
1992 Harrison - Fossil Catarrhines from Fort Ternan, Kenya
22
PRIMATES, 33(4): 501-522, October 1992 501 A Reassessment of the Taxonomic and Phylogenetic Affinities of the Fossil Catarrhines from Fort Ternan, Kenya TERRY HARRISON New York University ABSTRACT. This paper presents a reassessment of the taxonomic and phylogenetic affinities of the fossil catarrhine primates from the important middle Miocene site of Fort Ternan in Kenya. Although the sample of specimens is rather small, the material can be attributed to at least five different species, identified here as Kenyapithecus wickeri, Proconsul sp., a large species of oreopithecid, Simiolus sp., and a small species of catarrhine of indeterminate status. Kenyapithecus wickeri probably represents a conservative sister-taxon of the extant large hominoids. It is more derived than "Sivapithecus" africanus from Maboko Island, from which it can be distinguished at the generic level. A small species of catarrhine from Fort Ternan can be attributed to Simiolus. It is probably a different species from Simiolus enjiessi from the early Miocene of East Africa, but additional material is needed to confirm its taxonomic distinctiveness. The occurrence of at least five species of catarrhine primates at Fort Ternan confirms that species diversity levels were as high during the middle Miocene as they had been during the early Miocene. However, the overall taxonomic and ecological composition of the middle Miocene catarrhine community was quite different, evidently due to a significant change in the local ecological setting. Taxonomic differences between the catarrhine faunas at Fort Ternan and Maboko Island can probably be explained as a consequence of a chronological separation between the two sites, and, to a lesser degree, to paleoecological differences. Key Words: Catarrhines; Hominoids; East Africa; Miocene; Fort Ternan; Taxonomy; Phylogenetic relationships; Kenyapithecus; Simiolus; Proconsul; Oreopithecids. INTRODUCTION The middle Miocene site of Fort Ternan in Kenya, the type locality of Kenyapithecus wickeri, has received a good deal of attention from palaeoanthropologists since it was first excavated by L.S.B. LEAKEY in 1961. During the 1960s and 1970s Kenyapithecus wickeri was considered to be closely related to, and possibly even synonymous with, the purported early hominid Ramapithecus from the Siwalik Hills of Indo-Pakistan. However, by the early 1980s, Kenyapithecus had suffered the same fate as its Asian counterpart, and was deposed as a potential candidate for human ancestry (see WOLPOFV, 1983). Nevertheless, the paucity of hominoid-bearing sites from the later part of the Miocene in Africa, espe- cially those as well-dated as Fort Ternan, and with faunas as intensively studied, has meant that the site has remained one of the most important Neogene localities in sub-Saharan Africa. In recent years, with the cessation of active fieldwork at Fort Ternan, the focus of interest has shifted away from the study of the fossil primates themselves, to one encom- passing more broad-based attempts to understand the paleoecology at Fort Ternan and its significance for interpreting changes in the catarrhine community structure in East Africa during the Miocene (e.g. ANDREWS• NESBIT EVANS, 1979; SHIPMANet al., 1981; SHIPMAN, 1986; PICKFORD, 1983a, 1985a, 1986a, 1987; KAPPELMAN, 1991).
Transcript of 1992 Harrison - Fossil Catarrhines from Fort Ternan, Kenya

PRIMATES, 33(4): 501-522, October 1992 501
A Reassessment of the Taxonomic and Phylogenetic Affinities of the Fossil Catarrhines from Fort Ternan, Kenya
TERRY HARRISON New York University
ABSTRACT. This paper presents a reassessment of the taxonomic and phylogenetic affinities of the fossil catarrhine primates from the important middle Miocene site of Fort Ternan in Kenya. Although the sample of specimens is rather small, the material can be attributed to at least five different species, identified here as Kenyapithecus wickeri, Proconsul sp., a large species of oreopithecid, Simiolus sp., and a small species of catarrhine of indeterminate status. Kenyapithecus wickeri probably represents a conservative sister-taxon of the extant large hominoids. It is more derived than "Sivapithecus" africanus from Maboko Island, from which it can be distinguished at the generic level. A small species of catarrhine from Fort Ternan can be attributed to Simiolus. It is probably a different species from Simiolus enjiessi from the early Miocene of East Africa, but additional material is needed to confirm its taxonomic distinctiveness. The occurrence of at least five species of catarrhine primates at Fort Ternan confirms that species diversity levels were as high during the middle Miocene as they had been during the early Miocene. However, the overall taxonomic and ecological composition of the middle Miocene catarrhine community was quite different, evidently due to a significant change in the local ecological setting. Taxonomic differences between the catarrhine faunas at Fort Ternan and Maboko Island can probably be explained as a consequence of a chronological separation between the two sites, and, to a lesser degree, to paleoecological differences.
Key Words: Catarrhines; Hominoids; East Africa; Miocene; Fort Ternan; Taxonomy; Phylogenetic relationships; Kenyapithecus; Simiolus; Proconsul; Oreopithecids.
INTRODUCTION
The middle Miocene site of Fort Ternan in Kenya, the type locality of Kenyapithecus wickeri, has received a good deal of attention from palaeoanthropologists since it was first excavated by L.S.B. LEAKEY in 1961. During the 1960s and 1970s Kenyapithecus wickeri was considered to be closely related to, and possibly even synonymous with, the purported early hominid Ramapithecus from the Siwalik Hills of Indo-Pakistan. However, by the early 1980s, Kenyapithecus had suffered the same fate as its Asian counterpart, and was deposed as a potential candidate for human ancestry (see WOLPOFV, 1983). Nevertheless, the paucity of hominoid-bearing sites from the later part of the Miocene in Africa, espe- cially those as well-dated as Fort Ternan, and with faunas as intensively studied, has meant that the site has remained one of the most important Neogene localities in sub-Saharan Africa. In recent years, with the cessation of active fieldwork at Fort Ternan, the focus of interest has shifted away from the study of the fossil primates themselves, to one encom- passing more broad-based attempts to understand the paleoecology at Fort Ternan and its significance for interpreting changes in the catarrhine community structure in East Africa during the Miocene (e.g. ANDREWS • NESBIT EVANS, 1979; SHIPMAN et al., 1981; SHIPMAN, 1986; PICKFORD, 1983a, 1985a, 1986a, 1987; KAPPELMAN, 1991).

5 0 2
Table 1. A ca t a logue o f fossil c a t a r r h i n e s f r o m For t T e r n a n a).
Accession No. Specimen Taxon
T. HARRISON
Previous references b)
KNM-FT 7
KNM-FT 8
KNM-FT 11
KNM-FT 12
KNM-FT 13
KNM-FT 14
KNM-FT 15
KNM-FT 16
KNM-FT 17
KNM-FT 18
KNM-FT 19
KNM-FT 20 c)
KNM-FT 21 c)
KNM-FT 23 c)
KNM-FT 24 r
KNM-FT 25 c)
KNM-FT 28
KNM-FT 29
KNM-FT 34
KNM-FT 35
KNM-FT 36
Right mandibular fragment with P4-MI. Dentition damaged and worn. Left C 1 germ preserved in a maxillary fragment. 9
Left I 1. Unworn; slightly abraded.
Edentulous mandibular symphysis.
Edentulous mandibular symphysis of an immature individual. Left mandibular fragment of an immature individual with MI exposed in its crypt. Left C 1. Worn. 9
Left M 1. Slightly worn.
Left Ml. Unworn.
Right P4.
Left M 3. Moderately worn.
Left mandibular fragment with CI-M3. Dentit ion moderately worn. 9
Right M2. Moderately worn.
Right M3. Moderately worn.
Right P4. Moderately worn.
Right 12. Slightly worn.
Right Cb Unerupted.
Left p3. Slightly worn.
Left M3. Heavily worn.
Right P3. Slightly worn; damaged.
Right M 1. Badly weathered.
K. wickeri
K. wickeri
Indet. sp.
Indet. sp.
Simiolus sp.
Simiolus sp.
Indet. sp.
Proconsul sp.
Indet. sp.
Indet. sp.
Simiolus sp.
Simiolus sp.
Simiolus sp.
Simiolus sp.
Simiolus sp.
Simiolus sp.
K. wickeri
Proconsul sp.
K. wickeri
K. wickeri
Oreopithecid
R. wickeri (5,6) K. wickeri (8) R. wickeri (6) K. wickeri (8) cf. L. legetet (6) L. legetet (7) cf. L. legetet (6) L. legetet (7)
cf. L. legetet (6) L. legetet (7) cf. L. legetet (6) L. legetet (7) Dryopithecus cf. africanus (6) P. africanus (7) ?P. africanus? (8) cf. L. legetet (6) L. legetet (7) cf. L. legetet (6) L. legetet (7) cf. L. legetet (6) L. legetet (7) cf. Limnopithecus (3) cf. L. legetet (6) L. legetet (7) cf. Limnopithecus (3) cf. L. legetet (6) L. legetet (7) cf. Limnopithecus (3) cf. L. legetet (6) L, legetet (7) cf. Limnopithecus (3) cf. L. legetet (6) L. legetet (7) cf. L. legetet (6) L. legetet (7) Dryopithecus cf. nyanzae (6) R nyanzae (7) K. wickeri (8) Dryopithecus cf. africanus (6) P. africanus (7) ?P.. africanus? (8) Dryopithecus cf. nyanzae (6) P. nyanzae (7) K. wickeri (8) Dryopithecus cf. nyanzae (6) R nyanzae (7) K. wickeri (8) cf. Oreopithecus (3) Suidae (6) Oreopithecus sp. (9)
(continued)

Fossi l Ca t a r rh ine s f rom For t Ternan 503
Table 1. ( con t inued)
Accession No. Specimen Taxon Previous references b)
KNM-FT 37 Right p4. Moderately-heavily worn. Oreopithecid cf. Oreopithecus (3) Suidae (6) Oreopithecus sp. (9)
Right M3. Badly weathered. Oreopithecid cf. Oreopithecus (3) Suidae (6) Oreopithecus sp. (9)
Right C x. Moderately-heavily worn. cr K. wickeri Dryopithecus cf. nyanzae (6) R nyanzae (7) K. wickeri (8) Dryopithecus cf. nyanzae (6) P. nyanzae (7) K. wickeri (8) Dryopithecus cf. africanus (6) cf. Rangwapithecus gordoni (8) cf. Dryopithecus (3) R. wickeri (4,6) K. wickeri (8) K. wickeri (1,8) R. wickeri (6) K. wickeri (1) R. wickeri (6) ?K. wickeri (8) K. wickeri (1,8) R. wickeri (6) K. wickeri (1,8) R. wickeri (6) K. wickeri (2,8) Dryopithecus cf. nyanzae (6) P. nyanzae (7) Dryopithecus cf. nyanzae (6) P. nyanzae (7) Kenyapithecus? (10) hominoid? (10) K. wickeri (8) K. wickeri (8)
KNM-FT 38
KNM-FT 39
KNM-FT 40
KNM-FT 41
KNM-FT 45
KNM-FT 46a d)
KNM-FT 46b d)
KNM-FT 47 d)
KNM-FT 48 d)
KNM-FT 49
Right M3 in mandibular fragment. M3 heavily worn and damaged.
Left C 1. Slightly worn; tip of crown missing. (7
Left mandibular fragment with P3-P4. Dentition slightly to moderately worn.
Left maxilla with P4-M2. Dentition slightly worn. Left C I. Moderately worn. 9
Right maxilla with MI-M 2. Dentition slightly worn; M ~ damaged. Right MI. Slightly worn.
Left I I. Unerupted.
K. w&keri
Indet. sp.
K. wickeri
K. wickeri
K. wickeri
K. wickeri
K. wickeri
K. wickeri
KNM-FT 2751 Right distal humerus. Indet. sp.
Right proximal ulna. Left C~. Heavily worn. 9 Right 12 . Slightly worn. Left I2 in a mandibular fragment. Crown slightly worn.
KNM-SO 1102 c) e) Right C1. Slightly worn. 9 Simiolus sp. L. legetet (7)
KNM-FT 3381 Indet. sp. KNM-FT 3636 K. wickeri KNM-FT 3637 K. wickeri KNM-MB 124 c) e) Simiolus sp.
a) Several specimens previously identified as fossil catarrhines from Fort Ternan are excluded from this list: KNM-FT 22 is now missing from the collections of the National Museums of Kenya; KNM-FT 3318, a right dC1, is almost certainly from the early Miocene locality of Songhor, rather than Fort Ternan, and can be attributed to Proconsul major (see also PICKrORD, 1985a); KNM-FT 3376, an isolated incisor, does not belong to a primate; b) the numbers in parentheses refer to the following primary references: (1) LEAKEY, 1962; (2) LEAKEY, 1967; (3) LEAKEY, 1968; (4) ANDREWS, 1971; (5) WALKER • ANDREWS, 1973; (6) ANDREWS & WALKER, 1976; (7) ANDREWS, 1978; (8) PICKFORD, 1985a; (9) HARRISON, 1986; (10) SENUT, 1989; C) KNM-FT 20, 21, 23, 24, 25, KNM-MB 124, and KNM-SO 1102 are almost certainly attributable to a single individual; d) KNM-FT 46a and KNM-FT 47 are clearly attributable to a single individual. However, the isolated canine, KNM-FT 46b, which is usually associated with KNM-FT 46a, is more likely to represent a different individual based on its differential preservation and its more pronounced degree of wear (see also PICKFORD, 1985a). KNM-FT 48 was considered by LEAKEY (1962) to belong to the same individual, but there is insufficient evidence to support such a claim (see also PICKEORD, 1985a); e) KNM-MB 124 and KNM-SO 1102 have been wrongly accessioned as having been recovered from Maboko and Songhor respectively. They are quite evidently associated with KNM-FT 20-25.

504 T. HARRISON
The intention of this paper is to provide an updated assessment of the taxonomic and phylogenetic status of the fossil catarrhine primates from Fort Ternan. In addition to Kenyapithecus wickeri, several other species of catarrhines have been identified from the small sample of fossil primates available. Although no new specimens have been recovered from Fort Ternan since the last expedition to the site in 1974, new discoveries from other Miocene localities in East Africa, as well as changes in our appreciation of the evolutionary history of Miocene catarrhines in general over the past few years, have necessitated a rein- terpretation of the fossil primates from the site. In addition to simply providing an updated taxonomy of the Fort Ternan primates, the results of this study have wider implications for understanding the phylogenetic, biogeographical, and ecological relationships of the catarrhine primates in East Africa during the middle Miocene.
THE FOSSIL CATARRH1NES FROM FORT TERNAN
During the course of excavations at Fort Ternan, between 1961 and 1974, a total of 38 catarrhine primates was recovered from the site (Table 1) 1). These can be attributed to at least five different species, identified here as Kenyapithecus wickeri, Proconsul sp., a large species of oreopithecid, Simiolus sp., and a species of small catarrhine primate of uncertain affinity. Most of the individual specimens discussed here have been described in previous publications (LEAKEY, 1962, 1967, 1968; ANDREWS, 1971, 1978; WALKER & ANDREWS, 1973; ANDREWS & WALKER, 1976; PICKFORD, 1985a; HARRISON, 1986; see Table 1), so no further detailed descriptions are required.
Kenyapithecus wickeri
The hypodigm of Kenyapithecus wickeri LEAKEY, 1962 currently comprises 15 cranio- dental specimens, all derived from Fort Ternan, of which the majority are isolated teeth (Table 1). Detailed descriptions and good illustrations of the most important specimens have been presented previously by ANDREWS and WALKER (1976) and by PICKFORD (1985a). The convoluted taxonomic history of Kenyapithecus (LEAKEY, 1962, 1967, 1968; SIMONS, 1963, 1969, 1972; SIMONS & PILBEAM, 1965; PILBEAM, 1969; ANDREWS, 1971, 1973, 1978; AGUIRRE, 1975; CONROY & PILBEAM, 1975; FRAYER, 1976; ANDREWS & WALKER, 1976; GREENFIELD, 1979; SZALAY & DELSON, 1979; MADDEN, 1980) has been succinctly summa- rized elsewhere (ANDREWS & MOLLESON, 1979; PICKFORD, 1985a).
The purported sister-species to K. wickeri, "Sivapithecus" africanus LE GROS CLARK and LEAKEY, 1950 (commonly included in the genus Kenyapithecus), from the middle Miocene localities of Maboko Island, Majiwa and Kaloma 2), is now much better known than the type species (PICKFORD, 1982, 1985a, b, 1986b, c; BENEFIT & MCCROSSIN, 1989). Further recent discoveries of Kenyapithecus have been reported from other middle Miocene sites
1) Abbreviations used in the text and Tables are as follows: FT: Fort Ternan; KNM: National Museums of Kenya; KR: Kapsibor; M: British Museum Natural History, London; MB: Maboko Island; M J: Majiwa and Kaloma; SO: Songhor; WK: Kalodirr. 2) The type specimen of Sivapithecus africanus (M 16649) was identified by LE GROS CLARK and LEAKEY (1950, 1951) tO have been recovered from Rusinga Island, although, in the initial description of the specimen, MACINNES (1943) made no reference to a specific locality. However, there is good circumstantial evidence to support the contention that M 16649 was, in fact, recovered from Maboko Island, rather than Rusinga Island (ANDREWS & MOLLESON, 1979).

Fossil Catarrhines from Fort Ternan 505
in Kenya, including Emuruilem in the Samburu Hills (PICKFORD, 1983b, 1986c; ISHIDA et al., 1984), Kaimogool North and Chepetet West, Nyakach (PICKFORD, 1986d) and Muruyur in the Baringo Basin (PICKFORD, 1988; BROWN et al., 1991), but no detailed infor- mation on this material is yet available. Preliminary reports suggest, however, that the material is morphologically most similar to "Sivapithecus" africanus from Maboko Island. Clearly, a reassessment of the taxonomic and phylogenetic status of Kenyapithecus must await the detailed comparison of these extensive new collections. In the meantime, however, it is worthwhile here to report briefly on my own analysis of the K. wickeri material.
The most recent review of the status of Kenyapithecus has been presented by PICKFORD (1982, 1985a, b, 1986b, c), and a number of important conclusions reached by him have been confirmed by the present study. These are as follows: (1) Kenyapithecus is best regard- ed as a valid genus, morphologically distinct from the more conservative Proconsul, as well as from the Eurasian genera, Dryopithecus and Sivapithecus (=Ramapithecus), with which it has been commonly synonymized; (2) all of the specimens from Fort Ternan previously attributed to Proconsul nyanzae (ANDREWS 8Z WALKER, 1976; ANDREWS, 1978) can more appropriately be assigned to K. wickeri; (3) K. wickeri is strongly sexually dimorphic, at least in canine size and morphology; (4) K. wickeri is specifically distinct from "S . " africanus, and may even require separation at the generic level; and (5) K. wickeri appears to be more specialized cranio-dentally than "S. "' africanus, with the former possibly shar- ing significant synapomorphies with modern large hominoids.
The last two of these conclusions imply that a major rethinking of the taxonomic and phylogenetic position of Kenyapithecus is called for. However, the implications of these finding have been largely overlooked by subsequent workers, who have preferred to recog- nize K. wickeri and "S. "" africanus as congeneric (or even conspecific), and to regard Kenyapithecus as a conservative thick-enamelled large hominoid of uncertain affinities (KELLEY • PILBEAM, 1986; MARTIN, 1986; ANDREWS 8~; MARTIN, 1987; DELSON, 1988; ANDREWS, 1988). Although I agree with PICKFORD that the inadequacy of the material dictates an element of caution in making phylogenetic statements about Kenyapithecus, I feel that there is sufficient evidence to confirm the conclusions outlined above, and even to develop them one step further.
The major differences in the dentition and lower facial morphology between the material from Fort Ternan and Maboko certainly provide justifiable grounds for a taxonomic sepa- ration between them (see Table 2 for details). The nature and distribution of the differences effectively rules out the possibility that they can be explained as the result of individual variation or sexual dimorphism (contra GREENFIELD, 1979; KAY & SIMONS, 1983). In my view, the differences, especially in the lower facial morphology, are profound enough to merit a generic separation between Kenyapithecus wickeri and "Sivapithecus" africanus. A more precise statement of their taxonomic status and affinities will have to await detailed comparisons with other Miocene hominoids, especially the thick-enamelled hominoids from Turkey and Central Europe (ABEL, 1902; STEININGER, 1967; ANDREWS t~ TOBIEN, 1977; ALPAGUT et al., 1990). Nevertheless, some preliminary conclusions can be presented here that may help in resolving the relationships of Kenyapithecus wickeri.
Kenyapithecus wickeri and "Sivapithecus" africanus share a number of derived charac- ters that serve to distinguish them from the more conservative pattern seen in Proconsul. These include a relatively thick mandibular corpus, a mandibular symphysis with a well- developed inferior transverse torus, large and quite narrow upper premolars with the

506
Table 2. Morphological characters distinguishing Kenyapithecus africanus*.
wickeri from
Y. HARRISON
" Sivapithecus"
Element Specimens Distinguishing features of K. wickeri I l FT 49 Relatively broader and lower-crowned; more pronounced lingual cingulum;
lack of a distinct lingual pillar; more deeply furrowed lingual face. 12 FT 3 6 3 7 Relatively broader; more coarsely wrinkled lingual surface. C 1 FT 8, 39, 46b Larger in females (the basal area of FT 46b is 2507o larger than the largest
female canine from Maboko); crown less bilaterally compressed. p4 FT 46a Relatively slightly broader; better-defined transverse crests linking the para-
cone and protocone; no lingual cingulum; less-complex wrinkling of the enamel surface.
Upper molars FT 46a, 47 Taper slightly distally, giving the crown a more trapezoidal (rather than rectangular) shape; relatively narrower (the length-breadth proportions for M 1 and M 2 are 95.5 and 99.2 respectively, while those from Maboko range from 87.6 - 94.7 and 90.7 - 92.6 respectively); lack of secondary wrinkling; almost complete suppression of the lingual cingulum; slightly larger average size.
C1 FT 28, 3636 Female canines slightly larger in size. Lower molars FT 7, 34, 48 Slightly larger in size; relatively narrower (at least in M0; M3 hypoconulid
arranged more in line with other buccal cusps. Maxilla FT 46a, 47 Root of the zygomatic process originates higher on the face, and is more
strongly laterally flaring and upwardly sweeping; canine fossa more distinct; maxilla much less pneumatized in the region above the premolars; maxillary sinus more elevated, with its floor situated well-above the apices of the molar roots; palate relatively deeper.
*For a more detailed discussion of many of these features, see ANDREWS and WALKER (1976), WARD and PILBEAM (1983), KAY and SiraONS (1983), and PICKFORD (1985a, 1986c).
pa racone on ly s l ight ly more elevated than the pro tocone , mola r s with th ick enamel , low occlusal rel ief and reduced deve lopment o f the cingula. However, this c o m b i n a t i o n o f shared der ived features is also seen in o ther la ter Miocene homino ids , such as Sivapithecus, Gigantopithecus, Lufengpithecus, and Ouranopithecus, as well as in ear ly homin ids , and may poss ib ly represent a suite o f features pr imi t ive ly re ta ined f rom the ancestral large h o m i n o i d m o r p h o t y p e (MARTIN, 1983, 1986; ANDREWS & MARTIN, 1987).
Whi le "Sivapithecus" africanus retains some conservat ive t rai ts in its m o l a r m o r p h o l o g y and facial pa t t e rn tha t more closely approx imates the more general ized ca ta r rh ine m o r p h o l o g y exempli f ied by Proconsul, K. wickeri has deve loped some dist inct ive special i- za t ions which it shares with la ter large homino ids . As no ted by a number o f authors , the den t i t ion o f K. wickeri is very s imi lar to tha t o f Sivapithecus f rom the Siwalik Hil ls (PILBEAM, 1969; SIMONS, 1981; KAY t~r SIMONS, 1983; KELLEY t~ PILBEAM, 1986), and it is d i f f icul t to iden t i fy consis tent characters tha t would serve to d is t inguish them. Moreover, K. wickeri shares the dist inct ive der ived uppe r canine m o r p h o l o g y o f Sivapithecus, in which the roo t is more med ia l ly incl ined, external ly ro ta ted and q u a d r a n g u l a r in cross- sect ion (ANDREWS t~ WALKER, 1976; WARD • PILBEAM, 1983; PICKFORD, 1985a). In the p l acemen t and o r i en ta t ion o f the zygomat ic process o f the maxi l la , the presence o f a well- developed canine fossa, the lack o f p n e u m a t i z a t i o n o f the an te r io r a lveolar region o f the maxi l la , the conf igura t ion o f the max i l l a ry sinus and the dep th o f the palate, K. wickeri confo rms closely to the der ived pa t t e rn typica l o f Sivapithecus and Pongo. It seems l ikely tha t these specia l iza t ions in facial m o r p h o l o g y in K. wickeri relate to increasing the robus t ic i ty o f the max i l l a in the region o f the cheektee th and to improving the mechan ica l effectiveness o f the mas t i ca to ry muscula ture , bo th associa ted func t iona l ly wi th powerful chewing. A l t h o u g h the der ived h o m i n o i d features shared by K. wickeri and Sivapithecus

Fossil Catarrhines from Fort Ternan 507
may possibly be interpreted as functional convergence, it seems most parsimonious at present to assume that the detailed similarities in the dentition and facial morphology are homologous. These shared characteristics could be used to support the claim that Kenyapithecus represents a conservative member of the Pongo-Sivapithecus clade. However, it would appear more likely, based on comparisons with other extant hominoids, that the features shared by Kenyapithecus and Sivapithecus are, in fact, primitive retentions from the ancestral large hominoid morphotype (see also BROWN & WARD, 1988). Clearly, more detailed comparisons between Kenyapithecus and other later Miocene large homi- noids, such as Ouranopithecus, Dryopithecus, and Lufengpithecus are needed in order to resolve this issue. At present, I would favor retaining Kenyapithecus as a distinct genus that represents the conservative sister-taxon of the extant large hominoids. "Sivapithecus" africanus, which appears to share only primitive traits with the latter clade, should there- fore be included in a separate genus. However, the decision over the correct taxonomic sta- tus of "S. "' africanus must await careful comparative studies of the Maboko material with the samples from Pasalar in Turkey and the conservative thick-enamelled hominoids from the Vienna Basin (ABEL, 1902; STEININGER, 1967; ALPAGUT et al., 1990).
Proconsul sp.
A number of workers have noted that several isolated teeth from Fort Ternan are similar in their morphology to the corresponding teeth of Proconsul africanus, and that they may be attributed provisionally to this species (LEAKEY, 1968; ANDREWS & WALKER, 1976; ANDREWS, 1978; PICKFORD, 1985a, 1986b). My comparisons confirm that at least two isolated teeth, a left M 1 (FT 16) and a left p3 (FT 29) are morphologically and metrically consistent with Proconsul africanus from the early Miocene of East Africa. The upper molar can be distinguished from those of K. wickeri in retaining a well-developed lingual cingulum and what appears to be relatively thin enamel, while the premolar exhibits such conservative traits as a relatively broad crown and a marked difference in height between the protocone and paracone. Clearly, these teeth belong to a medium-sized proconsulid rather than Kenyapithecus, but beyond this there is insufficient evidence to provide a reasonable basis for the assessment of their specific affinities. They are certainly most like the early Miocene species Proconsul africanus, but as the hypodigm of this latter taxon is currently in a state of flux (KELLEY, 1986; KELLEY & PILBEAM, 1986; PICKFORD, 1986e; TEAFORD et al., 1988; WALKER & TEAFORD, 1988, 1989; RUFF et al., 1989; CAMERON, 1991), it may be preferrable to consider the Fort Ternan specimens as belonging to an indeter- minate species of Proconsul.
Oreopithecids
LEAKEY (1967, 1968, 1969) noted the presence of a large species of oreopithecid at Fort Ternan, based on three isolated teeth, a right M 1 (FT 36), a right p4 (FT 37), and a right M3 (FT 38). LEAKEY suggested that the specimens closely resembled Oreopithecus bambolii from the late Miocene of Italy, and tentatively referred the material to the same genus. Additional comparisons by SIMONS (1969, 1972) led him to conclude that the Fort Ternan material may even be attributable to the same species as the European taxon. However, the occurrence of oreopithecids at Fort Ternan was contested by later workers, who concluded that the specimens probably belonged to a suid (ANDREWS & WALKER, 1976). A re-examination of the material by the author has confirmed earlier assessments that these specimens do in fact represent isolated teeth of an oreopithecid primate

508 T. HARRISON
(HARRISON, 1986). In addition, a single isolated upper molar (KR 9755) recovered by PICKEORD at the neighboring and contemporaneous locality of Kapsibor (HARRISON, 1986; PICKFORD, 1986d) provides additional evidence that a large oreopithecid occurred in the Fort Ternan sediments. The concordance in size and morphology of these isolated teeth in- dicates that they can all be referred to a single species. The Fort Ternan specimens are larger and more derived than the middle Miocene oreopithecid, Nyanzapithecus pickfordi from Maboko Island (HARRISON, 1986), and are much more similar to the corresponding teeth of O. bambolii from Italy. HARRISON (1986) has argued that the degree of resemblance to 0. bambolii is sufficient to place the Fort Ternan material in the same genus, at least provi- sionally, until more complete material is obtained from the middle Miocene of East Africa.
Simiolus sp.
In 1968, LEAKEY announced the discovery of two mandibular fragments and some isolated teeth belonging to a small catarrhine primate from Fort Ternan (LEAKEu 1968). Although he was uncertain of its true affinities, suggesting a possible relationship to Pliopithecus from Europe, LEAKEr tentatively assigned the material to an unknown species of Limnopithecus. SIMONS (1969), who provided a brief review of the Fort Ternan primates, regarded LEAKEY'S cautious approach as unnecessary, owing to SIMONS' earlier synonymy of Limnopithecus with Pliopithecus (SIMONS, 1963; SIMONS & PILBEAM, 1965). S~MONS (1969) concluded that the differences between the Fort Ternan small catarrhine and Pliopithecus antiquus were likely to be at a level no greater than that between species of a single genus, and consequently referred the material to Pliopithecus sp.
ANDREWS and WALKER (1976) presented a more detailed discussion of the small catarrhine primate specimens from Fort Ternan. They suggested that the dentition was less specialized than that of Dendropithecus rnacinnesi and Pliopithecus antiquus, and that, on the basis of its size and more generalized morphology, it should tentatively be referred to Lirnnopithecus legetet. This assessment was further supported by ANDREWS (1978) in his revision of the East African Miocene hominoids, and followed by a number of subsequent authors (e.g. SZALAV & DELSON, 1979; SHIPMAN et al., 1981). More recently, however, ANDREWS (1980) has concluded that the affinities of the material may be with Den- dropithecus rather than Limnopithecus.
A detailed study of the small catarrhines from Fort Ternan by the author suggests that more than one species is represented in the collections. Unfortunately, the material is much too fragmentary to be certain just how many taxa are actually represented, but there are clear indications that at least two species are present. One species is quite well-represented, including several partial mandibles (FT 13 and FT 14), an almost complete mandibular toothrow (FT 20-25, MB 124, and SO 1102), and a single isolated tooth (FT 19) (see Table 1 for details).
As discussed above, several authors have made reference to this material (LEAKEr, 1968; ANDREWS & WALKER, 1976; ANDREWS, 1978), but no details have yet been published. A brief description of each specimen is therefore provided.
FT 20-25 is the most complete specimen, consisting of a crushed and fragmentary mandi- ble with C-M3 on the left side and some associated lower teeth from the right side (I2, P4, M2, and M3). In addition, an isolated left I2, MB 124, and a right lower canine, SO 1102, incorrectly accessioned as having been recovered from Maboko Island and Songhor respec- tively, are identical in morphology and preservation to the antimeres in the associated

Fossil Catarrhines from Fort Ternan 509
mandibular toothrow, and almost certainly belong with this individual. The dentition is moderately worn and is generally well-preserved.
Unfortunately, the mandibular fragment, KNM-FT 20, is too incomplete and too poorly- preserved to yield any pertinent information about the morphology of the mandible.
The I2 is narrow and gracile, and moderately high-crowned. The crown is bilaterally strongly asymmetrical, with a straight and steep mesial margin and an angular distal mar- gin. In addition, the apico-basal axis of the crown is set at a slightly oblique angle to the long-axis of the root, thereby producing a noticeable curvature of the root and crown towards the mesial aspect. The incisive apex is relatively narrow. A poorly-defined and rounded cingulum is present at the base of the lingual face of the crown. Originating from the mesial portion of the lingual cingulum, and terminating close to the incisive apex of the crown, is a distinct lingual pillar.
The lower canine is moderately low-crowned and gracile. The mesial ridge is long and sharp, and runs from the apex of the crown to the mesial termination of the lingual cingu- lure. The latter is narrow, rounded, and rather ill-defined. The distal ridge, which descends from the apex of the crown, is low and rounded, and it continues basally on to a short distal heel.
The P3 is low-crowned, mesio-distally quite elongated and slightly bilaterally com- pressed. The honing face on the mesiobuccal aspect of the crown, for occlusal contact with the upper canine, is short and steeply inclined, with only a slight extension of enamel onto the base of the mesial root. The morphology and size of the canine and P3 confirm that this individual can be identified as a female.
The P4 is mesiodistally long and narrow, relatively low-crowned and ovoid in occlusal outline. The protoconid is low and voluminous, while the metaconid is restricted to a small, marginally placed tubercle. The two cusps are set close together and are joined by an elevat- ed transverse crest, the protocristid. The mesial fovea is quite small and restricted. Posteri- orly there is a shallow and poorly-defined talonid basin. Only a slight trace of a buccal cingulum is present on the distobuccal margin of the crown.
The M1 is long and narrow, and ovoid in occlusal outline. The cusps are quite low and rounded, and their bases restrict the extent of the occlusal basins. The cristids linking the main cusps are rather poorly-developed, and are interrupted between the cusps by fine grooves. The protoconid is situated slightly more mesially than the metaconid, so that the protocristid is directed somewhat obliquely relative to the long-axis of the crown. The mesial fovea is confined to a small pit-like depression. The talonid basin is shallow, triangu- lar in shape, and quite restricted in its extent. The hypoconulid is well-developed and is situ- ated slightly towards the buccal side of the median-axis of the crown. The distal fovea is quite large and well-defined. The buccal cingulum is narrow and rounded, forming a con- tinuous ledge around the buccal margin of the crown, but is best developed in the intervals between the buccal cusps.
The M2 is similar in morphology to M1, but differs in being larger in size, and in having a more extensive talonid basin and broader foveae. As in Ml, the crown is long and nar- row, and is ovoid in occlusal outline. The cusps are low, rounded, and voluminous, and the cristids are poorly-developed. The mesial fovea is relatively broader and more transversely aligned than in M1. The extent of the buccal cingulum is similar to that seen in M j, except that it is better-developed around the protoconid and hypoconid. The groove system dissecting the talonid basin is simple, and there is no development of secondary wrinkling of the enamel surface.

510 T. HARRISON
The M3 is smaller than M2 (its occlusal area is 16.7% smaller than M2), but is slightly larger (9.2% larger) than M1. It is long and narrow, ovoid in occlusal outline, and tapers posteriorly to a bluntly rounded distal margin. The cusps are low, rounded, and voluminous. The mesial fovea is broad and transversely aligned. The talonid basin is shal- low, and quite long, and narrow. The three main buccal cusps are arranged linearly. The distal fovea is well-defined, but relatively small in size. The buccal cingulum is moderately well-developed, forming an almost continuous ledge around the buccal aspect of the crown.
A single isolated M 3, FT 19, is the only upper molar that can be attributed to this species. The crown is relatively short and very broad, with a length-breadth index of 73.8. The protocone is low and voluminous, while the paracone is higher and more conical. The metacone and hypocone are relatively reduced in size. The crista obliqua, linking the proto- cone to the metacone, is low but well-developed. A low and poorly-defined subsidiary crest runs anterior to and parallel with the crista obliqua. There are no crests linking the hypo- cone to the protocone or metacone. The lingual cingulum is moderately well-developed and forms an L-shaped shelf of enamel around the protocone. There is no evidence to confirm the suggestion that FT 19 belongs to the same individual as FT 20-25, although the wear and preservation are certainly consistent with such an association (ANDREWS & WALKER, 1976).
FT 14 consists of a fragment of a mandible of an immature individual, in which the germ of the first permanent molar has been exposed in its crypt. With M1 unerupted, the speci- men is likely to have retained a full deciduous dentition at the time of its death. The molar crown is incompletely formed, but well-preserved. The morphology of M1 is very similar to that in FT 20-25, and it is fairly certain that these specimens belong to the same species.
FT 13 comprises an edentulous symphyseal fragment of an immature individual, preserv- ing the alveoli for the lower deciduous incisors and canines. Small openings for the gubenacular canals are retained along the lingual side of the alveolar margin of the mandi- ble. The symphysis of the mandible is relatively low and quite robust. The internal surface of the symphysis is buttressed by a rounded superior transverse torus only. An ill-defined mental spine is preserved at the inferior margin of the midline of the symphysis. As the mandible is comparable in size and morphology with FT 14, which is presumably of similar dental age, it seems reasonable to assign FT 13 to the same species.
Initial comparisons of the small catarrhine primates from Fort Ternan led the author to conclude that they could readily be distinguished from all other taxa from the Miocene of East Africa (HARRISON, 1982). However, renewed study has shown that the material is very similar in morphology to the recently described early Miocene species, Simiolus enjiessi, from Kalodirr in northern Kenya (LEAKEY & LEAKEY, 1987). The distinctive features of the Fort Ternan material that it shares uniquely or in combination with Simiolus enjiessi are as follows: (I) the lower incisors are relatively high and narrow, and are quite small in relation to the size of the molars; (2) P3 is low-crowned, moderately bilaterally com- pressed, with a short and steep honing face extending only a short distance onto the lingual aspect of the mesial root; (3) P4 is moderately elongated, with the two main cusps closely approximated; and (4) the lower molars are relatively long and quite narrow, and ovoid rather than rectangular in occlusal outline, with moderately well-developed cristids (some- what obscured by wear in the Fort Ternan specimens), well-defined, but restricted mesial and distal foveae and talonid basins, and distinct buccal cingula. It is these characteristics of the lower dentition, especially the distinctive features of the lower molars, that provide support for the attribution of the Fort Ternan specimens to the genus Simiolus (Fig. 1).
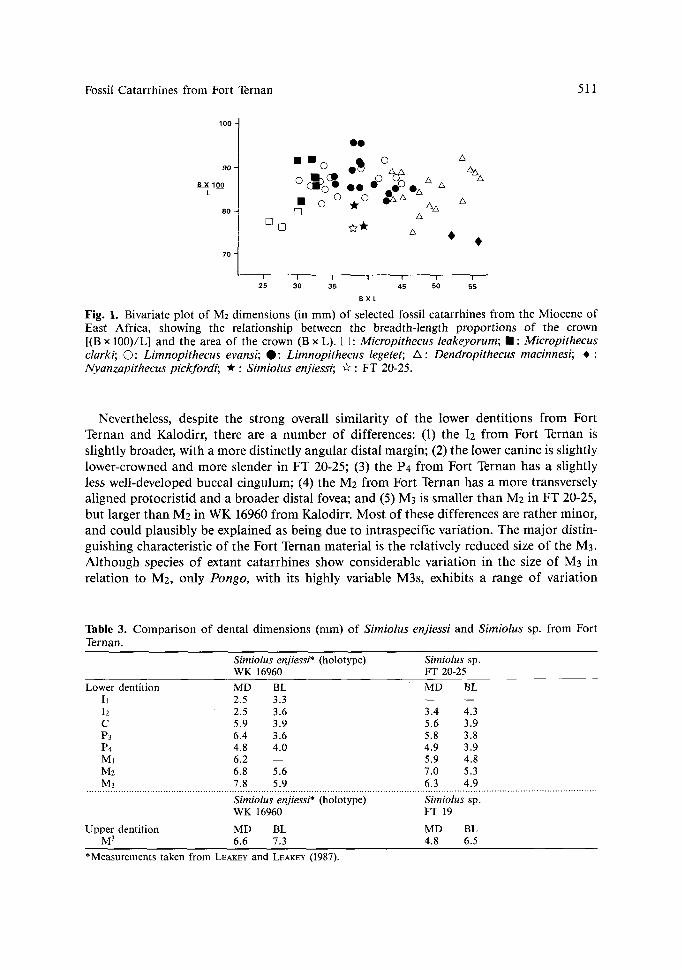
Fossil Catarrhines from Fort Ternan 51 1
100
9 0 "
s x 100 L
80-
7 0 -
oo z~
�9 �9 o o %,
�9 0 0 . 0 ~ a [] Zx
i I
25 3'0 ;5 ' "5 5o B X L
Fig. 1. Bivariate plot of M2 dimensions (in mm) of selected fossil catarrhines from the Miocene of East Africa, showing the relationship between the breadth-length proportions of the crown [(B x 100)/L] and the area of the crown (B x L). [] : Micropithecus leakeyorum; �9 : Micropithecus clarki; �9 Limnopithecus evansi; O: Limnopithecus legetet; A : Dendropithecus macinnesi; * : Nyanzapithecus pickfordi; * : Simiolus enjiessi; ~" : FT 20-25.
Nevertheless, despi te the s t rong overall s imi lar i ty o f the lower dent i t ions f rom For t Ternan and Kalodirr , there are a number o f differences: (1) the I2 f rom For t Ternan is s l ight ly broader , wi th a more d is t inc t ly angu la r dis ta l margin; (2) the lower canine is s l ightly lower-crowned and more s lender in F T 20-25; (3) the P4 f rom For t Ternan has a sl ightly less wel l -developed buccal c ingulum; (4) the M2 f rom For t Ternan has a more t ransversely a l igned p ro toc r i s t id and a b r o a d e r dis tal fovea; and (5) M3 is smal ler t han M2 in F T 20-25, bu t larger t han M2 in W K 16960 f rom Kalodirr . Mos t o f these differences are ra ther minor, and could p l aus ib ly be expla ined as being due to in t raspeci f ic var ia t ion . The m a j o r dist in- guishing character is t ic o f the For t Ternan mate r ia l is the relat ively reduced size o f the M3. A l t h o u g h species o f extant ca ta r rh ines show cons iderab le va r ia t ion in the size o f M3 in re la t ion to M2, on ly Pongo, with its h ighly var iable M3s, exhibits a range o f var ia t ion
Table 3. Comparison of dental dimensions (mm) of Simiolus enjiessi and Simiolus sp. from Fort Ternan.
Simiolus enjiessi* (holotype) Simiolus sp. WK 16960 FT 20-25
Lowerdentition MD BL MD BL I1 2.5 3.3 - - - - I2 2.5 3.6 3.4 4.3 C 5.9 3.9 5.6 3.9 P3 6.4 3.6 5.8 3.8 P4 4.8 4.0 4.9 3.9 MI 6.2 - - 5.9 4.8 M2 6.8 5.6 7.0 5.3 M3 7.8 5.9 6.3 4.9
Simiolus enjiessi* (holotype) Simiolus sp. WK 16960 FT 19
Upper dentition MD BL MD BL M 3 6.6 7.3 4.8 6.5
*Measurements taken from LEAKEY and LEAKEY (1987).

512 T. HARRISON
Table 4. Relative size of M3 a) in selected extant catarrhines and in Simiolus from Kalodirr and Fort Ternan.
Species N X Range Pan troglodytes 19 107.1 88.0 - 116.5 Gorilla gorilla 28 101.2 81.8 - 110.5 Pongo pygmaeus b) 25 104.0 79.2- 127.2 Hylobates lar 36 93.0 77.2 - 107.5 Symphalangus syndactylus 24 94.7 71.0- 108.7 Cercopithecus aethiops 26 95.3 84.1 - 107.0 Colobus abyssinicus 36 117.0 94.0 - 129.4 Fort Ternan 1 120.1 Simiolus enjiessi 1 82.8
M2 Breadth x Length x 100 a) Index of relative M3 size = M3 Breadth x Length ing individuals with pathological or supernumary molars.
; b) using data obtained from HOO1JER (1948), exclud-
sufficient to encompass the magnitude of difference exhibited by FT 20-25 and WK 16960 (see Tables 3 & 4). This difference probably provides a good basis for recognizing at least a species distinction between the two samples. Since most early fossil catarrhines, with the notable exception of Micropithecus clarki, have an M3 that is quite considerably larger than M2, as in Simiolus enjiessi, its small size in the Fort Ternan specimen would appear to represent a derived condition with respect to its earlier sister taxon.
Unfortunately, comparisons with the upper dentition of Simiolus from the early Miocene is confined to an isolated molar from Fort Ternan. The M 3 differs from the corresponding tooth from Kalodirr in being mesiodistally shorter and smaller in size. However, this rela- tionship is not unexpected given the relative size of M3. The reduction of the third molar in the Fort Ternan specimen, in conjunction with the other minor morphological differ- ences, probably provides sufficient grounds for recognizing a species distinction. However, as the samples from both Kalodirr and Fort Ternan are rather limited, it seems most judi- cious at present to await the recovery of more material before establishing a new taxon.
It can be noted in passing that, apart from a superficial similarity in the proportions of the lower molars, the material from Fort Ternan bears none of the distinctive features of the European pliopithecids to indicate that a close phyletic relationship may exist between them (HARRISON, 1982).
Indeterminate Material
As discussed above, comparisons suggest that a second species of small catarrhine, slight- ly larger than Simiolus sp., is present at Fort Ternan. This species is represented by three specimens only, an isolated upper incisor (FT 11), an upper canine (FT 15), and an eden- tulous mandibular symphysis (FT 12), that can be grouped together largely on the basis of size.
The isolated I 1, FT 11, is high-crowned and slender. Lingually there is a narrow cingu- lum at the base of the crown, which is uplifted centrally at its junction with a fine lingual pillar. The buccal surface is evenly convex and finely crenulated. FT 11 appears to be too large to be included in the same species as FT 20-25, and is morphologically inconsistent with the narrow and relatively small lower incisors in the latter. It is most similar in size and morphology to upper central incisors of Kalepithecus songhorensis from the early Mio-

Fossil Catarrhines from Fort Ternan 513
cene of East Africa, but differs in being slightly narrower and somewhat higher-crowned (see HARRISON, 1989a).
Similarly, the isolated upper canine, FT 15, is also too large to be included in the same species as FT 20-25. As it is a relatively low-crowned and robust canine, quite clearly from a female individual, the metrical differences cannot be explained as due to sexual dimor- phism. Just as in FT 11, the canine is similar to corresponding canines attributed to Kalepithecus songhorensis.
FT 12 consists of an edentulous mandibular symphyseal fragment. The symphysis is deep and robust, and is buttressed internally by a prominent superior transverse torus, with no development of an inferior torus. Judging from the size of the mandibular fragment, the specimen would have belonged to a catarrhine primate of similar cranio-dental size to K. songhorensis, and is therefore metrically consistent with FT 11 and FT 15. The size differ- ence between this specimen and those from Fort Ternan attributed to Simiolus are unlikely to be a function of sexual dimorphism. When compared with the alveoli for the incisors, the canine roots in FT 12 are relatively smaller than in FT 20-25. This relationship between incisor and canine size is inconsistent with the proposal that the two specimens may represent a male and female of the same species.
It seems likely, therefore, that FT 11, FT 12, and FT 15 belong to a single species that can be distinguished from the taxon typified by FT 20-25 mainly by its larger size. Morpho- logically and metrically the former specimens are very close to the material from the early Miocene localities of Koru and Songhor attributed to Kalepithecus songhorensis (see HARRISON, 1989a). This may indicate that Kalepithecus, like Simiolus, extended temporal- ly into the middle Miocene. However, until further material is available to reliably assign these specimens to a particular taxon they are regarded here as of indeterminate status.
In addition, there are two isolated teeth, FT 17 and FT 18, which cannot at present be attributed to either of the two species of small catarrhines so far discussed. FT 17 consists of a well-preserved and unworn isolated M1. It differs from the M1 in FT 20-25 in the following respects: (1) it is slightly larger in size; (2) it is higher-crowned and relatively broader; (3) the cusps are more elevated and more conical; (4) the mesial fovea is larger and more transversely aligned; (5) the distal fovea is more restricted; (6) the talonid basin is broader and deeper; (7) the cristids are less elevated; and (8) the buccal cingulum is not continuous around the hypoconid. These differences are sufficient to indicate that FT 17 does not belong to the same species as FT 20-25. A comparison of the isolated molar with other fossil East African catarrhines shows that it is comparable in size, and has its closest affinities, with Dendropithecus macinnesi from Rusinga Island. It is conceivable that the specimen belongs with FT II, FT 12, and FT 15, which are of the appropriate size. If this is the case, then the suggested possible relationship with Kalepithecus would need to be reconsidered.
FT 18 consists of an isolated P4 of uncertain taxonomic status. The crown is small, almost circular in occlusal outline, with a simple morphology. The protoconid is voluminous, with short, but well-developed mesial and distal cristids. The metaconid is much smaller than the protoconid, more distally placed and lacks cristids mesially and distally. The two cusps are connected by a low transverse crest. The buccal cingulum is moderately well-developed. The distal margin of the crown is raised and bears a small stylid at its junction with the distal crest of the protoconid. The specimen is smaller in size and morphologically quite distinct from the P4 in FT 20-25, and is unlikely to belong to the same species. It is also much too small to assign to the Kalepithecus-sized taxon from Fort

514 T. HARRISON
Ternan. Metrically the specimen is comparable to the corresponding tooth in Limnopithe- cus spp. and M. clarki from the early Miocene of East Africa, but it differs in the shape of the crown and in its detailed morphology.
An isolated left upper canine, FT 41, of a medium-sized catarrhine has proved difficult to assign to any of the species currently recognized from Fort Ternan. It is a large, distally recurved and blade-like tooth, with a strongly bilaterally compressed crown, and a double mesial groove. It is of the appropriate size to belong to a male individual of Kenyapithecus wickeri or Proconsul sp., but its rather specialized morphology would tend to argue against such an attribution. It is similar in overall size and morphology to Rangwapithecus gordoni, but differs in having a double mesial groove, a feature seen consistently only in male canines of D. macinnesi among the early Miocene catarrhines (HARRISON, 1982, 1986, 1989a; KELLEY, 1986). A single upper canine from Rusinga Island traditionally included in R. gordoni, does possess a double mesial groove, but in the absence of other specimens clearly attributable to Rangwapithecus from Rusinga, this isolated specimen is more suita- bly assigned to Nyanzapithecus vancouveringorum (see also KELLEY, 1986). The Fort Ternan canine is structurally very similar to those of male individuals of D. macinnesi, but it is considerably larger. It is also very similar in size and morphology to two isolated canines from Maboko Island (MB 9763 and MB 11643) of uncertain taxonomic status (HARRISON, 1986), but which are conceivably attributable to Nyanzapithecus pickfordi. The similarity of FT 41 to male canines of Rangwapithecus, and possible also to Nyan- zapithecus, may suggest that the Fort Ternan specimen belongs to the large species of oreopithecid that is already known from the site. Such an assignment would be reasonable on the basis of size. However, until the upper canine morphology of male individuals of Nyanzapithecus is ascertained with some confidence, or until more complete material is obtained from Fort Ternan, I prefer to regard this specimen as of indeterminate status.
A few rather fragmentary postcranial remains of primates are known from Fort Ternan, including a right distal humerus (FT 2751) and a right proximal ulna (FT 3381). The func- tional and phylogenetic affinities of the distal humerus have already been considered in some detail by previous workers (PREUSCHOFT, 1973; MCHENRY & CORRUCCINI, 1975; CORRUCCINI et al., 1976; ANDREWS & WALKER, 1976; FELDESMAN, 1982; MORBECK, 1983; ROSE, 1983; SENUT, 1982, 1989; PICKFORD, 1985a). The specimen was identified initially as belonging to Proconsul (PREUSCHOFT, 1973; ANDREWS & WALKER, 1976; SENUT, 1982), but most subsequent authors have favored attribution to Kenyapithecus (PICKFORD, 1985a; KELLEY & PILBEAM, 1986; SENUT, 1989). This latter assessment seems most likely given the size and morphology of the specimen, but it cannot be entirely ruled out that it may belong to the oreopithecid from the site, which is of comparable general size (see also PICKFORD, 1985a). The proximal ulna has been briefly discussed by a number of authors (RosE, 1983; KELLEY & PILBEAM, 1986; SENUT, 1989). Its larger size and its distinctive morphology are inconsistent with its attribution to the same species as FT 2751. It is possible that FT 3381 may be attributed to an oreopithecid, but I am more inclined to agree with PICKFORD (cited in SENUT, 1989) that the specimen probably belongs to one of the large creodonts represented by cranio-dental remains at Fort Ternan (see Table 5).
It should also be noted that LEAKEY (1968) briefly made reference to the occurrence of isolated teeth of at least two types of cercopithecids at Fort Ternan. However, this claim was presumably based on mistaken field identifications as no monkeys have been recog- nized subsequently in the collections from the site.

Fossil Catarrhines from Fort Ternan 515
ECOLOGICAL AND BIOCHRONOLOGICAL RELATIONSHIPS OF THE CATARRHINE COMMUNITY FROM FORT TERNAN
Comparisons of the primate faunas from the early and middle Miocene sites in East Africa indicate that the structure of the catarrhine community changed quite dramatically during the early part of the Miocene (ANDREWS, 1981; HARRISON, 1989b). This change appears to be correlated with a local habitat shift from one dominated by tropical forest and closed woodlands to one that included a greater proportion of more seasonal open woodland. Several authors have proposed that this change in the community structure of the East African catarrhines can be linked directly to the rise of the Old World Monkeys (KRANTZ, 1975; ANDREWS, 1981, 1982; TEMERIN • CANT, 1983; MALONE, 1988). They con- tend that increased levels of competition from cercopithecids, which were apparently much better suited ecologically to take advantage of the expanding open country conditions dur- ing the middle Miocene, led to a sharp decline in the species diversity of early hominoids and other non-cercopithecoid catarrhines. However, the community transformation appears to be much more complex than a simple replacement by more competitive primate taxa. As I have argued previously, based on a detailed taxonomic analysis of the primate fauna from Maboko Island, the middle Miocene catarrhine community differs from the early Miocene community in its overall taxonomic and ecological composition, rather than in its species diversity (HARRISON, 1989b). In fact, the actual number of catarrhine species at middle Miocene localities is comparable to that at early Miocene sites, and there is no direct evidence to indicate that competition from early cercopithecids had any appreciable impact on species diversity at this time. Based on my previous analysis of the fossil primates from Maboko Island (HARRISON, 1989b), I have suggested that the middle Miocene primate community comprised a greater number of species that were capable of processing leaves and other fibrous materials, and that the replacement (or at least partial replace- ment) of the early Miocene community was primarily the result of changes in the ecology from a rainforest/woodland mosaic to one consisting of more seasonal open woodland.
The reassessment of the taxonomy of the Fort Ternan primates presented here certainly provides further evidence to confirm this model of community change. Although only 38 primate specimens are known from Fort Ternan they belong to at least 5 different species. This is a level of diversity that is comparable to or approaches that at the major early Miocene localities from which much larger samples are available. In addition, the primate species at Fort Ternan, like those at Maboko Island, were apparently adapted to exploit leaves and other fibrous plant materials, as well as more seasonally available food resources.
In the past, like most previous authors, I have considered the Fort Ternan primate fauna to show strong similarities to the Maboko fauna, when compared with the early Miocene primate communities. In a very general sense, this still remains valid. At both sites thick- enamelled hominoids and oreopithecids are quite common, while proconsulids have under- gone a relative decline. However, the revised taxonomy of the Fort Ternan primates present- ed here clearly shows that the detailed composition of the community (i.e. at the alpha-taxonomic level) is strikingly different from that from Maboko Island. In fact, there are no species or genera in common (see Table 5). Moreover, the Fort Ternan primate fauna contrasts with that from Maboko Island in the complete absence of cercopithecids, which are extraordinarily common at Maboko, and in the rarity of primates in general, relative

516 T. HARRISON
Table 5. Comparison of the mammalian faunas from Fort Ternan and Maboko Island, Kenya. Orders Maboko island Shared species & genera Fort ternan
Insectivora Amphechinus rusingensis Amphechinus Macroscelidea
Primates Micropithecus leakeyorum Limnopithecus sp. Nyanzapithecus pickfordi Mabokopithecus clarki Victoriapithecus macinnesi "Sivapithecus" africanus
Lagomorpha Kenyatagomys sp. Rodentia Megapecletes sp.
Zenkerella Paraphiomys
Creodonta Anasinopa leakeyi Dissopsalis pyroclasticus Hyaenodon andrewsi Hyainailouros nyanzae
Carnivora
Hyracoidea Tubulidentata Proboscidea
Perissodactyla
Artiodactyla
Megalohyrax championi Orycteropus chemeldoi Prodeinotherium hobleyi Protanancus macinnesi Choerolophodon kisumuensis Chilotheridium pattersoni Dicerorhinus leakeyi Brachypotherium heinzelini ?Kenyapotamus sp. Diamantohyus africanus Libycochoerus khinzikebirus Listriodon akatikubas Lopholistriodon moruoroti Dorcatherium spp. Canthumeryx sirtensis Nyanzameryx pickfordi Climacoceras africanus
Amphechinus sp. Miorhynchocyon rusingae Myohyrax oswaldi Pronasilio ternani Simiolus sp. Proconsul sp. Kenyapithecus wickeri
Megapedetes sp.
Paraphiomys ?Anasinopa leakeyi
Hyainailouros
Orycteropus chemeldoi Prodeinotherium hobleyi
Choerolophodon Chilotheridium
? Kenyapotamus
Listriodon akatikubas
Dorcatherium
Climacoceras
Megapedetes sp. ? Paranomalurus Paraphiomys ?Anasinopa leakeyi
Hyainailouros sp. ? Megistotherium Percrocuta tobieni Genetta sp.
Orycteropus chemeMoi Deinotherium sp.
Choerolophodon ngorora Chilotheridium sp. Paradiceros mukirii
Kenyapotamus ternani
Listriodon akatikubas
Dorcatherium spp.
Climacoceras gentryi Palaeotragus primaevus Samotherium africanum Protragoceras labidotus Oioceras tanycerus Caprotragoides gentryi Gazella sp.
Sources: LAVOCAT (1978); SAVAGE (1978); HARR/S (1978); SnWMAN et al. (1981); BUTLER (1984); P/CKFORD (1986d); SHIPMAN (1986).
to other mammals at the site (0.9% of all mammal specimens at Fort Ternan compared with 37.5% of all mammals at Maboko Island). These differences may imply that there is a significant ecological and/or chronological distinction between the two sites.
The suggestion of a temporal/ecological difference between Maboko and Fort Ternan is further supported by comparisons of the entire mammalian faunas. Of the 25 species and 31 genera definitively identified from Maboko Island, only 2 species (8.0%) and 10 genera (32.3%) are shared with Fort Ternan (see Table 5). This represents a very low level of faunal similarity between the two sites. In fact, Maboko Island is much more similar faunally to

Fossil Catarrhines from Fort Ternan 517
the early Miocene Hiwegi Formation of Rusinga Island, with which it shares 9 species (36%) and 17 genera (54.8070), than it is to Fort Ternan.
Chronological and biostratigraphical evidence provides strong support that Fort Ternan and Maboko Island can be estimated to be dated at 14 Ma and 15 - 16 Ma respectively. The fossil beds at Fort Ternan are bracketed by radiometric dates of the overlying Kericho phonolite of 12.5 - 12.6 Ma (which is consistent with a series of radiometric dates obtained for the Plateau Phonolites, of which the Kericho Phonolite is part, from other parts of Kenya) and the underlying Baraget phonolite of 14.9-15.1 Ma (BISHOP et al., 1969; BAKER et al., 1971; LIPPARD, 1973; FITCH et al., 1978; SHIPMAN et al., 1981). Furthermore, the Fort Ternan fauna is located stratigraphically in superposition to the Nyakach fauna, which is almost identical in composition to the Maboko fauna, and has been dated to 13.4 Ma by fission-track methods (PICKEORD, 19860. These faunal differences between Maboko Island and Fort Ternan have been used as a basis by PICKVORD (1983a) to include the sites in different faunal sets (Maboko is included in Faunal Set III and Fort Ternan is included in Faunal Set IV). The Fort Ternan mammalian fauna is noticeably more progressive than that from Maboko Island, and the same general tendency is also reflected in the fossil primates. For instance, Kenyapithecus wickeri at Fort Ternan is appreciably more derived towards modern large hominoids than is "Sivapithecus" africanus, and the oreopithecid is more specialized than its antecedent, Nyanzapithecus, from the early and middle Mio- cene. These profound differences between the faunas from Maboko Island and Fort Ternan could indicate that the slender estimated age difference between the sites actually requires revising, with Fort Ternan, perhaps, being somewhat younger than 14 Ma (see PICKFORD, 1986f). Alternatively, the differences could reflect facies or ecological differences at the two sites.
A number of authors, employing contrasting techniques, have attempted to reconstruct the paleoecological setting at Fort Ternan. Although these studies have yielded somewhat contradictory results, the weight of the evidence would favor a habitat type that was predominantly open to closed woodland/bushland, with scattered grassy glades and more densely vegetated areas consisting of woodland-forest mosaics (ANDREWS & NESBIT EVANS, 1979; SHIPMAN et al., 1981; PICKFORD, 1983a, 1986a, b, 1987; BONNEFILLE, 1984, 1985; SHIPMAN, 1986; RETALLACK, 1988; DUGAS & RETALLACK, 1988; CERLING & QUADE, 1989; CZRLXNG et al., 1989, 1991; RETALLACK et al., 1990; KAPPELMAN, 1991). A generally similar mosaic habitat has also been inferred for Maboko Island (PIcKVORD, 1982, 1983a, 1986a, 1986d). However, Maboko was probably more densely wooded than Fort Ternan, with a thick gallery forest bordering the main watercourses, and with relatively few open grassy areas. The riverine setting at Maboko, with an abundance of hydrophilic vertebrates in the fauna, also provides a dramatic contrast with the subaerial depositional environment at Fort Ternan, in which crocodiles and turtles are almost entirely lacking. In fact, the general proximity of Maboko to a major water source may have been one of the most important factor contributing to the inferred vegetational differences between the two sites. In conclu- sion, although there appear to be some significant differences between the reconstructed paleoecologies at Fort Ternan and Maboko, that might account for differences in their respective faunas, it seems unlikely that these would have been profound enough to account for the extent of the differences observed. We may deduce from this that the faunal differ- ences, including the marked contrast in the composition of the respective primate commu- nities, between Fort Ternan and Maboko Island primarily reflect a significant time difference between the sites.

518 T. HARRISON
CONCLUSIONS
This paper provides a revised assessment of the taxonomic and phylogenetic affinities of the fossil catarrhine primates from the middle Miocene site of Fort Ternan in Kenya. The material can be attributed to at least five different species, which are identified here as Kenyapithecus wickeri, Proconsul sp., a large species of oreopithecid, Simiolus sp., and a small species of catarrhine of indeterminate status. Based on this reassessment, the major conclusions are as follows: (1) Kenyapithecus wickeri from Fort Ternan should be separated at the generic level from "Sivapithecus" africanus from Maboko Island. In its dentition and lower face, K. wickeri has developed a complex of derived traits that it shares with later large hominoids, that are not present in the more conservative "'S. '" africanus. Compari- sons of K. wickeri with other extant and fossil hominoids have revealed a marked similarity in a number significant respects to Pongo and Sivapithecus. It is conceivable that these shared characteristics may indicate a close phyletic relationship between Kenyapithecus and the Sivapithecus-Pongo clade, but they are best interpreted as shared primitive features retained from the ancestral large hominoid morphotype. The evidence therefore supports the inference that Kenyapithecus is a conservative sister-taxon of the extant large homi- noids. (2) Two isolated teeth from Fort Ternan are very similar to those of Proconsul africanus from the early Miocene of East Africa. However, given the present confusion surrounding the alpha-taxonomy of the Proconsul group, the teeth from Fort Ternan are considered to belong to an indeterminate species of Proconsul. (3) Several isolated teeth can be identified as belonging to a large species of oreopithecid. They are larger and more derived in their morphology than the early and middle Miocene oreopithecid, Nyanzapithe- cus, but are quite similar to Oreopithecus bambolii from the late Miocene of Italy. (4) A small species of catarrhine from Fort Ternan is morphologically and metrically similar to Simiolus enjiessi from Kalodirr in northern Kenya, and is attributed here to the same genus. It differs from the type species in a number of minor morphological difference of the denti- tion and in having a reduced M3. The differences are probably sufficient to recognize a separate species, but, as the sample sizes from Kalodirr and Fort Ternan are relatively small, it seems wise to await the recovery of more extensive material before proposing a new taxon. (5) A second species of small catarrhine, slightly larger than Simiolus, is represented at Fort Ternan. Metrically and morphologically the material is most similar to Kalepithecus songhorensis from the early Miocene localities of Koru and Songhor. However, until fur- ther material is available to reliably assign these specimens, they are regarded here as of indeterminate status. (6) The occurrence of at least five species of catarrhine primates at Fort Ternan confirms earlier claims (HARRISON, 1989b) that primate species diversity levels were as high in the middle Miocene as they had been in the early Miocene. The middle Miocene catarrhine community differs from the early Miocene community in its overall taxonomic and ecological composition, rather than in its species diversity. (7) The revised taxonomy of the Fort Ternan catarrhines has revealed that the detailed composition of the community is strikingly different from that from the middle Miocene locality of Maboko Island. This difference, also reflected in the wider mammalian faunas, can probably be explained as a consequence of a major chronological separation between the two sites, and, to a lesser degree, to paleoecological differences.
Acknowledgments. I am grateful to RICHARD LEAKEY, MEAVE LEAKEY, and the Trustees of the National Museums of Kenya for permission to study the fossil primates in their care. I would like

Fossil Catarrhines from Fort Ternan 519
to thank PETER ANDREWS, ERIC DELSON, TERRI HARRISON, MARTIN PICKFORD, and MIKE ROSE for their invaluable advice and comments. The research was supported by grants from the L.S.B. Leakey Foundation and the Boise Fund, Oxford University.
R E F E R E N C E S
ABEL, O., 1902. Zwei neue Menschenaffen aus den Leithakalkbildungen des Weiner Beckens. S. Ber. Akad. Wiss. Wien, Math.-nat. KL, 3:1171-1207.
AGUIRRE, E., 1975. Kenyapithecus and Ramapithecus. In: Paleoanthropology, Morphology and Paleoecology, R. TUTTLE (ed.), Mouton, The Hague, pp. 99-104.
ALPAGUT, B., P. ANDREWS, & L. MARTIN, 1990. New hominoid specimens from the middle Miocene site at Pasalar, Turkey. J. Human Evol., 19: 397-422.
ANDREWS, P. J., 1971. Ramapithecus wickeri mandible from Fort Ternan, Kenya. Nature, 231: 192 - 194.
1973. Miocene Primates (Pongidae, Hylobatidae) of East Africa. Ph.D. Dissertation, The Univ. of Cambridge.
1978. A revision of the Miocene Hominoidea of East Africa. Bull. Brit. Mus. Nat. Hist. (Geol.), 30: 8 5 - 224.
1980. Ecological adaptations of the smaller fossil apes. Z. Morph. Anthropol., 71: 164 - 173.
1981. Species diversity and diet in monkeys and apes during the Miocene. In: Aspects of Human Evolution, C. B. STRINGER (ed.), Taylor & Francis, London, pp. 2 5 - 61.
1982. Ecological polarity in primate evolution. Zool. J. Linn. Soc., 74: 2 3 3 - 244. - - , 1988. Hominoidea. In: Encyclopedia of Human Evolution and Prehistory, I. TATTERSALL,
E. DELSON, & J. VAN COUVERING (eds.), Garland, New York, pp. 248 - 255. - - & L. MARTIN, 1987. Cladistic relationships of extant and fossil hominoids. J. Human
Evol., 16: 101-118. - - t~ T. MOLLESON, 1979. The provenance of Sivapithecus africanus. Bull. Brit. Mus. Nat.
Hist. (Geol.), 32: 19-23. - - & E. NESBIT EVANS, 1979. The environment of Ramapithecus in Africa. Paleobiology, 5:
22 - 30. & H. TOBIEN, 1977. New Miocene locality in Turkey with evidence on the origin of
Ramapithecus and Sivapithecus. Nature, 268: 699-701. - - • A. WALKER, 1976. The primate and other fauna from Fort Ternan, Kenya. In: Human
Origins, G. L. ISAAC & E. R. McCowN (eds.), W. A. Benjamin, Menlo Park, California, pp. 274 - 304.
BAKER, B. H., L. A. J. WILLIAMS, J. A. MILLER, & E J. FITCH, 1971. Sequence and geochronology of the Kenya Rift volcanics. Tectonophysics, 11" 191-215.
BENEFIT, B. R. & M. L. MCCRoSSIN, 1989. New fossil primates from the middle Miocene of Maboko Island, Kenya. J. Human EvoL, 18: 493 - 497.
BISHOP, W. W., J. A. MILLER, & E J. FITCH, 1969. New Potassium-Argon age determinations relevant to the Miocene fossil mammal sequence in East Africa. Amer. J. Sci., 267: 669- 699.
BONNEFILLE, R., 1984. Cenozoic vegetation and environments of early hominids in East Africa. In: Evolution of the East Asian Environment, VoL II, R. O. WHYTE (ed.), Univ. of Hong Kong Press, Hong Kong, pp. 579-612.
- - , 1985. Evolution of the continental vegetation: the palaeobotanical record from East Africa. S. Afr. J. ScL, 81: 267-270.
BROWN, B., A. HILL, & S. WARD, 1991. New Miocene large hominoids from the Tugen Hills, Baringo District, Kenya. Amer. J.. Phys. Anthropol. (Supplement), 12: 55.
- - & S. C. WARD, 1988. Basicranial and facial topography in Pongo and Sivapithecus. In: Orang-utah Biology, J. H. SCHWARTZ (ed.), Oxford Univ. Press, Oxford, pp. 247- 260.
BUTLER, P. M., 1984. Macroscelidea, Insectivora and Chiroptera from the Miocene of East Africa. Palaeovertebrata, 14: 117-200.
CAMERON, D. W., 1991. Sexual dimorphism in the early Miocene species of Proconsul from the Kisingiri Formation of East Africa: a morphometric examination using multivariate statistics. Primates, 32: 329-343.

520 T. HARRISON
CERL1NG, Y. E. & J. QUADE, 1989. Carbon isotopes in paleosol carbonates as paleoecological indica- tors. Amer. J. Phys. Anthropol., 78: 203.
- - , S. H. AMBROSE, & N. E. SIKES, 1991. Fossil soils, grasses, and carbon isotopes from Fort Ternan, Kenya: grassland or woodland? J. Human Evol., 21: 295-306.
, - - , Y. WANG, & J. R. BOWMAN, 1989. Carbon isotopes in soils and paleosols as ecology and paleoecology indicators. Nature, 341: 138-139.
CONROY, G. C. & D. R. PILBEAM, 1975. Ramapithecus: a review of its hominid status. In: Paleoan- thropology, Morphology, and Paleoanthropology, R. TUTTLE (ed.), Mouton, The Hague, pp. 59-86.
CORRUCCINI, R. S., R. L. CIOCHON, 8~ H. M. McH~NRY, 1976. The postcranium of Miocene homi- noids: were dryopithecines merely "dental apes"? Primates, 17: 205-223.
DELSON, E., 1988. Catarrhini. In: Encyclopedia of Human Evolution and Prehistory, I. TATTERSALL, E. DELSON, & J. VAN COUVERING (eds.), Garland, New York, pp. 111-116.
DUGAS, D. P. & G. J. RETALLACK, 1988. Fossil grasses of the Kenyapithecus locality near Fort Ternan, Kenya. Amer. J. Phys. AnthropoL, 75: 205-206.
FELDESMAN, M. R., 1982. Morphometric analysis of the distal humerus of some Cenozoic catarrhines: the late divergence hypothesis revisited. Amer. J. Phys. AnthropoL, 59: 7 3 - 95.
FITCH, E J., P. J. HOOKER, & J. A. MILLER, 1978. Geochronological problems and radioisotopic dating of volcanic rocks in the Gregory Rift Valley of East Africa. In: Geological Background to Fossil Man, W. W. BISHOP (ed.), Scottish Academic Press, Edinburgh, pp. 441-461.
FRAYER, D. W., 1976. A reappraisal of Ramapithecus. Yrbk. Phys. Anthropol., 18: 19- 30. GREENFIELD, L. O., 1979. On the adaptive pattern of "'Ramapithecus'" Amer. J. Phys. AnthropoL,
50: 527- 548. HARRIS, J. M., 1978. Deinotheroidea and Barytherioidea. In: Evolution of African Mammals, V. J.
MAGLIO & H. B. S. COOKE (eds.), Harvard Uni~: Press, Cambridge, Massachusetts, pp. 315- 332. HARRISON, T., 1982. Small-bodied apes from the Miocene of East Africa. Ph.D. Dissertation, Univ.
of London. - - , 1986. New fossil anthropoids from the middle Miocene of East Africa and their bearing
on the origin of the Oreopithecidae. Amer. J. Phys. Anthropol., 71: 265-284. - - , 1989a. A taxonomic revision of the small Catarrhine primates from the early middle
Miocene of East Africa. Folia Primatol., 50: 59-108. - - , 1989b. A new species of Micropithecus from the middle Miocene of Kenya. J. Human
Evol., 18: 537-557. HOOIJER, D. A., 1948. Prehistoric teeth of man and of the orang-utan from central Sumatra, with
notes on the fossil orang-utan from Java and southern China. Zool. Med., 29:175-293. IsnmA, H., M. PICKFORD, H. NAr:AVA, & Y. NAKANO, 1984. Fossil anthropoids from Nachola and
Samburu District, Kenya. Afric. Stud. Monogn (Supplementary Issue), 2: 7 3 - 85. KAPPELMAN, J., 1991. The paleoenvironment of Kenyapithecus at Fort Ternan. J. Human Evol., 20:
95 - 129. KAY, R. E & E. L. SIMONS, 1983. A reassessment of the relationship between later Miocene and
subsequent Hominoidea. In: New Interpretations of Ape and Human Ancestry, R. L. CIOCHON & R. S. CORRt:CCINI (eds.), Plenum, New York, pp. 577-624.
KELLEY, J., 1986. Species recognition and sexual dimorphism in Proconsul and Rangwapithecus. J. Human Evol., 15:461-495.
- - & D. PILBEAM, 1986. The dryopithecines: taxonomy, comparative anatomy, and phylogeny of Miocene large hominoids. In: Comparative Primate Biology, Vol. 1: Systematics, Evolution, and Anatomy, D. R. SWINDLER & J. IRWIN (eds.), Alan R. Liss, New York, pp. 361-411.
KRANTZ, G. S., 1975. The double descent of man. In: Paleoanthropology, Morphology, and Paleo- ecology, R. H. TCTTLE (ed.), Mouton, The Hague, pp. 131-152.
LAVOCAT, R., 1978. Rodentia and Lagomorpha. In: Evolution of African mammals, V. J. MAGLIO & H. B. S. COOKE (eds.), Harvard Univ. Press, Cambridge, Massachusetts, pp. 69-89.
LEAKEY, L. S. B., 1962. A new lower Pliocene fossil primate from Kenya. Ann. Mag. Nat. Hist., 4: 689 - 696.
- - , 1967. An early Miocene member of Hominidae. Nature, 213: 155-163. - - - , 1968. Upper Miocene primates from Kenya. Nature, 218: 527-528. LEAKEY, R. E. & M. G. LEAKEY, 1987. A new Miocene small-bodied ape from Kenya. J. Human
Evol., 16: 369-387.

Fossil Catarrhines from Fort Ternan 521
LE GROS CLARK, W. E. & L. S. B. LEAKEY, 1950. Diagnoses of East African Miocene Hominoidea. Quart. J1. Geol. Soc. Lond., 105: 260-262.
& , 1951. The Miocene Hominoidea of East Africa. Foss. Mamm. Afr., 1:1-117. LIPPARD, S. J., 1973. Plateau phonolite lava flows, Kenya. Geol. Mag., 110: 543- 549. MACINNES, D. G., 1943. Notes on the East African Miocene primates. J1. East Aft. Nat. Hist. Sot.,
17:141 - 181. MADDEN, C. T., 1980. East African Sivapithecus should not be identified as Proconsul nyanzae.
Primates, 21:133 - 135. MALONE, D., 1988. Mechanisms of hominoid dispersal in Miocene East Africa. J. Human Evol., 16:
469 - 481. MARTIN, L., 1983. The relationships of the later Miocene Hominoiclea. Ph.D. dissertation, Univ. of
London. - - , 1986. Relationships among extant and extinct great apes and humans. In: Major Topics
in Primate and Human Evolution, B. WooD, L. MARTIN, & P. ANDREWS (eds.), Cambridge Univ. Press, Cambridge, pp. 161-187.
McHENRY, H. M. & R. S. CORRUCCIN1, 1975. Distal humerus in hominoid evolution. Folia Primatol., 23: 227- 244.
MORBECK, M. E., 1983. Miocene hominoid discoveries from Rudabanya. In: New Interpretations of Ape and Human Evolution, R. L. CIOCHON & R. S. CORRUCClN1 (eds.), Plenum, New York, pp. 369 - 404.
PICKFORD, M., 1982. New higher primate fossils from the middle Miocene deposits at Majiwa and Kaloma, western Kenya. Amer. J. Phys. Anthropol., 58: ! - 19.
- - , 1983a. Sequence and environments of the lower and middle Miocene hominoids of western Kenya. In: New Interpretations of Ape and Human Ancestry, R. L. CIOCHON & R. S. CORRUCCINI (eds.), Plenum, New York, pp. 421 -439.
- - , 1983b. An account of the new Kenyan fossil discoveries. Interim Evidence, 5 : 1 - 6 . - - , 1985a. A new look at Kenyapithecus based on recent discoveries in western Kenya. J.
Human Evol., 14: 113-143. - - , 1985b. Kenyapithecus: a review of its status based on newly discovered fossils from Kenya.
In: Hominid Evolution: Past, Present and Future, P. TOBIAS (ed.), Alan R. Liss, New York, pp. 107 - 112.
- - , 1986a. Snails. Anthroquest, 36: 3 - 5. - - , 1986b. Hominoids from the Miocene of East Africa and the phyletic position of
Kenyapithecus. Z. Morph. Anthropol., 76: 117-130. - - , 1986c. The geochronology of Miocene higher primate faunas of East Africa. In: Primate
Evolution, J. G. ELSE & P. C. LEE (eds.), Cambridge Univ. Press, Cambridge, pp. 19-33. - - , 1986d. Cainozoic paleontological sites of western Kenya. Munchner Geowiss. Abh., A, 8:
1-151. - - , 1986e. Sexual dimorphism in Proconsul. Human Evol., I: 111-148.
, 1986f. The geochronology of Miocene higher primate faunas of East Africa. In: Primate Evolution, J. G. ELSE & P. C. LE~ (eds.), Cambridge Univ. Press, Cambridge, pp. 19-33.
- - , 1987. Fort Ternan (Kenya) paleoecology. J. Human Evol., 16: 305 - 309. - - , 1988. Geology and fauna of the middle Miocene hominoid site at Muruyur, Baringo
District, Kenya. Human Evol., 3: 381- 390. PILBEAM, D., 1969. Tertiary Pongidae of East Africa: evolutionary relationships and taxonomy. Bull.
Peabody Mus. Nat. Hist., 31:1-185. PREUSCHOFT, H., 1973. Body posture and locomotion in some East African Miocene Dryopitheci-
nae. In: Human Evolution, M. DAY (ed.), Symposium of the Society for the Study of Human Biology, Vol. 11, pp. 13-46.
RETALLACK, G. J., 1988. Paleopedological evidence for the Miocene habitat of Kenyapithecus at Fort Ternan National Monument, Kenya. Amer. J. Phys. Anthropol., 75: 260.
- - , D. P. DUGAS, & E. A. BESTLAND, 1990. Fossil soils and grasses of a middle Miocene East African grassland. Nature, 247: 1325-1328.
ROSE, M. D., 1983. Miocene hominoid postcranial morphology: monkey-like, ape-like, neither, or both? In: New Interpretations of Ape and Human Evolution, R. L. CIOCHON & R. S. CORRUCCINI (eds.), Plenum, New York, pp. 405- 417.

522 T. HARRISON
RUFF, R. B., A. C. WALKER, • M. F. TEAFORD, 1989. Body mass, sexual dimorphism and femoral proportions of Proconsul from Rusinga and Mfangano Islands, Kenya. J. Human Evol., 18: 515- 536.
SAVAGE, R. J. G., 1978. Carnivora. In: Evolution of African Mammals, V. J. MAGLIO t~; H. B. S. COOKE (eds.), Harvard Univ. Press, Cambridge, Massachusetts, pp. 249-267.
SENUT, B., 1982. R6flexions sur la brachiation et l'origine des hominid6s A la lumi6re des hominoides mioc~nes et des hominid6s Plio-P16istocenes. Geobios, M~moire spdcial, 6: 335- 344.
- - , 1989. Le Coude des Primates Homino'tdes." Anatomie, Fonction, Taxonomie, Evolution. Cahiers de Pal6oanthropologie, CNRS, Paris.
SHIPMAN, P., 1986. Paleoecology of Fort Ternan reconsidered. J. Human Evol., 15: 193- 204. - - , A. WALKER, J. A. VAN COUVERING, P. J. HOOKER, & J. A. MILLER, 1981. The Fort Ternan
hominoid site, Kenya: geology, age, taxonomy and paleoecology. J. Human Evol., 10: 49-72 . SIMONS, E. L., 1963. Some fallacies in the study of hominid phylogenies. Science, 141: 879-889. - - , 1969. Late Miocene hominid from Fort Ternan Kenya. Nature, 221: 448-451. - - , 1972. Primate Evolution: An Introduction to Man's Place in Nature, MacMillan, New
York. - - , 1981. Man's immediate forerunners. Phil. Trans. R. Soc. London, B292:21-41.
& D. R. PILBEAM, 1965. Preliminary revision of the Dryopithecinae (Pongidae, Anthropoidea). Folia Primatol., 3: 81-152.
STEININGER, E, 1967. Ein Weiterer zahn von Dryopithecus (Dry.)fontani darwini Abel, 1902 (Mammalia, Pongidae) aus dem Mioz~in des Weiner Beckens. Folia Primatol., 7: 243- 275.
SZALAY, E S. & E. DELSON, 1979. Evolutionary History o f the Primates. Academic Press, New York. TEAFORD, M. E, K. C. BEARD, R. E. LEAKEY, t~r A. WALKER, 1988. New hominoid facial skeleton
from the early Miocene of Rusinga Island Kenya, and its bearing on the relationship between Proconsul nyanzae and Proconsul africanus. J. Human Evol., 17:461 -477.
TEMERIN, L. A. t~r J. G. H. CANT, 1983. The evolutionary divergence of Old World monkeys and apes. Amer. Naturalist, 122: 335- 351.
WALKER, A. t~r P. ANDREWS, 1973. Reconstruction of the dental arcades of Ramapithecus wickeri. Nature, 244: 313- 314.
- - 8r M. TEAFORD, 1988. The Kaswanga primate site: an early Miocene hominoid site on Rusinga Island, Kenya. J. Human Evol., 17: 539-544.
- - & - - , 1989. The hunt for Proconsul. Sci. Amer. January, 1989: 76-82 . WARD, S. C. t~r D. R. PILBEAM, 1983. Maxillo-facial morphology of Miocene hominoids from Africa
and Indo-Pakistan. In: New Interpretations o f Ape and Human Ancestry, R. L. CIOCHON & R. S. CORRUCCINI (eds.), Plenum, New York, pp. 325- 351.
WOLPOEF, M., 1983. Ramapithecus and human origins: an anthropologist's perspective of changing interpretations. In: New Interpretations o f Ape and Human Ancestry, R. L. CIOCHON & R. S. CORRUCC1N1 (eds.), Plenum, New York, pp. 651- 676.
- - Received January 7, 1992; Accepted June 16, 1992
Author's Name and Address: TERRY HARRISON, Department of Anthropology, New York University, 25, Waverly Place, New York, New York 10003, U. S. A.



















