 1997).
Eukaryotes in the Black Sea’s sulfidic sediments
Nine out of 17 successfully sequenced DGGE bands recov-
ered from the upper 8 cm of sulfidic sediments that underlie
830 m of sulfidic waters represented unique phylotypes and
grouped with copepods (phylotype 27), rotifers (phylotype
24), haptophytes (phylotype 22), dinoflagellates (phylotypes
21 and 23), and ciliates (four phylotypes; 19, 20, 25, 26)
(Fig. 4, Table 1). Figure 3b displays the position of the
sequenced DGGE bands corresponding to these phylotypes.
The phylogenetic affiliation with other eukaryotes, their
relative abundance, and whether they represent species
indigenous to the sediments or allochthonous species de-
rived from the water column above, will be discussed next.
Copepods
The upper 4 cm of sediment contained both DNA and RNA
from a copepod distantly related to Tortanus sp. New
Caledonia (phylotype 27; Fig. 4, Table 1). In the surface
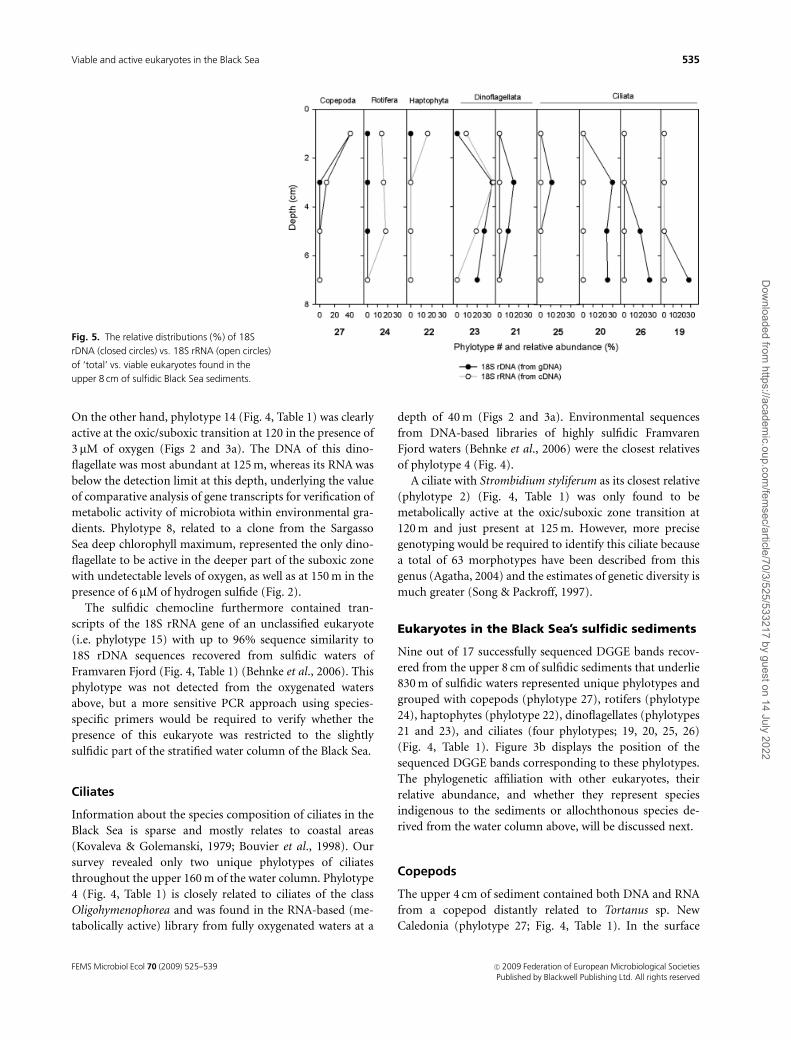
Fig. 5. The relative distributions (%) of 18S
rDNA (closed circles) vs. 18S rRNA (open circles)
of ‘total’ vs. viable eukaryotes found in the
upper 8 cm of sulfidic Black Sea sediments.
FEMS Microbiol Ecol 70 (2009) 525–539 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
535Viable and active eukaryotes in the Black Sea
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

sediment (0–2 cm), 18S rRNA of this copepod represented
40% of the total RT-PCR-amplified 18S rRNA pool (Fig. 5).
Most likely, the 18S rRNA of this Tortanus-related copepod
was extracted from resting eggs derived from the overlying
oxygenated water column. Assuming an absence of a post-
depositional redistribution of the exquisitely laminated
sulfidic sediments (Fig. 1b) and an average sedimentation
rate of 0.2 mm year�1 (M.J.L. Coolen, unpublished data), the
sediment interval at 4-cm depth with detectable Tortanus
18S rRNA gene transcripts would correspond to a deposi-
tional age of 200 years before present. This finding indicates
that copepod eggs (depending on the species) could remain
viable with detectable levels of 18S rRNA gene transcripts in
highly sulfidic environments for up to 200 years after
sedimentation. This is significantly longer than the previous
reported recovery of viable copepod eggs from up to
40-year-old sulfidic sediments from the stratified Petta-
quamscutt River Basin (RI) in the presence of 2 mM of
hydrogen sulfide (Marcus et al., 1994), which is approxi-
mately seven times the concentration of sulfide in the Black
Sea surface sediments (Zopfi et al., 2004). Because the
primary targets of sulfide are aerobic respiration enzymes,
for example cytochrome c oxidase, it was suggested that
copepod eggs may gain protection from sulfide by under-
going anaerobic respiration (Marcus et al., 1997).
Rotifers
The rotifer Mytilina mucronata was the closest relative of
phylotype 24 (Fig. 4). Its 18S rRNA, probably extracted from
viable resting eggs, comprised up to 20% of the total 18S
rRNA pool within up to 300-year-old (i.e. the upper 6 cm)
sulfidic sediments (Figs 3b and 5). 18S rDNA of the rotifer
was also identified but resulted only in a faint DGGE band
(band 24; Fig. 3b) at 2–4 cm, which was also the sample with
the most intense corresponding band with RT-PCR ampli-
fied 18S rRNA. Our data suggest that rotifer eggs in the
Black Sea sediments remained longer viable than copepod
eggs because the copepod 18S rRNA rapidly declined and
reached undetectable levels o 4 cm (i.e. �200-year-old
sediments; Fig. 5). This is in agreement with the results from
Marcus et al. (1994) who found that a higher percentage of
eggs from a rotifer of the genus Brachionus could be hatched
from up to 40-year-old sulfidic Pettaquamscutt River Basin
sediments as compared with copepod eggs.
Haptophytes
Interestingly, a unique haptophyte (phylotype 22; Figs 3b
and 4, Table 1) with E. huxleyi as its closest relative was
found to be active in the top 2 cm of sediment, whereas its
DNA was too faint to be detected by DGGE. This haptophyte
was not detected from the water column at the time of
sampling. These results imply that this haptophyte has
developed a strategy to remain metabolically active for
substantial periods of time (at least weeks and up to
decades) in the presence of �175 mM of sulfide. Previously,
it was shown that haptophytes were among various protists
that were found to proliferate from incubated Swedish
coastal sediments (Persson, 2002) but it is unclear to what
extent those protists were exposed to hydrogen sulfide.
Dinoflagellates
Nucleic acids of two dinoflagellate phylotypes were recov-
ered from the sediments. Phylotype 23 is related to dino-
flagellates of the cyst-forming genus Scrippsiella with an
identical sequence to that of Scrippsiella sp. MBIC11164
(Fig. 4, Table 1). Phylotype 23 comprised up to 40% of the
total 18S rRNA pool in the upper 6 cm of sulfidic sediment
(Fig. 5), which suggests that cysts of this species remain
viable for up to 300 years.
On the other hand, phylotype 21 with 99.8% sequence
similarity to species of the genus Pentapharsodinium (Fig. 4,
Table 1) was only recovered from the sedimentary 18S rDNA
pool. Its DNA was most likely protected inside organic-
walled cysts of Pentapharsodinium, which have been
described from recent Black Sea sediments (Marret et al.,
2004). However, the absence of Pentapharsodinium RNA
suggests that cysts of Pentapharsodinium were no longer
viable in the sulfidic sediments.
Ciliates
Phylotypes 19, 20, 25, and 26 were recovered from up to
8 cm of sulfidic Black Sea sediments (Figs 3b and 5) and
were closely related to 18S rRNA clones from unnamed
ciliates previously retrieved from the sulfidic waters (i.e.
�50 mM of sulfide) of the Danish Mariager Fjord (Fig. 4,
Table 1) (Zuendorf et al., 2006). The absence of 18S rRNA
from the four ciliates implied that they were not viable in the
subsurface Black Sea sediments, which exhibit a sulfide
concentration three times higher than in the Mariager Fjord
waters. Whether the sedimentary ciliate DNA was derived
from cells stemming from the less sulfidic chemocline could
not be confirmed because the same phylotypes were not
detected in the two analyzed sulfidic waters at 150 and
160 m.
Summary and conclusions
We have shown that analysis of gene transcripts has enabled
the identification of species that were active but escaped
identification in a parallel 18S rDNA survey. In addition, the
above examples also indicate that gene transcripts do not
always provide unequivocal proof that metabolically active
FEMS Microbiol Ecol 70 (2009) 525–539c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
536 M.J.L. Coolen & G. Shtereva
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

microorganisms are indigenous to a certain position in an
environmental gradient, because certain species stemming
from the oxygenated part of the photic zone can remain
viable in the presence of high sulfide concentrations. This is
especially true when environmental sequences represent
novel taxonomic clades with unknown phylogenetic affilia-
tion. The 18S rRNA gene survey from surface sediments
yielded taxa of pelagic origin, but none of these were
identified in the water column at the time of sampling. Vice
versa, none of the species, which were thriving in the water
column at the time of sampling, could be detected in the
fossil DNA fraction. There are various possible explanations
for this discrepancy: (1) the filtered POM only represents a
snapshot of the total annual plankton community, (2) not
all cellular material is equally well transported to the
sediment, or (3) there are species- or cell-specific variations
in the level of (post)depositional degradation of intracellular
DNA. Although the DGGE method represents a relatively
fast approach in screening a large number of samples along
the environmental gradient for major shifts in the eukaryote
community structure, it is insensitive to less dominant
species, or those with low genomic rDNA copy numbers.
Future research will require application of other molecular
biological approaches, notably tag-encoded FLX amplicon
pyrosequencing, in order to determine the diversity and
relative abundance of total vs. active eukaryotes.
Acknowledgements
We would especially like to thank Ognyana Hristova and
Tatyana Nikolova, IOBAS, for the analysis of the water
column chemistry and Dr Cornelia Wuchter, Dr Angela
Dickens, Alan Gagnon, Daniel Montlucon, Chris Ward
(WHOI), and the R/V Akademik staff, in particular Delcho
Solakov, for their extensive organizational and participatory
help with the cruise. We thank Dr Timothy Eglinton
(WHOI) and two anonymous reviewers for suggestions to
improve the manuscript. We are grateful for the financial
support from the US National Science Foundation grant
OCE 0602423, as well as funding from WHOI’s Access to the
Sea program, and a grant from the Andrew W. Mellon
Foundation Endowed Fund for Innovative Research.
References
Agatha S (2004) A cladistic approach for the classification of
oligotrichid ciliates (Ciliophora: Spirotricha). Acta Protozool
43: 201–217.
Alexander E, Stock A, Breiner HW, Behnke A, Bunge J, Yakimov
MM & Stoeck T (2009) Microbial eukaryotes in the
hypersaline anoxic L’Atalante deep-sea basin. Environ
Microbiol 11: 360–381.
Behnke A, Bunge J, Barger K, Breiner HW, Alla V & Stoeck T
(2006) Microeukaryote community patterns along an O2/H2S
gradient in a supersulfidic anoxic Fjord (Framvaren, Norway).
Appl Environ Microb 72: 3626–3636.
Besiktepe S, Kideys AE & Unsal M (1998) In situ grazing pressure
and diet vertical migration of female Calanus euxinus in the
Black Sea. Hydrobiologia 363: 323–332.
Bouvier T, Becquevort S & Lancelot C (1998) Biomass and
feeding activity of phagotrophic mixotrophs in the
northwestern Black Sea during the summer 1995.
Hydrobiologia 363: 289–301.
Buckley BA & Szmant AM (2004) RNA/DNA ratios as indicators
of metabolic activity in four species of Caribbean reef-building
corals. Mar Ecol-Prog Ser 282: 143–149.
Chicharo MA & Chicharo L (2008) RNA : DNA ratio and other
nucleic acid derived indices in marine ecology. Int J Mol Sci 9:
1453–1471.
Cline JD (1969) Spectrophotometric determination of hydrogen
sulfide in natural waters. Limnol Oceanogr 14: 454–458.
Codispoti LA, Friederich GE, Murray JW & Sakamoto CM (1991)
Chemical variability in the Black Sea: implications of
continuous vertical profiles that penetrated the oxic/anoxic
interface. Deep-Sea Res 38: S691–S710.
Coolen MJL, Boere A, Abbas B, Baas M, Wakeham SG &
Sinninghe Damste JS (2006) Fossil DNA derived from
alkenone-biosynthesizing haptophytes and other algae in
Holocene sediment from the Black Sea. Paleoceanography 21
DOI: 10.1029/2005PA001188.
Coolen MJL, Volkman JK, Abbas B, Muyzer G, Schouten S &
Sinninghe Damste JS (2007) Identification of organic matter
sources in sulfidic late Holocene Antarctic fjord sediments
from fossil rDNA sequence analysis. Paleoceanography 22
DOI: 10.1029/2006PA001309.
Coyne KJ & Cary SC (2005) Molecular approaches to the
investigation of viable dinoflagellate cysts in natural sediments
from estuarine environments. J Eukaryot Microbiol 52: 90–94.
Dawson SC & Pace NR (2002) Novel kingdom-level eukaryotic
diversity in anoxic environments. P Natl Acad Sci USA 99:
8324–8329.
Dıez B, Pedros-Alio C, Marsh TL & Massana R (2001)
Application of denaturing gradient gel electrophoresis
(DGGE) to study the diversity of marine picoeukaryotic
assemblages and comparison of DGGE with other molecular
techniques. Appl Environ Microb 67: 2942–2951.
Edgcomb VP, Kysela DT, Teske A, Gomez AD & Sogin ML (2002)
Benthic eukaryotic diversity in the Guaymas Basin
hydrothermal vent environment. P Natl Acad Sci USA 99:
7658–7662.
Eker-Develi E & Kideys AE (2003) Distribution of phytoplankton
in the southern Black Sea in summer 1996, spring and autumn
1998. J Marine Syst 39: 203–211.
Fleminger A & Hulsemann K (1977) Geographical range and
taxonomic divergence in North Atlantic Calanus (C.
helgolandicus, C. finmarchicus and C. glacialis). Mar Biol 40:
233–248.
FEMS Microbiol Ecol 70 (2009) 525–539 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
537Viable and active eukaryotes in the Black Sea
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

Grasshoff K, Ehrhardt M & Kremmling K (1983) Methods of
Seawater Analysis. Verlag Chemie Weinheim, New York.
Hamilton AK, Lovejoy C, Galand PE & Ingram RG (2008) Water
masses and biogeography of picoeukaryote assemblages in a
cold hydrographically complex system. Limnol Oceanogr 53:
922–935.
Hawkins SA, Robinson KG, Layton AC & Sayler GS (2008)
Response of Nitrobacter spp. ribosomal gene and transcript
abundance following nitrite starvation and exposure to
mechanistically distinct inhibitors. Environ Sci Technol 42:
901–907.
Houdan A, Probert I, Zatylny C, Veron B & Billard C (2006)
Ecology of oceanic coccolithophores. I. Nutritional preferences
of the two stages in the life cycle of Coccolithus braarudii and
Calcidiscus leptoporus. Aquat Microb Ecol 44: 291–301.
Hurt RA, Qiu X, Wu L, Roh Y, Palumbo AV, Tiedje JM & Zhou J
(2001) Simultaneous recovery of RNA and DNA from soils
and sediments. Appl Environ Microb 67: 4495–4503.
Jones HLJ, Leadbeater BSC & Green JC (1994) Mixotrophy in
haptophytes. The Haptophyte Algae. Systematics Association
Spec, Vol. 51 (Green JC & Leadbeater BSC, eds), pp. 247–264.
Clarendon Press, Oxford.
Jørgensen BB, Fossing H, Wirsen CO & Jannach HW (1991)
Sulfide oxidation in the anoxic Black Sea chemocline. Deep-
Sea Res 38: S1083–S1103.
Kawachi M, Inouye I, Maeda O & Chihara M (1991) The
haptonema as a food-capturing device – observations on
Chrysochromulina hirta (Prymnesiophyceae). Phycologia 30:
563–573.
Kovaleva VG & Golemanski VG (1979) Psammobiotic ciliates of
the Bulgarian coast of the Black Sea. Acta Protozool 18:
265–284.
Lopez-Garcia P, Vereshchaka A & Moreira D (2000) Eukaryotic
diversity associated with carbonates and fluid-seawater
interface in Lost City hydrothermal field. Environ Microbiol 9:
546–554.
Lopez-Garcia P, Philippe H, Gail F & Moreira D (2003)
Autochthonous eukaryotic diversity in hydrothermal sediment
and experimental microcolonizers at the Mid-Atlantic Ridge.
P Natl Acad Sci USA 100: 697–702.
Ludwig W, Strunk O, Westram R et al. (2004) ARB: a software
environment for sequence data. Nucleic Acids Res 32:
1363–1371.
MacGregor BJ, Moser DP, Baker BJ, Alm EW, Maurer M, Nealson
KH & Stahl DA (2001) Seasonal and spatial variability in Lake
Michigan sediment small-subunit rRNA concentrations. Appl
Environ Microb 67: 3908–3922.
Marcus NH, Lutz R, Burnett W & Cable P (1994) Age, viability,
and vertical-distribution of zooplankton resting eggs from an
anoxic basin – evidence of an egg bank. Limnol Oceanogr 39:
154–158.
Marcus NH, Lutz RV & Chanton JP (1997) Impact of anoxia and
sulfide on the viability of eggs of three planktonic copepods.
Mar Ecol-Prog Ser 146: 291–295.
Marret F, Leroy S, Chalie F & Gasse F (2004) New organic-walled
dinoflagellate cysts from recent sediments of Central Asian
seas. Rev Palaeobot Palyno 129: 1–20.
Mills HJ, Martinez RJ, Story S & Sobecky PA (2004) Identification
of members of the metabolically active microbial populations
associated with Beggiatoa species mat communities form Gulf
of Mexico cold-seep sediments. Appl Environ Microb 70:
5447–5458.
Moon-van der Staay S, De Wachter R & Vaulot D (2001) Oceanic
18S rDNA sequences from picoplankton reveal unsuspected
eukaryotic diversity. Nature 409: 607–610.
Murray JW & Yala-del-Rio HL (2006) The suboxic transition
zone in the Black Sea. Past and Present Marine Water Column
Anoxia, NATO Science Series: IV – Earth and Environmental
Sciences (Neretin LN, ed), pp. 105–138. Springer Verlag,
Dordrecht.
Murray JW, Jannasch HW, Honjo S et al. (1989) Unexpected
changes in the oxic/anoxic interface in the Black Sea. Nature
338: 411–413.
Muttray AF & Mohn WW (1998) RNA/DNA ratio as an indicator
of metabolic activity in resin acid-degrading bacteria. Water
Sci Technol 37: 89–93.
Niermann U & Greve W (1997) Distribution and fluctuation of
dominant zooplankton species in the southern Black Sea in
comparison to the North Sea and Baltic Sea. Sensitivity to
change: Black Sea, Baltic Sea and North Sea (Mikaelyan AS, ed),
pp. 65–78. Kluwer Academic Publishers, Dordrecht.
Norris TB, Wraith JM, Castenholz RW & McDermott TR (2002)
Soil microbial community structure across a thermal gradient
following a geothermal heating event. Appl Environ Microb 68:
6300–6309.
Overmann J, Cypionka H & Pfennig N (1992) An extremely low-
light-adapted phototrophic sulfur bacterium from the Black
Sea. Limnol Oceanogr 37: 150–155.
Persson A (2002) Proliferation of cryptic protists and
germination of resting stages from untreated sediment samples
with emphasis on dinoflagellates. Ophelia 55: 151–166.
Pruesse E, Quast C, Knittel K, Fuchs BM, Ludwig WG, Peplies J &
Glockner FO (2007) SILVA: a comprehensive online resource
for quality checked and aligned ribosomal RNA sequence data
compatible with ARB. Nucleic Acids Res 35: 7188–7196.
Schafer H & Muyzer G (2001) Denaturing Gradient Gel
Electrophoresis in Marine Microbial Ecology, pp. 425–468.
Academic Press, San Diego.
Seo HC, Kube M, Edvardsen RB et al. (2001) Miniature genome
in the marine chordate Oikopleura dioica. Science 294: 2506.
Shiganova T (2005) Changes in appendicularian Oikopleura
dioica abundance caused by invasion of alien ctenophores in
the Black Sea. J Mar Biol Assoc UK 85: 477–494.
Sogin ML, Morrison HG, Huber JA et al. (2006) Microbial
diversity in the deep sea and the underexplored ‘rare
biosphere’. P Natl Acad Sci USA 103: 12115–12120.
Song W & Packroff G (1997) Taxonomy and morphology of
marine ciliates from China with description of two new
species, Strombidium globosaneum nov. spec. and S. platum
FEMS Microbiol Ecol 70 (2009) 525–539c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
538 M.J.L. Coolen & G. Shtereva
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022

nov. spec. (Protozoa, Ciliophora). Arch Protistenkd 147:
331–360.
Stoeck T, Taylor GT & Epstein SS (2003) Novel eukaryotes from
the permanently anoxic Cariaco Basin (Caribbean sea). Appl
Environ Microb 69: 5656–5663.
Stoeck T, Zuendorf A, Breiner HW & Behnke A (2007) A
molecular approach to identify active microbes in
environmental eukaryote clone libraries. Microb Ecol 53:
328–339.
Svetlichny LS, Hubareva ES, Erkan F & Gucu AC (2000)
Physiological and behavioral aspects of Calanus euxinus
females (Copepoda: Calanoida) during vertical migration
across temperature and oxygen gradients. Mar Biol 137:
963–971.
Svetlichny LS, Kideys AE, Hubareva ES, Besiktepe S & Isinibilir M
(2006) Development and lipid storage in Calanus euxinus
from the Black and Marmara seas: variabilities due to habitat
conditions. J Marine Syst 59: 52–62.
Takishita K, Miyake H, Kawato M & Maruyama T (2005)
Genetic diversity of microbial eukaryotes in anoxic sediment
around fumaroles on a submarine caldera floor based on
the small-subunit rDNA phylogeny. Extremophiles 9:
185–196.
Takishita K, Yubuki N, Kakizoe N, Inagaki Y & Maruyama T
(2007) Diversity of microbial eukaryotes in sediment at a
deep-sea methane cold seep: surveys of ribosomal DNA
libraries from raw sediment samples and two enrichment
cultures. Extremophiles 11: 563–576.
Tillmann U (1998) Phagotrophy by a plastidic haptophyte,
Prymnesium patelliferum. Aquat Microb Ecol 14: 155–160.
Tiselius P, Petersen JK, Nielsen TG et al. (2003) Functional
response of Oikopleura dioica to house clogging due to
exposure to algae of different sizes. Mar Biol 142: 253–261.
Unal E, Frost BW, Armbrust V & Kideys AE (2006)
Phylogeography of Calanus helgolandicus and the Black Sea
copepod Calanus euxinus, with notes on Pseudocalanus
elongatus (Copepoda, Calanoida). Deep-Sea Res Pt II 53:
1961–1975.
Wagner R (1994) The regulation of ribosomal-RNA synthesis and
bacterial-cell growth. Arch Microbiol 161: 100–109.
Ward BB & Kilpatrick KA (1990) Nitrogen transformations in the
oxic layer of permanent anoxic basins: The Black Sea and
Cariaco Trench. Black Sea Oceanography (Izdar E & Murray
JW, eds), pp. 111–124. Kluwer Academic Publishers,
Dordrecht.
Zettler E, Amils R, Theroux S, Palacios C, Sogin M & Amaral-
Zettler L (2007) Protistan distribution in relation to spatial
variations of extreme geochemical parameters in the Rio
Tinto, Spain. J Phycol 43: 121.
Zettler LAA, Gomez F, Zettler E, Keenan BG, Amils R & Sogin ML
(2002) Eukaryotic diversity in Spain’s River of Fire – this
ancient and hostile ecosystem hosts a surprising variety of
microbial organisms. Nature 417: 137.
Zopfi J, Ferdelman T & Fossing H (2004) Distribution and fate of
sulfur intermediates-sulfide, tetrathionate, thiosulfate, and
elemental sulfur in marine sediments. Sulfur Biogeochemistry
(Amend JP, Edwards KJ & Lyons TW, eds), pp. 97–116. The
Geological Society of America, Boulder.
Zuendorf A, Bunge J, Behnke A, Barger KJA & Stoeck T (2006)
Diversity estimates of microeukaryotes below the chemocline
of the anoxic Mariager Fjord, Denmark. FEMS Microbiol Ecol
58: 476–491.
FEMS Microbiol Ecol 70 (2009) 525–539 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
539Viable and active eukaryotes in the Black Sea
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/70/3/525/533217 by guest on 14 July 2022




















