
3D Models of Epithelial-Mesenchymal Transition in Breast Cancer Metastasis: High-Throughput...
15
http://jbx.sagepub.com/ Journal of Biomolecular Screening http://jbx.sagepub.com/content/16/2/141 The online version of this article can be found at: DOI: 10.1177/1087057110392995 2011 16: 141 J Biomol Screen Qun Li, Chaoyu Chen, Amit Kapadia, Qiong Zhou, Mary Kay Harper, Jerome Schaack and Daniel V. Labarbera Assay Development, Validation, and Pilot Screen 3D Models of Epithelial-Mesenchymal Transition in Breast Cancer Metastasis : High-Throughput Screening Published by: http://www.sagepublications.com can be found at: Journal of Biomolecular Screening Additional services and information for http://jbx.sagepub.com/cgi/alerts Email Alerts: http://jbx.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011 jbx.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of 3D Models of Epithelial-Mesenchymal Transition in Breast Cancer Metastasis: High-Throughput...

http://jbx.sagepub.com/Journal of Biomolecular Screening
http://jbx.sagepub.com/content/16/2/141The online version of this article can be found at:
DOI: 10.1177/1087057110392995
2011 16: 141J Biomol ScreenQun Li, Chaoyu Chen, Amit Kapadia, Qiong Zhou, Mary Kay Harper, Jerome Schaack and Daniel V. Labarbera
Assay Development, Validation, and Pilot Screen3D Models of Epithelial-Mesenchymal Transition in Breast Cancer Metastasis : High-Throughput Screening
Published by:
http://www.sagepublications.com
can be found at:Journal of Biomolecular ScreeningAdditional services and information for
http://jbx.sagepub.com/cgi/alertsEmail Alerts:
http://jbx.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

© 2011 Society for Laboratory Automation and Screening www.slas.org 141
IntroductIon
In the United StateS, the probability of developing invasive breast cancer for women is 1 in 8 from birth to
death, and there are ~180,000 new cases of invasive breast cancer diagnosed each year.1 the 5-year survival rate for women diagnosed with ductal carcinoma in situ (dCiS) or localized breast cancer is 98%, which decreases significantly
for women diagnosed with invasive metastatic breast cancer to only 27%.1 therefore, metastatic dissemination remains the primary cause of mortality in breast cancer patients. in par-ticular, 15% to 20% of invasive ductal carcinomas display a basal-like (bl) phenotype defined by positive expression of human epidermal growth factor receptor 1 and vimentin and/or triple-negative (tn) phenotype, which is negative for pro-gesterone, estrogen, and human epidermal growth factor 2 receptors. bl/tn phenotypes are associated with aggressive metastasis, resistance to chemotherapy, and an overall poor prognosis.2,3
the pathogenesis of bl/tn breast cancers is still poorly understood, and it remains unclear as to what mechanisms drive these tumor cells to proliferate and metastasize.3 as a result, there are no specific targeted therapeutic systemic regimens for the treatment of these types of cancer, which ultimately contributes to the overall poor prognosis.3 however, growing evidence indi-cates that these types of cancer undergo epithelial-mesenchymal transition (eMt) prior to metastasis.2 therefore, the mechanisms that govern eMt can be targeted for the discovery of novel sys-temic therapies to treat metastatic breast cancer.
1department of pharmaceutical Sciences, School of pharmacy.2division of Medical oncology.3department of Microbiology, School of Medicine, University of Colorado denver anschutz Medial Center, aurora, Co, USa.4department of Medicinal Chemistry, University of Utah, Salt lake City, Ut, USa.
received Jul 2, 2010, and in revised form nov 8, 2010. accepted for publica-tion nov 9, 2010.
Supplementary material for this article is available on the Journal of Biomolecular Screening Web site at http://jbx.sagepub.com/supplemental.
Journal of biomolecular Screening 16(2); 2011doi: 10.1177/1087057110392995
3d Models of Epithelial-Mesenchymal transition in Breast cancer Metastasis: High-throughput Screening Assay development,
Validation, and Pilot Screen
Qun LI,1 cHAoyu cHEn,1 AMIt KAPAdIA,1 QIong ZHou,2 MAry KAy HArPEr,4 JEroME ScHAAcK,3 and dAnIEL V. LABArBErA1
despite advancements in therapies developed for the treatment of cancer, patient prognosis and mortality rates have improved minimally, and metastasis remains the primary cause of cancer mortality worldwide. an underlying mechanism promoting metastasis in many types of cancer is epithelial-mesenchymal transition (eMt). here the authors report a novel 3d model of eMt and metastatic breast cancer suitable for high-throughput screening (htS) drug discovery. the primary assay incor-porates the expression of the prognostic biomarker vimentin, as a luciferase reporter of eMt, in basil-like/triple-negative Mda-Mb-231 breast carcinoma spheroids. Using this model, the authors developed a number of known antitumor agents as control modulators of eMt. U0126, pKC412, pF2341066, dasatinib, and axitinib downregulated vimentin expression by 70% to 90% as compared to untreated spheroids. Counterassays were developed to measure spheroid viability and the inva-sive potential of Mda-Mb-231 spheroids after small-molecule treatment and used to confirm hits from primary screening. Finally, the authors conducted a pilot screen to validate this model for htS using a purified library of marine secondary metabolites. From 230 compounds screened, they obtained a Z′ score of 0.64, indicative of an excellent assay, and confirmed 4 hits, including isonaamidine b, papuamine, mycalolide e, and jaspamide. this htS model demonstrates the potential to identify small-molecule modulators of eMt that could be used to discover novel antimetastatic agents for the treatment of cancer. (Journal of Biomolecular Screening 2011;16:141-154)
Key words: eMt, Vimentin, 3d spheroid culture, high-throughput screening, marine natural products, anti-metastatic agents
original Articles
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

Li et al.
142 www.slas.org Journal of Biomolecular Screening 16(2); 2011
eMt is characterized by loss of expression of epithelial biomarkers, such as epithelial (e)-cadherin and cytokeratins, accompanied by increased expression of mesenchymal biomar-kers such as neuronal (n)-cadherin and vimentin.2,4-6 importantly, bl/tn cancers are characterized by high vimentin expression, and when vimentin expression is suppressed in these cell types, the inhibition of carcinoma cell migration and invasion has been observed.2,4,7 this demonstrates that vimentin expression not only correlates with the mesenchymal phenotype but may also play a functional role in eMt, invasion, and migration.
it is well established that cues from the microenvironment play a significant role in cellular events related to tumor progres-sion and metastasis.8 this is particularly true in the signaling events that govern eMt.9,10 despite our growing understanding of the importance of the tumor microenvironment, 2d cell-based models remain the mainstay of preclinical cancer drug discovery efforts for reasons of simplicity and cost. however, 3d culture models prove to be superior at recapitulating in vivo–like growth and differentiation of tissues as compared to 2d models.11
herein, we report a novel 3d model of eMt incorporating the biomarker vimentin into Mda-Mb-231 breast carcinoma multicellular tumor spheroids (spheroids). in this model, vimentin expression is a direct readout of the mesenchymal phenotype and is used to monitor phenotypic modulation of eMt after treatment with small molecules. this model has been adapted for high-throughput screening (htS) as a primary assay in 96- to 1536-well microplates incorporating single uni-form spheroids and extracellular matrix (eCM). in parallel, we have developed a spheroid viability counterassay and a second-ary cell invasion assay to confirm primary hits. Finally, we have conducted a pilot screen using a library of pure secondary metabolites isolated from marine organisms.
MAtErIALS And MEtHodS
Antibodies and reagents
Monoclonal mouse antivimentin antibody and mouse anti-α-tubulin antibody were purchased from Cell Signaling (danvers, Ma). peroxidase-conjugated affinipure goat anti-mouse igG (h+l) was purchased from Jackson immuno research laboratory (baltimore, pa). Matrigel™ basement membrane matrix growth factor reduced and mouse anti-e-cadherin were purchased from bd biosciences (bedford, Ma). 1-(4, 5-dimethoxy-2-nitrophenyl) ethyl ester (dMnpe-caged luciferin, spheroid-permeable firefly luciferase substrate) was purchased from invitrogen (eugene, or).
Vimentin promoter firefly luciferase cloning
We obtained the vimentin promoter firefly luciferase (Vimpro-Fluc) reporter plasmid as a kind gift from professor
Christine Gilles from the University of liege, belgium.12 the Vimpro-Fluc cassette was removed and cloned into a lentiviral vector plasmid (pCdh1-CMV-MCS1-eF1-puro), which con-tains a puromycin resistance gene. restriction endonucleases were used to digest the dna flanking both the 5′ (Kpni fol-lowed by blunt ending) and 3′ (Xbai) sides of the Vimpro-Fluc cassette. Compatible digestion sites were not possible on both the 5′ ends of the pGl3 vector or the pCdh1-CMV-MCS1-eF1-puro vector; thus, the noncompatible (Clai) end was con-verted to a blunt end. this fragment was purified from a 0.8% agarose gel and ligated into the lentiviral pCdh1-CMV-MCS1-eF1-puro vector replacing the cytomegalovirus (CMV) pro-moter creating pCdh1-Vimpro-Fluc-eF1-puro. after ligation, the plasmid was amplified using chemical transformation into dh5α Escherichia coli competent cells and then purified by Qiagen Maxiprep (Valencia, Ca).
Stable VimPro-Fluc-MDA-MB-231 cell line engineering
human 293t cells cultured in dulbecco’s modified eagle’s medium (dMeM) + 10% fetal bovine serum (FbS) were seeded at 7.5 × 106 cells in t-75 flasks for 24 h prior to transfec-tion (to be 70% confluent on day of transfection). plasmids (pCMV-VSV-G, phr-8.2 Δr, and pCdh1-Vimpro-Fluc-eF1-puro lentiviral vector) were transfected using liposomal lt1 transfection reagent as described by the manufacturer (Mirus, pittsburgh, pa). transfected cells were incubated (37%°C, 5% Co2) for 48 h. the resulting supernatant containing virus was collected and filtered through a 0.45-µm filter and supple-mented with polybrene (8 µg/ml). the supernatant containing viral particles was added to Mda-Mb-231 or pMC42-la cells (50% confluent) in 100-mm culture dishes and incubated for 24 h. the medium was removed and then replaced with fresh medium (rpMi-1640 + 10% FbS). after 3 days, transduced Mda-Mb-231 and pMC42-la cells were selected with 1 µg/ml or 0.5 µg/ml of puromycin, respectively (previously deter-mined with kill curves), over 7 days.
Vimentin expression in 2D cell culture
all cells were cultured in 100-mm tissue culture dishes in royal park Memorial institute–1640 medium (rpMi-1640; invitrogen) supplemented with 10% FbS (Sigma-aldrich, St. louis, Mo) and maintained in a 5% Co2 incubator at 37°C. Vimpro-Fluc-Mda-Mb-231 and Vimpro-Fluc-pMC42-la were seeded at 5000 cells/well into 96-well white plates and incubated overnight for 16 h. Vimpro-Fluc-pMC42-la and Vimpro-Fluc-Mda-Mb-231 cells were treated as described in Figure 1 for 72 h. the treatment medium was replaced with 50 µl fresh medium, then 50 µl of one-Glo luciferase reagent (promega, Madison, Wi), and incubated at room temperature for 10 min. in parallel, after small-molecule treatment, cells
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

3d Models of EMt in Breast cancer Metastasis
Journal of Biomolecular Screening 16(2); 2011 www.slas.org 143
were subjected to the Celltiter-Glo® (promega) luminescent assay to assess cell viability. the Celltiter-Glo® reagent was added to each well in equal volume to the treatment medium and incubated for 10 min at room temperature. For both assays, luminescence was recorded with an integration time of 1 s/well using a Clarity™ luminescence Microplate reader (biotek® instruments, Winooski, Vt).
3D culture of uniform single spheroids
Uniform single-spheroids of Mda-Mb-231 and Vimpro-Fluc-Mda-Mb-231 breast carcinoma cells were cultured as follows. the 96-well flat-bottom plates were coated with 50 µl of a 1.5% agarose (weight/volume) solution in serum-free rpMi-1640 (no phenol red) medium (freshly autoclaved at 121°C for 15 min). during the coating process, the agarose/rpMi-1640 solution is maintained at ≥60°C followed by cool-ing and setting at room temperature for 40 min. then cells were plated at a density of 10,000 cells/well in 50 µl of rpMi-1640 (10% FbS, no phenol red) and centrifuged at 1000 rpm for 15 min to induce cell aggregation. the aggregated cells were then coated with 50 µl of a 10% solution of growth factor–reduced Matrigel™ (protein concentration: 7.1 mg/ml) in cell culture medium, resulting in a final volume of 100 µl with 5% Matrigel™. Spheroids were cultured for 4 days to reach an average diameter of 500 µm under standard tissue culture con-ditions (37°C, 5% Co2) without changing or adding medium. the method used to obtain spheroid growth kinetics and culture spheroids in 1536-well plates is provided in the supplementary data.
Control modulators of EMT and treatment concentrations
the following known modulators of eMt were purchased: axitinib and pF-2341066 (Selleck, houston, tX); dasatinib and pKC412 (lC laboratory, Woburn, Ma); plX4720 (eMd Chemicals, San diego, Ca); U0126 (Upstate Cell Signaling Solutions; lake placid, ny); 1,5-isoquinolinediol, Sb-505124, wortmannin, and y-27632 (Sigma-aldrich); aiib2 blocking antibody (University of iowa developmental Studies hybrid oma bank, iowa City, ia); and antihuman iGFir blocking antibody (r&d Systems, Minneapolis, Mn). Spheroids were cultured as described above in the appropriate 96-well microplates for 4 days. on day 5 (average spheroid diameter ~500 µm), the spheroids were treated with 15-µl aliquots of the modulators in rpMi-1640, resulting in a final concentration/well as follows: 10 µM U0126, axitinib, y-27632, plX4720, wortmannin, Sb-505124, pKC412, 1,5-isoquinolinediol, and pF-2341066; 1 and 10 nM dasat-inib; and 10 µg/ml iGFir and aiib2. the spheroids were treated for 72 h followed by quantitative measurement in various assays as described below.
Primary assay
Vimentin expression in VimPro-Fluc MDA-MB-231 sphe-roids. Vimpro-Fluc-Mda-Mb-231 spheroids were cultured in 96-well optical bottom white plates and treated as described above. on day 8, after 72-h treatment with control eMt modu-lators, live spheroid vimentin expression was measured by add-ing 10 µl (final [12.5 µM]) of dMnpe-caged luciferin to each well and incubating at 37°C, 5% Co2 for 2 h. Untreated day 5 spheroids were used as an additional negative control, and day 8 spheroids treated with 10% triton X-100 at 37°C for 2 h were used as a control to induce spheroid death. luminescence was recorded as described above.
Multiplexing VimPro-Fluc-MDA-MB-231 spheroid viability. to access spheroid viability, the acid phosphatase (aph) coloro-metric (od405) assay13 was adapted to be multiplexed with vimentin expression in Vimpro-Fluc spheroids as follows: directly after measuring live Vimpro-Fluc-Mda-Mb-231 sphe-roid vimentin expression, cell viability was measured by adding 125 µl of aph assay buffer (0.1 M sodium acetate, 0.1% triton X-100, supplemented with 10 mM p-nitrophenyl phosphate; pierce, rockford, il) to each well followed by incubating for 90 min at 37°C. the optical density (od405) was measured directly after adding 10 µl of 1n naoh to each well using a Spectra Max 190 microplate reader (Molecular devices, Sunnyvale, Ca). then, 10% triton X-100 treatment at 37°C for 2 h before adding aph buffer was used as positive control to induce cell death.
Spheroid staining and sectioning. Spheroids were cultured and treated with U0126 and axitinib for 72 h as described above. they were then transferred into a new 96-well plate and washed with phosphate-buffered saline (pbS) 2×. Spheroids were fixed with 50 µl of 3.7% formaldehyde for 10 min at room temperature. they were then washed once with pbS, and 50 µl of 0.2% triton X-100 in pbS (v/v) was added for 10 min at room temperature to permeate cells. next they were blocked with 50 µl of cell culture medium (10% FbS) for 1 h at room temperature. blocking medium was then aspirated, and sphe-roids were treated with mouse antivimentin at 1:100 in tris-buffered saline tween-20 (tbSt) containing 20% cell culture medium (v/v) with shaking at 4°C overnight. they were then washed (3× tbSt) and stained with FitC green fluorescent conjugated goat antimouse secondary antibody at 1:1000 and with dapi (4,′ 6-diamidino-2-phenylindole) at 1:1000 in tbSt containing 20% cell culture medium (v/v) for 1 h at room tem-perature and washed with tbSt 4×. after fixing with formal-dehyde, some spheroids were sectioned using a leica CM 1950 cryostat (leica Microsystems, Wetzlar, Germany) with a thick-ness of 10 µm. Spheroid sections were washed with pbS and stained as described above. Spheroid sections on slides were covered with mounting medium and cover slips. imaging of
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

Li et al.
144 www.slas.org Journal of Biomolecular Screening 16(2); 2011
whole spheroids was done with an UltraVieW® VoX 3d confo-cal microscope (perkinelmer, Waltham, Ma) using a 20× objec-tive. Spheroid sections were imaged using a nikon d-eclipse C1 confocal microscope system (nikon instruments, Melville, ny). Vimentin protein expressions in spheroid sections were quantitated from multiple replicates using imageJ software (national institutes of health, bethesda, Md) and 5 random fields per spheroid section.
3D spheroid Western blot analysis. protein for Western blot analysis was obtained by using 10 spheroids per sample group. the spheroids were collected in 1.5-ml tubes, centrifuged at 10,000 rpm for 5 min at 4°C, and then washed with ice-cold pbS (2×). the spheroids were lysed with 66.6 µl spheroid lysis buffer containing 50 mM tris (ph 8.0), 150 mM naCl, 1% nonidet p-40, 20 mM naF, 2 mM sodium orthovanadate, 20 mM β-glycerophosphate, 1 µg/ml antipain, 1 µg/ml aprotinin, 10 µg/ml leupeptin, 1 µg/ml pepstatin a, and 20 µg/ml phenyl-methylsulfonyl fluoride (pMSF). Following lysis, each sample group was treated with 33.4 µl of 3× loading buffer (no β-mercaptoethanol). the protein content (10 µl) equivalent of 1 spheroid for each sample group was subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Subsequently, pro-tein was transferred to immobilon-p membranes (Millipore, billerica, Ma) in transfer buffer (50 mmol/l tris base, 40 mmol/l glycine, 0.04% sodium dodecyl sulfate, 10% metha-nol) using an owl semidry transfer apparatus from thermo Scientific (rochester, ny). the membranes were blocked in 5% nonfat dry milk–tbSt (50 mM tris hCl, 150 mM naCl, 0.05% tween-20 [ph 8.0]) and treated with mouse antivimentin at 1:1000 while shaken at 4°C overnight. Membranes were then washed (3× tbSt), probed with antimouse horseradish peroxi-dase (hrp)–conjugated secondary antibody (1:10,000 for 1 h at room temperature [rt]), washed again as before, and developed with eCl plus (amersham, pittsburgh, pa).
Secondary assay
Cell invasion from 3D spheroid culture. Vimpro-Fluc-Mda-Mb-231 spheroids were treated with control eMt mod-ulators as described using 15 spheroids per sample group. the spheroids for each sample group were pooled into 1.5-ml micro tubes and collected by centrifugation at 10,000 rpm for 10 min at rt. each sample group was then treated with 200 µl of spheroid dissociation solution (0.25% trypsin-edta) and incubated at 37°C for 30 min. Single-cell suspensions of each group in rpMi-1640 medium containing 0.1% bovine serum albumin (bSa) were seeded at 35,000 cells/chamber (50 µl) into the top chambers of a 96-well invasion plate (bd biosciences, bedford, Ma) previously coated with 50 µl of Matrigel™ (40 µg/ml protein in rpMi-1640 medium).
Chemoattractant (10% FbS in rpMi-1640 medium, 150 µl) was added to each bottom chamber, and the plate was incubated (37°C, 5% Co2) for 24 h. Following incubation, the insert was removed and the medium aspirated from both top and bottom chambers. Cells that did not invade were carefully removed from the top chamber using a cotton swab. each side of the membrane insert was washed 2× by adding 200 µl pbS-CMF (Ca/Mg free) to the bottom chamber, replacing the insert, and adding 50 µl pbS-CMF to the top chamber. the pbS-CMF was removed, and the invading cells were fixed with 200 µl of 4% paraformaldehyde in the bottom chambers at 37°C for 15 min. the insert was washed 2× with pbS-CMF as described above. the invading cells were subsequently stained with 200 µl dapi (3 µg/ml) in pbS-CMF for 30 min at rt and washed 4×, as described above, in 15-min intervals. invading cells were quantified using 5 random fields per filter insert in triplicate at 20× magnification using a fluorescent microscope equipped with metamorph image analysis and quantification software (Molecular devices, downingtown, pa).
Marine natural products pilot screening
a marine natural products library of 230 pure compounds was formatted into 3-mM stock solutions in dMSo and screened using a final concentration of 10 µM with a final volume of 0.3% dMSo. these compounds were screened alongside 10 µM axitinib, a positive control that downregulates vimentin expression, and 0.3% dMSo as negative control and vehicle percent used that does not modulate vimentin expression (for plate density map, see Supplemental Figure S2).
Uniform single spheroids were cultured in 96-well optical bottom white plates as described above and treated on day 5. to avoid edge effects, the outer wells were unused and filled with sterilized water. then, 15 µl of controls and marine natural products was added to each well at final concentrations described above. after 72 h, vimentin expression was measured by adding 10 µl (final 12.5 µM) of dMnpe-caged luciferin to each well and incubated for 2 h (37°C, 5% Co2). luminescence was recorded with an integration time of 1 s/well using a Clarity™ luminescence Microplate reader (biotek® instruments). directly following the primary assay measuring vimentin expression, spheroid viability was multiplexed using the aph assay. hits were confirmed using the cell invasion assay.
Statistical analysis and assay validation
data were subjected to unpaired 1-tailed t-test statistical analysis using prism (Graphpad Software, San diego, Ca), and p-values ≤0.01 (**) or ≤0.001 (***) were considered statisti-cally significant. Fluorescence intensity and Western blot quan-tification were carried out using imageJ software (national
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

3d Models of EMt in Breast cancer Metastasis
Journal of Biomolecular Screening 16(2); 2011 www.slas.org 145
institutes of health). the marine natural product pilot screen and 3d spheroid assay were validated using the statistical parameter Z′ factor.14
rESuLtS
Primary assay development
Vimentin promoter firefly luciferase cloning and stable cell line engineering. We cloned the Vimpro-Fluc cassette into the lentiviral vector pCdh1-MCS1-eF1-puro generating the pCdh1-Vimpro-Fluc-eF1-puro plasmid. this plasmid was used to generate a lentiviral vector used to stably transduce pMC42-la and Mda-Mb-231 breast carcinoma cell lines to create the Vimpro-Fluc-pMC42-la and Vimpro-Fluc-Mda-Mb-231 cell lines, respectively (Fig. 1).15,16
We then cultured these cell lines as monolayers in 96-well microplates and treated them with U0126, the specific inhibitor of mitogen-activated protein kinase kinase (MeK1). Vimpro-Fluc-pMC42-la cells were induced through eMt with 10 ng/ml epidermal growth factor (eGF) treatment for 72 h, resulting in upregulation of vimentin expression with concomitant down-regulation of e-cadherin (Fig. 1A,B).15 eGF-induced eMt was significantly inhibited by U0126 (p ≤ 0.001) as well as the control aG1478, an epidermal growth factor receptor (eGFr) kinase domain inhibitor. Vimpro-Fluc-Mda-Mb-231 cells, which highly express vimentin and do not express e-cadherin, showed significant downregulation of Vimpro-Fluc activity after treatment with U0126 (Fig. 1B). however, aG1478 had no statistically significant effect on Vimpro-Fluc activity (p > 0.05). importantly, downregulation of vimentin gene expres-sion correlated with the downregulation of vimentin protein expression in both pMC42-la and Mda-Mb-231 cells (Fig. 1A-d). Finally, both Mda-Mb-231 and eGF-treated pMC42-la cells expressing vimentin displayed a mesenchy-mal phenotype marked by a high invasive potential, and this invasive potential was significantly diminished after U0126 treatment (Fig. 1E,F). taken together, this indicates that the Vimpro-Fluc gene stably integrated in Mda-Mb-231 and pMC42-la cells offers a simplified system in which readily measured and quantitated luciferase expression acts as a sur-rogate for endogenous vimentin expression and eMt.
3D culture of uniform single MDA-MB-231 spheroids for HTS. Methods describing the standardized 3d culture of multi-cellular tumor spheroids with or without eCM (Matrigel™) have been published.17,18 We used and modified these methods to develop our model. agarose was used as a nonadherent sur-face to coat 96-well plates, facilitating tumor cell aggregation. Coating aggregated cells with Matrigel™ induces spheroid formation, which is not achieved with agarose alone (Fig. 2A).
in addition, we have characterized the spheroid growth kinet-ics, which demonstrates a uniform growth rate over 30 days for both Mda-Mb-231 and Vimpro-Fluc-Mba-Mb-231 cells (Fig. 2c). Under these conditions, we can accurately control spheroid growth, ensuring uniformity of spheroids in 96-well plates (Fig. 2B). Furthermore, we have adapted this methodol-ogy to obtain uniform single spheroids cultured in 1536-well htS plates (see Supplemental Figure S5).
Vimentin expression in VimPro-Fluc MDA-MB-231 sphe-roids. the Vimpro-Fluc Mda-Mb-231 cells were used in the 3d culture model and developed as a primary assay for htS. to test the 3d culture model and identify potential positive controls, 12 known antitumor agents were selected for their ability to inhibit signaling pathways known to control or modu-late eMt (Fig. 3A).19,20 Vimpro-Fluc-Mda-Mb-231 spheroids were treated with control eMt modulators on day 5. after a 72-h treatment, live spheroid Vimpro-Fluc activity was meas-ured by adding dMnpe-caged luciferin. Vimpro-Fluc activity was significantly downregulated by 70% to 90% after U0126, dasatinib, axitinib, pF2341066, or pKC412 treatment (Fig. 3B).
VimPro-Fluc-MDA-MB-231 spheroid viability multiplex assay. after 72-h treatment with control modulators and meas-uring live Vimpro-Fluc-Mda-Mb-231 spheroid vimentin pro-moter activity, cell viability was measured using a modified aph assay adapted to multiplex with the primary assay (Fig. 3c).13 Control modulators of eMt showed no statistically sig-nificant changes in spheroid viability (p > 0.05) compared to untreated spheroids on day 8. in addition, none of the control modulators displayed spheroid viability less than control sphe-roids on day 5 before treatment. a 2-hour treatment on day 8 with 10% triton X-100 was used as a control to identify sig-nificant spheroid death. thus, downregulation of Vimpro-Fluc activity by the control modulators was not due to spheroid death. Furthermore, the Vimpro-Fluc activity in day 5 sphe-roids was higher than all of the control modulator–treated spheroids, which indicates that these compounds have a spe-cific effect on Vimpro-Fluc activity.
Vimentin protein expression in MDA-MB-231 spheroids. to determine if the downregulation of Vimpro-Fluc activity cor-relates with functional protein expression, spheroids were treated as in the primary assay with control modulators of eMt followed by Western blot analysis. Vimpro-Fluc activity was normalized to spheroid viability and compared to vimentin protein expression (Fig. 4A). based on these data, there appears to be a threshold of 50% downregulation of Vimpro-Fluc activ-ity that must be reached before protein downregulation occurs. When Vimpro-Fluc activity is downregulated between 50% and 70%, there is significant downregulation of vimentin protein
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

Li et al.
146 www.slas.org Journal of Biomolecular Screening 16(2); 2011
FIg. 1. Vimentin promoter firefly luciferase reporter activity (Vimpro-Fluc). (A) Vimpro-Fluc-pMC42-la cells were induced through epithelial-mesenchymal transition (eMt) with epidermal growth factor (eGF) resulting in the upregulation of Vimpro-Fluc activity with concomitant down-regulation of e-cadherin protein expression (inset). eGF-induced eMt inhibited by treatment with U0126 or aG1478. (B) Western blot analysis after eGF-induced eMt showing gain of vimentin protein expression that is inhibited by U0126 or aG1478. (c) Vimpro-Fluc-Mda-Mb-231 mesenchymal cells displaying high vimentin expression determined by the Vimpro-Fluc reporter activity that can be inhibited by U0126 treatment. (d) Western blot analysis of Vimpro-Fluc cells showing high vimentin expression that is inhibited by U0126 treatment. (E) the invasive potential of eGF-induced pMC42-la is significantly inhibited by U0126 treatment. (F) the invasive potential of Mda-Mb-231 cells is significantly inhib-ited by U0126 treatment. the t-test was used for statistical analysis. **p ≤ 0.01. ***p ≤ 0.001. rlU, relative luminescence units.
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

3d Models of EMt in Breast cancer Metastasis
Journal of Biomolecular Screening 16(2); 2011 www.slas.org 147
expression. When Vimpro-Fluc downregulation reaches ≥70%, then downregulation of vimentin protein expression is more pronounced. of the modulators tested—U0126, dasatinib,
axitinib, and pF2341066—all downregulated Vimpro-Fluc activity by ≥70%, which correlated with a significant down-reglation of vimentin protein expression. Finally, dose-response
FIg. 2. Mda-Mb-231 spheroid culture methodology suitable for high-throughput screening (htS). (A) the top panel depicts Mda-Mb-231 cell aggregation on 96-well plates coated with agarose. When aggregated cells are coated with 5% Matrigel™, robust spheroid formation occurs, resulting in spheroid diameters of 500 µm (day 6) and 1 mm (day 12) (bottom panel). eCM, extracellular matrix. (B) digital camera view depict-ing uniform single spheroids in a 96-well plate. (c) Spheroid growth kinetics over 32 days.
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

Li et al.
148 www.slas.org Journal of Biomolecular Screening 16(2); 2011
curves were generated with both U0126 (iC50 = 2.5 µM) and axitinib (iC50 = 0.25 µM), demonstrating that these small mol-ecules modulate Vimpro-Fluc activity in a dose-dependent manner, indicating that their respective target(s) and signaling pathways play a significant role in maintaining vimentin expres-sion and possibly mesenchymal homeostasis (Fig. 4B).
to further assess vimentin expression in Vimpro-Fluc-Mda-Mb-231 spheroids, we conducted immunofluorescent staining of whole spheroids treated with U0126. Significant down-regulation of vimentin protein expression was observed through-out the outer surface of U0126-treated spheroids as visualized by
3d confocal microscopy (Fig. 5A). to measure the extent of pen-etration into the spheroid by U0126 or axitinib, spheroids were sectioned and stained for vimentin (Fig. 5B,c). these results dem-onstrate that both U0126 and axitinib penetrated into the spheroid, resulting in downregulation of vimentin throughout the whole spheroid by more than 80% as compared to untreated spheroids.
Secondary assay development
as a secondary assay to confirm hits from the primary assay, the boyden chamber cell invasion method was used and
FIg. 3. primary assay development. (A) Known signaling pathways that control or modulate epithelial-mesenchymal transition (eMt), includ-ing small-molecule inhibitors. (B) live spheroid Vimpro-Fluc activity after treatment with control modulators of eMt for 72 h compared to untreated control spheroids: day 5 (d5), day 8 (d8), and triton X-100–induced spheroid death. (c) Spheroid viability measured by multiplexing the acid phosphatase (aph) assay directly after measuring Vimpro-Fluc activity. the t-test analysis showed no statistical difference between spheroids treated with control eMt modulators and untreated d8 spheroids. rlU, relative luminescence units.
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

3d Models of EMt in Breast cancer Metastasis
Journal of Biomolecular Screening 16(2); 2011 www.slas.org 149
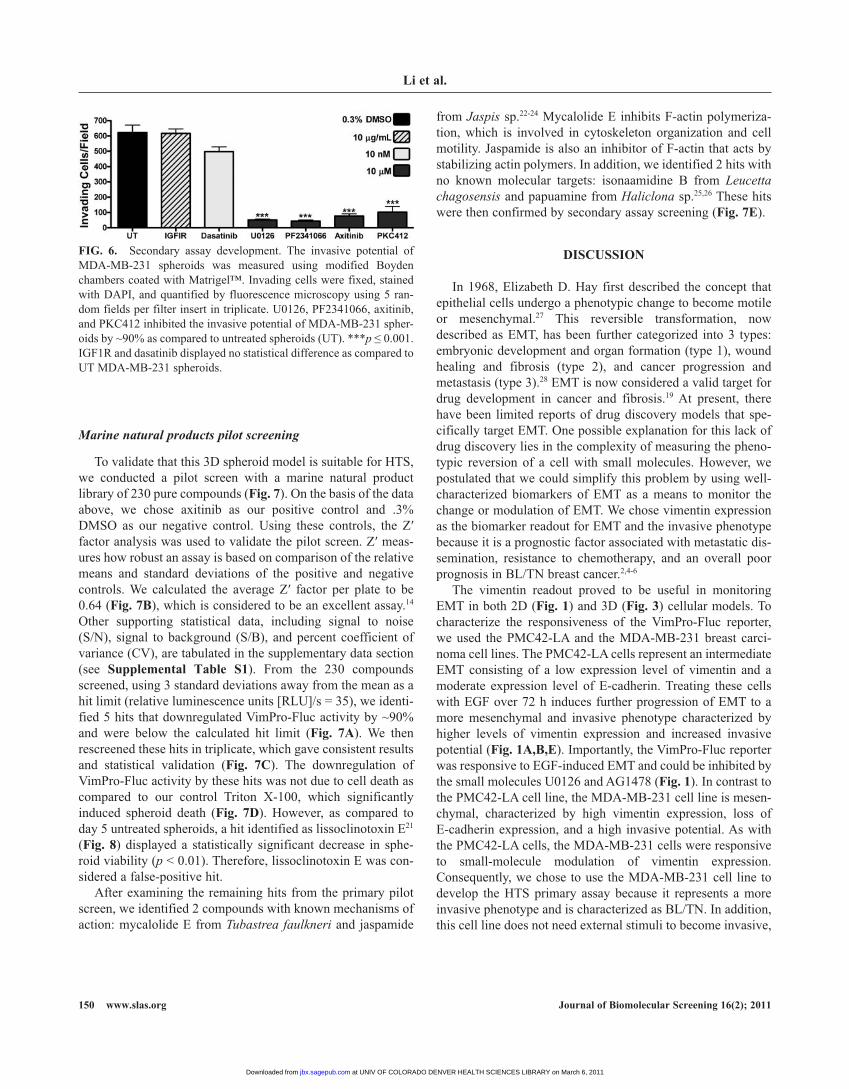
modified to measure the invasive potential of cells isolated from spheroids after treatment with control modulators of eMt. Spheroids were treated as in the primary assay. Following treatment, each sample group was disassociated into single-cell suspensions and seeded into invasion chambers followed by a 24-h incubation period. Under these conditions, the control modulators iGFir, dasatinib, U0126, pF2341066, axitinib, and pKC412 were tested (Fig. 6). U0126, pF2341066, axitinib, and pKC412 caused significant inhibition of the invasive potential of Mda-Mb-231 spheroids. Conversely, dasatinib, a potent
inhibitor of vimentin gene expression, did not significantly alter the invasive potential of Mda-Mb-231 spheroids. therefore, cell invasion is a viable assay that can be used to rule out false-positive hits identified in the primary screen.
FIg. 4. (A) Vimpro-Fluc activity in spheroids after 72-h treatment with control modulators of epithelial-mesenchymal transition (eMt) normalized to spheroid viability and compared to vimentin protein expression using Western blot analysis. (B) dose-response curves for both U0126 and axitinib control modulators of eMt. rlU, relative luminescence units. FIg. 5. (A) 3d confocal image (20× objective) of individual whole
spheroids fixed and stained for vimentin (green) and dapi (blue). (B) representative confocal image (10× objective) of individual spheroids fixed, sectioned, and then stained for vimentin (green) and dna (dapi, blue). (c) Vimentin protein expression from sections quanti-tated from multiple replicate spheroid treatments with U0126 and axitinib using imageJ software.
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from
Li et al.
150 www.slas.org Journal of Biomolecular Screening 16(2); 2011
Marine natural products pilot screening
to validate that this 3d spheroid model is suitable for htS, we conducted a pilot screen with a marine natural product library of 230 pure compounds (Fig. 7). on the basis of the data above, we chose axitinib as our positive control and .3% dMSo as our negative control. Using these controls, the Z′ factor analysis was used to validate the pilot screen. Z′ meas-ures how robust an assay is based on comparison of the relative means and standard deviations of the positive and negative controls. We calculated the average Z′ factor per plate to be 0.64 (Fig. 7B), which is considered to be an excellent assay.14 other supporting statistical data, including signal to noise (S/n), signal to background (S/b), and percent coefficient of variance (CV), are tabulated in the supplementary data section (see Supplemental table S1). From the 230 compounds screened, using 3 standard deviations away from the mean as a hit limit (relative luminescence units [rlU]/s = 35), we identi-fied 5 hits that downregulated Vimpro-Fluc activity by ~90% and were below the calculated hit limit (Fig. 7A). We then rescreened these hits in triplicate, which gave consistent results and statistical validation (Fig. 7c). the downregulation of Vimpro-Fluc activity by these hits was not due to cell death as compared to our control triton X-100, which significantly induced spheroid death (Fig. 7d). however, as compared to day 5 untreated spheroids, a hit identified as lissoclinotoxin e21 (Fig. 8) displayed a statistically significant decrease in sphe-roid viability (p < 0.01). therefore, lissoclinotoxin e was con-sidered a false-positive hit.
after examining the remaining hits from the primary pilot screen, we identified 2 compounds with known mechanisms of action: mycalolide e from Tubastrea faulkneri and jaspamide
from Jaspis sp.22-24 Mycalolide e inhibits F-actin polymeriza-tion, which is involved in cytoskeleton organization and cell motility. Jaspamide is also an inhibitor of F-actin that acts by stabilizing actin polymers. in addition, we identified 2 hits with no known molecular targets: isonaamidine b from Leucetta chagosensis and papuamine from Haliclona sp.25,26 these hits were then confirmed by secondary assay screening (Fig. 7E).
dIScuSSIon
in 1968, elizabeth d. hay first described the concept that epithelial cells undergo a phenotypic change to become motile or mesenchymal.27 this reversible transformation, now described as eMt, has been further categorized into 3 types: embryonic development and organ formation (type 1), wound healing and fibrosis (type 2), and cancer progression and metastasis (type 3).28 eMt is now considered a valid target for drug development in cancer and fibrosis.19 at present, there have been limited reports of drug discovery models that spe-cifically target eMt. one possible explanation for this lack of drug discovery lies in the complexity of measuring the pheno-typic reversion of a cell with small molecules. however, we postulated that we could simplify this problem by using well-characterized biomarkers of eMt as a means to monitor the change or modulation of eMt. We chose vimentin expression as the biomarker readout for eMt and the invasive phenotype because it is a prognostic factor associated with metastatic dis-semination, resistance to chemotherapy, and an overall poor prognosis in bl/tn breast cancer.2,4-6
the vimentin readout proved to be useful in monitoring eMt in both 2d (Fig. 1) and 3d (Fig. 3) cellular models. to characterize the responsiveness of the Vimpro-Fluc reporter, we used the pMC42-la and the Mda-Mb-231 breast carci-noma cell lines. the pMC42-la cells represent an intermediate eMt consisting of a low expression level of vimentin and a moderate expression level of e-cadherin. treating these cells with eGF over 72 h induces further progression of eMt to a more mesenchymal and invasive phenotype characterized by higher levels of vimentin expression and increased invasive potential (Fig. 1A,B,E). importantly, the Vimpro-Fluc reporter was responsive to eGF-induced eMt and could be inhibited by the small molecules U0126 and aG1478 (Fig. 1). in contrast to the pMC42-la cell line, the Mda-Mb-231 cell line is mesen-chymal, characterized by high vimentin expression, loss of e-cadherin expression, and a high invasive potential. as with the pMC42-la cells, the Mda-Mb-231 cells were responsive to small-molecule modulation of vimentin expression. Consequently, we chose to use the Mda-Mb-231 cell line to develop the htS primary assay because it represents a more invasive phenotype and is characterized as bl/tn. in addition, this cell line does not need external stimuli to become invasive,
FIg. 6. Secondary assay development. the invasive potential of Mda-Mb-231 spheroids was measured using modified boyden chambers coated with Matrigel™. invading cells were fixed, stained with dapi, and quantified by fluorescence microscopy using 5 ran-dom fields per filter insert in triplicate. U0126, pF2341066, axitinib, and pKC412 inhibited the invasive potential of Mda-Mb-231 spher-oids by ~90% as compared to untreated spheroids (Ut). ***p ≤ 0.001. iGF1r and dasatinib displayed no statistical difference as compared to Ut Mda-Mb-231 spheroids.
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

3d Models of EMt in Breast cancer Metastasis
Journal of Biomolecular Screening 16(2); 2011 www.slas.org 151
FIg. 7. Marine natural product pilot screen. (A) Vimpro-Fluc activity results from primary screening of 230 marine secondary metabolites. the dashed black line represents the hit limit calculated as 3 standard deviations away from the mean. (B) the Z′ statistical validation per plate screened is indicated, and the dashed black line represents the Z′ limit. (c) the Vimpro-Fluc activity of 5 identified hits rescreened in triplicate. (d) Spheroid viability of 5 hits identified from primary screening. With the exception of lissoclinotoxin e (p ≤ 0.001), all the hits identified showed no statistical difference in spheroid viability as compared to untreated spheroids on day 5. Spheroids treated with 10% triton X-100 two hours prior to acid phosphatase (aph) analysis represent a control that significantly induces spheroid death. (E) Secondary assay screening confirmed the hits identified also inhibit the invasive potential of Mda-Mb-231 spheroids by ~90% (p ≤ 0.001) as compared to untreated day 8 spheroids. rlU, relative luminescence units.
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

Li et al.
152 www.slas.org Journal of Biomolecular Screening 16(2); 2011
and it proved very responsive to small-molecule modulators of vimentin expression, which is indicative of reversion or partial reversion of eMt. during the course of our studies, we found that the conventional 2d cell culture models were significantly different from the 3d cell culture models. the most notable
differences were a much higher sensitivity to cytotoxic agents and reduced effect on eMt as compared to the 3d cell culture models. in particular, we characterized this differential cytotox-icity with the hits identified from the marine natural product pilot screen (see Supplemental Figure S4). Consequently, we
FIg. 8. the chemical structures of the 5 hits indentified from primary screen.
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

3d Models of EMt in Breast cancer Metastasis
Journal of Biomolecular Screening 16(2); 2011 www.slas.org 153
developed and adapted a 3d spheroid model of breast cancer using Mda-Mb-231 cells and extracellular matrix, mim-icking aspects of in vivo–like growth. We then optimized and adapted methods to culture uniform single spheroids in 96- to 1536-well microplates, which are suitable for htS (Fig. 2). to identify hits that explicitly modulate eMt, we modified the known aph cell viability assay, previously shown to be useful in 3d spheroid models, and developed this as a counterassay to mul-tiplex with Vimpro-Fluc activity in the 3d spheroid model (Fig. 3B).13 thus, we could eliminate false-positive hits that proved to induce spheroid death resulting in downregulation of vimentin.
Using this primary assay model, we then tested known anti-tumor agents as control modulators of eMt.19 a number of these compounds were efficient at downregulating vimentin gene expression without inducing spheroid death, most notably U0126, dasatinib, axitinib, pF2341066, and pKC412 (Fig. 3). all of these inhibitors directly or indirectly affect the Mapkinase, c-Met, and vascular endothelial growth factor receptor signaling pathways. importantly, the downregulation of vimentin gene expression by these control eMt modulators also correlated with the downregulation of vimentin protein expression (Figs. 4 and 5). a recent article, which used analysis of 421 pathways using breast cancer genome-wide association studies (GWaS), implicated the Mapkinase and c-Met path-ways as 2 of 4 total signaling pathways/cascades linked to breast cancer susceptibility through genetic aberration.29 this not only supports our results but also indicates the power of our assay to identify relevant targets and pathways related to breast cancer metastasis using biomarker expression. We next devel-oped a secondary assay to confirm viable hits from primary screening (Fig. 6). here we definitively show that hits from the primary assay can be confirmed as potential lead antimetastatic agents. interestingly, we also show that vimentin expression alone is not sufficient for cell invasion. For example, dasatinib is a c-SrC kinase inhibitor that potently downregulates Vimpro-Fluc activity by 80% at 10 nM (Fig. 3A). however, dasatinib-treated spheroids maintained their invasive potential as compared to other control eMt modulators such as U0126, pF2341066, and axitinib. this result is significant because it shows that the secondary assay has the ability to rule out false-positive hits.
Finally, we conducted a pilot screen to validate this model for htS using a library of pure marine secondary metabolites. in total, 230 compounds were screened in triplicate alongside con-trols, resulting in an average Z′ factor for 7 plates of 0.64 and the identification of 5 hits: lissoclinotoxin e, isonaamidine b, papua-mine, mycalolide e, and jaspamide (Fig. 8). lissoclinotoxin e was eliminated as a hit because of induced spheroid toxicity in the aph assay (Fig. 7d). both isonaamidine b and papuamine are antifungal agents with little known antitumor activity.25,26 Mycalolide e and jaspamide are known to be cytotoxic in con-ventional 2d cell culture models through the inhibition or stabi-lization of F-actin polymerization, respectively.22-24 a recent
article by Shankar et al.30 describing the role of pseudopodial actin dynamics as a molecular target to induce mesenchymal-epithelial transition (Met) showed that agents that destabilize F-actin revert eMt but that jaspamide stabilization of actin fila-ments, under short-term exposure (2 h) at a concentration of 100 nM, blocked the reversion of eMt in pseudopodia using con-ventional 2d cell culture. in our 3d spheroid model, we observed no spheroid toxicity with either jaspamide or mycalolide e (Fig. 7B) but saw significant downregulation of Vimpro-Fluc activity by greater than 90% (Fig. 7c). Furthermore, both jaspamide and mycalolide e abrogated the invasive potential of Mda-Mb-231 spheroids by 90%, indicative of reversion or partial reversion of eMt. in addition to the differences between 2d and 3d models, we attribute the difference in biological activity by jaspamide to long-term exposure (72 h) and treatment concentration of 10 µM.
in conclusion, we have developed a novel 3d spheroid-based model of eMt suitable for htS drug discovery. We have validated this model through pilot screening using a focused library of pure marine antitumor agents. in addition to identifying novel small-molecule modulators of eMt, this model system demonstrates the ability to identify new uses for existing antitumor agents such as jaspamide and mycalolide e. Furthermore, we demonstrate that this model allows for the discovery of known and novel molecular targets that control or modulate eMt. Finally, this study shows that cytotoxicity is relative to the model being used. this prompts the following question: is cytotoxicity in 2d cell culture a relevant endpoint for the discovery of novel antitumor agents?
AcKnowLEdgMEntS
the authors thank professor Chris M. ireland from the University of Utah for providing the marine natural product library. We also thank professor Christine Gilles from the University of liege, belgium, for the Vimpro-Fluc-pGl3 plas-mid and professor Margaret l. ackland, deakin University, Melbourne, australia, for the pMC42-la breast carcinoma cell line. this work was funded in part by a seed grant from the University of Colorado Cancer Center.
rEFErEncES
1. Jemal a, Siegel r, Ward e, hao y, Xu J, Murray t, et al: Cancer statistics, 2008. CA Cancer J Clin 2008;58:71-96.
2. Sarrio d, rodriguez-pinilla SM, hardisson d, Cano a, Moreno-bueno G, palacios J: epithelial-mesenchymal transition in breast cancer relates to the basal-like phenotype. Cancer Res 2008;68:989-997.
3. rakha ea, ellis io: triple-negative/basal-like breast cancer: review. Pathology 2009;41:40-47.
4. blick t, Widodo e, hugo h, Waltham M, lenburg Me, neve rM, et al: epithelial mesenchymal transition traits in human breast cancer cell lines. Clin Exp Metastasis 2008;25:629-642.
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from

Li et al.
154 www.slas.org Journal of Biomolecular Screening 16(2); 2011
5. Christofori G: new signals from the invasive front. Nature 2006;441:444-450.
6. Kokkinos Mi, Wafai r, Wong MK, newgreen dF, thompson eW, Waltham M: Vimentin and epithelial-mesenchymal transition in human breast cancer: observations in vitro and in vivo. Cells Tissues Organs 2007;185:191-203.
7. Mcinroy l, Maatta a: down-regulation of vimentin expression inhibits carcinoma cell migration and adhesion. Biochem Biophys Res Commun 2007;360:109-114.
8. bissell MJ, labarge Ma: Context, tissue plasticity, and cancer: are tumor stem cells also regulated by the microenvironment? Cancer Cell 2005;7:17-23.
9. Wang F, hansen rK, radisky d, yoneda t, barcellos-hoff Mh, petersen oW, et al: phenotypic reversion or death of cancer cells by altering signaling path-ways in three-dimensional contexts. J Natl Cancer Inst 2002;94:1494-1503.
10. Weaver VM, petersen oW, Wang F, larabell Ca, briand p, damsky C, et al: reversion of the malignant phenotype of human breast cells in three-dimensional culture and in vivo by integrin blocking antibodies. J Cell Biol 1997;137:231-245.
11. pampaloni F, reynaud eG, Stelzer eh: the third dimension bridges the gap between cell culture and live tissue. Nat Rev Mol Cell Biol 2007;8:839-845.
12. Gilles C, polette M, Mestdagt M, nawrocki-raby b, ruggeri p, birembaut p, et al: transactivation of vimentin by beta-catenin in human breast can-cer cells. Cancer Res 2003;63:2658-2664.
13. Friedrich J, eder W, Castaneda J, doss M, huber e, ebner r, et al: a reli-able tool to determine cell viability in complex 3-d culture: the acid phos-phatase assay. J Biomol Screen 2007;12:925-937.
14. Zhang Jh, Chung td, oldenburg Kr: a simple statistical parameter for use in evaluation and validation of high throughput screening assays. J Biomol Screen 1999;4:67-73.
15. ackland Ml, newgreen dF, Fridman M, Waltham MC, arvanitis a, Minichiello J, et al: epidermal growth factor-induced epithelio-mesenchy-mal transition in human breast carcinoma cells. Lab Invest 2003;83:435-448.
16. price Jt, tiganis t, agarwal a, djakiew d, thompson eW: epidermal growth factor promotes Mda-Mb-231 breast cancer cell migration through a phosphatidylinositol 3'-kinase and phospholipase C–dependent mechanism. Cancer Res 1999;59:5475-5478.
17. Friedrich J, Seidel C, ebner r, Kunz-Schughart la: Spheroid-based drug screen: considerations and practical approach. Nat Protoc 2009;4:309-324.
18. ivascu a, Kubbies M: rapid generation of single-tumor spheroids for high-throughput cell function and toxicity analysis. J Biomol Screen 2006;11:922-932.
19. Chua K-n, Ma J, thiery J-p: targeted therapies in control of eMt in carcinoma and fibrosis. Drug Discov Today Dis Mech 2007;4:261-267.
20. normanno n, Morabito a, de luca a, piccirillo MC, Gallo M, Maiello Mr, et al: target-based therapies in breast cancer: current status and future perspectives. Endocr Relat Cancer 2009;16:675-702.
21. davis ra, Sandoval it, Concepcion Gp, da roCha rM, ireland CM: lissoclinotoxins e and F, novel cytotoxic alkaloids from a philippine didemnid ascidian. Tetrahedron 2003;59:2855-2859.
22. Zabriskie t, Klocke J, ireland CM, Markus a, Molinski t, Faulkner d, et al: Jaspamide, a modified peptide from a Jaspis sponge with insecticidal and antifungal activity. J Am Chem Soc 1986;108:3123-3124.
23. Senderowicz aM, Kaur G, Sainz e, laing C, inman Wd, rodriguez J, et al: Jasplakinolide’s inhibition of the growth of prostate carcinoma cells in vitro with disruption of the actin cytoskeleton. J Natl Cancer Inst 1995;87:46-51.
24. rashid Ma, Gustafson Kr, Cardellina Jh ii, boyd Mr: Mycalolides d and e, new cytotoxic macrolides from a collection of the stony coral Tubastrea faulkneri. J Nat Prod 1995;58:1120-1125.
25. Carmely S, ilan M, Kashman y: 2-amino imidazole alkaloids from the marine sponge Leucetta chagosensis. Tetrahedron 1989;45:2193-2200.
26. baker bJ, Scheuer pJ, Shoolery Jn: papuamine, an antifungal pentacyclic alkaloid from a marine sponge, Haliclona sp. J Am Chem Soc 1988;110:956-966.
27. hay ed: organization and fine structure of epithelium and mesenchyme in the developing chick embryo. in Fleischmajer r, billingham re (eds): Epithelial-Mesenchymal Interactions. baltimore: Williams & Wilkins, 1968:31-55.
28. Zeisberg M, neilson eG: biomarkers for epithelial-mesenchymal transi-tions. J Clin Invest 2009;119:1429-1437.
29. Menashe i, Maeder d, Garcia-Closas M, Figueroa Jd, bhattacharjee S, rotunno M, et al: pathway analysis of breast cancer genome-wide asso-ciation study highlights three pathways and one canonical signaling cas-cade. Cancer Res 2010;70:4453-4459.
30. Shankar J, Messenberg a, Chan J, Underhill tM, Foster lJ, nabi ir: pseudopodial actin dynamics control epithelial-mesenchymal transition in metastatic cancer cells. Cancer Res 2010;70:3780-3790.
address correspondence to:Daniel V. LaBarbera, Ph.D.
Department of Pharmaceutical Sciences School of Pharmacy University of Colorado
Denver Anschutz Medical Center 12700 East 19th Avenue, Room P15-4001
Aurora, CO 80045
E-mail: [email protected]
at UNIV OF COLORADO DENVER HEALTH SCIENCES LIBRARY on March 6, 2011jbx.sagepub.comDownloaded from




















