
บทที่5 การพัฒนาระบบสารสนเทศ และ ระบบสารสนเทศสาหรับองค์กร ...
Upload
khangminh22Category
view
1download
0
คณะวทยาศาสตรมหาวทยาลยมหดล
วทยาศาสตรเพอการพฒนา รวมบทความวชาการฉลองครบรอบ 50 ป
แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
Wisdom of the Landปญญาของแผนดน


ตราสญลกษณ 50 ป คณะวทยาศาสตร มหาวทยาลยมหดล
ตวเลข 50 ป ครบรอบแหงการสถาปนาคณะวทยาศาสตร
สนำเงน คอสประจำมหาวทยาลยมหดล
สเหลองทอง คอสประจำคณะวทยาศาสตร
สแดง เงาตวเลขคอความสดใสกาวหนาสรางสรรคผลงานและพลงแฝง
เสนโคงสนำเงน แสดงถงองคประกอบโดยรวมความรวมมอรวมใจ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป
แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
บรรณาธการผทรงคณวฒ ศาสตราจารยดร.มรว.ชษณสรรสวสดวตน
ศาสตราจารยดร.ประพนธวไลรตน
ศาสตราจารยดร.นททพยกฤษณามระ
ศาสตราจารยดร.ยงควมลเลณบร
ศาสตราจารยดร.ศกรณมงคลสข
ProfessorDr.DavidJohnRuffolo
รองศาสตราจารยดร.รศมดาราหนสวสด
Dr.JamesDubbs
บรรณาธการ ศาสตราจารยดร.ชโลบลอยสข
ผจดทำ งานนโยบายและแผนและพฒนาคณภาพ
คณะวทยาศาสตรมหาวทยาลยมหดล
272ถนนพระรามท6เขตราชเทวกรงเทพฯ10400
โทรศพท0-2201-5000
www.sc.mahidol.ac.th
จำนวนพมพ 500เลม
พมพท บรษทเอเอนทออฟฟศเอกซเพรสจำกด
78อาคารมกดาชนGถนนสาทรเหนอ
แขวงสลมเขตบางรกกรงเทพฯ10500
โทร.0-2876-1440-1
เดอนปทจดพมพ สงหาคม2552
ISBN 978-974-11-1180-0
ออกแบบและสรางสรรค รจเรขาวทยาวฑฒกล
อภชยอารยะเจรญชย
วรษยาสนทรศารทล
สงวนลขสทธตามพระราชบญญตลขสทธพ.ศ.2537โดยคณะวทยาศาสตรมหาวทยาลยมหดล

คำนำ
ในโอกาสทคณะวทยาศาสตร มหาวทยาลยมหดล ไดกาวขามกงศตวรรษเขาสปท 51 แหง
การสถาปนาคณะฯ และการเปนคณะวทยาศาสตรชนนำของประเทศจงนบเปนโอกาสอนดทจะมการจดทำ
หนงสอ “วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะ
วทยาศาสตร มหาวทยาลยมหดล” เพอรวบรวมบทความทางวชาการตางๆ และเผยแพรความรจากผลงาน
วจยบางสวนของคณะวทยาศาสตรทกำลงดำเนนการอยในปจจบน ออกสสงคมเพอใหสาธารณชนสามารถ
นำไปใชประโยชน บทความทงหมดนสวนใหญเขยนขนโดยคณาจารย นกวจยของคณะวทยาศาสตร
มหาวทยาลยมหดลซงเปนคนรนใหมทมความรความสามารถในศาสตรแขนงตางๆและเปนทรพยากรบคคล
ททรงคณคา เปนกำลงสำคญในการขบเคลอนและพฒนาคณะวทยาศาสตรมหาวทยาลยมหดล ใหกาวตอไป
สการเปนคณะวทยาศาสตรชนนำในระดบสากล
หนงสอเลมน ประกอบไปดวยบทความทกลาวถงพฒนาการของงานวจยบางสาขาจากอดต
ถงปจจบนงานวจยเกยวกบสขภาพและการแพทยจลชพกอโรคการจดการคลงยาและการใชยาเทคนคและ
วธการตรวจสอบพชอาหารพชเศรษฐกจและเชอรากอโรคพชนเวศวทยาและสงแวดลอมเทคโนโลยและการ
ประยกตใช โดยลกษณะของการเขยนบทความ มทงบทความทใหความรสำหรบประชาชนทวไป และ
บทความทใหความรในเชงลกของศาสตรแขนงนนๆ
ในนามของคณะวทยาศาสตรมหาวทยาลยมหดลขอขอบคณกองบรรณาธการผทรงคณวฒ
บคลากรทชวยจดทำหนงสอ คณาจารย นกวจยทกทานทมสวนรวมในการเขยนบทความ จำนวน 41 เรอง
และหวงเปนอยางยงวา ความรจากบทความทางวชาการภายในหนงสอเลมน จะกอประโยชนตอการพฒนา
วงการวทยาศาสตรและเทคโนโลยของประเทศไทยอยางยงยน ใหสมดงคำปณธานของคณะวทยาศาสตร
มหาวทยาลยมหดล ในการเปนผนำทางวชาการและวจย เพอพฒนาวทยาศาสตร เทคโนโลย เศรษฐกจ
และสงคมของประเทศตลอดไป
(ศาสตราจารยศกรณมงคลสข)
คณบดคณะวทยาศาสตรมหาวทยาลยมหดล
สงหาคม2552


สารบญ
จากอดตถงปจจบน
Vitamin E Research at the Faculty of Science, Mahidol University
Prapon Wilairat .......................................................................................................................................... 1
Chemical Biology: Addressing Biological Questions from Chemical Perspectives
Chutima Jiarpinitnun .................................................................................................................................. 7
Forensic Science at the Faculty of Science, Mahidol University, in the International
Forensic Science Arena
Nathinee Panvisavas ................................................................................................................................... 17
สขภาพและการแพทย
การพฒนาเซลลสบพนธและกระบวนการปฏสนธในกงกลาดำ
รพพรรณ วานชวรยกจ และ ประเสรฐ โศภน ......................................................................................................... 25
การพฒนาวคซนตอโรคพยาธใบไมตบ
กลธดา ชยธระยานนท และ ประเสรฐ โศภน ........................................................................................................... 33
ความเครยดกระทบกระเทอนระบบภมคมกนของรางกายอยางไร
สพชา คมเกต .......................................................................................................................................................... 41
สารตานอนมลเสรกบทางสายกลาง
ระพ บญเปลอง ........................................................................................................................................................ 49
โรคฉหน: ภยทมากบนำ
กลลยาน ดวงฉว ...................................................................................................................................................... 59
Microglial Activation in Parkinson’s Disease
Rattanavijit Vijitruth ................................................................................................................................... 67
Sex Hormones and the Activation of Cardiac Machinery
Tepmanas Bupha-Intr and Jonggonnee Wattanapermpool ........................................................................ 73
ATP-binding Cassette Transporters in Biomarker Research
Tavan Janvilisri .......................................................................................................................................... 81
จลชพกอโรค
Leptospira: Overview and Genetics
Padungsri Dubbs ........................................................................................................................................ 89
How Malaria Genes are Regulated
Ruchanok Cheotacha, Thanat Chookajorn, Krittikorn Kumpornsin,
Patrath Ponsuwanna and Kanjana Wittayacom ......................................................................................... 99

การจดการคลงยาและการใชยา
คณตศาสตรในการจดการคลงยาในโรงพยาบาล
ระว สวรรณเดโชไชย ............................................................................................................................................ 105
การรกษาผวหนงอกเสบระคายสมผส
วรรณจรส รงพสทธพงษ และ ธรวฒ คลงเจรญชย ............................................................................................... 113
Antiviral Drugs for the Control of Influenza Virus Infections: Present and Future Perspectives
Radeekorn Akkarawongsapat ................................................................................................................... 119
เทคนคและวธการตรวจสอบ
ชดเครองวดภาคสนามสยามแกโซฮอล หนงในนวตกรรมครบรอบ 50 ป คณะวทยาศาสตร
กาญจนา อไรสนธว และ ดวงใจ นาคะปรชา ....................................................................................................... 125
Loop-mediated Isothermal Amplification (LAMP) Method: an Easy and Fast Gene Amplification Method
for Detection of Malaria Parasites
Sureemas Buates ...................................................................................................................................... 131
Reporter Gene Assays for Anti-TB Drug Screening
Chartchai Changsen ................................................................................................................................. 139
Oligomer Extrapolation Techniques: An Economic Approach to the Study of Relationships between
Polymeric Structures and Band Gaps
Withoon Chunwachirasiri ........................................................................................................................ 147
พชอาหารและพชเศรษฐกจ และเชอรากอโรคพช
เนอในเมลดมะมวง ของดทถกมองขาม
ปราณ อนประโคน .............................................................................................................................................. 153
ทรพยากรพนธกรรมกลวยในประเทศไทย...ทรพยแหงสวรรณภม
ศศวมล แสวงผล และ จามร สมณะ....................................................................................................................... 163
เทคโนโลยชวภาพพชเพอการปรบปรงพนธยางพารา
จรญญา ณรงคะชวนะ ........................................................................................................................................... 171
สารพษจากเชอรากบบทบาทตอความสามารถในการกอโรคพช
ทพา อศวรกษ ...................................................................................................................................................... 179
นเวศวทยาและสงแวดลอม
“ยงลาย” รายกวาทเคยคด
จนรภา โพธกสกร ระพ บญเปลอง วไล หนนภกด และ สภาวด ราชมณ ............................................................. 187
นเวศวทยาเชงอนรกษ...การเชอมโยงวทยาศาสตรกบสงคมศาสตร และ กรณศกษาปลาเฉพาะถน
ทใกลสญพนธ
เจนจต คดำรงสวสด ............................................................................................................................................. 195
แบบจำลองทางคณตศาสตรสำหรบระบบนเวศวทยา
กรกนก บญวงษ ................................................................................................................................................... 201

Phytoremediation of Heavy Metals and Bioaccumulation in Food Plants
Pahol Kosiyachinda .................................................................................................................................. 207
เทคโนโลยและการประยกตใช
แนวโนมการพฒนาอปกรณเปลงแสงจากสารอนทรย
สมศกด แดงตบ ธนากร โอสถจนทร และ สมบญ สหสทธวฒน .......................................................................... 215
เซลลแสงอาทตยแบบสยอมไวแสง
สภาวด เกยรตเสว และ ยทธนา ตนตรงโรจนชย ................................................................................................... 225
การประยกตใชกลองจลทรรศนแรงอะตอมในการศกษาทางชววทยา
ธราพร พนธธรานรกษ .......................................................................................................................................... 233
การประยกตใชสมการเลเวลเซตในศาสตรดานอนๆ
พลลภ ฮวบสมบรณ .............................................................................................................................................. 241
การควบคมการตายของเซลลสตวเพาะเลยงทใชในอตสาหกรรมเทคโนโลยชวภาพ
อษฎา มงสนตสข .................................................................................................................................................. 247
The Power of DNA Technology in Forensic Science
Patiya Phengon and Nathinee Panvisavas ............................................................................................... 257
Gold Nanoparticles: Applications and Synthetic Methodology
Jirarut Wongkongkatep, Pravit Wongkongkatep, Woraphoj Lappenpuntrap,
Runchuan Ladadat and Pranee Phinyocheep ........................................................................................... 265
Transparent Oxygen-Barrier Coating Based on Plasma Deposition of SiOx Thin Film on
Polymeric Substrate
Supa Wirasate ........................................................................................................................................... 271
Supported Planar Lipid Membranes and Their Applications
Pallop Karnchanaphanurach .................................................................................................................... 279
Whole-cell Biocatalysts from Organic Solvent-tolerant Bacteria
Thunyarat Pongtharangku and Alisa S Vangnai ..................................................................................... 287
Friction Stir Weld in High Strength Aluminium Alloys 2024
Manthana Jariyaboon ............................................................................................................................... 295
Geometry and Mechanics of Carbon Nanostructures
Duangkamon Baowan ............................................................................................................................... 303
A Survey of Approaches to Digital Image Matting
Rawesak Tanawongsuwan ........................................................................................................................ 311
Keyword Index .................................................................................................................................................. 321
ดชนคำสำคญ ................................................................................................................................................................. 325
ดชนผนพนธ .................................................................................................................................................................. 327


1
Vitamin E Research at the Faculty of Science, Mahidol University
Prapon Wilairat Department of Biochemistry, Faculty of Science, Mahidol University.
Abstract
During the past 50 years, research at the Faculty of Science, Mahidol University on vitamin E function in man and experimental animals have led to a clearer understanding of this fat-soluble vitamin, both as an antioxidant and in novel non-antioxidant roles.
Keywords: cholesterol; Macaca fascicularis; oxidative stress; thalassemia; vitamin E
Research on vitamin E at the Faculty of
Science could be considered to have started in 1964 when Dr. Serene (Vimokesant) Piboon-niyom joined the Department of Biochemistry, Faculty of Science. Dr. James S. Dinning, the Rockefeller Foundation representative at the Faculty of Science at that time, had previously studied the effects of vitamin E deficiency in rhesus monkey and showed that the first sign of deficiency of this fat-soluble vitamin in primates is anemia, followed later by muscular atrophy and abnormal excretion of urinary nitrogenous compounds(4). Dr. Serene was soon able to demonstrate that anemia associated with protein-calorie malnutrition in man also responds to vitamin E therapy(24). It was subsequently demonstrated (by others) that vitamin E improves anemia in premature children(2).
In order to understand the role of vitamin E in regulating erythroid development in man, a colony of young crab-eating monkeys, Macaca fascicularis, was established at the Faculty of Science. Monkeys were fed a basal vitamin-E depleted diet in the form of baked cookies. After a period of 11 months, plasma vitamin E level decreases from a normal value of 3-7 μg/ml to an undetectable level (less than 1 μg/ml using a fluorescence assay), and signs of anemia appeared after 15 months (but growth rates of vitamin E-deficient monkeys are comparable with control)(19). Bone marrow picture of vitamin E-deficient monkeys revealed the presence of multinucleated erythroid cells
and an accumulation of orthochromatophilic normoblasts. Extracted bone marrow cells contain an increased amount of 4-5 RNA species, which are synthesized during the late stages of erythroid development(1). Supple-mentation with vitamin E (80 mg/day of tocopheryl acetate for 10 days) resulted in reticulocytosis and a return to normal bone marrow picture. These results indicated that, in addition to the accepted function as an anti-oxidant, vitamin E may also play a role in differentiation of certain cells, possibly in the control of cytokinesis or nuclear division.
Red cells from vitamin E-deficient Macaca fascicularis, as expected, are sensitive to oxidant stress (hydrogen peroxide-induced lysis)(19). Although frank vitamin E deficiency is rare in humans, individuals with β-thalassemia, a hereditary anemia caused by point mutations in the β-globin gene resulting in diminished or complete absence of β-globin chain synthesis, have low plasma levels of vitamin E and have red cells that are also sensitive to hydrogen peroxide-induced lysis(5). A more serious consequence of the globin chain imbalance in β-thalassemia erythroid cells is a severe anemia due to ineffective erythropoiesis (reduced production of erythroid progenitor cells in bone marrow) and hemolysis (reduced lifespan of circulating red cells). Both phenomena stem from the inability of unmatched α-hemoglobin molecules to form homotetrameric α-hemo-globin (in contrast to the situation in α-thalas-semia where homotetrameric β-hemoglobin, Hb

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
2
H, exists), and the unstable monomeric α-hemoglobin precipitates onto membranes of erythroid cells where they produce oxidative damage to both membrane lipid and protein components. This produces membrane defects that result in apoptosis of erythroid precursor cells in bone marrow and accelerated clearance of effete red cells by the body’s reticulo-endothelial system. Chronic oxidative stress in β-thalassemia individuals consumes the body’s store of the natural lipid-soluble antioxidant, vitamin E, and hence accounts for the hypo-vitamin E status.
Unlike anemia in premature children or protein-calorie malnutrition, vitamin E supple-mentation (525 IU for 3 months) does not improve anemia of β-thalassemia but can restore plasma vitamin E to (near) normal level(6,8,21). However, plasma vitamin E level returns to the original avitaminosis state upon withdrawal of the vitamin supplement. Nevertheless, various oxidative damages to the lipid components of red cell membrane, viz. lipid peroxidation, exposure of phosphatidylserine (PS) at the cell surface (due to loss of phospholipid asym-metry), activation of prothrombinase activity, are corrected. Disruption to the asymmetric organization of phospholipids in thalassemic red cell membrane bilayer allowing PS to be exposed on the outer surface affords an explanation for enhanced erythrophagocytosis and thrombotic episodes and cardiopulmonary anomalies frequently observed in β-thalassemic patients. Long term studies of the possible beneficial role of vitamin E supplementation in β-thalassemia are clearly needed. It is worth pointing out that there are about 600,000 thalassemic patients (1% of the population) in Thailand, with 12,000 new cases born each year (about one thalassemic child per hour), which will increase (if there are no control measures) due to the increase in population.
Although studies in vitamin E-deficient monkey can be directly translated to man, maintenance of a primate colony at the Faculty of Science proved untenable due to both health and ethical concerns. When Dr. Prayad Koma-ratat returned from her PhD studies in Canada in
1974, Dr. Serene encouraged her to develop an alternative experimental animal model. Vitamin E deficiency requires the feeding a basal diet devoid of vitamin E to a rapidly growing animal, so as to allow depletion of endogenous vitamin E store. Literature indicated that rodents, such as mouse and rat, are rather refractory to depletion of this fat soluble vitamin (due to a relative small increase in body size), and so the choice fell on the rabbit.
One-month old weanling rabbits, 1 kg in weight, when fed a vitamin E-depleted basal diet (in powder form) have growth rates similar to control animals for the first 40 days, but thereafter cease to grow and their body weights decline(22). Rabbits were considered deficient in vitamin E when plasma vitamin E fell to an undetectable level, red cells were sensitive to hydrogen peroxide lysis and animals showed signs of muscular weakness.
As a tool to probe possible perturbation to membrane function resulting from depletion of vitamin E, incorporation of radiolabeled fatty acids (oleic and palmitic) into red cells were compared between experimental and control rabbits, revealing that uptake of both kinds of fatty acids is 2-fold higher in vitamin E-deficient red cells(20). This is not due to a decrease in fatty acid pool size or an increase in acyltransferase activity, and the phenomena were ascribed to a higher turnover of phospho-lipids, supporting a role of vitamin E in maintaining membrane stability postulated by Professor Jack Lucy(13).
It is well recognized that properties of membrane-bound enzymes are influenced by the nature of membrane lipids, which would have been affected by vitamin E depletion. In order to examine the effects of vitamin E deficiency on such enzymes, Arrhenius profiles (plot of log of enzyme activity versus reciprocal of absolute temperature) were obtained for 2 mitochondrial (oligomycin-sensitive ATPase, β-hydroxybutyrate dehydrogenase), 2 microsomal (glucose-6-phos-phatase, NADH cytochrome C reductase) and one sarcoplasmic reticulum (SR Ca2+-ATPase) membrane-bound enzymes. In control rabbit, all 5 enzymes exhibit a single

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
3
discontinuity in the Arrhenius plot over the range 16-19°C, whereas those from vitamin E-deficient animal are linear or have multiple discontinuities(22). These results were interpreted as reflecting alterations in the vitamin E-depleted lipid microenvironment, which in turn impact on membrane-associated enzymes.
However, we concluded the paper with the statement: “These modifications to the membrane lipid components may not be amenable for detection by the usual chemical analysis”(22). This was to take into account the prevailing concept at that time of “annular lipids”, which provide a proper micro-environment to integral membrane proteins, an attractive notion, but evidences for their existence were equivocal(17). Professor Dennis Chapman and his colleagues had employed a number of physical techniques demonstrating that the discontinuity in the Arrhenius plot of SR Ca2+-ATPase as measured using rate of ATP hydrolysis can be rationalized in terms of a sudden change in conformational and aggre-gational states of the enzyme and independent of the type of lipids in which it is dissolved(7). When delipidated SR Ca2+-ATPase from vitamin E-deficient rabbit was reconstituted with lipids from control SR, it still retains the original properties, namely, low ATPase activity and a lack of discontinuity in the Arrhenius plot, whereas in the converse experiment delipidated normal SR Ca2+-ATPase in the presence of lipids from vitamin E-deficient SR has reduced ATPase activity but retains the Arrhenius discontinuity(14). These results demonstrated that the altered (Arrhenius) property of SR Ca2+-ATPase is due to direct changes in Ca2+-ATPase and not from the lipid environment. The reduction in ATPase activity was interpreted as owing to inhibition by cholesterol, which is elevated in SR from vitamin E-deficient atrophic muscle(11).
In order to obtain a more consistent preparation of vitamin E-deficiency SR Ca2+-ATPase (it was difficult to judge when a rabbit had become sufficiently vitamin E-deficient to manifest the changes in SR Ca2+-ATPase properties), a chemically-induced enzyme
mimic was successfully produced by treating normal rabbit SR with ascorbic acid and ferrous sulfate in the cold(16). This allowed more extensive biochemical investigations of the altered enzyme to be conducted. Both iron-ascorbate-treated and vitamin E-deficiency Ca2+-ATPase contain half the normal sulfhydryl content, but disulfide and free amino contents are unaltered, indicating that loss of sulfhydryl groups (possibly through lipid peroxidation) is responsible for the altered properties(12). The extent of protein crosslinking employing dimethyl suberimidate and copper-phenan-throline reagent at above and below the Arrhenius discontinuity temperature is similar for the three SR Ca2+-ATPase preparations (iron-ascorbate-treated, vitamin E-deficiency and control), ruling out changes to protein aggregational state in the experimental samples as an explanation for the aberrant Arrhenius profiles.
Access to an autopsied muscle of a young woman with severe β-thalassemia allowed preliminary studies to be conducted on human SR Ca2+-ATPase. Surprisingly the enzyme preparation is similar to that of vitamin E-deficient rabbit enzyme(25). Muscle wasting is a common feature in the terminal stage of thalassemia, but how much of this stems from the consequence of low vitamin E status remains unclear. (Unfortunately a case control autopsy sample was not available at that time.)
The observation that muscle from vitamin E-deficient rabbit contain higher levels of cholesterol than control animal led to a series of studies on the relationship between vitamin E and cholesterol metabolism. Within 10 weeks on a vitamin E-depleted diet, weanling rabbits become hypercholesterolemic, with a plasma cholesterol some 1.6-folds higher than control animals, and this elevation in cholesterol is found in LDL and VLDL fractions, whereas HDL-cholesterol remains unchanged(3). Rabbits fed the basal diet supplemented with vitamin E (21 to 2100 IU per week for 9 weeks) have no signs of vitamin E deficiency, but there are dose-dependent decreases in plasma LDL- and VLDL-cholesterol with increase in vitamin E

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
4
dosage, whereas there is an elevation in the level of (beneficial) HDL-cholesterol(10).
These results suggested that vitamin E supplementation should be able to ameliorate hypercholesterolemia. Rabbits were fed a diet containing from 0.25-0.5% (w/w) cholesterol, with and without 2100 IU vitamin E per week(15). In the first 4 weeks plasma cholesterol increases 10-fold in all experimental groups, but declines in rabbits given vitamin E, reaching a level half of that of the unsupplemented group in the 8th week. Again the decline in plasma cholesterol is associated with LDL and VLDL fractions and HDL-cholesterol is unchanged.
Plasma cholesterol is taken up by the liver, via chylomicron and LDL receptors, where it utilized in a number of ways: used for steroid synthesis, incorporated into membrane, stored in esterified form, exported in VLDL and excreted into gall bladder. Activity of liver microsomal cholesterol 7α-hydroxylase (now known as CYP7A1, a member of the large family of cytochrome P450 enzymes), the rate limiting enzyme in the conversion of cholesterol to bile acids, is reduced in vitamin E-deficient rabbit (9th or 10th week) to 20% of control value(3). Liver microsomal cholesterol content in diet-induced hypercholesterolemic rabbit is reduced when supplemented with vitamin E, concomitant with an increase in cholesterol 7α-hydroxylase activity, and furthermore, there is a rise in bile acid concentration with improved lithogenic indices(15).
The explanation afforded is that the rapid onset of hypercholesterolemia in the rabbit is due to an inability to shunt the abnormally excessive dietary cholesterol (which is normally minimal for a herbivore) into the bile. Pharma-cological concentration of vitamin E elevates liver cholesterol 7α-hydroxylase activity, allowing cholesterol to be excreted and thereby reducing the amount that is exported into plasma. To eliminate the possibility that the action of vitamin E is also due to enhanced LDL-mediated liver uptake, pharmacokinetic studies were conducted of radiolabeled (human) LDL injected into the experimental animals, which demonstrated no differences between
cholesterol-fed rabbits with and without vitamin E dosing (Porntadavity C, Komaratat P, Wilairat P, unpublished). It is not clear how vitamin E regulates liver cholesterol 7α-hydroxylase activity. The original notion was that vitamin E protects against oxidative damage of the redox-sensitive cytochrome P450 enzyme. More recently, cultured human hepatocytes treated with vitamin E [(RRR)-R-tocopherol] have altered gene expression that is distinct from that elicited by another antioxidant, N-acetyl cysteine(23). In particular, vitamin E causes a significant decrease in the levels of mRNAs encoding HMG-CoA reductase, the rate limiting enzyme in cholesterol de novo biosynthesis, and LDL receptor. How vitamin E modulates rabbit hepatic genes expression awaits future dis-covery.
Vitamin E deficiency in subjects with chronic cholestasis is sometimes associated with areflexia, peripheral neuropathy, cerebellar involvement with gait and limb ataxia and decreased propioperception and vibration sense, all of which can be reversed by correcting the avitaminosis-E status(18). Rabbits fed a high vitamin E diet (300 mg/100 g of diet for 10 weeks) have a higher density of glutamate receptors in cerebellar membrane than those fed a lower vitamin E diet (0.56 and 3 mg/100 g diet) over the same period(9), suggesting that this vitamin is able to influence glutamate binding sites in this region of brain, either by affecting membrane structure directly and/or through modulating expression of the receptors.
What is the future of vitamin E research? Much current attention is being focused on non-antioxidant properties of vitamin E (and metabolites), including transcriptional regula-tion, apoptosis, cell adhesion and inhibition of enzymes(26). This is rather satisfying given the prescient statement 30 years ago by neophytes in vitaminology that “vitamin E may also play a role in differentiation of certain cells”(19).
Acknowledgements
The author thanks all colleagues who were involved in the research, especially Associate Professor Prayad Komaratat, the principal investigator of the rabbit studies. The

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
5
author is indebted to the skill and dedication of the graduate students who made the work possible. Parts of the article have previously appeared in Settasatian NC, Wilairat P, Komaratat P. Effect of vitamin E status on hepatic cholesterol 7α-hydroxylase. In CRC Handbook of Free Radicals and Antioxidants in Biomedicine, Vol II (Miquel J, Quintanilha AT, Weber H, ed). CRC Press Inc, Boca Raton, Florida 1989; p. 161-5; and Wilairat P, Koma-ratat P. Hypocholesterolemic effect of vitamin E. Biochem (Life Sci Adv) 1993;12: 53-5.
References 1. Boonjawat J, Wilairat P, Vimokesant SL.
Alteration in bone marrow RNA of vitamin E-deficient monkey, Macaca fascicularis. Am J Clin Nutr 1979;32:2065-7.
2. Chadd MA, Fraser AJ. Vitamin E deficiency in premature infants. Inter J Vit Res 1970;40:604-9.
3. Chupukcharoen N, Komaratat P, Wilairat P. Effects of vitamin E deficiency on the distribution of cholesterol in plasma lipoproteins and the activity of cholesterol 7α-hydroxylase in rabbit liver. J Nutr 1985;115:468-72.
4. Dinning JS, Day PL. Vitamin E deficiency in the monkey. I. Muscular dystrophy, hematolo-gic changes, and the excretion of urinary nitrogeneous constituents. J Exp Med 1957;105: 305-402.
5. Fucharoen S, Winichagoon P. Hemoglobinopa-thies in Southeast Asia: molecular biology and clinical medicine. Hemoglobin 1997;21:299-319.
6. Fucharoen S, Chantharaksri U, Wilairat P, Bunyaratvej A, Pootrakul P. Oxidative stress in thalassemia: a reappraisal. Trends Biomed Finland 1998; 1 Suppl :15-6.
7. Hoffmann W, Sarzala MG, Chapman D. Rotational motion and evidence for oligomeric structures of sarcoplasmic reticulum Ca2+-activated ATPase. Proc Natl Acad Sci USA 1979;76:3860-4.
8. Kasemsant S, Wilairat P. Effect of vitamin E on red blood cell procoagulant activity in β-thalassemia/Hb E disease. In: Packer L, Traber MG, Xin W, editors. Proceedings of the International Symposium on Natural Anti-oxidants: Molecular Mechanisms and Health Effects. USA: AOCS Press; 1996. p. 162-6.
9. Kittiwatanapaisan W, Wilairat P, Kotchabhakdi N, Govitrapong P. Effect of dietary vitamin E on L-[3H]glutamate binding in rabbit cerebellum. J Nutr Sci Vitaminol 1993;39:81-8.
10. Komaratat P, Chupukcharoen N, Wilairat P. Effect of vitamin E on cholesterol plasma lipoprotein distribution and metabolism in rabbit. Intern J Vit Nutr Res 1985;55:167-71
11. Liewsaree P, Yuthavong Y, Wilairat P, Koma-ratat P. Protein and lipid composition of sarcoplasmic reticulum from dystrophic muscles of vitamin E-deficient rabbits. Nutr Reports Intern 1980; 22: 853-62.
12. Nirdnoy W, Komaratat P, Wilairat P. Compari-son of sarcoplasmic reticulum Ca2+-adenosine triphosphatase from vitamin E-deficient dystrophic rabbit skeletal muscle with iron-ascorbate-treated and untreated enzyme. J Biochem 1988;103:309-12.
13. Lucy JA. Functional and structural aspects of biological membranes: a suggested structural role for vitamin E in the control of membrane permeability and stability. Ann NY Acad Sci 1972:203:4-11.
14. Patipaporn K, Wilairat P, Komaratat P. Altered property of sarcoplasmic Ca-ATPase from vitamin E-deficient dystrophic rabbit is associated with the protein and not the lipid component. Biochem Intern 1983;6:334-8.
15. Phonpanichrasamee C, Komaratat P, Wilairat P. Hypocholesterolemic effect of vitamin E on cholesterol-fed rabbit. Intern J Vit Nutr Res 1990; 60:240-4.
16. Promkhatkaew D, Komaratat P, Wilairat P. Ascorbic acid-Fe2+ treatment mimics effect of vitamin E deficiency on sarcoplasmic Ca-ATPase of rabbit muscle. Biochem Intern 1985;10:937-43.
17. Quinn PJ, Chapman D. The dynamics of membrane structure. CRC Crit Rev Biochem 1980;8:1-117.
18. Ricciarelli R, Argellati F, Pronzato MA, Domenicotti C. Vitamin E and neuro-degenerative diseases. Mol Aspects Med 2007;28:591-606.
19. Santiyanont R, Yaipimol C, Wilairat P. Accu-mulation of orthochromatophilic normoblasts in bone marrow of vitamin E-deficient monkey, Macaca fasicularis [sic]. J Nutr 1977;107:2026-30.
20. Ucchin P, Komaratat P, Wilairat P. Effect of vitamin E deficiency on fatty acid uptake of rabbit erythrocyte. In: Khor HT, Ong KK, Oo KC, editors. Food and nutritional biochemistry. Kuala Lumpur: Malaysian Biochemical Society, Malaysia; 1980. p. 237-47.
21. Unchern S, Laoharuangpanya N, Phumala N, Sipankapracha P, Pootrakul P, Fucharoen S, Wanachivanawin W, Chantharaksri U. The effects of vitamin E on platelet activity in β-

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
6
thalasaemia patients. Br J Haematol 2003;123: 738-44.
22. Vajanamarhutue C, Wilairat P, Komaratat P. Effects of vitamin E deficiency on the activities of lipid-requiring enzymes in rabbit liver and muscle. J Nutr 1979;109:848-55.
23. Valastyan S, Thakur V, Johnson A, Kumar K, Manor D. Novel transcriptional activities of vitamin E: inhibition of cholesterol biosynthesis. Biochemistry 2008;47:744-52.
24. Whitaker JA, Fort EG, Vimokesant S, Dinning SJ. Hematologic response to vitamin E in the anemia associated with protein-calorie malnu-trition. Am J Clin Nutr 1967;20;783-9.
25. Wilairat P, Komaratat P. Sarcoplasmic reticulum Ca-ATPase in vitamin E deficiency. In: Hayaishi O, Mino M, editors. Clinical and nutritional aspects of vitamin E. Elsevier Science Publishers; 1987. p. 85-8.
26. Zingg JM, Azzi A. Non-antioxidant activities of vitamin E. Curr Med Chem 2004;11:1113-33.

7
Chemical Biology: Addressing Biological Questions from Chemical Perspectives
Chutima Jiarpinitnun Department of Chemistry and Center for Innovation in Chemistry, Faculty of Science, Mahidol University.
Abstract
To unravel complex biological problems, multidisciplinary collaborative efforts, particularly, between chemists and biologists are highly requisite. For the past decade, chemical biology has emerged as a new vibrant scientific venue. Its origin involves scientific interest at the interface between chemistry and biology. Chemical biology prospers as the results of complemented skills and unique chemical and biological perspectives. The field is built on its foundation in chemistry, and the chemical biology community is extending its reach to understand and manipulate sophisticated biological systems. This review aims to highlight few chemical biology researches in order to illustrate the significance of fundamental chemistry in embarking upon the unaddressed biological questions. Multidisciplinary research has recently been center of attention in Thai scientific community. Chemical biology is emerged as a new scientific field of interest. The perspective on the research transition and the opportunity of conducting chemical biology research in Thailand will be discussed.
Keywords: chemical biology; natural product chemistry
Introduction
Chemical biology is difficult to define. Like other disciplines that concerned chemistry-biology interface such as biochemistry, chemical biology is a highly interdisciplinary field that requires skills and knowledge in chemistry and biology to solve scientific questions. While biochemistry can be described as the study of chemical interactions and reactions of biological molecules in biological systems, chemical biology research, on the other hand, can be less directed toward understanding nature and focuses more on exploiting chemical principle to manipulate biological components or systems. Therefore, chemical biology research can take advantage of chemical thinking to go beyond the molecules or systems offered by nature since the unique materials can be generated via chemical synthesis. These synthetic materials allow researchers to ask biological questions that would be difficult or impossible to address by using natural compounds or natural systems. Perhaps, the difference between chemical
biology and other disciplines that concerned with chemistry-biology interface is that, in the broadest sense, chemical biology applies chemical approaches to address biological questions. New compounds and new chemistry are developed and exploited as toolbox to facilitate in tackling the problem. Accordingly, in chemical biology, chemistry and biology cannot be advanced without one another. As the celebration of the 50th anniversary of the Faculty of Science, Mahidol University, several past and current researches at the Faculty of Science, Mahidol University, would be highlighted.
Addressing Biological Questions with Chemical Perspective
Chemical biology research is highly interdisciplinary, containing broad scope of knowledge and techniques from many scientific disciplines such as chemical sciences, medicinal chemistry, molecular biology, biochemistry, structural biology, bioinformatics, proteomics, genetics, and many more. As mentioned prior,

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
8
the definition of chemical biology is quite difficult to encapsulate, probably depending on the person defining it. As a result, the re-searches in this field cover very broad extent; yet, primarily, the focus is on developing of small molecules as research tools and their potential applications for solving biological problems that particularly concerned with advancing therapeutics.
Small molecules have been associated with biological discoveries for long period of time. It was considered to play a crucial role in the ‘central dogma’ of molecular biology(22), where the sequential information is flowing through the biological macromolecules: from DNA to RNA to proteins. Cells do not survive with only macromolecules; indeed, cells produce natural small molecules and trigger the signaling cascades in response to their recognitions to the small molecules in their environments. Chemical biologists focus on the design and development of small molecules that further be exploited in exerting controls over biological functions or probing the function of the biomolecules. Numerous chemical biology researches have significantly advanced the chemistry and biology. However, only few examples can be highlighted in this review.
Controlling Functions of Biological Macromolecules with Synthetic Small Molecules
Since the completion of Human Genome Project in 2003, the main challenge is to decode the information encoded in the functional genome. The number of human protein-coding genes is estimated to be around 20,000(12). With the processes such as splicing and post-translational modification, the number of functional proteome is expanded. It has been estimated that at least 106 biological molecules are required to maintain the function of human cells(3). To understand the roles of these biological molecules in cellular processes, the general approach is to perturb or disrupt the target biological molecules and evaluate the consequences. Molecular genetics can be used to eliminate specific proteins by knocking out genes of interest, or to alter the function of a
protein by making specific mutations in the genes. However, these approaches would be difficult for large genomes.
Chemical biologists utilize small mole-cules as a tool to study the roles of these biological molecules and cellular processes. The small molecules can bind to protein, modulate its functions, and perturb the cellular systems; therefore, molecular mechanisms underlying the biological processes can be revealed. The main idea is similar to molecular genetic approach; yet, instead of permanently alter or disrupt the function of a protein, small molecules can exert temporal or spatial controls over protein functions and biological processes, and these molecules can be useful for further therapeutic development. The small molecule approach is referred to as “chemical genetics.”
To discover the small molecules for each specific protein function, chemical biologists have exploited high-throughput approaches to synthesize chemical libraries for screening(21). Solid phase combinatorial synthesis using split and pool strategy is generally employed as synthetic methods. The approach is not only ease the purification step but also increase the ability of organic synthesis of produce huge collections of small molecules in short period of time. For instance, a library of 10,000 small molecules can be synthesized via three to four chemical reactions, of which each step using approximately ten chemical building blocks, and completed within days. The key factor for finding a small molecule for a particular protein is the composition of the library used in screening. For the proteins that their structures are solved or their endogenous ligands are known, the chemical library can be rationally designed, where the small molecule collections share common structural features that facilitate in binding to the preselected protein target. However, most proteins do not have the luxury of accessible structural information or of known ligand. To identify a small molecule selective for these proteins, the chemical library has to compose of structurally diverse compounds. Diversity oriented synthesis (DOS) is developed as a strategy for rapid synthesis of a collection of compounds that are complex and diverse in

structuresimultantheir smseveral moleculewith the insights discover
Idprotein olead butFor exFK BP1synthesinatural psmall mBy usingSchreibeFK506 of the complexcyclophicovery, iby Profcerning (NFAT) tion of pathwaya phospresponseimmunosystem aof organ
Tindicatedof bindforming small mThe smacrosslinkintracelluor reguThese cregulate pathwaybring thproximitapproachmanipula
es(21). DOSneously idenmall molec
institutionses libraries fcollaboratiointo biolog
red using sma
dentification of interest nt also can r
xample, the12, was discs focusing product FK5molecule 50g small moler and co-and cyclospphosphatase
xes of FKBPilin-cyclospoin combinati
fessor Crabtrthe nuclear signaling p
f calcium-cay that accounphatase can
e. FK506 suppressive after organ t
n rejection.
The discoveryd that small
ding to twoa protein-lig
molecule dimall molecule k the hybrid pular signalin
ulated by pchemical di
transcriptiys in engineehe corresponty. The bifh has beenation of th
วทยาศาส
can be usentify the protcule effectos have pfor screeningon between rgical procesall molecule
of a small not only can reveal signale FK506-bicovered viaon the imm
506, rapamy06BD, and lecules as pr-workers dporine inhibe clacineurP12-FK506-corin-calcineuion with the nree and co-factor of a
pathway, led alcineurin-Nnts for how tn suppressis currentlydrug to redu
transplant, lo
y of FK506 bmolecules h
o proteins gand complemerizers werdimerizers wproteins, the
ng that are naprotein-proteimerizers wion and mered cells bynding hybridfunctional s
n further exhe extensiv
สตรเพอการพฒนา :
ed as a tootein targets ors. Curren
provided smg; in conjuncresearchers, nsses have bs as probes.
molecule foprovide a d
ling networkinding prota target-orienmunosupprescin, non-natcyclosporin
robes, Profedetermined bit the actirin thoughcalcineurin,
urin. Thisnotable findi-workers(5) cactivated T-c
to the elucNFAT signa
the inhibition the immy used as uce the immowering the
binding partnhave the abisimultaneou
ex. As the resre develope
were designeereby controlaturally occurin interactio
were shownmany signay their abilityd proteins small molecxploited in ve network
รวมบทความวชาการ
l to and
ntly, mall tion new been
or a drug k(26). tein, nted sant tural n(20). ssor that vity the and dis-ings con-cells ida-
aling n of
mune an
mune risk
ners ility
usly, sult, ed(4). d to
lling rred ons. to
aling y to into cule the of
intefunnorprosma
impprofunNevcormostudSevdevcallmurecligawhmoare recproproof bioligatooKiethaattreluintesign
moselesmathe
F
ร ฉลองครบรอบ 50 ป
eracting prnctions are emrmally do nooximity withall molecule(
Protein-pportant in oteins do nonction in dimvertheless, rresponding olecule to itsdied on averal chemicveloping a uled “multiv
ultivalent inteptor and tands are genich numbers
olecule that battached, t
eptor simultoximity (Figoach can be in
potent effecomolecules(11
ands have alol to probe sessling and t the linearactants coulcidate the eractions in tnals.
These eolecules canectively contall molecule
dynamics o
Figure 1 Mu into
ป แหงการสถาปนาค
oteins. In merged whenot interact, a one anothe(7).
protein intersignal tran
t operate inmeric or olithe bindinbiological a
s bimoleculaa single ligcal biologisunique type
valent liganteractions btheir stimulinerally comps of recognitbinds to thethereby can taneously or gure 1). Thncorporated ctors or inh). Remarkab
lso been empsignal transd
her co-worar polymers ld be used
role of transducing b
examples aren be used trol the bioloes, new insiof ligand-pr
ultivalent ligano proximity.
คณะวทยาศาสตร มหา
some casn two proteinare forced inr by the bifu
ractions are nsductions.
ndividually bigomeric co
ng events activity of tar target are gand-receptosts have foce of small nds” to mibetween celli(10). The muposed of a sction epitopes receptor of
potentially r bring recephe mutlivaleinto the dev
hibitors of thbly, these muployed as a ductions(11). Prkers(6) dem
displaying as chemicalinter-chemo
bacterial che
e shown thto reversi
ogical networight informarotein interac
nd brings rece
าวทยาลยมหดล
es, new ns, which nto close
functional
critically Many
but rather omplexes. and the
the small typically
or basis. cused on molecule imic the l surface ultivalent caffold to s, a small f interest,
bind to ptors into ency ap-elopment he target ultivalent chemical Professor
monstrated chemo-
tools to oreceptor emotactic
hat small ibly and rk. Using
ation into ctions or
eptors
9

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
10
protein-protein interactions and their importance in biology could potentially be revealed.
Small Molecules as Tools to Probe Biological Signaling
In the post-genomic era, one of the main challenges is to determine how proteins function together in complex networks that direct a diverse set of cellular processes. Only manipulating or disrupting the function of biomolecule is not adequate to illuminate the complicated signaling network. To dissect complex cellular processes, the ability to visualize biomolecules within their native physiological condition or to track the small molecule modulators is additionally required.
In the past decade, the studies of protein functions have been heavily based on auto-fluorescent proteins; in fact, the discovery and development of green fluorescent protein (GFP) was awarded the Nobel Prize in Chemistry in 2008(2). Despite the impressive discoveries based on auto-fluorescent proteins, genetically encoded tag such as GFP can cause significant perturbations to the structure of labeled protein. In addition, some crucial biomolecules or some enzymatic activities cannot be detected by auto- fluorescent proteins. Fundamental chemistry has been playing an important role in the development of tools for biomolecular imaging. Synthetic fluorescent small molecules have been developed as complement approaches to auto-
developed as
developed as complement approaches to auto-fluorescent proteins to gain new insights into cellular networks. For example, the sensor, based on a Cu(II) complex of fluorescein derivative, has been developed to directly detect the second messenger, nitric oxide(14) (Figure 2A). The fluorescent based sensor is cell permeable and shows high specificity for nitric oxide over other reactive nitrogen or oxygen species. This fluorescent small molecule allows the visualization of nitric oxide formation in living cells. Another great example demon-strated how chemical thinking can be cleverly incorporated into the design and synthesis of chemical probes is the development of latent-fluorophores(13) (Figure 2B). The ‘trimethyl lock’ fluorogenic probe is converted from a non-fluorescent to highly fluorescent species by cytosolic esterases. Esterases cleave the ester functional group on the trimethyl lock, leading to rapid lactonization, subsequently releasing the highly fluorescent fluorophore. With this synthetic small molecule, the internalization of labeled biomolecules can be visualized with temporal and spatial resolution in living cells.
Networks of interacting biomolecules, ions, and metabolites are driving a complex array of cellular processes in living cells. Few biomolecules naturally possess unique features that allow the direct detection in the complex milieus; yet, many of them do not. The chemical
Figure 2. Small molecules as fluorescent based sensors.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
11
tool for tagging biomolecules has been developed from the chemical biology com- munity. Antibody conjugates have been widely used to track biomolecules in living cells; however, the large size of antibody conjugates hinders its access. Small molecules provide a better access to intracellular compartments. However, to use a small molecule to track biomolecules in the living cells, specificity is crucial. Most types of biomolecules contain nucleophilic functionality that can be readily conjugated to most types of chemical probes such as biotin or fluorophores. Thus, it is chemically challenging to develop the site-specific chemical modification of biomolecules within their native environment. Chemical biologists pioneered the development of bioorthogonal chemical reporter strategy. In brief, the chemical reporter is incorporated into the target biomolecules(17) (Figure 3). The incorporation can be achieved via the cellular machinery. For example, unnatural amino acids bearing bioorthogonal functional group can be incorporated into target proteins in residue-specific manner using auxotrophic strains of E.coli(23) or in site-specific manner using non-sense suppression approach(15); likewise, mono-saccharide derivatives bearing bioorthogonal functional groups can be introduced into glycans via enzymes in the glycan biosynthetic path-ways. Once the bioorthogonal chemical reporter is introduced into a biomolecular target, the
reporter can be covalently reacted with the chemical probe bearing a complementary chemical moiety. The reaction between the chemical reporter and the chemical probe requires high specificity; the chemical moieties on both the chemical reporter and chemical probe must be inert to the non-target molecules
in the cellular milieu. Beyond that specific re-quirement, the reaction must be done in a physiological condition. With the reaction condition constraints, only a small number of chemical transformations can be employed.
However, these handful examples can evidently illustrate the impact of chemical thinking in biological discoveries. For instance, Professor Tsien and his co-workers(8) have cleverly taken advantage of unique combination of amino acid side chains to create new functionality that suit the criteria of bio-orthogonal chemical reporter. The sequence containing tetracysteine motif –CCXXCC– can react selectively to membrane permeable biarse-nical dyes, which contain ethanedithiol substi-tuents to prevent the labeling of any bio-molecules bearing isolated cysteine residues. This tetracysteine-biarsenical dye system pro-vides an alternative route to visualize protein of interest. However, this strategy is limited to proteins since the tetracysteine tag is unlikely to be tolerated by biosynthetic enzymes of other biomolecules and metabolites. Smaller chemical reporter is desired. Azides have emerged as an alternative chemical reporter as they are hardly found in any natural biomolecules and do not re act with water as well as are resistant to oxidation. The bioorthogonal reactions with azides have been developed. These include the Staudinger ligation of azides with the chemical probes bearing phosphines(19) and the copper-catalyzed [3+2] cycloaddition between azides with terminal alkynes(1); both reactions can readily proceed in physiological conditions. These chemical reporters have been exploited in monitoring biomolecules and enzyme activities in cellular systems.
-
Figure 3 Bioorthogonal chemical reporter.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
12
By developing small molecule to potently exert control over biological functions and expanding its use of many discovered bio-logically active compounds, the chemical biology community not only provides chemical probes to investigate the biological signaling pathways and also new candidates for therapeu-tic development. As chemical tools, chemical biologists have shown their creativity to innovate the strategies and techniques, based on fundamental chemistry, to extend the study of complicate biological systems that would be difficult or impractical to accomplish otherwise. Highlighted in this review are only few examples that underscore the great impact that chemical biology has had on life sciences.
From Thai Medicinal Herbs to Chemical Biology
As many exciting unaddressed questions in science lie at the interfaces between scientific disciplines, the multidisciplinary research has recently been center of attention in Thai scientific community. Clearly, the traditional boundaries between scientific disciplines have become seamless. Many Thai scientists have put in substantial effort to interweave ideas from chemistry and biology to solve trivial problems.
Chemistry research in Thailand has roots in natural product chemistry. At the beginning, the natural product research mainly focused on the isolation and characterization of Thai medicinal herbs. Several bioactive molecules, isolated and characterized from Thai medicinal herbs, have been making significant contribution to therapeutic development. Excellent example is the finding of gambogic acid constitution by Professor Stang Mongkolsuk(16), the founder of the Faculty of Science at Mahidol University, and his co-workers. Gambogic acid was found to possess significant anti-cancer activity both in vitro and in vivo(9). Professor Vichai Reutrakul and co-workers(18) further carried out the structure-activity relationship of the gambogic acid; the finding facilitates the design and development of potent gambogic acid deri-vatives. Following on these accomplishments, part of the current natural product chemistry research is still focusing on the isolation and
characterization of complicated unknown mole-cules from the unique natural sources, including plants and microorganisms, found locally. Another main area of natural product research is focusing on the bioactive natural products and the establishment of arrays of enzyme and cell-based, and in vivo assays, fostering the develop-ment of new synthetic strategies to gain under-standing of molecular structure-function rela-tionship. These research advances provide a golden opportunity for chemical biology researchers in Thailand in utilizing the natural product molecules to expose biological path-ways.
The chemistry-biology interface in Thailand had been initiated in the pursuing bioactive natural products. Collaborative efforts between Thai chemists and biologists drive the success of natural product research in Thailand. As demonstrated in the discovery of VR-3848, a novel natural cyclic peptide isolated from Euphobiaceae (locally known as Takhe khum- wang), subsequent to the isolation and charac- terization of the structurally complicated com-pound, VR-3848 was tested for its ability to inhibit tumor growth in tumor cell lines(29). The compound was shown to exert potent growth-inhibitory effects in many tumor cell lines. With the effective collaboration with the biologists, the mechanism underlying the ability of VR-3848 to kill tumor cells was investigated. The study revealed that VR-3848 potently induced the activation of multiple apoptotic pathways in mitochondria-dependent manner, with the IC50 of low nanomolar(28).
Identification of the biomolecular target of the bioactive small molecules would facilitate in the elucidation of the signaling pathways that lead toward the observed in the corresponding biological activities. The binding partner of many of the bioactive natural products found in Thai natural sources have not yet been iden-tified. Natural products can be used as a tool to probe for their biological target; this aspect, which is using small molecules to explore biological pathways, is heavily emphasized in chemical biology research. Laboratory of Associate Professor Palangpon Kongsaeree at the Faculty of Science, Mahidol University,

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
13
immobilized natural product of interest to the magnetic nanoparticle. Taking advantage of the magnetic property, the natural product-target protein complex can be pulled out, leaving the unbound molecules behind. The complex is then cleaved from the magnetic nanoparticle and subsequently analyzed by mass spectroscopy. The identification of the natural product target would potentially open up more research venues in chemistry and biology and provide new therapeutic targets.
Regarding to the therapeutic develop-ment, the great hallmark of antimalarial thera-peutic developments in Thailand has empha-sized the accomplishment when chemistry and biology are merged. Antifolate antimalarial small molecules function by inhibiting dihydro-folate reductase (DHFR), thereby reducing amount of the essential folate cofactor of the parasite(30). The resistance of these antifolate antimalarials is resulted from the change in DHFR, decreasing the binding ability of the inhibitor. Professor Yongyuth Yuthavong and his co-workers(24) employed molecular genetics and structural biology to obtain the useful insights for the development of antifolates to overcome the resistance. In collaboration with Professor Yodhathai Thebtaranonth and his co-workers, new antifolate antimalarial small mole-cules are rationally designed(25). In addition, the team has developed a high-throughput method for screening inhibitor of DHFR mutants as well as synthetic methodologies that are applicable for synthesizing chemical libraries(27, 31).
It should also be highlighted that the cutting edge research in chemical biology highlighted above is made possible by the investment in the state of the arts facilities and equipment by The Faculty of Science Mahidol University, Center for Innovation in Chemistry (PERCH-CIC), and National Center for Genetic Engineering and Biotechnology (BIOTEC).
Conclusions
The examples mentioned above well illustrate how chemical biologist utilize chemical thinking to innovatively create small molecules to investigate complex biological systems. The discoveries have been making
significant advances in both fields of chemistry and biology. In Thailand, natural product chemistry has traditionally been the field of interest and now provided a basis to advance of chemical biology research in the country. Chemical biology research offers the ways to expand the application of natural compounds in life sciences. The precious small molecules isolated from unique natural sources in Thailand provide an invaluable opportunity for chemical biology research. Utilizing these molecules to exert controls over biological functions or developing chemical tools to expand the use of these molecules could potentially make signi-ficant impact. At the present, Thai scientific community has actively initiated the field of chemical biology. Many chemical biologists have been invited to give lectures in Thailand. The funding agencies have provided financial supported interdisciplinary research grant. The research and postgraduate training program in the area of chemical biology has also been supported by the Center for Innovation in Chemistry (PERCH-CIC). Hence, Thai re- searchers are equipped to prosper in the chemical biology research.
Acknowledgements
The author would like to acknowledge Professor Vichai Reutrakul, Professor Manat Pohmakotr, and Associate Professor Palangpon Kongsaeree for scientific discussions and critical reading of the manuscript.
References 1. Agard NJ, Prescher JA, Bertozzi CR. A strain-
promoted [3+2] azide-alkyne cycloaddition for covalent modification of biomolecules in living systems. J Am Chem Soc 2004;126:15046-7.
2. Awad AI, Awad IA. Chemistry of life: green fluorescent protein and the 2008 Nobel Prize in Chemistry. Neurosurgery 2008;63:11.
3. Bohacek RS, McMartin C, Guida WC. The art and practice of structure-based drug design: a molecular modeling perspective. Med Res Rev 1996;16:3-50.
4. Crabtree GR, Schreiber SL. Three-part inventions: intracellular signaling and induced proximity. Trends Biochem Sci 1996;21:418-22.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
14
5. Crabtree GR, Olson EN. NFAT signaling: choreographing the social lives of cells. Cell 2002;109 Suppl :S67-79.
6. Gestwicki JE, Kiessling LL. Inter-receptor communication through arrays of bacterial chemoreceptors. Nature 2002;415:81-4.
7. Gestwicki JE, Marinec PS. Chemical control over protein-protein interactions: beyond inhibitors. Comb Chem High Throughput Screen 2007;10:667-75.
8. Griffin BA, Adams SR, Tsien RY. Specific covalent labeling of recombinant protein mole-cules inside live cells. Science 1998;281: 269-72.
9. Gu H, You Q, Liu W, Yang Y, Zhao L, Qi Q et al. Gambogic acid induced tumor cell apoptosis by T lymphocyte activation in H22 transplanted mice. Int Immunopharmacol 2008;8:1493-502.
10. Kiessling LL, Gestwicki JE, Strong LE. Synthetic multivalent ligands in the exploration of cell-surface interactions. Curr Opin Chem Biol 2000;4:696-703.
11. Kiessling LL, Gestwicki JE, Strong LE. Synthetic multivalent ligands as probes of signal transduction. Angew Chem Int Ed Engl 2006; 45:2348-68.
12. Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J et al. Initial sequencing and analysis of the human genome. Nature 2001; 409:860-921.
13. Lavis LD, Chao TY, Raines RT. Fluorogenic label for biomolecular imaging. ACS Chem Biol 2006;1:252-60.
14. Lim MH, Wong BA, Pitcock WH, Jr., Mokshagundam D, Baik MH, Lippard SJ. Direct nitric oxide detection in aqueous solution by copper(II) fluorescein complexes. J Am Chem Soc 2006;128:14364-73.
15. Noren CJ, Anthony-Cahill SJ, Griffith MC, Schultz PG. A general method for site-specific incorporation of unnatural amino acids into proteins. Science 1989;244:182-8.
16. Ollis WD, Ramsay MVJ, Sutherland IO, Mongkolsuk S. Constitution of gambogic acid. Tetrahedron 1965;21:1453-70
17. Prescher JA, Bertozzi CR. Chemistry in living systems. Nat Chem Biol 2005;1:13-21.
18. Reutrakul V, Anantachoke N, Pohmakotr M, Jaipetch T, Sophasan S, Yoosook C et al. Cytotoxic and anti-HIV-1 caged xanthones from the resin and fruits of Garcinia hanburyi. Planta Med 2007;73:33-40.
19. Saxon E, Bertozzi CR. Cell surface engineering by a modified Staudinger reaction. Science 2000;287:2007-10.
20. Schreiber SL. Chemistry and biology of the immunophilins and their immunosuppressive ligands. Science 1991;251:283-7.
21. Schreiber SL. Target-oriented and diversity-oriented organic synthesis in drug discovery. Science 2000;287:1964-9.
22. Schreiber SL. Small molecules: the missing link in the central dogma. Nat Chem Biol 2005;1:64-6.
23. Sharma N, Furter R, Kast P, Tirrell DA. Efficient introduction of aryl bromide functionality into proteins in vivo. FEBS Lett 2000;467:37-40.
24. Sirawaraporn W, Sathitkul T, Sirawaraporn R, Yuthavong Y, Santi DV. Antifolate-resistant mutants of Plasmodium falciparum dihydrofolate reductase. Proc Natl Acad Sci USA 1997;94: 1124-9.
25. Sirichaiwat C, Intaraudom C, Kamchonwong-paisan S, Vanichtanankul J, Thebtaranonth Y, Yuthavong Y. Target guided synthesis of 5-benzyl-2,4-diamonopyrimidines: their antima-larial activities and binding affinities to wild type and mutant dihydrofolate reductases from Plasmodium falciparum. J Med Chem 2004; 47:345-54.
26. Spencer DM, Wandless TJ, Schreiber SL, Crabtree GR. Controlling signal transduction with synthetic ligands. Science 1993;262:1019-24.
27. Thongpanchang C, Taweechai S, Kamchon-wongpaisan S, Yuthavong Y, Thebtaranonth Y. Immobilization of malarial (Plasmodium falciparum) dihydrofolate reductase for the selection of tight-binding inhibitors from combinatorial library. Anal Chem 2007;79: 5006-12.
28. Ubol S, Kramyu J, Masrinoul P, Kachangchaeng C, Pittayanurak P, Sophasan S et al. A novel cycloheptapeptide exerts strong anticancer activity via stimulation of multiple apoptotic pathways in caspase-3 deficient cancer cells. Anticancer Res 2007;27:2473-9.
29. Uthaisang W, Reutrakul V, Krachangchaeng C, Wilairat P, Fadeel B. VR-3848, a novel peptide derived from Euphobiaceae, induces mito-chondria-dependent apoptosis in human leukemia cells. Cancer Lett 2004;208:171-8.
30. Vilaivan T, Saesaengseerung N, Jarprung D, Kamchonwongpaisan S, Sirawaraporn W, Yuthavong Y. Synthesis of solution-phase combinatorial library of 4,6-diamino-1,2-

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
15
dihydro-1,3,5-triazine and identification of new leads against A16V+S108T mutant dihydro-folate reductase of Plasmodium falciparum. Bioorg Med Chem 2003;11:217-24.
31. Yuthavong Y. Basis for antifolate action and resistance in malaria. Microbes Infect 2002;4: 175-82.


17
Forensic Science at the Faculty of Science, Mahidol University in the International Forensic Science Arena
Nathinee Panvisavas Department of Plant Science and Forensic Science Program, Multidisciplinary Unit, Faculty of Science, Mahidol University.
Abstract
The Master of Science in Forensic Science International Program at the Faculty of Science, Mahidol University was established in 2004, with the aim of producing forensic scientists with professional skills and ethics. The program was designed to fulfill the country’s needs for forensic science personnel by providing postgraduate education for existing or future law enforcement personnel and forensic laboratory analysts. This article discusses the establishment of the program, its activities, the curriculum and training scheme, and university’s role in developing forensic practitioners.
Keywords: forensic science; SCFS
Introduction
Forensic Science is a popular field for the Thai postgraduate study market during the past several years. The re-trial of the high profile case of MP Hangthong Thamwattana, the renown of the Thai forensic scientist Dr. Porntip Rojanasunan, together with the “CSI” television series, has made Forensic Science became more familiar and better known to the Thai community. The Hangthong case created an alert to the Thai nation on the criminal justice procedure. The high-profile case was re-trialed twice due to the lack of knowledge, understanding, and competent forensic science skill in justice. In this case, requests had been made for foreign forensic experts to testify and give expert opinions. These made the nation questioned the law enforcement and the criminal justice system and it was realized since then that Thai judicial and law enforcement authority lacks forensic practitioners to scientifically clarify the matters and to give expert testimony. Taking these matters into consideration, the Faculty of Science, Mahidol University launched the first and only Master of Science in Forensic Science International Program in Thailand(17).
Crimes have no borders. Forensic labora-tories work to a common high standard to enable cross-laboratory communication. Laboratory accreditation is a platform to maintain standards across different forensic laboratories. Indeed, ‘good forensic science’ is needed for achieving accreditation. Forensic science involves the selection, recovery, analysis, interpretation and reporting of physical evidences(12). Although there is no room for error in forensic activity, mistakes can still happen in forensic science processes that are systematically designed. It is important for the forensic scientist on duty to understand what needs to be done and why. So, good forensic science requires that a competent forensic scientist properly applies good science to produce “quality” work, and provides adequate explanations to assist the court(12). This would certainly help minimize chance of miscarriage of justice or wrongful convictions.
From Past to Present
In 2004, Faculty of Science, Mahidol University launched the Master of Science in Forensic Science (International Program) to fulfill the country’s need for qualified forensic practitioners, experts, and to help drive Thai forensic science into the international arena(17,18).

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
18
Establishment of the program was a colla-borative effort between the Faculty of Science and the Faculty of Graduate Studies, Mahidol University, with close connection to the Centre for Forensic Science (former Forensic Science Unit), University of Strathclyde in UK, with support from the two major government forensic science agencies: the Royal Thai Police Office of Forensic Science (RTPOFS), Royal Thai Police, and Central Institute of Forensic Science (CIFS), Ministry of Justice. Every academic year, the program committee makes 20 offers to potential candidates. The average class size is 15 post-graduate students.
To date, there have been five classes, in which 45 students have completed their M.Sc.
staff development programs of RTPOFS and CIFS were continuously assisted by SCFS. Since 2005, CIFS continuously supported two forensic officers to continue postgraduate study. More recently, the Australian Federal Police (AFP), which has been working closely to RTPOFS and CIFS, endorsed a supportive scheme to develop Thai forensic staff through SCFS. Generally, foreign scholarships are usually offered for students to study at a university located in the country offering the scholarships. However, in this case, six scholarships valued at 300,000 THB each were granted to Thai Government Forensic staff to study the Master of Science degree in Forensic Science (International Program) at the Faculty of Science, started in May 2008. The scholar-
degree, with an average period of study of one and a-half years. The second semester is tough for the M.Sc. candidates, as they have to complete the thesis research, as well as the coursework. Within the second semester, all coursework are finished and students are guided through the thesis research process. The minimum duration of study is 12 months. In addition, adjunct staffs from forensic agencies and visiting professors/lecturers from well established and accredited forensic courses are invited to participate in the program to ensure that the content of the curriculum is international and serves the need of the country.
Education is a key component in the development of a profession. The educational process familiarizes practitioners with the knowledge and skills to enable them to develop and be recognized as ‘professionals’(3). The
ship covers tuition, research and thesis fees for one academic year, as well as English courses. Moreover, AFP continues to support another six scholarships for the academic year, 2009. This certainly shows that the program is well accepted at international level.
Research is an essential component of the postgraduate degree course. Since 2006, each M.Sc. candidate is required to publish his/her own research to satisfy the M.Sc. graduation requirements(23) of the Commission on Higher Education for assuring the quality of post-graduates. This is also compatible with the Forensic Science Education Program Accre-ditation Committee (FEPAC) accreditation standards guide that each graduate student is required to complete an independent research project, the results of which shall be presented orally in a public forum for evaluation. In addition, contributions should be made to the
M.Sc. (Forensic Science) class activity involves lectures, seminars, laboratory practices, mock case scenario, and visits to professional offices and laboratories.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
19
knowledge base of forensic science, including research directed at improving the practice of forensic science(8). Currently, the program has resulted in three international publications, six national publications and nine proceedings articles.
A number of research questions were developed in collaboration with the RTPOFS and CIFS forensic staffs. For example, three research theses were conducted to clarify questions concerning DNA profiling of the Tsunami victims in 2004. Bandhaya et al.(2) designed research on “Optimization and recovery of DNA from toothbrushes” in order to establish an optimized working protocol for obtaining reference DNA profiles for comparison. Toothbrush is a personal item from which DNA can be extracted from the buccal cells deposited in the bristles. In this work, the number of bristle bundles and period of usage were determined, as well as comparison of two DNA extraction methods(2). Phengon et al.(22) designed experiments on DNA analysis from degraded tissue by using porcine meat as a model. The experiments related the chance of obtaining DNA profiles from materials exposed to three conditions, i.e. left dried in an open environment, immersed in salt water and immersed in fresh water, in relation to time period. More recently, there are research from two theses involve forensic work on the terrorist problems in Thailand. Boonpanya et al.(4), reported forensic investigation of the explosion cases in the four south provinces of Thailand, namely Pattani, Narathiwat, Yala, and Songkla, which are targets for terrorist activities. The study revealed a partial signature
of bomber(s) using 14 variables in evidence classification. However, the officer’s safety was the major limitation to evidence collection in these continuing terrorist activities. Jaijarat et al.(11) determined appropriate collection methods for oil-contaminated fingerprints and fingerprints on oil-coated surfaces which were developed by different techniques. The outcome of Jaijarat’s research provides technical support to the crime scene and fingerprint examiners. Latent fingerprint is another area of colla-borative research. Studies were conducted on enhancement, development, and collection techniques and methods. Examples of these are “Detection of latent fingerprint on wet non-porous surface”(25), “Latent fingerprint recovery using scenescope-RUVIS UV imager”(16), “Study of appropriate concentration of ninhydrin for latent fingerprints on various papers”(10), “The study and comparison of chemical reagents for visualization of bloody fingerprint on different surfaces”(15), etc. In the area of questioned documents, research was conducted on the alteration of ink compositions with time(24). Concerning quality issues, a survey on the current status of forensic DNA laboratories in Bangkok was conducted in 2007. It is reported that Thai forensic DNA labora- tories are aware and alert to work towards the international accreditation scheme, though none are as yet accredited(21). This collaborative research provides mutual benefits to Mahidol University and the two forensic agencies; RTPOFS and CIFS.
Forensic Science research at the Faculty of Science has also been linked to the Office of Narcotics Control Board (ONCB), another stakeholder in the Thai forensic industry. Research theses contributing to the analysis of Cannabis sativa L. using chemical and biological methods to discriminate the ‘fiber-type’ from ‘drug-type’ Cannabis(26), analysis of Cannabis sativa L. traces (14), and a survey of Cannabis sativa L. ‘fiber-type’ cultivated in Thailand (using gas chromatography-mass spectrophotometry data) was recently pub- lished(13).
The Australian Government Forensic Science Scholarship presentation ceremony, May 2008.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
20
All of the above examples showed that the linkage between university and the end user has been established, providing a channel to enhance the professional competency of forensic practitioners and boost the quality of Thai forensic science.
Extracurricular activities are scheduled throughout the academic year. There are two main types of activities; one is to promote forensic science itself, and the other is to use forensic science as a media to encourage students to become interested in science. During the past few years, forensic science was promoted through “Special Lectures” and the internal seminar “Science Café” hosted by the Faculty of Science. These events opened up the opportunity for faculty members, including students, to interact with forensic science stake-holders and stimulated internal collaborations on multidisciplinary research. Annual events hosted by the faculty such as the “Open House”, the Faculty of Science’s open day, and “Science Avenue”, target to promote science at school level. Current M.Sc. students are the key players. Activities organized demonstrate the integration of various science disciplines and introduce the use of scientific methods, deductive and inductive logic in a simple crime
scene reconstruction scenario. We found that the most attractive activity was demonstration of evidence search and collection in a mock case, with two-way interaction between the audience and demonstrators. This resulted in a very stimulating event, making students interested in science and wanting to ‘learn’ and ‘do’ more science.
Forensic Science graduates are employed by the two main government agencies, universities, and the private sector. A small number of graduates pursue their further study at the Ph.D. level. In addition, employment opportunities in the private sectors for M.Sc. graduates in Forensic Science also depend on the first degree which they obtained. This suggests that there may be a need in particular forensic specialty areas in near future.
Future Directions
The university plays the major role in training, education, and research. University also provides specialist advice and services to forensic science users and providers(19). In a conference, Professor G. Henderson outlined the key findings from the “Forensic Science on Trial” report of the Parliament Select Committee on Science and Technology, UK,
A part of extra curricular activities of the Forensic Science Program.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
21
regarding role of university in providing a way for “better management of the technology transfer process to facilitate the exploitation of academic research with potential for applications in crime prevention and detection technologies”(6). This can also be applied in the Thai situation.
An aim of the educational program is to develop new forensic practitioners. However, the job market must also be considered. Fryer, K. mentioned in his talk that “employers look for ‘best value’ and ‘work-ready graduates’(6,19). Interaction between the educational institute and employer is an essential factor for developing the prospective competency of forensic practi-tioners to suite the employer’s need. Students benefit from integrating the experience of forensic practitioners into the academic area. However, precautions need to be taken in this manner of learning; scientific must be distinguished from unscientific matters. As Evett(7) expressed in the FSS spring meeting, in 1995, that “Science is not so much about common sense as about ‘challenging’ common sense. Believing something to be so just because of something called ‘experience’ is, in my view, essentially unscientific” and he also quoted Buam quotation “the trouble with ‘experience’ as a way of approximating to reliable knowledge is that all of us tend to reinterpret each individual experience in the light of a previously held conceptual framework”.
The quality of forensic science education will be reflected by the competency of graduates. The Forensic Education Programs Accreditation Commission (FEPAC) of the American Academy of Forensic Science (AAFS) accreditation standards for forensic science education program was recently revised in July, 2008(8). It is essential to ensure that each student meets the general curriculum requirements, i.e. to under-stand the underlying scientific knowledge that is essential to forensic science, be able to apply basic forensic science concepts and specialty knowledge to problem solve scenarios, be oriented in professional values, concepts and ethics; and successfully demonstrate the ability to integrate the acquired knowledge and skills
through a capstone experience such as compre-hensive examinations, thesis, and/or research projects. On the UK side, 28 undergraduate and four postgraduate courses delivered by 11 (out of 60-70 institutions offering courses entitled with “forensic”) universities, not all, preceded accre-ditation through the Forensic Science Society (FSS). Nevertheless, the quality of these unac-credited programs still reaches the standards of UK degree programs(5). Although foreign univer-sities are out of the scope of FEPAC and FSS accreditations, their accreditation guides can be useful resources for revision, self-evaluation, and adjustment of existing forensic science degree programs.
There is an opportunity for universities to co-ordinate formal training courses for forensic professionals and law enforcement agencies in Thailand. Practitioners/scientific investigators with “broad knowledge” covering a consi-derable part of the work are more competent to carry out assignments(3). In addition, because science and technology evolve fast, there are needs for continuing education to develop and maintain the competency of forensic practi-tioners. There are different forms of continuing education; training courses and workshops, seminars, meetings and conferences, and involvement in research(3,9,19). If the university is able to co-ordinate formal training courses, competency of participants may then be evaluated by the university system. In this way, the quality of the training would be assured, at least in some form, according to the university system. However, it is necessary to emphasize that training and workshops do not substitute for formal educational programs.
University is the key player to carry out pure and applied research, focusing on the application of science and technology to forensic science(19). There is a need to co-ordinate research and development through a proper program, as well as to increase the level of funding for forensic science research. Research is a requirement of the degree program and research experience is a recommendation included within any forensic graduate degree program(1,8). External funding to support

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
22
research is essential to any university degree program, so that future forensic practitioners can be adequately educated. The quality of Thai Forensic Science would then be sustainably maintained through research.
Conclusions
Forensic Science is a discipline that is becoming more mature in the Faculty Science, Mahidol University. From past to present, the program has developed and gained international acceptance. In the future, there is opportunity for university to play a critical role in providing academic support for developing and maintaining the ‘competency’ of forensic practitioners and experts through education, training and workshops, and research. These developments would then be the driving force for Thai forensic science to have a sustainable place in the international arena.
Acknowledgements
The author would like to acknowledge forensic science advisors, program committee and staff, Faculty of Science, Mahidol University.
References 1. Almirall J, Furton K. Trends in forensic science
education: expansion and increased accounta-bility. Anal Bioanal Chem 2003;376:1156-9.
2. Bandhaya A, Panvisavas, N. Optimization of DNA recovery from toothbrushes. Forensic Sci Int Genet 2008; 1 Suppl :9-10.
3. Barnett P. The role of forensic science pro-fessional organizations in the new millennium of accreditation, certification, registration, and standardization. Sci Justice 2000;40:138-42.
4. Boonpanya J, Chaikum N, Kontrong P, Teinsuwan M. Forensic investigation of explo-sive cases in the restive south. Roy Arm Med J 2008; 60:1-5.
5. Cassella J. Forensic science on trial-still! Response to “educating the next generation” [Sci Justice, 2008; 48: 59-60]. Sci Justice 2008;48: 197-9.
6. Centre for Forensic Investigation, School of Science & Technology, University of Teesside “The future of forensic and crime scene science”, June 2005. Forensic Sci Int 2006; 157 Suppl :2-6.
7. Evett I. Expert evidence and forensic miscon-ceptions of the nature of exact science. Sci Justice 1996;36:118-22.
8. Forensic Science Education Accreditation Com-mission (FEPAC). Accreditation Standards. [Online]. 2008 Jul 26. [cited 2008 Dec 14];[13 screens]. Available from: URL:http:// www.aafs. org/pdf/FEPAC%20AccreditationStandards072608.pdf
9. Gough T. Quality assurance in forensic science: the UK situation. Accredit qual Assur 1997;2: 216-23.
10. Homkrajang P. Study of appropriate concentra-tion of ninhydrin for latent fingerprints on various papers. Master of Science Thesis. Mahidol University; 2005.
11. Jaijarat A, Riengrojpitak S, Chalermsooksant, S, Suwansamrith W. Study of appropriate collection method for oil-contaminated fingerprints. Pro-ceedings of the 8th National Graduate Research Conference, Mahidol University, Salaya, 2007.
12. Jamieson, A. Why do we need good forensic science? Sci Justice 2002;42:45-9.
13. Kallawicha K, Bunyapraphatsara N, Panvisavas N, Wilairat P. Survey of Cannabis sativa L. ‘fiber type’ cultivated in Thailand (using gas chromato-graphy-mass spectrometry data). Srinakharinwirot Sci 2008;24:153-64.
14. Kitpipit T, Bunyapraphatsara N, Panvisavas N. Forensic detection of marijuana traces. Forensic Sci Int Genet 2008; 1 Suppl :600-2.
15. Lektrakul W. The study and comparison of chemi-cal reagents for visualization of bloody fingerprint on different surfaces. Master of Science Thesis. Mahidol University; 2006.
16. Lohitsiri P. Latent Fingerprint recovery using scenescope-RUVIS UV imager. Master of Science Thesis. Mahidol University; 2005.
17. Mahidol University is preparing to launch the Forensic Science Program (in Thai). [Online]. [cited 2008 Dec 14];[1 screen]. Available from: URL:http://www.bangkokbiznews.com/scitech/2003/0807/index.php?news=p1.html
18. Master of Science Program in Forensic Science Curriculum, Faculty of Science Mahidol Univer-sity; 2004.
19. Munnell, J. The future of forensic science and crime scene science part II. A UK perspective on forensic science education. Forensic Sci Int 2003; 157 Suppl :13-20.
20. Nic Daeid, N. Educating the next generation. Sci Justice 2008;48:59-60.
21. Panthuen S, Panvisavas N, Wilairat P. Current status of forensic DNA laboratories in Bangkok,

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
23
Thailand. Proceedings of the 8th National Graduate Research Conference. Mahidol Univer-sity, Salaya Campus, Bangkok; 2007.
22. Phengon P, Wongwiggran S, Panvisavas N. Analysis of DNA in degraded tissue. Forensic Sci Int Genet 2008; 1 Suppl :439-41.
23. Regulations of thesis publishing for graduation in a Master's Degree Program B.E.2549; A Faculty of Graduate Studies Announcement. [Online]. [2006?] [cited 2008 Dec 14]. Available from: URL: http://www.grad.mahidol.ac.th/grad/down load/pdf/publication_en.pdf
24. Sanplirm K. Study of the alteration of ink com- positions with time. Master of Science Thesis. Mahidol University; 2005.
25. Sukkasem S. Detection of latent fingerprint on wet nonporous surface. Master of Science Thesis. Mahidol University; 2008.
26. Wattansiri S. Forensic detection and discrimi-nation of Cannabis types. Master of Science Thesis. Mahidol University; 2006.


25
การพฒนาเซลลสบพนธและกระบวนการปฏสนธในกงกลาดา
รพพรรณ วานชวรยกจ1,2 และ ประเสรฐ โศภน1 1ภาควชากายวภาคศาสตร 2ศนยวจยเพอความเปนเลศเทคโนโลยชวภาพกง คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
การผสมพนธของกงกลาดาเกดภายหลงทกงตวเมยลอกคราบ และอยในภาวะเปลอกออน โดยกงตวผจะทาการสอดใสถงอสจเขาไปเกบไวในอวยวะสาหรบเกบอสจ (thelycum) ของกงตวเมยเปนระยะเวลาหนง หลงจากทกงตวเมยพฒนารงไขจนมความสมบรณเตมทแลว แมกงจะวางไขโดยปลอยไขและอสจออกมาผสมกนภายนอกตว หลงการปฏสนธ ไขทไดรบการผสมจะพฒนาเปนตวออนกงตอไป การศกษาการพฒนาของเซลลสบพนธในกงกลาดาพบวา กระบวนการสรางเซลลไขเกดในรงไขของแมกงวยเจรญพนธ โดยสามารถแบงการพฒนาเซลลไขไดเปน 5 ขนตอน ไดแก oogonia และ oocyte ขนท 1-4 ในเซลล ไขขนท 4 นนมการปรากฏของแทงวน (cortical rods) ซงวางตวอยดานนอกของเซลลไข สวนกระบวนการสรางเซลลอสจนนเกดขนในทอสรางอสจ (seminiferous tubules) ในอณฑะ หลงจากนนเซลลอสจจะถกสงไปเกบไวในทอนาอสจซงสวนปลายพองออกเปนถงอสจ (spermatophore) อยางไรกตามเซลลอสจทเกบจากกงตวผนนไมมความสามารถในการปฏสนธกบเซลลไข เซลลอสจจาเปนตองผานกระบวนการพฒนาความพรอมสาหรบการปฏสนธ (capacitation) กอน ซงมหลกฐานบงชวากระบวนการนเกดเมอถงอสจถกกกเกบไวใน thelycum ของกงตวเมย ในกระบวน การปฏสนธระหวางเซลลไขและเซลลอสจนนจะมการเปลยนแปลงโครงสรางของเซลลสบพนธทงสองชนดขน เซลลไขจะมการเปลยนแปลงโครงสรางเมอสมผสกบนาทะเล โดย cortical rods หลดออก กลายเปนชนวนทอยรอบไข นอกจากนยงมการสรางเยอหมเหนยวรอบไข (hatching envelope) ขนมาอกดวย สวนของเซลลอสจนนจาเปนจะตองถกกระตนใหเกด กระ บวนการ acrosome reaction โดยโมเลกลจากเซลลไขเปนตวกระตนใหเซลลอสจเกดกระบวนการน ความรความเขาใจถงชววทยาการสบพนธของกงนเปนองคประกอบพนฐานสาหรบการนาไปศกษาเพอพฒนาและประยกตใชในอตสาหกรรม
การเพาะเลยงกงตอไป
คาสาคญ: กงกลาดา; เซลลสบพนธ; การปฏสนธ
Abstract
In black tiger shrimp, Penaeus monodon, male mates with a soft-shell newly molted female. During mating, the male inserts sperm sacs (spermatophores) into female sperm storage organ thelycum. At the time of spawning, female containing fully mature ovary spawns its eggs and sperm together into the surrounding seawater where fertilization takes place. Differentiating of female germ cells in the ovary can be divided into five steps: oogonia and four steps of oocytes (Oc1-4), based on changes in their morphological characteristics. The unique characteristic of the mature oocyte (Oc4) is presence of cortical rods, egg jelly precursors, which are located at the periphery of the oocyte. Spermatogenesis in P. monodon occurs in seminiferous tubules within the testis which is connected to the vas deferens, where spermatophores containing mature sperm are formed and located at its distal end that opens at the base of the 5th walking legs. The male germ cells in the testis can be classified into spermatogonia, spermatocytes, spermatids, and spermatozoa. The structure of spermatozoa can be divided into three parts: 1) main body containing highly decondensed chromatin fibers surrounded by a thin cytoplasmic rim, 2) acrosomal region containing a sac-like structure and an anterior spike, and 3) subacrosomal region. Spermatozoa of close-typed thelycum, such as that of P. monodon undergo both morphological and biochemical changes and gain their fertilizing ability during storage in the female thelycum, a process referred to as capacitation. During spawning, sperm and egg undergo a series of structural changes to achieve the fertilization process. Spawned eggs are activated upon contact with seawater, which causes such changes as protrusion and solubilization of cortical rods from egg periphery into the seawater, and formation of the hatching envelope. In addition, the spermatozoa undergo acrosome reaction, which is a prerequisite step

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
26
for successful fertilization. This acrosome reaction can be induced by substances dissociated from the spawned egg. Morphological changes of spermatozoa during acrosome reaction include spike disappearance, swelling of acrosomal cap, exocytosis and spherical mass formation. Finally, fertilization is completed when there is fusion of the egg and spermatozoon nuclei.
Keywords: Penaeus monodon; spermatogenesis; oogenesis; fertilization; acrosome reaction
บทนา
กงกลาดาเปนกงทมขนาดใหญ โตเรว และมราคาด เปนทตองการของตลาดโลก อกทงยงเปนกงทมถนกาเนดในแถบเอเชยตะวนออกเฉยงใต จงเหมาะสมในการเพาะเลยงในประเทศไทย โดยทวไปการเพาะเลยงกงกลาดานนตองอาศยพอแมพนธจากธรรมชาตซงมอยอยาง
จากดและจานวนลดลงอยางมาก อกทงประสทธภาพการสบพนธของกงทเลยงแบบกกขงในโรงเพาะฟกยงมการ
ลดลงอกดวย นอกจากนเกษตรกรผ เลยงกงยงประสบปญหาการทลกกงโตชาและโรคระบาด ทาใหเกดความไมแนนอนในการเลยง อยางไรกตาม ไดมความพยายามในการแกปญหาเหลาน โดยผลตพอแมพนธจากบอเลยง และพฒนาสายพนธ กงกลาดาทสามารถสบพนธไดดและ
ปราศจากการตดเชอขน ซงในปจจบนไดมการผลตพอแมพนธจากกงเลยงไดเปนผลสาเรจ(21) โดยศนยวจยและพฒ- นาสายพนธกง (ศวพก.) ซงเปนหนวยงานของศนยพนธ-วศวกรรมและเทคโนโลยชวภาพแหงชาต (ศช.) และยงมการดาเนนงานเพอคดเลอกและพฒนาสายพนธกงกลาดา
อกดวย นอกจากการคดเลอกสายพนธทโตเรวและมความตานทานสงตอโรคระบาดแลว สายพนธทมประสทธภาพในการสบพนธสง คอ มจานวนครงของการวางไขและใหปรมาณลกกงจานวนมาก ยงเปนลกษณะสบสายพนธ (trait) ทสาคญในการคดเลอกอกดวย
อยางไรกตาม ยงมขอจากดอยมากในการบงชวาสายพนธใดมประสทธภาพในการสบพนธสง และใหเซลล สบพนธ (ไขและอสจ) ทมคณภาพด อกทงยงขาดขอมลในเชงลกถงกระบวนการสรางและพฒนาเซลลสบพนธ
ตลอดจนกระบวนการปฏสนธของกงกลาดา ซงความร
ความเขาใจในขอมลเหลานเปนองคประกอบพนฐานใน
การพฒนาสายพนธกงกลาดาและการพฒนาประสทธภาพ
การผสมพนธของกง ทงนหนวยงานทสนบสนนงานวจย หลายแหงเชน สวทช. และสกว.ไดเหนถงความสาคญของการศกษาในเรองน จงใหทนเพอดาเนนการศกษา งานวจยในเรองนบางสวนดาเนนการโดยอาจารย นกศกษา และนกวจยของมหาวทยาลยมหดลรวมกบมหาวทยาลยอนๆ
ทงในและตางประเทศ ในชวงระยะเวลา 5-6 ปทผานมาน จงมความรความเขาใจเกยวกบกระบวนการตางๆ ในระบบสบพนธของกงมากขน และไดมการเผยแพรผลงานระดบนานาชาตจานวนหนง บทความนจงรวบรวมความรความเขาใจตางๆ ทไดจากการศกษาคนควา และงานวจยทางดานชววทยาการสบพนธของกงกลาดาไวเพอเปน
ประโยชนสาหรบผทสนใจและใชในการศกษาตอไป การผสมพนธและการวางไข การผสมพนธตามธรรมชาตของกงกลาดานน เกดภายหลงทกงแมพนธลอกคราบ ซงจะมเปลอกหมลาตวทอยในภาวะเปลอกออน กงพอพนธจะผสมพนธกบกงแมพนธทอยในภาวะเปลอกออนน โดยทาการสอดถงอสจทเรยกวา spermatophore (รปท 1A) ซงประกอบดวยเซลลอสจทถกหมดวยสารเมอกใส เขาไปในกงตวเมยทลอกคราบใหมและยงอยในภาวะเปลอกออน ถงอสจนจะถกเกบไวระยะหนงในอวยวะสาหรบเกบอสจของตวเมยท
เรยกวา thelycum ซงเปนชองทมฝาปดอยบรเวณหนาอกทอยระหวางขาเดนคทหาของกงตวเมย (รปท 1B) เมอกงตวเมยพฒนารงไขจนถงระยะสมบรณเตมท แมกงจะวางไขโดยปลอยไขและอสจมาผสมกน(12,17) หลงจากการปฎสนธจะไดตวออนของกงในระยะทเรยกวา nauplius ตวออนนจะพฒนาไปเปนระยะ protozoea , mysis และ postlarva ตามลาดบ
ปจจบนไดมการใชวธการผสมเทยมกงกลาดาใน
การเพาะเลยง ซงมขอดคอมความแนนอนของการผสมระหวางพอแมพนธ และยงสามารถเลอกพอพนธตวทตองการมาผสมกบแมพนธได การผสมเทยมนทาไดโดยการสอดใส spermatophore จากกงพอพนธเขาไปเกบไวใน thelycum ของกงแมพนธทอยในภาวะเปลอกออน ซงกคอภายหลงจากแมพนธลอกคราบ การรดเอาถงอสจออกมา
จากกงพอพนธนนทาไดโดยการบบดวยปลายนวมอลงไป
บนบรเวณโคนขาเดนคท 5 ของกงพอพนธ หรอการกระตนดวยไฟฟาจากแบตเตอร (~ 25 มลลแอมแปร) เขาไปบรเวณดานขางลาตวของกงพอพนธ กงพอพนธจะปลอยถงอสจออกมา ในกงทม thelycum ชนดปดน เชน

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
27
กงกลาดา กงแมพนธจะสามารถวางไขไดหลายครงทงๆ ทมการสอดใสถงอสจเพยงครงเดยว(3) นนคอเซลลอสจสามารถมชวตอยใน thelycum ไดนานเปนสปดาหถงเปนเดอน แตเมอกงแมพนธลอกคราบ thelycum ทมอสจอยนนจะหลดออกมาพรอมคราบเกาทงหมดและมการสราง thelycum ใหมขนมาทดแทน ดงนนหลงจากกงแมพนธ
ลอกคราบ หากไมมการสอดใสถงอสจเขาไปใหมจะไมมการปฏสนธของไขกบอสจขน เนองจากไมเหลออสจภาย ใน thelycum อยเลย อยางไรกตาม กงแมพนธไมไดสญเสยความสามารถในการวางไข โดยจะสามารถวางไขไดอกเมอรงไขมการพฒนาตวเตมท ถงแมวาจะไมไดรบการสอดใสถงอสจกตาม แตไขทถกปลอยออกมาจะไมมการพฒนาเปนตวออน
ระบบสบพนธของกงเพศผและการพฒนาเซลลสบพนธ ระบบสบพนธของกงกลาดาตวผวางตวอยในสวน
หวของกง ประกอบดวย อณฑะ (testis) วางตวอยบนตบ (hepatopancreas) มลกษณะแบนคลายนวมอแผยนออกไปเปนพ สามารถแบงไดเปนพคหนา 1 ค และพทางดานขาง 6 ค ดานบนของอณฑะมหวใจวางทบอย ทางสวนทายของอณฑะตดตอกบทอนาอสจ (vas deferens)(2,16) (รปท 2A-B) เมอกงโตจนถงวยเจรญพนธได เซลลอสจจะถกพฒนา ขนในอณฑะและสงผานมายงทอเกบและนาสงเซลลอสจ
สทอนาอสจน โดยเซลลอสจจะถกเคลอบดวยสารเมอกใสตลอดความยาวของทอนาเซลลอสจ ทอนาอสจนม 1 ค ซงเปนทอยาววางตวออมไปทางดานหนาของตวกง ทสวนปลายสดของทอนพองออกเปนกระเปาะเปนถง spermato-phore ซงภายในบรรจเซลลอสจทถกลอมรอบดวยสาร
เมอก spermatophore มสองขางอยทบรเวณโคนขาเดนคท
5 ซงพรอมสาหรบการสอดใสเขาไปเกบไวใน thelycum ของกงแมพนธเมอมการผสมพนธ
กระบวนการพฒนาเซลลสบพนธของกงกลาดาตว
ผเกดในอณฑะซงภายในบรรจทอสรางอสจ (seminiferous tubules) อยมากมาย ภายในมเซลลสบพนธระยะตางๆ ทมความแตกตางกนทงขนาดและลกษณะของเซลล รวมถงลกษณะการเรยงตวของเสนใยโครมาทนภายในนวเคลยส
ของเซลลระยะตางๆ เรมตงแตเซลลตงตน (spermatogo-nia) เซลลระยะ spermatocyte, spermatid และเซลลอสจ (spermatozoa) ลกษณะของเซลลทปรากฏภายในทอสรางอสจตางๆ นนแสดงถงลกษณะการพฒนาของเซลลแบบพรอมกนทงทอ (synchronization) กลาวคอ ภายในทอสรางอสจชวงหนงๆ จะพบเซลลสบพนธเพศผระยะเดยว กนเกอบทงหมด ยกเวนกลมของเซลลตงตนทรวมตวกนอยกลมหนงตดกบผนงทอสรางอสจเสมอ เมอเซลลอสจถกสรางภายในทอสรางแลวจะถกสงออกไปยงทอนาอสจ
ยอยซงทอสวนปลายจะรวมกนตอกบทอนาอสจ ภายในทอนาอสจนจะพบวาเซลลอสจถกลอมรอบดวยสารเมอก
หนาทยอมตดสชมพของส hematoxylin-eosin ทอนาอสจนมผนงคอนขางหนาในสวนตน และบางลงในสวนปลายดานนอกถกลอมรอบดวยชนของกลามเนอ ทสวนปลายของทอพองออกเปนกระเปาะ spermatophore ซงภายใน
บรรจดวยอสจทถกลอมรอบดวยสารเมอกทยอมตดสชมพ
เชนกน
สาหรบการพฒนาขนสดทายของเซลลอสจนน พบวาในขนเซลล spermatid กอนทจะเปนเซลลอสจมการพฒนา acrosomal vesicle ซงเปนถงทม granule อยภายใน ถงนเปนโครงสรางเรมตนของ acrosome ถง acrosomal vesicle นวางตวอยทางดานหนาสดของ spermatid ระยะ
รปท 1 รปถายแสดงถงอสจ (spermatophore) ในกงกลาดา ตวผ (A) และอวยวะสาหรบเกบอสจ (thelycum) ของกงกลาดาตวเมย
รปท 2 รปถาย ( A ) และแบบจาลอง ( B ) แสดงระบบสบพนธของกงกลาดาตวผ Ant. Tes, anterior t e s t i s ; L a t . T e s , l a t e r a l t e s t i s ; H e p , hepatopancreas; Vas, vas deferens; Spp, s p e r m a t o p h o r e ; C D , c o l l e c t i n g d u c t

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
28
ทายๆ ยงสงเกตเหนโครงสรางทพฒนาเปนสวนของแทงปลายแหลม (spike) ยนออกไปอกดวย นอกจากนยงพบวาเสนใยโครมาทนภายในนวเคลยสมการเปลยนแปลง
โครงสรางอยางมาก กลาวคอมการคลและกระจายออกมากเปนลาดบจากใน spermatid ระยะตนๆ จนถงขนเซลลอสจ ซงมการคลออกของใยโครมาทนอยางเตมท
เมอศกษาดวยกลองจลทรรศนอเลกตรอนแบบสอง
ผาน เซลลอสจของกงกลาดานนมลกษณะเปนเซลลรปรางกลม มเสนผานศนยกลางประมาณ 7-8 ไมโครเมตร มสวนแทงทมปลายแหลมยนออกไปทางดานหนาหนงอน สวนตวเซลลอสจทมลกษณะกลมนน ประกอบดวย นว- เคลยสทมขนาดใหญเกอบเตมเซลล ซงบรรจดวยเสนใยโครมาทนทคลกระจายออกเตมนวเคลยส แสดงถงลกษณะการคลออกของใยโครมาทนในนวเคลยส สวนของไซโท- พลาซมทอยโดยรอบนวเคลยสปรากฏเปนเพยงแถบบางๆ เทานน ทางดานหนาตรงบรเวณฐานของ spike มโครง สรางคลายถงอย 2 ขาง ซงเชอกนวานาจะเปนตาแหนงของ acrosome (รปท 3A) และบรเวณใต acrosome (บรเวณรอยตอระหวาง acrosome กบนวเคลยส)เปนสวนของ sub-acrosome ซงพบวามใย cytoskeleton ทประกอบดวยแอกทนบรรจอย(16) ในเซลลอสจของกงกลาดาทพฒนาเตมทแลว ไมพบเยอหมนวเคลยส เนองจากอาจมการสลายของเยอหมนวเคลยสของเซลล เพอทาใหโครมาทนจากอสจมความพรอมทจะรวมกบโครมาทนของเซลลเดมทนทหลง
การปฏสนธกบไข
เซลลอสจทพบในทอสรางอสจและภายในทอนา
อสจรวมถงอสจทพบภายใน thelycum มลกษณะโครง สรางทคลายคลงกนเมอเปรยบเทยบโครงสรางหลกของ
เซลล อยางไรกตาม เมอพจารณาอยางละเอยดจะพบวา
เซลลอสจนนมการเปลยนแปลงของโครงสรางละเอยด
บางประการ กลาวคอ มการเพมขนของระดบการคล กระจายของเสนใยโครมาทนภายในนวเคลยส นอกจากน ยงพบ
ยงพบวาบรเวณ subacrosome ของเซลลอสจทพบใน
อณฑะ ทอนาอสจ และถงเกบสวนปลายยงพฒนาไมสม- บรณ คอ ยงคงสงเกตเหนเปนชองวางภายในอยบาง ขณะทเซลลอสจทพบใน thelycum แสดงลกษณะเตมโดยไมม
ชองวางอยในสวน subacrosome ระบบสบพนธของกงตวเมยและการพฒนาเซลลไข ระบบสบพนธของกงกลาดาตวเมยประกอบดวยรง
ไข ซงวางตวอยเหนอตบเชนเดยวกบอณฑะ ทอนาไข ซงมรเปดอยทบรเวณโคนขาเดนคท 3 และอวยวะสาหรบเกบอสจหรอท เรยกวา thelycum ซง เปนชองทอยบร เวณหนาอกทโคนขาเดนคท 5 (รปท 1B) thelycum ของกงก-ลาดานเปนชนดปด ประกอบดวยโครงสรางทเปนเปลอกแขงภายนอก ภายในมลกษณะเปนชองทมของเหลวบรรจอย ฝาปดของ thelycum ม 3 แผน ประกอบดวยฝาทอยตรงกลาง 1 ฝา (median plate) และฝาดานขาง 2 ฝา (lateral plates) )15 ( ทฝาปดดานขางของ thelycum นนมกลามเนออยภายใน และมโครงสรางลกษณะคลายตอมอยจานวนมาก ซงอาจมหนาทหลงสารทเปนสวนประกอบของของเหลว
ทเปนองคประกอบของ thelycal fluid ซงนาจะมบทบาทสาคญในการเตรยมความพรอมของเซลลอสจใหมความ สามารถในการผสมพนธ ในกงบางชนดนน thelycum ม
ลกษณะเปนแบบเปด กลาวคอไมมชองสาหรบเกบเซลลอสจทมฝาปด โดย thelycum เปนเพยงแองทบรเวณ
หนาอกของกงเพศเมย ซงมไวสาหรบใหถงเซลลอสจมาแปะตดไวเทานน กงทม thelycum แบบเปดนพบไดในกงขาวแปซฟก (Penaeus vannamei) และกงกามกราม (Ma-crobrachium rosenbergii) )4( เปนตน
การพฒนารงไขของกงกลาดาสามารถแบงไดเปน 5 ระยะตามลกษณะและการปรากฏของรงไขซงสงเกตได
จากภายนอก(8) คอ
รปท 3 แสดงเซลลอสจของกงกลาดาปกต (A) และ
เซลลอสจ เมอเกดกระบวนการ acrosome reaction (B)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
29
ระยะท 1: รงไขปรากฏเหนเปนเสนบางๆ ทางดาน หลงของสวนหวกง (ระยะ spent) ระยะท 2: รงไขปรากฏเหนเปนเสนทหนามากขน (ระยะ developing) ระยะท 3: รงไขปรากฏเหนเปนเสนทหนาขนอยางชดเจน มการขยายใหญของรงไขไปทางสวนลาตวทางดาน หลง และมสเขมขนเปนสเขยว (ระยะ nearly ripe) ระยะท 4: รงไขปรากฏเปนเสนหนาใหญ ขยายขนาดไปทางดานหลงจนถงหาง และสวนหวทางดานหนา ในระยะนรงไขมสเขมมากขนเปนสเขยวเขม กงจะวางไขในระยะท 4 น โดยปลอยไขออกมาทางทอนาไข ซงมรเปดอยทโคนขาเดนคท 3 ตามทกลาวมาแลว (ระยะ ripe) ระยะท 5: ระยะหลงการวางไข ในระยะน รงไขจะปรากฏลกษณะคลายกบระยะท 1 คอเปนเพยงเสนบางๆ หรอถามการวางไขเพยงบางสวน (partial spawning) อาจพบเหนสวนของรงไขทปรากฏเปนเสนหนาเปนชวงๆ (ระยะ post-spawning)
นอกจากนเมอศกษาการพฒนาของไขภายในรงไข
พบวา เซลลไขแบงออกเปน 5 ขน(9) ไดแก oogonia และเซลลไข oocyte ขน 1 ถง 4 (Oc1, Oc2, Oc3 และ Oc4) ในรงไขระยะท 2 นน เซลลไขสวนมากเปนเซลลไขขนท 1 และ 2 (Oc1, Oc2) เปนเซลลขนาดเลก เสนผานศนยกลางประมาณ 36-49 ไมโครเมตร ไซโทพลาซมตดสนาเงนเขม เนองจากเซลลประกอบดวยไรโบโซมจานวนมาก สวนในรงไขระยะท 3 เซลลไขจะมขนาดใหญขน โดยประกอบ ดวยเซลลไขขน Oc3 เปนสวนใหญ (เสนผานศนยกลางประมาณ 142 ไมโครเมตร) ไซโทพลาซมตดสแดง ซงอาจเกดจากกระบวนการสรางโปรตนจาเพาะภาย ในเซลล เชน vitellogenin รงไขในระยะท 4 ซงเปนระยะเจรญเตมทพรอมวางไข เซลลไขเกอบทงหมดเปนขน Oc4 ซงมขนาดใหญ ข นอ ก ( เ ส นผ านศ นย กล างประมาณ 2 2 1 . 5 ไมโครเมตร) และมโครงสรางใหมปรากฏขนทบรเวณรมเซลลไข มลกษณะเปนแทงวางตวอยโดยรอบเซลลไข เปนโครงสรางทอยนอกเยอหมเซลล โครงสรางทเปนแทงนเรยก cortical rods (รปท 4A-B) ซงจะหลดออกกลายเปนชนวนทหมรอบไขเมอไขถกปลอยออกมาภายนอกตวแม
กง สวน cortical rods นมไกลโคโปรตนเปนสวนประกอบ และอาจมสวนประกอบสาคญทกระตนใหเซลลอสจเกด
การพฒนาความพรอมกอนการปฏสนธกบเซลลไข ทเรยกวา acrosome reaction (AR) ซงเปนกระบวนการทจาเปนและสาคญอยางยงในกระบวนการปฏสนธ (1 1 ) ชนนอกสดของไขถกลอมรอบดวยเยอ vitellin บางๆ(14)
การปฏสนธ การปฏสนธของไขกบเซลลอสจเปนกระบวนการ
ทเกดขนอยางรวดเรวมาก และมความซบซอนอยางยง ไดมการศกษากระบวนการปฏสนธของไขและเซลลอสจ
อยางละเอยดในกง Sicyonia ingentis ซงเปนกงทม thely-cum ชนดปดเชนเดยวกบกงกลาดา(6) พบวาเซลลอสจของกง S. ingentis นนจะจบกบเซลลไขโดยใชสวนของ spike ไปแตะกบเปลอกหมดานนอกของไขทประกอบดวยชน vitellin ในระยะน เซลลอสจจะถกกระตนดวยปจจย จากเซลลไขใหเกดกระบวนการ AR ผลจากการนทาใหถง acrosome ซงอยทสวนหวของเซลลอสจแตกออก (exocy-tosis)(10,13) ภายในถงบรรจเอนไซมทจาเปนในการยอยสารตางๆ ทหมรอบไข(22) ทาใหเซลลอสจสามารถแทรกตวเขาไปปฏสนธกบเซลลไขทอยภายใตสารเคลอบไขได แต ถาหากวาเซลลอสจไมไดรบการกระตนใหเกด AR กอน เซลลอสจนนจะไมสามารถเขาไปปฏสนธกบเซลลไขได
เปนผลสาเรจ
กระบวนการเกด AR ในเซลลอสจของกงกลาดา
นนเกดขนอยางรวดเรวภายในเวลาเพยง 1 นาท(11) และ
สามารถเหนยวนาใหเกดไดดวยการกระตนโดยของเหลวท
ปลอยออกมาพรอมกบการวางไข (egg water)(1,11,13) แสดงใหเหนวาองคประกอบทหลงออกมาจากไขเปนตวกระตน
ใหเกดการพฒนากระบวนการ AR ของเซลลอสจ นอก จากน ยงมการศกษาเพมเตมพบวาโมเลกลของ cortical rods ทหลดออกมาขณะทไขสมผสกบนาทะเลเพอกลาย เปนชนวนรอบไขอาจเปนตวกระตนใหเซลลอสจเกด AR(11) การเปลยนแปลงของเซลลอสจในระหวางเกด AR นน ประกอบดวย การหดหายไปของ spike การบวมขน
และแตกออกของสวน acrosome และการปรากฏตวของ
กอนกลมเนอทบแสงขนมาทดแทนตาแหนงของ spike
รปท 4 (A) เซลลไขของกงกลาดายอมดวยส hematoxylin-eosin(9) (B) เซลลไขหลงจากแมกงวางไข แสดงการหลดออกของ cortical rods (CR) และ (C) เซลลไขหลงจากแมกงวางไข แสดง hatching envelope (HE)(14)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
30
(รปท 3B)(11,13) กอนกลมใหมทปรากฏตวขนนมโปรตน แอกทนเปนสวนประกอบ และอาจเกดมาจากการรวมและเรยงตวกนใหมของโปรตนแอกทนทอยภายใน sub-acrosome ของเซลลอสจเอง(16)
ไขมการเปลยนแปลงทางโครงสรางขนเชนกน เมอไข ถกปลอยออกมาขณะแมกงวางไข มนจะเกดการเปลยนแปลงอยางรวดเรวเมอสมผสกบนาทะเล กระบวน การดงกลาวเรยกวา egg activation การเปลยนแปลงทเกด ขน ไดแก มการหลดออกของแทงวนทอยรอบไขซงจะละลายกลายเปนชนวนทลอมรอบไข หลงจากนนจะเกดกระบวนการสรางแผนเยอเหนยวมาหมรอบไข (hatching หรอ fertilization envelope) (รปท 4C) หลงจากทมตวอสจตวแรกเขาไปผสมกบไข ซงชนนจะชวยปองกนไมใหอสจเขาไปผสมกบไขมากกวาหนงตว และยงชวยปกปองตวออนทเจรญอยภายใน อยางไรกตาม การพฒนาโครงสรางแผนเยอเหนยวรอบไขนเกดขนไดโดยไมตองอาศยอสจ
เปนตวกระตน ในแมกงทไมมการสอดใสถงอสจเขาไปเลย แตมการพฒนารงไขจนพรอมวางไขไดนน พบวาไขทถกปลอยออกมานน cortical rods มกระบวนการหลดออกและสราง hatching envelope ได แตจะไมมการพฒนาตอ ไปเปนตวออน กระบวนการดงกลาวนเกดขนอยางรวด เรว เมอสงเกตดความเปลยนแปลงของไขภายใตกลองจล- ทรรศนทระยะเวลาตางๆ พบวา cortical rods หลดออก
และละลายกลายเปนวนรอบไขภายในเวลาเพยง 1 นาทหลงจากทไขสมผสกบนาทะเล หลงจากนน hatching envelope เรมสรางขนในเวลา 5 นาท และเสรจสนภายใน 13-15 นาท(14) การทไขมการเปลยนแปลงทางโครงสรางอยางรวดเรวน เปนสาเหตหนงททาใหการพยายามทาการผสมเทยมในกงกลาดา โดยใชไขและอสจมาปฏสนธกนในหลอดทดลองยงไมประสบความสาเรจ สาหรบปจจยทเปนตวกระตนใหเกดการเปลยนแปลงโครงสรางของไขน ในการศกษาเบองตนพบวาอาจเกยวของกบความเขมขน
ขององคประกอบของแรธาตในนาทะเล โดยเฉพาะความเขมขนของแมกนเซยมอออน
จงเหนไดวาการพฒนาของไขในระหวางการปฏ- สนธนน ไมจาเปนตองอาศยเซลลอสจเปนตวกระตนเลย แตการเกด AR ของเซลลอสจนนจาเปนตองอาศยสารจากไขเปนตวกระตน ความสามารถของเซลลอสจในการแสดงการเกด AR น สามารถใชเปนตวบงชถงความพรอมของเซลลอสจในการปฏสนธได การศกษาในกงกลาดาและกง S. ingentis พบวาเซลลอสจทเกบมาจากกงพอพนธไมสามารถกระตนใหเกด AR ไดโดย egg water ในขณะท
เซลลอสจทเกบจาก thelycum ของกงแมพนธถกเหนยวนาใหเกด AR ไดมากถง 80-90%(5,17) นอกจากนยงพบวาเซลลอสจมการเปลยนแปลงโครงสรางละเอยดเมอถงอสจถก
สอดใสไวใน thelycum กลาวคอ เมอศกษาดวยกลองจลทรรศนอเลกตรอนพบวา สวนของ spike ของเซลลอสจทนามาจากพอพนธมลกษณะเปนเกลยว ในขณะทเซลลอสจทนามาจาก thelycum ของแมพนธจะไมพบลกษณะ
ดงกลาว นอกจากนพบวาภายในสวนของ subacrosome ของเซลลอสจมการพฒนาโครงสรางตางๆทสมบรณเมอ
เซลลอสจถกบรรจอยภายใน thelycum(5,20) ขอมลนแสดงใหเหนวาการพฒนาความพรอมของเซลลอสจเพอใหม
ความสามารถในการปฏสนธไดนอาจเกดขนภายใน thelycum นอกจากน ยงมขอมลบงชวา เซลลอสจมการเปลยนแปลงในระดบโมเลกลเมออยใน thelycum อกดวย กลาวคอ มการเปลยนแปลงของโปรตนทผวเซลลอสจ โดยมการหลดออกของโปรตนบางตวทผวของเซลลอสจท
ถกกกเกบไวภายใน thelycum และมการดดซบโปรตนจากนา thelycum บางตวไวทผวเซลลอสจเพมขน อกทงยงพบวาเซลลอสจมระดบของการเปลยนแปลงกระบวนการ tyrosine phosphorylation ของโปรตนในเซลลเพมมากขน ซงกระบวนการทกลาวถงนเปนดชนบงชทสาคญอกอน หนงของกระบวนการเตรยมความพรอมของเซลลอสจใน
การปฏสนธทเรยกวากระบวนการ capacitation(7,18)
กระบวนการ capacitation นมการศกษาอยางมาก
ในเซลลอสจของสตวเลยงลกดวยนานม โดยเปนกระบวน การทพฒนาเซลลอสจใหพรอมปฏสนธซงเกดขนภายใน
ทางเดนอวยวะสบพนธของสตวเพศเมยภายหลงจากการ
ผสมพนธเสรจสนแลว กระบวนการเกด capacitation ของเซลลอสจในสตวเลยงลกดวยนานมดงกลาว สามารถเหนยวนาใหเกดขนไดภายนอกในหลอดทดลอง (in vitro) โดยใช capacitating medium ทเหมาะสมและจาเพาะกบเซลลอสจในสตวแตละชนด ในสภาวะท เหมาะสม โดยทวไปสารทใชในการเหนยวนาใหเกด capacitation น ประกอบดวย bovine serum albumin, HCO3
-, KCl(19) เปนตน แตทวาปจจยทสงผลใหเกดกระบวนการ capacitation ในกงนนยงไมมการศกษาวาปจจยทเกยวของกบกระบวน การนคออะไร ซงนาจะเกดขนใน thelycum ของกงแมพนธ โดยเฉพาะในกงทม thelycum ชนดปด อยางไรกตาม การพฒนาใหเกด in vitro capacitation ในกงอาจทาได ทงนตองมการศกษาเพมเตมถงปจจยจาเพาะตางๆ ทจาเปนตอการพฒนาเซลลอสจใหมความพรอมในการปฏสนธ และพฒนา capacitating medium จาเพาะตอเซลลอสจของกงซงนาจะมประโยชนในอตสาหกรรมการเพาะเลยงกง

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
31
กระบวนการสรางและพฒนาเซลลสบพนธ ตลอด จนกระบวนการปฏสนธของกงกลาดานนมความซบซอน
เปนอยางมาก เซลลอสจตองมการพฒนาความพรอมโดยอาศยอทธพลของสารคดหลงจาก thelycum ซงกระบวน การนเกดเมอถงอสจถกกกเกบไวใน thelycum ระหวาง
การผสมพนธ เซลลไขกมการพฒนาความพรอมเชน เดยวกน โดยเฉพาะการพฒนาของ cortical rods ซงเมอ
ละลายในนาทะเลขณะทถกปลอยออกไปสามารถปลอย
สารทกระตนกระบวนการ AR ในเซลลอสจทจะนาไปส
การรวมตวระหวางเซลลสบพนธทงสองชนด ถงแมวาในปจจบนจะมความรความเขาใจในเรองนระดบหนงแลว แตยงคงมคาถามทตองหาคาตอบอกมากเพอความเขาใจและ
นาไปสการพฒนาวธการใหมๆ ทนาไปใชไดจรงในอตสาหกรรมการเลยงกง และการพฒนาคดเลอกสายพนธกงกลาดา เชน ทาอยางไรจงจะเพมประสทธภาพการสบ พนธของกงในโรงเพาะฟก และตวบงชใดทบงชคณภาพของทงไขและอสจของกงพอแมพนธ เปนตน ดงนนการวจยเกยวกบเรองนจงมโจทยวจยอยอกมากทรอใหนกวจย
เขามาศกษาหาคาตอบ
กตตกรรมประกาศ คณะผวจยขอขอบคณสานกงานกองทนสนบสนน
การวจย สานกงานพฒนาวทยาศาสตรและเทคโนโลยแหงชาต และ สานกงานคณะกรรมการการอดมศกษา ทไดจดสรรทนสนบสนนงานวจยทางดานชววทยาการสบพนธ
ของกงกลาดา
เอกสารอางอง 1. Afzelius BA, Murray A. The acrosomal reaction of
spermatozoa during fertilization or treatment with egg water. Exp Cell Res 1957;12:325-37.
2. Bauer RT, Min LJ. Spermatophores and plug substance of the marine shrimp Trachypenaeus similes (Crustacea: Dacapod: Penaeidae): formation in the male reproductive tract and disposition in the inseminated female. Biol Bull 1993;185:174-85.
3. Browdy CL. A review of the reproductive biology of Penaeus species: Perspectives on controlled shrimp maturation systems for high quality nauplii production. Proceedings of the Special Session on Shrimp Farming. Baton Rouge; 1992. p. 22-51.
4. Chow S, Dougherty MM, Dougherty WJ, Sandifer PA. Spermatophore formation in the white shrimps Penaeus setiferus and P. vannamei. J Crustacean Biol 1991;11:201-6.
5. Clark WH Jr, Griffin FJ. The morphology and physiology of the acrosome reaction in the sperm of
the decapod, Sicyonia ingentis. Dev Growth Differ 1988;30:451-62.
6. Clark WH Jr., Kleve MG, Yudin AI. An acrosome reaction in natantian sperm. J Exp Zool 1981;218: 279-91.
7. Galantino-Homer HL, Visconti PE, Kopf GS. Regulation of protein tyrosine phosphorylation during bovine sperm capacitation by a cyclic adenosine 3′5′-monophosphate-dependent pathway. Biol Reprod 1997;56:707-19.
8. Hall MR, Young N, and Kenway M. Manual for determination of egg fertility in Penaeus monodon. AIMS Research: Australia; 2000.
9. Kruevaisayawan H. Structure of developing oocytes, biosynthesis and role of cortical rods, and factors involved in the acrosome reaction induction in Penaeus monodon. Ph.D. Dissertation. Faculty of Graduate Studies, Bangkok, Thailand; 2006.
10. Kruevaisayawan H, Vanichviriyakit R, Weerachat-yanukul W, Iamsaard S, Withyachumnarnkul B, Basak A, Tanphaichitr N, Sobhon P. Induction of the acrosome reaction of black tiger shrimp, Penaeus monodon, requires sperm trypsin-like enzyme activity. Biol Reprod 2008;79:134-41.
11. Kruevaisayawan H, Vanichviriyakit R, Weera-chatyanukul W, Withyachumnarnkul W, Sobhon P. Biochemical characterization and physiological role of cortical rods in black tiger shrimp, Penaeus monodon. Aquaculture 2007;270:289-98.
12. Perez Farfante I. The rock shrimp genus Sicyonia (Crustacea: Decapoda: Penaeoidea) in the eastern Pacific. US Natl Mar Fish Serv Fish Bull 1985;83:1-78.
13. Pongtippatee P, Vanichviriyakit R, Chavadej J, Plodpai P, Pratoomchat B, Sobhon P, Withya-chumnarnkul B. Acrosome reaction in the sperm of the black tiger shrimp Penaeus monodon (Decapoda, Penaeidae). Aqua Res 2007; 38:1635-44.
14. Pongtippatee-Taweepreda P, Chavadej J, Plodpai P, Pratoomchart B, Sobhon P, Weerachatyanukul W et al. Egg activation in the black tiger shrimp Penaeus monodon. Aquaculture 2004;234:183-98.
15. Taweepreda P. Natural fertilization of the black tiger shrimp, Penaeus monodon. Ph.D. Dissertation. Faculty of Graduate Studies, Mahidol University, Bangkok, Thailand; 2004.
16. Vanichviriyakit R. Development of male germ cells, sperm modifications, and sperm capacitation in black tiger shrimp, Penaeus monodon. Ph.D. Dissertation. Faculty of Graduate Studies, Mahidol University, Bangkok, Thailand; 2007.
17. Vanichviriyakit R, Kruevaisayawan H, Weerachat-yanukul W, Tawipreeda P, Withyachumnarnkul B, Pratoomchat B et al. Molecular modification of Penaeus monodon sperm in female thelycum and its

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
32
consequent responses. Mol Reprod Dev 2004;69: 356-63.
18. Visconti PE, Kopf GS. Regulation of protein phosphorylation during sperm capacitation. Biol Reprod 1998;59:1-6.
19. Visconti PE, Westbrook VA, Chertihin O, Demarco I, Sleight S, Diekman AB. Novel signaling pathways involved in sperm acquisition of fertilizing capacity. J Reprod Immunol 2002;53:133-50.
20. Wikramanayake AH, Uhlinger KR, Griffin FJ, Clark WH, Jr. Sperm of the shrimp Sicyonia ingentis undergo a bi-phasic capacitation accompanied by morphological changes. Dev Growth Differ 1992; 34:347-55.
21. Withyachumnarnkul B, Plodphai P, Nash G, Fegan D. Growth rate and reproductive performance of F4 domesticated Penaeus monodon broodstock. In: the 3rd National Symposium of Marine Shrimp; 2001 November 8-9; Bangkok: Thailand; 2001. p. 33-40.
22. Yudin AI, Clark WH, Jr, Kleve MG. Gamete binding in Sicyonia ingentis: primary and secondary. Am Zool 1980;20:875.

33
การพฒนาวคซนตอโรคพยาธใบไมตบ
กลธดา ชยธระยานนท และ ประเสรฐ โศภน ภาควชากายวภาคศาสตร คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
โรคพยาธใบไมตบเกดจากการตดเชอพยาธใบไมตบ Fasciola hepatica หรอ F. gigantica ในสตวเลยงและสตวเศรษฐกจโดยเฉพาะวว ควาย แพะ แกะ และสามารถตดเชอในมนษยไดโดยบงเอญ การรกษาโดยการใชยามคาใชจาย
คอนขางสงและยงไมคอยมประสทธภาพเนองจากเชอมกเกดการดอยา นอกจากนยงทาใหเกดการปนเปอนสารเคมทตกคางในผลตภณฑสตวทนามาประกอบเปนอาหารและในสงแวดลอม ทางเลอกอกทางหนงในการปองกนการตดเชอคอการใชวคซน ซงนาจะควบคมโรคพยาธใบไมตบไดดโดยใหผลเปนระยะเวลายาวนานกวา ปจจบนนไดมการศกษาถงศกยภาพของวคซนแตละชนด โดยศกษาในสตวทดลองขนาดเลกและสตวใหญ วคซนทมการทดสอบในปจจบน คอโปรตนทสาคญตอการดารงชพของพยาธ ไดแก fatty acid binding protein, cathepsin L/B, glutathione-S-transferase, leucine aminopeptidase และ saposin-like protein ศกยภาพของวคซนเหลานขนอยกบความสามารถในการกระตนระบบภมคมกนของโฮสต การศกษาสวนใหญในการพฒนาวคซนไดกระทาในพยาธ F. hepatica มากกวาใน F. gigantica โดยใชโปรตนเชงเดยวและแบบรวม นอกจากนยงมการแสวงหาวคซนตวอนทมประสทธภาพยงขน เชน การใชแอนตเจนจากผวของพยาธโดยใชวธทาง proteomic และ microarray รวมทงศกษาการตอบสนองทางภมคมกนของโฮสตตอวคซนตวเลอกเหลานดวย
คาสาคญ: F. hepatica; F. gigantica; โรคพยาธใบไมตบ; การพฒนาวคซน; แอนตเจน
Abstract
Fasciolosis is caused by either Fasciola hepatica or F. gigantica in ruminants or accidentally in humans. Control by drug treatment is not totally effective due to the development of drug resistance by the parasites, cost of treatment and contamination of drug residues in food and environment. As an alternative and more sustainable approach, vaccination can confer long-term protection. Currently, several major vaccine candidates have been identified and tested in the laboratory as well as in large animal models. These proteins, all of which are essential for maintaining parasite metabolism and survival, including fatty acid binding protein, cathepsin L/B, glutathione-S-transferase, leucine aminopeptidase and saposin-like protein stimulate significant host immune responses, and thus confer some degree of protection against the parasites. Vaccinations have been studied more in F. hepatica than F. gigantica. Efficacy of the vaccine may be increased using combinations of these proteins in a cocktail vaccination. Furthermore, more effective vaccine candidates, viz, those derived from the tegument, should be explored through proteomic and microarray approaches as well as by studying host immune responses to these candidate vaccines.
Keywords: fasciolosis; F. hepatica; F. gigantica; vaccine development; antigens
บทนา
โรคพยาธใบไมตบ (fasciolosis) เกดจากพยาธ Fasciola hepatica ในประเทศเขตหนาว และ F. gigantica ในประเทศเขตอบอน พบในแถบทวปแอฟรกาและเอเชย ตะวนออกเฉยงใตรวมทงไทย(44) และเวยดนาม พยาธใบ- ไมตบเปนปรสต และเจรญเปนตวเตมวยในสตวเคยวเออง
เชน วว ควาย แพะ แกะ สวนมนษยอาจตดเชอปรสตนไดแตพบไมบอยนก การตดเชอเกดจากการรบประทานผกจาพวกพชนาหรอดมนาจากลาหวย หนองนา หรอ ทงนา ทมตวออนระยะเมตาเซอรคาเรยอยโดยไมไดตมใหสก
กอน มรายงานการตดเชอพยาธชนดนในมนษยตงแตป

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
34
ค.ศ. 1980 ซงพบวามมากกวา 40 ประเทศ ทงในทวปแอฟรกา อเมรกา เอเชย และยโรป โดยเฉพาะแถบอเมรกา ใต พบการระบาดของพยาธ F. hepatica เปนจานวนมาก คาดการณวามมนษยทตดเชอพยาธชนดนราว 2.4-17 ลานคนทวโลก(48) และประมาณ 180 ลานคนจดอยในกลมเสยงตอการตดเชอ(19,30) ดงนนโรคพยาธใบไมตบจงกอใหเกดความเสยหายตอเศรษฐกจ สขภาพของสตวเลยง และ
มนษยทวโลก คดเปนมลคาความสญเสยประมาณ 3.2 ลานเหรยญสหรฐตอป(29,49)
วงจรชวตของพยาธใบไมตบ (รปท 1) เรมขนเมอ สตวเคยวเอองหรอมนษยกนหญาหรอผกหรอดมนาทมตว
ออนระยะเมตาเซอรคาเรย ตวออนระยะนสามารถมชวตอยไดยาวนานถง 1 ปขนอยกบสภาพอากาศ และความชน เมอเขาสรางกายของโฮสตแลว พยาธตวออนจะถกยอย โดยนายอยจากกระเพาะอาหารและออกจากซสตบรเวณ ลาไสเลกสวนตน หลงจากนนพยาธตวออนระยะท 1 จะไชทะลผานผนงลาไสเลกสชองทองแลวเขาสเนอตบ ทาใหเนอตบของโฮสตทมพยาธอาศยอย ถกทาลายและม เลอดออกมาก จงทาใหเกดภาวะซดและหากมการกนพยาธ เขาไปจานวนมาก อาจเปนสาเหตหนงททาใหสตวถงตาย ไดโดยเฉพาะในสตวทอายยงนอยและมขนาดเลก หลงจาก นนเมอตวออนมอายประมาณ 8-12 สปดาห มนจะไชเขา ไปอยในทอนาดและถงนาด เพอเจรญไปเปนพยาธตวเตม วย พยาธ 1 ตวสามารถผลตไขไดวนละประมาณ 20,000-50,000 ฟอง โดยปลอยไขออกมากบนาดแลวผานเขาส ระบบทางเดนอาหารแลวออกมากบอจจาระ ไขจะลอยอย ในนาประมาณ 9-15 วนจงเจรญเปนไมราซเดยม วายไป ในนาและไชเขาสโฮสตตวกลางทหนง คอ หอย ในโฮสตน ไมราซเดยมเจรญเตบโตตอไปเปนสปอรโรซสต เรเดย และ เซอรคาเรย ตามลาดบ ในทสดเซอรคาเรยจะออกจาก หอย วายไปเกาะพชนาซงเปนโฮสตตวกลางทสอง แลว สลดหางทงเพอสรางผนงหมซสต (encyst) กลายเปนพยาธตวออนระยะเมตาเซอรคาเรย ซงเปนระยะทตดเชอใน
โฮสตจาเพาะ คอสตวเคยวเอองหรอมนษย(31)
วธการปองกนและรกษาโรคตดเชอพยาธใบไมตบ ในสตวเลยงทเปนสตวเศรษฐกจในภาคเกษตรกรรมไดม การใชยาฆาพยาธ(16) เชน closantal, clorsulon, rofoxanide, nitroxynil และ triclabendazole ซงสวนใหญใชรกษาพยาธ ใบไมเลอด (Schistosoma spp.) และนามาใชรกษาโรค พยาธใบไมตบในสตวเคยวเอองดวย แตการใชยาฆาพยาธ ในสตวอาจกอใหเกดสารเคมตกคางในสตวและสงแวด ลอม นอกจากนมรายงานวา ยาฆาพยาธบางชนดสามารถ
ใชไดผลดกบพยาธตวแกเทานน และใหผลนอยลงใน พยาธตวออนเชน ยา ABZ (methyl-[(5-propylthio)-1H-benzimidazol-2-yl] carbamate) ทใชควบคมพยาธทมอาย มากกวา 12 สปดาหขนไป อยางไรกตามวธการรกษาโดย การใชยาอาจไมไดผลเมอตองใชยาซาอกหลายครง เนอง จากพบการดอยาเกดขน การใชยา triclabendazole รกษา
โรคพยาธ F. hepatica ในหลายๆ ประเทศ เชน ออสเตร-เลย(40) สกอตแลนด(32) เวลล(55) เนเทอรแลนด(18,33) และสเปน (5) พบวาพยาธมการดอยาเพมขน ดงนนจงเรมมแนวคดในการพฒนาวคซนขนโดยใชโปรตนจากพยาธ
ชนดตางๆ ทมคณสมบตเปนแอนตเจนและเปนโปรตนทจาเปนตอการดารงชพของพยาธมาพฒนาเปนวคซน
ทดสอบในสตวทดลอง เชน fatty acid binding protein (FABP), cathepsin L/B (CatL/B), glutathione-S-transferase (GST), leucine aminopeptidase (LAP) และ saposin-like protein (SAP) ฯลฯ ขอมลพนฐานสวนใหญเกยวกบการศกษาวคซนมาจากงานวจยพยาธ F. hepatica สวนงานวจยในทานองเดยวกนของ F. gigantica ยงมนอย ทงๆ ทพยาธทงสองชนดกอใหเกดความเสยหายในระดบใกลเคยงกน
รปท 1 วงจรชวตของพยาธใบไมตบ F. hepatica และ F. gigantica
Fatty Acid-binding (FABP) Proteins
โปรตน FABP พบมากในเนอเยอ parenchyma ของพยาธ ทาหนาทขนสง long-chain fatty acids สายยาว acyl-CoA esters และ cholesterol เขาสเซลลพยาธ เนอง จากพยาธไมสามารถสรางโมเลกลดงกลาวได ดงนน จง
ตองนาโมเลกลเหลานจากเลอดของโฮสตเพอเขาไปใชใน
กระบวนการเมทาบอรซมของไขมนและการซอมสรางเยอ
หมผวทมการผลดเปลยนทรวดเรวมาก การใช FABP ผลตเปนวคซนจะมผลไปขดขวางการขนสงกรดไขมนจาก
โฮสตเขาสพยาธซงอาจจะทาใหพยาธตายได(51)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
35
ในป ค.ศ. 1977 Hillyer และคณะ(20) สกดโปรตน FABP ซงมนาหนกโมเลกลประมาณ 12 kDa (nFh12) จากพยาธใบไมตบ F. hepatica ตวเตมวย พบวา FABP สามารถทาปฏกรยาขาม (cross reaction) กบแอนตบอด ของพยาธใบไมเลอด Schistosoma mansoni ได FABP ทถกคนพบนเมอนาไปทดสอบการใชเปนวคซนในววโดย
ใชปรมาณโปรตน 500 μg ผสมกบ Freund’s complete adjuvant (FCA) สามารถลดจานวนพยาธไดถง 55%(23) และยงใชไดผลดในหน mice ทดลอง โดยสามารถลด จานวนพยาธลงถง 69-78%(22) สวนในกระตายสามารถ ปองกนการตดเชอไดถง 40%(37) ตอมาในป ค.ศ. 1992 คณะของ Rodriguez-Perez(47) ตองการเพมปรมาณโปรตน ใหมากขน จงใชการสงเคราะหตามวธทางพนธวศวกรรม เนองจาก FABP ทไดจากธรรมชาตโดยการสกดจากตว พยาธไมเพยงพอตอการนาไปใชเพอผลตเปนวคซน ดงนน จงไดทาการโคลนยนนโดยวธ immunoscreening จากการใชแอนตเซรมตอ nFh12 ทาปฏกรยากบ cDNA expression library ของพยาธตวเตมวย F. hepatica ซงทาใหคนพบ rFh15 ทมนาหนกโมเลกลประมาณ 14.8 kDa และมลาดบการเรยงตวของกรดอะมโนคลายกบ FABP ทพบจาก S. mansoni (Sm14) ประมาณ 44%(35) หลงจากทาการผลตโปรตนลกผสม FABP (rFh15) จงนามาทดสอบการเปน วคซนในกระตาย ผลปรากฏวา สามารถปองกนการตดเชอ พยาธใบไมในตบไดถง 40-76% โดยทงขนาดของพยาธ จานวนไขและรอยโรคในตบลดลง(6) ตอมาจงมการทดลอง ในสตวทมขนาดใหญ เชน แกะ โดยผสม rFh15 กบ FCA พบวาวคซนนไมไดผลในการลดจานวนพยาธ โดยพบแต เพยงการลดขนาดของพยาธและปรมาณไขของพยาธใน อจจาระเทานน(45) แตเมอไดมการพฒนา adjuvant เพอทาใหโปรตน FABP อยไดเปนระยะเวลานานขนในสตว ทดลอง เชน ADAD ซงประกอบดวยสาร saponin ทสกด จาก Quillaja saponaria เปน adjuvant และม hydroalco-holic extract จาก Polypodium leucotomos เปน immuno-modulator รวมกบ non-mineral oil (montanide) เปนสารท ใชละลายแอนตเจน พบวาวคซนนใชไดผลดเมอทดลอง กบแกะและหน mice อตราการมชวตรอดของหนเพมขน ไดเปน 40% สวนในแกะพบจานวนพยาธเพยง 24.5% การเตบโตของพยาธชาลงถง 35% พบจานวนไขในนาด 58.1% และในอจจาระ 40.3% เมอเปรยบเทยบกบกลม ควบคม(28) งานวจยนสอดคลองกบการนา rFh15 มาใช รวมกบ adjuvant ใหมนในหน mice(38) พบวาอตราการม ชวตรอดประมาณ 50-62.5% เมอเปรยบเทยบกบการ ทดลองในอดตทใช nFh12 และพบวาโปรตนลกผสม
rFh15 ใหผลดกวาโปรตนธรรมชาต nFh12 สวนในแกะ วคซนทใช rFh15 ใหผลดอยกวาการใช nFh12 ทงๆ ทใช adjuvant และวธการทดลองแบบเดยวกน โดยพบจานวน พยาธ 43% จานวนไขในอจจาระลดลงเหลอประมาณ 38%(26,27) นอกจากนในบางการทดลองไดนา FABP ทง nFh12 และ rFh15 ของ F. hepatica และ FABP ทไดจาก S. mansoni (Sm 14) มาทดสอบการเปนวคซนขามสาย พนธพบวา nFh12 และ rFh15 สามารถปองกนการตดเชอ ของพยาธ S. bovis ในหน mice ไดสงถง 96% และ 72% ตามลาดบ(1,2) สวน Sm 14 กสามารถใหผลดในการปองกน การตดเชอพยาธทง S. mansoni และ F. hepatica(4,54,58)
ในกรณของพยาธใบไมตบ F. gigantica มการ ทดสอบความเปนวคซนของ FABP โดยนกวจยเพยง 2 กลมเทานน คอ Estuningsih และคณะ(15) ไดทดสอบความ เปนวคซนของ FABP ในวว โดยใชทงโปรตนธรรมชาตและโปรตนลกผสม พบวา FABP ธรรมชาตทสกดจาก พยาธใบไมตบ F. gigantica สามารถลดจานวนพยาธในวว ไดเพยง 31% แตวคซนโปรตน FABP ลกผสมใชไมไดผล ตอมาในป ค.ศ. 2005 Nambi และคณะ(39) ไดทดสอบวคซนนในควาย (Bubalus bubalis) โดยใชโปรตน FABP ลกผสมทสงเคราะหจาก bacterial system ผสมกบ FCA พบวาจานวนพยาธทอยในควายลดลงประมาณ 35.8% สวนนาหนก ขนาดความกวางและความยาวของพยาธลด ลงเพยง 13.4%, 12.6% และ 9.8% ตามลาดบ เมอเปรยบ เทยบกบกลมควบคม
Cathepsin L-protease (CatL) Cysteine proteases เปนเอนไซมทพบในพยาธ
เกอบทกชนด เอนไซมชนดนถกหลงออกมาจากพยาธตวเตมวย เพอใชยอยเซลลเมดเลอดและเซลลตบของโฮสต
และท าล าย เมทร กซ ภ าย ใน เซลล ของโฮสต เ ช น fibronectin, laminin และ คอลลาเจนธรรมชาต หลงจาก นน เอนไซม dipeptidyl peptidase และ aminopeptidase จะยอยเปปไทดเสนสนๆ ตอเพอใหไดกรดอะมโนซงจะ ถกดดซมผานลาไสเขาตวพยาธ และเอนไซมนยงถกนามา ใชในการยอยเนอเยอของโฮสต เชน ยอยผนงลาไส เนอ ตบและผนงของถงนาด เพอเคลอนเขาสอวยวะเปาหมาย ตามแตละชวงอายของพยาธ และยงใชเอนไซมชนดน เสรมการปองกนตวจากระบบภมคมกนของโฮสต โดย การยอยอมมโนโกลบลนของโฮสต(9) นอกจากนนแลวยง มหลกฐานทแสดงวาเอนไซม CatL ยงมบทบาทในการ สรางไขอกดวย (9,56) ในปจจบนพบวาพยาธใบไม ตบ F. hepatica ม CatL 5 แบบ (isoforms) แตนกวจยสวน

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
36
ใหญมงศกษาศกยภาพการเปนวคซนเฉพาะ CatL1 และ CatL2
Wijffels และคณะ(60) ไดนา CatL ทไดจากการ สกดพยาธตวเตมวย F. hepatica มาทดสอบการเปนวคซน ในแกะ พบวาจานวนของพยาธไมลดลง มแตเพยงจานวน ไขเทานนทลดลง และการเจรญเตบโตของพยาธตวออน เปนตวเตมวยมอตราใกลเคยงกบกลมทไมไดใชวคซน หลงจากนนในป ค.ศ. 1999 Piacenza และคณะ(42) ไดทา การสกด CatL1 and CatL2 จากสารคดหลง (excretory-secretory antigen) ของพยาธแลวใชทดสอบการเปนวคซน ในแกะ พบวาเอนไซมทง 2 ชนดสามารถลดจานวนพยาธ ไดใกลเคยงกนประมาณ 33% แตเมอใหพรอมกนทง 2 ชนด ปรากฎวาไดผลสงถง 60% รวมทงจานวนไขกลดลง ถง 81% แตงานวจยนไมไดผลดในแกะพนธ Australian Merino ทไมสามารถปองกนการตดเชอได มเพยงจานวน ไขเทานนทลดลงประมาณ 69.4%(52) สวนในววไดผลดใน การปองกน คอ สามารถลดจานวนพยาธลงไดมากกวา 40%(8,10) ในป ค.ศ. 2005 มการคนพบ procatL (FhCL3) จากพยาธตวออน พบวาวคซน FhCL3 ทสงเคราะหจาก baculovirus system สามารถปองกนการตดเชอพยาธใน หน rat ไดประมาณ 52% แตวคซนนใหผลไมดเมอใชโปร- ตนชนดเดยวกนนทสงเคราะหจากยสต (Saccharomyces cerevisiae) อาจเปนเนองจากการเกด posttranslational modifications ทไมเหมอนกน โดยระบบทสงเคราะหจาก ยสตจะให glycosylation มากกวา(46) อยางไรกตาม มการ ทดสอบวคซนชนดน พบวาสามารถปองกนการตดเชอ พยาธใบไมตบ F. hepatica ในหน rat ไดถง 78-80%(24) ในวว 54.2% และ ในแกะ 56.5% ทงทโปรตนทสงเคราะหนไดจาก inclusion bodies โดยระบบการสงเคราะหใน แบคทเรยท CatL อยในรป mature protein ซงใหผลดกวา ในรป proprotein(59)
นอกจากนแลวมนกวจย(42) ไดทดสอบวคซนทม แอนตเจนหลายๆ ชนดรวมกน (cocktail vaccine) พบวา วคซนทมองคประกอบของ FhCL1, FhCL2 และ leucine amino peptidase (LAP) สามารถปองกนการตดเชอพยาธ ได ถง 79% นอกจากนนยงมการนาแอนต เจนท เปน haemoglobin (Hb) ทมนาหนกโมเลกล >200 kDa ซงพยาธ ใชโปรตนชนดนในการขนสงออกซเจนหรอเกบออกซเจน ไวในชวงทสมผสกบสภาวะทมออกซเจนรวมทงยงชวย สรางไขดวยมาผสมกบ CatL2 วคซนรวมดงกลาว สามารถลดจานวนพยาธไดประมาณ 72.4% มากกวา CatL1 ผสมกบ Hb (51.9%) รวมทงจานวนไขกลดลงถง 98% ทงนอาจ
เปนเพราะวาวคซนทาให Hb ไมสามารถขนสงออกซเจนไดและทาใหเกดการยบยงการสรางไขซงเปนกระบวน การทตองอาศยออกซ เจน (8) สวนพยาธใบไม ตบ F. gigantica มการทดลองในควายโดย CatL ทสกดจากสารคดหลงของพยาธพบวา จานวนพยาธไมลดลง(15)
Glutathione-S-transferase (GST) เอนไซม GST เกยวของกบขบวนการทาลายพษ จากสงแปลกปลอมทเขามาสรางกายและสารทกอใหเกด พษ โดยเฉพาะอนมลเสรทเกดในรางกาย GST ม 5 isoforms นาหนกโมเลกลประมาณ 23-26.5 kDa โดยแตก ตางกนทลาดบเบสบรเวณ N-terminal พบ GST ในเนอเยอ parenchyma ของผวพยาธและเยอบผวลาไส มการนา GST มาทดสอบเปนวคซน ในป ค.ศ. 1990 โดย Sexton และ คณะ(50) พบวา GST ทสกดออกมาจากพยาธผสมกบ FCA สามารถลดจานวนของพยาธในแกะทตดเชอ F. hepatica ได 57% และปองกนการตดเชอในววได 49-69%(34) โดย ใช Quil A/SM เปน adjuvant หลงจากนนในป ค.ศ. 1999 Spithill และคณะไดทาการทดลองอกครงหนง แตไดผล การทดลองทไมสอดคลองกบผลเดม(53) ตอมาในป ค.ศ. 2003 De Bont และคณะ(11) ไดนา S. bovis GST ลกผสม (rSb28GST) ผสมกบ aluminium hydroxide, Quil A, หรอ FCA แตการทดลองนไมประสบความสาเรจเนองจากไม สามารถปองกนการตดเชอ F. hepatica ในววได
การศกษาใน F. gigantica ไมสามารถแสดงไดวา GST มศกยภาพเปนวคซน(15,41) อาจเปนเพราะวาไมไดไช FCA เปน adjuvant หรออาจเกดจากความแตกตางระหวาง พยาธทง 2 ชนด แตจากงานวจยลาสดเมอป ค.ศ. 2008(43) พบวาเมอนา GST ลกผสม (rFgGST26) มาทดสอบความ เปนวคซนในหน mice พบวาใชไดผลเมอฉดเขาสตว ทดลองทง 3 ทาง คอ ใตกลามเนอ ใตผวหนง และชนหนง แท โดยสามารถปองกนการตดเชอไดถง 77-84%
Leucine aminopeptidase (LAP) เอนไซม LAP เปน exopeptidase ทยอยกรดอะม-
โนในเปปไทดจากดาน NH2 terminal เอนไซมชนดน สามารถวดปรมาณไดดวยการตดซบสเตรตทตาแหนง leucine-7-amino-4-methlycoumarin (NHMec) เอนไซม
กระจายอยในเซลลเยอบผวลาไสของพยาธใบไมตบ ไดม การนา LAP ทสกดจากพยาธ F. hepatica มาทดสอบการ เปนวคซน พบวา LAP สามารถปองกนการตดเชอไดถง 89% และเมอนามาใชรวมกบ CatL1 หรอ CatL2 สามารถ เพมการปองกนการตดเชอไดมากกวาการใช CatL1 หรอ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
37
CatL2 เพยงอยางเดยว(42) ในป ค.ศ. 2008 มผสงเคราะห LAP ลกผสม แลวนาไปทดลองในกระตาย พบวาสามารถ ลดการตดเชอ F. hepatica ไดถง 78%(3)
Saposin-like protein (SAP) Saposin-like protein เปนโปรตนในลาไสของ
พยาธใบไมตบ F. hepatica มนาหนกโมเลกลประมาณ 11.5 kDa มลกษณะคลายกบ NK-lysin(12) ทาหนาทสลาย เมดเลอดแดงและเมดเลอดขาวของโฮสต มการทดลองใช โปรตนลกผสม FhSAP2 ทดสอบการเปนวคซนใน กระตาย ซงใหผลการปองกนการตดเชอไดถง 81.2% และ จานวนไขทพบในนาดและอจจาระ รวมทงพยาธสภาพ ของการตดเชอในตบของกระตายกลดลงเชนกน(13,14)
สรป
การทดสอบศกยภาพการเปนวคซนของโปรตน
ชนดตาง ๆ ของพยาธใบไมในตบ ใช F. hepatica มากกวา F. gigantica ซงปรากฎวาวคซนบางตวสามารถลดจานวน พยาธ ยบยงการเจรญเตบโตของพยาธไมใหกลายเปนตว เตมวย ลดจานวนไขหรอลดพยาธสภาพของโรคทตบได(17) แตอยางไรกตาม ยงไมมวคซนชนดใดทสามารถนามาผลตเพอใชงานจรงได เนองจากมอปสรรคหลายๆ ประการ เชน การทดลองสวนใหญกระทาในสตวทดลองขนาดเลก เชน หน mice หรอ rat หรอกระตาย เพอทดสอบประ- สทธภาพความเปนวคซนกอน เนองจากตองการประหยดงบประมาณ และเมอไดผลการทดลองทดจงคอยมาทดสอบวคซนอกครงในสตวขนาดใหญ เชน วว ควาย แตการใชสตว เลกเปนตนแบบทดสอบวคซนมขอจากด เนองจากสตวแตละชนดมระบบภมคมกนตอการตดเชอ พยาธใบไมตบไมเหมอนกน เชน หน mice ตดเชอพยาธ ไดงายกวาหน rat และมโอกาสทหน mice จะตายหลงจาก การตดเชอพยาธดวยเมทาเซอรคาเรยจานวนเพยง 2 เมดภายใน 28 วน(31) การใช adjuvant ระยะเวลาและวธการบรหารวคซนทไมเหมอนกน อกทงวธทเหมาะสมทสด คอการใหวคซนทางปากโดยการปนกบอาหาร ถงแมวาไดมการพฒนา adjuvant ขนมาใหมแทนการใช FCA ซงไม
เปนทยอมรบในทางปฎบ ต แตขณะนย งไมประสบ ความสาเรจเนองจากทดสอบกบแอนตเจนเพยงชนดเดยว คอ FABP ซงพบวามประสทธภาพในการลดการตดเชอ พยาธเพยง 43% ในแกะ ซงใหผลทใกลเคยงกบการใช FCA และตองใหวธฉดเขาใตผวหนง(27)
ปจจบนไดมการนาเทคโนโลยทางชวภาพมาชวย
เพมการผลตโปรตนแอนตเจนทจะใชเปนวคซน โดยหวง วาการสงเคราะหโปรตนลกผสมจะทาใหไดโปรตน จานวนมาก แทนการใชการสกดโปรตนจากตวพยาธซง ยากกวา เนองจากตองผานกระบวนการในการทาให โปรตนบรสทธหลายขนตอน และโดยมากจะใหปรมาณ โปรตนทตา แตการทาโปรตนลกผสมนตองอาศยการผลตผานระบบแบคทเรยหรอยสต ซงในบางครงอาจไมประสบ ความสาเรจ โดยเฉพาะโปรตนทมองคประกอบเปน
คารโบไฮเดรตอยดวย และคณสมบตทสาคญทสดในการ เปนวคซนคอ โปรตนตองเปนสารทกระตนการสรางภม ตานทานไดดและควรเปนโปรตนทพบไดตงแตพยาธตว ออน เชน Cathepsin B (CatB)(25) ทหลงออกมาจากลาไส ของพยาธทระยะ newly excysted juvenile ใชในการสลาย เนอเยอโฮสตเพอพยาธตวออนเคลอนไปทตบได การใช CatB เปนวคซนอาจปองกนการตดเชอได เนองจากพยาธ ตวออนทเพงออกมาจากซสต มขนาดเลกและถกทาลายได งายโดยเซลลหรอแอนตบอดจากระบบภมคมกนของ โฮสต อกทงพยาธตวออนยงไมพฒนาระบบหลกหนจากระบบภมคมกนของโฮสตได การใหวคซนในชวงน จะ
ประสบความสาเรจมาก นอกจากนการใชวคซนทเปน โปรตนจากผวพยาธเชน โปรตน 28.5 kDa(7) ซงพบในปรมาณมากและกระจายอยในชนผวและอวยวะระบบสบ พนธเพศเมยเทานน จากการตรวจสอบโดยใชโมโนโคล- นลแอนตบอด (รปท 2) ซงโปรตนนกเปนอกแนวทาง หนงของการพฒนาวคซน เนองจากผวพยาธเปนแนวท สมผสกบเซลลของระบบภมคมกนของโฮสตอยตลอด เวลา การเสาะแสวงหาโปรตนอนๆ ทอาจจะนามาใชเปน วคซนทมประสทธภาพมากขนกควรจะทา แตปจจบน ยงขาดขอมลรหสพนธกรรมของโปรตนแตละชนดทอาจจะ
นามาพฒนาเปนวคซนได ขอมลพนฐานสวนใหญไดจากพยาธใบไมเลอด (Schistosoma spp.) จงตองมการศกษารหสพนธกรรมของยนแตละชนด จาก cDNA library ของ F. hepatica และ F. gigantica ตอไป เชน การศกษา expressed sequence tag ทดาเนนการโดยสถาบน Sanger (http://www.sanger.ac.uk/ Projects/Helminths/) เพอเปน รากฐานในการศกษาระดบ proteomics และ genomics ของพยาธชนดนตอไป
ในปจจบนแอนตเจนทนาจะเหมาะสมทสดคอ FABP, LAP, CatB และ SAP เนองจากสามารถลดจานวน พยาธหรอปองกนการเจรญเตบโตของพยาธ ลดปรมาณ ไข และลดพยาธสภาพของโรคทตบได ซงควรจะนามา ทดลองทงแบบเชงเดยวและการใหพรอมกนแบบเปน

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
38
cocktail vaccine ในสตวใหญ ๆ ทเปนสตวเศรษฐกจ ทสาคญของประเทศ
รปท 2 กลมควบคมทใชเซรมจากหนทไมไดตดเชอพยาธใบไมตบ (ภาพ A) เมอเปรยบเทยบกบพยาธใบไมตบตวเตมวย F. gigantica ยอมดวยโมโนโคลนลแอนตบอดตอโปรตนนาหนกโมเลกล 28.5 kDa(7) พบการกระจายของแอนตเจน ทผว (Te) เซลลสรางผว (Tc) และระบบอวยวะสบพนธ เพศเมยคอ ไข (Ov), Mehlis gland (Mg) แตไมพบใน อณฑะ (Ti)) (ภาพ B-G)
กตตกรรมประกาศ การวจยทชวยสรางองคความรในเรองนไดรบการ
สนบสนนเงนทนจากสานกงานกองทนสนบสนนการวจย และมหาวทยาลยมหดล
เอกสารอางอง 1. Abáné JL, Ramajo V, Arellano JL, Oleaga A,
Hillyer GV, Muro A. A fatty acid binding protein from Fasciola hepatica induced protection in C57/BL mice from challenge infection with Schistosoma bovis. Vet Parasitol 1999;15:107-21.
2. Abáné JL, Oleaga A, Ramajo V, Casanueva P, Arellano JL, Hillyer GV et al. Vaccination of mice against schistosoma bovis with a recombinant fatty acid binding protein from Fasciola hepatica. Vet Parasitol 2000;24:33-42.
3. Acosta D, Cancela M, Piacenza L, Roche L, Carmona C, Tort JF. Fasciola hepatica leucine aminopeptidase, a promising candidate for vaccina-tion against ruminant fasciolosis. Mol Biochem Parasitol 2008;158:52-64.
4. Almeida MS, Torloni H, Lee-Ho P, Vilar MM, Thaumaturgo N, Simpson AJ et al. Vaccination against Fasciola hepatica infection using a Schistosoma mansoni defined recombinant antigen, Sm14. Parasite Immunol 2003;25:135-7.
5. Alvarez-Sánchez MA, Mainar-Jaime RC, Pérez-García J, Rojo-Vázquez FA. Resistance of Fasciola hepatica to triclabendazole and albendazole in sheep in Spain. Vet Rec 2006;159: 424-5.
6. Casanueva R, Hillyer GV, Ramajo V, Oleaga A, Espinoza EY, Muro A. Immunoprophylaxis against Fasciola hepatica in rabbits using a recombinant Fh15 fatty acid-binding protein. J Parasitol 2001; 87:697-700.
7. Chaithirayanon K. Cloning, characterization and expression of genes encoding 28-29 kDa tegumental antigen in Fasciola gigantica. Ph.D Dissertation. Faculty of Graduate Studies, Mahidol University, Bangkok, Thailand; 2005.
8. Dalton JP, McGonigle S, Rolph TP, Andrews SJ. Induction of protective immunity in cattle against infection with Fasciola hepatica by vaccination with cathepsin L proteinases and with hemoglobin. Infect Immun 1996;64:5066-74.
9. Dalton JP, Brindley PJ, Knox DP, rady CP, Hotez PJ, Donnelly S et al. Helminth vaccines: from mining genomic information for vaccine targets to systems used for protein expression. Int J Parasitol 2003;33:621-40.
10. Dalton JP, Neill SO, Stack C, Collins P, Walshe A, Sekiya M et al. Fasciola hepatica cathepsin L-like proteases: biology, function, and potential in the development of first generation liver fluke vaccines. Int J Parasitol 2003;33:1173-81.
11. De Bont J, Claerebout E, Riveau G, Shacht AM, Smets K, Conder G et al. Failure of a recombinant Schistosoma bovis-derived glutathione S-transferase to protect cattle against experimental Fasciola hepatica infection. Vet Parasitol 2003;113:135-44.
12. Espino AM, Hillyer GV. Molecular cloning of a member of the Fasciola hepatica saposin-like protein family. J Parasitol 2003;89:545-52.
13. Espino AM, Hillyer GV. A novel Fasciola hepatica saposinlike recombinant protein with immuno-prophylactic potential. J Parasitol 2004; 90:876-9.
14. Espino AM, Osuna A, Gil R, Hillyer GV. Fasciola hepatica: humoral and cytokine responses to a member of the saposin-like protein family following delivery as a DNA vaccine in mice. Exp Parasitol 2005;110:374-83.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
39
15. Estuningsih SE, Smooker PM, Wiedosari E, Widjajanti S, Vaiano S, Partoutomo S et al. Evaluation of antigens of Fasciola gigantica as vaccines against tropical fasciolosis in cattle. Int J Parasitol 1997;27:1419-28.
16. Fairweather I, Boray JC. Mechanisms of fasciolicide action and drug resistance in Fasciola hepatica. In: Dalton JP, editor. Fascioliasis. CAB International; 1999. p.225-76.
17. Hillyer GV. Fasciola antigens as vaccines against fascioliasis and schistosomiasis. J Helminthol 2005; 79:241-7.
18. Gaasenbeek CP, Moll L, Cornelissen JB, Vellema P, Borgsteede FH. An experimental study on triclabendazole resistance of Fasciola hepatica in sheep. Vet Parasitol 2001;95:37-43.
19. Garcia HH, Moro PL, Schantz PM. Zoonotic helminth infections of humans: echinococcosis, cysticercosis and fascioliasis. Curr Opin Infect Dis 2007;20:489-94.
20. Hillyer GV, del Llano de Diaz A, Reyes CN. Schistosoma mansoni: acquired immunity in mice and hamsters using antigens of Fasciola hepatica. Exp Parasitol 1977:42;348-55.
21. Hillyer GV. Schistosoma mansoni: reduced worm burdens in mice immunized with isolated Fasciola hepatica antigens. Exp Parasitol 1979: 48;287-95.
22. Hillyer GV. Induction of immunity in mice to Fasciola hepatica with a Fasciola/Schistosoma cross-reactive defined immunity antigen. Am J Trop Med Hyg 1985;34:1127-31.
23. Hillyer GV, Haroun EM, Hernandez A, De Galanes MS. Acquired resistance to Fasciola hepatica in cattle using a purified adult worm antigen. Am J Trop Med Hyg 1987;37:363-9.
24. Kesik M, Jedlina-Panasiuk L, Kozak-Cieszczyk M, Pucienniczak A, Wedrychowicz H. Enteral vaccination of rats against Fasciola hepatica using recombinant cysteine proteinase (cathepsin L1). Vaccine 2007;25:3619-28.
25. Law RH, Smooker PM, Irving JA, Piedrafita D, Ponting R, Kennedy NJ et al. Cloning and expression of the major secreted cathepsin B-like protein from juvenile Fasciola hepatica and analysis of immunogenicity following liver fluke infection. Infect Immun 2003;71:6921-32.
26. López-Abán J, Casanueva P, Nogal J, Arias M, Morrondo P, Diez-Baños P et al. Progress in the development of Fasciola hepatica vaccine using recombinant fatty acid binding protein with the adjuvant adaptation system ADAD. Vet Parasitol 2007;145:287-96.
27. López-Abán J, Nogal-Ruiz JJ, Vicente B, Morrondo P, Diez-Baños P, Hillyer GV et al. The addition of a new immunomodulator with the adjuvant adaptation
ADAD system using fatty acid binding proteins increases the protection against Fasciola hepatica. Vet Parasitol 2008;153:176-81.
28. Martínez-Fernández AR, Nogal-Ruiz JJ, López-Abán J, Ramajo V, Oleaga A, Manga-González Y et al. Vaccination of mice and sheep with Fh12 FABP from Fasciola hepatica using the new adjuvant/ immunomodulator system ADAD. Vet Parasitol 2004;126:287-98.
29. Mas-Coma MS, Esteban JG, Bargues MD. Epidemiology of human fascioliasis: a review and proposed new classification. Bull World Health Organ 1999;77:340-6.
30. Mas-Coma S. Epidemiology of fascioliasis in human endemic areas. J Helminthol 2005;79:207-16.
31. McManus DP, Dalton JP. Vaccines against the zoonotic trematodes Schistosoma japonicum, Fasciola hepatica and Fasciola gigantica. Parasitology 2006;133:S43-61.
32. Mitchell GB, Maris L, Bonniwell MA. Triclabendazole-resistant liver fluke in Scottish sheep. Vet Rec 1998;143:399.
33. Moll L, Gaasenbeek CP, Vellema P, Borgsteede FH. Resistance of Fasciola hepatica against triclaben-dazole in cattle and sheep in The Netherlands. Vet Parasitol 2000;91:153-8.
34. Morrison CA, Colin T, Sexton JL, Waddington J, Wilson L, Thompson C et al. Protection of cattle against Fasciola hepatica infection by vaccination with glutathione S-transferase. Vaccine 1996;14: 1603-12.
35. Moser D, Tendler M, Griffiths G, Klinkert MQ. A 14kDa Schistosoma mansoni polypeptide is homologous to a gene family of fatty acid binding proteins. J Biol Chem 1991;266:8447-54.
36. Mulcahy G, Dalton JP. Cathepsin L proteinases as vaccines against infection with Fasciola hepatica (liver fluke) in ruminants. Res Vet Sci 2001;70:83-6.
37. Muro A, Ramajo V, López J, Simón F, Hillyer GV. Fasciola hepatica: vaccination of rabbits with native and recombinant antigens related to fatty acid binding proteins. Vet Parasitol 1997;69:219-29.
38. Muro A, Casanueva P, López-Abán J, Ramajo V, Martínez-Fernández AR, Hillyer GV. Identification of Fasciola hepatica recombinant 15-kDa fatty acid-binding protein T-cell epitopes that protect against experimental fascioliasis in rabbits and mice. J Parasitol 2007;93:817-23.
39. Nambi PA, Yadav SC, Raina OK, Sriveny D, Saini M. Vaccination of buffaloes with Fasciola gigantica recombinant fatty acid binding protein. Parasitol Res 2005;97:129-35.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
40
40. Overend DJ, Bowen FL. Resistance of Fasciola hepatica to triclabendazole. Aust Vet J 1995;72:275-6.
41. Paykari H, Dalimi A, Madani R. Immunization of sheep against Fasciola gigantica with glutathione S-transferase. Vet Parasitol 2002;105:153-9.
42. Piacenza L, Acosta D, Basmadjian I, Dalton JP, Carmona C. Vaccination with cathepsin L proteinases and with leucine aminopeptidase induces high levels of protection against fascioliasis in sheep. Infect Immun 1999;67:1954-61.
43. Preyavichyapugdee N, Sahaphong S, Riengrojpitak S, Grams R, Viyanant V, Sobhon P. Fasciola gigantica and Schistosoma mansoni: vaccine potential of recombinant glutathione S-transferase (rFgGST26) against infections in mice. Exp Parasitol 2008;119:229-37.
44. Pholpark M, Srikitjakara L. The control of parasitism in swamp buffalo and cattle in north-east Thailand. In: International Seminar on Animal Health and Production Services for Village Livestock, Khonkhen: Thailand; 1989. p.244-9.
45. Ramajo V, Oleaga A, Casanueva P, Hillyer GV, Muro A. Vaccination of sheep against Fasciola hepatica with homologous fatty acid binding proteins. Vet Parasitol 2001;97:35-46.
46. Reszka N, Cornelissen JB, Harmsen MM, Bieńkowska-Szewczyk K, de Bree J, Boersma WJ et al. Fasciola hepatica procathepsin L3 protein expressed by a baculovirus recombinant can partly protect rats against fasciolosis. Vaccine 2005;23: 2987-93.
47. Rodriguez-Perez J, Rodriguez-Medina JR, Garcia-Blanco MA, Hillyer GV. Fasciola hepatica: molecular cloning, nucleotide sequence and expression of a gene encoding a polypeptide homologous to a Schistosoma mansoni fatty acid binding protein. Exp Parasitol 1992; 74:400-7.
48. Roig GV. Hepatic fascioliasis in the Americas: A new challenge for therapeutic endoscopy. Gastrointest Endosc 2002;56:315-7.
49. Saba R, Korkmaz M, Inan D, Mamikoğlu L, Turhan O, Günseren F et al. Human fascioliasis. Clin Microbiol Infect 2004;10:385-7.
50. Sexton JL, Milner AR, Panaccio M, Waddington J, Wijffels G, Chandler D et al. Glutathione S-transferase: Novel vaccine against Fasciola hepatica infection in sheep. J Immunol 1990;145:3905-10.
51. Spithill TW, Dalton JP. Progress in development of liver fluke vaccines. Parasitol Today 1998;14:224-8.
52. Spithill TW, Smooker PM, Copeman DB. Fasciola gigantica: epidemiology, control, immunology, and molecular biology. In: Dalton JP, editor. Fascioliasis. CAB International: Wallingford, Oxon; 1999. p.465-526.
53. Spithill TW, Smooker PM, Sexton JL, Bozas E, Nirrusib CS, Creany J et al. Development of vaccines against Fasciola hepatica. In: Dalton JP, editor. Fascioliasis. CAB International: Wallingford, Oxon, 1999. p.377-410.
54. Tendler M, Brito CA, Vilar MM, Serra-Freire N, Diogo CM, Almeida MS et al. A Schistosoma mansoni fatty acid-binding protein, Sm14, is the potential basis of a dual-purpose anti-helminth vaccine. Proc Natl Acad Sci USA. 1996;93:269-73.
55. Thomas I, Coles GC, Duffus K. Triclabendazole-resistant Fasciola hepatica in southwest Wales. Vet Rec 2000;146:200.
56. Tort J, Brindley PJ, Knox D, Wolfe KH, Dalton JP. Proteinases and associated genes of parasitic helminths. Adv Parasitol. 1999;43:161-266.
57. Tran MH, Pearson MS, Bethony JM, Serra-Freire N, Diogo CM, Almeida MS et al. Tetraspanins on the surface of Schistosoma mansoni are protective antigens against schistosomiasis. Nat Med 2006;12: 835-40.
58. Vilar MM, Barrientos F, Almeida M, Thaumaturgo N, Simpson A, Garratt R et al. An experimental bivalent peptide vaccine against schistosomiasis and fascioliasis. Vaccine 2003;22:137-44.
59. Wedrychowicz H, Kesik M, Kaliniak M, Kozak-Cieszczyk M, Jedlina-Panasiuk L, Jaros S et al. Vaccine potential of inclusion bodies containing cysteine proteinase of Fasciola hepatica in calves and lambs experimentally challenged with metacercariae of the fluke. Vet Parasitol 2007;147: 77-88.
60. Wijffels GL, Salvatore L, Dosen M, Waddington J, Wilson L, Thompson C et al. Vaccination of sheep with purified cysteine proteinases of Fasciola hepatica decreases worm fecundity. Exp Parasitol 1994;78:132-48.

41
ความเครยดกระทบกระเทอนระบบภมคมกนของรางกายอยางไร
สพชา คมเกต ภาควชาชววทยา คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
การศกษาผลของความเครยดทมตอระบบภมคมกนของรางกาย ใชความรในสาขาวชาจตวทยา ประสาทวทยา และวทยาภมคมกน โดยอาศยทงการวจยในสตวทดลอง และในกลมคนทมความเครยด แสดงใหเหนการตอบสนองของรางกายตอความเครยดทเชอมโยงกนอยางนอย 3 ระบบ ไดแก ระบบประสาท ตอมไรทอ และภมคมกน ผานทางการผลตฮอรโมนทเกยวของกบความเครยด การเปลยนแปลงหนาทของเซลลและกลไกชนดตางๆ ในระบบภมคมกนซงจะทาใหผทมความเครยดและอยในภาวะกดดนมโอกาสในการตดเชอสงขน มความรนแรงของโรคมากขน หรอประสทธภาพในการสรางภมตานทานโรคหลงไดรบวคซนตาลง
คาสาคญ: ความเครยด; ฮอรโมน; ภมคมกน
Abstract
Stress and negative emotions have strong impact on immune functions. Knowledge gained from the field of psychoneuroimmunology, using both animal and human models, has shown that coping with stress induces responses involving three systems: central nervous, endocrine and immune. Experiencing stressful events generates stress hormones, which alter the functions of several immune cells, thereby increasing risk of infection, enhancing disease severity as well as reducing immune resistance following vaccination.
Keywords: stress; endocrine; hormone; immunity
บทนา ความเครยดสามารถเกดขนไดกบผคนทกระดบใน
สงคมโดยไมจากดวาอยในชวงวยใด เชน ในวยทางาน ความเครยดมาจากปญหาการเงน การมหนสน ปญหาการงาน ความผดหวงจากการเลอนขนหรอตาแหนง ปญหาครอบครว การหยารางหรอสญเสยสมาชกในครอบครวอยางกระทนหน ปญหาสขภาพทเกดจากโรคเรอรงทงของตนเองหรอการทตองดแลผเจบปวยในครอบครว สวนในเดกวยเรยนและวยรนอาจมความเครยดจากการแขงขนใน
เรองการเรยน การสอบ และความสมพนธในหมเพอน สถานการณ ‘เครยด’ นนจะกระตนความคดและอารมณทางลบ เกดจากการทผรบรประเมนสภาวะแวดลอมท
เผชญอยวา กาลงอยในระดบเกนทตนจะสามารถควบคมหรอแกไขได หรอหากตองการผานพนสถานการณนนกตองใชความพยายามหรออดทนในขนทสงกวาปกต (7) การบรณาการความรในเชงจตวทยา ประสาทวทยา และวทยาภมคมกน (psychoneuroimmunology) โดยอาศยผล การวจยทงในคนและสตวแสดงใหเหนวา ภาวะความ
เครยดสงและบอยๆ หรอมความกดดนทางอารมณยาวนานสงผลกระทบตอระบบภมคมกนของรางกาย ทาใหมโอกาสเปนโรคทเกดจากเชอไวรสและแบคทเรยสงขน มแนวโนมทโรคจะรนแรงเพมขน รวมทงการสรางภมตานทานตอเชอโรคหลงไดรบวคซนดอยประสทธภาพลง
ดวย(15) รปแบบความเครยดและการตรวจวดความเครยด
ลกษณะความเครยดท เกดขนแบงไดโดยอาศย
ระยะ เวลาสน-ยาว และความตอเนองของสงกระตน(32) เกดเปนความเครยด 2 ระดบ ดงน
1. แบบเฉยบพลน เปนความเครยดทเกดขนทนทและคงอยในชวงระยะเวลาสนๆ เมอสงกระตนความ เครยดหมดไปรางกายจะเขาสภาวะปกต เชน ความเครยดทเกดในชวงการสอบ ในภาวะคบขน อนตราย ในสถานทมด แคบ สง หรออณหภมรอนหรอเยนเกนไป เปนตน
2. แบบเรอรง เปนความเครยดทเกดขนยดเยอยาว นาน เนองจากสาเหตแหงความเครยดคงอยอยางตอเนอง

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
42
ตวอยางเชน การเจบปวยจากโรคเรอรง หรอการดแลผปวยเรอรง ตกงาน มหนสน หรอปญหาความสมพนธระหวางบคคล เปนตน
โดยสวนใหญ การตรวจวดระดบความเครยดสามารถกระทาไดโดย 1) วธทางจตวทยา ไดแก การใชแบบประเมนความเครยด โดยผตอบตองใหขอมลความถของอาการทเกยวของกบความเครยด เชน นอนไมหลบ วตกกงวล ซมเศรา ฟงซ าน ขาดสมาธ หรอปวดตงกลามเนอทเกดขนในชวงระยะเวลาทกาหนด (สปดาห-เดอน) รวมทงใหนาหนกความรนแรงของอาการไดวามาก-นอยเปนคะแนน แลวนามาว เคราะหหาระดบความเครยดตามวธมาตรฐานไดแก global assessment of recent stress scale(27) และ the stress response inventory scale(23,24) เปนตน และ 2) วธทางชววทยา ไดแก การตรวจวดอตราการเตนของหวใจ การวดความดนโลหต อตราการหายใจ และการเปลยนแปลงระดบสารเคมภายในรางกาย เชน ฮอรโมนและไซโทไคนท เกยวของกบความเครยดในเลอดหรอสารคดหลงตางๆ(11)
กลไกการตอบสนองตอความเครยด ความเครยดสงผลกระทบตอการทางานของ
รางกายของเราอยางนอย 3 ระบบ ไดแก ระบบประสาทสวนกลาง ตอมไรทอ และภมคมกน(31) โดยสวนใหญการรบรถงความเครยดจะกระตนการทางานทเปนโครงขาย
เชอมโยงของระบบสมองสวนไฮโพทาลามส-ตอมใตสมอง -ต อมหมวกไต (hypothalamic-pituitary-adrenal axis; HPA) และระบบประสาทอตโนมตซมพาเทตก-ตอมหมวกไตสวนใน (sympathetic-adrenal medullary axis; SAM)(6,11,37)
สมองสวนไฮโพทาลามส-ตอมใตสมอง-ตอมหมวกไต
ไฮโพทาลามสเปนสมองสวนหนาประกอบดวย
กลมเซลลประสาททางานแบบนอกเหนออานาจจตใจ ทาหนาทควบคมการแสดงออกทางอารมณ และความตองการพนฐาน เชนความหว กระหายนา และความตองการทางเพศ บรเวณสวนหนาของไฮโพทาลามสมสวนเชอมตอกบตอมใตสมอง เมอไฮโพทาลามสรบสญญาณความเครยดจะหลงสาร corticotrophin releasing factor สงตอมใตสมองสวนหนาใหผลตฮอรโมนอะดรโนคอรตโค-โทรฟน (adrenocorticotrophic hormone; ACTH) ไป
ควบคมเปลอกตอมหมวกไต (adrenal cortex) ใหหลง
ฮอรโมนกลโคคอรตคอยด ซงมบทบาทตอการทางานของเซลลหลายชนดในระบบภมคมกน (15)
ระบบประสาทอตโนมตซมพาเทตก-ตอมหมวกไตสวนใน
ระบบประสาทอตโนม ต ประกอบดวย เสน ประสาทไขสนหลงเหนอกระดกกระเบนเหนบขนไป ทางานในภาวะฉกเฉนขณะเกดอนตรายหรอตกใจ เรงการทางานของหวใจ ซงทาใหเราตองตดสนใจทนทวาจะสหรอถอย (‘fight or flight’) ระบบประสาทอตโนมตซมพา-เทตกกระตนตอมหมวกไตสวนในใหหลงฮอรโมน
อะดรนาลนหรอเอพเนฟรน และ นอรอะดรนาลน หรอ นอรเอพเนฟรน(2) โดยฮอรโมนทผลตจากทง 2 ระบบนจะหลงออกมาเมอรางกายอยในสถานการณตงเครยด จงอาจกลาวไดวาเปนฮอรโมนทเกยวของกบความเครยด(2, 15)
การทางานของระบบภมคมกน รางกายของเรามกลไกการตานทานสงแปลก
ปลอมและเชอจลนทรยทอาจกอใหเกดโรคอย 2 ระบบ ไดแก 1) innate immunity เปนระบบทพรอมทางานไดทนท ปองกนตวเราไดตงแตเกด สงมชวตแทบทกชนด
อาศยกลไกทมอยในระบบนในการดารงชวต และ 2) acquired/adaptive immunity พบเฉพาะในสตวทมกระดกสนหลง เปนระบบทพฒนาขนใหมความจาเพาะเจาะจงสง มประสทธภาพ สามารถจดจาเชอโรคไดอยางแมนยา แตอาศยระยะเวลาในการกระตนการทางานนานหลายวน โดยเซลลททาหนาทหลกในระบบน ไดแก เซลลลมโฟ-ไซตชนดท (T-lymphocyte) และชนดบ (B-lymphocyte) ผานทางโปรตนททาหนาทจบแอนตเจนทผวของลมโฟ-ไซตชนดท (T-cell receptor) และในรปของแอนตบอดทถกหลงออกมาจากเซลลพลาสมาซงเปนเซลลทเปลยน แปลงมาจากลมโฟไซตชนดบ
Innate Immunity
สงแปลกปลอมและเชอจลนทรยสามารถผานเขาส
รางกายของเราไดหลายทาง ไดแก ผวหนง ทางเดนหายใจ ทางเดนอาหาร ระบบสบพนธ ซงตามธรรมชาต บรเวณเหลานมกลไกทปองกน ยบย ง กาจดและทาลายส งแปลกปลอมและเชอจลนทรยในเบองตน โดยอาศยเซลลเยอบผวทมขนโบกพด (cilia) และขบเมอก สภาพเปนกรดในกระเพาะอาหาร ไขมนจากตอมไขมนทผวหนง และเหงอจากตอมเหงอทม pH 3-5 ชวยลดการเจรญของจลน- ทรยไดหลายชนด และรางกายยงสรางโปรตนททาหนาท

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
43
เฉพาะในการทาลายเชอจลนทรยทผานเขาสรางกายหรอ
อวยวะตางๆ เรยก antimicrobial peptide ซงมหลายชนด
อยางไรกตาม หากสงแปลกปลอมหรอเชอสามารถรกรานเขาสรางกายตอไปได ยงมเซลลในระบบภมคมกน ไดแก กลมเซลลกลนกน (phagocytes) ทาหนาทจบกน
และทาลายสงแปลกปลอมเหลานโดยกระบวนการกลนกน นาเชอนนเขาเซลลสภายในแวควโอล กอนทหลอมรวมกบไลโซโซมซงบรรจเอนไซมและสารพวก oxidative free radical เพอยอยหรอทาลายจลนทรย เซลลในกลมนทกระจายอยทวไปในเนอเยอตางๆ ไดแก เซลลแมโครเฟจ และเซลลเดนดรทก นอกจากน เซลลกลมกลนกนชนดนวโทรฟลทมอย เปนจานวนมากภายในหลอดเลอดยง
สามารถแทรกตวผานเซลลเยอบผนงหลอดเลอดออกมายง
เนอเยอโดยรอบไดโดยอาศยกลไกการอกเสบ(19)
เมอเชอจลนทรยสามารถหลดผานเขาไปในเนอเยอ
ของรางกาย เซลลแมโครเฟจและเดนดรทก ซงตามปกตมอยแลวจานวนหนงจะเรมกระบวนการกลนกน พรอมกนนนเซลลทเปนสวนประกอบหลกของเนอเยอบรเวณนน เชน keratinocyte บรเวณผวหนง และ hepatocyte ทตบ
สามารถตรวจจบสงแปลกปลอมไดเชนกน เซลลเหลานจะรวมกนสงสญญาณในรปของไซโทไคนออกไปเพอ
กระตนเซลลทมทรบจาเพาะกบไซโทไคนชนดนนๆ ใหทางาน ในกรณของการอกเสบ ไซโทไคนทถกผลตออกมาจะเปนชนด pro-inflammatory ซงมหลายชนด เชน inter-leukin (IL)-1, IL-6, tumor-necrosis factor ทาใหมเลอดไหลมาเลยงบรเวณนนในปรมาณมากขน จงเพมปรมาณใหเซลลกลนกนทงนวโทรฟลและโมโนไซตออกจาก
เลอดแทรกเขาสเนอเยอทมสงแปลกปลอมอยมากขน โมโนไซตในเลอดทแทรกเขาไปในเนอเยอเปลยนเปนเซลล
แมโครเฟจ สภาวะนจงมทงเซลล ของเหลวและโปรตนตางๆ มาสะสมในเนอเยอพรอมกน ทาใหเกดอาการรวมกบการอกเสบทพบไดบอย คอ ปวด บวม แดง รอน (19)
เซลลกลนกนทงชนดแมโครเฟจและเดนดรทก นอกจากทาหนาท กลนกน เพ อยอยหรอท าลายเช อ จลนทรยแลว ยงสามารถเดนทางจากบรเวณเนอเยอทมการตดเชอไปยงตอมนาเหลองหรอมาม ซงเปนอวยวะในระบบนาเหลองทมกลมของเซลลลมโฟไซตชนดทและบ อยเปนจานวนมาก หากเซลลลมโฟไซตเหลานไดรบการกระตน จะเปนการเรมการตอบสนองแบบ acquired immunity
Acquired/Adaptive Immunity ชนดตอบสนองแบบอาศยเซลล (Cell-mediated Immune Response)
เซลลลมโฟไซตชนดท สวนใหญกระจายอยในบรเวณเปลอกนอกของตอมนาเหลอง บนผวเซลลมทจบแอนตเจน จงสามารถจบแอนตเจนทเปนโปรตนในรป เปปไทดทจบมากบโมเลกล histocompatibility complex (MHC) บนผวเซลลทตดเชอไดแบบจาเพาะ โดยเซลลลม-โฟไซตแตละตวจะมทจบแตกตางกนไป ทาใหมความ จาเพาะตอแอนตเจนตางกน ลมโฟไซตชนดนยงสามารถแบงออกเปน 2 กลม คอ ลมโฟไซตชนดททแสดงโมเลกล CD8 บนเยอหมเซลล (CD8+ T lymphocyte) และ ลมโฟ-ไซตชนดททแสดงโมเลกล CD4 (CD4+ T-lymphocyte)
CD8+ T-lymphocyte มหนาทหลกในการทาลาย
เซลลทตดเชอไวรส โดยการหลงสารทสะสมในแกรนล ผานเขาไปยงเซลลทตดเชอ กระตนใหเซลลทตดเชอไวรสนนตายแบบอะพอพโทสสโดยไมทาลายเซลลอนๆ ทไมตดเชอขางเคยงเลย ดงนนเซลลชนดนจงมชอเรยกอกชอหนงวา cytotoxic หรอ cytolytic T-lymphocyte (CTL) เซลลทกชนดทมนวเคลยสเมอตดเชอไวรสสามารถนาสาย
เปปไทดของไวรสจบไวกบโมเลกล MHC class I แลวแสดงออกมาทผวเซลล ซงโมเลกล MHC class I รวม
กบเปปไทดนจะกระตน CD8+ T-lymphocyte ใหทางาน
CD4+ T-lymphocyte ทาหนาทหลกในการกระตนลมโฟไซตชนดบใหเปลยนเปนเซลลพลาสมาแลวหลง
แอนตบอดออกมา ดงนน จงเรยกลมโฟไซตชนดทกลมนวา helper T-lymphocyte (Th) CD4+ T-lymphocyte จะมทรบแอนตเจนทจาเพาะกบเปปไทดทจบมากบ MHC class II ทผวของเซลลพเศษอกกลมหนง ทเรยกวา antigen presenting cell (APC)
Antigen presenting cell มอย 3 ชนดเทานน ไดแก เซลลแมโครเฟจ เดนดรทก และ ลมโฟไซตชนดบ บทบาทของ APC นอกจากสงเปปไทดรวมกบ MHC class II ไปยงทจบจาเพาะบน Th แลวยงสามารถกากบให Th มการเปลยนแปลงตอไปอก (บางครงเรยก Th ขณะนวา Th0) โดย APC หลงไซโทไคน ชนด IL-12 ทาให Th0 กลาย เปน Th1 ทาหนาทหลกในการตอบสนองการตดเชอโรคภายในเซลล และเมอ Th0 กลายเปน Th1 จะผลตไซโท-ไคนชนดแกมมาอนเตอรเฟยรอน (γ-interferon; IFN-γ) ออกมาในปรมาณสง แตถา APC ผลตไซโทไคนชนดอน ไดแก IL-4 หรอ IL-5 จะทาให Th0 กลายเปน Th2 ทาหนาทตอบสนองตอเชอโรคทอยภายนอกเซลล ไดแก การตดเชอพยาธ รวมทงการไดรบสารพษและสารกอภมแพตางๆ(1)

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
44
Acquired/Adaptive Immunity ชนดตอบสนองแบบอาศยแอนตบอด (Humoral Immune Response)
การตอบสนองแบบนอาศยลมโฟไซตชนดบ ซงรวมตวกนอยภายในถงเลก (follicle) ในตอมนาเหลอง
ลมโฟไซตชนดบรบสญญาณกระตนจาก CD4+ T-lymphocyte ทกลาวขางตน แลวพฒนาตอไปเปนเซลลพลาสมาเพอทาหนาทผลตแอนตบอดหลงออกมาในกระแสเลอด และสวนทเปนของเหลวนอกเซลลได แอนต- บอดม 5 ชนดไดแก อมมโนโกลบลน (immunoglobulin, Ig) ชนด IgD, IgM, IgE, IgA และ IgG แอนตบอดแตละชนดมคณสมบตและหนาทแตกตางกนไป สญญาณท ลมโฟไซตชนดบรบจาก Th นนมสวนกาหนดชนดของแอนตบอด กลาวคอ หากเปนสญญาณจาก Th1 แอนตบอดท ถกผลตขนมหนาทโดดเดนในการเพมศกยภาพของขบวนการกลนกนโดยเซลลกลนกน แตถาสญญาณมาจาก Th2 จะไดแอนตบอดทใชทาลายพษ ยบยงเชอโรคไมใหเขาสเซลลได รวมทงกระตนเซลลจาพวกอโอสโนฟลและเซลลมาสตใหหลงสารภายในแกรนลออกมาทาลายเชอโรคทอยภายนอกเซลล (1) ความสมพนธระหวางความเครยดและการตอบ สนองของระบบภมคมกน
ความเชอมโยงโดยตรงท เดนชดระหวางความ เครยดและระบบภมคมกนของรางกาย คอ เมอรางกายรบร
ถงความเครยด จะกระตนโครงขาย HPA และ SAM ใหหลงฮอรโมนท เกยวของกบความเครยดออกมา และเนองจากเซลลหลายชนดททาหนาทในระบบภมคมกน
ดงทกลาวมาขางตนนนมทรบฮอรโมนเหลานนอย เมอรบแลวจะมผลใหเซลลทาหนาทเปลยนแปลงไป ดงตวอยางแสดงในตารางท1
นอกจากน ไซโทไคนทเซลลหลายชนดของระบบภมคมกนผลตขนยงสามารถยอนกลบขนไปควบคม ไฮโพทาลามสใหผลต corticotrophin releasing factor มากขน เพมการหลงฮอรโมนความเครยดตอไปอก จงอาจกลาวไดวาความสมพนธระหวางระบบประสาท ตอมไรทอ และภมคมกน ทางานผานโครงขาย HPA และ SAM ซงเปนไปดงแสดงในรปท 1 ความเครยดเพมโอกาสในการตดเชอโรคและเพมความเสยงทโรคมความรนแรง
งานวจยในสตวทดลองแสดงใหเหนวาความเครยด
มผลตอการตดเชอไวรสไขหวดใหญ ศกษาโดยการจากดบร เวณของหนทดลองใหอยในขอบเขตเฉพาะทหน
สามารถเดนหนาและถอยหลงไดเทานนโดยไมสามารถ
หมนตวกลบได จนเกดความเครยด (restraint stress) แลวใหหนรบเชอไวรสไขหวดใหญ
หลงจากนนตรวจการตอบ
ตารางท 1 ความสมพนธระหวางเซลลทสามารถตอบสนองตอฮอรโมนทเกยวของกบความเครยดชนดตางๆ
ฮอรโมน เซลลทมทรบฮอรโมน ผลทมตอเซลล เอกสารอางอง
ACTH โมโนไซต เปลยนแปลงการเคลอนทของเซลล (12)
ลมโฟไซตชนดท กลม CD4+Th หลง IL-4 สงขน มการพฒนา Th2 มาก (30)
กลโคคอรตคอยด เซลลเดนดรทก ยบยงการเจรญของเซลลเดนดรทก ลดการผลตไซโทไคน IL-6, IL-12 และลดการนาแอนตเจนเปปไทดบน MHC class I
(10,36)
นวโทรฟล ลดประสทธภาพในการกลนกน และลดการสราง oxidative free radicals
(21,35)
โมโนไซต ควบคมการหลงไซโทไคนและการอกเสบ (38)
ลมโฟไซตชนดท กลม CD4+Th กระตนการพฒนา Th2 (28,39)
อะดรนาลน (เอพเนฟรน)
เซลลเดนดรทก ลดการหลงไซโทไคน IL-6, IL-12 , IL-23 (17)
นอรอะดรนาลน (นอรเอพเนฟรน)
ลมโฟไซตชนดท กลม CD8+ CTL
ลดการสราง TNF-α และ การทางานของ CTL (8,20)
ลมโฟไซตชนดท กลม CD4+Th กระตนการผลต IFN-γ และการพฒนา Th1 (34)

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
45
รปท 1 ความสมพนธเชอมโยงของระหวางระบบประสาท ตอมไรทอ และภมคมกน เมอไดรบความเครยดผานโครงขาย HPA และ SAM
สนองของระบบภมคมกน เปรยบเทยบกบหนกลมทไมไดถกจากดบรเวณ พบวา หนทเครยดมระดบ IL-1 กลไกการอกเสบ และการแทรกตวของเซลลกลนกนเพอทาลายไวรสทเนอเยอปอดตากวากลมทดลอง(25) และมปรมาณเชอไวรสไขหวดใหญในปอดสงกวากลมทดลอง(18) การตานทานเชอไวรสไขหวดใหญทลดลงนยงพบไดในหนทมความเครยด
อกลกษณะหนง คอ หนทมประวตการถกแยกจากแมในชวง 1-14 วนหลงคลอดอกดวย(3) อยางไรกตาม เมอใหสาร glucocorticoid receptor antagonist เพอยบยงการ
ทางานของฮอรโมนกลโคคอรตคอยด พบวาการตอบ สนองท ผดปกตดงกลาวลดลง และปรมาณเชอไวรส
ไขหวดใหญลดลงจนใกลเคยงกบกลมควบคม(33)
การทดลองในสตว เชน ลงวอก (rhesus macaque) ซงมความใกลเคยงกบคน การสลบสบเปลยนสมาชกในกลมลงบอยๆ จะทาใหเกดภาวะเครยดทางสงคม การศกษาผลการตอบสนองตอเชอ simian immunodeficiency virus เปรยบเทยบระหวางลงกลมทมความเครยดนกบลงทไมม
การสลบสบเปลยนสมาชกในกลมซงไมมภาวะดงกลาว
พบวา ลงกลมแรกมปรมาณไวรสในเลอดสงกวาและม
อตราการอยรอดตากวากลมทไมมความเครยด(4,5) ผล
การศกษาผปวยตดเชอ human immunodeficiency virus หรอไวรสกอโรคเอดสกแสดงใหเหนวาภาวะเครยดทาง
สงคมสงกระตนใหมการแสดงอาการในผปวยทไมเคย
แสดงอาการมากอนและทาใหพฒนาการของโรครนแรง
ขนเมอเปรยบเทยบในระยะเวลาการศกษาประมาณ 5 ป(26)
การตดตามนกศกษาแพทยทตดเชอไวรส Epstein-Barr ซงเปนไวรสทพบไดทวไปและมกแอบแฝงอยโดยไมแสดงอาการ พบวา ภาวะความเครยดในชวงการสอบกระตนใหไวรสมการเจรญและเพมจานวนออกจากเซลล
มากขน ระดบลมโฟไซตชนดท กลม CD8+ CTL ทจาเพาะกบไวรสลดลงกวาในภาวะทไมมความเครยด(16) ผลการ
วเคราะหเพมเตมพบวา ฮอรโมนความเครยดกลโคคอรต-คอยด เปนตวการหลกในการสงเสรมการเจรญของไวรสดงกลาว(13)
ความเครยดทาใหประสทธภาพในการสรางภมตานทานโรคชาลงหลงไดรบวคซน
การรบวคซนเปนการกระตน acquired immunity ทงแบบอาศยแอนตบอดและอาศยเซลลใหรจกและจดจา
เชอทออนกาลงหรอแอนตเจนจากเชอในวคซน แลว

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
46
สามารถตานทานการตดเชอโรคไดจรงอยางมประสทธ- ภาพโดยผรบวคซนจะไมเปนโรค
ความเครยดสงผลกระทบตอการสรางภมตานทาน หลงไดรบวคซน การศกษาในกลมคนทมความเครยด เรอรงจากภาระหนาทตองดแลคสมรสทปวยดวยโรคสมอง
เสอม (dementia) พบวา หลงไดรบวคซนปองกนไขหวดใหญจะมการสรางแอนตบอดและไซโทไคน IL-2 ใน
ระดบตากวากลมคนเพศและชวงอายเดยวกนทไมตองรบ
หนาทดงกลาว(22) ซงจะสงผลตอการตานเชอ รายงานลกษณะคลายคลงกนยงพบไดกบการรบวคซนปองกนโรค
หด(29) และไวรสตบอกเสบชนดบ(14) โดยกลมคนทมความวตกก งวลหรอมภาวะซม เศร าจะมระดบการสร าง
แอนตบอดและปรมาณลมโฟไซตชนดท ลดลงกวากลมคนควบคมทไมมความเครยด
สรป ความเครยดมผลรบกวนการทางานของรางกายท
เ ชอมโยงกน ต งแตระบบประสาท ตอมไรทอ และภมคมกน การมความเครยดเรอรงทาใหรางกายหลงฮอรโมนทมผลเปลยนแปลงการทางานของเซลลในระบบ
ภมคมกนทมหนาททง innate และ acquired immunity บนทอนการมสขภาพทด โดยทาใหภมตานทานตอเชอโรคตาลง การศกษาเพมเตมเพอหากลไกการยบยงผลกระทบดงกลาวและการหาวธลดความเครยดอยางมประสทธภาพ
เปนแนวทางในการคนควาทจะชวยใหมนษยมชวตใน
สงคมปจจบนไดอยางมความสขมากขน
กตตกรรมประกาศ ขอขอบคณ ดร.ณฐพล ออนปาน ในการวาดภาพ
ประกอบบทความ
เอกสารอางอง 1. Abbas A, Lichtman A. Basic immunology: function
and disorders of the immune system. Pennsylvania: WB Saunders Company; 2001.
2. Arms K, Camp P. Biology. Florida: Saunders College Publishing; 1995.
3. Avitsur R, Hunzeker J, Sheridan JF. Role of early stress in the individual differences in host response to viral infection. Brain Behav Immun 2006; 20:339-48.
4. Capitanio JP, Lerche NW. Social separation, housing relocation, and survival in simian AIDS: a retrospective analysis. Psychosom Med 1998;60: 235-44.
5. Capitanio JP, Mendoza SP, Lerche NW, Mason WA. Social stress results in altered glucocorticoid
regulation and shorter survival in simian acquired immune deficiency syndrome. Proc Natl Acad Sci USA 1998; 95:4714-9.
6. Chen E, Miller GE. Stress and inflammation in exa- cerbations of asthma. Brain Behav Immun 2007;21: 993-9.
7. Cohen S, Tyrrell DA, Smith AP. Psychological stress and susceptibility to the common cold. N Engl J Med 1991;325:606-12.
8. Cook-Mills JM, Cohen RL, Perlman RL, Chambers DA: Inhibition of lymphocyte activation by cate-cholamines: evidence for a non-classical mechanism of catecholamine action. Immunology 1995; 85:544-9.
9. Elenkov IJ, Chrousos GP. Stress hormones, proin-flammatory and antiinflammatory cytokines, and autoimmunity. Ann NY Acad Sci 2002; 966:290-303.
10. Elftman MD, Norbury CC, Bonneau RH, Trucken-miller, M.E. Corticosterone impairs dendritic cell maturation and function. Immunology 2007;122: 279-90.
11. Frankenhaeuser M Stress. In: Underwood G, editor. Oxford guide to the mind. New York: Oxford University Press; 2001. p. 183-5.
12. Genedani S, Filaferro M, Carone C, Oston R, Bucci L, Cevenini E et al. Influence of f-MLP, ACTH(1-24) and CRH on in vitro chemotaxis of monocytes from centenarians. Neuroimmunomodulation 2008; 15:285-9.
13. Glaser R, Kutz L, MacCallum RC and Malarkey, WB. Hormonal modulation of Epstein-Barr virus replication. Neuroendocrinology 1995;62:356-61.
14. Glaser R, Kiecolt-Glaser JK, Bonneau, RH, Malarkey W, Kennedy S, Hughes J. Stress-induced modulation of the immune response to recombinant hepatitis B vaccine. Psychosom Med 1992;54:22-9.
15. Glaser R, Kiecolt-Glaser JK. Stress-induced immune dysfunction: implications for health. Nat Rev Immunol 2005;5:243-51.
16. Glaser R, Padgett DA, Litsky ML, Baiocchi RA, Yung EV, Chen M et al. Stress-associated changes in the steady-state expression of latent Epstein-Barr virus: implications for chronic fatigue syndrome and cancer. Brain Behav Immun 2005;19:91-103.
17. Goyarts E, Matsui M, Mammone T, Bender AM, Wagner JA, Maes D, Granstein RD. Norepinephrine modulates human dendritic cell activation by altering cytokine release. Exp Dermatol 2008;17:188-96.
18. Hunzeker J, Padgett DA, Sheridan PA, Dhabhar FS, Sheridan JF. Modulation of natural killer cell activity by restraint stress during an influenza A/PR8 infection in mice. Brain Behav Immun 2004;18:526-35.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
47
19. Janeway C, Travers P, Walport M, Shlomchik M. Immunobiology: The immune system in health and disease. New York: Garland Science Publishing; 2005.
20. Kalinichenko VV, Mokyr MB, Graf LH, Jr, Cohen RL; Chambers DA. Norepinephrine-mediated inhibition of antitumor cytotoxic T lymphocyte generation involves a beta-adrenergic receptor mechanism and decreased TNF-alpha gene ex-pression. J Immunol 1999;163:2492-9.
21. Kaufmann I, Eisner C, Richter P, Huge V, Beyer A, Chouker A et al. Psychoneuroendocrine stress response may impair neutrophil function in complex regional pain syndrome. Clin Immunol 2007;125: 103-11.
22. Kiecolt-Glaser JK, Glaser R, Gravenstein S, Malarkey WB, Sheridan J. Chronic stress alters the immune response to influenza virus vaccine in older adults. Proc Natl Acad Sci USA 1996;93:3043-7.
23. Koh KB, Park JK, Kim CH, Cho S. Development of the stress response inventory and its application in clinical practice. Psychosom Med 2001;63:668-78.
24. Koh KB, Lee YJ, Beyn KM, Chu SH; Kim DM. Counter-stress effects of relaxation on proin-flammatory and anti-inflammatory cytokines. Brain Behav Immun 2008;22:1130-7.
25. Konstantinos AP, Sheridan JF. Stress and influenza viral infection: modulation of proinflammatory cytokine responses in the lung. Respir Physiol 2001; 128:71-7.
26. Leserman J, Jackson ED, Petitto JM, Golden RN, Silva SG, Perkins DO et al. Progression to AIDS: the effects of stress, depressive symptoms, and social support. Psychosom Med 1999;61:397-406.
27. Linn MW. A global assessment of recent stress (GARS) scale. Int J Psychiatry Med 1985;15:47-59.
28. Miller AH, Spencer RL, Pearce BD, Pisell TL, Azrieli Y, Tanapat P et al. Glucocorticoid receptors are dif-ferentially expressed in the cells and tissues of the immune system. Cell Immunol 1998;186:45-54.
29. Morag M, Morag A, Reichenberg A, Lerer B, Yirmiya R. Psychological variables as predictors of rubella antibody titers and fatigue: a prospective, double blind study. J Psychiatr Res 1999;33:389-95.
30. Okada K, Sugiura T, Kuroda E, Tsuji S, Yamashita U. Phenytoin promotes Th2 type immune response in mice. Clin Exp Immunol 2001;124:406-13.
31. Padgett DA, Glaser R. How stress influences the immune response. Trends Immunol 2003;24:444-8
32. Segerstrom SC, Miller GE. Psychological stress and the human immune system: a meta-analytic study of 30 years of inquiry. Psychol Bull 2004; 130:601-30.
33. Sheridan JF, Dobbs C, Jung J, Chu X, Konstantinos A, Padgett D et al. Stress-induced neuroendocrine modulation of viral pathogenesis and immunity. Ann NY Acad Sci 1998;840:803-8.
34. Swanson MA, Lee WT, Sanders VM. IFN-gamma production by Th1 cells generated from naive CD4+ T cells exposed to norepinephrine. J Immunol 201; 166:232-40.
35. Trabold B, Gruber M, Frohlich D. Functional and phenotypic changes in polymorphonuclear neutrophils induced by catecholamines. Scand Cardiovasc J 2007;41:59-64.
36. Truckenmiller ME, Princiotta MF, Norbury CC, Bonneau RH. Corticosterone impairs MHC class I antigen presentation by dendritic cells via reduction of peptide generation. J Neuroimmunol 2005;160: 48-60.
37. Yang EV, Glaser R. Stress-associated immunomo- dulation and its implications for responses to vacci-nation. Expert Rev Vaccines 2002;1:453-9.
38. Yeager MP, Pioli PA, Wardwell K, Beach ML, Martel P, Lee HK et al. In vivo exposure to high or low cortisol has biphasic effects on inflammatory response pathways of human monocytes. Anesth Analg 2008;107:1726-34.


49
สารตานอนมลเสร กบทางสายกลาง
ระพ บญเปลอง ภาควชาชววทยา คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
ปจจบน มการนาสารตานอนมลเสรมารบประทานเพอเสรมสขภาพกนเปนจานวนมาก เกดจากแนวคดทวา สารตานอนมลเสรมคณสมบตในการกาจดสารอนมลเสรและสารไวปฏกรยาตางๆ ซงเปนสาเหตของโรคนานาชนด สารตานอนมลเสรมทงชนดทเปนเอนไซม เชน ซเปอรออกไซดดสมวเทส แคทาเลส และกลตาไธโอนเพอรออกซเดส เปนตน และสารตานอนมลเสรชนดทไมใชเอนไซม เชน วตามนอ วตามนซ และบทาแคโรทน เปนตน การไดรบสารตานอนมลเสรในปรมาณมากเกนไป นอกจากจะไมกอประโยชนแตอยางใดแลว อาจเปนอนตรายตอรางกายกเปนได
คาสาคญ: อนมลเสร; สารตานอนมลเสร; วตามนอ; วตามนซ; แคโรทนอยด; เอนไซมซเปอรออกไซดดสมวเทส; เอนไซมแคทาเลส; เอนไซมกลตาไธโอนเพอรออกซเดส
Abstract
Nowadays, consumption of antioxidant-containing food supplements has become a common practice in hope of ridding the body of free radicals and reactive compounds, the main cause of aging and various diseases. Antioxidant agents are categorized into two main groups: enzymatic antioxidants, including superoxide dismutase, catalase and glutathione peroxidase, and non-enzymatic antioxidants, including tocopherol (vitamin E), ascorbic acid (vitamin C) and β-carotene. Over consumption of these antioxidants could lead to undesirable adverse effects.
Keywords: free radicals; antioxidant; vitamin E; vitamin C; carotenoid; superoxide dismutase; catalase; glutathione peroxidase; adverse effect
บทนา เมอกลาวถง อนมลเสร (อนมลอสระ หรอ free radi-
cals) และสารตานอนมลเสร (antioxidant) คนทวไปมกคดวาอนมลเสรเปนสงเลวราย เพราะเปนสาเหตสาคญของการแก(25,32) และการเกดโรคนานาชนด เชนพารกนสน (Parkin- son’s disease)(34) อลไซเมอร (Alzheimer’s disease)(31,42,49) ฮนตงตน (Huntington’s disease)(26) ความดนโลหตสง(56) โรคมะเรง(72) และโรคไต(10) เปนตน รวมทงยงเกยวของกบการเปนหมนในเพศชาย(71) แตคนทวไปมกเชอวา สารตานอนมลเสรเปนสงทด มคณประโยชนตอรางกาย จงมการใชสารตานอนมลเสรเพอชะลอการแกรวมทงปองกนและ
รกษาโรคตางๆ ซงไดผลดในระดบหนง แตการใชสารตานอนมลเสรในปรมาณมากเกนไป อาจนามาซงผลรายทคาดไมถง
อนมลเสร คอ อะตอมหรอโมเลกลทมอเลกตรอนจา นวนค (1 1) ทาใหไม เสถยร และจดเปนสารไวปฏกรยาเนองจากพยายามแยงอเลกตรอนจากโมเลกลอน ๆ เพอให
ตนเองเสถยร จงทาใหเกดความเสยหายกบชวโมเลกล อนๆ หลายชนด(46) เชน ดเอนเอ (DNA) อารเอนเอ (RNA) โปรตน และไขมน เปนตน(1,15,29)
นอกจากสารอนมลเสรแลว ยงมสารอกกลมหนงทคนทวไปอาจไมคอยคนเคย คอ สารไวปฏกรยา ซงแบง เปน 2 กลมใหญๆ ไดแก สารไวปฏกรยาทประกอบดวยออกซเจน (reactive oxygen species; ROS) และสารไวปฏกรยาทประกอบดวยไนโตรเจน (reactive nitrogen species; RNS) สารในสองกลมนมทงชนดทเปนและไม
เปนอนมลเสร ดงแสดงในตารางท 1
แหลงกาเนดทสาคญของสารไวปฏกรยาทง 2 ชนดทกลาวขางตน พบไดทงภายในและภายนอกรางกาย เราจงหลกเลยงสารเหลานไดยาก ตวอยางของแหลงกาเนดของสารไวปฏกรยาทพบภายนอกรางกาย ไดแก รงสแกมมา และรงสอลตราไวโอเลต ซงสามารถแยกโมเลกลของนาใหกลายเปน OH• (อนมลไฮดรอกซล)(29) และยงสามารถ

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
50
ตารางท 1 สารไวปฏกรยาทประกอบดวยออกซเจน (reactive oxygen species หรอ ROS) และสารไวปฏกรยาทประกอบ ดวยไนโตรเจน (reactive nitrogen species หรอ RNS)(29)
สารไวปฏกรยา สารอนมลเสร สารทไมเปนอนมลเสร ROS Superoxide radical, O2•
- Hydrogen peroxide, H2O2 Hydroxyl radical, OH• Hypochlorous acid, HOC1 Peroxyl radical, RO2• Ozone, O3 Alkoxyl radical, RO• Hydroperoxyl radical, HO2•
RNS Nitric oxide radical, NO• Nitrous acid, HNO2 Nitrogen dioxide radical, NO2• Dinitrogen tetroxide, N2O4 Dinitrogen trioxide, N2O3 Nitrous acid, HNO2 Peroxynitrite, ONOO- Peroxynitrous acid, ONOOH Nitronium cation, NO2
+
Alkyl peroxynitrites, ROONO
กระตนใหเกดการสราง OH• และ O2•- (อนมลซเปอร
ออกไซด) ภายในเซลล(63) จงสามารถทาใหโปรตนภายในเซลลเสยหายไดอกดวย(18) สวนแหลงกาเนดภายในรางกายของสารไวปฏกรยานน มอย 4 แหลงดวยกน(1,72) คอ
1 . กระบวนการขนส งอ เ ล กตรอน (electron transport chain) เพอสรางพลงงานในไมโทคอนเดรย หากเกดการผดพลาดในการถายโอนอเลกตรอน (ดงแสดงในรปท 1) จะทาใหออกซเจนประมาณรอยละ 2-3 ทเซลลใชในการสรางพลงงาน ถกเปลยนเปน O2•
- OH• และ H2O2
(55,68) มการประมาณวา ในเซลลหนแตละเซลลนน มการสราง O2•
- และ H2O2 มากถง 2 หมนลานโมเลกลตอ วน(1) ในขณะทในรางกายมนษยแตละคนนน เฉลยแลวมการสราง O2•
- มากถง 2 กโลกรมตอป (29 ) และสารไวปฏกรยาทสรางขนเหลาน กอใหเกดความเสยหายตอสารพนธกรรมภายในเซลลไดมากถง 1 แสนครงตอวนตอเซลลหนหนงเซลล และมากถง 1 หมนครงตอวนตอเซลลมนษยหนงเซลล(1)
รปท 1 กระบวนการถายโอนอเลกตรอนใหกบออกซเจน ซงทาใหเกดสารไวปฏกรยาในแตละขนได(1)
2. เซลลเมดเลอดขาวในระบบภมคมกน ซงปกตทาหนาทกาจดสงแปลกปลอม เชน ไวรสและแบคทเรย ดวยวธการกลนกน (phagocytosis) และสงแปลกปลอมเหลานจะถกทาลายดวย NO• (อนมลไนตรกออกไซด) O2•
- และ
H2O2 ทถกสรางขนภายในเมดเลอดขาวเอง หากมการตดเชอแบคทเรยอยางตอเนอง จะทาใหเกดการอกเสบเรอรง และมการผลตสารไวปฏกรยาอยางตอเนอง ซงอาจกอให เกดความเสยหายของสารพนธกรรมและเปนสาเหตสาคญ
ของโรคมะเรงได(1)
3. เพอรออกซโซม (peroxisome) เปนออรแกเนลล ทพบในเซลลทมเยอหมนวเคลยส หรอเซลลยแครโอต ทาหนาทยอยสลายกรดไขมนโดยม H2O2 เปนผลพลอยได
(1) นอกจากน ยงพบวา มการสราง NO• และ O2•
- ภายใน เพอรออกซโซมอกดวย(17,19) ซงหากสารไวปฏกรยาเหลานหลดรอดออกมาจากเพอรออกซโซมได ยอมจะกอใหเกดความเสยหายตอชวโมเลกลอนๆ ภายในเซลลไดเชนกน(1)
4. การทางานของเอนไซมในกลมไซโทโครมพ-450 (cytochrome P-450) ในกระบวนการกาจดสารพษภายในเซลล มผลทาใหจานวนอเลกตรอนของออกซเจนลดลง กลายเปน O2•
- ซงเปนสาเหตของความเสยหายของโมเลกลอนๆ ภายในเซลล(1,22)

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
51
ถงแมเราไมอาจปองกนการสรางสารไวปฏกรยาได แตในรางกายของสงมชวตมกลไกในการควบคมสารไว
ปฏกรยาเหลาน เพอไมใหกอความเสยหายแกเซลลไดโดยผานการทางานของสารตานอนมลเสร ซงเปนสารทปองกน กาจด รวมทงซอมแซมความเสยหายทเกดจากสารอนมลเสรและสารไวปฏกรยา สารตานอนมลเสรเหลานมทงชนดทเปนเอนไซม ไดแก ซเปอรออกไซดดสมวเทส (superoxide dismutase หรอ SOD) แคทาเลส (catalase หรอ CAT) และกลตาไธโอนเพอรออกซเดส (glutathione peroxidase หรอ GPx) เปนตน(30,67) และชนดทไมใชเอน- ไซม เชน โทโคฟรอล (tocopherols หรอ วตามนอ) กรดแอสคอบค (ascorbic acid หรอ วตามนซ) และแคโรท-นอยด (carotenoid) เปนตน
สารตานอนมลเสรในรางกาย หากมปรมาณไมเพยงพอทจะกาจดหรอควบคมปรมาณสารอนมลเสร และสารไวปฏกรยาไมใหกอความเสยหายได จะเกดสภาวะทเรยกวา ออกซเดทฟสเตรส (oxidative stress) ซงจะทาใหสารพนธกรรม โปรตน และไขมนภายในเซลลเสยหาย และอาจทาใหเซลลตายไดในทสด(1,38,67) อนเปนสาเหตสาคญของการแก(25,32) และโรคดงกลาวขางตน
สารตานอนมลเสรชนดทเปนเอนไซม
สารตานอนมลเสรทเปนเอนไซม ไดแก ซเปอร-ออกไซดดสมวเทส แคทาเลส และกลตาไธโอนเพอรออก-ซเดส เมอทางานรวมกน จะเปลยน O2•
- ใหกลายเปน H2O2 และตอมาเปลยนเปนนาและออกซเจนในทสด (ดงแสดงในรปท 2) โดยในคน ซเปอรออกไซดดสมวเทส ซงเปนเอนไซมตานอนมลเสรททรงประสทธภาพทสดภายใน
เซลล มอย 3 ชนดหลกๆ ไดแก 1) แมงกานสซเปอรออก- ไซดดสมวเทส (Mn-SOD) ในไมโทคอนเดรย 2) คอป-เปอร/ซงคซเปอรออกไซดดสมวเทส (Cu/Zn-SOD) ในไซ-โทซอล และ 3) ซเปอรออกไซดดสมวเทสชนดทพบภาย นอกเซลล (extracellular SOD หรอ EC-SOD) หนาทหลกของซเปอรออกไซดดสมวเทสเหลาน คอ เปลยน O2•
- ใหกลายเปน H2O2 ซงมความไวปฏกรยานอยกวา
(72)
แคทาเลสเปนเอนไซมตานอนมลเสรทพบไดใน
เพอรออกซโซม มประสทธภาพมากในการเปลยน H2O2
ใหเปนนาและออกซเจน มการประเมนวา แคทาเลส 1 โมเลกล สามารถกาจด H2O2 ไดถง 6 ลานโมเลกล (7 2 ) ในขณะทกลตาไธโอนเพอรออกซเดสเปนกลมเอนไซม
ตานอนมลเสรทมอยดวยกน 5 ชนดยอย ( 8 ) ทาหนาทเชนเดยวกนกบแคทาเลส คอ เปลยน H2O2ใหเปนนา แต
เกดควบคกบปฏกรยาออกซเดชนทเปลยนรดวซกลตาไธ-โอน (reduced glutathione หรอ GSH) ใหเปนออกซไดซ-กลตาไธโอน (oxidized glutathione หรอ GSSG) ดงแสดงในรปท 2
รปท 2 การทางานของเอนไซมตานอนมลเสรในการเปลยน O2•- ให
กลายเปนนาและออกซเจน (O2•-:อนมลซเปอรออกไซด, H2O2: ไฮโดรเจนเพอรออกไซด, SOD: ซเปอรออกไซดดส-มวเทส, GPx: กลตาไธโอนเพอรออกซเดส, CAT: แคทาเลส, GSH: รดวซกลตาไธโอน, GSSG: ออกซไดซกลตาไธโอน, NO•:อนมลไนตรกออกไซด, ONOO-: เพอรออกซไนไตรท, NO2
-:ไนไตรทอออน )(22,72)
หากปรมาณของเอนไซมตานอนมลเสรเหลานมไม
เพยงพอ O2•- และ H2O2 สามารถทาปฏกรยากบอออนของ
โลหะ เชน เหลก (Fe2+) หรอทองแดง (Cu+) ผานปฏกรยาเฟนตน (Fenton reaction ปฏกรยาท 2 ในรปท 3) และปฏกรยาฮารเบอรไวซ (Harber-Weiss reaction ปฏกรยาท 3 ในรปท 3)(38) เกดเปน OH• ซงเปนอนมลเสรทไวปฏกรยามาก และกอใหเกดความเสยหายตอชวโมเลกลภายในเซลลไดเชนเดยวกบสารไวปฏกรยาชนดอนๆ(22,72)
รปท 3 การสราง OH• (อนมลไฮดรอกซล) จาก O2•
- (อนมลซ-เปอรออกไซด) และ H2O2 โดยมโลหะ (Mn+ เชน เหลก (Fe2+) หรอทองแดง (Cu+) เปนตวเรงปฏกรยา(22,38,72)
มรายงานผลการทดลองจานวนมากทแสดงใหเหน
วา การเพมปรมาณของเอนไซมตานอนมลเสรทงโดยวธ การทางพนธวศวกรรม และโดยวธอนๆ ทาใหสตวทดลองเพมขดความสามารถในการควบคมและกาจดสารอนมล
เสรและสารไวปฏกรยาได
มผลการทดลองในหนดงน ปรมาณแคทาเลสทเพม ขนโดยวธทางพนธวศกรรม ชวยลดความเสยหายทเกดขนแกอวยวะภายในของหนทมอายมากขนได และยงลดความเสยงตอโรคมะเรงลงดวย(69) อกทงสามารถเพมอายขยเฉลยของหนขนไดถงรอยละ 21(65) เมอเพมปรมาณซเปอรออก- ไซดดสมวเทสโดยวธทางพนธวศวกรรม พบวา สามารถ

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
52
ชวยปองกนความเสยหายของจอตาของหนทเปนโรคเบา หวานได(35,39) ชวยลดพษของควนบหร(52) และชวยชะลอการสญเสยความสามารถในการเรยนรและการจดจาในหน
ทมอายมากขน(33) และการเพมปรมาณกลตาไธโอนเพอร-ออกซเดสดวยวธทางพนธวศวกรรม สามารถปกปองเซลลตบออนทผลตอนซลน (47) กลามเนอหวใจ (66 ,7 7) เซลลประสาท(23,74,75) ทนทานตอสภาวะออกซเดทฟสเตรส(45) รวมทงยงชวยใหเซลลตายลดลง(27) และตานทานการเจรญ เตบโตของเซลลมะเรงอกดวย(40)
การศกษาสมองของนกและหนพบวา เมอมอายมากขน จะมการสะสมของสารอนมลเสรและสารไวปฏกรยาในสมองมากขนตามไปดวย และพบวาสารทมฤทธแบบเด ยว กบซ เปอร ออกไซดดสม ว เทสและแคทา เลส (SOD/catalase mimetic) สามารถลดปรมาณของสารอนมลเสรและสารไวปฏกรยาในสมองของสตวเหลานลงได(64)
เมอใหสารทมฤทธแบบเดยวกบซเปอรออกไซด-ดสมวเทสและแคทาเลสกบหนอนปลอง Caenorhabditis elegans สายพนธปกต พบวา สามารถลดปรมาณของสารอนมลเสรและสารไวปฏกรยาลงไดเชนกน(62) และหนอนปลองเหลานนมอายขยเฉลยมากกวาหนอนทไมไดรบสาร
ดงกลาวถงรอยละ 54 และยงพบอกวาในหนอนปลอง C. elegans ทแกเรวผดปกต เปนเพราะขาดยน mev-1 ซงเปนยนทผลตไซโทโครม บ (cytochrome b) ในกระบวน การถายโอนอเลกตรอน เปนผลทาใหมอนมลเสรและสารไวปฏกรยาสะสมภายในเซลลจานวนมาก จงทาใหมอายขยเฉลยสนกวาหนอนสายพนธปกตถงรอยละ 37 แตเมอไดรบสารทมฤทธแบบเดยวกบซเปอรออกไซดดสมวเทส
และแคทาเลส พบวาหนอนเหลานมอายขยเฉลยเพมขนมาเทากบหนอนสายพนธปกตทไมไดรบสารดงกลาวได(43)
นอกจากนยงพบวา สารทมฤทธแบบเดยวกบซเปอรออกไซดดสมวเทสและแคทาเลสสามารถปกปอง
เซลลทสรางฮอรโมนอนซลนในตบออนของมนษยจาก
สารไวปฏกรยาได(48) สวนสารเอบซลน (ebselen) ซงเปนสารทมฤทธแบบเดยวกบซ เปอรออกไซดดสมวเทส สามารถปองกนหนทดลองจากการคกคามของสารอนมล
เสรและสารไวปฏกรยาได(28,70)
จากตวอยางทงหมดขางตนทาใหสรปไดวา อนมลเสรและสารไวปฏกรยาเปนสาเหตสาคญของการแก และโรคภยไขเจบตางๆ ดงนน หากสามารถควบคมและกาจดสารเหลานได อาจจะทาใหคนเรามชวตยนยาวและปลอดโรคบางชนดได(43) อยางไรกตาม มรายงานจานวนมากเชนกนทระบวา สารตานอนมลเสรเหลาน ถงแมวาจะ
ปองกน กาจด และลดความเสยหายจากการคกคามโดยสารอนมลเสรและสารไวปฏกรยาไดกจรง แตไมไดมผลทาใหสตวทดลองมอายยนยาวขนแตประการใด
การทดลองในแมลงวนบาน หนอนปลอง C. elegans และแมลงหว เมอใหรบสารมฤทธแบบเดยวกบซเปอร-ออกไซดดสมวเทสและแคทาเลส พบวา สารเหลานชวยลดปรมาณสารอนมลเสรและสารไวปฏกรยาลงไดจรง แตไมมผลทาใหสตวทดลองเหลานนมอายยนยาวขนแตอยาง ใด(2,20,36,37,41)
มรายงานทบงชวา การมปรมาณสารตานอนมลเสรมากเกนไป อาจเปนอนตรายตอชวตกได เชน หนทมปรมาณกลตาไธโอนเพอรออกซเดสชนดทอยในพลาสมา (plasma glutathione peroxidase หรอ GPx-P) เพมขนดวยวธทางพนธวศวกรรม ทนตอสภาวะอณหภมสงไดนอยกวาหนสายพนธปกตทอยในสภาวะเดยวกน(44) และหนทดลองทมปรมาณกลตาไธโอนเพอรออกซเดสชนดทพบใน ไซโทซอล (cytosolic glutathione peroxidase หรอ GPx-1) เพมขนดวยวธทางพนธวศวกรรม มความออนแอตอสภาวะออกซเดทฟสเตรสทเกดจากการไดรบยาแกปวด อะเซตามโนเฟนหรอพาราเซตามอลเกนขนาด(45) หรอการไดรบสารทเปนพษตอระบบประสาท(8) มากกวาหนสายพนธปกต จงอาจเปนไปไดวา การทมปรมาณกลตา-ไธโอนเพอรออกซเดสมากกวาปกตถง 4-6 เทา จะเขาไปรบกวนสมดลของระบบตานสารอนมลเสร ทาใหระบบดงกลาวลมเหลวและออนแอตอสภาวะออกซเดทฟสเตรส
ในทสด(8,44,45)
สารตานอนมลเสรชนดทเปนเอนไซม แคโรทนอยด เปนกลมของสารรงควตถทส ง
เคราะหในพชและจลนทรย ประกอบดวย บทาแคโรทน (β-carotene) และแอลฟาแคโรทน (α-carotene) พบมากในผกผลไมสเหลอง และสสม ไลโคปน (lycopene) พบมากในผลมะเขอเทศ และลทน (lutein) พบมากในผกใบเขยว
เขม(57) สารจาพวกแคโรทนอยดมความสามารถในการกาจดสารอนมลเสรโดยการสญเสยอเลกตรอนใหกบสาร
อนมลเสร ทาใหกลายเปนแคโรทนอยดทเปนสารอนมลเสร แตสารแคโรทนอยดทเปนอนมลเสรน มความเสถยรกวาอนมลอสระอน (รปท 4) จงไมทาใหเกดปฏกรยาตอเนองทจะทาใหเกดความเสยหายแกชวโมเลกลอนๆ เชน ไขมนและโปรตน เปนตน(22,57,72) มรายงานวาบทาแค-โรทน ซงเปนสารในกลมแคโรทนอยดสามารถลดความเสยงการเปนโรคหลอดเลอดหวใจ(76) ในขณะทไลโคปนซงเปนสารในกลมแคโรทนอยดมผลในการลดความดน

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
53
โลหต ลดความเสยงจากภาวะกระดกพรน และลดภาวะมบตรยากในเพศชายลงได(57)
ในแตละประเทศ มการกาหนดปรมาณไลโคปนทควรไดรบในแตละวนแตกตางกนไป เชน 3 .7-16.2 มลลกรมในประเทศสหรฐอเมรกา 25.2 มลลกรมในประเทศแคนาดา 1.3 มลลกรมในประเทศเยอรมน และ 1.1 มลลกรมในประเทศองกฤษ แตอยางไรกตาม สาหรบผทมสขภาพดแลว ไลโคปนปรมาณ 5-7 มลลกรม เพยงพอทจะปองกนโรคภยไขเจบไดด ในขณะผทปวยเปนโรคตางๆ เชน โรคมะเรง และโรคหลอดเลอดหวใจ อาจตองการ ไลโคปนสงถง 35-70 มลลกรมตอวนเลยทเดยว(57)
รปท 4 การทางานของสารตานอนมลเสรชนดทไมใชเอนไซม (R•:
สารอนมลเสร, R: สารทไมเปนอนมลเสร, Vit E: วตามนอ, Vit E •: วตามนอทเปนสารอนมลเสร, Vit C: วตามนซ, Vit C •: วตามนซทเปนสารอนมลเสร, Car : แคโรทนอยด, Car •: แคโรทนอยดทเปนสารอนมลเสร)(14,22,72)
อยางไรกตาม มรายงานวา การรบประทานบทา- แคโรทนมไดลดความเสยงของการเกดโรคจอประสาทตา
เสอม (aged related macular degeneration; AMD)(21) และยงพบอกวา บทาแคโรทนปรมาณสง แทนทจะชวยลดปรมาณสารอนมลเสรและสารไวปฏกรยา กลบทาใหเกดสารอนมลเสรและสารไวปฏกรยาเพมมากขน และไปยบยงการตายของเซลลมะเรงบางชนด รวมทงยงอาจเพมความเสยงตอโรคมะเรงปอดมากขนดวย ทงนอาจเปนไปไดวา ในสภาวะทมระดบออกซเจนสง บทาแคโรทนปรมาณสงจะเปนตวกระตนสภาวะออกซเดทฟสเตรสเสยเอง (72) นอกจากน แคโรทนอยดยงเปนองคประกอบสาคญในการสงเคราะหวตามนเออกดวย โดยเหตท วตามนเอนนมความสาคญตอสายตาและการเจรญเตบโตเปนอยางยง(57) จงทาใหมการบรโภควตามนเอในปรมาณสง เพอหวงใหมผลดตอสายตาและลดการตายของทารกในครรภ(73) มการกาหนดปรมาณวตามนเอทผใหญควรไดรบในแตละวนไว
ท 700-900 ไมโครกรม(5) สวนหญงตงครรภควรไดรบวตามนเอประมาณวนละ 1,200-1,300 ไมโครกรม แตใน
วตามนรวม 1 เมด อาจมวตามนเอสงถง 5,000 หนวยสากล ซงเทยบไดกบ 1,500 ไมโครกรมวตามนเอโดยประมาณ(50) ซงอาจมากเกนไป เนองจากการรบประทานวตามนเอในปรมาณทสงกวาปรมาณทแนะนาในแตละวน อาจจะรบกวนการทางานของวตามนดในการดดซมแคลเซยมจาก
อาหาร(24,50) และทาใหเกดภาวะกระดกพรนมากขนได(51,58)
วตามนอเปนกลมสารซงมคณสมบตละลายไดใน
นามน แบงเปน 2 กลมใหญๆ ไดแก กลมโทโคฟรอล ประกอบดวย แอลฟาโทโคฟรอล (α-tocopherol) บทาโท-โคฟรอล (β-tocopherol) แกมมาโทโคฟรอล (γ-toco-pherol) และเดลทาโทโคฟรอล (δ-tocopherol) สวนอก
กล มหน ง ไดแก กลมโทโคไทรอนอล (tocotrienol) ประกอบดวย แอลฟาโทโคไทรอนอล (α-tocotrienol) บทาโทโคไทรอนอล (β-tocotrienol) แกมมาโทโคไทรอ-นอล (γ- tocotrienol) และเดลทาโทโคไทรอนอล (δ-toco-trienol) แอลฟาโทโคฟรอลเปนวตามนอสวนใหญทพบไดในธรรมชาต เชน ในนามนพช ในเมลดถว และในผกใบเขยว เปนตน ในแตละวน รางกายควรไดรบวตามนอ ประมาณ 15-30 มลลกรม(5,59)
วตามนอ มบทบาทสาคญในการกาจดอนมลเสร(78) โดยการสญเสยอเลกตรอนใหกบสารอนมลเสร ทาใหกลายเปนวตามนอทเปนสารอนมลเสร ซงในทสดจะถกวตามนซ เปลยนใหกลบมาเปนวตามนออกคร ง (รป ท 4)(22,59,72) ถงกระนนกตาม มรายงานวา วตามนอไมสามารถลดภาวะความเสยงตอโรคมะเรง โรคหลอดเลอดหวใจและโรคเบาหวานลงได และยงไปกวานน การบรโภควตามนอมากเกนไป อาจกอใหเกดอนตรายอกดวย เชน เพมความเสยงของอาการหวใจวาย และทาใหการตดเชอในระบบหายใจเรอรงยาวนานขน(9,16)
วตามนซ หรอ กรดแอสคอรบค พบไดทวไปในผลไมหลากหลายชนด เชน ฝรง สม มะเขอเทศ มนฝรง เปนตน วตามนซเปนสารทมคณสมบตละลายนา ทาหนาทสาคญในการสงเคราะหคอลลาเจน ซงเปนสวนสาคญของเนอเยอเกยวพนในรางกาย ชวยในการสงเคราะหนาด การดดซมธาตเหลก(12) และสาคญอยางยงในการกาจดสารอนมลเสร โดยการสญเสยอเลกตรอนใหกบสารอนมลเสร ทาใหกลายเปนวตามนซทเปนสารอนมลเสร (รปท 4) ซงไมทาใหเกดปฏกรยาลกโซทจะทาลายชวโมเลกลอนๆ รวมทงยงทางานรวมกบวตามนอในการกาจดอนมลเสรอก
ดวย(22,59,60,72,78) แตเนองจากรางกายมนษยไมสามารถสงเคราะหวตามนซไดเอง จงมความจาเปนตองไดรบวตามนซใหพอเพยง ในแตละประเทศมการกาหนดปรมาณวตามนซทควรไดรบในแตละวนแตกตางกนออกไป ผใหญควรไดรบวตามนซประมาณ 30-70 มลลกรม(5) และตองสงขนกวานอกหากเปนหญงทกาลงตงครรภหรออย
ในชวงใหนมบตร(60) การไดรบวตามนซอยางพอเพยง จะชวยลดความเสยงตอการเปนโรคหลอดเลอดหวใจลงได(76) แตการไดรบวตามนซมากเกนไป ( เชน สงถง 6,000 มลลกรมตอวน) อาจทาใหเกดอาการทองรวง และเพม

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
54
ความเสยงตายในทารกกอนคลอดหรอแรกเกดได(60,61) จงมการกาหนดไววา หญงมครรภควรไดรบวตามนซไมเกน 2,000 มลลกรมตอวน(60) นอกจากนยงมรายงานวา วตามนซอาจลดประสทธภาพของยารกษามะเรงบางชนดอก ดวย(79) สวนการใหวตามนซและวตามนอในหญงตงครรภเพอเปนการปองกนภาวะความดนโลหตสงจากการ
ตงครรภ (pre-eclampsia)(13) อาจไมไดผลและยงเสยงตอการททารกทเกดมาโดยมนาหนกนอยกวาปกตอกดวย(54)
สรป ในปจจบน มการนาสารตานทานอนมลเสร เชน
วตามนตางๆ ในรปแบบผลตภณฑเสรมอาหารมาใชกนอยางแพรหลาย เพอปองกนโรคตางๆ แตวตามนเหลานนอาจไมใชยาอาย วฒนะอยางท เ ชอถอกน ตามรายงานการศกษาจานวนมากทสามารถนามาสรปไดวา การไดรบวตามนตางๆ ในปรมาณสงไมไดชวยลดความเสยงในการเปนโรคมะเรงในทางเดนอาหาร โรคทเกยวกบหลอดเลอดหวใจ ระบบประสาท สายตา ผวหนง หรอไขขอ(3,5,7,53) ทงยงอาจเปนการเพมความเสยงตอโรคตางๆ และเพมอตราการเสยชวตได(4,6) ดงนน หากตองการรบวตามนเสรมเพอบารงสขภาพ ควรรบประทานในปรมาณทพอเหมาะ กลาวคอ ในแตละวน ใหไดรบวตามนเอไมเกน 3,000 ไมโครกรม วตามนซไมเกน 2,000 มลลกรม และวตามนอไมเกน 1,000 มลลกรม (5) มฉะนนอาจไดรบผลเสยตอสขภาพแทนกเปนได
เอกสารอางอง 1. Ames BN, Shigenaga MK, Hagen TM. Oxidants,
antioxidants, and the degenerative diseases of aging. Proc Natl Acad Sci USA 1993;90:7915-22.
2. Bayne AC, Sohal RS. Effects of superoxide dismutase/catalase mimetics on life span and oxidative stress resistance in the housefly, Musca domestica. Free Radic Biol Med 2002;32:1229-34.
3. Bjelakovic G, Nikolova D, Simonetti RG, Gluud C. Antioxidant supplements for prevention of gastro-intestinal cancers: a systematic review and meta-analysis. Lancet 2004;364:1219-28.
4. Bjelakovic G, Nikolova D, Gluud LL, Simonetti RG, Gluud C. Mortality in randomized trials of anti-oxidant supplements for primary and secondary prevention: systematic review and meta-analysis. JAMA 2007;297:842-57.
5. Bjelakovic G, Nikolova D, Gluud LL, Simonetti RG, Gluud C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database Syst Rev 2008: CD007176.
6. Bjelakovic G, Nikolova D, Simonetti RG, Gluud C. Systematic review and meta-analysis: primary and secondary prevention of gastrointestinal cancers with antioxidant supplements. Aliment Pharmacol Ther 2008;28:689-703.
7. Bjelakovic G, Nikolova D, Simonetti RG, Gluud C. Antioxidant supplements for preventing gastro-intestinal cancers. Cochrane Database Syst Rev 2008:CD004183.
8. Boonplueang R, Akopian G, Stevenson FF, Kuhlenkamp JF, Lu SC, Walsh JP et al. Increased susceptibility of glutathione peroxidase-1 transgenic mice to kainic acid-related seizure activity and hippocampal neuronal cell death. Exp Neurol 2005;192:203-14.
9. Brigelius-Flohe R. Adverse effects of vitamin E by induction of drug metabolism. Genes Nutr 2007;2: 249-56.
10. Cachofeiro V, Goicochea M, de Vinuesa SG, Oubina P, Lahera V, Luno J. Oxidative stress and inflamma-tion, a link between chronic kidney disease and cardiovascular disease. Kidney Int 2008;74:S4-9.
11. Carey FA. Organic chemistry, 7th ed. New York: McGraw-Hill; 2008.
12. Catley L, Anderson KC. Velcade and vitamin C: too much of a good thing? Clin Cancer Res 2006;12:3-4.
13. Chappell LC, Seed PT, Briley AL, Kelly FJ, Lee R, Hunt BJ et al. Effect of antioxidants on the occurrence of pre-eclampsia in women at increased risk: a randomised trial. Lancet 1999;354:810-6.
14. Chaudiere J, Ferrari-Iliou R. Intracellular antioxi-dants: from chemical to biochemical mechanisms. Food Chem Toxicol 1999;37:949-62.
15. Cheng FC, Jen JF, Tsai TH. Hydroxyl radical in living systems and its separation methods. J Chromatogr B Analyt Technol Biomed Life Sci 2002;781:481-96.
16. Cook NR, Albert CM, Gaziano JM, Zaharris E, MacFadyen J, Danielson E et al. A randomized factorial trial of vitamins C and E and beta carotene in the secondary prevention of cardiovascular events in women: results from the women's antioxidant cardiovascular study. Arch Intern Med 2007;167: 1610-8.
17. Corpas FJ, Barroso JB, Carreras A, Quiros M, Leon AM, Romero-Puertas MC et al. Cellular and subcellular localization of endogenous nitric oxide in young and senescent pea plants. Plant Physiol 2004; 136:2722-33.
18. Davies MJ. Singlet oxygen-mediated damage to proteins and its consequences. Biochem Biophys Res Commun 2003;305:761-70.
19. del Rio LA, Corpas FJ, Sandalio LM, Palma JM, Gomez M, Barroso JB. Reactive oxygen species,

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
55
antioxidant systems and nitric oxide in peroxisomes. J Exp Bot 2002;53:1255-72.
20. Doonan R, McElwee JJ, Matthijssens F, Walker GA, Houthoofd K, Back P et al. Against the oxidative damage theory of aging: superoxide dismutases protect against oxidative stress but have little or no effect on life span in caenorhabditis elegans. Genes Dev 2008;22:3236-41.
21. Evans JR, Henshaw K. Antioxidant vitamin and mineral supplements for preventing age-related macular degeneration. Cochrane Database Syst Rev 2008:CD000253.
22. Fang YZ, Yang S, Wu G. Free radicals, antioxidants, and nutrition. Nutrition 2002;18:872-9.
23. Furling D, Ghribi O, Lahsaini A, Mirault ME, Massicotte G. Impairment of synaptic transmission by transient hypoxia in hippocampal slices: improved recovery in glutathione peroxidase transgenic mice. Proc Natl Acad Sci USA 2000; 97:4351-6.
24. Genaro Pde S, Martini LA. Vitamin A supplementa-tion and risk of skeletal fracture. Nutr Rev 2004; 62:65-7.
25. Gerschman R, Gilbert DL, Nye SW, Dwyer P, Fenn WO. Oxygen poisoning and x-irradiation: a mecha-nism in common. Science 1954;119:623-6.
26. Gil JM, Rego AC. Mechanisms of neurode- generation in huntington's disease. Eur J Neurosci 2008;27:2803-20.
27. Gouaze V, Andrieu-Abadie N, Cuvillier O, Malagarie-Cazenave S, Frisach MF, Mirault ME et al. Glutathione peroxidase-1 protects from cd95-induced apoptosis. J Biol Chem 2002;277:42867-74.
28. Guven A, Tunc T, Topal T, Kul M, Korkmaz A, Gundogdu G et al. Alpha-lipoic acid and ebselen prevent ischemia/reperfusion injury in the rat intestine. Surg Today 2008;38:1029-35.
29. Halliwell B. Antioxidants in human health and disease. Annu Rev Nutr 1996;16:33-50.
30. Halliwell B, Gutteridge J. Free radicals in biology and medicine, 3rd ed. New York: Oxford University Press; 1999.
31. Harman D. Alzheimer's disease pathogenesis: Role of aging. Ann NY Acad Sci 2006;1067:454-60.
32. Harman D. Free radical theory of aging. An update: increasing the functional life span. Ann NY Acad Sci 2006;1067:10-21.
33. Hu D, Serrano F, Oury TD, Klann E. Aging- dependent alterations in synaptic plasticity and memory in mice that overexpress extracellular superoxide dismutase. J Neurosci 2006;26:3933-41.
34. Jenner P. Oxidative stress in parkinson's disease. Ann Neurol 2003;53 (suppl 3):S26-36; discussion S36-38.
35. Kanwar M, Chan PS, Kern TS, Kowluru RA. Oxidative damage in the retinal mitochondria of diabetic mice: possible protection by superoxide dismutase. Invest Ophthalmol Vis Sci 2007;48:3805-11.
36. Keaney M, Gems D. No increase in lifespan in caenorhabditis elegans upon treatment with the superoxide dismutase mimetic euk-8. Free Radic Biol Med 2003;34: 277-82.
37. Keaney M, Matthijssens F, Sharpe M, Vanfleteren J, Gems D. Superoxide dismutase mimetics elevate superoxide dismutase activity in vivo but do not retard aging in the nematode Caenorhabditis elegans. Free Radic Biol Med 2004;37:239-50.
38. Kehrer JP. The haber-weiss reaction and mecha-nisms of toxicity. Toxicology 2000;149:43-50.
39. Kowluru RA, Kowluru V, Xiong Y, Ho YS. Over-expression of mitochondrial superoxide dismutase in mice protects the retina from diabetes-induced oxi-dative stress. Free Radic Biol Med 2006;41:1191-6.
40. Liu J, Hinkhouse MM, Sun W, Weydert CJ, Ritchie JM, Oberley LW et al. Redox regulation of pancrea-tic cancer cell growth: role of glutathione peroxidase in the suppression of the malignant phenotype. Hum Gene Ther 2004;15:239-50.
41 Magwere T, West M, Riyahi K, Murphy MP, Smith RA, Partridge L. The effects of exogenous antioxi-dants on lifespan and oxidative stress resistance in drosophila melanogaster. Mech Ageing Dev 2006; 127:356-70.
42. Martin LJ. DNA damage and repair: relevance to mechanisms of neurodegeneration. J Neuropathol Exp Neurol 2008;67:377-87.
43. Melov S, Ravenscroft J, Malik S, Gill MS, Walker DW, Clayton PE et al. Extension of life-span with superoxide dismutase/catalase mimetics. Science 2000;289:1567-9.
44. Mirochnitchenko O, Palnitkar U, Philbert M, Inouye M. Thermosensitive phenotype of transgenic mice overproducing human glutathione peroxidases. Proc Natl Acad Sci USA 1995;92:8120-4.
45. Mirochnitchenko O, Weisbrot-Lefkowitz M, Reuhl K, Chen L, Yang C, Inouye M. Acetaminophen toxicity. Opposite effects of two forms of glutathione peroxidase. J Biol Chem 1999;274:10349-55.
46. Möller MN, Lancaster JR Jr, Denicola A, Sadis M. The interaction of reactive oxygen and nitrogen species with membranes. New York: Academic Press; 2008. p.23-42. (Current topics in membranes; vol 61).
47. Moriscot C, Richard MJ, Favrot MC, Benhamou PY. Protection of insulin-secreting ins-1 cells against oxidative stress through adenoviral-mediated gluta-thione peroxidase overexpression. Diabetes Metab 2003;29:145-51.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
56
48. Moriscot C, Candel S, Sauret V, Kerr-Conte J, Richard MJ, Favrot MC et al. Mntmpyp, a metallo-porphyrin-based superoxide dismutase/catalase mimetic, protects ins-1 cells and human pancreatic islets from an in vitro oxidative challenge. Diabetes Metab 2007;33:44-53.
49. Nunomura A, Castellani RJ, Lee HG, Moreira PI, Zhu X, Perry G et al. Neuropathology in alzheimer's disease: awaking from a hundred-year-old dream. Sci Aging Knowledge Environ 2006;2006:p10.
50. Penniston KL, Tanumihardjo SA. Vitamin A in dietary supplements and fortified foods: too much of a good thing? J Am Diet Assoc 2003;103:1185-7.
51. Penniston KL, Tanumihardjo SA. The acute and chronic toxic effects of vitamin A. Am J Clin Nutr 2006;83:191-201.
52. Petrache I, Medler TR, Richter AT, Kamocki K, Chukwueke U, Zhen L et al. Superoxide dismutase protects against apoptosis and alveolar enlargement induced by ceramide. Am J Physiol Lung Cell Mol Physiol 2008;295:L44-53.
53. Plummer M, Vivas J, Lopez G, Bravo JC, Peraza S, Carillo E et al. Chemoprevention of precancerous gastric lesions with antioxidant vitamin supplemen-tation: a randomized trial in a high-risk population. J Natl Cancer Inst 2007;99:137-46.
54. Poston L, Briley AL, Seed PT, Kelly FJ, Shennan AH. Vitamin C and vitamin E in pregnant women at risk for pre-eclampsia (vip trial): randomised placebo-controlled trial. Lancet 2006;367:1145-54.
55. Powers SK, Jackson MJ. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol Rev 2008;88:1243-76.
56. Puddu P, Puddu GM, Cravero E, Rosati M, Muscari A. The molecular sources of reactive oxygen species in hypertension. Blood Press 2008;17:70-7.
57. Rao AV, Rao LG. Carotenoids and human health. Pharmacol Res 2007;55:207-16.
58. Ribaya-Mercado JD, Blumberg JB. Vitamin A: is it a risk factor for osteoporosis and bone fracture? Nutr Rev 2007;65:425-38.
59. Rumbold A, Crowther CA. Vitamin E supplementa-tion in pregnancy. Cochrane Database Syst Rev 2005:CD004069.
60. Rumbold A, Crowther CA. Vitamin C supplemen-tation in pregnancy. Cochrane Database Syst Rev 2005: CD004072.
61. Rumbold A, Middleton P, Crowther CA. Vitamin supplementation for preventing miscarriage. Cochrane Database Syst Rev 2005:CD004073.
62. Sampayo JN, Olsen A, Lithgow GJ. Oxidative stress in Caenorhabditis elegans: protective effects of superoxide dismutase/catalase mimetics. Aging Cell 2003;2:319-26.
63. Sarma L, Devasagayam TP, Mohan H, Mittal JP, Kesavan PC. Mechanisms of protection by buthionine sulphoximine against gamma-ray-induced micro-nuclei in polychromatic erythrocytes of mouse bone marrow. Int J Radiat Biol 1996;69:633-43.
64. Sasaki T, Unno K, Tahara S, Shimada A, Chiba Y, Hoshino M et al. Age-related increase of superoxide generation in the brains of mammals and birds. Aging Cell 2008;7:459-69.
65. Schriner SE, Linford NJ, Martin GM, Treuting P, Ogburn CE, Emond M et al. Extension of murine life span by overexpression of catalase targeted to mitochondria. Science 2005;308:1909-11.
66. Shiomi T, Tsutsui H, Matsusaka H, Murakami K, Hayashidani S, Ikeuchi M et al. Overexpression of glutathione peroxidase prevents left ventricular remodeling and failure after myocardial infarction in mice. Circulation 2004;109:544-9.
67. Sies H. Oxidative stress: from basic research to clinical application. Am J Med 1991;91:31S-38S.
68. Sohal RS, Weindruch R. Oxidative stress, caloric restriction, and aging. Science 1996;273:59-63.
69. Treuting PM, Linford NJ, Knoblaugh SE, Emond MJ, Morton JF, Martin GM et al. Reduction of age-associated pathology in old mice by overexpression of catalase in mitochondria. J Gerontol A Biol Sci Med Sci 2008;63:813-22.
70. Tripathi DN, Jena GB. Ebselen attenuates cyclo-phosphamide-induced oxidative stress and DNA damage in mice. Free Radic Res 2008;42:966-77.
71. Turner TT, Lysiak JJ. Oxidative stress: a common factor in testicular dysfunction. J Androl 2008;29: 488-98.
72. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact 2006;160: 1-40.
73. Van DE, Kulier R, Gulmezoglu AM, Villar J. Vitamin A supplementation during pregnancy. Cochrane Database Syst Rev 2002:CD001996.
74. Wang H, Cheng E, Brooke S, Chang P, Sapolsky R. Over-expression of antioxidant enzymes protects cultured hippocampal and cortical neurons from necrotic insults. J Neurochem 2003;87:1527-34.
75. Weisbrot-Lefkowitz M, Reuhl K, Perry B, Chan PH, Inouye M, Mirochnitchenko O. Overexpression of human glutathione peroxidase protects transgenic mice against focal cerebral ischemia/reperfusion damage. Mol Brain Res 1998;53:333-8.
76. Ye Z, Song H. Antioxidant vitamins intake and the risk of coronary heart disease: meta-analysis of cohort studies. Eur J Cardiovasc Prev Rehabil 2008; 15:26-34.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
57
77. Yoshida T, Watanabe M, Engelman DT, Engelman RM, Schley JA, Maulik N et al. Transgenic mice overexpressing glutathione peroxidase are resistant to myocardial ischemia reperfusion injury. J Mol Cell Cardiol 1996;28:1759-67.
78. Zaidi SM, Banu N. Antioxidant potential of vitamins A, E and C in modulating oxidative stress in rat brain. Clin Chim Acta 2004;340:229-33.
79. Zou W, Yue P, Lin N, He M, Zhou Z, Lonial S et al. Vitamin C inactivates the proteasome inhibitor ps-341 in human cancer cells. Clin Cancer Res 2006; 12:273-80.


59
โรคฉหน: ภยทมากบนา
กลลยาน ดวงฉว ภาควชาพยาธชววทยา คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
โรคฉหนมผลกระทบตอชวตผคนทอยใกลชดสงแวดลอมซงปนเปอนเชอกอโรคโดยไมรตว จากวถการดารงชวตของเกษตรกรและชาวนาไทยซงสวนใหญมการใชแหลงนาธรรมชาตรวมกบสตว และจากภาวะภยพบตทางธรรมชาตหรอภมอากาศทแปรปรวนเกอบทกแหงทวโลกรวมทงประเทศไทย ทาใหเกดพายรนแรง ฝนตกผดฤดกาลหรอภาวะนาทวมขงยาวนาน จากแนวโนมการระบาดของโรครบจากสตวสคน เชนโรคฉหน ทพบบอยขนและกออนตรายตอชวตผปวยสงขน อะไรเปนปจจยทกระตนใหโรคทวความรนแรง ในการระบาดแตละครงของแตละภมภาคกอคาถามสาคญตามมาคอ อะไรคอสาเหตของการแพรเชอโรคมาสคนทแทจรงและเกดจากเชอสายพนธใด เหลานเปนปญหาทยงรอการศกษาวจย ทงนความสมพนธระหวางคน สตว และสงแวดลอมจะเกยวโยงกบโรคนในประเทศไทยอยางไร ตลอดจนการพฒนาเครองมอสาหรบการตรวจตดตามสถานการณของโรคฉหนทเหมาะสมไดสอดคลองกบความเปนจรงจะมประโยชนตอการปองกน
และควบคมการระบาดหรอรกษาโรคไดตรงเปาหมายทสด
คาสาคญ: เชอฉหน; โรครบจากสตว; การแพรเชอ
Abstract
Leptospirosis is an important zoonotic disease and a global public health problem. Epidemics of leptospirosis have increased unexpectedly with environmental changes. A number of agricultural workers in Thailand develop undifferentiated febrile illness, and some cases recover without treatment while a small number have severe manifestations that can be fatal. Factors that affect disease severity and the zoonotic sources of leptospirosis transmission to humans in Thailand remain unknown. In order to control transmission, an integrated approach is needed including appropriate and effective diagnostic tools to help identify leptospirosis cases.
Keywords: leptospira; zoonosis; transmission
บทนา
โรคฉหนหรอเลปโตสไปโรซส (leptospirosis) เปนโรคทรบจากสตว (zoonosis) และรายแรงถงแกชวตได มกเกดในชวงฤดฝนและในทมนาทวมขง ถอเปนปญหาสา-ธารณสขสาคญของนานาประเทศทวโลก(20,22,28,30) รวมทงประเทศไทย จากสถตเมอป พ.ศ. 2543 ไทยเปนประเทศทมอบตการณสงเปนอนดบ 7 รองจากประเทศเซเชลล ตรนแดด และโตเบโก บารเบโดส จาไมกา คอสตารกา และศรลงกา (ขอมลจาก The International Leptospirosis Society, ILS, worldwide survey ป ค.ศ. 2003) กลมอาชพเสยงไดแก ชาวนา เกษตรกร ททางานไถหวาน งานปศสตว ตลอดจนกจกรรมทสมผสนา พบโรคในคนหลงการแชนานานๆ หรออยในบรเวณพนทชนแฉะทมการปนเปอนเชอฉหนและไมไดสวมใสรองเทาปองกน หรอเกดบาดแผลทผว
หนงจากการเหยยบยาเปลอกหอย (รายงานการสอบสวนโรคผปวยเสยชวตดวยโรคเลปโตสไปโรซส ของจงหวดบรรมย วนท 27 กรกฎาคม พ.ศ. 2551 โดย Surveillance and Rapid Response Team สานกระบาดวทยา กรมควบคมโรค กระทรวงสาธารณสข)
ผทตองสงสยวาเปนโรคฉหน ไดแก ผทแสดงอาการไขเฉยบพลนโดยไมทราบสาเหตชดเจน ไมม โรคประจาตวเรอรงหรอไขนานไมเกน 3 สปดาห ไมม ลกษณะทางคลนกทบงชโรคตดเชออนๆ เชน ไขหวด มาลาเรย ไขเลอดออก ไขสมองอกเสบ สครบไทฟส หรอ เมลออยโดสส สวนผปวยอาจมอาการแสดงไมจาเพาะ
หรอไมมอาการใดๆ จนกระทงรนแรงเสยชวตดวยภาวะตบและไตวาย(16) แตสวนมากผปวยมกมอาการไมรนแรงและหายไดเอง ภายใน 2 สปดาห ในกรณผปวยมอาการ

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
60
แสดง มกปวดศรษะ ปวดตามกลามเนอ ตาแดง หรอเลอด ออกทเยอบตา เปนตน ซงอาการทางคลนกดงกลาวไมใชลกษณะจาเพาะ พบไดกบโรคทมสาเหตอนๆ เชน กน(40) จงจาเปนตองอาศยผลการตรวจทางหองปฏบตการ หากวนจฉยโรคไดทนการ จะสามารถรกษาผปวยใหหายไดดวยยาปฏชวนะ ผปวยท เสยชวตมกเกดจากไดรบการวนจฉยลาชาและการรกษาทไมเหมาะสม การเสยชวตเฉยบพลนเกดเนองจากอาการแทรกซอน เชน ไอเปนเลอด ความดนโลหตตา อาการตวเหลอง ตาเหลอง ตบวาย ไตวาย ปสสาวะนอย ระบบหายใจลมเหลว ดงนนโรคฉหนจงเปนปญหาทางสาธารณสขทเราควรตระหนกและรบรไว
เพอปองกนตน เนองจากมหนตภยนาทวมทเกดขนทวทกหนแหงบนโลกขณะนไมอาจรบประกนไดวาในอนาคตจะ
ไมแฝงดวยภยตางๆ รวมทงเชอฉหนทมาไดพรอมกบนา แหลงเกบกกเชอในธรรมชาต เชอฉหนแยกไดครงแรกเมอป ค.ศ. 1916 โดยนก วจยชาวญปน เปนเวลาเกอบศตวรรษท เราร จก เช อแบคทเรยชนดนเพมกวา 250 สายพนธมทงชนดทกอโรคและไมกอโรคในคน มชอเรยกทางวทยาศาสตรตามการจดกลมตามลกษณะทางพนธกรรมหรอจโนไทป เปน genomospecies หรอจาแนกโดยวธการทางวทยาเซรม (microscopic agglutination test; MAT) เปน serovar ดงตว
(microscopic agglutination test; MAT) เปน serovar ดงตวอยางแสดงในตารางท 1 และ 2 เชอถกทาลายไดหลายวธทงการใชยาฆาเชอ ความรอน ความแลง หรอสภาพความเปนกรด-ดาง (pH) ตากวา 6 หรอสงกวา 8 และพบไดตามแหลงนาธรรมชาต พนดนทชนแฉะหรอมรงโรคในสตวเลยงลกดวยนมหรอสตวเลอดอน ซงเลยงตามบานเรอนหรอฟารม ไดแก ววควาย สกร สนข แมว กวาง แกะ และ มา(6,9,13,39) สตวเหลานมกพบความผดปกต เชน ลกตายในทอง แทงลก โรคไตและตาอกเสบ เปนตน หรอพบไดในสตวอาศยตามธรรมชาตแตมโอกาสใกลชดกบ
คนหรอสตวเลยงเชน สกงค แรคคน คางคาว กระรอก หนนา และหนทอ (18,31,37 ) ทสาคญคอ การสมผสเชอจากปสสาวะของสตวดงกลาวโดยไมรตว โดยเฉพาะอยางยงหนทอทมกพบชกชมตามแหลงขยะ ตลาดสด หรอชมชนแออด ทมระบบจดการสงแวดลอมและสาธารณสขไม ด (2 4 ,29 ,3 5 ,3 6 ) ท งน เปนททราบวาหน เปนแหลงรงโรค (reservoir) ทสาคญ หนเหลานสะสมเชอทไตโดยไมเปนอนตรายตอตวมนเองและยงปลอยเชอทางปสสาวะได
ตลอดเวลา ตราบใดทเรายงไมสามารถควบคมมใหหนแพรขยายพนธอยางรวดเรวหรอขจดเชอฉหนทถกเกบกก
ในตวหนจนตลอดอายขยของมนได คนจะมโอกาสเสยงตอการรบเชอหรอเกดโรคนจากสงแวดลอมทปนเปอนได
ตลอดเวลา
ตารางท 1 ตวอยางชอเรยกและชนดของเชอฉหน (The Committee on the Taxonomy of Leptospira, The International Union of Microbiological Societies, IUMS, 2002, แหลงขอมล: Institute Pasteur Genetics Server)
Genus and Genomospecies Serovar Strain
Leptospira interrogans Leptospira kirshneri Leptospira noguchii Leptospira santarosai Leptospira borgpetersenii Leptospira weilii Leptospira wolbachii Leptospira inadai Leptospira meyeri Leptospira biflexa Turneria parva (proposed) Leptonema illini Leptospira genomospecies 1 Leptospira genomospecies 2 Leptospira genomospecies 3 Leptospira genomospecies 4 Leptospira genomospecies 5
Icterohaemorrhagiae Cynopteri Panama Shermani Javanica Ranarum Codice Lyme Semaranga Patoc Parva Illini Pingchang Manhao3 Holland Hualin Saopaulo
RGA 3522 C CZ 214 1342 K Veldrat Batavia 46 Iowa City Frog (ICF) CDC 10 Veldrat Semarang 173 Patoc I H 3055 80-412 L 60 Waz Holland (P438) LT 11-33 Sao Paulo

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
61
ตารางท 2 การจาแนกเชอฉหนโดยวธทางวทยาเซรม
Genomospecies Serogroup Recommended serovar **
1. L. interrogans 1 Australis Australis [pathogenic] 2 Autumnalis Autumnalis 3 Ballum Castellonis 4 Bataviae Bataviae 5 Canicola Canicola 6 Celledoni 7 Cynopteri Cynopteri 8 Djasiman 9 Grippotyphosa Grippotyphosa 100 Hebdomadis Hebdomadis 11 Icterohaemorrhagiae Icterohaemorrhagiae Copenhageni 12 Javanica Javanica 13 Louisiana 14 Lyme 15 Manhao 16 Mini 17 Panama Panama 18 Pomona Pomona 19 Pyrogenes Pyrogenes 20 Ranarum 21 Sarmin 22 Sejroe Sejroe Wolffi Hardjo 23 Shermani 24 Tarassovi Tarassovi 25 New or undesignated 2. L. biflexa 1 Andamana [non-pathogenic] 2 Codice 3 Semaranga Patoc 3. L. borgpetersenii 4. L. weilii 5. L. kirshneri 6. L. meyeri 7. L. wolbachii
** สายพนธทควรมรวมอยในชดตรวจ MAT
นอกจากสตวและคนทอาศยอยบนบกสามารถตด
เชอไดจากแหลงนาจด หนองนาธรรมชาตหรอสมผสดน ชนแฉะทมการปนเปอนปสสาวะจากสตวทเปนรงโรคแลว ยงพบวามสงโตทะเลปวยและเสยชวตดวยโรคไตจากเชอฉ
หน(18) ในการศกษาตอมา ผสารวจใหขอมลวาสนขนาจะเปนแหลงแพรเชอตามชายฝง(33) จากขอสงเกตพบวาสตว
ทะเลประเภทนจานวนมากปวยดวยอาการปอดบวมและ
ตรวจพบเชอฉหนเพมขนและถขนทก 4-5 ป ตามแนวชายฝงอเมรกาแถบ California-Oregon(19,34,38) อาจเปนเพราะสภาพภมอากาศทเปลยนแปลงอยางรวดเรวและบอยขน ดงนน นกวทยาศาสตรจงรวมมอเพอสารวจปจจยอนๆ ทอาจเกยวโยงถงสตวจาพวก แมวนา ปลาวาฬ ตลอดจนกลไกทเชอแพรกระจายลงสทะเลไดอยางไร เพอหาทางปองกนม
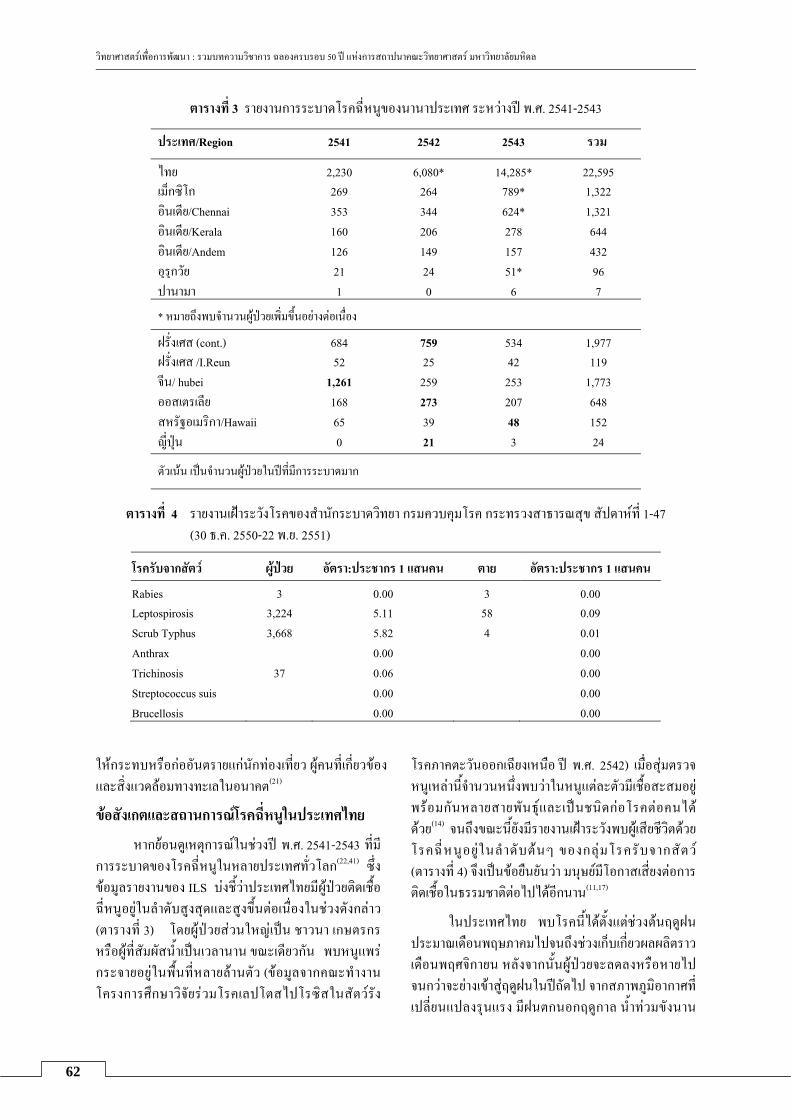
วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
62
ตารางท 3 รายงานการระบาดโรคฉหนของนานาประเทศ ระหวางป พ.ศ. 2541-2543
ประเทศ/Region 2541 2542 2543 รวม
ไทย 2,230 6,080* 14,285* 22,595 เมกซโก 269 264 789* 1,322 อนเดย/Chennai 353 344 624* 1,321 อนเดย/Kerala 160 206 278 644 อนเดย/Andem 126 149 157 432 อรกวย 21 24 51* 96 ปานามา 1 0 6 7
* หมายถงพบจานวนผปวยเพมขนอยางตอเนอง
ฝรงเศส (cont.) 684 759 534 1,977 ฝรงเศส /I.Reun 52 25 42 119 จน/ hubei 1,261 259 253 1,773 ออสเตรเลย 168 273 207 648 สหรฐอเมรกา/Hawaii 65 39 48 152 ญปน 0 21 3 24
ตวเนน เปนจานวนผปวยในปทมการระบาดมาก
ตารางท 4 รายงานเฝาระวงโรคของสานกระบาดวทยา กรมควบคมโรค กระทรวงสาธารณสข สปดาหท 1-47 (30 ธ.ค. 2550-22 พ.ย. 2551)
โรครบจากสตว ผปวย อตรา:ประชากร 1 แสนคน ตาย อตรา:ประชากร 1 แสนคน
Rabies 3 0.00 3 0.00 Leptospirosis 3,224 5.11 58 0.09 Scrub Typhus 3,668 5.82 4 0.01 Anthrax 0.00 0.00 Trichinosis 37 0.06 0.00 Streptococcus suis 0.00 0.00 Brucellosis 0.00 0.00
ใหกระทบหรอกออนตรายแกนกทองเทยว ผคนทเกยวของและสงแวดลอมทางทะเลในอนาคต(21)
ขอสงเกตและสถานการณโรคฉหนในประเทศไทย
หากยอนดเหตการณในชวงป พ.ศ. 2541-2543 ทมการระบาดของโรคฉหนในหลายประเทศทวโลก(22,41) ซงขอมลรายงานของ ILS บงชวาประเทศไทยมผปวยตดเชอฉหนอยในลาดบสงสดและสงขนตอเนองในชวงดงกลาว (ตารางท 3) โดยผปวยสวนใหญเปน ชาวนา เกษตรกร หรอผทสมผสนาเปนเวลานาน ขณะเดยวกน พบหนแพร กระจายอยในพนทหลายลานตว (ขอมลจากคณะทางานโครงการศกษาวจยรวมโรคเลปโตสไปโรซสในสตวรง
โรคภาคตะวนออกเฉยงเหนอ ป พ.ศ. 2542) เมอสมตรวจหนเหลานจานวนหนงพบวาในหนแตละตวมเชอสะสมอย
พรอมกนหลายสายพนธและเปนชนดกอโรคตอคนได ดวย(14) จนถงขณะนยงมรายงานเฝาระวงพบผเสยชวตดวยโรคฉหนอยในลาดบตนๆ ของกลมโรครบจากสตว (ตารางท 4) จงเปนขอยนยนวา มนษยมโอกาสเสยงตอการตดเชอในธรรมชาตตอไปไดอกนาน(11,17)
ในประเทศไทย พบโรคนไดตงแตชวงตนฤดฝน
ประมาณเดอนพฤษภาคมไปจนถงชวงเกบเกยวผลผลตราว
เดอนพฤศจกายน หลงจากนนผปวยจะลดลงหรอหายไปจนกวาจะยางเขาสฤดฝนในปถดไป จากสภาพภมอากาศทเปลยนแปลงรนแรง มฝนตกนอกฤดกาล นาทวมขงนาน

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
63
ขน ยงเพมโอกาสเสยงของการเกดโรคนในพนททไมเคยมการระบาดไปยงคนกลมใหม (ขอมลรายงาน-จงหวดบรรมย ยงไมไดตพมพ) ดงนน โรคฉหนทพบจงไมไดจากดเฉพาะพนทและอาชพเทานน แตมการระบาดในกลมนกทองเทยวในตางประเทศ(9) และประเทศไทย(1-4) (สตล-พงงาพ.ศ. 2550-2551) ในกลมผใชแรงงาน ขดไถดนนอกฤดกาล (รอยเอด พ.ศ. 2549) กลมทหารเกณฑทสมผส
โคลน (นครนายก พ.ศ. 2550) และกลมผรบผลกระทบจากอทกภยทงในพนททเคยมการระบาดมากอนและกลม
ระบาดใหมตงแตทางภาคเหนอของประเทศ (เชยงราย นาน พะเยา และแพร) ภาคตะวนออกเฉยงเหนอ (ชยภม กาฬสนธ และบรรมย) ภาคกลาง (อยธยา) หรอภาคใต (พงงา) รวมทงในกลมผทจบและนาหนมาประกอบเปน
อาหารซงไดรบเชอโดยไมรตวและปวยเขารบการรกษาใน
โรงพยาบาล กระทรวงสาธารณสขไทยไดตระหนกถงปญหาดง กลาว จงมการประกาศพนท เสยงใหประชาชนทราบ รณรงคใหประชาชนหลกเลยงการสมผสสตวปวยทสงสย
หรอสงแวดลอมทปนเปอนปสสาวะของสตวรงโรค ใหรจกการปองกนตนเอง เชน การใช ถงมอ ถงเทา รองเทา ใหความรความเขาใจเกยวกบโรคเพอปองกนความตน
ตระหนกของคนในพนททไมเคยมการระบาดมากอน สาหรบกรง เทพมหานครไมมรายงานผปวยชดเจน นอกจากการสารวจพบหนทอและสตวคลายหน (shrew) ทมเชอฉหนซงอาจกออนตรายตอคนได(14,25,31)
ทกประเทศทวโลกไดเฝาระวงควบคมโรคอยาง
ตอเนอง มการสรางเครอขายแลกเปลยนขอมลขาวสารและรายงานสถานการณทกครงทพบการระบาด มองคกรประสานงานระหวางชาต (ILS) ซงกาหนดใหมการ
ประชมแลกเปลยนประสบการณเปนระยะทก 3 ป เพอเรงแกไขและควบคมปญหาความรนแรงของโรค ดวยเหตทแตละภมภาคบนโลกมปจจยเสยงและเชอกอโรคแตกตาง
กนไป ทาใหขอมลการระบาดทวโลกประมาณ 350,000-500,000 คนตอป นาจะตากวาความเปนจรง(41) เนองจากระบบเฝาระวงและตดตามโรคฉหนยงไมครอบคลมทวถง
และปญหาการขาดแคลนชดตรวจวนจฉยทเหมาะสม
การพฒนาวธตรวจวนจฉยโรค ในประเทศไทยใชการยนยนวนจฉยโรคฉหนตาม
มาตรฐานสากลทมอยหลายวธ ทงนแตละหนวยงานจะเลอกใชวธใดขนอยกบปจจยและความพรอม โดยสรปม 2 แนวทางคอ
1. การตรวจหาเชอ spirochaete ทมลกษณะเปนเกลยว โดยใชกลองจลทรรศนพนมด (dark field micro-scope) แมการเพาะแยกเชอจะใหผลแนนอน แตเปนท
ทราบดวาปญหาและความยงยากของเทคนคมหลาย
ประการ เชน การเพาะเลยงเชอจากผปวยในหลอดเลยงมกไมประสบผลสาเรจ หากเชอมการเจรญเตบโตไดกมกใชเวลานานเปนสปดาหจนถงหลายเดอน จงไมสามารถใชในการวนจฉยโรค สวนการตรวจหาแอนตเจนหรอสารพนธกรรมของเชอยงอยระหวางการวจยพฒนาและมการ
ใชตรวจในสตว (34) และส งแวดลอมบาง (17 ) กาลงรอประยกตใชกบโรคในคน(26,27)
2. วธทนามาใชเพอการวนจฉยไดบางแตไมเหมาะ สมกบทกราย คอการตรวจวดระดบไตเตอรของภมคมกนหรอแอนตบอดตอเชอโรคฉหนดวยวธ MAT และวธ ELISA(10,12) สาหรบวธ MAT ยงคงเปนทนยมใช แมจะมขอจากดและความยงยากจากการเตรยมเชอหลายสายพนธ
(ตารางท 2) พรอมกน ซงควรเปนเชอกลมทเคยกอโรคประจาถนและกลมทคาดวาจะเกดระบาดใหม วธนจงเหมาะสาหรบหองปฏบตการทมความพรอมดานบคลากร เครองมอและเชอในชดทดสอบ รวมทงการแปลผลการตรวจขนกบเงอนไขตางๆ เพอการวนจฉยโรค คอ อยางนอยตวอยางสงตรวจครงแรกทไดมาควรใหผล MAT ไตเตอรมากกวา 1:400 หรอคาเปรยบเทยบระหวางตวอยางสงตรวจ 2 ครงซงเกบหางกน 1-2 สปดาห มไตเตอรเพมขนตงแต 4 เทาขนไป ซงวธ MAT จะยนยนผลไดในผปวยทมอาการแสดงเกนสปดาหหรออยางนอย 8-10 วน(39) แตอาจพบปญหาทตางจากขอกาหนดขางตน คอ ผปวยทม ไตเตอร ตากวา เกณฑกาหนด (1:50-1:200) ตองมการประเมนและพจารณาปจจยอนๆ เพมเตม หรอในกรณตรวจพบไตเตอรในผทไมมอาการแสดง(5,9,32) เปนตน
นอกจากวธดงกลาวขางตนแลว ยงมวธตรวจคดกรองอนๆ เพอขอมลทางระบาดวทยา เชน Lepto-Dipstick, indirect hemagglutination, indirect immuno-fluorescence, macroscopic slide agglutination(7), micro-capsule agglutination, Lepto Tek, Lateral Flow, Lepto Tek Dri-Dot เปนตน ซงแตละวธมขอจากดแตกตางกนไปและเนองจากเปนชดทดสอบทนาเขาจากตางประเทศ เชอทใชเตรยมมกไมครอบคลมและแตกตางจากเชอกอโรค
ของประเทศไทย ผลทไดไมอาจรองรบสถานการณจรง ขอมลทไดจงคลาดเคลอนหรอตากวาความจรง(9) จากการศกษาวจยและพฒนาชดทดสอบแบบ IgM immunoblot(15) ใหผลการประเมนทยนยนโรคได และรวดเรวกวาวธ

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
64
มาตรฐาน MAT อยางไรกตามยงตองการพฒนาเปนชด
ทดสอบใหเหมาะกบการใชในภาคสนามตอไป
กลาวโดยสรป ปจจยจากสงแวดลอมและนาในธรรมชาตมการเปลยนแปลงอยางตอเนอง จงไมอาจคาด การณไดวาโรคฉหนจะเปนปญหาระบาดใหมและระบาด
ซามากนอยอยางไร แตสงสาคญอนดบแรกทตองเรง
พฒนาในปจจบนคอ วธการตรวจวนจฉยทสะดวก รวดเรว สะทอนปญหาการระบาดของโรคได ถกตอง เพ อประโยชนในการลดอตราการเสยชวตหรอปวยโดยไม
ทราบสาเหตใหนอยลง และเมอภยนหมนกลบมาอกครงกจะควบคมไดทนหรอไมรนแรงตอไป
กตตกรรมประกาศ
ผเขยนขอขอบคณ ศ. ดร.รชนย อดมแสงเพชร ทกรณา ใหคาแนะนาในการเขยนบทความน
เอกสารอางอง 1. ดเรก สดแดน, ถนอม นอยหมอ, วราลกษณ ตงคณะกล,
มนศลป ศรมาตย, ไพบลย ทนนไชย, สลกจต ชตพงษวเวท และคณะ. การระบาดครงใหญทสดของโรคฉหนในประเทศไทยจากอทกภย เดอนสงหาคม-กนยายน ปพ .ศ . 2549. รายงานการเฝ าระวงทางระบาดวทยา ประจ าสปดาห 2550;38:885-90.
2. ดเรก สดแดน, วราลกษณ ตงคณะกล, มนศลป ศรมาตย, ไพบลย ทนนไชย, สลกจต ชตพงษวเวท, พจมาน สรอารยา-ภรณ และคณะ. ปจจยทมผลตอการเปนโรคฉหนหลงจากอทกภยจงหวดนาน เดอนสงหาคม-กนยายน พ.ศ. 2549. รายงานการเฝ าระวงทางระบาดวทยา ประจ าสปดาห 2551;39:161-5.
3. เบญจวรรณ ระลก, พจมาน สรอารยาภรณ, มณ ผลภาษ, ธรศกด ชกนา, ดารนทร อารยโชคชย, ดเรก สดแดน และคณะ. การระบาดของโรคเลปโตสไปโรซสในกลมทหารประจาการในคายทหารแหงหนง จงหวดนครนายก วนท 27 พฤศจกายน - 28 ธนวาคม พ.ศ. 2550. รายงานการเฝาระวงทางระบาดวทยา ประจาสปดาห 2552;40:21-5.
4. โรม บวทอง , ททตะ ชลามาตย , วราลกษณ ตงคณะกล , โสภณ ศรเอยมถาวร, วรรณา หาญเชาวกล, คานวณ องชศกด. โรคฉหนกบการทองเทยว: การสอบสวนโรคฉหนในนกทองเทยวชาวฮองกง ภายหลงจากการลองแกง อาเภอเมอง จงหวดพงงา ปพ.ศ. 2550. รายงานการเฝาระวงทางระบาดวทยา ประจาสปดาห 2550;38:541-6.
5. Ashford DA, Kaiser RM, Spiegel RA, Perkins BA, Weyant RS, Bragg SL et al. Asymptomatic infection and risk factors for leptospirosis in Nicaragua. Am J Trop Med Hyg 2000;63:249-54.
6. Ayanegui-Alcerreca MA, Wilson PR, Mackintosh CG, Collins-Emerson JM, Heuer C, Midwinter AC et al. Leptospirosis in farmed deer in New Zealand: a review. NZ Vet J 2007;55:102-8.
7. Brandão AP, da Silva Camargo ED, Silva MV, Abrão RV. Macroscopic agglutination test for rapid
diagnosis of human leptospirosis. J Clin Microbiol 1998;36:3138-42.
8. Cachay ER, Vinetz JM. A global research agenda for leptospirosis. J Postgrad Med 2005;1:174-8.
9. Ciceroni L, Stepan E, Pinto A, Pizzocaro P, Dettori G, Franzin L et al. Epidemiological trend of human leptospirosis in Italy between 1994 and 1996. Eur J Epidemiol 2000;16:79-86.
10. Cumberland PC, Everard COR, Levett PN. Assess-ment of the efficacy of the IgM enzyme-linked im-munosorbent assay (ELISA) and the microscopic agglutination test (MAT) in the diagnosis of acute leptospirosis. Am J Trop Med Hyg 1999;61:731-4.
11. De Faria MT, Calderwood MS, Athanazio DA, McBride AJ, Hartskeerl RA, Pereira MM et al. Carriage of Leptospira interrogans among domestic rats from an urban setting highly endemic for leptospirosis in Brazil. Acta Trop 2008;108:1-5.
12. Diagnosis of Leptospirosis - Role of MAT.[Online]. 2006 ;[2 screens]. Available from: URL:http:// www.japi.org/april2006/Corr3.pdf
13. Dorjee S, Heuer C, Jackson R, West DM, Collins-Emerson JM, Midwinter AC et al. Prevalence of pathogenic Leptospira spp. in sheep in a sheep-only abattoir in New Zealand. NZ Vet J 2008;56:164-70.
14. Doungchawee G, Phulsuksombat D, Naigowit P, KhoaprasertY, Sangjun N, Kongtim S et al. 2005. Survey of leptospirosis of small mammals in Thailand. Southeast Asian Trop Med Pub Health 2005;36:1516-22.
15. Doungchawee G, Kositanont U, Niwetpathomwat A, Invisai T, Sagarasaeranee P, Haake DA. Early diagnosis of leptospirosis by IgM immunoblot testing. CVI 2008;15:492-8.
16. Feigin RD, Anderson DC. Human leptospirosis. CRC Crit Rev Clin Lab Sci 1975;5:413-67.
17. Ganoza CA, Matthias MA, Collins-Richards D, Brouwer KC, Cunningham CB, Segura ER et al. Determining risk for severe leptospirosis by molecular analysis of environmental surface waters for pathogenic Leptospira. PLoS Med 2006;3:e308.
18. George W, Gorman SMcKeever, Roger DG. Leptospirosis in wild mammals from southwestern Georgia. Am J Trop Med Hyg 1962;11:518-24.
19. Gulland FM, Koski M, Lowenstine LJ, Colagross A, Morgan L, Spraker T. Leptospirosis in California sea lions (Zalophus californianus) stranded along the central California coast, 1981-1994. J Wildlife Dis 1996;32:572-80.
20. Hogan MC, Pate G, McConkey SJ, O'Flanagan D, Mongan C, Mellotte GJ. Leptospirosis in the republic of Ireland: 1985 to 1996. Commun Dis Rep CDR Rev 1997;7:186-9.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
65
21. Hunt TD, Ziccardi MH, Gulland FM, Yochem PK, Hird DW, Rowles T et al. Health risks for marine mammal workers. Dis Aquat Organ 2008;81:81-92.
22. ILS World Wide Survey. First ILS worldwide survey on human leptospirosis, 1999 [Online]. 1999; [8 screens]. Available from: URL:http://www. leptonet.net/html/ils_worldwide_survey.asp
23. Jansen A, Schneberg I, Frank C,Alpers K, Schneider T, Stark K. Leptospirosis in Germany, 1962-2003. Emerging Infect Dis 2005;11:1048-54.
24. Ko AI, Galvao RM, Ribeiro DC, Johnson WD Jr, Riley LW. Urban epidemic of severe leptospirosis in Brazil. Salvador Leptospirosis Study Group. Lancet 1999;354:820-5.
25. Kositanont U, DoungchaweeG, Kongtim S, Pulsawat W, Singchai C, Puthavathana P. Inves-tigation of leptospira carrier potential among rodents in Bangkok. Oral Presentation, 4th Scientific Meeting of the International Leptospirosis Society (ILS); 2005 November 14-16; Chiang Mai, Thailand.
26. Kositanont U, Rugsasuk S, Leelaporn A, Phulsuk- sombati D, Tantitanawat S, Naigowit P. Detection and differentiation between pathogenic and saprophytic Leptospira spp. by multiplex poly- merase chain reaction. Diag Microbio&Infect Dis 2007;57:117-22.
27. Kositanont U, Saetun P, Krittanai C, Doungchawee G, Tribuddharat C, Thongboonkerd V. Application of immunoproteomics to leptospirosis: toward clinical diagnostics and vaccine discovery. Proteomics Clin Appl 2007;1:400-9.
28. Leal-Castellanos CB, Garcia-Suarez R, Gonzalez-Figueroa E, Fuentes-Allen JL, Escobedo-de la Pens J. Risk factors and the prevalence of leptospirosis infection in a rural community of Chiapas, Mexico. Epidemiol Infect 2003;131:1149-56.
29. Maciel EA, de Carvalho AL, Nascimento SF, de Matos RB, Gouveia EL, Reis MG, Ko AI. Household transmission of leptospira infection in urban slum communities. PLoS Negl Trop Dis 2008;2:e154.
30. Matthias MA, Ricaldi JN, Cespedes M, Diaz MM, Galloway RL, Saito M et al. Human leptospirosis caused by a new, antigenically unique leptospira associated with a rattus species reservoir in the Peruvian Amazon. PLoS Negl Trop Dis 2008;2: e213.
31. Niwetpathomwat A, Doungchawee G. An investi-gation of rodents leptospirosis in Bangkok,Thailand.
Online J Vet Res 2005;9:95-100. Available from: URL:http://users.comcen.com.au/~journals/jvet196a. htm.
32. Niwetpathomwat A, Niwatayakul K, Doungchawee G. Surveillance of leptospirosis after flooding at Loei province, Thailand by year 2002. Southeast Asian J Trop Med Pub Health 2005;36 Suppl 4 :185-8.
33. Norman SA, Digiacomo RF, Gulland FM, Meschke JS, Lowry MS. Risk factors for an outbreak of leptospirosis in California sea lions (Zalophus californianus) in California, 2004. J Wildl Dis 2008;44:837-44.
34. OSU Vet Diagnostic Lab, Stranding Network Testing Sea Lions for Leptospirosis [Online]. 2007 [2007 Jan 17];[6 screens]. Available from: URL:http://www.oregonstate.edu/dept/ncs/newsarch/2007/Jan07/lepto.html
35. Pate GE, Hogan MC, FitzSimon N, Mellotte GJ. A review of the epidemiology of leptospirosis in the Republic of Ireland. Ir Med J 2000;93:114-7.
36. Reis RB, Ribeiro GS, Felzemburgh RD, Santana FS, Mohr S, Melendez AX et al. Impact of environment and social gradient on leptospira infection in urban slums. PLoS Negl Trop Dis 2008;2:e228.
37. Richardson Dennis J, Gauthier Julie L. A serosurvey of leptospirosis in Connecticut peridomestic wildlife. Vector-borne Zoo Dis 2003;3:187-93.
38. The Marine Mammal Center begins new lepto-spirosis study in California [Online]. 2008 [2008 Oct 22]; screen]. Available from: URL:http://www. eurekalert.org/pub_releases/2008-10/tmmc-tmm102 208.php
39. Thiermann AB, Frank RR. Human leptospirosis in Detroit and the role of rats as chronic carriers. Int J Zoonoses 1980;7:62-72.
40. Trevejo RT, Rigau-Perez JG, Ashford DA, McClure EM, Jarquin-Gonzalez C, Amador JJ et al. Epidemic leptospirosis associated with pulmonary hemorrhage-Nicaragua, 1995. J Infect Dis 1998; 178:1457-63.
41. Weekly Epidemiological Record. [ Online]. 1999 [1999 Jul 23]; [8 screens]. Available from: URL:http://www.http://www.who.int/docstore/wer/74_27_52.html


67
Microglial Activation in Parkinson’s Disease
Rattanavijit Vijitruth Department of Biology, Faculty of Science, Mahidol University.
Abstract
Parkinson’s disease is a severe movement disorder with a mysterious cause. It is a puzzle that challenges all researchers in the field. Nevertheless, the end result is established, the severe degeneration of the dopamine releasing neurons in the substantia nigra pars compacta. It has become more evident that microglial involvement is crucial in the process of neurodegeneration. This review introduces Parkinson’s disease and microglia and discusses related research findings on the effect of activated microglia in Parkinson’s disease. An unidentified insult may cause acute primary damage to neurons leading to the activation of microglia, which can further induce secondary damage to neurons through chronic inflammation. However, this process can be blocked by selectively inhibiting or knocking out cyclooxygenase-2, suggesting a potential therapeutic intervention for Parkinson’s disease by reducing the progressive loss of dopaminergic neurons.
Keywords: microglia; inflammation; Parkinson; oxidative stress; COX-2; neurodegeneration; neurons; dopamine; LPS; cytokines; SNpc
Introduction
Parkinson’s disease (PD) is a prolonged and age-related locomotor impairment. There are over 1.5 million North Americans diagnosed with PD. Parkinson’s disease patients comprise about one percent of all sixty five year olds and five percent by age eighty-five. The classic symptoms are tremor, rigidity, and postural instability. Parkinson’s disease was first described in an 1817 essay by the British physician James Parkinson. He called it “the shaking palsy”. The disease was named after him who identified the major symptoms. For about 150 years, scientists could only define its range of symptoms, distribution among the population, and prospects for cure. Finally in the early 1960s, they identified the pathological basis of the motor defect, which is due to the loss of the dopamine (DA) producing neurons in the SNpc. The neurotransmitter dopamine helps direct muscle activity. The diagnosis of PD is still generally based on clinical symptoms and confirmed with neuropathological examination post-mortem. The most prevalent treatment is replacing the dopamine normally produced in the substantia nigra pars compacta (SNpc) by
supplying the DA precursor, L-dopa. Unfortunately, the treatment is only effective for a short time. Eventually, patients develop an unusual movement (dyskinesia) as one of the major side effects. Psychotic episodes also occur in patients taking L-dopa. In addition, L-dopa treatment does not stop or slow the progression of the disease. As a result, there is a need to find new treatments for PD. Nevertheless, finding a cure remains a difficult task because the etiology of PD is still unknown.
Microglia in Neuroinflammation
There have been advances in research in this field and several factors, both genetic and environmental, have been shown to induce PD or increase the risk of developing PD. Genetic factors account for only 20% of the causes of PD. With the exception of a small number of patients with familial PD, most patients have idiopathic PD. Several lines of evidence suggest that inflammation may be involved in the etiology of several neurodegenerative diseases, such as: amyotrophic lateral sclerosis (ALS), Alzheimer’s disease and PD(5,9,29). Major participants in neuroinflammation are

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
68
microglia. These glia are resident macrophages of the brain sharing similar properties with macrophages(39,43) and constituting 10% of brain cells(26). Once activated, microglia undergo a transformation from striated bodies into large round, amoeboid bodies with short thick processes. In PD, activated microglia in the SNpc have been found to express the major histocompatibility (MHC) II complex HLA-DR(29), the macrophage marker EMB11(3), tumor necrosis factor-α (TNF-α)(6,19), interleukins (e.g. IL-1β, IL-6), interferon-γ, the low affinity immunoglobulin E receptor CD23(19), inducible nitric oxide synthase (iNOS)(17,22), cyclo-oxygenase-2(22), complement 3 receptor(3,33) and increased levels of ferritin(33).
Cyclooxygenase-2 and Activation of Microglia
There are two isoforms of the enzyme cyclooxygenase (COX), namely COX-1 and COX-2, both of which limit the prostaglandin production rate, and use arachidonic acid as their substrate(40, 53). Most tissues of the body regularly express a constant level of COX-1, and only COX-2 is increased in response to pathophysiological inflammatory stimuli (e.g. bacterial lipopolysaccharide (LPS), interleukin-1β (IL-1β), and multiple growth factors(24,37). Reactive oxygen species (ROS) are the by-products of the enzymatic reaction catalyzed by COXs(38), and they can induce microglial activation as can endotoxin (LPS), mitogens, cytokines, and many inflammatory mediators(36). Generally, oxidative stress can activate microglia(8). The widely used indicators for microglial activation in research are the up-regulation of macrophage antigen complex-1, phagocytosis, and the production of cytotoxic molecules including: ROS, nitric oxide (NO), and various proinflammatory cytokines, e.g. IL-1β(2,12,16). Activated microglia are able to phagocytose other cells, physically help in the immunological process, or release substances that can directly or indirectly promote survival or death of nearby, or distant cells. They constitutively express complement receptors and transform into activated immunocompetent antigen-presenting cells in response to various
stimuli(7). Activated microglia cluster around dopaminergic neurons in phagocyto- sis(3,22,25,28,29,50). Vulnerable dopaminergic neu-rons display nitrosylation of proteins(13), express receptors for TNF-α(6) and produce downstream intracellular effector molecules like the nuclear transcription factor NFκB(18). These data indicate an aggressive, selective and active microglia-related inflammatory process pro-moting cell death in PD.
Linking Microglia to PD
Accumulating evidence from studies in PD patient(8,14) and animal models(41,46) indicates increased activation of microglia as has been observed in other diseases with neuro-degeneration(11,23,27). First of all, there are increased levels of inflammatory cytokines, including TNF-α, IL-6, and IL-1β in PD patients’ brains(6,34,35). In addition, many investigators have found that there is a high density of microglia in the SNpc(21,26). Our previous research(49) supports the importance of microglial activation in the degeneration of neurons, revealing a strong correlation between selective COX-2 inhibition or deficiency of COX-2 and a reduction in activated microglia. Another group of investigators had shown reduced microglial activation with selective COX-2 inhibitor treatment in a rat 6-hydroxy dopamine-induced PD model(20). They used this model to induce PD in rats while our group used another toxin of the nervous system, 1-methy-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP), to imitate human PD in mice. Our group demon-strated that MPTP induced a high number of activated microglia in wild type mice, while heterozygous mice having only one functional COX-2 allele had lower numbers of microglia. In homozygous COX-2 knock-out mice the level of activated microglia was the lowest. As a result, we postulated that COX-2 may be instrumental in sustaining several years of inflammation and activation of microglia in people or monkeys exposed to MPTP(4,25,30). Inhibition of microglial activation should reduce the secondary neuronal loss due to pathogenic inflammation. Indeed, reduction in COX-2 activity, through inhibition or knocking out

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
69
COX-2, correlated well with the numbers of activated microglia and the death of dopa-minergic neurons after MPTP intoxication. Thus, our previous study demonstrated that treatment with valdecoxib (a selective COX-2 inhibitor) and COX-2 deficiency decreases MPTP-induced behavioral impairments.
Microglial Involvement in Neuronal Injury
Microglial activation seems to happen before the loss of the neurons producing dopamine(45). Reactive oxygen species (ROS) derived from COX-2 probably mediate excitation of microglia(8). So COX-2 inhibition should affect secondary microglial incitation that perpetuates long-term inflammation in PD induced by MPTP. Our group and others have discovered that the death of dopaminergic neurons resulted from microglial release of NO(1,19,32,52), superoxide free radicals(10), or proinflammatory cytokines like TNF-α(6). Other mechanisms include beading of neurites(47) or stripping dendritic synapses(42) thus breaking neuronal connections and trophic factors(20,51). Secondary injury is likely mediated by microglia releasing NO, ROS, and cytokines. This process is significant with respect to progressive neuronal death and the conti-nuation of the inflammation uncovered in PD. Studies in cultured microglia have found that inhibition of COX-2 decreases the level of iNOS in microglia stimulated by LPS(31). This, in turn, reduces NO production, suggesting a positive feedback effect of COX-2 in releasing toxic molecules from induced microglia. The metabolism of DA by dopaminergic neurons in the SNpc and autoxidation produce ROS, making them susceptible to oxidative stress from inflammatory processes(44). As a consequence, COX-2 involvement in dopa-minergic neurodegeneration is due to the oxidation of DNA(38) or dopamine (to toxic dopamine quinone)(15) and the augmentation of activated microglia. Both parts lead to prolonged inflammation.
Time and space of microglia
Our studies suggest that activation of microglia and degeneration of neurons are related, temporally and spatially(49). The involvement of these related events along with COX-2 up-regulation, have been supported by others(46,48,54). Apparently, COX-2 may modulate stimulation of microglia and magnify the inflammation and other toxic effects, which exacerbate degeneration of dopaminergic neurons. Since MPP+ (MPTP metabolite) causes initial death of dopaminergic neurons, and increases in COX-2 levels and microglial activation were observable a few days after the MPP+ exposure(46,48), COX-2 and activated microglia must be significantly involved in causing the secondary damage of the neurons and in the perpetuation of inflammation. Injured nerve cells can stimulate microglia, and inhibiting microglia by minocycline stops neurotoxicity by blocking the production micro-glial IL-1β, NADPH-oxidase and iNOS(54). So, the excitation of microglia and the release of their toxins must happen prior to the progressive secondary dopaminergic neurodegeneration. Thus, chronic oxidation and inflammation following the primary insult can lead to the death of neurons with reversible injury(46). Inhibition of microglial activation by COX-2 inhibition or gene disruption can, directly or indirectly, reduce the MPTP-induced degenera- tion of dopaminergic neurons and the develop-ment of motor impairments.
Conclusions
The initiation and progression of PD seems to involve the activation of microglia. Thus, inhibition of microglial activation with a selective COX-2 inhibitor would be an effective therapeutic approach alleviating the progression of PD. Continued exploration of the mecha-nisms of action underlying the involvement of microglia in neurodegeneration and the inhi-bitory effect of a selective COX-2 inhibitor on microglial activation is highly justified.
Acknowledgements
The author thanks all people who made the collection of review articles celebrating the

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
70
fiftieth anniversary of the Faculty of Science, Mahidol University.
References 1. Arimoto T, Bing G. Up-regulation of induci-ble
nitric oxide synthase in the substantia nigra by lipopolysaccharide causes microglial activation and neurodegeneration. Neurobiol Dis 2003; 12:35-45.
2. Banati RB, Gehrmann J, Schubert P, Kreutz-berg GW. Cytotoxicity of microglia. Glia 1993; 7:111-8.
3. Banati RB, Daniel SE, Blunt SB. Glial pathology but absence of apoptotic nigral neurons in long-standing Parkinson's disease. Mov Disord 1998;13:221-7.
4. Barcia C, Sanchez BA, Fernandez-Villalba E, Bautista V, Poza MPY, Fernandez-Barreiro A et al. Evidence of active microglia in substantia nigra pars compacta of parkinsonian monkeys 1 year after MPTP exposure. Glia 2004;46:402-9.
5. Bojinov S. Encephalitis with acute Parkin-sonian syndrome and bilateral inflammatory necrosis of the substantia nigra. J Neurol Sci 1971;12:383-415.
6. Boka G, Anglade P, Wallach D, Javoy-Agid F, Agid Y, Hirsch EC. Immunocytochemical ana-lysis of tumor necrosis factor and its receptors in Parkinson's disease. Neurosci Lett 1994;172: 151-4.
7. Cross AK, Woodroofe MN. Immunoregu- lation of microglial functional properties. Microsc Res Tech 2001;54:10-7.
8. Czlonkowska A, Kurkowska-Jastrzebska I, Czlonkowski A, Peter D, Stefano GB. Immune processes in the pathogenesis of Parkinson's disease - a potential role for microglia and nitric oxide. Med Sci Monit 2002;8:RA165-77.
9. Furukawa Y, Nishi K, Kondo T, Tanabe K, Mizuno Y. Significance of CSF total neopterin and biopterin in inflammatory neurological diseases. J Neurol Sci 1992;111 65-72.
10. Gao HM, Jiang J, Wilson B, Zhang W, Hong JS, Liu B. Microglial activation-mediated delayed and progressive degeneration of rat nigral dopaminergic neurons: relevance to Parkinson's disease. J Neurochem 2002;81:1285-97.
11. Gebicke-Haerter PJ. Microglia in neurode-generation: molecular aspects. Microsc Res Tech 2001;54:47-58.
12. Gehrmann J, Matsumoto Y, Kreutzberg GW. Microglia: intrinsic immuneffector cell of the brain. Brain Res Brain Res Rev 1995;20: 269-87.
13. Giasson BI, Ischiropoulos H, Lee VM, Trojanowski JQ. The relationship between oxidative/nitrative stress and pathological inclusions in Alzheimer's and Parkinson's diseases. Free Radic Biol Med 2002;32:1264-75.
14. Hartmann A, Hunot S, Hirsch EC. Inflam-mation and dopaminergic neuronal loss in Parkinson's disease: a complex matter. Exp Neurol 2003;184:561-4.
15. Hastings TG. Enzymatic oxidation of dopa-mine: the role of prostaglandin H synthase. J Neurochem 1995;64:919-24.
16. Hopkins SJ, Rothwell NJ. Cytokines and the nervous system. I: Expression and recognition. Trends Neurosci 1995;18:83-8.
17. Hunot S, Boissiere F, Faucheux B, Brugg B, Mouatt-Prigent A, Agid Y et al. Nitric oxide synthase and neuronal vulnerability in Parkin-son's disease. Neuroscience 1996;72:355-63.
18. Hunot S, Brugg B, Ricard D, Michel PP, Muriel MP, Ruberg M et al. Nuclear translocation of NF-kappaB is increased in dopaminergic neurons of patients with Parkinson's disease. Proc Natl Acad Sci USA 1997;94:7531-6.
19. Hunot S, Dugas N, Faucheux B, Hartmann A, Tardieu M, Debre P et al. Fcepsilon-RII/CD23 is expressed in Parkinson's disease and induces, in vitro, production of nitric oxide and tumor necrosis factor-alpha in glial cells. J Neurosci 1999;19:3440-7.
20. Isacson O, Brundin P, Gage FH, Bjorklund A. Neural grafting in a rat model of Huntington's disease: progressive neuro-chemical changes after neostriatal ibotenate lesions and striatal tissue grafting. Neuroscience 1985;16:799-817.
21. Kim WG, Mohney RP, Wilson B, Jeohn GH, Liu B, Hong JS. Regional difference in susceptibility to lipopolysaccharide-induced neurotoxicity in the rat brain: role of microglia. J Neurosci 2000;20:6309-16.
22. Knott C, Stern G, Wilkin GP. Inflammatory regulators in Parkinson's disease: iNOS, lipocortin-1, and cyclooxygenases-1 and -2. Mol Cell Neurosci 2000;16:724-39.
23. Koutsilieri E, Scheller C, Tribl F, Riederer P. Degeneration of neuronal cells due to oxidative stress-microglial contribution. Parkinsonism Relat Disord 2002;8:401-6.
24. Kujubu DA, Fletcher BS, Varnum BC, Lim RW, Herschman HR. TIS10, a phorbol ester tumor promoter-inducible mRNA from Swiss 3T3 cells, encodes a novel prostaglandin synthase/ cyclooxygenase homologue. J Biol Chem 1991; 266:12866-72.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
71
25. Langston JW, Forno LS, Tetrud J, Reeves AG, Kaplan JA, Karluk D. Evidence of active nerve cell degeneration in the substantia nigra of humans years after 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine exposure. Ann Neurol 1999; 46:598-605.
26. Lawson LJ, Perry VH, Dri P, Gordon S. Heterogeneity in the distribution and morpho-logy of microglia in the normal adult mouse brain. Neuroscience 1990;39:151-70.
27. Liu B, Hong JS. Role of microglia in inflam-mation-mediated neurodegenerative diseases: mechanisms and strategies for therapeutic intervention. J Pharmacol Exp Ther 2003;304:1-7.
28. McGeer PL, Itagaki S, Akiyama H, McGeer EG. Rate of cell death in parkinsonism indicates active neuropathological process. Ann Neurol 1988;24:574-6.
29. McGeer PL, Itagaki S, Boyes BE, McGeer EG. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson's and Alzhei-mer's disease brains. Neurology 1988;38:1285-91.
30. McGeer PL, Schwab C, Parent A, Doudet D. Presence of reactive microglia in monkey substantia nigra years after 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine administration. Ann Neurol 2003;54:599-604.
31. Minghetti L, Nicolini A, Polazzi E, Creminon C, Maclouf J, Levi G. Inducible nitric oxide synthase expression in activated rat microglial cultures is downregulated by exogenous prosta-glandin E2 and by cyclo-oxygenase inhibitors. Glia 1997;19:152-60.
32. Minghetti L, Levi G. Microglia as effector cells in brain damage and repair: focus on prostanoids and nitric oxide. Prog Neurobiol 1998;54:99-125.
33. Mirza B, Hadberg H, Thomsen P, Moos T. The absence of reactive astrocytosis is indicative of a unique inflammatory process in Parkinson's disease. Neuroscience 2000; 95:425-32.
34. Mogi M, Harada M, Kondo T, Riederer P, Nagatsu T. Interleukin-1 beta, interleukin-6, epidermal growth factor and transforming growth factor-alpha are elevated in the brain from parkinsonian patients. Neurosci Lett 1994; 180:147-50.
35. Mogi M, Harada M, Riederer P, Narabayashi H, Fujita K, Nagatsu T. Tumor necrosis factor-alpha (TNF-alpha) increases both in the brain and in the cerebrospinal fluid from parkinsonian patients. Neurosci Lett 1994; 165:208-10.
36. Nakamura Y. Regulating factors for micro-glial activation. Biol Pharm Bull 2002; 25:945-53.
37. Needleman P, Isakson PC. The discovery and function of COX-2. J Rheumatol 1997; 24 Suppl 49:6-8.
38. Nikolic D, van Breemen RB. DNA oxidation induced by cyclooxygenase-2. Chem Res Toxicol 2001;14:351-4.
39. Perry VH, Bell MD, Brown HC, Matyszak MK. Inflammation in the nervous system. Curr Opin Neurobiol 1995;5:636-41.
40. Samuelsson B. Arachidonic acid metabolism: role in inflammation. Z Rheumatol 1991; 50 Suppl 1: 3-6.
41. Sanchez-Pernaute R, Ferree A, Cooper O, Yu M, Brownell AL, Isacson O. Selective COX-2 inhibition prevents progressive dopamine neuron degeneration in a rat model of Parkinson's disease. J Neuroinflammation 2004;1:6.
42. Schiefer J, Kampe K, Dodt HU, Ziegl-gansberger W, Kreutzberg GW. Microglial motility in the rat facial nucleus following peripheral axotomy. J Neurocytol 1999;28: 439-53.
43. Sedgwick JD, Schwender S, Imrich H, Dorries R, Butcher GW, ter Meulen V. Isolation and direct characterization of resident microglial cells from the normal and inflamed central nervous system. Proc Natl Acad Sci USA 1991;88:7438-42.
44. Stokes AH, Hastings TG, Vrana KE. Cytotoxic and genotoxic potential of dopamine. J Neurosci Res 1999;55:659-65.
45. Sugama S, Cho BP, Degiorgio LA, Shimizu Y, Kim SS, Kim YS et al. Temporal and sequential analysis of microglia in the substantia nigra following medial forebrain bundle axotomy in rat. Neuroscience 2003; 116: 925-33.
46. Sugama S, Yang L, Cho BP, Degiorgro L, Lorenzl S, Albers D et al. Age-related microglial activation in 1-methyl-4-phenyl-1,2,3,6-tetra-hydropyridine (MPTP)-induced dopaminergic neurodegeneration in C57BL/6 mice. Brain Res 2003;964:288-94.
47. Takeuchi H, Mizuno T, Zhang G, Wang J, Kawanokuchi J, Kuno R et al. Neuritic beading induced by activated microglia is an early feature of neuronal dysfunction toward neuronal death by inhibition of mitochondrial respiration and axonal transport. J Biol Chem 2005;280: 10444-54.
48. Teismann P, Tieu K, Choi D-K, Wu D-C, Naini A, Hunot S et al. Cyclooxygenase-2 is instrumental in Parkinson's disease neuro-

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
72
degeneration. Proc Natl Acad Sci USA 2003; 100:5473-8.
49. Vijitruth R, Liu M, Choi DY, Nguyen XV, Hunter RL, Bing G. Cyclooxygenase-2 mediates microglial activation and secondary dopa-minergic cell death in the mouse MPTP model of Parkinson's disease. J Neuroinflammation 2006;3:6.
50. Vila M, Jackson-Lewis V, Guegan C, Teismann P, Wu DC, Choi DG et al. The role of glial cells in Parkinson's disease. Curr Opin Neurol 2001; 14:483-9.
51. Volpe BT, Wildmann J, Altar CA. Brain-derived neurotrophic factor prevents the loss of nigral neurons induced by excitotoxic striatal-pallidal lesions. Neuroscience 1998; 83:741-8.
52. Wang MJ, Lin WW, Chen HL, Chang YH, Ou HC, Kuo JS et al. Silymarin protects dopa-minergic neurons against lipopolysac charide-induced neurotoxicity by inhibiting microglia activation. Eur J Neurosci 2002;16: 2103-12.
53. Wenzel SE. Arachidonic acid metabolites: mediators of inflammation in asthma. Pharma-cotherapy 1997;17:3S-12S.
54. Wu DC, Jackson-Lewis V, Vila M, Tieu K, Teismann P, Vadseth C et al. Blockade of microglial activation is neuro-protective in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson disease. J Neurosci 2002;22: 1763-71.

73
Sex Hormones and the Activation of Cardiac Machinery
Tepmanas Bupha-Intr and Jonggonnee Wattanapermpool Department of Physiology, Faculty of Science, Mahidol University.
Abstract
Cardiovascular diseases top the list of causes of mortality in Western and developing countries including Thailand (www.trcdreducerisk.com). Many risk factors clearly correlate with the disease incidence with the factor of gender receiving the highest attention in most studies. The significantly lower prevalence of cardiovascular disease in young women compared to that in age-matched men raises the issue of differential effects of sex hormones on cardiovascular function. The theme of this review is therefore focused on a series of investigations from our laboratory exploring the hypothesis that sex hormones directly regulate the myocardial physiology. Following deprivation of ovarian sex hormones, we have identified changes in cardiac contractile activation that lead to cardiac failure. On the other hand, male sex hormones possibly exert a biphasic influence on myocardial functions.
Keywords: estrogen; testosterone; myofilament activation; calcium handling; extracellular matrix; cardiac hypertrophy
Introduction
The direct effect of female sex hormones on the heart is undoubtedly supported by the presence of both estrogen and progesterone receptors in cardiac myocytes and fibro-blasts(17,20). Dysfunctions of both systolic and diastolic activities of the heart have been reported in female sex hormone-deprived patients(12,35,36,42). As compared to age-matched premenopausal women, cardiac systolic dys-function, indicated by a greater left ventricular wall thickness with a lower mid-wall fractional shortening(42), and diastolic dysfunction, sug- gested by a lower left ventricular filling, were detected in menopausal women(12). The bene-ficial role of female sex hormones in cardiac systolic and diastolic functions was further confirmed by increases in the ejection fraction and the filling of blood in the ventricle after hormone replacement therapy(1,35). Also, the presence of estrogen receptors in male cardiac myocytes strongly supports a direct role of female sex hormones in cardiac activation(34).
It is still arguable whether testosterone exerts a beneficial or adverse effect on the heart. The presence of androgen receptors in cardiac
myocytes imply a direct effect of male sex hormones on contractile activities(31,39). A high incidence of cardiac sudden death in men receiving androgen therapy or in men taking high doses of testosterone has been docu-mented(11,30). A recent clinical study has also demonstrated a close correlation between varia-tions in androgen receptor genes and the incidence of hypertrophic cardiomyopathy(28). These human studies thus point to the poten-tially harmful effects of testosterone on cardiac performance. By contrast, studies in castrated animals found suppressed contractile function with reductions in many subcellular activi- ties(14,15,40), which could be restored by testo-sterone supplementation. Based on this body of evidence, testosterone appears to have a role in the maintenance of normal cardiac activities. It is therefore possible that testosterone might have a biphasic influence on the heart.
In this review, the impact of both female and male sex hormones on various aspects of the heart, including contractile machinery activation, intracellular Ca2+ handling, and extracellular matrix components, is examined. The protective effect of estrogen and the

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
74
adverse effect of overdoses of testosterone are also discussed.
Estrogen and the Cardiac Myofilament Activation
The role of estrogen in maintaining the systolic function of the heart has been the focus of cardiovascular research for decades. Beneficial effects of estrogen on cardiac contractile performance have been demonstrated in clinical studies in postmenopausal women as well as in an ovariectomized (OVX) animal model(40-42). It was firstly reported by Schiable et al.(40) and Scheuer et al.(41) that estrogen prevented reductions in the stroke volume and the ejection fraction of hearts isolated from OVX rats. The preserved systolic function in OVX rat hearts by estrogen was found to correlate with the myosin Ca2+-ATPase activity(40,41). As summarized in Figure 1, a series of experiments have been performed by our group exploring the role of estrogen in cardiac contractile activation. A decrease in maximum Ca2+-dependent actomyosin Mg-ATPase activity combined with an increase in the Ca2+-responsiveness of the myofilaments were clearly shown in OVX rat hearts and these changes could be reversed by estrogen supple- mentation (Figure 1, No.1)(52,54). Strikingly, the increased myofilament Ca2+-sensitivity detected in the hearts of OVX rats was similar to changes reported in both human and animal cardio-myopathic hearts(49,55). Interestingly, mechanical measurements of the contractile machinery demonstrated a specific induction of Ca2+ hypersensitivity in heart but not skeletal muscle after ovariectomy(53). These similar results strongly suggest an increased risk of developing heart disease after estrogen deprivation.
Many underlying mechanisms have been hypothesized to explain the estrogen effect on cardiac function. The significant effect of estrogen on cardiac myofilament activation has been one an active area of investigation. Although possible changes in the myofibrillar regulatory proteins were implied from the Ca2+ hypersensitive response, so far no evidence of changes in the level of myofibrillar protein in
response to the presence or absence of estrogen has been demonstrated. However, qualitative changes like isoform shifts and phosphorylation of myofilament proteins might be occuring. Similar to that observed in the failing hearts, an isoform shift of the cardiac myosin heavy chain (MHC) from a fast α-isoform to a slow β-isoform ATPase activity has been clearly shown in OVX rats(53). This isoform shift of MHC supports the regulatory effect of estrogen on maximum myofilament ATPase activity. Using the combined conditions of estrogen and insulin deprivation, we further found that estrogen not only affects MHC expression but also has a significant impact on the interaction among myofilament proteins(46).
The possibility that estrogen may alter phosphorylation levels of many cardiac contractile proteins was indirectly indicated from the upregulation of β-adrenoceptors in OVX rats, which returned to normal levels following estrogen supplementation (Figure 1, No.5)(47). Alterations of the phosphorylation status either by protein kinase A (PKA) or protein kinase C (PKC) can affect cardiac myofilament activities under physiological conditions and in many heart diseases(48,49). Phosphorylation of troponin I by PKA and PKC results in a decreased sensitivity of troponin C to Ca2+(44). While the effects of PKC phospho-rylation on troponin T (TnT) vary depending on the site of TnT phosphorylation and the type of PKC isoforms(45), an increased Ca2+ sensitivity at submaximal levels of cardiac contraction was detected after myosin light chain (MLC) phosphorylation by PKC(16). There is so far no direct evidence of an effect of estrogen on PKA activation but the upregulation of β-adreno-ceptors in OVX rat hearts has been reported(47). There is also indirect evidence showing the inhibition of PKCε translocation from the cytosol to the nucleus after ovariectomy(19) that might be associated with increased MLC phosphorylation, and therefore, modification of the myofilament Ca2+ sensitivity(16,48). Taken together, it is highly possible that estrogen plays a positive role in the regulation of a number of

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
75
Figure 1 Impacts of sex hormones on cardiac myofilament Ca2+ activation, intracellular Ca2+ handling, and
extracellular matrix components. Ca2+-induced Ca2+ release (CICR) from the sarcoplasmic reticulum (SR) activates myofilament contraction. Ca2+ is then pumped into the SR and extruded by a Na+-Ca2+ exchanger (NCX) leading to myofilament relaxation. The activation can be modulated by β1-adrenoceptors but stabilized by heat shock protein (HSP). Effects of estrogen and testosterone on myofilament Ca2+ activation, intracellular Ca2+ handling, and extracellular matrix are summarized in the boxes. and represent with and without regulatory effect, respectively. ? represents no or controversial information.
Effects of Androgen
1. Myofilament Ca2+ activation: Cross-bridge kinetic Ca2+ sensitivity
2. CICR (Golden et al., 2002) L-type Ca2+ channel expression
? RyR-CRC expression 3. SR Ca2+ uptake:
? SERCA activity & expression ? Phospholamban phosphorylation
4. Ca2+ extrusion: ? Na+-H+ exchange activation
? NCX protein expression 5. Adrenergic stimulation:
β1-Adrenergic receptor expression β1-Adrenoceptor sensitivity 6. Protective proteins:
? Heat shock protein 70 expression 7. Extracellular matrix:
? Collagen production
Effects of Estrogen
1. Myofilament Ca2+ activation: Cross-bridge kinetic Ca2+ sensitivity
2. CICR (Chu et al., 2006) L-type Ca2+ channel expression RyR-CRC expression
3. SR Ca2+ uptake: SERCA activity & expression Phospholamban phosphorylation 4. Ca2+ extrusion:
Na+-H+ exchange activation ? NCX protein expression 5. Adrenergic stimulation:
β1-Adrenergic receptor expression β1-Adrenoceptor sensitivity
6. Protective proteins: Heat shock protein 70 expression
7. Extracellular matrix: ? Collagen production

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
76
proteins on the contractile machinery of the heart.
Estrogen and the Intracellular Ca2+ Handling
The role of estrogen in the regulation of cardiac contractility involves mobilization of intracellular Ca2+. It is the amount of Ca2+ binding to troponin C on the thin filament that determines the number of cross-bridge inter-actions underlying cardiac contractility. An effect of estrogen on contractility is indicated by the observation that a reduction in the amplitude of the intracellular Ca2+ transients with a prolonged Ca2+ decay time was detected in the hearts of the 8-week and 10-week OVX rats by our group and others(7,37). By contrast, one recent study in 6-week OVX rats(24) reported an increased amplitude with a faster decay of cardiac Ca2+ transients. The time-dependent responses from these controversial data suggest both direct and indirect regulatory effects of estrogen on the cardiac Ca2+ handling activity.
Estrogen may regulate the Ca2+ handling activity by acting on only one or both of the two processes of intracellular Ca2+ mobilization, Ca2+-induced Ca2+ release (CICR) and cytosolic Ca2+ removal (Figure 1, No. 2 and 3). Although the evidence is not yet conclusive, estrogen seems to be involved in both regulatory processes of the Ca2+ handling. A direct regulatory effect of estrogen on CICR has been shown by an increased expression of L-type Ca2+ channels in the hearts of estrogen receptor knockout mice and OVX rats(9,22). Increases in the activities of both L-type Ca2+ channels and ryanodine receptor Ca2+ release channels were also detected in 6-week OVX rat hearts(23,24). The data suggest an increase in the amplitude of Ca2+ transients during the early phase of estrogen deprivation. By contrast, there is so far no report concerning the effect of long-term estrogen deprivation on the CICR process.
Estrogen also regulates the handling of intracellular Ca2+ by affecting cytosolic Ca2+ removal. Reuptake of Ca2+ by the sarcoplasmic reticulum (SR) Ca2+-ATPase (SERCA) pump and extrusion of Ca2+ by Na+-Ca2+ exchanger (NCX) result in muscle relaxation and therefore
diastole of the ventricles(3). Suppressed SERCA expression is considered a standard marker to indicate a pathological state of the heart(33,50). While there was no change in SERCA expression in 4- and 6-week OVX rat hearts(9,24), significant reductions in both protein and mRNA levels of SERCA were clearly detected in 10-week OVX rats with decreases in maximum SR Ca2+ uptake and SERCA activities(6). In as much as SERCA levels are directly correlated with the amount of heat shock protein 72 (HSP72)(29), the reduced SERCA in OVX rats might have resulted from decreased HSP72 as detected in our study (Figure 1, No.6)(5). The reduced quantity and quality of SERCA in 10-week OVX rat hearts are also similar to that found in failing hearts and thus supports a possible risk of pathological induction after chronic deprivation of estrogen.
Results regarding estrogen effects on NCX activity are rather controversial (Figure 1, No.4). While the mRNA expression of NCX was decreased in 4-week OVX rats(9), there was no change in the protein level. In contrast, a three-fold increase in NCX activity was reported in 6-week OVX rats(24), which might underlie the faster Ca2+ decay observed. However, NCX upregulation is expected in most heart diseases in conjunction with a suppression of SERCA in order to compensate for the Ca2+ overloading of the heart(18,33). It is presently unknown how NCX activity and expression are altered after chronic deprivation of estrogen. However, our laboratory has shown an increased Na+-H+ exchange (NHE1) activity in 10-week OVX rat hearts(7) which is probably associated with intracellular Na+ accumulation, and subsequent activation of the reversed-mode NCX activity. As such, estrogen may indirectly regulate NCX activity via modification of NHE1 and thus slow the process of Ca2+ extrusion, resulting in a prolonged Ca2+ transient decay.
Estrogen and the Cardiac Extracellular Matrix
Estrogen has significant effects on the extracellular matrix components of many tissues and may be able to prevent cardiac fibrosis (Figure 1, No.7)(1,35). Physiologically, the extra-

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
77
cellular matrix is a significant contributor to cardiac diastolic function. Decreased ventricular blood filling appears when collagen is over-expressed(25). Given the presence of estrogen receptors in cardiac fibroblasts(17), there is no doubt that estrogen can exert a direct impact on the synthesis of the extracellular matrix. In in vitro studies, estrogen inhibits the growth of cardiac fibroblasts and therefore decreases collagen synthesis in a dose-dependent manner(56). The data raise the possibility of a correlation between the effect of estrogen deficiency and collagen-induced myocardial stiffness, and therefore ventricular filling. While no change in cardiac collagen was reported in 6-week OVX rats(26,32), myocardial infarction-induced collagen deposition was significantly greater in OVX mice compared to sham(8,43). Despite clear evidence of some estrogen dependent effect on collagen synthesis, it seems to exert a significant effect only under pathological insults. The hypothesis that this estrogen effect is pathology-dependent is currently under investigation by our group.
Testosterone Deficiency and Cardiac Contractile Activation
Testosterone has also been found to play a significant role in the maintenance of cardiac function. Castration in either pre- or post-pubertal male rats is associated with reduced ventricular functions, including decreases in cardiac output, peak systolic pressure and ejection fraction. The depressed cardiac functions are well correlated with a reduced myosin ATPase activity and a shift in myosin isoenzymes from a predominant V1 to V3
form(40,41). Suppression of the amplitude of the intracellular Ca2+ transient along with a prolonged Ca2+ decay was also observed after castration(13,15) in synchronization with the down-regulations of the L-type Ca2+ channel and NCX. These changes somewhat resemble the characteristics of heart failure. All of the cardiac changes in castrated rats disappear after testosterone supplementation, thus validating the crucial role of testosterone in contractile performance (Figure 1). As mentioned earlier, androgen receptors are expressed in both male
and female hearts. The use of testosterone derivatives in females has also been found to increase the risk of heart failure. Sudden death with cardiac ischemia has been reported in transsexual women receiving testosterone supplementation(21). Diastolic dysfunction of the heart was reported in women with polycystic ovaries who have marked increases in the plasma testosterone level(51). Taken together, these lines of evidence suggest both a significant positive impact as well as some conditionally adverse effects of testosterone on cardiac contractile activity.
Testosterone and Cardiac Hypertrophy
Consumption of testosterone to increase the strength of skeletal muscle is prohibited in sport competition and in body building due mainly to its detrimental effects. Induction of muscle hypertrophy in both skeletal and cardiac tissues is a well recognized effect of testo-sterone. It is the anabolic steroid action of testosterone that directly mediates cardiac hypertrophy through androgen receptors(4,31). It is presently unclear whether the cardiac hypertrophy effect of testosterone is physiologic or pathologic.
The impacts of testosterone on the heart are mostly accomplished by pathological adaptation. Chronic treatment of rats with androgenic steroids induced cardiac hyper-trophy together with cardiac fibrosis(38). In transgenic cardio-myopathic mice, the hyper-trophy and fibrosis of the heart can be reduced by androgen inhibition or castration and enhanced by high doses of testosterone(27). The involvement of pathological signals, such as MAPKs, has been suggested in many studies to potentially be the cause of testosterone-induced cardiac hypertrophy. While injection of testosterone, before ischemic-reperfusion, in castrated male rats induced further increases in the phosphorylation of p38-MAPK and JNK in the heart(10), an androgen receptor antagonist suppressed apoptotic activity and reduced p38-MAPK phosphorylation. Moreover, acute testosterone exposure in guinea pig cardio-myocytes causes an increased phosphorylation of Akt, a well-accepted physiologic hypertrophy

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
78
signal(2). While this evidence clearly points to the involvement of physiologic signals in testosterone-induced cardiac hypertrophy, their exact role has not been well defined.
Conclusions
Sex hormones directly regulate various aspects of cardiac contraction/relaxation, from activation of the contractile machinery to the regulation of intracellular Ca2+ and the compo-nents of the extracellular matrix. Many mecha-nistic roles, as assessed by our group, underlie the positive regulatory impact of estrogen on cardiac contraction. The significance of estrogen in cardiac relaxation is also being investigated in our laboratory. In addition, the mechanism of action of testosterone on the induction of cardiac hypertrophy is being evaluated in our laboratory.
Acknowledgements We thank Mahidol University and the
Thailand Research Fund for supporting grant.
References 1. Alecrin IN, Aldrighi JM, Caldas MA, Gebara
OC, Lopes NH, Ramires JA. Acute and chronic effects of oestradiol on left ventricular diastolic function in hypertensive postmenopausal women with left ventricular diastolic dysfunction. Heart 2004;90:777-81.
2. Bai CX, Kurokawa J, Tamagawa M, Nakaya H, Furukawa T. Nontranscriptional regulation of cardiac repolarization currents by testosterone. Circulation 2005;112:1701-10.
3. Bers DM. Cardiac excitation-contraction coup-ling. Nature 2002;415:198-205.
4. Beutel A, Bergamaschi CT, Campos RR. Effects of chronic anabolic steroid treatment on tonic and reflex cardiovascular control in male rats. J Steroid Biochem Mol Biol 2005;93:43-8.
5. Bupha-Intr T, Wattanapermpool J. Cardioprotec- tive effects of exercise training on myofilament calcium activation in ovariectomized rats. J Appl Physiol 2004;96:1755-60.
6. Bupha-Intr T, Wattanapermpool J. Regulatory role of ovarian sex hormones in calcium uptake activity of cardiac sarcoplasmic reticulum. Am J Physiol Heart Circ Physiol 2006;291:H1101-8.
7. Bupha-Intr T, Wattanapermpool J, Pena JR, Wolska BM, Solaro RJ. Myofilament response to Ca2+ and Na+/H+ exchanger activity in sex hormone-related protection of cardiac myocytes
from deactivation in hypercapnic acidosis. Am J Physiol Regul Integr Comp Physiol 2007;292: R837-43.
8. Cavasin MA, Sankey SS, Yu AL, Menon S, Yang XP. Estrogen and testosterone have opposing effects on chronic cardiac remodeling and function in mice with myocardial infarction. Am J Physiol Heart Circ Physiol 2003;284: H1560-9.
9. Chu SH, Goldspink P, Kowalski J, Beck J, Schwertz DW. Effect of estrogen on calcium-handling proteins, beta-adrenergic receptors, and function in rat heart. Life Sci 2006;79:1257-67.
10. Crisostomo PR, Wang M, Wairiuko GM, Morrell ED, Meldrum DR. Brief exposure to exogenous testosterone increases death signaling and adversely affects myocardial function after ischemia. Am J Physiol Regul Integr Comp Physiol 2006;290:R1168-74.
11. Fineschi V, Baroldi G, Monciotti F, Paglicci Reattelli L, Turillazzi E. Anabolic steroid abuse and cardiac sudden death: a pathologic study. Arch Pathol Lab Med 2001;125:253-5.
12. Gokce M, Karahan B, Erdol C, Kasap H, Ozdemirci S. Left ventricular diastolic function assessment by tissue Doppler echocardiography in relation to hormonal replacement therapy in postmenopausal women with diastolic dysfunc-tion. Am J Ther 2003;10:104-11.
13. Golden KL, Marsh JD, Jiang Y. Castration reduces mRNA levels for calcium regulatory proteins in rat heart. Endocrine 2002;19:339-44.
14. Golden KL, Marsh JD, Jiang Y. Testosterone regulates mRNA levels of calcium regulatory proteins in cardiac myocytes. Horm Metab Res 2004;36:197-202.
15. Golden KL, Marsh JD, Jiang Y, Brown T, Moulden J. Gonadectomy of adult male rats reduces contractility of isolated cardiac myocytes. Am J Physiol Endocrinol Metab. 2003;285:E449-53.
16. Goldspink PH, Montgomery DE, Walker LA, Urboniene D, McKinney RD, Geenen DL et al. Protein kinase Cepsilon overexpression alters myofilament properties and composition during the progression of heart failure. Circ Res 2004; 95:424-32.
17. Grohe C, Kahlert S, Lobbert K, Stimpel M, Karas RH, Vetter H et al. Cardiac myocytes and fibroblasts contain functional estrogen receptors. FEBS Lett 1997;416:107-12.
18. Houser SR, Piacentino V, 3rd, Mattiello J, Weisser J, Gaughan JP. Functional properties of failing human ventricular myocytes. Trends Cardiovasc Med. 2000;10:101-7.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
79
19. Hunter JC, Kostyak JC, Novotny JL, Simpson AM, Korzick DH. Estrogen deficiency decreases ischemic tolerance in the aged rat heart: Roles of PKCdelta, PKCepsilon, Akt, and GSK3beta. Am J Physiol Regul Integr Comp Physiol 2007; 292:R800-9.
20. Ingegno MD, Money SR, Thelmo W, Greene GL, Davidian M, Jaffe BM et al. Progesterone receptors in the human heart and great vessels. Lab Invest 1988;59:353-6.
21. Inoue H, Nishida N, Ikeda N, Tsuji A, Kudo K, Hanagama M et al. The sudden and unexpected death of a female-to-male transsexual patient. J Forensic Leg Med 2007;14:382-6.
22. Johnson BD, Zheng W, Korach KS, Scheuer T, Catterall WA, Rubanyi GM. Increased expression of the cardiac L-type calcium channel in estrogen receptor-deficient mice. J Gen Physiol 1997;110: 135-40.
23. Kam KW, Kravtsov GM, Liu J, Wong TM. Increased PKA activity and its influence on isoprenaline-stimulated L-type Ca2+ channels in the heart from ovariectomized rats. Br J Pharmacol 2005;144:972-81.
24. Kravtsov GM, Kam KW, Liu J, Wu S, Wong TM. Altered Ca(2+) handling by ryanodine receptor and Na(+)-Ca(2+) exchange in the heart from ovariectomized rats: role of protein kinase A. Am J Physiol Cell Physiol 2007;292:C1625-35.
25. Kuwahara F, Kai H, Tokuda K, Takeya M, Takeshita A, Egashira K et al. Hypertensive myocardial fibrosis and diastolic dysfunction: another model of inflammation? Hypertension 2004;43:739-45.
26. Lekgabe ED, Royce SG, Hewitson TD, Tang ML, Zhao C, Moore XL et al. The effects of relaxin and estrogen deficiency on collagen deposition and hypertrophy of nonreproductive organs. Endocrinology 2006;147:5575-83.
27. Li Y, Kishimoto I, Saito Y, Harada M, Kuwahara K, Izumi T et al. Androgen contributes to gender-related cardiac hyper-trophy and fibrosis in mice lacking the gene encoding guanylyl cyclase-A. Endocrinology 2004;145:951-8.
28. Lind JM, Chiu C, Ingles J, Yeates L, Humphries SE, Heather AK et al. Sex hormone receptor gene variation associated with phenotype in male hypertrophic cardiomyopathy patients. J Mol Cell Cardiol 2008;45:217-22.
29. Liu J, Kam KW, Borchert GH, Kravtsov GM, Ballard HJ, Wong TM. Further study on the role of HSP70 on Ca2+ homeostasis in rat ventricular
myocytes subjected to simulated ischemia. Am J Physiol Cell Physiol 2006;290:C583-91.
30. Luke JL, Farb A, Virmani R, Sample RH. Sudden cardiac death during exercise in a weight lifter using anabolic androgenic steroids: pathological and toxicological findings. J Forensic Sci 1990; 35:1441-7.
31. Marsh JD, Lehmann MH, Ritchie RH, Gwathmey JK, Green GE, Schiebinger RJ. Androgen receptors mediate hypertrophy in cardiac myocytes. Circulation 1998;98:256-61.
32. Mercier I, Pham-Dang M, Clement R, Gosselin H, Colombo F, Rouleau JL et al. Elevated mean arterial pressure in the ovariectomized rat was normalized by ET(A) receptor antagonist therapy: absence of cardiac hypertrophy and fibrosis. Br J Pharmacol 2002;136:685-92.
33. Mishra S, Sabbah HN, Rastogi S, Imai M, Gupta RC. Reduced sarcoplasmic reticulum Ca2+ uptake and increased Na+-Ca2+ exchanger expression in left ventricle myocardium of dogs with pro-gression of heart failure. Heart Vessels 2005;20: 23-32.
34. Nordmeyer J, Eder S, Mahmoodzadeh S, Martus P, Fielitz J, Bass J et al. Upregulation of myocardial estrogen receptors in human aortic stenosis. Circulation 2004;110:3270-5.
35. Ozdemir K, Celik C, Altunkeser BB, Icli A, Albeni H, Duzenli A et al. Effect of postmeno-pausal hormone replacement therapy on cardio-vascular performance. Maturitas 2004;47:107-13.
36. Palmiero P, Maiello M, Passantino A, Antoncecchi E, Deveredicis C, DeFinis A et al. Correlation between diastolic impairment and lipid metabolism in mild-to-moderate hyper-tensive postmenopausal women. Am J Hypertens 2002;15:615-20.
37. Ren J, Hintz KK, Roughead ZK, Duan J, Colligan PB, Ren BH et al. Impact of estrogen replacement on ventricular myocyte contractile function and protein kinase B/Akt activation. Am J Physiol Heart Circ Physiol 2003;284:H1800-7.
38. Rocha FL, Carmo EC, Roque FR, Hashimoto NY, Rossoni LV, Frimm C et al. Anabolic steroids induce cardiac renin-angiotensin system and impair the beneficial effects of aerobic training in rats. Am J Physiol Heart Circ Physiol 2007;293:H3575-83.
39. Ruizeveld de Winter JA, Trapman J, Vermey M, Mulder E, Zegers ND, van der Kwast TH. Androgen receptor expression in human tissues: an immunohistochemical study. J Histochem Cytochem 1991;39:927-36.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
80
40. Schaible TF, Malhotra A, Ciambrone G, Scheuer J. The effects of gonadectomy on left ventricular function and cardiac contractile proteins in male and female rats. Circ Res 1984;54:38-49.
41. Scheuer J, Malhotra A, Schaible TF, Capasso J. Effects of gonadectomy and hormonal replace-ment on rat hearts. Circ Res 1987;61:12-9.
42. Schillaci G, Verdecchia P, Borgioni C, Ciucci A, Porcellati C. Early cardiac changes after meno-pause. Hypertension. 1998;32:764-9.
43. Smith PJ, Ornatsky O, Stewart DJ, Picard P, Dawood F, Wen WH et al. Effects of estrogen replacement on infarct size, cardiac remodeling, and the endothelin system after myocardial infarction in ovariectomized rats. Circulation 2000;102:2983-9.
44. Sumandea MP, Burkart EM, Kobayashi T, De Tombe PP, Solaro RJ. Molecular and integrated biology of thin filament protein phosphorylation in heart muscle. Ann NY Acad Sci 2004; 1015:39-52.
45. Sumandea MP, Pyle WG, Kobayashi T, de Tombe PP, Solaro RJ. Identification of a functionally critical protein kinase C phospho-rylation residue of cardiac troponin T. J Biol Chem 2003;278:35135-44.
46. Thawornkaiwong A, Pantharanontaga J, Wattana-permpool J. Hypersensitivity of myo-filament response to Ca2+ in association with mala-daptation of estrogen-deficient heart under diabetes complication. Am J Physiol Regul Integr Comp Physiol 2007;292:R844-51.
47. Thawornkaiwong A, Preawnim S, Wattana-permpool J. Upregulation of beta 1-adrenergic receptors in ovariectomized rat hearts. Life Sci 2003;72:1813-24.
48. van der Velden J, Papp Z, Boontje NM, Zaremba R, de Jong JW, Janssen PM et al. The effect of myosin light chain 2 dephosphorylation on Ca2+-sensitivity of force is enhanced in failing human hearts. Cardiovasc Res 2003; 57:505-14.
49. van der Velden J, Papp Z, Zaremba R, Boontje NM, de Jong JW, Owen VJ et al. Increased Ca2+-sensitivity of the contractile apparatus in end-stage human heart failure results from altered phosphorylation of contractile proteins. Cardiovasc Res 2003;57:37-47.
50. Vanderheyden M, Mullens W, Delrue L, Goethals M, de Bruyne B, Wijns W et al. Myocardial gene expression in heart failure
patients treated with cardiac resynchronization therapy responders versus nonresponders. J Am Coll Cardiol 2008;51:129-36.
51. Vrtovec B, Meden-Vrtovec H, Jensterle M, Radovancevic B. Testosterone-related shorten-ing of QTc interval in women with polycystic ovary syndrome. J Endocrinol Invest 2008;31: 653-5.
52. Wattanapermpool J. Increase in calcium respon-siveness of cardiac myofilament activation in ovariectomized rats. Life Sci 1998;63:955-64.
53. Wattanapermpool J, Reiser PJ. Differential effects of ovariectomy on calcium activation of cardiac and soleus myofilaments. Am J Physiol 1999;277:H467-73.
54. Wattanapermpool J, Riabroy T, Preawnim S. Estrogen supplement prevents the calcium hypersensitivity of cardiac myofilaments in ova-riectomized rats. Life Sci 2000;66:533-43.
55. Wolff MR, Buck SH, Stoker SW, Greaser ML, Mentzer RM. Myofibrillar calcium sensitivity of isometric tension is increased in human dilated cardiomyopathies: role of altered beta-adrener-gically mediated protein phosphorylation. J Clin Invest 1996;98:167-76.
56. Zhou L, Shao Y, Huang Y, Yao T, Lu LM. 17 beta-estradiol inhibits angiotensin II-induced collagen synthesis of cultured rat cardiac fibroblasts via modulating angiotensin II receptors. Eur J Pharmacol 2007;567:186-92.

81
ATP-binding Cassette Transporters in Biomarker Research
Tavan Janvilisri Department of Biology, Faculty of Science, Mahidol University.
Abstract
ATP-binding cassette transporters are ubiquitous transmembrane proteins that bind ATP and use the energy of its hydrolysis to translocate various molecules across biological membranes, a process pivotal for most aspects of cell physiology. Their dysfunction is associated with a number of human disorders as well as with multidrug resistance to antibiotics and cancer therapeutic drugs. In this paper, the function of human ABC transporters is reviewed, highlighting genetic defects that may lead to biomarker applications for human diseases.
Keywords: ATP-binding cassette; ABC transporter; multidrug resistance; biomarker
Introduction
ATP-binding cassette (ABC) transporters belong to a superfamily of ATPases that are integrated in cellular membranes. They usually share a common basic structure formed by two nucleotide-binding domains, which contain conserved sequences involved in ATP binding and hydrolysis, and two transmembrane domains that form the pathway through which substrates cross the membrane and are believed to determine the substrate specificity of the transporters. ABC transporters are ubiquitous in biological systems, and are present in virtually all living organisms ranging from archaea to humans(12,15). These transporters mediate uni-directional transport under physiological conditions, either inwards or outwards, and therefore are either importers (permeases or uptake systems) or exporters (efflux-pumps or extruders), respectively. They are fundamental to the membrane transport of a broad variety of substrates including lipids, amino acids, peptides, proteins, saccharides, lipopolysaccha- rides, inorganic ions, metals, and drugs(15). These proteins not only move a variety of substrates into and out of the cell, but also are involved in intracellular compartmental transport. They therefore play a vital role in numerous cellular functions including the transport of metabolic products, lipids and sterols across cell membranes(6).
Several ABC transporters account for multidrug resistance (MDR) traits in both human cancer cells and pathogenic micro-organisms. Furthermore, mutation(s) and/or deletion(s) in genes encoding ABC transporters in humans are the molecular bases of several diseases involving transport deficiency(6). At present, there are 49 ABC transporters that have been identified in humans, which are divided into seven subfamilies (denoted ABCA to ABCG) based on genomic organization, order of domains and sequence conservation. Table 1 displays a list of all 49 known human ABC transporters indicating their distribution, trans-porting substrates and related diseases or phenotypes. To date, there are many initiatives that aim to accelerate the verification and validation of ABC transporters as biomarkers for early disease diagnosis and as therapeutic targets, particularly of those that participate in MDR. This review provides an overview of the function of ABC proteins in each subfamily with an emphasis on their potential application as biomarkers for certain diseases.
ABCA Subfamily
The human ABCA subfamily comprises 13 transporters, which are found only in multi-cellular eukaryotes. Most of ABCA proteins exert their functions in the control of cellular lipid transport. Loss-of-function mutations in

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
82
Table 1 Human ABC transporters, their expression, transporting substrates and related diseases.
Name Alias Expression Transporting substrate(s) Disease(s)/Phenotype(s)
Subfamily A
ABCA1 ABC1 Ubiquitous Cholesterol Tangier disease, familial hypoapoproteinemia
ABCA2 ABC2 Brain, kidney, lung, heart Steroid derivatives, lipids Alzheimer’s disease
ABCA3 ABC3 Lung Unknown Neonatal surfactant deficiency
ABCA4 ABCR Retina N-retinylidiene-phosphatidylethanolamine
Stargardt’s disease, retinitis pigmentosa, cone-rod dystrophy,
age-related macular degeneration
ABCA5 ABC13 Muscle, heart, testis Unknown -
ABCA6 Ubiquitous Unknown -
ABCA7 ABCX Spleen, thymus Phospholipid Sjögren syndrome
ABCA8 Heart, liver, ovary Unknown -
ABCA9 Heart, brain, fetal tissue Unknown -
ABCA10 Heart, brain, muscle Unknown -
ABCA12 Stomach Lipids Lamellar ichtyosis type 2, Harlequin ichtyosis
ABCA13 Trachea, testes, bone marrow Unknown -
Subfamily B
ABCB1 MDR1, PGP
Many tissues especially with barrier functions: liver, blood-brain barrier, kidney, intestine
Hydrophobic xenobiotics, amphipathic drugs
Drug hypersensitivity, ulcerative colitis
ABCB2 TAP1 Ubiquitous Peptides Immune deficiency
ABCB3 TAP2 Ubiquitous Peptides Immune deficiency
ABCB4 MDR2 Brain, liver Phosphatidylcholine Progressive familial intrahepatic cholestasis type 3
ABCB5 Ubiquitous Unknown -
ABCB6 MTABC3 Mitochondria Iron in heme, porphyrin -
ABCB7 ABC7 Mitochondria Iron-sulfate complexes X-linked sideroblastosis anemia and ataxia
ABCB8 MABC1 Mitochondria Unknown -
ABCB9 TAPL Testes, heart, brain Peptides -
ABCB10 MTABC2 Mitochondria Peptides -
ABCB11 BSEP, SPGP
Liver Bile salts Progressive familial intrahepatic cholestasis type 2
Subfamily C
ABCC1 MRP1 Ubiquitous Leukotriene C4, anionic drug conjugates, GSH, GSSG
Drug hypersensitivity
ABCC2 MRP2 Liver, intestine, kidney, blood-brain barrier, placenta, lung
Bilirubin glucuronides, bile salts, organic anions
Dubin-Johnson syndrome
ABCC3 MRP3 Liver, intestine, kidney, placenta
Sulfate and glucuronide metabolites
-
ABCC4 MRP4 Ovary, testes, kidney, lung Nucleotide analogues -
ABCC5 MRP5 Ubiquitous Nucleotide analogues -

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
83
Name Alias Expression Transporting substrate(s) Disease(s)/Phenotype(s)
ABCC6 MRP6 Kidney, liver Unknown Pseudoxanthoma elasticum
ABCC7 CFTR Exocrine tissues Chloride ion Cystic fibrosis, congenital bilateral absence of
the vas deferens
ABCC8 SUR1 Pancreas Sulfonylurea Familial persistent hyperinsulinemic hypoglycemia
of infancy
ABCC9 SUR2 Heart, muscle Sulfonylurea Dilated cardiomyopathy with ventricular tachycardia
ABCC10 MRP7 Low in all tissues except pancreas
E(2)17betaG -
ABCC11 MRP8 Kidney, spleen, colon, brain E(2)17betaG, steroid sulfate, glutathione conjugates
-
ABCC12 MRP9 Brain, breast, skeletal, ovary Unknown -
ABCC13 PRED6 Fetal liver, bone marrow Unknown -
Subfamily D
ABCD1 ALD Peroxisomes Very long chain fatty acids Adrenoleukodystrophy
ABCD1 ALDR Peroxisomes Very long chain fatty acids -
ABCD1 PMP70 Peroxisomes Very long chain fatty acids Zellweger syndrome 2
ABCD1 PMP69 Peroxisomes Very long chain fatty acids -
Subfamily E
ABCE1 OABP, RNS41
Ubiquitous Oligoadenylate -
Subfamily F
ABCF1 ABC50 Ubiquitous Unknown -
ABCF2 Ubiquitous Unknown -
ABCF3 Ubiquitous Unknown -
Subfamily G
ABCG1 ABC8, white
Ubiquitous Cholesterol, phospholipid -
ABCG2 BCRP, MXR
Placenta, intestine, liver, breast, stem cells, blood-brain barrier
Sterols, steroids, polyphenols, porphyrins, amphipathic drugs
Drug hypersensitivity, protoporphyria IX
ABCG4 White2 Brain, thymus, lung, eye Cholesterol -
ABCG5 Sterolin1 Liver, intestine Sterols Sitosterolemia
ABCG8 Sterolin2 Liver, intestine Sterols Sitosterolemia
the ABCA proteins reveal their pivotal roles in specialized lipid secretory pathways of the cell including high-density lipoprotein (HDL) bio-synthesis (ABCA1), lung surfactant production (ABCA3), retinal integrity (ABCA4) and skin lipid barrier formation (ABCA12). ABCA1 mediates the transport of cholesterol and phospholipids to apolipoprotein acceptors in the
bloodstream(36), a crucial step for HDL forma-tion. ABCA1 mutations can cause Tangier disease, a rare inherited disorder of severe HDL-deficiency characterized by cholesterol deposition in tissue macrophages and prevalent atherosclerosis. Several studies have also shown that ABCA1 polymorphisms are associated with an increased risk of coronary heart disease, metabolic syndrome and type-2

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
84
diabetes(36). Another well-studied ABCA protein, ABCA4, transports retinoid-lipid com-plexes out of photo-receptor cells. Mutations in the ABCA4 gene can lead to an autosomal recessive form of juvenile macular degeneration known as Stargardt’s disease(2). ABCA4 dys-function also causes cone-rod dystrophy, retinitis pigmentosa, and age-related macular degeneration(3).
ABCB Subfamily
The ABCB subfamily comprises both full and half transporters. ABCB1, the first cloned and characterized human ABC transporter, mediates the efflux of a broad range of xenobiotic compounds. ABCB1 over-expression is responsible for decreased drug accumulation and often mediates a multidrug resistance phenotype in cancer cells. In normal tissues, ABCB1 is found in the apical border of many secretory cell types such as those in the kidney, gastrointestinal tract, and liver, suggesting its role in drug uptake, distribution, and elimination at these sites. Thus, ABCB1 often mediates the bioavailability of, and the level and duration of exposure to, its substrate drugs(21). ABCB1 also plays a protective role in biological barriers such as the blood-brain barrier, the blood-testes barrier, and the placenta(21). Several polymor- phisms have been identified in both the ABCB1 gene as well as its promoter. They alter the gene’s transcriptional level which may lead to different drug susceptibility in each indivi-dual(31). The heterodimers of the half-transporters ABCB2 and ABCB3 transport peptides across the endoplasmic reticulum into the membrane-bound compartment for presenta-tion by the major histocompatibility complex (MHC) class I molecules. Mutations in these genes are associated with immune deficiency due to a defective antigen presenting process(13). ABCB9 is localized in lysosomal compartments and forms homodimers which share structural and functional similarities to ABCB2/3; however, its biological role seems to be unrelated to MHC I antigen processing(13). ABCB4 and ABCB11 are both expressed in the liver, where they transport components of the bile. Mutations in ABCB4 and ABCB11 are
responsible progressive familial intrahepatic cholestasis type 3 and 2, respectively. ABCB4 is the canalicular membrane transporter of phosphatidylcholine, a major and exclusive component of mammalian bile(30). ABCB11 facilitates the secretion of monoanionic bile salts, including glycocholate and taurocholate, through the canalicular membrane(10). These transporters provide the major driving force for the generation of bile flow. Alterations in the function of ABCB4 and ABCB11 by either inherited mutations or acquired processes, such as inhibition by drugs or disease-related down-regulation, may lead to a wide spectrum of mild to severe forms of liver disease. Furthermore, many population based studies have revealed that single nucleotide polymorphisms in the ABCB4 and ABCB11 genes serve as biomarker candidates for predicting the progression and prognosis of hepatic disorders such as primary biliary cirrhosis and intrahepatic cholestasis(1). The half transporters ABCB6, ABCB7, ABCB8 and ABCB10 are located in mitochondria, where they function to maintain iron homeo-stasis and are necessary for the formation of iron-sulfur protein complexes. Two distinct missense mutations in ABCB7 are found in patients with X-linked sideroblastic anemia and ataxia(28).
ABCC subfamily
Members of the ABCC subfamily are full transporters that function in a wide range of cellular activities including ion transport, cell surface reception, and toxin secretion. ABCC1 transports a diverse spectrum of therapeutic agents and physiological substrates such as conjugated organic anions and its over-expression confers drug resistance to several types of cancer cells(5). For example, in our laboratory, we identified MRP1 expression as a marker of prognostic significance and a potential target for chemotherapeutic schemes in patients with advanced stages of nasopharyngeal carcinoma(24). The role of ABCC1 polymor-phisms in normal cellular physiology as well as clinical drug resistance and toxicity is in-creasingly evident based primarily on in vitro studies using site-directed mutant proteins(8).

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
85
Similar to ABCC1, ABCC2-5 proteins are all organic anion pumps, but they differ in substrate specificity, transport kinetics and tissue distribu-tion (Table 1). Acquired or hereditary defi-ciency of ABCC2 results in Dubin-Johnson syndrome, which is characterized by an increased concentration of bilirubin glucuro-nosides in the blood. The association of this rare but benign condition of the liver with the loss of expression and function of ABCC2 led to the identification of a number of naturally occurring disease-causing genetic variations in this transporter(26). The substrate specificity of ABCC6 is still unknown; however, defects in ABCC6 cause pseudo-xanthoma elasticum, a heritable disorder characterized by calcification of connective fibers in the skin, retina, and arteries(16). ABCC7 is a unique ABC transporter as it functions as a chloride-ion channel that plays a role in exocrine secretion. Mutations in ABCC7 introduce a range of clinical symptoms from congenital bilateral aplasia of the vas deferens to severe cystic fibrosis(27). Cystic fibrosis is an autosomal recessive disease that causes the production of abnormally thick and sticky mucus in several different parts of the body, predominantly in the lungs and other parts of the respiratory system. Genetic variations in ABCC7 and combinations of particular alleles at several polymorphic loci, yielding ABCC7 proteins with functional differences, have been extensively investigated in order to improve the capacity to predict cystic fibrosis outcomes(27). Unlike other ABCC members, ABCC8 and ABCC9 proteins have no identified transport function, but instead function as sulfonylurea receptors that regulate potassium channels involved in the modulation of insulin secretion. Mutations in either subunit can alter the potassium balance. In the case of ABCB8, mutations found in neurons and insulin-secreting pancreatic beta cells are the cause of familial persistent hyperinsulinemic hypogly-cemia during infancy and neonatal diabetes(11). ABCC9 dysfunction has been found to be associated with dilated cardiomyopathy with ventricular tachycardia(32).
ABCD Subfamily
All ABCD subfamily members are half transporters that are located in peroxisomes. They are proposed to form a functional homodimeric or heterodimeric transporter, which can potentially mediate the peroxisomal transport or catabolism of very long chain fatty acids(14). Mutations in the ABCD1 gene result in elevated levels of very long-chain fatty acids and reduced β-oxidation in peroxisomes. These mutations are the underlying cause of a severe neurodegenerative disorder called X-linked adrenoleukodystrophy, which is characterized by adrenal deficiency and progressive demye-lination of the nervous system(34). Defects in the ABCD3 gene have been associated with Zell- weger syndrome, a genetically heterogeneous disorder of peroxisomal biogenesis(34). The ABCD proteins exhibit overlapping substrate specificities, however, their detailed physio- logical functions are still largely unknown.
ABCE and ABCF subfamilies
The ABCE and ABCF proteins possess two nucleotide-binding domains, which are con- served throughout other ABC superfamily proteins, but lack the transmembrane domains. They are ubiquitously expressed and have not been associated with any membrane transport processes. The ABCE subfamily consists of one single member, ABCE1 (alternatively referred to as 2'-5'-oligoadenylate-binding protein, that inhibits the activity of RNase L, an essential part of the cellular innate immune response. Activation of RNase L leads to the inhibition of protein synthesis through the degradation of cellular RNAs(4). There are three members in the ABCF subfamily. The best characterized member, ABCF1, serves as a component of ribosomal complexes, which are involved in translation initiation and elonga-tion(19). Although these proteins are highly conserved in eukaryotic genomes, there are no human diseases/phenotypes linked to these subfamilies.
ABCG Subfamily
The ABCG subfamily contains five half-transporters with a single nucleotide-binding

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
86
domain at the amino terminus followed by a transmembrane domain. They form homo- or obligate heterodimers to become fully active membrane transporters. ABCG1 is predomi-nantly localized in intracellular compartments including the endoplasmic reticulum and the Golgi membranes. ABCG1 plays a significant role in the efflux of excess cholesterol to HDL in macrophages, the most relevant step with respect to atherosclerosis(35). Excess cholesterol in the body must be exported from macrophage foam cells, and then converted or dissolved into bile acids in the liver, followed by excretion in the feces. Over-expression of ABCG1 has been found in macrophages from patients with Tangier disease(25). Recently, we found that ABCG1 is expressed in cancerous cholangio- cytes, but not in normal cells, supporting its role in cholesterol homeostasis(23). Even though there is significant evidence for the involvement of ABCG1 in cholesterol and phospholipid transport, no loss-of-function mutations in the ABCG1 gene have been identified that would help to clarify the physiological function of ABCG1. ABCG2 was initially cloned from the placenta and multidrug-resistant cancer cell lines that did not express either ABCB1 or ABCC1. Over-expression of ABCG2 has been observed in several human cancer cell lines that had been selected for drug resistance as well as in tumor samples of cancer patients. Several studies demonstrated that ABCG2 transports a wide range of clinical agents as well as endogenous substrates such as sterols, porphy-rins and a variety of dietary compounds(9,17,18,33). ABCG2 expression in the apical membrane of cells in tissues with excretory functions, such as hepatocytes, enterocytes, endothelial and syncy- tiotrophoblast cells, suggests a role in forming a protective barrier or in the secretion of environ- mental or clinically administered compounds. Variations in the ABCG2 gene appear influential in affecting inter-patient drug disposition and the bioavailability of certain chemotherapeutic drugs(7,18). Several reports on ABCG2 pharma-cogenetics have identified prevalent ABCG2 polymorphisms in many populations, however, there are many conflicting results regarding the functional effects of these polymorphisms(7).
ABCG4 shares approximately 70% similarity in amino acid sequence to the human ABCG1. Similar to ABCG1, ABCG4 is able to transfer cholesterol to HDL in vitro(35); however, its physiological role has not yet been elucidated. ABCG5 and ABCG8 expression is confined to the liver and intestines. ABCG5/ABCG8 are obligate heterodimers and are dependent on one another for sterol transport across the endoplasmic reticulum and for the excretion of hepatic sterol into the bile. Mutations in either ABCG5 or ABCG8 cause sitosterolemia, an autosomal recessive disorder characterized by impaired absorption and retention of both plant-derived (primarily sitosterol) and animal-derived (cholesterol) sterols in plasma and tissues(29). Certain polymorphisms in the ABCG5 or ABCG8 genes are also associated with several facets of cholesterol metabolism, including the baseline cholesterol level, cholesterol kinetics, the individual respon-siveness of plasma cholesterol to dietary and pharmaceutical interventions for hypercholes-terolemia, and an increased risk of gallstones(29).
ABC Transporters in Non-human Organisms
Homologs and orthologs of seven ABC subfamilies are found across eukaryotic genomes. Conveniently, ABC transporters in animal models have been identified and used to probe the function and regulation of selected genes. Characterization of the ABC transporters from model organisms as well as the human genome will certainly provide additional insights into normal physiology and human disease. Recently, we reported the results of our studies on the ABC multidrug transporters in the rock oyster(20). The levels of ABC proteins in oysters can be useful as indicators in monitoring the effects of general environmental stress. We also attempted to characterize the ABCB1, ABCB11, ABCC1 and ABCG2 ortho- logs in the giant liver fluke and found that they potentially play a role in bile salt detoxification and resistance to anthelmintics(22). As anthel-mintic resistance is emerging in some countries, understanding the resistance mechanisms, especially in regard to the role of the ABC

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
87
transporters, may provide a glimpse for fascio-licidal targets.
Conclusions
The ABC superfamily is one of the largest protein families and is found across a broad range of organisms. Members of this superfamily transport a diverse set of substrates across membranes. Several techniques have been used to identify substrates and inhibitors of these proteins and subsequently assess the effect of genetic mutations on transport, both in vitro and in vivo. There is a significant variation in the ABC encoding genes, resulting in alterations at the transcriptional and translational level as well as protein function. Such genetic varia-bility in the ABC transporters underlines the features of the aforementioned diseases or phenotypes in humans as well as the inter-individual variability in the absorption, distribution, metabolism, and excretion of compounds, ultimately resulting in differences in clinical endpoints. Therefore, investigation of these variations including polymorphisms, mutations, expression levels and protein functions will provide us a window into the usefulness of the ABC transporters as biomarkers for human diseases.
Acknowledgements
I am grateful to Mahidol University for financial support of the research on ABC proteins.
References 1. Alissa FT, Jaffe R, Shneider BL. Update on
progressive familial intrahepatic cholestasis. J Pediatr Gastroenterol Nutr 2008:46;241-52.
2. Allikmets R. A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat Genet 1997:17;122.
3. Allikmets R. Simple and complex ABCR: genetic predisposition to retinal disease. Am J Hum Genet 2000:67;793-9.
4. Bisbal C, Salehzada T, Silhol M, Martinand C, Le Roy F, Lebleu B. The 2-5A/RNase L pathway and inhibition by RNase L inhibitor (RLI). Methods Mol Biol 2001:160;183-98.
5. Borst P, Evers R, Kool M, Wijnholds J. A family of drug transporters: the multidrug
resistance-associated proteins. J Natl Cancer Inst 2000;92: 1295-302.
6. Borst P, Elferink RO. Mammalian ABC trans-porters in health and disease. Annu Rev Biochem 2002;71:537-92.
7. Cervenak J, Andrikovics H, Ozvegy-Laczka C, Tordai A, Német, K, Váradi A et al. The role of the human ABCG2 multidrug transporter and its variants in cancer therapy and toxicology. Cancer Lett 2006;234:62-72.
8. Conseil G, Deeley RG, Cole SP. Polymorphisms of MRP1 (ABCC1) and related ATP-dependent drug transporters. Pharmacogenet Genomics 2005;15:523-33.
9. Cooray HC, Janvilisri T, Van Veen HW, Hladky SB, Barrand MA. Interaction of the breast cancer resistance protein with plant polyphenols. Biochem Biophys Res Comm 2004;317:269-75.
10. Gerloff T, Stieger B, Hagenbuch B, Madon J, Landmann L, Roth J et al. The sister of P-glycoprotein represents the canalicular bile salt export pump of mammalian liver. J Biol Chem 1998;273:10046-50.
11. Gloyn AL, Siddiqui J, Ellard S. Mutations in the genes encoding the pancreatic beta-cell KATP channel subunits Kir6.2 (KCNJ11) and SUR1 (ABCC8) in diabetes mellitus and hyper-insulinism. Hum Mutat 2006;27:220-31.
12. Gottesman MM. Mechanisms of cancer drug resistance. Annu Rev Med 2002;53:615-27.
13. Herget M, Tampé R. Intracellular peptide trans-porters in human--compartmentalization of the "peptidome". Pflugers Arch 2007;453:591-600.
14. Hettema E H, Tabak HF. Transport of fatty acids and metabolites across the peroxisomal mem-brane. Biochim Biophys Acta 2000;1486: 18-27.
15. Higgins CF. ABC transporters: from micro-organisms to man. Annu Rev Cell Biol 1992;8: 67-113.
16. Iliás A, Urbán Z, Seidl TL, Le Saux O, Sinkó E, Boyd CD et al. Loss of ATP-dependent transport activity in pseudoxanthoma elasticum-associated mutants of human ABCC6 (MRP6). J Biol Chem 2002;277:16860-7.
17. Janvilisri T, Venter H, Shahi S, Reuter G, Balakrishnan L, Van Veen HW. Sterol transport by the human breast cancer resistance protein (ABCG2) expressed in Lactococcus lactis. J Biol Chem 2003;278:20645-51.
18. Janvilisri T, Shahi S, Venter H, Balakrishnan L, Van Veen HW. Arginine-482 is not essential for transport of antibiotics, primary bile acids and unconjugated sterols by the human breast cancer

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
88
resistance protein (ABCG2). Biochem J 2005; 385:419-26.
19. Kerr ID. Sequence analysis of twin ATP binding cassette proteins involved in translational control, antibiotic resistance, and ribonuclease L inhibition. Biochem Biophys Res Commun 2004;315:166-73.
20. Kingtong S, Chitramvong Y, Janvilisri T. ATP-binding cassette multidrug transporters in Indian-rock oyster Saccrostrea forskalii and their roles in the export of an environmental organic pollutant tributyltin. Aquat Toxicol 2007;85: 124-32.
21. Kipp H, Arias IM Intracellular trafficking and regulation of canalicular ATP-binding cassette transporters. Semin Liver Dis 2000;20:339-51.
22. Kumkate S, Chunchop S. Janvilisri T. Expression of ATP-binding cassette multidrug transporters in the giant liver fluke Fasciola gigantica and their possible involvement in the transport of bile salts and anthelmintics. Mol Cell Biochem 2008;317:77-84.
23. Laohasinnarong S, Larbcharoensub N, Kumkate, S, Janvilisri T. Expression of lipid transporters -ABCA1, ABCB11 and ABCG1 in intrahepatic cholangiocarcinoma. Abstract. 3rd Annual Symposium of Protein Society of Thailand: Frontiers in Protein Research, Chulabhorn Research Institute, Bangkok, Thailand. 2008.
24. Larbcharoensub N, Leopairat J, Sirachainan E, Narkwong L, Bhongmakapat T, Rasmeepaisarn K et al. Association between multidrug resis-tance-associated protein 1 (MRP1) and poor prognosis in patients with nasopharyngeal carcinoma treated with radiotherapy and concurrent chemotherapy. Hum Pathol 2008;39: 837-45.
25. Lorkowski S, Kratz M, Wenner C, Schmidt R, Weitkamp B, Fobker M et al. Expression of the ATP-binding cassette transporter gene ABCG1 (ABC8) in Tangier disease. Biochem Biophys Res Commun 2001;283:821-30.
26. Nies AT, Keppler D. The apical conjugate efflux pump ABCC2 (MRP2). Pflugers Arch 2007; 453:643-59.
27. Riordan JR CFTR function and prospects for therapy. Annu Rev Biochem 2008;77:701-26.
28. Rouault TA, Tong, WH. Iron-sulfur cluster bio-genesis and human disease. Trends Genet 2008; 24:398-407.
29. Rudkowska I, Jones PJ. Polymorphisms in ABCG5/G8 transporters linked to hypercholes-terolemia and gallstone disease. Nutr Rev 2008; 66:343-8.
30. Ruetz S, Gro, P. Phosphatidylcholine translo-case: a physiological role for the mdr2 gene. Cell 1994;77:1071-81.
31. Schwab M, Eichelbaum M, Fromm MF. Genetic polymorphisms of the human MDR1 drug transporter. Annu Rev Pharmacol Toxicol 2003; 43:285-307.
32. Solbach TF, König J, Fromm MF, Zolk O. ATP-binding cassette transporters in the heart. Trends Cardiovasc Med 2006;16:7-15.
33. Velamakanni S, Wei SL, Janvilisri T, van Veen HW. ABCG transporters: structure, substrate specificities and physiological roles: a brief overview. J Bioenerg Biomembr 2007;39:465-71.
34. Wanders RJ, Visser WF, van Roermund CW, Kemp S, Waterham HR. The peroxisomal ABC transporter family. Pflugers Arch 2007;453:719-34.
35. Wang N, Lan D, Chen W, Matsuura F, Tall AR. ATP-binding cassette transporters G1 and G4 mediate cellular cholesterol efflux to high-density lipoproteins. Proc Natl Acad Sci USA 2004;101:9774-9.
36. Zarubica A, Trompier D, Chimini G. ABCA1, from pathology to membrane function. Pflugers Arch 2007;453:569-79.

89
Leptospira : Overview and Genetics
Padungsri Dubbs Department of Microbiology, Faculty of Science, Mahidol University.
Abstract
Since a major outbreak in Nicaragua in 1995, leptospirosis has been recognized as one of the important infectious diseases that affects human populations worldwide. In Thailand, according to a recent report from Center of Epidemiological Information, Bureau of Epidemiology, Ministry of Public Health, there were 3,437 reported cases of leptospirosis, of which 59 patients died from the disease. The severity of leptospirosis ranges from a relatively mild flu-like illness to a severe, potentially fatal syndrome of multi-organ involvement resulting in clinical manifestations such as jaundice, renal failure and/or pulmonary hemorrhage. Little knowledge of the pathogenic mechanisms of this organism exists. One of the key reasons for this is the lack of genetic tools. During the past decade, tools for gene manipulation in Leptospira spp. have been progressively established. Introduction of a gene into Leptospira can be achieved via transformation by electroporation or conjugation. In addition, techniques for random and targeted mutagenesis have been developed. The availability of these new tools for genetic analysis in Leptospira provides a basis for future molecular studies of the physiology and pathogenic mechanisms in this organism.
Keywords: leptospirosis; genetic tools; virulence factors
Introduction
Leptospirosis is now regarded as one of the important emerging infectious diseases with the latest outbreaks occurring in Nicaragua, Brazil, India, Malaysia, Thailand and the United States(31). In Thailand, a major outbreak in 2000 resulted in a peak of 14,285 reported cases of leptospirosis, since then the annual number has declined to about 3000-4000 reported cases(18,19,56). This disease is caused by spirochetal bacteria in the genus Leptospira. Reservoir hosts of the infection are wild, peridomestic and domestic animals, especially rodents. Leptospires persist and proliferate in the proximal renal tubules of these maintenance hosts and the organisms are shed in the urine of asymptomatic carriers throughout their life-time(4). Humans are accidental hosts in the transmission cycle. The infection is usually through either direct or indirect contact with infected animals, contaminated urine or urine-contaminated water or moist soil(20). Clinical manifestations of leptospirosis range from subclinical illness to severe manifestations like
jaundice, renal failure and/or pulmonary hemorrhage(58). The mortality rate can be up to 23%(61).
Many leptospiral proteins such as: haemo-lysins(30,44), sphingomyelinase(44,55), phospholi- pase(44), haemolysis-associated protein-1 (Hap1 or LipL32)(23) and a leptospiral immunoglo-bulin-like protein (Lig)(36) have been implicated as potential virulence factors. However, there is little evidence directly linking these factors to pathogenesis. One factor that has impeded progress in this area is the lack of appropriate genetic tools. This review will focus on current development of new methods for genetic mani-pulation in Leptospira.
Taxonomy and Classification
Leptospires are members of the class, Spirochaetes, in the order, Spirochaetales, which contains the families; Spirochaetaceae, Serpuli-naceae and Leptospiraceae(46). The family Spirochaetaceae contains two genera of medical importance; Borrelia, the causative agent of Lyme disease/relapsing fever and

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
90
Treponema, the causative agent of syphilis(26). The family, Leptospiraceae, is divided into three genera: Leptonema, Turneria (previously Leptospira parva) and Leptospira, the only genus of the group containing members that cause disease(21). Two systems currently exist for the classification of members of genus, Leptospira. One is serology based and the other is based on DNA relatedness. In the older serology based system the genus Leptospira consists of two species, the free-living, saprophytic, nonpathogenic Leptospira biflexa (L. biflexa) and the pathogenic, L. interrogans. Both species are classified into serovars defined by agglutination after cross-absorption with anti-sera to homologous antigens(31). Anti-genically related serovars are grouped into serogroups. This serological classification divides the “biflexa” complex (later L. biflexa sensu lato) into over 60 serovars and the “interrogans” complex (later L. interrogans sensu lato) into more than 200 serovars comprising 24 serogroups(4). More recently, a genotypic system, which is more taxonomically consistent, has been used to classify Leptospira based on DNA relatedness. This method divides the genus Leptospira into 16 genomospecies (excluding L. parva): L. interrogans (referred to as L. interrogans sensu stricto), L. santarosai, L. borgpetersenii, L. kirschneri, L. noguchii, L. weilii, L. inadai, L. fainei, L. biflexa (referred to as L. biflexa sensu stricto), L. meyeri, L. alexanderi, L. wolbachii, and genomospecies 1, 3, 4, 5(8). However, this genotypic classification is incompatible with the older system of serogroups and serovars that has been used for many years(31). Therefore, strains of Leptospira are still generally referred to by serovar.
Microbiology
Leptospires are thin, flexible, helically shaped, Gram-negative bacteria with pointed ends that are usually bent or hooked (Figure 1).
The size of the organisms is approxi-mately 0.1 μm in diameter and 6-20 μm in length, thus leptospires can pass through 0.2 μm pore-size filters(59). The organisms can be directly visualized using a dark-field or phase-contrast microscope. Leptospires are obligately
aerobic bacteria with an optimum growth tem-perature between 28 and 30°C and an optimal pH range for growth of 7.2-7.6(21). The bacteria are chemoheterotrophs and use long-chain fatty alcohols as carbon and energy sources. They cannot use carbohydrates or amino acids as major sources of energy(27). Leptospires can be cultured in simple media containing long-chain fatty acids as a source of carbon, ammonium salts as a nitrogen source, and vitamins B2 and B12 as growth factors(31). A commonly used growth medium is Ellinghausen-McCullough-Johnson-Harris (EMJH) medium containing Tweens as a source of fatty acids and BSA as a detoxifier(21). When grown in a solid medium, the bacteria will form colonies just beneath the surface of the medium.
Figure 1 A darkfield micrograph of leptospires. Arrows indicate the hooked ends.
Spirochetes have a distinct double-membrane structure consisting of a cytoplasmic membrane that is closely associated with a peptidoglycan layer followed by a periplasmic space and an outer membrane-like structure called an outer membrane sheath (Figure 2).
The outer membrane sheath of spirochetes appears to be fluid and labile(23) consisting of lipopolysaccharide (LPS) and outer membrane proteins (OMPs). LPS extracted from L. interrogans serovar Canicola had lower endotoxic activity than E. coli LPS(25). It has been shown that leptospiral LPS activates human cells through the Toll-like receptor 2 (TLR2)(41,60), whereas it activates murine cells through TLR2 and TLR4(41). Leptospiral OMPs

Figure 2
are comtransmemproteins
Spstructureflagella periplasmaround which enclosedmembraperiplasmthe celltoward overlappimparteddriven bcell cyliclockwisoppositeother enthat promotion(3
through that foumotility stalled(1)
flagella (both clThis nonmotions the cell nvibrating
Schematic leptospiral relation to cell wall.
mposed of at lmbrane prot and lipoprot
pirochetes poe called p
or axial fimic space (Fthe helical comprises
d by the ane complexmic flagelluml cylinder.
the centeping(27). Mod by rotationby the protoinder in eithse directione directions nd counter-copels the ce32). This a
highly viscund in con
of most ). Flexing at both ends
lockwise or n-translationof a spiroch
near the endg etc.(10). Th
วทยาศาส
illustration operiplasmic
a cytoplasmic
least three tyteins, periphteins(16).
ossess a uniqeriplasmic ilaments thaFigure 2). Th
protoplasmithe cytoplapeptidoglyc
x. Leptospim attached n Each flag
er of the ovement ofn of the peripon motive foher a clockwn(35). Flagel(one end cloclockwise) gell to move llows spiroccous environ
nnective tissother bactemovement
s rotate in theboth counte
nal motility ihetal cell sucds or the middhe compositi
สตรเพอการพฒนา :
of a location c flagellum c membrane a
ypes of proteheral membr
que filamentflagella, en
at reside in he flagella wic cell cylinasmic contcan-cytoplasires have near each endgellum exte
cell withf the cell plasmic flageorce, around wise or counlla rotationockwise andgenerates thin a screw
chetes to mnments suchsues whereeria would
occurs we same direcer-clockwise)includes varich as bendingdle, looping ion of perip
รวมบทความวชาการ
of a in
and a
eins: rane
tous ndo-
the wrap nder ents
smic one d of ends hout
is ella, the
nter-in
the hrust wlike move h as
the be
when tion )(11). ious g of and
plas-
micextof oof flagothprofilaThesimbac
thapalThibotinsiconabokb(
repto CBouandposLairep
EpMa
diseanddisecouAmRisoccwowatbutdevareandwatOuflootheorg
ร ฉลองครบรอบ 50 ป
c flagellar fiternal flagellother bacterifilament progellar filame
her bacteriaotein sheath mament coree spirochete
milarities tocteria(32).
The leptan the 1.1 andllidum and Bis reflects thth as free livide animal hnsists of twout 3.6-4.39,44,51,53,62). In
plicon denoteCI and CII(5
urhy et al.(7)
d CII, L. ssesses a 5iGI I that
plicating plas
pidemiologyanifestation
Leptospirease that occd developingease is huntries, suchmerica, than csks for incupations thrkers and anter, especialtchers, soldveloped coun
either assocd adventure ter(24) or
utbreaks of oding or rec possibility
ganisms in c
ป แหงการสถาปนาคณ
filaments is dar filamentsia are compootein(32). Spients are thiand are com
made up of Fcomposed oFlaB filame
o filament
tospiral genod 1.5 Mb gen
Borrelia burghe ability oving organismhosts(4). Theo circular cMb and CIn L. biflexa s
ed p74 was a51). Moreove) revealed thinterrogans
54-kb genomcan exist amid.
y and Clinins
rosis is an curs worldwig countries. igher in th as Southecountries in tnfection arhat promot
nimal urine olly: farmers
diers and ntries, most cciated with r
travel thatconditions leptospirosi
creational evy of humancontaminated
ณะวทยาศาสตร มหา
distinct from. Flagellar f
osed of only irochetal pericker than mprised of aFlaA that surof FlaB(11)
ent core has sproteins o
ome is mucnomes of Tr
gdorferi, respf leptospiresms and as pe leptospiralchromosomeII of about serovar Pato
also found iner, a recent hat, in additis serovar Lmic island as an auton
ical
emerging ide in both dThe incidentropical deeast Asia antemperate cle associate
te contact or urine conta, livestock sewer workcases of leptrecreational t are involv
of slum is often arivents which n exposure d water and
าวทยาลยมหดล
9
m that of filaments one type
riplasmic those of a unique rrounds a (Fig. 2). sequence of other
ch larger eponema
pectively. s to live athogens
l genome s: CI of 300-350 c, a third
n addition study by
ion to CI Lai also
denoted nomously
zoonotic eveloped
nce of the eveloping nd Latin imates(4). ed with between
aminated workers, kers. In tospirosis activities
ved with living(4).
ise after increase to the
d soil(52).
91

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
92
For example, an outbreak in 1998 was reported after triathlons in Wisconsin and Illinois(38). In 2000, an outbreak occurred in Thailand that was associated with heavy flooding, while a second outbreak in Malaysia was linked to the EcoChallenge Sabah 2000 competition in Borneo(31).
Leptospira infection in animals such as dogs, marsupials, bats and rodents often results in renal carriers. Transmission to humans is usually from direct or indirect contact with urine of infected animals or exposure to urine-contaminated water or soil(21). The excreted leptospires can survive as free-living organisms for months in a moist and relatively neutral to alkaline (pH 6.2-8) environment at temperatures exceeding 22°C(59). The organisms cannot survive in dry and acidic environments. It is believed that the organisms enter their host through abrasions or breaches of the surface integument such as those that can occur during prolonged immersion of the skin in water. The organisms are also able to enter through the mucosal surfaces of the eye, mouth and nasopharynx(21).
The majority of cases of Leptospira infection may result in a mild form of disease with severity ranging from subclinical or very mild symptoms to a self-limited febrile illness. Nevertheless, some cases can be severe and possibly fatal and these cases are of major concern. Two severe disease forms, icteric leptospirosis (also known as Weil’s disease) and severe pulmonary haemorrhage syndrome (SPHS) have been reported(37). Weil’s disease, found in about 5-10% of all leptospirosis patients, is characterized by jaundice and renal failure and has a 5-15% mortality rate(31). Leptospirosis-associated SPHS has been recognized since a major outbreak in Nicaragua in 1995(58) and is now a major cause of death with over 50% case fatality(37). The illness usually starts, after an average incubation period of 10 days, with headache, fever and severe muscular pain, particularly in the thighs, calves, lumbosacral region and abdomen(59). In some studies, it has been reported that the majority of leptospirosis patients display conjunctival suffusion(31). The clinical presentation of this
disease is biphasic with an acute, septicemic or leptospiremic phase lasting about a week, followed by an immune or leptospiruric phase that is characterized by the production of antibodies and the excretion of leptospires in the urine. During the acute phase, leptospires can be recovered from the blood and cerebrospinal fluid (CSF). The immune phase is usually accompanied by the disappearance of lepto-spires in the blood(31). In addition to its importance as a human pathogen, Leptospira infection is also a significant problem for domestic and commercial livestock that can lead to abortion, stillbirth, infertility, decreased milk production, and death(21).
Leptospiral Virulence Factors
The various pathologies of Leptospira infection are thought to be the result of the direct effects of the organism on the host and the effects of the host's immune response to infection(4). Up until now, little is known about the pathogenesis of this disease. After host infection, pathogenic leptospires must rapidly get into the bloodstream and quickly disseminate to target organs in order to evade the host immune response. Thus, motility and chemotaxis may be crucial during initial infection and dissemination of spirochetes(4). Leptospiral outer membrane proteins (OMPs) are thought to be important for host interactions(16). Examples of these are two groups of lipoproteins that have been identified as potential afimbrial adhesins. The first group consists of three Lig (leptospiral immuno-globulin-like repeat) proteins, LigA, LigB and LigC, belonging to the superfamily of bacterial immunoglobulin-like (Big) repeat domain proteins(36). The Lig proteins are detected only from leptospires isolated from infected animals and are lost in highly passaged, culture-attenuated, leptospires(36). It was shown that LigA and LigB bind to fibronectin and fibrinogen(13,33). The second leptospiral adhesin is LfhA (Lsa 24), a putative factor H binding protein that binds the extracellular matrix protein laminin(2,57). Evidence suggesting that the OMPs are involved in pathogenesis includes the observations that genes encoding the Lig proteins and LfhA are absent in nonpathogenic

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
93
L. biflexa(51). Moreover, it was shown that the expression of some surface-exposed outer membrane proteins like Q1p42(42), LipL36(43), pL50(15), pL24(15) were regulated in response to environmental signals similar to those encountered during host infection. These included: growth phase, growth temperature, iron availability and osmolarity. It is possible that the response to these changes in the environment might allow virulent leptospires to adapt to the new environments they encounter inside or outside the host.
Leptospiral invasion may be mediated by the secretion of enzymes that degrade host cell such as: sphingomyelinase C (sphA)(55), haemo-lysin (sphH)(30) phospholipase D, collagenase and metalloprotease(44). Genes encoding proteins with homology to animal proteins that have functions involved in haemostasis such as: pafAH (encoding the mammalian platelet-activating factor acetyl hydrolase), vwa (encoding von Williebrand factor type A domains) and pon (encoding paraoxonase), have also been identified in the genomes of L. interrogans(53).
Genetic Tools for Use in Leptospira
A breakthrough for genetic exchange in Leptospira began with the discovery that the temperate bacteriophage, LE1, in L. biflexa serovar Patoc strain Patoc1 replicates in the prophage state as a circular plasmid(54). This led to the construction of a L. biflexa-E. coli shuttle vector, pGKLep1, which contains a 5.2-kb DNA fragment spanning the LE1 origin of replication that allows maintenance of the plasmid in leptospires and a selectable kanamycin resis-tance cassette from Enterococcus faecalis(54). Using electrotransformation, this shuttle vector could be stably transformed into L. biflexa serovar Patoc strain Patoc1 and another saprophytic species, L. meyeri serovar Semaranga strain Veldrat Semarang 173 with high efficiency(54). Unfortunately, attempts to transformation of pGKlep1 into three patho-genic species, L. interrogans, L. inadai and L. fainei, were unsuccessful(54). Subsequently, a first L. biflexa flaB knockout mutant was generated by the same group of researchers
from the Institut Pasteur using allelic gene exchange techniques(48). The critical step for successful gene replacement in leptospires was the use of UV irradiated or alkaline treated suicide plasmid DNA to enhance the frequency of homologous recombination(48). The flaB mutant did not possess periplasmic flagella and was non-motile providing direct evidence that the periplasmic flagella function in motility(48). Moreover, the fact that the flaB mutant retained its helical shape suggested that leptospiral periplasmic flagella did not determine the shape of the cell. These results were in contrast to those observed in a Borrelia flaB mutant that lost its flat-wave morphology and adopted a rod-like shape(39). Following this study, a L. biflexa recA mutant(28), a L. biflexa hemH mutant(22), a L. meyeri trpE mutant(3) and L. meyeri metW, metX, metY mutants(49) were constructed to gain insight into the functions of their respective gene products. Successful complementation was then performed in the L. meyeri trpE mutant using plasmid, pGKLStrpE, a derivative of pGKLep1 containing a 2.2-kb LE1 origin of replication, a spectinomycin cassette from Staphylococcus aureus as a second selectable marker and L. meyeri trpE(3).
Another major advance for genetic analysis in Leptospira is the use of the Himar1 mariner transposon to perform random inser-tional mutagenesis in L. biflexa(34) as well as in pathogenic L. interrogans(6,40). Transposon mutagenesis allows the identification of novel genes that might be involved in a process of interest. This is particularly useful since a comparative whole genome analysis of three pathogenic leptospires: L. interrogans serovars Lai and Copenhageni as well as L. borgpeter-senii, and the nonpathogenic leptospire, L. biflexa, revealed that 893 of the 1431 pathogen specific genes encode proteins of unknown function(51). The random mutagenesis approach would help in identifying those genes that play a role in pathogenesis by looking for transposon insertion mutants that exhibit a decrease in virulence.
The Himar1 mariner transposon, ori-ginally isolated from the horn fly Haematobia irritans, can function in diverse organisms that

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
94
can be either eukaryotes or prokaryotes because transposition does not require host-specific factors(29). Although the original plasmid, pSC189(12), carrying the hyperactive transposase C9 and transposon terminal inverted repeats flanking a kanamycin resistance gene, was successfully utilized to deliver the kanamycin-resistant Himar1 transposon in both L. biflexa and L. interrogans; however, lower numbers of L. interrogans mutants were obtained(6). In order to increase the yield of L. interrogans insertion mutants, two derivatives of pSC189, pMKL and pMSL, were constructed by replacing the original promoter of the C9 transposase with the B. burgdorferi flgB promoter. This results in an increase in transposase expression and thus, an increase in the numbers of transposon insertion mutants. Plasmid pMKL carries transposon terminal inverted repeats flanking an Enterococcus faecalis kanamycin resistance cassette, whereas pMSL carries transposon terminal inverted repeats flanking a Staphylococcus aureus spectinomycin cassette(6). Eventually, targeted mutagenesis was successfully performed in L. interrogans using an allelic gene exchange method similar to that previously described in L. biflexa. Although it was shown that proteins in the Big superfamily, such as intimin from enteropathogenic E. coli(45) and invasin from Yersinia pseudotuberculosis(47), are associated with virulence, analysis of a L. interrogans ligB insertion-mutant revealed that the ligB deficient strain retained its ability to cause acute leptospirosis in a hamster model system with an LD50 similar to that of the wild-type strain(14). The efficiency of the L. interrogans ligB mutant to induce renal colonization in experimental rats (Rattus norvegicus) was comparable to that of the wild-type. In addition, the disruption of ligB had no effect on the organism’s ability to attach to MDCK cells.
Besides electroporation, the introduction of foreign DNA to Leptospira spp. can be achieved by conjugation(50). A recent report revealed evidence of conjugal DNA transfer between a donor strain, E. coli β2163(17), carrying RP-based broad-host-range plasmids, to a recipient, either L. biflexa or L.
interrogans(50). This offers one more available tool to facilitate genetic manipulation in Leptospira.
Conclusions
As compared to other well studied bacteria, genetic study in Leptospira is still in its early phase. Characterization of most lepto-spiral genes was performed by complementa- tion studies in E. coli mutants. However, whole genome analysis of L. interrogans serovars Lai(53), Copenhageni(44), L. borgpetersenii(9) and L. biflexa(51) suggested that many leptospiral genes are distinctive from those of E. coli. Therefore, the availability of new genetic tools will make possible functional studies of those unique leptospiral proteins. Moreover, since leptospires are facultative parasites that can live as free-living organisms for months prior to infection of mammalian hosts, the organisms must have the ability to regulate gene expression in response to drastic changes in their environment. To date, it is not known how leptospires regulate gene expression in response to environmental cues. After host infection, virulent leptospires must adapt to the new environment inside the host and evade the host immune response in order to survive and proliferate within host tissues. The new genetic tools will make it possible to better understand the regulatory network(s) controlling gene expression that allow Leptospira to sense and adapt to growth as either a parasitic or free-living organism.
Acknowledgements
The author would like to thank Dr. James Dubbs for editing the manuscript and Benjapron Mokmake for her help in figure preparation.
References 1. Armitage JP. Behavioral responses in bacteria.
Annu Rev Physiol 1992;54:683-714. 2. Barbosa AS, Abreu PAE, Neves FO, Atzingen
MV, Watanabe MM, Vieira ML et al. A newly identified leptospiral adhesin mediates attach-ment to laminin. Infect Immun 2006;74:6356-64.
3. Bauby H, Saint Girons I, Picardeau M. Con-struction and complementation of the first auxo-

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
95
trophic mutant in the spirochaete Leptospira meyeri. Microbiology 2003;149:689-93.
4. Bharti AR, Nally JE, Ricaldi JN, Matthias MA, Diaz MM, Lovett MA et al. Leptospirosis: a zoonotic disease of global importance. Lancet Infect Dis 2003;3:757-71.
5. Bourhy P, Saint Girons I. Localization of the Leptospira interrogans metF gene on the CII secondary chromosome. FEMS Microbiol Lett 2000;191:259-63.
6. Bourhy P, Louvel H, Saint Girons I, Picardeau M. Random insertional mutagenesis of Lepto-spira interrogans, the agent of leptospirosis, using a mariner transposon. J Bacteriol 2005; 187:3255-8.
7. Bourhy P, Salaun L, Lajus A, Medigue C, Boursaux-Eude C. A genomic island of the pathogen Leptospira interrogans serovar Lai can excise from its chromosome. Infect Immun 2007;75:677-83.
8. Brenner DJ, Kaufmann AF, Sulzer KR, Steigerwalt AG, Rogers FC, Weyant RS. Further determination of DNA relatedness between serogroups and serovars in the family of Leptospiraceae with a proposal for Leptospira alexanderi sp. nov. and four new Leptospira genomospecies. Int J Syst Bacteriol 1999;49: 839-58.
9. Bulach DM, Zuerner RL, Wilson P, Seemann T, McGrath A, Cullen PA et al. Genome reduction in Leptospira borgpetersenii reflects limited transmission potential. Proc Natl Acad Sci USA 2006;103:14560-5.
10. Canale-Parola E. Motility and chemotaxis of spirochetes. Annu Rev Microbiol 1978;32:69-99.
11. Charon NW, Goldstein SF. Genetics of motility and chemotaxis of a fascinating group of bacteria: The spirochetes. Annu Rev Genet 2002;36:47-73.
12. Chiang SL, Rubin EJ. Construction of a mariner-based transposon for epitope-tagging and genomic targeting. Gene 2002;296:179-85.
13. Choy HA, Kelly MM, Chen TL, Moller AK, Matsunaga J, Hakke DA. Physiological osmotic induction of Leptospira interrogans adhesion: LigA and LigB bind extracellular matrix proteins and fibrinogen. Infect Immun 2007; 75:2441-50.
14. Croda J, Figueira CD, Wunder JEA, Santos CS, Reis MG, Ko AI et al. Targeted mutagenesis in pathogenic Leptospira species: disruption of the LigB gene does not affect virulence in animal models of leptospirosis. Infect Immun 2008;76: 5826-33.
15. Cullen PA, Cordwell SJ, Bulach DM, Hakke DA, Adler B. Global analysis of outer membrane proteins from Leptospira interrogans serovar Lai. Infect Immun 2002;70:2311-8.
16. Cullen PA, Hakke DA, Adler B. Outer membrane proteins of pathogenic spirochetes. FEMS Microbiol Rev 2004;28:291-318.
17. Demarre G, Guerout AM, Matsumoto-Mashimo C, Rowe-Magnus DA, Marliere P, Mazel D. A new family of mobilizable suicide plasmids based on broad host range R388 plasmid (IncW) and RP4 plasmid (IncPα) conjugative machineries and their cognate Escherichia coli host strains. Res Microbiol 2005;156:245-55.
18. Division of Epidemiology, Ministry of Public Health, Thailand. Reported cases and deaths of priority diseases under surveillance. Wkly Epidemiol Surveill Rep 2003-2008;34-9.
19. Division of Epidemiology, Ministry of Public Health, Thailand. Reported cases and deaths of priority diseases under surveillance. Wkly Epidemiol Surveill Rep 2008;39:856.
20. Faine S. Silver staining of spirochetes in single tissue sections. J Clin Pathol 1965;18:381-2.
21. Faine S, Adler B, Bolin C, Perolat P. Leptospira and Leptospirosis. 2nd ed. Melbourne: MediSci; 1999.
22. Guegan R, Camadro J-M, Saint-Girons I, Picardeau M. Leptospira spp. possess a complete haem biosynthetic pathway and are able to use exogenous haem sources. Mol Microbiol 2003; 49:745-54.
23. Haake DA. Spirochaetal lipoproteins and patho-genesis. Microbiology 2000;146:1491-504.
24. Haake DA, Dundoo M, Cader R, Kubak BM, Hartskeerl RA, Sejvar JJ et al. Leptospirosis, water sports, and chemo-prophylaxis. Clin Infect Dis 2002;34:e40-3.
25. Holt SC. Anatomy and chemistry of spirochetes. Microbiol Rev 1978;42:114-60.
26. Indest KJ, Ramamoorthy R, Phillip MT. Transcriptional regulation in spirochetes of medical importance. In: The Spirochetes: mole-cular and cellular biology. Wymondham: Horizon Scientific Press; 2001. p. 159-69.
27. Johnson RC. The spirochetes. Annu Rev Microbiol 1977;31:89-106.
28. Kameni A-PT, Couture-Tosi E, Saint Girons I, Picardeau M. Inactivation of the spirochete recA gene results in a mutant with low viability and irregular nucleoid morphology. J Bacteriol 2002; 184:452-8.
29. Lampe DJ, Akerley BJ, Rubin EJ, Mekalanos JJ, Robertson HM. Hyperactive transposase

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
96
mutants of the Himar1 mariner transposon. Proc Natl Acad Sci USA 1999;96:11428-33.
30. Lee SH, Kim KA, Park YK, Seong IW, Kim MJ. Identification and partial characterization of a novel hemolysin from Leptospira interrogans serovar lai. Gene 2000;254:19-28.
31. Levette PN. Leptospirosis. Clin Microbiol Rev 2001;14:296-326.
32. Li C, Motaleb MA, Sal M, Goldstein SF, Charon NW. Gyrations, rotations, periplasmic flagella: the biology of spirochete motility. In: The spirochetes: molecular and cellular biology. Wymondham: Horizon Scientific Press; 2001. p. 11-22.
33. Lin Y-P, Chang Y-F. A domain of the Leptospira LigB contributes to high affinity binding of fibronectin. Biochem Biophys Res Comm 2007; 362:443-8.
34. Louvel H, Saint Girons I, Picardeau M. Isolation and Characterization of FecA- and FeoB-mediated iron acquisition systems of the spirochete Leptospira biflexa by random inser-tional mutagenesis. J Bacteriol 2005;187:3249-54.
35. Lux R, Moter A, Shi W. The role of chemotaxis in pathogenesis spirochetes. In: The spirochetes: molecular and cellular biology. Wymondham: Horizon Scientific Press; 2001. p. 23-34.
36. Matsunaga J, Barocchi MA, Croda J, Young TA, Sanchez Y, Siqueira I et al. Pathogenic Leptospira species express surface-exposed proteins belonging to the bacterial immuno-globulin superfamily. Mol Microbiol 2003;49: 929-45.
37. McBride AJA, Athanazio DA, Reis MG, Ko AI. Leptospirosis. Curr Opin Infect Dis 2005;18: 376-86.
38. Morgan J, Bornstein SL, Karpati AM. Outbreak of leptospirosis among triathlon participants and community residents in Springfield, Illinois, 1998. Clin Infect Dis 2002;34:1593-9.
39. Motaleb MA, Corum L, Bono JL, Elias AF, Rosa P, Samuels DS et al. Borrelia burgdorferi periplasmic flagella have both skeletal and motility functions. Proc Natl Acad Sci USA 2000;97:10899-904.
40. Murray GL, Ellis KM, Lo M, Adler B. Leptospira interrogans requires a functional heme oxygenase to scavenge iron from hemoglobin. Microbes Infect 2008;10:791-7.
41. Nahori MA, Fournie-Amazouz E, Que-Gewirth NS. Differential TLR recognition of leptospiral lipid A and lipopolysaccharide in murine and human cells. J Immunol 2005;175:6022-31.
42. Nally JE, Artiushin S, Timoney JF. Molecular characterization of thermoinduced immunogenic proteins Qlp42 and Hsp15 of Leptospira inter-rogans. Infect Immun 2001;69:7616-24.
43. Nally JE, Timoney JF, Stevenson B. Tempera-ture-regulated protein synthesis by Leptospira interrogans. Infect Immun 2001;69:400-4.
44. Nascimento ALTO, Ko AI, Martins EAL, Monteiro-Vitorello CB, Ho PL, Hakke DA et al. Comparative genomics of two Leptospira interrogans serovars reveals novel insights into physiology and pathogenesis. J Bacteriol 2004; 186:2164-72.
45. Nougayrede JP, Fernandes PJ, Donenberg MS. Adhesion of enteropathogenic Escherichia coli to host cells. Cell Microbiol 2003;5:359-72.
46. Paster BJ, Dewhirst FE. Phylogenetic founda-tion of spirochetes. In: The Spirochetes: molecular and Cellular Biology. Wymondham: Horizon Scientific Press; 2001. p. 5-9.
47. Pepe JC, Miller VL. Yersinia enterocolitica invasin: a primary role in the initiation of infec-tion. Proc Natl Acad Sci USA 1993;90:6473-7.
48. Picardeau M, Brenot A, Saint Girons I. First evidence for gene replacement in Leptospira spp. inactivation of L. biflexa flaB results in non-motile mutants deficient in endoflagella. Mol Microbiol 2001;40:189-99.
49. Picardeau M, Bauby H, Saint Girons I. Genetic evidence for the existence of two pathways for the biosynthesis of methionine in the Leptospira spp. FEMS Microbiol Lett 2003;225:257-62.
50. Picardeau M. Conjugative transfer between Escherichia coli and Leptospira spp. as a genetic tool. Appl Environ Microbiol 2008;74:319-22.
51. Picardeau M, Bulach DM, Bouchier C, Zuerner RL, Zidane N, Wilson PJ et al. Genome sequence of the saprophyte Leptospira biflexa provides insights into the evolution of Leptospira and the pathogenesis of leptospirosis. PLoS ONE 2008;3:1-9.
52. Plank R, Dean D. Overview of the epide-miology, microbiology, and pathogenesis of Leptospira spp. in humans. Microbes Infect 2000;2:1265-76.
53. Ren SX, Fu G, Jiang XG, Zeng R, Miao YG, Xu H et al. Unique physiological and pathogenic features of Leptospira interrogans revealed by whole-genome sequencing. Nature 2003;422: 888-93.
54. Saint Girons I, Bourhy P, Ottone C, Picardeau M, Yelton D, Hendrix RW et al. The LE1 bacteriophage replicates as a plasmid within Leptospira biflexa: Construction of an

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
97
L. biflexa-Escherichia coli shuttle vector. J Bacteriol 2000; 182:5700-5.
55. Segers RP, van der Drift A, de Nijs A, Corcione P, van der Zeijst BA, Gaastra W. Molecular analysis of a shingomyelinase C gene from Leptospira interrogans serovar hardjo. Infect Immun 1990;58:2177-85.
56. Tangkanakul W, Smits HL, Jatanasen S, Ashford DA. Leptospirosis: An emerging health problem in Thailand. Southeast Asian J Trop Med Public Health 2005;36:281-8.
57. Verma A, Hellwage J, Artiushin S, Zipfel PF, Kraiczy P, Timoney JF et al. LfhA, a novel factor H-biding protein of Leptospira interro-gans. Infect Immun 2006;74:2659-66.
58. Vinetz JM. Leptospirosis. Curr Opin Infect Dis 2001;14:527-38.
59. Watt G. Leptospirosis. In: Strickland GT, editor, Hunter's tropical medicine and emerging infectious diseases. 8th ed. Pensylvania: WB Saunders Company; 2000. p. 453.
60. Wert C, Tapping RI, Mathison JC, Chuang T-H, Kravchenko V, Saint Girons I et al. Leptospiral lipopolysaccharide activates cells through a TLR2-dependent mechanism. Nature Immunol 2001;2:346-52.
61. World Health Organization. Wkly Epidemiol Rec 1999;74:237-42.
62. Zuerner RL, Herrmann JL, Saint Girons I. Comparison of genetic maps for two Leptospira interrogans serovars provides evidence for two chromosomes and interspecies heterogeneity. J Bacteriol 1993;175:5445-51.


99
How Malaria Genes are Regulated
Ruchanok Cheotacha, Thanat Chookajorn, Krittikorn Kumpornsin, Patrath Ponsuwanna and Kanjana Wittayacom Department of Biochemistry, Faculty of Science, Mahidol University.
Abstract
The malaria pathogen has been a great evolutionary force throughout the natural history of mankind. Its footprint on our genome is palpable through the widespread presence of thalassemic traits in the world’s population. It has already been the subject of two Nobel prizes. Still, the dream of eradicating malaria is unrealized. Within the last decade, the parasite’s genome was sequenced, which has made available a plethora of genetic information. Instead of revealing the secrets behind its biology, the parasite’s genetic repertoire is packed with uncommon codes, which cannot be deciphered based on our current knowledge of gene regulation. The aim of this review is to provide a general overview of our understanding of malaria gene regulation and to attract newcomers to join this exciting field.
Keywords: malaria; gene regulation; antigenic variation; rRNA switching
Malaria Genes and Genomes
Plasmodium falciparum, the deadliest malaria pathogen, was identified more than a hundred years ago, and it has been the subject of extensive research ever since. The parasites can switch between mosquito and human hosts. They are also adept in handling challenges from host immune responses, environmental stresses, therapeutic drugs, etc. Hence, it caught many well-informed scientists by surprise to learn that little is known about how the genes of this deadly parasite are regulated, and how the parasites alter their expression patterns to adapt to the human host. Even though P. falciparum displays stage-specific gene expression patterns, the parasite does not show stress-induced changes in its transcription profile. Studies in yeast, plants, and humans showed changes in gene expression to adjust to environmental perturbations. Challenges by stresses such as anti-malarial drugs do not significantly alter malaria gene expression. It is believed that the P. falciparum transcription machinery is ‘hard-wired’ and fundamentally different from those of other organisms(14).
The whole-genome sequencing of P. falciparum and its sibling species has made
possible comparative genome analyses which have revealed similarities in the gene organi-zation among several Plasmodium species(4,15,19). The P. falciparum genome is about 23 Mbp in size and in its blood stage is haploid with fourteen chromosomes. It is estimated that there are approximately 5,400 genes in P. falciparum. One of the distinct features of the P. falciparum genome is its high AT content (>80%)(15). The high AT content obstructs bioinformatic searches for regulatory sequences such as the TATA box. The obscurity of putative cis-regula- tory elements hints at the unique gene control mechanisms of P. falciparum. Successful trans-cription requires cooperation between general and specific transcription factors. General transcription factors comprise the minimum components necessary for the basal level of gene expression. Specific transcription factors act on a limited set of genes to make them express only at certain times and in specific locations. P. falciparum has a complicated life cycle with stage-specific gene expression patterns during its infection of mosquito and human hosts. However, it does not contain close homologues of known specific transcription factors based on several search attempts using sequence similarity algorithms(1,7,15). Callebaut et al.(3)

10
วทยาศาสตรเพ
00
analyzedfalciparucalled tanalysis search identity, such as:subunitsusing ttranscripApicomphomologProteins to bind troles in g
Evmalaria mechaniwith a controllegene regantigen antigeniccontrolleexampledifferentfeature factive ripoints. intriguinmalaria g
Malaria
Mswitchinproteins system. protein erythrocencodedlocated somal rchromosinteractiovarious hin pathcontrollegene famgroups,
พอการพฒนา : รวมบ
d the tranum using atwo-dimensi(HCA). HC
for homoloand several
: TBP, TFII of the TFIIthis methodption factorplexan AP2gue of the
in the ApiAto GC-rich egene regulati
ven though wgene regu
isms in P. faglimpse of
ed. Malariagulation meproteins to c variation med at the trane is a switt stages of Pfor protozoa ibosomal RNOur review
ng examples gene regulati
a Antigenic
Malaria anting the expr
that are ex The family
antigens, yte membra
d by about at subtelom
regions in somes(15). Pons betweenhuman cells
hogenesis(18).ed at the tramily can bnamely: up
บทความวชาการ ฉลอ
nscription a more senional hydroCA is a usefogues with general tranIA, TFIIE, H core have
d(3). Recentr was ide2 (ApiAP2)
plant AP2Ap2 gene famelements in vion are not y
we do not knulation, twalciparum haf how mala parasites uechanism to
avoid host mechanism wnscription letch of ribos
P. falciparumto switch b
NA genes aw will rely
to discuss thion.
c Variation
igenic variression of xposed to th
of highly dPlasmodiu
ane protein 60 var ge
meric and cthirteen ou
PfEMP1 prn infected red
which play Antigenicanscription le divided i
psA, upsB, u
องครบรอบ 50 ป แหง
factors in nsitive progophobic cluful techniqu
low sequescription facTFIIF, and
e been identitly, a specentified ca), which is
2/ERF familmily were shovitro(8), but t
yet clear.
now much abwo well-stud
ave providedaria genes
use an elaboswitch surfdetection(6).
was found tovel(21). Anosomal RNA
m. It is a unibetween exisat different t
on these he complexity
n
ation invovariant surf
he host immivergent surf
um falcipa1 (PfEMP1)
enes which central chromut of fourtoteins med
d blood cells important r
switching level. The nto four m
upsC and up
งการสถาปนาคณะวท
P. gram uster e to ence ctors
the ified cific alled s a ly(8). own their
bout died d us
are orate face . Its o be other A at ique sting time two y of
lves face
mune face rum ), is
are mo-teen
diate and oles
is var
major psE,
basqueencshoacidenc
singgendesis shosurpsterexoneitranmigmu
Fig
speidean hommigteloescsilelati
ทยาศาสตร มหาวทยา
sed on thences(16). Tcodes an extort trans-memdic terminacoded by exo
An indivgle var genene is activesignated to bcalled mutu
own to be coprisingly hasrile RNA tron 2 region(2
ther transponslated in tght play a
utual exclusio
gure 1 Mutuagenesgene directpromopromoparasivar geswitch
Even thecific for thntified, histoimportant r
mologue of ght cause vomeric regiocape of suencing(10). Fion at an acti
าลยมหดล
eir non-codThe first exotracellular bimbrane domaal segment on 2(16).
vidual parase at a time. ely transcribbe silenced(6)
ual exclusionontrolled by ts promoter aranscripts c,9,22). These orted out othe cytoplasphysical rol
on.
ally exclusivs. Parasites exat a time. Th
tion of transoter or bidoter. In the nites can continene using the h to another v
hough a trahe var geneone modificarole in var
yeast histovar gene s
on. Sir2 disruubtelomeric
Furthermore, ive var gene
ding upstreon of each vinding domaain. The cyt
(ATS) do
ite expresse When onlybed, the ot). This phenn (Figure 1the var introactivity and corresponding
sterile transcof the nucsm(11). Henle in the co
ve expressionxpress only a he arrows repcription from
directional vanext erythrocynue to expressmemory mecar gene.
anscription es has not yation is knowgene regula
one deacetylsilencing wiuption result
var genehistone hy
promoter in
eam se-var gene ain and a toplasmic omain is
es only a y one var thers are nomenon ). It is
on, which produces g to the cripts are leus nor
nce, they ontrol of
n of var single var
present the m the var ar intron ytic cycle, s the same chanism or
factor(s) yet been
wn to play ation. A lase Sir2 ithin the ted in the es from ypoacety-n the sub-

telomeriof PfSirthe subteof var unaffecte
Anvar genegene is mbe recogsuch thaagain innism keparasitestypes ofrecognizparasite
Epmajor rHowevemutual designedin mutuscientistsnew theantigenicvulnerab
Ribosom
AnexpressioThe prribosomemachinemoleculeeukaryotrRNA ggene is tare procmoleculeribosomacompone
Intcontain Asexual in mammsites andS-type rrRNA g
c region is r2. Howeveelomeric var
genes neaed by the del
nother exame expressionmarked with gnized by that the mark
n the next cyeps the var s switch surff surface anzed resultingby the host i
pigenetic corole in conr, the exaexclusion
d to identify ual exclusios around theerapies can c variation ble to malaria
mal DNA S
nother impon can be coocess requies to work
ery. Ribosomes and ribotic organism
genes arrangtranscribed icessed into es. The 18Sal subunit, aents of the la
terestingly, Ptwo types
ribosomal gmalian hostsd mating cellribosomal gegenes found
วทยาศาส
correlated wer, the changr genes sincear centromeletion of PfS
mple of epigen memory(5).
epigenetic ce transcriptioed gene wiycle. This mswitching r
face antigensntigens willg in the cimmune syst
ontrol is likntrolling muact mechani
is still unregulatory f
on are beine world with
be developand make
a vaccines.
Switching
portant stepontrolled is prires mRNAtogether in
mes are comosomal prote
ms contain mged in tandeinto rRNA p18S, 5.8S a
S rRNA is pand 28S and arge ribosom
P. falciparums of ribosgenes (A-typs. Sexually-cls in mosquienes. The in P. falcip
สตรเพอการพฒนา :
with the absege is specifice the expresseric regionsSir2(13).
enetic contro An active
codes, which onal machinll be expresmemory mecrate low. If s too quicklyl eventuallyclearing of em(6).
kely to playutual exclusism underlynclear. Stufactors involng pursued h the hope ped to pert
e the paras
p where grotein synthe
A, tRNA, the transla
mposed of rReins. Genera
multiple identem arrays. Eprecursors whand 28S rR
part of the sm5.8S rRNAs
mal subunit(23)
m was foundomal genese) are exprescommitted pitoes expressnumbers of arum are fe
รวมบทความวชาการ
ence c to sion s is
ol is var can
nery, ssed cha-
f the y, all y be
the
y a sion. ying dies lved
by that turb sites
gene esis. and
ation RNA ally, tical
Each hich
RNA mall s are ).
d to s(20). ssed
para- s the
the ewer
tharRNtwoThediffHotrancircthe gamrRNrRNviva
Fig
migmenmenneematrantrancomtranthadiffgluindswirRN
ร ฉลองครบรอบ 50 ป
n those fouNA genes wo genes eache parasites ferent devewever, onenscripts thaculation migh
parasites metocytes(24)
NA was repoNA was fouax oocysts.
gure 2 Stageparasiribosoand thumaand commswitch
It is highght play a rnt. Due to dnts between
ed to dramchinery. Eanslation ofnscripts, anmpensate fonscription fat rRNA swiferent cultu
ucose concenduces gametoitching. TheNA gene is
ป แหงการสถาปนาค
und in otherswere identifieh for A-type
switch betelopmental caveat is th
at are deteht have comcommitted . In additiorted in P. v
und to be hig
-specific rRNites can switchomal RNA gethe S-type. n host propagexpress A
mitted to the h to the S-type
hly possible role in maladrastic differ
its two hostmatically adach type of rRf certain d this rRN
or the lack actors. Recitching could
ure conditionntration in Pocyte formatie transcriptiodecreased af
คณะวทยาศาสตร มหา
s species. Oed in P. faland S-type r
tween two stages (Fig
hat the S-typected in the from the frto become
ion, a third vivax(17). Thighly express
RNA expressh between tw
enes, namely tThe parasite
gate by asexuA-type rRNA
sexual stage,e rRNA expre
that rRNA saria parasite rences in the ts, the parasdapt its trRNA might groups of
NA switchinof known
cently, it wad be observns(12). LoweP. falciparumion and caus
on level of thfter lowering
าวทยาลยมหดล
1
Only four lciparum: rRNA(12). types at
gure 2). pe rRNA he blood raction of e mating
type of is O-type sed in P.
ion. The wo types of the A-type es in the ual mitosis A. Once , parasites ession. switching develop-environ-
ite might ranslation favor the
mRNA ng could
specific as shown ved under ering the m culture ses rRNA he A-type g glucose
01

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
102
concentration and is replaced by S-type rRNA transcripts(12). The switch might coincide with the change from the glucose-enriched blood to the mosquito gut. The mechanism of rRNA switching is not known. The 5′ upstream regions of A-type and S-type rRNA genes are different from one another, which might indicate the binding of different transcription factors(12). It is worth noting that rRNA switching is a novel example of how transcription and translation can work together to regulate gene expression.
Conclusions
Here, we provided a brief summary of the state of the malaria gene regulation field. As one might notice, there is much unknown territory in regard to the study of malaria gene regulation. New findings will likely improve our chance in eradicating the disease and will provide a more complete picture of the transcription machinery in parasitic protozoa. The unique biology of malaria gene regulation reflects the parasite’s complex life cycle and its elaborated survival strategy. The field is wide open for newcomers to make their mark in the fight against one of the deadliest foes of the human race.
References 1. Altschul SF, Gish W, Miller W, Myers EW,
Lipman DJ. Basic local alignment search tool. J Mol Biol 1990;215:403-10.
2. Calderwood MS, Gannoun-Zaki L, Wellems TE, Deitsch KW. Plasmodium falciparum var genes are regulated by two regions with separate promoters, one upstream of the coding region and a second within the intron. J Biol Chem 2003;278:34125-32.
3. Callebaut I, Prat K, Meurice E, Mornon JP, Tomavo S. Prediction of the general transcription factors associated with RNA polymerase II in Plasmodium falciparum: conserved features and differences relative to other eukaryotes. BMC Genomics 2005;6:100.
4. Carlton JM, Adams JH, Silva JC, Bidwell SL, Lorenzi H, Caler E et al. Comparative genomics of the neglected human malaria parasite Plasmodium vivax. Nature 2008;455:757-63.
5. Chookajorn T, Dzikowski R, Frank M, Li F, Jiwani AZ, Hartl DL et al. Epigenetic memory at malaria virulence genes. Proc Natl Acad Sci USA 2007;104:899-902.
6. Chookajorn T, Ponsuwanna P, Cui L. Mutually exclusive var gene expression in the malaria parasite: multiple layers of regulation. Trends Parasitol 2008;24:455-61.
7. Coulson RM, Hall N, Ouzounis CA. Compa-rative genomics of transcriptional control in the human malaria parasite Plasmodium falciparum. Genome Res 2004;14:1548-54.
8. De Silva EK, Gehrke AR, Olszewski K, Leon I, Chahal JS, Bulyk ML et al. Specific DNA-binding by apicomplexan AP2 transcription factors. Proc Natl Acad Sci USA 2008;105: 8393-8.
9. Deitsch KW, Calderwood MS, Wellems TE. Malaria. Cooperative silencing elements in var genes. Nature 2001;412:875-6.
10 Duraisingh MT, Voss TS, Marty AJ, Duffy MF, Good RT, Thompson JK et al. Heterochromatin silencing and locus repositioning linked to regulation of virulence genes in Plasmodium falciparum. Cell 2005;121:13-24.
11. Epp C, Li F, Howitt CA, Chookajorn T, Deitsch KW. Chromatin associated sense and antisense noncoding RNAs are transcribed from the var gene family of virulence genes of the malaria parasite Plasmodium falciparum. Rna 2008;15: 116-27.
12. Fang J, Sullivan M, McCutchan TF. The effects of glucose concentration on the reciprocal regulation of rRNA promoters in Plasmodium falciparum. J Biol Chem 2004;279:720-5.
13. Freitas-Junior LH, Hernandez-Rivas R, Ralph SA, Montiel-Condado D, Ruvalcaba Salazar OK, Rojas-Meza AP et al. Telomeric heterochromatin propagation and histone acetylation control mutually exclusive expression of antigenic variation genes in malaria parasites. Cell 2005; 121:25-36.
14. Ganesan K, Ponmee N, Jiang L, Fowble JW, White J, Kamchonwongpaisan S et al. A genetically hard-wired metabolic transcriptome in Plasmodium falciparum fails to mount protective responses to lethal antifolates. PLoS Pathog 2008;4:e1000214.
15. Gardner MJ, Hall N, Fung E, White O, Berriman M, Hyman RW et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002;419:498-511.
16. Kyes SA, Kraemer SM, Smith JD. Antigenic variation in Plasmodium falciparum: gene organization and regulation of the var multigene family. Eukaryot Cell 2007;6:1511-20.
17. Li J, Gutell RR, Damberger SH, Wirtz RA, Kissinger JC, Rogers MJ et al. Regulation and trafficking of three distinct 18 S ribosomal

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
103
RNAs during development of the malaria parasite. J Mol Biol 1997;269:203-13.
18. Miller LH, Baruch DI, Marsh K, Doumbo OK. The pathogenic basis of malaria. Nature 2002; 415:673-9.
19. Pain A, Bohme U, Berry AE, Mungall K, Finn RD, Jackson AP et al. The genome of the simian and human malaria parasite Plasmodium knowlesi. Nature 2008;455:799-803.
20. Rogers MJ, Gutell RR, Damberger SH, Li J, McConkey GA, Waters AP et al. Structural features of the large subunit rRNA expressed in Plasmodium falciparum sporozoites that dis-tinguish it from the asexually expressed subunit rRNA. Rna 1996;2:134-45.
21. Scherf A, Hernandez-Rivas R, Buffet P, Bottius E, Benatar C, Pouvelle B et al. Antigenic variation in malaria: in situ switching, relaxed and mutually exclusive transcription of var genes during intraerythrocytic development in Plasmo-dium falciparum. Embo J 1998;17:5418-26.
22. Su XZ, Heatwole VM, Wertheimer SP, Guinet F, Herrfeldt JA, Peterson DS et al. The large diverse gene family var encodes proteins involved in cytoadherence and antigenic variation of Plasmodium falciparum-infected erythrocytes. Cell 1995;82:89-100.
23. van Spaendonk RM, Ramesar J, van Wigcheren A, Eling W, Beetsma AL, Van Gemert GJ et al. Functional equivalence of structurally distinct ribosomes in the malaria parasite, Plasmodium berghei. J Biol Chem 2001;276:22638-47.
24. Waters AP, van Spaendonk RM, Ramesar J, Vervenne RA, Dirks RW, Thompson J et al. Species-specific regulation and switching of transcription between stage-specific ribosomal RNA genes in Plasmodium berghei. J Biol Chem 1997;272:3583-9.


105
คณตศาสตรในการจดการคลงยาในโรงพยาบาล
ระว สวรรณเดโชไชย ภาควชาคณตศาสตร คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
ในปจจบนคณตศาสตรมบทบาทอยางแพรหลายในธรกจตางๆ โดยการนาคณตศาสตรไปประยกตใชในการแกปญหาทเกดขนจรง รวมทงการนาไปใชในการจดการคลงยาในโรงพยาบาล ซงมความสาคญเปนอยางยง เนองจากมผลกระทบโดยตรงตอตนทนของโรงพยาบาลและการขาดแคลนยาของผปวย การจดการคลงยาทเหมาะสมจะชวยลดมลคายาทจดเกบในคลง ลดคาใชจายในการสงซอยา เพมประสทธภาพในการใหบรการแกผปวย ลดขยะและลดการทาลายสงแวดลอม การจดการคลงยาทดจะตองอาศยคณตศาสตรในการหาจดสงซอและปรมาณยาทตองสงซอทเหมาะสมเพอใหสอดคลองกบรปแบบของปรมาณการใชยาแบบตางๆ บทความนเสนอตวอยางกรณศกษาโรงพยาบาลของรฐ 1 แหงในกรงเทพมหานคร ซงใชสตรในการคานวณหาจดสงซอและปรมาณการสงซอสาหรบยาชนดตางๆ เพยงสตรเดยว และขอมลทใชในการคานวณหาจดสงซอและปรมาณการสงซออาจไมไดสะทอนถงปรมาณการใชยาทเกดขนจรง หากตองการใหการจดการคลงยามประสทธภาพ สตรทใชในการคานวณควรจะแตกตางกนตามรปแบบของปรมาณความตองการใชยา โรงพยาบาลหลายแหงมการนาโปรแกรมสาเรจรปมาใชในการบรหารจดการคลงยา อยางไรกตาม การประยกตใชโปรแกรมตางๆ ในการจดการคลงยาในโรงพยาบาลเพอใหทางานไดอยางมประสทธภาพจะตองเลอกใชสตรใหสอดคลองกบรปแบบปรมาณการใชยา ในการประยกตใชสตรตางๆ จะตองทาความเขาใจถงขอจากดและทาการศกษาอยางละเอยด หากเลอกสตรไมเหมาะสม ผลลพธจะไมเปนไปตามเปาหมาย
คาสาคญ: การจดการคลงยา; จดสงซอ; ปรมาณสงซอยาทเหมาะสม; รปแบบปรมาณการใชยา
Abstract
Mathematics currently plays an important role in business; especially in applications to real world problems. The article describes how mathematics has been applied to inventory management in a hospital. Managing hospital warehouse is very important as it has a high impact on hospital investment cost and drugs supply. Proper inventory management not only reduces average inventory in warehouse and ordering cost, but also increases the quality of services for patients, reduces waste and helps preserve the environment. The goal of inventory management in a hospital warehouse is to identify reorder points so as to allow the order of quantities that balances effectively demand and supply. This case study at one government hospital in Bangkok, indicated that the hospital considered only information on reorder points and order quantities, so that results did not reflect the real demand. Importantly, only one formula has been used. Different policies should be used for each demand pattern. Although many hospitals have bought ‘Enterprise Resource Planing’ packages to manage their inventory, problems can occur if they do not match each formula provided in the package to the right demand pattern.
Keywords: inventory management; reorder point; order quantity; demand patterns
บทนา การจดการคลงยาในโรงพยาบาลมความสาคญเปน
อยางมาก เนองจากหากมการจดการทไมมประสทธภาพเทาทควรอาจจะทาใหยาบางชนดมไมเพยงพอใหบรการ
แกผปวย หากผปวยเปนโรครายแรงอาจจะทาใหถงแกชวต
ได นอกจากน ยาบางชนดอาจมมากเกนไป ทาใหตองเสยคาใชจายในการเกบรกษาเกนความจาเปนซงสงผลให
ตนทนของโรงพยาบาลสงขน การเกบยามากเกนไปอาจจะทาใหยาบางสวนหมดอายกอนจะนาออกมาใช โดยเฉพาะยาหรอเวชภณฑทมอายคอนขางสน เชน

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
106
tuberculin ทมอายการเกบเพยง 5 เดอน การเกบยามากเกนไปอาจทาใหนาออกมาใชไมทน
ยาบางชนดเมอหมดอาย โรงพยาบาลจะทาการสง คนบรษทยา สวนยาทเหลอจะตองนาไปทาลาย ซงอาจใช บรการของบรษทก าจ ดสาร เคมหรอโรงพยาบาล
ดาเนนการเอง ซงทาไดหลายวธแลวแตชนดของยา เชน นาไปฝงดน เผา หรอละลายนากอนทงลงในบอกาจดนาเสย การทาลายยานอกจากจะทาใหเสยคาใชจายในการท าลายและเสยทรพยากร ย งอาจส งผลกระทบตอ มนษยและสงแวดลอมทงทางตรงและทางออม สรฉตร งอสรเชษฐ และคณะ(1) ไดทาการศกษาผลการดาเนนงานของโรงพยาบาลตางๆ พบวาโรงพยาบาลขนาดใหญสวนใหญจะประสบปญหาขาดทน อยางไรกตาม โรงพยาบาลเหลานสามารถอาศยการบรหารจดการในดานการจดสรร
บคลากรและดานการจดการคลงยาทดมาชวยปรบปรง
แกไขเพอชวยลดตนทนได ดงนนการจดการคลงยาทดจะชวยลดตนทนของโรงพยาบาล ลดมลคายาทจดเกบในคลง เพมประสทธภาพในการใหบรการแกผปวย ลดขยะและลดการทาลายสงแวดลอม
การจดการคลงยาในโรงพยาบาลเปนเรองของฝายบรหารจดการ แลวคณตศาสตรเกยวของกบการจดการคลงยาอยางไร ในความเปนจรงแลว การจดการคลงยาเปนสงทเกยวของกบคณตศาสตรโดยตรง เนองจากการจดการคลงยาจะตองทราบวาควรจะสงซอยาแตละชนดเมอใดและ
ควรสงซอปรมาณเทาใด การวเคราะหขอมลของยาแตละชนดและสรางสตรในการคานวณจดสงซอและปรมาณท
ควรจะสงซอเปนการนาคณตศาสตรมาประยกตใชโดยตรง อยางไรกตาม การนาคณตศาสตรไปประยกตใชในการแกปญหานน นอกจากการนาขอมลมาวเคราะหแลวจะตองศกษาขอจากดตางๆ ทเกดขนดวย
“โครงการวจยและพฒนาการจดการโลจสตกสและโซอปทานตนแบบในอตสาหกรรมบรการ:โรงพยาบาล” เปนโครงการความรวมมอระหวางคณะวทยาศาสตร คณะวศวกรรมศาสตร และคณะเภสชศาสตร มหาวทยาลย มหดล โดยทาการศกษาการจดการโซอปทานและโลจส- ตกสภายในโรงพยาบาลแหงหนง เพอใหไดตนแบบการ
บรหารจดการโลจสตกสและโซอปทานของโรงพยาบาล
ในประเทศไทย งานวจ ยโครงการนสวนหน ง เปนการศกษาการจดการวสดคงคลงยา โดยทาการศกษาคลงยาใหญและคลงยายอยรวมทงสน 7 หอง แบงเปนหองยาผปวยนอกจานวน 5 หอง ไดแก หองยากลาง หองยาหนวยฉกเฉน หองยาตา หองยาสตนารเวช และหองยาเดก และ
หองยาของหอผปวยในจานวน 2 หองซงตางแผนกกน ซงหองยายอยแตละหองรบภาระจายยาใหกบผปวยตามแผนก
ตางๆ ทหองยารบผดชอบ ดงนนหองยายอยแตละหองจะมจานวนและรายการยาทแตกตางกนโดยมเภสชกรแตละ
หองยาเปนผดแล และหองยายอยเหลานจะทาการเบกยาจากคลงยาใหญ
ในการสงซอยา คลงยาจะพจารณาคา Msale หรอ monthly sale ซงเปนคาทใชในการชวยตดสนใจปรมาณการสงซอและจดสงซอ Msale คอ ปรมาณการใชโดยเฉลยในแตละเดอน โดยในทกๆ วน คลงยาใหญจะทาการพจารณาปรมาณยาในคลง หากปรมาณยาในคลงยาใหญตากวา 70% ของ Msale คลงยาจะสงขอมลใหฝายจดซอเพอท าการตดต อบรษทยาและด า เนนการส งซ อ โดยกาหนดการสงซอแบบเตมเตม นนคอ สงซอเพอใหปรมาณยารวมในคลงยามประมาณ 100-150% ของ Msale คาของ Msale จะมการปรบทกๆ เดอน เพอใหสอดคลองกบปรมาณความตองการยา โดย Msale ในเดอนปจจบนจะคานวณไดจากคาเฉลยระหวาง Msale ในเดอนทผานมากบคาเฉลยของปรมาณการใชยาในเดอนทผานมา ดงน
2-nD-nMsale
nMsale 11+=
โดย Msalen แทนคา Msale ในเดอนท n และ D n แทน คาเฉลยของปรมาณการใชในเดอนท n จะเหนไดวาทาง
โรงพยาบาลใหการสงซอแบบ (s,S) โดย s แทนระดบของการสงซอซงมคาเทากบ 70% ของ Msale และ S แทนจดเตมเตมซงคอระดบของปรมาณยาในคลงเมอทาการเตม
เตมแลว หากกาหนดให Q แทนปรมาณการสงซอ ดงนน
ปรมาณการสงซอ Q = S-s
นโยบายของโรงพยาบาลในการสงซอยา คอ ในทกๆ วนจะตรวจสอบปรมาณยาในคลง ถาปรมาณยาในคลงลดลงตากวา s จะทาการสงซอดวยปรมาณ Q
ปญหาทเกดขนในคลงยาใหญ คอ สตรการคานวณ หาจดสงซอและปรมาณการสงซอมเพยงสตรเดยวซงคด
จากคา Msale และการปรบคา Msale ทาเพยงเดอนละครง ดงนนหากปรมาณความตองการเปลยนไป คา Msale จะไมปรบตามจนถงเดอนถดไป จงสงผลใหเกดปญหาปรมาณยาในคลงใหญเนองจากพนทการจดเกบยามคอนขางจากด และยาบางชนดอาจมการสงซอมากเกนไปทาใหใชไมทน สงผลใหมยาบางสวนหมดอายและจาเปนตองทาลายทง ในขณะเดยวกนยาบางชนดมปรมาณไมเพยงพอตอความ
ตองการใช หากพจารณาจากสตรท ใชพบวา Msale

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
107
เปรยบเสมอนคาการพยากรณปรมาณการใชในเดอนหนา
โดยใชขอมลของความตองการใชยาตงแตมการเรมใชยา
จนถงการใชยาในเดอนทผานมาและเปนการพยากรณแบบ
ถวงนาหนก
( ) ( ) ( ) ∑−
−−++−
+=
==1
0111
10
n
i iDnwnD21Dn
21Dn
21nMsale L
โดย ( ) ininw −
− = 21
การสงซอยาในโรงพยาบาลของรฐมหลายขนตอน โดยเฉลยจะใชเวลาประมาณ 10-12 วน คอใชเวลาในการขออนมตการสงซอ 5-7 วน จากนนอกประมาณ 5 วนสาหรบการจดสง ดงนนการคานวณจดสงซอยาจงม
ความสาคญสาหรบโรงพยาบาลของรฐหรอโรงพยาบาลท
ใชเวลาในการสงซอคอนขางนานมาก หากสงซอยาชาเกนไปอาจทาใหยาขาดหรอมยาไมเพยงพอใหบรการ
ในชวงการสงซอ หรอหากสงซอยาเรวเกนไปจะทาใหส น เปล องค า ใช จ า ย ด ง นน การจ ดการคล งย าในโรงพยาบาลของรฐจะตองคานงถงการพยากรณความ
ตองการลวงหนาเพอใชในการคานวณจดสงซอ เนองจากใชเวลาในการสงซอทคอนขางนานดงกลาว ซงตางจากโรงพยาบาลเอกชนบางแหงทโดยปกตจะใชเวลาการสงซอ
ยาเพยง 1 วน และในกรณเรงดวนสามารถไดรบยาภายในวนทสงซอได
รปท 1 แสดงถงการเชอมโยงในการระบบการซอขายยา ระหวาง คนไข คลงยายอย คลงยาใหญ ฝายจด ซอและบรษทยา โดยแสดงการไหลของยาและการไหลของขอมล การเบกยาของหองยายอยจะคานวณจาก
ปรมาณการซอยาของคนไขในแตละหอง และปรมาณการสงซอยาโดยคลงยาใหญจากบรษทยาจะใชตวเลขของ
ปรมาณการเบกยาของหองยายอย บรษทจะทาการจดหายาโดยใชขอมลการซอยาจากโรงพยาบาลตางๆ จะเหนไดวาการทคลงยาใหญและบรษทยาไมทราบปรมาณการใชจรง
ทาใหเกดปรากฏการณ bullwhip effect ซง Lee และคณะ(3) อธบายไววาเปนปรากฏการณทคลงยายอยสงยาเผอโดยม
การเกบมลภณฑนรภย (safety stock) ไวสวนหนง
เนองจากปรมาณการใชยามความไมแนนอนในแตวน ในขณะเดยวกนคลงยาใหญกจะสงยาจากบรษทโดยเกบมล
ภณฑนรภยเพมขนอก ซงในโซอปทานของยานหากผทเกยวของอยหางจากผบรโภคมากเทาไหร จะพบวาขอมลปรมาณความตองการจะแปรปรวนมากทาใหตองมการเกบ
มลภณฑนรภยมากขนซงจะทาใหตนทนของผเกยวของทก
ภาคสวนเพมขนตามกนไปดวย การศกษาของ Eppen(2) บงชวาคาใชจายเหลานสามารถลดลงไดหากคลงยาใหญ
สามารถทราบถงปรมาณความตองการใชจรงจากคลงยา
ยอย และหากทางบรษทยาสามารถทราบถงขอมลปรมาณความตองการใชจรงของผปวยจะทาใหคาใชจายของ
บรษทลดลงดวยเชนกน การคดสตรในการคานวณหาจดสงซอและปรมาณการใชยาทเหมาะสมจงตองเรมจาก
การศกษารปแบบความตองการใชยาท เ กดขนจร ง เนองจากยาในโรงพยาบาลมไมตากวา 2,000 ชนด การทจะศกษารปแบบของความตองการยาทกชนดจงทาได
คอนขางยาก ดงนน จงตองทาการจาแนกยาออกเปนกลมๆ เพอใหงายตอการบรหารจดการและเพอเลอกชนดของยาท
จะศกษา
รปท 1 การเชอมโยงในการระบบการซอขายยา แสดงการไหลของยาและ แสดงการไหลของขอมล
คนไข
คลงยาใหญ
คลงยายอย
บรษทยา
คลงยายอย คลงยายอย คนไข คนไข
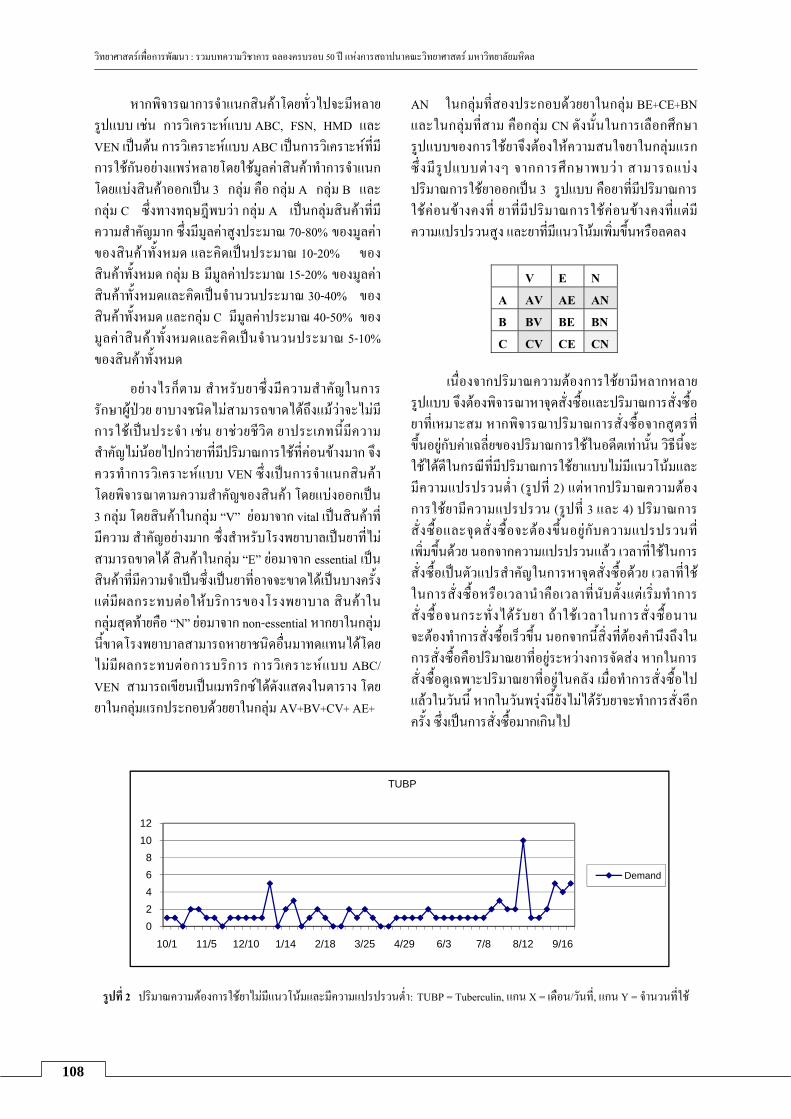
วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
108
02468
1012
10/1 11/5 12/10 1/14 2/18 3/25 4/29 6/3 7/8 8/12 9/16
TUBP
Demand
หากพจารณาการจาแนกสนคาโดยทวไปจะมหลาย
รปแบบ เชน การวเคราะหแบบ ABC, FSN, HMD และVEN เปนตน การวเคราะหแบบ ABC เปนการวเคราะหทมการใชกนอยางแพรหลายโดยใชมลคาสนคาทาการจาแนก โดยแบงสนคาออกเปน 3 กลม คอ กลม A กลม B และกลม C ซงทางทฤษฎพบวา กลม A เปนกลมสนคาทม
ความสาคญมาก ซงมมลคาสงประมาณ 70-80% ของมลคาของสนคาทงหมด และคดเปนประมาณ 10-20% ของ
สนคาทงหมด กลม B มมลคาประมาณ 15-20% ของมลคาสนคาทงหมดและคดเปนจานวนประมาณ 30-40% ของ
สนคาทงหมด และกลม C มมลคาประมาณ 40-50% ของมลคาสนคาทงหมดและคดเปนจานวนประมาณ 5-10% ของสนคาทงหมด
อยางไรกตาม สาหรบยาซงมความสาคญในการรกษาผปวย ยาบางชนดไมสามารถขาดไดถงแมวาจะไมมการใชเปนประจา เชน ยาชวยชวต ยาประเภทนมความ สาคญไมนอยไปกวายาทมปรมาณการใชทคอนขางมาก จงควรทาการวเคราะหแบบ VEN ซงเปนการจาแนกสนคา โดยพจารณาตามความสาคญของสนคา โดยแบงออกเปน 3 กลม โดยสนคาในกลม “V” ยอมาจาก vital เปนสนคาทมความ สาคญอยางมาก ซงสาหรบโรงพยาบาลเปนยาทไมสามารถขาดได สนคาในกลม “E” ยอมาจาก essential เปนสนคาทมความจาเปนซงเปนยาทอาจจะขาดไดเปนบางครง แตมผลกระทบตอใหบรการของโรงพยาบาล สนคาในกลมสดทายคอ “N” ยอมาจาก non-essential หากยาในกลมนขาดโรงพยาบาลสามารถหายาชนดอนมาทดแทนไดโดย
ไมมผลกระทบตอการบรการ การวเคราะหแบบ ABC/ VEN สามารถเขยนเปนเมทรกซไดดงแสดงในตาราง โดยยาในกลมแรกประกอบดวยยาในกลม AV+BV+CV+ AE+
AN ในกลมทสองประกอบดวยยาในกลม BE+CE+BN และในกลมทสาม คอกลม CN ดงนนในการเลอกศกษารปแบบของการใชยาจงตองใหความสนใจยาในกลมแรก ซงมรปแบบตางๆ จากการศกษาพบวา สามารถแบงปรมาณการใชยาออกเปน 3 รปแบบ คอยาทมปรมาณการใชคอนขางคงท ยาทมปรมาณการใชคอนขางคงทแตม
ความแปรปรวนสง และยาทมแนวโนมเพมขนหรอลดลง
V E N A AV AE AN B BV BE BN C CV CE CN
เนองจากปรมาณความตองการใชยามหลากหลาย
รปแบบ จงตองพจารณาหาจดสงซอและปรมาณการสงซอยาทเหมาะสม หากพจารณาปรมาณการสงซอจากสตรทขนอยกบคาเฉลยของปรมาณการใชในอดตเทานน วธนจะใชไดดในกรณทมปรมาณการใชยาแบบไมมแนวโนมและ
มความแปรปรวนตา (รปท 2) แตหากปรมาณความตอง การใชยามความแปรปรวน (รปท 3 และ 4) ปรมาณการสงซอและจดสงซอจะตองขนอยกบความแปรปรวนท
เพมขนดวย นอกจากความแปรปรวนแลว เวลาทใชในการสงซอเปนตวแปรสาคญในการหาจดสงซอดวย เวลาทใชในการสงซอหรอเวลานาคอเวลาทนบตงแตเรมทาการ
สงซอจนกระทงไดรบยา ถาใชเวลาในการสงซอนานจะตองทาการสงซอเรวขน นอกจากนสงทตองคานงถงในการสงซอคอปรมาณยาทอยระหวางการจดสง หากในการสงซอดเฉพาะปรมาณยาทอยในคลง เมอทาการสงซอไปแลวในวนน หากในวนพรงนยงไมไดรบยาจะทาการสงอกครง ซงเปนการสงซอมากเกนไป
รปท 2 ปรมาณความตองการใชยาไมมแนวโนมและมความแปรปรวนตา: TUBP = Tuberculin, แกน X = เดอน/วนท, แกน Y = จานวนทใช

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
109
0
500
1000
1500
2000
10/1 11/1 12/1 1/1 2/1 3/1 4/1 5/1 6/1 7/1 8/1 9/1
DORM
Demand
0
20000
40000
60000
80000
100000
120000
10/1 11/20 1/9 2/28 4/19 6/8 7/28 9/16
OMPZ
Actual
Linear (Actual)
รปท 3 ปรมาณความตองการใชยาไมมแนวโนมแตมความแปรปรวนสง: DORM = Dormicum, แกน X = เดอน/วนท, แกน Y = จานวนทใช
รปท 4 ปรมาณความตองการใชยามแนวโนมและมความแปรปรวนสง: OMPZ = Omeprazole, แกน X = เดอน/วนท, แกน Y = จานวนทใช
กลาวโดยสรป ปรมาณการสงซอและจดสงซอยาแตละชนดควรจะขนอยกบคาเฉลยและความแปรปรวนของ
ปรมาณความตองการใชยา เวลาท ใช ในการส งซ อ แนวโนมของปรมาณความตองการใชยาในอนาคต และปรมาณยาทอยระหวางการจดสง การคานวณปรมาณการสงซอและจดสงซอยาควรจะทาการปรบคาทกวนและ
ขอมลทใชในการคานวณควรจะตองสะทอนถงปรมาณการ
ใชจรง การสรางสตรในการคานวณปรมาณการสงซอและหาจดสงซอยาทสามารถนาไปใชไดจรง ควรจะกาหนดเปาหมายอยางชดเจนและจะตองไมซบซอนมากเพอความ
สะดวกรวดเรวในการคานวณเมอนาไปใชจรง เนองจากยาทใชในโรงพยาบาลมเปนจานวนมาก นอกจากนขอมลทใชควรสะทอนปรมาณความตองการทแทจรง
เนองจากรปแบบความตองการของการใชยาทม 3 รปแบบดงแสดงในรปท 2-4 จงสามารถทาการสรางสตร
การคานวณสาหรบยาในรปแบบตางๆ โดยกาหนดให r แทนจดสงซอ μ และ σ2 แทนคาเฉลยและความแปรปรวน
ของปรมาณความตองการใชยาในแตละวน L แทนเวลานา
N แทนเวลาทตองการเกบยาไวในคลง S แทนการสงซอสาหรบปรมาณความตองการททราบลวงหนา (แบบพเศษ) และ y แทนปรมาณยาทกาลงดาเนนการจดสง k แทนระดบการใหบรการโดยท 0 < k < 1 เมอ k เขาใกล 1 หมายถง อตราความตองการยาของผปวยทงหมดไดรบยา min order แทนปรมาณตาสดททางบรษทยากาหนดในการสงซอแต
ละครง P (DL ≤ r) แทนความนาจะเปน
สตรท 1: สาหรบปรมาณการใชยาไมมแนวโนมและมความแปรปรวนตา
r = μL
order}min,boxperunit
ySNmax{Q -+=
μ สตรท 2: สาหรบปรมาณการใชยาไมมแนวโนมแต
มความแปรปรวนสง P (DL ≤ r) = k
order}min,boxperunit
ySNmax{Q -+=
μ

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
110
โดย DLแทนตวแปรสมของปรมาณความตองการ
ในระหวางเวลานา ซงมคาเฉลยเทากบ μ(L+1) และคา
แปรปรวนเทากบ (L+1)σ2
สตรท 3: สาหรบปรมาณการใชยาทมแนวโนมและมความแปรปรวนสง
P (DL ≤ r) = k P (DN ≤ Y) = k
order}min,boxperunitySYmax{Q -+
=
สาหรบปรมาณความตองการทมแนวโนมจะหา
ปรมาณการใชยาในอนาคต ใชสมการการถดถอยเชงเสนในการพยากรณ ซงใชสญลกษณ f(x) แทนคาพยากรณ
สาหรบวนท x bxaf(x) ˆˆ +=
โดยท a และ b แทนพารามเตอรของสมการถดถอยเชงเสนซงคานวณไดจากขอมลในอดต M วน DL แทนปรมาณความตองการใชยาลวงหนาในชวงเวลานา มคาเฉลยเทากบ
∑+
=
LM
Mii )f(x และคาความแปรปรวน (L+1)MSE และ DN
แทนปรมาณความตองการลวงหนา N วน ซงมการแจกแจง
แบบปกตและมคาเฉลยเทากบ ∑+
=
NM
Mii )f(x และคาความ
แปรปรวน N*MSE
นโยบายในการสงซอยา
กาหนดให I แทนปรมาณยาในคลง ถา I +y < r จะทาการสงซอดวยปรมาณ Q
ผลจากการวเคราะหเชงตวเลขแสดงใหเหนถง
ประสทธภาพของสตรและนโยบายทเสนอแนะ นอกจากนย งพบว าหากสามารถลด เวลาในการส งซ อ ย า ใน
โรงพยาบาลของรฐจะสามารถลดมลคาสนคาในคลง
ไดมากขน นอกจากน ขอมลของยาบางชนดชใหเหนวาปรมาณการใชยาบางชนดสามารถทราบลวงหนาได เชน การใชยาฉดใหกบนกศกษาแพทยกอนขนหอผปวยในชวง
การเรยนการสอน การทแพทยนดคนไขทตองการใชยาเปนจานวนมากใหมารบยาทกๆ เดอน เปนตน ดงนนในการคานวณปรมาณการสงซอ หากแยกขอมลของปรมาณความตองการใชยาททราบลวงหนากบปรมาณความตองการใช
ยาทมความไมแนนอนจะทาใหผลทไดมความแมนยามาก
ขน
โรงพยาบาลหลายแหง ไดนาโปรแกรมสาเรจรปสาหรบระบบ ERP (Enterprise Resource Planning) เขามาใชในการจดระบบการจดซอยา ERP เปนเครองมอทนามา ใชในการบรหารธรกจ สาหรบการวางแผนการและบรหารทรพยากรตางๆ โดยรวมระบบการจดซอจดจาง การผลต การขาย การบญช และการบรหารบคคลเขาดวยกนและเชอมโยงขอมลตางๆ อยางทนททนใด การซอโปรแกรมสาเรจรปสาหรบระบบ ERP ทงระบบเพอใชในการบรหารคลงยาใหญและคลงยายอยจะเสยคาใชจายสงมากไมคมคา
กบการลงทน โรงพยาบาลสามารถลดคาใชจายโดยการซอโปรแกรมสาเรจรปเฉพาะสวนทเกยวของกบการจดการ
สนคาคงคลงสาหรบคลงยาใหญได
อยางไรกตาม การใชโปรแกรมสาหรบคลงยาใหญเพยงแหงเดยวอาจทาใหเกดปญหาในการจดซอสาหรบ
หองยายอยเนองจากโปรแกรม ERP เปนโปรแกรมทสรางสาหรบการบรหารธรกจทวไป เมอนามาใชกบโรงพยาบาลจะตองทาการปรบโปรแกรมทมอยใหเขากบระบบทางาน
ของโรงพยาบาล สตรการคานวณหาปรมาณการสงซอและจดสงซอในโปรแกรม ERP มหลายรปแบบ การเลอกใชสตรเพยงสตรเดยวอาจจะไมสอดคลองกบปรมาณความ
ตองการใชยาสาหรบยาทกชนด ดงนน การเลอกสตรตางๆ ใหเหมาะสมกบรปแบบปรมาณความตองการใชยาควร
ตองพจารณาอยางระมดระวง ทางเภสชกรสามารถเปนผตดสนใจเนองจากทราบถงรปแบบการใชยาแตละชนดเปน
อยางด อยางไรกตามในการประยกตใชสตรควรจะตองทาการเขาใจถงขอจากดของสตรตางๆ อยางละเอยดเสยกอน หากเลอกสตรทไมเหมาะสมผลลพธจะไมเปนไปตาม
เปาหมาย
นอกจากระบบ ERP ทใชในการจดการภายใน
โรงพยาบาลแลว องคการเภสชกรรมซงเปนหนงในบรษทยาไดเรมใชระบบ VMI (Vendor Managed Inventory) ซงเปนระบบทผขายสนคาเขาไปชวยดแลคลงยาของลกคา ในการน องคการเภสชกรรมจะเขาไปชวยดแลคลงยาใหกบโรงพยาบาลผานระบบคอมพวเตอร เพอใหโรงพยาบาลมปรมาณยาทเหมาะสม โดยจะนายาไปเตมเตมคลงยาในโรง พยาบาลเมอถงจดสงซอ ในทางปฏบต โรงพยาบาลจะสงขอมลปรมาณยาชนดตางๆ ในคลง ททาสญญากบองคการเภสชกรรม ในทางทฤษฏนนการททางโรงพยาบาลไมตองบรหารคลงยาเองทาใหตนทนในการสงซอสนคาและความ
ผดพลาดเกยวกบการสงขอมลลดนอยลง การจดซอทาไดสะดวกและรวดเรวมากขน ลดเวลาในการจดซอ นอกจาก นการทองคการเภสชกรรมสามารถทราบขอมลปรมาณยา

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
111
ในคลงของโรงพยาบาลทาใหสามารถทาการพยากรณ
ความตองการยา เตรยมพรอมในวางแผนการผลตและแผนการกระจายสนคา ชวยลดการขาดแคลนยาได ทาใหประสทธภาพในการบรการยาแกผปวยสงขน ในทาง
ปฏบตระบบ VMI จะเปนไปไดอยางมประสทธภาพหาก
ปรมาณยาในคลงสะทอนถงปรมาณความตองการยาจรง ดงนน หากองคการเภสชกรรมเหนปรมาณยาในคลงใหญเพยงแหงเดยวการเพมหรอลดของยาในคลงจะไมสะทอน
ถงความตองการใชจรง หากตองการทราบปรมาณความตองการยาทแทจรงควรจะใชปรมาณยาในคลงรวมทงหมด
จงเหนไดวาในปจจบนการจดการดานโลจสตกส
และหวงโซอปทานอยางมประสทธภาพเปนปจจยสาคญท
จะทาใหธรกจตางๆ ดาเนนไปได ซงในการดาเนนการตองเรมตงแตตนนาถงปลายนา นนกคอตงแตผผลตจนถงผบรโภค ตองมกระบวนการวางแผนการผลต การจดหาวตถดบ การจดการสนคาคงคลงอยางมประสทธภาพและสอดคลองกบความตองการของลกคา คณตศาสตรสามารถนาไปประยกตใชในกระบวนการตางๆ เหลาน ซงบทบาทของคณตศาสตรประยกตในการจดการคลงยาเปนเพยง
ตวอยางหนงเทานน
สรป คณตศาสตรประยกตมบทบาททสามารถนาไปใช
ในการจดการคลงยาในโรงพยาบาลเพอลดตนทนของ
โรงพยาบาล ลดมลคายาทจดเกบในคลง ลดคาใชจายในการสงซอยา เพมประสทธภาพในการใหบรการแกผปวย ลดขยะและลดการทาลายสงแวดลอม บทความนไดยกตวอยางกรณศกษาของโรงพยาบาลของรฐแหงหนง โดยไดพจารณาคลงยาใหญและหองยายอย การคานวณจดสงซอและปรมาณการสงซอใชตวเลขจากการเบกยาของ
หองยายอยซงมไดสะทอนถงปรมาณความตองการท
แทจรง
สตรทใชในการคานวณจดสงซอและปรมาณการ
สงซอมเพยงสตรเดยวและขนอยกบคาเฉลยของการเบกยา
ของแตละเดอนเทานน ในการสรางสตรทางคณตศาสตรในการหาจดสงซอและปรมาณการสงซอท เหมาะสม
จะตองศกษาถงองคประกอบตางๆ ทมผลตอจดสงซอและปรมาณการสงซอ เนองจากยาทหมนเวยนในคลงมไมตากวา 2,000 ชนด การเลอกกลมยาทจะศกษาไดจากการวเคราะหแบบ ABC/VEN ในการจดกลมยา รปแบบปรมาณความตองการใชยาสามารถแบงออกเปน 3 กรณ คอ ความตองการใชยาแบบไมมแนวโนมและมความแปรปรวนตา ความตองการใชยาแบบไมมแนวโนมแตม
ความแปรปรวนสง และความตองการใชยาแบบมแนวโนมและมความแปรปรวนสง คณะผวจยจงเสนอสตรในการคานวณจดส งซอและปรมาณการส งซอท เหมาะสม สอดคลองกบรปแบบความตองการใชยาซงสรางจาก
แนวโนมของปรมาณความตองการใชยา คาเฉลยความแปรปรวนและเวลาทใชในการสงซอยา ในปจจบนโปรแกรมระบบ ERP และ VMI ไดถกนามาใชในการจดการคลงยาในโรงพยาบาล อยางไรตาม การเลอกใชสตรการคานวณจะตองทาการศกษาอยางละเอยดและเขาใจ
ขอจากดของสตรตางๆ เพอทาการเลอกใหเหมาะสมกบรปแบบความตองการใชยาจรง
กตตกรรมประกาศ ขอขอบคณบคลากรฝายคลงยาใหญและฝายจดซอ
ของโรงพยาบาลททาการศกษาทไดใหขอมลตางๆ และ รศ. ดร.ดวงพรรณ กรชชาญชย ศฤงคารนทร ศนยการจดการโลจสตกส หวหนาโครงการวจยทผเขยนไดมสวนรวม
เอกสารอางอง 1. สรฉตร งอสรเชษฐ, อรรถพล ศรเลศลาวาณช. การศกษา
ลกษณะของโรงพยาบาลในประเทศไทยทมผลดาเนนการ
ขาดทนหรอกาไรหลกจากปแรกของการมหลกประกนสขภาพ
ถ วนหน า . สถาบนว จ ย ระบบสาธารณสข กระทรวงสาธารณสข; 2547.
2 Eppen GD. Effects of centralization on expected costs in a multi-location newsboy problem. Manage Sci 1979;25:498-501.
3. Lee HL, Padmanabhan V, Whang S. Information distortion in a supply chain: the bullwhip effect. Manage Sci 1997;43:546-58.


113
การรกษาผวหนงอกเสบระคายสมผส
วรรณจรส รงพสทธพงษ1 และ ธรวฒ คลงเจรญชย2 1ภาควชาอายรศาสตร วทยาลยแพทยศาสตรกรงเทพมหานครและวชรพยาบาล
2ภาควชาเภสชวทยา คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
โรคผวหนงอกเสบระคายสมผสมสาเหตเกยวเนองกบการทางานถงรอยละ 80 ของโรคผวหนงอกเสบจากการ
สมผสทงหมด การรกษาจาเปนตองทราบสารทกอใหเกดการระคายเคองและหลกเลยงการสมผสสารนน ในเบองตน ตองลางบรเวณทสมผสดวยนาซงเจอจางดวยกรดหรอดางออนทมฤทธตรงขามกบสารทกอการระคายสมผส ยาทาทนยมและใชบอย ไดแก คอรตโคสตรอยด เนองจากเปนยาทมประสทธภาพ ในกรณทผวหนงอกเสบเรอรงหรอรนแรงอาจบาบดดวยแสง หรอใชยากดภมคมกน
คาสาคญ: ผวหนงอกเสบระคายสมผส; ยาทาคอรตโคสตรอยด
Abstract
Irritant contact dermatitis is a common occupational-related and accounts for 80 percents of all cases of contact dermatitis. Identification and avoidance of potential irritants are the mainstay of treatment. Early treatment is to wash with large amounts of water and weak acid or base depending on type of irritant. Topical corticosteroid is effective and commonly used in irritant contact dermatitis. In severe or chronic cases, phototherapy and immunosuppressive drugs may be effective.
Keywords: irritant contact dermatitis; topical corticosteroid
บทนา
โรคผ วหน ง อ ก เสบจ ากการส มผ ส (contact dermatitis) แบงเปน 2 ประเภท คอ ผวหนงอกเสบระคายสมผส (irritant contact dermatitis) และผวหนงอกเสบ
ภมแพสมผส (allergic contact dermatitis) บทความน
กลาวถงโรคผวหนงอกเสบระคายสมผส เนองจากพบไดบอยถงรอยละ 80 ของโรคผวหนงอกเสบจากการสมผส
ทงหมด โดยพบไดบอยในบคลากรททางานทางสายวทยาศาสตรและดานอตสาหกรรมเนองจากตองทางาน
เก ยวกบสารเคม การหาสาเหตและหลกเล ยงสารทกอใหเกดการระคายเคองและแพจงเปนสงสาคญ แต
บางครงกไมสามารถหลกเลยงได ดงนนเมอตองทางานทเกยวของกบสารเคม จงจาเปน ตองใชอปกรณปองกน หากมการสมผสจาเปนตองรกษาหรอบรรเทาอาการเบองตน
โรคผวหนงอกเสบจากการสมผส โรคผวหนงอกเสบระคายสมผสสามารถเกดไดใน
การสมผสสารเปนครงแรกโดยไมตองอาศยปฏกรยาผาน
ระบบภมคมกน อาจเกดในเวลาเปนนาท หรอใน 2-3 ชวโมงหลงสมผสสาร ตอมาผวหนงอาจอกเสบรนแรงจนเปอยยย สาหรบผปวยทมผวบอบบางจะเกดอาการทรนแรงกวาผทมผวหนาได ความรนแรงยงขนกบระยะเวลาทสมผสสารและความเขมขนของสาร อาการเดน คอ รสกเจบปวดและแสบรอน(8) สารททาใหเกดการระคายสมผสชนดนไดแก
1. สารทมฤทธ เปนดาง เชน สบ นายาทาความสะอาดเตาอบ และนายาลางหองนา เปนตน การใชกรดออนๆ ประเภทนาสมสายช นามะนาว หรอกรดมดออนๆ ลางบรเวณผวหนงทสมผสสาร จะชวยบรรเทาความรนแรงลงได(8)
2. สารทมฤทธเปนกรด เชน กรดไฮโดรฟลออรก ซงใชในการละลายตะกรนในทอประปา เปนสวนผสมในนายาฆาเชอโรค ใชอตสาหกรรมพลาสตก และงานแกะ สลกแกว เปนตน ทาใหเนอเกดแผลและเนาได กรดไนตรก ซงมฤทธการกดกรอนสง ทาใหเนอเยอเปนสเหลอง

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
114
กรดซลฟรก ซงใชในวงการอตสาหกรรมอยางแพร หลาย ทาใหผวหนงไหมเปนสนาตาล กรดออกซาลกทาใหปลายนวชา เขยวและเนาได และยงทาใหเลบเปนสเหลอง วธการสะเทนกรดขางตนทดทสดคอ การใช milk of mag-nesia สาหรบ ฟนอล หรอ กรดคารบอลก ซงในปจจบน มการนามาใชในการลอกหนาเพอความงามกนคอนขาง
แพรหลาย ทาใหเกดรอยดางขาวบนผวหนง ไตอกเสบ หวใจเตนผดจงหวะ และเกดอาการชาเปนบางครงบางคราว หากสะเทนดวย 65% ethyl หรอ isopropyl alcohol จะชวยได
นอกจากนยงมสารทมฤทธเปนกรดอกหลายชนดท
สามารถทาใหเกดผวหนงอกเสบเชนนได ในการรกษาใหลางดวยนาปรมาณมากๆ ทนท และลางตอดวยสารทมฤทธเปนดาง เชน sodium bicarbonate, calcium hydroxide และนาสบ(8)
3. สารเคมบางชนด ตองใชการดแลพเศษ เชน fluorine ใหสะเทนดวย magnesium oxide สาหรบกรณทมรอยไหมบรเวณรอบๆ เลบ ควรใช 10% calcium gluconate เพอลดความรนแรงของเนอเยอทจะถกทาลาย กรดไฮโดร-ฟลออรก ทาใหระดบแคลเซยม แมกนเซยม และโพแทส- เซยมในเลอดตาลงได หากเปนฟอสฟอรส ควรใชนา จานวนมากลางออกและใช copper sulfate รวมดวย แตถาสมผสสาร titanium hydrochloride หากใชนาลางจะทาใหเกดการเผาไหมทรนแรงขน ดงนนจงควรใชการเชดสารออกเทานน(8)
4. ผวหนงอกเสบจากถงลมนรภย เมอมการเบรกรถยนตอยางกระทนหน สาร sodium azide และ supric oxide ทอยในหลอดเลกๆ ทขบเคลอนไดจะถกกระตนและปลอยแกสไนโตรเจนออกมา ทาใหถงลมนรภยพองตวออกในอตรา 160 กโลเมตรตอชวโมง สารตางๆ ภายในไดแก ผงแปง sodium hydroxide และ sodium carbonate จะถกปลอยออกมาในถงลม การเสยดสและสมผสกบเคมเหลานจะเกดผวหนงอกเสบระคายสมผสได การใชยาทา สตรอยดอาจไดผลดกบการอกเสบแดงตนๆ แตหากเปนลกษณะผวหนงเปนรอยไหมนนหนา อาจตองรกษาดวยการผาตด และการปลกถายเนอเยอเพมเตม(2)
5. ผวหนงอกเสบจากใยแกว พบในผปวยทมอาชพทเกยวของกบใยแกว ซงมทใชในอตสาหกรรม แผนมาน ฉนวนความรอนในรถยนต ถงเชอเพลงและยานอวกาศเปนตน ผปวยมประวตสมผสโดยขาดความระมดระวง ใยแกวเลกๆจะซมผานผวหนงทาใหผวหนงอกเสบ มลกษณะนนแดง มรองรอยเกาและคนมาก ในกรณทรนแรงจะเกดรขม
ขนอกเสบลกษณะเปนผน บางครงคนมากจนมลกษณะ
คลายกบตดเชอหดหรอถกตวไรกด การรกษาเบองตน ตองลางบรเวณทสมผสดวยนาปรมาณมากๆ การทาแปงตามบรเวณรอยพบตางๆ บนรางกายจะทาใหใยแกวไมเกาะตดตามผวหนง ในการทดสอบเพอประเมนคนงานทจะเรมทางานเกยวของกบอตสาหกรรมใยแกวควรใชแผนแปะตอ epoxy resins(7)
6. ฝนหรอแกสบางชนดอาจระคายเคองผวหนงในสภาวะทความรอนและชน เชน ขณะมเหงอออก ฝนทสามารถกอสภาวะนได ไดแกฝนไม ฝนจากสาร arsenic สงกะส ตนไลมและเปลอกตนควนน เปนตน ฝนจากเปลอกผลไมเปลอกแขง เชนเกาลด อาจทาใหเปลอกตาบวม มผนทใบหนา แขน และลาคอ ในผปวยททางานในโรงงานอาจเกดการระคายเคองมาก ทาใหเกดผนทวรางกายได (5)
7. พรก/capsicin ทาใหเกดการระคายเคองทมอ มกเกดจากอาหารเผดรอนทมสวนประกอบของพรกไทย สเปรยพรกไทยซงมความเขมขนสงททางการตารวจใช
กอใหเกดรอยโรคทแสบไหมได การใชนาเยนรกษามกไมชวยเนองจากพรกไทยไมละลายนา ควรใชนาสมสายช/กรดแอซตก 5% หรอ ยาลดกรด สามารถบรรเทาอาการ
แสบรอนไหมไดดแมใชหลงสมผสพรกนานเปนชวโมง
แลวกตาม และควรใชอยางตอเนองจนกวารอยโรคจะหายด (9)
8. ผวหนงอกเสบจากแกสนาตา สารในแกสนาตาทเคยใชเพอควบคมการจลาจล ไดแก chloroacetophenone (แกส CN; mace) หรอ trichloroethane ซงใชในรปสเปรย ในปรมาณสงอาจกอใหเกดผวหนงอกเสบระคายสมผส การรกษาใหถอดเสอผาทเปอนออก ลางบรเวณผวหนงดวยนานม CN เคยใชในเวยตนาม ในปจจบนสารทใชเปนแกสนาตาอยางแพรหลาย คอ chlorobenzylidone malononitrile (แกส CS) ซงมความเปนพษตากวาและไมอนตรายเทา CN โดยผสมกบ methylene chloride การไดรบแกส CS นอกจากทาใหนาตาไหล แสบตา มผลตอระบบหายใจและอนๆ แลว ยงทาใหระคายผว และแสบรอนบรเวณสมผส ใหผสมผสออกไปอยในทโลง ลางบรเวณสมผสดวยนาและใชกรดบอรกลางตา(6)
สารทเคยใชเปนอาวธเคม ไดแก sulfur mustard gas (yperite) ซงใชในสงครามอรกและอหราน เนองจากมคณสมบตเปนของเหลวทระเหยงาย (ไมใชแกส) จงใชในรปฉดพนเปนละอองฝอย หากมการสมผสรนแรงอาจเกดผนทผวหนงและเยอบคลายผนแพยาชนดรนแรง (toxic

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
115
epidermal necrolysis) ใหถอดเสอผาออก ฟอกบรเวณปน เปอนดวยสบและนาและสะเทนดวยนาคลอรนเจอจาง(8)
9. ไฮโดรคารบอน สารประกอบประเภทไฮโดร คารบอนมคณสมบตละลายไดดในไขมนรวมถงคณสมบต
ความหนดทแปรผกผนกบความสามารถในการละลายใน
ไขมน นามนดบเปนหนงในสารประเภทน กอใหเกดอาการคน รขมขนอกเสบและเกดสวได สวนนามนทกลนแลวจะเกดความระคายเคองตอผวไดนอยกวานามนทยง
ไมไดผานการกลน กลมไฮโดรคารบอนททาใหเกดผวหนงอกเสบชนดรนแรงนอย ไดแก benzene และ naptha เปนตน กลมสาร napthalene สามารถกออาการไดมากกวากลมพาราฟน นอกจากน พยาธสภาพบรเวณผวหนงอาจเกดจากการสมผสกบนามนทมเกลอแรตางๆ นามนปโตรเลยมทมคณภาพตา และครโอโซต
คนททางานเกยวของกบนามนหลอลนจะพบวาเกด
ผวหนงอกเสบมากกวาผอน นามนหลอลนนแบงเปน 2 ประเภทไดแก นามนหลอลนชนดไมละลายในนา ทาใหเกดผวหนงอกเสบคลายสวบรเวณหลงมอ แขนทอนลาง ใบ หนา หนาขา และหลงลาคอ การสมผสสารเหลานนอกจากทาใหเกดรอยคลาและผวหนงหนาแลว ยงมความ สมพนธกบการเกดโรคมะเรงทอณฑะ สวนนามนหลอลนชนดละลายในนาและนามนสงเคราะหตางๆ ทใชในอตสาหกรรมเกยวกบโลหะ จะเกดผวหนงอกเสบคลายผนแพ พบบอยบรเวณหลงแขนทอนลางและมอ
นามนจากหนเฌลทาใหเกดผนแดงและการอกเสบ
ของรขมขนจนกระทงผวหนงหนาตวขน ซงจะเปนบรเวณทกลายเปนมะเรงตอไปได จากสถตพบวาคนงานททางานเกยวกบนามนหนเฌล มประมาณครงหนงทมปญหา
ผวหนงอกเสบจากการสมผสถานหนทใชเปนเชอเพลง ทาใหเกดผวหนงอกเสบจากสวนของนามนดน (coal tar) ทเปนผลพลอยไดจากอตสาหกรรมถานหน การสมผสยาง มะตอย และผลตภณฑอนๆ ทมาจากนามนดน สามารถกอใหเกด melanoderma ได การสมผสนามนดบ นามนดน และ paraffin ทไมบรสทธหรอทมคณภาพตาบอยๆ อาจทาใหรขมขนหนาตวและสเขมขน มกเกดบรเวณดานหลง
ของนวมอ แขน ขา และอก โดยเรมจากรขมขนอกเสบ ตอมาจะเปนสคลาออกสนาตาลหรอสมวง ถารนแรงขนจะเปนผนกระจายออกไป โดยเฉพาะบรเวณอวยวะเพศ อาการแสดงนจะคลายคลงกบ pityriasis rubra pilaris หรอ lichen spinulosus นอกจากนยงมโรคมะเรงทมกเกดบรเวณอณฑะซงเกดจากการสมผสเขมาควนสะสมทผวหนง จนผวหนงเกดหนาตวเรยก soot wart และเปลยนแปลงเปน
มะเรงในเวลาตอมา เรยกอกชอหนงวา chimney sweep’s cancer(8,10)
10. ตวทาละลาย (solvents) เปนสาเหตของผวหนงอกเสบระคายสมผสทเกดจากการทางานประมาณรอยละ 10 สาเหตจากการใชตวทาละลายลางมอเพอขจดคราบนามนตางๆ อาจเกดผวหนงอกเสบ มรอยแตกเปนรองลก นอกจากน บรเวณปลายประสาท และระบบนาเหลองอาจอกเสบไดดวย การทตองสดดมสารเหลานจะทาใหปากและจมก อกเสบบวมแดง สารละลายทเปอนบนผาเชดหนาทาใหผใชเกดเปนผวหนงอกเสบระคายสมผสได การหายใจ/สดดม trichloroethylene ซงใชในอตสาหกรรม
เครองเยนและซกแหง ทาใหผวหนงอกเสบแดง เยอบผวหนงกดกรอนและหลดลอก มเซลลเมดเลอดขาวชนด eosinophil ขนสงและตบอกเสบ กรณตวทาละลายเปนแอลกอฮอลชนด cetyl และ stearyl สามารถกอใหเกดผวหนงอกเสบระคายสมผสแบบผนลมพษได(8)
ผวหนงอกเสบภมแพสมผส ผวหนงอกเสบภมแพสมผสเกดจากปฏกรยาตอบ
สนองทางอมมนชนดท 4 (delayed type/cell-mediated hypersensitivity) ผปวยตองเคยสมผสสารทกอใหเกด
อาการแพมากอน แตไมจาเปนตองพบในทกคนทสมผสสาร กลไกการเกดเรมดวยรางกายดดซมสารประเภท hapten เขาทางผว หนงกอน เมอ haptenไปจบกบโปรตนทผวหนงแลว จะผานกระบวนการโดย Langerhans cell ทอยบรเวณชนผวหนงกาพรา และตอมานาเสนอตอ CD4+ T-cell ในรปสารเชงซอน (complex) กบ HLA-DR ทผวนอกของเซลล สกระบวนการสราง sensitized T-cell ทม
ความสามารถจดจาสาร hapten ทแพได กระบวนการนจะใชเวลาประมาณ 7-10 วนเรยกระยะนวา sensitized phase จากนน sensitized T-cell จะกลบสกระแสโลหต บางสวนไปอยบรเวณผวหนงทวรางกาย เมอผปวยสมผส hapten ทแพ แลวสารซมเขาสผวหนงไปจบกบโปรตนกลายเปนแอนตเจน เมอเกดปฏกรยาระหวาง sensitized T-cell กบ receptor-CD3 complex สารทกอการอกเสบจะถกปลอย
ออกมา เรยกระยะนวา elicitation phase ซงจะเกดภายใน 24 ชวโมง อาการแสดงของผวหนงจะเปนตม บวมแดง และคนมาก ความรนแรงของการอกเสบขนอยกบปจจยตางๆ ไดแก พนธกรรม เพศ อาย เชอชาต โรคทางผวหนงทผปวยมอยเดม และระยะเวลาทสมผสสาร เปนตน(8)
การปองกน การหาสาเหตและปองกนสงระคายเคองททาใหเกด

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
116
ผวหนงอกเสบระคายสมผสเปนสงทสาคญทสด แตในบางครงผปวยไมสามารถหลกเลยงสงระคายเคองดงกลาว
ได ดงนน จงจาเปนตองใชอปกรณปองกนเชนถงมอผาสาหรบงานทไมเปยกหรอถงมอยางททาจาก polyvinyl chloride (PVC) หรอ polyethylene (PE) สาหรบงานทเปยกนา นอกจากนการใชครมทาปองกน (barrier cream) ทมสวนผสมของ ceremide หรอ dimethicone กสามารถปอง กนการสมผสสารไดบางเชนกน(1)
การรกษา
ยาทมประสทธภาพในการรกษาโรคผวหนงอกเสบ
ระคายสมผส ไดแก ยาทาคอรตโคสตรอยด ซงเปนยาทใชบ อย และอาจร วมกบการใหสาร เพ มความช ม ชน (emollient) ในกรณทผนเปนแบบเรอรงหรอรนแรง อาจใชการรกษาดวยแสง (phototherapy) หรอยากดภมคมกน เชน azathiopine และ cyclosporin เปนตน ยาทาทเปน calcineurin inhibitor เชน pimecrolimus อาจใชแทนยาทาคอรตโคสตรอยดทมความแรงตา ยาปฏชวนะสามารถใชในกรณทตดเชอแทรกซอนบรเวณผวหนง(1)
ยาคอรตโคสตรอยด มฤทธตานการอกเสบ กดภม คมกน ยบยงการแบงเซลล และทาใหหลอดเลอดหดตว โดยยาไปจบเปนสารประกอบเชงซอนกบ receptor ใน
ไซโทพลาซมของเซลลแลวเคลอนยายเขาสนวเคลยส ซง drug-receptor complex จะจบกบ corticosteroid responsive element ซงอยบนยนตางๆ สงผลใหลดการสราง activator protein 1 และ nuclear factor kappa B ซงเปนโปรตนทเกยวของกบการอกเสบ และยงไปเพมการสราง lipocortinและp11/calpactin-binding protein รวมทงลดการหลง arachidonic acid นอกจากน lipocortin ยงยบยงการทางานของเอนไซม phospholipase A2 ทาให prostaglandin และ leukotriene ลดลง เปนผลใหลดการอกเสบทเกดขนทผวหนงและเยอบ คอรตโคสตรอยดยงสามารถกดภมคมกนโดยทาใหจานวน lymphocyte, neutrophil, fibroblast และ mast cell บรเวณทเกดการอกเสบลดลง สามารถยบยงการแบงเซลลโดยกระบวนการไมโทซสและสงเคราะหดเอนเอ และยงทาใหหลอดเลอดหดตวเนองจากทาใหสารทมฤทธ
ขยายหลอดเลอด เชน histamine, bradykinin และ prostaglandin ลดลง
ยาทาคอรตโคสตรอยดทมใชในประเทศไทยแบง
ไดเปน 7 กลม ดงแสดงตวอยางในตารางท 1(3,4) ยาทา คอรตโคสตรอยดชนดเดยวกน เมออยในรปแบบทแตกตางกน อาจใหความแรงไมเทากน โดยทวไปแบบขผงแรงกวาครม และแบบครมแรงกวาโลชน
ตารางท 1 การจาแนกยาทาคอรตโคสตรอยดตามความแรงเรยงลาดบจากมากไปหานอยโดยวธ vasoconstriction assay
ยาทาคอรตโคสตรอยด ชอสามญ ชอทางการคา Class 1 - Superpotent Class 2 - Potent Class 3 - Potent, upper mid-strength Class 4 - Mid-strength Class 5 - Lower mid-strength Class 6 - Mild strength Class 7 - Least potent
Clobestasol propionate 0.05% Augmented betamethasone dipropionate 0.05% Betamethasone dipropionate 0.05% Desoximetasone 0.25% Betamethasone dipropionate 0.05% Amcinonide 0.01% Triamcinolone acetonide 0.1% Mometasone fuorate 0.1% Prednicarbae 0.1% Betamethasone dipropionate 0.05% Betamethasone valerate 0.1% Triamcinolone acetonide 0.1% Triamcinolone acetonide 0.02% Aclometasone dipropionate 0.05% Hydrocortisone 0.5%, 1.0%, 2.5%
Dermovate cream Diprotop cream, ointment Diprosone ointment Topicort/esperson cream Diprosone cream Visderm cream/lotion TA/aristocort ointment Elomet cream Dermatop cream Diprosone lotion Betnovate cream TA/aristocort cream/lotion TA/aristocort cream Alcovate cream/ointment Hydrocortisone cream

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
117
หลกในการใชยาทาคอรตโคสตรอยด ในกรณทมการอกเสบแบบเฉยบพลน ไมหนา เปนทบรเวณใบหนาและบรเวณทอบชน ควรเลอกยาทความแรงตา หรอปานกลาง แตในกรณทเปนผนหนาเรอรงหรอบรเวณทผวหนงหนา เชนฝามอและฝาเทา ควรเลอกยาทความแรงสงหรอสงมาก หากรอยโรคเปนบรเวณกวางหรอในเดกและผสง อายควรใชยาทความแรงตาหรอปานกลาง เนองจากยา
สามารถถกดดซมเขาสกระแสเลอดไดงายและอาจไปกด
การทางานของตอมหมวกไต ยาทมความแรงสงมากไมควรใชตอเนองนานเกน 2 ถง 3 สปดาห และไมควรใชเกนสปดาหละ 45 กรม และยาทมความแรงปานกลางถงสงไมควรใชตอเนองเกน 3 เดอน เมอผปวยอาการดขนตองรบลดความแรงของยาลง แตถาจาเปนตองใชยาเปนเวลานานควรจะหยดยาบ า ง เปนระยะๆ เพ อป องกนการลดการตอบสนองตอยา (tachyphylaxis) และยาทความแรงตาหรอปานกลางไมควรใชเกนสปดาหละ 100 กรม
นอกจากน ควรระมดระวงในการใชยาทาบรเวณผนผาออมในเดก ควรใชยาทมฤทธออน และไมควรทายาทหวนมและเตานมกอนใหนม ผลขางเคยงเฉพาะทของยา ไดแก ผวหนงเหยว ผวบางลง แตกลาย หลอดเลอดขยาย สผวบรเวณททาจางลง มขนขน เกดสว และผวหนงตดเชอตางๆ ไดงาย นอกจากนอาจเกดผวหนงอกเสบแพสมผสจากยาทาคอรตโคสตรอยดเองหรอสารทใชกนบดในตวยา เชน paraben, polyethylene glycol และ benzyl alcohol เปนตน ในกรณททายาบรเวณกวางและเปนเวลานาน หรอใชยาทมความแรงสงมาก โดยเฉพาะในเดกและผสงอาย อาจเกดผลขางเคยงทวรางกาย เชน ตอกระจก ตอหน กดการทางานของตอมหมวกไต หรอในเดก ทาใหการเจรญเตบโตชาลงได เปนตน(3)
เอกสารอางอง 1. Antoine A, James ST, Apra S. Irritant contact
dermatitis. In: Fitzpatrick’s dermatology in general medicine. 7th ed. New York: The McGraw-Hill Company; 2008. p.395-401.
2. Corazza M, Trincone S, Zampino MR, Virgili A. Air bags and the skin. Skinmed 2004;3:256-8.
3. Isabel CV, Francisco AK. Topical corticosteroids. In: Fitzpatrick’s dermatology in general medicine. 7th ed. New York: The McGraw-Hill Company; 2008. p.2102-6.
4. Lindy PF, Hans FM, David RB. Dermatology pharmacology. In: Goodman & Gilman’s the pharmacological basis of therapeutics. 11th ed. New York: The McGraw-Hill Company; 2006. p.1798-1801.
5. Spiewak R, Skorska C, Dutkiewicz J. Occupational airborne contact dermatitis caused by thyme dust. Contact Dermatitis 2001;44:235-9.
6. Varma S, Holt PJ. Severe cutaneous reaction to CS gas. Clin Rxp Dermatol 2001;26:248-50.
7. Wang BJ, Lee JY, Wang RC. Fiberglass dermatitis report of two cases. J Formos Med Assoc 1993;92: 755-8.
8. William DJ, Timothy GB, Dirk ME. Contact derma-titis and drug eruptions. In: Andrew’s diseases of the skin clinical dermatology. 10th ed. Philadelphia: WB Saunders Company; 2006. p.91-6.
9. Williams S, Clark R, Dunford J. Contact dermatitis associated with capsaicin: hunan hand syndrome. Ann Emerg Med 1995;25:713-5.
10. Yokota K, Johyama Y, Matsumoto N, Yamaguchi K. Occupational dermatitis from a one-component naphthalene type epoxy adhesive. Ind Health 2002; 40:63-5.


119
Antiviral Drugs for the Control of Influenza Virus Infections: Present and Future Perspectives
Radeekorn Akkarawongsapat Department of Microbiology, Faculty of Science, Mahidol University.
Abstract
Influenza A viruses are infectious respiratory pathogens that continue to cause widespread morbidity and mortality. Cross-species transmission of a highly pathogenic H5N1 avian influenza A virus directly from birds to humans has raised concern that this may give rise to the next influenza pandemic. Administration of antiviral drugs is thus an important first line of defense against the virus. Current antiviral drugs are effective against seasonal influenza virus infection, and some have been used in the treatment of patients infected with the avian influenza virus. However, the emergence of drug-resistant viruses could become a major problem in the future. Identification of new antiviral drugs is clearly needed in order to better control influenza infection. In this article, some new antiviral strategies are reviewed and proposed, which may be developed as future therapies.
Keywords: influenza; avian influenza; drug resistance; antivirals; adamantanes; neuraminidase inhibitors
Introduction
Influenza A viruses are significant respiratory pathogens causing seasonal flu epidemics each year and potentially infrequent pandemics. In the United States, 5-20% of the population acquires the flu and about 36,000 people die from flu each year(6), indicating that the viruses remain a major cause of morbidity and mortality in the United States. In the past 100 years, there have been three major influenza A virus pandemics: Spanish flu in 1918, Asian flu in 1957, and Hong Kong flu in 1968; all claimed several millions of lives. Currently, there is an influenza virus pandemic among birds, predominantly involving the highly pathogenic avian influenza (HPAI) H5N1 virus, occuring throughout many parts of the world. It is now clear that the avian influenza virus is able to be directly transmitted from birds to humans. Although the prevalence of direct transmission is low, the mortality rate in infected individuals is high (~60%, according to the data from World Health Organization). The emergence of an avian influenza H5N1 virus that aquires the capacity to effectively infect humans would pose a serious public health
threat that could result in the next pandemic. In fact, a recent study has shown that an H5N1 virus isolated from a fatal human case can bind to a human-type receptor, suggesting the selection of mutant virus strains that have adapted to more easily infect humans(1). Although annual vaccination is the primary strategy for preventing infections, current vaccines are not effective against avian influenza viruses. The use of influenza antiviral drugs therefore remains to be a key approach to control illness and transmission. Several pieces of evidence have demonstrated that the appearance of antiviral drug-resistant strains of influenza viruses are increasing, highlighting the urgent need for novel antiviral drugs(9,13,16). In this article, the current influenza antiviral drugs are reviewed and recent developments pointing to potential new antiviral drugs against influenza infections are described.
Antigenic Variations: a Reason for the Emergence of Influenza Virus Variants
The major antigenic determinants of the influenza A virus are the viral surface glyco-proteins, hemagglutinin (H) and neuraminidase

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
120
(N). The different subtypes of both H (H1-H16) and N (N1-N9) were identified in wild birds, the natural reservoir of influenza A viruses. The proofreading activity of viral RNA polymerase is not efficient and this creates a high rate of transcription errors in the viral genome, including genes that encode antigenic deter-minants. Some of these virus variants have mutations in the surface glycoproteins, such as in the antibody binding sites of the hemagglutinin, thus enabling the virus to evade pre-existing host humoral immunity. This process is termed “antigenic drift,” and generally results in epidemics. Some virus variants have mutations in proteins that are targets of antiviral drugs, generating drug-resistant virus strains. The second form of antigenic variation is “antigenic shift.” The segmented RNA genome of the virus allows a process called gene reassortment, that occurs when different subtypes of influenza viruses infect the same cell. RNA gene segments are then mixed, resulting in new viruses that contain new combinations of the viral RNA segments. Thus, a pandemic may arise if the virus with a new subtype of hemagglutinin and/or neurami- nidase has emerged(18).
Factors that Contribute to the Appearance of Drug-resistant Viruses
The error rate of viral RNA polymerase is not the only factor that contributes to the emer-gence of drug-resistant influenza viruses. The concentration and duration of exposure to antiviral drugs at the site of infection are also important factors contributing to the emergence of drug resistance. In order to survive, the viruses have the capacity to adapt themselves to fit better in the environment. Moreover, the type and virulence of the influenza strain, the nature of the host immune response, and pharmacokinetics differences between patients all appear to play a role in determining the emergence of drug-resistant viruses. Therefore, an influenza virus infected individual has to be treated with antiviral drugs early and at the appropriate dose in order to help minimize the likelihood of the emergence of drug resistant
virus. In addition, the use of combination antiviral therapy may be useful as well(11).
The Current Antiviral Drugs against Influenza A Virus Infections
Two classes of anti-influenza drugs have been approved for treatment and prophylaxis of influenza. They are adamantane derivatives (amantadine and rimantadine) and neurami- nidase inhibitors (NAIs; zanamivir and oselta-mivir)(14), which block the function of the viral matrix-2 (M2) ion channel protein and neurami-nidase surface glycoprotein, respectively. Both classes need to be promptly administered within 48 hours after the onset of symptoms in order to obtain optimum drug efficacy.
The influenza A virus M2 protein is an ion channel protein that plays an important role in the viral uncoating process. Once the virus is taken up by the cells through receptor-mediated endocytosis, the virus in the endosome requires an acidic environment in order to release its genome into the cytosol and subsequently the nucleus for viral replication. The M2 protein transports hydrogen ions (H+) into the inter-nalized virus particle, providing an acidic en- vironment inside the particle. Adamantane derivatives bind to the transmembrane region of the viral M2 protein; thus, blocking the membrane pore and preventing the viral uncoating process.
The use of adamantanes could be limited in the future due to the high occurrence rate of adamantane-resistant viral isolates. Several studies have demonstrated that adamantane-resistant viruses are generated during treatment with the drug both in tissue culture and infected individuals(20). Resistant viruses were found to have acquired a single point mutation in the M2 transmembrane region, thus preventing adaman-tane binding(19)
. These resistant viruses were shown to remain as pathogenic and transmissi-ble as wild type viruses(10). In the United States, from 1995 to 2002, the rate of adamantane resistance was 2%. That number has subse-quently increased significantly to 92.3% in the 2005-2006 flu seasons(3). Moreover, the emer-gence of adamantane-resistant virus was observed in avian influenza virus isolates(23),

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
121
posing an added concern. Therefore, alternative antiviral strategies are clearly needed to treat influenza virus infection.
The second class of anti-influenza virus drugs is the neuraminidase inhibitors (NAIs). The NAIs inhibit the enzymatic activity of the viral neuraminidase (N ) protein, which plays an important role in the viral replication cycle, specifically in viral release from the infected cell. Neuraminidase removes sialic acid from cell-surface receptors that allows viral progeny to be released to infect new cells. The influenza virus that is deficient in N activity would result in inhibition of virus propagation. Since the crystal structure of the N is solved(8), structure-based design of NAIs is one of the most common approaches in the development of anti-influenza virus drugs. These efforts have resulted in the licensing of the drugs zanamivir and oseltamivir. Both drugs target the active site of the N protein and have been shown to be effective inhibitors both in vitro and in vivo(7). Although zanamivir and oseltamivir are similar in their mode of action, oseltamivir is more commonly prescribed to treat influenza virus infection. A major reason for this is due to differences in route of administration. The ease of drug administration becomes an important factor for current drug development. The bioavailability of zanamivir is poor, thus the drug has to be administered by inhalation. By contrast, oseltamivir is in the form of a pro-drug that is administered orally. As a consequence, many organizations in many countries stockpile oseltamivir for the prevention of influenza virus infection and in preparation for the next pandemic.
The occurrence of resistance to NAIs is not zero; however, it is much fewer than that resistance to adamantanes. Several studies demonstrated that influenza viruses that are resistant to the NAIs have been isolated in vitro and in vivo. These mutant viruses were found to exhibit amino acid changes either in the active site of the N or in the H(14). The alteration in the H protein reduces the affinity of the protein for the sialic acid receptor, thus presumably facilitating more viral exit which depends
mainly on N protein. More studies also showed that the NAI-resistant viruses often have their growth impaired in cell culture and have their infectivity and transmissibility reduced in animal infections(12). This may explain the few emergences of NAI-resistant viruses that are recovered from patients treated with NAIs. Nevertheless, drug resistance in patients infected with avian H5N1 virus was reported in Vietnam(9). Thus, it would be prudent to prepare or develop additional antiviral drugs in order to better treat and prevent influenza virus infections.
Ongoing Researches and Future Perspectives for Anti-influenza Drugs Development
Several strategies have been pursued in order to identify new anti-influenza drugs. Some of these show promising results suggesting that they may become effective in the clinic. Each strategy provides both benefits and drawbacks; as a result, careful evaluation of using each potential antiviral drug is needed.
RNA interference (RNAi) is a well known pathway that involves specific inhibition of target mRNA expression by introducing sequence-specific double-stranded RNA duplex, or short interfering (si)RNA. RNAi has been shown to display antiviral activity both in vitro and in vivo against several viruses, including influenza A virus(4). For anti-influenza virus, siRNAs were designed to target degradation of viral nucleoprotein, polymerase, and matrix genes. The results showed that viral replication was suppressed in tissue culture and virus yields in the lungs of infected mice were reduced significantly. The antiviral effect was demon-strated to be mRNA specific and effective against different subtypes of human and avian influenza viruses(22). The benefits of RNAi-based therapeutics are that siRNA can be synthesized within short period of time and the cost of synthesis is relatively low. However, a concern is raised because there have been reports showing that non-specific degradation of some host genes was observed when using some antiviral siRNA molecules. Therefore, speci-

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
122
ficity for the target gene degradation has to be carefully examined.
Influenza viruses, similar to many RNA viruses, replicate their virus genome via a replicative intermediate (RI), a double-stranded RNA intermediate. Because host cells do not produce such molecules, the presence of RI in mammalian cells stimulates antiviral response. For the search of new antiviral drug, molecules that mimic the structure of RIs have been synthesized. Studies have shown that synthetic dsRNA molecules composed of polyribo-inosinic polyribocytidylic acid (poly[I:C]) are effective in battling influenza virus infection in cell culture(25). However, in vivo data are not available. This approach is still needed to be tested for its effectiveness in preventing influenza virus infection in human patients.
Structure-based approach has been successfully used to identify effective neurami-nidase inhibitors. Similar strategy then could also be conducted using other viral proteins that are essential for virus replication cycle as a new target. The other influenza virus surface glyco-protein, H, serves as a potential target for antiviral drugs because it is required for the initial stage of virus infection, receptor recog-nition and membrane fusion of virus and cell membranes; and the crystal structures of H at neutral- and fusion-pH are available. After receptor binding, virus is in the endosome. Acidic pH in the endosome then triggers conformational change in H structure such that the protein is fusion-competent, mediating membrane fusion of virus envelope and cell endosomal membrane. Therefore, a molecule that prevents H rearrangement, thus inhibiting membrane fusion, is expected to be a powerful antiviral drug.
Indeed, several small molecules, such as tert-butyl hydroquinone (TBHQ), that block conformational change in H required for membrane fusion have been identified(17)
. However, problems persist because of numerous subtypes of H. Limitations involve the lack of crystal structures of relevant H complexes and H group specificity (i.e. the compounds are effective against certain subtypes of H). Perfect
instances are the Hs of current seasonal influenza viruses (H1 and H3) and the Hs of the highly pathogenic avian influenza viruses (H5 and H7). Each member in each pair is grouped into different phylogenetic groups. Thus, understanding the structural basis of drug sensitivity and group specificity would aid in designing more potent inhibitors of anti-influenza drugs.
Protein-protein interaction is a funda-mental process that is important in all biological activities. Identification of peptides that mimic the protein-protein interaction sites, thus inter-fering the protein interaction, has been proved to be successful for antiviral drug development and useful tools for elucidating the mechanisms underlying the involving process. For instance, the anti-human immunodeficiency virus (HIV) drug Fuzeon is a peptide derived from HIV membrane fusion protein and acts by blocking HIV entry(24). Recently, a 20-amino acid peptide, EB, derived from the signal sequence of fibroblast growth factor 4 was shown to exhibit antiviral activity against influenza viruses in vitro and in vivo. The antiviral activity is also active against the H5N1 subtype in vitro(12). The EB peptide was demonstrated to bind to the viral H protein, blocking viral binding to cell surface receptor, thus preventing infection. Therefore, it is of interest to obtain peptides corresponding to the H protein and test for antiviral activity against influenza viruses. The benefits are two-fold. The peptides that exhibit antiviral activity can not only be developed as potential antiviral drugs, it can also aid us in understanding the underlying mechanisms of how the virus enters cells. In addition, the availability of crystal structures of H should assist further development of antiviral drugs that target H protein for the treatment and prevention of influenza virus infection.
Many current antiviral strategies focus on drugs that target the virus proteins that are essential for virus replication cycle. With an increasing knowledge on influenza virus-host interactions, it is also possible that targeting critical cellular factors that play a role in the virus replication cycle can be another potential

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
123
antiviral strategy. Chloroquine is a lysosomo-tropic agent that elevates the endosomal pH. Previous studies have demonstrated that treatment of cells with chloroquine exhibits inhibitory effect on influenza virus replication in tissue cultures(15). Several studies also report the importance of cellular lipid-rafts in the virus maturation process(5,21). Lipid-rafts are specia-lized microdomains in the cell membrane that are rich in cholesterol. Treatment of cells with statin, which removes membrane-bound choles-terol and thus disrupts lipid-raft membranes, was shown to inhibit the assembly process of influenza virus in tissue culture(2). Although the available data is encouraging, both chloroquine and statin are needed to be further evaluated for anti-influenza drugs in humans. Extra consi-deration must be taken when working with antiviral drugs that target cellular factors. Since the cellular factors can be critical to many cellular processes, it is necessary to confirm that inhibition of the cellular factors would not have any adverse effects on cells.
Conclusions
Prevention for the next influenza virus pandemic has to be promptly prepared. Since the last influenza virus pandemic, numerous studies were done on how the influenza virus interacts with its host. The increase in knowledge greatly enhances our understanding of the biology of the influenza virus, and thus facilitating the development of new antiviral drugs. Many laboratories are currently working on antiviral drugs development, and it is clear that this process will not be a major limitation when the next pandemic arrives. Rather, the challenge is the availability of anti-influenza drugs to the hands of affected individuals that live in many parts of the world. The antiviral drugs must be easily mass produced with affordable cost and distributed to all parts of the world. The challenge is difficult, but is realistic with the cooperation among researchers, government, and pharmaceutical industry.
Acknowledgements
The author would like to thank Assistant Professor Arunee Thitithanyanont for valuable discussion and advice.
References 1. Auewarakul P, Suptawiwat O, Kongchanagul A,
Sangma C, Suzuki Y,Ungchusak K et al. An avian influenza H5N1 virus that binds to a human-type receptor. J Virol 2007;81:9950-5.
2. Barman S, Nayak DP. Lipid raft disruption by cholesterol depletion enhances influenza A virus budding from MDCK cells. J Virol 2007;81: 12169-78.
3. Bright RA, Shay DK, Shu B, Cox NJ, Klimov AI. Adamantane resistance among influenza A viruses isolated early during the 2005-2006 influenza season in the United States. JAMA 2006;295: 891-4.
4. Bumcrot D, Manoharan M, Koteliansky V, Sah DW. RNAi therapeutics: a potential new class of pharmaceutical drugs. Nat Chem Biol 2006; 2: 711-9.
5. Carrasco M, Amorim MJ, Digard P. Lipid raft-dependent targeting of the influenza A virus nucleoprotein to the apical plasma membrane. Traffic 2004;5:979-92.
6. Centers for Disease Control and Prevention. [cited 2008 12 December]; Available from: http://www.cdc.gov/flu/about/disease/index.htm
7. Collins PJ, Haire LF, Lin YP, Liu J, Russell RJ, Walker PA et al. Crystal structures of oseltamivir-resistant influenza virus neurami-nidase mutants. Nature 2008;453:1258-61.
8. Colman PM, Laver WG, Varghese JN, Baker AT, Tulloch PA, Air GM et al. Three-dimensional structure of a complex of antibody with influenza virus neuraminidase. Nature 1987;326:358-63.
9. de Jong MD, Tran TT, Truong HK, Vo MH, Smith GJ, Nguyen VC et al. Oseltamivir resis-tance during treatment of influenza A (H5N1) infection. N Engl J Med 2005;353:2667-72.
10. Hayden FG. Respiratory viral threats. Curr Opin Infect Dis 2006;19:169-78.
11. Ilyushina NA, Bovin NV, Webster RG, Govorkova EA. Combination chemotherapy, a potential strategy for reducing the emergence of drug-resistant influenza A variants. Antiviral Res 2006;70:121-31.
12. Jones JC, Turpin EA, Bultmann H, Brandt CR, Schultz-Cherry S. Inhibition of influenza virus infection by a novel antiviral peptide that targets

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
124
viral attachment to cells. J Virol 2006;80:11960-7.
13. Le QM, Kiso M, Someya K, Sakai YT, Nguyen TH, Nguyen KH et al. Avian flu: isolation of drug-resistant H5N1 virus. Nature 2005;437: 1108.
14. Nicholson KG, Wood JM, Zambon M. Influenza. Lancet 2003;362:1733-45.
15. Ooi EE, Chew JS, Loh JP, Chua RC. In vitro inhibition of human influenza A virus replication by chloroquine. Virol J 2006;3:39.
16. Puthavathana P, Auewarakul P, Charoenying PC, Sangsiriwut K, Pooruk P, Boonnak K et al. Molecular characterization of the complete genome of human influenza H5N1 virus isolates from Thailand. J Gen Virol 2005;86 (Pt 2) :423-33.
17. Russell RJ, Kerry PS, Stevens DJ, Steinhauer DA, Martin SR, Gamblin SJ et al. Structure of influenza hemagglutinin in complex with an inhibitor of membrane fusion. Proc Natl Acad Sci USA 2008;105:17736-41.
18. Stephenson I, Democratis J. Influenza: current threat from avian influenza. Br Med Bull 2005; 75-76:63-80.
19. Sugrue RJ, Hay AJ. Structural characteristics of the M2 protein of influenza A viruses: evidence that it forms a tetrameric channel. Virology 1991; 180:617-24.
20. Sugrue RJ, Tan BH, Yeo DS, Sutejo R. Antiviral drugs for the control of pandemic influenza virus. Ann Acad Med Singapore 2008;37:518-24.
21. Takeda M, Leser GP, Russell CJ, Lamb RA. Influenza virus hemagglutinin concentrates in lipid raft microdomains for efficient viral fusion. Proc Natl Acad Sci USA 2003;100:14610-7.
22. Tompkins SM, Lo CY, Tumpey TM, Epstein SL. Protection against lethal influenza virus challenge by RNA interference in vivo. Proc Natl Acad Sci USA 2004;101:8682-6.
23. Wainright PO, Perdue ML, Brugh M, Beard CW. Amantadine resistance among hemag-glutinin subtype 5 strains of avian influenza virus. Avian Dis 1991;35:31-9.
24. Wild CT, Shugars DC, Greenwell TK, McDanal CB, Matthews TJ. Peptides corresponding to a predictive alpha-helical domain of human immunodeficiency virus type 1 gp41 are potent inhibitors of virus infection. Proc Natl Acad Sci USA 1994;91:9770-4.
25. Wong JP, Yang H, Nagata L, Kende M, Levy H, Schnell G et al. Liposome-mediated immuno-therapy against respiratory influenza virus
infection using double-stranded RNA poly ICLC. Vaccine 1999;17:1788-95.

125
ชดเครองวดภาคสนามสยามแกโซฮอล หนงในนวตกรรมครบรอบ 50 ปคณะวทยาศาสตร
กาญจนา อไรสนธว และ ดวงใจ นาคะปรชา ภาควชาเคม คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
แกโซฮอลเปนพลงงานเชอเพลงทดแทนทใชสาหรบยานยนต ไดจากการผสมเอทานอลทมความบรสทธสงลงไปในนามนแกสโซลน ในระหวางการเกบรกษา สภาวะแวดลอมอาจสงผลใหระดบความเขมขนของเอทานอลเปลยนแปลงไปได เปนผลใหคาออกเทนของเชอเพลงไมไดมาตรฐาน บรษทผผลตจงไดใหความสาคญในการออกตรวจและตดตามวดคาตางๆ รวมทงระดบความเขมขนของเอทานอลอยางใกลชดและเปนประจา เพอควบคมคณภาพของพลงงานเชอเพลงชนดน ใหอยในมาตรฐานของกรมธรกจพลงงานดวย ซงตามมาตรฐานแลวชวงทยอมใหคาคลาดเคลอนไดนนอยในชวงแคบเพยง 1 % (v/v) ชดเครองวดภาคสนามสยามแกโซฮอล เปนชดเครองวดแบบเคลอนทสาหรบควบคมคณภาพแกโซฮอล เปนผลงานนวตกรรมเพอใชในการตรวจวดภาคสนามสาหรบวดระดบความเขมขนของเอทานอลในทตองทาเปนประจา เครองวดนไดรบการออกแบบพเศษใหงายตอการใชงานและอานผลไดภายในเวลา 2 นาท และยงมคณสมบตทดในดานของความเทยง (3% RSD) ความแมนยาในการวด (98-100%) และวดคาเอทานอลในแกโซฮอลไดละเอยดในระดบ 0.3 % (v/v)
คาสาคญ: เอทานอล; แกโซฮอล; ชดตรวจวด; การวดภาคสนาม
Abstract
Gasohol, a biofuel for automobiles, contains gasoline supplemented with ethanol of high purity. During storage, the ethanol content may vary due to some environmental conditions and as a result gasohol specification is changed as ethanol content determines ‘octane number’ of the fuel, an issue of concern for petrol companies producing gasohol. After distribution to gas stations, petrol producers need to monitor this blended fuel closely and regularly to ensure that the ethanol content is within 1 % (v/v) of the specification designated by the Department of Business Energy. Siam Gasohol-Kit (SG-Kit) is a portable test kit for quality control of gasohol. SG kit is an innovation tool suitable for routine on-site measurement of ethanol in gasohol and has been designed specifically to offer a simple method for users. Reading of ethanol content appears in 2 minutes. SG-Kit provides high precision (3% RSD) and accuracy (98-100%). As SG-Kit provides a resolution of 0.3 % (v/v) ethanol in gasohol, the kit is suitable for use as part of quality control scheme in the gasoline industry.
Keywords: ethanol; gasohol; test kit; on-site analysis
บทนา
แกโซฮอลเปนหนงในพลงงานทางเลอกเพอใชใน
การคมนาคมและการขนสง นอกเหนอจากพลงงานทดแทนอนๆ เชน แกสเอนจว แอลพจ และไบโอดเซล ในปจจบน รฐบาลกาลงสงเสรมและรณรงคใหประชาชนหนมาใชพลงงานทดแทน เพอลดการนาเขานามนเชอเพลงทราคาผนผวน และพรอมรบมอกบพลงงานเชอเพลงทเรมจะขาดแคลน ซงในปจจบน มแกโซฮอลเปนนามนเชอเพลงทเกดจากการผสมนามนเบนซนพนฐานกบเอทานอลแปลง
สภาพ สามารถนามาใชทดแทนนามนเบนซนพนฐานทไดจากนามนดบโดยตรง โดยการใชนามนทดแทนประเภทนเปนทนยมกนในประเทศแถบอเมรกา และยโรป เชน บราซล และสวเดน เปนตน สาหรบในประเทศไทย นามนแกโซฮอลเรมเปนทรจกในฐานะทเปนโครงการตามแนว
พระราชดารในพระบาทสมเดจพระเจาอยหว ททรงรเรมเมอป พ.ศ. 2528 เพอสงเสรมการแปรรปผลตผลจากภาค
การเกษตรใหเปนเอทานอลและเปนพลงงานทดแทน พรอมทงสามารถแกปญหาราคาสนคาเกษตรตกตาในคราว

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
126
เดยวกน จนกระทงในป 2543 การปโตรเลยมแหงประเทศไทย (ปตท .) ดาเนนการทดสอบการใชแกโซฮอลในรถยนต และไดทดลองจาหนายครงแรกเมอป พ.ศ. 2544(2)
การผสมเอทานอลในสดสวนทแตกตางกนลงไปใน
นามนเบนซนแสดงไดจากคา ‘E-ตวเลข’ ซงตวเลขเปนสงทกาหนดคารอยละโดยปรมาตรของเอทานอลทผสมกบ
นามนเบนซนพนฐาน ตามขอกาหนดลกษณะและคณภาพของนามนเชอเพลงแกโซฮอลโดยกรมธรกจพลงงาน กระทรวงพลงงานนน นามนแกโซฮอลชนด E10 ตองม เอทานอลผสมอยรอยละ 9-10 โดยปรมาตร นามนแกโซฮอลชนด E20 ตองมเอทานอลผสมอยรอยละ 19-20 และนามนแกโซฮอลชนด E85 ตองมเอทานอลผสมอยไมตากวารอยละ 75 โดยระดบความเขมขนเปนรอยละตอปรมาตรนน
ตองไมเกนชวงทระบไวดงกลาว หากระดบ เอทานอลไมเปนไปตามน จดวาน ามนเชอ เพลงนนต ากวา เกณฑ
มาตรฐานทกาหนดไว ดงประกาศของกระทรวงฯ ปรากฎในราชกจจานเบกษา เรอง กาหนดลกษณะและคณภาพ
ของนามนแกโซฮอล พ.ศ. 2551(1)
บทความวชาการนนาเสนอภาพรวม เหตผล และความจาเปนในการวดระดบเอทานอลในนามนแกโซฮอล วธการโดยทวไปสาหรบการควบคมคณภาพน ามน แกโซฮอล พรอมทงการเสนอผลงานนวตกรรมทพฒนาขนโดยคณะวทยาศาสตร มหาวทยาลยมหดล เพอการวดระดบเอทานอลในนามนแกโซฮอลแบบภาคสนาม
บทบาทของเอทานอลกบการควบคมคาออกเทน ของแกโซฮอล
สาเหตทตองควบคมใหระดบเอทานอลเปนไปตาม
มาตรฐาน โดยไมตากวาหรอสงกวารอยละทกาหนดไว คอเนองจากเอทานอลเปนสารทนอกจากจะเตมลงไปเพอเปน
พลงงานทดแทนแลว ยงทาหนาท เปนตวเพมและหรอควบคมคาออกเทนของนามนแกโซฮอลดวย ซงเอทานอลสามารถควบคมคาออกเทนทดแทนสารควบคมตวเกาคอ methyl tertiary butyl ether (MTBE) เพอควบคมคาออกเทนใหได 91 หรอ 95 ตามฉลากทระบเพอจาหนาย ซงนนหมายถง ระดบเอทานอลตองเปนไปตามทระบไว มฉะนนคาออกเทนจะผดเพยนไปจากทกาหนดไวได และมผลทางกฎหมายดวย
เอทานอลมากหรอนอยไปสงผลกบเครองยนตอยางไร?
เนองจากเอทานอลเปนตวเพมคาออกเทนใหกบเชอ
เพลงเพอใหไดตามมาตรฐานทระบ และใหออกเทนตรงกบฉลากทตดไวตามสถานบรการขณะจาหนาย ดงนนหากระดบเอทานอลเปลยนแปลงไปยอมสงผลกบคาออกเทน ของนามนขณะจาหนายดวย และสดทายทาใหนามน แกโซฮอลท เ ตมให กบรถยนตน นไม เหมาะสมกบ
เครองยนตเนองจากมเอทานอลตากวาทระบไว สงผลใหมคาออกเทนตากวามาตรฐาน
แตในกรณทมเอทานอลสงกวาทระบไว เชน สงกวารอยละ 10 โดยปรมาตร แตวาเอทานอลไดถกเตมลงไปเกนกวามาตรฐานของ E10 เชน มการเตมลงไปเพราะราคา เอทานอลตากวาราคานามนเชอเพลง เปนตน เอทานอลทมระดบสงเกนมาตรฐานนจะสงผลกบรถยนตโดยเฉพาะรน
เกา ทาใหเกดความฝดในทอเชอเพลง สงผลใหเครองยนตกระตกหรออาจดบไดเปนบางครง
สาเหตทอาจทาใหระดบเอทานอลเปลยนแปลงไปจนไมไดมาตรฐานหลงการผลต
ระดบเอทานอลสามารถเกดการเปลยนแปลงไปได
เอง ภายหลงผสมกบนามนเบนซนพนฐานและผลตเปน แกโซฮอลแลว และแนนอนยอมทาใหคาออกเทนเปลยน แปลงไดดวย โดยอาจสรปสาเหตไดดงน
1. การระเหย เนองจากทงนามนเบนซนพนฐานและเอทานอล
ตางกเปนสารระเหยงาย และระเหยไดตลอดเวลาหลงผสม ดงนนระหวางการเกบรกษากอนจาหนาย หากตองเกบไวเปนเวลานาน ระดบเอทานอลกเปลยนแปลงไปจากทระบไวได
2. การปนเปอนดวยนาในถงเกบทาใหเอทานอลลดลง
ภายในถงเกบนามนของสถานบรการ หรอแมแตคลงนามน จะมปรากฎการณทมอาจหลกเลยงไดและเกด ขนเอง คอ มการปนเปอนของนาเขาไปอยภายในถงเกบนามน เกดจากการควบแนนของไอนาภายในถง ซงจะเกดไดดยงขนหากมการเปลยนแปลงอณหภมทเยนตวลง ทาใหเกดการควบแนนของไอนาเปนหยดนา หรอเหงอนามนภายหลงการเกดนาแลว สวนทเปนนาจะอยทระดบลางสดของถงเกบเชอเพลง และการทมนาอยในถงแบบนทาให เอทานอลในชนนามนเชอเพลงลดลงเนองจากเอทานอล
ละลายในนาไดดกวาในนามนมาก เอทานอลจะเคลอนตวมาละลายในชนนา ซงหากเกดนาขนภายในถงเกบ ระดบ เอทานอลจะลดลงกวามาตรฐาน จงทาใหคาออกเทนของ แกโซฮอลนนลดลงได

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
127
3. การปลอมปน เนองจากประเทศไทยเปนประเทศเกษตรกรรมและ
มวตถดบตลอดจนมนโยบายผลกดนใหมการผลตเอทา-นอลมากเพอเปนพลงงานทดแทน จนกระทงในทสด ราคา เอทานอลจะต ากว าน ามน เบนซนพนฐานท ใช เปน
สวนผสมในการผลตแกโซฮอล ดงนน หากมการปลอมปนโดยการเตมเอทานอลลงไปเองภายหลงการสงแกโซฮอล
ใหกบสถานบรการในเครอขายของบรษทแลว กจะทาใหมระดบเอทานอลมากกวาปกต ซงสงผลเสยกบเครองยนตทประชาชนใชอย ตลอดจนเสยภาพพจนของบรษทผผลตได
การควบคมคณภาพนามนแกโซฮอล โดยภาครฐและบรษทผผลต
1. การออกตรวจดวยรถตรวจคณภาพเคลอนท โดยปกตแลวบรษทผผลตนามนเชอเพลงมการดแลและควบคมคณภาพนามนเชอเพลงอยางเปนระบบ อกทงประเทศไทยยงมกรมธรกจพลงงาน กระทรวงพลงงาน คอยควบคมดแลคณภาพอยโดยตลอด ซงทงภาครฐและบรษทผผลตจะมรถออกตรวจสอบคณภาพนามนเชอเพลง
ทกประเภทตามสถานบรการตาง ๆ โดยบรษทนามนนนจะออกตรวจสอบทกๆ สถานบรการทมทวประเทศ สวนภาครฐจะใชวธสมตรวจทวประเทศ รปท 1 แสดงภาพรถตรวจสอบของกระทรวงพลงงาน
รปท 1 รถตรวจสอบคณภาพเชอเพลง ซงภายในมหองปฏบตการ ภาคสนาม
2. วธการตรวจวดระดบเอทานอลดวยรถตรวจ สอบเคลอนท
วธการทภาครฐและบรษทนามนสวนใหญใชตรวจ
เมอออกตรวจดวยรถตรวจคณภาพเคลอนทมกใชหลกการ
ผสมแกโซฮอลกบนาในกระบอกตวง (ดงแสดงในรปท 2) ซงผวดจะวดระดบนาทเพมขน แลวนาระดบทเพมขนไปคานวณเทยบกลบมาเปนระดบเอทานอล การวดแบบนมคาความถกตองตา เนองจากสเกลของกระบอกตวงนนหยาบมาก จงไมเหมาะสมอยางยงสาหรบการตรวจวดซงตองการความละเอยดสงมาก เชน E10 ตองวดไมใหมเอทานอลตา
กวารอยละ 9 และไมเกนรอยละ 10 ซงเครองมอวดทเหมาะสมนนควรตองอานไดอยางนอยใหแยกความ
แตกตางไดในชวงรอยละ 0.5 เชน วดคาเอทานอลได 9.5 + 0.5 สาหรบ E10 และวดคาเอทานอลได 19.5 + 0.5 สาหรบ E20 เปนตน
รปท 2 แสดงการใชกระบอกตวงในภาคสนาม (สาหรบรถตรวจ คณภาพเพอวดเอทานอลในแกโซฮอล
3. การวดระดบเอทานอลในหองปฏบตการและ แบบวธมาตรฐาน (เคลอนทไมได)
ตงแตอดตจนถงปจจบน ไดมนกวจยทาการพฒนาวธการตรวจวดคณภาพเอทานอลในหองปฏบตการหลาย
วธดวยกน ตวอยางเชน การตรวจวดระดบเอทานอลโดยเทคนค liquid chromatography(8) การตรวจวดการดดกลนแสงอนฟาเรดของเอทานอลในนามนแกโซฮอลดวย
อปกรณ attenuated total reflectance (ATR)(3) และ fourier transform-near infrared (FT-NIR)(4)การตรวจวดคณ สมบตทางกายภาพเกยวกบการวดคาความตานทาน
โดยรวมของนามนแกโซฮอลซงมปรมาณเอทานอลตางๆ(6) อยางไรกด วธการขางตนทอาศยคณสมบตทางกายภาพ
อาจเกดความผดพลาดไดงาย เนองจากนามนแกโซฮอลของแตละบรษทผผลตนามนมองคประกอบทแตกตางกน
จงสามารถสงผลตอความถกตองและแมนยาของการ
ตรวจวดได และในป พ.ศ. 2551 ไดมผลงานวจยของคนไทยทอาศยหลกการวเคราะหแบบไหลตอเนองเพอ
ตรวจวดคณภาพนามนแกโซฮอล(5) โดยอาศยปฏกรยาเคมทมความเฉพาะเจาะจงตอเอทานอลและองคประกอบตางๆ ในนามนแกโซฮอล ไมรบกวนการวเคราะห
วธการมาตรฐานสาหรบการตรวจวดปรมาณเอทา
นอลในแกโซฮอลทใชกนในหองปฏบตการของบรษท ผผลตรวมทงหนวยงานทรบผดชอบในการตรวจสอบ
คณภาพนามนของกรมธรกจพลงงานกระทรวงพลงงานคอ
After Before

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
128
การใชเครองมอขนาดใหญ ไดแก Gas chromatograph (GC) เพอทาการวดระดบเอทานอล วธนเปนการวดแบบมาตรฐานสากล ASTM D4815(7) เครองมอ GC นใช
สาหรบวดสารระเหยไดทวๆ ไปในปโตรเคมและเหมาะ สมทจะใชวดเอทานอลในแกโซฮอลดวย การวดดวย GC ใหคาความถกตองสง มความละเอยดเพยงพอ อยางไรกด เครองมอนไมสามารถเคลอนยายได การวดดวยวธนจงตองเกบรวบรวมตวอยางจากสถานบรการทวประเทศแลวสง
เขาวดในหองปฏบ ตการเทานน แตหลงจากการเกบตวอยางและรอเขาวดดวย GC นนระดบเอทานอลจะ
เปลยนไปไดอก ดงนน กวาตวอยางจะไดรบการตรวจสอบระดบเอทานอลทอานไดกเปลยนแปลงไปจากเดมแลว
Siam Gasohol Kit (SG-Kit): ชดเครองมอแบบเคลอนทสาหรบวดเอทานอลในแกโซฮอล ทแมนยาและถกตองเทยบเทา GC มาตรฐาน ASTM
คณะนกวจยจาก FIRST Labs ภาควชาเคม คณะวทยาศาสตร มหาวทยาลยมหดล เลงเหนถงปญหาน จงไดทาการคดคนและประดษฐชดเครองวดคณภาพแกโซฮอล
สยามขน (Siam Gasohol Kit หรอ SG-Kit) โดยในการวจยและพฒนาไดรบความอนเคราะหคาปรกษาจากทมฝายวจย
เชอเพลงและหลอลน สถาบนวจยและเทคโนโลย ปตท. และปรกษาหารอกบกรมธรกจพลงงาน กระทรวงพลงงาน เพอขอความเหนในการพฒนาจนประสบความสาเรจใน
การผลตเปนชดเครองมอวด (รปท 3) ผลงานชดนไดยนเสนอเพอขอจดสทธบตรในนามมหาวทยาลยมหดลแลว เมอป พ.ศ. 2550
หลกการ SG-Kit
SG-Kit ใชหลกการทมกระบวนการเกดขน 2 กระบวนการพรอมๆ กน คอ การสกดเอทานอลลงในชนนายาเคม และการเกดปฏกรยาระหวางเอทานอลทสกดไดกบนายาจนสเปลยนไป ซงระดบความเขมของสนนถกอานดวยเครองมอวด และแสดงผลทหนาจอของเครองเปนระดบรอยละของเอทานอล เชน 10%v/v เปนตน ซงกระบวนการทงหมดนถกควบคมดวยโปรแกรมอตโนมต
เพอใหมความแมนยาสง และการเทยบกบสารมาตรฐานกอนการวเคราะหในลกษณะเดยวกบการใชงานของ pH meter
คณลกษณะเดนของ SG-Kit
1. ความถกตองและแมนยาสงมากเทยบเทา GC ผลการเปรยบเทยบกบการวดดวย GC (ASTM D4815) ซงทมฝายวจยเชอเพลงและหลอลน สถาบนวจยและเทคโนโลย
รปท 3 แสดงภาพภายนอกและภายในชดเครองวด SG-Kit รน Exclusive class
ปตท. ใหความอนเคราะหทาการวดเพอเปรยบเทยบ ดงแสดงในรปท 4 ขอเดนพเศษของเครองมอน คอ อานคาไดละเอยดในชวงรอยละ 0.3
รปท 4 แผนภมแสดงปรมาณเอทานอลในแกโซฮอลทวเคราะห ดวย SG-Kit เปรยบเทยบกบวธมาตรฐาน (GC-FID) จานวน
12 ตวอยาง ซงทง 2 วธไดผลไมแตกตางกนอยางมนยสาคญทระดบความเชอมน 95% (Paired t-Test)
2. อานคาไดรวดเรวมาก คอภายใน 2 นาท จงเหมาะกบการใชงานภาคสนามทตองทราบคาทนท
3. ชดเครองมอมขนาดเลก บรรจในกระเปาเหลกบกนกระแทก ภายในกระเปามเครองมอ นายาและอปกรณครบครน เมอเปดกระเปา จงพรอมใชงานไดทนท
4. ขนตอนการใชงานงาย ผใชสามารถฝกการใชงานไดภายใน 2-3 ชวโมง
5. ชดเครองมอมราคายอมเยาและคาใชจายในการวดตากวาเมอใช GC 10 เทา คอคาใชจายเฉลยประมาณ 250 บาท/ตวอยางแกโซฮอล/สถานบรการ
คณลกษณะอนๆ ของเครอง SG-Kit แสดงในตารางท 1
SG-Kit เปนนวตกรรม
SG-Kit เปนผลงานคดคนวจยโดยคณะอาจารยและนกศกษา เปนผลงานใหม ยงไมมทใดนาเสนอมากอน ถอ
1
2
3
0
2
4
6
8
10
12
14
16
1 2 3 4 5 6 7 8 9 10 11 12Sample number
Etha
nol c
once
ntra
tion
% (v
/v) GC
SG-Kit

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
129
เปนนวตกรรมในวงการตรวจสอบคณภาพนามนเชอเพลง และเปนผลงานจากประเทศไทย ซงคาดวาตอไปจะสรางชอเสยงใหกบประเทศไทยในเรองของการสนบสนน
นโยบายพลงงานทดแทนในระดบชาตและนานาชาตตอไป
การใชSG-Kit วดคณภาพ ณ โรงผลตแกโซฮอล
นอกจาก SG-Kit เหมาะสาหรบการมเพอไวใชในรถออกตรวจคณภาพนามนภาคสนามแลว ยงเหมาะสมกบการตรวจคณภาพเพอวดระดบเอทานอลภายหลงการผสม
กบนามนเบนซนพนฐาน ณ โรงผลตแกโซฮอลดวยเพอควบคมคณภาพการผลต เพราะอานผลไดทนทหลงการผสม กอนทจะสงเขายนยนดวย GC อกครง หากตองการเรงการใชพลงงานทดแทน SG-Kit จงสอดรบกบนโยบายพลงงานแหงชาต
จดออนในการประชาสมพนธพลงงานทดแทนในปจจบน
แกโซฮอลถอเปนกาวแรกของการใชพลงงาน
ทดแทนในประเทศไทย ไดมการประชาสมพนธขาวสารเพอกระตนใหเกดความเชอมนในพลงงานทดแทนตวนมา
โดยตลอดตงแตเรมใหมการจาหนายเชงพาณชยเปนครง
แรกในป พ.ศ. 2544
การประชาสมพนธ เรมตนจากรณรงคการใช
พลงงานแกโซฮอล เพอใหประชาชนมความเชอมนกบ
พลงงานแกโซฮอล เพอใหประชาชนมความเชอมนกบคณภาพวา สามารถใชแกโซฮอลไดโดยไมมผลเสยตอเครองยนต เรมโดยเนนประชาสมพนธการใช E10 โดยเฉพาะการใชสอทางโทรทศน ทดาเนนการทงภาครฐและเอกชน ซงจะเหนวาตองใช เวลานานมากกอนทประชาชนจะยอมรบ
ตอมาเรมจงประชาสมพนธ E20 ซ งปจจบนผ บรโภคซงมรถยนตรนเกาและตองไปปรบเครองยนต
เสยกอนจงจะใช E20 ไดนน ยงมทไมมนใจอยเปนจานวนมากอยางไรกด จะเหนไดวาในการประชาสมพนธทกครง มเพยงการยนยนจากหลายฝายวาใชได แตยงขาดการประชาสมพนธในเรองวธการควบคมคณภาพแกโซฮอลท
ภาครฐและบรษทผผลตนามาใชวา มาตรฐานเชงปฏบตนนเปนอยางไร นบจากวนทผลตจนกระทงวนทเตมลงในรถยนต
ดงนน หากมการประชาสมพนธทด โดยเพมเตมดวยว า ปจจบนไดมการผลต เคร องมอ เพ อ ตดตามตรวจสอบคณภาพ นบตงแตผลตจนออกจากหวจายนามนจนกระทงถงรถยนตของผบรโภควา มการใชเครองวดตดตามโดยตลอด ประชาชนผใชรถกจะมนใจมากขนวา พลงงานแกโซฮอลสามารถใชได เครองยนตไมเสย ไมกระตก
ตารางท 1 คณลกษณะและประสทธภาพของชดเครองวดภาคสนามสยามแกโซฮอล
คณลกษณะ การตรวจวด ใชหลอด LED วดการดดกลนแสงทชวงความยาวคลนจาเพาะ
ประสทธภาพ ความแมนยา 98 % ความละเอยดในการวเคราะห + 0.3 % (v/v) ethanol ความเทยงตรง 3 % RSD ชวงการวด 5 – 20 % (v/v) ethanol ปรมาตรแกโซฮอลทใช 200 ไมโครลตร เวลาในการอานผล 2 นาท หลกการและผลการวด Single standard calibration. (อานผลในหนวย % (v/v) ethanol) นาหนก 8 กโลกรม หนาจอแสดงผล LCD 2 บรรทด แหลงจายไฟ 220 V และแบตเตอร

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
130
SG-Kit จดแขง กบนโยบายของกระทรวงพลงงาน หากกระทรวงพลงงาน เหนประโยชนจากนวต-
กรรม SG-Kit และเหนพองตอไปอกวาควรใหมการ
ผลกดนเชงนโยบายในแงของเทคนคการวดทคดคนดวย
ทมนกวจยระดบมหาวทยาลย โดยผลกดนใหมการใชงาน SG-Kit เพอตรวจสอบคณภาพแกโซฮอลทวประเทศ นอกจากจะเปนการคมครองผบรโภคในแงคณภาพ
เชอเพลงแลว ยงสามารถใช SG-Kit เปนสวนหนงในการประชาสมพนธโดยสอทางโทรทศน วทย และสงพมพไดดวย เนองจากประชาชนจะใหความสนใจยงขนทกๆ ครงทมการใชองคความรและบคลากรทางวชาการมาเกยวของ
ในการประชาสมพนธเรองตางๆ
SG-Kit กบการประชาสมพนธคณภาพแกโซฮอลใหมชนด E20 ของ บรษท ปตท. จากด (มหาชน)
ในการคดคนและวจย SG-Kit นน มหาวทยาลย มหดลไดรบคาปรกษาทดมาโดยตลอดจากฝายวจย
เชอเพลงและหลอลน สถาบนวจยและเทคโนโลย ปตท. ตลอดจนใหความรวมมอในการประเมนประสทธภาพของ SG-Kit ซงพบวามความแมนยาและถกตองใกลเคยงกบเทคนคมาตรฐานโดยเทยบเทาเครองมอใหญแบบ GC นอกจากน ยงไดใหการอนเคราะหเพมเตม โดยวาจางผลตแลว 1 ชด เพอใหฝายตรวจสอบคณภาพ คลงนามนพระโขนง ปตท. ทดลองใชงาน (เรมตงแต 27 ธนวาคม 2550)
ทงน ฝายประชาสมพนธ บรษท ปตท. อาจสนใจทจะใช SG-Kit เปนสวนหนงในการประชาสมพนธเชงรก สาหรบ E20 ซงเชอวาจะมประสทธภาพมากขน เนองจากประชาชนเรมมนใจการใชพลงงานทดแทนแกโซฮอลทม
เอทานอลทดแทนมากกวา E10 เดม โดยในครงน สามารถเนนเนอหาการประชาสมพนธในเรองทมการดาเนนการ
ตรวจสอบคณภาพอยางใกลชดดวยเครองมอ SG-Kit ซงมประสทธภาพดานความถกตองและแมนยาสง สามารถออกตรวจสอบไดอยางสมาเสมอดวยรถตรวจคณภาพของ ปตท. ทเปนรถเคลอนท ซงประชาชนสวนนอยเทานนท
ทราบวามการใชรถตรวจคณภาพออกตรวจอยแลวเปน
ประจา
สรป ชดเครองวดภาคสนามสยามแกโซฮอล (Siam Gasohol Kit) เขามามบทบาทชวยใหภาครฐ บรษทผผลต รวมถงผบรโภค ไดตระหนกถงการเขามามสวนรวมในการตรวจสอบคณภาพนามนเชอเพลง ซงผลงานนนบเปนหนงนวตกรรมในรอบ 50 ปแหงการสถาปนาคณะวทยาศาสตร
มหาวทยาลยมหดล โดยนวตกรรมนเนนใหเหนวาผลงานวชาการทพฒนาขนในระดบมหาวทยาลยสามารถนามา
ประยกตใชงานไดจรง ไมใชเปนผลงานขนหงอกตอไป อกทงยงมสวนชวยภาครฐในการประชาสมพนธใหผบรโภค
เกดความมนใจในการใชพลงงานทางเลอกอกดวย
กตตกรรมประกาศ งานวจยน ไดรบการสนบสนนจากสานกงาน
กองทนสนบสนนการวจย (สกว.)และศนยความเปนเลศดานนวตกรรมทางเคม (PERCH-CIC) สานกงานคณะ กรรมการการอดมศกษา กระทรวงศกษาธการ
เอกสารอางอง 1. กรมธรกจพลงงาน กระทรวงพลงงาน Available from:
URL: http://www.doeb.go.th/law/data/e85.pdf 2. บรษท ปตท. จากด (มหาชน) Available from: URL:
http://www.pttplc.com/Files/Document/Pdf/energy/ขอมลนามนแกโซฮอล.pdf
3. Battiste DR, Fry SE, White FT, Scoggins MW, Mc Williams TB. Determination of ethanol in gasohol by infrared spectrometry. Anal Chem 1981;53:1096-9.
4. Mendes LS, Oliveira FCC, Suarez PAZ, Rubim JC. Determination of ethanol in fuel ethanol and beverages by fourier transform (FT)-near infrared and FT-Raman spectrometries. Anal Chim Acta 2003;493:219-31.
5. Muncharoen S, Sitanurak J, Tiyapongpattana W, Choengchan N, Ratanawimarnwong N, Motomizu S et al. Quality control of gasohol using a micro-unit for membraneless gas diffusion. Microchim Acta 2009;164:203-10.
6. Rocha MS, Sim, Õ es-Moreira JR. A simple impedance method for determining ethanol and regular gasoline mixtures mass contents. Fuel 2005; 84:447-52.
7. The American Society of Testing and Materials. Designation: D 4815-03 Standard test method for determination of MTBE, ETBE, DIPE, tertiary-amyl alcohol and C1 to C4 alcohols in gasoline by gas chromatography. West Conshohocken, USA; 2003. p. 854.
8. Zinbo M. Determination of one-carbon to three-carbon alcohols and water in gasoline/alcohol blends by liquid chromatography. Anal Chem 1984;56:244-7.

131
Loop-mediated Isothermal Amplification (LAMP) Method: an Easy and Fast Gene Amplification Method for Detection of Malaria Parasites Sureemas Buates Department of Microbiology, Faculty of Science, Mahidol University.
Abstract
Malaria is one of the world’s most important parasitic infections in humans. A sensitive, rapid and accurate diagnostic test for malaria is not only crucial for proper treatment of the individual patient but also essential for malaria control programs and epidemiological surveillance, which is needed to observe trends in malaria prevalence and anti-malarial drug resistance. Conventional microscopy is the gold standard for malaria diagnosis. Although it is an accurate tool, it requires time and a well-trained staff. Alternative methods [e.g. nested polymerase chain reaction (PCR), real-time quantitative PCR and immunochromatographic tests] have had several limitations making it difficult to adopt them for routine testing. Currently, a novel, fast, simple, sensitive and inexpensive method called loop-mediated isothermal amplification (LAMP) has been used successfully to amplify the target gene for detection of malaria parasites in both humans and mosquitoes. This method is easier and faster than other methods, in addition to being as sensitive and dependable as PCR. The use of LAMP for malaria detection is reviewed in this paper,
Keywords: loop-mediated isothermal amplification (LAMP); malaria
Introduction
Malaria is one of the world’s most important parasitic infections and is the third leading cause of death attributable to an infectious disease worldwide. Throughout the world, malaria causes disease in 300-500 million people and kills approximately 1.1-2.7 million people each year predominately in sub-Saharan Africa(20). The causative agent of malaria is an intracellular protozoan parasite of the genus Plasmodium. At least four species of malaria infect humans, P. falciparum, P. vivax, P malariae, and P. ovale. Among the four species, P. falciparum is the major cause of severe clinical malaria and death. The transmission of malaria to humans is via the bite of an infected female Anopheles mosquito. Once inside the human host, parasites multiply exponentially in the liver hepatocytes and then in infected red blood cells. When a mosquito ingests human blood containing the sexual stages of Plasmodium (an infective stage to mosquitoes), the parasites undergo another reproductive phase inside the mosquito before being passed on to a
new human host to continue the cycle. A unique characteristic of P. vivax and P. ovale is the production of dormant liver stages called hypnozoites that are responsible for relapse(2).
A key to effective management of malaria is rapid and accurate diagnosis to guide prompt and effective therapy. This is crucial to avoid non-target effects, to delay the develop-ment of resistance, and to avoid the additional cost of alternative drug therapies. Therefore, a cost-effective, simple, and rapid method for malaria diagnosis is an urgent need. In this paper, the drawbacks of existing malaria diagnosis including conventional microscopy, nested polymerase chain reaction (PCR), real-time quantitative PCR and immunochroma- tographic tests (ICT) are mentioned. The advantages of loop-mediated isothermal ampli-fication (LAMP), a new and outstanding gene amplification procedure for the diagnosis of malaria infection, are reviewed in terms of: simplicity, rapidity, cost-effectiveness, high sensitivity and specificity.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
132
Malaria Diagnosis
Conventional diagnosis: For malaria diagnosis, light microscopy of thick and thin blood smears stained with Giemsa, Wright’s or Field’s stain is the gold standard method. It is cheap, rapid and enables identification of Plasmodium species and parasite quantification, both of which are essential to assess disease severity and to prescribe adequate therapy. However, it has several disadvantages, i.e. it is time-consuming and requires a microscope. Interpretation of the result requires considerable expertise, especially in the case of low levels of parasitemia or sequestered parasites. Miss interpretation potentially leads to false negative results or unreliable species determination leading to therapeutic delays. Its sensitivity is only moderate, with a detection limit of 50 to 500 parasites/μl (thick versus thin film smears)(6). Additionally, mixed infections are often missed, particularly with the presence of P. malariae or P. ovale(4).
Alternative diagnosis: In recent years, new technological methods have been evaluated as alternatives to microscopy including nucleic acid amplification methods, e.g. nested PCR and real-time quantitative PCR approaches. They have shown greater sensitivity as compared to traditional microscopy, are able to detect parasitaemia in samples containing as low as 1-5 parasites/μl, and are more effective in detecting mixed infections(4,5,11,14-18). However, in addition to their high cost the use of these tests is limited by the need for: specialized instruments, many costly reagents, skilled operators, long assay run times, and poor applicability and/or availability in the field. A recently developed immuno- chromatographic test represents a very simple, rapid and reliable tool for malaria diagnosis. It is commonly relied on for the detection of the P. falciparum histidine-rich protein-2 (HRP-2) or Plasmodium-specific lactate dehydrogenase (pLDH) antigen. Its sensitivity is only moderate, able to detect 50 to 100 parasites/μl, but its use has some limitations(6,13). It cannot be used to determine the parasitaemia level. It shows rather low sensitivity in the detection of malarial species other than P. falciparum. It cannot be
used to differentiate between non-P. falciparum species and cannot discriminate accurately between a mixed infection and a P. falciparum infection.
Requirements for LAMP
LAMP is an easy, fast, specific and inexpensive nucleic acid amplification method. It is easier and faster, besides being as sensitive and dependable as compared to PCR. From its feature, LAMP represents an evolving techno-logy that can be applied in countries where cost-effectiveness is a crucial factor and simplicity of diagnostic test performance and training of personnel are also major consi-derations.
The LAMP Method
Principle of LAMP: LAMP, a sensitive strand displacement technique, was originally described by Notomi et al.(9) It can be per-formed at an isothermal temperature using one type of enzyme as strand denaturation is done by strand displacement. Its specificity is ex-tremely high since it uses four primers which are constructed from six areas on the target DNA as shown in Figure 1. LAMP amplifies the target DNA from a few copies to 109 copies in less than an hour with no special reagents required. The LAMP reaction mechanism is briefly illustrated in Figure 2. LAMP reaction proceeds through two steps: 1) starting structure-producing step (Figure 2a) and 2) cycling amplification step (Figure 2b), using DNA polymerase with high strand dis-placement activity. The first step requires all four basic primers. As shown in Figure 2a, at the starting structure-producing step, one of the inner primers (i.e. FIP) anneals to the target region and primes the replicative DNA synthesis by Bst DNA polymerase. Then, the outer primer, F3, hybridizes to the area outside FIP of the target DNA and starts strand displacement DNA synthesis, releasing newly synthesized DNA strand. With the same process of BIP and B3, this generates the single-stranded dumbbell-DNA structure (structure 5) which is looped out at each end. This DNA construct is the starting substrate for

the cyclprogressBIP. In converteprimed Dsingle stinitiates thereforestructureis made.5 and 7,made breactionseries oare prodcan be utranscripand DNAenzyme DNA poless thancan be primer c
Figure 1
ing amplificses using onlthis step, th
ed to a stem-DNA synthetranded regio
strand dise, the come (structure 7. In the cycl, the elongat
by the elon. At the enf stem-loop
duced. Besidused to detecption-LAMPA amplificatmix (AMV
olymerase) an an hour in
further acccalled loop pr
1 Loop-medconstructefrom the the F2 sePrimer) issequence composed
วทยาศาส
cation step (Fly two inner e starting sub-loop DNA sesis. Next, Fon of the stemsplacement mplement o7) or the ‘turle reaction beted structurengation andnd of the aut
DNA with es detection ct target RN
P (RT-LAMPtion can be areverse tran
at a constann one step. celerated usrimers. Usi
diated isotheed from 6 dis5' end). C is
equence at thes composed oat the 3' end
d of the B3 se
สตรเพอการพฒนา : ร
Figure 2b) wprimers, FIPbstrate is qustructure by FIP hybridizm-loop DNADNA synthof the starn over’ struetween struces (9-12) ared recycling to-cycling stdifferent lenof DNA, LA
NA by the revP) reaction. achieved usinnscriptase andnt temperatur LAMP rea
sing an optng loop prim
ermal amplifistinct regionsa complemene 3' end and of the F3 sequd and the B1equence.
รวมบทความวชาการ
which P and ickly self-
zes to A and hesis, arting cture
ctures also step
ep, a ngths AMP verse RT
ng an d Bst re in ction ional
mers
incDNreaoneffno
prPCLApareqseacpr(bpeatemThExexso
ication (LAMs on the targentary sequencthe F1c sequ
uence. BIP (B1c sequence
ร ฉลองครบรอบ 50 ป
creases the nNA synthesaction; the re
ne-third to officient methostics(8)
.
Design rimers is muCR and apprAMP amplifair of inner aquirement fots of loop p
cceleration(8).rimers are usp), respectiv
ach primer mperatures ahe primers xplorer versxplorer.jp/elaoftware to des
MP) primer det DNA (F3, ce. FIP (Forwuence at the Backward Innat the 5' end
ป แหงการสถาปนาคณ
number of tsis and theeaction time
one-half, mahod to be us
of primers: uch more coropriate desigfication. Fo
and outer primfor LAMP rprimers can . The lengthsually over 4vely. The tec
and the oare explainedcan be dession 4 sofamp4.0.0/indsign LAMP p
design. Four F2, F1, B1c,
ward Inner Pr5' end. F3 Prner Primer) i
d. B3 (Backw
ณะวทยาศาสตร มหา
the starting pe specificity can be shor
aking LAMPsed in disea
Designing oomplex thangn is a cruciour basic prmers, are a mreaction. Abe used for h of inner a40 and 25 bchnical condoptimized ad by Notomsigned usingftware (httpdex.html), a primer.
types of pri B2c and B3rimer) is comrimer (Forwais composed oward Outer P
าวทยาลยมหดล
13
point for y of the rtened by P a more ase diag-
of LAMP n normal al key in rimers, a minimum dditional reaction
and outer ase pairs
ditions of annealing
mi et al.(9) g Primer p://primer
special
imers are c regions
mposed of ard Outer of the B2
Primer) is
33

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
134
Figure 2 Schematic representation of the mechanism of Loop-mediated isothermal amplification (LAMP) reaction. LAMP progresses through two steps, (1) starting structure-producing step (F.2a) and (2) cycling amplification step (F.2b).
B1
B1c
B2
F1c
F1
F2c3’ 5’
F1
F1c
F2
3’5’
B1c
B2
BIP
B2c
B1c
B1
3’5’F1c F2 F1 B1c B2c B3c
3’5’F1c F2 F1 B1c B2c B3c
Target DNAF3 F2 F1 B1c B2c B3c
F3c F2c F1c B1 B2 B3
5’
3’ 5’
3’Target DNAF3 F2 F1 B1c B2c B3c
F3c F2c F1c B1 B2 B3
5’
3’ 5’
3’
F2
F1
F1c B2
B1
B2c
B1c
B1c
B1
F1
F1c
F1c
F1 F2c
F2c
F2
F1
F1c B2
B1
B2c
B1c
B1c
B1
F1
F1c
F1c
F1 F2c
F2c
F2
F1
F1c B2B2cB1c
B1
F1
F1c
F1c
F1 F2c
F1c
F2 F2c
B1
B1c
F2
F1
F1c B2B2cB1c
B1
F1
F1c
F1c
F1 F2c
F1c
F2 F2c
B1
B1c
F2
F1c F1
F2c F1cB1c
B2c
B1
F1
B1
B1c B2c
B2
B1
B1cF2
F1c F1
F2c F1cB1c
B2c
B1
F1
B1
B1c B2c
B2
B1
B1c
F1
F2c F1cB1c
B2c
B1
F1
B1
B1c B2c
B2
B1
B1c
F2F1F2c
F1c
B1c
B2c
B1
F1
B1
B1c B2c
B2
B1
B1cF1c
B2
B1c
F2F1F2c
F1c
B1c
B2c
B1
F1
B1
B1c B2c
B2
B1
B1cF1c
B2
B1cB1
B1c
B2
F1c
F1
F2c3’ 5’
F2
F1c
FIP
B1
B1c
B2
F1c
F1
F2c3’ 5’
F2
F1c
FIP
F2
F1c
FIP
F1
F2
F1c
B1c B2c B3c
5’
3’
BIP 5’
3’
B3 Primer
F1
F2
F1c
B1c B2c B3c
5’
3’
BIP 5’
3’
B3 Primer
FIP
3’
5’
F3F3 Primer
Bst DNA polymerase
3’ 5’
F3c F2c F1c B1 B2 B3
FIP
3’
5’
F3F3 Primer FIP
3’
5’
F3F3F3 Primer
Bst DNA polymerase
3’ 5’
F3c F2c F1c B1 B2 B3
3’B1c B2c B3c
5’B1 B2 B3
F2 F1F1c5’
F3 Primer
F3
3’F3c F2c F1c
3’B1c B2c B3c
5’B1 B2 B3
F2 F1F1c5’
F3 Primer
F35’
F3 Primer
F3
3’F3c F2c F1c
F.2a
F.2b
10.
9.
11.
8. 6.
5.
12. 7.
5.
2.
3.
1.
4.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
135
Requirements for LAMP reaction: For DNA amplification, LAMP reaction consists of primer mixture, 2x reaction buffer with betaine to facilitate DNA strand separation, Bst DNA polymerase with strand displacement activity, dNTPs, DW, and the target DNA (Eiken Genome Co. Ltd, Tokyo, Japan). For RNA amplification, an enzyme mix (AMV reverse transcriptase and Bst DNA polymerase) was used. LAMP reaction does not need to denature double stranded DNA into a single stranded form. The reaction mixture is incubated at isothermal conditions (normally between 60-65°C) within less than 1 hr relying on the effectiveness of the developed primers and template DNA. The reaction is inactivated at 80°C for 5 min (Figure 3). The major advantage of LAMP is that only a water bath or heating block is used to maintain the required temperature. The reaction can also be done using the real-time or the end point turbid-meter, a specific device for LAMP.
Detection of Amplified Products
Visual detection: LAMP amplified products can easily be assessed by visualization the amount of white precipitate of magnesium pyrophosphate (an insoluble salt) produced during DNA elongation(7). The production of magnesium pyrophosphate is made from combination of the pyrophosphate ion released from the substrate [deoxyribonucleotide triphos- phates (dNTPs)] as a by-product and magnesium ion (a divalent metallic ion) in LAMP reaction buffer. The turbidity of magnesium pyrophosphate is produced in proportion to the amount of amplified products. LAMP can produce a remarkably large amount of DNA (as high as 400 μg/ml) allowing the detection of turbidity by only the naked eye. LAMP products can be seen on a UV transilluminator or a black light by incor-porating intercalating agents (ethidium bromide, etc.) directly into LAMP-amplified tubes as shown in Figure 3(9). The visual detection can be improved by adding manganese ion and calcein, a fluorescent metal indicator, to the reaction to increases the sensitivity and to help
to visual discriminate the results as recently described by Tomita et al.(19)
Electrophoresis detection: The ampli-fied product can be observed by an agarose gel stained with an intercalating agent e.g. ethidium bromide or SYBR Green I. LAMP amplified products are not observed as a single band as in PCR. As LAMP generates products of different lengths of stem loop structures, the product sizes vary from 300 bp upto the loading well, which can be seen as the smear and the ladder pattern of the band at the bottom of the gel as shown in Figure 3. This is a typical pattern for LAMP. Because of the amplified product is extremely high at the end, detecting LAMP products by gel electrophoresis can cause contamination in subsequent amplifications. Therefore, accurate prevention of contamination is essential.
Figure 3 Schematic representation of the LAMP
procedure. The LAMP reaction is done at a constant temperature. The amplified product can be detected by an agarose gel stained with an intercalating agent or by visualization of the turbidity or the fluorescence in the reaction.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
136
Real-time detection: The turbidity and fluorescent detection of LAMP products can be real-time monitored by a real-time turbidimeter and by using this machine; the quantitative LAMP analysis of the target genes can be determined.
Application of LAMP for Detection of Malaria Parasites
To date, there are only 4 publications regarding the use of LAMP to detect malaria parasites. The first report was described by Poon et al.(12) In this study, the authors developed a novel strategy using LAMP to detect P. falciparum from heat-treated blood samples from 102 patients with P. falciparum infection and 100 healthy volunteers. LAMP primers were designed by targeting the highly conserved 18S ribosomal RNA (rRNA) gene of falciparum malaria. As compared to PCR, the sensitivity and specificity of LAMP were 95% and 99%, respectively. LAMP detection limit was ∼ 5 parasites/μl. Interpreting LAMP reac-tion with the naked eye gave consistent results with those deduced from the real-time turbid-meter. Poon et al.(12) calculated that the running cost for LAMP is ∼ 1/10 of that of normal PCR.
Paris et al.(10) evaluated LAMP for the detection of P. falciparum in 115 patients with fever using two different DNA template preparation methods, i.e. QIAamp DNA MiNi Kit and heating of samples, and three different endpoints [1) commercial DNA extraction method and gel electrophoresis, 2) heat-treated samples and gel electrophoresis, and 3) heat-treated samples and turbidity visual detection]. The sensitivity of LAMP using conventional DNA extraction and gel electrophoresis showed the highest agreement with nested PCR [a reference method (81.7%, κ = 0.64)], with sensitivity of 76.1% and specificity of 89.6%. These results were in contrast with a higher accuracy in the study conducted by Poon et al.(12) These two studies used the same primer sets for the detection of P. falciparum. Regarding the discrepancy between the two studies, Paris et al.(10) mentioned that it could have been from the use of the same primer set between PCR and LAMP in the study by Poon
et al(12). This could have resulted in an overestimation of the quality of LAMP.
The use of LAMP to detect genus and species specificity of all four species of human malaria, P. falciparum, P. vivax, P. malariae, and P. ovale, for clinical diagnosis has been validated by Han et al.(3) This study was conducted using 68 samples positive for malaria and 53 samples negative for malaria by conventional microscopy which was used as the gold standard to compare the sensitivity and specificity with nested PCR and LAMP. The target sequence of LAMP and nested PCR primers for Plasmodium genus specificity consists of conserved regions of 18S rRNA gene only and for species specificity consists of both conserved and variable regions of the same gene. LAMP demonstrated that a detection limit of the target 18S rRNA genes for P. malariae and P. ovale is 10 copies and for the genus Plasmodium, P. falciparum, and P. vivax is 100 copies. The results from this study showed that LAMP had a resemble sensitivity and a greater specificity as compared to the reference method and LAMP produced results resemble to those of nested PCR in a shorter turnaround time.
Recently, Aonuma et al.(1) demonstrated that LAMP was appropriate for diagnosis of Plasmodium in both humans and mosquitoes. LAMP was applied to detect Plasmodium-carrying mosquitoes to survey pathogen-transmitting vectors. In this study, a Plasmodium-transmitting experimental model was performed using P. berghei and Anopheles stephensi. The detection sensitivity limit of the target P. berghei SPECT2 gene was 1 × 102 of
purified Plasmodium parasites collected from both the blood and vector stages (sporozoites). This detection limit was enough for reliable identification of infectious mosquitoes. It was revealed that LAMP has its ability to detect both Plasmodium oocysts and sporozoites from an “all-in-one” template using whole mosquito bodies. Moreover, LAMP successfully detected even just one oocyst in an infectious mosquito, a level which can easily be missed by a conventional microscopy.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
137
Conclusions
LAMP for detection of malaria parasites is simple; it does not require thermal cyclers, expensive reagents (time-consuming steps) to purify DNA for efficient DNA amplification, down-stream processing for product detection, or skilled technicians. The assay positivity can be verified by visual interpretation. It is highly specific since the reaction progresses using four different highly specific primers. As compared to normal PCR, LAMP has higher ability to amplify target DNA, shorter assay time only one third, and lower cost for running the assay ∼ 1/10. As compared to nested PCR, LAMP is at least equal or more sensitive and similar specificity for detection of target DNA. From its feature, LAMP is a promising technique, which the first time enables a molecular diagnosis of malaria parasites in both humans and mosquitoes in a setting with limited resources. Therefore, LAMP can be useful for clinical diagnosis and active surveillance of malaria parasites in malaria endemic countries.
Acknowledgements
The author thanks Mr. Sirasate Bantuchai for his kind assistance in drawing the figures presented in this review.
References 1. Aonuma H, Suzuki M, Iseki H, Perera N, Nelson
B, Igarashi I et al. Rapid identification of Plasmodium-carrying mosquitoes using loop-mediated isothermal amplification. Biochem Biophys Res Commun 2008;376:671-6.
2. Greenwood BM, Fidock DA, Kyle DE, Kappe SH, Alonso PL, Collins FH et al. Malaria: progress, perils, and prospects for eradication. J Clin Invest 2008;118:1266-76.
3. Han ET, Watanabe R, Sattabongkot J, Khuntirat B, Sirichaisinthop J, Iriko H et al. Detection of four Plasmodium species by genus- and species-specific loop-mediated isothermal amplification for clinical diagnosis. J Clin Microbiol 2007; 45:2521-8.
4. Kilian AH, Metzger WG, Mutschelknauss EJ, Kabagambe G, Langi P, Korte R et al. Reliabi- lity of malaria microscopy in epidemiological studies: results of quality control. Trop Med Int Health 2000;5:3-8.
5. Kimura K, Kaneko O, Liu Q, Zhou M, Kawa-moto F, Wataya Y et al. Identification of the
four species of human malaria parasites by nested PCR that targets variant sequences in the small subunit rRNA gene. Parasitol Int 1997;46: 91-5.
6. Moody A. Rapid diagnostic tests for malaria parasites. Clin Microbiol Rev 2002;15:66-78.
7. Mori Y, Nagamine K, Tomita N, Notomi T. Detection of loop-mediated isothermal amplifi-cation reaction by turbidity derived from magnesium pyrophosphate formation. Biochem Biophys Res Commun 2001;289:150-4.
8. Nagamine K, Hase T, Notomi T. Accelerated reaction by loop-mediated isothermal amplifica-tion using loop primers. Mol Cell Probes 2002; 16:223-9
9. Notomi T, Okayama H, Masubuchi H, Yonekawa T, Watanabe K, Amino N et al. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res 2000;28:E63.
10. Paris DH, Imwong M, Faiz AM, Hasan M, Yunus EB, Silamut K et al. Loop-mediated isothermal PCR (LAMP) for the diagnosis of falciparum malaria. Am J Trop Med Hyg 2007; 77:972-6.
11. Perandin F, Manca N, Calderaro A, Piccolo G, Galati L, Ricci L et al. Development of a real-time PCR assay for detection of Plasmodium falciparum, Plasmodium vivax, and Plasmodium ovale for routine clinical diagnosis. J Clin Microbiol 2004;42:1214-19.
12. Poon LL, Wong BW, Ma EH, Chan KH, Chow LM, Abeyewickreme W et al. Sensitive and inexpensive molecular test for falciparum mala-ria: detecting Plasmodium falciparum DNA directly from heat-treated blood by loop-mediated isothermal amplification. Clin Chem 2006;52: 303-6.
13. Proux S, Hkirijareon L, Ngamngonkiri C, McConnell S, Nosten F. Paracheck-Pf: a new, inexpensive and reliable rapid test for P. falcipa-rum malaria. Trop Med Int Health 2001;6:99-101.
14. Rougemont M, Van Saanen M, Sahli R, Hinrikson HP, Bille J, Jaton K. Detection of four Plasmodium species in blood from humans by 18S rRNA gene subunit-based and species-specific real-time PCR assays. J Clin Microbiol 2004;42:5636-43.
15. Singh B, Bobogare A, Cox-Singh J, Snounou G, Abdullah MS, Rahman HA. A genus- and species-specific nested polymerase chain reaction malaria detection assay for epidemio-logic studies. Am J Trop Med Hyg 1999;60:687-92.
16. Singh B, Kim Sung L, Matusop A, Radhakrish-nan A, Shamsul SS, Cox-Singh J et al. A large focus of naturally acquired Plasmodium

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
138
knowlesi infections in human beings. Lancet 2004;363: 1017-24.
17. Snounou G, Viriyakosol S, Jarra W, Thaithong S, Brown KN. Identification of the four human malaria parasite species in field samples by the polymerase chain reaction and detection of high prevalence of mixed infections. Mol Biochem Parasitol 1993;58:283-92.
18. Snounou G, Viryakosol S, Zhu X P, Jarra W, Pinheiro L, Rosario VE et al. High sensitivity of detection of human malaria parasites by the use of nested polymerase chain reaction. Mol Biochem Parasitol 1993;61:315-20.
19. Tomita N, Mori Y, Kanda H, Notomi T. Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products. Nat Protoc 2008;3:877-82.
20. WHO Expert Committee on Malaria: 20th Report (World Health Organization, Geneva, 2000). Available from: URL: http://www.rbm. who.int/docs/ecr20.pdf

139
Reporter Gene Assays for Anti-TB Drug Screening
Chartchai Changsen
Department of Microbiology, Faculty of Science, Mahidol University.
Abstract
Tuberculosis continues to be a major global public health problem, since it is now the leading infectious disease in the world in addition to the increasing prevalence of multidrug-resistance Mycobacterium tuberculosis (MDR-TB). Moreover, the need for a prolonged current anti-TB chemotherapy, which is a consequence of the presence of persistent M. tuberculosis under hypoxic milieu within granulomas and increasing emergence of drug-resistant strains, poses a significant problem for effective control. These situations have stimulated much scientific interest in developing new antimycobacterial agents. Therefore, treatment of patients with M. tuberculosis strains resistant to existing drugs, and shortening of the duration of short-course treatment to improve patient compliance should be established. Many emerging fluorescence-technologies for high-throughput applications are being developed for both in vitro and cell-based high-throughput screening (HTS). Several types of reporter gene assays are also available for use in mycobacteria. The majority of such tools are based on classical reporter genes such as lacZ, cat, and luciferase genes. As a result of the importance of the intracellular lifestyle for many pathogenic mycobacterial species, it seems desirable to develop approaches for visualization of mycobacteria infecting inside macrophages and monitoring their gene expression in intracellular environment. Along with an assay that is capable of non-invasive detection and is convenient, these concepts encouraged the recent development of green fluorescent protein (GFP) reporter systems. Recent reporter-based applications for anti-TB drug screening will be reviewed and limitations associated with the detection of fluorescence signal in fluorescence-based HTS assays demonstrated by previous investigations will be summarized. A recent approach to achieve a high-level GFP expression directed by an acetamidase promoter in mycobacteria will be illustrated. Potential directions of development of reporter-based system for anti-TB drug discovery will also be discussed.
Keywords: GFP; luciferase; reporter gene; acetamidase promoter; drug screening; mycobacteria
Introduction
A number of approaches may be taken to identify lead compounds that inhibit cellular activity or a particular target of an infectious agent. The most comprehensive approach is high throughput screening (HTS) and indeed this may be the only way forward when there are no other clues to guide discovery. HTS have provided a way for the identification of primary hits. It is common to screen a massive number of compounds, maximizing the structural diversity by including synthetic compounds and natural products. One of main challenges faced in the process of drug discovery is the development of the efficient screening of large libraries of these compounds and the identi-
fication of new lead drug candidates. In the case of tuberculosis, the target will differ according to whether the final drug will be used to treat actively growing or persisting organisms. Experimental evidence to validate the target may include the demonstration that the target is essential at a particular stage during the infection process(23). In addition, the compound will be evaluated for cytotoxicity in mammalian cultures. The next stage of screening uses a macrophage infection assay to determine if the compound can inhibit the growth of Mycobacterium tuberculosis in its intracellular environment. Generally, fluorescence-based assay technologies can offer equivalent sensitivity to other detection systems (radioactive-based methods), but have the added

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
140
advantages of greater versatility, stability, capability of HTS, safe handling, and ease of disposal.
Reporter Gene Assays
The term reporter gene is used to describe a gene with a readily quantifiable phenotype that can be distinguished easily over a background of endogenous proteins. Reporter gene-based assays have been established as popular tools in a variety of applications, such as studying of gene expression and promoter analysis(6), gene delivery system(26), drug screening(20), cyto-toxicity(14) and other cell-based assays. The main advantages of the reporter gene assays are higher sensitivity, reliability, easy detectable system, convenience, and adaptability to large-scale screening. A number of reporters are available in either naturally occurring or genetically engineered form. These include β-galactosidase(20), chloramphenicol acetyl trans-ferase (CAT)(27), firefly or bacterial luciferase (luminescent protein), and green fluorescent protein (GFP) or red fluorescent protein (RFP) as well as their mutants(11,19). Excellent reporter genes should be environmentally safe, not toxic to cells, easy to use and highly sensitive. They should support a variety of detection methods, yield high signal-to-noise ratio and have wide dynamic response. Only luminescent and fluorescent proteins fulfill most of these criteria.
Luminescent and Fluorescent Proteins
Luminescent and fluorescent molecules are widely used as probes for monitoring growth and other cellular activities in a number of applications in diverse research areas. Because they are proteins that can be genetically engineered and expressed continuously in living cells, they have been applied to a majority of biological studies to investigate cellular actions, such as localizing specific proteins in viruses, organelles, cells as well as multicellular orga-nisms.
Luciferase refers to a family of enzymes that catalyze the oxidation of various substrates (e.g. luciferin), resulting in light emission. The most commonly used luciferases for reporter gene assays are bacterial luciferases (Vibrio
harveyi and V. fischeri)(15), firefly luciferase (Photinus pyralis)(7), and more recently Renilla luciferase (from the bio-luminescent sea pansy, Renilla reniformis)(29). The development of luciferase assays was investigated in which cell metabolism (ATP and O2) is determined using either bacterial or firefly luciferase. The bacterial luciferase uses reduced riboflavin phosphate (produced by viable cells) to oxidize an added aldehyde substrate, which is accompanied by the production of light at 490 nm. Firefly luciferase is dependent upon ATP generated by cells to decarboxylate luciferin, resulting in the production of light at 562 nm. Light production may be measured easily using a luminometer in a high-throughput system. However, the application of the luminescent proteins is somewhat restricted by the require-ment of addition of a substrate (i.e. luciferin or η-decacyl aldehyde).
Fluorescence assays based on Aequorea victoria gfp gene encoding GFP or its mutants have been used widely. GFPs have several properties, which are advantageous for the use as a reporter for bacterial viability and growth, including no substrate requirement, low toxicity, continuous production during replication, ease of imaging and quantification with a variety of techniques including fluorescence microscopy(1), confocal scanning laser microscopy(24), flow cytometry(12), and spectrofluorometry(2,4).
GFP Applications in Mycobacteria
Several types of reporter gene assays are available for use in mycobacteria (table 1). The majority of such tools are based on the classical reporter genes such as lacZ, cat, and luciferase genes. As a result of the importance of the intracellular lifestyle for many pathogenic mycobacterial species, it is desirable to develop approaches for visualization of mycobacteria infected inside macrophages and for monitoring their gene expression in intracellular environ-ment. Along with an assay that is capable of non-invasive detection and is convenient, these concepts have encouraged recent developments of GFP reporter systems. Like with other bacteria, GFP-based assay is widely used in a

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
141
Table 1 Commonly used reporter genes in mycobacteria.
variety of mycobacterial studies including moni-toring in vivo gene expression(8), in vivo traffick- ing(22), promoter isolation(30), protein locali-zation(5), antiseptic screening(31), and drug screening(2,4,9).
In Vitro Drug Screening Methods for Mycobacterium tuberculosis
The recent worldwide increase in the incidence of tuberculosis has caused a renewed interest in evaluating the activity of new compounds(25) and new analogs of existing drugs(16) to combat tuberculosis and the significant emergence of multidrug-resistance M. tuberculosis (MDR-TB) cases. Also, there is an urgent need to generate new lead compounds and drugs rationally designed for new specific targets in mycobacteria and to establish rapid and reliable screening methods to evaluate large numbers of compounds generated from natural biodiversity and combinatorial chemistry. The most straightforward method of identifying lead compounds is to screen for agents that have activity against M. tuberculosis in vitro. However, M. tuberculosis has the disadvantage of being a slow grower. As a result, screening tests can take several weeks to finish. Biosafety level 3 containment facilities are required because this pathogen is a highly infectious microorganism. These impede the ability to
execute screening of vast numbers of compounds. In general, conventional broth and agar-based culture methods for susceptibility testing are laborious, time consuming and expensive. In broth dilution methods, which are more common used for screening minimum concentrations required to inhibit bacterial growth, a series of tubes or wells containing serial dilutions of the extracts in inoculated broth is required.
Established Standard Methods for Drug Screening in Mycobacteria
Rapid radiometric methods utilizing measurement of generated radioactive carbon dioxide (BACTEC™ 460 TB system; Becton Dickinson, Sparks, Md.) have been established for in vitro testing of antimycobacterial compounds or for susceptibility testing of clinical isolates(17,21) and can provide results within 4 to 8 days compared with 3 to 4 weeks for conventional methods. The greater sensitivity of this method allows rapid diagnosis and convenient quantitation of the therapeutic effect of anti-TB drugs. Nevertheless, the BACTEC system has limited throughput due to its high cost and technical difficulties involved in measuring radioactive carbon dioxide, and the disposal of radiolabeled materials. Also the
Reporter gene Advantage Disadvantage
Chloramphenicol acetyltransferase( CAT)
No endogenous activity; automated ELISA available.
Narrow application range; use of radioisotopes.
β-Galactosidase
Well-characterized, simple colorimetric readout; sensitive bio- or chemilumi- nescent assays.
Endogenous activity in mammalian cells; stability.
Luciferase (bacterial or firefly)
High specific activity; no endogenous activity; broad dynamic range; well-characterized and convenient assays.
Requires substrate (luciferin or aldehyde) and presence of O2 and ATP; not suitable for mammalian cells.
Green fluorescent protein (GFP) (jellyfish)
Autocatalytic fluorescence (no substrate needed); a variety of detection techniques available; mutants with altered spectral and intensity qualities available.
Requires presence of O2 for chromophore formation; low to moderate sensitivity (no signal amplification); quite stable and detectable background fluorescence.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
142
large volume of BACTEC™ media requires large amounts of tested compounds. Another widely-used established method utilizes a redox dye, Alamar blue (resazurin), which changes color from blue to pink in the presence of viable M. tuberculosis(18). This assay has the advantage of being simple and does not require radioactive substrate, but takes several days (from 4 to 7) to obtain antimycobacterial results. The need to preload the dye, its expense as well as laborious and invasive procedures involved have impeded its optimal performance and development into a high throughput screening.
Reporter Gene Assay as an Alternative Tool for Anti-TB Drug Screening
A number of alternative HTS assays have been developed using reporter genes such as β-galactosidase, luciferase and GFP as a viability marker. β-Galactosidase has been used for rapid in vitro screening of antimycobacterial drugs and a good correlation was found between inhibition of β-galactosidase production and viability of recombinant rapidly growing M. aurum in the presence of anti-TB drugs(10). However, the assay has not been applied for M. tuberculosis because of the requirement for cell lysis. Thus this reporter assay is not amenable for the large-scale drug screening.
The use of luciferase reporter strains of mycobacteria expressing Vibrio (luxAB) or firefly (FFlux) luciferase for rapid susceptibility testing or rapid screening of antimycobacterial compounds has been widely reported(13) due to its rapid turnaround time (2 to 4 days). Despite the fact that it is a well-characterized radio-isotope-free system whose susceptibility results showed a good correlation with that of standard BACTEC™ method, it has been pointed out that while luminescence provides an immediate measure of physiological status in bacteria, the ability to form colonies is influenced by drug-induced changes that occur during the subsequent weeks in culture and that front-line anti-TB drugs exert variable effects on luminescence output in short-term assays(28). These limitations of the luciferase readout as a marker for drug activity and cell growth in addition to requirement of preloaded substrates
(luciferin and aldehyde) indicate potential roles of other alternative reporter constructs in further optimization of the reporter assay for drug screening.
Previously, recombinant M. tuberculosis H37Ra strain carrying Phsp60::gfp construct has been utilized in GFP microplate assay (GFPMA) to assess minimal inhibitory concentration (MIC) of known antimyco-bacterial drugs(4). The degree of fluorescence of M. tuberculosis during the growth phase appears to correlate well with the increase of cell growth in culture. The GFPMA MICs result could be obtained within 7 days. A good correlation of MIC results was found between BACTEC 460 and GFPMA method. With its non-invasive procedure that precludes substrate addition, GFP can be utilized as a reporter for rapid in vitro screening for anti-TB compounds, which is labor-efficient and cost-effective. However, there are unique problems associated with the detection of fluorescence signal, which have compromised many potential fluores-cence-based HTS assays in the past. The problems are: 1) the signal can be subject to quenching by compounds, plastics, and media; 2) fluorescence emission can be scattered by particulates; and 3) the signal can be masked by autofluorescence (from proteins or compounds).
Screening for antituberculosis compounds using GFP driven by a heat shock promoter (Phsp60) has been of limited utility due to the low signal to noise ratio. Recently, we demonstrated the evaluation of a newly identified promoter from M. smegmatis MC2155 with acetamide-independent high-level constitu-tive promoter activity to enhance the sensitivity of GFP detection during microplate-based culture as well as the assessment for response to 18 established antimicrobial agents using GFPMA(2). A GFP reporter plasmid that contains a gfp gene driven by an unusual acetamidase promoter (Pace) of M. smegmatis was constructed and transformed into M. tuberculosis H37Ra and M. smegmatis MC2155. By using flow cytometric analysis, GFP-mediated fluorescence of recombinant strains of mycobacteria with the Pace::gfp construct was

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
143
8-fold higher than the strain bearing Phsp60::gfp construct.(2)
This remarkable acetamidase promoter can direct GFP protein production in mycobacteria of up to 8-20% of total protein, while the hsp60 promoter achieved only 3% total protein. In addition, fluorescence curves of M. tuberculosis strains H37Ra, H37Rv, and Erdman harboring the Pace::gfp construct correlated well with growth curves of the corresponding strains
(shown in Figure 1) and the kinetic response to twofold serial dilutions of antimycobacterial agents was easily detected by fluorescence reading of the microplate cultures after 1 week (shown in Figure 2 for M. tuberculosis H37Rv Pace::gfp against isoniazid or rifampin). MICs of known antimycobacterial drugs determined by GFPMA against laboratory M. tuberculosis gfp strains are in excellent agreement with those determined by the reference microplate Alamar blue assay(2).
Incubation tim e (days)0 1 2 3 4 5 6 7 8
Rel
ativ
e Fl
uore
scen
ce U
nit (
RFU
)
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
LOG
cfu
per
ml
5 .0
6.0
7.0
8.0 B
Figure 1 GFP fluorescence (bar) and growth curve (line) of M. tuberculosis H37Rv strain harboring either
Pace::gfp (green) or Phsp60::gfp (red) construct. H37Rv wild type strain containing no vector was included (black). Cells were grown in 7H12 minimal broth at 37oC.
0 1 2 3 4 5 6 7 80
250
500
750
1000
1250
1500
1750
2000
2250
2500
2750
3000
3250
3500
0.75 μg/ml
0.375 μg/ml
0.188 μg/ml0.094 μg/ml
0.047 μg/ml
0.023 μg/ml0.012 μg/ml
0.006 ug/mlDrug-free control cut-off line
Isoniazid
I b i
Rel
ativ
e Fl
uore
scen
ce U
nit (
RFU
)
0 1 2 3 4 5 6 7 80
250
500
750
1000
1250
1500
1750
2000
2250
2500
2750
3000
3250
3500
0.375 μg/ml
0.188 μg/ml
0.094 μg/ml0.047 μg/ml0.023 μg/ml
0.012 μg/ml0.006 μg/ml0.003 ug/mlDrug-free control cut-off line
Rifampin
Figure 2 Isoniazid and rifampin dose-response curves for M. tuberculosis H37Rv gfp determined by fluorescence.
Incubation time (day)
Re
lati
ve F
luo
resc
en
ce
Un
it (
RF
U)
Incubation time (day)
Re
lati
ve F
luo
resc
en
ce
Un
it (
RF
U)
LO
G c
fu p
er
ml

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
144
Conclusions
The recently developed GFPMA using Pace::gfp construct represents the most sensitive fluorescence-based assay available for anti-tuberculosis drug screening in vitro. However, screening for new antimicrobial agents is rou-tinely conducted only against actively repli-cating bacteria. In addition, it is now widely recognized that a physiological state of non-replicating persistence is responsible for anti-microbial tolerance in many bacterial infections. In tuberculosis, the key to shortening the 6-month regimen is based on targeting this dormant sub-population. Preliminary study of the application of gfp strain in drug screening under low oxygen condition resembling non-replicating dormancy state of M. tuberculosis showed that fluorescence signals are highly detectable in such low oxygen condition where bacteria were expected to become non-replicating and dormant, indicating the unusual stability of GFP that accumulates inside the cells. Such limitation in utilization of stable gfp strains as described previously demands future development of unstable or de-stabilized gfp strains of M. tuberculosis in order to expand the application of gfp stains for detection of viability of dormant or non-replicating cells. Recently, a high-throughput, luminescence-based low-oxygen recovery assay (LORA) has been developed to screen antimicrobial agents against non-replicating persistence (NRP) mycobac-teria(3). M. tuberculosis H37Rv containing a plasmid with the acetamidase promoter driving a bacterial luciferase gene was adapted to low oxygen conditions and was then exposed to drugs under anaerobic conditions followed by an oxygen recovery phase. Luminescent signals were found to be proportional to the numbers of colony forming units determined prior to the recovery phase. Thus, the LORA method is sufficiently robust for use for primary high-throughput luciferase-based screening of com-pounds against non-replicating persistent M. tuberculosis. The acetamidase-driven luxAB construct also showed promising application for TB research in macrophage and mice infection models.
Acknowledgements
The author thanks Dr. Prasit Palittapon- garnpim for helpful discussion and critical reading of the manuscript.
References 1. Castro-Garza J, King CH, Swords WE, Quinn
FD. Demonstration of spread by Mycobacterium tuberculosis bacilli in A549 epithelial cell mono-layers. FEMS Microbiol Lett 2002;212:145-9.
2. Changsen C, Franzblau SG, Palittapongarnpim P. Improved green fluorescent protein reporter gene-based microplate screening for anti-tuberculosis compounds by utilizing an aceta-midase promoter. Antimicrob Agents Chemother 2003;47:3682-7.
3. Cho SH, Warit S, Wan B, Hwang CH, Pauli GF, Franzblau SG. Low-oxygen-recovery assay for high-throughput screening of compounds against nonreplicating Mycobacterium tuberculosis. Antimicrob Agents Chemother 2007;51:1380-5.
4. Collins LA, Torrero MN, Franzblau SG. Green fluorescent protein reporter microplate assay for high-throughput screening of compounds against Mycobacterium tuberculosis. Antimicrob Agents Chemother 1998;42:344-7.
5. Cowley SC, Av-Gay Y. Monitoring promoter activity and protein localization in Mycobac-terium spp. using green fluorescent protein. Gene 2001;264:225-31.
6. Croston GE. Functional cell-based uHTS in chemical genomic drug discovery. Trends Bio-technol 2002;20:110-5.
7. Dusthackeer A, Kumar V, Subbian S, Sivara-makrishnan G, Zhu G, Subramanyam B et al. Construction and evaluation of luciferase reporter phages for the detection of active and non-replicating tubercle bacilli. J Microbiol Methods 2008;73:18-25.
8. Ewann F, Jackson M, Pethe K, Cooper A, Mielcarek N, Ensergueix D et al. Transient requirement of the PrrA-PrrB two-component system for early intracellular multiplication of Mycobacterium tuberculosis. Infect Immun 2002;70:2256-63.
9. Gangopadhyay JP, Jiang SQ, Paulus H. An in vitro screening system for protein splicing inhibitors based on green fluorescent protein as an indicator. Anal Chem 2003;75:2456-62.
10. Gupta N, Singh BN. Deciphering kas operon locus in Mycobacterium aurum and genesis of a recombinant strain for rational-based drug screening. J Appl Microbiol 2008;105:1703-10.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
145
11. Gurskaya NG, Fradkov AF, Terskikh A, Matz MV, Labas YA, Martynov VI et al. GFP-like chromoproteins as a source of far-red fluorescent proteins. FEBS Lett 2001;507:16-20.
12. Harris NB, Zinniel DK, Hsieh MK, Cirillo JD, Barletta RG. Cell sorting of formalin-treated pathogenic Mycobacterium paratuberculosis expressing GFP. Biotechniques 2002;32:522-4,6-7.
13. Hazbon MH, Guarin N, Ferro BE, Rodriguez AL, Labrada LA, Tovar R et al. Photographic and luminometric detection of luciferase reporter phages for drug susceptibility testing of clinical Mycobacterium tuberculosis isolates. J Clin Microbiol 2003;41:4865-9.
14. Hunt L, Batard P, Jordan M, Wurm FM. Fluorescent proteins in animal cells for process development: optimization of sodium butyrate treatment as an example. Biotechnol Bioeng 2002;77:528-37.
15. Karsi A, Howe K, Kirkpatrick TB, Wills R, Bailey RH, Lawrence ML. Development of bioluminescent Salmonella strains for use in food safety. BMC Microbiol 2008;8:10.
16. Langdon G, Wilkins JJ, Smith PJ, McIlleron H. Consecutive-dose pharmacokinetics of rifapen- tine in patients diagnosed with pulmonary tuber- culosis. Int J Tuberc Lung Dis 2004;8:862-7.
17. Laszlo A, Rahman M, Espinal M, Raviglione M. Quality assurance programme for drug suscep-tibility testing of Mycobacterium tuberculosis in the WHO/IUATLD Supranational Reference Laboratory Network: five rounds of proficiency testing, 1994-1998. Int J Tuberc Lung Dis 2002; 6:748-56.
18. Luna-Herrera J, Martinez-Cabrera G, Parra-Maldonado R, Enciso-Moreno JA, Torres-Lopez J, Quesada-Pascual F et al. Use of receiver operating characteristic curves to assess the performance of a microdilution assay for determination of drug susceptibility of clinical isolates of Mycobacterium tuberculosis. Eur J Clin Microbiol Infect Dis 2003;22:21-7.
19. Matz MV, Lukyanov KA, Lukyanov SA. Family of the green fluorescent protein: journey to the end of the rainbow. Bioessays 2002;24:953-9.
20. Okuno T, Goto Y, Matsumoto Y, Otsuka H. Applications of recombinant Leishmania amazonensis expressing egfp or the beta-galactosidase gene for drug screening and histopathological analysis. Exp Anim 2003;52: 109-18.
21. Parrish NM, Houston T, Jones PB, Townsend C, Dick JD. In vitro activity of a novel anti-mycobacterial compound, N-octanesulfonylacet-
amide, and its effects on lipid and mycolic acid synthesis. Antimicrob Agents Chemother 2001; 45:1143-50.
22. Quinn A, Jiang W, Velaz-Faircloth M, Cobb AJ, Henry SC, Frothingham R. In vivo protein expression and immune responses generated by DNA vaccines expressing mycobacterial antigens fused with a reporter protein. Vaccine 2002;20: 3187-92.
23. Rosenkrands I, Slayden RA, Crawford J, Aagaard C, Barry CE, 3rd, Andersen P. Hypoxic response of Mycobacterium tuberculosis studied by metabolic labeling and proteome analysis of cellular and extracellular proteins. J Bacteriol 2002;184: 3485-91.
24. Russo A, Filippi C, Tombolini R, Toffanin A, Bedini S, Agnolucci M et al. Interaction between gfp-tagged Pseudomonas tolaasii P12 and Pleurotus eryngii. Microbiol Res 2003;158:265-70.
25. Sood R, Rao M, Singhal S, Rattan A. Activity of RBx 7644 and RBx 8700, new investigational oxazolidinones, against Mycobacterium tubercu-losis infected murine macrophages. Int J Anti-microb Agents 2005;25:464-8.
26. Sorensen SJ, Sorensen AH, Hansen LH, Oregaard G, Veal D. Direct detection and quantification of horizontal gene transfer by using flow cytometry and gfp as a reporter gene. Curr Microbiol 2003 ;47:129-33.
27. Sung ND, Lee CY. Coregulation of lux genes and riboflavin genes in bioluminescent bacteria of Photobacterium phosphoreum. J Microbiol 2004; 42:194-9.
28. Turner DJ, Hoyle SL, Snewin VA, Gares MP, Brown IN, Young DB. An ex vivo culture model for screening drug activity against in vivo phenotypes of Mycobacterium tuberculosis. Microbiology 2002;148:2929-36.
29. Wang Y, Yu YA, Shabahang S, Wang G, Szalay AA. Renilla luciferase-Aequorea GFP (Ruc-GFP) fusion protein, a novel dual reporter for real-time imaging of gene expression in cell cultures and in live animals. Mol Genet Genomics 2002;268:160-8.
30. Wattiau P, Springael D, Agathos SN, Wuertz S. Use of the pAL5000 replicon in PAH-degrading mycobacteria: application for strain labeling and promoter probing. Appl Microbiol Biotechnol 2002;59:700-5.
31. Zafer AA, Taylor YE, Sattar SA. Rapid screening method for Mycobactericidal activity of chemical germicides that uses Mycobacterium terrae expressing a green fluorescent protein gene. Appl Environ Microbiol 2001;67:1239-45.


147
Oligomer Extrapolation Techniques: An Economic Approach to the Study of Relationships between Polymeric Structures and Band Gaps
Withoon Chunwachirasiri Department of Physics, Faculty of Science, Mahidol University.
Abstract
A calculation technique for the band gaps in polymeric systems using the oligomer extrapolation techniques is presented as a viable approach for studies of structure/property relationships, and specifically the relationships between polymer main chain conformations and energy gaps, in solid-state polymers. This technique is computationally inexpensive, provides flexibility in altering the main chain conformations, and gives accurate results as compared with other techniques with higher levels of sophistication. The calculations have been completed using a personal computer with modest specifications within a reasonable amount of computing time.
Keywords: oligomers; band gap; density functional theory, DFT
Introduction
Understanding the relationship between molecular structures and electronic properties has always been a focus of material studies and plays a crucial role in material design and fabri-cation. This is especially true for polymers due to their unique characteristics of combined physical and chemical properties, including flexibility, chemical processibility and inherited physical properties of semiconducting materials. In general, polymers are composed of a number of atoms and have complex molecular struc-tures. These complexities are further enhanced in the case of polymers in a solid phase due to the effects of molecular packing. As a result, a good understanding of structure/property rela- tionships serves as guidance as to which new materials one should be constructing.
One out of many physical properties that is fundamental to the new material fabrication is the electronic band gap. The band gaps are found in many solid-state crystals and polymeric systems that exhibit structural periodicity. In molecular orbital terminology, the band gap separates the highest occupied molecular orbital (HOMO) and lowest unoccupied molecular orbital (LUMO). Electrons in a LUMO can make transitions to
the higher energy levels when the excitation energy is equal to or greater than the “energy gap,” which is the energy difference between the HOMO and LUMO levels. The excitations are commonly in the form of photo-excitation. In contrast to the absorption process, if the excited electrons in the HOMO level or above are to relax back to lower states, the transition will occur across the band gap and we have an emission process. There will be photons asso-ciated with emission processes. The energy of the emitting photons, once again, depends on the size of the energy gap. Consequently, absorbance and emission profiles obtained experimentally from photo-absorption and photo-emission measurements are direct observations of the magnitude of the energy gap.
The importance of controlling the band gap is apparent, especially in optical related applications such as for polymer light emitting diodes, solar cells and photocells. Each application requires photons to be absorbed or emitted at some particular wavelengths. Some may require the absorbing/emitting region to be very narrow, while some may require broad spectra. The way to manipulate the band gap is through variations of the atomic construction of the material. Different atomic constructions

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
148
will have different distributions and arrange-ments of electrons among constituent atoms which ultimately affect the band gap of the system as a whole. Because there is a countless number of ways how a material can be constructed, a trial and error approach is very inefficient and wasteful. More systematic approaches are preferred. This is how molecular modeling comes into the picture.
There are many levels of sophistication in molecular modeling, ranging from purely theoretical (high level of sophistication) to purely experimental (low level of sophistica- tion). Each level has its own advantages and disadvantages. One key comparison between different levels of sophistication is the computa-tional cost. The costs include hardware, software, implementation and computing time. As a general rule, a higher the level of sophistication involves higher computational costs. Thus most popular models at a time when computers are expensive are ones intermediate between the two, i.e. semi-empirical models. However, even today when the computing technology has improved tremendously from decades ago, the calcu-lations with a high level sophistication are still not easily carried out. This is because the materials of interest have also grown in numbers and complexities. As a result, the optimization between the computational cost and accuracy is still an ongoing research topic.
In many circumstances, computing re-sources are a rare commodity. Not every researcher can gain access to high-end computing facilities. However, PC computers are ubiquitous nowadays and these PCs are powerful enough to perform moderately complex calculations within a respectable amount of time. This suggests that if some questions can be answered using an approach that a single PC can handle, then this could help reducing some reliance on high-end computing facilities and give the available PCs effective uses.
Motivation and Problem
The main problem to be answered here is the relationship between the conformations of
the polymer main chain and the associated energy band gaps. The polymers to be considered here are conjugated polymers. Electrons in conjugated polymer chains are considered to be able to move freely along the chains. This is referred to as “electron delocalization”. Because of the electron delocalization, this class of polymer has an ability to conduct electricity. The extent of electron delocalization is called the “conju-gation length.” It is well-known that the energy gap increases with an increase in the main chain torsion angle. This is because a high torsion angle decreases the electronic coupling between adjacent molecular orbitals, which leads to shorter conjugation lengths. As a consequence, the associated energy gaps increase.
The sensitivity to which the torsion angle affects the energy gap depends on the molecular construction of the individual conjugated polymer. If a polymer chain under consi-deration is a single molecule in the gas phase, and the temperature is absolute zero, the optimized structure will be the one that has the lowest energy. This is usually a tran-planar conformation (the torsion angle of 180°) in the case of unbranched polymers. However, the calculations for these conditions do not reflect real world situations where the polymers are commonly cast as solid films at room temperature. Under solid-state conditions the steric repulsions from chain-to-chain packing are very likely to force the polymer main chain into conformations that deviate from the lowest energy conformation. As a result, the bulk solid polymers could end up with mixtures of various main chain conformations. Because the energy gap depends on the main chain conformation, this will affect the photoabsorption and photo-emission behaviors of the solid polymers. If the conformation/energy gap relationship shows a strong dependence, this would suggest that control of the polymer conformation is crucial to the final behavior of the material to be fabri-cated. On the other hand, if the dependence is weak, this leads to a material that is con-formation-insensitive and could make the material fabrication less structurally demanding. Being able to determine the relationship

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
149
between the polymer main chain conformation and their corresponding energy gap will provide a deeper insight to the structure/property relationships along with technological implica- tions.
A standard way to calculate band gaps of polymeric materials is by finding their band structures. There are many molecular modeling packages readily available to perform the calculations. However, in this particular study of chain conformations and energy gaps there are two main reasons why the conventional approaches are not ideal.
First, to obtain a band structure of a polymeric system, periodic boundary conditions must be imposed on a unit cell, the material’s primitive building block. Both ends of the unit cell must match exactly for the polymeric system to be constructed. This makes the study of various main chain conformations somewhat difficult since at some torsion angles the unit cell has to be made very large to accommodate the boundary condition requirements. The large unit cell lengthens the computing time required to complete the calculations.
Second, because the goal is to be able to determine the energy gaps of conformations quickly and cost effectively, the calculation of the band structures just to gain information on the energy gap can be considered excessive. To solve the current problem, an energy gap calculation technique that provides great accuracy and does not demand high computing resources is needed.
Computational Approach
A computational approach that fits current demands follows Kertesz’s scheme,(10) for which great success was the prediction of band gaps of conjugated polymers based on extrapolating the calculated band gaps of related oligomers. The process starts with geometry optimization followed by the energy level calculation of a short-chain oligomer. The calculations are then repeated on subsequently longer-chain oligomers until the asymptotic behavior is observable; in their report the behavior was a linear relationship between the
band gap and the reciprocal of the oligomer chain length. The choice of the theoretical level and basis set used in the geometry optimization and energy level calculation is crucial. The combinations that provide accurate results in comparison with the experimental data are B3LYP/PM3 and B3LYP/B3LYP, where x/y refers to geometry optimization at the “y” theoretical level followed by energy level calculations at the “x” level. B3LYP represents the hybrid density functional theory (DFT) method that includes Becke’s three parameter hybrid functional(1) with the Lee-Yang-Parr(5) correlation functional. PM3 represents para-metric method 3(9) which is a parameterized self-consistent field, the Hartree-Fock method. The basis set used in the geometry optimization step is 6-31G(d) and the basis set in the energy level calculation step is 6-311G(d,p). The later basis set is expected to enhance the quantitative accuracy of the computed energy gaps. The energy gap of each individual oligomer is extracted directly from the energy difference between the HOMO and LUMO energy levels.
Trial calculations using a less sophis-ticated level of theory, specifically the HF method, have been performed. Although the calculation times were considerably lower, all of the calculated band gaps were substantially overestimated. This is to be expected because of the lack of electron correlations in the HF method. On the other hand, the PM3 method is considered expensive from a computational cost perspective. At this theoretical level, the geometry optimization of a moderate-size oligomer takes longer than a week to converge on the available PC. As a consequence, the method used here as a compromise between computational cost and accuracy was chosen to be B3LYP/B3LYP. At this theoretical level the geometry optimization of the oligomer with 19 primitive units was completed in less than 2 days.
All calculations described in this work were performed using the Gaussian 03 package(3). The computer used in trials is a PC with a Dual Core Processor running at 2.8 GHz with 2 GB of accessible RAM.

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
150
To explore the applicability of the stated approach, a newly discovered class of materials, polytitasilanes, was chosen for study. In a primitive configuration, polytitasilane is com-posed of silicon (Si) and titanium (Ti) along the polymer main chain bonded solely to hydrogen atoms without branching. Polytitasilanes have emerged recently among significant efforts toward the design and synthesis of polymers with a narrow band gap (a few eV) to achieve the fabrication of light emitting diodes operating in the IR region. The study reports for polytitasilanes are still scarce. However, the band gap calculated by DFT has been reported by Feng and coworkers(2) to range from 2.7 eV to 3.4 eV depending on the Si-Ti sequence and unit cell used in the calculations. Although the synthesis of polytitasilane has not been realized, TiH4 has been experimentally identified(4) while the pure silicon polymers, polysilanes, have been studied extensively for more than two decades(6-8). So this is one of many classes of polymer that is worth investigating.
The atomic sequencing of oligotitasilane that gives the lowest energy gap is the one with alternating Si and Ti atoms throughout the polymer chain, terminated with an extra Si atom. Thus the oligomers, in this communi-cation, are Sin+1TinH4(n+1), where n is the number of Ti atoms. The oligotitasilanes having n = 1 to n = 9 were used as examples. As a prelimi-nary calculation, the main chain conformations in all of the oligotitasilanes are constrained to the trans-planar configuration.
Extrapolation of Band Gaps
The obtained energy gaps E, in eV are shown in Table 1. As aforementioned, one of the well-known characteristics of conjugated polymers is the reduction of the band gap in longer polymer chain.
The data in Table 1 clearly show this characteristic. Furthermore, the calculated band gaps exhibit asymptotic behavior. This behavior can be observed more succinctly by plotting the calculated band gap as a function of 1/N. The plot is shown in Figure 1. The downward trend toward smaller 1/N, i.e. longer chain lengths,
reaches a linear regime at approximately N = 7. Following the analysis suggested by Kertesz, this linear trend can be expressed by a linear equation:
( ) ,gaE N bN
= +
where the value of a and b are determined by linear regression. The value of b or the y-intercept represents the prediction of the polymer band gap where N is very large (1/N approaches zero). Therefore b is the predicted band gap of polytitasilane. The value of a is a scaling factor specific to each individual class of polymers. Different classes of polymers are expected to have different values of a when computed by the oligomer extrapolation technique.
Table 1 Calculated HOMO-LUMO energy gaps (Eg) of oligotitasilanes of varying length. n is the number of Ti atoms. N is the total number of Ti and Si atoms.
n N = 2n+1 Eg (eV) 1 3 3.876981 2 5 3.242699
3 7 3.043789
4 9 2.971136
5 11 2.935762
6 13 2.912633
7 15 2.879708
8 17 2.871817
9 19 2.861477
The last two data points (indicated by triangles) in Figure 1 were excluded from the linear regression analysis to ensure that only data within the linear regime are being used in the regression. From the included points (indicated by circles), the values of a and b are 1.9339 and 2.7581, respectively. This means that the estimated band gap of polytitasilane is 2.76 eV. The results agree reasonably well with the reported value of 2.73 eV using the periodic boundary condition DFT method. The asso- ciated coefficient of determination (R2) of

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
151
98.93% implies a high goodness of fit and provides strong confidence in the extrapolation.
Figure 1 Plot between calculated HOMO-LUMO
energy gaps (Eg) in eV and 1/N, where N is the total number of constituent atoms in oligotitasilanes.
The value of “a” obtained here was found to be much smaller than those for other π-conjugated polymers, including poly (p-phy-lene) polythiophene, poly (p-phenyleneviny-lene), polyacetylene, polypyrrole, and poly (thienylvinylene), as previously reported. The values of a for those π-conjugated polymers ranged from 17.2 to 18.4 while the value of a for polytitasilane is 1.9. The smaller value of a is, however, expected since the variation in energy gaps among oligomers having a very narrow band gap is limited within a narrower range than those of materials with large band gaps.
The computing time to complete the calculations to obtain the energy gaps from oligomers ranges from a couple of hours for small n to approximately 2 days for the longest oligomers used in the current example. The complete extrapolation as shown in Figure 1 was completed within 4 days. The total calcu-lation time could be reduced further by exploiting the fact that the progression of data points reaches the linear regime very quickly. Therefore, the computation of very large chains can be excluded in actual application of the technique. This could result in the reduction of the computing time by approximately 20%. Additionally, the PC used in the present examples is equipped with AMD Athlon X2
5600 (2.8 GHz) CPU with 2 GB of memory which can be classified as a very modest speed and specifications by today standard.
By adopting a PC with a faster CPU speed and better hardware configurations that are available today, the total computation time could be reduced even further.
Conclusions
The calculation of polymer energy gaps by the oligomer extrapolation technique was presented and suggested as a way to facilitate the study of the relationships between polymer main chain conformations and energy gaps. The technique offers many advantages over conven-tional approaches. First, this technique performs the calculation on oligomers so there are no requirements on periodic boundary conditions to be met. Second, the oligomers are small enough for any modest PC to handle the computation. Thus this technique can be simply applied using already available PCs. When applied to polyti-tasilane, a newly discovered class of narrow band gap materials, the results agree well with those from the calculations using higher levels of sophistication. The benefits from the low computational cost, the flexibility in altering molecular conformations and accessibility of the computing machines make this technique a very attractive alternative for the study of the rela-tionships between polymer chain conformations and their energy gaps.
Acknowledgements
This work was supported by Mahidol University under the Young Scientist Research Fund. The author gratefully acknowledges the Physics Department, Faculty of Science, Mahidol University for continuing support.
References 1. Becke AD. Density-functional thermochemistry.
III. The role of exact exchange. J Chem Phys 1993;98:5648-52.
2. Feng D, Ding Y, Feng S, Zhang J, Xie J. The DFT study on the electronic structures of poly-titasilanes: several new classes of narrow band gap polymers. Polymer 2006;47;3681-8.
y = 1.9339x + 2.7581R2 = 0.9893
2.52.7
2.93.1
3.3
3.53.7
3.9
0 0.1 0.2 0.3
E'g
(eV
)
1 / N
N = 7

วทยาศาสตรเพอการพฒนา : รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
152
3. Frisch MJ, Trucks GW, Schlegel HB, Scuseria G E, Robb MA, Cheeseman JR et al. Pittsburgh: Gaussian Inc; 2003.
4. Hood DM, Ptizer RM, Schaefer III HF. Homo-nuclear 3d transition-metal diatomics: a sys-tematic density functional theory study. J Chem Phys 1979;71:705.
5. Lee C, Yang W, Parr, RG. Development of the Colle-Salvetti correlation-energy formula into a functional of the electron density. Phys Rev B 1988:785-9.
6. Michl J, West R. Silicon-containing polymers: The science and technology of their synthesis and applications. New York: Springer 2001. p. 499-530.
7. Miller RD, Michl J. Polysilane high polymers. J Chem Rev 1989;89,1359-1410.
8. Sakurai Y, Yoshimura D, Ishii H, Ouchi Y, Isaka H, Teramae H et al. Electronic structure of poly-carbosilane studied by UV photo-electron spectroscopy. J Phys Chem B 2001; 105:5626-9.
9. Stewart JJP. Optimization of parameters for semiempirical methods. I. Method. J Comp Chem 1989;10:209-20.
10. Yang S, Olishevski P, Kertesz M. Bandgap calculations for conjugated polymers. Synthetic Met 2004;141:171-7.

153
เนอในเมลดมะมวง ของดทถกมองขาม
ปราณ อนประโคน ภาควชาเทคโนโลยชวภาพ คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
ในแตละป การบรโภคและการแปรรปมะมวงในประเทศไทย กอใหเกดขยะเนอในเมลดมะมวงทถกทงและทาใหเกดปญหาแกสภาพแวดลอมในปรมาณทสงถง 57,200 ตน ขยะเมลดมะมวงเหลานเปนของดทถกมองขามจากสงคมไทยอยางนาเสยดาย ผลงานวจยททนสมยในปจจบนไดชใหเหนถงประโยชนของเมลดมะมวง ซงเปนแหลงอาหารทใหคารโบไฮเดรต ไขมน และโปรตนทมคณคาทางโภชนาการสง และสามารถนามาใชเปนวตถดบในการผลตไขมนทดแทนเนยโกโกและเนยขาวปราศจากไขมนทรานสซงมมลคาสงและไมสามารถผลตไดในประเทศเขตรอนชน นอกจากน เนอในเมลดมะมวงยงอดมไปดวยสารตานอนมลเสรทมฤทธสง การรบประทานเนอในเมลดมะมวงเปนอาหารเสรมจะชวยเสรมสรางภมคมกน และปองกนรางกายจากพยาธสภาพตางๆ ทเกดจากอนมลเสร เชน โรคความจาเสอม โรคหวใจ และมะเรง เนอในเมลดมะมวงมคณสมบตทางเครองสาอางเพราะชวยใหความนมชมชนและชะลอการแกของเซลลผวหนงและสามารถใชเปนอาหารเลยงสตวทมราคาถกไดอกดวย
คาสาคญ: เนอในเมลดมะมวง; สารตานอนมลเสร; สารกนเสยชวภาพ; เครองสาอาง
Abstract
In Thailand, at least 57,200 tonnes of mango kernels are discarded annually as waste from consumption and industrial processing of mango and thus have become a source of environmental pollution. Unfortunately, high value-added use of this mango seed waste has not realized. Recent research has indicated the usefulness of mango seed as a potential food source which contains high nutritional carbohydrate, fat and protein. The fat portion can be used as a cocoa-butter substitute as well as raw material for the production of trans-fatty acid-free shortenings which are not available in tropical countries. Moreover, mango seed kernel is a rich source of potent antioxidants. Food supplementation with mango kernel extract, therefore, helps to increase our immunity as well as preventing free radical induced-diseases such as loss of memory, atherosclerosis and cancer. Due to its emollient and anti-ageing properties, mango seed kernel is suitable for cosmetics. In addition, mango seed kernel can also be utilized as alternative source of animal feed stuff.
Keywords: mango seed kernel; antioxidants; biopreservative; cosmetics
บทนา
“มะมวง” นอกจากเปนผลไมทคนไทยรจกกน
แพรหลายแลว ยงเปนทรจกเปนอยางดสาหรบนกวจย ทศกษาเกยวกบสารตานอนมลเสรวา มคณคาสงมาก ทกสวนของมะมวง ไมวาจะเปนใบ เปลอกของตน ดอก ผล และเมลด มสารกลมแทนนน (tannin) ฟลาโวนอยด (flavonoids) พอลฟนอล (polyphenols) และฟนอลก (phenolics) (รปท 1) ซงมฤทธตานอนมลเสรในปรมาณทสงมาก(26-27) มะมวงเปนผลไมทอยคกบคนไทยมาชานาน
และเปนผลไมทมผนยมบรโภคมากทสดชนดหนงใน
ประเทศ มพนธใหเลอกรบประทานหลากหลาย ทงแบบผลดบ เชน เขยวเสวย แรด หนองแซง พมเสนมน และรบประทานผลสก เชน อกรอง นาดอกไม หนงกลางวน หรอในรปของมะมวงแปรรป เชน นามะมวง มะมวงกวน มะมวงแชอม มะมวงดอง โดยเฉพาะมะมวงพนธทนยมนามาดองนน คอ แกว สามป และตลบนาค มะมวงนบเปนพชเศรษฐกจทสาคญอกชนดหนงทประเทศไทยมปรมาณ
การผลตมากเปนอนดบท 4 ของโลกรองจากอนเดย จนและเมกซโก โดยมปรมาณการผลตประมาณ 572,000

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
154
รปท 1 สารตานอนมลเสร ประเภทพอลฟนอล และ สารฟนอลก ทพบในเนอในเมลดมะมวง
เมตรกตนตอป มะมวงทวโลกมอยประมาณ 793 พนธ โดยชนด (species) ทพบในประเทศไทยคอ มะมวงกะลอนหรอมะมวงปา (Mangifera duperrana Chochinchina), M. indica, M. caloneura, M. siamensis, M. oblongifolia Malaca, M. camptosperma และ M. lagenifera การบรโภคและการแปรรปมะมวงในระดบอตสาหกรรมไดกอใหเกด
ปญหาทางสภาพแวดลอมทสงมาก ทงนเปนผลมาจากการใชเฉพาะสวนเนอมะมวงเทานน สวนอนๆ ของผลมะมวง จะถกทงเปนขยะสงถง 40-60% โดยไมมการนามาใช ประโยชน ทงนแบงเปนเปลอก 12-15% เศษเนอมะมวง 5-10% และเปนเมลดมะมวง 15-20% ในประเทศอนเดย
ประเทศเดยวมเมลดมะมวงทถกทงจากการบรโภคและ
อตสาหกรรมการแปรรปมะมวงสงถง 300,000 ตน/ป เพอลดปญหาของขยะเมลดมะมวงจานวนมหาศาลน จงมการวจยเพอนาเอาเมลดมะมวงมาใชประโยชนทงเปนอาหาร อาหารเสรม เครองสาอาง ยา และอาหารสตว(17) ในขณะทประเทศไทยซงมขยะเมลดมะมวงในปรมาณสงถง 95,000 ตน/ป โดยคดเปนเนอในเมลดมะมวงทมคณคาทถกทงสง
ถง 57,200 ตน/ปนน ยงไมมการทาวจยหรอการใหขอมลถงคณคาของเมลดมะมวงอนจะเปนการกระตนใหวงการ
วจยเกดความสนใจในการนาเอาขยะเมลดมะมวงกลบมา
ใชประโยชนแตอยางใด ขยะเมลดมะมวงจงเปนของดทถกมองขามอยางนาเสยดายและกลบถกทงใหกลายเปนปญหา
ตอสภาพแวดลอม จงสมควรทหนวยงานทเกยวของจะตองใหความรทเกยวกบคณคาและประโยชนของเมลดมะมวง
เพอกระตนใหนกวจยหนมาสนใจศกษาอยางจรงจงและ
เพอใหคนไทยหนมาใหความสนใจตอของดใกลตวและ
นามาใชใหเกดประโยชนอยางสงสดตอไป องคประกอบของเนอในเมลดมะมวง
การวจยในปจจบนยงคงจากดอยทเนอในเมลด
ของมะมวงสายพนธ M. indica จากประเทศอนเดย อยปต และไนจเรยเทานน สวนมะมวงสายพนธอนๆ ยงไมมรายงานการศกษาวจย ทงน ผลการวจยใหความรว า องคประกอบของเนอในเมลดมะมวงมความแตกตางกน
เลกนอย ขนอยกบแหลงภมศาสตรทเพาะปลกและสภาพ
OH
HO O
OH
OH
OH
HO O
OH
OH
OH
OH
OH
HO O
OH
OH
OH
n
Condense Tannin
Hydrolyzable Tannin
OH
HO
Gallic acidOH
HO
HO
O
OH
O
OO
O
OH
OH
Ellagic acid
HO
OH
O
OH
OH
OH
Epicatechin
HO
OH
O
OH
OH
OH
Gallocatechin
OH

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
155
ภมอากาศ เมอกะเทาะเปลอกเมลดออกจะเหนเนอในสขาว เนอในของเมลดมะมวงสดนนมความชนประมาณ 51% มรสขม เมอนาไปละลายนาประมาณ 50 เทาจะมความเปนกรดเลกนอย (pH 5) และเปลยนเปนสนาตาลอยางรวดเรวเมอสมผสอากาศ แสดงถงปฏกรยาออกซเดชนของสาร ฟนอลกดวยเอนไซมพอลฟนอลออกซเดส (polyphenol oxidase)(4) เมอทาใหแหงดวยวธตางๆ เชน ตากแดด หรอ อบดวยความรอน จะเหลอความชนประมาณ 9-12% นอกจากน เนอในเมลดมะมวงแหงยงประกอบดวยโปรตน 5-6% ไขมน 9-16% คารโบไฮเดรต 73-80% เสนใย 2-3% เถา 2-3% และแทนนน 1%(7,20,29)
การประยกตใชในผลตภณฑอาหาร
เนอในเมลดมะมวงมคณคาทางโภชนาการสง
เพราะอดมไปดวยโปรตน ไขมน แปง และเสนใย และม
ความปลอดภยสงหากใชบรโภค ชนพนเมองและคนยากจนของอนเดยและทวปแอฟรกาบรโภคเนอในเมลด
มะมวงโดยนามาควหรอตม เพอทดแทนอาหารจาพวกแปงมาแตสมยโบราณ และอาหารอนเดย เชน จาปาตและสพาร ในบางทองถนมแปงเนอในเมลดมะมวงเปนสวนผสม
สาคญ แปงทไดจากเนอในเมลดมะมวงเปนแปงจากผลไมทมปรมาณของแปงซงไมถกยอย (resistance starch) ใน
ปรมาณทสงมาก (75-80%) ดงนนหลงจากการรบประทานแปงชนดนจงทาใหคาดชนนาตาล (glycemic index) ตา(24)
โดยองคประกอบของแปงนมปรมาณแอมโลส (amylose) 40% และปรมาณแอมโลเพกทน (amylopectin) 61% คณสมบตของเมดแปง คณสมบตในการละลาย ความหนด และการขนรปคลายคลงกบแปงจากธญพชทนยมบรโภค
กนทวไป เชน แปงมนฝรง แปงขาวเจา และแปงสาล จงสามารถนาแปงเมลดมะมวงมาใชทดแทนหรอผสมกบแปง
เหลานได(15) โดยเฉพาะแปงสาลซงเปนสวนประกอบของขนมหลายประเภทนน ตองพงพาการนาเขาและมราคาแพง
การทดแทนแปงสาลดวยแปงทมราคาถกกวา เชนแปงเมลดมะมวงนบวานาสนใจอยางยงเพราะแปงเมลด
มะมวงมความสามารถในการดดนาสง จงเหมาะทจะ
นามาใชในการทาโดและขนมอบ จากการศกษาผลกระทบตอคณภาพของขนมปงกรอบเมอทดแทนแปงสาลดวยแปง
เมลดมะมวงบางสวน พบวาแปงเมลดมะมวงสามารถใชแทนแปงสาลในสตรการผลตขนมปงกรอบไดสงถง 50% โดยทผบรโภคไมสามารถบอกความแตกตางจากขนมปง
กรอบสตรทใชแปงสาล 100%ได ทงในดานรสชาต ความกรอบ ส และกลน จากการวเคราะหองคประกอบของแปงเมลดมะมวง แปงสาล แปงผสมเมลดมะมวงกบแปงสาล และขนมปงกรอบทผลตจากแปงผสมดงกลาว พบวามองคประกอบและคณคาทางโภชนาการทไมแตกตางกน
อยางมนยสาคญ ดงแสดงในตารางท 1(5)
ตารางท 1 องคประกอบของแปงเมลดมะมวง แปงสาล แปงผสม เมลดมะมวง:แปงสาล (50:50) และ ขนมปงกรอบทผลตจากแปงผสม(5)
องคประกอบ(กรม/กโลกรม) ชนดของแปง ขนมปงกรอบทผลตจาก
แปงผสม เนอในมะมวง สาล แปงผสม ความชน 91.0 136.8 122.9 77.1 โปรตน 66.1 134.0 93.0 68.1 ไขมน 94.0 10.0 65.1 182.0 เสนใย 28.0 3.8 13.8 10.3 คารโบไฮเดรตทละลายนา (ในรปนาตาลซโครส) 27.5 31.9 57.0 179.0 แปง 500.0 657.0 594.7 336.1 คารโบไฮเดรตทงหมด 182.6 20.4 46.3 136.5 แคลอร (kJ/กโลกรม) 16.6 14.5 15.4 18.9 pH 4.5 4.4 4.4 4.4 ความเปนกรด (เทยบกบกรดซตรก) 17.5 7 10.5 10.5 กรดแอสคอบก 0.17 0.08 0.09 0.00 แทนนน (เทยบกบ แคทเทชน) 23.4 0.6 15.1 1.3

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
156
สาหรบแปงบางชนดทมคณคาทางโภชนาการตา
เชน แปงมนสาปะหลง แปงขาวโพด แปงขาวฟาง การผสมกบแปงเมลดมะมวงจะทาใหปรมาณไขมนและ
โปรตนของแปงเหลานสงขน(3) ปรมาณของคารโบไฮเดรตทสงทาใหแปงชนดนเหมาะทจะนามาใชเปนสารตงตนใน
การยอยดวยเอนไซมแอลฟาแอมเลส (α-amylase) และ
กลโคแอมเลส (glucoamylase) เพอผลตนาเชอมกลโคส(28)
แปงเนอในเมลดมะมวงนถงแมวาจะมปรมาณ
โปรตนคอนขางตาเมอเปรยบเทยบกบปรมาณโปรตนใน
แปงทใชบรโภคทวไป (ตารางท 1) แตนบวาเปนแหลงโปรตนทสาคญเนองจากมกรดอะมโนทจาเปนตอรางกาย
เปนสวนประกอบในอตราสวนสงถง 32% ของกรดอะม-โนทงหมด และกรดอะมโนจาเปนทพบในเนอในเมลดมะมวงนมคาสงกวาคามาตรฐานทกาหนดโดย FAO/WHO
(ตารางท 2)(1) ยกเวน methionine, threonine และ tyrosine
เมอพจารณาในแงปรมาณและคณคาเชงพาณชย
เพอการผลตนามนสาหรบบรโภค นามนจากเนอในเมลดมะมวงมศกยภาพในการนามาผลตในระดบอตสาหกรรม
เพราะมปรมาณไขมนในระดบ 11-18% ซงนบวาเปนปรมาณทสงเมอเปรยบเทยบกบพชนามนชนดอนๆ ทมการผลตอยในปจจบน เชน ขาวโพดม 5% ถวเหลอง 18% เมลดฝาย 15%(10) สวนกระบวนการผลตและการทาใหนามนบรสทธนนไมยงยากและมตนทนตา เพราะนามนเมลดมะมวงทนตอการเกดออกซเดชน และมปรมาณของเอนไซมลเพส (lipase) คอนขางนอย ทาใหนามนมความเปนกรดตา นามนพชทผลตจากประเทศเขตรอนชนทวไปนนประกอบดวยกรดไขมนสายโซยาวปานกลางและอมตว เชน กรดลอรก (lauric acid) ทมปรมาณมากในนามน
มะพราว หรอกรดปาลมตก (palmitic acid) ในนามนปาลม
ตารางท 2 องคประกอบของกรดอะมโนของแปงเมลดมะมวง (mean± SD, n=4) เปรยบเทยบกบโปรตนมาตรฐานท กาหนดโดย FAO/WHO (กรมของกรดอะมโน/100 กรมของโปรตน)(1)
กรดอะมโน เนอในเมลดมะมวง มาตรฐาน FAO/WHO
Leucine Isoleucine Methionine Phenylalanine Lysine Threonine Tyrosine Valine
6.9± 0.2 4.4± 0.2 1.2± 0.1 3.4± 0.1 4.3± 0.2 3.4± 0.2 2.7± 0.1 5.8± 0.3
4.8 4.2 2.2 2.8 4.2 4.0 4.1 4.2
กรดอะมโนทจาเปนทงหมด 32.1± 2.2
Aspartic acid Glutamic acid Serine Proline Glycine Alanine Histidine Arginine
6.5± 0.4 18.2± 0.8 3.3± 0.1 3.5± 0.2 4.0± 0.4 4.2± 0.4 5.5± 0.6 7.3± 0.8
กรดอะมโนทไมจาเปนทงหมด 52.2± 2.1

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
157
แตนามนเมลดมะมวงมคณคาทางโภชนาการสง และแตกตางจากนามนพชทวไปในเขตรอนชนเนองจากม
ปรมาณกรดไขมนจาเปน คอ กรดโอเลอก (oleic acid) และกรดลโนเลอก (linoleic acid) ในปรมาณทสงคลาย คลงกบนามนมะกอก (ตารางท 3) มสารตานอนมลเสรกลมทละลายในนามน คอฟอสโฟลปด (phospholipids) และกลมไฟโทสเตอรอล (phytosterol) เชน สควาลนสเตอรอล (squalene sterol) และโทโคฟรอล (tocopherol) ถง 0.34 % นบเปนปรมาณทสงเมอเทยบกบนามนทบรโภค
ทวไปซงมปรมาณของไฟโทสเตอรอล 0.1-0.5%(16)
เมอพจารณาถงชนดและปรมาณของกรดไขมนท
เปนองคประกอบของไตรกลเซอไรดในนามนเมลดมะมวง
นน พบวามลกษณะพเศษมากเพราะมความคลายคลงกบนามนทหายาก คอเนยโกโก ทงชนดของกรดไขมน และตาแหนงของกรดไขมนบนไตรกลเซอไรด ดงแสดงในตารางท 3(11)
มะมวงกจะเปนสงทตรวจสอบไดยากมาก(8) ในปจจบน ทวปแอฟรกาผกขาดการผลตเนยโกโก และปรมาณทผลต
ไดกไมแนนอนขนอยกบสภาพอากาศ จงทาใหเนยโกโกมราคาคอนขางแพง ดงนน ในประเทศทไมสามารถผลตเนยโกโกได เชนประเทศไทย การผลตนามนเมลดมะมวงมาทดแทนเนยโกโกจงนาสนใจ เปนความทาทาย และจะทาใหการผลตชอกโกแลตมราคาถกลง ในแถบเอเชย ปจจบนมการผลตนามนเมลดมะมวงในระดบอตสาหกรรมใน
ประเทศอนเดยเทานน
เนยขาว (shortening) เปนสวนผสมทสาคญของ
ขนมปง ขนมปงกรอบ และ เคก ซงเนยขาวนนมไขมนทมกรดไขมนสายโซยาวและอมตว เชน กรดปาลมตก และกรดสเตอรกเปนองคประกอบ เนยขาวทางการคาผลตโดยกระบวนการเตมไฮโดรเจน (hydrogenation) ซงมขอเสย
คอกระบวนการนจะกอใหเกดกรดไขมนทรานส (trans-fatty acid) สงถง 20-40% ปจจบนมรายงานทางการแพทย
ตารางท 3 ปรมาณของกรดไขมน (%) ของนามนเนอในเมลดมะมวง เนยโกโก และ นามนมะกอก(11)
ชนดของกรดไขมน ปรมาณของกรดไขมน (%) ของนามน
เนอในเมลดมะมวง เนยโกโก มะกอก กรดไมรสตก (C14:0) 0.5± 0.1 - 0.0-1.5 กรดปาลมตก (C16:0) 5.8± 0.3 23-26 7.5-20 กรดสเตอรก (C18:0) 38.3± 1.2 35-37 0.5-5.0 กรดโอเลอก (C18:1) 46.1± 2.3 34-37 55.0-83.0 กรดไลโนเลอก (C18:2) 8.2± 2.3 1-3 3.5-21.0 กรดไลโนเลนก (C18:3) 1.2± 0.2 - 0.0-1.5
เนยโกโกม ลกษณะพ เศษมากและ เปนองค ประกอบหลกของชอกโกแลต มนามนเพยงไมกชนดทมคณสมบตเชนน คอมจดหลอมเหลวท 35oC ใกลเคยงกบอณหภมของรางกายมนษย จงทาใหหลอมละลายอยางรวดเรวในปากขณะทกาลงเคยว คณสมบตเชนนเปนเพราะเนยโกโกมสวนประกอบของกรดปาลมมตก กรดสเตอรก และกรดโอเลอกในอตราสวนทเหมาะสม และกรดโอเล-อกจะตองอยทตาแหนง sn-2 ของไตรกลเซอไรด ฉะนนนามนเมลดมะมวงนบวาเปนนามนเพยงไมกชนดทม
ปรมาณของกรดปาลมมตก กรดสเตอรก และกรดโอเลอกทตาแหนง sn-2 ซงคลายคลงกบเนยโกโกมากจนถงขนาดทหากมการปลอมแปลงเนยโกโก โดยการเตมนามนเมลด
วากรดไขมนทรานสเปนตวการสาคญทกอใหเกดโรค หวใจ ทาใหปรมาณของไขมน HDL ในเลอดลดลงในขณะ ทปรมาณของไขมน LDL สงขน การบรโภคกรดไขมน ทรานสในอตราสวนทสงกวา 4% ของแคลอรทงหมดจะ
เพมปรมาณไขมนชนดเลวในเลอดใหสงขน ดงนน จงไดมความพยายามทจะผลตเนยขาวทปราศจากกรดไขมน ทรานสขน และน ามนเมลดมะมวงมองคประกอบทเหมาะสมเพราะมกรดสเตอรกในปรมาณสง การผลตเนยขาวนนจะตองสกดนามนเมลดมะมวงเพอแยกเอาสวนทม
กรดสเตอรกสงดวยตวทาละลาย และพบวาสวนดงกลาว
นนซงปราศจากกรดไขมนทรานส มปรมาณสงถง 55% ทสามารถใชทดแทนเนยขาวทางการคาในการผลตขนมปง เคกและขนมปงกรอบได ถงแมเนยขาวจากเนอในเมลด

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
158
มะมวงจะมราคาแพงกวาเนยขาวทผลตโดยกระบวนการ
เตมไฮโดรเจน แตมขอดทประมาณคามไดคอความ
ปลอดภยและสขภาพของผบรโภค(22)
การใชเนอในเมลดมะมวงเปนอาหารจะตองคานง ถงรสชาตและความปลอดภยควบคกนไป ตามปกตเนอในเมลดมะมวงมรสฝาดเนองจากมสารแทนนนเปนสวน ประกอบในปรมาณสง แตสารแทนนนละลายนาไดด ดงนนการลดปรมาณแทนนนจงทาไดงายโดยแชเนอใน
เมลดมะมวงในนาหรอนาไปอบหรอตมทอณหภมสง วธการเหลานสามารถลดปรมาณของแทนนนได ถง 48% เชอวาการบรโภคเนอในเมลดมะมวงนนนาจะมความปลอดภยสง เพราะมการรบประทานมาตงแตสมยโบราณ สวนการศกษาทางวทยาศาสตรนนพบวาคนทมนาหนกตว 70 กโลกรมสามารถรบประทานเนอในเมลดมะมวง 400 กรม หรอแปงของเนอในเมลดมะมวงทผานกระบวนการ 800 กรมไดโดยไมกอใหเกดอนตราย(23)
การประยกตใชในผลตภณฑทไมใชอาหาร สารกนเสย และสารกนหนธรรมชาต
เนอในของเมลดมะมวงนนอดมไปดวยสารทม
ฤทธตานอนมลเสรสง ประเภทพอลฟนอล เชน แทนนนทสลายไดเมอทาการแยกสลายดวยนา (hydrolyzable tannin) และแกลโลแทนนน (gallotannin) แทนนนทแตกตวหรอสลายดวยนาไดยาก (condensed tannin) หรออกนยหนงคอ โปรแอนโทไซยานดน (proanthocyanidin) และสารฟนอ-ลก เชน กรดแกลลก (gallic acid) กรดแอลลาจก (ellagic acid) และเอพแคทเทชน (epicatechin) (รปท 1)(6, 20) กรดแกลลกทพบในเมลดมะมวงมจานวนประมาณ 23-838 มลลกรม/100 กรมของนาหนกแหง สวนกรดแอลลาจกมปรมาณตงแต 3-126 มลลกรม/100 กรม ขนกบวธการสกด สายพนธมะมวงและแหลงทปลก กรดแกลลกและกรดแอลลาจกอาจอยในรปสารอสระหรอสารประกอบโดย
การสรางพนธะเอสเทอร (ester bond) กบนาตาลในรปของแกลโลแทนนน(17)
เปนททราบกนวา สารพอลฟนอลและสารฟนอลก เปนสารทมฤทธแรงทงในการตานอนมลเสรและยบยงการ
เจรญของเชอจลนทรย (2) ดงนน สารสกดจากเนอในเมลดมะมวงจงเปนทงสารกนหนและสารกนเสยชวภาพทชวย
ยดอายผลตภณฑตางๆ ได นอกจากน ในนามนของเนอในเมลดมะมวงยงพบสารกลมฟอสโฟลปดอก 8 ชนดไดแก phosphatidyl serine, lyso-phosphatidyl choline, phospha-tidyl inositol, sphingomyelin, phosphatidyl choline,
phosphatidyl ethanolamine, phosphatidic acid และ
สารประกอบของ glycerophosphatidyl ทมฤทธตานอนมลเสรดวย ในประเทศอนเดย มการทดลองนาสารสกดจากเนอในเมลดมะมวงทมทงสารฟนอลกและฟอสโฟลปด
เปนสวนประกอบมาใชเปนสารกนหนในเนยก (buffalo gee) ซงเปนอาหารทบรโภคกนมากในประเทศ จากการ ศกษาพบวา ปรมาณของสารสกด 5% มคณสมบตตานการหนในเนยกไดดกวา butylated hydroxyanisole (BHA) ซงเปนสารกนหนสงเคราะหในปรมาณทอนญาตใหเตมได 0.02% โดยนาหนก/ปรมาตร สารสกดจากเนอในเมลดมะ- มวง 5% ยดอายการเกบเนยกไดนานถง 928 ชวโมงใน ขณะท BHA 0.02% ยดอายการเกบเนยกไดเพยง 210.6 ชวโมงเทานน(20)
นอกจากมคณสมบตเปนสารตานอนมลเสรแลว ในหลายประเทศยงใชสารสกดจากเนอในเมลดมะมวงเปน
สารกนเสยอกดวย ในประเทศญปนไดอนญาตใหใชกรดแอลลาจกเปนสารกนเสยในอาหาร ในขณะทแกลโลแทนนนถกจดใหเปนสารเตมแตงอาหารทคอนขางปลอดภย (GRAS; generally recognized as safe) เนองจากในรางกายของสงมชวต แกลโลแทนนนถกยอยงายไดเปนกรดแกล-ลกหรอกรดแอลลาจก การศกษาโดยสกดสารพอลฟนอลดวยแอลกอฮอล 95% ไดสารสกดทมปรมาณของสาร พอลฟนอลเปนสวนประกอบสงถง 80% สารสกดนมฤทธยบยงการเจรญของเชอจลนทรยทกอโรคในอาหารทงกลม
แกรมบวกและลบททาการทดสอบรวม 43 สายพนธ(14) โดยมคา MIC (minimum inhibitory concentrations) ตงแตนอยกวา 50 ถง 2500 ppm แตสามารถยบยงการเจรญของแบคทเรยแกรมบวกไดดกวาแบคทเรยแกรมลบ ดงนน การใชสารสกดเนอในเมลดมะมวงเปนสารกนเสยจง
จาเปนตองใชรวมกบสารกนเสยชนดอนเพอใหความ สามารถในการยบยงแบคทเรยแกรมลบสงขน
เครองสาอาง
เนอในเมลดมะมวงใช เปนสวนประกอบของ
เครองสาอางทชวยชะลอความแกได เพราะประกอบดวยนามนทใหความชมชนพเศษตอผวหนง และสารพอลฟ-นอล และสารฟนอลก ซงเปนสารตานอนมลเสรทมฤทธแรงดงทกลาวขางตน นามนเมลดมะมวงมกรดโอเลอกเปนองคประกอบ เมอผสมในเครองสาอางจะทาหนาทเปนสารใหความนมชมชนเชนเดยวกบเนยโกโกและนามนมะกอก นอกจากน การทนามนชนดนมกรดสเตอรกในปรมาณสงจงทาใหเหมาะทจะใชเปนวตถดบในการผลตสบกอนทม
เนอสมผสแขง มผวมนวาวเหมอนกบนามนปาลม แตมขอ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
159
ดกวานามนปาลมคอสบทผลตจากนามนเมลดมะมวงทม
กรดโอเลอกเปนสวนประกอบหลกนนจะใหฟองสบทม
ขนาดเลกกวา ทนตอการฟอก และมลกษณะเปนครมนม ทาใหระคายเคองตอผวหนงนอยกวาสบจากกรดปาลมตก
โดยปกต อนมลเสรในรางกายเกดขนตลอดเวลาทงจากมลภาวะภายนอกและความเครยดภายในรางกาย อนมลเสรทเกดขนมผลในการทาลายเซลล เปลยนแปลงโครงสรางทางเคมและหนาททางชวภาพของสารชว-โมเลกลตางๆ เชนโปรตน ไขมน กรดนวคลอก ทาใหเกดการเสอมสภาพและการทาลายเซลลตางๆ เกดเปนพษเรอรงตอเนอเยอและพยาธสภาพตอรางกาย เชน ทาใหความจาเสอม หลอดเลอดหวใจอดตน ผวหนงเหยวยน สญเสยนาและความยดหยนเปนตน(12) สารตานอนมลเสรในเมลดมะมวงมฤทธแรงและจบกบอนมลเสรไดอยางม
ประสทธภาพ ดงนน ผลตภณฑทมสารสกดจากเนอใน
เมลดมะมวงเปนสวนผสมจงสามารถใชเปนเครองสาอาง
ชะลอความแก เปนสารบารงผว(9) และใชเปนอาหารเสรมทชวยเสรมสรางภมคมกนและปองกนรางกายจากพยาธ
สภาพตางๆ ทเกดจากอนมลเสร เชน โรคความจาเสอม โรคหวใจ มะเรง เปนตน(27) บรษท Snow Brand Milk Products ประเทศญปน มลขสทธการใชสารสกดเนอในเมลดมะมวงเปนสวนผสมในผลตภณฑตางๆ ซงสามารถยบยงการเจรญของเชอ Propionibacterium acnes ททาใหเกดสว และ Streptococcus sanguinis ซงเปนจลนทรย
ประจาถนในคราบฟน ดงนนสบ แชมพ ครม และยาทาผวทมสารสกดเนอในเมลดมะมวงเปนสวนประกอบจง
อาจจะชวยปองกนหรอรกษาสวได ในขณะทอาหารเสรม ยาสฟนและนายาบวนปากทมสารสกดเนอในเมลดมะมวง
เปนสวนประกอบนาอาจจะชวยปองกนการเกดคราบ
หนปนได โดยปรมาณทแนะนาใหเตมในผลตภณฑคอ 0.001-5%(18)
การใชสารสกดในผลตภณฑตางๆ จะตองคานงถงความเสถยรของสารสกดนนเมอผานกระบวนการผลต หรอเมอนาไปเตมในผลตภณฑทมคณสมบตทางฟสกส/เคมทอาจจะทาใหสารสกดเสยสภาพ เชน ทอณหภม หรอ pH สงหรอตามากๆ จากการศกษาความเสถยรของสารสกดเนอในเมลดมะมวง พบวาเปนสารทเสถยรตอทงกระบวนการผลตและตอคณสมบตของผลตภณฑอาหาร
และเครองสาอางทวไป (pH 3-9 อณหภม -20 ถง 121oC) ทาใหสามารถเตมสารสกดเนอในเมลดมะมวงในอาหาร
และเครองสาอางทวไปได
อาหารสตว
เนอในเมลดมะมวงซงประกอบดวยแปง โปรตน ไขมน และสารพอลฟนอล ทาใหสามารถใชเปนทงแหลงอาหารหลกเพอการเจรญเตบโต และเสรมสรางภมตานทานของปศสตวได ไดมการทดลองใชเนอในเมลดมะม วง เปนอาหารทดแทนอาหารของลกปลาน ล (Oreochromis niloticus)(19) หรอใชแทนขาวโพดในการเลยงไกเนอ(13) พบวาเนอในเมลดมะมวงดบใชทดแทนอาหารหลกไดในปรมาณไมเกน 10% แตถาใชเนอในเมลดมะมวงทตมสกแลวจะใชไดในปรมาณทสงขนถง 20% โดยไมมผลกระทบตอการเจรญเตบโตของสตวเลยง ไดมการศกษาการเตมเนอในเมลดมะมวงในอาหารหลกเพอ
เสรมสรางภมตานทานในระบบฟารมทมการเลยงสตวนา
อยางหนาแนนในบอ ซงเปนวธการเลยงทประสบปญหามาก เนองจากการเลยงอยางหนาแนนจะทาใหสตวนาตายงายเนองจากการตดเชอ ดงนน จงจาเปนตองใชสารเคม และยาปฏชวนะเพอปองกนโรคตดเชอในปรมาณมาก กอใหเกดการตกคางของยาปฏชวนะทงในรางกายของ
ผบรโภคและในสภาพแวดลอม และอนตรายเนองจากยาปฏชวนะทตกคางอาจจะกอใหเกดการคดเลอกเชอดอยา เปนอนตรายตอสขภาพและไมเปนทยอมรบของผบรโภค ปจจบนจงมความพยายามทดแทนการใชยาปฏชวนะดวย
วธทางธรรมชาต เชน การใช สารเสรมชวนะ (probiotics) สารเคมจากพชหรอพชสมนไพร(21) ซงมฤทธชวยกระตนและเพมภมตานทานทาใหสตวนามสขภาพแขงแรง
เน อในเมลดมะมวงใหสารท เปนประโยชน
มากมายจงเปนทสนใจอยางมากเนองจากเปนของเหลอ
และทงไป หากคดเปนมลคาจงมราคาถก ทงยงมฤทธในการยบยงการเจรญของแบคทเรย รา ไวรส สามารถใชรกษาอาการอกเสบ รกษาโรคอหวาตกโรค เปนสารตานอนมลเสรปองกนการเกดเนองอก จากการทดลองผสมผงเนอในเมลดมะมวง 0.5%ในอาหารเลยงลกปลายสกเทศ (Labeo rohita) และใชเปนเวลา 60 วน จากการวเคราะหเลอดปลาพบวา ภมต านทานแบบไมจ า เพาะ (innate response) สงขน โดยการสราง superoxide anion และ ไลโซไซม ซงมฤทธในการทาลายเชอโรคทเขามาในตวปลาไดดขน นอกจากนพบวาระดบของโปรตนแอลบมนและโกลบลนในเซรมสงขนดวย เมอนาปลายสกมาทดสอบโดยการใหเชอ Aeromonas hydrophila ซงปกตแลวจะทาใหปลาตายดวยโรค motile aeromonas แตพบวาหลงจากปลาไดรบเชอ 10 วน ปลาซงไดกนอาหารทมสวนผสมเนอในเมลดมะมวง 0.5% มอตราการรอดชวต

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
160
จากโรคดงกลาวถง 98% เมอเปรยบเทยบกบปลายสกกลมควบคม ดงนน เนอในเมลดมะมวงจงเหมาะสมทจะนามา ใชผสมในอาหารปลาเพอเพมภมตานทานในการเลยงปลา
ยสกเทศแทนยาปฏชวนะโดยไมมผลกระทบตอการ
เจรญเตบโต ขอดของเนอในเมลดมะมวงนนอกจากจะเปนสารจากธรรมชาตทยอยสลายได ไมตกคางและปลอดภยทงตอสภาพแวดลอมและผบรโภคแลวยงเปนสารทมราคา
ถก(25)
สรป ประโยชนของเมลดมะมวงไมควรมองขามอกตอ
ไป ในยคทอาหารกาลงมจานวนจากด พลงงานกาลงขาดแคลน ขณะทสารจากธรรมชาตกาลงเปนผลตภณฑมลคาสงเปนทตองการของสงคมโลกเปนอยางมาก นอกจากขยะเนอในเมลดมะมวงทเกดจากการบรโภคและการแปร
รปในระดบอตสาหกรรมแลว ในประเทศไทยเรายงมขมทรพยของเนอในเมลดมะมวงจานวนมหาศาลทยง
ไมไดมการศกษาวจยเพอนามาใชประโยชนใหครบถวน เชนเนอในของเมลดมะมวงอกหลายสายพนธทไมเปนท
นยมของผบรโภค เพราะมเนอนอย รสชาตไมอรอย และมเมลดขนาดใหญเกนไป เชน มะมวงปาและมะมวงกะลอน นกวจยไทยจงควรจะใหความสนใจศกษาวจยเนอในเมลด
มะมวงและประยกตใชอยางครบถวน อนจะเปนประโยชนในการเพมมลคาวตถดบและลดปญหาสภาพแวดลอมของ
ประเทศจากขยะเมลดมะมวงตอไป
เอกสารอางอง 1. Abdalla AEM, Darwish SM, Ayad EHE, El-
Hamahmy RM. Egyptian mango by-product 1. Compositional quality of mango seed kernel. Food Chem 2007;103:1134-40.
2. Ahmed I, Aquil F, Owais M. Modern phytomedicine; turning medicinal plants into drugs. Weinheim: Wiley-VCH Verlag GmbH&co KgaA; 2006.
3. Arogba SS. Physical, chemical and functional properties of nigerian mango (Mangifera indica) kernel and its processed Flour. J Sci Food Agric 1997;73: 321-8.
4. Arogba SS, Ajiboye OL, Ugboko LA, Essiennette SY, Afolabi PO. Properties of polyphenol oxidase in mango (Mangifera indica) kernel. J Sci Food Agric 1998;77:459-62.
5. Arogba SS. The performance of processed mango (Mangifera indica) kernel flour in a model food system. Bioresource Technol 1999;70:277-81.
6. Arogba SS. Mango kernel: chromatographic analysis of the tannin and stability study of the associated
polyphenols oxidase activity. J Food Compos Anal 2000;13:149-56.
7. Bhatanagar HC, Subramanyam H. Some aspects of preservation, processing and export of mango and its products. Expert group Meeting; Vienna; UNIDO, ID/WG 88/15;1971.
8. Crews C. Authentication of cocoa butter; in oils and fats authentication. Michael J, editor. New York: Blackwell Publishing Ltd; 2002. p.66-89.
9. Fox C. Advance in the cosmetics science and technology of topical bioactive materials. Cosmet Toillet Magazine 1997;112:67-84.
10. Gunstone FD. Production and trade of vegetable oil. In: Vegetable oils in food technology, composition, properties and uses. Oxford: Blackwell Publishing Ltd; 2002. p.1-17.
11. Gunstone FD, Padley FB. Lipid technologies and applications. New York: Marcel Dekker Inc; 1997.
12. Gutterridge JMC, Halliwell B. Antioxidants in nutrition, health, and disease. Oxford: Oxford University Press; 1996.
13. Joseph JK, Abolaji J. Effect of replacing maize with graded levels of cooked Nigerian mango seed kernels (Mangifera indica) on the performance, carcass yield and meat quality of broiler chickens. Bioresource Technol 1997;61:99-102.
14. Kabuki T, Nakajima H, Arai M, Ueda S, Kuwabara Y, Dosako S. Characterization of novel anti-microbial compounds from mango (Mangifera indica L.) kernel seeds. Food Chem 2000;71:61-6.
15. Kaur M, Singh N, Sandhu KS, Guraya HS. Physicochemical, morphological, thermal and rheological properties of starches separated from kernels of some Indian mango cultivars (Mangifera indica L.). Food Chem 2004;85:131-40.
16. Lercker G, Rodriguez-Estrada MT. Chromatographic analysis of unsaponifiable compound of olive oils and fat-containing foods. J Chromat A 2000;881: 105-29.
17. Masibo M, He Q. Major mango polyphenols and their potential significance to human health. Comp Rev Food Sci Food Safety 2008;7:309-19.
18. Nakajima H, Tadokoro H, Hashiba H, Ito F, Furuya H, Kabuki T et al. Bacteriostatic and antibacterial agent containing mango kernel component. US patent 2009 [cited Feb 2009]. Available from: URL: http://www.patentstorm.us/patents/6063382.html
19. Omoregie E, Ufodike EBC, Umara MS. Growth and feed utilization of Orechromis niloticus fingerlings fed diets containing cassava peeling and mango seeds. Aquabute 1991;4:6-7.
20. Puravankara D, Boghra V, Sharma RS. Effect of antioxidant principles isolated from mango (Mangifera indica L) seed kernels on oxidative

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
161
stability of buffalo gee (butter-fat). J Sci food Agric 2000;80:522-6.
21. Rao YV, Das BK, Pradhan, J, Chakrabarti R. Effect of achyranthes aspera on the immunity and survival of labeo rohita infected with Aeromonas hydrophila. Fish Shellfish Immonol 2006;20:263-73.
22. Reddy SY, Jeyarani T. Trans-free bakery shortening from mango kernel and mahua fats by fractionation and blending. J Am Oil Chem Soc 2001;78:635-40.
23. Rukmini C, Vijayaraghavan M. Nutritional and toxicological evaluation of mango kernel oil. J Am Oil Chem Soc 1984;61:780-92.
24. Sandhu KS, Lim S-T. Structural characteristic and in vitro digestibility of mango kernel starches (Mangifera indica L.). Food Chem 2008;107:92-7.
25. Sahu S, Kumar Das B, Pradhan J, Mohapatra BC, Mishra BK, Sarangi N. Effect of Mangifera indica kernel as feed additive on immunity and resistance to Aeromonas hydrophila in Labeo rohita fingerlings. Fish Shellfish Immunol 2007;23:109-18.
26. Schieber A, Ullrich W, Carle R. Characterization of polyphenols in mango puree concentrate by HPLC with diode array and mass spectrometric detection. Innovat Food Sci Emerg Tech 2000;1:161-6.
27. Shahidi F, Naczk M. Phenolics in food and nutraceuticals. Florida: CRC press; 1951.
28. Velan M, Krishanan RV, Lakshmanan CM. Conversion of mango kernel starch to glucose syrups by enzymatic hydrolysis. Bioprocess Bioeng 1995; 12:323-6.
29. Zein RE, El-Bagoury AA, Kassab HE. Chemical and nutritional studies on mango seed kernels. J Agric Sci Mansoura University 2005;30;3285-99.


163
ทรพยากรพนธกรรมกลวยในประเทศไทย...ทรพยแหงสวรรณภม
ศศวมล แสวงผล1 และ จามร สมณะ2 1ภาควชาพฤกษศาสตร 2ภาควชาชวเคม คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
ประเทศไทยตงอยบนดนแดนสวรรณภมในใจกลางภาคพนทวปของภมภาคเอเชยตะวนออกเฉยงใตซงเปน
แหลงกาเนดกลวย จากการสารวจพนธกรรมกลวยทวทกภาคของประเทศไทย พบพชในวงศกลวย (Musaceae) 13 ชนดจาก 3 สกล ในจานวนน ม 2 ชนดทนาจะเปนชนดใหมทยงไมเคยมรายงานมากอน และ 2 ชนดเปนพอแมของกลวยพนธปลกไมมเมลดสวนใหญในโลก หนงในสองชนดนนคอ กลวยปา (Musa acuminata Colla) ซงพบวามถง 4 ชนดยอย (subspecies) ทพบในปาธรรมชาต นอกจากนในทองถนตางๆ ยงมการเกบรกษากลวยพนธปลก (cultivar) อกมากกวา 100 พนธทมความแตกตางกนในรปลกษณ และการใชประโยชน การพบความหลากหลายเชนนเนนยาความสาคญของการอนรกษทงสภาพพนทปาสมบรณ และทรพยากรพนธกรรมกลวย ซงจะมความสาคญตอการนาลกษณะทางพนธกรรมบางประการ เชน สายพนธทนแลง ทนนาทวม ทนเคม ผลผลตสง หรอการมคณคาพเศษทางโภชนาการ มาใชประโยชนเพอความมนคงทางอาหารและการเกษตรของประเทศไทยในอนาคต
คาสาคญ: สกลกลวย; สกลกลวยโทน; สกลกลวยดารารศม; ความหลากหลายทางชวภาพ
Abstract Thailand, located at the heart of mainland Southeast Asia, is among the cradles of bananas. Our extensive surveys throughout Thailand have revealed 13 species in three genera of the banana family, Musaceae. Among these, two species are probably new and another two species have been parents of most of the world seedless edible cultivars. One of the latter species Musa acuminata Colla has as many as four subspecies found in natural habitats. More than 100 banana cultivars have been found in several native localities. Species and cultivar diversity, i.e. unique morphology and utility, indicate the importance of conservation of natural forest areas and the banana genetic resources. Such genetic traits as draught, flood, and salt tolerance, high yield and specific nutritional value would, in the near future, ensure food and agricultural security for the nation. Keywords: Musa; Ensete; Musella; biodiversity
บทนา กลวยเปนพชเศรษฐกจสาคญ เปนผลไมทมการ
ผลตเพอรบประทานผลเปนการคามากทสดในโลก คอราว 73 ลานตนตอป (ขอมลป พ.ศ. 2548) มลคาของผลผลตกลวยในตลาดโลกสงทสดเปนอนดบทสรองจากขาว ขาวสาล และขาวโพด โดยประเทศอนเดยมการผลตกลวยมากทสด(16) นอกจากมลคาในตลาดโลกแลว กลวยยงเปนผลไมสาคญในแงทเปนแหลงอาหารหลกและแหลงธาต
อาหารสาคญในทองถนสาหรบประชากรหลายรอยลาน
คนซงมฐานะยากจนทสวนใหญอาศยอยในเขตรอนใน
ทวปเอเชย แอฟรกา อเมรกากลาง และอเมรกาใต(14)
กลวยผกพนกบคนไทยมานานแสนนานและม
ความสาคญในวถชวตของคนไทยตงแตเกด นอกจากนานมแลว เดกทารกสวนใหญมกรบประทานกลวยสกบดเปนอาหารชนดแรก เพราะยอยงายและมคณคาทาง
โภชนาการสง กลวยไมเปนเพยงผลไมเพอรบประทานผลสกและแปรรปเทานน คนไทยยงสามารถใชประโยชนจากกลวยทงตนอกมากมาย เชน หยวก ปล ผลออนเปนผก กาบใชทาเชอก และกระดาษ ใบใชหอขนมและอาหาร รวมทงนาลาตน กาบ หรอใบมาใชในงานศลปหตถกรรมตางๆ นอกจากนน กลวยยงมความสาคญในการประกอบพธกรรมของผคนทกชนชนมาตงแตโบราณ เชน ใชในพธขนอ บายศรสขวญ บชาแมโพสพ บวชนาค ยกขนหมาก แมกระทงในพธศพ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
164
ในประเทศไทยมแหลงปลกกลวยเพอการคาหลาย
แหง เชน กลวยหอมในอาเภอทายาง จงหวดเพชรบร กลวยไขในจงหวดกาแพงเพชร กลวยเลบมอนางในจงหวดชมพร กลวยหนในจงหวดยะลา และกลวยตานในอาเภอ สวรรคโลก จงหวดสโขทย เปนตน ในป พ .ศ . 2550 ผลผลตเพอสงออกซงสวนใหญเปนกลวยไขและกลวย
หอม มปรมาณราว 15 และ 3 ลานตน ตามลาดบ(1) สวนกลวยชนดอนๆ มกใชบรโภคภายในทองถน
ประเทศไทยตงอยในบรเวณรอยตอของเขตพช
พรรณสาคญของโลก ไดแก อนโดจน และซนดาแลนด ซงมภมอากาศแตกตางกนมาก ทาใหภมภาคแถบนเปนแหลงทมความหลากหลายทางชวภาพสงทสดใน 8 ลาดบแรกของโลก มพชพรรณรวมแลวมากกวา 10% ของพชพรรณทงหมดในโลก(17,22)
ผลการสารวจกลวยในอดต แสดงใหเหนวาดนแดนอนโดจนและปาปวนวกนเปนแหลงกาเนดกลวยทสาคญ
ทสดของโลก โดยเฉพาะในประเทศไทย มการคนพบกลวยปา Musa acuminata Colla ถง 4 ชนดยอยจากทง หมด 5 ชนดยอย(20) และมกลวยพนธปลกไมมเมลดเกอบครบทกกลมจโนมจากกลวยกวา 60 พนธทรวบรวมพนธเมอป พ.ศ. 2527(9) อยางไรกตาม ประเทศไทยไมไดเขารวมการเกบรกษาพนธกรรมกลวยในธนาคารเชอพนธ
กลวยระหวางประเทศ (ITC-INIBAP, International Transit Center-International Network for the Improve-ment of Banana and Plantain) ซงดาเนนการในประเทศเบลเยยมรวมกบประเทศอนๆ อก 18 ประเทศ อกทงมตวอยางกลวยจากประเทศไทยในบญชรวมนอยมาก เพยง 8 ตวอยางจากจานวนกลวยจากทวโลกทงหมดถง 5,347 ตวอยาง(15) นอกจากน ยงไมมรายงานใดทแสดงผลการสารวจกลวยทครอบคลมทงวงศกลวยในประเทศไทย
กลวยแหงอษาคเนย
กลวยมการกระจายพนธทวไปในบรเวณเอเชย
ตะวนออกเฉยงใต เอเชยใต และเอเชยตะวนออก และมหลกฐานวามการนากลวยไปปลกในประเทศเขตรอนทว
โลกมาตงแตยคหนใหม ซงมการอพยพยายถนจากแผนดน ใหญ ใน เอ เช ยตะวนออกเฉ ยงใต ไปย งหม เ กาะใน
มหาสมทรแปซฟกและแอฟรกา(11,18) นกวชาการบางทานเสนอวากลวยอาจเปนพชปลกชนดแรกๆ ของมนษย(10) โดยมการนาหนอกลวยมาปลกใกลบานเพอเกบผลและใช
ประโยชนจากสวนอนๆ มารโคโปโล นกเดนทางทม
โอกาสไดเหนตนกลวยและความสาคญของกลวยตอชน
ชาตตางๆ ในภมภาคแถบนยงเรยกกลวยวา แอปเปลของ
อดม และ Linnaeus นกพฤกษศาสตรชาวสวเดนยงไดตง
ชอกลวยจากเอเชยตะวนออกเฉยงใตเปนภาษาละตนวา Musa sapientum และ M. paradisiaca แปลวาตนไมของนกปราชญ และตนไมจากสรวงสวรรค ตามลาดบ(10)
นอกจากหลกฐานทางโบราณคดและมานษยวทยา
แลว นกภาษาศาสตร(5-7) ชแนะวา คาเรยกเผอก มน กลวย สาเก [breadfruit--Artocarpus altilis (Parkinson) Fosberg] มะพราว ออย และคาเรยกเครองมอเพาะปลก รวมทงสตวเลยง เชน ควาย หม และไก เปนคาเกาแก ซงมใชกนแพร หลายมานานแลวในกลมคนทใชภาษากลมออสโตร-ไต (Austro-Tai) ซงเปนตนกาเนดของภาษาไทย ลาว และออสโตรนเชยน ไดแก ภาษามาเลย อนโดนเซย และภาษาของชาวเกาะในมหาสมทรแปซฟก โดยเฉพาะกลวยและสวนตางๆ ของตนกลวย มคาเรยกในภาษาตางๆ หลายภาษา เชน ภาษายวย อนโดนเซย ธเบโต-พมา เขมร และกะเหรยง ฯลฯ
อนกรมวธานของกลวย
พชในวงศกลวย (Musaceae) อยในอนดบขง ขา (Zingiberales) ซงเปนอนดบของพชใบเลยงเดยวทม
จานวนสมาชกกวา 2,500 ชนด อนดบนมวงศสาคญอนอก 7 วงศ ไดแก วงศกลวยพด (Steritziaceae) ธรรมรกษา (Heliconiaceae) ขง ขา (Zingiberaceae) เอองหมายนา (Costaceae) พทธรกษา (Cannaceae) คลา (Marantheceae) และวานดอกเหมน (Lowiaceae) ทงนหลกฐานทางชวโม- เลกลระบวาวงศกลวยเปนวงศทดกดาบรรพทสดในอนดบ
เดยวกน(4)
พชในวงศกลวยจาแนกไดเปน 3 สกล คอ สกลกลวย (Musa) สกลกลวยโทน (Ensete) และสกลกลวยดารารศม (Musella) พชในสกลกลวยและสกลกลวยดารา-รศมเปนพชลมลก อายหลายป สวนสกลกลวยโทนมอายเพยงปเดยว มลาตนใตดนเรยกวาเหงา [ลกษณะเปน rhizome หรอ rootstock หรอบางชนดเปนหว (corm)] เหนอดนเปนลาตนเทยม (pseudostem) อวบนา ใบเรยงเวยน กานใบเปนรอง เสนใบขนานแบบขนนก ชอดอก เรยกวาปลหรอหวปล (เมอเจรญเปนผลแลวเรยกวาเครอ) เจรญจากลาตนใตดนผานแกนกลางลาตนเทยมขนมา ชอดอกประกอบดวยดอกทออกเปนกลมเรยงเวยนบนแกนชอ
ดอก แตละกลมมใบประดบ 1 ใบรองรบ ดอกยอยแตละกลม (เมอเจรญเปนผล เรยกวาหว) เรยงเปนแถว 1-2 แถว แถวละ 3-10 ดอก กลมดอกทเกดกอนใกลกานชอดอกเปนดอกเพศเมยหรอดอกสมบรณเพศ ปลายชอดอกเปนกลมกลมดอกเพศผ ดอกมรงไขใตวงกลบ กลบรวม (compound

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
165
tepal) สครม อาจมแตมสชมพ หรอมวงแดง ปลายกลบรวมแยกเปน 5 กลบ กลบแยก (free tepal) สครมโปรงใส เกสรเพศเมย 1 เกสรเพศผ 5 ผลกลวยปามเมลดประมาณ 60-200 เมลด ในพนธปลกกนผลเปนผลลม (partheno-carpic fruit) ไมมเมลดหรอมเมลดนอย 1-5 เมลดเทานน รปท 1 แสดงปลกลวยตานและผลกลวยปาซงมเมลดจานวนมาก
รปท 1 ปลกลวยตาน (ซาย) ปลและผลกลวยปา (กลางและขวา)
ทรพยากรพนธกรรมกลวยแหงสวรรณภม
การสารวจและวจยกลวยโดยอาจารยและนกศกษา
ร ะด บป ร ญญ าต ร แ ล ะ โทขอ งคณะว ท ย า ศ าสตร มหาวทยาลยมหดล ตงแตป พ.ศ. 2548 เปนตนมาจนถง
ปจจบน พบกลวยปาในปาธรรมชาตในประเทศไทย 2 สกล รวม 10 ชนด (species) 4 ชนดยอย (subspecies) และกลวยประดบนาเขา 4 ชนด และ 1 ชนดยอย (ตารางท 1) และกลวยปลกกนผลซงไมมเมลดมประมาณ 100 พนธ
ปลก (cultivar) ลกษณะของกลวยบางชนดแสดงในรปท 2
รปท 2 แถวบน: กลวยดารารศม (ซาย) กลวยนวล (กลาง) กลวยผา (ขวา) แถวลาง: กลวยบวสชมพ (ซาย) กลวยบวสสม (ขวา)
กลวยพนธปลกทไมมเมลดเกอบทงหมดเปนกลวย
ลกผสม หรอกลายจากกลวยปา 2 ชนดในสกลกลวย ไดแกกลวยปาหรอกลวยแข (M. acuminata Colla) และกลวยตาน (M. balbisiana Colla) กลวยทงสองชนดมโครโม- โซม 2 ชด (diploid: 2n=2x=22) ในขณะทกลวยปลกซงไมมเมลดสวนใหญมโครโมโซม 3 ชด (triploid: 2n=3x=33) ทงนพนธปลกบางพนธมลกษณะคลายกลวยปา บางพนธมลกษณะคลายกลวยตาน เมอนกวชาการ(9,19) ใหคะแนนลกษณะสณฐาน 15 ลกษณะของพนธปลก และกาหนดใหพนธทไดคะแนนตาเปนพนธกลม A (acuminata) พนธทไดคะแนนสงเปนกลม B (balbisiana) ประกอบกบการนบจานวนโครโมโซมและการทดสอบดวยโมเลกลคลอโรพ
ลาสต(21) ทาใหสามารถจาแนกพนธปลกในประเทศไทยไดเปนกลม ดงน
กลมกลวยไข (AA) เชน กลวยเลบมอนาง กลวยไข กลมกลวยหอม (AAA) เชน กลวยหอม กลวยนาก กลม AAB เชน กลมแพลนเทน (plantain หรอ
cooking banana) ไดแก กลวยกลาย กลวยงาชาง และกลมกลวยเปรยวหรอไมซอร (mysore-เรยกตามชอเมองในรฐทางตะวนตกเฉยงใตของประเทศอน เด ยทมความ
หลากหลายของกลวยกลมน) เชน กลวยจน กลวยทองสม กลม ABB/BBA เชน กลมกลวยหกมก เชน กลวย
หกมกสวน หกมกสม หกมกนวล หกมกเขยว กลวยนมหม กลมกลวยนาวา ซงเปนกลมทพบความหลากหลายมากท สดในประเทศไทย(21) ทาใหสนนษฐานวานาจะมกาเนดในประเทศไทย เชน กลวยนาวาสวน นาวาคอม นาวาไสดา นาวาไสเหลอง นาวานวล เปนตน และกลมกลวยเทพรส-ทพรส ซงเดมเขาใจวาเปนกลวยทมโครโมโซม 4 ชดแบบ BBBA(9) แตในภายหลงเมอใชเทคโนโลยวเคราะหขนาดจโนมและปรบวธการตรวจสอบจานวนโครโมโซม ทาใหพบวากลวยในกลมนมโครโมโซม 3 ชด เท านน (13 ) นอกจากกลวยเทพรส-ทพรส ตอมาไดพบอก 1 พนธปลกทมลกษณะคลายกนคอ กลวยชาง
กลมกลวยเลบชางกด (BBB) พบไดนอยมากในโลก เชน กลวยเลบชางกดในประเทศไทย และกลวยซาบา (Saba) ในประเทศฟลปปนส
ทงนนกวชาการนยมระบกลมจโนมประกอบกบชอ
พนธทเปนชอทองถน เชน กลวยนางพญา เขยนวา Musa (ABB) ‘Nang Phaya’ และกลวยนวนางรา คอ Musa (AA) ‘Nio Nang Ram’ เปนตน
การจาแนกกลมจโนมกลวยมากกวา 100 พนธปลกทเกบรวบรวมจากทวประเทศไทย ดวยลกษณะสณฐานวทยาและวธการวเคราะหขนาดจโนมดวยเครองมอ flow

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
166
cytometer(3) ไดตวอยางกลวยดงในตารางท 2 และลกษณะผลและ/หรอเครอของกลวยพนธปลกบางชนดในประเทศไทยแสดงในรปท 3 และ 4
กลวย...ทรพยคาอนนต กลวยนบเปนอาหารหลกของประชากรทยากจน
กวา 400 ลานคนทวโลก ในทวปแอฟรกา คนทองถนนยมปลกกลวยกลมแพลนเทนซงมผลขนาดใหญและมแปง
มากโดยตมรบประทานสกแบบเดยวกบการรบประทาน
หวเผอก หวมน และมแพลนเทนบางพนธทเหมาะนามาหมกทาเบยร(23)
ตารางท 1 กลวยทพบในประเทศไทย
# ชอสกล ชอพฤกษศาสตร ชอไทย ชออน สถานภาพ
1.1 Musa M. acuminata Colla subsp. malaccensis (Ridl.) Simmonds
กลวยปา กลวยเถอน พบในภาคใต
1.2 Musa M. acuminata Colla subsp. microcarpa (Becc.) Simmonds
กลวยปา - พบใน จ. ประจวบครขนธ
1.3 Musa M. acuminata Colla subsp. siamea Simmonds
กลวยปา กลวยแข พบทกภาค ยกเวนภาคใต
1.4 Musa M. acuminata Colla subsp. truncata (Ridl.) Kiew
กลวยปา - นาจะเปนการรายงานครงแรกในประเทศไทย
1.5 Musa M. acuminata Colla subsp. zebrina (Van Houtte ex Planch.) Simmonds
กลวยทหารพราน กลวยเสอพราน, กลวยเลอด ไมประดบนาเขาจากประเทศอนโดนเซย
2 Musa M. balbisiana Colla กลวยตาน กลวยพองลา, กลวยพงลา (ใต), กลวยกบมา, ตานใน, ชะนใน, เมลด (สรนทร), ง (พจตร)
พบนอยมากในธรรมชาต แตปลกแพรหลายทวประเทศ
3 Musa M. itinerans Cheesman กลวยหก กลวยไหล, กลวยแดง พบในภาคเหนอ
4 Musa M. ornata Roxb. กลวยบวสชมพ กลวยบว กลวยบวชมพ (กลาง)
ไมประดบนาเขาจากประเทศอนเดย
5 Musa M. laterita Cheesman กลวยบวสสม กลวยบวสม, กลวยบว (กรงเทพ), กลวยแข (เหนอ), กลวยแวก, ยะมอง, ยะโม (ปกากะญอ แมฮองสอน)
พบในภาคเหนอและภาคตะวนตก
6 Musa M. velutina H. Wendl. & Drude กลวยรงอรณ - ไมประดบนาเขาจากประเทศอนเดย
7 Musa M. gracilis Holttum กลวยศรนรา ปซงกะแต, ปซงเวก, ปซงโอนก (นราธวาส)
พบใน จ. นราธวาส
8 Musa M. coccinea Andrews รตกทล กทล ไมประดบนาเขาจากประเทศอนเดย-จน
9 Musa Musa sp. 1 กลวยลานนา - นาจะเปนชนดใหม
10 Musa Musa sp. 2 กลวยบวสกหลาบ - นาจะเปนชนดใหม
11 Ensete E. glaucum (Roxb.) Cheesman กลวยนวล กลวยญวน, กลวยโทน (นาน), กลวยหวโต (กรงเทพ), กลวยศาสนา (เชยงใหม), อะแพละ (ปกากะญอ แมฮองสอน)
พบในภาคเหนอและภาคตะวนตก ปลกแพรหลายในภาคตะวนออกเฉยงเหนอ
12 Ensete E. superbum (Roxb.) Cheesman กลวยผา กลวยครก พบบนภเขา-หนาผา ทกภาค
13 Musella Musella lasiocarpa (Franch.) H. W. Li
กลวยดารารศม กลวยคนหมง (กรงเทพ) ไมประดบนาเขาจากประเทศจน

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
167
กลวยเปนพชสารพดประโยชน ผลกลวยแตละพนธ มรสชาต กลน ลกษณะเนอและแปง รวมทงคณคาทาง โภชนาการแตกตางกน ซงนอกจากรบประทานผล สกแลว คนไทยยงมกรรมวธในการแปรรปดวยการ ตม ปง ทอด กวน เชอม ตากแหง อบแหง ทาเปนผง หนเปนแผนแลวทอดเปนกลวยฉาบ บดเปนอาหารเหลวสาหรบเดก หรอผสมในขนมอบ ปน กรองเปนนา ทาเปนนาหวานเขมขน และไวน สาหรบหยวก ปลและผลออนนน คนในเอเชยตะวนออกเฉยงใตนยมนามารบประทานเปนผก และใชเปนอาหารสตว ใบกลวยตานและกลวยนาวาใชสาหรบหอของ หอขนม แหนม หมยอ และใชทางานประดษฐพวกบายศร สาหรบเสนใยจากกาบกลวยยงอาจใชทากระดาษและเชอก ทงนในฟลปปนสและเอควาดอร มการผลตเสนใยปานมะนลา (abaca หรอ manila hemp) จากกาบกลวย อะบาคา (M. textilis Nee: จานวนโครโมโซม 2n=2x=20) ซงมคณภาพดมากเหมาะสาหรบใชทากระดาษทชช เทป สายเคเบลโทรศพท ฉนวนลวด กระดาษกรอง ถงชากาแฟ ปลาสเตอรปดแผล ฟลมเอกซเรย กระดาษ พมพธนบตร เชอกสมอเรอ แผนวสดเพอใชบฝาผนงและเพดาน(12)
สาหรบในการใชกลวยเปนยานน ปรากฏหลกฐาน
เรองการใชสวนตางๆ ของกลวยเปนยารกษาโรคและอาการตางๆ(2) รวมทงการใชยางกลวยรกษาอาการทเกดจากพษง(8) นอกจากนนยงมการปลกกลวยหลายชนดเปนไมประดบเพอการคา โดยเฉพาะกลวยบวสชมพ กลวยบวสสม และรตกทล รวมทงกลวยแคระ กลวยทมใบ-ผลสแดงหรอแตมแดง เชน กลวยนาก กลวยทหารพราน และทมใบดาง เชน กลวยนาวาดาง เปนตน
ในป พ.ศ. 2549 องคการอาหารและเกษตรแหง
สหประชาชาต (FAO) แสดงความกงวลวากลวยจะสญ
พนธ โดยเฉพาะในอนเดยซงเปนแหลงพนธกรรมสาคญของกลมกลวยหอมและกลมกลวยกลาย มผลผลตกลวยลดลงอยางเหนไดชดและกลวยปาซงเปนพอแมของกลวย
เหลาน กลายเปนพชหายากจากการแผวถางทาลายปาธรรมชาต ทงนแมวาจะมโครงการปรบปรงพนธกลวยในทวปแอฟรกา อเมรกากลาง และอเมรกาใต แตพอแมพนธมาจากสายตน (clone) เพยง 1-2 สายตนเทานน ทาใหรนลกทไดอาจมความออนแอตอโรค ดงทเคยเกดการระบาดของ Panama disease ในกลมกลวยหอม ‘Gros Michel’ และเกษตรกรในทวปอเมรกากลางและใตซงเปนผผลต
สาคญตองสญเสยผลผลตกลวยไปเปนจานวนมาก16)
ตารางท 2 ตวอยางกลวยพนธปลกในประเทศไทย
กลมท กลมจโนม ชอกลมพนธ ตวอยางชอพนธปลก
1 AA กลวยไข กรน, ไข, ไขสวนผง, ทองขแมว, เนอทอง, สาวกระทบหอ
กลวยเลบมอนาง ไขทองเงย, เลบมอนาง, สา, หอมจนทน, หอมจาปา
กลวยอนๆ แซลอ, ทองดอกหมาก, นาหมาก, นวนางรา, แสมา
2 AAA กลวยหอม เขยวปากชอง, ไช, มน, หอมกะเหรยง, หอมเขยวคอม, หอมเขยวตนสง, หอมคอม, หอมทอง, หอมโปรย, หอมศรสะเกษ
กลวยนาก กงเขยว, นาก, นากกะเหรยง, นากคอม, นากทองผาภม
กลวยหอมทพย ไขทองรวง, ไขหอม, ตานวล, ตนเตา, ทองแขก, สายนาผง, หอมทองปา, หอมทพย, หอมผลสน
3 AAB กลวยกลาย กลาย, กาไน, งาชาง, จ, ตะโหลน
กลวยเปรยว กอกหมาก, ไขศรสะเกษ, จน, ทองสม, ลงกานครสวรรค
กลวยหวาน กรบร, ขม, ไขดา, ไขโบราณ, นมนาง, นมสาว, นมสาวตะนาวศร, รอยหว
กลวยนา นา, นาเชยงราย, นาไทย, นานม, นาฝาด
4 ABB/BBA กลวยตบ-หกมก ไขมหาสารคาม, ตบ, ตบคา, นมหม, พมาแหกคก, สมมง, หกมกเขยว, หกมกทอง, หกมกนวล, หกมกสม
กลวยนาวา
นาวากาบขาว, นาวาเขยว, นาวาคอม, นาวาดา, นาวาตะนาวศร, นาวานวล, นาวามะลออง, นาวาลกไสดา, นาวาสวน, นาวาไสแดง, นาวาไสเหลอง
กลวยเทพรส ชาง, เทพรส-ทพรส
กลวยอนๆ นางกราย, นางพญา, นวมอนาง, สามเดอน, หน
5 BBB กลวยเลบชางกด เลบชางกด

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
168
รปท 3 (บน) แถว 1: กลวยหกมก (ABB; ซาย) กลวยหอม (AAA; ขวา)
แถว 2: กลวยนานม (AAB; ซาย) กลวยเทพรส (BBA; ขวา) แถว 3: กลวยกลาย (AAB; ซาย) กลวยนาฝาด (AAB; ขวา)
แถว 4: กลวยกาไน (AAB; ซาย) กลวยเลบชางกด (BBB; ขวา)
รปท 4 (ขวา) กลวยเลบมอนาง (AA) จ. ชมพร (บน) กลวยไข (AA) จ. กาแพงเพชร (กลาง) กลวยนาวาไสเหลอง (BBA; ลางซาย) กลวยหกมกนวล (ABB; ลางขวา)
หนทางเดยวทจะรกษาความมนคงทางอาหารและ
เกษตรของประชากรทจาเปนตองพงพากลวยเปนอาหาร
และเปนรายไดสาคญของครอบครว คอ การนาพนธกรรมทมคณคา เชน การทนแลง ทนนาทวม ทนเคม รวมทงทนทานตอโรคตางๆ มาใชในการปรบปรงพนธ และหนทางส เป าหมายนจะตองเรมตนทการสารวจเชอ
พนธกรรมในแหลงกาเนดดงเดมทยงมความหลากหลายสง
อย างกว างขวางครอบคลม โดยไมปลอยให พนททเปราะบางตองถกทาลายไปกอนทเราจะคนพบอญมณมคา
อนนต...ทรพยแหงสวรรณภม
หมายเหต: เนอหาในบทความน ปรบปรงเพมเตมจากศศวมล แสวงผล และคณะ(3) (สวนบทนา)
กตตกรรมประกาศ
ขอขอบคณคณะส ารวจและว จ ยกล วย คณะ
วทยาศาสตร มหาวทยาลยมหดล น.ส.กนกพร อะทะวงษา นายสรภพ วงษเนยม นายกาสพงษ กจดารงสนต นาย พงศกร โพธถาวร น.ส.วลาวณย ชาวหงษา น.ส.พรประวณ โชคสวรรณเลศ น.ส.หยาดนภา เชยโต นายณรงคศกด สกแกวมณ และ น.ส.พจนา เกยรตประไพ
เอกสารอางอง 1. กรมสงเสรมการเกษตร. ปรมาณและมลคาการสงออก
ผลไมป พ.ศ. 2546 - 2550 [ขอมลในอนเตอรเนต]. กรงเทพ: กรมสงเสรมการเกษตร; 2551 [อางองวนท 17 ธนวาคม 2551]. จาก: URL: www.agriinfo.doae. go.th

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
169
2. มหาวทยาล ย เกษตรศาสตร . เ ร องกลวย กลวย [บทความในอนเตอรเนต]. กรงเทพ: มหาวทยาลย เกษตรศาสตร; 2551 [อางองวนท 17 ธนวาคม 2551]. จาก: URL: http://ku-iged.project.ku.ac.th/032/ banana/drug.htm
3. ศศวมล แสวงผล จามร สมณะ สมรรถชย ฉตราคม. 108 พนธกลวยไทย. กรงเทพ: มลนธสวนสมเดจพระนางเจาสรกต พระบรมราชนนาถ; 2552.
4. Andersson L. Musaceae. In: Kramer KU, Kubitzki K, Hxber H, Green PS, editors. The families and genera of vascular plants. Vol. 4. Springer; 1990. p. 296-301.
5. Bellwood P. The Austronesian dispersal and the origin of languages. Sci Am 1991;265:88-93.
6. Benedict PK. Austro-Thai language and culture, with a glossary of roots. New Haven: HRAF Press; 1975.
7. Robert B. The prehistory of the Austronesian-speaking peoples: a view from language. J World Prehist 1995;9:453-10.
8. Borges MH, Alves DLF, Raslan DS, Piló-Veloso D, Rodrigues VM, Homsi-Brandeburgo MI, de Limaa ME. Neutralizing properties of Musa paradisiaca L. (Musaceae) juice on phospholipase A2, myotoxic, hemorrhagic and lethal activities of crotalidae venoms. J Ethnopharmacol 2005;98:21-9.
9. Chomchalow N; Silayoi B. Banana germplasm in Thailand. IBPGR/SEAP Newsletter 1984;8:23-8.
10. De Langhe E, De Maret P. Tracking the bananas: its significance in early agriculture. In: Gosden C, Hather J, editors.). The prehistory of food. London: Routledge; 1999. p. 377-96.
11. Gray RD, Jordan FM. Language trees support the express-train sequence of Austronesian expansion. Nature 2000;405:1052-5.
12. Gonzal LR, Capuno OB. Musa textilis Nee in PROSEA: A Selection. In: Westphal E, Jansen, PCM, editors. Prosea: Bogor; 1993.
13. Horry J-P, Dolezel J, Dolezelova M, Lysak MA. Do natural AxB tetraploid bananas exist? Infomusa 1999;7:5-6.
14. INIBAP Global Conservation Strategy for Musa (Banana and Plantain). Belgium: International Network for the Improvement of Banana and Plantain; 2006.
15. INIBAP. International collection [database on the internet]. Belgium: The International Network for the Improvement of Banana and Plantain [cited 2008 Dec 17]. Available from: URL: http://bananas. bioversityinternational.org/content/view/12/31/lang, en/
16. Matthews C. Concern at vanishing bananas, Newsroom [article on the internet]. Rome: The Food
and Agriculture Organization (FAO); 2006 May 3 [cited 2008 Dec 17]. Available from: URL: www.fao.org/newsroom/en/news/2006/1000285/ index.html
17. Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J. Biodiversity hotspots for conservation priorities. Nature 2000;403:853-8.
18. Oppenheimer S. Eden in the east: the drowned continent of Southeast Asia. Great Britain: Weidenfeld & Nicolson; 1998.
19. Simmonds NW, Shepherd, K. The taxonomy and origins of the cultivated bananas. J Linn Soc Bot 1955;55:302-12.
20. Simmonds NW. Botanical results of the banana collecting expedition, 1954-5. Kew Bull 1956;3:463-89.
21. Swangpol S, Volkaert V, Sotto RC, Seelanan T. Utility of selected non-coding chloroplast DNA sequences for lineage assessment of Musa interspecific hybrids. J Biochem Mol Biol 2007;40: 577-87.
22. Tougard C. Biogeography and migration routes of large mammal faunas in South-East Asia during the late middle Pleistocene: focus on the fossil and extant faunas from Thailand. Palaeogeogr Palaeocl 2001; 168:337-58.
23. Verheij EWM, Coronel RE. Musa. In: Verheij EWM, Coronel RE, editorss. PROSEA No. 2: Edible fruits and nuts. Prosea: Bogor; 1992.


171
เทคโนโลยชวภาพพชเพอการปรบปรงพนธยางพารา
จรญญา ณรงคะชวนะ ภาควชาเทคโนโลยชวภาพ คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
ยางพาราเปนพชเศรษฐกจหลกทสาคญมากในประเทศไทย นารายไดเขาสประเทศประมาณ 3.5 แสนลานบาท/ป โดยทความตองการใชยางธรรมชาตในอตสาหกรรมตางๆ ทวโลกมปรมาณสงขนทกป ในขณะทพนทปลกมอยจากด ดงนนงานวจยเพอเพมผลผลตนายาง ตลอดจนลดปญหาททาใหผลผลตนายางตกตาจงมความจาเปนมาก สาหรบประเทศไทย
ปจจบนไดมการขยายพนทปลกยางพาราจากภาคใตไปทางภาคตะวนออก ภาคตะวนออกเฉยงเหนอ และภาคเหนอ ซงเปนพนทใหมคอนขางแหงแลงสาหรบการปลกยางพารา จงมความจาเปนเรงดวนในการคดเลอกและปรบปรงสายพนธยางท
สามารถปรบตวใหผลผลตสงในแตละพนท ปจจบนเทคโนโลยชวภาพพชมความกาวหนารวดเรวมาก กอใหเกดประโยชนทงในดานการพฒนาพชพนธดตลอดจนการพฒนาผลตภณฑจากพชมากมาย เทคโนโลยทเกยวของทงดานการเพาะเลยง
เนอเยอพช การพฒนาเครองหมายโมเลกลเพอการปรบปรงพนธ การคนหายน และการทาพนธวศวกรรมพชสามารถนามาประยกตใชในการปฏรปการผลตยางธรรมชาตทงในระยะสน เพอชวยรนระยะเวลา แรงงาน และเพมความแมนยาใน
ขนตอนการคดเลอกพนธยางใหม รวมทงใชพฒนาเครองหมายโมเลกลสาหรบการเฝาตรวจระวง และจดการระบบเพอใหไดผลผลตสมาเสมอและคณภาพดในปรมาณสงอยางมประสทธภาพ และในระยะยาว การคนหายนทเปนประโยชนตอการปรบปรงพนธยางจะสามารถนาไปประยกตใชดดแปลงพนธกรรมตนยางพาราเพอการปรบปรงพนธใหมคณสมบตตรงตาม
ความตองการ และ/หรอจดการใหตนยางพาราสรางผลตภณฑใหมได
คาสาคญ: ยางพารา; เทคโนโลยชวภาพพช; การเพาะเลยงเนอเยอพช; เครองหมายโมเลกลเพอการปรบปรงพนธ; การคนหายน; พนธวศวกรรม
Abstract
Rubber tree is one of the major cash crops in Thailand and due to a continuous increase of the world industrial utilization of natural latex and its products, the future of rubber tree looks very promising. Thailand earns about 350,000 millions bath annually from rubber export. Rubber tree plantation area has expanded from the traditional southern to the more marginal northeast and north areas. Thus, selection of rubber clones with high yield potential, as well as stress-tolerance, and of suitable growth conditions for the various plantation areas are needed. Plant biotechnology is a rapidly expanding field within modern biotechnology and has resulted in crop improvement and production of novel products through plant molecular farming. Such approaches as plant tissue culture, molecular marker-assisted breeding, gene tagging, and genetic engineering will contribute to solve production problems so as to ensure the country's position as the world's leading producer and exporter of natural rubber. Marker-assisted selection plays an important role in selection of genotypes, which will greatly increase efficiency and effectiveness compared to conventional breeding methods. Rubber clones suitable for each plantation area will help increase natural rubber latex production and its market value, which would have a high financial benefit for all involved, both at agricultural and end-user levels. In addition, emphasis also is being directed towards genetic modification of latex for the production of commercially valuable proteins. As one of the main suppliers of natural rubber latex and products, Thailand stands to benefit the most from basic and applied researches in rubber tree. Keywords: rubber tree; plant biotechnology; plant tissue culture; molecular marker assisted breeding;
gene tagging; genetic engineering

วทยาศาสตรเพอการพฒนา: รวทบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
172
บทนา
ตนยางพาราจดอยใน Family Euphorbiaceae, Genus Hevea, Species brasiliensis แมใน Genus Hevea มตนยางมากกวา 20 ชนด แตยางพารา (Hevea brasiliensis) เปนชนดทใหนายางมากทสด และเนอยางมคณสมบตในการนาไปใชประโยชนดกวายางทไดจาก Hevea ชนด อน ๆ จงมการปลกตนยางพาราใชประโยชนอยาง
แพรหลาย ในป พ.ศ. 2550 สานกงานเศรษฐกจการเกษตร ประเทศไทย รายงานวา พนทปลกยางธรรมชาตทงโลกมประมาณ 63 ลานไร ใน 24 ประเทศ โดยประเทศ
อนโดนเซยมพนทปลกยางมากทสด รองลงมาคอ ประเทศไทย และมาเลเซย รวมพนทปลกยางทง 3 ประเทศ เปนรอยละ 73 ของพนทปลกยางของโลก(9) นบตงแตป พ.ศ. 2534 เปนตนมาจนถงปจจบน ประเทศไทยไดกาวขนเปนผผลตและสงออกยางพาราเปนอนดบ 1 ของโลก การทปจจบนประเทศไทยมปรมาณการผลตยางธรรมชาตสงถง 3 ลานตนตอป เปนผลสบเนองจากปจจยแรกคอ การขยายพนทปลกยางเพมขน จากแหลงปลกยางเดมใน 14 จงหวดภาคใต และใน 4 จงหวดภาคตะวนออก ไปยงแหลงปลกยางใหมในภาคตะวนออก ภาคกลาง ภาคเหนอ และภาคตะวนออกเฉยงเหนอ และปจจยทสองซงมบทบาทสาคญมาก คอ การเพมผลผลตตอหนวยพนทโดยการปลกทดแทนดวยยางพนธด ประเทศไทยสามารถทารายไดจากการสงออกในรปยางแผนดบ ผลตภณฑยางแปรรป และผลตภณฑจากไมยางพารา คดเปนมลคาทงสนกวา 3.5 แสนลานบาทตอป ยางพาราจงเปนพชทมความสาคญทางเศรษฐกจของประเทศไทยในอนดบตนๆ อกทงยงมความสาคญตอชวตความเปนอยของชาวสวนยางซงคด
เปนรอยละ 10 ของจานวนประชากรทงประเทศ ปจจบนรฐบาลไดใหการสงเสรมการปลกยางพาราในทกภาคของ
ประเทศ ยางพาราจงเปนพชเศรษฐกจท เกษตรกรทวประเทศไทยใหความสนใจตองการปลกเพมขน นบเปน
พชทมอนาคตสดใส อกทงทวโลกยงมความตองการใชยางธรรมชาตอยเปนจานวนมาก คาดการณวาปรมาณการใชยางธรรมชาตของโลกในอก 7 ปขางหนา จะสงขนถง 10.6 ลานตน กอปรกบในปจจบนโลกไดเขาสยคขาดแคลนนามน ความสามารถในการผลตยางสงเคราะหททามาจากนามนจงมแนวโนมลดนอยลง ในขณะทยางธรรมชาตทผลตไดจากพชจะมความสาคญเพมขน เพอเปนการรกษาสถานภาพและจดเดนความเขมแขงดานการผลตยางพารา
ของประเทศไทย จงตองมงเนนการเพมผลผลตตอหนวยพนท โดยการสงเสรมเรองการจดการเลอกใชพนธยางให
เหมาะสมกบพนทปลกทวทกภาคของประเทศ วทยาการดานเทคโนโลยชวภาพพชไดเขามามบทบาทอยางมาก บทความนเปนการรวบรวมความรและงานวจยทเกยวของ
ในการนาเทคโนโลยชวภาพพชมาประยกตใชชวยในการ
ปฏรปการผลตยางธรรมชาต ในระยะสนใชในการพฒนาเครองหมายโมเลกลทเกยวของกบลกษณะพงประสงคของ
ตนยาง เพอชวยรนระยะเวลา แรงงาน และเพมความแมนยาในขนตอนการคดเลอกพนธยางใหมทมคณสมบต
ตามตองการในโปรแกรมการปรบปรงพนธยางทดาเนนอย รวมทงใชพฒนาโมเลกลเครองหมายสาหรบการเฝาตรวจ
ระวง และจดการระบบการปลก การกรด การดแลตนยางเพอใหไดผลผลตสมาเสมอ และสงตามเปาหมายอยางมประสทธภาพ นอกจากนน สามารถใชระบ จาแนกพนธเพอการอนรกษพนธ เพอจดการเรองตนพนธสาหรบการปลกและจดการทางการคาไดเปนอยางด สวนในระยะยาวการคนหายนทเปนประโยชนตอการปรบปรงพนธยาง ตลอดจนการศกษาใหเขาใจองคความรพนฐานทเกยวของ
กบตนยางพาราในทกดาน จะสามารถนาไปประยกตใชดดแปลงพนธกรรมตนยางพาราเพอการปรบปรงพนธใหม
คณสมบตตรงตามความตองการ และ/หรอจดการใหตนยางพาราสรางผลตภณฑใหมไดอยางมประสทธภาพ
การทาสวนยางพาราในประเทศไทย การทาสวนยางในประเทศไทยเรมจากภาคใต
พฒนาการของการทาสวนยางในประเทศไทยแบงไดตาม
การปรบตวของระบบการทาสวน การตลาด และ การอตสาหกรรม สามารถจดแบงออกไดเปน 5 ชวงเวลา คอ ยคระบบการผลตแบบดงเดม หรอ ปายาง ตงแตเรมปลกยางพาราในประเทศไทย จนถง พ.ศ. 2502 สวนมากใชพนธซงมตนกาเนดจากประเทศอนโดนเซย ยคนเปนการปลกยางแทนปาไมโดยการโคนลมพชพนธเกา แลวทาการปลกยางโดยตนกลาทเพาะจากเมลด ปลอยใหยาง
เจรญเตบโตแขงกบพชชนดอน ยคปฏวตเขยว หรอ สวนยางพนธด ในชวงป พ.ศ. 2503-2512 เมอยางถกจดเปนพชเศรษฐกจทสาคญ ทใหผลตอบแทนทางเศรษฐกจไดอยางเปนรปธรรมแกเกษตรกร กอปรกบมการพฒนาวทยา
การเกษตรแผนใหมภายใตการสนบสนนของรฐผานทาง
สถาบนวจยยาง และสานกงานกองทนสงเคราะหการทา
สวนยาง เกดการทาสวนยางพนธดขนโดยการปลกจาก
เมลดยางทผานการคดเลอกแลววาใหผลผลตด เรมใชยางพนธใหมทใหผลผลตสง ยคระบบการผลตยางสมยใหม ป พ.ศ. 2513-2522 มการขยายเนอทปลกยางอยางตอเนอง และเกษตรกรเรมมการรวมกลมกนขายยางเพอสราง

วทยาศาสตรเพอการพฒนา: รวทบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
173
อานาจตอรองดานการตลาด ยคระบบการผลตยางพาราทางเลอก ในชวงปพ.ศ. 2523-2532 มวชาการดานการ
ปรบปรงพนธยางเพอใหผลผลตสงขน เรมมการขยายพนธโดยการตดตาเพอปองกนการกลายพนธ มการดาเนนการสงเสรมการปลกโดยกองทนสงเคราะหการทาสวนยาง เกดการโคนลมปายาง ปลกทดแทนยางเกาดวยยางพนธด มสวนยางสงเคราะหภายใตการสนบสนนปจจยการผลต
จากกองทนสงเคราะหการทาสวนยาง ยคระบบการผลตเชงอตสาหกรรม ยคนเรมตงแตป พ.ศ. 2533 ถงปจจบน สวนยางสวนมากจะเปนสวนยางสงเคราะห มการสงเสรมการปลกยางในภาคอนๆ ของประเทศ ยคนรฐบาลมงเนนใหเกดอตสาหกรรมยางพาราในประเทศมากขนเพอสราง
มลคาเพมในประเทศ นบตงแตป พ.ศ. 2534 เปนตนมา
จนถงปจจบน ประเทศไทยไดกาวขนเปนผผลตและ
สงออกยางพาราเปนอนดบ 1 ของโลก มปรมาณการผลต การสงออกและการใชในประเทศเพมมากขนทกป จากการสารวจในป พ.ศ. 2550 ประเทศไทยมพนทปลกยางรวมประมาณ 15 ลานไร ในจานวนดงกลาวมพนททตนยางสามารถกรดไดแลวประมาณ 11 ลานไร ปรมาณการผลตยางของไทยมทงสนประมาณ 3 ลานตนตอป คดเปนสดสวนรอยละ 34 ของผลผลตทงหมดของโลก ทารายไดใหประเทศคดเปนมลคาทงสนกวา 3.5 แสนลานบาทตอป
การปรบปรงพนธยางพารา ยางพาราไดถกนาเขามาปลกในทวปเอเชยครงแรก
ในราว ป พ.ศ. 2419 โดยเปนเมลดยางท Henry Wickham รวบรวมมาจากรมฝงแมนา Tapajos ในมลรฐ Para ของประเทศบราซล ซงเปนแหลงของยางชนดทใหผลผลตดทสด ตอมามผลการศกษาของบรษทฟอรดมอเตอรยนยนวา พนธยางพาราทถกนามานมผลผลตสงและคณภาพ
ดกวาพนธอนๆ การปรบปรงพนธยางในแถบเอเชยดาเนนไปอยางตอเนองชาๆ ยางบางพนธนยมปลกกนมานานกวา 30 ปแลว มงานวจยเพอพฒนายางพนธใหมดาเนนการอย แตกวาจะไดพนธใหมทดกวาเดมนนตองใชเวลานานหลาย
สบป ส าหรบประเทศไทยมผลงานส าคญจากการ
ด า เนนงานด านปรบปร งพนธ ย างของนก วชาการ
สถาบนวจยยางของไทยทเหนชดเจน คอ สามารถทาใหผลผลตจากเดม 96 กโลกรม/ไร/ปในป พ.ศ. 2530 เพมขนเปน 218 กโลกรม/ไร/ปในป พ.ศ. 2540 เนองจากปจจบนการเพมพนทปลกจะมขอจากดขนเรอยๆ จงควรมงเนนการเพมผลผลตตอหนวยพนท โดยการสงเสรมเรองการจดการเลอกใชพนธยางใหเหมาะสมกบพนทปลกทวทก
ภาคของประเทศ ชวยลดปญหาทบนทอนผลผลตคอ โรค
หรอแมลงศตรยางพารา ซงเขาทาลายทใบ กงกาน ลาตน หรอราก ขนอยกบชนด ความรนแรงของโรค แหลงปลก พนธยาง และสภาพอากาศในแตละป นอกจากนยงมอ าการ เปลอกแห งของตนยางท ไม ผ ลตน า ย างอน
เนองมาจากการเลอกใชพนธยาง รวมกบสภาพแวดลอมไมเหมาะสม เชน สภาพทดนไมมความอดมสมบรณเพยงพอ ความชนในดนไมเหมาะสม หรอเกดจากระบบการกรด การใชสารเคมไมเหมาะสม และอนๆ จงควรมการสงเสรมเรองการจดการพนธยาง ระบบปลก ระบบกรด และการดแลสวนยางทเหมาะสม เพอใหไดผลผลตสงสมาเสมอ และมคณภาพดสาหรบอตสาหกรรมตอเนอง
สาหรบการปรบปรงพนธยางของประเทศไทย หนวยงานหลก คอ สถาบนวจยยาง กรมวชาการเกษตร กระทรวงเกษตรและสหกรณ ไดแบงการทางานดานการปรบปรงพนธยาง ออกเปน 2 ระยะ คอ ระยะแรก ตงแตป พ.ศ. 2476-2507 สวนใหญเปนการคดเลอกยางพนธพน เมองทมลกษณะดมารวบรวมไว เพอทดลองและคดเลอกพนธดไวสาหรบสงเสรมแนะนาใหชาวสวนยางปลกและ
ใชเปนพอแมพนธสาหรบผสมพนธยางตอไป ระยะท 2 ตงแตป พ.ศ. 2508 จนถงปจจบน เปนชวงทมการปรบปรงพนธยางอยางจรงจง มการคดเลอกพนธ ผสมพนธ และนาความรทางวชาการใหมๆ เขามาใชในการปรบปรงพนธยาง ขณะเดยวกนกมโครงการแลกเปลยนพนธยางกบ
ตางประเทศดวย สาหรบขนตอนในการปรบปรงพนธยาง เพอใหไดยางพนธด ถงมอเกษตรกรนน จะตองผานขนตอนตางๆ เปนเวลายาวนานตามวธการ คอ การผสมพนธยางโดยการนาเกสรตวผจากตนทมลกษณะดเดน
ตามทตองการ ไปผสมกบเกสรดอกตวเมยบนตนแมทม
ลกษณะดเดนเชนกนในสภาพแปลงปลกตามธรรมชาต ซงวธการเปนทนยม เนองจากสามารถทราบประวตของพนธพอ-แม และสามารถตดตามการถายทอดลกษณะตางๆ ในลกผสมไดเมอนาไปปลกทดสอบ การคดเลอกพนธยางทไดจากการผสมพนธจะตองผานหลายขนตอน ทงน
เนองจากยางพาราเปนพชยนตนกวาจะปลกใหไดขอมล
ผลผลตและลกษณะอนๆ ทตองตรวจสอบในแตละขน ตอน ตองใชเวลายาวนาน กลาวคอหลงจากผสมพนธแลว ตองนาเมลดยางลกผสมทไดไปปลกคดเลอกในแปลง
คดเลอกพนธยางซงจะใชเวลาประมาณ 3 ป เพอคดเลอกสายพนธลกผสมทมลกษณะดตามทตองการ สาหรบนาไปปลกในแปลงเปรยบเทยบพนธขนตนซ งใช เวลาอก
ประมาณ 10 ป หลงจากนนจะตองทาการคดเลอกสายพนธจากแปลงเปรยบเทยบพนธขนตน ไปปลกทดสอบใน
แปลงเปรยบเทยบพนธขนปลายในทองทตางๆ โดยใช

วทยาศาสตรเพอการพฒนา: รวทบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
174
เวลาอกประมาณ 13 ป ดงนนกวาจะไดยางพนธแนะนาชนหนงเพอแนะนาใหเกษตรกรปลกโดยทวไปอยางมนใจ ตองใชเวลาดาเนนการประมาณ 30 ป
เนองจากยางพาราทปลกอยในประเทศตางๆ ในแถบเอเชย ไดรบการพฒนามาจากพนธยางจากแหลง กาเนดในประเทศบราซลเพยง 22 ตน ท Henry Wickham นาเขามาเมอป พ.ศ. 2419 จงนบวายางพาราในแถบนมฐานพนธกรรมแคบ ทาใหการปรบปรงพนธเพอเพมผลผลตทาไดยาก หากยางพารามฐานพนธกรรมกวางขนจะทาให
ประสบความสาเรจในการพฒนาพนธทปรบตวไดกบ
สภาพแวดลอมไดดขน ยางสายพนธปามความแปรปรวนทางพนธกรรมสง แมวาสายพนธปาสวนใหญใหผลผลต
นายางตา แตมลกษณะทดอนๆ อกหลายลกษณะทมประโยชนตอแผนการปรบปร งพนธ ย างระยะส น โดยเฉพาะอยางยงมการเจรญเตบโตด แขงแรง ลกษณะลาตนตรง เรยบ แตกกงในระดบสง และตานทานตอโรคทางใบ เหมาะสาหรบใชเปนแหลงพนธกรรมสาหรบการปรบปรงพนธยางเพอผลตเนอไม สวนการปรบปรงพนธเพอเพมผลผลตนายางจาเปนตองใชแผนระยะยาวโดยการ
ปรบปรงประชากรพนธปาใหดขนกอน เพอเปนการขยายฐานพนธกรรมของยางในประเทศไทย สาหรบลกษณะหลกของพนธยางทพงประสงค คอ ใหผลผลตนายางและเนอไมสง สวนลกษณะรอง คอ การเจรญเตบโตเรวและ
สมาเสมอทงกอนกรดและหลงกรด เปลอกลาตนมความ
หนา เปลอกทถกกรดไปแลวมการสรางใหมไดเรว และ
หนาใกลเคยงกบเปลอกเดม มลกษณะโครงสรางของตนทด ลาตนตรง เรยบ แตกกงขนาดปานกลาง-เลก ทรงพมสมดลย ตานทานโรคและแมลง เกดอาการเปลอกแหงนอย ตานทานลมแรง ปรบตวไดดในสภาพแวดลอมตางๆ เชน ทนแลง และอณหภมสง มคณภาพไมทเหมาะสมกบการแปรรปในอตสาหกรรมไม มคณสมบตของนายางทเหมาะ สมกบอตสาหกรรมยาง และความสามารถทจะปลกในพนทลาดชน และ/หรอในพนทหนาดนตน เทคโนโลยชวภาพเพอการปรบปรงพนธพช
เทคโนโลยชวภาพพช เปนเครองมอทใชในการปรบปรงพนธ พช และใชพชใหเกดประโยชน โดยมวตถประสงคหลกเพอเพมปรมาณอาหารและผลตภณฑ
อนๆ ใหเพยงพอกบประชากรทเพมขน ตลอดจนชวยในการรกษาสงแวดลอมอนจะนาไปสสขภาพและความ
เปนอยทดขนของประชากร ในอดตไดมการใชเทคโนโลย ชวภาพพชแบบพนฐานในการปรบปรงพนธพช จนถงยคปฏวตเขยว (green revolution) มการผลตสายพนธพชท
ใหผลผลตสง หลงจากนน จงเขาสยคของการปฏวตยน (gene revolution) เรมรจกยนและใชเทคโนโลยยนในการศกษาวจยในแนวลกมากขน มการศกษาขอมลลาดบเบสของยนตางๆ ตลอดจนทงจโนมของพชและสงมชวตตางๆ รจกการถายโอนยน (gene transfer) เขาสพช จนกระทงในป พ.ศ. 2537 ไดมการผลตและจาหนายพชถายโอนยน (transgenic plant) ชนดแรก และเมอเขาสยคหลงจโนม (post-genomic) การศกษาวจยทาใหมความเขาใจเกยวกบการทางานของยนมากขน ทาใหสามารถนาความรมาใชในการปรบปรงพนธพชใหตรงตามความตองการอยางแมนยา
มากยงขน(6)
เทคโนโลยชวภาพพนฐาน เชน การเพาะเลยงเนอเยอยางพารามประโยชนทงในแงการอนรกษพนธ การสรางตนกลาปลอดโรค การขยายพนธเพอการปรบปรงพนธ การขยายพนธ เดยวในเวลาอนรวดเรวใหไดตนสมาเสมอเพอการเกษตรกรรมและการคา เปนประโยชนรองรบการปรบปรงพนธยางโดยวธการดดแปลงพนธ กรรม และอนๆ สวนเทคโนโลยยนสามารถเขามาชวยในการปฏรปการผลตยางธรรมชาตไดโดย 1) ใชสรางแผนททางพนธกรรมและเครองหมายโมเลกล เพอใชระบ จาแนกพนธ เพอการอนรกษพนธ เพอจดการเรองตนพนธสาหรบการปลก และจดการทางการคา สามารถใชชวยรนระยะเวลา แรงงาน และเพมความแมนยาในขนตอนการคดเลอกพนธยาง ในการปรบปรงพนธยาง อกทงยงใชในการคนหายนทเปนประโยชนตอการปรบปรงพนธ 2) ใชศกษาเพอใหเขาใจองคความรพนฐานทเกยวของกบตน
ยางพาราในทกดาน เพอการประยกตใช 3) ใชพฒนาโมเลกลเครองหมายในการเฝาตรวจระวงและจดการระบบ
การปลก การกรด การดแล ตนยางเพอใหไดผลผลตสมาเสมอ และสงตามเปาหมายอยางมประสทธภาพ 4) ใชดดแปลงพนธกรรมตนยางเพอการปรบปรงพนธใหมคณสมบตตามตองการ และ 5) ใชเพอสรางตนยางพนธใหมทสามารถใชเปนแหลงผลตเวชภณฑหรอสารอนๆ ทมคณคาทางอตสาหกรรม และ/หรอ เกษตรกรรม
การเพาะเลยงเนอเยอยางพารา ปกตฝกยางประกอบดวย 3 เมลด ซงในธรรมชาต
จะตองไดรบการผสมครบทกเมลดกอน จงจะสามารถม
การพฒนาตอเปนเมลดแกได หากไดรบการผสมเพยง 1 หรอ 2 เมลด กจะมการพฒนาตออกระยะสนๆ แลวรวงไป ไมไดเมลดสมบรณในทสด ซงตางจากพชไมผลทวไปท
แมผสมไมครบทกเมลดกสามารถพฒนาเปนเมลดแกได ปจจบนสามารถเพาะเลยงเนอเยอจากลกผสมยางทมอาย

วทยาศาสตรเพอการพฒนา: รวทบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
175
เพยง 3-5 วนหลงจากผสมพนธ โดยเนอเยอจากสวนเมลดออนทเรยกวา คพภะของเมลดยาง มความเหมาะสมทสดในการนามาเพาะเล ยงในหลอดแกว สามารถนามาเพาะเลยงบนอาหารเทยมแลวชกนาใหพฒนาเปนตน
สมบรณได โดยจาก 1 เมลดสามารถเพาะเลยงใหได 1 ตน หรออาจเพาะเลยงใหเปนหลายตนกได ดงนน เมอนาเทคนคการเพาะเลยงคพภะนมาใช ฝกยางพาราทมการ
ผสมตดแมเพยง 1 เมลด กสามารถนามาเพาะเลยงใหเปนตนสมบรณไดมากมาย ซงจะเพมประสทธภาพของการ
ผสมพนธยางไดมากขน นอกจากนตนยางใหมทไดรบก
สามารถขยายพนธตอในหองปฏบตการใหไดปรมาณมาก
ตามทตองการ ดกวาวธการเดมทจะไดเพยง 1 ตนตอ 1 เมลดเทานน(9) ขอดของการทสามารถเพาะเลยงคพภะของเมลดยาง คอ สามารถมนใจในทมาของพอและแมพนธทใช ทาใหสามารถตดตามลกษณะเดนและลกษณะดอยท
สาคญในลกผสมทได ในการผสมพนธยางทผานมา นยมใชวธผสมเกสรดวยแรงคน โดยมรายงานวา อตราความ สาเรจ หรออตราการผสมตดอยในระดบตามาก คออย
ระหวาง 2-5 % เทานน และยงมขนตอนทยงยาก และมปจจยหลายอยางทมสวนเกยวของกบผลสาเรจทงทควบคม
ไดและไมได เพราะเมลดทผสมนอยในสภาพธรรมชาต และเมลดจะใชเวลาถง 5 เดอนจงจะพฒนาเปนเมลดแก ซงในชวงดงกลาว เมลดกตองเสยงอยในสภาพแวดลอมตามธรรมชาต จงมผลใหความสาเรจอยในอตราตา นอกจากนปจจบนไดมการพฒนาวธการผสมพนธยางในหลอดแกว เปนการควบคมสภาพแวดลอมใหเหมาะสม โดยนาเอา
เกสรจากดอกตวผทใชเปนพอพนธไปผสมกบดอกตวเมย
ทใชเปนแมพนธ ดาเนนการในหลอดแกวในหอง ปฏบตการ เมอผสมตดกนาไปเพาะเลยงตามวธการทกลาวขางตน ซงนอกจากจะชวยเพมประสทธภาพการผสมพนธยางแลว ยงสามารถดาเนนการไดในปรมาณมากใน
ระยะเวลาสนกวาและใชพนทนอยกวาดวย
แผนททางพนธกรรมและเครองหมายโมเลกลเพอการปรบปรงพนธยางพารา พนธยางทใชปลกทวไปในปจจบนบางพนธม
ลกษณะสณฐานวทยาภายนอกทคอนขางแตกตางกน
ชดเจน สามารถจาแนกดวยวธการงายๆ และทนทในแปลงปลก ไมตองมการเกบตวอยางพช และไมตองอาศยวธการทางวทยาศาสตรทยงยากซบซอนแตอยางใด แตในงาน
ทดลองทมวตถประสงคเพอการจาแนกพนธยางลกผสมท
มสายพนธ ใกล เคยงกนมากจนมอาจจาแนกลกษณะ
สณฐานวทยาภายนอกได หรอเนนการรบรองการขน
ทะเบยนพนธพช และพระราชบญญตคมครองพนธพช การจาแนกลกษณะประจาพนธยาง โดยวธการจดทาลายพมพดเอนเอ (DNA fingerprint) ซงเปนวธการจาแนกพนธทมความแมนยา
การสรางแผนททางพนธกรรมมหลายวธการ ซงวธการตางๆ นนกจะใหขอมลทแตกตางกนทระดบของรายละเอยด สามารถนาไปประยกตใชในการตอบปญหาทางพนธศาสตรทแตกตางกน ในการสรางแผนทจโนม (linkage map) การเลอกประชากรสาหรบการสรางแผนทโมเลกลเครองหมายสมพนธนน จะเกยวของกบการเลอก พอ แม วธการผสม และชนดของโมเลกลเครองหมายทจะใชกขนอยกบจดประสงคของงานทดลอง ซงองคประกอบขอมลทสาคญในการสรางแผนทจโนมม 3 ประการ คอ 1) ประชากรทมการกระจายตว (segregating population) อาจจะเกดจากการผสมระหวางพนธแท (inbred line) 2 พนธ ซงนยมใชในพชผสมตวเอง และ พชผสมขาม หรออาจจะเกดจากการผสมระหวางพนธทาง 2 พนธ หรอพนธ ทางผสมพนธแทกได ตวอยางของประชากรทสามารถใชในการสรางแผนทจโนม คอ ประชากรของ doubled haploid ประชากรของลกผสมรนท 2 ประชากรของลก ผสมกลบ (backcross) หรอ ประชากรของ recombinant inbred lines 2) ขอมลพนธลกษณะ (genotype) ของประชากร ซงสามารถใชเครองหมายทงทางกายภาพและระดบโมเลกลเปนตวบงบอก สามารถสรางขนโดยใชเทคนคตางๆ เชน restriction fragment polymorphism (RFLP), random amplified length polymorphism (RAPD), amplified fragment length polymorphism (AFLP), simple sequence repeat (SSR), sequence tag site (STS), express sequence tag (EST) และยงมโมเลกลเครองหมายลกษณะอนๆ ทกาลงมการพฒนาขนอกเรอยๆ 3) ขอมลลกษณะทแสดงออกมาภายนอก (phenotype) ของประชากร เปนองคประกอบทมความสาคญอยางยงตอการวเคราะหหาจานวน ตาแหนง และบทบาทของยน ซงลกษณะตางๆ ทเราเหนอยนนเปนผลของการแสดงออกของยนทควบคมลกษณะนนๆ และสภาพแวดลอมทเปนตวสนบสนนสงเสรมหรอยบยงการแสดงออก (phenotype = genotype + environment) คณลกษณะทางกายวภาคและสรรวทยาทแสดงออกมาภายนอกเปนผลจากกรรมพนธ
และสงแวดลอม ลกษณะทางกายวภาคและสรรวทยาอาจแบงตามการแสดงออกไดเปน 2 กลม คอ ลกษณะทางคณภาพ (qualitative traits) เปนลกษณะทควบคมดวยยนไมกตว การแสดงออกในประชากรทมการกระจายตวสามารถแบงเปนกลมไดชดเจน และการแสดงออกนนไม

วทยาศาสตรเพอการพฒนา: รวทบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
176
ขนอยกบสภาพแวดลอม เชน สดอก สวนลกษณะทาง
ปรมาณ (quantitative trait) เปนลกษณะทควบคมดวยยนจานวนมาก การแสดงออกในประชากรทมการกระจายตวไมสามารถแบงเปนกลมไดชดเจน หรอเปนการกระจายตวแบบตอเนอง การแสดงออกของลกษณะนนมกจะม
อทธพลของสภาพแวดลอมมาเกยวของ เชน ผลผลต ความสง ความตานทานโรค(10)
การสรางแผนทจโนมอาศยหลกการทยนหรอ
โมเลกลเครองหมายทมตาแหนงอยใกลกนบนจโนมจะถก
ถายทอดไปดวยกนจากรนหนงไปยงรนถดไป ยงใกลกนมากเทาไรโอกาสทจะพบโมเลกลเครองหมายทงสองใน
รนลกตนเดยวกนกมมากเทานน หลกการนใชมาเปนตว กาหนดระยะหางระหวางโมเลกลเครองหมาย โดยระยะ หางระหวางโมเลกลเครองหมายทงสองคานวณจากความถ
ของการเกด recombination ในรนลก การวเคราะหหาจานวน ตาแหนง และบทบาทของยนควบคมลกษณะ
ปรมาณ (quality trait loci) เปนการหาความสมพนธระ หวางลกษณะทางปรมาณทสามารถชงตวงวด และลกษณะทางกายภาพทกระจายตวในประชากรกบโมเลกล
เครองหมายดเอนเอทกระจายตวในประชากรนนๆ ซงการวเคราะหความสมพนธดงกลาวจะตองอาศยองคประกอบ
หลก คอ ประชากรทมการกระจายตว และขอมลพนธลกษณะของประชากรรวมกบแผนทจโนมทมขอมล
ลกษณะทแสดงออกของประชากร คอ ลกษณะเชงคณภาพและลกษณะเชงปรมาณทสาคญทางการเกษตร เชน ผล ผลตและองคประกอบของผลผลต คณภาพของผลผลต ลกษณะตางๆ ทเกยวของกบการเขตกรรม ความตานทานโรค ความตานทานตอความเครยด ในการศกษาทางดานพนธศาสตรของลกษณะตางๆ เหลาน เพอใชในการปรบปรงพนธ นกปรบปรงพนธจะตองศกษาวา มยนจานวนเทาไรทควบคมลกษณะนน ยนเหลานนอยทใดบนโครโมโซม แตละยนมผลตอการแสดงออกเทาเทยมกน
หรอไม การทางานรวมกนของยนเหลานนเปนอยางไร การกระจายตวของลกษณะดงกลาวเปนผลมาจากยนเทาไร และสภาพแวดลอมเปนอยางไร เพอชวยใหการปรบปรง
พนธพชเปนไปอยางแมนยาและไดผล
เนองจากการปรบปรงพนธแบบดงเดมใชเวลา นานและขนกบสภาพแวดลอม ลกษณะตางๆ ทสาคญในการปรบปรงพนธอาจตรวจวดไดยาก หรอเปลยนไปตามสภาพแวดลอม ดงนน การใชโมเลกลเครองหมายทาใหการคดเลอกพนธไมตองขนกบปจจยเหลาน สามารถทาไดตงแตพชยงมขนาดเลก พอทจะแยกด เอนเอออกมา
ตรวจสอบได ทาใหเพมความแมนยาในขนตอนการคดเลอกพนธ สาหรบใชทดสอบในขนตอๆ ไปอยางมประสทธภาพ ซงชวยรนระยะเวลาในการคดเลอกไดมากในกรณของยางพารา นอกจากนน โมเลกลเครองหมายทพฒนาขนนน อาจนาไปสการคนพบยนทควบคมหรอเกยวของกบลกษณะพงประสงคตางๆ ทจะใชประโยชนตอไปไดมากมาย ทงการเผยองคความรเกยวกบยางพารา และการดดแปลงพนธใหมลกษณะพงประสงค ยางพาราม 18 โครโมโซม (2n = 36) ปจจบนหนวยงานวจย CIRAD ประเทศฝรงเศส สามารถสราง saturated genetic linkage map ของยางพารา ทง 18 โครโมโซม แผนทจโนมของยาง พาราดงกลาว ไดแสดง 717 โมเลกลเครองหมาย กระจายตวบน 18 linkage groups แผนทนประกอบดวย 301 RFLP, 388 AFLP, 18 microsatellite และ 10 isozyme markers ศกษาโดยใชประชากรจาก RRIM600 x PB217(3) และโดยความรวมมอระหวาง CIRAD กบนกวจย
สถาบนวจยยางของไทย ไดพยายามทาแผนทพนธกรรมยางใหละเอยด มการตรวจสอบความถกตอง แมนยามากขน(4) และปจจบนมการขยายความรวมมอพยายามพฒนาโมเลกลเครองหมายทเกยวของกบผลผลตนายางและการ
เจรญเตบโต รวมถงลกษณะอนๆ ซงคาดวาจะมประโยชนมากในระบบการคดเลอกเพอปรบปรงพนธยางพาราใน
อนาคตอนใกล
การคนหายนทเปนประโยชนตอการปรบปรงพนธยางพารา
เครองหมายโมเลกลทางดเอนเอถกนามาใชในการ
สบหายนควบคมลกษณะตางๆ ของพช ทาใหการปรบปรงพนธพชมประสทธภาพสงยงขน การใชเครองหมาย
โมเลกลชวยในกระบวนการหายนควบคมลกษณะตางๆ ทางพนธกรรมนน สามารถทาไดหลายวธ เชน การใชวธสบหายน (gene tagging) และการวางตาแหนงของยน (gene mapping) การคนหายนในพชมเปาหมายสงสด คอ การคนใหพบยนทควบคมความแตกตางของลกษณะ
ดงกลาว การสบหาเครองหมายโมเลกลทอยใกลกบยน เปนวธการคนหายนทมความเกยวพนกบลกษณะทาง
กายภาพ ลกษณะทางสรรวทยาหรอทางเคม โดยอาศยเครองหมายโมเลกล การสบหายนนนสามารถสบหาใน
กลมของประชากรชนดตางๆ ท เกดจากการผสมพนธระหว างพอและแม พนธ ท ม ความแตกต า งกนทาง
พนธกรรมของลกษณะทตองการ โดยหลกการสาคญ คอ การสรางกลมตวแทนสองกลมทมความแตกตางของ
ลกษณะทจะทาการศกษาอยางชดเจน เครองหมายโมเลกล

วทยาศาสตรเพอการพฒนา: รวทบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
177
จะถกนามาใชเพอสบหาความแตกตางระหวางกลมทงสอง โดยท เครองหมายโมเลกลใดกตามทใหความแตกตาง
ระหวางกลม ถอวาเปนเครองหมายทมความเกยวพนกบ
ความแตกตางของลกษณะในกลมทงสองนน และนาจะอยใกลกบยนทควบคมลกษณะนน การคนหายนมหลาย
แนวทางดวยกนขนกบวาเราทราบขอมลจาเพาะอะไรบาง
ของยนนน เชน ลาดบเบสของยน ตาแหนงยนบน
โครโมโซม ความจาเพาะในการแสดงออกของยน ทราบหนาทการทางานของยน การคนหายนอาจทาไดเมอทราบขอมลลาดบเบส/ลาดบกรดอะมโน และถอเปนการคนหายนทตรงไปตรงมาทสด เนองจากลาดบเบส/กรดอะมโนจะถกนามาใชในการออกแบบ primers ในการขยายปรมาณและโคลนยน หรออาจนามาใชออกแบบ probe เพอใชในการคดเลอก clone จาก genomic libraries หรอ cDNA libraries ไดโดยตรง ในกรณทสามารถแยกโปรตนบรสทธได โปรตนดงกลาวอาจใชในการสราง polyclonal antibody จาเพาะเพอใชในการตรวจหาโคลนจาก expression library ไดโดยตรง เมอไดโคลนยนแลวจงนามาทดสอบหนาทตอไป อกแนวทางหนงในการคนหายน คอ การคนหายนเมอทราบความจา เพาะในการแสดงออกของยน ยนบางชนดมการแสดงออกอยาง
จาเพาะเจาะจง (differential gene expression) กบชวงเวลา (temporal) และบรเวณทแสดงออก (spacial) ทงน
เนองจากยนเหลานมลาดบเบสทควบคมการแสดงออกของ
ยน (regulatory sequence) ทตอบสนองตอสงเรา หรอ ระยะการเจรญเตบโต หรอชนดของเนอ เยอ ( tissue specific gene expression) การโคลนยนในกรณนตองม cDNA library ทสรางจากตวอยางทยนนนๆ กาลงแสดงออก นอกจากน เรายงสามารถคนหายนแบบ insertional mutagenesis โดยทยนปกตเมอมดเอนเอเขามาแทรกในตาแหนง coding sequence หรอ regulatory sequence อาจทาใหการแสดงออกเปลยนไปมาก จนทาใหลกษณะทยนเหลานควบคมอยเปลยนไปจนสงเกตไดดวย
ตาเปลา อาศยระบบการถายยนททรงประสทธภาพ ยนทกยนบนจโนมกอาจถกแทรก จนทาใหเราสามารถศกษาทราบหนาทของแตละยนนนได หนวยงานวจย IRD ประเทศฝรงเศสรวมกบนกวจยของมหาวทยาลยมหดล และนกวจยสถาบนวจยยางไดเนนในเรองการคนหายน
และศกษาหนาทของยนในยางพาราทมความสาคญในการ
ปรบปรงพนธและการพฒนาเปนโมเลกลเครองหมาย สาหรบการคดเลอกพนธ การจดการระบบปลกระบบกรด ยาง รวมทงมการสรางและเกบรวบรวม genomic library, cDNA library และ expressed sequence tags ของยางพารา
เพอใชสาหรบการคนหายนทมประโยชนในการปรบปรง
พนธยางซงคาดวาจะมประโยชนมากในอนาคตอนใกล(2,8)
การดดแปลงพนธกรรมยางพารา การเปลยนแปลงพนธกรรมพชสามารถทาไดโดย
การผสมพนธแบบดงเดม การทาใหเกดการเปลยนแปลง ทางพนธกรรมโดยวธการทางฟสกส เคม และชววทยาซงมกเปนแบบไมจาเพาะเจาะจง การทาใหยนทมอยเดมไมแสดงออก หรอการเพมยนทพชนนไมมอยเดมแบบ
จาเพาะเจาะจง สวนเทคโนโลยทเกยวของในการปรบปรงพนธพชมหลายรปแบบ ไดแก เทคโนโลยการควบคมดวยวธการทางฟสกสและเคม การเพาะเลยงเซลล เนอเยอ และอวยวะ การเปลยนจานวนชดโครโมโซม protoplast การกลายพนธ และการถายโอนยนซงเปนการสรางพช
ดดแปลงพนธกรรม (genetically modified plants or transgenic plants) เพอการปรบปรงพนธใหมคณสมบต
ตามตองการ ขนตอนพนฐานในการสรางพชดดแปลง
พนธกรรม คอ เลอกยนทสนใจและนาไปใสในดเอนเอ
พาหะ เตรยมเซลลหรอเนอเยอเพอทาหนาทเปนตวรบยน การนาพาดเอนเอเขาไปในเซลลหรอตวรบ และวธการคดเลอกเซลลหรอเนอเยอทไดรบยนตามตองการ นอกจากการเลอกยนทถกตองแลว ยงตองคานงถงการจดการใหเกดการแสดงออกของยนเปาหมายอยางถกตาแหนง ถกเวลา การกาหนดการแสดงออกทาไดโดยการเลอกบรเวณ promoter ทควบคมการถอดรหสใหแมนยาซงม
ความสาคญมาก หากตองการถายยนจากสงมชวตชนดอนทไมใชพช จะตองมการปรบใชรหสการแปลยน (codon usage) ใหถกตองตรงกบพชตวรบยนอกดวย สาหรบในยางพารา มความพยายามทจะคนหา latex cell-specific promoters เพอใชในการควบคมใหยนแสดงออกเฉพาะในทอนายางเทานน ซงจะมประโยชนในการประยกตใชตนยางพาราผลตสารสาคญๆ ซงนกวจยจากสถาบนวจย IRD/CIRAD ประเทศฝรงเศส รวมกบมหาวทยาลยมหดล ไดรายงานการคนพบ latex cell-specific promoter ชนดหนงทพสจนวาสามารถใชงานได นอกจากนนยงอยในระหวางการศกษา ethylene-inducible promoters ใน
ยางพาราเพอใชในการควบคมการสงใหยนแสดงออกเมอ
ตองการไดอกดวย(5)
การใช พช เปนแหล งผลตสาร เวชภณฑและ
อตสาหกรรม (plant molecular farming) เปนงานวจยทกาลงไดรบความสนใจและนาจะไดรบการยอมรบสงกวา
พชดดแปลงยนเพออาหาร ในชวงเวลาทผานมา การผลตสารสาคญทางเวชกรรมในสงมชวตจะทาในแบคทเรยหรอ

วทยาศาสตรเพอการพฒนา: รวทบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
178
ยสต รวมไปถงเซลลเพาะเลยงของสตวเลยงลกดวยนม เซลลเพาะเลยงของแมลง และสตวถายโอนยน สาหรบการศกษาวจยในระบบพชถายโอนยน มรายงานวามขอไดเปรยบหลายประการ เชน ราคาถกกวา เมอเปรยบเทยบกบระบบทใชยสตหรอแบคทเรย ขยายปรมาณไดงาย ปลอดภยเนองจากไมมเชอกอโรคของคนเมอเปรยบเทยบ
กบความเสยงของการผลตในสตว และงายตอการสกดและนาสารมาใช ขณะนมประมาณ 11 บรษททวโลกทกาลงจะมผลงานวจยในดานนออกมาสตลาด(7) สาหรบยางพารา ไดมหนวยงานวจยหลายแหงทวโลกมแนวคดการใช
ยางพาราเปนแหลงผลตสารสาคญทางการแพทยและ
อตสาหกรรม โดยตองการบงคบใหตนยางผลตสารสาคญออกมากบนายางแบบเดยวกบทแมววใหนานม เพอความสะดวกในการเกบเกยวไปใชประโยชน นอกจากนยงมแนวคดในการปรบปรงคณภาพเนอไมของยางพาราให
สอดคลองกบความตองการอกดวย ความกาวหนาขณะนมการพฒนาระบบการเพาะเลยงเนอเยอยางพาราบางพนธ
อยางมประสทธภาพ มการพฒนาระบบการถายยน การเลอก regulatory sequence ควบคมการแสดงออกของยนเปาหมายและอนๆ โดยมหลายหนวยงานทวโลกรายงานความสาเรจในการสรางยางพาราดดแปลงพนธกรรมใน
หองปฏบตการ เชน สถาบนวจยยางของประเทศมาเลเซย ไดรายงานความสาเรจการผลตตนยางพาราดดแปลง
พนธกรรมใหสามารถผลตสารสาคญทางการแพทยบาง
ชนดไดแลวรวมทงมการจดสทธบตรระบบนไวดวย(1) จงเปนไปไดวาแนวคดการใชตนยางพาราในการผลตสาร
เวชภณฑและอตสาหกรรมอาจจะถกนามาใชจรงใน
อนาคตอนใกล
เอกสารอางอง 1. Boffey SA, Jones H, Slater RJ, Arokiaraj P, Cheong
KF, Wan Abdul Rahaman WY et al. Method for the production of proteins in plant fluids. United States Patent 5580768;1996.
2. Charoenwut C, Kongsawadworakul P, Pichaut JP, Nandris D, Sookmark U, Narangajavana J et al. Cloning and characterization of a specific molecular marker of rubber tree trunk phloem necrosis. Proceedings of the IRRDB Annual Meetings and Conference 2007 on "Natural Rubber Industry: R&D Achievements and Challenges"; 2007 November 12-16; Siem Reap, Cambodia. 2007. p.64-72.
3. Lespinasse D, Rodier-Goud M, Grivet L, Leconte A, Legnate H, Seguin M. A saturated genetic linkage map of rubber tree (Hevea spp.) based on RFLP, AFLP, microsatellite, and isozyme markers. Theoret Appl Genetics 2000;100:127-38.
4. Prapan K, Clement-Demange A, Teerawatanasuk K, Rodier-Goud M, Seguin M. Genetic mapping and field study of a full-sib family (RRIM600 x PB217) in Hevea brasiliensis (Genmap project). KU-RRIT-DOA-INRA-PIAF-CIRAD Seminar; 2004 May 27-28; Bangkok, Thailand. 2004.
5. Pujade-Renaud V, Sanier C, Cambillau L, Pappusamy A, Jones H, Ruengsri N et al. Molecular characterisa-tion of new members of the Hevea brasiliensis hevein multigene family and analysis of their promoter region in rice. Biochim Biophys Acta 2005;1727: 151 -61.
6. Shewry PR, Jones HD, Halford NG. Plant biotechno-logy: transgenic crops. Adv Biochem Eng Biotechnol 2008;111:149-86.
7. Sun SS. Application of agricultural biotechnology to improve food nutrition and healthcare products. Asia Pac J Clin Nutr 2008;17:87-90.
8. Tungngoen K, Sakr S, Kongsawadworakul P, Sook-mark U, Narangajavana J, Chrestin H. Aquaporin gene expression in the trunk phloem and in the laticifers of rubber tree. Proceedings of the IRRDB Annual Meetings and International Rubber Con-ference; 2006 November 13-17; Ho Chi Minh City, Vietnam; 2006. p.125-34.
9. Venkatachalam P, Jayashree R, Rekha K, Sushma-kumari S, Sobha S, Jayasree PK et al. Rubber tree (H. brasiliensis Muell. Arg). Methods Mol Biol 2006;344:153-64.
10. Xu Y, Crouch JH. Marker-assisted selection in plant breeding: from publications to practice. Crop Sci 2008;48:391-407.

179
สารพษจากเชอรากบบทบาทตอความสามารถในการกอโรคพช
ทพา อศวรกษ ภาควชาเทคโนโลยชวภาพ คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
เชอรามบทบาทสาคญในทางการเกษตรในฐานะของเชอกอโรคพชททาใหเกดความเสยหายตอพชเศรษฐกจไดอยาง
มาก ในจานวนเมแทบอไลตหลายชนดทเชอราใชในกระบวนการกอโรคพชนน สารพษของเชอรามบทบาทสาคญในการปฏสมพนธระหวางเชอรากอโรคและพชใหอาศย สารพษทเกยวของกบกระบวนการกอโรคนมทงทเปนสารพษจาเพาะตอพชใหอาศย (host-selective toxins) ซงออกฤทธตอพชใหอาศยเฉพาะสายพนธหรอบางจโนไทปเทานน และสารพษไมจาเพาะตอพชใหอาศย (non host-selective toxins) ซงออกฤทธตอพชหลายชนด สารพษจาเพาะตอพชใหอาศยสวนใหญเปนสารทตยภมทมขนาดโมเลกลตาและมเพยงไมกชนดเทานนทเปนโปรตน สารพษจาเพาะหลายชนดมบทบาทสาคญตอความสามารถในการกอโรคพชของเชอราหรอตอความรนแรงของโรคพช ตวอยางของสารพษเหลานรวมถงสารพษจาเพาะทตยภมทรจกกนด เชน T-toxin, victorin, HC-toxin และสารพษทเปนโปรตน เชน Ptr-toxin เปนตน ในปฏสมพนธระหวางเชอราทสรางสารพษจาเพาะและพชใหอาศยนน ความไวตอสารพษมกขนอยกบยนเพยงหนงยนในพชใหอาศย ในขณะทเชอรามกอาศยยนหลายยนรวมกนในการสงเคราะหโมเลกลของสารพษทเปนสารทตยภม โดยยนทใชในการสงเคราะหเหลานมกเปนกลมยน
คาสาคญ: เชอรา; เชอรากอโรคพช; สารพษ; สารพษจาเพาะตอพชใหอาศย
Abstract
Filamentous fungi are known for their ability to cause devastating plant diseases. During the pathogenesis process, fungi utilize several metabolites in order to penetrate and successfully colonize host plants. Among these metabolites, toxins are known to be involved in several fungal-host plant interactions. Phytotoxins produced by fungal pathogens include host-selective toxins (HSTs), which target only host of specific genotypes, and non host-selective toxins, which are active against a wide range of hosts. Most known HSTs are low molecular weight secondary metabolites, although a few protein toxins have been described. In some pathosystems, HSTs play essential roles in pathogenesis or virulence. Examples of such HSTs include the well known secondary metabolites T-toxin, victorin and HC-toxin, and protein toxins, Ptr-toxin. In most interaction between a HST producer and its host plant, sensitivity to HST is determined by a specific host plant gene. Biosynthesis of HST, however, commonly involves several genes that are usually clustered together.
Keywords: fungi; fungal plant pathogen; toxin; host-selective toxin
บทนา
เชอรามบทบาทสาคญในทางการเกษตรในฐานะ
ของเช อกอโรคพชทท าให เกดความเสยหายตอพช
เศรษฐกจไดอยางมาก กระบวนการกอโรคพชของเชอราอาจแบงออกไดเปน 2 กระบวนการหลกคอ การชอนเขาสพช (penetration) และการยดครอง (colonization) เรมตงแตการทเชอราสมผส และยดเกาะกบผวของพช เมอ
เจรญและอยในบรเวณทเหมาะสมแลว กจะชอนลกเขาส
ภายในพชและเจรญอยภายใน จนในทสดจะเจรญออกมาภายนอกตนพชอกครงเพอสรางและแพรกระจายสปอร(29) ในกระบวนการดงกลาวนเชอราจะสรางเมแทบอไลตหลาย
ชนดเพอชวยในขนตอนตางๆ ของกระบวนการกอโรคพช ความสาคญของเมแทบอไลตจากเชอราในการกอโรคพช
นนแตกตางกนไปในแตละปฏสมพนธ บางชนดอาจม
ความจาเปนตอการกอโรค หรออาจมผลตอความรนแรงของโรคทเกด หรออาจไมมความจาเปนในการกอโรคก

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
180
ได(29) นกวจยใหความสาคญกบการศกษาปฏสมพนธ
เหลานอยางมากเนองจากความเขาใจถงกระบวนการการ
กอโรคของเชอราอาจนาไปสการปองกนและควบคมโรค
พชทสรางความเสยหายตอพชเศรษฐกจได ในจานวน เมแทบอไลตทมบทบาทในการกอโรคพชนน สารพษนบเปนอาวธสาคญของเชอราทใชในการกอโรคพชใน
หลายปฏสมพนธ สารพษของเชอราอาจมผลโดยตรงตอเซลลของพชซงมกเกยวของกบการรบกวนการเลอกผาน
ของเยอหมเซลล หรออาจไปรบกวนการสรางหรอการทางานของเมแทบอไลตของพช(10)
สารพษของเชอราทมความเกยวของกบความ สามารถในการกอโรคพชอาจแบงออกไดเปน 2 กลม คอ สารพษจาเพาะตอพชใหอาศย (host-selective toxins) ซงเปนพษตอพชบางสายพนธเทานนและสารพษไมจาเพาะ
ตอพชใหอาศย (non host-selective toxins) ซงมผลตอพชไดหลายชนด(5,25) ในบทความนจะกลาวถงตวอยางของ
สารพษทมบทบาทสาคญในปฏสมพนธระหวางเชอรากอ
โรคพชและพชใหอาศย โดยมงเนนทตวอยางตนแบบของสารพษจาเพาะตอพชใหอาศยทมการศกษาถงการออกฤทธ
และการสงเคราะหในระดบชวโมเลกลแลว
สารพษจาเพาะตอพชใหอาศย
สารพษจาเพาะทสรางโดยเชอรากอโรคพชสวน
ใหญเปนสารทตยภมทมนาหนกโมเลกลตาและมความเปน
พษจาเพาะตอพชบางพนธหรอบางจโนไทปเทานน มเพยงสวนนอยท เปนโปรตน (27) สารพษจาเพาะหลายชนดมความสาคญตอความสามารถกอโรคพชของเชอราและ
ความรนแรงของโรคในพชใหอาศย(12,25) แมจะมรายงานถงเชอราหลายสกลทสามารถสรางสารพษจาเพาะได แตสารพษจาเพาะทเปนสารทตยภมสวนใหญสรางโดยเชอรา
ในสกล Alternaria และ Cochliobolus(26) ทนาสนใจคอเชอรามกใชยนหลายยนรวมกนในการสงเคราะหสารพษทตย
ภมแตละชนด โดย ยนเหลานมกรวมกลมกนอยเปนกลมยน (gene cluster) ในขณะทพชจะมเพยงยนเดยวททาใหเกดความไวตอสารพษจาเพาะ (27) ตวอยางตนแบบของสารพษจาเพาะทเปนสารทตยภมทรจกกนด ไดแก T-toxin, victorin และ HC-toxin นอกจากสารทตยภมแลวยงมราย งานถงสารพษจาเพาะบางชนดทเปนโปรตน เชน Ptr-toxin อกดวย(21,27)
T-Toxin T-toxin เปนสารพษในกลมพอลคไทดสายตรง
ทมความยาวประมาณ 35-49 คารบอน สรางโดยเชอรา
Cochliobolus heterostrophus ซงกอโรคใบไหม (southern corn leaf blight) ในขาวโพด(3) โดยทวไปแลว C. heterostrophus กอโรคใบไหมในขาวโพดอยางไมรนแรงนก(3) จนตอมามการระบาดของโรคใบไหมขาวโพดทประเทศสหรฐอเมรกาซงกอใหเกดความเสยหายอยางมาก จากการวจยพบวาเชอรา C. heterostrophus สายพนธทกอใหเกดการระบาดอยางรนแรงในชวงเวลานน คอ C. heterostrophus race T ซงแตกตางจากสายพนธเดมคอ race O ตรงท race T สามารถสราง T-toxin ไดและมความ สามารถกอโรคในขาวโพดสายพนธ Texas male sterile cytoplasm (T-cytoplasm) ไดอยางรนแรง(25,28)
ความไวตอ T-toxin ของขาวโพดสายพนธ T-cytoplasm เกดจากยน T-urf13 ซงใหผลผลตเปนโปรตนขนาด 13 กโลดอลตนเรยกวา URF13 ซงอยทเยอหมชนในของไมโทคอนเดรย โดยโปรตนนนอกจากจะทาใหเกดการเปนหมนทางไซโทพลาซมของเพศผในขาวโพด T-cytoplasm แลว ยงทาหนาทเปนตวรบของ T-toxin อก ดวย(23) ในขาวโพด T-cytoplasm นน T-toxin จะจบกบโปรตน URF13 ทาใหเกดการขยายตวของรเปดในเยอหมชนในของไมโทคอนเดรยและรบกวนการเลอกผานของ
เยอหมชนในของไมโทคอนเดรย นาไปสการรวไหลของอเลก-โทรไลต(23) การแสดงออกของ URF13 ในสงมชวตชนดตางๆ เชน Escherichia coli และ Saccharomyces cerevisiae ทาใหเกดความไวตอ T-toxin ในสงมชวตนนๆ ซงแสดงใหเหนถงความสาคญของโปรตนนในฐานะตวรบ
จาเพาะของ T-toxin(4,15) สารพษชนดนไมมผลตอขาวโพดสายพนธ normal cytoplasm (N-cytoplasm) ซงไมม URF13 ในเยอหมชนในของไมโทคอนเดรย ดงนน C. heterostrophus race T จงไมกอโรคอยางรนแรงใน
ขาวโพด N-cytoplasm เชนเดยวกบ race O(28)
การศกษาทางพนธศาสตรแสดงใหเหนวาเชอรา C. heterostrophus สายพนธ race O และ race T นเกอบจะเปนสายพนธ isogenic โดย race T ตางจาก race O ท 2 โครโมโซม ซงคาดวาเกดจากการแลกเปลยนชนสวนของโครโมโซม 6 และโครโมโซม 12 ของ race O เกดเปนโครโมโซม 6;12 และโครโมโซม 12;6 ใน race T นอก จากน ยงมการแทรกของชนสวนดเอนเอแปลกปลอมขนาดรวม 1.2 เมกะเบสเขาตรงบรเวณตาแหนงทมการแลก เปลยนบนโครโมโซมทงสองทเกดใหม(3,17) ยนของ C. heterostrophus ท เ ก ย ว ข อ ง ก า ร ส ง เ ค ร า ะห T-toxin ประกอบดวยยนหลายยนอยบน Tox1 loci ซงประกอบ ดวย Tox1A และ Tox1B loci โดย loci ทง 2 นอยใน
ชนสวนดเอนเอขนาด 1.2 เมกะเบสทพบเฉพาะใน race T

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
181
เทานน(3) จากการศกษาทางชววทยาระดบโมเลกลพบวามยนบน Tox1 loci อยางนอย 7 ยน ไดแกยนสาหรบ poly-ketide synthases (PKS1 และ PKS2) ยนทมความคลายคลงกบยนสาหรบ Lambda crystalline (LAM1) บน Tox1A และยนสาหรบ decarboxylase (DEC1) และ reductases (RED1, RED2 และ RED3) บน Tox1B(25) โดยยน PKS1, PKS2, LAM1, DEC1, RED2 และ RED3 มความจาเปนตอการสงเคราะห T-toxin และตอความรนแรงของโรคใน
ขาวโพด T-cytoplasm(3,25,28)
PM-toxin
นอกจาก T-toxin แลว URF13 ยงทาใหเกดความไวตอ PM-toxin ซงเปนสารพษจาเพาะทสงเคราะหโดยเชอรา Didymella zeae-maydis ซงทาใหเกดโรคใบไหมเหลอง (yellow leaf blight) ในขาวโพด(15) เปนทนาสนใจวาถงแมเชอรา D. zeae-maydis จะมลกษณะสณฐานวทยาและวงจรชวตทแตกตางจาก C. heterostrophus แต PM-toxin มทงการออกฤทธและโครงสรางทคลายกบ T-toxin อยางมาก โดยโครงสรางโพลคไทดสายตรงของ PM-toxin มกสน
กวา T-toxin คอมความยาวประมาณ 33-35 คารบอน(25)
ยนทเกยวของกบการสงเคราะห PM-toxin อยรวม กนบน MzTox1 locus ยนบน locus นทเกยวของกบ
กระบวนการสงเคราะหสารพษ ไดแก polyketide synthase (MzPKS1) และ reductases (MzRED1 และ MzRED2)(25) เมอเปรยบเทยบกบยนทอยบน Tox1 loci ของ C. hetero-strophus แลวพบวา MzPKS1 ของ D. zeae-maydis นาจะเปนยนทมตนกาเนดเหมอนกบ PKS1 ของ C. hetero-strophus(3) ในขณะท MzRED1 และ MzRED2 ของ D. zeae-maydis และ RED1, RED2 และ RED3 ของ C. heterostrophus มลาดบกรดอะมโนและโครงสรางของยนทแตกตางกน(25)
Victorin
Victorin อยในกลมของ cyclic pentapeptides สรางโดย C. victoriae ซงกอใหเกดโรคไหม (victoria blight) ในขาวโอต(27) สารพษชนดนมความจาเปนตอการกอโรคของ C. victoriae ในขาวโอต และพบเฉพาะในสายพนธทสามารถกอโรคได เทานน (27) ความจาเพาะตอ victorin พบในขาวโอตสายพนธ Victoria ซงมอลลลเดนของยน Vb(8) ซงขาวโอตสายพนธนมยน Pc-2 ซงทาใหเกดความทนทานตอโรคราสนม Puccinia coronata(19) เนอง จากความทนทานตอโรคราสนมและความไวตอ victorin ไมสามารถแยกจากกนไดในการศกษาทางพนธศาสตร ทา
ใหเปนทคาดกนวา Pc-2 และ Vb อาจเปนยนเดยวกนหรอเปนยนทยดเรยงกน(19)
ทนาสนใจคอสารพษชนดนมผลกระตนใหเซลล
ของพชมปฏกรยาคลายคลงกบปฏกรยาการปองกนตนเอง
ของพช เชน การสรางสารตานจลชพอยาง phytoalexin การเปลยนแปลงทางกายภาพทมกพบในกระบวนการ
ตานทานโรค และการตายของเซลลอยางรวดเรวคลายคลงกบการเกดอะพอพโทสส เปนตน ซงปฏกรยาเหลานอาจเปนผลมาจากความไวตอสารพษชนดน และความตานทานตอโรคราสนมถกควบคมโดยยนเดยวกนหรอยนทยดเรยง
กน (9,19) นอกจากนสารพษชนดนยงจบอยางจาเพาะกบโปรตนซงเปน subunit ของ glycine decarboxylase complex ของไมโทคอนเดรย และมผลยบยงกจกรรมของกลมเอนไซมนซงมบทบาทสาคญในการหายใจเชงแสง
ของพชดวย(8) โดยคาดวากระบวนการออกฤทธของ victorin นน เรมจากปฏสมพนธของสารพษชนดนกบเปา หมายซงเกยวของกบผลผลตของยน Vb ในพช นาไปสการตายของเซลลในลกษณะคลายกบอะพอพโทสส ซงม
ขนตอนททาใหเกดการเปลยนแปลงการเลอกผานของเยอ
หมไมโทคอนเดรยทาใหสารพษชนดนสามารถเขาไปจบ
กบ P-subunit ของ glycine decarboxylase complex ได (8)
สาหรบกระบวนการสงเคราะหสารพษชนดนยง
ไมมงานวจยทบง ชถงยนท เกยวของกบกระบวนการ
สงเคราะหมากนก แตเปนทคาดกนวายนบน Tox3 locus ทใชในการสงเคราะหนาจะประกอบดวย nonribosomal peptide synthetase เนองจากโครงสรางของสารพษชนดนเปนแบบ cyclic pentapeptides(25, 26, 27)
HC-toxin
HC-toxin เปน cyclic tetrapeptide ทสงเคราะหโดยเชอรา C. carbonum race 1 ซงกอโรคใบไหม (northern corn leaf blight) ในขาวโพด(26) ความจาเพาะของ HC-toxin พบในปฏสมพนธระหวาง C. carbonum race 1 กบขาวโพดทมยนดอยแบบฮอโมไซกสท Hm1 และ Hm2 loci โดยสารพษนจะยบยงเอนไซม histone deacetylase ซงคาดวามบทบาทสาคญเกยวของกบการปองกนตนเองของ
พชจาก C. carbonum(26)
ถงแมวา HC-toxin จะถกจดอยในกลมของสารพษจาเพาะ แตสารพษชนดนมกลไกความจาเพาะตอพชใหอาศยแตกตางไปจากสารพษจาเพาะชนดอนๆ ทกลาวมาแลวอยางมาก เมอเปรยบเทยบกบ T-toxin ซงความไว
ของพชใหอาศยขนอยกบการมโปรตนทสรางโดยยน T-urf13 และ victorin ซงความไวขนอยกบการมอลลลเดน

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
182
ของยน Vb แลว จะเหนไดวาความไวตอสารพษทงสองชนดดงกลาวขนอย กบการมเปาหมายพเศษในพชให อาศย(25,27) ในขณะทความไวตอ HC-toxin ขนอยกบการขาดหายไปของผลผลตของยน Hm1 และ Hm2 ทปกตมอยในขาวโพด แตสญเสยไปจากการผาเหลาในขาวโพดสายพนธทไวตอ HC-toxin(26) ขาวโพดสายพนธทมอลลลเดนท Hm1 หรอ Hm2 จะมความตานทานตอ HC-toxin เนองจากยนทงสองนตางใหผลผลตเปนเอนไซม HC-toxin reduc-tase (HCTR) ซงจะขจดพษของ HC-toxin โดยการรดวซ กลมคารบอนลให HC-toxin อยในรปไมมการออกฤทธ
ทางชวภาพ(2) แมการมอลลลเดนท Hm1 หรอ Hm2 เพยงยนเดยวจะทาใหเกดความตานทานตอ HC-toxin ได แต Hm1 จะใหความตานทานทสมบรณกวา Hm2(24) นอกจากขาวโพดแลวยงพบ Hm1 ในธญพชหลายชนดซงอาจเนอง จากวาพชตระกลนสวนใหญรกษายนนไวเพอปองกนพช
จากการทาลายของ HC-toxin(24,26)
การสงเคราะห HC-toxin อยภายใตการควบคม
ของ Tox2 locus ซงเปนกลมยนขนาดรวมมากกวา 0.5 เมกะเบส(1) ประกอบดวยยนอยางนอย 7 ยน ไดแก ยนสาหรบ nonribosomal peptide synthetase เรยกวา HC-toxin synthetase (HST1), transport protein (TOXA), beta subunit ของ fatty acid synthase (TOXC), dehydrogenase (TOXD), transcription factor (TOXE), amino acid transminase (TOXF) และ alanine racemase (TOXG)(1) ซงยน HST1, TOXA, TOXC, TOXE, TOXF และ TOXG มบทบาทในการสงเคราะห HC-toxin(1) นอกจากนยงมยนอกหลายยนบน Tox2 ทอาจจะเกยวของกบกระบวนการสง- เคราะห HC-toxin เชน ยนทเขารหสเปน alpha subunit ของ fatty acid synthase เปนตน(26) โดยยนบน Tox2 นพบอยหลายชดในจโนมของเชอรา C. carbonum สายพนธทสราง HC-toxin (race 1) เทานนและไมพบในสายพนธอนทไมสราง HC-toxin(26)
Ptr-toxin ตามทกลาวแลววาสารพษจาเพาะสวนใหญทสราง
โดยเชอรากอโรคพชเปนสารทตยภม และมไมกชนดทเปนโปรตน(27) ตวอยางทมการศกษาถงการออกฤทธและการ
สงเคราะหแลว ไดแก Ptr-toxin สรางโดย Pyrenophora tritici-repentis ซงกอโรคใบจด (tan spot หรอ yellow leaf spot) ในขาวสาล โดยทาใหเกดแผลจากการตายของเนอ เยอหรอเกดภาวะพรองคลอโรฟล(11) เปนทนาสนใจวา
ความสามารถในการกอโรคและอาการของโรคนนขนอย
กบจโนไทปของพชใหอาศย และสายพนธของเชอราซงม
อยางนอย 8 สายพนธ (race 1 - race 8)(18) โดยเชอรา P. tritici-repentis แตละสายพนธจะกอโรคและใหอาการของโรคในขาวสาลจโนไทปตางๆ แตกตางกนไป แสดงวาเชอรานอาจสรางสารพษจาเพาะหลายชนด(7, 18)
มการศกษาถงสารพษจาเพาะของ P. tritici-repentis แลวอยางนอย 3 ชนด ไดแก Ptr ToxA, Ptr ToxB และ Ptr ToxC(22) โดย Ptr ToxA และ PtrToxB เปนโปรตน ในขณะท PtrToxC เปนสารขนาดโมเลกลตา(11,20) นอก จากน ยงมรายงานเบองตนถงสารพษจาเพาะอก 2 ชนดในชอ Ptr ToxD(22) เชอราทง 8 สายพนธจะสรางสารพษ
จาเพาะทแตกตางกนไปดงแสดงในตารางท 1 อาการของโรคพชทเปนผลจาก Ptr-toxin นมทงการตายของเนอเยอ (Ptr ToxA) และภาวะพรองคลอโรฟล (Ptr ToxB และ Ptr ToxC)(22) อาการทเกดจะเปนชนดใดขนอยกบยนพเศษของพชใหอาศย โดยความไวตอสารพษทนาไปสอาการตายของเนอเยอนนกาหนดโดยอลลลเดนของยน Tsn1 ในขณะทความไวตอสารพษทนาไปสภาวะพรองคลอโรฟล
กาหนดโดย quantitative trait loci (QTL)(21)
ในการสงเคราะหสารพษ Ptr ToxA ซงเปนโปรตนขนาด 13.2 กโลดอลตน สรางโดยยน ToxA ซงใน P. tritici-repentis race 2 จะมยนนเพยงชดเดยว(7,20) สวน Ptr ToxB ซงเปนโปรตนขนาดประมาณ 6.6 กโลดอลตน สงเคราะหโดยยน ToxB ซงใน P. tritici-repentis race 5 มยน ToxB อยหลายชดในจโนม(22)
สารพษจาเพาะตอพชใหอาศยชนดอนๆ นอกจากทกลาวมาขางตนแลว ยงมสารทตยภมท
จดอยในกลมสารพษจาเพาะตอพชใหอาศยอกมากกวา 20 ชนด ซงหลายชนดมบทบาทในความสามารถกอโรคพชหรอตอความรนแรงของโรคพช(26,28) ตวอยางทนาสนใจ เชน Alternaria alternata ซงมถง 7 pathotypes ทสรางสารพษทมความจาเพาะตอพชใหอาศยแตกตางกน ไดแก tomato pathotype, strawberry pathotype, tangerine pathotype, Japanese pear pathotype, rough lemon pathotype, apple pathotype และ tobacco pathotype ซงสรางสารพษ AAL-toxin, AF-toxin, ACT-toxin, AK-toxin, ACR-toxin, AM-toxin และ AT-toxin ตามลาดบ(16) โดยสารพษทง 7 ชนดนมโครงสรางทแตกตางกนไป ยกเวน AF-toxin, ACT-toxin และ AK-toxin ซงม 9,10-epoxy-8-hydroxy-9-methyl-decatrienoic acid moiety ทเหมอนกนในโครงสรางของโมเลกล(14,16) นอกจากน ยงมสารพษจาเพาะตอพชใหอาศยทเปนสารทตยภม ซงสรางโดยเชอราในสกลอนๆ อก เชน Periconia circinata ซง

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
183
ตารางท 1 ตวอยางสารพษจาเพาะตอพชใหอาศยทมบทบาทในการกอโรคพชของเชอรา
สารพษ โครงสรางหลก เชอราสายพนธทสงเคราะหสารพษ ตาแหนงยน (locus) ทเกยวของกบการสงเคราะหสารพษ (และยนททราบบนตาแหนงยนนนๆ)
ยนทใหความ จาเพาะในพช ใหอาศย
เอกสาร อางอง
T-toxin พอลคไทดสายตรง C. heterostrophus race T Tox1A (PKS1, PKS2 และ LAM1) และ Tox1B (DEC1, RED1, RED2 และ RED3)
T-urf13 (3, 25, 28)
PM-toxin พอลคไทดสายตรง D. zeae-maydis MzTox1 (MzPKS1, MzRED1, MzRED2)
T-urf13 (3, 25)
Victorin Cyclic pentapeptide
C. victoriae Tox3 (ยงไมมรายงานในระดบยน)
อลลลเดนท Vb locus
(8, 26, 27)
HC-toxin Cyclic tetrapeptide C. carbonum race 1 Tox2 (HST1, TOXA, TOXC, TOXD, TOXE, TOXF และ TOXG)
อลลลดอยท Hm loci (Hm1 และ Hm2)
(1, 26)
Ptr-toxin โปรตน (Ptr ToxA และ Ptr ToxB) และสารโมเลกลตา (Ptr ToxC)
P. tritrici-repentis race 1 (Ptr ToxA, Ptr ToxC); race 2 (Ptr ToxA); race 3 (Ptr ToxC); race 5 (Ptr ToxB); race 6 (Ptr ToxB, Ptr ToxC); race 7 (Ptr Tox A, Ptr ToxB) และ race 8 (Ptr ToxA, Ptr ToxB, Ptr ToxC)
ยน ToxA สาหรบ PtrToxA หรอ ยน ToxB สาหรบ PtrToxB
Tsn1 (Ptr ToxA) และ QTLs จาเพาะ (PtrToxB)
(18, 21, 22)
สายพนธทกอโรคพชสามารถสราง peritoxins ทออกฤทธเฉพาะตอขาวฟางบางจโนไทปได(6) เปนตน
โปรตนทเปนสารพษจาเพาะตอพชใหอาศยนนม
การรายงานและศกษาในระดบชวโมเลกลอยนอยมาก ชนดหนงในกลมนคอ Sn toxin ของเชอรา Stagonospora nodorum ซงกอโรค S. nodorum blotch ในขาวสาล เชอราชนดนสรางสารพษจาเพาะไดอยางนอย 3 ชนด คอ Sn ToxA, Sn Tox1 และ Sn Tox2(13) ทนาสนใจคอ Sn ToxA มความจาเพาะกบผลผลตของยน Tsn1 ในขาวสาล ซงเปนตวรบของ Ptr-toxin เชนกน สวนความไวตอ Sn tox1 และ Sn Tox2 นนเกดจากยน Ssn1 และ Ssn2 ในขาวสาล ตามลาดบ(13)
สรป
โรคพชทเกดจากเชอรากอใหเกดความเสยหายตอ
ผลผลตทางการเกษตรเปนอยางมากในแตละฤดกาล การศกษาปฏสมพนธระหวางเชอรากอโรคและพชใหอาศย
นบวามความสาคญอยางย ง เนองจากความเขาใจถงกระบวนการกอโรคของเชอราอาจนาไปสการปองกนและ
ควบคมโรคพชทสรางความเสยหายตอพชเศรษฐกจได สารพษจาเพาะตอพชใหอาศยเปนหนงในจานวนเมแทบอ-
ไลตทมความสาคญในการกอโรคหรอตอความรนแรงของ
โรคในหลายกรณ(12, 25) สารพษจาเพาะตอพชใหอาศยเปนสารพษกลมหนงทสรางโดยเชอรากอโรคพชโดยออกฤทธ
เฉพาะตอพชบางสายพนธหรอบางจโนไทปเทานน (27) สารพษกลมนมบทบาทสาคญตอความสามารถในการกอ
โรคหรอตอความรนแรงของโรคในหลายปฏสมพนธ
ระหวางเชอราและพชใหอาศย (16, 18) สารพษจาเพาะสวนใหญเปนสารทตยภมและมเพยงไมกชนดเทานนทเปน
โปรตน(13, 27) ถงแมจะมการศกษาถงสารพษจาเพาะตอพชใหอาศยมาเปนเวลานานแตมจานวนนอยทมการศกษา
อยางละเอยดและมหลกฐานทแสดงใหเหนบทบาทสาคญ
ในการกอโรคพชอยางชดเจน บทความนไดกลาวถงตว อยางตนแบบของสารพษจาเพาะหลายชนดทมความสาคญ
ในการกอโรคพชซงมการศกษาถงความจาเพาะในการออก
ฤทธและยนทเกยวของกบกระบวนการสงเคราะหของ
สารพษดงกลาว เมอเปรยบเทยบปฏสมพนธระหวางเชอราทสรางสารพษจาเพาะกบพชใหอาศยทไวตอสารพษแตละ
ชนดในตวอยางขางตน จะเหนไดวาความจาเพาะของสารพษในกลมนมกขนอยกบยนเพยงยนเดยวในพชให
อาศย (27) โดยสวนใหญแลวยนททาใหเกดความไวตอสารพษจาเพาะจะพบเฉพาะในพชใหอาศยสายพนธหรอ จโนไทปทไวตอสารพษเทานนและไมพบในพชสายพนธ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
184
อน โดยผลผลตของยนนมกจะทาหนาทเปนตวรบเฉพาะของสารพษ(24) ยกเวนในกรณของ HC-toxin ทความไวตอสารพษเกดจากการขาดหายไปของยนซงมอยโดยทวไปใน
พชหลายชนด ทาใหไมสามารถตานทานตอสารพษได(26) การสงเคราะหสารพษจาเพาะทเปนสารทตยภมนนมก
อาศยยนหลายยนทมกจะอยรวมกนเปนกลม(25) นอกจากการออกฤทธผานตวรบจาเพาะในพชใหอาศยแลว สารพษบางชนด เชน victorin ยงกระตนใหพชเกดปฏกรยาคลาย คลงกบทเกดในกระบวนการตานทานโรค(19, 27) ซงเปนทนาสนใจศกษาถงความสาคญของปฏกรยาดงกลาวตอการ
กอโรคของเชอราทสรางสารพษนนๆ
เอกสารอางอง 1. Ahn JH, Cheng YQ, Walton JD. An extended
physical map of the TOX2 locus of Cochliobolus carbonum required for biosynthesis of HC-toxin. Fungal Genet Biol 2002;35:31-8.
2. Baidyaroy D, Brosch G, Graessle S, Trojer P, Walton JD. Characterization of inhibitor-resistant histone de-acetylase activity in plant-pathogenic fungi. Eukaryot Cell 2002;1:538-47.
3. Baker SE, Kroken S, Inderbitzin P, Asvarak T, Li BY, Shi L et al. Two polyketide synthase-encoding genes are required for biosynthesis of the polyketide virulence factor, T-toxin, by Cochliobolus hetero-strophus. Mol Plant Microbe Interact 2006;19:139-49.
4. Braun CJ, Siedow JN, Levings CS, 3rd. Fungal toxins bind to the URF13 protein in maize mitochondria and Escherichia coli. Plant Cell 1990;2:153-61.
5. Chen HQ, Lee MH, Chung KR. Functional characterization of three genes encoding putative oxidoreductases required for cercosporin toxin biosynthesis in the fungus Cercospora nicotianae. Microbiology 2007;153:2781-90.
6. Churchill AC, Dunkle LD, Silbert W, Kennedy KJ, Macko V. Differential synthesis of peritoxins and precursors by pathogenic strains of the fungus Periconia circinata. Appl Environ Microbiol 2001; 67:5721-8.
7. Ciuffetti LM, Tuori RP, Gaventa JM. A single gene encodes a selective toxin causal to the development of tan spot of wheat. The Plant cell 1997;9:135-44.
8. Curtis MJ, Wolpert TJ. The oat mitochondrial permeability transition and its implication in victorin binding and induced cell death. Plant J 2002;29:295-312.
9. Curtis MJ, Wolpert TJ. The victorin-induced mitochondrial permeability transition precedes cell shrinkage and biochemical markers of cell death, and
shrinkage occurs without loss of membrane integrity. Plant J 2004;38:244-59.
10. Daub ME, Herrero S, Chung KR. Photoactivated perylenequinone toxins in fungal pathogenesis of plants. FEMS microbiology letters. 2005;52:197-206.
11. Effertz RJ, Meinhardt SW, Anderson JA, Jordahl JG, Francl LJ. Identification of a chlorosis-inducing toxin from Pyrenophora tritici-repentis and the chromosomal location of an insensitivity locus in wheat. Phytopathology 2002;92:527-33.
12. Friesen TL, Faris JD, Solomon PS, Oliver RP. Host-specific toxins: effectors of necrotrophic patho-genicity. Cell Microbiol 2008;10:1421-8.
13. Friesen TL, Meinhardt SW, Faris JD. The Stagonospora nodorum-wheat pathosystem in-volves multiple proteinaceous host-selective toxins and corresponding host sensitivity genes that interact in an inverse gene-for-gene manner. Plant J 2007;51: 681-92.
14. Hatta R, Ito K, Hosaki Y, Tanaka T, Tanaka A, Yamamoto M et al. A conditionally dispensable chromosome controls host-specific pathogenicity in the fungal plant pathogen Alternaria alternata. Genetics 2002;161:59-70.
15. Huang J, Lee SH, Lin C, Medici R, Hack E, Myers AM. Expression in yeast of the T-urf13 protein from Texas male-sterile maize mitochondria confers sensitivity to methomyl and to Texas-cytoplasm-specific fungal toxins. EMBO J 1990; 9:339-47.
16. Ito K, Tanaka T, Hatta R, Yamamoto M, Akimitsu K, Tsuge T. Dissection of the host range of the fungal plant pathogen alternaria alternata by modi-fication of secondary metabolism. Mol Microbiol 2004;52:399-411.
17. Kodama M, Rose MS, Yang G, Yun SH, Yoder OC, Turgeon BG. The translocation-associated tox1 locus of Cochliobolus heterostrophus is two genetic elements on two different chromosomes. Genetics 1999;151:585-96.
18. Lamari L, Strelkov SE, Yahyaoui A, Orabi J, Smith RB. The identification of two new races of Pyrenophora tritici-repentis from the host center of diversity confirms a one-to-one relationship in tan spot of wheat. Phytopathology. 2003;93:391-6.
19. Lorang JM, Sweat TA, Wolpert TJ. Plant disease susceptibility conferred by a "resistance" gene. Proc Nat Acad Sci USA 2007;104:14861-6.
20. Manning VA, Ciuffetti LM. Localization of Ptr ToxA produced by Pyrenophora triticirepentis reveals protein import into wheat mesophyll cells. Plant cell 2005;17:3203-12.
21. Manning VA, Hardison LK, Ciuffetti LM. Ptr ToxA interacts with a chloroplast-localized protein. Mol Plant Microbe Interact 2007;20:168-77.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
185
22. Martinez JP, Oesch NW, Ciuffetti LM. Charac-terization of the multiple-copy host-selective toxin gene, ToxB, in pathogenic and nonpathogenic isolates of Pyrenophora tritici-repentis. Mol Plant Microbe Interact 2004;17:467-74.
23. Siedow JN, Rhoads DM, Ward GC, Levings CS. The relationship between the mitochondrial gene T-urf13 and fungal pathotoxin sensitivity in maize. Biochim Biophy Acta 1995;1271:235-40.
24. Sindhu A, Chintamanani S, Brandt AS, Zanis M, Scofield SR, Johal GS. A guardian of grasses: specific origin and conservation of a unique disease-resistance gene in the grass lineage. Proc Nat Acad Sci USA 2008;105:1762-7.
25. Turgeon BG, Baker SE. Genetic and genomic dissection of the cochliobolus heterostrophus Tox1 locus controlling biosynthesis of the polyketide virulence factor T-toxin. Adv Genet 2007;57:219-61.
26. Walton JD. HC-toxin. Phytochemistry 2006;67: 1406- 13.
27. Wolpert TJ, Dunkle LD, Ciuffetti LM. Host-selective toxins and avirulence determinants: what's in a name? Ann Review Phytopathol 2002;40:251-85.
28. Yang G, Rose MS, Turgeon BG, Yoder OC. A polyketide synthase is required for fungal virulence and production of the polyketide T-toxin. Plant Cell 1996;11:2139-50.
29. Yoder OC, Turgeon BG. Molecular genetic evaluation of fungal molecules for roles in pathogenesis to plants. J Genet 1996;75:425-40.


187
“ยงลาย” รายกวาทเคยคด
จนรภา โพธกสกร1 ระพ บญเปลอง2 วไล หนนภกด3 และ สภาวด ราชมณ4 1ภาควชาจลชววทยา 2ภาควชาชววทยา 3ภาควชาชวเคม คณะวทยาศาสตร มหาวทยาลยมหดล 4ศนยควบคมโรคตดตอนาโดยแมลงท 4.1 กาญจนบร
บทคดยอ ยงลายบาน (Aedes aegypti) และ ยงลายสวน (Ae. albopictus) มรปรางลกษณะคลายคลงกน และทง 2 ชนดสามารถ
นาโรคไขเลอดออก ไขเหลอง และโรคชคนกนยา มาสคนได รวมทงสามารถนาโรคพยาธฟลาเรยของสตวไดดวย นอกจากน ยงพบวายงลายสวนออกหากนไดตลอด 24 ชวโมง ขณะทยงลายชนดอนๆ ในปากสามารถนาโรคบางอยางมาสคนไดเชนกน เชนโรคทเกดจากพยาธเทาชางชนด Wuchereria bancrofti เปนตน จงควรพฒนาการวจยทางชววทยาและศกษาความสามารถในการนาโรคของยงลาย
คาสาคญ: ยงลายบาน; ยงลายสวน; พาหะนาโรค
Abstract Aedes aegypti and Ae. albopictus are morphologically similar and are vectors of dengue
hemorrhagic, yellow and chikungunya fevers in humans, and of dirofilariosis in dogs. These mosquitoes are widespread in several locations of Thailand. Ae. albopictus has been recognized as a diurnal-nocturnal feeder. Many species of mosquitoes in the genus of Aedes are not well characterized and some could act as vectors of diseases caused by filarial parasites, viz. Wuchereria bancrofti. Research on the biology of Aedes mosquitoes and the disease they transmit should be pursued.
Keywords: Ae. aegypti; Ae. albopictus; vectors
บทนา “ยง” เปนแมลงขนาดเลก มปกบางใส 1 ค รปรางบอบบาง ลาตวแบงเปน 3 สวนอยางชดเจนคอ หว อก และทอง ลวดลายทปรากฏบนตวของยงเกดจากการเรยงตวกนของเกลดขนาดเลก ซงนามาใชในการจาแนกชนดของยงได จากหลกฐานทางซากดกดาบรรพพอจะสนนษฐานไดวา ยงถอกาเนดขนในโลกตงแตปลายยค Paleozonic ซง
เปนยคดกดาบรรพประมาณ 200 ลานปมาแลว โดยเชอกนวายงกนปลอง (Anopheles) เปนยงทเกดขนกอนเปนพวกแรก ซงคณลกษณะพเศษบางประการททาใหยงสามารถปรบตวเพอการอยรอดมาไดยาวนานและเพมจานวน
มากมายถงทกวนนไดกคอ การทยงมรางกายขนาดเลก จงไมตองการอาหารมาก สามารถอาศยอยไดทกทและมความวองไวสงในการหลบหลกศตร
หากนบรวมกนทวโลก ยงมมากมายหลากหลายสายพนธและอาจนบไดถงหมนชนด อาจแบงยงตามคณสมบตในการเปนพาหะออกเปน 2 กลมใหญๆ ไดแกกลมทเปนพาหะนาโรค และกลมทไมเปนพาหะนาโรค
สาหรบยงในประเทศไทยนนพบวามถง 436 ชนด (species) จาก 23 สกล (genus) และ 48 สกลยอย (sub- genus)(20) แตทเปนพาหะนาโรคทสาคญมเพยง 4 สกลเทานน คอ ยงลาย (genus Aedes) พาหะในการนาโรคไขเลอดออก ยงราคาญ (genus Culex) พาหะนาโรคไขสมองอกเสบ ยงกนปลอง (genus Anopheles) พาหะนาโรคมาลาเรย และ ยงเสอหรอยงเหลอง (genus Mansonia) พาหะนาโรคเทาชาง เปนตน โดยทในแตละสกลทกลาวมานตางกยงประกอบไปดวยชนดตางๆ อกนบรอยชนด ดงเชน ยงในสกลของยงลายในประเทศไทยมทงสน 45 ชนด(21) และทกชนดกเรยกวา “ยงลาย” เพราะตางมสสนของเกลดบนลาตว มองเหนเปนลายขาวสลบดา
อยางไรกตาม ยงลายทเปนทรจกของคนทวๆ ไป โดยเฉพาะอยางยงในเขตชมชนสงคมเมองนนมเพยงชนด
เดยวคอ ยงลายบาน (Ae. aegypti) สรางความเดอดรอนตอชวตความเปนอยของมนษยพอสมควรโดยกอใหเกด
อาการ คน ราคาญจากการถกกดเพอดดกนเลอดของยงเพศเมย และยงสามารถนาโรครายแรงถงชวตและทรพยสนมาสผทถกกดไดอกดวย เพราะเปนตวการสาคญในการนา

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
188
โรคไขเดงกและไขเลอดออก ซงตอมานกวจยพบวายงมยงลายสวน (Ae. albopictus) อกชนดหนง ทสามารถนาโรคไขเลอดออกไดเชนกน โดยทยงชนดนเปนยงทมแหลงทอยตามชนบทและปาเขา จงนบเปนการแบงเขตทองทนาโรคแยกจากยงลายบาน
บทความนไดรวบรวมความนารเพมเตมเกยวกบ
ยงลายไว ท งจากการศกษาคนควาและจากงานวจยบางสวน เพอนาเสนอใหทราบวาแททจรงแลวยงลายยงมเรองรายๆ ทนารซอนไวอกมากมายกวาทเราเคยทราบมากอนมใชนอย จนอาจกลาวไดเลยวา “ยงลาย” รายกวาทเคยคด
ประวตของยงลาย ยงลาย Aedes aegypti พบเปนครงแรกในแถบ
เอเชยตะวนออกเฉยงใต ตงแตกอนป พ.ศ. 2443(10) หรอในรชสมยของพระบาทสมเดจพระจอมเกลาเจาอยหวรชกาล ท 4 โดยเชอกนวายงลายบาน มแหลงกาเนดในทวป แอฟรกา แลวเขาสผนแผนดนของทวปเอเชยดวยการตดมากบการขนสงระหวางประเทศ ทงทางนาและทางบก เพราะมหลกฐานในการพบยงลายในบรเวณชายฝงทะเลกอน โดยเฉพาะบรเวณทาเรอขนสงสนคา จากนนจงเรมพบลกเขามาในเขตแผนดนในเวลาตอมา(30) ในป พ.ศ.2463 เกดการระบาดของโรคไขเหลองในเขตกรงเทพมหานครฯ และจากการสารวจยงในบรเวณทาเรอขนสงสนคา พบวายงลาย Ae. aegypti เปนพาหะทสาคญในการนาโรคไข
เหลอง(9,12) ตอมาในป พ.ศ. 2480 มรายงานจากการศกษาวาพบยงลายชนดเดยวกนนในอกหลายจงหวด
นบตงแตมการระบาดของโรคไขเลอดออกเปน
ครงแรกในประเทศไทย เมอป พ.ศ. 2497(1) และการระบาดครงใหญของโรคไขเลอดออกในเขตกรงเทพมหานครฯ ในป พ.ศ. 2500(15) เปนตนมา ยง Ae. aegypti ซงไดรบการพสจนยนยนวาเปนพาหะนาโรคไขเลอดออก(23) กเปนท
รจกของคนทวไปในประเทศไทย ในชอเรยกสามญวา “ยงลาย” และดวยเหตทพบทงเชอกอโรคและตวยงเปนครงแรกในตวเมองกรงเทพฯ จงทาใหในชวงแรกๆ นกวจยคดวาโรคไขเลอดออกเปนโรคทเกดขนเฉพาะกบ
คนทอาศยอยในเขตเมองเทานน แตตอมาอกไมนานม
รายงานการพบผปวยดวยโรคไขเลอดออกหลายรายใน
ตางจงหวดทหางไกล และแมแตในหมบานเลกๆ ตามชนบท ทาใหตองมการเรมตนศกษากนอก และจากการศกษาถงเขตการแพรระบาดของโรคควบคไปกบการ
ปรากฏตวของยงลายพบวา นอกจากยงลาย Ae. aegypti ทเคยเชอวาเปนพาหะเพยงชนดเดยวแลว ยงมยงลายอก
ชนดหนง คอ Ae. albopictus ซงมลกษณะคลายคลงกนกสามารถนาโรคนไดเชนกน(7,8,22-24) ชนดหลงนพบมากในเขตพนทชนบทและปาเขา จากเหตผลดงกลาวทาใหยงลาย ตองมชอขยายความเพอแบงออกจากกนเปน 2 ชนดใหชดเจนคอ ยงลายบาน (Ae. aegypti) และยงลายสวน (Ae. albopictus) ตงแตนนมา
ยงลายสวน เชอกนวามจดกาเนดอยในเอเชย เปนยงทแพรพนธไดรวดเรว หากนเกงจนไดชอวา “ยงเสอแหงเอเชย” หรอ “Asian tiger mosquito” เพราะมความดราย กดเจบ เขาจโจมเหยอเกง มการแพรระบาดมาแลวมากกวา 20 ประเทศทวโลกในชวงเวลาเพยง 26 ป (ขอมลป ค.ศ. 1979-2004) และเชอวายงคงจะแพรระบาดไดตอไปอกอยางกวางขวางโดยยากทจะปองกนได เนองจากความทนทานของไขยงทมนมกวางไวในยางรถยนตเกาๆ ทถกทงไว หรอยางรถยนตทเกบไวเพอการคา ถงแมวานาทเคยขงอยจะแหงไปนานแลวหรอยางนนถกทงทามกลางแดด
รอนจดหรอแลงจดเพยงใดกตาม ไขของยงลายสวนสามารถท จะม ช วตอย รอดได ยงลายสวน หรอ Ae. albopictus จงถกจดไวเปน “หนงในรอยชนดของยงทพบแพรระบาดมากทสดในโลก” ชนดหนง(5)
อนกรมวธาน และการจาแนกลกษณะ ยงลาย จดอยในสกล Aedes วงศ Culicidae อนดบ Diptera ไฟลม Arthropoda ยงลายทพบในกลมประเทศ
แถบเอเชย มทงสน 24 สกลยอย ประมาณ 252 ชนด(4) ในจานวนนพบในประเทศไทย 14 สกลยอย จานวนสมาชกรวม 45 ชนด (ในวงเลบ)(20,21) ไดแก
1. Aedimorphus (9) 2. Alanstonea (1) 3. Bothaelia (2) 4. Cancraedes (2) 5. Christophersiomyia (3)
6. Diceromyia (5) 7. Edwardsaedes (1) 8. Fredwardsius (1) 9. Isoaedes (1) 10. Lorrainea (2)
11. Neomelaniconion (1) 12. Paraedes (2) 13. Scutomyia (1) 14.Stegomyia (14)
ยงลายทนาโรคตางๆ เชน โรคไขเลอดออก ไขเหลอง โรคเทาชาง และมเปนจานวนมากในประเทศไทย

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
189
นน เปนยงลายบานและยงลายสวน ทอยในสกลยอย Stegomyia โดยชอเตมตามหลกอนกรมวธาน คอ Aedes (Stegomyia) aegypti และ Aedes (Stegomyia) albopictus แตโดยทวไปในตาราตางๆ เราจะพบเพยงชอทเขยนยอไวเปน Aedes aegypti (Ae. Aegypti) และ Aedes albopictus (Ae. albopictus) เทานน
ทนาสนใจคอ ผลจากการสารวจในชวงป พ.ศ. 2550-2551 ในทองทปาสวนยาง เขตตาบลหวยเขยง อาเภอทองผาภม จงหวดกาญจนบร ของผเขยนและคณะ พบการปรากฏตวของยงลายตางๆ (เฉพาะทชอบกนเลอดมนษย) มเพยง 7 ชนด จาก 2 สกลยอย ดงตอไปน
ลาดบ สกลยอย ชนด 1 Aedimorphus vexans 2 Stegomyia aegypti
albopictus annandalei desmotes gardnerii sub species imitator pseudalbopictus
พบวา ในจานวนยงลาย 7 ชนดน ม 4 ชนด ไดแก Ae. annandalei, Ae. desmotes, Ae. imitator และ Ae. albopictus (รปท 1) เปนยงทสามารถนาโรคพยาธฟลาเรยระยะตดตอในธรรมชาต (natural vector) ไดอกดวย(6,10,18)
การจาแนกระหวางยงลายบานและยงลายสวนนน หากมองโดยผวเผนแลว จาแนกออกจากกนไดยาก เนองจากยงทงสองชนดมลกษณะทคลายคลงกน กลาวคอมลายสขาวดาบนสวนหว อก ทอง และขาคลายๆ กน (รปท 2) แตเมอมองภายใตกลองจลทรรศนแบบสามมต (stereomicroscope) หรอเลนสขยาย จะพบวา ลวดลายจากการเรยงตวของเกลดสขาวบนสวนบรเวณดานหลงของ
สวนอกของยงลายบาน เปนรปเคยว 2 เสนขนาบขางเสนตรงเสนบางๆ 2 เสน สวนลวดลายในบรเวณเดยวกนของยงลายสวนนนจะมเพยงเสนตรง 1 เสนเทานน (ดงแสดงในรปท 3)
เพอความสะดวกในการจาแนก บางครงอาจใชเพยงภาพวาดแสดงลกษณะเดนคราวๆ เมอมองภายใตกลองจลทรรศนแบบสามมต(14) กได อยางไรกตาม ยงทมลวดลายบนอกคลายคลงกบยงลายบานและยงลายสวน ยงมอกหลายชนด ดงนนในการจาแนกชนดยง จงตองใชความระมดระวงและละเอยดรอบคอบเปนอยางมาก และจาเปนตองใชรายละเอยดจากกญแจยงตามทกลาวขางตน
มาประกอบดวยจงจะสามารถสรปไดวาเปนยงชนดใด
ชววทยาของยงลาย ยงลายพบไดเกอบทวโลก แตจะพบมากในเขตรอนชนและเขตอบอน ชกชมในฤดฝน มการเปลยนรปรางแบบสมบรณ เชนเดยวกบยงชนดอนๆ วฏจกรชวต
การเจรญเตบโตแบงเปน 4 ระยะ คอ
Aedes annandalei Aedes desmotes Aedes imitator Aedes albopictus
รปท 1 ภาพถายยงลายทง 4 ชนดทพบวาสามารถนาพยาธฟลาเรยระยะตดตอในธรรมชาตได
การจาแนกชนดยง หลกในการจาแนกชนดของยงคอ การใชรปราง
ลกษณะตางๆ ลวดลายการเรยงตวของเกลดบนตวยงและลกษณะของอวยวะสบพนธเปนหลก โดยดเทยบกบกญแจยง (mosquitoes key) ตางๆ เพอการตดสนทถกตอง สาหรบในประเทศไทยนน กญแจยงทนยมและใชอยในปจจบนไดแกกญแจยงของ Rattanarithikul และคณะ(20)
1. ระยะไข (egg) ยงเพศเมยจะวางไขหลงจากการดดกนเลอดเตมทแลวประมาณ 2-3 วน จานวนไขทวางมตงแต 10-300 ฟอง ไขมลกษณะวางเดยวๆ เกาะตดตามภาชนะทมนาขง บรเวณเหนอผวนาเลกนอย ไขใหมจะมสขาว จากนนประมาณ 1-2 ชวโมงจะเปลยนเปนสดา ระยะฟกตวประมาณ 2-4 วน(3) เมอมนาทวมถง ไขจะฟกเปนตวลกนายงภายในเวลา 10 นาท ถง 2 วน แลวแตความสมสมบรณของไข( 1 8 ) และอยในทแหงไดนานเปนปในสภาพ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
190
ยงลายบาน
ยงลายสวน
รปท 2 ลกษณะรปรางโดยทวไปของยงลายบาน และยงลายสวน
ยงลายบาน (Aedes. aegypti)
ยงลายสวน (Aedes. albopictus)
รปท 3 รปรางลกษณะเฉพาะของยงลายบาน และยงลายสวน เมอมองภายใตกลองจลทรรศนแบบสามมต
แวดลอมทพอเหมาะ
2. ระยะลกนายง (larva) หลงจากฟกออกจากไข เรยกลกนาระยะท 1 (first instar larva หรอ larva stage 1) มขนาดประมาณ 0.1-0.15 ซม. เรมกนอาหาร มการเจรญ เตบโตโดยการลอกคราบทกคร งท เปลยนขนาดของ
รางกาย รวมทงสน 4 ครง การลอกคราบแตละครง จะแยกออกเปนระยะตางๆ ไดแก ระยะ 2 ระยะ 3 และระยะ 4 ตามลาดบ ใชเวลาในการเจรญเตบโตประมาณ 7-10 วน จงจะลอกคราบครงสดทายเพอเปนตวโมงหรอดกแด (pupa stage) แตเมอเลยงในหองปฏบตการ พบวาลกนายงลายสวนจากปายางพารา สามารถอยในระยะ 3 และ 4 ไดนานถง 2 เดอน และสามารถทยอยกนเตบโตเปนตวเตมวยไดเกอบทงหมด(18)
3. ระยะดกแด หรอตวโมง (pupa stage) เปนระยะทมการเปลยนแปลงรปราง ลาตวหดสนลง โดยสวนหวจะมาเชอมตอกบสวนอก และโคงงอ รปรางคลายเครองหมายจลภาค ไมกนอาหาร เคลอนไหวรวดเรว มทอหายใจ 1 คอยทสวนหว เรยก ทรมเปต (trumpets) จงมกลอยตว เอาสวนหวและหลงตดผวนาใหทอหายใจสมผสผวนา เพอใช
ในการหายใจ ระยะนใชเวลา 1-3 วน จงลอกคราบเปนตวเตมวย
4. ระยะตวเตมวย (adult stage) ยงเพศผมกจะโผลออกจากคราบดกแดกอนยงเพศเมย และเรมผสมพนธไดเมออายประมาณ 24 ชวโมง เมอยงเพศเมยโผลออกจากคราบดกแดแลว จะพรอมผสมพนธไดในเวลา 1-2 วน โดยผสมพนธเพยงครงเดยว แตจะวางไขไดหลายครงจนตลอดอายขย (ประมาณ 2 ถง 5 เดอน หรออาจมากกวานนถาจาศล) ในขณะทยงเพศผตวหนงๆ สามารถผสมพนธไดหลายครง โดยทวไปยงเพศผจะมอายประมาณ 1 สปดาห ยกเวนในกรณทมอาหารและความชนพอเหมาะ กจะสามารถอยไดนานเปนเดอนหรอมากกวานน(2, 3) ยงเพศเมยจะออกหาเหยอเพอดดกนเลอด หลงจากกนเลอดอมแลว จะเกาะพกในบรเวณทมด อบลม และอยไมไกลจากแหลงอาหารและแหลงเพาะพนธเดมเปนเวลาประมาณ 3-4 วน หลงจากนน เมอไขเจรญเตมทแลว จะบนไปหาทวางไขตามภาชนะทมนาสะอาดขงตอไป
แหลงเพาะพนธ
ในป พ.ศ. 2509 องคการอนามยโลก (WHO) ไดกอตง The Aedes Research Unit (ARU) ซงเปนหนวยงานเฉพาะกจ เกยวกบการวจยยงลายขนในประเทศไทย โดยมจดมงหมายเพอศกษาแหลงเพาะพนธและการปองกนกาจด
ยงลายทง 2 ชนดดงกลาว นกวจยผทาการสารวจไดเรมตนโดยการใชวธการของ Sheppard(25) ในการเกบลกนายง
เพยงตวเดยวจากภาชนะทพบวามนาขงอยทงในและนอก
บานเรอน รวมทงตามแหลงในธรรมชาตอนๆ(30) จากผล การวจยพบวา ยง Ae. Aegypti เปนยงทพบไดเกอบทกทองท และเกอบทกภาชนะทมนาขง แตมเปนจานวนมากในแถบชมชนเมองมากกวาในแถบชนบท สวนบรเวณปาเขานนพบนอยมากหรอไมพบเลย ขณะทยง Ae. albopictus พบไดตามชานเมอง ชนบท หรอตามปาเขา มากกวาในเขตตวเมอง(30) ผลการวจยในระยะเวลาตอมาจนถงปจจบน กยงคงยนยนผลการสารวจวา ยง Ae. aegypti มแหลงเพาะ พนธทสาคญอยในเมองมากกวาในชนบท โดยมกพบลกนายงลายบานในภาชนะขงนาทมนษยกอขน เชน ตมใสนา ถงซเมนตใสนา บอคอนกรตในหองนา จานรองขาตกบขาว กระปอง แจกน กะลามะพราว ฯลฯ ซงสวนมากจะอยภายในบรเวณบานคนและอยในทรม ขณะทมกพบ
ลกนายงลายสวน Ae. albopictus ตามแหลงทมนาขงตามธรรมชาตและอยหางจากบรเวณตวบานมากกวา เชน ในสวนผลไม สวนยาง ตามโพรงไม กระบอกไมไผ กะลามะพราว กาบใบกลวย กระปอง ถงพลาสตกทถกวาง

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
191
ทงในปา และแมแตถวยรองนายางจากตนยางทวางหงายไว อยางไรกตาม สามารถพบยงทง 2 ชนดอยรวมกนไดบอยในบางพนททอยระหวางเมองกบชนบท(3,11,13,19,26,29) โดยเฉพาะอยางยงในยางรถยนตเกาทถกทงไว สามารถพบลกนายงลายทง 2 ชนดได แตจะพบลกนายงลายสวนมากกวาลกนายงลายบานอยางมนยสาคญ(5,26, 29) อยางไรกตาม ไขของยงลายทง 2 ชนดตางกมความคงทนตอสภาพ แวดลอมไดดกวาไขของยงอนๆ โดยในสภาวะแวดลอมทเหมาะสม คอ ไมแหงแลงหรอรอนจนเกนไป ไขของยง ลายสามารถรอดชวตอยไดโดยปราศจากนานานเปนป และมรายงานวา ไขของ Ae. vexans และ Ae. sticticus มชวตรอดไดนานถง 3 ป(2)
อาหาร และพฤตกรรม
อาหารของยงลายทง 2 เพศ ไดแก นาหวานจากเกษรดอกไมและผลไมเพอใชเปนพลงงาน แตยงลายเพศเมยเกอบทกชนดยงตองการโปรตนจากเลอดมนษยหรอ
สตวเพอชวยในการเจรญของไขและเปนแหลงพลงงาน
ดวย ยกเวนยงบางชนดเชน Ae. togoi ทไมจาเปนตองกนเลอด ไขกสกไดโดยใชอาหารทสะสมไวในตวเทานน เรยกวาเปนพวก autogeny(2,3) ยงแตละชนดชอบกนเลอดตางกน พวกทชอบกนเลอดสตวเรยกวา zoophilic พวกท
ชอบกนเลอดคนเรยก anthrophilic แตยงลายเกอบทกชนดกนทงเลอดคนและเลอดสตวได จากการวจยพบวายงลายบานรอยละ 99 ชอบกนเลอดคนมากกวาเลอดสตว โดยอาจกนเลอดสตวอนๆ เชน หมและสนขไดบางในยามอดอยาก แตไมพบวากนเลอดววและเลอดไก ในขณะทยงลายสวนนน สวนใหญชอบกนแตเฉพาะเลอดคน(17)
สถานทหาอาหารของยงลายบานมกอยภายใน
บานเรอนของมนษย เพอฉวยโอกาสในการดดกนเลอด มกออกหากนอยในชวงเวลากลางวน อาจมบางในชวงยารงและพลบคา แตพบจานวนมากในชวงเวลา 09.00-11.00 น.และ 13.00-14.30 น.(1) และเมอกนอมแลว กจะเกาะพกอยตามมมมดภายในบานนนๆ เชนมมมดในหองนา ในตเสอผา มง มาน เพอรอใหไขเจรญเตบโต เรยกชวงนวา gonotrophic cycle จากนนจะบนหาภาชนะทมนาขงในบรเวณบานเพอวางไขเพาะพนธตอไป ขณะทยงลายสวนจะมแหลงทอยและหาอาหารอยตามสมทมพมไมนอกบาน เชนในสวนผลไม สวนยาง คอยดกผคนทเดนทางผานในบรเวณนนๆ และเขาโจมตเพอดดกนเลอดแลวบนหนไปไดอยางรวดเรว แหลงเพาะพนธกจะอยในบรเวณใกลเคยง สาหรบชวงเวลาออกหากนนน เคยมรายงานวา ยงลายสวนมกออกหากนเฉพาะชวงกลางวน คอระหวาง 06.00-
18.00 น. เทานน(1) แตจากผลการศกษาวจยในชวงป พ.ศ. 2550-2551 ของผเขยนและคณะพบวา ยงลายสวนออกหากนตลอด 24 ชวโมง โดยพบยงลายสวนออกหากนสงสดในชวง 13.00 น. (ชวโมงท 8) ถง 18.00 น. (ชวโมงท 13) ดงแสดงในรปท 4
การบน ย งล ายบ านม กจะบ นไปไม ไกลจากแหล ง
เพาะพนธมากนก โดยทวไปจะมรศมการบนหาอาหารประมาณ 100 เมตรจากแหลงเพาะพนธ(1) และสามารถบนไดไกลประมาณ 30-300 เมตรเทานน ในขณะทยงลายสวนบนไดไกลกวา คอประมาณ 400-600 เมตร สาหรบยงชนดอนๆ เชน ยงกนปลอง พบวามรศมการบนไดไกลระหวาง 200 เมตรถงหลายกโลเมตร(3)
การผสมพนธ
การผสมพนธตามธรรมชาต สวนใหญเกดขนในอากาศ (แตในหองปฏบตการ พฤตกรรมการผสมพนธของยงลายเกดขนไดเสมอ แมจะอยในทจากด)โดยยงเพศผจะบนวนเปนกลม เรยกวาเกด swarming โดยเฉพาะในชวง
เวลาหวคา และใกลรง ตามพมไม ทงโลง หรอบรเวณใกลกบเหยอ เชน เหนอศรษะคนเปนตน ยงเพศเมยจะบนเขาไปเพอผสมพนธ ซงสวนใหญจะผสมพนธเพยงครงเดยวโดยจะเกบนาเชอของยงเพศผไวในถงเกบ และสามารถใชไปไดตลอดชวตของมน จงมกพบวายงเพศเมยทจบไดในธรรมชาตมกมนาเชออยในถงเกบแลวเสมอ สวนยงเพศผนนสามารถผสมพนธไดหลายครงตลอดอายขย
รปท 4 แผนภมแทงแสดงชวงเวลาการออกหากนของ ยงลายสวน จากชวโมงท 1-24 (06.05-06.00น.ของวนรงขน)(18)
ความสาคญของยงลาย ยงลาย นอกจากเปนพาหะนาโรคทสาคญหลาย
โรคดงทกลาวมาแลว คนยงไดรบผลกระทบจากการถกกดโดยตรง โดยนาลายของยงจะเขาไปกระตนใหรางกายเกดปฏกรยาตอบสนอง เกดเปนผนคนทผวหนง เปนตม
บวม หลงจากโดนกดประมาณ 15-30 นาท ในกรณทแพ ผ
0
100
200
300
400
500
600
700
800
900
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
ชวโมงท
จานวนยงทพบในแตละชวโมง

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
192
ถกกดจะคนมาก และแผลทเกดจากการเกาอาจมการตดเชอแบคทเรย กลายเปนแผลพพอง เปนหนองได และยงกอความราคาญเนองจากเปนยงทหากนกลางวนซงเปนเวลา
เดยวกนกบการทางานของมนษย
สรป “ยงลาย” เปนแมลงขนาดเลกทกอปญหาใหญ
เพราะนามาซงการสญเสยทงชวตและทรพยสนของมวล
มนษยชาตทเกอบทวโลกยงหาทางกาจดใหหมดสนไป
ไมได ยงลายในประเทศไทยมประมาณ 45 ชนด แตทรจกกนเปนอยางดม 2 ชนด คอยงลายบาน (Ae. aegypti) และ ยงลายสวน (Ae. albopictus) ขณะทยงมอกหลายชนดทยงขาดผใหความสนใจทจะทาการศกษาคนควาทางชววทยา
รวมทงความสาคญในการนาโรคตางๆ แมแตบทบาทของยงลายสวนทถกจดวาเปนทรจกกนดแลวกตาม ความรตางๆ เกยวกบยงลายชนดนกยงเพงเรมพฒนาในชวงเวลา 10 กวาปทผานมาน โดยพบวา ยงลายสวนกสามารถนาโรคสาคญตางๆ ไดไมนอยไปกวายงลายบานเชนกน หรออาจจะมากกวาเสยอกหากพยาธระยะตดตอทพบในยงลาย
สวนทจบไดในปาสวนยางพาราดงกลาวขางตนเปนระยะ
ตดตอของพยาธ Wu. Bancrofti ซงกอโรคเทาชางในคนเนองจากทองททพบพยาธในยงดงกลาวเปนทองททสานก
ควบคมโรคตดตอนาโดยแมลง กระทรวงสาธารณสข จดไววาเปนเขตระบาดของโรคเทาชางชนดนมาหลายปแลว
กตตกรรมประกาศ ผลจากงานวจยในรายงานนไดรบความชวยเหลอ
ดานทนวจยจากโครงการพฒนาองคความรและศกษา
นโยบายการจดการทรพยากรชวภาพในประเทศไทย โดยสานกงานกองทนสนบสนนการวจยรวมกบศนยพนธ
วศวกรรมและเทคโนโลยชวภาพแหงชาต (BRT, TRF/ BIOTEC) และการสนบสนนจากภาควชาจลชววทยา คณะวทยาศาสตร มหาวทยาลยมหดล รวมทงความรวมมอในดานตางๆ จากเจาหนาทของสานกควบคมโรคตดตอนาโดยแมลง ท 4.1 จงหวดกาญจนบร กระทรวงสาธารณสข
เอกสารอางอง 1. บญญต สขศรงาม . ย งลายกบการแพรระบาดของโรค
ไขเลอดออก. สานกบรการวชาการ มหาวทยาลยบรพา; 2546. 2. อาคม สงขวรานนท. กฏวทยาทางสตวแพทย. ภาควชาพยาธ
วทยา คณะสตวแพทยศาสตร มหาวทยาลยเกษตรศาสตร. นนทบร: โรงพมพสหมตรพรนตง; 2538.
3. อษาวด ถาวระ: ยงพาหะ (Mosquito Vectors). ฝายชววทยาและนเวศวทยา กลมงานกฏวทยา สถาบนวจยวทยาศาสตร
ส าธารณส ข กรมวทย าศาสตร ก า รแพทย กระทรวงสาธารณสข; 2544. หนา 1-21.
4. Apiwathnasorn C. A List of Mosquito Species in Southeast Asia. Bangkok Museum and Reference Centre, SEAMEO-TROPMED National Centre of Thailand, Mahidol University. 1986. p.73.
5. Boase N. Asian tiger mosquito: current status and challenges. Int Pest Cont 2006;48:80-5.
6. Gould DJ, Bailey CL, Vongpradist S. Implication of forest mosquitoes in the transmission of Wuchereria bancrofti in Thailand. Mosquito News; 1982;42:560-4.
7. Gould DJ, Mount GA, Scanlon JE, Ford HR, Sullivan M. Ecology and control of dengue vectors on an island in the Gulf of Thailand. J Med Entomol 1970;7:499-508.
8. Gould DJ, Yuill TM, Moussa MA, Simasathien P, Rutlidge LC. An insular outbreak of dengue haemorrhagic fever. III. Identification of vectors and observation on vector ecology. Am J Trop Med Hyg 1968;17:609-18.
9. Gubler DJ. Dengue. In: Monarth T, editor. The arboviruses. Vol II. Epidemiology and ecology. Florida: CEC Press; 1988. p. 223-6.
10. Harinasuta C, Charoenlarp P, Sucharit S, Deesin T, Surathin K, Vutikes S. Studies on Malayan filariasis in Thailand. Southeast Asian J Trop Med Pub Health 1970;1:29-31.
11. Harrington LC, Scott TW, Lerdthausnee K, Coleman RC, Costero A, Clark GG et al. Dispersal of the dengue vector Aedes aegypti within and between rural communities. Am J Trop Med Hyg 2005;72:209-20.
12. Jatanasen S. Environmental manipulation and health education in Aedes aegypti control in Thailand. Bull Wld Hlth Org 1967;36:636-8.
13. Kittayapong P, Strickman D. Distribution of container-inhabiting Aedes larvae (Diptera: Culici- dae) at a dengue focus in Thailand. J Med Entomol 1993;30:601-5.
14. Mahadevan G, Cheung WH, Wamin M. Chart to the identification of Aedes (Stegomyia) group. I.M.R. Medical illustration XII. Division of Entomology Institute for Medical Research, USA; 1974.
15. Nymmannitya S. Dengue haemorrhagic fever in Thailand. Southeast Asian J Trop Med Pub Health 1987;18:291-4.
16. Pant CP, Jatanasen S, Yasuno M. Prevalence of Aedes aegypti and Aedes albopictus and observations on the ecology of dengue haemorrhagic fever in several areas of Thailand. Southeast Asian J Trop Med Public Health 1973;4:113-21.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
193
17. Ponlawat A, Harrington LC. Blood feeding patterns of Aedes aegypti and Ae. albopictus in Thailand. J Med Entomol 2005;42: 845-9.
18. Pothikasikorn J, Boonplueang R, Noonpakdee W, Ratchamanee S. Survey of diversity of mosquitoes susceptible to filarial parasites in the endemic area of Wuchereria bancrofti at the Thai-Myanmar border of Thong Pha Phum district, Kanchanaburi Province, Thailand. (unpublished data).
19. Preechaporn W, Jaroensutasinee M, Jaroensutasinee K. The larval ecology of Aedes aegypti and Ae. albopictus in three topographical areas of southern Thailand. Dengue Bulletin 2006;30:204-13.
20. Rattanarithikul R, Harrison BA, Panthusiri P, Coleman RE. Illustrated keys to the mosquitoes of Thailand. 1. Background; geographic distribution; list of genera, subgenera, and species; and a key to the genera. Southeast Asian J Trop Med Public Health 2005;36:1-79.
21. Reinert JF, Harbach RE, Kitching IJ. Phylogeny and classification of Aedini (Diptera: Culicidae), based on morphological characters of all life stages. Zool J Linn Soc 2004;142:289-368.
22. Rudnick A, Eu Tan E, Lucas JK, Omar MB. Mosquito-borne haemorrhagic fever in Malaya. Brit Med J 1965;1:1269-72.
23. Rudnick A, Hammon WM. Newly recognized Aedes aegypti problems in Manila and Bangkok. Mosquito News. 1960;20:247-9.
24. Rudnick A, Hammon WM. Entomological aspects of Thai haemorrhagic fever epidemics in Bangkok, the Philippines and Singapore 1956-1961. SEATO Medical Research Monograph No. 2. Symposium on haemorrhagic fever; 1961.
25. Sheppard PM, Macdonald WW, Tonn RJ. The single larva method. Bull Wld Hlth Org 1969;40:467-8.
26. Suwonkerd W, Tsuda Y, Takagi M, Wada Y. Seasonal Occurrence of Aedes aegypti and Ae. albopictus in used tires in 1992-1994, Chiangmai, Thailand. Trop Med 1996; 38:101-5.
27. Tiawsirisup S, Kaewthamasorn M. The potential for Aedes. Albopictus (Skuse) (Diptera: Culicidae) to be a competent vector for canine heartworm, Dirofilaria immitis (Leidy). Southeast Asian J Trop Med Pub Health 2007;38:208-14.
28. Tiawsirisup S, Nithiuthai S. Vector competent of Aedes aegypti (L) and Culex quinquefasciatus (Say) for Dirofilaria immitis (Leidy). Southeast Asian J Trop Med Pub Health 2006;37:110-4.
29. Tsuda Y, Suwonkerd W, Chawprom S, Prajakwong S, Takagi M. Different spatial distribution of Aedes aegypti and Ae. albopictus along an urban-rural gradient and the relating environmental factors examined in three villages in Northern Thailand. J Am Mosq Con Assoc 2006;22:222-8.
30. Yasuno M, Tonn RJ, Jatanasen S. Distribution of Aedes aegypti and Aedes albopictus in Thailand. I. initial survey. 1969. p. 1-10. (WHO/VBC/69; vol 157).


195
นเวศวทยาเชงอนรกษ...การเชอมโยงวทยาศาสตรกบสงคมศาสตร และ กรณศกษา ปลาเฉพาะถนทใกลสญพนธ
เจนจต คดารงสวสด ภาควชาชววทยา คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
นเวศวทยาเชงอนรกษเปนศาสตรทอาศยความรและทกษะดานวทยาศาสตรและสงคมศาสตรประกอบเขาดวยกน รวมทงตองใชขอมลภายในหองปฏบตการตลอดจนถงการเกบขอมลภาคสนาม การดาเนนงานวจยในดานนจงไมไดจากดอยแตในหองปฏบตการหรอพนทในธรรมชาตเทานน แตยงตองครอบคลมไปถงชมชนเมองทมนษยสรางขน เพราะการกระทาของมนษยทาใหเกดผลกระทบตอสงมชวตตางๆ ทงโดยทางตรงและทางออม การอนรกษสงมชวตโดยเฉพาะสตวทใกลสญพนธทพบอยในบรเวณชมชนเมองและชนบทจงจาเปนตองอาศยความรวมมอจากมนษยดวย ผลจากการดาเนนงานวจยเชงอนรกษสามารถกอใหเกดประโยชนทางดานวชาการและตอชมชนอกดวย
คาสาคญ: นเวศวทยาเชงอนรกษ; สตวทใกลสญพนธ; วทยาศาสตร; สงคมศาสตร; ชมชน
Abstract
Conservation ecology is a discipline that requires knowledge and skills from both natural and social sciences. Information obtained from both laboratory and field work is necessary. Research in this discipline is not only confined to indoor spaces or natural habitats, but also extends to human communities since anthropogenic activities directly and indirectly affect living organisms. Thus, conservation of these biotic components, especially endangered animals that co-exist with humans, unavoidably involves cooperation from humans. Results of such research can contribute to basic knowledge and also provide some benefits to mankind.
Keywords: conservation ecology; endangered species; science; social science; community
บทนา
นเวศวทยาเชงอนรกษ (conservation ecology) เรมเปนทรจกแพรหลายและมบทบาทอยางมากในปจจบน
เนองมาจากผลกระทบของการกระทาของมนษยตอ
ธรรมชาต ศาสตรนไมใชเพยงแคการศกษาดานสงแวดลอม (environment study) หรอเปนการลงพนทเพอสารวจชนดของส งม ช วตในพนทหน งๆ หรองานอนกรมวธาน (taxonomy) ตามทคนสวนมากเขาใจ การคนพบสงมชวตชนดใหมด เหมอนจะเปนแรงจงใจททาใหคนสนใจ
การศกษาธรรมชาต ถงแมวาการคนพบนเปนจดเรมตนทสาคญจดหนงของงานดานนเวศวทยา แตไมใชหวใจของนเวศวทยาดงทคนสวนมากเขาใจ
งานวจยเชงอนรกษโดยเฉพาะในสงมชวตทใกลสญ
พนธจาเปนตองอาศยความรทางวทยาศาสตร ซงจะใหขอมลเกยวกบสงมชวต ระบบนเวศ ถนทอยอาศย และอนๆ ทมความสมพนธกบสงมชวตททาการศกษา นอกจากนการ ศกษาในหองปฏบตการยงไดขอมลเชงโมเลกล สรรวทยา และววฒนาการของสงมชวตนนๆ ขอมลเหลานเปนสวนเรมตนของการอนรกษทถกวธ แตเปนทนาเสยดายทองคความรมกจะถกเกบไวโดยไมมการนามาใชประโยชนอยาง
เตมท เชนเดยวกบความรดานสงคมศาสตรซงมความสาคญตองานอนรกษ แตมกจะถกมองขามจากบคลากรสายวทยาศาสตร หากมการนาความรทงสองดานมาประยกตใชในงานอนรกษ จะชวยใหงานอนรกษดาเนนไปไดอยางมประสทธภาพมากขน

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
196
วทยาศาสตรกบงานวจยเชงอนรกษ เมอกลาวถงการอนรกษสงมชวต องคความรทาง
ชววทยามบทบาทอยางมาก โดยเฉพาะทางสายนเวศวทยาและสงแวดลอมซงจะใหขอมลทสาคญเกยวกบสงมชวต เชน โครงสรางประชากร ลกษณะถนทอยอาศย อาหาร การสบพนธ การยายถนทอย การเปลยนแปลงเชงพนท เปนตน การเปลยนแปลงขององคประกอบเหลานอาจสงผลกระทบตอสงมชวตนนๆ งานวจยในลกษณะนสวนใหญเปนงานในภาคสนาม และนกอนรกษตองทางานในสภาพแวดลอมทมตวแปรทไมสามารถควบคมได
ในการอนรกษสงมชวตทใกลจะสญพนธ การทราบจานวนประชากรทยงคงเหลออยในธรรมชาต มความสาคญตอการประเมนสถานภาพของสงมชวตนนๆ อยางไรกตาม จานวนประชากรทปรากฏอยในธรรมชาตในชวงเวลาททาการศกษา มไดบงชถงจานวนประชากรในอนาคต จงไมสามารถใชเปนแนวทางในการวางแผนงานอนรกษอยางมประสทธภาพได(1) ดงนน จงมความจาเปนทตองศกษาในระดบสรรวทยาและโมเลกลดวย ขอมลเหลานตองอาศยการวจยในหองปฏบตการ โดยอาจมการทดลองสกดสารพนธกรรมหรอสารอนทรยตางๆ จากตวอยางของสงม ช วตทศกษา ความรทไดอาจมประโยชนในการวเคราะหปญหาและอาจนาไปสการแกปญหาไดตรง
เปาหมาย
ตวอยางงานอนรกษสงมชวตทใกลสญพนธ เชน หมแพนดา เปนทรจกแพรหลาย งานสวนใหญเปนการพยายามเพมจานวนประชากรและการฟนฟถนทอยอาศย
ของหมแพนดา แมวาแผนงานอนรกษมมานานแลว แตหมแพนด าก ย งม แนวโนมท จะสญพนธ งานว จ ยของ Gittleman(6) ทศกษาลกษณะทางกายภาพและววฒนาการ
ของหมแพนดาบงชวา หมแพนดาอยในกลมเดยวกบสตวเลยงลกดวยนมประเภทกนเนอ แตหมแพนดา กลบกนแตใบไผเปนอาหารเพยงอยางเดยว นอกจากนตวออนของหมแพนดายงมอตราการเกดและการรอดชวตตา เมอรวมกบลกษณะการกนทเฉพาะเจาะจงและการทาลายปาไผโดย
ฝมอมนษย จงไมแปลกทหมแพนดามความเสยงสงตอการสญพนธในขณะทสตวอนๆ ทมสายววฒนาการใกลเคยงกนกลบมความเสยงนอยกวาเพราะมการปรบตวไดดกวา ขอมลเหลานสามารถนาไปประยกตในการบรหารจดการ
กลมประชากรของหมแพนดาทยงเหลออยใหถกวธ โดยอาจเพมการดแลตวออนในระยะแรกเปนพเศษเพอใหม
อตราการอยรอดเพมขน และใหมการอนรกษและฟนฟปาไผเพอเปนทอยอาศยของหมแพนดา เนองจากการจบหม
แพนดาหรอสงมชวตมาอยในสวนสตว หรอสถานอนรกษนน ไมเพยงพอตอการเพมจานวนประชากรแบบยงยน
อกตวอยางหนงทแสดงใหเหนถงการนาความรทาง
วทยาศาสตรมาใช ไดแก การศกษาปลาทใกลสญพนธ(4) คอ ปลา boulder darter (Etheostoma wapiti) ในวงศ Percidae ซงอยรวมกลมกบปลา darters อนๆ อก 5 ชนด ในสกลยอย Nothonotus (subgenus Nothonotus) ปลาเหลาน เปนสตวเฉพาะถนทางตอนใตของทวปอเมรกา
เหนอ จากการศกษาพบวา ปลาในกลมนสวนใหญมกจะวางไขในบรเวณแหลงนาทพนลาธารมหนขนาดใหญและ
ขนาดกลาง จงพบไดทวไปในปรมาณทมากพอสมควรตามลาธารตนนา ยกเวนปลา boulder darter ทลดจานวนลงอยางรวดเรว ขอมลดานนเวศวทยาของปลาชนดนมอยอยางจากด การศกษาเพมเตมทาไดยากเพราะจานวนประชากรมนอยและพบไมบอย แตเนองจากปลาชนดนมสายสมพนธทางววฒนาการใกลชดกบปลาอนๆ อก 5 ชนดในสก ล ย อ ย Nothonotus จ ง อ า จน า ม าอ า ง อ ง ได ว าพฤตกรรมและลกษณะทางนเวศวทยามความคลายคลงกน ซงหมายความวา boulder darter นาจะวางไขในบรเวณลาธารทมพนเปนหนขนาดใหญและขนาดกลาง เมอพจารณาถงทอยอาศยของ boulder darter ในชวงเวลาทประสบปญหากพบวากอนหนขนาดใหญและขนาดกลางไดมการ
ถกเคลอนยายจากลาธารเพอไปทาเปนแนวรวรมตลง การกระทาดงกลาวอาจเปนสาเหตใหจานวนประชากรของ boulder darter ลดลงเนองจากไมมภมสภาพทเหมาะสมสาหรบการวางไข ดงนนแนวทางทเหมาะสาหรบการอนรกษและฟนฟประชากร boulder darter จงควรเปนการปรบปรงพนทในลาธารใหเหมาะสมกบการดารงชวตของ
ปลาชนดน
ตวอย า งท งหมดข างตนแสดงได เหนถ งการ
ดา เนนการอนรกษทมแบบแผนโดยมการใชความร
ภาคสนาม ไดแก การสารวจจานวนประชากรและลกษณะทอยอาศยของสงมชวต รวมกบความรทไดจากการศกษาระดบโมเลกลของสายววฒนาการของสงมชวต จงทาใหการวางแผนมทศทางทชดเจนมากขน ดงนนการเชอมโยงความรหลายดานเขาดวยกนจะทาใหการวเคราะหขอมล
และการวางแผนการแกปญหามโอกาสสาเรจมากขน(14) อยางไรกตาม ตวอยางขางตนแสดงใหเหนไดวา ขอมลทางวทยาศาสตรเพยงอยางเดยวไมเพยงพอตอการอนรกษใน
ระยะยาว การฟนฟและปกปองถนอาศยของหมแพนดาตองใช ก าล งทรพย และก าล งคนอย างมหาศาล ซ งจาเปนตองอาศยความรวมมอจากหลายฝาย เชนเดยวกบการฟนฟสภาพแหลงนาเพอใหเหมาะสมตอการดารงชวต

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
197
ของ boulder darter กตองใชความรวมมอของผคนทอาศยอยบรเวณลาธาร การทาใหมวลชนกลมนเขาใจถงความ สาคญและใหความรวมมอในการอนรกษสงมชวตจง
ตองการขอมลดานอนๆ นอกเหนอจากวทยาศาสตร
สงคมศาสตรกบงานวจยเชงอนรกษ ความรทางสงคมศาสตรไมวาจะเปนดานรฐศาสตร
มนษยวทยา เศรษฐศาสตร สงคมศกษา กฎหมาย ปรชญาและศาสนา มความสาคญตอการอนรกษสงมชวตไมนอยกวาขอมลทางวทยาศาสตรเพราะเปนการศกษาความ สมพนธระหวางหนวยตางๆ ของสงคมรวมทงพฤตกรรมของมนษยในแงมมของมนษยดวย เนองจากงานอนรกษมความสมพนธและสาคญตอชมชนมนษย โดยเฉพาะอยางยงปญหาเรองการสญพนธของสงมชวตสวนใหญมสาเหต
มาจากการกระทาของมนษย(5) ไมวาจะเปนการทาลายถน อาศยของสงมชวต การทาใหเกดมลพษตอสงมชวต การหาประโยชนเกนควร การนาสงมชวตตางถนไปปลอยในททไมใชถนเดม เปนตน(13) ซงการกระทาเหลานมกจะมสาเหตมาจากปรมาณประชากรมนษยทเพมขนอยางรวดเรวและ
ความตองการทจะใชทรพยากรธรรมชาตเพอการอยรอด
ของตนและครอบครว หรอในบางกรณอาจเปนความมกงายและความตองการทไมมจดสนสด
ตวอยางงานวจยเชงอนรกษสายสงคมศาสตร ไดแก การอนรกษชะนมอขาวในลมนาลาง จงหวดแมฮองสอน การศกษาจานวนประชากรชะนมอขาวทเหลออยระหวางป ค.ศ. 2004-2005 พบวา บรเวณทประชากรชะนมความหนาแนนสงอยใกลกบชมชนกะเหรยงทม วฒนธรรม
อนรกษชะน(18) การศกษาบทบาทของชมชนชาวเขาใน
จงหวดแมฮองสอนยนยนเรองน เพราะชาวเขาเหลานเชอวาการบรโภคเนอชะนจะทาใหประสบโชคราย(17) จงไมนยมลาชะน งานวจยนแสดงใหเหนถงพลงของชมชนในการอนรกษสงมชวต อยางไรกตาม สงทนาเปนหวงคอความเชอเหลานคอยๆ หมดไป ในขณะเดยวกนวฒนธรรมของสงคมเมองทตองการอยรอดโดยตกตวงประโยชนจาก
ทรพยากรธรรมชาตใหมากทสดเรมเพมขน การลาชะนเพอบรโภคและการคารวมทงการทาลายปาเพอเพาะปลกทาให
การอนรกษ เปนไปอยางยากลาบากมากขน กฎหมายคมครองสตวปาไมสามารถบงคบใชไดอยางมประสทธ- ภาพในพนทดงกลาว ดงนน แผนการอนรกษชะนใน กรณนจงจาเปนตองอาศยชมชนในการสรางและฟนฟ
คานยมเกยวกบการอนรกษชะนและปาใหแกคนรนใหม
ตวอยางอนๆ พบไดจากงานอนรกษชางปาซงเปนสตวคบานคเมองของไทยมานาน ปจจบนชางปาประสบ
ปญหาจานวนประชากรทลดลงอยางรวดเรวเพราะมการลา
ชางเพอการทองเทยว เพอนางาไปจาหนาย และการขาดแคลนพนทหากนเนองจากถกรกรานพนทปาเพอการ
เพาะปลก การแกปญหาการลดลงของจานวนประชากรยงไมประสบความสาเรจเทาทควร ขอมลดานวทยาศาสตรเกยวกบชางทงในระดบโมเลกล สรรวทยา กลมประชากร และพฤตกรรมนนมอยจากด จงยงตองใชเวลาพอสมควรในการศกษาและเกบขอมล แตการอนรกษไมสามารถรอวทยาศาสตรได เพราะคนกบชางปามความสมพนธใกลชด เนองจากอาศยอยในแหลงหรอบร เวณเดยวกน และถงแมวามการบงคบใชกฎหมายอนรกษสตวปาแลว แตกไมเพยงพอเพราะไมมการระบถงการอนรกษทอยอาศยของ
ชาง เมอพนทหากนของชางลดลงจงเกดปญหาชางปาเขามาในเขตชมชน ทาลายพชผลการเกษตร(15) เปนปญหาเรอรงมายาวนานกวา 10 ป แตยงไมสามารถดาเนนการแกไข
ระยะยาวได การแกปญหาเฉพาะหนาคอการไลชางออกจากบรเวณพนทเพาะปลกโดยมการวางยามเฝาระวง การใชอปกรณทไม เปนอนตรายถงชวต เชน การฉายไฟสปอรตไลท การจดประทด การเปดสญญาณไซเรนเพอใหชางตกใจไมกลาเขาใกลพนทการเกษตร เปนตน(12) ซงเปนเพยงการแกปญหาชวคราวเทานน และยงทาใหสนเปลองกาลงงานมากขนโดยชาวบานตองเสยสละเวลาและ
แรงงานมาเปนอาสาสมครเฝายาม การยายททากนหรอปลกพชทไมใชอาหารชางกเปนการแกปญหาทไมตรงจด เพราะการยายทเพาะปลกอาจนาไปสปญหาเดมในทแหง
ใหม สวนชางปากยงคงตองออกจากปาเพอหาอาหารดงเดม ดงนนการฟนฟสภาพปาใหเหมาะแกการอยอาศยของชางนาจะเปนทางเลอกทดกวา การแกปญหาระยะยาวจาเปนตองอาศยขอมลทางวทยาศาสตรมาเปนแนวทางใน
การวางแผน ในขณะเดยวกนควรใหชมชนมสวนรวมในการแกปญหาดวย รวมทงสรางจตสานกในการอนรกษสงมชวตคบานคเมองและการดแลรกษาปาควบคกนไป
ตวอยางขางตนแสดงใหเหนวางานดานการอนรกษ
มความจาเปนทตองอาศยพลงและความรวมมอจากชมชน
ในการด า เนนงาน และศาสตรสาขาต างๆ ในสายสงคมศาสตร ลวนมเครองมอทสามารถนามาใชวเคราะหและสรางองคความรในการอธบายพฤตกรรมของมนษยทง
ในระดบทองถน ระดบชาต และนานาชาต(11) โดยเฉพาะในการวางแผนการจดการพนทอนรกษรวมไปถงการปฏบต
จรง เนองจากนกวชาการสวนใหญไมไดเปนผปฏบตและไมไดอยในพนทตลอดเวลา การปลกฝงใหคนทองถนและผทมสวนเกยวของรกและรจกรกษาทรพยากรธรรมชาตจง
มความสาคญตอการอนรกษแบบยงยน โดยมนกวชาการ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
198
รวมในการสรางองคความร ผลตสอ และถายทอดความรแกชมชนเพอนาไปใชปฏบต ในขณะเดยวกนควรมการ
วางรากฐานการศกษาเกยวกบธรรมชาตใหนาสนใจเพอคน
รนใหมจะไดเขาใจ มจต สานกทดตอสงมชวตและตอการอนรกษทรพยากรธรรมชาต
กรณศกษา: การใชวทยาศาสตรและสงคมศาสตรในการอนรกษปลาเฉพาะถนในประเทศสหรฐอเมรกา
การศกษาปลาเฉพาะถน (endemic species) ทกาลงใกลสญพนธ ทชอวา Vermilion darter (Etheostoma chermocki) ปลาชนดนอยในกลมสกลยอย Ulocentra (subgenus Ulocentra) รวมกบปลาอนๆอกประมาณ 20 ชนด(3) ถนอาศยของ Vermilion darter พบในลาธารเพยงแหงเดยวในชวงเพยง 11.6 กโลเมตรนอกเมองเบอรมง- แฮม มลรฐอลาบามา ประเทศสหรฐอเมรกา การศกษาการเปลยนแปลงของจานวนปลาในชวงป ค.ศ. 1995-1997 พบวาจานวนลดลงอยางรวดเรวจนถงขนทไมพบปลาชนด
นในบรเวณทเคยพบเปนประจา(2) ดงนน ในป ค.ศ. 2001 Vermilion darter จงไดถกขนทะเบยนคมครองเปนสงม ชวตทใกลสญพนธ โดยหนวยงาน United States Fish and Wildlife Service ในเบองตน หนวยงานนมแผนการซอทดนของเอกชนในบรเวณรอบลาธารทเคยพบ Vermilion darter แตเนองจากไมมงบประมาณเพยงพอและการขาด
แนวทางในการอนรกษและฟนฟประชากรแบบยงยน ทางหนวยงานจงขอความรวมมอจากมหาวทยาลยอลาบามาใน
การวางแผน ซงงานอนรกษและฟนฟประชากรปลาชนดนจาเปนตองอาศยขอมลทางชววทยาเพอเปนแนวทาง แตไมพบวามขอมลบนทกไวมากนก จงตองมการศกษาเพมเตม(8)
เน องจากปลาชนดน ได รบการค มครองตาม
กฎหมายแลว จงไมสามารถไปเกบตวอยางจากแหลงนาธรรมชาตได จงมการนาตวอยางจากพพธภณฑมาใชศกษา แตจานวนทมในพพธภณฑยงคงไมเพยงพอตอการศกษา จงตองอาศยขอมลจากการศกษาปลาชนดอนทมสาย
สมพนธทางววฒนาการใกลชดกน(7) จากขอมลความสม- พนธทางววฒนาการของปลากลมนระบวา Warrior darter เปนปลาทมความใกลชดทางววฒนาการกบ Vermilion darter มากทสด จงนาจะใหขอมลทใกลเคยงมากทสด Warrior darter พบไดทวไปตามลาธารสามแหงในมลรฐ อลาบามาเชนกน(16) ซงหนงในสามนเปนลาธารคขนานกบลาธารทเปนแหลงอาศยของ Vermilion darter ดงนน ปลา Warrior darter จากลาธารนจงถกสมตวอยางเพอนามา
ศกษาเพราะอย ในสภาพภมประเทศและภมอากาศท
เหมอนกน นอกจากนตวอยางของ Warrior darter ทเกบรกษาไวในพพธภณฑกถกนาใชประกอบการศกษาดวย
ผลการศกษายนยนวา ลกษณะของ Vermilion darter ไมแตกตางจาก Warrior darter และปลาชนดอนๆ ทอยในสายววฒนาการใกลเคยงกนอยางมนยสาคญ(10) แตสงทแตกตางกนอยางชดเจนคอสภาพของถนอาศยของ Vermilion darter ทถกคกคามอยางหนกจากการพยายาม “พฒนา” พนทโดยการสรางถนน สะพานและศนยการคาผลของการพฒนาคอ ทอยของปลาชนดนเสอมสภาพลงเพราะมตะกอนและเศษขยะเปนจานวนมากไหลลงสลาธาร
ทาใหเกดการตนเขน บรเวณรมตลงทเคยปรากฏพชนา
หลายชนดกถกทาลาย ซงสงผลโดยตรงตอการดารงชวตและแพร พนธ ส ง เหล าน น าจะ เปนสา เหตท ท า ให Vermilion darter มแนวโนมทจะสญพนธในเวลาอนใกล ในขณะ เดยวกนทอยอาศยของ Warrior darter ในบรเวณใกลเคยงยงคงอยในสภาพทเหมาะสมตอการดารงชวต
อยางไรกตาม การศกษาความหลากหลายทางพนธ กรรมของ Vermilion darter และ Warrior darter โดยใช microsatellite DNA(9) พบวาปลาทงสองชนดมปรมาณ
ความหลากหลายทางพนธกรรมใกลเคยงกบปลานาจด
ทวไปทไมไดถกคกคาม จงอาจกลาวไดวา มโอกาสสงทจะฟนฟและอนรกษ Vermilion darter(7) แตจาเปนตองอาศยความรวมมอจากรฐและชมชนบรเวณนนในการดแลรกษา
ลาธารแหงนนใหอยในสภาพทสมบรณ นอกจากนการตรวจสอบจานวน โครงสรางประชากร และความหลาก หลายทางพนธกรรมของ Vermilion darter เปนระยะๆ จะทาใหสามารถประเมนแผนงานอนรกษได โดยเปาหมายจานวนประชากรทควรมเพอใหปลาชนดนคงอยไดโดยไม
ตองเสยงกบการสญเสยความหลากหลายจากความไมแน นอนทางพนธกรรม (genetic drift) สามารถคานวณไดจากขอมลทางพนธกรรม ตวเลขทไดจากการคานวณจะถกใชเปนตวอางองของสถานภาพของกลมประชากร แตตวเลขนเพยงอยางเดยวไมไดเปนตวบงบอกถงความสมบรณของ
สงมชวตหากแตเปนเพยงการประมาณในกรณทไมมดชน
ทสามารถนามาใชในการประเมนสถานภาพของสงมชวต
ได การตรวจสอบสถานภาพของสงมชวตนนควรดจากลกษณะอนดวย นอกเหนอไปจากจานวนประชากรและความหลากหลายทางพนธกรรม เชน ลกษณะทางกายภาพ อตราการเจรญเตบโต ขนาดของตวเตมวย เปนตน
การไดมาซงขอมลเหลานตองไดรบความรวมมอ
จากชมชน การเกบตวอยางมาศกษาจาเปนตองผานเขาไปบรเวณทดนสวนบคคล แมวาพนนาจะเปนของรฐกตามแต

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
199
เอกชนสามารถถอครองทดนบรเวณรมตลงได ถาปราศจากคาอนญาตใหผานเขาไปบรเวณทสวนบคคล งานวจยกไมอาจทาสาเรจได เอกชนบางรายไดปฏเสธการอนญาตใหเขาพนทสวนบคคล เนองจากเกรงวาการสารวจพบสงมชวตทไดรบการคมครองตามกฎหมายอาจนาไปส
ความยงยากในการใชทดนสวนตวเพราะตองมการยนเรอง
ตอศาลใหพจารณาอนมตการดาเนนกจกรรมใดๆ ทอาจสงผลกระทบตอสงมชวตในทดนของตนเองได อยางไรกตาม ยงมผคนอกจานวนหนงทใหความรวมมอเปนอยางดเพราะเขาใจถงความสาคญของการอนรกษ
ขอมลจากการศกษาลกษณะการดาเนนชวตของ Vermilion darter ถกนาไปแสดงเปนหลกฐานตอศาลยตธรรมเรองการพจารณาอนมตโครงการกอสรางบาน
จดสรรขนาดกลางทกาลงจะมขนบนพนทตดรมตลงของลา
ธารท Vermilion darter อาศยอย งานกอสรางโครงการนทาใหชาวบานทอาศยอยบรเวณโดยรอบไมพอใจอยแลว เนองจากเกรงวาอาจกอใหเกดปญหาดานการจราจรและ
มลพษทางนา อากาศ และเสยง โดยทฝายผกอสรางไมไดวางระบบการจดการเรองเหลานไว การยกเรองกฎหมายคมครองสตวทกาลงจะสญพนธ ซงอาจไดรบผลกระทบ
จากการกอสราง ทาใหศาลตองพจารณาอยางรอบคอบ โครงการกอสรางบานจดสรรเปนโครงการทชวยให
ประชากรไดมทอยอาศยเปนของตนเอง แตผรบเหมากควรเพมความระมดระวงในการกอสรางไมใหสงผลกระทบตอ
ลาธาร ซงขอเสนอแนะกไดถกยนใหมการพจารณาดวย ศาลจงส งใหมการสอบพยานหลกฐานอกคร ง ฝ ายผรบเหมากอสรางไมพอใจและไมเชอถอขอมลจากงานวจย จงไดวาจางบรษทเอกชนใหทาการสารวจบรเวณดงกลาว บรษทนไดทาการสารวจ แตในรายงานไมปรากฏวามสตวทใกลสญพนธอาศยอยในลาธาร การสารวจจงถกทาอกครงโดยทมวจยจากมหาวทยาลยในจดเดยวกบทบรษททา แตเนองจากเวลาไดผานมาระยะหนง ปลาไดเปลยนแหลงหากนตามฤดกาล ณ จดทมการสมตวอยาง นาในลาธารไดแหงจนเกอบหมดซงกเปนปกตตามธรรมชาตของลาธารใน
แถบ น อณหภมของน า เพ มข นส ง เกนกว า ระดบท Vermilion darter จะอาศยอยได อยางไรกตาม หางไปเพยง 200-300 เมตร ลาธารกมความลกมากขน พนทเรมมตนไมอยรมตลงเปนจานวนมาก อณหภมของนาในจดนตากวาจดทบรษทไปสารวจเกอบ 10oC ซงมผลตอการดารงชวตของสตวอกหลายอยาง แมวาจะไมพบ Vermilion darter แตกพบปลาอกชนดหนงทไดถกขนทะเบยนคมครองอาศยอย
ในลาธารเชนกน ดงนนขอมลนจงถกนาไปใหศาลพจารณา พรอมทงเสนอใหทางผรบเหมากอสรางและบรษททศกษา
สงแวดลอม ไปสารวจแหลงนาอกครง ในทสดศาลมมตใหผรบเหมากอสรางปฏบตตามขอเสนอแนะดงกลาวเพอ
ปองกนไมใหเกดผลกระทบตอลาธาร หลงจากนน 3-4 เดอนโครงการกอสรางกถกยกเลก เนองจากผรบเหมาไมสามารถปรบโครงการกอสรางใหสอดคลองกบขอเสนอได ดงนนพนทนนจงถกขายใหแกรฐเพอใชสรางโรงเรยนแทน ซ งการกอสร างนสามารถปรบไม ให เกดผลเสยตอ
สภาพแวดลอมได นอกจากน ประชาชนในชมชนนนมความยนดทจะใหมการสรางโรงเรยนมากกวาอาคารทอย
อาศย เพราะลกหลานจะไดมสถานศกษาใกลบานเปนการลดปญหาการจราจรและปญหาสงคมอนๆ
งานวจยชนนมทงสวนทเปนการสรางองคความร
ทางวชาการและไดประสบการณการนาความรไปใชใหเกด
ประโยชนตอชมชน การทางานดานอนรกษทดนนไมสามารถดาเนนการไดเพยงลาพงดวยตวคนเดยว ตองอาศยทมงานทสามารถเขากนไดด และจาเปนตองมความรความสามารถหลายๆ ดานนอกเหนอจากทไดรบจากในหองเรยน เชน ความรดานนตศาสตร ในหลายประเทศรวมทงประเทศไทย มกฎหมายคมครองสตว แตยงไมมการบงคบใชกฎหมายอยางจรงจง โดยเฉพาะในประเทศทกาลงพฒนาและพงพาทรพยากรธรรมชาต เปนหลกมกให
ความสาคญกบการใชประโยชนจากทรพยากรธรรมชาต
มากกวาการอนรกษระบบน เวศและสงม ช วต แตในประเทศทพฒนาแลว อยางเชน สหรฐอเมรกา มการบงคบใชกฎหมายคมครองสตวอยางจรงจง และมกจะไดรบการพจารณาเปนพเศษเมอถกอางอง ดงนนงานอนรกษในประเทศทกาลงพฒนาทมแนวโนมประสบผลสาเรจจงควร
มการประนประนอมระหวางนกวชาการทตองการปกปอง
รกษาทรพยากรธรรมชาตและชมชนทตองการอยอาศย
อยางสงบและยงยน เพอใหเกดประโยชนแกทงสองฝาย
สรป นเวศวทยาเชงอนรกษเปนศาสตรทเกยวของกบการ
ศกษาความสมพนธระหวางสงมชวตและสภาพแวดลอม
ซ งมการ เปล ยนแปลงอยตลอดเวลา การจะอนรกษทรพยากรเหลาน ใหคงอย ตองอาศยความรทางดานวทยาศาสตรและสงคมศาสตร ตองมการทาวจย เกบขอมล และใชขอมลให เปนประโยชน เปนแนวทางในการแกปญหา ในขณะเดยวกนความรดานสงคมศาสตรทาใหงานอนรกษเกดขนได การใหความรแกชมชนเพอใหคนในชมชนมสวนรวมในการอนรกษ รวมทงการบงคบใชกฎหมายในการอนรกษสตวใกลสญพนธ เชน หมแพนดา ชางปา ชะนมอขาว และปลานาจดหลายชนด จะทาให

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
200
ระบบนเวศคงอยอยางยงยนและเกดประโยชนรวมกนทง
ฝายมนษยและสงแวดลอม
กตตกรรมประกาศ
ผนพนธขอขอบคณทมวจย (Dr. Bernie Kuhajda, Mr. Micah G. Bennett, Mr. J. Heath Howell และสมาชกของ Harris Laboratory) มหาวทยาลยอลาบามา ประเทศสหรฐอเมรกาทรวมแรงรวมใจในการทางานอนรกษปลา
เฉพาะถนทกาลงจะสญพนธ นอกจากนยงขอขอบคณ รศ.สมโภชน ศรโกสามาตร และสมาชกหอง N505 สาหรบโลกทศนในเรองชมชนกบงานอนรกษ
เอกสารอางอง 1. Abrams PA. Will small population sizes warn us of
impending extinctions? Amer Nat 2002;160:293-305.
2. Blanco CC, Mayden RL. Status of the Vermilion darter, Etheostoma chermocki, in Turkey Creek, Jefferson County, Alabama, 1996-1997. Final Report. Montgomery (AL): Dept. of Conservation and Natural Resources; Jackson (MS): United States Fish and Wildlife Service (US); 1997.
3. Boschung HT, Mayden RL, Tomelleri JR. Etheostoma chermocki, a new species of darter (Teleostei: Percidae) from the black warrior river drainage of Alabama. Bull Alabama Mus Nat Hist 1992;13:11-20.
4. Brooks DR, Mayden RL, McLennan DA. Phylo-genetics and conservation. Trends Ecol Evolut 1992; 7:353.
5. Carson RL. Silent Spring. New York (NY): Houghton Mifflin; 1962.
6. Gittleman JL. Are the pandas successful specialists or evolutionary failures? BioSci 1994;44:456-64.
7. Khudamrongsawat J. Comparative study of life history and population structure of the endangered Vermilion darter (Etheostoma chermocki) and its sister species, the Warrior darter (E. bellator) [Dissertation]. Tuscaloosa (AL): Univ Alabama; 2007.
8. Khudamrongsawat J, Arrington DA, Kuhajda BR, Rypel AL. Life history of the endangered Vermilion darter (Etheostoma chermocki) endemic to the Black Warrior river system, Alabama. J Fresh Ecol 2005; 20:469-77.
9. Khudamrongsawat J, Heath LS, Heath HE, Harris PM. Microsatellite DNA primers for the endangered
Vermilion darter, Etheostoma chermocki, and cross-species amplification in other darters (Percidae: Etheostoma). Mol Ecol Notes 2007;7:811-3.
10. Khudamrongsawat J, Kuhajda BR. Life history of the Warrior darter (Etheostoma bellator) and comparison with the endangered Vermilion darter (E. chermocki). J Fresh Ecol 2007;22:241-8.
11. Mascia MB, Brosius JP, Dobson TA, Forbes BC, Horowitz L, McKean MA et al. Conservation and the social sciences. Conserv Biol 2003;17:649-50.
12. Noonto B. Managing human-elephant conflict: a case study at Thong Pha Phum National Park, Kanchanaburi Province, Thailand. In: Balmford A, Green R, Trevelyan R, Prior S, editors. Student Conference on Conservational Science; 2008 Mar 25-27; University of Cambridge, United Kingdom; 2008.
13. Primack RB. Essentials of conservation biology. 4th ed. Sunderland (MA): Sinauer Associates, Inc; 2006.
14. Soulé ME. What is conservation biology? A new synthetic discipline addresses the dynamics and problems of perturbed species, communities, and ecosystems. BioSci 1985;35:727-34.
15. Stewart-Cox B, Ritthirat J, editors. Mitigating human-elephant conflict in Asia: a new initiative in Thailand. In. Mitigating Human-Elephant Conflict: Case Studies from Africa and Asia. 2007. p. 13-22.
16. Suttkus RD, Bailey RM. Etheostoma colorosum and Etheostoma bellator, two new darters, subgenus Ulocentra, from southeastern United States. Tulane Studies Zool Bot 1993;29:1-28.
17. Yimkao P, Naksamrit J, Srikosamatara. Roles of communities and impassioned individuals in conservation of gibbons (Hylobates lar L.) in upper Mae Hong Son Province, northern Thailand. Nat Hist Bull Siam Soc (in press).
18. Yimkao P, Srikosamatara S. Ecology and site-based conservation of the white-handed gibbon (Hylobates lar L.) in human-use forests in Mae Hong Son Province, northern Thailand. Nat Hist Bull Siam Soc 2006;54:109-38.

201
แบบจาลองทางคณตศาสตรสาหรบระบบนเวศวทยา
กรกนก บญวงษ ภาควชาคณตศาสตร คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
พลศาสตรประชากรอธบายไดวา ประชากรในรนหนงจะเปลยนแปลงไปเปนประชากรในอกรนหนงอยางไร องค ประกอบพนฐานทเกยวของกบการเปลยนแปลงของประชากรทงหมดสามารถแบงไดเปน 5 องคประกอบ ดงน การเกด การตาย อตราสวนเพศ โครงสรางอาย และการกระจายตว แตทงหมดนจะไรความหมายทนท ถาไมนาเรองของเวลามารวมพจารณาดวย ไมวาจะเปนเวลาแบบวยตหรอเวลาแบบตอเนองกตาม ดวยเหตน กระบวนการศกษาพลศาสตรประชากรจงมประโยชนมากกวาการศกษาเฉพาะเวลาใดเวลาหนง และในหลายกรณ สงมชวตแตละหนวยมอนตรกรยากบสงมชวตหนวยอนเปนจานวนจากด (เฉพาะทเปนเพอนบาน) จงทาใหปรากฏการณทางนเวศวทยาเชงพนท อนเนองมาจากพนท การเฟนสม และความวยตของสงมชวตแตละหนวยมบทบาทสาคญ เนองจากไมสามารถจาลองไดดวยระบบแบบคาเฉลย ดงนน จงควรมการพฒนาแบบจาลองเชงพนทโดยปรยายและแบบจาลองเชงพนทชดแจง โดยใชวธการทงแบบเชงตวเลขและเชงวเคราะห
คาสาคญ: พลศาสตรประชากร; แบบจาลองเชงพนทโดยปรยาย; แบบจาลองเชงพนทชดแจง; การประมาณแบบคาเฉลย; การประมาณแบบค
Abstract
Population dynamics describe how one generation can change to the next generation. All changes in population can be divided into five basic components namely birth, death, sex ratio, age structure and dispersal. These processes become meaningless when they are divorced from their continuous or discrete temporal context. Therefore, population dynamics provides a more useful picture rather than single snapshots. Moreover, in many circumstances individuals only interact with a limited number of other individuals (their neighbors), and then spatial ecological phenomena due to space, stochasticity and discreteness of individuals play a key role. This cannot be modeled by mean-field systems and spatially implicit and explicit models need to be developed using both numerical and analysis approaches. Keywords: population dynamics; spatially implicit model; spatially explicit model; mean-filed and pair approximation
บทนา
แบบจาลองทางคณตศาสตร คอ การนาเซตของสมการและสตรมาใชจาลองระบบทสนใจ หรออกนยหนง แบบจาลองทางคณตศาสตร กคอ การอธบายระบบทสนใจในรปของตวแปร ตวแปรเสรมหรอพารามเตอร และคาคงทซงทงหมดนลวนเปนตวแทนคณสมบตบางอยาง
ของระบบนนเอง(7) แบบจาลองทางคณตศาสตรทดตองสามารถดลระหวางความยงยากซบซอนของระบบในโลก
แหงความเปนจรงททาใหยากตอการแกสมการ และการลดทอนความซบซอนในแบบจาลอง ซงทาใหงายและสะดวกตอการแกสมการได แบบจาลองทางคณตศาสตรสามารถแบงประเภทไดหลากหลายรปแบบ ทงนขนอยกบ
คณสมบตของระบบและเครองมอทใชในการศกษา เชน
แบบจาลองพลศาสตร (dynamic models) หรอแบบ จาลองสถต (static models) ซงใชการเปลยนแปลงของเวลามาพจารณา โดยแบบจาลองสถตใชอธบายระบบ ณ เวลาใดเวลาหนง ในขณะทแบบจาลองพลศาสตรเหมาะทจะใชอธบายระบบทเวลามการเปลยนแปลง Prévost และคณะ(13) ไดนาทงแบบจาลองพลศาสตรและแบบจาลองสถตมาใช
คะเนความชกชมของปลากอนทจะทาการประมง
แบบจาลองทเวลาเปนแบบวยต (discrete time models) หร อแบบจ าลองท เ วล า เปนแบบต อ เน อ ง (continuous time models) ซงใชรปแบบของการเปลยน แปลงของเวลาเปนตวพจารณา โดยเวลาทใชในแบบจาลองแบบเวลาวยตนน จะเพมขนทละขน หรอจะมองเวลาในรป

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
202
ของจานวนเตมกได สวนเวลาทใชในแบบจาลองแบบเวลาตอเนองนนจะอยในรปของจานวนจรง ตวอยางเชน ประชากรทมฤดการผสมพนธชดเจน เชน ดอกทานตะวน ซงใหเมลดในชวงเวลาใกลเคยงกน หากผานพนชวงเวลาดงกลาวนไปแลว กจะไมมการใหกาเนดเมลดอก จนกวาจะถงฤดการผสมพนธอกครงหนง ดงนนจานวนประชากรจงสามารถพจารณาในรปแบบเปนรนๆ เชน รนทวด รนปยาตายาย รนพอแม รนลก และรนหลานได ซงเหมาะทจะนาแบบจาลองแบบเวลาวยตซงนยมเขยนอยในรปของ
สมการเชงผลตางมาใชอธบายได อยางไรกตาม ถาประชากรมลกษณะเปนแบบไมมฤดการผสมพนธทชดเจน เชน คน แตละคนสามารถใหกาเนดลกหลานเมอใดกได ดงนน จงจาเปนตองนาจานวนประชากรมาพจารณา
ตลอดเวลา โดยเวลาอยบนเสนจานวนจรง แบบจาลองท
เหมาะสมจงเปนแบบจาลองแบบเวลาตอเนองซงนยม เขยนอยในรปของสมการเชงอนพนธ Nedorezov และ Nedorezova(11) ไดทาการศกษาแบบจาลองทงสองในเชง
เปรยบเทยบ
แบบจาลองเชงกาหนด (deterministic models) หรอ แบบจาลองสโทแคสตก (stochastic models) ใชสถานะของระบบ ณ เวลาถดไปเปนตวพจารณา โดยสถานะของระบบ ณ เวลาถดไปในแบบจาลองเชงกาหนดจะขนอยกบสถานะของระบบ ณ เวลาปจจบนและเวลาในอดต ดงนน ภายใตคาตวแปรเสรมชดเดยวกน ผลทไดจากการคานวณแบบจาลองจะเหมอนกนทกครง ไมวาจะทาซาอกกครงกตาม ในขณะทสถานะของระบบ ณ เวลาถดไป สาหรบแบบจาลองสโทแคสตกสามารถเปนไปไดหลายรปแบบ
เพราะการเฟนสม ดงนน การคานวณซาอาจใหผลท
แตกตางกนได ทงนแบบจาลองเชงกาหนดเหมาะสมกบการอธบายพฤตกรรมของประชากรทมขนาดใหญ สวนแบบจาลองสโทแคสตกเหมาะสมกบการอธบายพฤตกรรม
ของประชากรทมขนาดเลก West และ Thompson(18) ไดสรางแบบจาลองทงสองขนเพอใชอธบายการแพรระบาด
อยางงาย และเพอศกษาในเชงเปรยบเทยบ
นเวศวทยา (ecology) มรากศพทมาจากคาในภาษากรก คอ oikos หมายถง บาน และ logy หมายถง การศกษา ดงนน นเวศวทยา จงหมายถงศาสตรทศกษาถงความ สมพนธระหวางสงมชวตและสงแวดลอมรอบตว รวมทงอนตรกรยาทสงมชวตแตละหนวยมตอกนดวย โดยนก สตววทยาชาวเยอรมน ชอ Ernst Haeckel เปนคนแรกทนาค าน มาใช ในป พ .ศ . 2413 Hofbauer( 5 )และ Rough-garden(15) แบงการศกษานเวศวทยาออกเปน 3 ระดบ ดงน
1. ระดบรายหนวย รวมถงนเวศวทยาทางสรรวทยา ซงเปนการศกษาเกยวกบการแลกเปลยนระหวางสงมชวต
แตละหนวยและสงแวดลอมรอบตว โดยเฉพาะอยางยง การศกษากลไกทสงมชวตแตละหนวยใชเพอความอยรอด
ของตนเอง ไมวาจะเปนกลไกทางพฤตกรรมหรอทางสรรวทยา
2. ระดบประชากร เปนการศกษาเกยวกบการเตบโตและอนตรกรยาของประชากรทงหมดเมอเวลาแปร
เปลยนไป โดยประชากรทสนใจอาจมเพยงกลมเดยวหรอหลายกลมกได
3. ระดบระบบนเวศหรอชมนมสงมชวต เปนการ ศ กษาท เ ก ย วข อ งก บ ชมน มส ง ม ช ว ตท งหมดและ
สงแวดลอมรอบตว เชน ทะเล ทะเลทราย และปา เปนตน
แบบจาลองทางคณตศาสตรไดรบการพฒนาขน เพ อชวยศกษาน เวศ วทยาในทกระดบ ท งน ทฤษฎประชากรมประวตศาสตรทเกยวของกบแบบจาลองยาว นานทสด โดยแบบจาลองทางคณตศาสตรทใช มชอเรยกเฉพาะวา แบบจาลองพลศาสตรประชากร
แบบจาลองพลศาสตรของประชากร (Population Dynamics Models)
แบบจาลองทเกาแกทสดแบบหนงสรางขนในป พ.ศ. 1745 โดย Fibonacci นกคณตศาสตรชาวอตาล เขยนลงในหนงสอ ชอ Liber Abaci เนอหาเปนเรองแบบจาลองของประชากรกระตาย เรมตนในเดอนแรกดวยกระตายวยออน 1 ค เปนเพศผและเพศเมยอยางละ 1 ตวเสมอ ภายใตเงอนไขทวา กระตายวยออนจะเตบโตเปนกระตายตวเตมวยในเดอนถดมา และกระตายตวเตมวย 1 ค จะใชเวลาในการตงครรภ 1 เดอนและใหกาเนดกระตายวยออน 1 ค ทกเดอน นอกจากน กระตายทกตวไมมวนตาย ดงนน จานวนกระตายทนบเปนคในแตละเดอน นบตงแต เดอนท 1 จงสามารถเขยนใหอยในรปของลาดบไดดงน 1, 1, 2, 3, 5, 8, ..... ลาดบน มชอเรยกวา Fibonacci sequence ตอมาในป พ.ศ. 2341 Malthus ไดสรางแบบจาลองทแสดงใหเหนวา ประชากรมการ เตบโตแบบเลขช ก าล ง ( exponential growth) นนคอ
[ ] [ ]NrdtNd
=
เมอ [ ])(tN แทนขนาดของประชากร ณ เวลา t และ r แทนอตราการเจรญเตบโต ตามลาดบ ซงสอดคลองกบผล การทดลองในสภาวะทมอาหารและปจจยสาคญสาหรบ
การดารงชวตทเพยบพรอมสมบรณ อยางไรกตาม ในโลก

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
203
แหงความเปนจรงนน อาหาร ทอยอาศย และปจจยตางๆ มกมอยอยางจากด ในป พ.ศ. 2378 Verhulst จงไดสรางแบบจาลองขนใหม โดยเพมตวแปรเสรม K แทนความจทส งแวดลอมจะรองรบส งม ช วตไดสงสด (carrying capacity) ในสมการ ดงน
[ ] [ ] [ ]⎟⎠⎞
⎜⎝⎛ −=
KNNr
dtNd 1
การเตบโตของประชากรตามสมการนเรยกวา การเตบโตตามทรพยากร และเรยกแบบจาลองนวา แบบจาลองตามทรพยากร อยางไรกตาม แบบจาลองทงสองนยงคงเปนแบบจาลองสาหรบประชากรเพยงชนดเดยว จนกระทงในป พ.ศ. 2468 Lotka และในป พ.ศ. 2469 Volterra ไดคดคนแบบจาลองพลศาสตรของประชากร 2 ชนด นนคอ แบบจาลองผลา-เหยอ ดงน
[ ] [ ] [ ][ ] [ ] [ ]221
2
2111
NmNNbdtNd
NNaNrdtNd
−=
−=
เมอ [ ])(1 tN แทนจานวนของเหยอ ณ เวลา t , [ ])(2 tN แทนจานวนของผลา ณ เวลา t , [ ]1 2N N แทนจานวน ของผลาและเหยอทมอนตรกรยาตอกน r แทนอตราการ เตบโตในตวหรออตราการเกดตามธรรมชาตของเหยอ b แทนอตราการแพรพนธของผลาตอเหยอ 1 ตวทถกกน a แทนอตราการตายของเหยอจากการถกลา และ mแทนอตราการตายตามธรรมชาตของผลา ตามลาดบ เนองจากระบบสมการเชงอนพนธสามญในระยะแรกๆ ยงเปนสมการแบบคาเฉลยทสนใจขอมลระดบประชากร แตไมใหความสาคญในเรองของพนท โดยใชเงอนไขทวาสมาชกแตละหนวยมโอกาสพบปะกนไดอยางทวถงราว
กบโมเลกลของแกสทสามารถแพรกระจายไดทวไป
ภายในภาชนะปด ซงเมอเปรยบเทยบกบปฏกรยาเคม จะพบวา เทอมทประชากรมอนตรกรยาตอกนนน สามารถใชกฎการกระทาของมวล(12) อธบายไดดงน
[ ] [ ] [ ][ ][ ] [ ][ ] [ ]221
2
2111
NmNNbdtNd
NNaNrdtNd
−=
−=
ในความเปนจรง อนตรกรยาเชงนเวศวทยา อาทเชน การลาเหยอ ภาวะปรสต การแพรระบาด การแกงแยงทรพยากร และการสบพนธมกพบในกลมทมมาตราสวนเชงพนทเลกกวาประชากรทงหมด และขอมลเชงพนทจะ
ยงมความสาคญมากขนไปอกถาสงมชวตเคลอนทไดใน
แวดวงจากด ในกรณเชนน การอธบายอนตรกรยาของประชากรดวยกฎการกระทาของมวลจงไมคอยเหมาะสม
แบบจาลองเชงพนทโดยปรยาย (Spatially Implicit Models)
ในป พ.ศ. 2512 Levins(4) ไดรเรมสรางแบบจาลองเชงพนทโดยปรยายขนในชอ ประชากรกลม แบบจาลองนแมไมมตวแปรทเกยวของกบพนทปรากฏในสมการ แตกไดมการแบงประชากรออกเปนกลมยอยหลายๆ กลม โดยมการยายเขาและยายออกเปนตวเชอมระหวางกลมดงน
euuudtdu
−−= )1(λ
เมอ u แทนสดสวนของพนททถกครอบครอง λ แทนอตราการยดครองพนทวาง และ e แทนอตราการสญพนธเฉพาะท ตามลาดบ ถาจดรปสมการใหม จะเปนดงน
⎟⎟⎟⎟
⎠
⎞
⎜⎜⎜⎜
⎝
⎛
−−−=
λ
λe
uuedtdu
11)(
หากเปรยบเทยบกบแบบจาลองตามทรพยากร พบวาม
โครงสรางคลายกน โดยท e−λ แทนอตราการเพมขนของ u ในภาวะการเขาครอบครองตา หรอ
u มขนาด
เลก และ λe
−1 เทยบเทากบความจทสงแวดลอมสามารถ
รองรบสงมชวตไดสงสด นอกจากน ประชากรทอาศยอยในพนทเลกๆ แตละแหงยงสามารถใชสมการชดเดยวกนได และยงยอมใหมการยายเขาและ/หรอออกจากพนทหนงไปยงอกพนทหนงไดเลกนอย มแบบจาลองสาหรบพนทเลกๆ ท Morris(10) นาเสนอซงเปนสมการ ดงน
∑+−−−==
N
iiiiii
i uN
ueuuudtdu
1)1( μμλ
โดย iu แทนจานวนของสงมชวตทอยในกลมประชากร
ยอย i , μ แทนการคควบระหวางกลมของประชากรยอย และคาเฉลยวงกวาง ตามลาดบ และ N แทนจานวนกลมทถกคควบ ทงน มนกวจยหลายทาน(3,5,8) ไดนาแนวคดแบบประชากรกลมนไปใชศกษาปรากฏการณทางชววทยา
ทพบในนเวศวทยาเชงประชากร นเวศวทยาเชงภมทศน และชววทยาเชงอนรกษ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
204
แบบจาลองเชงพนทชดแจง (Spatially Explicit Models)
แบบจาลองเชงพนทชดแจงกาเนดขนในเวลาตอมา
โดยแบงเปน 2 แบบ คอ แบบพนทตอเนอง และแบบพนทไมตอเนอง สาหรบแบบจาลองแบบพนทตอเนองนน มกถกนาเสนอในรปของสมการเชงอนพนธยอย ยกตวอยางเชน แบบจาลองการแพรปฏกรยา ซงนาเสนอในป พ.ศ. 2480 โดย Fisher เพออธบายการแพรกระจายของยน
),(2 tufuDtu
+∇=∂∂
เมอ ),( txu แทนความหนาแนนของประชากรเฉพาะท ณ ตาแหนง x และเวลา t ใดๆ, 2∇ แทนลาปลาเซยน (Laplacian), D แทนสภาพแพร และ ),( tuf แทนฟงกชนไมเชงเสนทอธบายการเปลยนแปลงของประชากร
สทธจากการเกดและการตาย ตามลาดบ ขอจากดของแบบ จาลองประเภทน คอ หาคาตอบเชงวเคราะหไดยาก จงมกตองใชคอมพวเตอรชวยในการวเคราะหเชงตวเลข ทาใหตองปรบความตอเนองของพนทใหเปนแบบไมตอเนอง ตามขอจากดของสมรรถนะของคอมพวเตอร สวนแบบ จาลองทพนทไมตอเนองนนนยมนาเสนอในรปของแบบ จาลองแลตทซ (lattice model) ในกรณทตวแปรสถานะมความตอเนอง แบบจาลองทใชจะเปนแบบคพเพลแมพ แลตทซ (coupled map lattice: CML) แตถาตวแปรสถานะมความไมตอเนอง แบบจาลองทใชจะเปนแบบเซลลลาร ออโตเมตอน (cellular automaton: CA) แบบจาลองแลต- ทซนน ประกอบดวยจด เสน และโครงสรางของประชากรทเนนความสมพนธของสมาชกแตละตว ดงน .
จะเหนไดวา แตละจดอาจถกสงมชวตชนด A หรอ N ครอบครองหรอไมถกสงมชวตใดๆ ครอบครองเลยกได โดย [A] คอ จานวนของจดเดยวทถก A ครอบครอง ซงมคาเทากบ 6 และ [AN] หรอ [NA] คอ จานวนของค AN หรอ NA ทมความสมพนธตอกน ตามลาดบ ซงมคาเทากนคอเทากบ 5 สวน [AA] คอ จานวนของค AA ทมความสม พนธตอกน ซงมคาเทากบ 2 (ตองนบทง 2 ทศทาง) [NAN] คอ จานวนของสามจด NAN ทมความสมพนธตอกน ซงม
คาเทากบ 2 และ [AANA] หรอ [ANAA] คอ จานวนของสจด AANA หรอ ANAA ทมความสมพนธตอกน ตาม ลาดบ ซงมคาเทากนและเทากบ 1
จากตวอยางขางตน สามารถเรยงสญลกษณตางๆ ตามอนดบ ไดดงน [A] มอนดบเทากบ 1 [AN] [NA] และ[AA] เทากบ 2 [NAN] เทากบ 3 [AANA] และ [ANAA] เทากบ 4 นอกจากน การครอบครองจดยงสามารถเปลยน แปลงไดตามเหตการณ อาทเชน การเกด การตาย และการยายเขา-ยายออก เมอเวลาแปรเปลยนไป ขอจากดของการวเคราะหแบบจาลองแลตทซกคอ จาเปนอยางยงทตองพงพาคอมพวเตอร ซงมกใช เวลาในการคานวณนาน โดยเฉพาะอยางยงเมอระบบประกอบดวยสมาชกจานวน
มาก และ/หรอเมอเหตการณมการปรบเปลยนแบบเฟนสม
ในป พ.ศ. 2535 นกวจยชาวญปนไดบกเบกนาแนวคดการประมาณแบบคมาใช กบแบบจาลองทาง
คณตศาสตรทใชอธบายพฤตกรรมของสงมชวตในระบบ
นเวศ โดย Matsuda และคณะ(9) นาแนวคดของการประ- มาณแบบคไปประยกตใชกบแบบจาลองของ Lotka และ
Volterra ดวยวธสรางระบบสมการเชงอนพนธทมความ
หนาแนนทงหมดและความหนาแนนเฉพาะถนเปนตวแปร โดยความหนาแนนทงหมดประกอบไปดวยความหนาแนน
ของจดเดยวและความหนาแนนของค สวนความหนาแนนเฉพาะถนเปนตวบงชความหนาแนนรอบๆ จด Sato และคณะ(16) เรยกวธการดงกลาวนวา การประมาณแบบคสามญ และไดทาการปรบปรงวธการประมาณแบบคดงกลาว เพอใหเหมาะสมกบสถานการณททงผใหอาศยและจลชพ
กอโรคตางกใหกาเนดรนลกหลาน ณ บรเวณพนทใกลๆ กบตวผใหกาเนด ซงเปนผลทาใหสงมชวตชนดเดยวกนมแนวโนมทจะอาศยอยใกลๆ กน โดยเรยกวธการประมาณแบบนวา การประมาณแบบคทปรบปรงแลว ในขณะท Rand(14) เรมตนดวยการสรางสมการครอบ คลม
ตวอยางเชน แบบจาลองเอสไออาร (SIR model)
[ ]
M
][2])[]([2][
][])[(][
][])[(
SSISRSIdtSSd
SIIdtId
SISNdtSd
βμ
βγμ
βμ
−+=
++−=
−−=
เมอ [ ])(tS คอจานวนของสงมชวตทอาจตดเชอได ณ เวลา t , [ ])(tI จานวนของสงมชวตทตดเชอ ณ เวลา t ,[ ])(tR จานวนของสงมชวตทหาย/ปลอดเชอแลวและม

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
205
ภมคมกน ณ เวลา t , มจดเดยวทงหมด N จดในระบบ, [ ]ij เปนจานวนของค ij ทมความสมพนธตอกน, [ ]ijk เปนจานวนของสามจด ijk ทมความสมพนธตอกน, β คออตราการตดเชอ, γ คออตราการหายปวย และ γ คออตราการตายและเกดในเวลาเดยวกนของ R จะเหนไดวา สมการครอบคลมจะประกอบดวยสมการเชงอนพนธทใช
จานวนของจดเดยว จานวนของค จานวนของสามจด และจานวนของจดทมากขนซงมความสมพนธตอกนเปนตว
แปร โดยการเปลยนแปลงของจดเดยวจะเกยวกบคและการเปลยนแปลงของคกจะเกยวของกบสามจด ดงนน สมการครอบคลมจงประกอบดวยสมการจานวนอนนต จงตองใชการประมาณแบบโมเมนตโคลสเชอะ (moment closure approximation) มาชวยประมาณปรมาณทมอนดบสงกวาใหอยในรปของปรมาณทมอนดบตากวา โดยการประมาณแบบคาเฉลยเปนการประมาณแบบโมเมนตโคลสเชอะ อนดบท 0 และการประมาณแบบคเปนการประมาณแบบโมเมนตโคลสเชอะอนดบท 1 ซงถานามาใชประมาณสมการครอบคลมกจะทาใหไดระบบสมการเชงอนพนธ
ใหม มเฉพาะจานวนของจดเดยวและจานวนของคเทานนทเปนตวแปรและมจานวนสมการลดลง ซงสมการชดนมชอเรยกวา สมการสหสมพนธ แนวคดนถกนาไปใชอธบายปรากฏการณตางๆ เชน การคดสรรญาต(2,17) วทยาการ
ระบาด(1) และภาวะปรสต(6) เปนตน สรป
แบบจาลองทางคณตศาสตรมรปแบบทหลากหลาย บทความนมงเนนใหเหนความแตกตางระหวางแบบจาลอง
ทไมคานงถงพนทและแบบจาลองทคานงถงพนท พรอมทงประโยชนใชสอย การเลอกใชแบบจาลองทเหมาะสมจาเปนตองพจารณาจากระบบทตองการศกษา และปญหาทตองการหาคาตอบ หากระบบทสนใจนนมงศกษากลมประชากรทมขนาดใหญ การสรางแบบจาลองในระดบรายหนวยยอมทาใหการหาคาตอบเตมไปดวยความยงยาก
ซบซอนและใช เวลาในการค านวณนาน ในขณะท
แบบจาลองเชงกาหนดมขนตอนทงายกวา และใหภาพโดยรวมไดด เพราะเปนการศกษาพฤตกรรมของประชากรโดยเฉลยอยแลว แตถากลมประชากรทกาลงสนใจนนมขนาดเลกหรอสนใจศกษาความสมพนธของสงมชวต
ระดบรายหนวย จะละทงอนตรกรยาของประชากรไปไมได แบบจาลองทเหมาะสมจงตองนาแนวคดเชงพนทเขามาเกยวของดวยเสมอ
กตตกรรมประกาศ
ขอขอบคณผรวมงานและบคลากรในคณะวทยา- ศาสตร มหาวทยาลยมหดลทกทานทใหการสนบสนนในการเขยนบทความน ตลอดจนสานกงานคณะกรรมการการอดมศกษาและสานกงานกองทนสนบสนนการวจย ทเหนความสาคญและใหทนสนบสนนเพอพฒนาการประมาณ
แบบคสาหรบแบบจาลองเชงพนท
เอกสารอางอง 1. Bauch CT. Moment closure approximations in
epidemiology. Ph.D. Thesis. Coventry: University of Warwick; 2000. 2. Bunwong K. Spatial Modeling in evolutionary eco-
logy. Ph.D. Thesis. Coventry: University of Warwick; 2006.
3. Ferreras P. Landscape structure and asymmetrical inter-patch connectivity in a metapopulation of the endangered Iberian Iynx. Biol Conserv 2001;100: 125-36.
4. Hanski I, Gilpin M. Metapopulation dynamics: brief history and conceptual domain. Biol J Linn Soc 1991;42:3-16.
5. Hofbauer J, Sigmund K. The theory of evolution and dynamical systems. Cambridge: Cambridge Univer-sity Press; 1988.
6. Keeling MJ. The ecology and evolution of spatial host-parasite systems. Ph.D. thesis. Coventry: University of Warwick; 1998.
7. Ljung L, Glad T. Modeling of dynamic systems. New Jersey: Prentice Hall; 1994.
8. Lundquist JE, Klopfenstein NB. Integrating concepts of landscape ecology with the molecular biology of forest pathogens. Forest Ecol Manag 2001;150:213-22.
9. Matsuda H, Ogita N, Sasaki A, Sato K. Statistical-mechanics of population - the lattice Lotka-Voltera model. Prog Theor Phys 1992;88:1035-49.
10. Morris AJ. Representing spatial interactions in simple ecological models. PhD thesis. Coventry: University of Warwick; 1997.
11. Nedorezov LV, Nedorezova BN. Correlation between models of population dynamics in continuous and discrete time. Ecol Model 1995;82: 93-7.
12. Neuhauser C. Mathematical challenges in spatial ecology. Not AMS 2001;48:1304-14.
13. Prévost E, Crozier WW, Schön PJ. Static versus dynamic model for forecasting salmon pre-fishery abundance of the River Bush: a Bayesian compare-son. Fish Res 2005;73:111-22.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
206
14. Rand DA. Correlation equations and pair approxi-mations for spatial ecologies. Warwick Preprint; 1998.
15. Roughgarden J. Theory of population genetics and evolutionary ecology: an introduction. New Jersey: Prentice Hall; 1996.
16. Sato K, Matsuda H, Sasaki A. Pathogen invasion and host extinction in lattice structured populations. J Math Biol 1994;32:251-68.
17. Van Baalen M, Rand DA. The unit of selection in viscous populations and the evolution of altruism. J Theor Biol 1998;193:631-648.
18. West RW, Thompson JR. Models for the simple epidemic. Math Biosci 1997;141:29-39.

207
Phytoremediation of Heavy Metals and Bioaccumulation in Food Plants
Pahol Kosiyachinda Department of Biology, Faculty of Science, Mahidol University.
Abstract
Heavy metals are major constraints of plant growth and development, and heavy metal stress can affect agricultural productivity. Heavy metal accumulation in food plants creates risks for human exposure. Thus, there are growing concerns over environmental and ecological safety. Several approaches have been used to reduce hazards, including regulations to restrict anthropogenic activities in contaminated areas and promoting awareness in the general population. The hazards can be limited in several ways; nevertheless, biological methods are believed to be more sustainable and environmentally friendly. Various plant species are being studied for their potential to remediate heavy metal contamination. Since each species responds differently to heavy metal stress, some have become useful tools for assessing environmental quality, while others have been used for phytoremediation. This paper aims to present broad overview of the evidence and current knowledge on phytoremediation of heavy metals and its implication for food plants.
Keywords: heavy metal; phytotoxicity; phytoremediation; plant development; physiological response
Introduction
Contamination of heavy metals is one of the major environmental and public health problems. It is generally believed that this problem is restricted to industrial areas. However, in fact, contamination of heavy metals is a problem for everyone because heavy metals in soil can be taken up by food plants and incorporated into the food chain(12,24), poisoning animals(9,12) and humans(11,31,45). These heavy metal contaminants have been attributed to both geogonic and anthropogenic sources. Then, both abiotic and biotic processes in the environment affect heavy metals in their distribution, precipitation, adsorption onto and desorption from mineral surfaces, dissolution, interactions with living organisms, and bioavailability(25,29,39). Unlike organic contami-nants, heavy metals do not degrade and are not generally metabolized by microorganisms; so, they can persist in the environment for long periods of time(38). Consequently, heavy metal contaminations may have a major impact on human and animal life, and exposure to heavy metals may cause diseases(7,45).
Learning from many occurrences of catas-trophic heavy metal contaminations, scientists have developed an assortment of technologies to clean up contaminated sites, or to reduce the impacts of such contaminations. These technologies include land filling, soil flushing, solidification, vitrifaction, and phytoreme-diation. While biological approaches provide alternatives to efficient, but costly, physico-chemical approaches, it is important to select the most appropriate approach for each contaminated location carefully. The method of remediation is often determined by the desired rate of the cleanup process, the scale of the contamination, the financial situation, the nature of the contaminated sites, and the relevant hazards.
Phytoremediation may be a promising technology for large scale remediation. This is a plant-based environmental technology for cleaning up contaminated sites by means of degradation, extraction, containment, immobili- zation, or a combination of these approaches(50). Phytoremediation takes advantage of the fact that a living plant acts as a solar-driven

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
208
mechanism, which can extract and concentrate certain heavy metals from the environment. Since heavy metals are often non-essential for plants and a high concentration of heavy metals may be lethal to the plant, several plant species have evolved mechanisms to cope with metals(8,33,54). Those plants with constitutive and adaptive mechanisms for accumulating or tolerating high contaminant concentrations may be used to remediate contaminated soil. Phyto-remediation is a remediation technology that maintains the biological properties and physical structure of the soil; as a result, it is considered environmentally friendly, and potentially cost-effective. Moreover, phytoremediation may not only be a tool for remediation but may also help restore the ecosystem in situ(13).
Mechanisms for Phytoremediation of Heavy Metals
Phytoremediation can be classified based on the mechanisms involved, namely extraction of contaminants from the environment, concentration of the contaminants in plant tissues, and immobilization of contaminants in the root zone of the plants, etc. Since heavy metals cannot be degraded with biological mechanisms, whether in planta or by micro-organisms associated with the plants(22,38), plants with phytoremediation potential take up heavy metals and accumulates them within its tissues. This is called phytoextraction or phytomining, which offers the possibility of bio-recovery of the heavy metals. Nevertheless, specific mechanisms responsible for these processes still need to be discovered.
A good plant candidate for phyto-extraction requires high translocation power, as the plant needs to accumulate high levels of heavy metal to achieve the phytoremediation objective. A number of success stories on phytoextraction of lead, cadmium, zinc, and arsenic have been reported(26,34,44,52). Many plant species use compartmentalization to prevent heavy metal toxicity; thus, the plant does not distribute the accumulated metals evenly in its body. There are certainly mechanisms that control accumulation in the root and in the shoot. In the laboratory, there have been
attempts to elucidate these mechanisms and even to engineer plants with an improved mechanism for phytoremediation purposes(15,49). In field studies investigating uptake and translo-cation of cadmium, chromium, lead, arsenic, and mercury by different parts of rice (Oryza sativa), the results showed that rice grain contained significantly lower amounts of the five metals than straw and root in all samples, whereas the root part accumulated cadmium, arsenic, and mercury at a significantly higher levels than other plant parts(23).
In addition, a plant species may not perform phytoextraction equally well with all heavy metals. For example, vetiver grass (Vetiveria zizanioides) showed a poor efficiency for chromium and copper (both less than 0.1% in shoots and roots in a 30-day experiment), but showed high capability for lead and zinc uptake (0.4% in shoots and 1% in roots for lead, and 1% both in shoots and in roots for zinc in a 30-day experiment)(4). Moreover, different species of related plants performed differently in phytoextraction. For example, V. zizanioides could accumulate more lead from soil than V. nemoralis. Although lead toxicity was observed from the decrease in biomass of both V. zizanioides and V. nemoralis when the lead concentration was increased, V. zizanioides appeared to be more tolerant, as it showed greater biomass than V. nemoralis(6).
A plant can accumulate very high levels of heavy metals, and in the case of hyperaccumulator plants, the concentration of metal accumulated exceeds the concentration in the environment. Several plant species have been reported to hyperaccumulate lead(30,48), arsenic(20,34,52), and cadmium(40,46,47,51). Attempts to understand the molecular basis of hyperaccumulation and differences in accu-mulation potential have been made by transcriptome technology(8,15). Like the example of vetiver abovementioned, it is unlikely that different hyperaccumulator plants use the same molecular mechanism for hyperaccumulation. Nevertheless, that should not be generalized either as the similarities in molecular plant

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
209
responses of Thlaspi caerulescens to cadmium and lead compounds had been reported(21).
The other mechanism for phytoreme-diation of heavy metals is phytostabilization, which immobilizes the heavy metals in the contaminated sites. A plant absorbs the heavy metals and accumulates them in or on its roots; thus, preventing the contaminant from spreading by wind and water erosion, leaching, and soil dispersion. Phytostabilization often involves microorganisms in the root zone of the plants. To achieve the full potential, it is important to understand microenvironment conditions of the soil, pertaining soil microbiology and soil chemistry, since the physico-chemical proper-ties of the soil, such as soil pH and presence of certain compounds may alter the outcome of phytoremediation(16,41). Microenvironment con-ditions of the soil can change metal solubility and mobility or impact dissociation of organic compounds. Application of plant growth-promoting rhizobacteria, including nitrogen-fixing bacteria, and phosphate and potassium solubilizers, may help us understand and further develop phytoremediation techniques. The presence of these beneficial bacteria may stimulate plant growth and provide protection against metal toxicity to achieve higher efficiency of phytoex-traction(5,10,17).
In addition, plants with heavy metal hyperaccumulation ability show a physiological difference in many of their enzyme activities when exposed to heavy metals. Several of these activities have been reported as protection mechanisms for plant cells from environmental stresses including intrinsic oxidative stress reactions from heavy metals. Elevated levels of peroxidase activities, and changes in the production of phytochelatins (PCs), non-protein thiols (NP-SH), glutathione, and antioxidative chemicals which are parts of the plant defense system are among major processes that indicate detoxification mechanisms(19,28,32).
Limitations for Phytoremediation
Even though phytoremediation appears to be one of the cleanest and most economical cleanup technologies for heavy metal conta-minants, phytoremediation has several limita-
tions. First, the plants must be able to take up the heavy metal contaminants from their substrate. Whether the substrate is soil, sediment, or water, the primary limitation for phytoremediation is at the root contact. Without a good contact here, efficiency of the uptake will be compromised. Secondly, the heavy metals must be available for the absorption. This largely involves the physico-chemical conditions of the heavy metals in the substrate. Then, the plants must be able to translocate metals into their shoots.
To overcome these limitations, scientists have devised various approaches to enhance phytoextraction efficiency. An example of induced phytoextraction is the chemically-assisted mobilization of heavy metals with chelating agents. Several chelating agents have been shown to increase bioavailability of heavy metals from the soil solid phase to soil pore water. Common ethylenediaminetetraacetic acid, S,S-ethylenediaminedissucinic acid, and other organic acids have been reported for their application for enhanced phytoextraction of heavy metals(27,36).
Furthermore, when phytoextraction is selected as the remediation approach, the extent of the contamination, the metal concentration that can accumulate in the biomass, and the amount of biomass must be considered. Most hyperaccumulators, studied or reported so far, generally possess low biomass and slow growth rate. This means that the total amount of heavy metals removed might be low, and the remediation process may take a greater amount of time to restore the ecosystem, as compared to other more traditional cleanup technologies. In this case, a plant that extracts a lower concentration of metals, but has a much greater biomass than the hyperaccumulators, may be more desirable because of their greater rate of heavy metal removal(43,53). Even though sites with widespread but low contamination level within the root zone may be the best candidates for the phytoremediation processes, high concentrations of the contaminants may inhibit plant growth and affect development of the plants(37). As a result, this could limit the

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
210
application of phytoremediation on some locations.
Heavy Metal Contamination and Effects on Food Plants
Heavy metals can be absorbed and accumulated in different parts of the plant. Excessive amount of heavy metals can cause a number of toxicity symptoms in plants, e.g. stunted growth, chlorosis, blackening of the root system, and even death. Heavy metals can interfere with plant physiological processes, such as photosynthesis, mineral nutrition and water balance, hormonal status, and integrity of membrane structure and its permeability. These effects are strongly governed by soil and plant factors, and differ significantly with plant species, as mentioned previously. At molecular level, several heavy metals are known to be effective in displacing various cationic metals from roots, suggesting that they may also play a role in destabilizing physiological barriers to solute movement in the root system(8,42,53).
Our research group has been interested in biology of plants in response to heavy metal stress, with projects ranging from studies of heavy metal homeostasis in plants to bioaug-mentation. We have been inspired to work on this major environmental problem when we had an opportunity to visit another campus of Mahidol University in Kanchanaburi Province, Thailand a few years ago. Near this campus, is a field site in which Professor Dr. Maleeya Kruatrachue from Department of Biology, Faculty of Science, Mahidol University located and had established a project to remediate the Bo Ngarm Lead Mine, near Kriti Village where lead contamination was found at very high concentrations. Lead toxicity has manifested itself in local villagers, although the Bo Ngarm Lead Mine had ceased its operation for many years (Figure 1).
On our visit, we observed that many villagers, who had been employed to work in the mine, refused to relocate and were farming to feed their families on the very same land as the lead mine. Workers, remaining in the Kriti Village, became vegetable farmers to earn their living. For those who own a larger piece of
land, vegetables may be grown for commercial purposes (Figure 2). Many of these vegetables would reach the market and become parts of someone’s dinner. We were shocked by this finding, since plants grown in lead contaminated soils can accumulate lead from the adherence of dust and translocation into the plant tissue(31,33). The amount of lead compounds taken up and stored in these plants will vary, depending on the plant species, the type of soil, agricultural practices, and the concentration of lead in the soil. This chain reaction could have been minimized, if not prevented. At this point, this lead contamination problem is no longer restricted to the local villagers of the Kriti. This is just one example of the problems of using former mining areas as agricultural land.
There have been attempts to elucidate heavy metal accumulation levels and their per-taining health risk due to dietary intake of the food plants grown in contaminated areas(1,3,13,55,56). However, there have been few studies in Thailand. A report on accumulation of heavy metals in water spinach (Ipomoea aquatica) cultivated in the Bangkok region of Thailand has suggested that the plants accumulated mercury compounds, but not lead and cadmium, to the levels exceeding its safety standard(13). Other reports of situations in Thailand were mostly performed in animal systems, such as fish(2,18), and crustacean(35).
Figure 1 Lead contamination manifests its toxic effect on the surrounding area of the former mine.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
211
Figure 2 Field of collard in the Kriti Village.
As mentioned above, different plant species respond differently to heavy metals. Therefore, it is important that crop plant candidates be tested for their tolerance to the heavy metal contamination, for their potential to be grown as crop plants in the contaminated areas, and for their safety to be consumed and incorporated into the food chain. Of no less importance, such information should be made public to raise awareness and concerns on heavy metal toxicity.
Several of our projects have been supported by Thailand Research Fund and Mahidol University, especially under the higher education development projects under the Science and Technology Postgraduate Education and Research Development Office (PERDO), of the Commission on Higher Education, Ministry of Education, Thailand. Such support includes scholarships for graduate students who are interested in and concerned by environmental problems.
Conclusions
Plants with phytoremediation potential should have the desirable characteristics of accumulation or tolerance to high level of heavy metals, which may occur through the mecha-nisms of phytoextraction and phytostabili- zation. Meanwhile, scientists are making discoveries of hyperaccumulator plants and trying to understand basic mechanisms of heavy metal hyperaccumulation. In other cases,
phytostabilization may be more practical when there are attempts to carry on agriculture in a contaminated area, especially where access cannot be restricted. Nevertheless, the actual and not just perceived risks of health hazards must not be compromised with possible economic or agricultural uses of the contami-nated sites to ensure that the vegetables comply with the food quality standards on the heavy metal contamination.
Acknowledgements
We would like to thank Professor Dr. Maleeya Kruatrachue for her valuable advice and support throughout our studies and in the preparation of this article. We would like to extend our gratitude to the Center for Toxicology, Environmental Health and Manage-ment of Toxic Chemicals, Mahidol University for their support. Also, parts of this review derive from the experimental results from a research project supported by the Thailand Research Fund (No. MRG4880100) and the Commission of Higher Education, Ministry of Education, Thailand.
References 1. Agbenin JO, Danko M, Welp G. Soil and
vegetable compositional relationships of eight potentially toxic metals in urban garden fields from northern Nigeria. J Sci Food Agr 2009; 89:49-54.
2. Agusa T, Kunito T, Sudaryanto A, Monirith I, Kan-Atireklap S, Iwata H et al. Exposure assessment for trace elements from consumption of marine fish in Southeast Asia. Environ Poll 2007;145:766-77.
3. Alam MGM, Snow ET, Tanaka A. Arsenic and heavy metal contamination of vegetables grown in Samta village, Bangladesh. Sci Total Environ 2003;308:83-96.
4. Antiochia R, Campanella L, Ghezzi P, Movassaghi K. The use of vetiver for reme-diation of heavy metal soil contamination. Anal Bioanal Chem 2007;388:947-56.
5. Braud A, Jezequel K, Bazot S, Lebeau T. Enhanced phytoextraction of an agricultural Cr- and Pb-contaminated soil by bioaugmentation with siderophore-producing bacteria. Chemo-sphere 2009;74:280-6.
6. Chantachon S, Kruatrachue M, Pokethitiyook P, Upatham S, Tantanasarit S, Soonthornsarathool

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
212
V. Phytoextraction and accumulation of lead from contaminated soil by vetiver grass: Laboratory and simulated field study. Water Air Soil Poll 2004;154:37-55.
7. Citterio S, Aina R, Labra M, Ghiani A, Fumagalli P, Sgorbati S et al. Soil genotoxicity assessment: A new strategy based on bio-molecular tools and plant bioindicators. Environ Sci Technol 2002;36:2748-53.
8. Clemens S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 2006;88:1707-19.
9. DalCorso G, Farinati S, Maistri S, Furini A. How plants cope with cadmium: Staking all on metabolism and gene expression. J Integr Plant Biol 2008;50:1268-80.
10. Di Gregorio S, Barbafieri M, Lampis S, Sanangelantoni AM, Tassi E, Vallini G. Combined application of Triton X-100 and Sinorhizobium sp Pb002 inoculum for the improvement of lead phytoextraction by Brassica juncea in EDTA amended soil. Chemosphere 2006;63:293-9.
11. Dorea JG, Donangelo CM. Early (in uterus and infant) exposure to mercury and lead. Clin Nutr 2006;25:369-76.
12. Gal J, Hursthouse A, Tatner P, Stewart F, Welton R. Cobalt and secondary poisoning in the terrestrial food chain: Data review and research gaps to support risk assessment. Environ Int 2008;34:821-38.
13. Gaur A, Adholeya A. Prospects of arbuscular mycorrhizal fungi in phytoremediation of heavy metal contaminated soils. Curr Sci India 2004; 86:528-34.
14. Gothberg A, Greger M, Bengtsson BE. Accumulation of heavy metals in water spinach (Ipomoea aquatica) cultivated in the Bangkok region, Thailand. Environ Toxicol Chem 2002; 21:1934-9.
15. Hammond JP, Bowen HC, White PJ, Mills V, Pyke KA, Baker AJM et al. A comparison of the Thlaspi caerulescens and Thlaspi arvense shoot transcriptomes. New Phytol 2006;170:239-60.
16. Jankong P, Visoottiviseth P, Khokiattiwong S. Enhanced phytoremediation of arsenic conta-minated land. Chemosphere 2007;68:1906-12.
17. Jankong P, Visoottiviseth P. Effects of arbuscular mycorrhizal inoculation on plants growing on arsenic contaminated soil. Chemosphere 2008; 72:1092-7.
18. Jankong P, Chalhoub C, Kienzl N, Goessler W, Francesconi KA, Visoottiviseth P. Arsenic accumulation and speciation in freshwater fish
living in arsenic-contaminated waters. Environ Chem 2007;4:11-7.
19. Jonak C, Nakagami H, Hirt H. Heavy metal stress. Activation of distinct mitogen-activated protein kinase pathways by copper and cadmium. Plant Physiol 2004;136:3276-83.
20. Koller CE, Patrick JW, Rose RJ, Offler CE, MacFarlane GR. Arsenic and heavy metal accumulation by Pteris vittata L. and P. umbrosa R. Br. B Environ Contam Tox 2008;80:128-33.
21. Kovalchuk I, Titov V, Hohn B, Kovalchuka O. Transcriptome profiling reveals similarities and differences in plant responses to cadmium and lead. Mut Res-Fund Mol M 2005;570:149-61.
22. Lasat MM. Phytoextraction of toxic metals: A review of biological mechanisms. J Environ Qual 2002;31:109-20.
23. Liu WX, Shen LF, Liu JW, Wang YW, Li SR. Uptake of toxic heavy metals by rice (Oryza sativa L.) cultivated in the agricultural soil near Zhengzhou City, People's Republic of China. B Environ Contam Tox 2007;79:209-13.
24. McLaughlin MJ, Parker DR, Clarke JM. Metals and micronutrients - food safety issues. Field Crop Res 1999;60:143-63.
25. McLaughlin MJ, Hamon RE, McLaren RG, Speir TW, Rogers SL. Review: A bioavailability-based rationale for controlling metal and metalloid contamination of agri-cultural land in Australia and New Zealand. Aust J Soil Res 2000;38:1037-86.
26. Manousaki E, Kadukova J, Papadantonakis N, Kalogerakis N. Phytoextraction and phyto-excretion of Cd by the leaves of Tamarix smyrnensis growing on contaminated non-saline and saline soils. Environ Res 2008;106:326-32.
27. Meers E, Ruttens A, Hopgood MJ, Samson D, Tack FMG. Comparison of EDTA and EDDS as potential soil amendments for enhanced phyto-extraction of heavy metals. Chemosphere 2005; 58:1011-22.
28. Memon AR, Schroder P. Implications of metal accumulation mechanisms to phytoremediation. Environ Sci Poll Res 2009;16:162-75.
29. Miretzky P, Fernandez-Cirelli A. Phosphates for Pb immobilization in soils: a review. Environ Chem Lett 2008;6:121-33.
30. Munn J, January M, Cutright TJ. Greenhouse evaluation of EDTA effectiveness at enhancing Cd, Cr, and Ni uptake in Helianthus annuus and Thlaspi caerulescens. J Soil Sediment 2008;8: 116-22.
31. Mushak P. Lead remediation and changes in human lead exposure: some physiological and

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
213
biokinetic dimensions. Sci Total Environ 2003; 303:35-50.
32. Nouairi I, Ben Ammar W, Ben Youssef N, Ben Miled DD, Ghorbal M, Zarrouk M. Antioxidant defense system in leaves of Indian mustard (Brassica juncea) and rape (Brassica napus) under cadmium stress. Acta Physiol Plant 2009; 31:237-47.
33. Olguin EJ, Sanchez-Galvan G, Perez-Perez T, Perez-Orozco A. Surface adsorption, intracellu- lar accumulation and compartmentalization of Pb(II) in batch-operated lagoons with Salvinia minima as affected by environmental conditions, EDTA and nutrients. J Ind Microbiol Biot 2005; 32:577-86.
34. Ouyang Y. Phytoextraction: Simulating uptake and translocation of arsenic in a soil-plant system. Int J Phytoremediat 2005;7:3-17.
35. Panutrakul S, Khamdech S, Kerdthong P, Senanan W, Tangkrock-Olan N, Alcivar-Warren A. Heavy metals in wild banana prawn (Fenneropenaeus merguiensis de Man, 1888) from Chantaburi and Trat provinces, Thailand. J Shellfish Res 2007;26:1193-202.
36. Parra R, Ulery AL, Elless MP, Blaylock MJ. Transient phytoextraction agents: Establishing criteria for the use of chelants in phytoextraction of recalcitrant metals. Int J Phytoremediat 2008; 10:415-29.
37. Patra M, Bhowmik N, Bandopadhyay B, Sharma A. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ Exp Bot 2004;52:199-23.
38. Ramteke DD, Nagrale VD, Ramteke MD. In situ phytoremediation of heavy metals: A sustainable approaach for waste water monitoring in India. Res J Biotechnol 2008:305-8.
39. Rieuwerts JS. The mobility and bioavailability of trace metals in tropical soils: a review. Chem Spec Bioavailab 2007;19:75-85.
40. Roosens NH, Leplae R, Bernard C, Verbruggen N. Variations in plant metallothioneins: The heavy metal hyperaccumulator Thlaspi caeru-lescens as a study case. Planta 2005;222:716-29.
41. Rotkittikhun P, Kruatrachue M, Chaiyarat R, Ngernsansaruay C, Pokethitiyook P, Paijit-prapaporn A et al. Uptake and accu-mulation of lead by plants from the Bo Ngam lead mine area in Thailand. Environ Poll 2006;144:681-688.
42. Shen MJ, Hu SY, Blaha U, Yan HT, Roesler W, Hoffmann V. A magnetic study of a polluted soil profile at the Shijingshan industrial area, Western Beijing, China. Chinese J Geophys-Ch 2006;49: 1665-73.
43. Shen ZG, Li XD, Wang CC, Chen HM, Chua H. Lead phytoextraction from contaminated soil with high-biomass plant species. J Environ Qual 2002;31:1893-900.
44. Solhi M, Shareatmadari H, Hajabbasi M. Lead and zinc extraction potential of two common crop plants, Helianthus annuus and Brassica napus. Water Air Soil Poll 2005;167:59-71.
45. Strydom C, Robinson C, Pretorius E, Whitcutt JM, Marx J, Bornman MS. The effect of selected metals on the central metabolic pathways in biology: A review. Water Sa 2006;32:543-54.
46. Sun YB, Zhou QX, Diao CY. Effects of cadmium and arsenic on growth and metal accumulation of Cd-hyperaccumulator Solanum nigrum L. Bioresource Technol 2008;99:1103-10.
47. Sun YB, Zhou QX, Wang L, Liu WT. Cadmium tolerance and accumulation characteristics of Bidens pilosa L. as a potential Cd-hyperaccu-mulator. J Hazard Mater 2009;161: 808-14.
48. Tanhan P, Kruatrachue M, Pokethitiyook P, Chaiyarat R. Uptake and accumulation of cadmium, lead and zinc by Siam weed Chromolaena odorata (L.) King & Robinson. Chemosphere 2007;68:323-9.
49. Tong YP, Kneer R, Zhu YG. Vacuolar compartmentalization: a second-generation approach to engineering plants for phyto-remediation. Trends Plant Sci 2004;9:7-9.
50. US-EPA. Introduction to Phytoremediation. Cincinnati, Ohio, USA: United States Environ-mental Protection Agency; 2000. Report No.: EPA/600/R-99/107.
51. Van TK, Kang YM, Fukui T, Sakurai K, Iwasaki K, Aikawa Y et al. Arsenic and heavy metal accumulation by Athyrium yokoscense from contaminated soils. Soil Sci Plant Nutr 2006; 52:701-10.
52. Visoottiviseth P, Francesconi K, Sridokchan W. The potential of Thai indigenous plant species for the phytoremediation of arsenic conta-minated land. Environ Poll 2002;118:453-61.
53. Weiss J, Hondzo N, Biesboer D, Semmens N. Laboratory study of heavy metal phytoreme- diation by three wetland macrophytes. Int J Phytoremediat 2006;8:245-59.
54. Wilson B, Pyatt FB. Heavy metal bioaccu-mulation by the important food plant, Olea europaea L., in an ancient metalliferous polluted area of Cyprus. B Environ Contam Tox 2007; 78:390-4.
55. Zheng N, Wang QC, Zheng DM. Health risk of Hg, Pb, Cd, Zn, and Cu to the inhabitants around

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
214
Huludao Zinc Plant in china via consumption of vegetables. Sci Total Environ 2007;383:81-9.
56. Zhuang P, McBride MB, Xia HP, Li NY, Lia ZA. Health risk from heavy metals via consumption of food crops in the vicinity of Dabaoshan mine, South China. Sci Total Environ 2009;407:1551-61.

215
แนวโนมการพฒนาอปกรณเปลงแสงจากสารอนทรย
สมศกด แดงตบ ธนากร โอสถจนทร และ สมบญ สหสทธวฒน ภาควชาฟสกส คณะวทยาศาสตร มหาวทยาลยมหดล และ ศนยเทคโนโลยโลหะและวสดแหงชาต
บทคดยอ
อปกรณเปลงแสงจากสารอนทรย (โอแอลอด) ถอกาเนดขนเมอเกอบ 20 ปทแลว และเรมมความสาคญเชงพาณชยในบทบาทเปนจอแสดงผลแบบแบน ศกยภาพของอปกรณเปลงแสงจากสารอนทรยยงไมอมตว ซงมแนวโนมทจะทวบทบาทและความสาคญเชงพาณชยไดอกมาก การพฒนาและวจยในเชงลกจงมสวนสาคญจะชวยเพมประสทธภาพการเปลงแสง ความเขมการสองสวาง รวมถงชวยลดกาลงไฟทตองจายใหอปกรณลง โครงสรางของอปกรณเปลงแสงประกอบไปดวย ซบสเตรต แอโนด ชนฟลมบางของสารอนทรยเปลงแสง และแคโทด เมอผานกระแสไฟฟาให อปกรณจะเปลงแสง สทเปลงออกมาขนอยกบชนดหรอโครงสรางของสารอนทรย ประสทธภาพของการเปลงแสงถกกาหนดจากระดบพลงงานสมพทธระหวางชนตางๆ ในโครงสรางของอปกรณ ทศทางการเปลงแสงเลอกจากความโปรงแสงของชนอเลกโทรด และสถาปตยกรรมดานโครงสรางในตวอปกรณ อปกรณเปลงแสงจากสารอนทรยแตละดานยงสามารถพฒนาตอไปไดอกมาก
คาสาคญ: อปกรณเปลงแสงจากสารอนทรย; โอแอลอด; สารอนทรยเปลงแสง; จอแสดงผลแบบแบน
Abstract
Organic light emitting device (OLED) was developed some 20 years ago and has already been utilized commercially, mainly in flat panel display. However, OLED has not reached its full potential and its commercial role continues to grow. In-depth research and development will further improve OLED performance in terms of luminous efficiency, luminance and reduction in power consumption. OLED consists of substrate, anode, electroluminescence organic layer and cathode. When a current is injected into the device, light is emitted and the emission wavelength or colour depends on the nature of the organic material. Luminous efficiency is governed by the electronic structures between neighbouring layers. Direction of emission depends on transparency of electrodes and structure of the device. Trends in research and development of OLED are discussed.
Keywords: organic light emitting devices; OLED; electroluminescence organic materials; flat panel display
บทนา
ในปจจบนอปกรณ เปลงแสงจากสารอนทรย (organic light emitting devices) หรอไดโอดเปลงแสงจากสารอนทรย (organic light emitting diodes; OLED) ไดรบความนยมกนอยางแพรหลายในอปกรณตาง ๆ ไมวาจะเปนโทรศพทมอถอ โทรทศน แผนไฟสองสวาง หรออปกรณแสดงผลอยางอนๆ และคาดวา จะเปนอปกรณ
หลกเพอการแสดงผลทจะมาทดแทนการแสดงผลแบบ
ผลกเหลวหรอแบบพลาสมา ชนสารอนทรยเปลงแสงเปนแหลงกาเนดแสงโดยตรงไมตองอาศยแหลงกาเนดแสงอน
OLED จงกนพลงงานไฟต า และแสงท เปลงออกมาสามารถครอบคลมมมการมองเกอบรอบทศทาง 180 องศา ชนสารอนทรยทใชเปนชนฟลมบางเปลงแสงมความหนา
เพยง 100 นาโนเมตร เมอรวมกบความหนาของชนฟลมบางอนๆ แลว ทาใหสามารถพฒนาอปกรณใหมขนาดทบาง ตลอดจนควบคมสทเปลงออกมาโดยการควบคมชนดหรอโครงสรางของโมเลกลของสารอนทรย ทาใหสามารถเลอกการเปลงสแดง เขยว นาเงน หรอขาว จากโมเลกลทเหมาะสมได นอกจากน OLED สามารถตอบสนองไดรวดเรวกวาจอแสดงผลแบบอนมากถง 100 เทา ปจจบนเรมมการนา เครองรบโทรทศนทม จอแสดงผลจาก

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
216
สารอนทรยเปลงแสงทหนาไมเกนสามมลลเมตรออก
วางขายในทองตลาดแลว
ศกยภาพของเทคโนโลย OLED เปนสงทไมควรมองขาม ถงแมวาอปกรณชนดนจะมการพฒนามายาวนานพอสมควร แตเทคโนโลยนยงตองการการวจยและพฒนา เชงลกตอไปอกเพอหาจดทเหมาะสม ใหกาวขามปญหาเรองอายการใชงาน ประสทธภาพของการสองสวาง กอนทจะกาวเขามาทดแทนเปนอปกรณแสดงผลหลกได
อยางสมบรณ
ประเทศไทยมศกยภาพและความแขงแกรงทางเคม
อนทรยพอล เมอร และเคมสง เคราะห รวมถงความ สามารถดานเครองมอและประสบการณดานการประกอบ
อปกรณ จงนาสนใจทจะพฒนาศกยภาพของเทคโนโลย OLED ใหเขมแขงขน บทความนไดกลาวถงเสนทางการพฒนา หลกการการทางาน แนวโนมของการพฒนา OLED ทงในดานวสดและกระบวนการตลอดจนนาเสนอผลงานวจยบางสวนทเกยวเนอง
เสนทางการพฒนาอปกรณเปลงแสงจากสาร อนทรย
ประวตศาสตรของ OLED เรมมาจากงานวจยของ Bernanose และคณะ(4) ในชวงตนป พ.ศ. 2500 เปนกลมแรกทศกษาการเปลงแสงจากสารอนทรย โดยการกระตนดวยกระแสไฟฟาจากการจายไฟฟากระแสสลบความตาง
ศกย สงใหแกผลกของ acridine orange และ quinacrine ทเปนฟลมบางๆ หลงจากนน ในป พ.ศ. 2508 Helfrich และ Schneider(8) พบว า เม อ ใหกระแสไฟฟ าผ าน เข าผลก anthracene ทเชอมตอเซลลเคมไฟฟา ผลกนจะเปลงแสงสฟาออกมาได ตอมา Vincett และคณะ(13) ไดทาการปรบ ปรงการสราง OLED โดยการทา anthracene เปนฟลมบางดวยวธระเหยผง anthracene ในสญญากาศและใชขวโลหะ สงผลให OLED เปลงแสงไดทความตางศกยทตาลงมาก
OLED ทมประสทธภาพสงขนไดถกสรางขนโดย Tang และคณะ(11) แหงบรษท Kodak ในป พ.ศ. 2530 ไดทา OLED ทประกอบดวยโครงสรางสารอนทรยสองชนโดยแยกเปนชนเปลงแสงและนาอเลกตรอนจาก Alq3 และชนนาโฮลจากสาร aromatic diamine พบวา ประสทธภาพของ OLED สงขนจากเดมหลายสบเทา ทงนสวนหนงเนองจากสมดลระหวางอเลกตรอนและโฮลทถกฉดเขาส
ชนเปลงแสงถกทาใหดขนดวยโครงสรางเชนน ผลงานนถอเปนจดเปลยนททาใหการวจยและพฒนาดาน OLED เตบโตและกาวหนามาจนถงในปจจบน
แนวความคดนถกนาไปตอยอดกบดานโพลเมอร โดย Burroughes และคณะ (5) ไดตพมพผลงานเกยวกบ พอลเมอรท เปลงแสงสเขยวทประสทธภาพสงมากใน
วารสาร Nature ในป พ.ศ. 2533 จากนนความสนใจในงาน วจยและพฒนาดาน OLED ไดเพมขนอยางรวดเรว ตลอด จนมการพฒนาและสงเคราะหสารอนทรยชนดใหมๆ ทชวยเพมประสทธภาพของ OLED ใหสงขนดวย
การใชประโยชนเชงพาณชย เทคโนโลย OLED ไดรบการยอมรบและประยกต ใชในเชงพาณชย(10) เชน จอแสดงผลขนาดเลกของโทร- ศพทมอถอ เครองเลน MP3 แบบพกพา วทยในรถยนต กลองถายรปดจตอล และจอแสดงผลความละเอยดสงในอปกรณแสดงผลแบบสวมศรษะ การนาไปใชงานสะทอนใหเหนถงขอดของ OLED ทใหความคมชดแมในทมแสงสวางจาและความประหยดไฟ การใชงานอปกรณแสดงผลแบบพกพาเปนแบบชวงๆ ทาใหอายการใชงานทจากดของ OLED ไมใชอปสรรค อปกรณแสดงผลตนแบบทบดงอและมวนเรมมใหเหนบางแลว บรษทผลตโทรศพทมอถอ เชน โมโตโรลา และซมซง รวมไปถง โซน อรกสน เชนรน Z610i และโซนวอลคแมนบางรนไดนา OLED ไปใชในสวนแสดงผล นอกจากนน ยงพบไดในเครองเลน MP3 รนสราง สรรค เชน v/v plus โนเกยเรมนา OLED ไปประยกตใชในโทรศพทมอถอรนปรซม 7900 และอารต 8800 ทงน ในเดอนสงหาคม พ.ศ.2551 บรษทโนเกยได
เปดตวสมาชกใหม ซรเอน รน N85 ทใชจอแสดง ผลแบบ AMOLED (active matrix OLED) ขนาด 2.6 นว ซงถอวามขนาดใหญทสดในกลมโทรศพทมอถอ GSM ขณะทในเดอนตลาคม พ.ศ. 2550 โซนเปนบรษทแรกทประกาศสรางโทรทศน OLED รน XEL-1 ขนาด 11 นว และเรมวางจาหนายในเดอนธนวาคม พ.ศ. 2550 เปนตนมา และในเดอนเมษายน พ.ศ. 2551 บรษทไดนาเสนอจอ แสดงผลขนาด 3.5 นว ความละเอยด 320×200 จด ทมความหนาเพยง 0.2 มม. และจอแสดงผลความละเอยด 960×540 จด ความหนา 0.3 มม. ซงเปนขนาดความหนาของจอเพยงหนงในสบของโทรทศนรน XEL-1 ทความกวางและความละเอยดเทากน
โครงสรางและการทางานของอปกรณเปลงแสงจากสารอนทรย โครงสราง OLED ประกอบดวยซบสเตรต ชนแอโนด ชนนาไฟฟา ชนเปลงแสง และ แคโทด ในอปกรณแสดงผลทใชงานจรงจะมกาวอพอกซและกระจกชนบน เพอหมหออปกรณทงหมดไว (รปท 1)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
217
(ก) (ข)
รปท 1 (ก)โครงสรางของจอแสดงผล (ข) ภาพขยายของโครงสรางของแตละพกเซลในจอแสดงผล (ดดแปลงจาก1)
เมอมกระแสไฟฟาไหลผาน ชนฟลมบางสาร อนทรยหรอสารประกอบโลหะอนทรยจะปลดปลอยแสง
ออกมาผานทะลขวไฟฟาดานหนงหรอทงสองดาน ดงแสดงในรปท 2 ซบสเตรตในโครงสราง OLED เปนวสดยอมใหแสงผานได เชน แผนกระจกแกวหรอพลาสตกใส ซบสเตรตจะถกเคลอบดวยขวไฟฟาแอโนดโปรงแสง ซงมกจะใชออกไซดของอนเดยมทเจอดวยดบก (tin-doped indium oxide; ITO) ชนแอโนดมความหนาประมาณ 100 นาโนเมตร จากนนจะมชนฟลมบางทชวยการนาไฟฟาของพาหะโฮล ความหนาอยระหวาง 10 ถง 100 นาโนเมตร และตอมาจงทาการเคลอบชนสารอนทรยททาหนาทเปลงแสง ความหนาไมเกน 100 นาโนเมตร ชนสาร อนทรยท เปลงแสงเปนสารอนทรยทมชองวางระดบ
พลงงานในชวงแสงทตามองเหนและสามารถเกดปรากฏ การณวาวแสง (fluorescence) ได เชน สารประกอบ Alq3 หรอสารพอลเมอรทมพนธะคอนจเกต อาทเชน poly-phe- nylene vinylene (PPV) และ polyfluorene (PF)
ดานบนของฟลมเปลงแสงเคลอบดวยขวไฟฟาทมคา
ฟงกชนงานใกลเคยงกบคา lowest unoccupied mole- cular orbit (LUMO) ในฟลมเปลงแสงเพอลดระดบกาแพงศกยทอ เลกตรอนตองขามจากชนแคโทดไปยงชนฟลมสาร อนทรย โดยทวไปอาจเลอกใชโลหะแคลเซยมหรอลเธยมเปนแคโทด แลวเคลอบทบดวยอลมเนยมเพอปองกนการทาปฏกรยากบนาและออกซเจน ความหนารวมของชนแค- โทดไมตากวา 100 นาโนเมตร รปท 2 แสดงระดบพลงงานสมพทธระหวางแตละชนฟลม อยางไรกตาม อาจมการ
ปรบปรงโครงสราง OLED โดยการเพมชนชวยการนาอ- เลกตรอนระหวางชนฟลมเปลงแสงและแคโทดเพอชวย
ปรบระดบพลงงานของอเลกตรอนระหวางชนแคโทดและ
ฟลมเปลงแสง ในการออกแบบโครงสราง OLED จงตองพยายามปรบปรมาณการเคลอนทของพาหะโฮลและอเลก- ตรอนจากชนแอโนดและแคโทดใหเหมาะสม เพอใหเกดการรวมตวกนของอเลกตรอนจากชน LUMO และโฮล
รปท 2 โครงสรางและระดบพลงงานสมพทธระหวางฟลมชนตาง ๆ ในOLED ลกศรสนาเงนแสดงทศทางการเคลอนทของอเลกตรอนและโฮล
และลกศรสเหลองแทนทศทางของแสงทเปลงออกมา (ดดแปลงจาก1)
Transparent substrate
Conducting polymer
LUMO
HOMO
h+ h+
แอโนด(ITO)
Emissive organic
LUMO
HOMO
e– e–
h+
แคโทด
แสง
ความหนา ~ 100 nm <100 nm ของชนฟลม 10 – 100 nm >100 nm
แอโนด (ITO) พอลเมอรนาไฟฟา สารอนทรยเปลงแสง
แคโทด (Li/Al, Al)
กระจกทบหลง
กาวอพอกซ
พอลเมอร นาไฟฟา
ซบสเตรตกระจก
แอโนด
แคโทด
สารอนทรย เปลงแสง

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
218
จากชน highest occupied molecular orbit (HOMO) อยางเหมาะสมและพอดในชนของฟลมสารอนทรยเปลงแสง
โดยทวไปสารอนทรยเปลงแสงจะมการนาไฟฟาท
ตา คอ มคาสภาพเคลอนทไดของพาหะโฮลในชวง 10-7 ถง 10-3cm2/Vs ในขณะทคาสภาพเคลอนทไดของอเลกตรอนจะนอยกวา 10-100 เทา ซงการนาไฟฟาตาเปนผลมาจากความไมเปนระเบยบของการเรยงตวของโมเลกลในฟลม และการทสารอนทรยนาโฮลไดดกวาอเลกตรอน เพราะโฮลมศกยออกซเดชนตา(3) กลไกการเคลอนทของพาหะ
อสระในสารอนทรยน เปนการกระโดด (hopping) ของพาหะระหวางสถานะในแตละโมเลกล จากผลการวดสภาพเคลอนทไดของพาหะในโครงสราง OLED ดวยวธตางๆ พบวาคานจะเปลยนไปตามคาสนามไฟฟาทปอนใหในแตละชวง ซงสามารถอธบายไดดวยการเกดโพลารอน โดยพาหะจะมอนตรกรยาคอนขางแรงกบการสนของ
โมเลกลและโมเลกลใกลเคยง นนคอ ความสามารถในการเคลอนทของพาหะขนกบอณหภม โดยทวไปถาอณหภมสงขนการเคลอนทของพาหะจะดขน
ทบรเวณรอยตอระหวางขวไฟฟาและสารอนทรย โดยปกตระดบพลงงานของพาหะในโลหะและสารอนทรย
จะมกาแพงศกย เนองจากความตางกนของระดบพลงงานอย แตเมอมการถายเทพาหะไปยงสถานะกบดก (trapped state) ในสารจะเกดศกยทสามารถลดกาแพงศกยนอยาง
มากในบรเวณรอยตอ ซงเรยกวาศกยแรงเสมอน ทาใหพาหะสามารถถกฉดเขาสสารอนทรยได จากนนพาหะจะกระโดดเขาไปในสารตามแนวของสนามไฟฟาภายนอกท
ปอนให
เพอให OLED เปลงแสงออกมาใหมความเขมมากนน จาเปนตองใหมการฉดกระแสทความหนาแนนตอหนวยพนทสงมาก เขาไปยงสารอนทรยทมสภาพเคลอนทไดของพาหะนอยมาก จงจะทาใหมการสะสมของประจ
จานวนมาก ดวยสนามไฟฟาทปอนใหกบโครงสราง OLED จะทาใหประจเหลานปรบการกระจายตวใหม และความสมพนธระหวางกระแสและความตางศกยทไดจงม
ลกษณะคลายกบกระแสไฟฟาทไหลในกระบวนการจากด
กระแสดวยประจ ซงกระแสจากกระบวนการนจะขนกบความตางศกยยกกาลงคาตาง ๆ และยงขนกบลกษณะและปรมาณของสถานะกบดก
การพฒนาให OLED มประสทธภาพทางพลงงานสง หรอทาใหแปลงพลงงานไฟฟาเปนพลงงานแสงใหเปลงออกมามากทสดนน นอกจากจะเปนประโยชนตอการใชงานแลว ยงชวยยดอายการใชงานของอปกรณ OLED ดวย เพราะชวยลดการเกดพลงงานความรอนในอปกรณใหนอยลง ประสทธภาพของ OLED นขนกบประสทธภาพการเปลงแสงของโมเลกล อตราสวนการปลดปลอยโฟตอนจากผวของอปกรณ อตราสวนการเกดเอกซตอน (exciton) ตอการไหลของกระแสไฟฟา และอตราสวนการเกดเอกซตอนเดยวตอเอกซตอนทไมเกดการเปลงแสง จากการประมาณทางทฤษฎ ประสทธภาพสงสดของ OLED จะถกจากดอยประมาณ 25% ของประสทธภาพการเปลง แสงจากโมเลกล
คณลกษณะของอปกรณเปลงแสงจากสารอนทรย ลกษณะเฉพาะทดของ OLED วดไดจากหลายพารา
มเตอร เชน ความหนาแนนกระแสทสงเมอผานความตางศกยไฟตรงให แตนอยลงมากเมอผานความตางศกยยอน กลบ [รปท 3(ก)] คาสงสดของความสองสวางทไดเมอผานความตางศกยให [รปท 3(ข)] และคาความตางศกยทางานทตาเมอเรมใหความสองสวางอยางนอย 100 cd/m2 ซงเพยง พอสาหรบการนาไปใชเปนจอแสดงผล แตสาหรบการใชเพอเปนแหลงกาเนดแสงสวาง เชน หลอดไฟ จะตองใชความสองสวางเพมมากขนเปน 1000 cd/m2
(ก)
(ข)
(ค)
(ง)
รปท 3 ความสมพนธระหวางคณลกษณะของอปกรณ (ก) ความหนาแนนกระแสกบความตางศกย (ข) ความสองสวาง กบความตางศกย (ค) ประสทธภาพกบความสองสวาง และ (ง) ประสทธภาพกบความตางศกย (ดดแปลงจาก1)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
219
สถาปตยกรรมดานโครงสรางของ OLED การวางเรยงชนฟลมใน OLED อางองกบซบ-สเตรต โดยใหซบสเตรตเปนดานลาง การกาหนดทศทาง การเปลงแสงยดการอางองแบบเดยวกน อปกรณในยคตนๆ เปนแบบเปลงแสงทางดานลางผานแอโนดและซบ-สเตรตทโปรงแสง รปท 4 เปนตวอยาง OLED ทเปลงแสงทางดานลางโดยใชกระจกเปนซบสเตรต ITO เปนแอโนดและอะลมเนยมเปนแคโทด
ตอมาไดมการปรบใชวสดโปรงแสงเปนชนแค- โทดดวย ทาใหอปกรณ เปลงแสงสามารถใหแสงทงดานบนและดานลาง เปนอปกรณเปลงแสงแบบโปรงใส (transparent organic light-emitting device; TOLED) [รปท 5(ก)] ซงเมอยงไมทางานจะสามารถมองทะลผานชนฟลมบางทงหมดทกชน เหนขอความทอยทางดานหลงของอปกรณได และเมอทางานแสงทเปลงออกมาจะบงขอ ความทอยทางดานหลงไป [รปท 5(ข)] TOLED มขอดคอ ชวยเพมความเปรยบตางส (contrast) ใหมากขน ทาใหการแสดงผลในบรเวณทมแสงจาดขน เชน การแสดงผลบนหนากากสวมหวหรอบนเลนสแวนตา
ผลแบบผลกเหลว การวางโครงสรางแบบเปนชนๆ ชวยเพมอานาจการจาแนกสอกอยางนอย 3 เทา และยงชวยการแสดงผลแบบสเตม อปกรณเปลงแสงแบบกลบโครงสราง (inverted OLED; IOLED) เปนโครงสรางทแตกตางจาก OLED ปกตทใหแผนแอโนดอยบนซบสเตรต IOLED จะวางข วแคโทดบนซบสเตรตแทนเพอใหตอกบดาน สายระบายของทรานซสเตอรแบบฟลมบางชนดเอน (n-channel TFT) ทาใหสามารถนาเอาซลกอนแบบอสณ-ฐานมาใชเปนระนาบหลงของทรานซสเตอรแบบฟลมบาง
ท ใชประกอบการแสดงผลแบบแอกทฟเมตรกซได(AMOLED) ซงชวยลดราคาตอหนวยอปกรณลง
แนวโนมการพฒนาดานขวไฟฟาโปรงใส ออกไซดของอนเดยมทเจอดวยดบกหรอ ITO จด
เปนวสดหลกทใชทาแอโนด เนองจากนาไฟฟาไดด มสภาพตานทานไฟฟาเพยง 10-4 Ω-cm ขณะทยอมใหแสงสองผานไดมากกวา 85% ITO มคาฟงกชนงานสงประมาณ 4.7-5.0 eV จงชวยเสรมการฉดโฮลเขาสชนพอลเมอร ITO เมอถกเจอดวยซเซยม จะมคาฟงกชนงานทลดลง สามารถทจะนามาใชเปนชนแคโทด(12) ชวยใหอปกรณทงหมด
(ก)
(ข)
(ค)
(ง)
รปท 4 OLED แบบเปลงแสงผานซบสเตรตดานลาง (ก) ขณะปด และขณะเปด (ข) สแดง (ค) สเขยว และ (ง) สนาเงน (ตวอกษรภาษาญปนเปน ชอมหาวทยาลย Tokyo Polytechnic University)
(ก)
(ข)
รปท 5 อปกรณเปลงแสงสเขยวแบบโปรงใส (ก) ขณะปด และ (ข) เปด
อปกรณ เปลงแสงแบบซอนเปนชนๆ (stacked OLED) มโครงสรางทนาเอาสวนแสดงผลทเปนแมสแดง เขยว และนาเงนมาซอนกนเปนชน แทนการวางเรยงขางๆ ซงเปนเทคโนโลยทใชในหลอดภาพแสดงผลและจอแสดง
เปนอปกรณโปรงใส (TOLED) การเคลอบฟลมบาง ITO ทาโดยการฉาบดวยโลหะ เมอตองการใช ITO ชนแคโทดบนซบสเตรตทมชนสารอนทรยเปลงแสงเคลอบอยอาจ
ประสบปญหาเรองการเสยสภาพของชนสารอนทรย เมอใชระบบฉาบดวยโลหะทมเปา ITO และซบสเตรตวางอยตรงขามกน อาจตองมการปรบ ปรงระบบไปใชโครงสรางแบบ facing target ทใชเปา ITO ควางตรงขามกน แลววางซบสเตรตดานขางแทน(6)
จากการประมาณการมลคาการตลาดของอเลก- โทรดโปรงใส ITO ในปจจบน อยท 8 พนลานเหรยญ สหรฐฯ(2) ขณะทมลคาอเลกโทรดโปรงแสงจากวสดอนอย

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
220
ท 1.4 พนลานเหรยญสหรฐฯ เทานน เชอกนวา ITO จะยงครองสวนแบงหลกนไปอกอยางนอยประมาณ 10 ป คาดกนวามลคาการตลาดของ ITO จะอยท 9.4 ลานเหรยญ
สหรฐฯ ในป พ.ศ. 2558 ทวาปญหาของอนเดยมทมราคาแพงและปรมาณแหลงแรสารองทมจากด จงเรมมการมองหาวสดทดแทน คณสมบตพนฐานทสาคญของวสดทจะนา มาใชเปนอเลกโทรดโปรงใสแทน ควรมสมบตทางไฟฟาและทางแสงใกลเคยงหรอดกวา ITO มเสถยรภาพทด และตองการการอบผสานเนอดวยอณหภมทไมสงนก ไมเสยสภาพการทางานเมอถกโคงงอ เพอใหรองรบการเคลอบบนซบสเตรตทยดหยนได เชน พลาสตกทนรอนและทคงรปเมออณหภมไมเกน 200°C เทานน
แนวโนมการพฒนาดานสารอนทรย นบแตมการนาพอลเมอรมาประยกตใชเปนสาร อนทรยเปลงแสง ตงแตป พ.ศ. 2533 เปนตนมา นกวจย
หลายกลมได เรมหนมาใหความสนใจในการนาสาร อนทรยและสารประกอบโลหะอนทรยชนดใหมหลาก หลายชนดมาใชสราง OLED โดยมการทดลองนาสารเหลานนมาใชเปนสารเปลงแสง สารเจาบาน สารนาโฮล หรอสารนาอเลกตรอนตามแตคณสมบตของสารนนๆ
สารทใชเปนสารเปลงแสงตองมความสามารถใน
การเปลยนพลงงานไฟฟาเปนพลงงานแสงไดด และ/หรอใหสทเปลงออกมาตามความตองการ เชน สแดง เขยว หรอนาเงน เปนตน รปท 6 แสดงตวอยางสารทถกใชเปนสารเปลงแสง
ตามทฤษฏแลว การเปลงแสงจากกระบวนการวาว
แสงดวยการใหพลงงานไฟฟา สามารถมประสทธภาพสง
ไดถง 25% ทงนเพราะการวาวแสงเกดจาก singlet exciton
ซงมอตราการเกดเทากบ 1:3 เมอเทยบกบการเกด triplet exciton ดงนน หากตองการเพมประสทธภาพของ OLEDตองเพมกระบวนการเรองแสง (phosphorescence) เขามาดวยการเตมสารท เรองแสงดวยกระบวนการเรองแสง ตวอยางของสารประกอบโลหะอนทรยทเรองแสงไดดวย
กระบวนการเรองแสงนแสดงในรปท 7
สของแสงทเปลงออกมาจาก OLED อาจปรบแตงไดโดยการเลอกใชสารเปลงแสงทใหสทตางกนไป หรอจากการผสมสารเปลงแสงทเปนสทแตกตางกนเขาดวยกน เชน การผสมสารทเปลงแสงในชวงสแดงกบนาเงนในอตราสวนทเหมาะสม ทาใหไดแสงสขาวเปลงออกมาจาก OLED ซงเหมาะสมกบการใชเปนแหลงกาเนดแสงตางๆ เชน ใชเปนแหลงกาเนดแสงพนหลงในจอ LCD หรอใชแทนหลอดไฟได
ในกรณทถกใชเปนสารเจาบาน สารอนทรยหรอสารประกอบโลหะอนทรยตองมแถบตองหามกวางกวา
สารเปลงแสงทถกผสมเขาไป เพอใหพลงงานสามารถถาย เทจากสารเจาบานทมระดบพลงงานในสภาวะเราทสงกวา
ไปยงสารเปลงแสงได ทงนแสงทเปลงออกมาจะขนกบสารเปลงแสงทผสมเขาไป เชน สาหรบ OLED เปลงแสงสแดง อาจใชสารทมแถบตองหามไมกวางนก เชน Alq3 เปนสารเจาบานได แตสาหรบ OLED เปลงแสงสนาเงน จา เปนตองใชสารทมแถบตองหามกวางมากขน เชน AND
วธการหลกในการเพมประสทธภาพของ OLED ทาไดโดยการปรบสมดลระหวางอเลกตรอนและโฮลทถกฉด
เขาสชนสารเปลงแสง ซงเปนททพาหะทงคจะสามารถรวมตวกนเปนเอกซตอน และอาจสลายตวใหพลงงานแสงเปลงออกมาได สารอนทรยสวนใหญมการนาโฮลไดดกวา
NO
N
O
NO
Al
Alq 3, green ADN, Blue
O
S
N
ON
Coumarin dye, green
N
CH3
CH3
O CH3
CNNC
DCM, red
รปท 6 ตวอยางสารอนทรยและสารประกอบ โลหะอนทรยทถกใชเปนชนเปลงแสงแบบวาวแสง

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
221
N
NEu
O
O
3 Eu(dbm)3(Phen) , Red
IrO
ON
S
3 Ir(btp)2(acac) , Red
Ir
N
NN
Ir(ppy)3 , Green
IrN
O
O
2 Ir(ppy)2(acac) , Green
Ir
N
N
OO
N
F
F
F
F
FIrPic , Blue
IrN
N
3 mer-Ir(Pmb)3 , Blue
รปท 7 ตวอยางสารประกอบโลหะอนทรยทถกใชเปนชนเปลงแสงแบบฟอสฟอเรสเซนท
อเลกตรอน สงผลใหจานวนโฮลทเขาสชนสารเปลงแสงมจานวนมากกวาอเลกตรอน โฮลสวนเกนจงไหลออกจากชนเปลงแสงไปโดยเปลาประโยชน ดงนน การเพมชนนาอเลกตรอน เชน Alq3 หรอชนปองกนโฮล เชน BCP แทรกระหวางชนสารเปลงแสงกบแคโทดจงสามารถเพมประ- สทธภาพของ OLED ได นอกจากน ยงพบวาการเพมชนนาโฮล เชน TPD, NPB หรอพวก phthalocyanines เปนตน แทรกระหวางชนสารเปลงแสงกบขวแอโนด สามารถเพมประสทธภาพและอายการใชงานของ OLED ได ทงนเนอง จากวสดโปรงแสงทใชทาขวแอโนดมกเปน ITO ซงมฟงก ชนงานสง แตสารอนทรย เปลงแสงสวนใหญม HOMO ทตากวานน ดงนน การเลอกใชสารนาโฮลทม HOMO ใกลเคยงกบฟงกชนงานของ ITO ทาใหโฮลถกฉดจาก ITO เขามายงชนสารอนทรยนาโฮลเพอสงตอไปยง
ชนสารอนทรยเปลงแสงไดสะดวกขน อกทงชนสารนาโฮลนยงสามารถปองกนไมใหอนเดยมจาก ITO แพรผานเขาไปยงชนสารเปลงแสง ซงทาใหอายการใชงานและประสทธภาพของ OLED ตาลงได
โดยปกตสารอนทรยหรอสารโลหะอนทรยทใชใน
การสราง OLED มคณสมบตเปนฉนวน มความสามารถในการนาพาหะทางไฟฟาตา การเตมสารบางชนดเขาไปในสารอนทรยเหลานเพอใหเปนสารกงตวนาและนาพาหะ
ทางไฟฟาไดดขนเปนอกวธหนงในการเพมประสทธภาพ
ของ OLED การทาใหชนสารนาโฮลนาพาหะบวกไดดขนหรอการเจอแบบ p (p-type doping) ทาไดโดยการเตมสาร เจอ(14) ตวอยางเชน F4-TCNQ ซงโมเลกลของสารนมหม
ดงอเลกตรอนอยจานวนมาก เมอเจอเขากบสารนาโฮลจะทาใหสารนาโฮลเกดสภาวะพรองอเลกตรอนและสามารถ
นาโฮลไดดขน ตวอยางสารอนทรยและสารประกอบ
โลหะอนทรยท ถกใชในหนาทอนๆ นอกจากเปนตวเปลงแสง แสดงในรปท 8
การทาใหชนนาอเลกตรอนนาอเลกตรอนไดดขน
หรอการเจอแบบเอน (n-type doping) มกทาโดยการเจอชนสารนดวยไอออนโลหะ เชน ล เธยมหรอซ เซยม ไอออน(14) ซงเปนตวใหอเลกตรอนทดลงในชนสารนาอเลกตรอน เชน Alq3 กรณทใชลเธยมไอออน ทาโดยเคลอบชน LiF หนาประมาณ 1 นาโนเมตร ลงบนชนสารนาอ เลกตรอน จากนนดวยการเคลอบขวโลหะ เชน อลมเนยมทบชน LiF น พลงงานความรอนจากการเคลอบขวโลหะนจะสงผลใหลเธยมไอออนแพรเขาสชนสารนา
อเลกตรอนได ขอเสยของการใชลเธยม คอ มขนาดอะตอมทเลก ลเธยมจงแพรเขาไปยงชนสารอนทรยไดงาย เมอแพรเขาไปยงชนสารอนทรยเปลงแสง ลเธยมเปนตวระงบการเปลงแสงได สงผลใหประสทธภาพและอายการใชงานของ OLED ลดลงในทสด ตรงกนขาม ซเซยมมขนาดอะตอมทใหญกวาลเธยมมาก จงยากตอการแพรเขาสชนสารอนทรย ในการเจอสารอนทรยดวยซเซยมมกทาไปพรอมกบการเคลอบชนสารอนทรย ทงนผลจากการทดลอง พบวาการใชซเซยมเจอแบบเอนในสารอนทรยนาอเลกตรอนไมไดสงผลลบตออายการใชงานของ OLED(9)
นอกจากสารอนทรยโมเลกลเลกทกลาวมาแลว สารอนทรยประเภทพอลเมอรกไดถกศกษาเพอนามา ใชในการสราง OLED เชนกน โดยเรมจากงานวจยของ Burroughs และคณะในป พ.ศ. 2533 ซงใช poly(p-phenylene vinylene) (PPV) เปนสารเปลงแสง นบเปนการเปดทางสการศกษา OLED ทใชพอลเมอรเปนสารเปลง แสง และยงมการศกษาความเปนไปไดในการใชพอลเมอร เปนชนนาโฮลดวย โดย Gustafsson และคณะ(7) ในป พ.ศ.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
222
NN
TPD
NN
NPB
CuN
NN N
N
N
N
N
CuPC
AND
NN BCP
F4-TCNQ
รปท 8 ตวอยางสารอนทรยและสารประกอบโลหะอนทรยทถกใชในการสราง OLED โดยทาหนาทอน ๆ ทไมใชเปนตวเปลงแสง
2535 นา polyaniline ทถกโดปดวย camphor sulphonic acid ใหนาไฟฟาได มาใชเปนชนนาโฮลสาหรบ OLED ทม poly[2-methoxy,5-(2′-ethyl-hexoxy)1,4-phenylene- vinylene] เปนชนเปลงแสง
พอลเมอรทนยมนามาใชเปนชนเปลงแสงมาจาก
สาร 3 กลมไดแก 1) PPV และอนพนธของมน 2) polythio-phene และอนพนธของมน และ 3) polyfluorene และ
อนพนธของมน นอกจากโฮโมพอลเมอรเหลานจะถก
สงเคราะหและนามาใชในการสราง OLED แลว ยงมการสงเคราะหและนาโคพอลเมอรมาใชดวย ตวอยางพอล-เมอรทถกใชในการสราง OLED แสดงในรปท 9
อายการใชงานของ OLED ขนกบเสถยรภาพของสารอนทรยทใชในการสราง OLED และโลหะทใชทาขว ไฟฟา โดยทวไปสารอนนทรยทใชในการสราง OLED มกวองไวตอการเกดปฏกรยาเคม กลาวคอ งายตอการถกออกซไดซดวยออกซเจนและไฮโดรไลซดวยนา เชนเดยว กบโลหะทใชทาชนแคโทด ซงเพอความเหมาะสมควรมฟงกชนงานทใกลเคยงกบ LUMO ของสารอนทรย ทมคาในชวง -2 ถง -3 eV เพอใหการฉดอเลกตรอนจากขวโลหะเขาส ชนสารอนทรยเปนไปไดโดยงาย แตโลหะทมคาฟงก ชนงานในชวงน วองไวตอการเกดปฏ กรยากบ
ออกซเจนและนาเชนกน ดงนนในการยดอายการใชงานของOLED จาเปนตองมการปองกนไมใหออกซเจนและนาเขาไปสมผสกบสารอนทรยและขนโลหะ โดยการหอหม OLED ซงอาจทาไดดวยการใชแผนโลหะหรอกระจกปดหมดานหลงของ OLED และดวยการหอหมทดพบวา OLED สามารถมอายการใชงานไดสงถง 65,000 ชวโมง (Kodak claimed, www.oled-info.com) ซงเพยงพอสาหรบการนาไปใชในการทาจอแสดงผลสาหรบอปกรณ
อเลกทรอนกสทวไป หรอนาไปใชเปนแหลงกาเนดแสงทดแทนหลอดไฟ ซงคาการสองสวางทมากขนจะสงผลให OLED มอายการใชงานสนลง โดยนกวจยของบรษท Universal Display Corporation อางวา สามารถสราง OLED เปลงแสงสขาวทมพอลเมอรเปนชนเปลงแสงซงมประสทธภาพในการใชพลงงานสงถง 75 lm/w และมอายการใชงานมากกวา 35,000 ชวโมง ทความสองสวางเรมตน 1,000 cd/m2 แนวโนมการพฒนาดานอน ๆ
เปนททราบกนดวา ระบบสญญากาศมราคาท
คอนขางแพง ดงนนแนวโนมหนงของการพฒนา OLED จงเปนการเปลยนหรอทดแทนบางสวนของการเคลอบ
ฟลมดวยระบบเคลอบแบบการหมนเหวยงทไมตองใช
ระบบสญญากาศ หรอใชการพมพแบบพนหมก หรอเทคโนโลยการพมพแบบสกรนซงจะชวยเพมความ
ไดเปรยบของ OLED เทยบกบการแสดงผลแบบอนๆ นอกจากน ยงพฒนาวสดทใชทาซบสเตรตเพอทดแทนกระจกโดยเพมคณสมบตความโคงงอได เพอใหสามารถประยกตเปนจอแบบมวนได หรอสามารถแสดงผลบนเสอผาหรอเครองนงหมได รปท 10 แสดงตวอยางอปกรณเปลงแสงททาบนพลาสตกทโคงงอได
สรป บทความนนาเสนอแงมมตางๆ ทเกยวของกบอป-
กรณเปลงแสงจากสารอนทรย รวมทงเสนทางการพฒนาหลกการทางาน คณลกษณะหลกๆ ของอปกรณ สถาปตย-กรรมดานโครงสราง ซงบรณาการศาสตรดานฟสกส เคม และศลปดานวศวกรรมโครงสรางของตวอปกรณ เพอการตระหนกและเขาถงความสาคญและเหนแนวโนมของการ
พฒนาอปกรณเปลงแสงจากสารอนทรย
CN
CN
NC
NC
FF
FF

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
223
.n
Poly(p-phenylene vinylene)
S
n Polythiophene
n Polyfluorene รปท 9 ตวอยางพอลเมอรทถกใชใน
การสราง OLED NH NH N N
x y Polyaniline
รปท 10 อปกรณเปลงแสงจาก Alq3 ทโคงงอได เกดจากการเคลอบ อปกรณบนซบสเตรตพลาสตก (ตวอกษรภาษาญปนเปนชอยอของ Japan Society for the Promotion of Science (JSPS))
กตตกรรมประกาศ
ผลงานวจยและพฒนาอปกรณเปลงแสงจากสาร อนทรยน เปนความรวมมอระหวางมหาวทยาลยมหดลและศนยเทคโนโลยโลหะและวสดแหงชาต ผเขยนขอขอบคณ นกวจย ผรวมวจยทกทานในโครงการ “การวจยและพฒนาอปกรณอเลกทรอนกสชนดออนรวมถงอปกรณแสดงผล
อนทรยและวสดทใชประกอบ” ภายใตทนอดหนนการวจยโครงการความรวมมอกบตางประเทศ (ไทย-ญปน) ของสภาวจยแหงชาต และ JSPS ประจาป พ.ศ. 2549-2551 และการสนบสนนของศนยนาโนเทคโนโลยแหงชาต ผานศนยเครอขายดานวศวกรรมวสดนาโนและสานกงาน
กองทนสนบสนนการวจย เอกสารอางอง 1. Antoniadis H. Overview of OLED Display techno-
logy. Available from: URL:http://www.ewh.ieee. org/ soc/cpmt/presentations/cpmt0401a.pdf, 2003
2. Aukkaravittayapun A. Toward wet route coating for flexible TC films: general review. 5th International Symposium of Center for Hyper Media Research; 2008 October 11; Tokyo, Japan. 2008.
3. Bassler H, Hadziioannou G, van Hutten PF, editors. Semiconducting polymers: chemistry, physics and engineering. Germany: Wiley-VCH; 2000.
4. Bernanose A, Comte M, Vouaux P. A new method of emission of light by certain organic compounds, J Chem Phys 1953;50:64-8.
5. Burroughes JH, Bradley DDC, Brown AR, Marks RN, Mackay K, Friend RH et al. Light-emitting diodes based on conjugated polymers, Nature 1990; 347:539-41.
6. Dangtip S, Hoshi Y, Kasahara K, Onai Y, Osotchan T, Sawada Y et al. Study of low power deposition of ITO for top emission OLED with facing target and RF sputtering systems. J Phys: Conference Series 2008;100:042011(1-6).
7. Gustafsson G, Cao Y, Treacy GM, Klavetter F, Colaneri N, Heeger AJ. Flexible light-emitting diodes made from soluble conducting polymers. Nature 1992;357:477-9.
8. Helfrich W, Schneider WG. Recombination radia-tion in anthracene crystals. Phys Rev Lett 1965; 140:229-31.
9. Lee JH, Wu MH, Chao CC, Chen HL, Leung MK. High efficiency and long lifetime OLED basedon a metal-doped electron transport layer. Chem Phys Lett 2005;416:234-7.
10. Organic light emitting diode. Available from: URL: http://en.wikipedia.org/wiki/Organic_light-emitting _diode
11. Tang CW, VanSlyke SA. Organic electrolumines-cent diodes. Appl Phys Lett 1987;51:913-5.
12. Uchida T, Mimura T, Ohtsuka M, Otomo T, Ide M, Shida A et al. Cesium-incorporated indium-tin-oxide films for use as a cathode with low work function for a transparent organic light-emitting device. Thin Solid Films 2006;496:75-80.
13. Vincett PS, Barlow WA, Hann RA, Roberts GG. Electrical conduction and low voltage blue electro-luminescence in vacuum-deposited organic films. Thin Solid Films 1982;94:171-83.
14. Walzer K, Maennig B, Pfeiffer M, Leo K. Highly efficient organic devices based on electrically doped transport layers. Chem Rev 2007;107:1233-71.


225
เซลลแสงอาทตยแบบสยอมไวแสง
สภาวด เกยรตเสว1 และ ยทธนา ตนตรงโรจนชย1,2 1ภาควชาเคม ศนยพลงงานทางเลอก และศนยนวตกรรมทางเคม คณะวทยาศาสตร มหาวทยาลยมหดล 2โครงการ Computational Nanoscience Consortium ศนยนาโนเทคโนโลยแหงชาต สานกงานพฒนาวทยาศาสตรและเทคโนโลยแหงชาต
บทคดยอ
เซลลแสงอาทตยชนดสยอมไวแสงไดรบความสนใจเปนอยางมากทงในวงการวจยเชงวชาการและเชงพาณชย เนองจากเซลลประเภทนมประสทธภาพในการเปลยนพลงงานแสงใหเปนพลงงานไฟฟาสงใกลเคยงกบเซลลแสงอาทตย
ชนดฟลมบางจากสารอนนทรย ประกอบกบไมจาเปนตองใชเทคโนโลยการผลตทสงนก ทาใหมแนวโนมคอนขางสงทจะสามารถผลตไดในราคาตนทนทตากวาเซลลแสงอาทตยชนดฟลมบางจากสารอนนทรย บทความนจงรายงานความเปนมาและประเภทของเซลลแสงอาทตย โครงสรางและหลกการทางาน รวมทงเสนอขอมลความกาวหนาของการพฒนาเซลลแสงอาทตยชนดสยอมไวแสงในปจจบน
คาสาคญ: เซลลแสงอาทตยชนดสยอมไวแสง; เซลลแสงอาทตย; สยอม; พลงงานทางเลอก
Abstract
Dye-sensitized solar cells (DSSCs) have gained recent significant interests from both academic and industrial researchers, as DSSCs have achieved high energy conversion efficiency comparable to that of thin-film solar cells based on inorganic materials. In addition, this type of cell can be manufactured with potentially more cost-effective compared to silicon- or inorganic-based solar cells. This report summarizes the history of DSSCs development, their structures and principles of operation, including the latest research and development.
Keywords: dye-sensitized solar cells; solar cells; dyes; alternative energy
บทนา
วกฤตนามนฟอสซลกาลงเปนปญหาใหญทสงผล
กระทบตอการดารงชวตและภาคเศรษฐกจของประเทศท
ตองพงพาการนาเขานามนเชอเพลงเพอการพฒนาประเทศ ประกอบกบการผลตพล ง ง านไฟฟ าซ งนบว า เปน
ปจจยพนฐานทสาคญในการพฒนาประเทศยงจาเปนตอง
ใชพลงงานจากเชอเพลงฟอสซลเปนหลก ทาใหประเทศไทยตองมงความสนใจไปทการพฒนาพลงงานทางเลอก
และพลงงานหมนเวยนเพอทดแทนการใชเชอเพลงจาก
ฟอสซลและเพอสรางเสถยรภาพดานพลงงานของประเทศ
อยางจรงจง
พลงงานแสงอาทตยเปนแหลงพลงงานทางเลอก
หนงทสาคญยง เพราะพลงงานแสงอาทตยเปนพลงงานทดเหมอนจะไมมวนหมดสน โลกของเราดดกลนพลงงาน
แสงอาทตยประมาณ 3,850 zettajoules (ZJ) ตอป (zeta = 1021) พลงงานจากดวงอาทตยเพยง 1 ชวโมงมปรมาณ
มากกวาพลงงานทมนษยโลกใชทงป ค.ศ. 2002(22) พลงงานแสงอาทตยเปนพลงงานสะอาดทไมกอใหเกดมลพษกบ
สงแวดลอม แตกตางจากแหลงพลงงานอน เชน เชอเพลงฟอสซลทปลดปลอย CO2 หรอพลงงานนวเคลยรฟชชนใน
ปจจบนซงอาจเกดปญหาการแพรกระจายสารกมมนต- ภาพรงส นอกจากน พลงงานแสงอาทตยยงเปนตนกาเนดของพลงงานทางเลอกอนๆ เชน พลงงานลม เปนตน ซงการดดซบพลงงานแสงอาทตยของบรรยากาศของโลก สงผลใหมการถายเทความรอน การระเหยและการควบแนนของไอนา สดทายทาให เกดกระแสลมและกระแสนา
ตงแตโบราณกาลมนษยใชประโยชนจากแสงและ
ความรอนจากดวงอาทตย ในปจจบนมนษยเราพยายาม

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
226
เปลยนพลงงานแสงอาทตยมาเปนพลงงานไฟฟาเพอใชกบ
เครองใชไฟฟาตางๆ แตปญหาสาคญทสดของการนา
พลงงานแสงอาทตยมาใช คอ ทาอยางไรจงจะสามารถเปลยนพลงงานแสงอาทตยมาเปนพลงงานไฟฟาไดอยางม
ประสทธภาพ ทาใหมการคดคนเซลลแสงอาทตยขน
เซลลแสงอาทตย เปนอปกรณท เปล ยนแปลง
พลงงานแสงอาทตยเปนกระแสไฟฟาโดยอาศยหลกการ
ของปรากฏการณโฟโตอเลกทรก ถอกาเนดเปนครงแรกในป ค.ศ. 1883 โดยนกวทยาศาสตรชอ Fritts ซงใชแวนผลกซ ล เนยมเปนองคประกอบ (21) ตอมาในป ค .ศ . 1954 Pearson, Fuller และ Chapin นกวจยจากหองปฏบตการ Bell ประเทศสหรฐอเมรกา ไดคดคนเซลลแสงอาทตยจากแวนผลกซลกอนขนเปนครงแรก และพฒนาโดยมแรงขบเคลอนจากโครงการสารวจอวกาศของ NASA และ
ปญหาวกฤตน ามนในป ค .ศ . 1973 หลงจากนนเซลลแสงอาทตยไดรบการพฒนาใหมประสทธภาพสงขนเรอยๆ จนในชวงทศวรรษท 80-90 กาลงการผลตไฟฟาจากเซลลแสงอาทตยสงขน 10-20 เปอรเซนตตอป และตงแตป ค.ศ. 2002 เปนตนมา อตราการเตบโตเทากบ 48 เปอรเซนตตอป โดยในชวงป ค.ศ. 2006 กาลงการผลตไฟฟาจากเซลลแสงอาทตยของทงโลกรวมกนมคามาก กวาในป ค.ศ. 2000 ถง 6 เทา(5,23)
เซลลแสงอาทตยในปจจบนมทงแบบดงเดมซงม
การใชแวนผลกซลกอน ทงแบบเปนผลกเดยวและผลกผสม และยงมเซลลแสงอาทตยแบบใหมทใชเทคโนโลยฟลมบางมาใชในการผลต ประเภททสองนใชวธสรางชนสารทไวตอแสงทบางมากๆ บนแผนรองซงอาจจะเปนแกว โลหะ หรอพลาสตก เพอลดตนทนการผลต เนองจากแวนผลกซลกอนนนมราคาสง แรงขบเคลอนสาคญสาหรบการพฒนาเทคโนโลยฟลมบางนเรมจากการขาดแคลนพอล-ซลกอนหรอซลกอนผลกผสมในป ค.ศ. 2004 ตอมาในป ค.ศ. 2006 พอลซลกอนมากกวาครงทผลตออกมาถกสงเขาไปสกระบวนการผลตเซลลแสงอาทตย แทนทจะถกนาไปผลตชปคอมพวเตอร ถงแมวาประสทธภาพในการเปลยนพลงงานแสงใหเปนพลงงานไฟฟาของเซลลแสงอาทตย
ชนดฟลมบางนจะยงไมสงเทยบเทากบเซลลประเภท
ซลกอนแบบดงเดม แตเซลลแสงอาทตยชนดฟลมบางสามารถถกนาไปใชประโยชนไดหลากหลายกวา เนองจากความยดหยนของวสดทใชและดวยราคาตนทนทตากวา เพราะเซลลชนดซลกอนมคาใชจายในการผลตทสง ในกระบวนการผลตมการใชพลงงานจานวนมากในการ
เตรยมซลกอนใหมความบรสทธตามตองการ
เซลลแสงอาทตยชนดฟลมบางมหลายประเภท วสดทนยมใชมาสรางฟลมบางกนมากทสดคอซลกอนแบบ อสณฐาน (a-Si:H) และยงมการพฒนาใชวสดใหมอนๆ อกดวย เชน CuInSe2 GaAs และ CdTe รวมทงฟลมบางของผลกซลกอนบนแผนรองราคาถก เปนตน ในกรณของฟลมบางของผลกซลกอนนน ความหนาของแผนฟลมอยท 10-30 μm ซงบางกวาความหนาของแวนผลกซลกอนถง 10 เทา(1) เซลลแสงอาทตยชนดฟลมบางทกลาวไปขางตนน
ลวนแตมสารอนนทรยเปนองคประกอบหลก นอกจากนยงมเซลลแสงอาทตยแบบอนทรยชนดฟลมบาง (organic thin film solar cells) และเซลลแสงอาทตยชนดสยอมไวแสง (dye-sensitized solar cells หรอ DSSCs หรอ DSCs)
เซลลแสงอาทตยแบบอนทรยชนดฟลมบางนนใช
สารกงตวนาทเปนสารอนทรยมาประกอบเปนฟลมบางๆ หนาประมาณ 100 nm(13) สารกงตวนาอนทรยเหลานน ไดแก สารโมเลกลเลก เชน copper phthalocyanin สาร ประกอบพอลเมอร เชน polyphenylene vinylene และสารโมเลกลใหญอยาง fullerene ในขณะทเซลลแสงอาทตย
ชนดสยอมไวแสงอาศยการดดกลนแสงของสยอม ซงสวนใหญเปนสารเชงซอนของรธเนยมทยดเกาะบนฟลมบาง
ของสารกงตวนา TiO2 ทเปนอนภาคเลก ๆ
ตารางท 1 สรปประสทธภาพของเซลลแสงอาทตยประเภทตางๆ พบวาเซลลแสงอาทตยชนดฟลมบางจาก สารอนนทรยใหประสทธภาพคอนขางสง อยางไรกตาม ขนตอนการผลตสารกงตวนาอนนทรยตองใชเทคโนโลย
ขนสงและยงกอใหเกดมลพษมากมาย ดงทไดกลาวมาแลว ในขณะทเซลลแสงอาทตยแบบอนทรยชนดฟลมบางม
ระยะการแพรของเอกซตอน (exciton เปนสภาวะทอเลก- ตรอนรวมตวกบ hole) คอนขางสน ซงไปจากดประสทธ- ภาพของตวเซลล ประสทธภาพการเปลยนพลงงานแสงใหเปนพลงงานไฟฟาสงสดทไดถกรายงานไวมคา 3-5% ตางจากเซลลประเภทสยอมไวแสงซงใหประสทธภาพทสง
กวา ขอดของเซลลชนดหลงนคอ สามารถประยกตใชกบวสดทหลากหลาย ในการผลตเซลลใชเพยงเทคโนโลยการพมพสกรนซงทาใหตนทนการผลตตากวาเซลลประเภทอน
ทตองใชวสดกงตวนาในปรมาณมาก จงทาใหมการวจยมากมายทพยายามพฒนาเซลลแสงอาทตยชนดสยอมไว
แสงในเชงพาณชย อยางไรกตาม ยงมอกหลายปจจยทตองพฒนาเซลลประเภทนใหดยงขนอก เชน ความคงทนของเซลลเมออยภายใตความรอนและแสงแดดเปนเวลานาน และการปดผนกเซลลเพอปองกนไมใหสารละลายภายใน
รวไหลออกมา

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
227
ตารางท 1 ประสทธภาพการเปลยนพลงงานแสงใหเปนพลงงานไฟฟาสงสดของเซลลแสงอาทตยประเภทตางๆ
ประเภทของเซลล ประสทธภาพรวม (η)* เอกสารอางอง
ซลกอนผลกเดยว (Single crystalline silicon) 24.7% (28) ซลกอนผลกผสม (Multicrystalline silicon หรอ Polysilicon) 20.3% (20) เซลลแสงอาทตยชนดฟลมบางของซลกอนอสณฐานและไมโครครสตลไลนซลกอน (a-Si/μc-Si)
11.7% (4)
เซลลแสงอาทตยชนดฟลมบางของ Cu(In,Ga)Se2 หรอทเรยกวา CIGS 18-19.4% (15) เซลลแสงอาทตยชนดฟลมบางของ CdTe 16.5% (2) เซลลแสงอาทตยแบบอนทรยชนดฟลมบาง 3-5% (3,12,19,27) เซลลแสงอาทตยชนดสยอมไวแสง 9.5-11.18% (6,14)
*η คอ ประสทธภาพการเปลยนพลงงานแสงอาทตยเปนพลงงานไฟฟา คานวณไดจาก
เมอ Voc คอ open circuit voltage, Jsc คอ ความหนาแนนกระแสไฟฟาลดวงจรตอหนงหนวยพนทของโฟโตอเลกโทรด (short circuit current density), I0 คอ ความเขมแสงทตกกระทบตอหนงหนวยพนทของโฟโตอเลกโทรด
และ ff คอ คา fill factor คานวณไดจาก โดย Pmax คอ คากาลงสงสดของเซลลแสงอาทตย
η ปกตจะถกวดท AM 1.5 โดย AM เปนตวยอของ air mass คอความเขมของแสงอาทตยทตกกระทบถงผวโลก สามารถอธบายไดในรปของมวลอากาศ โดยคา AM = 1/cosθZ เมอ θZ คอมม zenith มมนคอมมระหวางแนวดง (เสนตรงในแนวตง) กบดวงอาทตย เชน เมอดวงอาทตยอยตรงศรษะพอด มคา θZ = 0o ผลรวมระหวางมม Zenith กบมมมองดวงอาทตยมคาเทากบ 90°
เนองจากเซลลแสงอาทตยชนดสยอมไวแสงนนม
ประสทธภาพใกลเคยงกบเซลลแสงอาทตยชนดฟลมบาง
จากสารอนนทรย และในการผลตไมจ า เปนตองใชเทคโนโลยขนสงนก ทาใหมแนวโนมคอนขางสงทประเทศไทยจะสามารถแขงขนในการพฒนาเซลล
แสงอาทตยประเภทนกบนานาประเทศได บทความนกลาวถงหลกการทางาน และความกาวหนาของการพฒนาเซลลแสงอาทตยชนดสยอมไวแสง
โครงสรางและหลกการทางานของเซลลแสงอาทตยชนดสยอมไวแสง
เซลลแสงอาทตยชนดสยอมไวแสง (dye-sensitized solar cell; DSSC) ถกพฒนาในชวงตนทศวรรษท 90 โดยศาสตราจารย Grätzel(18) (Laboratory for Photonics and Interfaces, Swiss Federal Institute of Technology, Lausanne ประเทศสวตเซอรแลนด) เปนเซลลไฟฟาจากพลงงานแสง (photovoltaic cell) ทเลยนแบบกระบวนการสงเคราะหแสงในธรรมชาต มการใชสยอมเปนตวรบพลงงานแสงอาทตย (light sensitizer) เพอทาใหเกดการกระตนอเลกตรอนในสยอมไปอยในสถานะเรา พรอมจะสงถายตอไปยงสารกงตวนากลมไทเทเนยมไดออกไซด (TiO2) ขนาดโครงสรางระดบ 250-300 นาโนเมตร ทมพนทผวจานวนมากเพอเพมปรมาณโมเลกลสยอม หลงจากนนอเลกตรอนจะเคลอนทไปยงขวอเลกโทรดโปรงแสงท
เคลอบดวยชนสารนาไฟฟาอยาง fluorine-doped SnO2
(FTO) แลวเคลอนสวงจรภายนอก (load หรอ storage) อเลกตรอนจะกลบมายงขวอเลกโทรดอกดานทมตวเรง
ปฏกรยาทางไฟฟาเคม (Pt) เคลอบอย โดยม I3- รบ
อเลกตรอนหรอทเรยกวาถกรดวซไปเปน I - และเคลอนทสสยอม เพอคนอเลกตรอนตอสยอม จงครบกระบวนการเคลอนทของอเลกตรอน สามารถสรปกลไกการทางานของเซลลแสงอาทตยชนดสยอมไวแสง ดงแสดงในรปท 1
ความกาวหนาของการพฒนาเซลลแสงอาทตยชนดสยอมไวแสง
ความกาวหนาสาคญททาใหเซลลแสงอาทตยชนดส
ย อมไวแสง เป นท สน ใจอย า งม าก เ ก ด จ ากการทศาสตราจารย Grätzel ไดเปลยนแปลงการใชผลก TiO2 ทส
ยอมมายดจบไดในลกษณะฟลมชนเดยวมาเปนการใช
ไทเทเนยมไดออกไซดโครงสรางระดบนาโนเมตรทมร
พรนจานวนมาก มพนทใหสยอมจบเกาะไดเปนปรมาณมาก ทาใหเกดการปรบปรงคาประสทธภาพการเปลยนพลงงานแสงไปเปนกระแสไฟฟ าท ส งขนมาก ซ งศาสตราจารย Grätzel ไดรายงานถงระบบฟลม meso-porous TiO2 ทม Ru(2,2’-bipyridine-4,4’-dicarboxylic acid)3 [Ru(dcbpy)3] และ cis-Ru(dcbpy)2(H2O)2 เปนสยอม
ซงมคาของ incident photon to current conversion efficiency (IPCE) ถง 80% ทความยาวคลนทมการดดกลนแสงสงสดของ Ru(dcbpy)3 ตอมามการ เปล ยนมาใช Ru(dcbpy)2(NCS)2 หรอ N3 dye พบวาเมอทาการวดภายใต
η = J S C x V O C x f f / I 0 x 1 0 0
f f =P m a x
J S C x V O C

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
228
ความเขมแสงท AM 1.5 ใหคาประสทธภาพรวม (η) ท 10% ซงคาสงสดในปจจบนทมรายงานไวเทากบ 11.18%(6) IPCE คอประสทธภาพการเปลยนโฟตอนเปนกระแสไฟฟาของเซลล
แสงอาทตยซงขนกบความยาวคลนแสงทตกกระทบ คานวณไดจาก
IPCE[%] =1240[eV.nm] x JSC[μAcm-2]
λ[nm] x φ[μWcm-2]x 100
เมอ φ คอความเขมแสงทตกกระทบตอหนงหนวยพนทของโฟโตอเลก-
โทรด และ λ คอความยาวคลนของแสงทตกกระทบ
กระบวนการรบแสงโดยสยอมอาจถกพจารณาวา
เปนการสรางอเลกตรอนและ hole โดยจะมการเคลอนท
ของอเลกตรอนผาน TiO2 ขณะทการเคลอนทของ hole ในกรณน คอการเคลอนทของ I - ทนาอเลกตรอนคนกลบมายงสยอม ซงกระบวนการรบแสงโดย light sensitizer เชนนมขอดคอชวยลดกระบวนการรวมตวของอเลกตรอนและ hole (electron-hole recombination) ดงเชนทเกดในระบบซลกอน แตกทาใหเกดรอยตอ (interface) จานวนมาก ซงประสทธภาพโดยรวมจะขนกบอตราการถายเทอเลกตรอน
ภายในองคประกอบยอยและระหวางรอยตอตางๆ มการศกษาทดลองถงอตราการถายเทอเลกตรอน พบวากระบวนการถายเทอเลกตรอนจากสยอม N3 ไปยง TiO2 ใชชวงเวลาประมาณ 50 ± 25 fsec(8,24) ขณะทการถาย
อเลกตรอนยอนกลบใชเวลานานกวามากคออยในชวง μsec-msec และการถายอเลกตรอนยอนกลบไปจาก TiO2 สอเลกโทรไลตกใชเวลานานในชวง msec-1 sec การถายอเลกตรอนคนจากอเลกโทรไลตสสยอมทถกกระตนแลว
ใชเวลาประมาณ 110 nsec และเชอกนวาปจจยทนาจะ กาหนดอตราการถายเทอเลกตรอนนาจะอยในสวนของ
โครงสรางทเปนรพรนของ TiO2 เพราะจะเปนบรเวณทม
รอยตอระหวางกนมากทสด การเคลอนทของอเลกตรอนจะประพฤตตวตามกฎการแพร จงมแนวคดทจะพฒนา TiO2 ทมโครงสรางเปนระเบยบเพอชวยใหอตราการถายเท
อเลกตรอนดขน เชนเดยวกนกบการเคลอนทของอเลก-ตรอน การเคลอนทของ hole หรอ I -/ I3
- electrolyte กเปนปจจยสาคญตอประสทธภาพของระบบ เนองจากเปนกลไกการเคลอนทตามกระบวนการแพร ซงถกคาดวาเปนสวนทเปนขนกาหนดอตราเรวในการเพมประสทธภาพของเซลล
แสงอาทตยชนดสยอมไวแสง จงมความจาเปนตองเขาใจปจจยเหลานวามผลตอขนาดของปรมาณดงกลาวอยางไร เชน อะไรเปนปจจยใดตอการ photoharvest หรอการถายเทอเลกตรอนไปยงอเลกโทรดโปรงแสงซงเปนทมาของ
โจทยวจยทตองการพฒนา ปรบปรง และศกษาพนฐานของโครงสรางองคประกอบตางๆ ของเซลลแสงอาทตยชนดส
ยอมไวแสง เพอใหเขาใจความสมพนธของโครงสรางและคณสมบต ตลอดจนความสมพนธขององคประกอบตางๆ ทสงผลตออตราการเคลอนทบรเวณรอยตอ ดงนนการพฒนาเซลลแสงอาทตยชนดสยอมไวแสงจงตองศกษา
หลายองคประกอบควบคกนไป ซงสามารถสรปได 4 แนวทางหลก ดงน
1. การพฒนาและสงเคราะหโมเลกลสยอมไวแสงชนดใหมทมความทนทานตอแสงแดดเปนเวลานาน และมความสามารถในการดดกลนแสงตลอดชวงความยาวคลน
แสงตาเหนจนถงแสงชวงใกลอนฟราเรด 2. การพฒนาและศกษาดานไฟฟาเคมของอเลก-
โทรไลตเพอใหมประสทธภาพสง 3. การพฒนาโครงสรางของอนภาค TiO2 เพอใหม
ระยะทางการแพรของอเลกตรอนเพมขน 4. การพฒนาขวอเลกโทรดโปรงแสงใหมความ
ยดหยน เพอการใชงานไดหลายรปแบบ
การพฒนาและสงเคราะหโมเลกลสยอมไวแสง
สยอมเปนสวนทรบแสงโดยตรงมาจากแสงอาทตย สารพวกนเปนสารพวกทมส จงมกเรยกวาสยอม และเปนทมาของชอเซลลแสงอาทตยชนดน เพอใหประสทธภาพของเซลลแสงอาทตยมคามากทสด โมเลกลสยอมจะตองมสมบต 5 ประการ คอ
1. ประสทธภาพการดดกลนแสงสง บรเวณชวง
แสงจากดวงอาทตย (ชวงกวางทตามองเหนไดจนถงชวงใกลอนฟราเรด)
2. สามารถถายเทอเลกตรอนไปยง TiO2 ไดดโดยม
พลงงานพอดกบ energy gap ของมน 3. สามารถถายทอดอเลกตรอนกลบจากอเลกโทร-
ไลตไปยงสยอมทถกออกซไดสไดด 4. สามารถนามายดเกาะกบพนผวของสารกงตวนา
ไทเทเนยมไดออกไซดได 5. มความคงทนตอการใชงานหรอการถกแสง
อาทตยเปนเวลานาน
โมเลกลทถกนามาใชกนมากจงมกเปนสารประ-กอบเชงซอนของธาตโลหะทรานสชน (Fe Ru Os) กบทม pi-conjugation มากๆ เพราะมความสามารถในการถาย ทอดอเลกตรอนภายในโมเลกลไดด เชน porphyrin, bipyri-dine, terpyridine โดยชนดทมคณสมบตดทสดนนพฒนาโดยกลมของศาสตราจารย Grätzel ซงใชลแกนดอนทรย
กลมพอลไพรดลขนาดโมเลกลใหญยดจบกบโลหะรธ
เนยมตรงกลางในลกษณะสารเชงซอน สยอมทรจกกน ดคอ cis-dithiocyanatobis(4,4′-dicarboxy-2,2′-bipyridine)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
229
รปท 1 กลไกการทางานของเซลลแสงอาทตยชนดสยอมไวแสง (ดดแปลงจาก7)
ruthenium (II) (N3 หรอ red dye), cis-dithiocyanate-N,N′-bis-(4-carboxylate-4′-tetrabutyl-ammonium-carboxylate-2,2′-bipyridine) ruthenium (II) (หรอ N719), trithio-cyanato(4,4′,4′′-tricarboxy-1-2,2′:6′,2′-terpyridine) ruthe- nium(II) complex (หรอ black dye) และสยอม K19 โครง สรางดงแสดงในรปท 2
สยอม N3 มคาประสทธภาพ IPCE ทสงถง 80% แตดดกลนแสงไดเพยงในชวงความยาวคลน 400-600 nm เทานน และมคา molar extinction coefficient (ε )ทตา ใน ขณะทสยอม K19 มคา ε = 18.2×103
M-1cm-1 ซงนบวา
คอนขางสง สวน ε ของสยอม N719 มคาเทากบ 14.0×103
M-1cm-1) ในแงประสทธภาพ (η) ภายใตสภาวะเดยวกน N3 (10%) จะใหประสทธภาพสงกวา K19 (7.0%) และ N719 (6.7%) นอกจากนเซลลแสงอาทตยจากสยอม K19 สามารถคงทนอยภายใตแสงท 60°C ไดนานกวา 1,000 ชวโมง โดยไมเสยสภาพ แตเทาทมรายงานมา black dye คอสยอมทดทสด เนองจากมความสามารถในการดดกลนแสงตลอดชวงทตามองเหนจนถงชวงใกลอนฟราเรด คอท 920 nm และใหประสทธภาพ IPCE 80% และ η >10% ทสงกวาสยอมชนดอนๆ(16)
ในขณะเดยวกน มผใหความสนใจกบการพฒนาสยอมกลมสารอนทรยและสยอมจากธรรมชาต เชนในกลมของ cyanidine dyes, perylene pigments, xanthene ตลอด
จน coumarin derivatives ซงมรายงานถงคาประสทธภาพη ในยาน 5-6% โดยขอจงใจของสารสยอมกลมอนทรยมาจากราคาทถกกวาและสงเคราะหไดงายกวา(9-10) อยางไรกตาม สยอมกลมหลงนยงไมคงทนเทยบเทากบกลมสารเชงซอนของรธเนยมได
การพฒนาและศกษาดานไฟฟาเคมของอเลกโทร-ไลต
อเลกโทรไลตทาใหการเคลอนทของอเลกตรอน
ครบวงจร ระยะแรกๆ นยมใชระบบของคปฏกรยารดอกซ I-/I3
- ดงทไดกลาวไปแลวเนองจากเปนของเหลว ทาให
เตรยมไดงาย แตเมอระบบอยใตสภาวะการใชงานจรงทอาจมอณหภมสงถง 70-80°C พบวาความดนไอทสงขนทาให เกดแรงดนเพมขนสงผลให เกดการรวซม และมความชนเขามาปนเปอนในระบบ ทาใหเซลลแสงอาทตยชนดสยอมไวแสงมอายการทางานทสน จงมการพฒนานาของเหลวไอออนทมจดเดอดสงขนและสามารถนาไฟฟา
ไดดมาใชแทน(26) เชน เกลอของ imidazolium แตพบวายงมปญหาเรองการรวซมอย และมคา η ไมสงนก เนองจากของเหลวไอออนนมความหนดสง ทาให I- เคลอนทไดชา จากนนมการพฒนาใชของผสมระหวาง 1-methyl-3-pro- pylimidazolium iodide, 1-methyl-3-ethylimidazolium dicyanamide และ lithium iodide กบเซลลสยอมไวแสงทใชสารประกอบเชงซอนโลหะรธเนยม ซงสามารถใหคา η
Pt/แกว (อเลกโทรดเคานเตอร) FTO/แกว
สยอม TiO2
ขนท 1 สยอม (S) ดดกลนแสงอาทตย อเลกตรอนของสยอมถกกระตน S + hν --> S* ขนท 2 อเลกตรอนจากสยอมถกถายเทไปยง FTO โดยผาน TiO2 S* --> S+ + e- (ไปยง TiO2) ขนท 3 อเลกตรอนเคลอนไปยงอเลกโทรดเคานเตอรหลงจาก
เครองใชไฟฟาไดรบพลงงานไปแลว ขนท 4 ทอเลกโทรดเคานเตอร
1/2I3- + e- (จากขว Pt) --> 3/2I-
ขนท 5 ทสยอม 3/2I- --> 1/2I3- + e-
S+ + e- --> S

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
230
NN
TBAOOC
N
COOTBA
COOTBA
S C N
SCN
Ru
SCN
N3 dye
NN
HOOC
N
N
COOH
COOH
COOHS C N
SCN
Ru
black dye
NN
HOOC
N
N
COOTBA
COOTBA
COOHS C N
SCN
Ru
N719
NN
HOOC
N
N
COOH
S C N
SCN
Ru
O
O
K19
รปท 2 โครงสรางของสยอม N3, black dye, N719 และ K9 (TBA = Bu4N-)
ทสงถง 6.6% เพราะอตราเรวในการคนอเลกตรอนตอส
ยอมดงกลาวเพมขน
ขณะเดยวกนกเรมมการใชวสดแขงและกงเหลวกง แขง(17) ไดแก พอลเมอรเจล พอลเมอรอเลกโทรไลต หรอการนาของเหลวไอออนผสมกบพอล เมอร เจลท เปน อเลกโทรไลต ดงเชนทมรายงานการใช 1-methyl-3-propylimidazolium iodide กบ poly(vinylidenefluoride-co-hexa-fluoropropylene) ซงอยในรป quasi-solid state elec-trolyte พบวาใหคา η = 5.3% ลดลงกวาการใชสารละลาย I2/I3
- เพยงเลกนอย ภายใต AM 1.5(25)
นอกจากระบบทกลาวขางตน ยงมการใชสารอนๆ เชน นาโนคอมโพสท และอเลกโทรไลตทเปนฟลมบาง เปนตน ซงยงตองไดรบการพฒนาตอไปอก
การพฒนาโครงสรางของอนภาค TiO2
TiO2 อเลกโทรดเปนสวนประกอบทสาคญของ
เซลลแสงอาทตยชนดสยอม TiO2 มคณสมบตเปนสารกงตวนาทม band gap คอนขางกวาง (3.2 eV) เสถยร มราคาถก และ TiO2 ใน anatase phase มพนทผวทไวตอปฏกรยาสงทาใหสามารถซมซบสยอมลงบนพนผวไดมาก ผลกระดบนาโนของ TiO2 มหนาทรบอเลกตรอนตอจากสยอม และเปนทยดเกาะของสยอมดวยหมคารบอกซลกทเปน
สวนประกอบของสยอม หมฟงกชนนจงมอยในสยอมตางๆในรปท 2 ขนาดของผลกระดบนาโนนนมความ สาคญตอความสามารถในการถายเทอ เลกตรอนมาก เนองจากยงขนาดเลกยงมพนทผวมากสาหรบการถายเท นอกจากนสยอมทดยงตองระดบพลงงานในสภาวะเราท
สมพนธกบ energy gap ของ TiO2 โดยทวไปจะเตรยมจาก
สารคอลลอยด TiO2 โดยใชเทคนคการพมพสกรนลงบน
แผนแกวทเคลอบดวย ITO (indium tin oxide) หรอ FTO รายละเอยดของวธการเตรยม ความบรสทธ ขนาดความพรน ความเปนผลก และรปรางผลก ของ TiO2 รวมถงอณหภมเผา ความหนาของชนฟลม และสาร binder ทใชมผลตอการยดตดของสยอมและการเคลอนทของประจเปน
อยางมาก และถงแมจะมการศกษาถงอทธพลของปจจยเหลานมาบางแลว แตยงไมมขอสรปทแนนอน TiO2 จง
ยงคงตองไดรบการศกษาพฒนาตอไปอกอยางตอเนอง เพอเพมประสทธภาพรวมของเซลลแสงอาทตยสยอมไวแสง
ใหสงขน
การพฒนาขวอเลกโทรดโปรงแสง
ขวไฟฟาสาหรบเซลลแสงอาทตยชนดสยอมไวแสง
ตงแตระยะเรมแรกจนถงปจจบนยงนยมใชแกวทเคลอบ
ดวย ITO หรอ FTO แมวาขวไฟฟาประเภทนจะมการใชอยางแพรหลาย สามารถหาซอไดงายในอตสาหกรรม และใหเซลลทมประสทธภาพสง แตการประยกตใชงานมขอจากด เปราะแตกงาย แตกตางจากลกษณะของแผนรองพลาสตกโปรงแสงหรอขวอเลกโทรดทมพลาสตกเปน
องคประกอบทมความยดหยน นาหนกเบา พบงอได ไมเปนสนม ทนตอการกดกรอนไดด โดยทวไป อนภาคระดบนาโนสามารถยดเกาะเปนแผนฟลมบนพนผวพลาสตกได
โดยใชเทคนคตางๆ เชน การใชแรงกดอด การใชรงสไมโครเวฟ การตกผลกแบบไฮโดรเทอรมล electrophoretic deposition และ film transfer เปนตน อยางไรกตาม ปญหาสาคญของการพฒนาเซลลแสงอาทตยชนดสยอมไวแสง
บนแผนรองพลาสตกคอ การเผาเพอใหเปนเนอเดยว (sintering) ของ TiO2 บนพลาสตกนนตองทาทอณหภม

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
231
คอนขางตา เพราะพลาสตกไมอาจทนกระบวนการทอณหภมสงเชนแบบทใชเตรยมเซลลสยอมไวแสงปกตได (ประมาณ 400°C) จงกอใหเกดปญหาการไมเกาะตดของชน TiO2 บนแผนรองยดหยน ท าใหการบดงอและประสทธภาพของเซลลทไดมขอจากด (η ประมาณ 2%)(11) สงผลใหในปจจบนมงานวจยเกยวกบการเตรยมขวไฟฟา
โปรงแสงนบนพลาสตกออกมานอยมาก แตเมอไมนานมาน มรายงานชนแรกจากกลมวจยในประเทศสงคโปรเกยวกบการเตรยมลวดนาโนของสารกงตวนา ZnO ดวยวธไฮโดรเทอรมลบนแผนรองทเปน polyethylene terephta- late (PET)/ITO ทอณหภม 85°C เพอใชเปนขวอเลกโทรดสาหรบเซลลแสงอาทตยชนดสยอมไวแสง โดยทดสอบกบสยอม N719 พบวาเซลลทไดสามารถทนแรงบดไดด แผนฟลมไมเปราะเมอบดงอ และมประสทธภาพทดกวาเซลลยดหยนอนๆ ทไดเคยมรายงานมากอน ถงแมวาจะยงไมดเทากบเซลลปกตทใชแกวเปนอเลกโทรด ดงนน จงยงคงตองมการวจยพฒนาหาแนวทางการผลตเซลลบน
พลาสตกทยดหยนตอไป
สรป
เซลลแสงอาทตยชนดสยอมไวแสงสามารถไดรบ
การพฒนาใหมขดความสามารถทจะถกผลตในเชงพาณชย
และทดแทนเซลลแสงอาทตยแบบซลกอนทใชอยใน
ปจจบนได โดยตองพฒนาองคประกอบทงสควบคกนไป ไดแก สยอม ระบบอเลกโทรไลต อนภาคนาโน TiO2 และข วอ เล กโทรด เพ อ ให เหมาะกบการใช ง านจร ง มประสทธภาพสงและมความคงทน ซงจาเปนตองใชนกวทยาศาสตรหลายสาขาเขามาแกปญหาจากมมมองท
หลากหลาย ทงนกเคมอนทรย นกเคมอนนทรย นกเคมวเคราะห นกวทยาศาสตรและเทคโนโลยพอลเมอร นกเคมเชงฟสกส และนกฟสกส เพอการพฒนาใหเกดเปนเซลลแสงอาทตยจากองคความรของคนไทยทใชงานไดในขน
ตอไป
เอกสารอางอง 1. Birkmire RW, Shafarman WN, Eser E, Hegedus
SS, McCandless BE, Aparicio R et al. Processing materials, devices and diagnostics for thin film photovoltaics: fundamental and manufacturability issues. [Online] [2002?] [cited 2008 Dec 10]; [146 screens]. Available from: URL: http://www.udel. edu/iec/iec 2002 nrelan nual.pdf
2. Bosio A, Romeo N, Podestà A, Mazzamuto S, Canevari V. Why CuInGaSe2 and CdTe poly-crystalline thin film solar cells are more efficient
than the corresponding single crystal? Cryst Res Technol 2005;40:1048-53.
3. Campbell WM, Jolley KW, Wagner P, Wagner K, Walsh PJ, Gordon KC et al. Highly efficient porphyrin sensitizers for dye-sensitized solar cells. J Phys Chem C 2007;111:11760-2.
4. Debajyoti D, Madhusudan J. P-doped μc-Si:H films at a very low thickness and high deposition rate: suitable for application in solar cells. J Mater Res 2003;18:2371-8.
5. Dorn JG. Solar cell production jumps 50 percent in 2007. [Online] 2007 December 27 [cited 2008 Dec 10];[3 screens]. Available from: URL: http://www.earth-policy.org/Indicators/Solar/ 2007. htm
6. Grätzel M. Conversion of sunlight to electric power by nanocrystalline dye-sensitized solar cells. J Photochem Photobiol A 2004;164:3-14.
7. Grätzel M. Solar energy conversion by dye-sensitized photovoltaic cells. Inorg Chem 2005; 44:6841-51.
8. Hannappel T, Burfeindt B, Storck F, Willig F. Measurement of ultrafast photoinduced electron transfer from chemically anchored Ru-dye molecules into empty electronic states in a colloidal anatase TiO2 film. J Phys Chem B 1997;101:6799-802.
9. Hara K, Sayama S, Ohga Y, Shinpo A, Suga S, Arakawa H. A coumarin-derivative dye sensitized nanocrystalline TiO2 solar cell having a high solar-energy conversion efficiency up to 5.6%. Chem Commun 2001;6:569-70.
10. Hara K, Kurashige M, Ito S, Shinpo A, Suga S, Sayama K et al. Novel polyene dyes for highly efficient dye-sensitized solar cells. CHEM Commun 2003;2:252-3.
11. Jiang CY, Sun XW, Tan KW, Lo GQ, Kyaw KK, Kwong DL. High-bendability flexible dye-sensitized solar cell with a nanoparticlemodified ZnO-nanowire electrode. Appl Phys Lett 2008; 92:143101-3.
12. Kim JY, Kim SH, Lee HH, Lee K, Ma W, Gong X et al. New architecture for high-efficiency polymer photovoltaic cells using solution-based titanium oxide as an optical spacer. Adv Mater 2006;18:572-6.
13. Kim JY, Lee K, Coates NE, Moses D, Nguyen TQ, Dante M et al. Efficient tandem polymer solar cells fabricated by all-solution processing. Science 2007;317:222-5.
14. Kuang D, Klein C, Ito S, Moser JE, Humphry-Baker R, Evans N et al. High-efficiency and stable mesoscopic dye-sensitized solar cells based on a high molar extinction coefficient

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
232
ruthenium sensitizer and nonvolatile electrolyte. Adv Mater 2007;19:1133-7.
15. Metzger WK, Repins IL, Contreras MA. Long lifetimes in high-efficiency Cu(In,Ga)Se2 solar cells. App Phys Lett 2008;93:022110-3.
16. Nazeeruddin MK, Péchy P, Renouard T, Zakeeruddin SM, Humphry-Baker R, Comte P et al. Engineering of efficient panchromatic sensitizers for nanocrystalline TiO2-based solar cells. J Am Chem Soc 2001;123:1613-24.
17. Nogueira AF, Longo C, De Paoli MA. Polymers in dye sensitized solar cells: overview and perspectives. Coor Chem Rev 2004;248:1455-68.
18. O’Regan B, Grätzel M. A low-cost, high-efficiency solar cell based on dye-sensitized colloidal TiO2 films. Nature 1991;353:737-40.
19. Peumans P, Uchida S, Forrest SR. Efficient bulk heterojunction photovoltaic cells using small-molecular-weight organic thin films. Nature 2003; 425:158-62.
20. Schulz O, Glunz SW, Riepe S, Willeke GP. High-efficiency solar cells on phosphorus gettered multicrystalline silicon substrates. Prog Photo-voltaics Res Appl 2006;14:711-9.
21. Smith J. Solar energy: hope for a brighter future. Energy current news for the business of energy. [Online] 2007 Sep 11 [cited 2008 Dec 10];[5 screens]. Available from: URL: http://www. energycurrent.com/index.php?id=3&storyid=6620
22. Solar Power Set to Shine Brightly. [Online] 2007 May 22 [cited 2008 Dec 10];[4 screens]. Available from: URL: http://www.worldwatch. org/node/5086
23. Solar Power Set to Shine Brightly. [Online] 2007 May 22 [cited 2008 Dec 10];[4 screens]. Available from: URL: http://www.worldwatch. org/node/5086
24. Tachibana Y, Moser JE, Grätzel M, Klug DR, Durrant JR. Subpicosecond interfacial charge separation in dye-sensitized nanocrystalline titanium dioxide films. J Phys Chem 1996;100: 20056-62.
25. Wang P, Zakeeruddin SM, Exnar I, Grätzel M. High efficiency dye-sensitized nanocrystalline solar cells based on ionic liquid polymer gel electrolyte. Chem Commun 2002;24:2972-3.
26. Wang P, Zakeeruddin SM, Moser JE, Grätzel M. A new ionic liquid electrolyte enhances the conversion efficiency of dye-sensitized solar cells. J Phys Chem B 2003;107:13280-5.
27. Yamanari T, Taima T, Sakai J, Saito K. Highly efficient organic thin-film solar cells based on
poly(3-hexylthiophene) and soluble C70 fullerene derivative. Jpn J Appl Phys 2008;47: 1230-3.
28. Zhao J. Recent advances of high-efficiency single crystalline silicon solar cells in processing technologies and substrate materials. Sol Energy 2004;82:53-64.

233
การประยกตใชกลองจลทรรศนแรงอะตอมในการศกษาทางชววทยา
ธราพร พนธธรานรกษ ภาควชาชววทยา คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
กลองจลทรรศนแรงอะตอม (atomic force microscopy; AFM) ประดษฐขนในป ค.ศ. 1986 เพอใชในการศกษาพนผวของวสดแขงในระดบอะตอม ตอมาไดถกนาไปประยกตใชอยางแพรหลายในการศกษาทางวทยาศาสตรชวภาพ กายภาพ วสดศาสตรและอตสาหกรรม ประโยชนของเครอง AFM ทเหนอกวากลองจลทรรศนชนดความละเอยดสงประเภทอนๆ คอ สามารถศกษากระบวนการทางชววทยาในระดบโมเลกลในระบบทใกลเคยงกบภาวะทางสรรวทยา ขอมลทไดจากการศกษาดวยเครอง AFM มประโยชนในการนาไปประยกตใชไดหลากหลาย และการศกษาสภาพภมลกษณของพนผวไมไดเจาะจงเฉพาะตวอยางทเปนผลกเทานน หากแตยงสามารถศกษาโมเลกลชวภาพตางๆ รวมทงเซลลและอนๆ ตวอยางเชน สามารถใชเครอง AFM ศกษาคณสมบตการเขาจบกนแบบจาเพาะของโมเลกลชวภาพตางๆ ในระบบทางชววทยาได การนา AFM มาประยกตใชในงานวจยทาใหเกดการบรณาการความรและเปนการเปดโลกทศนสการนาไป
ประยกตใชศกษาปรากฏการณทางชววทยาทยากตอการศกษาโดยวธการอนๆ
คาสาคญ: กลองจลทรรศนแรงอะตอม; โมเลกลชวภาพ; การเขาจบกนของโมเลกลแบบจาเพาะ
Abstract
Atomic force microscopy (AFM), developed in 1986 to explore atomic details on hard material surfaces, has proven to be a powerful tool for biological studies. The numbers of AFM applications in life, physical and material sciences, and in industries have exploded. The particular advantage of AFM over other high-resolution methods commonly used in biological sciences is that AFM allows biomolecules to be investigated not only under physiological conditions but also while biological processes are at work. The information generated with AFM can provide several applications. The detailed topological information is not restricted to crystalline specimens, hence, single biomolecules, cells and others samples can be directly monitored. AFM can also provide insight into binding properties of biological systems. Application of AFM in biological research can pave a way for a better understanding of how to integrate AFM into biological studies, especially in observing various biological phenomena that are hard to observe in real space.
Keywords: atomic force microscopy; AFM; biomolecule; single-molecule recognition บทนา
ปจจบนองคความรแบบบรณาการของวทยาศาสตร
สาขาตางๆ เชน ฟสกส เคม ชววทยา และคณตศาสตร สามารถกอประโยชนอยางมากตอความกาวหนาทางดาน
งานวจยพนฐานและประยกต และใชอธบายปรากฏการณตางๆ ในธรรมชาตไดเปนอยางด อยางไรกตาม ในโลกทางชววทยายงคงมคาถามอกมากมายทรอนกวจยมาชวยกน
คนควาหาคาตอบ การพฒนาเทคนควธวจยใหมๆ จงมความสาคญเปนอยางมาก โดยเฉพาะการพฒนาเพอนาไป สการตอบโจทยปญหาในระดบโมเลกล ในภาวะทางสรร-
วทยาใหมากทสด การเขาใจคณสมบตตางๆ ของสสารในระดบโมเลกลหรออะตอมนนจะทาใหเขาใจพฤตกรรมของ
สสารนนในระดบโครงสรางทใหญขนไดเปนอยางด ปจจบนศาสตรทางดานเทคโนโลยชวภาพระดบนาโนได
ถกนามาประยกตใช กนอยางแพรหลายมากขน เชน การศกษาชววทยาโครงสราง (structural biology) ของโมเลกล การตรวจวเคราะหโรค (clinical diagnostics) การใหยาเขารางกาย (drug delivery) เปนตน(13,14,17) การศกษาทางดานเทคโนโลย ชวภาพระดบนาโนนนสามารถ
ดาเนนการไดโดยใชเครองมอหลากหลายชนด อยางไรกด

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
234
การศกษาทภาวะใกลเคยงกบภาวะจรงทางสรรวทยายงม
ขอจากด กลองจลทรรศนแรงอะตอม (Atomic Force Microscopy; AFM)(1) เปนหนงในเครองมอทเหมาะสม
สาหรบการศกษาทางดานเทคโนโลยชวภาพระดบนาโน
ในภาวะทใกลเคยงกบภาวะทางสรรวทยา โดยเครองมอนสามารถตรวจวดสมบตของพนผวและแสดงภาพของ
พนผวตางๆ ไดในระดบอะตอม และจากการทเครอง AFM สามารถนาไปใชศกษาพนผวไดหลากหลายชนด และมขนตอนการเตรยมตวอยางเพอการศกษาทไมยงยาก ราคาไมแพง สามารถไดขอมลแบบสามมต (3-dimensional information) และยงมขอดอนๆ อกมากมาย จงทาใหมการประยกตใชเครอง AFM กนอยางแพรหลาย โดยเรมตนจากการนาไปใชศกษาทางดานวสดศาสตร (material science) และวทยาศาสตรกายภาพ (physical science) รวมถงนาไป ใชทางดานอตสาหกรรม และเมอประมาณ 10 ปทผานมา ไดมการประยกตใชกบการศกษาทางดานชววทยา(12,26)
เครอง AFM จดอยในกลมเครองมอ scanning probe microscopy (SPM) โดยเครองมอในกลมนจะใช
ปลายแหลมทมขนาดเลกระดบนาโนเมตรลากไปบนพนผว
และตรวจสอบคณสมบตของพนผวนนทงในเชงปรมาณ
และคณภาพ เชน วดแรงกระทาระหวางอะตอมทปลายเขมกบอะตอมหรอโมเลกลบนพนผว หรอวดการลอดผานไดแบบ ควอนตมของอเลกตรอน ปลายแหลมทมขนาดเลกระดบอะตอมหรอโมเลกลนนทาใหสามารถวดคณสมบต
ตางๆ ระดบอะตอมหรอโมเลกลไดอยางละเอยดเชนกน สงผลใหการศกษาวตถทมขนาดเลกระดบนาโนเมตร
สามารถทาไดอยางถกตองและแมนยา เครอง AFM ถก
พฒนาจากเครอง scanning tunneling microscopy (STM) ทประดษฐขนในป ค.ศ. 1981 โดยนกวทยาศาสตรสาขา
ฟสกส 2 ทาน คอ Binning และ Rohrer ซงเปนชาวเยอรมนและสวสเซอรแลนด ตามลาดบ ไดรวมกนพฒนาเครองมอนในขณะททางานอยทศนยวจย IBM เมองซรก ประเทศสวสเซอรแลนด และจากการคดคนประดษฐเครอง STM นนสงผลใหทงสองไดรบรางวลโนเบลสาขาฟสกสในป ค.ศ. 1986 และในปเดยวกนนนเอง Binning และคณะฯ อก 2 คนคอ Quate และ Gerber ไดรวมกนประดษฐเครอง AFM เครองแรกสาเรจ(1) จากความสาเรจในการประดษฐเครอง STM และ AFM ถอเปนการเปดประตสโลกของ
พนผวในระดบนาโนเทคโนโลย โดยสามารถทาการศกษาวสดตางๆ ในระดบอะตอมและทราบถงโครงสรางการจดเรยงตวของอะตอมไดเปนอยางด จงนามาสองคความรและความเขาใจทมประโยชนอยางมากในสาขาวชาตางๆ
เครองมอในกลม SPM ยงถกพฒนาอยางตอเนองอยเสมอ โดยทาใหเกดเครองมอทางเทคนคใหมๆ มากกวา 20 ชนดทอาศยหลกการใชปลายแหลมขนาดเลกเพอตรวจคณ สมบตดานตางๆ ของพนผวทแตกตางกน เชน เครอง electrostatic force microscopy (EFM), force modulation microscopy (FMM), Kelvin probe force microscopy (KPFM), magnetic force microscopy (MFM) เปนตน จากการพฒนาและวจยอยางกวางขวางประมาณ 20 ปทผานมา ทาใหเกดกลมวจยในประเทศตางๆ มากมาย ทงในแถบยโรป สหรฐอเมรกา และเอเชย และยงมการเชอมโยงระหวางมหาวทยาลยกบบรษทผผลตตางๆ เชน บรษท Agilent Technologies (www.nano.tm.agilent.com) และ Veeco Instruments Inc. (www.veeco.com) นอกจากนนยงมการประชมทางวชาการประจาปเกยวกบเครองมอ SPM อยางตอเนอง สาหรบประเทศไทย การศกษาวจยโดยใชเครองมอในกลม SPM ยงมอยคอนขางนอยมากและจากดอยเพยงการประยกตใชสาหรบการตรวจวดคณสมบตของ
วสดทางฟสกสและพอลเมอร บทความนกลาวถงการศกษาวจยทางชววทยาในระดบโมเลกล โดยใชเครอง AFM เนอหาครอบคลมหลกการทางานของเครองมอ รวมถงแนวทางการนาไปประยกตใชเพอการทางานวจย
ทางดานชววทยา เพอเปนการเปดโลกทศนสการนาไปประยกตใชใหกวางขวางมากขนในวงการวจยของประเทศ
ไทย
หลกการทางานของกลองจลทรรศนแรงอะตอม เครอง AFM เปนเครองมอสาหรบใชงานดาน
วทยาศาสตรระดบนาโน โดยสามารถศกษาแบบความคม ชดสง (high resolution) ทมความละเอยดมากอยในระดบถง 0.1 นาโนเมตร เครอง AFM ประกอบดวยอปกรณ
ตรวจจบหรอหวตรวจ (probe) ทมปลายแหลมเลกซงตดอยกบคานยนทโกงงอได (cantilever) (รปท 1) โดยปลายหว วดหรอ AFM tip นนมรศมทปลายเขม (end radius) อยในระดบเพยงไมกนาโนเมตร (โดยทวไปมขนาดประมาณ 5-20 นาโนเมตร) ดงนนกระบวนการผลตคานยนชนดนจะอาศยการผลตแบบ microlithography คลายคลงกบการ
ผลตชป (chip) ของคอมพวเตอร เมอเครอง AFM ลากสวนปลายเขมแหลมนไปบนพนผวของตวอยาง จะมแรงกระทาระหวางอะตอมทปลายเขมแหลมกบอะตอมหรอโมเลกล
บนพนผวเกดขนเชน แรง van der Waals และแรงพนธะเคมอนๆ จากแรงกระทานเองสามารถทาใหคานยนเบนหรอ โกงงอ (เกด deflection ซงเปนไปตาม Hooke’s law)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
235
รปท 1 ภาพแสดงหววด (probe) คานยน (cantilever) และปลาย
แหลม (AFM tip) (Veeco Instruments Inc.)
และจะถกตรวจวดไดดวยแสงเลเซอรทสองผานไปกระทบ
ทสวนปลายของคานยนและสะทอนมายงจดรบแสง (photodiodes หรอ laser detector) นนเอง (แสดงในรปท 2) ถงแมวาแรงกระทาจะมขนาดนอยมากในระดบพโคนวตน (picoNewton, pN) กตาม แตเครอง AFM กสามารถตรวจ วดขนาดของแรงปฏสมพนธระหวางความสมพนธเชง
ตาแหนงของสวนปลายเขมแหลมกบพนผวของวตถใน
ขณะทลากไปบนพนผวไดดวยความชวยเหลอของอปกรณ
ควบคมตาแหนง (piezoelectric tube) ซงจะถกนามาแปรสญญาณและประมวลผลดวยระบบคอมพวเตอรเพอนามา
สรางเปนภาพสามมตของพนผวทมกาลงขยายสงไปแสดง
บนมอนเตอร ทาใหสามารถศกษาลกษณะเชงโครงสราง
ของพนผวในระดบอะตอมหรอโมเลกล รวมทงขนาดและการจดเรยงตวได
รปท 2 หลกการทางานของเครอง Atomic force Microscopy (AFM)
ระบบการทางานของเครอง AFM สามารถแบงออกไดเปน 2 แบบ คอ แบบสมผส (contact mode) และแบบพลศาสตร (dynamic mode หรอ intermittent contact mode) ลกษณะการทางานของเครองแบบสมผสนน ปลายเขมแหลมจะสมผสพนผวตลอดเวลาในขณะทลากไปบน
ผวตวอยาง เครองจะควบคมใหแรงระหวางปลายเขมกบผวมคาคงทโดยอาศยสญญาณปอนกลบของคานยนทโกงงอ ขอเสยของวธนคอ จะทาใหเกดแรงตานในแนวของการเคลอนทซงขนานกบพนผวขน หากใชวธนจงควรใชหววดทมคานยนชนดทมคาคงทของสปรงนอยมาก สวนวธการทางานของเครองแบบพลศาสตร คานยนจะแกวงกวดหรอสนใกลเคยงความถเรโซแนนซหรอฮารมอนก ทาใหปลายเขมแหลมสมผสพนผวเปนระยะเวลาสนๆ ในแนวตงฉากกบพนผว เครองจะควบคมแอมพลจดการสนใหคงท ดวยลกษณะการสมผสแบบนแรงตานในแนวขนานกบพนผว
จงเกดนอยกวาวธสมผส เครองแบบพลศาสตรนจงใชกนมากกบตวอยางทางชวภาพ ซงบางครงอาจเรยกชอไดตางๆ กนตามแตวธการทาใหคานสน เชน tapping mode อาศยกระแสไฟฟาจาก piezoelectric element หรอ magnetic AC mode (MAC mode) ทอาศยสนามแมเหลก เปนตน
จากการตรวจวดคณสมบตของพนผวดวยเครอง AFM สามารถไดขอมลหรอสญญาณหลายชนดประกอบกน ทงน ขนกบวธการทางานของเครอง เชน วธสมผสจะไดสญญาณภาพรายละเอยดเฉพาะสวนของพนผว สญญาณความเบนหรอโกงงอของปลายเขม สญญาณความเสยดทานระหวางปลายเขมกบพนผว สวนวธพลศาสตรนนจะไดสญญาณภาพรายละเอยดเฉพาะสวนของพนผว สญญาณความเปลยนแปลงของแอมพลจด สญญาณความเปลยนแปลงของเฟส และยงสามารถวดสภาพยดหยนของพนผว การยดตดหรอประสาน และความแขงตงของพนผวตวอยางไดอกดวย การประยกตใชกลองจลทรรศนแรงอะตอมในการ ศกษาทางชววทยา
เครอง AFM สามารถนาไปประยกตใชในการ ศกษาพนผววสดไดมากมายหลากหลายชนดเชน พนผวโลหะ แกว พอลเมอร เซรามก และรวมถงโมเลกลชวภาพตางๆ เชน โปรตน เซลล ดเอนเอ และยงสามารถตรวจวดพนผวทงชนดทนาและไมนาไฟฟาไดอกดวย ซงถอเปนคณลกษณะจาเพาะพเศษของเครอง AFM ทมมากกวา
เครอง STM นอกจากนเครอง AFM ยงสามารถศกษา
คณสมบตของพนผวไดทงในสภาพแหงและในสภาพทอย
ในนา และไมจาเปนตองมการเคลอบพนผว ดวยโลหะหรอคารบอนใหเปนพนผวทนาไฟฟา ซงจาเปนตองทาในกรณการศกษาดวยกลองจลทรรศนอเลกตรอนแบบสองกราด (scanning electron microscope, SEM) นอกจากนยงไมจาเปนตองอาศยระบบสญญากาศซงมราคาแพงในการตด ตงอกดวย ดงนน เครอง AFM จงสามารถนามาประยกตใช
AFM Tip
Sample surface scanned back and forth
Mirror
Laser Laser detector
Display
Computer
AFM Tip
Sample surface scanned back and forth
Mirror
Laser Laser detector
Display
Computer

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
236
ในการศกษาโมเลกลชวภาพไดเปนอยางด โดยเฉพาะอยางยงสามารถศกษาในระบบทใกลเคยงกบภาวะทางสรรวทยา
ไดอกดวย อยางไรกด การประยกตใชเครอง AFM ยงคงมขอจากดอย คอตวอยางทตองการศกษาเชน เนอเยอ เซลล โปรตน หรอดเอนเอ นนจะตองยดตดกบพนผวรองรบ เชน แผนไมกา กระจกปดสไลด ซลคอนแผนบาง ไดเปนอยางด อยางไรกตาม นกวจยไดพฒนาวธการตางๆ ในการยดตดโมเลกลชวภาพซงโดยปกตนนจะอยในสภาพลองลอยใน
ของเหลวใหตดแนนบนพนผวรองรบได ตวอยางเชน การใชบฟเฟอรทมไอออนของโลหะชวยในการยดตดดเอนเอ
บนพนผวแกวหรอไมกา(11) การใชกรดอะมโน poly-L-lysine หรอการใชสารประกอบ Cell-Tax ในการชวยยดตดเซลล เมด เลอดแดงและเซลลชนดต างๆ บนกระจก สไลด (10,23) หรอการใชเจละตนในการชวยยดตดเซลลแบคทเรยบนแผนไมกา(7) เปนตน ยงไปกวานนยงสามารถใชปฏกรยาทางเคมหรอใชหมเคมของพนผวในการเชอม
ตดตวอยางกบพนผวรองรบดวยพนธะโคเวเลนซไดอกดวย ตวอยางเชน การยดตดแอนตบอดซงเปนโปรตนและถกดดแปลงโดยการเพม thiol group กบพนผวทองดวย disulfide bridge(9) อกวธหนงทสามารถใชยดตดโปรตนบนพนผวได เปนอยางดคอ การใชแรงยดตดระหวาง nitrilotriacetate (NTA) และ histidine 6 (His6) เรยกวา NTA-His6 system(6)
การศกษาทางสณฐานวทยาของเซลลและชววทยา
ระดบโมเลกลมประโยชนอยางมหาศาลตอการพฒนาดาน
การแพทย เทคนคการแพทย และการสาธารณสข เชนการพฒนายาเพอการรกษาโรคตางๆ และการพฒนาชดตรวจโรค(13,14) เปนตน ลกษณะตางๆ ของเซลลสงมชวตและกลไกการทางานของเซลล สณฐานวทยาและการเคลอนท รวมถงผลกระทบของเซลลตอโมเลกลชวภาพหรอสารเคม
ตางๆ ยงคงเปนเปาหมายทนกวจยตองการคนหาและทาความเขาใจใหมากทสด เทคนคการประยกตใชเครอง AFM ในการศกษาทางชววทยาจงมบทบาทสาคญอยางมากในวงการวจยเพอนาไปสเปาหมายดงกลาวขางตน เนองจากการศกษาดวยเครอง AFM สามารถศกษาเซลลไดหลากหลายชนดทงเซลลสตว แบคทเรยและไวรสในสภาพจรงหรออยในระบบทใกลเคยงกบภาวะทางสรรวทยา(12,26) การศกษาดวยเครอง AFM กบระบบทางชววทยาไดถกประยกตใชอยางแพรหลายในระยะเวลาไมกปทผานมา เชน การศกษาโครงสรางของเซลล เยอหมเซลล แขนงของเซลล เชน filopodia, lamellipodia, membrane ruffles, microspike และ microvilli รวมถงการศกษาโปรตนบนผวเซลลและเยอจาลอง เปนตน(2,15,19,20,24) ดงตวอยางการศกษาสณฐานวทยาของเซลลและการกระจายตวของโปรตนบน
เยอจาลองดงแสดงในรปท 3 และ 4 ตามลาดบ(19,20)
การประยกตใชเทคนค AFM รวมกบเทคนคการยอมดวยสารเรองแสงฟลออเรสเซนซและเทคนคการศกษา
การนาพาไอออน (patch-clamp technique) สามารถชวยในการระบโปรตนแบบจาเพาะได (24) อยางไรกตาม ไดมการพฒนาเทคนคใหมๆ ขนเสมอ ทาใหเครอง AFM สามารถ
รปท 3 ตวอยางการศกษาสณฐานวทยาของเซลล A. ภาพ deflection AFM ของเซลล Chinese hamster ovary (CHO) ทยงมชวตทถกเพาะเลยงบนพนผวกระจก จากภาพแสดงรปราง ขนาด และโครงรางของเซลล (cytoskeleton) B. ภาพ deflection AFM ของเซลล osteoblast (เซลลกระดกชนดหนง) ทถกคงสภาพดวย 4% paraformaldehyde จากภาพแสดงในเหนรายละเอยดของผวเซลลและแขนงของเซลลไปยงเซลลขางเคยงไดอยางชดเจน [ภาพโดย ธราพร พนธธรานรกษ]

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
237
รปท 4 ตวอยางการศกษาการกระจายตวของโปรตนบนเยอจาลอง A. ภาพ topographical AFM ของเยอจาลอง (lipid bilayer membrane) บน
พนผวไมกา B. ภาพ topographical AFM ของเยอจาลองภายหลงการใสโปรตนฆาลกนายง Cry4Ba toxin [ภาพโดย ธราพร พนธธรานรกษ] ระบโปรตนจาเพาะบนผวเซลลไดโดยไมจาเปนตองยอม
ดวยสารฟลออเรสเซนซอกตอไป การพฒนาเทคนคน
ดาเนนการโดยกลมวจยของ Prof. Hinterdorfer รวมกบ
บรษท Agilent Technologies โดยเรยกเทคนคนวา TREC (Topography and RECognition System) ซงไดรบรางวล R&D 100 Awards จากนตยสาร R&D ในป ค.ศ. 2004 อกดวย ตวอยางงานวจยทใชเทคนค TREC เชน การศกษาโปรตน vascular endothelial-cadherin บนผวเซลล mouse myocardium(5) และโปรตน human ether-a-go-go-related (hERG) K+ channel บนผวเซลล HEK 293(4) เปนตน
เครอง AFM ยงสามารถประกอบเขากบชดควบคมสภาพแวดลอมไดแก เครองควบคมอณหภมและปรมาณกาซคารบอนไดออกไซด และสามารถเปลยนถายอาหารเลยงเชอหรอใสสารทตองการศกษาเพมเตมในขณะทาการ
ทดลองไดอกดวย ทาใหสามารถศกษารปรางและขนาด
ของเซลลทยงมชวตอย และศกษาการเปลยนแปลงของเซลลหรอโมเลกลชวภาพนนๆ ตอสภาวะตางๆ เชน การเปลยนแปลงคาความเปนกรด-ดางของอาหารเลยงเชอหรอการเปลยนแปลงอณหภมแบบตอเนองตามเวลาจรง (real-time)ไดอกดวย(26) ตวอยางงานวจยไดแก การศกษาผล กระทบของสารสลายโปรตนโครงรางตอรปรางและขนาด
ของเซลล(25) การหลดออกของไวรส (exocytosis) จากเซลลทตดเชอ(16) และการเปลยนแปลงรปรางของเกลดเลอด(8) เปนตน
เครอง AFM สามารถวดแรงกระทาไดถงระดบ พโคนวตน จงสามารถใชในการศกษาคณสมบตการเขา จบกนแบบจาเพาะของโมเลกลชวภาพตางๆ เชน โปรตน-โปรตน ดเอนเอ-ดเอนเอ ดเอนเอ-โปรตน และแอนตบอด-
แอนตเจนไดเชนกน โดยยดตดโมเลกลชวภาพจาเพาะทปลายเขมแหลม จากนนนาไปเขาจบกบเซลลหรอโมเลกลชวภาพบนพนผวตวอยาง แลวจงดงปลายเขมแหลมออก (ดงในรปท 5) วธการนเรยกวา force-distance cycle ซงสามารถวดแรงกระทา (unbinding force, fu) ทใชในการแยกโมเลกลชวภาพบนปลายเขมออกจากเซลลหรอ
โมเลกลชวภาพบนพนผวแบบจาเพาะได ตวอยางการศกษาดวยวธน เชน การศกษาการเขาจบของแอนตบอดแบบจาเพาะกบโปรตนขนสงโซเดยมและกลโคสบนผว
เซลลทยงมชวต และศกษาการเขาจบของนาตาลชนดตางๆ ทาใหสามารถเขาใจโครงสรางและคณสมบตการขนสง
นาตาลของโปรตนขนสงนไดดยงขน(18,20,21) วธการนยง
สามารถประยกตใชในการศกษาคณสมบตการคลายตว (unfolding) ของโปรตน(3) และสามารถวดความยดหยนของเซลล(22) ไดอกดวย จงเหนไดวาเครอง AFM มประ- สทธภาพมากและสามารถนาไปประยกตใชในงานวจยทาง
ชววทยาไดเปนอยางด
สรป เครอง AFM เปนเครองมอทใชงานทางวทยาศาสตร
ระดบนาโน โดยถกพฒนาและประยกตใชในการศกษาคณสมบตทางโครงสราง รปรางและขนาดของโมเลกลแบบทมความละเอยดในระดบสงมากถง 0.1 นาโนเมตร และยงสามารถนามาประยกตใชในการศกษาทางชวภาพ
ไดเปนอยางด โดยระบบทางชวภาพทนามาศกษานนสามารถอยในรปแบบทใกลเคยงกบภาวะทางสรรวทยาได
อกดวย ทาใหสามารถนาไปสการเปดโลกทศนโดยประยกตใชใหกวางขวางมากยงขนในงานวจยดานตางๆ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
238
รปท 5 ภาพจาลองวธการศกษาการเขาจบกนแบบจาเพาะ ของโมเลกลดวยวธ force-distance cycle
กตตกรรมประกาศ ขอขอบคณภาควชาชววทยา และศนยนาโนศาสตร
และนาโนเทคโนโลย คณะวทยาศาสตร มหาวทยาลย มหดล สานกงานกองทนสนบสนนการวจย และศนย นาโนเทคโนโลยแหงชาต สานกงานพฒนาวทยาศาสตรและเทคโนโลยแหงชาต ทใหทนสนบสนนการวจย
เอกสารอางอง 1. Binnig G, Quate CF, Gerber C. Atomic force micro-
scope. Phys Rev Lett 1986;56:930-3. 2. Braet F, Seynaeve C, De Zanger R, Wisse E.
Imaging surface and submembranous structures with the atomic force microscope: a study on living cancer cells, fibroblasts and macrophages. J Microsc 1998; 190:328-38.
3. Carrion-Vazquez M, Oberhauser AF, Fowler SB, Marszalek PE, Broedel SE, Clarke J et al. Mecha- nical and chemical unfolding of a single protein: a comparison. Proc Natl Acad Sci USA 1999;96:3694-9.
4. Chtcheglova LA, Atalar F, Ozbek U, Wildling L, Ebner A, Hinterdorfer P. Localization of the ergtoxin-1 receptors on the voltage sensing domain of hERG K(+) channel by AFM recognition imaging. Pflügers Arch Eur J Physiol 2008;456:247-54.
5. Chtcheglova LA, Waschke J, Wildling L, Drenckhahn D, Hinterdorfer P. Nano-scale dynamic recognition imaging on vascular endothelial cells. Biophys J 2007;93:L11-3.
6. Conti M, Falini G, Samori B. How strong is the coordination bond between a histidine tag and Ni- nitrilotriacetate? An experiment of mechano-
chemistry on single molecules. Angew Chem Int Ed Engl 2000;39:215-8.
7. Doktycz MJ, Sullivan CJ, Hoyt PR, Pelletier DA, Wu S, Allison DP. AFM imaging of bacteria in liquid media immobilized on gelatin coated mica surfaces. Ultramicroscopy 2003;97:209-16.
8. Fritz M, Radmacher M, Gaub HE. Granula motion and membrane spreading during activation of human platelets imaged by atomic force microscopy. Biophys J 1994;66:1328-34.
9. Grandbois M, Beyer M, Rief M, Clausen-Schaumann H, Gaub HE. How strong is a covalent bond? Science 1999;283:1727-30.
10. Grandbois M, Dettmann W, Benoit M, Gaub HE. Affinity imaging of red blood cells using an atomic force microscope. J Histochem Cytochem 2000; 48:719-24.
11. Han W, Lindsay SM, Dlakic M, Harrington RE. Kinked DNA. Nature 1997;386:563.
12. Hoh JH, Hansma PK. Atomic force microscopy for high-resolution imaging in cell biology. Trends Cell Biol 1992;2:208-13.
13. Jain KK. Applications of nanobiotechnology in clinical diagnostics. Clin Chem 2007;53:2002-9.
14. Jain KK. Nanobiotechnology-based drug delivery to the central nervous system. Neurodegener Dis 2007;4:287-91.
15. Muller DJ, Schoenenberger CA, Schabert F, Engel A. Structural changes in native membrane proteins monitored at subnanometer resolution with the atomic force microscope: a review. J Struct Biol 1997;119:149-57.
16. Ohnesorge FM, Horber JK, Haberle W, Czerny CP, Smith DP, Binnig G. AFM review study on pox viruses and living cells. Biophys J 1997;73:2183-94.
17. Preiner J, Tang J, Pastushenko V, Hinterdorfer P. Higher harmonic atomic force microscopy: imaging of biological membranes in liquid. Phys Rev Lett 2007;99:046102.
18. Puntheeranurak T, Kasch M, Xia X, Hinterdorfer P, Kinne RK. Three surface subdomains form the vestibule of the Na+/glucose cotransporter SGLT1. J Biol Chem 2007;282:25222-30.
19. Puntheeranurak T, Stroh C, Zhu R, Angsuthana- sombat C, Hinterdorfer P. Structure and distribution of the Bacillus thuringiensis Cry4Ba toxin in lipid membranes. Ultramicroscopy 2005;105:115-24.
20. Puntheeranurak T, Wildling L, Gruber HJ, Kinne RK, Hinterdorfer P. Ligands on the string: single-molecule AFM studies on the interaction of antibodies and substrates with the Na+-glucose co-transporter SGLT1 in living cells. J Cell Sci 2006;119:2960-7.
fu
1
2
3 4
ระยะทางความเบนข
องคานย
น (ca
ntile
ver d
eflec
tion)
fu
1
2
3 4
ระยะทางความเบนข
องคานย
น (ca
ntile
ver d
eflec
tion)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
239
21. Puntheeranurak T, Wimmer B, Castaneda F, Gruber HJ, Hinterdorfer P, Kinne RK. Substrate specificity of sugar transport by rabbit SGLT1: single-molecule atomic force microscopy versus transport studies. Biochemistry 2007;46:2797-804.
22. Radmacher M. Measuring the elastic properties of living cells by the atomic force microscope. Methods Cell Biol 2002;68:67-90.
23. Schilcher K, Hinterdorfer P, Gruber HJ, Schindler H. A non-invasive method for the tight anchoring of cells for scanning force microscopy. Cell Biol Int 1997;21:769-78.
24. Smith PR, Bradford AL, Schneider S, Benos DJ, Geibel JP. Localization of amiloride-sensitive sodium channels in A6 cells by atomic force micro-scopy. Am J Physiol 1997;272:C1295-8.
25. Wada H, Kimura K, Gomi T, Sugawara M, Katori Y, Kakehata S et al. Imaging of the cortical cyto-skeleton of guinea pig outer hair cells using atomic force microscopy. Hear Res. 2004;187:51-62.
26. You HX, Yu L. Atomic force microscopy imaging of living cells: progress, problems and prospects. Methods Cell Sci 1999;21:1-17.


241
การประยกตใชสมการเลเวลเซตในศาสตรดานอนๆ
พลลภ ฮวบสมบรณ ภาควชาคณตศาสตร คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ
วธการเลเวลเซตเปนกลวธเชงตวเลขทพฒนาขนเพอใชตดตามววฒนาการของหนาสมผส มจดเดนคอ สามารถใชอธบายการเปลยนแปลงของหนาสมผสในกรณทวธการอนไมสามารถกระทาได เชน กรณทหนาสมผสแตกออกจากกน หรอกรณทหนาสมผสรวมตวกน เปนตน วธการเลเวลเซตไดถกประยกตใชในดานตางๆ มากมาย ตวอยางเชน การจาลองปรากฏการณ การปรบความชดเจนของภาพถาย การสรางเรขภาพคอมพวเตอร และการพยากรณภยพบตทางธรรมชาต เปนตน
คาสาคญ: วธการเลเวลเซต
Abstract Level set methods are numerical techniques for tracking a moving interface over time. As these methods can handle such topological changes as merging or breaking interfaces. They have many applications, e.g. simulation of various phenomena, improving image quality, computer graphics and prediction of natural disasters.
Keywords: level set methods
บทนา
วธการเลเวลเซต (level set methods) คดคนโดย Osher และ Sethian ในป ค.ศ. 1988(7) เปนกลวธเชงตวเลข (numerical techniques) ทพฒนาขนเพอใชตดตามววฒนา การของหนาสมผส (interface) สมการเลเวลเซตมรปแบบทวไปอยในรปของสมการอนพนธยอยดงน
0|| =∇+∂∂ φφ Ft
(1)
เมอ ),,( tyxφ คอฟงกชนเลเวลเซต และ F คอ
ฟงกชนความเรวของการเคลอนทของหนาสมผส โดยทตาแหนงของหนาสมผส ณ เวลา t ใดๆ จะถกนยามโดยเซต { }0),,(|),( =tyxyx φ (รปท 1)
วธการเลเวลเซตมจดเดนคอ ความสามารถในการจาลองการเปลยนแปลงของหนาสมผสในกรณทวธการอน
ไมสามารถทาได เชน กรณทหนาสมผสแตกออกจากกน หรอหนาสมผสรวมตวกน เปนตน จดเดนอกประการหนงของวธการเลเวลเซตคอ คาตางๆ ทเกยวของกบคณสมบตของหนาสมผสสามารถคานวณไดโดยตรงจากฟงกชน เลเวลเซต เชน ความโคงของหนาสมผสสามารถคานวณได
จากสมการ
⎟⎟⎠
⎞⎜⎜⎝
⎛∇∇
⋅−∇=|| φ
φκ
หรอทศทางท ต งฉากกบหนาสมผส สามารถคานวณไดจากสมการ ||/ φφ ∇∇=nr เปนตน
รปท 1 ฟงกชนเลเวลเซต (รปสแดง) และหนาสมผส (ขอบของรป
สเทาดานบน) ณ เวลาตางๆ กน กรอบสเหลยมสฟาคอระดบซงมคาเทากบศนย ดงนนรอยตดระหวางกรอบสฟากบรปสแดงคอตาแหนงซงฟงกชนเลเวลเซตมคาเทากบ
ศนย (ภาพจาก http://en.wikipedia.org/wiki/Level_set_ method)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
242
สมการเลเวลเซตถกพฒนาขนมาโดยใชพนฐานทาง
ทฤษฎของการหาผลเฉลยของสมการ Hamilton-Jacobi ซงมรปแบบทวไปของสมการ คอ
0),( =∇+∂∂ φφφ Ht
(2)
ซงทฤษฎนเปนทยอมรบอยางกวางขวางและมการพสจน
ความถกตองของผลเฉลยเชงตวเลขของสมการดงกลาว ดงนนเราจงสามารถประยกตใชกลวธในการหาผลเฉลยเชง
ตวเลขของสมการ Hamilton-Jacobi เขากบการหาผลเฉลยของสมการเลเวลเซตได
วธการเลเวลเซตไดถกประยกตใชในดานตางๆ จานวนมากเชน การบรณะภาพถาย การเตบโตของผลก เปนตน บทความนกลาวถงการประยกตใชสมการเลเวลเซตใน 4 ดาน คอ 1) การประยกตดานการจาลองปรากฏการณทางธรรมชาต 2) การประยกตทเกยวของกบภาพถาย 3) การประยกตในอตสาหกรรมภาพยนตร และ 4) การประยกตทเกยวกบภยทางธรรมชาต ซงการประยกตในแตละกลมมขนตอนทคลายคลงกน กลาวคอ เรมตนดวยการสรางสมการในรปแบบของสมการ (1) หรอ (2) โดยการเพมพจน (term) ลงไปในสมการ พจนทเพมเขาไปนจะมความหมายทางกายภาพและสวนใหญมฟงกชนเลเวลเซต
ปรากฏอย ในบางกรณเราอาจจะสรางสมการเลเวลเซตมากกวาหนงสมการเพออธบายเหตการณตางๆ พรอมกน ขนตอนตอมาคอ หาผลเฉลยเชงตวเลขของสมการทสรางขนมานโดยใชวธการหาผลเฉลยของสมการ Hamilton-Jacobi และขนตอนสดทายคอการสรางภาพจาลองหรอแสดงผลลพธโดยพจารณาเฉพาะตาแหนงทฟงกชนเลเวล
เซตมคาเทากบศนย ในบทความน จะแสดงเฉพาะสมการเลเวลเซตท เกยวของกบการประยกตใชในแตละกรณ จากนนจะแสดงผลลพธจากการคานวณ โดยไมกลาวถงการหาผลเฉลยของสมการ หรอกลาวถงคาของพารามเตอรตางๆ ทเกยวของกบสมการเลเวลเซตนนๆ
ขอมลเพมเตมเกยวกบวธการเลเวลเซต สามารถหาอานไดจากหนงสอ 2 เลมทนพนธโดย Osher และ
Fedkiw(5) และ Osher และ Sethian(9)สวนภาพเคลอนไหวท
สวยงามของการประยกตใชวธการเลเวลเซตในดานตางๆ สามารถดไดท http://physbam.stanford.edu/~fedkiw/
การประยกตใชสมการเลเวลเซตดานการจาลองปรากฏการณทางธรรมชาต การจาลองการกระจายตวของควน สมการทเกยวของมรปแบบของสมการ(1) คอ
( ) fpVVtV
+∇−∇⋅−=∂∂ (3)
เมอ p คอ แรงดนของแกส f คอ แรงทกระทาจากภายนอก และ V คอฟงกชนของ vu, และ w ในทน V คอฟงกชนเลเวลเซต ผลลพธจากการคานวณแสดงในรปท 2
รปท 2 ภาพตอเนองแสดงการจาลองการเคลอนทของควน โดยทควนถกพนออกมาจากทางซาย (ภาพมมบนซาย) และควนจะคอยๆ กระจายตวออกไปทางขวา (ภาพจาก1)
การจาลองการไหลของนา สมการทเกยวของมรป แบบของสมการ(1) คอ
0=∇⋅+∂∂ φφ Vt
(4)
เมอ ),,( wvuV = และ φ คอ ฟงกชนเลเวลเซตทถกใชเพอหารอยตอระหวางนากบอากาศ ผลลพธจากการคานวณแสดงในรปท 3
การจาลองการลกไหมของไฟ สมการทเกยวของมรปแบบของสมการ(1) คอ
( ) fpVVtV
+∇
−∇⋅−=∂∂
ρ (5)
รปท 3 ภาพจาลองการเทนาใสแกว (ภาพจาก1)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
243
รปท 4 ภาพจาลองการตดไฟของลกบอลเมอเคลอนผานเปลวไฟ (ภาพจาก1)
เมอ ),,( wvuV = p คอ แรงดนของแกส, ρ คอ ความหนาแนนของเชอเพลง และ f คอ แรงทกระทาจากภายนอก ผลลพธจากการคานวณแสดงในรปท 4
การประยกตใชสมการเลเวลเซตท เกยวของกบภาพถาย การหาขอบของวตถในภาพถาย เนองจากภาพของวตถทเราตองการหาขอบเขตจะมสแตกตางจากสของฉาก
หลงของภาพถายและสสองสทแตกตางกนจะมสนาม
เวกเตอรทมทศทางตรงขามกน ดงนน ตาแหนงทสนามเวกเตอรของสสองสชนกนจะมเวกเตอรทมคาเทากบศนย
และเนองจากวธการเลเวลเซตจะใหความสาคญเฉพาะคา
ของฟงก ชนเลเวลเซตทมคาเทากบศนย ดงนนเราจงสามารถประยกตใชวธการเลเวลเซตในการหารอยตอหรอ
ขอบเขตของวตถในภาพถายได
สมการเลเวลเซตทใชในการหาขอบเขตของวตถท
อยในภาพถายมรปแบบสมการ(4) คอ
( ) φβφφαφφ
∇⎥⎥⎦
⎤
⎢⎢⎣
⎡⎟⎠⎞
⎜⎝⎛ −
−++⎟⎟⎠
⎞⎜⎜⎝
⎛
∇∇
+Δ∇∇=∂∂
2)(),(, 21
21ccICcyxgdivIIsign
t (6)
และตวอยางการหาขอบเขตของวตถในภาพถายแสดงใน
รปท 5
รปท 5 ภาพของตวเลขทไมชดเจน (ภาพซาย) และขอบเขตของตว เลขของภาพทางซาย หลงจากใชวธการเลเวลเซตชวยในการหาขอบเขต (ภาพขวา) (ภาพจาก4)
รปท 6 ภาพถายตนฉบบทความชดเจนสง (ภาพหลงสด ) ภาพทถกรบกวน (noised image) ( ภาพกลาง) และภาพทไดรบการปรบความชดเจนโดยวธการเลเวลเซต (ภาพหนาสด) จะเหนวาภาพดานหนาสดมความชดเจนใกลเคยงกบภาพถาย
ตนฉบบ (ภาพจาก8)
การปรบความชดเจนของภาพ บอยครงทพบวาภาพถายจากดาวเทยมมความชดเจนคอนขางตาเนองจาก
ภาพถายไดถกรบกวนในระหวางทสญญาณจากดาวเทยม
สงภาพมายงสถานรบสญญาณดาวเทยม ดวยเหตน เราจาเปนตองปรบความชดเจนของภาพถาย สมการเลเวลเซตทใชในการปรบความชดเจนของภาพมรปแบบของ สมการ(8) คอ
20
3
20
2222 uu
uu
uu
uyuu
uxt
u
yx
y
yx
x μλ −−⎟⎟
⎠
⎞
⎜⎜
⎝
⎛
+∂∂
+⎟⎟
⎠
⎞
⎜⎜
⎝
⎛
+∂∂
=∂∂ (7)
และตวอยางของภาพถายทไดถกปรบความชดเจนแสดงใน
รปท 6

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
244
การประยกตใชสมการเลเวลเซตในอตสาหกรรมภาพยนตร ปจจบนเรขภาพคอมพวเตอร (computer graphics)เขามามบทบาทอยางมากในอตสาหกรรมภาพยนตร วธการเลเวลเซตมสวนชวยในการสรางรปทรงสามมตท
คอนขางซบซอน โดยเรมดวยการสรางจดทอยบนผวของรปทรงสามมต (รปท 7) จากนนใชวธการเลเวลเซตสรางผวหมจดเหลานน สมการเลเวลเซตในกรณนมรปแบบของสมการ(10) ดงน
φφφ
φφφ
∇⎥⎦
⎤⎢⎣
⎡
∇∇
⋅∇+∇∇
⋅∇=∂∂ ddt
(8)
เมอ ),()( Sxdistxd rr= คอ ฟงกชนระยะทาง และ S
คอ เซตของจดทเราสรางในตอนเรมตน
ตวอยางภาพยนตรทประยกตใชวธการเลเวลเซต
โดยใชกลวธขางตน คอ ภาพยนตรเรอง Pirates of the Caribbean: Dead Man’s Chest ซง Ronald Fedkiw (ผ พฒนาวธการเลเวลเซต) เปนผสรางเรขภาพคอมพวเตอร และภาพยนตรเรองนไดรบรางวลออสคารสาขาผลพเศษ
ทางภาพ (visual effects) ในป ค.ศ. 2007 อกดวย ในภาพยนตรเรองน ตวละครทชอ Davy Jones (แสดงโดย Bill Nighy) ไดถกสาปใหมหนาตาคลายหมก ในการสรางภาพใบหนาของ Davy Jones นน จดหลายจดจะถกเขยนบนใบหนาของนกแสดง จากนนภาพสามมตของหนวดหมกจะถกสรางโดยใชวธการเลเวลเซตทกลาวขางตน
อกตวอยางหนงของภาพยนตรทใชวธการเลเวล
เซตชวยจาลองปรากฏการณทางธรรมชาต คอ ภาพยนตรเรอง Poseidon มการจาลองการทะลกของนาทะเลเขามาในเรอเดนสมทร และภาพยนตรเรองนยงไดถกเสนอชอเพอชงรางวลออสคารสาขาผลพเศษทางภาพในป ค.ศ. 2007 เชนกน
นอกจากนยงมภาพยนตรเรองอนๆ ทใชวธการเล เวลเซตในการสร างผลพ เศษทางภาพ อาท เชน ภาพยนตรเรอง Terminator 3: Rise of the Machines เรอง Star Wars Episode III: Revenge of the Sith และเรอง Harry Potter and the Goblet of Fire เปนตน (ดไดท http:// en.wikipedia. org/wiki/Ronald_Fedkiw)
การประยกตใชสมการเลเวลเซตทเกยวกบภยทางธรรมชาต ไฟไหมปาเปนหนงในภยพบตทางธรรมชาต แบบ จาลองทางคณตศาสตรทใชคานวณหาตาแหนงของหนา
ไฟหรอรอยตอระหวางบร เวณท ถกไฟไหมแลวและ
บรเวณทยงไมไดถกไฟไหม มสมการซงอยในรปของ
สมการเลเวลเซต(3) ดงน
0|| =∇+∂∂ φφ Ft
(9)
โดยท F คอ ฟงกชนความเรวของการเคลอนทของหนาไฟและมรปแบบของสมการ(2) ดงน
⎪⎩
⎪⎨
⎧
>+
<+=
2||),cos()sin(
2||),sin()cos(
),(2 πθθεθβ
πθθβθθ
μ
μ
UU
UUvUF
f (10)
เมอ U คอ ความเรวของลม θ คอ มมระหวางทศทางตงฉากกบหนาสมผสและทศทางของลม (วดทวนเขมนา- ฬกา ) μ ≈ 2-4 คอ คาคงททใชกาหนดรปรางของไฟปา และ vf , β และ ε คอ ฟงกชนทกาหนดความเรวของไฟในตาแหนงดานหนา ดานขาง และดานหลง ตามลาดบ
การพยากรณหาตาแหนงของหนาไฟมอย 2 ขนตอน คอ เรมจากการหาตาแหนงของหนาไฟ ณ เวลาปจจบน และขนตอนตอไปคอการหาผลเฉลยของ สมการ (9) ในขนตอนแรก เปนการใชเทคนคของการหาขอบเขตของวตถในภาพถายดงทกลาวมาแลวขางตน เพอชวยในการหาขอบเขตของไฟปาจากภาพถายดาวเทยม (รปท 8) หลงจากนน นาคาของฟงกชนเลเวลเซตทไดจากขนตอนแรกมาเปนคาเรมตนสาหรบการคานวณหาผล
เฉลยเชงตวเลขในสมการ (9) (รปท 9)
สรป จงเหนไดวาสมการคณตศาสตรเพยงรปแบบเดยว คอ “สมการเลเวลเซต” สามารถนามาประยกตใหเกด
ประโยชนอยางมากในแตละสาขาวชาทแตกตางกน แตในวชาคณตศาสตร ยงมสมการอกหลายรปแบบทสามารถนามาสรางประโยชนในดานตางๆ ใหกบเรา ดวยเหตน
รปท 7 เซตของจดทอยบนพนผวของรปทรงสามมตทตองการ สราง (ภาพซาย ) และรปทรงสามมตทสรางขนมาโดยใชวธการเลเวลเซต (ภาพขวา) (ภาพจาก10)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
245
หากนกวจยหรอผปฏบตงานดานตางๆ มความรความเขาใจในวชาคณตศาสตร จะสามารถนาไปบรณาการใหเกดผลยงขนไปอกได
กตตกรรมประกาศ ผนพนธขอขอบคณ Springer Science & Business Media ทไดเออเฟอภาพประกอบรปท 2-7 ([email protected]; www.springer.com/rights)
เอกสารอางอง 1. Fedkiw R. Simulating Natural Phenomena. In:
Osher S, Paragios N, editors. Geometric level set methods in imaging, vision, and graphics. New York: Springer Verlag; 2003. p. 461-79.
2. Fendell FE, Wolff MF. Wind-aided fire spread. In: Johnson EA, Miyanishi K, editors. Forest fire: behavior and ecological effects. San Diego: Academic Press; 2001. p. 171-223.
3. Huabsomboon P. Implicit level set methods for a fire spread model. Proceedings of the 2nd International Conference on Asian Simulation and Modeling; 2007 Jan 9-11; Chiang Mai, Thailand.
4. Kimmel R. Fast edge integration. In: Osher S,
Paragios N, editors. Geometric level set methods in imaging, vision and graphics. New York: Springer Verlag; 2003. p. 59-77.
5. Osher S, Fedkiw R. Level set methods and dynamics implicit surfaces. New York: Springer Verlag; 2003.
6. Osher S, Paragios N. Geometric level set methods in imaging, vision and graphics. New York: Springer Verlag; 2003.
7. Osher S, Sethian JA. Fronts propagating with curvature dependent speed: algorithms based on Hamilton-Jacobi formulations. J Comp Physics 1988;79:12-49.
8. Rudin L, Lions PL, Osher S. Multiplicative denoising and deblurring: theory and algorithms. In: Osher S, Paragios N, editors. Geometric level set methods in imaging, vision, and graphics. New York: Springer Verlag; 2003. p. 103-19.
9. Sethian, JA. Level set methods and fast marching methods: evolving interfaces in computational geometry, fluid mechanics, computer vision, and materials science. Cambridge University Press; 1999.
10. Zhao H, Osher S. Visualization, analysis and shape reconstruction of sparse data. In: Osher S, Paragios N, editors. Geometric level set mehtods in imaging, vision, and graphics. New York: Springer Verlag; 2003. p. 361-80.
รปท 9 ภาพตวอยางแสดงขนตอนของการพยากรณหาตาแหนงของ หนาไฟ เสนสแดงแสดงถงตาแหนงของหนาไฟทพยากรณ
โดยใชวธการเลเวลเซต
รปท 8 ภาพตวอยางแสดงขนตอนของการหาตาแหนงของหนาไฟจากภาพถายดาวเทยม เสนโคงสนาเงนจะคอย ๆ หดตวเพอหาขอบเขตของบรเวณทถกไฟไหม (บรเวณทมสสม-แดง)


247
การควบคมการตายของเซลลสตวเพาะเลยงทใชในอตสาหกรรมเทคโนโลยชวภาพ
อษฎา มงสนตสข ภาควชาเทคโนโลยชวภาพ คณะวทยาศาสตร มหาวทยาลยมหดล
บทคดยอ วตถประสงคในการผลตโปรตนจากเซลลลกผสมทเพาะเลยงในถงชวปฏกรณสวนหนงมงเนนไปทการพฒนา
กระบวนการผลตเพอใหไดผลผลตโปรตนในปรมาณสงทสด แตอะพอพโทสสเปนกระบวนการตายทพบไดบอยในเซลลสตวเพาะเลยงทใชในอตสาหกรรม การตายของเซลลเพาะเลยงจากสาเหตนจงเปนอปสรรคทสาคญในการผลตโปรตนลกผสม ทาใหเกดการศกษาวจยในแงมมทนากลยทธตางๆ มาใชเพอเพมความอยรอดและลดอะพอพโทสสในเซลลเพาะเลยง
คาสาคญ: อะพอพโทสส; วฏจกรเซลล; โปรตนลกผสม; เซลลตดแตงพนธกรรม
Abstract
One of the aims for production of proteins from recombinant animal cells in bioreactors has been to develop processes to achieve the highest possible yield. However, apoptosis is the major cause of cell death in industrial animal cell lines and is a significant hindrance to large scale production of recombinant proteins. Many strategies have been devised to enhance cell viability and reduce apoptosis in cell cultures.
Keywords: Apoptosis; cell cycle; recombinant proteins; cell engineering
บทนา อตสาหกรรมทนาเทคโนโลยชวภาพมาประยกตใช
มการพฒนาและกาวหนาไปอยางรวดเรวเพอสนองตอ
ความตองการของตลาดในการผลตโปรตนชนดตางๆ โดยเฉพาะอยางยงโปรตนทใชในทางการแพทย ในปจจบน มการนาเทคโนโลยดเอนเอลกผสม (recombinant DNA technology) มาใชในการผลตโปรตนหลายชนด ซงผลผลตสวนใหญมาจากเซลลสตวเลยงลกดวยนม อยางไรกตาม ผลผลตโปรตนทไดนนถกจากดดวยการทเซลลเพาะเลยงม
กาหนดเขาสระยะการตาย ซงโดยมากมาจากการตายแบบ อะพอพโทสส (apoptosis) หรอการตายของเซลลแบบทมการโปรแกรมไวลวงหนาแลว (programmed cell death) การทเซลลเพาะเลยงมจานวนลดลงเนองจากอะพอพโทสส เปนผลใหปรมาณโปรตนทผลตไดมจานวนลดลงตามไป
ดวย นอกจากน ยงทาใหกระบวนการหาสภาวะทเหมาะสมสาหรบการแสดงออกของโปรตนในระดบอตสาหกรรมถก
จากด ดงนน จงมความจาเปนทจะตองหาวธการรกษาการเจรญเตบโตของเซลลไว เพอใหไดผลผลตโปรตนใน
ปรมาณสงๆ การควบคมอะพอพโทสสในเซลลเพาะเลยงจงมความสาคญเปนอยางมาก
อะพอพโทสสสามารถเกดไดในเซลลเพาะเลยงทใช
เปนโฮสตสาหรบการผลตโปรตนลกผสม (recombinant protein) เชน เซลล myeloma และ Chinese hamster ovary (CHO) เปนตน(14,26,28,30,40) ดงนน ความรทวากระบวนการตายของเซลลถกควบคมอยางไร จงมประโยชนอยางยงในอตสาหกรรมเทคโนโลยชวภาพเพอการพฒนาใหได
ผลผลตของเซลลในปรมาณสง
บทความนกลาวถงความสาคญของการใชเซลล
สตว เพาะเลยงในการผลตโปรตนหรอชวเภสชภณฑ คว ามส มพ น ธ ร ะหว า ง ว ฏ จ ก ร เ ซลล แ ล ะก า ร เ ก ด อะพอพโทสส รวมถงกลยทธการตดแตงพนธกรรมของเซลลซงเปนวศวกรรมดานเซลลในการปองกนหรอชะลอ
การเกดอะพอพโทสสในเซลลสตวเพาะเลยงทใชในระดบ
อตสาหกรรมเทคโนโลยชวภาพเพอเพมผลผลตโปรตนจาก
เซลล
การผลตโปรตนลกผสมทใชเปนเภสชภณฑ ปจจบนความตองการของโปรตนทใชในทางการ
แพทยมเพมมากขน นาไปสการเพมจานวนของโรงงานอตสาหกรรมเทคโนโลยชวภาพ มการนาเทคโนโลยดเอน-

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
248
เอลกผสมมาประยกตใชในชวงปลายทศวรรษท 1970 ซงไดมการตดตอยนในเซลลโฮสตทเปนจลชพ โดยเฉพาะใน แบคทเรย Escherichia coli เนองจากทาไดงาย และเซลลเจรญเตบโตไดรวดเรว โปรตนทมความแตกตางกน (heterologus protein) สามารถทจะแสดงออกไดโดยการใสยนลกผสมของสตวเลยงลกดวยนมในเซลลแบคทเรย ตงแตมการผลตอนซลนของมนษยใน E. coli ซงไดรบการยอมรบในป ค.ศ. 1982 การผลตยาชวภาพในรปแบบดง กลาวกมการดาเนนการมาอยางตอเนองและแพรหลาย ตลอดทศวรรษท 1980 มการนาเทคโนโลยดเอนเอลกผสมมาใชในการผลตโปรตนซงมความคลายกบโปรตนดงเดม เพอใชในทางการแพทยเปนจานวนมาก ระหวางทศวรรษท 1990 ไดมการใชเทคนคนเพอผลตโปรตนทลดความ สามารถในการกระตนภมคมกน มเวชภณฑหรอยาชวภาพมากกวา 165 ชนดทไดรบการยอมรบในประเทศสหรฐ- อเมรกาและทวปยโรป(44) ตวอยางของผลตภณฑเหลาน
ไดแก recombinant interferon (IFNs) interleukin (ILs), recombinant hormones, recombinant blood factors, haematopoietic growth factors, vaccines, mAb-based products, nucleic acid-based drugs, therapeutic enzymes และผลตภณฑอนๆ ตวอยางของโปรตนและโฮสตทใชในการผลตแสดงในตารางท 1 ในป ค.ศ. 2004 ผลตภณฑ
เหลานมคาการตลาดประมาณ 33 ลานเหรยญสหรฐ และคาดวาจะถง 70 ลานเหรยญสหรฐในปลายทศวรรษน โดยยาชวภาพทขายดทสดคอ erythropoietin (EPO) และ recombinant EPO ตางๆ ซงในป ค.ศ. 2005 มมลคาการขายมากถง 10.7 ลานเหรยญสหรฐ
การเพาะเลยงเซลลสตวเพอใชในการผลตโปรตน
ใหไดปรมาณตามความตองการของตลาดนนไดมการ
พฒนาในสวนของการขยายขนาดการเพาะเลยง การควบคมคณภาพ ความปลอดภยและราคา การผลตโปรตนจาเพาะใหไดปรมาณมากสามารถทาไดโดยการใชเทคนค ดเอนเอลกผสม โดยนายนทตองการหรอทสนใจมาตดแตงและวางไวภายใตการควบคมของ promoter และ enhancer เพอใหยนแสดงออกไดเพยงพอ และการใหไดมาซงโปรตนตามตองการนนจะตองเลอกโฮสตท เหมาะสมดวย ซงโปรตนลกผสมหลายชนดทไดรบการยอมรบกนโดยทวไป
แลว สามารถผลตไดในแบคทเรย เชน E. coli ในยสต เชน Saccharomyces cerevisiae และในเซลลสตวเลยงลกดวยนม เชน CHO, human embryonic kidney (HEK293), mouse myeloma derived (NS0), human retina derived (PerC6) และ baby hamster kidney (BHK) หรอในเซลลแมลง(1, 2, 7, 18, 47) เปนตน โดยทไดมการพจารณาปจจยตางๆ
ไดแก ความตองการใหโปรตนนนๆ มการดดแปลงหลง จากการแปลรหส (post-translational modification) โปรตนสามารถหลงออกมาจากเซลลได เสถยรภาพและการพบกลบไดของโปรตน รวมถงคาใชจายตางๆ อกดวย แม E. coli และเซลลจากสตวเลยงลกดวยนมถกนามาใชเปนโฮสตหลกในการผลตยาชวภาพ โดยโปรตน 9 ใน 31 ชนดทใชในการรกษาซงไดรบการยอมรบในป ค.ศ. 2003 ผลตใน E. coli และ 17 ชนดผลตในเซลลสตวเลยงลกดวยนม การผลตโปรตนทตองผานกระบวนการไกลโคสเลชนยงคง
ตองใชเซลลสตว หากตองการเพมผลผลตจาเปนตองมการพฒนา expression construct และตองมความเขาใจเกยวกบเมแทบอลซมและคณสมบตทางกายภาพของเซลลสตวดวย ในปจจบน สามารถผลตโปรตนลกผสมไดถง 5 กรมตอลตร ซงเปนปรมาณทมากกวาในสมยกอนถงสบเทา งาน วจยทกาลงดาเนนการในปจจบนสวนหนงจงมงเนนไปยง
การปรบปรงอาหารเลยงเซลลและการตดตอยนเพอปองกน
หรอยบยงการตายของเซลลโดยอะพอพโทสส สามารถยดอายการผลตโปรตน และการลดไกลโคสเลชนทตางแบบกน(3)
การควบคมการเจรญเตบโตและการแบงตวของเซลล
วฏจกรของเซลลแบง เปนระยะอน เตอร เฟส (interphase) คอการเตรยมตวใหพรอมทจะแบงเซลล และ ระยะการแบงตวของเซลล (M) โดยระยะอนเตอรเฟสน
สามารถแบงออกเปน 3 ระยะยอยๆ คอระยะกอนสราง ดเอนเอ (G1) ซงเซลลมการเจรญเตบโตเตมทและมการ
สรางสารบางอยางเพอใชในการสรางดเอนเอ ตอมาเปนระยะสรางดเอนเอ (S) ซงเปนชวงทถกควบคมโดยปจจยหลายอยาง ไดแก origin recognition complex, mini-chromosome maintenance proteins และ cyclin dependent kinases (Cdks) โดยปจจยเหลานชวยปองกนไมใหดเอนเอเพมจานวนขนใหมหลงจากทมการถอดแบบดเอนเอไปแลว และระยะหลงสรางดเอนเอ (G2) ซงเปนระยะทเซลลมการเจรญเตบโตและเตรยมความพรอมเพอจะแบงโครโมโซม
และไซโทพลาซม เซลลจะใชเวลามากทสดในระยะ G1 เพอเพมมวลของเซลลหลงจากทมการแบงเซลลไปแลว การเพมมวลของเซลลเปนผลมาจากการสงเคราะหสารโมเลกล
ใหญ การเจรญเตบโตของเซลลจะถกกระตนดวยปจจยการเจรญเตบโต (growth factor) ตางๆ อยางไรกตาม หากเซลลไมไดรบปจจยการเจรญเตบโตทเพยงพอจะทาใหเซลลออก
จากระยะ G1 เขาสระยะ G0 ซงจะไมมการเจรญเตบโต

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
249
ตารางท 1 ตวอยางโปรตนทใชเปนยาชวภาพทไดรบการยอมรบในยโรปและสหรฐอเมรกา(44)
Product Examplesa Sources of production Therapeutic indication
Recombinant interferons and interleukins
PEGinterferon α-2a PEGylated rIFN- α-2a rhIFN-α-2b rh IFN-β-1a r IFN-β-1b r IL-1 receptor antagonist r IL-11 r IL-2
E. coli E. coli E. coli CHO cells E. coli E. coli E. coli E. coli
Hepatitis B, C, Cancer Chronic hepatitis C Hepatitis B, C, Cancer Relapsing/remitting multiple sclerosis Relapsing/remitting multiple sclerosis Rheumatoid arthritis Prevention of chemotherapy-induced thrombocytopenia Renal-cell carcinoma
Recombinant hormones
Insulin, rh insulin rh FSH rh luteinizing hormone rh TSH rh GH
E. coli, S. cerevisae CHO cells CHO cells CHO cells E. coli, S. cerevisae
Diabetes mellitus Infertility, Anovulation and superovulation Some forms of infertility Thyroid cancer Growth failure
Recombinant growth factor
r EPO, rh EPO r GM-CSF GM-CSF rh PDGF
CHO cells BHK cells NS0 myeloma cells E. coli Namawa cells Endothelial cells Fibroblast S. cerevisae
Anemia Chemotherapy-induced neutropenia Lower-extremity diabetic neuropathic ulcers
Recombinant vaccines
r HBsAg r OspA, a lipoprotein Influenza Smallpox Mumps
S. cerevisae E. coli Chick embryo CHO cells Rabbit kidney cells Human diploid cells
Immunization against hepatitis A and B Lyme disease vaccine Influenza Smallpox Mumps
Recombinant blood factors
rh Factor VIII rh Factor IX Anticoagulant rh tPA r tPA
CHO cells, BHK cells CHO cells S. cerevisae CHO cells E. coli
Hemophilia A Hemophilia B Prevention of venous thrombosis Acute myocardial infraction
Monoclonal antibody-based products
Humanized mAbs NS0 myeloma cells Hybridoma cells CHO cells EBV-infected B-cells (human)
e.g. Non-Hodgkins lymphoma, Carcinoma of the colon, Prevention of acute kidney transplant rejection
Other recombinant products
Dornase-α, rDNase rh hyaluronidase rh TNF- α r TNFR-IgG
CHO cells CHO cells E. coli CHO cells
Cystic fibrosis Adjuvant to increase absorption and dispersion of other drugs Adjunct to surgery for subsequent tumor removal Rheumatoid arthritis
aAbbreviations: DNase, Deoxyribonuclease; EPO, Erythropoietin; FSH, Follicle-stimulating hormone; GM-CSF, Granulocyte-macrophage colony-stimulating factor; HBsAg, Hepatitis B surface antigen; GH, growth hormone; IgG, Immunoglobulin type G; IFN, Interferon; IL, Interleukin; mAbs, Monoclonal antibodies; OspA, Outer-surface protein A; PDGF, Platelet-derived growth factor; PEG, polyethylene glycol; r, Recombinant; rh, Recombinant human; TNF, Tumor necrosis factor; TNFR, TNF receptor; tPA, Tissue plasminogen activator.
การเปลยนแปลงรปรางระหวางการเจรญเตบโต
ของเซลลเกดขนเมอ Cdks ไดรบการกระตนใหทางาน ซงจะไปเตมหมฟอสเฟต (phosphorylation) ใหกบโปรตนเปาหมายทตาแหนงเซอรนและทรโอนน ปกตแลว Cdks

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
250
จะทางานไดกตอเมออยรวมกบ cyclin ซงการรวมกนนจะเกดขนในวฏจกรของเซลลทตอบสนองตอสงกระตนททา
ใหเกดการแบงตว(29) แต Cdk-cyclin complexs จะไม
สามารถทางานไดเมอไปจบกบ p16, p21 และ p27 (23,32) เมอเซลลอยในสภาวะทไมเหมาะสม เชน อาหารหมด เซลลมความหนาแนนมาก มสารพษสะสม หรอไดรบสารเคม รงสยว เปนตน เซลลจะหยดการเจรญเตบโตและอยในระยะนง (stationary phase) ไมแบงตวและเปน
ชวงเวลาททาการประมวลผลเพอตดสนใจวาจะทาอะไร
ตอไปด เชน หากดเอนเอถกทาลายหรอโปรตนมการพบทผดปกตเนองจากไดรบสารเคมหรอรงสยว กจะทาการซอมแซม หากซอมแซมไมไดกอาจจะทาลายตวเองโดยกระบวนการอะพอพโทสส อยางไรกตาม หากเซลลไดรบอาหารและปจจยกระตนการแบงตวตางๆ อยางเพยงพอ หรอไดรบอาหารใหมทปราศจากสารพษ เซลลจะกลบเขาสวฏจกรการเจรญและแบงเซลลใหมไดอก มเชนนนเซลลจะเขาสชวงระยะตาย (death phase)
การตายแบบอะพอพโทสสในเซลลสตวเพาะเลยง อะพอพโทสส(19) เปนกระบวนการตายของเซลลท
ถกควบคมโดยยน มความสาคญตอการพฒนาของตวออน ทาใหเกดภาวะธารงดล (homeostasis) ในเนอเยอของ
อวยวะในตวเตมวย การกาจดเซลลและการแกของเซลล ลกษณะของเซลลท เกดอะพอพโทสส คอ เซลลมการเปลยนแปลงรปรางทจาเพาะ ไดแก การหดตว เยอหมเซลลเกด blebling มการแตกหกของชนดเอนเอ การหดตวของโครมาทน และตอมาเยอหมเซลลเกด ruffling ซงจะแตก
เปนอะพอพโทตกบอด (apoptotic bodies) นอกจากน ยงมการเปลยนแปลงของโมเลกลบนผวเซลลเพอใหเซลลกลน
กน (phagocyte) ทอยขางเคยงสามารถจดจาและมากลนกน กระบวนการนไมนาไปสการอกเสบหรอมการอกเสบเพยง
เลกนอย ระหวางการเกดอะพอพโทสสนน สวนประกอบทเปนฟอสโฟลพดทผวเยอหมเซลลชนใน ทเรยกวา ฟอสฟาทดลเซอรน (phosphatidylserine) จะยายไปสเยอหม
เซลลชนนอก ทาใหเซลลเปนเปาหมายของการถกกลนกนตอไปได นอกจากน เซลลทเกดอะพอพโทสสยงเกดการเปลยนแปลงทางชวเคมตางๆ ภายในเซลลอกดวย
อะพอพโทสสเปนการตายทพบไดมากในเซลล
สตวเพาะเลยงทใชในการผลตโปรตนในอตสาหกรรม ทาใหการผลตโปรตนลดลง ดงนนการตรวจอะพอพโทสสตงแตระยะแรกๆ ของการเพาะเลยงเซลลจะทาใหทราบการเปลยนแปลงทงภายนอกและภายในเซลลกอนทเซลล
ทงหมดจะตายไป และยงชวยลดคาใชจายในการเพาะเลยง
เซลลในระดบอตสาหกรรมลงอกดวย ความรความเขาใจในการควบคมการเกดอะพอพโทสสจงมความสาคญอยาง
ยงตอการเพาะเลยงเซลลเพอใชผลตโปรตนในระดบ
อตสาหกรรม
ในเซลลของสตวเลยงลกดวยนม การตายแบบ อะพอพโทสสจะเกดขนเปนขนตอน โดยเซลลจะรบสญญาณจากสงเราทงภายนอกหรอภายในเซลลขนอยกบ
ความสมดลยของสญญาณความอยรอดและความตาย ซงการตดสนใจนจะนาไปสการเกดอะพอพโทสส สญญาณจะถกสงผานโดยตวรบ (receptor) บนผวเซลลหรอโมเลกลนาสญญาณภายในเซลล และขอมลท ถกสงมาจะถกวเคราะหในขนควบคม (control stage) โดยผานวถตวรบความตาย (death receptor pathway) และ/หรอวถไมโท-คอนเดรย วถเหลานสงผลใหมการขยายสญญาณและเกดการทางานของเอนไซมคาสเพส (เปนเอนไซมชนดโปรต-เอสทตดโปรตนหลงกรดอะมโนแอสปารตค) ในชวงกระทาการ (execution phase) และในทสด สญญาณทสรางจากอะพอพโทตกบอดจะทาใหเซลลนถกกลนกน โดยเนอเยอบรเวณดงกลาวไมเกดการอกเสบแตอยางใด
วถการเกดอะพอพโทสสโดยอาศยตวรบและ ลแกนดบนผวเซลล (รปท 1) เชน Fas-FasL และ TNFR1-TNFα จะสงสญญาณกระตนใหกลมเอนไซมคาสเพสตงตน (caspase-2, -8 และ -10) ทางานได เอนไซมคาสเพสจะทางานเปนขนตอนตอเนอง (caspase cascade) โดย คาสเพสตงตนจะกระตนใหคาสเพสปฏบตการ (caspase-3, -6 และ -7) ทางานโดยไปยอยโปรตนทจาเพาะ และยงพบ วาคาสเพสตงตนสามารถกระตนใหเซลลเกดอะพอพโท-สสผานวถไมโทคอนเดรยไดอกดวย โดยคาสเพส -8 สามารถตดโปรตน Bid ในไซโทพลาซม ซงเปนโปรตนในกลม Bcl-2 ออกเปน 2 สวน ขนาด 11 และ 15 กโลดาลตน โปรตน Bid ทมขนาด 15 กโลดาลตนจะจบกบโปรตน Bax และยายไปอยท เยอชนนอกของไมโทคอนเดรย ทาให ไซโทโครมซ (cytochrome C) และโปรตนอนๆ ทเกยว ของกบการเกดอะพอพโทสส เชน apoptotic inducing factor เปนตน(34) รวออกมา ไซโทโครมซจะทาใหอะพอพ-โทโซม (cytochrome c/Apaf-1/ATP/dATP/ procaspase-9) ทางาน คาสเพส-9 จะกระตนคาสเพสปฏบตการใหทางานตอ นอกจากน การเปลยนแปลงทไมโทคอนเดรยยงสามารถควบคมไดโดยโปรตนชนดอนๆ ในกลมของ Bcl-2 ทปองกนการเกดอะพอพโทสส (เชน Bcl-2, Bcl-xL, Bcl-w, Bag-1, Bag-3, Mcl-1, A1 และ Boo) โดยโปรตน ในกลมนอาศย Bcl-2 homology (BH)-3 domain ไปจบกบ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
251
รปท 1 วถการเกดอะพอพโทสสในเซลลสตวเลยงลกดวยนม
โปรตนในกลมของ Bcl-2 ททาใหเกดอะพอพโทสส (เชน Bax, Bak, Bid, Bad, Bok, Bik, Bim, Bcl-xS, Noxa, Hrk, NBIP3 และ NBIP3L)
ในป ค.ศ. 1996 Shimizu และคณะ(36) รายงานวา
การแสดงออกทมากเกนไปของยนในกลม Bcl-2 ทปองกน อะพอพโทสสนสามารถยบย ง เอนไซมคาสเพสและ อะพอพโทสสได และการเกดอะพอพโทสสยงขนกบปรมาณสมพนธของโปรตนในกลม Bcl-2 ททาใหเกด อะพอพโทสสและปองกนการเกดอะพอพโทสสอกดวย เชน การแสดงออกทมากเกนไปของ Bax ในเซลลทม Bcl-xL แสดงออกตา จะทาใหมการกระตนคาสเพสและการตายแบบอะพอพโทสส ในขณะทอะพอพโทสสไมเกดในเซลลทม Bcl-xL แสดงออกในระดบสง(37) นอกจากน การเกดอะพอพโทสสยงสามารถเกดผานวถร างแห เอนโดพลาซม (endoplasmic reticulum) จากการกระตนโดยคาสเพส-12 ไดอกดวย ซงวถนอาจเกดมาจากการมปรมาณของแคลเซยมไอออนทมากจนเกนไปหรอจากการ
กระตนของยนทแสดงออกเมอมการพบของโปรตนท
ผดปกตไป เปนตน
อะพอพโทสสและวฏจกรเซลล อะพอพโทสสและการเจรญเตบโตของเซลลเปน
กระบวนการพนฐานททาใหเซลลปกตสามารถดารงชวต
ตอไปได และเปนกระบวนการกมมนตทมการควบคมทซบซอน ซงตองอาศยปจจยททาใหเซลลเจรญเตบโตและอยรอดได กลไกทใชควบคมการเจรญเตบโตของเซลล
และอะพอพโทสสนนจะมสวนทเชอมโยงกน หากกลไกเหลานมความบกพรอง เซลลจะไมตายและอาจสามารถนาไปสการเกดเซลลมะเรงได หรออาจทาใหภมคมกนตว เองเกดผดปกต นอกจากนสญญาณทไมสามารถเขากนไดในสวนท เกยวกบการเจรญเตบโตและการเหนยวรง
ของวฏจกรเซลลจะทาใหเกดอะพอพโทสสได การเกด อะพอพโทสสสามารถเกดไดทกชวงของวฏจกรเซลล
ขนกบวาอยชวงใดในขณะทเกดเหตการณ หรอหากเซลลไดรบสารพษ เชน เซลล human promyelocytic leukemia (HL-60) เมอไดรบสารกอมะเรง (DW2282) จะกระตนใหเกดอะพอพโทสสหลงจากทชวง G2/M ถกเหนยวรง(31 ) หรอเซลลชนดเดยวกนจะเกดอะพอพโทสสหากไดรบการ
กระตนดวยไซโคลเฮกซาไมดในชวง G0/G1 และ S(9) เปนตน
ในขณะทเซลลเพาะเลยงมชวตและเจรญเตบโต
จะตองอาศยปรมาณอาหารทเพยงพอและสภาพแวดลอมท
เหมาะสม เมอสงแวดลอมเปลยนแปลงไป เชน อาหารจากด การเปลยนของคาความเปนกรดดางและออสโมลา- รตของอาหารเลยงเซลล การเกดรแอคทฟออกซเจนสปชส เซลลจะหยดเจรญและเกดการกระตนอะพอพโทสสซง
นาไปสการตาย นอกจากน การตายแบบอะพอพโทสสยงสามารถถกกระตนจากสารเคมบางชนด เชน cisplatin, tunicamycin และ hydrogen peroxide เปนตน(21) การขาดสญญาณอยรอด (survival signals) เชน ฮอรโมนไซโทไคน (cytokine) อนทกรน (integrin) และปจจยททาใหเจรญ เตบโต และยงถกกระตนไดดวยจลชพ เชน เชอไวรสและ

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
252
แบคทเรย(6,12) หรอดวยผลผลตสดทายจากเมแทบอลซม
ของเซลล เชน กรดแลคตกและแอมโมเนย ซงเปนผลผลตจากการใชนาตาลกลโคสและกลตามนในอาหารเพาะเลยง
เซลล ตามลาดบ(21)
การปองกนการเกดอะพอพโทสสในเซลลสตวเพาะเลยง วตถประสงคขอหนงในการผลตโปรตนจากเซลล
สตวคอการพฒนากระบวนการผลตใหไดผลผลตปรมาณ
สงหรอมากทสด อยางไรกตาม อะพอพโทสสทเกดขนนนเปนอปสรรคทสาคญของการผลต เนองจากเซลลทใชใน ทางการคา เชน BHK, CHO, hybridoma และ NS0 นนงายตอการเกดอะพอพโทสส จงมการศกษาถงกลยทธในการลดหรอปองกนการเกดอะพอพโทสส ดงตารางท 2 ซง
คาดหวงเพอทจะเพมผลผลตโปรตนลกผสม การปองกนการเกดอะพอพโทสสอาจทาไดโดยการปรบเปลยน
สภาพแวดลอมของการเพาะเลยง เชน เลยงเซลลแบบมการเตมอาหารสมาเสมอ เปนตน หรอโดยการตดแตงพนธ- กรรมของเซลลเพอใหมความตานทานหรอชะลอการ
เกดอะพอพโทสส เชน การทาใหยน bcl-2 แสดงออกได
มากขนในเซลล myeloma และ CHO(33,41,42) หรอการทาให hybridoma มความตานทานตอการขาดสารอาหารและ
สารพษทเกดจากการเลยงในระบบหรอสภาวะทไมเหมาะ สมหรอรนแรง(40)
การศกษาในเซลล hybridoma พบวาเซลลทม Bcl-2 แสดงออกมาก จะม ช วตอย รอดและมผลผลตทมาก ขน(38) ในป ค.ศ. 1999 Mercille และ Massie(27) ไดศกษา
เซลล myeloma ทตดแตงพนธกรรมเพอให E1B-19K (adenovirus Bcl-2 homologue) มการแสดงออก เปนผลใหเซลลเจรญไดหนาแนนเพมมากขนถงสองเทา และสามารถผลตโมโนโคลนลแอนตบอดเพมมากขนถงรอยละ 40 นอกจากน ยงมการใชเทคนคของ small interfering RNA เพอทาใหโปรตนสองชนดทชกนาใหเกดอะพอพโทสส คอ Bax และ Bak ไมทางาน(22) ทาใหสามารถขยายเวลาการอย
รอดของเซลลและเพมผลผลตของอนเตอรเฟยรอนแกมมา
ในเซลล CHO ไดรอยละ 35 การเพมหรอลดอตราการเจรญ เตบโตของเซลลเพอเพมผลผลตใหมากขนยงสามารถทาได
โดยการตดแตงยนทเกยวของกบวฏจกรเซลลเพยงอยาง
เดยว เชน c-myc, p21, interferon regulatory factor 1, p27 หรอตดแตงยนดงกลาวรวมกบยนทเกยวของกบอะพอพ-โทสส เชน ในเซลล CHO ทมการแสดงออกของยน c-myc และ bcl-2 ทมากนนจะทาใหเซลลเจรญเตบโตเพมขนและมความหนาแนนสงสด อกทงยงชวยลดอะพอพโทสสลงดวย(16) นอกจากน การตดแตงยนใหสามารถบอกถงการตายโดยการเรองแสงของเซลลยงสามารถชวยใหทราบ
สภาพของเซลลในชวงการเพาะเลยงไดกอนทเซลลจะตาย
ทงหมด เชน การตดตอยนทกระตนใหเกดการยบยงการเจรญเตบโตของเซลลและการทาลายดเอนเอ (GADD153) เขากบเวกเตอรทถอดรหสใหโปรตนเรองแสงฟลออเรส-เซนตในเซลล NS0 นน เมอ เซลลอย ในสภาวะท ไม เหมาะสม ยน GADD153 จะมการแสดงออกพรอมกบการเรองแสงของเซลล ทาใหสามารถทราบถงสถานการณของการเพาะเลยงไดอยางรวดเรว จงเหมาะกบการนาไปใชกบการเพาะเลยงเซลลขนาดใหญในถงชวปฏกรณ(21)
ตารางท 2 ตวอยางกลยทธในการปองกนหรอชะลอการเกดอะพอพโทสสในเซลลสตวเพาะเลยง
Events Cell types References 1. Manipulation of the culture environment
1.1 Supplement media with nutrients (fed-batch)
1.2 Use of anti-apoptosis chemicals, e.g.,
suramin, N-acetylcysteine and pancaspase zVAD-fmk
2. Manipulation (engineering) of the cell 2.1 Genetic modifications 2.1.1 Over-expression of e1B-19K, hsp70, bcl-2, bcl-xL, aven, XIAP, CrmA, htert, neo, etc. 2.1.2 RNAi to Bak, siRNA to Alg2 etc. 2.2 Use of cell-cycle arrest to avoid apoptosis e.g. expressing p27, IRF-1, p21, p53, c-myc and anti-sense c-jun
CHO, NS0 CHO, NS0 CHO, NS0, hybridoma, BHK, HEK293 CHO CHO, BHK, NS0
(21, 28) (4, 24, 25) (5, 8, 10, 16, 17, 20, 27, 35, 41, 42, 43) (22, 46) (11, 13, 15, 45)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
253
สรป การเกดอะพอพโทสสในเซลลเพาะเลยงสามารถ
ควบคมไดโดยการปรบเปลยนสภาวะแวดลอมในการ
เพาะเลยงเซลลใหเหมาะสมในระหวางการเพาะเลยง หรอการใช เซลลท ไดรบการตดแตงพนธกรรมเพอใหม
ความสามารถในการเจรญเตบโตไดยาวนานขน และชะลอการเกดอะพอพโทสสโดยการทาใหยนทปองกนการเกด อะพอพโทสสแสดงออกมากขน ถงแมการปองกนการเกดอะพอพโทสสจะชวยใหเซลลมการเจรญเตบโตมากขน การผลตโปรตนบางชนด เชน โมโนโคลนลแอนตบอดจะผลตไดดในชวงทเซลลอยในระยะนงเนองจากเซลลจะใช
พลงงานไปในการสรางโปรตนทตยภมมากกวาการแบง
เซลล ดงนน การศกษาเพอใหไดเซลลลกผสมทสามารถผลตโปรตนทตองการไดมากขนนนจาเปนตองมการ
คดเลอกเซลลทสามารถมชวตอยรอดไดนานและสามารถ
ใหผลผลตตอเซลลสงทสด นอกจากน การตดแตงยนทเกยวของในวถเมแทบอลซมของเซลลเพอใหเซลลใช
พลงงานเพมขนและลดความเปนพษของผลผลตจาก เมแทบอลซม ยงเปนอกกลยทธหนงทจะทาใหเซลลมชวตอยรอดไดนาน ลดการเกดอะพอพโทสสและเพมผลผลตโปรตนไดมากขน
เอกสารอางอง 1. Baldi L, Muller N, Picasso S, Jacquet R, Girard P,
Thanh HP et al. Transient gene expression in suspension HEK-293 cells: application to large-scale protein production. Biotechnol Prog 2005;21:148-53.
2. Barnes LM, Bentley CM, Dickson AJ. Charac-terization of the stability of recombinant protein production in the GS-NS0 expression system. Biotechnol Bioeng 2001;73:261-70.
3. Butler M. Animal cell cultures: recent achievements and perspectives in the production of biopharma- ceuticals. Appl Microbial Biotechnol 2005;68:283-91.
4. Chang KH, Kim KS, Kim JH. N-acetylcysteine increases the biosynthesis of recombinant EPO in apoptotic Chinese hamster ovary cells. Free Rad Res 1999;30:85-91.
5. Charbonneau JR, Furtak T, Lefebvre J, Gauthier ER. Bcl-xL expression interferes with the effects of L-glutamine supplementation on hybridoma cultures. Biotechnol Bioeng 2003;81:279-90.
6. Choi SS, Kim Y, Han KS, You S, Oh S, Kim SH. Effects of Lactobacillus strains on cancer cell proliferation and oxidative stress in vitro. Lett Appl Microbiol 2006;42:452-8.
7. Cockett MI, Bebbington CR, Yarranton GT. High level expression of tissue inhibitor of metallo-
proteinases in Chinese hamster ovary cells using glutamine synthetase gene amplification. Biotech-nology (NY) 1990;8:662-7.
8. Crea F, Sarti D, Faleiani F, Al-Rubeai M. Over-expression of hTERT in CHO KI results in decreased apoptosis and reduced serum dependency. J Biotechnol 2006;121:109-23.
9. Endresen PC, Prytz PS, Aarbakke J. A new flow cytometric method for discrimination of apoptotic cells and detection of their cell cycle specificity through staining of F-actin and DNA. Cytometry 1995;20:162-71.
10. Figueroa B Jr, Ailor E, Osborne D, Hardwick JM, Reff M, Betenbaugh MJ. Enhanced cell culture performance using inducible anti-apoptotic genes E1B-19K and Aven in the production of a monoclonal antibody with Chinese hamster ovary cells. Biotechnol Bioeng 2007;97:877-92.
11. Fussenegger M, Schlatter S, Datwyler D, Mazur X, Bailey JE. Controlled proliferation by multigene metabolic engineering enhances the productivity of Chinese hamster ovary cells. Nat Biotechnol 1998; 16:468-72.
12. Geiser V, Rose S, Jones C. Bovine herpesvirus type 1 induces cell death by a cell-type-dependent fashion. Microb Pathog 2008;44:459-66.
13. Geserick C, Bonarius HP, Kongerslev L, Hauser H, Mueller PP. Enhanced productivity during controlled proliferation of BHK cells in continuously perfused bioreactors. Biotechnol Bioeng 2000;69:266-74.
14. Hwang SO, Lee GM. Nutrient deprivation induces autophagy as well as apoptosis in Chinese hamster ovary cell culture. Biotechnol Bioeng 2008;99:678-85.
15. Ifandi V, Al-Rubeai M. Stable transfection of CHO cells with the c-myc gene results in increased proliferation rates, reduces serum dependency, and induces anchorage independence. Cytotechnology 2003;41:1-10.
16. Ifandi V, Al-Rubeai M. Regulation of cell proli-feration and apoptosis in CHO-K1 cells by the coexpression of c-myc and bcl-2. Biotechnol Prog 2005;21:671-7.
17. Ishaque A, Al-Rubeai M. Role of vitamins in determining apoptosis and extent of suppression by bcl-2 during hybridoma cell culture. Apoptosis 2002; 7:231-9.
18. Jones D, Kroos N, Anema R, Van Montfort B, Vooys A, van der Kratts S et al. High-level expression of recombinant IgG in the human cell line PER.C6. Biotechnol Prog 2003;19:163-8.
19. Kerr JFR, Wyllie AH, Currie AR. Apoptosis: a basic biological phenomenon with wide-ranging impli-cations in tissue kinetics. Br J Cancer 1972;26:239-57.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
254
20. Lasunskaia EB, Fridlianskaia II, Darieva ZA, da Silva MS, Kanashiro MM, Margulis BA. Transfection of NS0 myeloma fusion partner cells with hsp70 gene results in higher hybridoma yield by improving cellular resistance to apoptosis. Biotechnol Bioeng 2003;81:496-504.
21. Lengwehasatit I, Dickson AJ. Analysis of the role of GADD153 in the control of apoptosis in NS0 myeloma cells. Biotechnol Bioeng 2002;80:719-30.
22. Lim SF, Chuan KH, Liu S, Loh SO, Chung BY, Ong CC et al. RNAi suppression of Bax and Bak enhances viability in fed-batch cultures of CHO cells. Metab Eng 2006;8:509-22.
23. Mani S, Wang C, Wu K, Francis R, Pestell R. Cyclin-dependent kinase inhibitors: novel anticancer agents. Expert Opin Investig. Drugs 2000;9:1849-70.
24. Mastrangelo AJ, Zou S, Hardwick JM, Betenbaugh MJ. Antiapoptosis chemicals prolong productive lifetimes of mammalian cells upon Sindbis virus vector infection. Biotechnol Bioeng. 1999;65:298-305.
25. McKenna SL, Cotter TG. Inhibition of caspase activity delays apoptosis in a transfected NS/0 myeloma cell line. Biotechnol Bioeng 2000;67:165-76.
26. Mercille S, Massie B. Induction of apoptosis in nutrient-deprived cultures of hybridoma and mye-loma cells. Biotechnol Bioeng 1994;44:1140-54.
27. Mercille S, Massie B. Apoptosis-resistant E1B-19K-expressing NS/0 myeloma cells exhibit increased viability and chimeric antibody productivity under perfusion culture conditions. Biotechnol Bioeng 1999;63:529-43.
28. Murphy TC, Woods NR, Dickson AJ. Expression of the transcription factor GADD153 is an indicator of apoptosis for recombinant Chinese hamster ovary (CHO) cells. Biotechnol Bioeng 2001;75:621-9.
29. Murray A, Hunt T. The cell cycle: an introduction. New York: Freeman; 1993.
30. O'Connor KC, Muhitch JW, Lacks DJ, Al-Rubeai M. Modeling suppression of cell death by Bcl-2 over-expression in myeloma NS0 6A1 cells. Biotechnol Lett 2006;28:1919-24.
31. Piao W, Yoo J, Lee DK, Hwang HJ, Kim JH. Induction of G(2)/M phase arrest and apoptosis by a new synthetic anti-cancer agent, DW2282, in pro-myelocytic leukemia (HL-60) cells. Biochem Pharmacol 2001;62:1439-47.
32. Polyak K, Lee MH, Erdjument-Bromage H, Koff A, Roberts JM, Tempst P et al. Cloning of p27Kip1, a cyclin-dependent kinase inhibitor and a potential mediator of extracellular antimitogenic signals. Cell 1994;78:59-66.
33. Rabinder PS. Enhancement of survivability of mammalian cells by overexpression of the apoptosis-
suppressor gene bcl-2. Biotechnol Bioeng 1996; 52:166-75.
34. Roucou X, Rostovtseva T, Montessuit S, Martinou JC, Antonsson B. Bid induces cytochrome c-imper-meable Bax channels in liposomes. Biochem J. 2002;363:547-52.
35. Sauerwald TM, Oyler GA, Betenbaugh MJ. Study of caspase inhibitors for limiting death in mammalian cell culture. Biotechnol Bioeng 2003;81:329-40.
36. Simpson HS, Milner AE, Al-Rubeai M. Prevention of hybridoma cell death by bcl-2 during suboptimal culture conditions. Biotechnol Bioeng 1997;54:1-16.
37. Simpson NH, Singh RP, Perani A, Goldenzon C, Al-Rubeai M. In hybridoma cultures, deprivation of any single amino acid leads to apoptotic death, which is suppressed by the expression of the bcl-2 gene. Biotechnol Bioeng 1998;59:90-8.
38. Shimizu S, Eguchi Y, Kamiike W, Matsuda H, Tsujimoto Y. Bcl-2 expression prevents activation of the ICE protease cascade. Oncogene 1996;12:2251-7.
39. Shinoura N, Yoshida Y, Asai A, Kirino T, Hamada H. Relative level of expression of Bax and Bcl-XL determines the cellular fate of apoptosis/necrosis induced by the overexpression of Bax. Oncogene 1999;18:5703-13.
40. Tey BT, Al-Rubeai M. Suppression of apoptosis in perfusion culture of Myeloma NS0 cells enhances cell growth but reduces antibody productivity. Apoptosis 2004;9:843-52.
41. Tey BT, Singh RP, Piredda L, Piacentini M, Al-Rubeai M. Influence of bcl-2 on cell death during the cultivation of a Chinese hamster ovary cell line expressing a chimeric antibody. Biotechnol Bioeng 2000;68:31-43.
42. Tey BT, Singh RP, Piredda L, Piacentini M, Al-Rubeai M. Bcl-2 mediated suppression of apoptosis in myeloma NS0 cultures. J Biotechnol 2000; 79:147-59.
43. Veraitch FS, Al-Rubeai M. Enhanced growth in NS0 cells expressing aminoglycoside phosphotransferase is associated with changes in metabolism, pro-ductivity, and apoptosis. Biotechnol Bioeng 2005; 92:589-99.
44. Walsh G. Biopharmaceutical benchmarks 2006. Nat Biotechnol 2006;24:769-76.
45. Watanabe S, Shuttleworth J, Al-Rubeai M. Regu-lation of cell cycle and productivity in NS0 cells by the over-expression of p21CIP1. Biotechnol Bioeng 2002;77:1-7.
46. Wong DC, Wong KT, Nissom PM, Heng CK, Yap MG. Targeting early apoptotic genes in batch and fed-batch CHO cell cultures. Biotechnol Bioeng 2006;95:350-61.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
255
47. Wurm FM. Production of recombinant protein therapeutics in cultivated mammalian cells. Nat Biotechnol 2004;22:1393-8.


257
The Power of DNA Technology in Forensic Science
Patiya Phengon and Nathinee Panvisavas Forensic Science Graduate Programme, Multidisciplinary Unit, Faculty of Science, Mahidol University.
Abstract
DNA analysis is a powerful tool in forensic science. The technique provides information linking the suspect with the crime scene, identifies individuals, and enables kinship analysis. Moreover, DNA can be used for analysis of both human and non-human DNA specimens. In this article, applications of current DNA technology in forensic science and other promising technologies for the future are discussed.
Keywords: DNA technology; forensic science; human DNA; non-human DNA
Introduction
Biological evidence found at the crime scene can play a crucial role in forensic investigation. Deoxyribonucleic acid (DNA) from biological materials can be extracted and analyzed. The resulting DNA profiles can be utilized for identity testing to link or exclude an individual from involvement in a crime. DNA is found in nucleus, mitochondria and chloroplast. Both nuclear and mitochondrial DNA are found in human, animals, and plants, while chloroplast DNA is found only in plant. Due to advances in molecular biology techniques, some 628 genomes of living species have been charac-terized, 36 of these are eukaryotic genomes(29). This facilitates the study of DNA polymor- phisms, which are variations in the DNA sequence of the genome. The DNA polymor- phisms can be single base variations (single nucleotide polymorphisms or SNPs) or variable numbers of DNA repeats (satellite DNA). Because of these variations, they can be employed as DNA markers to differentiate one living organism from another(32). Hence, the analysis of DNA polymorphisms is an essential tool which has forensic dimensions, i.e. identification, individualization, and reconstruc-tion.
Application of DNA Technology in Forensic Science
Human DNA Analysis
In 1986, repetitive DNA markers were first used in forensic science to identify the perpetrator in a sexual assault case(19). Currently, autosomal DNA markers which have been used in human identification are “microsatellites” or “Short tandem repeats” (STR) markers, Gender marker (XY-homolo-gous amelogenin gene) and non-autosomal markers which are mitochondrial DNA and male-specific markers on Y-chromosome. Short Tandem Repeats (STRs) are length polymorphisms that are scattered throughout the human genome. They consist of 2-6 nucleotide units. The number of repeats varies between, and differs among individuals, except in identical twins(36). Forensic STR markers are usually 4 nucleotide repeats and their length varies between 100-400 bp. Because of their short length, STR is amenable to amplification using polymerase chain reaction (PCR).
PCR is an enzymatic reaction which target DNA can be rapidly amplified using optimal PCR components in a tube by using 3 different temperatures. PCR is a very sensitive technique, so that a small amount of starting DNA template can be analyzed. For example, trace amounts of skin cells on handled item or

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
258
latent fingerprints can provide sources of DNA(10,33). Moreover, DNA profiles can be obtained from a single cell containing less than 100 picogram of DNA, which can also be amplified by increasing the number of PCR cycles from 28 into 34 cycles. The technique is called Low Copy Number DNA analysis (LCN). Nonetheless, cross-contamination of other human DNA is possible in forensic analysis. Cellular components from other persons may be presented in crime scene samples. Laboratory technicians are also sources of DNA contamination. PCR inhibitors will also affect the enzymatic reaction. Heme compounds from blood and urea from urine, are potential inhibitors of Taq DNA polymerase. Therefore, it is necessary to perform controls including a negative control with no DNA and a positive control with DNA to make sure that PCR has no contaminants and inhibitors, respectively(8).
Multiplex-PCR enables multiple DNA markers to be analyzed in one reaction. The use of multi-color analysis, with fluorescent-based labeling of the amplified DNA product, thereby allows multiple STR loci to be simultaneously amplified in a tube. Such analyses may consist of 9-15 autosomal STR markers which will decrease the random match probability. Additionally, a gender marker, amelogenin gene, is added in commercial kits to indicate the gender of contributor. Amplified STR products would then be determined for a set of genotypes by capillary electrophoresis. At present, there are many commercial kits that are widely used for human identity testing.
A capillary electrophoresis instrument consists of narrow capillary where high separation voltage can be applied without overheating problems. Samples are consecu-tively injected, separated, and detected. DNA samples are automatically analyzed; so many samples can be loaded into the machine at the same time. A minute amount of DNA sample is then drawn into a capillary by electrokinetic injection. The run time of each sample is approximately 30 minutes, which is faster than conventional slab gel electrophoresis. How-
ever, the cost of instrument is still higher than that of gel-based electrophoresis(27).
A DNA profile, consisting of a combi-nation of genotypes of STR loci, from a crime scene sample must be compared with that from the reference sample. If the DNA profile generated from a crime scene sample and a reference sample match, the likelihood ratio would then be calculated to compare the probability of 2 competing hypotheses for DNA evidence; the prosecutor’s hypothesis and the defendant’s hypothesis. If likelihood ratio is greater than 1, it then supports the prosecutor’s hypothesis. On the other hand, if likelihood ratio is less than 1, this would support the defendant’s hypothesis.
DNA plays role as an investigative tool in forensic casework, human identity testing in mass disaster, and kinship testing(12). In 1983, a 15-year-old girl was found raped and murdered. A semen sample which was taken from the dead body presented type A blood group and an enzyme profile. However, the perpetrator had not been arrested at the time. Three years later, another 15-year-old girl was found sexually assaulted in the same town. Blood group from semen sample matched the previous case. It was believed that the perpetrator was the same man but he denied this. In 1985, DNA analysis was used to prove the case. The first suspect was exonerated by using DNA profiling. Then another suspect, Colin Pitchfork, was arrested because his DNA profile matched with the profiles from the semen of both murders. This was the first case in which DNA was used in criminal case.
DNA can not only identify an offender but also exonerate an innocent who is wrongly convicted. The INNOCENCE project has been established for re-investigating convicted crimi-nal cases by using DNA analysis. Thomas Webb, convicted as a sexual offender by hair evidence, was later shown to be wrongly convicted. Analysis of hair evidence collected from the crime scene was shown to be consistent with that from Mr. Webb. Then, in 1996, DNA testing revealed that Mr. Webb was innocent. He was exonerated after 13 years of

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
259
wrongful conviction. Now, at least 200 prisoners have been exonerated by DNA testing(20,37).
In mass disasters, dead bodies or human remains cannot be identified by positive identification methods. DNA analysis can be used as a method of choice for identification(39). Biological specimens from victims or human remains are sources of DNA for typing. Reference DNA profile can be typed from personal belongings, such as a toothbrush or inferred from kinship analysis (3). Two well known examples of human DNA identity testing are the terrorist attacks on the World Trade Center in 2001, and the devastating Tsunami hitting South East Asia in 2004. The high level of biological sample degradation challenges human remain identification. The remains were exposed to extreme conditions; light, tem-perature, humidity and microorganisms, which all affected the quality of DNA. The chance of obtaining a complete DNA profile would be reduced if the remains are highly degraded(14). Thus, miniSTR markers were developed to analyze highly degraded DNA. Large STR loci are re-designed by using primers to produce shorter STR fragments (miniSTRs)(2).
According to Mendelian inheritance, the biological father and mother pass on half of their genetic materials to the child. DNA can thus be used for parentage testing. In 1999, a medical student who disappeared was killed by her boyfriend. Bloodstains and decomposed muscle tissue were collected for DNA analysis. The DNA profiles matched DNA profile of victim’s putative parents. The result of pa-rentage analysis suggested that biological samples came from the victim(34). In some cases that few STR loci mismatch, if DNA mutation is not taken into account this could lead to false exclusion of paternity. Generally, at least 12 STR markers distributed on at least 10 different chromosomes would be examined in paternity cases. An unusual immigration case was reported in 2006. The alleged mother was excluded from maternity of a male child. This had been regarded as a deficiency case. It was then questioned if the 2 different STR loci was
an exclusion of the putative father or the existence of 2 mutations out of 22 autosomal STR loci. This case was then clarified by further analysis of Y-STR markers(21).
Mitochondrial DNA and Y-chromosomal DNA are also used as lineage markers for kinship analysis. Because the mother passes on mitochondrial DNA to both son and daughter, the children would then possess the same mitochondrial DNA as their mother and maternal relatives. Sequence-specific oligo-nucleotide typing and minisequencing are employed to examine the polymorphic site across the hypervariable regions of mito-chondrial DNA. The result is reported in term of their variation from the Anderson reference sequence. The father only passes on Y chromo-some to his son. The son will thus have the same Y-STR markers as his father and paternal relatives. Because mitochondrial DNA and Y-STR are lineage markers, their discrimination powers are lower than those of autosomal STR markers. Nevertheless, mitochondrial DNA is still useful when analysis of autosomal STR markers fail. Typing of mitochondrial DNA is useful for highly degraded forensic samples. Because of the high copy number of mito-chondrial DNA in a cell and mitochondria are resistance to extreme environment conditions. Under such conditions, it is more likely to obtain mitochondrial DNA profile than nuclear STR profile. Mitochondrial DNA can be recovered from minute amount of samples such as teeth, hair shaft, bone fragments, latent fingerprints, human feces, and highly degraded samples.
Linkage testing was demonstrated by classic “Romanov family” case using mitochon-drial DNA analysis. In 1994, human bones were discovered and suspected to be of the Romanov family. The bones were then subjected to analysis of gender markers, autosomal STR markers and mitochondrial DNA. The mitochondrial DNA sequence of putative Tsar Nicolas matched that of living maternal relatives. The mitochondrial DNA of putative Tsarina Alexandra also matched that of 3 skeleton remains and His Royal Highness

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
260
Prince Philip who is a maternal descent relative(6,18).
Heteroplasmy is defined as an event that more than one type of mitochondrial DNA is detected within an individual. There are 3 types of heteroplasmy in an individual: a) an individual may show more than one mito-chondrial DNA type in a single tissue- heteroplasmy, b) an individual may have heteroplasmy in one tissue sample and homo-plasmy in another tissue, and c) an individual may exhibit more than one homoplasmic types in different tissue types. Hence, heteroplasmy and tissue types must be considered when questioned and known samples are compared.
Y-STRs markers play an important role in criminal cases and paternal cases. In the most serious crimes such as sexual offences, perpetrators are often male. Therefore, Y-STRs are used to differentiate male from mixed evidence collected from sexual offences. It is difficult to resolve a mixed DNA profile if the female’s DNA content is higher than the male’s. However, the chance of detecting the male’s DNA would be increased if Y-STRs are analyzed. In case of the perpetrators are azoospermic or vasectomized, no sperm will be found. Thus, Y-STRs can be used to detect male components from biological fluids(8). All males in the same paternal lineage have identical Y-STR haplotypes. Because there is no recombination on the male-specific region, the biological father can pass genetic information in this region unchanged to his son unless mutations occurred(4). Y-chromosome analysis has been applied to help identify Saddam Hussein. In this case, a man had been captured in the village of Ad Dawr, Iraq in December 2003. Autosomal STR and Y-STR were ana-lyzed and compared to the profile of his son. The results showed that Y-STR profile of the captured man matched those of Saddam’s sons as well as sharing autosomal STR markers(5,8).
Non-human DNA Analysis
DNA analysis is also applied for identifying animals and plants in forensic casework. Forensic non-human DNA analysis
was applied not only for species identification but also for linkage analysis. The most common used DNA markers for animal species testing is cytochrome B (cytB) gene(26). Other DNA markers reported are such as 12S, 16S rDNA, cytochrome C oxidase subunit I (COI). For plants, trnL-trnF and rbcL regions on the chloroplast DNA, ITS gene on nuclear DNA are common DNA markers. In addition, short tandem repeat markers are also used for defining individuals. Forensic DNA analysis for non-human identification has also been applied to wildlife crimes, narcotic plants, domestic animal cases, food authentication, and bioterrorism(1,15,38).
Trade of endangered species is illegal. The Convention on the International Trade in Endangered Species of Flora and Fauna (CITES) is an agreement among governments listing 28,000 plant species and 5,000 animal species as protected species. For instance, Asian and African elephants are listed in Appendix I of CITES. Elephants are poached for ivory. Identification of protected elephant species from ivory has been developed by using the cytB gene. Lee et al. reported ivory identification by DNA profiling of the cytB gene (24). Moreover, cytB and microsatellites have been developed for identifying tiger species and indivi-duals(25,40). Tigers are endangered species and listed in Appendix I of CITES and their organs are illegally used as an ingredient in traditional Chinese medicine.
Cannabis sativa L. is classified into two types, “drug type” or “marijuana” and “fiber type” or “hemp”. The cultivation of drug type C. sativa L. is illegal because it contains a high level of narcotic chemical substance, δ-9-tetrahydrocannabinol (THC). Various C. sativa L. DNA markers have been studied for example the intergenic spacer sequence, the ITS region as well as short tandem repeats(16,17). Moreover, DNA markers from THC synthase gene has also been developed for marijuana identification in order to differentiate the drug type from fiber type(22).
Microbial DNA is also used as evidence to support forensic investigation. For example,

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
261
human immunodeficiency virus (HIV) DNA was used to prove a linkage between 2 persons. In 1998, an American physician injected the HIV into his girlfriend after taking a vial of blood from his HIV-infected patient. Results showed that the viral DNA from the victim and the patient matched almost exactly, though the HIV genome rapidly mutated(28).
Domestic animals may sometimes be involved in crime scenes, being as a witness, victim, or perpetrator. Both cytB and STR markers have been developed for domestic animal species testing and identity testing. An example of a dog attack, which dog saliva would be left as evidence, the dog DNA was analyzed by canine-STR typing. Identity of dog can be revealed though large amount of human blood was present(11,30).
DNA also plays role in food authentica-tion. It is necessary to verify food ingredients to prevent substitution in foodstuffs, fraud labeling and contamination. Analysis of pork DNA in halal food is an example of contaminant identification. Processed food and raw mate-rials are subjected to DNA analysis by using cytB gene markers to confirm species of origin(1).
New Technology in Forensic Science
Even though the available DNA markers and technology are still useful in forensic DNA testing, novel DNA markers and technology should be developed to overcome limitations as well as to provide fast, reliable and low cost analytical methods. Currently, single nucleotide polymorphisms (SNPs) markers have been developed for identity testing, lineage analysis, and phenotypic analysis(7,9). There are some 108 candidate SNPs that are currently reported for human identification, based upon at least 40 populations; however, validation by several laboratories and casework application must be tested in order to demonstrate robustness and reliability(31). To provide information for identifying people, informative SNPs for foren-sic identity testing should have high hetero-zygosity and low inbreeding coefficient (Fst) in all populations because there are only few SNPs
would that give high levels of discrimination power. At least 50 loci of SNPs must be used to give the same discrimination power as 10-13 STR markers. However, larger STR fragment, often fails to amplify from highly degraded DNA, resulting in an incomplete or no DNA profile. SNPs testing might be useful for highly degraded DNA sample. In addition, it is difficult to resolve DNA sample of mix origin by SNPs analysis. To date, there is no com- mercial kit available for identity testing(9). It is expected that core SNPs panel for identity testing will be soon launched into the forensic DNA community. Moreover, informative phe-notypic SNPs have been developed to associate physical traits, such as skin, eye color, etc of an individual. For example, SNPs on melanocortin 1 receptor gene and human pigmentation gene, were suggested for determination pigment variation of individuals(7).
Novel instruments are miniaturized into portable devices. This would allow DNA analysis to be performed in a mobile lab to immediately support the crime scene in-vestigation. Furthermore, instruments are also designed for high-throughput analysis. Exam-ples of emerging technologies are microchip capillary electrophoresis (microchip CE), STR hybridization array, microchip-based PCR, and mass spectrometry. The development of a microchip CE device has been ongoing since 1990, which allow reduced run time for DNA separation. It is expected that a microchip CE will be available for routine DNA analysis in the near future(13). STR hybridization array and microchip-based PCR device are miniaturized devices for DNA testing in the field. A microchip-based PCR device also reduces PCR reagents as well as the amount of precious forensic samples subjected to the analysis(23).
Matrix-assisted laser desorption ioniza-tion time-of-flight mass spectrometry (MALDI-TOF MS) is a high-throughput instrument so it has potential application in forensic science in terms of high-throughput analysis. Analysis time typically takes less than an hour for each sample. Unfortunately, the cost of MALDI-TOF MS system is still high, so it is still not

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
262
available for forensic STR-based DNA analysis. However, it may be available when SNPs markers are widely used for forensic purpose(35).
Conclusions
DNA markers and PCR technology are very powerful investigative tools. DNA has been used in forensic science to link the evidence to the suspect, and for identification of individuals and species. To date, both human and non-human DNA analysis has been reported to aid forensic investigation. DNA evidence not only links the suspect with the crime scene but also help exclude the innocent. It can be used to identify missing persons and victims in mass disasters when there is a lack of positive identification methods. In paternity testing, DNA of Putative parents can also be tested to see whether or not they are biological parents of the child. In addition, DNA provides informa-tion on kinship relation i.e. maternal lineage and paternal lineage of people who have different surnames. In non-humans, DNA plays im-portant role in species identification such as in wildlife crimes, narcotic plants, microbes, domestic animals, and food authentication and can also provide linkage of samples to an organism. However, there are some limits to DNA analysis of forensic samples. Highly degraded samples are difficult to type. Novel DNA makers such as autosomal SNP markers and phenotypic SNP markers will soon be introduced to forensic community and thus contribute information to forensic casework. Miniaturization of instrumentation was de-veloped to support on site analysis as well as high-throughput to perform analysis of large numbers of samples in the laboratory. Novel markers and emerging technology will enable “fast, reliable and low cost” DNA analysis in the future, thus, enhancing the power of DNA technology in forensic science.
References 1. Aida AA, Che Man YB, Wong CMVL, Raha
AR, Son R. Analysis of raw meats and fats of pigs using polymerase chain reaction for Halal authentication. Meat Sci 2005;69:47-52.
2. Andrade L, Bento AM, Serra A, Carvalho M, Gamero JJ, Oliveira C et al. AmpFlSTR®
MiniFiler(TM) PCR amplification kit: The new miniSTR multiplex kit. Forensic Sci Int Genet 2008;1 (Suppl) :89-91.
3. Bandhaya A, Panvisavas N. Optimization of DNA recovery from toothbrushes. Forensic Sci Int Genet 2008;1 (Suppl) :9-10.
4. Barbaro A, Cormaci P, Barbaro A. Complex family relationship reconstruction by Y-STRs test. Forensic Sci Int Genet 2008; 1 (Suppl) :494-5.
5. Bhattacharya S. Fast-track DNA tests confirm Saddam’s identity. [Online]. [2003 December 15] [cited 2008 Dec 14] Available from: URL: http://www.newscientist.com/article/dn4481-fasttrack-dna-tests-confirm-saddams-identity. html
6. Budowle B, Allard MW, Wilson MR, Chakra-borty R. Forensics and mitochondrial DNA: applications, debates, and foundations. Ann Rev Genomics Hum Genet 2003;4:119-41.
7. Budowle B, van Daal A. Forensically relevant SNP classes. Biotechniques. 2008;44:603-8,10.
8. Butler JM. Forensic DNA typing. 2nd ed. San Diego: Academic Press; 2005.
9. Butler JM, Budowle B, Gill P, Kidd KK, Phillips C, Schneider PM et al. Report on ISFG SNP Panel Discussion. Forensic Sci Int Genet (Suppl) 2008;1:471-2.
10. Djuric M, Varljen T, Stanojevic A, Stojkovic O. DNA typing from handled items. Forensic Sci Int Genet 2008;1 (Suppl) :411-2.
11. Eichmann C, Berger B, Reinhold M, Lutz M, Parson W. Canine-specific STR typing of saliva traces on dog bite wounds. Int J Legal Med 2004;118:337-42.
12. Goodwin W, Linacre A, Hadi S. An introduction to forensic genetics: Chichester: John Wiley; 2007.
13. Henry CS. Microchip capillary electrophoresis: an introduction. Method Mol Biol 2006;339:1-10.
14. Hoff-Olsen P, Jacobsen S, Mevag B, Olaisen B. Microsatellite stability in human post-mortem tissues. Forensic Sci Int 2001;119:273-8.
15. Hsieh H-M, Chiang H-L, Tsai L-C, Lai S-Y, Huang N-E, Linacre A et al. Cytochrome b gene for species identification of the conservation animals. Forensic Sci Int 2001;122:7-18.
16. Hsieh H-M, Hou R-J, Tsai L-C, Wei C-S, Liu S-W, Huang L-H et al. A highly polymorphic STR locus in Cannabis sativa. Forensic Sci Int 2003; 131:53-8.
17. Hsieh H-M, Liu C-L, Tsai L-C, Hou R-J, Liu K-L, Linacre A et al. Characterization of the

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
263
polymorphic repeat sequence within the rDNA IGS of Cannabis sativa. Forensic Sci Int 2005; 152:23-8.
18. Iwamura ES, Soares-Vieira JA, Muñoz DR. Human identification and analysis of DNA in bones. Rev Hosp Clin Fac Med S Paulo 2004; 59:383-8.
19. Jeffreys AJ. Genetic fingerprinting. Nature Med. 2005;11:1035-9.
20. Jobling MA, Gill P. Encoded evidence: DNA in forensic analysis. Nature Genet Rev 2004;5:739-51.
21. Junge A, Brinkmann B, Fimmers R, Madea B. Mutations or exclusion: an unusual case in paternity testing. Int J Legal Med 2006;120:360-3.
22. Kojoma M, Seki H, Yoshida S, Muranaka T. DNA polymorphisms in the tetrahydrocanna- binolic acid (THCA) synthase gene in "drug-type" and "fiber-type" Cannabis sativa L. Forensic Sci Int 2006;159:132-40.
23. Kricka L, Wilding P. Microchip PCR. Anal Bioanal Chem 2003;377:820-5.
24. Lee JC, Hsieh HM, Huang LH, Kuo YC, Wu JH, Chin SC et al. Ivory identification by DNA profiling of cytochrome b gene. Int J Legal Med 2009;123:117-21.
25. Linacre A, Tobe SS. On the trial of tigers-tracking tiger in traditional East Asian medicine. Forensic science international: Genetics 2008;1 (Suppl) :603-4.
26. Linacre A. The use of DNA from non-human sources. Forensic Sci Int Genet 2008;1 (Suppl) : 605-6.
27. McCord BR, Buel E, Jay AS. Analytical techniques: capillary electrophoresis in forensic biology. Encyclopedia of forensic sciences. Oxford: Elsevier; 2000. p. 127-35.
28. Metzker ML, Mindell DP, Liu XM, Ptak RG, Gibbs RA, Hillis DM. Molecular evidence of HIV-1 transmission in a criminal case. Proc Natl Acad Sci USA 2002;99:14292-7.
29. National Center for Biotechnology Information. Entrez Genome Project. [Online]. [cited 2008 Dec 14]. Available from: URL:http://www.ncbi. nih.gov
30. Pádár, Pádár Z, Egyed, Egyed B, Kontadakis, Kontadakis K et al. Canine STR analyses in forensic practice. Int J Legal Med 2002;116: 286-8.
31. Pakstis AJ, Speed WC, Kidd JR, Kidd KK. SNPs for individual identification. Forensic Sci Int Genet 2008;1 (Suppl) :479-81.
32. Passarge E. Color atlas of genetics. 2nd ed. New York: Thieme Stuttgart; 2001.
33. Phipps M, Petricevic S. The tendency of indivi-duals to transfer DNA to handled items. Forensic Sci Int 2007;168:162-8.
34. Rerkamnuaychoke B, Chantratita W, Jomsawat U, Thanakitgosate J, Pattanasak N, Rojanasunan P. Comparison of DNA extraction from blood stain and decomposed muscle in STR polymer-phism analysis. J Med Assoc Thailand 2000;83 Suppl 1:S82-8.
35. Sobrino B, BriÓn M, Carracedo A. SNPs in forensic genetics: a review on SNP typing methodologies. Forensic Sci Int 2005;154:181-94.
36. Tamaki K, Jeffreys AJ. Human tandem repeat sequences in forensic DNA typing. Legal Med 2005;7:244-50.
37. The Innocence Project-Know the cases. [Online]. [cited 2008 Dec 14 ]. Available from: URL:http://www.innocenceproject.org/Content/ 289.php
38. Wesselink M, Kuiper I. Species identification of botanical trace evidence using molecular markers. Forensic Sci Int Genet 2008;1 (Suppl) : 630-2.
39. Whitaker JP, Clayton TM, Urquhart AJ, Millican ES, Downes TJ, Kimpton CP et al. Short tandem repeat typing of bodies from a mass disaster: high success rate and characteristic amplification patterns in highly degraded samples. Biotech-niques 1995;18:670-7.
40. Xu YC, Li B, Li WS, Bai SY, Jin Y, Li XP et al. Individualization of tiger by using microsatel- lites. Forensic Sci Int 2005;151:45-51.


265
Gold Nanoparticles: Applications and Synthetic Methodology
Jirarut Wongkongkatep1, Pravit Wongkongkatep1, Woraphoj Lappenpuntrap1, Runchuan Ladadat1 and Pranee Phinyocheep2 1Department of Biotechnology 2Department of Chemistry, Faculty of Science, Mahidol University.
Abstract
This review introduces the useful applications of gold nanoparticles, particularly in biology. The synthesis method is focused on gold nanoparticles protected by small molecules and macromolecules. The classic and popular Frens method of gold nanoparticle synthesis is introduced in detail because it is a very useful method. Increase in the stability of gold nanoparticles by using Brust synthesis, and recent simplification of the Brust method, are then introduced. Finally, work on gold nanoparticles encapsulated with a polymer focusing on poly-N-isopropylacrylamide, is presented. The ability to carefully synthesize and tailor the physical or chemical properties of gold nanoparticles is essential for their applications in physics, chemistry, biology and medicine.
Keywords: gold; nanoparticle; nanomaterial; synthesis; polymer
Introduction
Nanoparticles have attracted great interest during the past decade due to their potential applications in biomedical, electronic and optical materials. Sizes, shapes, and compositions of metal nanoparticles can be systematically varied to produce materials with distinct light scattering properties. Nanomaterials are suitable, especially for bio-diagnostic assays because of their i) small size (1-100 nm) and correspondingly large surface-to-volume ratio, ii) chemically tailored physical properties, iii) unusual target binding properties, and iv) overall structural robust- ness (12). Only noble metals like gold are able to exist in the unoxidized state at the nanoscale and offer a unique surface chemistry that allows them to be used as platforms for self-assembling layers of organic molecules. Surprisingly, nanoparticles have been used in bioconjugation and as cellular labeling agents for the past four decades. However, new synthesis, fabrication, and characterization methods for gold nano- particles have evolved to the point that deliberate modulation of their size, shape, and composition is possible, therefore allowing exquisite control of their properties. The ability to carefully tailor
the physical properties of gold nanoparticles is essential for their application.
One example of the use of gold nano- particles is the detection of the missing base- pairs of oligonucleotides. Specifically, when 13-nm gold nanoparticles were modified by attaching oligonucleotides, sequence-specific particle assembly events induced by the target DNA occurred and the color of the solution changed from red to blue. The color change was due to the analyte-directed aggregation of gold nanoparticles, as a consequence of inter- acting particle surface plasmons and aggregate scattering properties. This phenomenon pointed out the use of gold nanoparticles as DNA detection agents in a type of “litmus test”(6). The modification with Raman active dyes reduced the detection limit down to 20 femtomolar (4).
In addition to the detection of nucleotides, Nam et al.(10) reported the ultrasensitive detection of proteins based on gold nano- particles. The system relies on microparticle probes with antibodies that specifically bind to a prostate-specific antigen, and nanoparticle probes that are encoded with DNA that is

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
266
unique to the protein target of interest and antibodies that can sandwich the target captured by the microparticle probes. This method can further reduce the sensitivity to 3 attomolar, six orders of magnitude better than for conventional methods that usually have sensitivity limits of about 3 picomolar.
Mixing polymers and nanoparticles can create surprising materials. By tailoring the nanoparticle coating and size, Balazs et al.(1) have created self-healing materials for improved sustainability and self-corralling rods for photovoltaic applications. Gold nanoparticles smaller than 8 nm in diameter are promising as low temperature catalysts for carbon monoxide oxidation. Using gold nanoparticles as catalysts can reduce air pollution because most pollution from automobiles is emitted in the first 5 min after start-up, but the conventional Pt- or Pd- based catalysts currently used in automobiles are inactive below 200oC. Therefore, gold nanoparticle-based catalysts present a potential solution to this cold-startup problem (3).
Therefore, the synthesis of gold nano- particles is particularly important because it opens the window for the use of gold nanoparticles in the areas of physics, chemistry, and biology. Methods for synthesis that enable us to control the properties of gold nanoparticles such as size, shape, dispersity and composition are introduced hereafter. Some of the useful methods as well as major advances focusing on methodology for the synthesis of gold nanoparticles are introduced. Because gold nanoparticles prepared by chemical reduction of the gold salts in a solution yield a small and narrow size distribution, this review will focus mainly on chemical reduction. A quantitative reduction method that is popular even now is presented, followed by the Brust method that greatly improves the stability of gold nano- particles. Recent advanced methodology for the synthesis of gold nanoparticles stabilized by small molecules and macromolecules focusing on poly-N-isopropylacrylamide is discussed.
Synthesis of Gold Nanoparticles Stabilized by Small Molecules
Among the several methods of synthesis of gold nanoparticles by reduction of gold deri- vatives, citrate reduction has been very popular and in use for a long time. Turkevitch(15) founded the method of synthesis of gold nanoparticles by citrate reduction in 1951.
Twenty-two years later, Frens(7) modified the Turkevitch method and introduced a promising procedure for the preparation of a set of monodisperse gold nanoparticles with widely different particle diameters. Frens reported that reduction of gold chloride with sodium citrate in aqueous solution could produce monodisperse gold nanoparticles with particle diameters varying from 16 nm to 150 nm. Two aqueous solutions are prepared: 0.01% HAuCl4 by weight (aqueous solution I) and 1% sodium citrate by weight (aqueous solution II). Then, 50 mL of solution I is heated to boiling before the addition of 0.5 mL of solution II. After approximately 2 min, the color of the solution changes to brilliant red, indicating the formation of monodisperse spherical gold nanoparticles. Reduction of gold chloride is practically completed after 5 min of boiling. By changing the amount of solution II, different diameter of gold nanoparticles can be obtained as demonstrated in Table 1. The Frens method is convenient and still being used even now when a rather loose shell of ligand is needed around the gold core in order to prepare a precursor to valuable gold nanoparticle-based materials. Figure 1 shows gold nanoparticle of a diameter 13 nm synthesized by Frens method.
While the gold nanoparticles produced by the Frens method are stable in water, they will aggregate at high concentrations and cannot be isolated in dried form. The Brust method, published in 1994, has had a considerable impact on the overall field because it allowed the facile synthesis of thermally stable and air-stable gold nanoparticles(2). Prepared by this method, gold nanoparticles can be repeatedly isolated and redissolved in common organic solvents without irreversible aggregation or decomposition. Brust’s strategy.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
267
Table 1 Relationship between amount of sodium citrate solution (1% by weight) and diameter of gold nanoparticles obtained after reaction with 50 mL gold chloride 0.01% by weight aqueous solution.
Amount of sodium citrate solution (mL) Particle diameter (nm) Color of particles
1.00
0.75
0.50
0.30
0.21
0.16
16
25
41
72
98
147
Orange
Red
Red
Dark red
Violet
Violet
consisted in growing the gold nanoparticles with the simultaneous attachment of thiol on the growing nuclei. AuCl4
- was transferred from aqueous solution to toluene using tetra- octylammonium bromide as the phase transfer reagent and reduced with aqueous sodium borohydride in the presence of dodecanethiol (C12H25SH). The preparation technique was as follows: An aqueous solution of hydrogen tetrachloroaurate (30 mL, 30 mM) was mixed with a solution of tetraoctylammonium bromide in toluene (80 mL, 50 mM). The two phase mixture was vigorously stirred until all the tetrachloroaurate was transferred into the organic layer and dodecanethiol (170 mg) was then added to the organic phase. A freshly prepared aqueous solution of sodium borohydride (25 mL, 0.4 M) was slowly added with vigorous stirring for 3 hours. The organic phase was separated and purified with ethanol and the brown precipitate was filtered off. Compared to the Frens method in which citrate acts both as the reducing agent and stabilizing ligand, Brust used sodium borohydride as a reducing agent and thiol as a ligand that strongly binds gold due to the soft character of both Au and S. This was the first time that gold nanoparticles stabilized by thiol, could be handled and characterized as a simple chemical compound. However, gold nano- particles stabilized by thiols are so stable that ligand exchange or further modification of the gold nanoparticles is comparatively difficult when compared with those using weak ligands such as citrate, carboxylate or amine.
Figure 1 Transmission electron microscope (TEM) image of gold nanoparticles synthesized by the Frens method.
Recent investigation was attemptd to simplify the synthesis method to a one-pot synthesis. Kim et al.(9) introduced a facile one- pot synthesis of gold nanoparticles using alcohol ionic liquids. The alcohol ionic liquids simulta- neously serve as both reductants and protective agents, so it significantly simplifies the process of nanoparticle preparation compared with Brust’s method. The concept is based on a simple idea because ionic liquids are literally ion compounds, and then the solvation and stabilization of gold ions by ionic liquids is superior. Alcohol is one of the best reducing agents. Therefore, gold nanoparticles can be easily obtained by simply mixing gold salt and alcohol ionic liquid in water without using a

26
วทยาศาสตรเพ
68
two-phasagents isin the smorphol[BF4]) (of gold nstraightfnanopartcondition[BF4]) isolution water unstirring astabilizePurificatunbound([HEMMthat reduserves aparticles
N-(2t
Figur
OtphosphinCappingamino aparticlesrenderedThis finlogically
SynthesStabiliz
Alachieve two diff
พอการพฒนา: รวมบท
se system, as not requiredstudy is N-linium tetrFigure 2). nanoparticlesforward andticles was ns. Then, 0in 10 mL o
of 0.02 mmnder nitrogenat 90oC for 2d gold nantion was pd ionic liquMor][BF4]) fuuces the Auas a stabiliz.
2-hydroxyethytetrafluorobor
re 2 Structur Kim et a
ther ligands ne, phosphin
g aqueous gacid lysine in solut
d them air-snding is a py relevant res
sis of Gold zed by Mac
lthough thergold nanop
ferent appro
ทความวชาการ ฉลอง
and the additd. The ioni-(2-hydroxyerafluoroborat
The one-phs in an aqueo
d purificatiocarried out 0.04 mmol of water wamol of HAuCn atmosphere24 hours, ([Hnoparticles performed buid with eunctions as au3+ ion to Azer in form
yl)-N-methylmrate ([HEMMo
re of ionic liqual.(9)
such as amine oxide can gold nanopa
stabilized ttion electrotable and wpromising stsearch (13).
Nanoparticromolecule
re are a varparticle-polymaches domin
งครบรอบ 50 ป แหง
tion of reduic liquid seleethyl)-N-mette ([HEMMhase preparaous solution
on of the gunder amb
of ([HEMMas added to Cl4 in 10 mLe. After vigoHEMMor][Bwere obtai
by removalethanol. Ha reducing aAu0, and it
ming gold n
morpholinium or][BF4])
uid used by
ine, carboxybe stabilizer
aticles with the gold nostatically
water-disperstep toward
icles es
riety of waymer composnate. The
การสถาปนาคณะวท
ucing ected thyl- Mor] ation was gold bient Mor]
the L of
orous BF4]) ined. l of Here, agent
also ano-
ylate, rs (5).
the ano- and
ible. bio-
ys to sites, first
one nanosecoaroublenpre-Thethanpolynanopartdispdepepoly
goldpon(PNtiontionlamof gpentreacsynt11-mmolby partPNIof Pis CPACPAand of Dremtionplacthe wasnoncentpolyFigu
goldtionnano
ยาศาสตร มหาวทยาล
consists ofoparticle inond one invound the gonding of pre-synthesized e advantage n small moleymers are soparticles fticle-polymerpersed, stablending on ymer.
Tenhu et d nanoparticsive polym
NIPAM) by n-fragmentatin. The polymide (NIPAMgold nanopatanoic acid ctions were thesis of gmercapto-1-ulecules wereesterificatio
ticles were uIPAM. ThePNIPAM on as follows.
ADB-gold nADB (0.0054
NIPAM (2.1DMF. Oxyg
moved by pen procedure thced in an oil polymerizat
s isolated by -grafted poltrifugation.ymer nanocoure 3.
Another d-polymer nan of ligand-oparticles.
ลยมหดล
f in situ syn the polymolves polymeold nanopare-made gold
polymer wof using m
ecules as stabsuperior for from coagur compositele, and isolthe propert
al.(11) reporcles grafteder (poly-N-surface-indu
ion chain-tmerization o
M) was initiaarticles mod
dithiobenzoin three s
old nanoparundecanol (e connected on; and 3) used in RAFe method fothe surface
. An amounanoparticles4 mmol), A1 mmol), wagen in the erforming a hree times. bath at 60oC
tion, the goldprecipitationlymer was r A TEM omposite m
approach foanocomposit-terminated PNIPAM n
ynthesis of mer matrix, erization of thrticles(5). H
nanoparticlwas also repomacromolecul
bilizing agenprotecting
ulation. Goles are usualable in drities of the
rted the synd with a th-isopropylacruced reversitransfer polof N-iso-pro
ated from thdified with oate (CPADsteps as folrticles stabi(MUD); 2) to the MUDCPADB-gol
FT polymerir the polymof gold nanount of 70 s, which cIBN (0.0004
as dissolved ireaction vefreeze-thaw Then the veC for 48 houd-polymer c
n in diethyl eremoved in image of t
material is s
or preparatite material ipolymers o
nanogels we
the gold and the
he matrix However, es into a orted(8,14). les rather nts is that the gold
ld nano- lly well- ed form,
e coating
nthesis of hermores- rylamide) ible-addi- lymeriza- opylacry- e surface 4-cyano-
DB). The llows: 1) ilized by
CPADB D ligands ld nano- zation of
merization oparticles
mg of contained 4 mmol), in 1.5 mL essel was w evacua-
essel was urs. After composite ether. Any
THF by the gold- shown in
ion of a s adsorp-
onto gold ere syn-

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
269
thesized by polymerization of a PNIPAM shell onto a pre-coated gold nanoparticle-PNIPAM seed. Etching of the gold core afterward with KCN results in hollow hydrogel nanoparticles (14). The product shows potential as a carrier for drug delivery and as a capsule for stabilizing the protein/enzyme. Citrate-stabilized gold nano- particles synthesized by the Frens method were used for the formation of polymerization seeds, due to the ease of their preparation and charac- terization. Then the gold nanoparticles were incubated with NH2-terminated PNIPAM for 10 min at room temperature. The solution was cleaned by centrifugation to remove any non- adsorbed NH2-terminated PNIPAM followed by resuspension in DI water. The polymer-adsorbed gold nanoparticles were then heated to 70oC under a nitrogen atmosphere and stirred in a glass vial pre-cleaned with aqua regia. An aqueous solution of NIPAM, the cross-linker N,N-methylenebis(acrylamide) (BIS), and 65 μL of 0.1 M aqueous solution of the surfactant sodium dodecyl sulfate was added to the reaction vial. After mixing for 1 hour, the reaction was initiated by injection of 130 μL of 0.1 M ammonium persulfate solution and the reaction was kept at 70oC for 4 hours. Nanogels obtained from this method can easily be tuned for particle size as well as the inner cavity by changing the thickness of the polymer shell or changing the size of the gold core. Therefore, the method is applicable for drug and gene delivery, for cellular imaging, as nanoreactors, as protective shells for enzymes, and as transfection vectors in gene therapy.
Conclusions
Several useful methods as well as recent advanced techniques for the synthesis of gold nanoparticles have been introduced. The classic Frens synthesis with citrate reduction is practical and still very much used to prepare precursors. The stabilization of gold nano- particles by thiols and macromolecules is also very impressive. However, new synthesis and fabrication methods for gold nanoparticles are still needed for modulation of the size, shape, and composition of the nanoparticles. The ability to carefully tailor the physical or chemical properties of gold nanoparticles is essential for
their applications in physics, chemistry, biology and medicine.
Figure 3 TEM image of gold nanoparticles coated by poly-N-isoproplyacrylamide.
Acknowledgements
Gold nanoparticle related research was supported by the Thailand Research Fund (TRF) and Commission on Higher Education, contract number MRG5080250. The authors dedicate this paper to Professor Aoyama Yasuhiro on the occasion of his retirement from Kyoto University in June 2009.
References 1. Balazs AC, Emrick T, Russell TP. Nanoparticle
polymer composites: where two small worlds meet. Science 2006;314:1107-10.
2. Brust M, Walker M, Bethell D, Schriffin DJ, Whyman RJ. Synthesis of thiol-derivatized gold nanoparticles in two-phase liquid-liquid system. J Chem Soc Chem Commun 1994; 801-2.
3. Campbell CT. The active site in nanoparticle gold catalysis. Science 2004;306:234-5.
4. Cao YC, Jin R, Mirkin CA. Nanoparticles with Raman spectroscopic fingerprints for DNA and RNA detection. Science 2002;297:1536-40.
5. Daniel MC, Astruc D. Gold Nanoparticles: assembly, supramolecular chemistry, quantum- size-related properties, and applications toward biology, catalysis and nanotechnology. Chem Rev 2004;104:293-346.
6. Elghanian R, Storhoff JJ, Mucic RC, Letsinger RL, Mirkin CA. Selective colorimetric detection of polynucleotides based on the distance- dependent optical properties of gold nanoparticles. Science 1997; 277:1078-81.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
270
7. Frens G. Controlled nucleation for the regulation of the particle size in monodisperse gold suspensions. Nat Phys Sci 1973;241:20-2.
8. Kang Y, Taton TA. Core/shell gold nanoparticles by self-assembly and crosslinking of micellar, block-copolymer shells. Angew Chem Int Ed 2005;44:409-12.
9. Kim KS, Choi S, Cha JH, Yeon SH, Lee H. Facile one-pot synthesis of gold nanoparticles using alcohol ionic liquids. J Mater Chem 2006;16: 1315-7.
10. Nam JM, Thaxton CS, Mirkin CA. Nanoparticle- based bio-bar codes for the ultrasensitive detection of proteins. Science 2003;301:1884-6.
11. Raula J, Shan J, Nuopponen M, Niskanen A, Jiang H, Kauppinen EI, Tenhu H. Synthesis of gold nanoparticles grafted with a thermo- responsive polymer by surface-induced reversible-addition-fragmentation chain-transfer polymerization. Langmuir 2003;19:3499-504.
12. Rosi NL, Mirkin CA. Nanostructures in bio- diagnostics. Chem Rev 2005;105:1547-62.
13. Selvakannan PR, Mandal S, Phadtare S, Pasricha R, Sastry M. Capping of gold nanoparticles by the amino acid lysine renders them water- dispersible. Langmuir 2003;19:3545-9.
14. Singh N, Lyon LA. Au nanoparticle templated synthesis of pNIPAm nanogels. Chem Mater 2007;19:719-26.
15. Turkevitch J, Stevenson PC, Hillier J. Nucleation and growth process in the synthesis of colloidal gold. Discuss Faraday Soc 1951;11:55-75.

271
Transparent Oxygen-Barrier Coating Based on Plasma Deposition of SiOx Thin Film on Polymeric Substrate
Supa Wirasate Department of Chemistry, Faculty of Science, Mahidol University.
Abstract
In order to prevent products from degradation due to oxidation process, O2-barrier property is required for packaging materials. Polymer-based packaging materials have long been developed for various applications including replacements of glass containers, which are impermeable to oxygen. In order to replace glass containers, improvement of the O2-barrier property of polymer is mandatory. Utilization of SiOx thin film to coat polymeric substrates is a promising application due to its transparency, high O2-barrier performance, recycling capability and tolerance to microwave radiation. O2-barrier performance of SiOx film depends strongly on such factors as chemical structure, surface morphology and defects, coating thickness and adhesion of deposited SiOx film. The purpose of this review is to provide background and an understanding of the parameters affecting O2-barrier property of SiOx film deposited by plasma-enhanced chemical vapor deposition techniques.
Keywords: oxygen-barrier; SiOx film; plasma deposition; plasma enhanced chemical vapor deposition; PECVD; packaging material
Introduction
Polymers have been used in many applica-tions, including adhesive bonding, protective coating, packaging, biomaterial, microelectronic device, and medical device. Usually, packaging materials are used to protect products from degradation, especially by oxidation process and to preserve product quality and hygiene. Therefore, gas barrier properties, especially oxygen (O2), are crucial for polymers used in this field. However, several polymers have high oxygen permeation while other properties are suitable for packaging applications. Examples of those polymers include polyethylene (PE), poly-propylene and poly(ethylene terephthalate) (PET). Their oxygen permeability is shown in Table 1.
Gas barrier properties of these polymeric films can be improved by various techniques, including utilization of novel microstucture to control polymer free volume, use of polymer composites containing barrier materials and laminating by a polymer film that has excellent gas barrier properties, or depositing a thin metal
film. Polymer films having excellent gas barrier properties, including poly(vinylidene chloride), poly(vinyl alcohol), ethylene-vinyl alcohol co-polymer and polyamide, have been applied by the laminating method (see Table 1). However, the thickness of these polymeric films used to laminate poorer gas barrier polymers is in the micron range and usually an adhesive layer is required to promote adhesion between the barrier layer and the polymeric substrate. In case of thin metal film deposition, such films provide excellent O2-barrier performance but it is not suitable for applications that require transparency and/or ability of the packaging to be microwaved.
It is known that bulk oxide materials including glass are impermeable to O2, H2O and a number of inert gases(1). Therefore, numerous attempts have been made to investigate the gas barrier properties of thin oxide films such as SiOx and AlyOx
(2-3,6-7,9-14) although these thin oxide films do not possess the same level of barrier properties as their bulk counterparts. SiOx film is of interest due to its similarity in chemical structure to silicate glass, its chemical

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองรบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
272
inertness, and the variety of commercially available monomers that can be used to produce the film. Various techniques have been used to deposit SiOx film, including sputtering, evaporation and plasma deposition (or sometimes called plasma polymerization or plasma enhanced chemical vapor deposition). Among these techniques, plasma deposition is most promising due to its low operational temperature, suitable for polymeric substrates. Furthermore, SiOx film produced by this method is flexible and has good adhesion to several polymers.
result from ionization, dissociation and excita-tion processes. Collisions in a plasma medium arise involving electrons accelerated by the electric field, which induce ions, electrons, metastables, neutral molecules, photons and free radicals in ground and excited states(5). Plasma constituents are associated with energy ranging from 0 to 40 eV while the bond energies of organic materials are in the range of 2 to 9 eV. Thus, the energies associated with a plasma or glow discharge are high enough to break chemical bonds during plasma processing. The difference between plasma polymerization and
Table 1 Oxygen permeability of some polymers used as packaging materials(15).
Polymer Oxygen permeability at 23oC, 50% or 0% relative humidity [cm3 mm/(m2⋅day⋅atm)]
Polyethylene (PE) 50-200
Polypropylene (PP) 50-100
Poly(ethylene terephthalate) (PET) 1-5
Polyamide (PA) 0.1-1
Poly(vinylidene chloride) (PVDC) 0.01-0.3
Poly(vinyl alcohol) (PVA) 0.02
Ethylene vinyl alcohol (EVOH) 0.001-0.01
O2-barrier performance of SiOx film is dependent on such parameters as chemical structure, surface morphology and defects, coating thickness and adhesion. Therefore, the background and an understanding of the effects of these parameters on O2-barrier property of SiOx films deposited by plasma deposition technique are reviewed in this article. Also, plasma technique is described briefly.
Plasma Techniques
Plasma is the fourth state of matter, which is an ionized gas. Since the late 1960s, plasma processing has been used primarily in micro-electronic industry for thin film deposition and for plasma etching of semi-conductors, metals and polymers.
The reactive species in a plasma process
plasma treatment is that plasma of a gas, which is not polymerizable (plasma treatment), will mainly interact with the polymeric substrate. On the other hand, plasmas of polymerizable gases mostly interact with monomer vapor(24). Film deposited by plasma processing usually contains branched and highly crosslinked structures.
Gas-Barrier Properties of SiOx Film
Gas permeation rate or gas transmission rate is defined as the volume of gas at standard temperature and pressure (STP) passing through a unit area of the membrane per second divided by the pressure difference across the membrane. For coated polymer membrane, gas transport can occur based on solubility-diffusion mechanism and gas flow through defects. In order to be

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
273
able to compare the effectiveness of coatings applied onto different polymers, barrier improvement factor (BIF) is utilized. BIF is equal to the permeation rate of uncoated polymer divided by the permeation rate of coated polymer(4).
Normally, oxygen transmission rate (OTR) of food packaging films should be in the order of 1 cm3/(m2⋅day⋅atm) at STP(4). As aforementioned, bulk silicate glass is impermeable to O2, due to the very tight interstitial spaces of Si-O lattice(1,17). It has been proposed that SiOx coating on polymer film acts as a defective block to oxygen transport, which results in an improvement of the O2-barrier property of a polymeric substrate(22).
Various factors affecting on oxygen-barrier properties of SiOx film produced from plasma deposition technique have been investigated. These factors include chemical composition, surface morphology, thickness, defect and adhesion of the deposited SiOx film.
Chemical Structure of Deposited SiOx Film
Based on the article of Lamendola and d’Agostino(16), high content of carbonaceous compounds in deposited SiOx film results in a low degree of cross-linking and thus high gas permeability. Numerous attempts have been performed to synthesize SiOx film with low carbon content and O to Si ratio close to that of silicate glass(6-8,10,13,21,23). Several approaches have been used to achieve this goal including selection of a monomer to produce SiOx film, addition of O2 gas to silane or siloxane monomer, and modification of plasma deposition conditions. Table 2 shows carbon contents, O/Si ratios of SiOx films deposited from various monomer/gasses and their oxygen transmission rates and BIF values. Several monomers, such as hexamethyldisiloxane (HMDSO), tetramethoxy silane (TMOS), trimethyl silane, and tetraethoxy silane (TEOS) have been used to produce SiOx films by plasma deposition technique. Inagaki et al.(12-14) employed TMOS instead of TEOS to prepare SiOx film as a gas barrier coating on
poly(ethylene terephthlate) (PET), polypropy-lene (PP) and poly(ethylene naphthalate) films. The selection of monomer was based on a lower C/Si atomic ratio of TMOS comparing to that of TEOS. By using X-ray photoelectron spectro-scopy (XPS) to determine the chemical compo-sitions of deposited SiOx films obtained from TEOS and TMSO, they found that C/Si atomic ratios of the SiOx films prepared form TMOS (C/Si ratio are 1.4-1.7 (depending on RF power used), which were lower than that prepared from TEOS (C/Si ratio was 3.2), suggesting that SiOx film obtained from TEOS contains a higher content of carbonaceous compounds than that obtained from TMOS(13). However, the O/Si atomic ratio of SiOx films deposited from both monomers are similar (1.7-1.9).
The second approach was to add O2 gas during plasma deposition of SiOx film since O2 gas plasma is known to create etching and oxygen incorporation effect to organic compounds. Thus, adding O2 (mixing with silane or siloxane monomer) during plasma deposition SiOx film would reduce carbona-ceous compounds in the final chemical structure of the SiOx film. Results obtained from several investigators showed that SiOx films obtained from silane or siloxane monomer alone contain high content of carbonaceous compounds as shown in Table 2(8,13). Inagaki et al.(13) found that C/Si atomic ratios of SiOx film deposited at a radio frequency (RF) power of 60 W from TMOS only and TMOS/O2 mixture is 1.5 and 0.39-0.49 (depending on O2 mol %) respect-tively. O/Si atomic ratios of SiOx film deposited at the RF power of 60 W from TMOS only and TMOS/O2 mixture are 1.8 and 1.4 (at all O2 mol %) respectively. Oxygen transmission rate of SiOx film deposited from TMOS alone is about 28-29 cm3/(m2⋅day⋅atm), while oxygen transmission rate of SiOx film deposited from TMOS/O2 mixture is 0.60 and 0.10 cm3/ (m2⋅day⋅atm) for 40 and 60 mol % O2 respec-tively. Teshima et al.(21) found that as the fraction of O2 increases during plasma deposition of TMOS, the content of carbona-ceous compound decreases. SiOx film contain-ing carbon-free and O/Si atomic ratio of 1.85

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองรบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
274
Table 2 Carbon content, O/Si ratio of SiOx film deposited from various gases and their oxygen transmission rate and BIF values.
Deposited gases Polymer/coating Characterization technique OTR cm3/(m2-atm-day)
BIF References
TMOS
TMOS/O2
TMOS/O2
PET/SiOx 100 nm XPS: C/Si atomic ratio = 1.5
O/Si atomic ratio = 1.8
XPS: C/Si atomic ratio = 0.42
O/Si atomic ratio = 1.4
XPS: C/Si atomic ratio = 0.49
O/Si atomic ratio = 1.4
27
0.6
0.1
2
73
440
(13)
(13)
(13)
TEOS
HMDSO
PE 40 μm/SiOx
PE 40 μm/SiOx
XPS: C/Si atomic ratio = 1.97, 2.04
O/Si atomic ratio = 1.55, 1.93
XPS: C/Si atomic ratio = 0.91-1.04
O/Si atomic ratio = 1.59-1.8
2
2
(8)
(8)
HMDSO/O2 PET 12 μm/SiOx XPS: C content = 41 atomic %
XPS: C content = 36 atomic %
XPS: C content = 8 atomic %
2.7
1.82
1.03
37
55
97
(10)
(10)
(10)
HMDSO/O2 PET 23 μm/SiOx 60 nm EDX*: C content = 49 %
O/Si = 1.0
EDX: C content = 18 %
O/Si = 1.5
EDX: C content = 0 %
O/Si = 1.7
70
70
1
1
1
70
(6)
(6)
(6)
* Energy dispersive X-ray analysis.
possesses an oxygen transmission rate as low as 1.3 cm3/(m2⋅day⋅atm). Walker et al.(23) also showed that without O2 during plasma deposi-tion of HMDSO the SiOx films produced contain many carbonaceous compounds as evidence in Fourier transform infrared spectrum of peaks appearing at 2,960, 2,900 and 1,260 cm-1, which correspond to C-H stretching of -CH3 group. Due to these carbonaceous compounds the deposited SiOx film shows no significant improvement in O2 barrier performance. At higher ratios of O2 to HMDSO used during plasma deposition, the SiOx film produced contains higher inorganic content and more crosslinks, which results in a reduction of diffusion coefficient of about 4 orders of magnitude. However, inhomogeneity and columnar growth with many micro-defects (pinholes) of the SiOx film are produced as
HMDSO:O2 ratio went to 1:40, which in turn lowers O2-barrier performance. Deshmukh and Shetty(8) modified polyethylene surface by depositing SiOx film from TEOS and HMDSO without adding O2 gas during plasma poly-merization process. Their results showed little improvement in barrier performance as compared to others. Their barrier improvement factor (BIF) is approximately 2 for SiOx films produced from HMDSO and TEOS on PE substrate.
Lastly, an adjustment of plasma process-ing has been used to produce carbon-free SiOx film. By using pulsed microwave plasma to deposit SiOx film from HMDSO/O2, Deilmann et al.(6-7) demonstrated a strong relationship between oxygen barrier property and chemical composition of SiOx film. They reported that SiOx film containing carbon-free and a small

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
275
amount of hydrogen (in silanol form) possesses a residual oxygen permeation of 1.0 ± 0.3 cm3/ (m2⋅day⋅atm). The best barrier film possesses an O to Si stoichiometry of 1.7 (measured by energy dispersive X-ray spectroscopy tech-nique). They reported BIF values of 65 and more than 1,000 for one-sided and two-sided coating respectively. Gruniger et al.(10) also reported an improvement of O2-barrier per-formance of PET substrate coated with SiOx film as the carbon content in the SiOx film is reduced from 41 atomic % to 8 atomic %, corresponding to BIF value of 37 and 97, respectively.
Surface Morphology and Defect of Deposited SiOx Film
It has been reported by several investi-gators that surface morphology and defect of deposited SiOx film play vital roles in O2-barrier performance(6,10,19,20). Generally, two types of defects are found in thin oxide film, namely, micro-defect and nano-defect. Micro-defect is composed of pinholes and micro-cracks, whereas nano-defect is caused by a non-equilibrium thermodynamic nature of the plasma (and/or vapor) process. Sobrinho et al.(19-20) successfully combined reactive ion etching in oxygen plasma and optical and/or electron microscopy techniques to detect micro- and sub-micro-defects on ultra-thin transparent SiOx layer on transparent polymeric substrate. They reported defect sizes ranging from 0.05 to 1.4 μm and demonstrated that the main sources of defects are silica micro-particles produced during plasma processing and dust particles. Erlat et al.(9) used atomic force microscopy to probe surface texture and morphology of deposited SiOx film and showed that good barrier properties are related to the presence of densely packed, small and spherical SiOx grains and a smooth surface of SiOx film. Once the barrier performance deteriorates it was noticed that the grains become irregular shaped and coalesce into larger features, which result in a rougher surface and more defect sites. In their work, intergranular defects were detected by
tunneling electron microscope. Gruniger et al.(10) reported that a dense and smooth SiOx film
is responsible for a high barrier improvement factor of 500 for PET substrate. Deilmann et al.(6) used a method described by Sobrinho et al.(19-20) to determine defects in their SiOx films and attributed the defect formation firstly, to limitation of surface mobility during plasma deposition; secondly, to dust particles formed during plasma process; and finally, to the additives in PET substrate. They also claimed that these defects were responsible for the residual permeation of O2.
Coating Thickness
Generally, the thickness of a barrier coating is 100-1000 times thinner than that of the polymeric substrate. In order for the deposited SiOx film to act as a gas barrier layer, a critical film thickness needs to be reached. Diverse values of critical film thickness have been reported, ranging from 15 nm-700 nm(3,7,20). Although all these SiOx films are deposited by plasma deposition method, deposition conditions such as plasma excitation modes, power and monomer gas flow rate are different. Thickness of the barrier coating depends partly on polymeric substrate surface roughness. Rougher polymer surface requires thicker coating in order to cover the whole area of the substrate otherwise barrier properties will drop. However, it has to keep in mind that thick coating tends to promote cracks.
Adhesion between Deposited SiOx Film and Polymer Substrate
Good adhesion between deposited barrier coating and polymeric substrate is crucial for packaging materials. In the case of PET substrate, good adhesion between silicon oxide and PET is obtained without surface modifi-cation of PET. Formation of Si-O-C and Si-C bonds between silicon oxide and PET substrate were reported in literature(18). However, for PE and PP, surface modification is required to promote interfacial adhesion. Rotger et al.(18) showed that without N2 plasma treatment prior to SiOx deposition, the SiOx coating tend to form islands on untreated surfaces of PE. The bonds linking silicon oxide and modified PE substrate were found to be Si-O-C and Si-N-C bonds.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองรบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
276
Inagaki et al.(14) demonstrated that poor adhesion between SiOx film and oriented polypropylene (OPP) substrate results in no improvement of oxygen permeation rate (> 2,000 cm3/(m2⋅day⋅atm) for both pure OPP and OPP coated with 100 nm thick SiOx film. However, after surface modification of OPP to improve adhesion, oxygen transmission rate is reduced to 37-52 cm3/(m2⋅day⋅atm).
Conclusions
Chemical structures of deposited SiOx film strongly influence O2 transmission rate. SiOx films containing low content of carbona-ceous compounds and O/Si ratio ranging from 1.4-1.85 show large improvements in O2-barrier property. Densely packed or highly cross-linked SiOx film possesses low O2 transmission rate than film having a lower density. Smooth surface with well connected intergranular SiOx grains results in an enhancement of O2-barrier property. Residual O2 transmission rate is caused by micro- and nano-defects. Micro-defects could be revealed by a combination of reactive oxygen etching and microscopy techniques. A critical SiOx thickness is needed to be reached in order to obtain an efficient barrier improvement. Without sufficient adhesion between the SiOx film and polymeric substrate, improvement in O2-barrier cannot be achieved.
References: 1. Barrer RM. Diffusion in and through solid.
Cambridge: Cambridge University Press; 1941. 2. Benmalek M, Dunlop HM. Inorganic coatings on
polymers. Surf Coat Tech 1995;76-77:821-6. 3. Bieder A, Gruniger A, von Rohr PR. Deposition
of SiOx diffusion barrier on flexible packaging materials by PECVD. Surf Coat Tech 2005;200: 928-31.
4. Chatham H. Oxygen diffusion barrier properties of transparent oxide coatings on polymeric substrates. Surf Coat Tech 1996; 78:1-9.
5. Clark DT, Dilks A, Shuttleworth D. The application of plasma to the synthesis and surface modification of polymers: polymer surfaces. New York: John Wiley & Sons; 1978. p. 188-99.
6. Deilmann M, Grabowski M, Theiβ S, Bibinov N, Awakowicz P. Permeation mechanisms of
pulsed microwave plasma deposited silicon oxide films for food packaging applications. J Phys D: Appl Phys 2008;4:135207.
7. Deilmann M, Theiβ S, Awakowicz P. Pulsed microwave plasma polymerization of silicon oxide films: Application of efficient permeation barriers on polyethylene terephthalate. Surf Coat Tech 2008;202:1911-7.
8. Deshmukh RR, Shetty AR. Modification of polyethylene surface using plasma polymeri-zation of silane. J Appl Polym Sci 2007;106: 4075-82.
9. Erlat AG, Spontak RJ, Clarke RP, Robinson TC, Haaland PD, Tropsha Y et al. SiOx gas barrier coatings on polymer substrates: morphology and gas transport considerations. J Phys Chem B 1999;103:6047-55.
10. Gruniger A, Bieder A, Sonnenfeld A, Rohr Ph, Muller U, Hauert R. Influence of film structure and composition on diffusion barrier per-formance of SiOx thin films deposied by PECVD. Surf Coat Tech 2006;200:4564-71.
11. Henry BM, Dinelli F, Zhao K-Y, Grovenor CR, Kolosov OV, Briggs GA et al. A micro-structure study of transparent metal oxide gas barrier films. Thin Solid Films 1999;355-356:500-5.
12. Inagagki N, Cech V, Narushima K, Takechi Y. Oxygen and water vapor gas barrier poly(ethylene naphthalate) films by deposition of SiOx plasma polymers from mixture of tetramethoxy silane and oxygen. J Appl Polym Sci 2007;104:915-25.
13. Inagagki N, Tasaka S, Hiramatsu H. Preparation of oxygen gas barrier poly (ethylene terephthlate) films by deposition of silicon oxide films plasma-polymerized from a mixture of tetramethoxy silane and oxygen. J Appl Polym Sci 1999; 71:2091-100.
14. Inagagki N, Tasaka S, Nakajima T. Preparation of oxygen gas barrier polypropylene films by deposition of SiOx films plasma-polymerized from a mixture of tetramethoxy silane and oxygen. J Appl Polym Sci 2000;78:2389-97.
15. Lang J, Wyser Y. Recent inovation in barrier tech-nologies for plastic packaging - a review. Packag Technol Sci 2003;16:149-58.
16. Lamendola R, d’Agostino R, Process control of organosilicon plasmas for barrier film preparations. Pure Appl Chem 1998;70:1203-8.
17. Leterrier Y. Durability of nanosized oxygen-barrier coatings on polymers. Prog Mat Sci 2003;48:1-55.
18. Rotger JC, Pireaux JJ, Thorne NA, Dunlop HM, Benmalek M. Deposition of silicon-oxide onto polyethylene and polyethylene-terephthalate: an

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
277
x-ray photoelectron-spectroscopy interfacial study. J Vac Sci Technol A 1995;13:260-7.
19. Sobrinho AS, Czermuszkin G, Latreche M, Dennler G, Wertheimer MR. A study of defects in ultra-thin transparent coatings on polymers. Surf Coat Tech 1999;116-119:1204-10.
20. Sobrinho AS, Latreche M, Czermuszkin G, Klemberg-Sapieha JE, Wertheimer MR. Transparent barrier coatings on polyethylene terephthlate by single and dual-frequency plasma-enhanced chemical vapor deposition. J Vac sci Technol A 1998;16:3190-8.
21. Teshima K, Inoue Y, Sugimura H, Takai O. Gas barrier properties of silicon oxide films prepared by plasma-enhanced CVD using tetramethoxy-silane, Vacuum 2002;66:353-7.
22. Tropsha YG, Harvey NG. Activated rate theory treatment of oxygen and water transport through silicon oxide poly (ethylene terephthalate) composite barrier structures. J Phys Chem B 1997;101: 2259-66.
23. Walker M, Meermann F, Schneider J, Bazzoun K, Feichtinger J, Schulz A et al. Investigations of plasma polymerized barrier films on polymeric materials. Surf Coat Tech 2005;200: 947-52.
24. Yasuda H. Plasma polymerization and plasma modification of polymer surfaces. In: Ebdon JR, editor. New methods of polymer synthesis. New York: Chapman and Hall; 1991. p.161-96.


279
Supported Planar Lipid Membranes and Their Applications
Pallop Karnchanaphanurach Department of Chemistry, Center for Innovation in Chemistry and Center for Alternative Energy, Faculty of Science, Mahidol University.
Abstract
Natural membranes provide key environments for many biological processes, including cellular communication and transport of small molecules. To understand the complex behaviors associated with such membranes, it is important to gain biophysical knowledge of the membranes themselves. Supported planar lipid membrane offers a crucial link towards the understanding of biomembranes and their related biological activities. This article presents a general overview of this type of model membrane, from preparation methods to exciting research arenas where supported membranes play an active role.
Keywords: supported planar lipid membrane; bilayer; vesicle; lateral mobility Introduction
Living cells and organelles could not have existed and maintained their functions without a natural mechanism to create a boundary from the environment and to produce compartmentalization within cells. Such mecha-nism is provided by biological membrane, which consists of a lipid bilayer. In addition to cholesterol (in mammalian cells) and phospho-lipids of various chain lengths and head groups, the bilayer also contains many proteins and carbohydrates inserted or attached to the membrane surface. Integral proteins such as ion channels span the membrane asymmetrically. Peripheral proteins associate with the lipid bilayer either by partially immersing in the lipids, by anchoring to the lipids, or by interacting with other proteins in the membrane. Many peripheral proteins, such as the human ubiquitously expressed protein decay-acce-lerating factor (DAF or CD55), anchor to exoplasmic membrane surface via a glycosyl-phosphatidylinositol (GPI) linkage, which in turn is partially inserted into the lipid bilayer(9,23,25). A high percentage of membrane proteins and lipids in human are also covalently linked to carbohydrates, which are thought to play a vital role in cell communication with the surroundings(34). All of these molecular compo-
nents are held together non-covalently in the membrane, allowing most of them to have lateral mobility. This ability of lipids and proteins to traverse in the two-dimensional plane is fundamental to many biological pro-cesses(19,33). Together with the structural complexity of the membrane, the dynamics of various membrane components adds up to an already complicated picture of the living system, often too complex to be dissect scientifically.
Supported planar lipid membranes have proven to be a successful model for the study of membrane structure and protein dynamics, owing to a number of advantages. Lipids and reconstituted proteins in supported membranes retain lateral mobility(3). Molecular properties of membrane constituents and their compo-sitions can be systematically varied. Moreover, these model membranes offer a thin, optically-penetrable, two-dimensional structures that can be studied using a wide range of interface-sensitive techniques that include total internal reflection fluorescence(18), atomic force spectro-scopy(29,31), X-ray and neutron scattering(17,21), fourier-transform infrared spectroscopy(42),
quartz crystal microbalance(20), surface Plasmon resonance(20,43), nuclear magnetic resonance(1), and even mass spectrometry(22). This article presents a general overview of the physical

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
280
properties of supported planar lipid membranes, along with key aspects of their research applications.
Supported Planar Lipid Membranes
The lipid bilayer is separated from the solid support by an ultrathin film of water molecules of thickness 5-20 Å(1,17) (see Fig 1). This water layer serves as a lubricant to relieve the membrane of motional friction with the underlying substrate surface, which is usually glass, mica, silicon, or quartz. The suppressed friction thus enables the lipids to move freely in a two-dimensional plane. The supported membranes are stably held onto the substrate by a combination of hydration, steric, electrostatic, and van der Waals forces(8). The thermal and mechanical stability of the membrane is sufficient to withstand high flow rate of solution(40), making them very useful particularly in flow-based experiments.
inability to incorporate membrane proteins into the bilayer because of the potential loss of proteins to the aqueous reservoir that suspends the monolayer. In addition, during the transfer of the monolayer, proteins may be exposed to air and consequentially dehydrate, resulting in the denaturation of the proteins.
The second approach to prepare supported bilayers involves spontaneous fusion of the unilamellar lipid vesicles on solid substrate. The fusion process is thought to initiate with low-density adsorption of the vesicles to the solid surface(20,49). At a critical density of the adsorbed vesicles, they coalesce to form larger vesicles(31). These vesicles ultimately rupture, fuse, and spread to form a lipid bilayer(31). To make supported membranes biologically functional, vesicles with recon-stituted integral proteins can be used to infuse
Figure 1 Schematic diagram showing the main structure of a supported planar lipid bilayer.
The supported lipid membranes are prepared by three general approaches: i) mono-layer transfer by Langmuir-Blodgett (LB) and Langmuir-Schaefer (LS) techniques (Fig 2A); ii) spontaneous lipid vesicle fusion (Fig 2B); and iii) combination of the two (Fig 2C).
In its simplest procedure, the first method involves vertically passing a solid substrate through a monolayer film of lipids suspended at air/water interface (LB) in order to deposit one layer of lipid membrane on the substrate. A bilayer is then formed by horizontally pressing the solid-supported monolayer to a suspended monolayer in a trough (LS). The advantage of this technique is its easy manipulation of lipid composition and density of the membrane. This means that the asymmetric properties of the membrane bilayer can be varied at will. However, the downside of this method is an
the membranes. Another powerful way to incorporate the proteins is to include some molecular anchors in the preformed planar bilayer and subsequently introduce proteins that have been tailored to specifically bind to those anchors. Since the vesicle fusion method primarily relies on the spontaneous fusion mechanism, a controlled variation of membrane compositions can be difficult to achieve. The advantages of the LB/LS technique and the vesicle fusion can be combined in the third method, where the lower leaflet is first deposited onto the substrate by the LB procedure, followed by spontaneous fusion of the vesicles to obtain the bilayer.
As noted earlier, the spacing between the bilayer and the solid support is only 5-20 Å(1,17). While this thin space may provide enough room to allow good lateral fluidity for lipids, it could

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
281
prove problematic for reconstituted transmem-brane proteins. Large integral proteins such as some ion channels likely protrude some distance from the membrane surface. If this protruded portion is located at the inner leaflet, the protein may contact directly the bare solid surface (Fig 3A), resulting in loss of protein mobility, functionality, and even complete denaturation. This problem can be overcome by adding a cushion between the membrane and the sub-strate as illustrated in Fig 3B(41). Polymer materials are good candidates for this purpose because of their generally hydrophilic proper-ties, chemical stability, and good wetting properties at the interface. A good example of a polymer cushion is regenerated cellulose, which has been shown to retain functionality of the incorporated proteins(12). An alternative strategy to increase the membrane-substrate gap makes use of lipopolymer linkers(30). The lipid-like ends of these linkers hook to the membrane while the macromolecular head groups act as a spacer (Fig 3C). The unique benefit of this approach comes from the fact that the spacer length, density, and viscosity of the linker layer can be systematically controlled. Another intriguing choice of spacer is bacterial cell-surface layer (S-layer) proteins(45). However, the use of S-layer proteins has so far been l imi t ed t o gold a nd s i lver s ubs t r a t es .
limited to gold and silver substrates. Protein Interactions at the Membrane Interface
Supported planar lipid membranes provide an excellent environment for preserving much of a protein function over a large two-dimensional macroscopic area. They are therefore a desirable model for investigations of molecular interactions at the membrane interface. The use of protein-functionalized supported membranes was first pioneered in the study of molecular recognition and associated dynamics in the immunological synapse(3). An important event in adhesion and com-munication of immune cells occurs when an antigen-presenting cell (APC) interacts with a T cell prior to activation of the immune response. This cell-cell interaction is strongly coupled with binding of T cell receptors (TCRs) on the T cell to peptide fragments carried by the major histocompatibility complex (MHC) on APC. The binding of multiple molecules creates a supramolecular activation cluster (SMAC). Upon binding, it is likely that the mobility of each protein will be altered. By using fluorescence video microscopy, Brian and McConnell(3) were able to visualize this crucial change in dynamics and demonstrate that
Figure 2 General procedures for the formation of supported bilayer. A, Langmuir-Blodgett (vertical dipping)
is used to deposit the monolayer, followed by Langmuir-Schaefer (horizontal contacting) to complete the bilayer structure. B, vesicle fusion directly deposits the bilayer on solid substrate. C, a combined method uses LB to prepare a single monolayer and sequentially introduces the upper leaflet via vesicle fusion

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
282
Figure 3. Degree of interaction of a transmembrane protein with three types of supports. A, bare solid support.
B, hydrated-polymer cushioned support. C, support cushioned by lipopolymer tethers. immobile peptide-MHC could activate the cytotoxic T cells. This sophisticated use of supported membranes had led since to many more immunological transmembrane communi-cation findings(5,13,39,44).
The dynamic trajectory for immunolo- gical synapse has been found to start with an initial diffuse protein distribution and end with a static protein organization. In addition to a better description of immunological synapse, accurate quantitative information has been accumulated over the years. For example, it has been reported that total MHC accumulation at the synapse has a strong correlation with the half-time of TCR-peptide-MHC binding(13), and that at least 60 peptide-MHC per m2 in SMAC is needed for proliferation(13). Together with theoretical modeling, it was further suggested that the increased accumulation of peptide-MHC is necessitated by the low TCR-peptide-MHC binding affinity and also helps level off the strong signal by inducing endocytosis of acti-vated TCRs(13).
The lenient membrane environment for proteins, the versatile geometry of the supported membrane systems, and the insulating properties of the bilayer all provide a suitable platform for electrochemical sensing, particularly in the study of ion channels(2). The set up is analogous to the traditional patch-clamp technique, which involves sealing a fine glass pipette tip with a cell or patch of membrane and monitoring the electric current activity as ions pass through the ion channels. In one such study, lipid bilayer with reconstituted gramicidin channels was polymer-cushioned on a semiconductor elec-trode(2). By using a combination of DC current recordings to observe channel activity and fluorescence imaging to simultaneously follow
co-localization of the proteins, a strong correlation was observed between channel activity and dimerization of gramicidin(2). An improvement of sensitivity of this approach to the level of single-molecule events has been achieved later by using membranes supported on indium-tin-oxide(15) or semicon-ductor materials(36,37) and analyzing with time-resolved Fourier transform impedance spectro-scopy.
The idea of a special form of membrane partitioning known as liquid-ordered, choles-terol-rich domains or lipid rafts has attracted considerable interest over the past decade(4,35). These domains are thought to provide crucial platforms for many biological processes, including cell signaling, cell adhesion, and pathogenic infection. Supported bilayers can serve as an indispensable tool for investigations of physical properties of domains that are normally difficult to study. Using nanometer scale-sensitive, integrated technique of atomic force microscopy and near-field scanning optical microscopy, phase separation and dynamic nature of such domains could be followed in real time(28). Particularly interesting is a stable existence of nanometer-sized domains, which were previously thought to be too thermo-dynamically unstable to occur in cell mem-branes(10,11). A more relevant system to cell membranes utilizes peptide- and protein-incorporated model bilayers, where it was found that -amyloid peptide A42 was preferentially localized in the domains(7). In addition to peptides, a protein called synapsin in the neural synapse system was also found to have domain-dependent activity(28).

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
283
Lipid Membrane Patterning
Due to the strong dependence on sup-porting materials, two-dimensional shapes of planar lipid bilayers can be controlled by directly manipulating the solid supports. If a substrate surface is overlaid with an array of lipid barriers, then an array of partitioned bilayers would be obtained. A variety of techniques, such as photolithography, electron-beam lithography and contact printing, can be used to create the barriers. Currently, two general schemes are used to generate the patterning. The first method directly confines bilayers to specific locations on the surface. This can be achieved either by using an optical mask to spatially control the cross-linking of some lipids such as diacetylene lipid(26) or printing onto/blotting out certain areas of the membrane by using lipid-specific stamp such as polydimethylsiloxane(16). Alternatively, lipid microarrays can be obtained by contact printing lipid-repellant barriers or hydrophobic proteins onto the substrate surface prior to the bilayer deposition(32). These barriers effectively confine the bilayers within the untreated areas, resulting in corrals of membranes on the substrate. Since the membrane fully retains its fluidity and bilayer structure in each corral(14), the micropatterned lipid membranes can potentially be used as platforms for combinatorial libraries of small molecules, proteins and other analytes. Parallel screening of candidates for drug discovery is also feasible.
In addition to biotechnological applica- tions, micropatterned lipid membranes offer new opportunities in cell membrane studies. In a study of immunological synapse, an array of 100-nm-linewidth chromium barriers was deposited onto glass substrate using electron-beam lithography(27). Artificial patterned APC membrane was then prepared on the substrate. The patterned chromium grids then acted as diffusion barriers to individual patches of membranes. When a T cell interacted with this patterned membrane, it was found that lipids, peptide-MHCs, as well as intercellular adhesion molecules (ICAMs) induced pattern formation of the TCRs and lymphocyte function-associated antigen 1 (LFA-1, a ligand of ICAM) on the T
cell membrane as well. The fact that the patterns on both membranes were governed by the arrangement of the chromium grids indicates that TCRs were not preclustered prior to contact with the membrane. An observation that each TCR was located in the grid corner nearest to the SMAC center also suggests that the TCR organization is linked to the cytoskeletal network of the T cell(27).
Hybrid Designs Using Supported Membranes
The ability to preserve protein functions of bilayers has found broad applications in new technologies, including the field of micro-fluidics. The inner surfaces of microfluidic channels that are coated with lipid bilayers protect enzymes from denaturation and give strong signal-to-noise ratio(46). When alkaline phosphatase is tethered to the lipid membrane-coated channel via biotin-streptavidin linkage, the result was a better signal-to-noise reading and only a small amount of protein was needed to run the assay(24). The application of fluidics to study enzymatic activity in supported membrane systems also resulted in greater sensitivity in data acquisition(24,46).
Supported planar lipid membranes also have been used as a biocompatible support for DNA-tethered vesicles(6). Two vesicles fuse together when placed in a close proximity. The rate of this fusion could be controlled by tweaking the degree of DNA hybridization(38), making this a potential tool for the study of membrane fusion. If the vesicles are composed of charged lipids, the distance between any two vesicles could be adjusted using electric field parallel to the bilayer surface(48). Vesicles can also be tethered to the bilayers with biotin-streptavidin coupling. It was demonstrated with single-molecule accuracy that yeast SNARE proteins incorporated in bilayer-tethered vesicles mediate fusion with free-standing SNARE-vesicles(47).
Conclusions
Biologically functional environment and versatile manipulation of supported planar lipid membranes have given researchers a powerful

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
284
tool to address important challenges in cell membrane phenomena. Applications of this class of model membranes in immunological and neural synapse studies and in the lipid raft characterizations have yielded a plethora of insightful information. Development of more sophisticated supported membranes is ongoing, and there is no reason not to expect more new discoveries on membrane interfaces. Coupling between supported membranes and new analytical techniques marks a promising possibility for future technological inventions. In microfluidics applications, bilayers with functional proteins may give nanodevices an immunologically-benign shield and molecular specificity needed to make successful thera-peutic agents. Patterning technology with greater precision and accuracy could pave the way for combinatorial biochemistry and membrane-active drug screening. Supported bilayers also serve as a platform not only for functionalized vesicles, but also for pathogenic studies. Model membranes may be constructed to resemble envelopes of viruses of interest. These membranes could then be used to reveal much of the unknown factors in viral infection. They can even be envisioned to provide core structures for new drug delivery systems.
Acknowledgements
The author would like to thank Asiya Mehtar and Pongsit Tangcananurak for their invaluable support. This article is financially supported in part by the Thailand Research Fund, and the Center for Innovation in Chemistry and the Center for Alternative Energy, Faculty of Science, Mahidol University.
References 1. Bayerl TM, Bloom M. Physical-properties of
single phospholipid-bilayers adsorbed to micro glass-beads-a new vesicular model system studied by H-2-nuclear magnetic-resonance. Biophys J 1990;58:357-62.
2. Borisenko V, lougheed T, hesse J, Füreder-Kitzmüller E, Fertig N, Behrendo JC et al. Simultaneous optical and electrical recording of single gramicidin channels. Biophys J 2003;84: 612-22.
3. Brian AA, McConnell HM. Allogeneic stimuli-tion of cytotoxic T cells by supported planar
membranes. Proc Natl Acad Sci USA 1984;8: 6159-63.
4. Brown DA, London E. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J Biol Chem 2000;275:17221-4.
5. Bruinsma R, Behrisch A, Sackmann E. Adhesive switching of membranes: experiment and theory. Phys Rev E 2000;61:4253-67.
6. Chan Y-HM, Lenz P, Boxer SG. Kinetics of DNA-mediated docking reactions between vesicles tethered to supported lipid bilayers. Proc Natl Acad Sci USA 2007;104:18913-8.
7. Cordy JM, Hooper NM, Turner AJ. The involvement of lipid rafts in Alzheimer’s disease (review). Mol Membr Biol 2006;23:111-22.
8. Cremer PS, Boxer SG. Formation and spreading of lipid bilayers on planar glass supports. J Phys Chem B 1999;103:2554-9.
9. Daniels G. Functions of red cell surface proteins. Vox Sang 2007;93:331-40.
10. De Almeida RFM, Loura LMS, Federov A, Prieto M. Lipid rafts have different sizes depending on membrane composition: a time-resolved fluorescence resonance energy transfer study. J Mol Biol 2005;346:1109-20.
11. Frolov VAJ, Chizmadzhev YA, Cohen FS, Zimmerberg J. “Entropic traps” in the kinetics of phase separation in multicomponent membranes stabilize nanodomains. Biophys J 2006;91:189-205.
12. Goennenwein S, Tanaka M, Hu B, Moroder L, Sackmann E. Functional incorporation of inte-grins into solid supported membranes on ultrathin films of cellulose: impact on adhesion. Biophys J 2003;85:646-55.
13. Grakoui A, Bormley SK, Sumen C, Davis MM, Shaw AS, Allen PM et al. The immunological synapse: a molecular machine controlling T cell activation. Science 1999;285:221-7.
14. Groves JT, Ulman N, Boxer SG. Micro-patterning fluid lipid bilayers on solid supports. Science 1997;275:651-3.
15. Hillebrandt H, Wiegand G, Tanaka M, Sackmann E. High electric resistance polymer/lipid composite films on indium-tin-oxide electrodes. Langmuir 1999;15:8451-9.
16. Hovis JS, Boxer SG. Patterning barriers to lateral diffusion in supported lipid bilayer membranes by blotting and stamping. Langmuir 2000;16: 894-7.
17. Johnson S, Bayer TM, McDermott DC, Adam GW, Rennie AR, Thomas RK et al. Structure of an adsorbed dimyristoylphosphatidylcholine bi-

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
285
layer measured with specular reflection of neutrons. Biophys J 1991;59:289-94.
18. Kalb E, Frey S, Tamm LK. Formation of supported planar bilayers by fusion of vesicles to supported phospholipid monolayers. Biochim Biophys Acta 1992;1103:307-16.
19. Karnchanaphanurach P,Mirchev R, Ghiran I, Asara JM, Papahadjopoulos-Sternberg B, Nicholson-Weller A et al. C3b deposition on human erythrocytes induces the formation of a membrane skeleton-linked protein complex. J Clin Invest 2009;119:788-801.
20. Keller CA, Glasmaestar K, Zhdanov VP, Kasemo B. The formation of supported mem-branes from vesicles. Phys Rev Lett 2000;84: 5443-6.
21. Kjaer K, Als-Nielsen J, Helm CA, Laxhuber LA, Mohwald H. Ordering in lipid monolayers studied by synchrotron X-ray-diffraction and fluorescence microscopy. Phys Rev Lett 1987; 58:2224-7.
22. Kraft ML, Weber PK, Longo ML, Hutcheon ID, Boxer SG. Phase separation of lipid membranes analyzed with high-resolution secondary ion mass spectrometry. Science 2006;313:1948-51.
23. Lukacik P, Roversi P, White J, Esser D, Smith GP, Billington J et al. Complement regulation at the molecular level: the structure of decay-accelerating factor. Proc Natl Acad Sci USA 2004;101:1279-84.
24. Mao H, Yang T, Cremer PS. Design and characterization of immobilized enzymes in microfluidic systems. Anal Chem 2002;74:379-85.
25. Medof ME, Lublin DM, Holers VM, Ayers DJ, Getty RR, Leykam JF et al. Cloning and characterization of cDNAs encoding the complete sequence of decay-accelerating factor of human complement. Proc Natl Acad Sci USA 1987;84:2007-11.
26. Morigaki K, Baumgart T, Offenhausser A, Knoll W. Patterning solid-supported lipid bilayer membranes by lithographic polymerization of a diacetylene lipid. Ang Chem Inter Ed 2001; 40:172-4.
27. Mossman KD, Campi G, Groves JT, Dustin ML. Altered TCR signaling from geometrically repatterned immunological synapses. Science 2005;310:1191-3.
28. Murray J, Cuccia L, Ianoul A, Cheetham JC, Johnston LJ. Imaging the selective binding of synapsin to anionic membrane domains. Chem Bio Chem 2004;5:1489-94.
29. Puntheeranurak T, Stroh C, Zhu R, Angsuthanasombat C, Hinterdorfer P. Structure
and distribution of the Bacillus thuringiensis Cry4Ba toxin in lipid membranes. Ultramicro-scopy 2005;105:115-24.
30. Purrucker O, Foertig A, Jordan R, Tanaka M. Supported membranes with well-defined polymer tethers-incorporation of cell receptors. Chem Phys Chem 2004;5:327-35.
31. Reviakine I, Brisson A. Formation of supported phospholipid bilayers from unilamellar vesicles investigated by atomic force microscopy. Langmuir 2000;16:1806-15.
32. Sapuri AR, Baksh MM, Groves JT. Electro-statically targeted intermembrane lipid exchange with micropatterned supported membranes. Langmuir 2003;19:1606-10.
33. Saxton MJ, Jacobson K. Single-particle tracking: applications to membrane dynamics. Annu Rev Biophys Biomol Struct 1997;26:373-99.
34. Sharon N, Lis H. Carbohydrates in cell recog-nition. Sci Am 1993;268:82-9.
35. Simons K, Vaz WL. Model systems, lipid rafts, and cell membranes. Annu Rev Biophys Biomol Struct 2004;33:269-95.
36. Steinhoff G, Purrucker O, Tanaka M, Stutzmann M, Eickhoff M. AlxGa1-xN-a new material system for biosensors. Adv Funct Mater 2003; 13:841-6.
37. Steinhoff G, Baur B, Wrobel G, Ingebrandt S, Offenhäusser A, Dadgor A et al. Recording of cell action potentials with AlGaN/GaN field-effect transistor. Appl Phys Lett 2005;86: 033901.
38. Stengel G, Zahn R, Hook F. DNA-induced programmable fusion of phospholipid vesicles. J Am Chem Soc 2007;129:9584-5.
39. Stoddart A, Dykstra ML, Brown BK, Song WX, Pierce SK, Brodsky FM. Lipid rafts unite signaling cascades with clathrin to regulate BCR internalization. Immunity 2002;17:451-62.
40. Tadini-Buoninsegni F, Bartolommei G, Moncelli MR, Fendler K. Charge transfer in p-type ATPases investigated on planar membranes. Arch Biochem Biophys 2008;476:75-86.
41. Tanaka M, Sackmann E. Polymer-supported membranes as models of the cell surface. Nature 2005;437:656-63.
42. Tatulian SA, Hinterdorfer P, Baber G, Tamm LK. Influenza hemagglutinin assumes a tilted con-formation during membrane-fusion as determined by attenuated total-reflection FTIR spectroscopy. EMBO J 1995;14:5514-23.
43. Terrettaz S, Stora T, Duschl C, Vogel H. Protein-binding to supported lipid-membranes-investiga-tion of the cholera-toxin ganglioside

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
286
interaction by simultaneous impedance spectro-scopy and surface-plasmon resonance. Langmuir 1993;9: 1361-9.
44. Watts TH, Gaub HE, McConnell HM. T-cell-mediated association of peptide antigen and major histocompatibility complex protein detected by energy-transfer in an evanescent wave-field. Nature 1986;320:179-81.
45. Wetzer B, Pum D, Sleytr UB. S-layer stabilized solid supported lipid bilayers. J Struct Biol 1997; 119:123-8.
46. Yang T, Jung SY, Mao H, Cremer PS. Fabrication of phospholipid bilayer-coated micro-channels for on-chip immunoassays. Anal Chem 2001;73:165-9.
47. Yoon T-Y, Okumus B, Zhang F, Shin Y-K, Ha T. Multiple intermediates in SNARE-induced membrane fusion. Proc Natl Acad Sci USA 2006;103:19731-6.
48. Yoshina-Ishii C, Boxer SG. Controlling two-dimensional tethered vesicle motion using an electric filed: interplay of electrophoresis and electro-osmosis. Langmuir 2006;22:2384-91.
49. Zhdanov VP, Keller CA, Glasmaestar K, Kasemo B. Simulation of adsorption kinetics of lipid vesicles. J Chem Phys 2000;112:900-9.

287
Whole-cell Biocatalysts from Organic Solvent-tolerant Bacteria
Thunyarat Pongtharangkul1 and Alisa S Vangnai2 1Department of Biotechnology, Faculty of Science, Mahidol University, 2Department of Biochemistry, Faculty of Science, Chulalongkorn University.
Abstract
Whole-cell biocatalyst is expected to be one of the most important tools in industrial synthesis of bulk chemicals as well as pharmaceutical and agrochemical intermediates and bioactive compounds. The use of whole cells not only provides a means for cost-effective production and regeneration of enzyme cofactors, such as NAD(P)H, but also allows the utilization of multi-component and membrane-bound enzymes in biotransformation of chemicals. As several potential substrates and products are, however, sparingly water-soluble or water-immiscible, a second phase of organic solvent is required. Development of effective solvent-tolerant microbial platform is, therefore, a necessary step to make whole-cell biocatalyst applicable for production of water-immiscible chemicals. Several examples of whole-cell biocatalysts based on organic solvent-tolerant bacteria are presented.
Keywords: biotransformation; organic-aqueous two-liquid phase system
Introduction Chemical industry not only consumes a tremendous amount of energy and non-renewable resources, but also produces toxic wastes and pollutants. Therefore, attempts have been carried out in order to develop more environmentally friendly industrial chemical processes. The implementation of ‘green chemistry’ concept in which a chemical should be synthesized in a safe and energy-efficient manner with the lowest impact on environ-ment(1) emphasizes the role of biocatalysis processes. The fact that a high stereo- or enantiomerically pure product can be synthesized in a mild condition makes bio-catalysts a promising alternative for production of several selective and complex chemicals(35).
Comparing to an isolated enzyme system, whole-cell biocatalyst is more cost-effective in terms of production, especially in systems that require regeneration of co-factors(16) and that involve multi-component or membrane-bound enzymes. Various potential substrates and products, however, are water-immiscible and thus require the use of solvents for solubilization. Although the solvent phase in
organic-aqueous two-phase system can be used as a means to overcome the toxicity of substrates and products as well as to recover products in situ, toxicity of the solvent phase itself has become a major drawback in the applications of whole-cell biocatalysts. For-tunately, recent discovery of solvent-tolerant bacteria has made whole-cell biocatalysis in an organic-aqueous two-phase system more promising.
Organic Solvent-tolerant Bacteria
Organic solvent-tolerant bacteria con-stitute a unique group of extremophilic microorganisms that can thrive in an environment with high concentrations of solvents(19). These bacteria have been consi-dered to be potentially useful in many applications of microbial transformation for environmental remediation as well as for production of valuable chemicals(32). Although most of the solvent-tolerant bacteria reported thus far are Gram-negative, a number of Gram-positive bacteria, sometimes with even higher solvent tolerance level, were also reported (Table 1). The adaptive mechanisms of solvent

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
288
Table 1 Examples of organic-solvent tolerant bacteria
Microorganism %(v/v) Solvent
Solvent (logPow)
Reference
1-B
utan
ol (0
.88)
Chl
orof
orm
(1.9
7)
Ben
zene
(2.1
3)
n-H
epta
nol (
2.62
)
Tol
uene
(2.7
3)
Xyl
ene
(3.1
6)
Cyc
lohe
xane
(3.2
)
Die
thyl
phth
alat
e (3
.3)
n-H
exan
e (3
.5)
n-H
epta
ne (4
.66)
n-D
ecan
e (5
.6)
Pseudomonas putida NRRL B-18435 25 ‐ ‐ ‐ + + + + + + + + (6)
Pseudomonas putida S12 1 * * ‐ + + + + + + * + (40)
Bacillus stearothermophilus BR325 Plate * * + * + * * * * * * (26)
Pseudomonas putida DOT-T1 10 ‐ ‐ ‐ + + + + + * + + (29)
Bacillus sp. SB1 Plate + + + * + + + * + * + (32)
Bacillus coagulans NBRC 12583 1 + ‐ * * ‐ ‐ ‐ * + ‐ + (24)
Leuconostoc fallax NRIC 0210 1 + ‐ * * ‐ ‐ + * + + + (24)
S. epidermis ATCC 14990 Plate * * + * + + + * * * * (27)
Rhodococcus opacus B-4 10 ‐ ‐ + ‐ + + + + + * + (25)
Pseudomonas aeruginosa 50 ‐ ‐ ‐ * ‐ + + * + + + (14)
Enterobacter aerogenes 20 ‐ * ‐ * ‐ ‐ + * + + * (15)
Burkholderia cepacia ZZ8 0.2-2 * * ‐ ‐ ‐ ‐ + + + * * (46)
Staphylococcus sp. ZZ1 0.2-2 * * + ‐ + + + + + * * (46)
Bacillus cereus ZZ3 0.2-2 * * + ‐ + + + + + * * (46)
Plate = growth on agar plate overlaid with solvents; *unavailable data; + and –, positive and negative solvent tolerance properties, respectively.
tolerance have been discussed in detail elsewhere(16).
The value of logPow in Table 1 represents a partition coefficient of the solvent to octanol from the water phase. Several studies have reported significant correlation between bio-compatibility of an organic solvent and its logPow
(2,6,20,38). The concentration of solvent present in the cell membrane actually depends on several factors including its concentration in the aqueous phase, partition coefficient into the membrane, and volume ratio of the two liquid phases. Physical disruption of the cytoplasmic membrane usually occurs when the solvent concentration in the membrane reaches a certain level(16). In general, solvents with logPow ranging between 1 and 4 are considered as being highly toxic. This is because a
compound with logPow < 1 would be too hydrophilic to be incorporated into a cell membrane, while a compound with logPow > 4 is barely soluble in water and thus cannot partition into the aqueous phase where the cell usually exists. It should be noted, however, that several solvent-tolerant bacteria previously reported could tolerate highly toxic solvents, such as benzene and toluene, at very high concentrations.
Organic-aqueous Two-liquid Phase Bio-transformation System
During the past decade, several whole-cell biotransformations have been implemented using organic-aqueous two-liquid phase systems in various scales(16,22). The most commonly used genetic engineering host,

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
289
Escherichia coli, was found to be inappropriate in several cases, especially when substrates or products-of-interest are highly toxic(4,33,39). Therefore, several bio-transformation systems have been developed based on organic solvent-tolerant bacteria (Table 2). As the solvent-tolerant bacteria do not always possess the desired properties, a number of genetically modified biocatalysts were created by intro-ducing gene(s)-of-interest. Solvent-tolerant Pseudomonas was the most popular choice due to its excellent solvent tolerance and availabi- lity of geneticrinformation.
Most of the solvent-tolerant biocatalysts discovered thus far, including Pseudomonas sp., are hydrophilic (Figure 1). They are suitable for biotransformation of substrates that
are somewhat soluble in water. This platform, however, will be less effective for water-immiscible substrates such as naphthalene. The need for more hydrophobic biocatalysts, with greater access to water-immiscible substrates, therefore, has created a niche for a unique group of hydrophobic solvent-tolerant bacteria, such as Rhodococcus opacus B-4(17) and R. erythropolis DCL14(9). These hydrophobic biocatalysts can act at the interface(17) as well as being able to disperse evenly in an anhydrous solvent(45). It should be noted, however, that the substrate and product of the hydrophobic platform should not be highly toxic because of the greater chance of a direct contact between the biocatalyst and the solvent. Having two different platforms, hydrophilic and hydro-
Table 2 Examples of whole-cell biocatalysts from organic solvent-tolerant bacteria.
Product Biocatalyst Organic Phase References
Hydrocortisone derivatives Arthrobacter simplex ATCC 6946 Octanol (13) Indandiol Pseudomonas putida ATCC 55687 Silicone oil (3) Rhodococcus sp. B264-1* Silicone oil (3) Toluene cis glycol Rhodococcus sp. MA 7249* Silicone oil (5) 3-Methylcatechol Pseudomonas putida S12* Octanol (42)
Pseudomonas putida MC2* Octanol (18) Pseudomonas putida DOT-T1E* Octanol (30)
5-Methylpyrazine-2-carboxylic acid
Pseudomonas putida ATCC 33015 p-xylene (34)
Carvone Rhodococcus erythropolis DCL14 n-dodecane (9) Indigo Acinetobacter sp. ST-550 Diphenylmethane (11)
Rhodococcus opacus B-4* bis(2-ethylhexyl)phthalate) (17, 45) 4-Androstene-3,17-dione Rhodococcus equi Tetrachloroethane (7)
Mycobacterium sp. NRRL B-3805 bis(2-ethylhexyl)phthalate) (41) Phenol Pseudomonas putida S12* Octanol (43) o-Cresol Pseudomonas putida T-57 Octanol (12)
Rhodococcus opacus B-4* bis(2-ethylhexyl)phthalate) (17) (-)-Perillyl alcohol Pseudomonas putida* bis(2-ethylhexyl)phthalate) (37) Decarboxycitrinin Moraxella sp. MB1 Ethyl acetate (10) (S)-Styrene oxide Pseudomonas sp. VLB120ΔC* bis(2-ethylhexyl)phthalate) (28) Vanillin Pseudomonas putida IE27 Dimethyl sulfoxide (44) Alkane hydroxylation Rhodococcus opacus B-4 bis(2-ethylhexyl)phthalate) (31)
* indicates recombinant or mutated biocatalyst.

29
วทยาศาสตรเพ
90
Fig
phobic, substrateincreaseschemica
SolventDevelop
Insecond biocompcoefficieOctanol bioconvesuch as noted, hobiocatalyliquid phphase. Fethylhexfrequentits low bility, acompareBEHP temulsioncomplicatwo-liquyeast extemulsify
พอการพฒนา: รวมบ
gure 1 Hydro biotra
greatly expaes and produs the potentl synthesis.
t Selection pment
n general, anphase is ch
patibility asent (Kp) o(logPow = 2.
ersion of pphenols andowever, that ysts would nhase systemFor more hyxyl) phthalatetly selected a
toxicity, poand relativelyed with othetends to fon, making tated, when y
uid phase sytract should
ying agents
บทความวชาการ ฉลอ
ophilic and hyansformation s
ands the ranucts, and thutial of biotra
and Proces
n organic solhosen with s well as
of the prod.9) is commo
products witd catechols(8)
commonly unot stand a ch
m with octanydrophobic pe (BEHP) (l
as a solvent-ooor flammay inexpensir pure solveorm a stabthe product yeast extractystem(4). In be omitted os (e.g. hy
งครบรอบ 50 ป แหง
ydrophobic biosystem.
nge of possius significanansformation
ss
lvent used aregards to the partit
duct-of-interonly chosen th low logP). It shouldused whole-chance in a tw
nol as a secoproducts, bislogPow = 9.6of-choice dueability, recycve price wh
ents. Howevble oil-in-wa
recovery mt was fed to
this case, or microbial ydrolases a
งการสถาปนาคณะวท
ocatalysts in a
ible ntly n in
as a its
tion est. for
Pow, d be cell wo-ond (2-) is e to cla-hen ver, ater
more the the de-and
proLecelR.redtraseclarbioforbioits
unforemconintadfrain of orgrec(frA indem
ทยาศาสตร มหาวทยา
an organic-aqu
oteases) shoueppchen et all de-emulsif
erythropoliduce the pansformationconds. This rge-scale dowotransformatrms stableoemulsifier(self.
Althougndesirable prrmation of
mulsion gennversion rateterface areadition of sur
action and agorder to redsubstrate
ganic phase commended rom oil-in-wrecent study
dicated the mulsion (at an
ลยมหดล
ueous two-liqu
uld be applial.(23) demonfication usinis DSM 43
phase separan media from
approach wwnstream ption broth,
emulsion s) produced
h a stableroperty for d
small solvnerally rese as a conseqa(21). Severarfactant, incrgitation speedduce the masand producfraction of
in order to awater to watey from Hond
practicalityn organic pha
uid phase
ed. Recent strated that
ng Bacillus s3297 can dation time
m 1 week towould greatly
rocess of wwhich fras a re
d by the bi
e emulsiondownstream
vent dropletults in aquence of thal approachreasing organd) have beenss transfer limct(36). Generf less than 0avoid phase rer-in-oil emuda et al.(17), y of a waase fraction
study by a whole-ubtilis or
drastically of bio-
o only 20 y simplify whole-cell frequently esult of iocatalyst
n is an process,
ts as an a higher he greater hes (e.g. nic phase n adopted mitations rally, an 0.6-0.7 is reversion
ulsion)(22). however,
ater-in-oil of 0.9) in

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
291
which stable water droplets can be easily separated by sedimentation. It should be noted that this system greatly depends on the ability of the biocatalyst to act at the interface. Irrespective of an oil-in-water or a water-in-oil system, a continuous phase should be employed where the reaction substrate can be added because a continuous feeding of a toxic substrate at a sub-lethal concentration was shown to enhance the biotransformation even further(5).
Conclusions
Recent discoveries of organic solvent-tolerant bacteria have opened up the possibility of using whole cells in biocatalysis of water-immiscible or toxic compounds. An organic-aqueous two-phase system not only helps to overcome poor substrate solubility, but also gives an opportunity to recover the product in situ. Several whole-cell biocatalysts based on organic solvent-tolerant bacteria have been developed for production of various complex chemicals. So far, they have proven the potential of this system and have attracted increased attention to the field.
References 1. Anastas PT, Warner JC. Green chemistry:
theory and practice. New York:Oxford University Press; 1998.
2. Bassetti L, Tramper J. Organic solvent toxicity in Morinda citrifolia cell suspensions. Enzyme Microb Technol 1994;16:642-8.
3. Buckland BC, Drew SW, Connors NC, Chartrain MM, Lee C, Salmon PM et al. Microbial conversion of indene to indandiol: A key intermediate in the synthesis of CRIXIVAN. Metabolic Eng 1999;1:63-74.
4. Buhler B, Bollhalder I, Hauer B, Witholt B, Schmid A. Use of the two-liquid phase concept to exploit kinetically controlled multistep biocatalysis. Biotechnol Bioeng 2003;81:683-94.
5. Chartrain M, Ikemoto N, Taylor C, Stahl S, Sandford V, Gbewonyo K et al. Production of cis-1,2-dihydroxy-3-methylcyclohexa-3,5-diene (toluene cis glycol) by Rhodococcus sp. MA 7249. J Biosci Bioeng 2000;90:321-7.
6. Cruden DL, Wolfram JH, Rogers RD, Gibson DT. Physiological properties of a Pseudomonas strain which grows with p-xylene in a two-phase
(organic-aqueous) medium. Appl Environ Microb 1992;58:2723-9.
7. Cruz A, Fernandes P, Cabral JMS, Pinheiro HM. Solvent partitioning and whole-cell sitosterol bioconversion activity in aqueous-organic two-phase systems. Enz Microbial Technol 2004; 34:342-53.
8. de Bont JAM. Solvent-tolerant bacteria in biocatalysis. Trends in Biotechnol 1998;16: 493-9.
9. de Carvalho CCCR, da Fonseca MMR. Influence of reactor configuration on the production of carvone from carveol by whole-cells of Rhodococcus erythropolis DCL14. J Mol Catalysis B: Enzymatic 2002;19-20:377-87.
10. Devi P, Naik CG, Rodrigues C. Biotrans-formation of citrinin to decarboxycitrinin using an organic solvent-tolerant marine bacterium, Moraxella sp. MB1. Marine Biotechnol 2006; 8:129-38.
11. Doukyu N, Nakano T, Okuyama Y, Aono R. Isolation of an Acinetobacter sp. ST-550 which produces a high level of indigo in a water-organic solvent two-phase system containing high levels of indole. Appl Microbiol Biotechnol 2002; 58:543-6.
12. Faizal I, Dozen K, Hong CS, Kuroda A, Takiguchi N, Ohtake H et al. Isolation and characterization of solvent-tolerant Pseudo-monas putida strain T-57, and its application to biotransformation of toluene to cresol in a two-phase (organic-aqueous) system. J Ind Micro-biol Biotechnol 2005;32:542-7.
13. Fernandes P, Cabral JMS, Pinheiro HM. Bio-conversion of a hydrocortisone derivative in an organic-aqueous two-liquid-phase system. Enz Microbial Technol 1995;17:163-7.
14. Gupta A, Khare SK. A protease stable in organic solvents from solvent tolerant strain of Pseudomonas aeruginosa. Biores Tech 2006; 97:1788-93.
15. Gupta A, Singh R, Khare SK, Gupta MN. A solvent tolerant isolate of Enterobacter aero-genes. Biores Tech 2006;97:99-103.
16. Heipieper HJ, Neumann G, Cornelissen S, Meinhardt F. Solvent-tolerant bacteria for bio-transformations in two-phase fermentation systems. Appl Microbiol Biotechnol 2007; 74:961-73.
17. Honda K, Yamashita S, Nakagawa H, Sameshima Y, Omasa T, Kato J et al. Stabili-zation of water-in-oil emulsion by Rhodococcus opacus B-4 and its application to biotrans-

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
292
formation. Appl Microbiol Biotechnol 2008;78: 767-73.
18. Husken LE, Dalm MCF, Tramper J, Wery J, de Bont JAM, Beeftink R. Integrated bioproduction and extraction of 3-methylcatechol. J Bio-technol 2001;88:11-9.
19. Inoue A, Horikoshi K. A Pseudomonas putida thrives in high concentration of toluene. Nature 1989;338:264-6.
20. Isken S, de Bont JAM. Bacteria tolerant to organic solvents. Extremophiles 1998;2:229-38.
21. Kim MK, Rhee JS. Lipid hydrolysis by Pseudomonas putida 3SK cultured in organic-aqueous two-phase system. Enz Microb Technol 1993;15:612-16.
22. Leon R, Fernandes P, Pinheiro HM, Cabral JMS. Whole-cell biocatalysis in organic media. Enz Microb Technol 1998;23:483-500.
23. Leppchen K, Daussmann T, Curvers S, Bertau M. Microbial de-emulsification: a highly efficient procedure for the extractive workup of whole-cell biotransformations. Org Proc Res Develop 2006; 10:1119-25.
24. Matsumoto M, Mochiduki K, Kondo K. Toxicity of ionic liquids and organic solvents to lactic acid-producing bacteria. J Biosci Bioeng 2004; 98:344-7.
25. Na K, Kuroda A, Takiguchi N, Ikeda T, Ohtake H, Kato J. Isolation and characterization of benzene-tolerant Rhodococcus opacus strains. J Biosci Bioeng 2005;99:378-82.
26. Natarajan MR, Lu Z, Oriel P. Cloning and expression of a pathway for benzene and toluene from Bacillus stearothermophilus. Biodegradation 1994;5:77-82.
27. Nielsen LE, Kadavy DR, Rajagopal S, Drijber R, Nickerson KW. Survey of extreme solvent tolerance in gram-positive cocci: membrane fatty acid changes in Staphylococcus haemo-lyticus grown in toluene. Appl Environ Micro-biol 2005; 71:5171-6.
28. Park JB, Buehler B, Panke S, Witholt B, Schmid A. Carbon metabolism and product inhibition determine the epoxidation efficiency of solvent tolerant Pseudomonas sp. strain VLB120DC. Biotechnol Bioeng 2007;98:1219-29.
29. Ramos JL, Duque E, Huertas MJ, Haidour A. Isolation and expansion of the catabolic potential of a Pseudomonas putida strain able to grow in the presence of high concentrations of aromatic hydrocarbons. J Bacteriol 1995;177: 3911-6.
30. Rojas A, Duque E, Schmid A, Hurtado A, Ramos JL, Segura A. Biotransformation in double-phase system: physiological responses of Pseudomonas putida DOT-T1E to a double phase made of aliphatic alcohols and bio-synthesis of substituted catechols. Appl Environ Microbiol 2004;70:3637-43.
31. Sameshima Y, Honda K, Kato J, Omasa T, Ohtake H. Expression of Rhodococcus opacus alkB genes in anhydrous organic solvents. J Biosci Bioeng 2008;106:199-203.
32. Sardessai Y, Bhosle S. Tolerance of bacteria to organic solvents. Res Microbiol 2002;153:263-8.
33. Schewe H, Kaup BA, Schrader J. Improvement of P450BM-3 whole-cell biocatalysis by integrating heterologous cofactor regeneration combining glucose facilitator and dehydro-genase in E. coli. Appl Microbiol Biotechnol 2008;78:55-65.
34. Schmid A, Hollman F, Park JB, Buhler B. The use of enzymes in the chemical industry in Europe. Curr Opinion Biotechnol 2002;13:359-66.
35. Schoemaker HE, Mink D, Wubbolts MG. Dispelling the myths-biocatalysis in industrial synthesis. Science 2003;299:1694-7.
36. Smolders AJJ, Pinheiro HM, Noronha P, Cabral JMS. Steroid bioconversion in a microemulsion system. Biotechnol Bioeng 1991;38:1210-7.
37. van Beilen JB, Holtackers R, Luscher D, Bauer U, Witholt B, Duetz WA. Biocatalytic produc-tion of perillyl alcohol from limonene by using a novel Mycobacterium sp. cytochrome P450 alkane hydroxylase expressed in Pseudomonas putida. Appl Environ Microbiol 2005;71:1737-44.
38. Vermue M, Sikkema J, Verheul A, Bakker R, Tramper J. Toxicity of homologous series of organic solvents for the Gram-positive bacteria Arthrobacter and Nocardia sp. and the Gram-negative bacteria Acinetobacter and Pseudo-monas sp. Biotechnol Bioeng 1993; 42:747-58.
39. Wahbi LP, Gokhale D, Minter S, Stephens GM. Construction and use of recombinant Escheri-chia coli strains for the synthesis of toluene-cis-glycol. Enz Microb Technol 1996;19:297-306.
40. Weber F, Ooijkaas L, Schemen R, Hartmans S, de Bont J. Adaptation of Pseudomonas putida S12 to high concentrations of styrene and other organic solvents. Appl Environ Microbiol 1993; 59:3502-4.
41. Wendhausen R, Frigato M, Fernandes P, Carvalho CCCR, Cruz A, Pinheiro HM et al. Chrysotile as a support for the immobilization

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
293
of Mycobacterium sp. NRRL B-3805 cells for the bioconversion of β-sitosterol in an organic-aqueous two-liquid phase system. J Mol Catalysis B: Enzymatic 2005;32:61-5.
42. Wery J, Mendes da Silva DI, de Bont JAM. A genetically modified solvent-tolerant bacterium for optimized production of a toxic fine chemical. Appl Microbiol Biotechnol 2000; 54:180-5.
43. Wierckx NJP, Ballerstedt H, de Bont JAM, Wery J. Engineering of solvent-tolerant Pseudo-monas putida S12 for bioproduction of phenol from glucose. Appl Environ Microbiol 2005; 71:8221-7.
44. Yamada M, Okada Y, Yoshida T, Nagasawa T. Biotransformation of isoeugenol to vanillin by Pseudomonas putida IE27 cells. Appl Micro-biol Biotechnol 2007;73:1025-30.
45. Yamashita S, Satoi M, Iwasa Y, Honda K, Sameshima Y, Omasa T et al. Utilization of hydrophobic bacterium Rhodococcus opacus B-4 as whole-cell catalyst in anhydrous organic solvents. Appl Microbiol Biotechnol 2007; 74:761-7.
46. Zahir Z, Seed KD, Dennis JJ. Isolation and characterization of novel organic solvent-tolerant bacteria. Extremophiles 2006;10:129-38.


295
Friction Stir Welding of High Strength Aluminium Alloy 2024
Manthana Jariyaboon Department of Chemistry, Faculty of Science, Mahidol University.
Abstract
Friction stir welding (FSW) is a relatively new solid-state joining technique that provides several advantages over conventional fusion welding such as being environmentally-friendly and energy and weight saving. In particular, this technique has been successfully used to join high strength aluminium alloys such as AA2024, which are difficult to weld with conventional fusion welding, to achieve excellent properties. In this article, the current state of understanding of microstructural evolution, hardness properties, corrosion behaviour and influence of welding parameters, in particular rotation and travel speeds for FSW AA2024 are discussed.
Keywords: friction stir welding; aluminium alloy; AA2024
Introduction
High strength 2XXX series aluminium alloys such as AA2024 are commonly used for airframe structures. They are regarded as difficult to weld using conventional fusion welding techniques such as TIG or MIG due to solidification cracking and grain boundary melting in the weld zone. Mechanical fastening is the current method used for joining aircraft structures. However, this technique is time-consuming, and suffers from a weight penalty, insufficient strength and corrosion problems.
Friction stir welding (FSW), a new solid-state joining technique invented and developed by The Welding Institute (TWI), UK in 1991(25), is a promising technique to overcome these problems and it is currently being used for joining high strength aluminium alloys. In the short period of time after the invention of FSW, it has become a very successful and sufficient solid-state joining technique for high strength aluminium alloys in real aircraft applications such as airframes of the Airbus A380 and lightweight space shuttle tanks. This is due to less generated heat, fewer microstructural changes and improved mechanical properties as well as corrosion behaviour relative to conventional fusion welding(27). The amount of generated heat, which can affect the micro-structural evolution, mechanical properties and
corrosion behaviour in the weld zone, is largely a function of welding parameters such as rotation speed and travel speed. The aim of this article is to address the current state of understanding of microstructure, hardness properties and corrosion behaviour of FSW in the high strength aluminium alloy AA2024. The effect of welding parameters (rotation speed and travel speed) on the microstructure and corrosion behaviour is also reviewed.
Friction Stir Weld Processing
FSW is produced by rotating and plunging a specially designed cylindrical and shouldered tool with a small diameter probe into the joint line, such as between two butted plates. The shoulder is pressed against the surface of the metal and the relative motion between the rotating tool and the metal generates frictional heat, which causes the metal to soften and allows the tool to traverse along the joint line. Friction stir welding is an asymmetrical technique, so the two sides of the weld are different. The “advancing side” of the weld is defined as that side of the weld on which the motion of the surface of tool owing to the tool rotation is in the same direction as the tool translation, while on the “retreating side” of the weld, the rotating motion of the tool is in the opposite direction to the tool translation(6,21). In addition, the heat input can be modified by

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
296
varying the welding parameters. For example, a higher rotation speed or a lower travel speed causes a higher heat input leading to a higher temperature associated with a longer period of time at an elevated temperature. In addition, the peak temperature also increases with an increase in applied vertical pressure(18,28).
Microstructure of FSW
The FSW process generates three unique microstructural zones: the nugget, the thermo-mechanically-affected zone (TMAZ) and the heat-affected zone (HAZ). The different zones in FSW are illustrated in Figure 1. The nugget is the region through which the toolpiece pin passes, and thus experiences high deformation and high heat. Generally, the nugget region consists of fine recrystallised equiaxed grains(12). The TMAZ adjacent to the nugget is the region where the metal is plastically deformed as well as heated, but this is not sufficient to cause recrystallisation, leading to grains that have been rotated along the TMAZ/nugget boundary. A more diffuse boundary is observed on the retreating side compared with that on the advancing side(20). The HAZ is the area located farther from the weld centre where the material experiences only a thermal cycle without plastic deformation. The HAZ region has a similar microstructure to the base metal. In addition to the main three zones described above, the FSW tool also creates a broader deformation zone with a shallow depth, which appears as the shoulder contact region. Dissolution and coarsening of strengthening precipitates as well as the formation of wide precipitate-free zones are found in the weld region.
Figure 1 Schematic of FSW zone(19).
Microstructure of FSW AA2024
The microstructure in the weld region of FSW AA2024 has been investigated by a number of authors(4,9-10,14,17,19,23-24,28) for butt welds produced with various rotation speeds (150-2400 rpm) and travel speeds (60-272 mm/min) as well as different plate thicknesses (1.5-25 mm).
Grain Structure in the Nugget Region
The nugget region contains fine recrys-tallised grains with sizes of 0.3-10 μm (1-3,5,16-
17,19,23-24,28) depending on the position in the nugget region and welding parameters. Across the thickness of the weld, there is a decrease in grain size in the nugget region from top to bottom due to a higher temperature near the top surface(3,19,24).
Sutton et al.(23) showed that the grain size in the nugget decreases when increasing the weld line energy. However, work done by Kristensen et al.(17) illustrated that even when the line energy is fixed, the grain size in the nugget can still decrease with lower rotation and lower travel speeds. It is likely that there is no relationship between the grain size in the nugget region and line energy. Comparing Sutton’s work and Kristensen’s work, there is one thing in common: the smallest grain size in the nugget region was observed in the weld produced with the lowest rotation and travel speeds. Thus, the grain size in the nugget region of weld is likely to be dependent on either the rotation speed or travel speed.
The investigation by Yang et al.(28)
showed that a slow travel speed weld gives a finer grain size in the nugget than a fast travel speed weld. However, the two welds were not produced at the same rotation speed (the slow weld was produced at a lower rotation speed), so there was still no conclusion as to whether rotation speed or travel speed controls the grain size of the weld in AA2024 in this study.
A systematic study on the effect of welding parameters on the grain structure in the nugget region was performed later by Jariyaboon and colleagues(14). They showed that the rotation speed has a primary role to
0.5 cm
Shoulder contact
Top surface
HAZ HAZ
(Thermo-mechanically affected zone)
(Heat affected zone)
Cross section
Nugget zone(Recrystallised
equiaxed grains)
HAZ zone(Elongated
grains)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
297
influence the grain size in the nugget region. At fixed travel speed, the higher rotation speed welds increase the grain size as a result of higher heat input while at fixed rotation speed, there is no significant change in grain size when changing the travel speed.
Particle Distribution and Hardness
Since friction stir welding is a thermo-mechanical process, the stirring action affects constituent intermetallic particles while the heat influences the precipitate particles. The temperature in the nugget region of FSW AA2024 is found to be typically in the range of 350-500oC(13). The temperature in the TMAZ and HAZ is lower than that in the nugget (e.g. 425oC in the TMAZ and below 350oC in the HAZ(13)). However, the temperature in each region can be varied depending on the welding parameters used.
Constituent Intermetallic Particles
The constituent intermetallic particles are found to be fragmented in the nugget region(1-
2,4,14,23,28) leading to an increase in particle number density(23,28). The average particle size in the nugget region is approximately 1-5 μm in diameter whereas outside the nugget region is approximately 7 μm(23,28). More fragmented constituent intermetallic particles can be observed when increasing rotation speed(14).
Hardness and Precipitate Particles
Nugget Region
Across the thickness of the weld in the nugget region, the maximum hardness is always seen at the top of the weld and the lowest hardness is at the bottom of the weld. Some authors(4-5) suggested that the recovery of strength in the nugget region is a result of the fine grains corresponding to the Hall-Petch relationship as illustrated in Equation 1:
σy = σo + kyd-1/2 Eq 1
where σy is the yield strength, σo and ky are constants for a particular material and d is the average grain diameter.
Considering the grain size distribution in the nugget region mentioned earlier and the
hardness behaviour in the nugget region across the thickness of weld in this section, the larger grains are found near the top surface where the hardness is higher compared with the bottom surface, where the grain size is smaller and the hardness is lower. Therefore, the reduction of grain size in the nugget region is unlikely to be the explanation for the variation in hardness.
It is well known that the hardness is strongly related to precipitate particles, so the hardness behaviour of weld is more likely to depend on the precipitate particles rather than on the grain size. According to TEM and DSC data(9,13), GPB zones and fine S′ precipitates are found in the nugget region. These precipitates form after welding by natural ageing and cause the recovery of hardness. This implies that the temperature in the nugget region is high enough to cause the partial dissolution of particles, which are then able to reprecipitate during natural ageing.
HAZ and TMAZ Regions
A high density of coarse S-phase precipitates at grain boundaries associated with precipitate-free zones (PFZ) as well as the precipitates within the grains are found in the minimum hardness region in the TMAZ(9) or HAZ (4,13). Thus, the minimum hardness occurs at the temperatures where precipitate particles grow until they lose their coherency.
A “W” pattern of hardness profiles across the FSW AA2024 can be observed in most published results, in which the lowest hardness is in either the TMAZ or HAZ region and a recovery in hardness occurs in the nugget region(4-5,9,13,17,28). However, Jariyaboon et al.(14) showed that the hardness profiles can have either a “W” shape or “U” shape depending on the heat input, which is controlled by the welding parameters, especially the rotation speed. For a low rotation speed weld, the lowest hardness area is in the nugget region where the coarse S-phase precipitate particles can be observed, whereas for the high rotation speed weld, the minimum hardness value is in the HAZ in association with a high density of the coarse S-precipitate particles.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
298
Corrosion of FSW AA2024 Studies of the corrosion behaviour of
FSW AA2024 have revealed that the weld zone is generally more reactive than the base metal(1-
2,4,7,10-11,26) in which the corrosion attack areas are found in the nugget, TMAZ or HAZ zone. There are only a few investigations showing either similar or better corrosion behaviour in the weld region compared with the base metal(8,22). The variation of preferential corrosion attack in the weld region as described above is likely to be due to the difference in welding parameters (rotation and travel speeds) as well as the thickness of base metal used to produce FSWs, which affects the heat distribution across the weld region differently in each work, leading to variation in the location of predominant corrosion attack. Thus, there was still no concrete explanation as to which factors affect the corrosion behaviour of welds. However, it was generally observed that one of the corrosion forms in the weld is intergranular corrosion.
Later work by Jariyaboon et al.(14)
provided a clear explanation of how the corrosion behaviour of FSW AA2024 can be related to microstructural changes, which are controlled by the welding parameters. They showed that rotation speed is a major parameter to control the corrosion behaviour of the weld. The welding parameters used in their work are shown in Figure 2. As seen from the gel visualisation test in Figure 2, the area most
susceptible to corrosion is the nugget when producing the weld with low rotation speed (215 rpm), whereas the main corrosion attack is in the HAZ region for the higher rotation speed (350-468 rpm) weld regardless of the travel speed.
The anodic reactivity is elevated in both the HAZ and nugget regions compared with the base metal for both low and high rotation speed welds, as shown by micro-electrochemical measurements (Figure 3). However, the higher anodic reactivity in the nugget region compared with that in the HAZ region can be observed in low rotation speed welds as illustrated in Figure 3(a), whereas for the higher rotation speed weld the highest anodic reactivity is in the HAZ region as shown in Figure 3(c). For the cathodic reactivity, there is an enhancement of cathodic reactivity in the nugget region for both low and high rotation speed welds as shown in Figures 3(b) and 3(d), respectively. The different locations of corrosion attack are due to the galvanic coupling between anodic and cathodic reactivities in the weld regions.
The maximum anodic reactivity in the nugget region of low rotation speed welds is a result of the presence of both grain boundary and matrix precipitation of S-phase in the nugget region as the suitable temperature (250-300oC) for S-phase precipitation is in this region as shown in Figure 4a. For the higher rotation
1 cm
A Rtoolpiece shoulder
nugget HAZ
top
bottom
Rotation speed (rpm)
Travel speed (mm)/min Note
215 154Low rotationspeed weld
215 75
350 95 Medium rotationspeed weld
468 154High rotation speed weld
468 75
Figure 2 Gel visualisation test on the transverse cross section of FSW AA2024-T351 with different Weldng parameters after 22 hours of exposure(14).

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
299
Figure 3 (a) Anodic and (b) cathodic polarisation curves in 0.1 M NaCl of FSW AA2024-T351 produced with rotation speed of 215 rpm and travel speed of 75 mm/min, (c) anodic and (d) cathodic polarisation curves in 0.1 M NaCl of FSW AA2024-T351 produced with rotation speed of 468 rpm and travel speed of 75 mm/min(14).
.
speed welds, a sufficient temperature for the growth of S-phase precipitates both in the grain boundary and matrix is found in the HAZ region where the highest anodic reactivity as well as the coarse grain boundary precipitation of S-phase are found as illustrated in Figure 4b. The grain boundary precipitation of S-phase will cause the precipitate-free zones adjacent to the grain boundary and this solute-depleted region acts as the anodic site, which leads to the intergranular corrosion attack as seen from the immersion test in Figure 5.
Figure 4 FEG-SEM (BSE mode) images in (a) nugget region of FSW AA2024-T351 produced with rotation speed of 215 rpm and travel speed of 75 mm/min and (b) HAZ region of FSW AA2024-T351 produced with rotation speed of 468 rpm and travel speed of 75 mm/min(14)
Figure 5 SEM (SE mode) images in (a) nugget region of FSW AA2024-T351 produced with rotation speed of 215 rpm and travel speed of 75 mm/min and (b) HAZ region of FSW AA2024-T351 produced with rotation speed of 468 rpm and travel speed of 75 mm/min after immersion test in 0.1 M NaCl for 24 h(14).
Conclusions
The microstructural changes in the weld region as a result of FSW processing affect both mechanical and corrosion properties. The generated frictional heat causes microstructural changes that can be controlled by welding parameters such as rotation speed and travel speed. The understanding of the relationship between heat distribution related to welding parameters and microstructural changes in the weld region will lead to predictions of mechanical and corrosion properties of FSW, which can then be used to produce tailor-made FSW structures and properties.
1 μm3 μm
(a) (b)
5 μm 50 μm
(a) (b)
(a) (b)
(c) (d)

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
300
Acknowledgements The author would like to thank the Royal Thai Government for funding and BAE SYSTEMS for technical support. The author would also like to acknowledge Trans Tech Publications and Elsevier for the permission of using the Figures.
References 1. Ambat R, Jariyaboon M, Davenport AJ,
Williams SW, Price D, Wescott A, Micro-electrochemical investigation of friction stir welds in aluminium aerospace alloy 2024. Proceeding of the 15th International Corrosion Congress; 2002 Sep 22-27; CD-ROM.
2. Ambat R, Jariyaboon M, Williams SW, Price Da, Wescott A, Davenport AJ, Corrosion protection of friction stir welds using laser surface melting. Proceeding of the International Symposium on Aluminium Surface Science and Technology; 2003 May 19-22; 258-63.
3. Benavides S, Li Y, Murr LE, Brown D, McClure JC. Low-temperature friction-stir welding of 2024 aluminum. Scripta Mater 1999;41:809-15.
4. Biallas G, Braun R, Dalle Donne C, Staniek G, Kaysser WA, Mechanical properties and corrosion behaviour of friction stir welded 2024-T3. Proceeding of the 1st International Symposium on Friction Stir Welding; 1999 Jun 14-16; CD-ROM.
5. Bussu G, Irving PE. The role of residual stress and heat affected zone properties on fatigue crack propagaion in friction stir welded 2024-T351 aluminium joints. Int J Fatigue 2003;25: 77-88.
6. Colligan K. Material flow behavior during friction stir welding of aluminum. Welding Research Supplement 1999 Jul;229s-38s.
7. Connolly BJ, Davenport AJ, Jariyaboon M, Padovani C, Ambat R, Williams SW et al, Localised corrosion of friction stir welds in aluminium alloys. Proceeding of the 5th International Friction Stir Welding Symposium; 2004 Sep 14-16; CD-ROM.
8. Corral J, Trillo EA, Li Y, Murr LE. Corrosion of friction-stir welded aluminum alloys 2024 and 2195. J Mater Sci Lett 2000;19:2117-22.
9. Genevois C, Deschamps A, Denquin A. Characterisation of the microstructural evolution during friction stir welding of aluminium alloys: a comparative study of 5251 and 2024 alloys. Proceeding of the 5th International Friction Stir Welding Symposium; 2004 Sep 14-16; CD-ROM.
10. Hannour F, Davenport A, Strangwood M. Corrosion of friction stir welds in high strength aluminium alloys. Proceeding of the 2nd International Symposium on Friction Stir Welding; 2000 Jun 27-28; CD-ROM.
11. Hannour F, Davenport A, Williams SW, Morgan PC, Figgures CC. Corrosion behaviour of laser treated friction stir welds in high strength aluminium alloys. Proceeding of the 3rd International Symposium on Friction Stir Welding; 2001 Sep 27-28; CD-ROM.
12. Hassan KAA , Prangnell PB, Norman AF, Price DA, Williams SW. Effect of welding parameters on nugget zone microstructure and properties in high strength aluminium alloy friction stir welds. Sci Technol Weld JOI 2003; 8:257-68.
13. Heurtier P, Jones MJ, Desrayaud C, Driver JH, Montheillet F. Thermomechanical conditions and resultant microstructures in friction stir welded 2024 aluminum. Mater Sci Forum 2003;426-432:2927-32.
14. Jariyaboon M, Davenport AJ, Ambat R, Connolly BJ, Williams SW, Price DA. The effect of welding parameters on the corrosion behaviour of friction stir welded AA2024-T351. Corros Sci 2007;49:877-909.
15. Jata KV, Semiatin SL. Continuous dynamic recrystallization during friction stir welding of high strength aluminum alloys. Scripta Mater 2000;43:743-9.
16. Karlsen M, Frigaard F, Hjelen J, Grong F, Norum H. SEM-EBSD characterisation of the deformation microstructure in friction stir welded 2024-T351 aluminium alloy. Mater Sci Forum 2003;426-432:2861-6.
17. Kristensen JK, Dalle Donne C, Ghidini T, Mononen JT, Norman A, Pietras A et al, Properties of friction stir weld joints in the aluminium alloys 2024, 5083, 6082/6060 and 7075. Proceeding of the 5th International Friction Stir Welding Symposium; 2004 Sep 14-16; CD-ROM.
18. Nandan R, Debroy T, Bhadeshia HKDH. Recent advances in friction-stir welding-process, weldment structure and properties. Prog Mater Sci 2008;53:980-1023.
19. Norman AF, Brough I, Prangnell PB. High resolution EBSD analysis of the grain structure in an AA2024 friction stir weld. Mater Sci Forum 2000;331-3:1713-8.
20. Paglia CS, Carroll MC, Pitts BC, Reynolds AP, Buchheit RG. Strength corrosion and environ-metally assisted cracking of a 7075-T6 friction stir weld. Mater Sci Forum 2002;396-402:1677-84.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลอง 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
301
21. Reynolds AP. Visualisation of material flow in autogenous friction stir welds. Sci Technol Weld JOI 2000; 5:120-4.
22. Squillace A, Fenzo AD, Giorleo G, Bellucci F. A comparison between FSW and TIG welding techniques: modifications of microstructure and pitting corrosion resistance in AA2024-T3 butt joints. J Mater Process Technol 2004;152:97-105.
23. Sutton MA, Reynolds AP, Yang B, Taylor R. Mode I fraction and microstructure for 2024-T3 friction stir welds. Mater Sci Eng A 2003;354:6-16.
24. Sutton MA, Yang B, Reynolds AP, Taylor R. Microstructural studies of friction stir welds in 2024-T3 aluminum. Mater Sci Eng A 2002;323: 160-6.
25. Thomas WM, Nicholas ED, Needham JC, Murch MG, Temple-Smith P, Dawes CJ. Friction Welding. United States Patent 1995; 5,460,317.
26. Williams S, Ambat R, Price D, Jariyaboon M, Davenport A, Wescott A. Laser treatment method for improvement of the corrosion resistance of friction stir welds. Mater Sci Forum 2003;426-32:2855-60.
27. Williams SW. Welding of airframes using friction stir. Air Space Eur 2001;3:64-6.
28. Yang B, Yan J, Sutton MA, Reynolds AP. Banded microstructure in AA2024-T351 and AA2524-T351 aluminum friction stir welds: part I metallurgical studies. Mater Sci Eng A 2004; 364:55-65.


303
Geometry and Mechanics of Carbon Nanostructures
Duangkamon Baowan Department of Mathematics, Faculty of Science, Mahidol University.
Abstract
This article underlines the importance of utilizing basic mathematical concepts to describe nanostructures, such as carbon nanotubes and C60 fullerenes. By employing classical geometry and elementary mechanical principles, mathematical modeling of nanoscale systems can be achieved. The chiral vector for carbon nanotubes is described in terms of fundamental vector analysis and the composition of polygonal rings on such carbon nanostructures is governed by Euler’s polyhedra theorem. Furthermore, the Lennard-Jones potential function for the non-bonded interaction energy between two molecules and the continuum approximation are examined to investigate the oscillatory behavior of double-walled carbon nanotubes. Finally, some further uses of mathematics in nanotechnology are mentioned briefly.
Keywords: mathematics; carbon nanotubes; C60 fullerenes; Euler’s polyhedra theorem; gigahertz oscillators
Introduction
Nanotechnology is multidisciplinary, drawing from a number of fields such as physics, chemistry, biology, materials science, and mechanical and electrical engineering. The underlying theme of nanotechnology is the control and assembly of matter at the atomic and molecular levels where physical properties are size dependent. The word “nano” is the prefix for 10-9, so one nanometer is one billionth of a meter. To put this scale in context, the radius of a human hair is around 100,000 nanometers and the size of a virus ranges from 15 to 600 nanometers. The word “technology” refers to the branch of knowledge that deals with device creation and implementation, such as in industrial arts, engineering, applied science, and pure science. Therefore, “nanotechnology” refers to the study of those small-scale objects that can be assembled to create novel devices. Examples of nano-technology in modern use are the present day manufacture of textiles, the design of computer chips, ingredients for cosmetics, and carbon fibers used in optical wires.
Worldwide nanotechnology is currently a major focus for scientific disciplines. Most research in this area deals with chemical, physical and biological issues or a combination of these areas, but to date very little work has been undertaken on mathematical modeling, which can significantly reduce the time taken in the trial-and-error processes leading to applications, and which in turn decreases the research cost. For example, as stated by Ferrari(10)
“Novel mathematical models are needed, in order to secure the full import of nanotechnology into oncology.”
Mathematical models are important not only for nanomedicine but in all areas of nano-technology, where it is important to fully comprehend phenomena that are often subtle and complex. In this article, some examples of utilizing basic mathematical principles and elementary mechanics are presented to model nanoscale systems.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
304
Geometry of Nanostructures
Carbon nanostructures including ful- lerenes, carbon nanotubes, nanocones and nanotori have received much attention because of their unique properties, such as their high flexibility and high thermal conductivity. In addition, carbon nanotubes presently constitute the strongest materials known (9). All the carbon nanostructures originate from the hexagonal graphene sheet, which may be rolled up to form a tube, or when adding pentagonal rings may be formed into a C60 fullerene, or twisted into a tube-like shape to form a torus. Accordingly, we first examine the hexagonal network. The discovery of carbon nanotubes in 1991 by Iijima(17) led to many scientific challenges for their adaptation to a wide variety of applications in nanotechnology. A single-walled carbon nanotube can be thought of as a graphene sheet that is rolled up to form a seamless cylinder. Multi-walled carbon nanotubes can be envisaged as multilayers of graphite sheets that are rolled up co-axially.
The helical represented by (n,m), where n and m are integers such that m ≤ n, and a1 and a2 are the basis structures of carbon nanotubes can be described by the chiral vector, C = na1 + ma2, usually vectors. Some parameters for the carbon nanotubes such as a circumference, diameter and chiral angle can be obtained directly from this chiral vector. To start, we determine the positions of vectors a1 and a2 in a Cartesian coordinate system. From Figure 1(a), the two vectors can be written as
where a0 is the length of the unit vector, and from experiments a0 is found as √3 ac-c = 2.46 Å. On substituting the two unit vectors into the equation for the chiral vector we obtain
0 00 0
33 1 3 1ˆ ˆ ˆ ˆ ˆ ˆ( ) ( )2 2 2 2 2 2
a ana ma n m n m⎛ ⎞ ⎛ ⎞
= + + − = + + −⎜ ⎟ ⎜ ⎟⎜ ⎟ ⎜ ⎟⎝ ⎠ ⎝ ⎠
C i j i j i j ,
Figure1 Diagram of (a) unit vectors and (b) the graphite plane of a graphene sheet that can be rolled up to form
a nanotube surface.
1 0 0
2 0 0
3 1ˆ ˆ ˆ ˆ(cos30 sin 30 ) ,2 2
3 1ˆ ˆ ˆ ˆ(cos30 sin 30 ) ,2 2
a a
a a
⎛ ⎞= + = +⎜ ⎟⎜ ⎟
⎝ ⎠⎛ ⎞
= − = −⎜ ⎟⎜ ⎟⎝ ⎠
a i j i j
a i j i j
o o
o o

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป คณะวทยาศาสตร มหาวทยาลยมหดล
305
so that the circumference of the tube is given by
2 2 2 2 2 2003( 2 ) ( 2 ) ( )
2C = + + + − + = + +
a n nm m n nm m a n nm m .
The radius of the tube is then determined from r = |C|/(2π). Moreover, the chiral angle α is also depicted in Figure 1(b). There are three types of carbon nanotubes: armchair, zigzag and chiral. The armchair is the structure that has n = m and α = 30° and the zigzag has m = 0 and α = 0°. A carbon nanotube which is neither armchair nor zigzag is termed chiral. The electronic structure of carbon nanotubes can also be determined by (n,m). If n-m is a multiple of 3, then the tube is metallic, otherwise, it is a semiconductor. Thus, for example, all armchair tubes are metallic.
The C60 fullerene consists of precisely 60 carbon atoms, and they are similar in structure to graphene, being composed of linked hexagonal and pentagonal rings such that the pentagonal rings are needed to close the surface into a spherical shape. The distance between two pentagons can also be determined from the chiral vector, which also identifies the symmetry group of the structure as well as the number of carbon atoms. We refer the reader to the work of Fujita et al.(11) for the full details of the honeycomb lattice of C60 fullerenes.
In terms of polygonal rings, all polygonal rings of carbon nanotubes are hexagons and in C60 fullerenes, a hexagonal lattice of any size or shape can only form a closed structure with exactly twelve pentagons, a result directly arising from Euler’s polyhedra theorem. Euler's polyhedra theorem, which can be applied to the topological structure of the molecule, states that
F + V – E = χ , (1)
where F, V and E denote the numbers of faces, vertices and edges for the given polyhedron and χ is the Euler characteristic. We note that any surface which is homeomorphic to a sphere has an Euler characteristic of 2, and further details of Euler's polyhedra theorem can be found in (7). The notation Pn is used to denote the number of n-gonal sides, e.g. P5 is the number of
pentagonal sides. Every atom is linked with three others in the sp2 structure, which is one of the hybridizations of carbon-carbon bonds; the details can be found in any first-year chemistry textbook. Thus, the three terms on the left-hand side of equation (1) can be expressed as
F = P4 + P5 + P6 + P7 + P8 + P9 + P10,
3V = 4P4 + 5P5 + 6P6 + 7P7 + 8P8 + 9P9 + 10P10,
2E = 4P4 + 5P5 + 6P6 + 7P7 + 8P8 + 9P9 + 10P10.
We note that one vertex is shared by three polygons and one edge is shared by two polygons. By substituting these three relations into (1) with χ = 2, Euler's polyhedra theorem simplifies to give
2P4 + P5 – P7 – 2P8 – 3P9 – 4P10 = 12. (2)
For example, a C60 fullerene is formed by pentagons and hexagons, and (2) implies that there are precisely twelve pentagons required to close the spherical shape, as illustrated in Figure 2. This is a result of the number of hexagonal sides P6 being an invariant of (2) and we note that there are always exactly twelve pentagons, irrespective of the number of atoms in the fullerene.
Mechanics of Nanostructures
Elementary mechanical principles and classical applied mathematical modeling techniques are employed to formulate explicit analytical criteria and ideal model behavior in nanotechnology for which previously were available only from experiments and molecular dynamics simulations. One aspect that has attracted much attention is the creation of nano-scale oscillators, the so-called gigahertz oscilla-tors. While there are difficulties for micro-mechanical oscillators, or resonators, to reach frequencies in the gigahertz range, because of their small size it is possible for nanomechanical systems to achieve such frequencies.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
306
Figure 2 There are twelve pentagons in the closed
structure of a C60 fullerene, where the dashed and solid lines represent bonds in the background and foreground, respectively.
First, we use the Lennard-Jones potential function for the non-bonded interaction potential energy between two atoms and the continuum approximation, which assumes that the interatomic interactions can be modeled by smearing the constituent atoms uniformly across surfaces, to investigate the mechanical properties of double-walled carbon nanotubes. Using this approach, we may determine analytic expressions for the van der Waals interaction energy, the suction energy and the oscillatory behavior.
The non-bonded interaction energy between two molecules can be obtained by summing the potential interaction for each atom pair
( )φ ρ= ∑∑ iji j
E , (3)
where φ (ρij) is a potential function for atom i on the first molecule and atom j on the second molecule a distance ρij apart. Following Girifalco et al.(12) and Hodak and Girifalco,(16) the continuum approach assumes that the atoms are uniformly distributed over the surfaces of the molecules, and its validity as compared to the discrete atom-atom model is discussed by Girifalco et al.,(12)
“from a physical point of view the discrete atom-atom model is not necessarily preferable to the continuum model. The
discrete model assumes that each atom is the center of a spherically symmetric electron distribution while the continuum model assumes that the electron distribution is uniform over the surface. Both of these assumptions are incorrect and a case can even be made that the continuum model is closer to reality than a set of discrete Lennard-Jones centers”.
Thus, instead of the double summation in (3), the interaction energy can be obtained by performing double surface integrals, averaged over the surface of each entity
1 2 1 2( )= ∫ ∫E dS dSηη φ ρ , (4)
where η1 and η2 are the mean atomic surface density of atoms on each molecule and ρ denotes the distance between two typical surface elements dS1 and dS2 on each molecule. In this study, the Lennard-Jones potential is employed, which is given by
6 12( )φ ρρ ρ
= − +A B ,
where A and B are the attractive and repulsive constants, respectively. The van der Waals interaction force between two typical atoms of two molecules is given by
= −∇vdWF E , (5)
where the energy E is defined by (4).
In terms of double-walled carbon nano-tubes, the interaction between the inner and outer tubes in the continuum approximation is obtained by averaging the atoms over the surface of each tube and integrating the Lennard-Jones function over two cylindrical surfaces. Figure 3 shows (a) the cross-sectional view and (b) the longitudinal view of a double-walled carbon nanotube. The distance between two surface elements is denoted by ρ and the distance between the centers in the longitudinal axis of the tubes is represented by Z. We refer readers to the work of Baowan and Hill(3) for the full derivation and the detailed mathematical expressions of the force distribution for double-walled carbon nanotubes.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป คณะวทยาศาสตร มหาวทยาลยมหดล
307
Figure 3 Illustrations of double-walled carbon nanotubes: (a) cross-sectional view and (b) longitudinal view.
The suction energy is defined as the total work performed by van der Waals interactions, given by (5), on a molecule entering a carbon nanotube. In certain cases, the van der Waals force becomes repulsive as the entering particle crosses the tube opening. In these cases, the acceptance energy is defined as the total work performed by van der Waals interactions on the particle entering the nanotube, up until the point that the van der Waals force once again becomes attractive. In Figure 4, we show an example for the acceptance condition of a particular molecule entering a carbon nanotube. It is rather like a roller-coaster; the molecule will be accepted if the interaction energy E inside the tube (positive Z) is less than the energy outside the tube. Once the inner tube is entered inside the outer tube, the oscillatory behavior is investigated.
Newton’s second law with any frictional effects neglected is used to describe the oscillatory behavior, to obtain
2
2
∂= −
∂d Z EMdt Z
,
where M is the mass of the inner tube. Following Cumings and Zettl(8), the inner tube is pulled out a distance d and then released. By using the equations of motion and for the particular case when the initial velocity is zero, the oscillation frequency can be written as
{ }2
2 1
,4 [2 ( )]
αα
=+ + −
dfd d L L
where α is the initial condition parameter given by α2 = |E(0)|/(ML1), E(0) is the potential energy at Z = 0, and L1 and L2 are half-lengths of the inner tube and the outer tube, respectively. The frequency of double-walled carbon nanotubes when the inner tube oscillates is in the gigahertz range, which cannot be reached by a micro-material. Furthermore, we observe that the longer the extrusion distance and the shorter the inner tube length, the higher the oscillatory frequency (3).
Other Mathematical Research in Nanotechnology
One of the most exciting applications of nanotechnology is the proposed use of nano-structures for drug delivery, and in particular the targeted delivery of drugs using nanotubes. Targeted drug delivery would have many significant implications for the future treatment of patients, particularly those suffering from cancer. Understanding the encapsulation and expulsion of drug molecules from nanocarriers
is vital for the development of nanoscale drug delivery. In particular, the suction and acceptance of the anticancer drug molecules cis platin, paclitaxel and doxorubicin into a carbon nanotube is examined by Hilder and Hill(13,15). They also investigate an alternative drug deli-very mechanism, namely a nano-syringe constructed from a double-walled carbon nanotube(14). In this case, rather than being directed to particular cells by chemical receptors attached to the nanotube surface, the nano-syringe pierces the cell membrane and injects molecules such as drugs or DNA directly into the cell. For each of these nanomedical applications specific nanotube radii are deter-mined for acceptance and maximum drug uptake, and some overall design guidelines are provided(13-15).
Cox and Hill(4) examine the conventional theory, which prescribes the geometric parameters of carbon nanotubes, and they find that it does not adequately account for the curvature inherent in the structure of nanotubes. By considering three postulates and then applying geometry and trigonometry, a poly-hedral model is proposed(4) and new formulae

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
308
Figure 4 Typical interaction energy, where a particular molecule is accepted into (----) and rejected from (____) nanotubes of two distinct radii.
are derived, which provide more precise expressions for geometric parameters such as the chiral angle, radius and unit length. Asymptotic expansions of the new expressions are also obtained to produce the conventional formulae as the leading order terms, which show that the polyhedral model is consistent with conventional theory for large values of (n, m).
Following this, Cox and Hill(5) and Baowan et al.(1) describe approximately toroidal molecules that are formed from two(5) and three(1) distinct carbon nanotubes, which have defects that cause a bend in an otherwise straight nanotube. The angle of the bend is determined using the variation in bond length method and then expressions are derived giving the dimensions for tori formed from a number of these bend defects. Representative formulae for the ideal toroidal parameters, the generating radius and tube radius, are determined from integral expressions for the circumference and surface area of the ideal surface. Furthermore, the perpendicular joining structures for carbon nanotubes and flat graphene is investigated. Two simple least squares approaches, which are the variation in bond length and the variation in bond angle, are employed by Baowan et al.(2) and the calculus of variations method is used by Cox and Hill(6). Such a combined structure
might constitute an electronic transmission platform for ultra-small computer chips.
Conclusions
Nanotechnology is currently a timely and diverse research area that ranges from the study of simple biological systems to the creation of novel nano-devices. Most research in this research area utilizes knowledge from physics, chemistry and biology, and very little mathematical modeling work has been undertaken. This article highlights the use of mathematics for this area, and primarily the application of geometry and mechanics to describe the behavior of both carbon nanotubes and C60 fullerenes. Our theoretical study provides simple mathematical models which give important insights into nanostructures, and which may also be exploited to guide future computational and experimental research.
Acknowledgements
The above summary is a synopsis of the work undertaken by both the author and her colleagues in the Nanomechanics Group at the University of Wollongong, Australia, and special thanks are due to Jim Hill, Ngamta Thamwattana, Barry Cox and Tamsyn Hilder, who are all participants in the work described. The Nanomechanics Group is funded entirely
Inside the tube
Outside the tube

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป คณะวทยาศาสตร มหาวทยาลยมหดล
309
by the Australian Research Council, and this financial support is gratefully acknowledged.
References 1. Baowan D, Cox BJ, Hill JM. Toroidal molecules
formed from three distinct carbon nanotubes. J. Math Chem 2007;44:515-27.
2. Baowan D, Cox BJ, Hill JM. Two least squares analyses of bond lengths and bond angles for the joining of carbon nanotubes to graphenes. Carbon 2007;45:2972-80.
3. Baowan D, Hill JM. Force distribution for double-walled carbon nanotubes and gigahertz oscillators. Z Angew Math Phys 2007;58:857-75.
4. Cox BJ, Hill JM. Exact and approximate geometric parameters for carbon nanotubes incorporating curvature. Carbon 2007;45:1453-62.
5. Cox BJ, Hill JM. New carbon molecules in the form of elbow-connected nanotori. J Phys Chem 2007;111:10855-60.
6. Cox BJ, Hill JM. A variational approach to the perpendicular joining of nanotubes to plane sheets. J Phys A: Math Theor 2008;41:125203.
7. Cromwell PR. Polyhedra. 1st ed. Cambridge: University Press; 1997.
8. Cumings J, Zettl A. Low-friction nanoscale linear bearing realized from multiwall carbon nanotubes. Science 2000;289:602-4.
9. Dresselhaus MS, Dresselhaus G, Eklund PC. Science of fullerenes and carbon nanotubes. 1st ed. California: Academic Press; 1995.
10. Ferrari M. Cancer nanotechnology: opportunities and challenges. Nat Rev Cancer 2005;5:161-71.
11. Fujita M, Saito S, Dresselhaus G, Dresselhaus MS. Formation of general fullerenes by their projection on a honeycomb lattice. Phys Rev B 1992;45:13834-6.
12. Girifalco LA, Hodak M, Lee RS. Carbon nano-tubes, buckyballs, ropes, and a universal graphitic potential. Phys Rev B 2000;62:13104-10.
13. Hilder TA, Hill JM. Carbon nanotubes as drug delivery nanocapsules. Curr Appl Phys 2008;8: 258-61.
14. Hilder TA, Hill JM. Modeling loading and unloading of drugs into nanotubes. Small 2008;5: 300-8.
15. Hilder TA, Hill JM. Modelling the encapsulation of the anti-cancer drug cisplatin into carbon nano-tubes. Nanotechnology 2007;18:275704.
16. Hodak M, Girifalco LA. Fullerenes inside carbon nanotubes and multi-walled carbon nanotubes: Optimum and maximum sizes. Chem Phys Lett 2001;350:405-11.
17. Iijima S. Helical microtubules of graphitic carbon. Nature 1991;354:56-8.


311
A Survey of Approaches to Digital Image Matting
Rawesak Tanawongsuwan Department of Computer Science, Faculty of Science, Mahidol University.
Abstract
Image matting is the operation concerned with extracting a foreground object from an image. The extracted foreground image, which is referred to as a matte, is used in image and video editing applications. In the current world of digital photography and digital movie special effects, compositing tasks often need mattes to combine two or more images or layers together into a final image. With the need for high-quality mattes in digital multimedia applications, image matting research has received much attention in the past decade. This paper provides an introductory survey to image matting, a review of methods in image-based and video-based matting proposed in the past few years, and opportunities and challenges in the image matting research.
Keywords: image matting; video matting; alpha channel; blue screen matte; trimap
Introduction
The extracted foreground image which is referred to as a matte is used in image and video editing applications. In the current world of digital photography and digital movie special effects, compositing tasks often need mattes to combine two or more images or layers together into a final image. The goals of image matting are to calculate the foreground and background color components along with the opacity values (alpha values). Figure 1 shows a typical step in the digital image composition task.
In the typical image matting framework, an image with a desired foreground object is given as an input. Normally we work with a color image. Therefore, each pixel contains a color component . The color in each pixel in the input image is modeled as a linear combination of a foreground color and a background color and is the alpha value in the range of [0, 1]. The typical matting equation can be written as
1 (1)
(a) (b) (c)
(d) (e)
Figure 1 (a) An input image. (b) An extracted matte. (c) A color version of the matte. (d) A new background image. (e) A new composite.

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
312
The matting problem is to separate a given input image into a foreground object image, a background image, and an alpha matte. In a normal situation, we will have a 3-channel color image (e.g. RGB color components). So for each pixel, we have 7 unknowns to recover (3 for , 3 for , and 1 for ) from 3 known values ( )(8). This problem is an under-constrained problem. Therefore, there were many methods proposed in the past several years to try to estimate these unknowns.
Image matting and image segmentation are related topics in the image processing, computer vision, and computer graphics research communities. Image segmentation is a low-level operation to partition images into disjoint regions. Each region will be given assigned a unique label. In the case of segmenting a foreground object from an image, each pixel will be assigned either 0 or 1. However, if the foreground object that we want to segment contains regions with small fine details or transparent materials, the pixels in those regions might contain both foreground and background parts. In this case, we cannot assign unique labels to them. Using equation (1), the image matting problem will assume an alpha value between 0 and 1; however, the image segmentation will assume an alpha value of either 0 or 1.
To obtain reasonable mattes, many techniques can pull the mattes directly from normal photographs taken in natural scenes (natural image matting) using image processing and computer vision techniques, assume a known background or environment that can be controlled (i.e. blue screen matting), use multiple input images, or even use multiple cameras. In real-life situations, extracting fore-ground objects can be quite tricky. Material properties (transparency, translucency, specu-larity, etc.) of the objects can play important roles in the outcome of the mattes. Occlusion and shadow effects commonly occur in real photographs. The colors of the foreground object may be similar to the colors of the background scene. All these example situations are challenges to matting and compositing research and at the same time are considered
good research opportunities to advance the state-of-the-art of image matting.
This paper is organized to 1) describe several interesting and challenging foreground objects for the matting problem, 2) explain the levels of user interaction that the users have to provide to the existing matting algorithms, and 3) provide a brief summary of the current state-of-the-art matting techniques.
Foreground Object
In digital image matting, desired fore-ground objects can have complex shapes, materials, color components, etc. Each type of foreground object may require different methods to extract a reasonable matte. In this section, we will consider various cases that illustrate the difficulty of some matting tasks. Figure 2 shows examples of typical photographs taken in natural settings. Matting of this type, for relatively arbitrary photographs, is known as natural image matting(3). Figure 3 shows foreground objects that need to be recovered not only for their color components, but also for their material properties. Figure 3(a) shows a non-trivial task to extract the shape of the bottles along with their transparent and refractive properties. Figure 3(b) illustrates a problem that occurs if we simply pull the foreground object with the reflective material and composite it to a new background image. The reflections of oranges will also be on the pot’s surface in the new composite. Figure 4 shows that sometimes a user might want to extract not only the foreground object, but also its shadow. However, overlaying the extracted foreground object and its shadow on a new background scene may be problematic when the shadow has to be in front of another object. The shape of the shadow must follow the background geometry in order to obtain a realistic composite. Figure 5 shows quite difficult cases in which the colors of the foreground objects are similar to the colors of the background scenes and their shapes are quite complex for even to the human eye to distinguish.
Levels of User Interaction
Since the matting problem is closely re-

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
313
lated to the segmentation problem and is considered an under-constrained problem, we should not expect to easily implement an automatic matting system. It is reasonable to expect some levels of user interaction to provide some initial constraints to the problem. One simple way to select a region in an image is to use the rectangular selection tool as shown in Figure 6(a). This selection mode can be thought of as generating another small image that may reduce some ambiguities between foreground and background. The “GrabCut”(12) system has shown some attempt to offer this simple selection feature and perform some initial “hard” segmentation and then followed by a border refinement procedure to get a reasonable matte. This work has shown that in many cases, the user needs to provide some additional hints to the system. Another popular user interaction in the current matting systems is to let the user draw a trimap to pre-define three separate regions in the image: background, foreground, and unknown. Figures 6(b)-(c) show an exam-ple of a trimap drawn on the input image using three different colors. An early Bayesian matting algorithm(3) has shown impressive results based on the user-supplied trimap. However, the manual creation of a trimap proves to be a quite tedious and time-consuming process. Thus, good trimap generation is subsequently becoming another important research topic. One of the state-of-the-art image matting algorithms(6,8) uses stroke-based inter- faces to let the users roughly indicate fore-ground and background regions using a few simple strokes. The current results in the stroke-based matting systems are quite impressive. However, the results depend also on the locations of the strokes. Users may have to refine their strokes to obtain the results that they want. Recently, the Soft Scissors system was introduced as an interactive tool for extracting high quality mattes(18). This system lets the user draw along the foreground boundary and shows the matte and the final composite in real-time.
Matte Extraction
In this section, we will present a survey of matting techniques and classify them into dif-
(a) (b)
Figure 2 (a)-(b) Photographs taken in natural settings where the challenges are in the extraction of hair.
(a) (b)
Figure 3 (a) Extracted foreground objects should contain not only their color components, but also their transparency and refraction. (b) The extracted stainless pot has specular material property, which needs to be recovered along with its color.
(a) (b)
Figure 4 (a) An image of a thermos cup taken on a sunny day with its shadow. (b) The shape of the shadow changes when it falls on to another 3D object.
(a) (b)
Figure 5 (a)-(b) A foreground tree in front of a background scene with similar color components.

31
วทยาศาสตรเพ
14
Figure 6
ferent scments.
B
Btechniquobject assumptitake a pbackgroucolors inremove color anmatte. Ibackgroucolors fOther cocertain foregrouimage us
E
FitranspareExtractineven modified topaths fforegrourefractionique byrecover ttwo pictubackdroptranspareapply thobject (scalled “
พอการพฒนา: รวมบท
(a)
6 Different levimage into tto roughly in
chemes base
Blue Screen M
Blue screen ue to extrac
from the ion of the ticture of theund color wn the foregrpixels that h
nd are left wiIt is conveund color isfor this purolors can bthat they
und object. sed in blue sc
Environment
igure 3 shoent, refractivng a good more difficult. o take into from the und object von. The “Bly Smith and the transpareures using twps. Howevency properthis techniqusuch as a hu
“Environmen
ทความวชาการ ฉลอง
vels of user ithree differentndicate foregr
ed on their s
Matting
matting ct the desi
backgroundtechnique is e object in frwhich is difround objechave the knoith the desir
enient to ass a constant rpose are be used as l
are differFigure 7 shocreen mattin
t Matting
ows examplve, and reflec
matte with the Equation (account thebackground
via the refllue Screen Blinn(14) offeency of the owo different er, it can oties and it is
ue to a movuman). A b
nt Matting”(2
งครบรอบ 50 ป แหง
(b)
interaction. (at regions: bacround and bac
specific requ
is a popred foregrod. The m
that if we ront of a knofferent from ct, then we own backgrorable foregrossume that color. Pop
blue and grelong as we rent from ows an exam
ng.
le objects wctive propertese propertie(1) must be me light-transpd through lection and Matting” te
fers a solutioobject by takbackgroundnly recover cumbersom
ving foregrobetter techni2,21) is propo
การสถาปนาคณะวท
a) Typical selckground, forekground regio
uire-
ular ound
major can
own the can
ound ound
the ular een. are our
mple
with ties. es is mo-port the the
ech-n to king
ds or the
me to ound ique osed
to betwaccof iin ThereflTo takmobacobtproof tobtbe of t
extrTheestiitsesitin
ทยาศาสตร มหาวทยาล
(c)
lection modeeground, and uons.
model the ween the ba
curately. Thimages of thefront of knese images alected and rget accurat
en in a cnitors are
ckdrop and ttain high-quaoperties. Thethe foregrountain, and it mrather smallthe backdrop
Figur
Shadow
In Figurracting the ere are at limate the celf. The secong task. W
ลยมหดล
. (b)-(c) A trunknown. (d)
complex ackground ais technique e desired forown variablare used to crefracted froe results, montrolled enset up arothe sidedropality mattes wey can be usnd object thamust be statil (a limitatiop).
re 7 Blue scre
Matting
re 4, we sshadow maeast two diolor of the ond one deaWhen we w
(d)
rimap that seg) Stroke-based
lighting intand foregrou
relies on a reground objle-pattern bacalculate howom the envi
many images nvironment und objects
p). These tewith realisticsed to preparat the users cic and its sizon of size av
een matting.
ee the diffiatte from anifficulties. O
shadow coals with thewant to ove
gments an d interface
teractions und more sequence ect taken
ackdrops. w light is ironment.
must be (usually
s as the echniques c material re mattes
can easily ze should vailability
ficulty of n image.
One is to omponent e compo-erlay the

extractedshadow resultingphysicaloverlaid composithe newproposedalgorithmwould bscene. shadow take a sebackgrouobject (uwarping displacemacceptabscene. task to oarbitrarytric infor
a
d foregrounonto a ne
g composite ly correct on top of a
ite relies on w background a shadowm where thbehave in th
The techniqscanning ste
equence of pund with a cusually a lon
function froment map.
ble if we canIt still rema
overlay the sy backgroundrmation.
arg max , ,
วทยาศา
d object anew backgrou
will not loif the sha
another objecthe underlyi
nd scene. Cw matting anhey learn hohe new desirque requiresep. They bpictures of thcasted shadowng stick). Thom these im
This techn access the nains a difficshadow mattd image with
, ,
าสตรเพอการพฒนา:
nd its extracund scene, ook realisticadow must ct. The shading geometryChuang et and composiow the shadred backgros the geomeasically havheir new desw of the knohey compute
mages to creahnique is onew backgroult composite on top of hout its geo
| arg m
(a) (b) (c)
รวมบทความวชาการ
cted the
c or be
dow y of al.(5) iting dow
ound etric e to ired own
e the ate a only ound iting any
ome-
phoTheverAt shomatpreregThethe for obsforethe attaby bes(MAtrieby
max , ,
pixthe solufromregmamatthenthe Fig
ร ฉลองครบรอบ 50 ป
Natural I
Natural otographs is e effective sry useful to d
least two ow impressitting. Both
e-segment angions: backgre goal is to cbackgroundeach pixel
served color eground andsurrounding
acks the undusing a Bay
st solution uAP) estimat
es to find th
| , ,
Figure 8xels used in
unknown ution is comm the know
gions. Anothtting is introtte from then reconstrucPoisson equ
ure 8 (a)-(b)and itscirclesthe oppixels
ป แหงการสถาปนาค
Image Matt
image maa more gene
solutions to digital imageearlier propive results techniques u
n input imaground, foregrcompute the
d color , anin the unkn
. The basd backgroung pixels. Thder-constrainyesian frameusing the mtion. The te optimal
8 illustratesthe calculatregions. T
mputed usingwn foregrouher techniq
oduced to come image withct the matte uations (15).
) A zoom-in s correspondins show the piptimal , , ain the unknow
ณะวทยาศาสตร มหา
ting
atting of eral matting this problem
e editing appposed techn
for naturause the trimage into three round, and u
e foreground nd the alphanown regionsic ideas ared distributio
he Bayesian mned matting ework and fi
maximum a ptechnique es, , and ,
s the neightion of two This way, thg the color
und and bacque called mpute the grh a given tri
solution by
portion of Fing trimap. (c)ixels used toand valueswn region.
าวทยาลยมหดล
3
arbitrary problem.
m can be lications. iques(3,13)
al image ap idea to
separate unknown.
color , a value n with its e to build ons using matting(3) problem
inding its posteriori ssentially given ,
(2)
hborhood pixels in he matte statistics
ckground Poisson
radient of imap and y solving
igure 1(a) ) Two red calculate s for two
15

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
316
The creation of a reasonable trimap is a time-consuming process. Also, the general matting problem is difficult to solve, especially when the background and foreground color distributions are similar. The GrabCut system(12) therefore attacks an easier problem by letting the user select the region of interest with a rectangular selection tool [similar to that in Figure 6(a)] and trying to compute a hard segmentation and followed by the border matting step.
Recently, more advanced algorithms have been proposed with a simpler user interaction. Users can now draw several strokes to indicate the foreground and background regions, Figure 6(d). The work based on the belief propagation technique(17) tries to estimate the alpha value for every pixel in the input image with user-drawn strokes. This method has proven to work well with images with significant semi-transparent foreground regions. However, due to the nature of its iterative non-linear optimization process, this technique is quite slow and may get stuck in a local minimum. A closed-form solution for the natural image matting is introduced in(8). Based on local smoothness assumptions for the foreground and background colors and , it is shown that one can analytically eliminate and
from a derived cost function leaving a quadratic cost function in . Therefore a global solution of this cost function can then be solved with a sparse linear system of equations. Subsequent work on Spectral Matting(9) extends ideas from spectral analysis for the hard
segmentation task to the extraction of soft matting components. This technique uses the eigen analysis of the matting Laplacian to automatically pull out the fore-ground matte by representing it in terms of fundamental matting components. Recent faster closed form optimization techniques called Easy Matting and Robust Matting(6,19) have been shown to outperform previous methods in terms of quality of mattes and performance. The Soft Scissors system(18) is the latest interactive matting tool that allows users to extract mattes as they paint or trace along the foreground edge.
Multiple-Image and Video Matting
Flash Matting
Extracting a good foreground from an input image is still a difficult problem to solve, especially when the foreground and background pixels have similar colors. In the Flash Matting technique(16), a high-quality matte of a (static) scene can be obtained by taking two pictures of the same scene, with flash and without flash (Figure 9), if we assume that the alpha value at the same pixel location of both pictures are the same. Then we have , , , , values to estimate from the observed and , where the superscript represents values from the flash version. Furthermore, it assumes that and are the same (the flash has a small effect on the intensity change of the background). The Bayesian matting is then extended to the so-called Joint Bayesian flash matting:
arg max , , , , , , | , arg max , , , | , , | ,
(3)
Figure 9 (a) An image taken without flash. (b) The same scene taken with flash.

Tmattes techniquflash phwith, suinstead. only for
V
Seinformatof the scThe deptinput imand the focus. Tcapturescamera amatting(
can captbut varytechniquthe caseshapes.
Figure 10
he techniquethat are ha
ues. Also thhotography uch as flash
However, static object
Variable-Foc
everal recenttion availablcene taken wth-of-field-b
mages: one wother one w
The variabl the scene maperture (Fig10) relies on ture images in focus, ex
ues yield reae of static
0 The first acalculated
วทยาศา
e produces mard to obtaere are severthat can behing only tthis techniq
ts.
cus-Based M
t matting tecle from two with a differased matting
where the objwhere the bale-aperture-b
multiple timesgure 10). Thspecially buthat share a
xposure, and asonable ma
objects an
and second rtrimap and th
าสตรเพอการพฒนา:
many impresain from oral variatione experimenthe backgroque works w
Matting
chniques expor more ima
rent focus(1,1
g(11) requires bject is in foackground i
based mattins by varying
he defocus viuilt cameras an optical ceaperture. Thatte qualitiesd non-comp
rows show imhe new compo
รวมบทความวชาการ
sive ther
ns of nted
ound well
lore ages 0,11).
two cus, s in ng(1)
g the ideo that
enter hese s in plex
Vid
duevidextrtechtechflowkeycancapthe usebotacc
arravidobjcamangobjbac
mages taken site.
ร ฉลองครบรอบ 50 ป
deo Matting
Video me to the spdeos. There hract movinhnique in(4)
hnique to uw trimaps yframes. Then also perforpture multipl
same opticed along witth types of curate video m
Recentlyay has pro
deo matting rects. The b
meras captugles(7) so thects in fro
ckground sce
with variable
ป แหงการสถาปนาค
g
matting is evpatial and thave been s
ng mattes extends th
use the opticbetween
e defocus vidrm video mae images fro
cal center. th the norm
informationmatting of th
y, a system duced veryresults on bobasic idea iure the scehat we can ont of diffene as shown
e aperture. Th
ณะวทยาศาสตร มหา
ven more chatemporal naseveral technfrom vide
he Bayesian cal flow mehand-drawn
deo matting satting becau
om cameras tThe depth
mal camera(20
n to obtain he foreground
based on ay impressiveoth static andis to have ene from
capture foferent parts n in Figure 11
he last row s
าวทยาลยมหดล
3
allenging atures of niques to os. The
matting ethods to n trimap system(10)
use it can that share sensor is
0) to fuse a more
d object.
a camera e natural d moving
multiple different
oreground of the
1.
shows the
17

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
318
Figure 11 This sequence of images was taken from different camera locations. The plant in the back is occluded by the one in front. Their colors are very similar. With images from different viewpoints, one can hope to recover an accurate matte of the foreground plant.
Conclusions
In this paper, an introductory survey of approaches in digital image matting and a review of proposed methods of image-based and video-based matting are presented. There are still plenty of opportunities and challenges in image matting research. Different types of foreground objects might require different types of matting algorithms. The close relationships between image segmentation and matting are worth exploring more as we can see from the important theoretical research on spectral matting(9). Using more than one image seems to be quite practical for extracting natural objects. Since there has been a lot of progress on natural image matting of a single image, one might imagine extending these state-of-the-art matting algo-rithms to multiple-image-based and video matting. Finally, with the advancement of computer hardware, parallel computation can be explored to create more interactive matting tools that have a real-time performance.
References 1. Boonsom N. Variable-aperture-based image
matting. Master’s Thesis, Computer Science, Mahidol University; 2008.
2. Chuang Y, Agarwala A, Curless B, Salesin DH, Szeliski, R. Video matting of complex scenes. Proceedings of the 29th Annual Conference on Computer Graphics and Interactive Techniques; 2002 July 23-26; New York, San Antonio: ACM; 2002. p.243-8.
3. Chuang YY, Curless B, Salesin DH, Szeliski R. A Bayesian approach to digital matting. Proceedings of the IEEE Computer Vision and Pattern Recognition, CVPR; 2001 December 9-14; Kauai Marriott, Hawaii: IEEE Computer Society; 2001. vol. 2, p. 264-71.
4. Chuang Y, Goldman DB, Curless B, Salesin DH, Szeliski R. Shadow matting and compositing. In ACM Transactions on Graphics. Proceedings of ACM SIGGRAPH; 2003 July 27-31; San Diego; 2003. vol. 22, p. 494-500.
5. Chuang Y, Zongker DE, Hindorff J, Curless B, Salesin DH, Szeliski R. Environment matting extensions: towards higher accuracy and real-time capture. Proceedings of ACM SIGGRAPH 2000, ACM Press/ACM SIGGRAPH, Computer Graphics Proceedings, Annual Conference Series. New York: ACM Press/Addison-Wesley Publishing Co; 2000. p. 121-130.
6. Guan Y, Chen W, Liang X, Ding Z, Peng Q. Easy matting: A stroke based approach for continuous image matting. Proceeding of Eurographics; Vienna; 2006. p. 567-76.
7. Joshi N, Matusik W, Avidan S. Natural video matting using camera arrays. Proceedings of ACM SIGGRAPH; 2006 July 30-August 03; Boston; 2006. p 779-86.
8. Levin A, Lischinski D, Weiss Y. A Closed-form solution to natural image matting. IEEE Transactions on Pattern Analysis and Machine Intelligence 2008:30;228-42.
9. Levin A, Rav-Acha A, Lischinski D. Spectral Matting. IEEE Transactions on Pattern Analysis and Machine Intelligence 2008:30;1699-1712.
10. McGuire M, Matusik W, Pfister H, Hughes JF, Durand F. Defocus video matting. Proceedings

วทยาศาสตรเพอการพฒนา: รวมบทความวชาการ ฉลองครบรอบ 50 ป แหงการสถาปนาคณะวทยาศาสตร มหาวทยาลยมหดล
319
of ACM SIGGRAP; 2004 July 31-August 4; Los Angeles; 2005. p. 567-76.
11. Reinhard E, Khan E. A. Depth-of-field-based alpha-matte extraction. Proceedings of the 2nd Symposium on Applied Perception in Graphics and Visualization; 2005 August 26-28; A Coroña, Spain; 2005. vol. 95, 95-102.
12. Rother C, Kolmogorov V, Blake A. GrabCut: interactive foreground extraction using iterated graph cuts. In ACM SIGGRAPH 2004 Papers. SIGGRAPH '04; 2004 August 8-12; Los Angeles; 2004. p. 309-14.
13. Ruzon MA, Tomasi C. Alpha estimation in natural images. Proceedings of the IEEE Computer Vision and Pattern Recognition, (CVPR'00); 2000 June 13-15; Hilton Head, South Carolina; 2000.
14. Smith AR, Blinn JF. Blue screen matting. Proceedings of the 23rd Annual Conference on Computer Graphics and interactive Techniques SIGGRAPH '96; New York, ACM; 1996. p. 259-68.
15. Sun J, Jia J, Tang C, Shum, H. Poisson matting. ACM Transactions on Graphics 2004:23;315-21.
16. Sun J, Li Y, Kang SB, Shum H. Flash matting. In ACM SIGGRAPH 2006 Papers. SIGGRAPH '06; 2006 July 30-August 03; New York, Boston: ACM; 2006. p. 772-8.
17. Wang J, Agrawala M, Cohen MF. Soft scissors: an interactive tool for realtime high quality matting. ACM SIGGRAPH 2007 Papers. SIGGRAPH '07. 2007 August 5-9; New York, San Diego: ACM; 2007.
18. Wang J, Cohen MF. An Iterative optimization approach for unified image segmentation and matting. Proceedings of the 10th IEEE inter-national Conference on Computer Vision; 2005 October 17- 20; Beijing, China., Washington DC: IEEE Computer Society; 2005. vol. 2, p. 936-43.
19. Wang, J. and Cohen, M. F. Optimized color sampling for robust matting. Proceedings of IEEE Computer Vision and Pattern Recognition, CVPR07, Minneapolis; 2007. p. 1-8.
20. Wang O, Finger J, Yang Q, Davis J, Yang R. Automatic natural video matting with depth. Proceedings of the 15th Pacific Conference on Computer Graphics and Applications. PG07; 2007 October 29-November 2; Washington DC, IEEE Computer Society; 2007. p. 469-72.
21. Zongker DE, Werner DM, Curless B, Salesin DH. Environment matting and compositing. Proceedings of the 26th Annual Conference on Computer Graphics and interactive Techniques.
New York, Los Angeles: ACM Press/Addison-Wesley Publishing; 1999. p. 205-14;


321
KEYWORD INDEX
AA2024, 295 ABC transporter, 81 acetamidase promoter,139 acrosome reaction, 25 adamantanes, 119 adverse effect, 49 Ae. Aegypti, 187 Ae. Albopictus, 187 AFM, 233 alpha channel, 311 alternative energy, 225 aluminium alloy, 295 antigenic variation, 99 antigens, 33 antioxidant(s), 49,153 antivirals, 119 apoptosis, 247 atomic force microscopy, 233 ATP-binding cassette, 81 avian influenza, 119 band gap, 147 bilayer, 279 biodiversity, 163 biomarker, 81 biomolecule, 233 biopreservative, 153 biotransformation, 287 blue screen matte, 333 C60 fullerenes, 303 calcium handling, 73 carbon nanotubes, 303 cardiac hypertrophy, 73 carotenoid, 49 catalase, 49 cell cycle, 247 cell engineering, 247 chemical biology, 7 cholesterol, 1 community, 195 conservation ecology, 195 cosmetics, 153 COX-2, 67
cytokines, 67 demand patterns, 105 density functional theory, 147 DFT, 147 DNA technology, 257 dopamine, 67 drug resistance, 119 drug screening, 139 dyes, 225 dye-sensitized solar cells, 225 electroluminescence organic materials, 215 endangered species, 195 endocrine, 41 Ensete, 163 estrogen, 73 ethanol, 125 Euler’s polyhedra theorem, 303 extracellular matrix , 73 F. gigantic, 33 F. hepatica, 33 fasciolosis, 33 fertilization, 25 flat panel display, 215 forensic science, 17, 257 free radicals, 49 friction stir welding, 295 fungal plant pathogen, 179 fungi, 179 gasohol, 125 gene regulation, 99 gene tagging, 171 genetic engineering, 171 genetic tools, 89 GFP, 139 gigahertz oscillators, 303 glutathione peroxidase, 49 gold, 265 heavy metal, 207 hormone, 41 host-selective toxin, 179 human DNA, 257

322
image matting, 311 immunity, 41 inflammation, 67 influenza, 119 inventory management, 105 irritant contact dermatitis, 113 lateral mobility, 279 leptospira, 59 leptospirosis, 89 level set methods, 241 loop-mediated isothermal amplification (LAMP), 131 LPS, 67 Luciferase, 139 Macaca fascicularis, 1 malaria, 99, 131 mango seed kernel, 153 mathematics, 303 mean-filed and pair approximation, 201 microglia, 67 molecular marker assisted breeding, 171 multidrug resistance, 81 Musa, 163 Musella, 163 mycobacteria, 139 myofilament activation, 73 nanomaterial, 265 nanoparticle, 265 natural product chemistry, 7 neuraminidase inhibitors, 119 neurodegeneration, 67 neurons, 67 non-human DNA, 257 OLED, 215 oligomers, 147 on-site analysis, 125 oogenesis, 25 order quantity, 105 organic light emitting devices, 215 organic-aqueous two-liquid phase system, 287 oxidative stress, 1, 67 oxygen-barrier, 271 packaging material, 271 Parkinson, 67 PECVD, 271
Penaeus monodon, 25 physiological response, 207 phytoremediation, 207 phytotoxicity, 207 plant biotechnology, 171 plant development, 207 plant tissue culture, 171 plasma deposition, 271 plasma enhanced chemical vapor deposition, 271 polymer, 265 population dynamics, 201 recombinant proteins , 247 reorder point, 105 reporter gene, 139 rRNA switching, 99 rubber tree, 171 SCFS, 17 science, 195 single-molecule recognition, 233 SiOx film, 271 SNpc, 67 social science, 195 solar cell, 225 spatially explicit model, 201 spatially implicit model, 201 spermatogenesis, 25 stress, 41 superoxide dismutase, 49 supported planar lipid membrane, 279 synthesis, 265 test kit, 125 testosterone, 73 thalassemia, 1 topical corticosteroid, 113 toxin, 179 transmission, 59 trimap, 311 vaccine development, 33 vectors, 187 vesicle, 279 video matting, 311 virulence factors, 89 vitamin C, 49

323
vitamin E, 1,49 vitamin E, 49 zoonosis, 59


325
ดชนคาสาคญ
กลองจลทรรศนแรงอะตอม, 233 การเขาจบกนของโมเลกลแบบจาเพาะ, 233 การคนหายน, 171 การจดการคลงยา, 105 การปฏสนธ, 25 การประมาณแบบคาเฉลย, 201 การประมาณแบบค, 201 การพฒนาวคซน, 33 การเพาะเลยงเนอเยอพช, 171 การแพรเชอ, 59 การวดภาคสนาม, 125 กงกลาดา, 25 แกโซฮอล, 125 ความเครยด, 41 ความหลากหลายทางชวภาพ, 163 เครองสาอาง , 153 เครองหมายโมเลกลเพอการปรบปรงพนธ, 171 แคโรทนอยด, 49 จอแสดงผลแบบแบน, 215 จดสงซอ, 105 ชดตรวจวด, 125 ชมชน, 195 เชอฉหน, 59 เชอรา, 179 เชอรากอโรคพช, 179 เซลลตดแตงพนธกรรม, 247 เซลลสบพนธ, 25 เซลลแสงอาทตย, 225 เซลลแสงอาทตยชนดสยอมไวแสง, 225 เทคโนโลยชวภาพพช, 171 นเวศวทยาเชงอนรกษ, 195 เนอในเมลดมะมวง, 153 แบบจาลองเชงพนทชดแจง, 201
แบบจาลองเชงพนทโดยปรยาย, 201 ปรมาณสงซอยาทเหมาะสม, 105 โปรตนลกผสม, 247 ผวหนงอกเสบระคายสมผส, 113 พลศาสตรประชากร, 201 พลงงานทางเลอก, 225 พนธวศวกรรม, 171 พาหะนาโรค, 187 ภมคมกน, 41 โมเลกลชวภาพ, 233 ยางพารา, 171 ยาทาคอรตโคสตรอยด, 113 ยงลายบาน, 187 ยงลายสวน, 187 รปแบบปรมาณการใชยา, 105 โรคพยาธใบไมตบ, 33 โรครบจากสตว, 59 วฏจกรเซลล, 247 วตามนซ, 49 วตามนอ, 49 วทยาศาสตร, 195 วธการเลเวลเซต, 241 สกลกลวย, 163 สกลกลวยดารารศม, 163 สกลกลวยโทน, 163 สงคมศาสตร, 195 สตวทใกลสญพนธ, 195 สารกนเสยชวภาพ, 153 สารตานอนมลเสร, 49, 153 สารพษ, 179

326
สารพษจาเพาะตอพชใหอาศย, 179 สารอนทรยเปลงแสง, 215 สยอม, 225 อนมลเสร, 49 อะพอพโทสส, 247 อปกรณเปลงแสงจากสารอนทรย, 215 เอทานอล, 125 เอนไซมกลตาไธโอนเพอรออกซเดส, 49 เอนไซมแคทาเลส, 49 เอนไซมซเปอรออกไซดดสมวเทส, 49 แอนตเจน, 33 โอแอลอด, 215 ฮอรโมน, 41

327
ดชนผนพนธ
กรกนก บญวงษ, อ. ดร., 201 Kornkanok Bunwong, Lect. Dr., 201
กลลยาน ดวงฉว, รศ. ดร., 59 Galayanee Doungchawee, Assoc. Prof. Dr., 59
กาญจนา วทยาคม, อ. ดร., 99 Kanjana Wittayacom, Lect. Dr., 99
กาญจนา อไรสนธว, อ. ดร., 125 Kanchana Uraisin, Lect. Dr., 125
กฤตกร คมพรสน, 99 Krittikorn Kumpornsin, 99
กลธดา ชยธระยานนท, อ. ดร., 33 Kulathida Chaithirayanon, lect. Dr., 33
จงกลณ วฒนาเพมพน, รศ. ดร., 73 Jonggonnee Wattanapermpool, Assoc. Prof. Dr., 73
จรญญา ณรงคะชวนะ, รศ. ดร., 171 Jarunya narangajavana, Assoc. Prof. Dr., 171
จามร สมณะ, อ. นพ. ดร., 163 Jamorn Somana, Lect. Dr., 163
จนรภา โพธกสกร, ดร., 187 Jinrapa Phothikasikorn, Dr., 187
จรารตน วงศคงคาเทพ, อ. ดร., 265 Jirarut Wongkongkatep, Lect. Dr., 265
เจนจต คดารงสวสด, อ. ดร.,195 Jenjit Khudamrongsawat, Lect, Dr., 195
ชาตชาย แจงเสน, อ. ดร., 139 Chartchai Changsen, Lect. Dr., 139
ชตมา เจยรพนจนนท ,อ. ดร., 7 Chutima Jiarpinitnun, Lect. Dr., 7
ณฏฐน พนธวศวาส, อ. ดร., 17, 257 Nathinee Panvisavas, Lect. Dr., 17, 257
ดวงกมล เบาวน, อ. ดร., 303 Duangkamon Baowan, Lect. Dr., 303
ดวงใจ นาคะปรชา, รศ. ดร., 125 Duangjai Nacapricha, Assoc. Prof. Dr., 125
ทพา อศวรกษ, อ. ดร., 179 Thipa Asvarak, Lect. Dr., 179
เทพมนส บบผาอนทร, อ. ดร., 73 Tepmanas Bupha-intr, Lect. Dr., 73
เทวญ จนทรวไลศร, อ. ดร., 81 Tavan Janvilisri, Lect. Dr., 81
ธนรรถ ชขจร, อ. ดร., 99 Thanat Chookajorn, Lect. Dr., 99
ธนากร โอสถจนทร, ผศ. ดร., 215 Tanakorn Osotchan, Assist. Prof. Dr., 215
ธญญารตน พงศทรางกร, อ. ดร., 287 Thunyarat Pongtharangkul, Lect. Dr., 287
ธรวฒ คลงเจรญชย, อ. นพ., 113 Theerawut Klanjareonchai, Lect. Dr.,113
ธราพร พนธธรานรกษ, อ. ดร., 233 Theeraporn Puntheeranurak, Lect. Dr., 233
ปฏญา เพงอน, 257 Patiya Phengon, 257
ประพนธ วไลรตน, ศ. ดร., 1 Prapon Wilairat, Prof. Dr., 1
ประวทย วงศคงคาเทพ, 265 Pravit Wongkongkatep, 265
ประเสรฐ โศภน, ศ. ดร., 25, 33 Prasert Sobhon, Prof. Dr., 25, 33
ปราณ ภญโญชพ, รศ. ดร., 265 Pranee Phinyocheep, Assoc. Prof. Dr., 265

328
ปราณ อนประโคน, อ. ดร., 153 Pranee Inprakhon, Lect. Dr., 153
ผดงศร ดบส, อ. ดร., 89 Padungsri Dubbs, Lect. Dr., 89
พหล โกสยจนดา, อ. ดร., 207 Pahol Kosiyachinda, Lect. Dr., 207
พทธรต พลสวรรณา, 99 Patrath Ponsuwanna, 99
พลลภ กาญจนภานรช, อ. ดร., 279 Pallop Karnchanaphanurach, Lect. Dr., 279
พลลภ ฮวบสมบรณ, อ. ดร., 241 Pallop Huabsomboon, Lect. Dr., 241
มณฑนา จรยาบรณ, อ. ดร., 295 Manthana Jariyaboon, Lect. Dr., 295
ยทธนา ตนตรงโรจนชย, ผศ. ดร., 225 Yuthana Tantirungrotechai, Assist. Prof., 225
รดกร อครวงษาพฒน, อ. ดร., 119 Radeekorn Akkarawongsapat, Lect. Dr., 119
รพพรรณ วานชวรยกจ, อ. ดร., 25 Rapeepan vanichvirikit, Lect. Dr., 25
รวศกด ธนวงศสวรรณ, อ. ดร., 311 Rawesak tanawongsuwan, Lect. Dr., 311
ระพ บญเปลอง, อ. ดร., 49, 187 Rapee Boonplueang, Lect. Dr., 49, 187
ระว สวรรณเดโชไชย, อ. ดร., 105 Rawee Suwandechochai, Lect. Dr., 105
รชนก เชอเตชะ, 99 Ruchanok Cheotacha, 99
รตนวจตร วจตรรตน, อ. ดร., 67 Rattanavijit Vijitruth, Lect. Dr., 67
รญจวน ละลาดาษ, 265 Runchuan Ladadat, 265
วรพจน ลาภเพมพนทรพย, 265 Woraphoj Lappenpuntrap, 265
วรรณจรส รงสทธพงษ, อ. พญ., 113 Wanjarus Roongpisuthipong, Lect. Dr., 113
วฑร ชนวชรศร, อ. ดร., 147 Withoon Chunwachirasiri, Lect. Dr., 147
วไล หนนภกด, รศ. ดร., 187 Wilai Noonpakdee, Assoc. Prof. Dr., 187
ศศวมล แสวงผล, อ. ดร., 163 Sasivimon Swangpol, Lect. Dr., 163
สมบญ สหสทธวฒน, ดร., 215 Somboon Sahasithiwat, Dr., 215
สมศกด แดงตบ, อ. ดร., 215 Somsak Dangtip, Lect. Dr., 215
สพชา คมเกต, อ. ดร., 41 Supeecha Kumkate, Lect. Dr., 41
สภา วรเศรษฐ, อ. ดร., 271 Supa Wirasate, Lect. Dr., 271
สภาวด เกยรตเสว, อ. ดร., 225 Supawadee Kiatsevi, Lect. Dr., 225
สภาวด ราชมณ, 187 Supawadee Ratchamanee, 187
สรยมาศ บวเทศ, อ. ดร., 131 Sureemas Buates, Lect. Dr., 131
อลสา วงใน, รศ. ดร., 287 Alisa S Vangnai, Assoc. Prof. Dr., 287
อษฎา มงสนตสข, อ. ดร., 247 Idsada Mungsantisuk, Lect. Dr., 247


คณะวทยาศาสตร มหาวทยาลยมหดล272 ถนนพระรามท 6 เขตราชเทว กรงเทพฯ 10400โทรศพท 0-2201-5000 โทรสาร 0-2201-5072
www.sc.mahidol.ac.th
Copyright © 2022 FDOKUMEN








