
Pengendalian Penyakit Busuk Pangkal Batang Kelapa Sawit...
36
Pengendalian Penyakit Busuk Pangkal Batang Kelapa Sawit secara Kultur Teknis Cultural Practices for Management of Basal Stem Rot Disease of Oil Palm Hari Priwiratama, Agus Eko Prasetyo, Agus Susanto* Pusat Penelitian Kelapa Sawit, Medan 20158 ABSTRAK Pengendalian penyakit busuk pangkal batang yang disebabkan oleh Ganoderma boninense secara kultur teknis dapat dilakukan melalui beberapa metode. Empat seri percobaan lapangan dilakukan untuk menentukan keefektifan pengendalian kultur teknis yang meliputi sanitasi sumber inokulum, sistem penanaman hole in hole, pembedahan dan pembumbunan, dan pembuatan parit isolasi. Percobaan sanitasi sumber inokulum terdiri atas sanitasi sisa-sisa akar dan penggunaan lubang tanam besar, penggunaan lubang tanam besar dan sistem tanam ulang standar sebagai pembanding. Percobaan sistem tanam hole in hole terdiri atas penanaman dengan sistem hole in hole dan penanaman standar sebagai pembanding. Percobaan pembedahan dan pembumbunan terdiri atas pembedahan dan pembumbunan disertai aplikasi teer dan Trichoderma, pembedahan dan pembumbunan disertai aplikasi Trichoderma, pembumbunan disertai aplikasi Trichoderma, dan sebagai pembanding ialah tanaman terserang tanpa perlakuan. Percobaan parit isolasi digunakan untuk membandingkan antara perlakuan parit isolasi dan tanpa parit isolasi. Insidensi penyakit busuk pangkal batang diamati setiap tahun. Hasil penelitian menunjukkan bahwa empat metode pengendalian secara kultur teknis dapat mencegah infeksi G. boninense pada tahap awal perkembangan kelapa sawit. Sanitasi inokulum mampu mencegah infeksi G. boninense hingga 2 tahun setelah perlakuan (TSP), sistem hole in hole mampu menekan perkembangan penyakit busuk pangkal batang hingga 7 TSP, pembedahan dan pembumbunan mampu memperpanjang masa hidup tanaman terinfeksi hingga 3, TSP dan parit isolasi mampu mencegah penularan penyakit hingga 2 TSP. Kata kunci: hole in hole, insidensi penyakit, parit isolasi, pembedahan, pembumbunan, sanitasi ABSTRACT Effort to manage basal stem rot disease (BSR) caused by Ganoderma boninense could be conducted through several cultural practice approaches. Four series of field experiments was conducted to observe the effectiveness of cultural practice methods for controlling the epidemic of basal stem rot due to G. boninense. Sanitation trial consisted of three treatments i.e root sanitation with big hole planting system, big hole without root sanitation, and standard replanting system as comparison. Hole in hole and standard planting system were compared in the hole in hole trial. Digging and mounding trial was conducted in four treatments, i.e. digging and mounding with teer and Trichoderma, digging and mounding with Trichoderma, mounding with Trichoderma, and infected palm without any treatment as comparison. Meanwhile, a 4 m × 4 m trench was used in the isolation trench trial. Annual observation was conducted to determine the disease incidence of BSR in each trial. Results showed that all methods could prevent palms from G. boninense infection in the early stage of oil palm development. Proper root sanitation delayed G. boninense infection until 2 years after treatment (YAT), hole in hole planting system was able to suppress disease incidence up to 7 YAT, digging and mounding was useful to prolong *Alamat penulis korespondensi: Pusat Penelitian Kelapa Sawit, Jalan Brigjen Katamso No 51, Medan 20158 Tel: 061-7862477, Faks: 061-7862488, Surel: [email protected] Volume 10, Nomor 1, Februari 2014 Halaman 1–7 DOI: 10.14692/jfi.10.1.1 ISSN: 2339-2479 1
Transcript of Pengendalian Penyakit Busuk Pangkal Batang Kelapa Sawit...

Pengendalian Penyakit Busuk Pangkal Batang Kelapa Sawit secara Kultur Teknis
Cultural Practices for Management of Basal Stem Rot Disease of Oil Palm
Hari Priwiratama Agus Eko Prasetyo Agus SusantoPusat Penelitian Kelapa Sawit Medan 20158
ABSTRAK
Pengendalian penyakit busuk pangkal batang yang disebabkan oleh Ganoderma boninense secara kultur teknis dapat dilakukan melalui beberapa metode Empat seri percobaan lapangan dilakukan untuk menentukan keefektifan pengendalian kultur teknis yang meliputi sanitasi sumber inokulum sistem penanaman hole in hole pembedahan dan pembumbunan dan pembuatan parit isolasi Percobaan sanitasi sumber inokulum terdiri atas sanitasi sisa-sisa akar dan penggunaan lubang tanam besar penggunaan lubang tanam besar dan sistem tanam ulang standar sebagai pembanding Percobaan sistem tanam hole in hole terdiri atas penanaman dengan sistem hole in hole dan penanaman standar sebagai pembanding Percobaan pembedahan dan pembumbunan terdiri atas pembedahan dan pembumbunan disertai aplikasi teer dan Trichoderma pembedahan dan pembumbunan disertai aplikasi Trichoderma pembumbunan disertai aplikasi Trichoderma dan sebagai pembanding ialah tanaman terserang tanpa perlakuan Percobaan parit isolasi digunakan untuk membandingkan antara perlakuan parit isolasi dan tanpa parit isolasi Insidensi penyakit busuk pangkal batang diamati setiap tahun Hasil penelitian menunjukkan bahwa empat metode pengendalian secara kultur teknis dapat mencegah infeksi G boninense pada tahap awal perkembangan kelapa sawit Sanitasi inokulum mampu mencegah infeksi G boninense hingga 2 tahun setelah perlakuan (TSP) sistem hole in hole mampu menekan perkembangan penyakit busuk pangkal batang hingga 7 TSP pembedahan dan pembumbunan mampu memperpanjang masa hidup tanaman terinfeksi hingga 3 TSP dan parit isolasi mampu mencegah penularan penyakit hingga 2 TSP
Kata kunci hole in hole insidensi penyakit parit isolasi pembedahan pembumbunan sanitasi
ABSTRACT
Effort to manage basal stem rot disease (BSR) caused by Ganoderma boninense could be conducted through several cultural practice approaches Four series of field experiments was conducted to observe the effectiveness of cultural practice methods for controlling the epidemic of basal stem rot due to G boninense Sanitation trial consisted of three treatments ie root sanitation with big hole planting system big hole without root sanitation and standard replanting system as comparison Hole in hole and standard planting system were compared in the hole in hole trial Digging and mounding trial was conducted in four treatments ie digging and mounding with teer and Trichoderma digging and mounding with Trichoderma mounding with Trichoderma and infected palm without any treatment as comparison Meanwhile a 4 m times 4 m trench was used in the isolation trench trial Annual observation was conducted to determine the disease incidence of BSR in each trial Results showed that all methods could prevent palms from G boninense infection in the early stage of oil palm development Proper root sanitation delayed G boninense infection until 2 years after treatment (YAT) hole in hole planting system was able to suppress disease incidence up to 7 YAT digging and mounding was useful to prolong
Alamat penulis korespondensi Pusat Penelitian Kelapa Sawit Jalan Brigjen Katamso No 51 Medan 20158 Tel 061-7862477 Faks 061-7862488 Surel marihat_agusyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 1ndash7
DOI 1014692jfi1011ISSN 2339-2479
1
J Fitopatol Indones Priwiratama et al
the life of the infected palms up to 3 YAT meanwhile the use of isolation trench was demonstrated to prevent G boninense infection to neighboring palms up to 2 YAT
Key words disease incidence hole in hole isolation trench sanitation digging and mounding
PENDAHULUAN
Busuk pangkal batang (BPB) kelapa sawit yang disebabkan oleh Ganoderma boninense merupakan penyakit yang paling destruktif di perkebunan kelapa sawit di Indonesia dan Malaysia Patogen ini tidak hanya menyerang tanaman tua tetapi juga yang masih muda Saat ini laju infeksi penyakit BPB berjalan semakin cepat terutama pada tanah dengan tekstur berpasir (Susanto et al 2013) Beberapa dekade lalu insidensi penyakit yang tinggi hanya dijumpai pada kebun dengan lebih dari dua kali tanam ulang namun saat ini insidensi penyakit sudah cukup tinggi Dengan demikian sekarang banyak ditemukan daerah perkebunan kelapa sawit dengan kriteria endemik G boninense dan mengalami kerugian yang besar
Upaya pengendalian penyakit BPB kelapa sawit telah banyak dilakukan oleh pekebun kelapa sawit Pengendalian secara kultur teknis dapat dilakukan sejak proses tanam ulang yaitu melalui sanitasi sisa-sisa batang dan akar yang terinfeksi Ganoderma Sanitasi sumber inokulum ini dapat meminimalkan kontak antara akar sehat dan sisa-sisa akar terinfeksi yang merupakan salah satu mekanisme utama penyebaran Ganoderma di lapangan (Paterson 2007 Naher et al 2013) Pada daerah endemik Ganoderma umumnya diterapkan sistem penanaman hole in hole dan parit isolasi
Pengendalian hayati dilakukan dengan pemanfaatan agens antagonis seperti cendawan Trichoderma sp (Priwiratama dan Susanto 2014) dan endomikoriza (Kartika et al 2006) Meskipun demikian hasil pengendalian secara hayati ini masih belum konsisten di lapangan Sementara itu teknik pengendalian secara kimiawi sintetik menggunakan beberapa bahan aktif fungisida juga dilaporkan kurang memuaskan Pengendalian penyakit BPB yang paling ideal ialah menggunakan tanaman toleran Ganoderma (Idris et al
2004 Durand-Gasselin et al 2005) tetapi untuk mendapatkan tanaman yang toleran masih membutuhkan waktu yang relatif lama Oleh karena itu tujuan penelitian ini ialah untuk menentukan strategi pengendalian secara kultur teknis
BAHAN DAN METODE
Penelitian terdiri atas empat seri percobaan lapangan yaitu sanitasi sumber inokulum pada waktu tanam ulang kultur teknis penanaman dan penyisipan kelapa sawit pembedahan dan pembumbunan tanaman terinfeksi Ganoderma dan penerapan parit isolasi untuk mengurangi penyebaran penyakit BPB Penelitian berlangsung dari tahun 2008 sampai tahun 2013 Pada masing-masing seri percobaan diamati insidensi penyakit Ganoderma atau kematian tanaman setiap tahunnya
Sanitasi Sumber Inokulum G boninase Saat Awal Tanam Ulang
Penelitian dilakukan sejak tahun 2011 pada areal endemik Ganoderma di kebun Sei Pancur Sumatera Utara Persiapan lahan selama proses tanam ulang dilakukan dengan tiga metode yaitu persiapan lahan standar dengan lubang tanam besar disertai pengumpulan dan pemusnahan sisa-sisa akar tanaman lama (A) persiapan lahan standar dengan lubang tanam besar tanpa pengumpulan sisa-sisa akar tanaman lama (B) dan persiapan lahan dengan prosedur standar (C) Tiga perlakuan ini dilakukan pada luasan 5 ha dengan tingkat insidensi penyakit BPB lebih dari 50 pada penanaman sebelumnya
Persiapan lahan standar dilakukan dengan beberapa tahap kegiatan umum yang terdiri atas bajak dan garu tanah penumbangan tanaman lama disertai sanitasi batang dan bonggol kelapa sawit dan pembuatan lubang tanam berukuran 60 cm times 60 cm dengan
2
J Fitopatol Indones Priwiratama et al
kedalaman 50 cm Pada perlakuan A dan B lubang tanam besar yang digunakan berukuran 3 m times 3 m dengan kedalaman 80 cm Sisa-sisa akar tanaman kelapa sawit lama dikumpulkan secara manual selama proses penumbangan dan pengolahan tanah berlangsung
Perlakuan disusun dalam rancangan acak kelompok (RAK) dengan 10 ulangan Pada masing-masing ulangan terdapat 70 tanaman sampel Pengamatan insidensi penyakit di-lakukan secara terus-menerus pada interval enam bulan
Sistem Penanaman Hole in HolePenelitian dilakukan di kebun Teluk
Dalam Sumatera Utara sejak tahun 2003 Perlakuan terdiri atas sistem penanaman hole in hole dan sistem penanaman standar sebagai pembanding Perlakuan disusun dalam RAK yang diulang dalam 17 blok penanaman Setiap blok memiliki luasan 25 ha dengan kerapatan 143 tanaman per ha Insidensi penyakit BPB diamati pada seluruh tanaman dengan interval 1 tahun
Sistem penanaman hole in hole dilakukan dengan membuat lubang tanam standar berukuran 60 cm times 60 cm dengan kedalaman 50 cm di dalam lubang tanam besar berdimensi 3 m times 3 m times 08 m sedangkan pada sistem penanaman standar hanya digunakan lubang tanam berukuran 60 cm times 60 cm Masing-masing perlakuan diberikan aplikasi sebanyak 400 g Trichoderma sp ke lubang tanam sebelum penanaman bibit kelapa sawit
Pembedahan dan Pembumbunan Tanaman Terinfeksi Ganoderma
Percobaan dilakukan pada tanaman kelapa sawit berumur 7 tahun di kebun Dalu-Dalu (Riau) kebun Bukit Sentang dan kebun Aek Pancur (Sumatera Utara) sejak 2010 Tanaman terserang Ganoderma dibedah dengan cara membuang seluruh jaringan pangkal batang yang telah mengalami pembusukan Sisa-sisa jaringan terinfeksi dari pembedahan dikumpulkan dan dimusnahkan dan jaringan pangkal batang setelah pembedahan dilapisi dengan senyawa fenolik yang berfungsi sebagai desinfektan (teer) Selanjutnya tanaman dibumbun dengan tanah yang telah
dicampur 400 g Trichoderma sp hingga menutupi seluruh jaringan tanaman yang dibedah
Perlakuan dibedakan menjadi empat yaitu pembedahan dengan aplikasi 415 mL teer dan pembumbunan (A) pembedahan dan pembumbunan (B) pembumbunan disertai aplikasi Trichoderma sp (C) dan sebagai pembanding adalah tanaman terserang tanpa perlakuan (D) Perlakuan disusun dalam RAK dengan 10 ulangan dan pada masing-masing ulangan terdapat 4ndash5 tanaman sampel
Parit IsolasiPercobaan dilaksanakan pada tanaman
kelapa sawit berumur 8 tahun di kebun Sampit Kalimantan Tengah sejak 2011 Parit isolasi individu dibuat mengelilingi tanaman ter-infeksi Ganoderma berukuran 4 m times 4 m dengan lebar dan kedalaman parit berturut-turut 40 cm dan 60 cm Sebagai pembanding ialah tanaman terinfeksi tanpa perlakuan parit individu Perlakuan disusun dalam rancangan acak kelompok dengan 64 ulangan dan pada setiap ulangan terdapat 7 tanaman sampel
HASIL
Sanitasi Sumber Inokulum Ganoderma Saat Awal Tanam Ulang
Berdasarkan hasil pengamatan sisa-sisa akar kelapa sawit yang dikumpulkan berbobot 7250 kg atau rerata 1450 kg ha-1 Insidensi penyakit BPB pada perlakuan persiapan lahan standar pada satu tahun setelah perlakuan (TSP) sudah mencapai 21 dan meningkat hingga 68 pada 2 TSP Sementara itu kelapa sawit yang memperoleh perlakuan dengan lubang tanam besar dan pengumpulan akar belum diserang oleh BPB sampai dengan 2 TSP (Gambar 1)
Sistem Penanaman Hole in HoleInsidensi penyakit BPB pada sistem tanam
standar dan hole in hole perlakuan sudah dapat diamati sejak 1 TSP dan terus meningkat hingga 10 TSP (Gambar 2) Secara umumtingkat insidensi penyakit BPB pada sistem penanaman hole in hole lebih rendah di-bandingkan dengan sistem tanam standar
3
J Fitopatol Indones Priwiratama et al
Hingga 7 TSP insidensi penyakit BPB pada sistem penanaman hole in hole meningkat dari 076ndash517 sedangkan insidensi penyakit pada sistem tanam standar meningkat dari 22ndash724 Pada 10 TSP insidensi penyakit BPB pada kedua perlakuan dijumpai pada tingkat yang relatif berimbang
Penerapan sistem penanaman hole in hole tidak memberikan pengaruh terhadap produksi tandan buah segar Hasil rerata produksi tandan buah segar per tahun selama 5 tahun terakhir tidak menunjukkan perbedaan yang nyata antara dua sistem tanam (Tabel 1) Produksi tandan buah segar per tahun berfluktuasi antara 1721ndash1924 ton ha-1 per tahun pada sistem penanaman hole in hole dan 1585 dan 1988 ton ha-1 per tahun pada sistem penanaman standar
Pembedahan dan Pembumbunan Tanaman Terinfeksi Busuk Pangkal Batang
Hingga 3 TSP tingkat kematian tanaman terinfeksi Ganoderma pada perlakuan kontrol secara nyata lebih tinggi dibandingkan de-ngan perlakuan pembedahan dengan aplikasi 415 mL teer dan pembumbunan pembedahan dan pembumbunan pembumbunan disertai aplikasi Trichoderma (Tabel 2) Untuk per-lakuan kontrol tingkat kematian tanaman terinfeksi pada 1 2 dan 3 TSP berturut-turut 233 2791 dan 3256 Sementara itu hingga 3 TSP kematian tanaman pada perlakuan A lebih rendah dibandingkan dengan perlakuan B dan C dengan tingkat kematian berturut-turut 233 465 dan 465
4
Gambar 1 Perkembangan insidensi penyakit busuk pangkal batang oleh Ganoderma boninense di kebun Sei Pencur Sumatra Utara dengan tiga perlakuan persiapan lahan standar lubang tanam besar dan pengumpulan sisa-sisa akar persiapan lahan standar dan lubang tanam besar dan persiapan lahan standar
Kej
adia
n pe
nyak
it (
) 8
6
4
2
01 2
Tahun setelah tanam
Gambar 2 Perkembangan insidensi penyakit busuk pangkal batang oleh Ganoderma boninense di kebun Teluk Dalam Sumatra Utara pada dua sistem tanam standar dan hole in hole
Kej
adia
n pe
nyak
it (
)
20
16
12
8
4
01 2 3 4 5 6 7 8 9 10
Tahun setelah tanam
J Fitopatol Indones Priwiratama et al
Parit IsolasiSecara umum insidensi penyakit BPB pada
perlakuan kontrol lebih tinggi dibandingkan dengan perlakuan parit isolasi (Tabel 3) Insidensi penyakit BPB pada perlakuan kontrol sudah teramati sejak 1 TSP (052) dan terus meningkat hingga 182 pada 3 TSP Sebaliknya pada perlakuan parit isolasi insidensi penyakit BPB baru teramati pada 3 TSP dengan tingkat yang lebih rendah (026)
PEMBAHASAN
Sanitasi sisa-sisa akar kelapa sawit saat tanam ulang memiliki peran penting untuk mencegah penularan dini G boninense Pada
sistem tanam ulang standar kelapa sawit memiliki risiko terinfeksi G boninense lebih tinggi sejak tahun pertama setelah penanaman yang dapat dilihat dari insidensi penyakit mencapai 2 Hal ini terjadi karena tingginya peluang terjadi kontak akar antara perakaran kelapa sawit sehat dan sisa-sisa akar terinfeksi yang lebih cepat pada sistem tanam ulang standar dibandingkan dengan sistem yang diujikan Virdiana et al (2012) melaporkan bahwa penanaman bibit kelapa sawit dengan jarak sekurang-kurangnya 2 m dari lubang tanam lama pada saat tanam ulang dapat menunda proses infeksi penyakit melalui kontak akar dan menekan tingkat insidensi penyakit secara nyata hingga 11 tahun setelah tanam Sebelumnya Flood et al (2005) juga telah melaporkan bahwa penanaman bibit kelapa sawit jauh dari tumpukan batang dan bonggol saat tanam ulang dapat menunda infeksi Ganoderma
Upaya menekan laju penyakit BPB dapat juga dilakukan melalui penggunaan sistem tanam hole in hole Sistem tanam ini bertujuan mengurangi sumber inokulum Ganoderma hingga seminimal mungkin pada titik pe-nanaman bibit kelapa sawit Berdasarkan percobaan jangka panjang yang dilakukan di daerah endemik penyakit BPB di kebun
5
Tabel 1 Rerata produksi tandan buah segar di kebun Teluk Dalam Sumatra Utara pada sistem tanam standar dan hole in hole
Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Perlakuan
Lubang tanam standarHole in hole
20102009 2011Rerata produksi TBS (tonhatahun)
201320121585 a1721 a
1810 a1927 a
1988 a1974 a
1924 a1853 a
1880 a1752 a
Tabel 2 Perkembangan kematian tanaman terinfeksi Ganoderma pada perlakuan pembedahan dan pembumbunan
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
PerlakuanPembedahan dengan aplikasi teer dan pembumbunanPembedahan dan pembumbunanPembumbunanKontrol
2 TSP1 TSP 3 TSPMortalitas tanaman ()
000 a000 a000 a233 a
233 a 000 a 000 a2791 b
233 a 465 a 465 a3256 b
Tabel 3 Insidensi penyakit busuk pangkal batang di kebun Sampit Kalimantan Tengah pada sistem parit isolasi
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Sistem Tanam
Parit isolasiKontrol
2 TSP1 TSP 3 TSPInsidensi penyakit ()
000 a052 a
000 a104 b
026 a182 b
J Fitopatol Indones Priwiratama et al
Teluk Dalam Sumatera Utara sistem tanam hole in hole dapat mengurangi tingkat G boninense pada masa awal penanaman kelapa sawit di lapangan Sistem penanaman hole in hole mampu menekan tingkat insidensi penyakit BPB hingga 7 TSP dibandingkan dengan sistem penanaman dengan lubang tanam standar Hal ini dapat disebabkan oleh perbedaan pola sebaran dan jangkauan akar kelapa sawit yang berbeda dengan pada kedua sistem penanaman Perakaran kelapa sawit berumur 3 tahun pada lubang tanam standar dapat menjangkau area sepanjang 467 m dengan sebaran merata sehingga meningkatkan potensi kontak dengan inokulum G boninense di lapangan Pada sistem hole in hole jangkauan akar hanya mencapai 301 m dengan pola sebaran akar terpusat di dalam lubang tanam besar sehingga memperkecil peluang terjadinya kontak akar dengan sumber inokulum Namun demikian setelah 7 TSP sistem tanam hole in hole tidak mampu menghambat laju perkembangan penyakit BPB Pada tahap ini perakaran kelapa sawit telah tumbuh berkembang melebihi area lubang sanitasi sehingga peluang terjadinya kontak dengan sumber inokulum G boninense di luar area hole in hole menjadi semakin tinggi
Penggunaan sistem hole in hole dianggap memberikan dampak negatif terhadap per-kembangan dan produktivitas kelapa sawit karena sebagian besar lapisan tanah bagian atas yang memiliki kandungan hara lebih tinggi terbuang dan hanya menyisakan lapisan tanah dalam yang miskin unsur hara Namun berdasarkan pengamatan di lapangan sistem tanam ini tidak memberikan pengaruh terhadap keragaan tanaman dan produktivitas kelapa sawit Data produksi TBS selama lima tahun terakhir (Tabel 1) memperlihatkan produktivitas kelapa sawit pada kedua sistem tanam relatif berimbang Dalam praktiknya penambahan bahan organik berupa tandan kosong kelapa sawit pada penerapan sistem hole in hole merupakan salah satu faktor penting agar asupan nutrisi kelapa sawit selama tahap awal perkembangannya dapat terpenuhi
sehingga keragaan dan produktivitasnya tidak tertinggal dari sistem tanam standar
Penerapan teknik pembedahan dan pem-bumbunan untuk mengendalikan penyakit BPB menunjukkan hasil yang cukup menjanjikan Meskipun tidak dapat menghindarkan tanaman dari infeksi G boninense pembedahan jaringan sakit yang disertai pembumbunan mampu memperpanjang masa hidup tanaman terinfeksi sekurang-kurangnya hingga 3 TSP dibandingkan dengan tanaman tanpa perlakuan Sedangkan pada tanaman dengan hanya pembumbunan mampu bertahan hingga 2 TSP Hal ini cukup penting mengingat dalam kurun waktu 2ndash3 tahun tanaman sakit masih terus memproduksi TBS Pembedahan dilaporkan dapat memperpanjang masa hidup kelapa sawit hingga 2ndash3 tahun (Ho dan Hashim 1997) Pembedahan di perkebunan rakyat mampu memperpanjang masa hidup kelapa sawit di lapangan (Marshall et al 2004 Cooper et al 2011) Berdasarkan pengamatan di lapangan tingkat keberhasilan teknik pembedahan ini sangat bergantung pada ketepatan jaringan sakit yang dibuang Bila jaringan sakit tidak seluruhnya dibuang umumnya kelapa sawit hanya akan bertahan selama satu tahun karena sisa jaringan sakit tersebut berperan sebagai sumber inokulum yang akan terus berkembang hingga menyebabkan pembusukan pada pangkal batang tanaman yang dibumbun
Teknik pengendalian kultur teknis lain yang dapat diterapkan di lapangan ialah pembuatan parit isolasi yang bertujuan mencegah terjadinya kontak akar Hasil pe-ngamatan menunjukkan bahwa parit isolasi mampu mencegah terjadinya infeksi pada tanaman tetangga di luar parit hingga 2 TSP Keberadaan parit isolasi akan mencegah terjadinya kontak akar sehingga tanaman sehat terhindar dari infeksi G boninense dari jaringan tanaman terinfeksi dalam area isolasi Meskipun cukup sulit ketepatan waktu dan titik pembuatan parit isolasi sangat mempengaruhi keefektifan pencegahan pe-nyebaran G boninense di lapangan Dengan akurasi waktu pembuatan yang baik parit isolasi berukuran 2 m times 2 m dilaporkan mampu
6
J Fitopatol Indones Priwiratama et al
mencegah penularan penyakit BPB hingga 14 tahun (Hasan dan Turner 1998 Chung 2011)
Dari uraian di atas dapat diketahui bahwa tujuan pengendalian penyakit BPB secara kultur teknis dititikberatkan untuk menekan adanya sumber inokulum potensial sehingga dapat mencegah terjadinya infeksi G boninense di lapangan Metode pengendalian kultur teknis yang digunakan dapat disesuaikan dengan tahap perkembangan kelapa sawit Di areal pengembangan atau penanaman baru pengendalian dilakukan sejak tahap awal penanaman melalui sanitasi sumber inokulum terutama sisa-sisa perakaran kelapa sawit dan penggunaan lubang tanam besar Sistem penanaman hole in hole dapat diterapkan untuk melakukan penyisipan tanaman baru terutama di areal endemik G boninense Sementara itu untuk tanaman terserang dapat dilakukan tindakan pembedahan diiringi pembumbunan dan bila sudah tidak memungkinkan dapat dilakukan isolasi tanaman terserang untuk mencegah penularan penyakit pada tanaman sehat di sekitarnya Beberapa penelitian di atas akan terus dilanjutkan untuk melihat pengaruh jangka panjang dari teknik-teknik yang diterapkan
DAFTAR PUSTAKA
Chung GF 2011 Management of Ganoderma diseases in oil palm plantations Planter 87(1022)325ndash339
Cooper RM Flood J Rees R 2011 Ganoderma boninense in oil palm plantations current thinking on epidemiology resistance and pathology Planter 87(1024)515ndash526
Durand-Gasselin T Asmady H Flori A Jacquemard Jc Hayun Z Breton F De Franqueville H 2005 Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq) to basal stem rot caused by Ganoderma boninensemdashprospects for future breeding Mycopathologia 159(1)93ndash100 DOI httpdxdoiorg10 1007s11046-004-4429-1
Flood J Keenan L Wayne S Hasan Y 2005 Studies on oil palm trunks as sources of infection in the field Mycopathologia
159(1)101ndash107 DOI httpdxdoiorg 101007s11046-004-4430-8
Hasan Y Turner P 1998 The comparative importance of different oil palm tissues as infection sources for basal stem rot in replantings Planter 74(864)119ndash135
Ho CT Hashim K 1997 Usefulness of soil mounding treatments in prolonging productivity of prime-aged Ganoderma infected palms Planter 73(854)239ndash244
Idris AS Kushairi A Ismail S Ariffin D 2004 Selection for partial tolerance in oil palm progenies to Ganoderma basal stem rot J Oil Palm Res 16(2)12ndash18
Kartika E Yahya S Wilarso S 2006 Isolasi karakterisasi dan pemurnian cendawan mikoriza arbuskular dari dua lokasi perkebunan kelapa sawit (bekas hutan dan bekas kebun karet) J Penelitian Kelapa Sawit 14(3)145ndash155
Marshall R Hunt R Pilotti C 2004 Low cost control for basal stem rotmdasha Poliamba initiative Planter 80(936)173ndash176
Naher L Yusuf UK Tan SG Ismail A 2013 Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq) Aus Sci 7(11)1723ndash1727
Paterson RRM 2007 Ganoderma disease of oil palmmdasha white rot perspective necessary for integrated control Crop Protect 26(2007)1369ndash1376 DOI httpdxdoiorg101016jcropro200611009
Priwiratama H Susanto A 2014 Utilization of fungi for the biological control of insect pests and Ganoderma disease in the Indonesian oil palm industry Agr Sci Tech A 4(2014)103ndash111
Susanto A Prasetyo AE Wening S 2013 Laju infeksi Ganoderma pada empat kelas tekstur tanah J Fitopatol Indones 9(2)39-46 DOI httpdxdoiorg1014692jfi9239
Virdiana I Flood J Sitepu B Hasan Y Aditya R Nelson S 2012 Integrated disease management to reduce future Ganoderma infection during oil palm replanting Planter 88(1305)383ndash393
7
Volume 10 Nomor 1 Februari 2014Halaman 8ndash16
DOI 1014692jfi1018
Pengelolaan Penyakit Kuning pada Tanaman Lada oleh Petani di Wilayah Bangka
Management of Yellow Diseases on Pepper by Farmers in Bangka Region
Abdul Munif Ita SulistiawatiInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit kuning pada tanaman lada yang disebabkan oleh nematoda parasit tumbuhan hingga kini masih menjadi masalah utama pada pertanaman lada di wilayah Bangka Kehilangan hasil berupa penurunan produksi lada yang diakibatkan oleh penyakit kuning cukup besar Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan Penelitian ini meliputi dua kegiatan yaitu survei tingkat kejadian penyakit kuning pada pertanaman lada di lapangan dan wawancara dengan petani lada Hasil survei menunjukkan bahwa tingkat kejadian penyakit kuning sudah merata dan menyerang hampir semua lahan yang diamati Hampir semua petani responden menyatakan bahwa penyakit kuning merupakan penyakit utama yang sangat merusak pertanaman lada Tingkat pemahaman petani terhadap penyebab gejala dan ekologi penyakit kuning dan cara-cara pengendaliannya masih sangat terbatas Tindakan pengendalian yang dilakukan oleh petani terhadap penyakit kuning ialah pencabutan dan pembakaran tanaman terserang dan pemberian kapur Beberapa petani tidak melakukan tindakan pengendalian namun sebagian besar petani menggunakan pestisida Penggunaan pestisida untuk pengendalian penyakit kuning oleh petani belum sepenuhnya mengikuti prinsip pengendalian hama terpadu Masih rendahnya pemahaman petani terhadap penyakit kuning mengakibatkan semakin sulit bagi petani untuk melakukan pengendaliannya
Kata kunci nematoda pengendalian penyakit pestisida Piper nigrum
ABSTRACT
Yellow disease caused by plant-parasitic nematodes are still a major problem on pepper production area in Bangka region Significant yield losses occurred due to yellow disease The objective of this research was to survey the yellow disease incidence and to study farmerrsquos knowledge attitude and practices on the management of yellow disease of black pepper in Bangka district Central Bangka and South Bangka This study consisted of a survey of the incidence of yellow disease in the field and interviews to the farmers The result showed that the yellow disease had infected nearly all of black pepper fields in the areas with incidence of as much as 20ndash60 Almost all of the respondents reported that the yellow disease was the major disease of black pepper in Bangka region Farmersrsquo level of knowledge about the causal of disease symptoms ecology and control methods of the yellow disease are still very limited Various control measures have been done by farmers to combat the disease especially by removing and burning the infected plants or by application of lime A few farmers did not even controlling the disease Pesticides have been used by a few farmers to control the yellow disease but the application of pesticide does not in line with integrated pest management principles
Key words disease control nematode pesticide Piper nigrumAlamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor
Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel abdulmunifipbacid
8
ISSN 2339-2479
J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Priwiratama et al
the life of the infected palms up to 3 YAT meanwhile the use of isolation trench was demonstrated to prevent G boninense infection to neighboring palms up to 2 YAT
Key words disease incidence hole in hole isolation trench sanitation digging and mounding
PENDAHULUAN
Busuk pangkal batang (BPB) kelapa sawit yang disebabkan oleh Ganoderma boninense merupakan penyakit yang paling destruktif di perkebunan kelapa sawit di Indonesia dan Malaysia Patogen ini tidak hanya menyerang tanaman tua tetapi juga yang masih muda Saat ini laju infeksi penyakit BPB berjalan semakin cepat terutama pada tanah dengan tekstur berpasir (Susanto et al 2013) Beberapa dekade lalu insidensi penyakit yang tinggi hanya dijumpai pada kebun dengan lebih dari dua kali tanam ulang namun saat ini insidensi penyakit sudah cukup tinggi Dengan demikian sekarang banyak ditemukan daerah perkebunan kelapa sawit dengan kriteria endemik G boninense dan mengalami kerugian yang besar
Upaya pengendalian penyakit BPB kelapa sawit telah banyak dilakukan oleh pekebun kelapa sawit Pengendalian secara kultur teknis dapat dilakukan sejak proses tanam ulang yaitu melalui sanitasi sisa-sisa batang dan akar yang terinfeksi Ganoderma Sanitasi sumber inokulum ini dapat meminimalkan kontak antara akar sehat dan sisa-sisa akar terinfeksi yang merupakan salah satu mekanisme utama penyebaran Ganoderma di lapangan (Paterson 2007 Naher et al 2013) Pada daerah endemik Ganoderma umumnya diterapkan sistem penanaman hole in hole dan parit isolasi
Pengendalian hayati dilakukan dengan pemanfaatan agens antagonis seperti cendawan Trichoderma sp (Priwiratama dan Susanto 2014) dan endomikoriza (Kartika et al 2006) Meskipun demikian hasil pengendalian secara hayati ini masih belum konsisten di lapangan Sementara itu teknik pengendalian secara kimiawi sintetik menggunakan beberapa bahan aktif fungisida juga dilaporkan kurang memuaskan Pengendalian penyakit BPB yang paling ideal ialah menggunakan tanaman toleran Ganoderma (Idris et al
2004 Durand-Gasselin et al 2005) tetapi untuk mendapatkan tanaman yang toleran masih membutuhkan waktu yang relatif lama Oleh karena itu tujuan penelitian ini ialah untuk menentukan strategi pengendalian secara kultur teknis
BAHAN DAN METODE
Penelitian terdiri atas empat seri percobaan lapangan yaitu sanitasi sumber inokulum pada waktu tanam ulang kultur teknis penanaman dan penyisipan kelapa sawit pembedahan dan pembumbunan tanaman terinfeksi Ganoderma dan penerapan parit isolasi untuk mengurangi penyebaran penyakit BPB Penelitian berlangsung dari tahun 2008 sampai tahun 2013 Pada masing-masing seri percobaan diamati insidensi penyakit Ganoderma atau kematian tanaman setiap tahunnya
Sanitasi Sumber Inokulum G boninase Saat Awal Tanam Ulang
Penelitian dilakukan sejak tahun 2011 pada areal endemik Ganoderma di kebun Sei Pancur Sumatera Utara Persiapan lahan selama proses tanam ulang dilakukan dengan tiga metode yaitu persiapan lahan standar dengan lubang tanam besar disertai pengumpulan dan pemusnahan sisa-sisa akar tanaman lama (A) persiapan lahan standar dengan lubang tanam besar tanpa pengumpulan sisa-sisa akar tanaman lama (B) dan persiapan lahan dengan prosedur standar (C) Tiga perlakuan ini dilakukan pada luasan 5 ha dengan tingkat insidensi penyakit BPB lebih dari 50 pada penanaman sebelumnya
Persiapan lahan standar dilakukan dengan beberapa tahap kegiatan umum yang terdiri atas bajak dan garu tanah penumbangan tanaman lama disertai sanitasi batang dan bonggol kelapa sawit dan pembuatan lubang tanam berukuran 60 cm times 60 cm dengan
2
J Fitopatol Indones Priwiratama et al
kedalaman 50 cm Pada perlakuan A dan B lubang tanam besar yang digunakan berukuran 3 m times 3 m dengan kedalaman 80 cm Sisa-sisa akar tanaman kelapa sawit lama dikumpulkan secara manual selama proses penumbangan dan pengolahan tanah berlangsung
Perlakuan disusun dalam rancangan acak kelompok (RAK) dengan 10 ulangan Pada masing-masing ulangan terdapat 70 tanaman sampel Pengamatan insidensi penyakit di-lakukan secara terus-menerus pada interval enam bulan
Sistem Penanaman Hole in HolePenelitian dilakukan di kebun Teluk
Dalam Sumatera Utara sejak tahun 2003 Perlakuan terdiri atas sistem penanaman hole in hole dan sistem penanaman standar sebagai pembanding Perlakuan disusun dalam RAK yang diulang dalam 17 blok penanaman Setiap blok memiliki luasan 25 ha dengan kerapatan 143 tanaman per ha Insidensi penyakit BPB diamati pada seluruh tanaman dengan interval 1 tahun
Sistem penanaman hole in hole dilakukan dengan membuat lubang tanam standar berukuran 60 cm times 60 cm dengan kedalaman 50 cm di dalam lubang tanam besar berdimensi 3 m times 3 m times 08 m sedangkan pada sistem penanaman standar hanya digunakan lubang tanam berukuran 60 cm times 60 cm Masing-masing perlakuan diberikan aplikasi sebanyak 400 g Trichoderma sp ke lubang tanam sebelum penanaman bibit kelapa sawit
Pembedahan dan Pembumbunan Tanaman Terinfeksi Ganoderma
Percobaan dilakukan pada tanaman kelapa sawit berumur 7 tahun di kebun Dalu-Dalu (Riau) kebun Bukit Sentang dan kebun Aek Pancur (Sumatera Utara) sejak 2010 Tanaman terserang Ganoderma dibedah dengan cara membuang seluruh jaringan pangkal batang yang telah mengalami pembusukan Sisa-sisa jaringan terinfeksi dari pembedahan dikumpulkan dan dimusnahkan dan jaringan pangkal batang setelah pembedahan dilapisi dengan senyawa fenolik yang berfungsi sebagai desinfektan (teer) Selanjutnya tanaman dibumbun dengan tanah yang telah
dicampur 400 g Trichoderma sp hingga menutupi seluruh jaringan tanaman yang dibedah
Perlakuan dibedakan menjadi empat yaitu pembedahan dengan aplikasi 415 mL teer dan pembumbunan (A) pembedahan dan pembumbunan (B) pembumbunan disertai aplikasi Trichoderma sp (C) dan sebagai pembanding adalah tanaman terserang tanpa perlakuan (D) Perlakuan disusun dalam RAK dengan 10 ulangan dan pada masing-masing ulangan terdapat 4ndash5 tanaman sampel
Parit IsolasiPercobaan dilaksanakan pada tanaman
kelapa sawit berumur 8 tahun di kebun Sampit Kalimantan Tengah sejak 2011 Parit isolasi individu dibuat mengelilingi tanaman ter-infeksi Ganoderma berukuran 4 m times 4 m dengan lebar dan kedalaman parit berturut-turut 40 cm dan 60 cm Sebagai pembanding ialah tanaman terinfeksi tanpa perlakuan parit individu Perlakuan disusun dalam rancangan acak kelompok dengan 64 ulangan dan pada setiap ulangan terdapat 7 tanaman sampel
HASIL
Sanitasi Sumber Inokulum Ganoderma Saat Awal Tanam Ulang
Berdasarkan hasil pengamatan sisa-sisa akar kelapa sawit yang dikumpulkan berbobot 7250 kg atau rerata 1450 kg ha-1 Insidensi penyakit BPB pada perlakuan persiapan lahan standar pada satu tahun setelah perlakuan (TSP) sudah mencapai 21 dan meningkat hingga 68 pada 2 TSP Sementara itu kelapa sawit yang memperoleh perlakuan dengan lubang tanam besar dan pengumpulan akar belum diserang oleh BPB sampai dengan 2 TSP (Gambar 1)
Sistem Penanaman Hole in HoleInsidensi penyakit BPB pada sistem tanam
standar dan hole in hole perlakuan sudah dapat diamati sejak 1 TSP dan terus meningkat hingga 10 TSP (Gambar 2) Secara umumtingkat insidensi penyakit BPB pada sistem penanaman hole in hole lebih rendah di-bandingkan dengan sistem tanam standar
3
J Fitopatol Indones Priwiratama et al
Hingga 7 TSP insidensi penyakit BPB pada sistem penanaman hole in hole meningkat dari 076ndash517 sedangkan insidensi penyakit pada sistem tanam standar meningkat dari 22ndash724 Pada 10 TSP insidensi penyakit BPB pada kedua perlakuan dijumpai pada tingkat yang relatif berimbang
Penerapan sistem penanaman hole in hole tidak memberikan pengaruh terhadap produksi tandan buah segar Hasil rerata produksi tandan buah segar per tahun selama 5 tahun terakhir tidak menunjukkan perbedaan yang nyata antara dua sistem tanam (Tabel 1) Produksi tandan buah segar per tahun berfluktuasi antara 1721ndash1924 ton ha-1 per tahun pada sistem penanaman hole in hole dan 1585 dan 1988 ton ha-1 per tahun pada sistem penanaman standar
Pembedahan dan Pembumbunan Tanaman Terinfeksi Busuk Pangkal Batang
Hingga 3 TSP tingkat kematian tanaman terinfeksi Ganoderma pada perlakuan kontrol secara nyata lebih tinggi dibandingkan de-ngan perlakuan pembedahan dengan aplikasi 415 mL teer dan pembumbunan pembedahan dan pembumbunan pembumbunan disertai aplikasi Trichoderma (Tabel 2) Untuk per-lakuan kontrol tingkat kematian tanaman terinfeksi pada 1 2 dan 3 TSP berturut-turut 233 2791 dan 3256 Sementara itu hingga 3 TSP kematian tanaman pada perlakuan A lebih rendah dibandingkan dengan perlakuan B dan C dengan tingkat kematian berturut-turut 233 465 dan 465
4
Gambar 1 Perkembangan insidensi penyakit busuk pangkal batang oleh Ganoderma boninense di kebun Sei Pencur Sumatra Utara dengan tiga perlakuan persiapan lahan standar lubang tanam besar dan pengumpulan sisa-sisa akar persiapan lahan standar dan lubang tanam besar dan persiapan lahan standar
Kej
adia
n pe
nyak
it (
) 8
6
4
2
01 2
Tahun setelah tanam
Gambar 2 Perkembangan insidensi penyakit busuk pangkal batang oleh Ganoderma boninense di kebun Teluk Dalam Sumatra Utara pada dua sistem tanam standar dan hole in hole
Kej
adia
n pe
nyak
it (
)
20
16
12
8
4
01 2 3 4 5 6 7 8 9 10
Tahun setelah tanam
J Fitopatol Indones Priwiratama et al
Parit IsolasiSecara umum insidensi penyakit BPB pada
perlakuan kontrol lebih tinggi dibandingkan dengan perlakuan parit isolasi (Tabel 3) Insidensi penyakit BPB pada perlakuan kontrol sudah teramati sejak 1 TSP (052) dan terus meningkat hingga 182 pada 3 TSP Sebaliknya pada perlakuan parit isolasi insidensi penyakit BPB baru teramati pada 3 TSP dengan tingkat yang lebih rendah (026)
PEMBAHASAN
Sanitasi sisa-sisa akar kelapa sawit saat tanam ulang memiliki peran penting untuk mencegah penularan dini G boninense Pada
sistem tanam ulang standar kelapa sawit memiliki risiko terinfeksi G boninense lebih tinggi sejak tahun pertama setelah penanaman yang dapat dilihat dari insidensi penyakit mencapai 2 Hal ini terjadi karena tingginya peluang terjadi kontak akar antara perakaran kelapa sawit sehat dan sisa-sisa akar terinfeksi yang lebih cepat pada sistem tanam ulang standar dibandingkan dengan sistem yang diujikan Virdiana et al (2012) melaporkan bahwa penanaman bibit kelapa sawit dengan jarak sekurang-kurangnya 2 m dari lubang tanam lama pada saat tanam ulang dapat menunda proses infeksi penyakit melalui kontak akar dan menekan tingkat insidensi penyakit secara nyata hingga 11 tahun setelah tanam Sebelumnya Flood et al (2005) juga telah melaporkan bahwa penanaman bibit kelapa sawit jauh dari tumpukan batang dan bonggol saat tanam ulang dapat menunda infeksi Ganoderma
Upaya menekan laju penyakit BPB dapat juga dilakukan melalui penggunaan sistem tanam hole in hole Sistem tanam ini bertujuan mengurangi sumber inokulum Ganoderma hingga seminimal mungkin pada titik pe-nanaman bibit kelapa sawit Berdasarkan percobaan jangka panjang yang dilakukan di daerah endemik penyakit BPB di kebun
5
Tabel 1 Rerata produksi tandan buah segar di kebun Teluk Dalam Sumatra Utara pada sistem tanam standar dan hole in hole
Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Perlakuan
Lubang tanam standarHole in hole
20102009 2011Rerata produksi TBS (tonhatahun)
201320121585 a1721 a
1810 a1927 a
1988 a1974 a
1924 a1853 a
1880 a1752 a
Tabel 2 Perkembangan kematian tanaman terinfeksi Ganoderma pada perlakuan pembedahan dan pembumbunan
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
PerlakuanPembedahan dengan aplikasi teer dan pembumbunanPembedahan dan pembumbunanPembumbunanKontrol
2 TSP1 TSP 3 TSPMortalitas tanaman ()
000 a000 a000 a233 a
233 a 000 a 000 a2791 b
233 a 465 a 465 a3256 b
Tabel 3 Insidensi penyakit busuk pangkal batang di kebun Sampit Kalimantan Tengah pada sistem parit isolasi
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Sistem Tanam
Parit isolasiKontrol
2 TSP1 TSP 3 TSPInsidensi penyakit ()
000 a052 a
000 a104 b
026 a182 b
J Fitopatol Indones Priwiratama et al
Teluk Dalam Sumatera Utara sistem tanam hole in hole dapat mengurangi tingkat G boninense pada masa awal penanaman kelapa sawit di lapangan Sistem penanaman hole in hole mampu menekan tingkat insidensi penyakit BPB hingga 7 TSP dibandingkan dengan sistem penanaman dengan lubang tanam standar Hal ini dapat disebabkan oleh perbedaan pola sebaran dan jangkauan akar kelapa sawit yang berbeda dengan pada kedua sistem penanaman Perakaran kelapa sawit berumur 3 tahun pada lubang tanam standar dapat menjangkau area sepanjang 467 m dengan sebaran merata sehingga meningkatkan potensi kontak dengan inokulum G boninense di lapangan Pada sistem hole in hole jangkauan akar hanya mencapai 301 m dengan pola sebaran akar terpusat di dalam lubang tanam besar sehingga memperkecil peluang terjadinya kontak akar dengan sumber inokulum Namun demikian setelah 7 TSP sistem tanam hole in hole tidak mampu menghambat laju perkembangan penyakit BPB Pada tahap ini perakaran kelapa sawit telah tumbuh berkembang melebihi area lubang sanitasi sehingga peluang terjadinya kontak dengan sumber inokulum G boninense di luar area hole in hole menjadi semakin tinggi
Penggunaan sistem hole in hole dianggap memberikan dampak negatif terhadap per-kembangan dan produktivitas kelapa sawit karena sebagian besar lapisan tanah bagian atas yang memiliki kandungan hara lebih tinggi terbuang dan hanya menyisakan lapisan tanah dalam yang miskin unsur hara Namun berdasarkan pengamatan di lapangan sistem tanam ini tidak memberikan pengaruh terhadap keragaan tanaman dan produktivitas kelapa sawit Data produksi TBS selama lima tahun terakhir (Tabel 1) memperlihatkan produktivitas kelapa sawit pada kedua sistem tanam relatif berimbang Dalam praktiknya penambahan bahan organik berupa tandan kosong kelapa sawit pada penerapan sistem hole in hole merupakan salah satu faktor penting agar asupan nutrisi kelapa sawit selama tahap awal perkembangannya dapat terpenuhi
sehingga keragaan dan produktivitasnya tidak tertinggal dari sistem tanam standar
Penerapan teknik pembedahan dan pem-bumbunan untuk mengendalikan penyakit BPB menunjukkan hasil yang cukup menjanjikan Meskipun tidak dapat menghindarkan tanaman dari infeksi G boninense pembedahan jaringan sakit yang disertai pembumbunan mampu memperpanjang masa hidup tanaman terinfeksi sekurang-kurangnya hingga 3 TSP dibandingkan dengan tanaman tanpa perlakuan Sedangkan pada tanaman dengan hanya pembumbunan mampu bertahan hingga 2 TSP Hal ini cukup penting mengingat dalam kurun waktu 2ndash3 tahun tanaman sakit masih terus memproduksi TBS Pembedahan dilaporkan dapat memperpanjang masa hidup kelapa sawit hingga 2ndash3 tahun (Ho dan Hashim 1997) Pembedahan di perkebunan rakyat mampu memperpanjang masa hidup kelapa sawit di lapangan (Marshall et al 2004 Cooper et al 2011) Berdasarkan pengamatan di lapangan tingkat keberhasilan teknik pembedahan ini sangat bergantung pada ketepatan jaringan sakit yang dibuang Bila jaringan sakit tidak seluruhnya dibuang umumnya kelapa sawit hanya akan bertahan selama satu tahun karena sisa jaringan sakit tersebut berperan sebagai sumber inokulum yang akan terus berkembang hingga menyebabkan pembusukan pada pangkal batang tanaman yang dibumbun
Teknik pengendalian kultur teknis lain yang dapat diterapkan di lapangan ialah pembuatan parit isolasi yang bertujuan mencegah terjadinya kontak akar Hasil pe-ngamatan menunjukkan bahwa parit isolasi mampu mencegah terjadinya infeksi pada tanaman tetangga di luar parit hingga 2 TSP Keberadaan parit isolasi akan mencegah terjadinya kontak akar sehingga tanaman sehat terhindar dari infeksi G boninense dari jaringan tanaman terinfeksi dalam area isolasi Meskipun cukup sulit ketepatan waktu dan titik pembuatan parit isolasi sangat mempengaruhi keefektifan pencegahan pe-nyebaran G boninense di lapangan Dengan akurasi waktu pembuatan yang baik parit isolasi berukuran 2 m times 2 m dilaporkan mampu
6
J Fitopatol Indones Priwiratama et al
mencegah penularan penyakit BPB hingga 14 tahun (Hasan dan Turner 1998 Chung 2011)
Dari uraian di atas dapat diketahui bahwa tujuan pengendalian penyakit BPB secara kultur teknis dititikberatkan untuk menekan adanya sumber inokulum potensial sehingga dapat mencegah terjadinya infeksi G boninense di lapangan Metode pengendalian kultur teknis yang digunakan dapat disesuaikan dengan tahap perkembangan kelapa sawit Di areal pengembangan atau penanaman baru pengendalian dilakukan sejak tahap awal penanaman melalui sanitasi sumber inokulum terutama sisa-sisa perakaran kelapa sawit dan penggunaan lubang tanam besar Sistem penanaman hole in hole dapat diterapkan untuk melakukan penyisipan tanaman baru terutama di areal endemik G boninense Sementara itu untuk tanaman terserang dapat dilakukan tindakan pembedahan diiringi pembumbunan dan bila sudah tidak memungkinkan dapat dilakukan isolasi tanaman terserang untuk mencegah penularan penyakit pada tanaman sehat di sekitarnya Beberapa penelitian di atas akan terus dilanjutkan untuk melihat pengaruh jangka panjang dari teknik-teknik yang diterapkan
DAFTAR PUSTAKA
Chung GF 2011 Management of Ganoderma diseases in oil palm plantations Planter 87(1022)325ndash339
Cooper RM Flood J Rees R 2011 Ganoderma boninense in oil palm plantations current thinking on epidemiology resistance and pathology Planter 87(1024)515ndash526
Durand-Gasselin T Asmady H Flori A Jacquemard Jc Hayun Z Breton F De Franqueville H 2005 Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq) to basal stem rot caused by Ganoderma boninensemdashprospects for future breeding Mycopathologia 159(1)93ndash100 DOI httpdxdoiorg10 1007s11046-004-4429-1
Flood J Keenan L Wayne S Hasan Y 2005 Studies on oil palm trunks as sources of infection in the field Mycopathologia
159(1)101ndash107 DOI httpdxdoiorg 101007s11046-004-4430-8
Hasan Y Turner P 1998 The comparative importance of different oil palm tissues as infection sources for basal stem rot in replantings Planter 74(864)119ndash135
Ho CT Hashim K 1997 Usefulness of soil mounding treatments in prolonging productivity of prime-aged Ganoderma infected palms Planter 73(854)239ndash244
Idris AS Kushairi A Ismail S Ariffin D 2004 Selection for partial tolerance in oil palm progenies to Ganoderma basal stem rot J Oil Palm Res 16(2)12ndash18
Kartika E Yahya S Wilarso S 2006 Isolasi karakterisasi dan pemurnian cendawan mikoriza arbuskular dari dua lokasi perkebunan kelapa sawit (bekas hutan dan bekas kebun karet) J Penelitian Kelapa Sawit 14(3)145ndash155
Marshall R Hunt R Pilotti C 2004 Low cost control for basal stem rotmdasha Poliamba initiative Planter 80(936)173ndash176
Naher L Yusuf UK Tan SG Ismail A 2013 Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq) Aus Sci 7(11)1723ndash1727
Paterson RRM 2007 Ganoderma disease of oil palmmdasha white rot perspective necessary for integrated control Crop Protect 26(2007)1369ndash1376 DOI httpdxdoiorg101016jcropro200611009
Priwiratama H Susanto A 2014 Utilization of fungi for the biological control of insect pests and Ganoderma disease in the Indonesian oil palm industry Agr Sci Tech A 4(2014)103ndash111
Susanto A Prasetyo AE Wening S 2013 Laju infeksi Ganoderma pada empat kelas tekstur tanah J Fitopatol Indones 9(2)39-46 DOI httpdxdoiorg1014692jfi9239
Virdiana I Flood J Sitepu B Hasan Y Aditya R Nelson S 2012 Integrated disease management to reduce future Ganoderma infection during oil palm replanting Planter 88(1305)383ndash393
7
Volume 10 Nomor 1 Februari 2014Halaman 8ndash16
DOI 1014692jfi1018
Pengelolaan Penyakit Kuning pada Tanaman Lada oleh Petani di Wilayah Bangka
Management of Yellow Diseases on Pepper by Farmers in Bangka Region
Abdul Munif Ita SulistiawatiInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit kuning pada tanaman lada yang disebabkan oleh nematoda parasit tumbuhan hingga kini masih menjadi masalah utama pada pertanaman lada di wilayah Bangka Kehilangan hasil berupa penurunan produksi lada yang diakibatkan oleh penyakit kuning cukup besar Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan Penelitian ini meliputi dua kegiatan yaitu survei tingkat kejadian penyakit kuning pada pertanaman lada di lapangan dan wawancara dengan petani lada Hasil survei menunjukkan bahwa tingkat kejadian penyakit kuning sudah merata dan menyerang hampir semua lahan yang diamati Hampir semua petani responden menyatakan bahwa penyakit kuning merupakan penyakit utama yang sangat merusak pertanaman lada Tingkat pemahaman petani terhadap penyebab gejala dan ekologi penyakit kuning dan cara-cara pengendaliannya masih sangat terbatas Tindakan pengendalian yang dilakukan oleh petani terhadap penyakit kuning ialah pencabutan dan pembakaran tanaman terserang dan pemberian kapur Beberapa petani tidak melakukan tindakan pengendalian namun sebagian besar petani menggunakan pestisida Penggunaan pestisida untuk pengendalian penyakit kuning oleh petani belum sepenuhnya mengikuti prinsip pengendalian hama terpadu Masih rendahnya pemahaman petani terhadap penyakit kuning mengakibatkan semakin sulit bagi petani untuk melakukan pengendaliannya
Kata kunci nematoda pengendalian penyakit pestisida Piper nigrum
ABSTRACT
Yellow disease caused by plant-parasitic nematodes are still a major problem on pepper production area in Bangka region Significant yield losses occurred due to yellow disease The objective of this research was to survey the yellow disease incidence and to study farmerrsquos knowledge attitude and practices on the management of yellow disease of black pepper in Bangka district Central Bangka and South Bangka This study consisted of a survey of the incidence of yellow disease in the field and interviews to the farmers The result showed that the yellow disease had infected nearly all of black pepper fields in the areas with incidence of as much as 20ndash60 Almost all of the respondents reported that the yellow disease was the major disease of black pepper in Bangka region Farmersrsquo level of knowledge about the causal of disease symptoms ecology and control methods of the yellow disease are still very limited Various control measures have been done by farmers to combat the disease especially by removing and burning the infected plants or by application of lime A few farmers did not even controlling the disease Pesticides have been used by a few farmers to control the yellow disease but the application of pesticide does not in line with integrated pest management principles
Key words disease control nematode pesticide Piper nigrumAlamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor
Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel abdulmunifipbacid
8
ISSN 2339-2479
J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Priwiratama et al
kedalaman 50 cm Pada perlakuan A dan B lubang tanam besar yang digunakan berukuran 3 m times 3 m dengan kedalaman 80 cm Sisa-sisa akar tanaman kelapa sawit lama dikumpulkan secara manual selama proses penumbangan dan pengolahan tanah berlangsung
Perlakuan disusun dalam rancangan acak kelompok (RAK) dengan 10 ulangan Pada masing-masing ulangan terdapat 70 tanaman sampel Pengamatan insidensi penyakit di-lakukan secara terus-menerus pada interval enam bulan
Sistem Penanaman Hole in HolePenelitian dilakukan di kebun Teluk
Dalam Sumatera Utara sejak tahun 2003 Perlakuan terdiri atas sistem penanaman hole in hole dan sistem penanaman standar sebagai pembanding Perlakuan disusun dalam RAK yang diulang dalam 17 blok penanaman Setiap blok memiliki luasan 25 ha dengan kerapatan 143 tanaman per ha Insidensi penyakit BPB diamati pada seluruh tanaman dengan interval 1 tahun
Sistem penanaman hole in hole dilakukan dengan membuat lubang tanam standar berukuran 60 cm times 60 cm dengan kedalaman 50 cm di dalam lubang tanam besar berdimensi 3 m times 3 m times 08 m sedangkan pada sistem penanaman standar hanya digunakan lubang tanam berukuran 60 cm times 60 cm Masing-masing perlakuan diberikan aplikasi sebanyak 400 g Trichoderma sp ke lubang tanam sebelum penanaman bibit kelapa sawit
Pembedahan dan Pembumbunan Tanaman Terinfeksi Ganoderma
Percobaan dilakukan pada tanaman kelapa sawit berumur 7 tahun di kebun Dalu-Dalu (Riau) kebun Bukit Sentang dan kebun Aek Pancur (Sumatera Utara) sejak 2010 Tanaman terserang Ganoderma dibedah dengan cara membuang seluruh jaringan pangkal batang yang telah mengalami pembusukan Sisa-sisa jaringan terinfeksi dari pembedahan dikumpulkan dan dimusnahkan dan jaringan pangkal batang setelah pembedahan dilapisi dengan senyawa fenolik yang berfungsi sebagai desinfektan (teer) Selanjutnya tanaman dibumbun dengan tanah yang telah
dicampur 400 g Trichoderma sp hingga menutupi seluruh jaringan tanaman yang dibedah
Perlakuan dibedakan menjadi empat yaitu pembedahan dengan aplikasi 415 mL teer dan pembumbunan (A) pembedahan dan pembumbunan (B) pembumbunan disertai aplikasi Trichoderma sp (C) dan sebagai pembanding adalah tanaman terserang tanpa perlakuan (D) Perlakuan disusun dalam RAK dengan 10 ulangan dan pada masing-masing ulangan terdapat 4ndash5 tanaman sampel
Parit IsolasiPercobaan dilaksanakan pada tanaman
kelapa sawit berumur 8 tahun di kebun Sampit Kalimantan Tengah sejak 2011 Parit isolasi individu dibuat mengelilingi tanaman ter-infeksi Ganoderma berukuran 4 m times 4 m dengan lebar dan kedalaman parit berturut-turut 40 cm dan 60 cm Sebagai pembanding ialah tanaman terinfeksi tanpa perlakuan parit individu Perlakuan disusun dalam rancangan acak kelompok dengan 64 ulangan dan pada setiap ulangan terdapat 7 tanaman sampel
HASIL
Sanitasi Sumber Inokulum Ganoderma Saat Awal Tanam Ulang
Berdasarkan hasil pengamatan sisa-sisa akar kelapa sawit yang dikumpulkan berbobot 7250 kg atau rerata 1450 kg ha-1 Insidensi penyakit BPB pada perlakuan persiapan lahan standar pada satu tahun setelah perlakuan (TSP) sudah mencapai 21 dan meningkat hingga 68 pada 2 TSP Sementara itu kelapa sawit yang memperoleh perlakuan dengan lubang tanam besar dan pengumpulan akar belum diserang oleh BPB sampai dengan 2 TSP (Gambar 1)
Sistem Penanaman Hole in HoleInsidensi penyakit BPB pada sistem tanam
standar dan hole in hole perlakuan sudah dapat diamati sejak 1 TSP dan terus meningkat hingga 10 TSP (Gambar 2) Secara umumtingkat insidensi penyakit BPB pada sistem penanaman hole in hole lebih rendah di-bandingkan dengan sistem tanam standar
3
J Fitopatol Indones Priwiratama et al
Hingga 7 TSP insidensi penyakit BPB pada sistem penanaman hole in hole meningkat dari 076ndash517 sedangkan insidensi penyakit pada sistem tanam standar meningkat dari 22ndash724 Pada 10 TSP insidensi penyakit BPB pada kedua perlakuan dijumpai pada tingkat yang relatif berimbang
Penerapan sistem penanaman hole in hole tidak memberikan pengaruh terhadap produksi tandan buah segar Hasil rerata produksi tandan buah segar per tahun selama 5 tahun terakhir tidak menunjukkan perbedaan yang nyata antara dua sistem tanam (Tabel 1) Produksi tandan buah segar per tahun berfluktuasi antara 1721ndash1924 ton ha-1 per tahun pada sistem penanaman hole in hole dan 1585 dan 1988 ton ha-1 per tahun pada sistem penanaman standar
Pembedahan dan Pembumbunan Tanaman Terinfeksi Busuk Pangkal Batang
Hingga 3 TSP tingkat kematian tanaman terinfeksi Ganoderma pada perlakuan kontrol secara nyata lebih tinggi dibandingkan de-ngan perlakuan pembedahan dengan aplikasi 415 mL teer dan pembumbunan pembedahan dan pembumbunan pembumbunan disertai aplikasi Trichoderma (Tabel 2) Untuk per-lakuan kontrol tingkat kematian tanaman terinfeksi pada 1 2 dan 3 TSP berturut-turut 233 2791 dan 3256 Sementara itu hingga 3 TSP kematian tanaman pada perlakuan A lebih rendah dibandingkan dengan perlakuan B dan C dengan tingkat kematian berturut-turut 233 465 dan 465
4
Gambar 1 Perkembangan insidensi penyakit busuk pangkal batang oleh Ganoderma boninense di kebun Sei Pencur Sumatra Utara dengan tiga perlakuan persiapan lahan standar lubang tanam besar dan pengumpulan sisa-sisa akar persiapan lahan standar dan lubang tanam besar dan persiapan lahan standar
Kej
adia
n pe
nyak
it (
) 8
6
4
2
01 2
Tahun setelah tanam
Gambar 2 Perkembangan insidensi penyakit busuk pangkal batang oleh Ganoderma boninense di kebun Teluk Dalam Sumatra Utara pada dua sistem tanam standar dan hole in hole
Kej
adia
n pe
nyak
it (
)
20
16
12
8
4
01 2 3 4 5 6 7 8 9 10
Tahun setelah tanam
J Fitopatol Indones Priwiratama et al
Parit IsolasiSecara umum insidensi penyakit BPB pada
perlakuan kontrol lebih tinggi dibandingkan dengan perlakuan parit isolasi (Tabel 3) Insidensi penyakit BPB pada perlakuan kontrol sudah teramati sejak 1 TSP (052) dan terus meningkat hingga 182 pada 3 TSP Sebaliknya pada perlakuan parit isolasi insidensi penyakit BPB baru teramati pada 3 TSP dengan tingkat yang lebih rendah (026)
PEMBAHASAN
Sanitasi sisa-sisa akar kelapa sawit saat tanam ulang memiliki peran penting untuk mencegah penularan dini G boninense Pada
sistem tanam ulang standar kelapa sawit memiliki risiko terinfeksi G boninense lebih tinggi sejak tahun pertama setelah penanaman yang dapat dilihat dari insidensi penyakit mencapai 2 Hal ini terjadi karena tingginya peluang terjadi kontak akar antara perakaran kelapa sawit sehat dan sisa-sisa akar terinfeksi yang lebih cepat pada sistem tanam ulang standar dibandingkan dengan sistem yang diujikan Virdiana et al (2012) melaporkan bahwa penanaman bibit kelapa sawit dengan jarak sekurang-kurangnya 2 m dari lubang tanam lama pada saat tanam ulang dapat menunda proses infeksi penyakit melalui kontak akar dan menekan tingkat insidensi penyakit secara nyata hingga 11 tahun setelah tanam Sebelumnya Flood et al (2005) juga telah melaporkan bahwa penanaman bibit kelapa sawit jauh dari tumpukan batang dan bonggol saat tanam ulang dapat menunda infeksi Ganoderma
Upaya menekan laju penyakit BPB dapat juga dilakukan melalui penggunaan sistem tanam hole in hole Sistem tanam ini bertujuan mengurangi sumber inokulum Ganoderma hingga seminimal mungkin pada titik pe-nanaman bibit kelapa sawit Berdasarkan percobaan jangka panjang yang dilakukan di daerah endemik penyakit BPB di kebun
5
Tabel 1 Rerata produksi tandan buah segar di kebun Teluk Dalam Sumatra Utara pada sistem tanam standar dan hole in hole
Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Perlakuan
Lubang tanam standarHole in hole
20102009 2011Rerata produksi TBS (tonhatahun)
201320121585 a1721 a
1810 a1927 a
1988 a1974 a
1924 a1853 a
1880 a1752 a
Tabel 2 Perkembangan kematian tanaman terinfeksi Ganoderma pada perlakuan pembedahan dan pembumbunan
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
PerlakuanPembedahan dengan aplikasi teer dan pembumbunanPembedahan dan pembumbunanPembumbunanKontrol
2 TSP1 TSP 3 TSPMortalitas tanaman ()
000 a000 a000 a233 a
233 a 000 a 000 a2791 b
233 a 465 a 465 a3256 b
Tabel 3 Insidensi penyakit busuk pangkal batang di kebun Sampit Kalimantan Tengah pada sistem parit isolasi
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Sistem Tanam
Parit isolasiKontrol
2 TSP1 TSP 3 TSPInsidensi penyakit ()
000 a052 a
000 a104 b
026 a182 b
J Fitopatol Indones Priwiratama et al
Teluk Dalam Sumatera Utara sistem tanam hole in hole dapat mengurangi tingkat G boninense pada masa awal penanaman kelapa sawit di lapangan Sistem penanaman hole in hole mampu menekan tingkat insidensi penyakit BPB hingga 7 TSP dibandingkan dengan sistem penanaman dengan lubang tanam standar Hal ini dapat disebabkan oleh perbedaan pola sebaran dan jangkauan akar kelapa sawit yang berbeda dengan pada kedua sistem penanaman Perakaran kelapa sawit berumur 3 tahun pada lubang tanam standar dapat menjangkau area sepanjang 467 m dengan sebaran merata sehingga meningkatkan potensi kontak dengan inokulum G boninense di lapangan Pada sistem hole in hole jangkauan akar hanya mencapai 301 m dengan pola sebaran akar terpusat di dalam lubang tanam besar sehingga memperkecil peluang terjadinya kontak akar dengan sumber inokulum Namun demikian setelah 7 TSP sistem tanam hole in hole tidak mampu menghambat laju perkembangan penyakit BPB Pada tahap ini perakaran kelapa sawit telah tumbuh berkembang melebihi area lubang sanitasi sehingga peluang terjadinya kontak dengan sumber inokulum G boninense di luar area hole in hole menjadi semakin tinggi
Penggunaan sistem hole in hole dianggap memberikan dampak negatif terhadap per-kembangan dan produktivitas kelapa sawit karena sebagian besar lapisan tanah bagian atas yang memiliki kandungan hara lebih tinggi terbuang dan hanya menyisakan lapisan tanah dalam yang miskin unsur hara Namun berdasarkan pengamatan di lapangan sistem tanam ini tidak memberikan pengaruh terhadap keragaan tanaman dan produktivitas kelapa sawit Data produksi TBS selama lima tahun terakhir (Tabel 1) memperlihatkan produktivitas kelapa sawit pada kedua sistem tanam relatif berimbang Dalam praktiknya penambahan bahan organik berupa tandan kosong kelapa sawit pada penerapan sistem hole in hole merupakan salah satu faktor penting agar asupan nutrisi kelapa sawit selama tahap awal perkembangannya dapat terpenuhi
sehingga keragaan dan produktivitasnya tidak tertinggal dari sistem tanam standar
Penerapan teknik pembedahan dan pem-bumbunan untuk mengendalikan penyakit BPB menunjukkan hasil yang cukup menjanjikan Meskipun tidak dapat menghindarkan tanaman dari infeksi G boninense pembedahan jaringan sakit yang disertai pembumbunan mampu memperpanjang masa hidup tanaman terinfeksi sekurang-kurangnya hingga 3 TSP dibandingkan dengan tanaman tanpa perlakuan Sedangkan pada tanaman dengan hanya pembumbunan mampu bertahan hingga 2 TSP Hal ini cukup penting mengingat dalam kurun waktu 2ndash3 tahun tanaman sakit masih terus memproduksi TBS Pembedahan dilaporkan dapat memperpanjang masa hidup kelapa sawit hingga 2ndash3 tahun (Ho dan Hashim 1997) Pembedahan di perkebunan rakyat mampu memperpanjang masa hidup kelapa sawit di lapangan (Marshall et al 2004 Cooper et al 2011) Berdasarkan pengamatan di lapangan tingkat keberhasilan teknik pembedahan ini sangat bergantung pada ketepatan jaringan sakit yang dibuang Bila jaringan sakit tidak seluruhnya dibuang umumnya kelapa sawit hanya akan bertahan selama satu tahun karena sisa jaringan sakit tersebut berperan sebagai sumber inokulum yang akan terus berkembang hingga menyebabkan pembusukan pada pangkal batang tanaman yang dibumbun
Teknik pengendalian kultur teknis lain yang dapat diterapkan di lapangan ialah pembuatan parit isolasi yang bertujuan mencegah terjadinya kontak akar Hasil pe-ngamatan menunjukkan bahwa parit isolasi mampu mencegah terjadinya infeksi pada tanaman tetangga di luar parit hingga 2 TSP Keberadaan parit isolasi akan mencegah terjadinya kontak akar sehingga tanaman sehat terhindar dari infeksi G boninense dari jaringan tanaman terinfeksi dalam area isolasi Meskipun cukup sulit ketepatan waktu dan titik pembuatan parit isolasi sangat mempengaruhi keefektifan pencegahan pe-nyebaran G boninense di lapangan Dengan akurasi waktu pembuatan yang baik parit isolasi berukuran 2 m times 2 m dilaporkan mampu
6
J Fitopatol Indones Priwiratama et al
mencegah penularan penyakit BPB hingga 14 tahun (Hasan dan Turner 1998 Chung 2011)
Dari uraian di atas dapat diketahui bahwa tujuan pengendalian penyakit BPB secara kultur teknis dititikberatkan untuk menekan adanya sumber inokulum potensial sehingga dapat mencegah terjadinya infeksi G boninense di lapangan Metode pengendalian kultur teknis yang digunakan dapat disesuaikan dengan tahap perkembangan kelapa sawit Di areal pengembangan atau penanaman baru pengendalian dilakukan sejak tahap awal penanaman melalui sanitasi sumber inokulum terutama sisa-sisa perakaran kelapa sawit dan penggunaan lubang tanam besar Sistem penanaman hole in hole dapat diterapkan untuk melakukan penyisipan tanaman baru terutama di areal endemik G boninense Sementara itu untuk tanaman terserang dapat dilakukan tindakan pembedahan diiringi pembumbunan dan bila sudah tidak memungkinkan dapat dilakukan isolasi tanaman terserang untuk mencegah penularan penyakit pada tanaman sehat di sekitarnya Beberapa penelitian di atas akan terus dilanjutkan untuk melihat pengaruh jangka panjang dari teknik-teknik yang diterapkan
DAFTAR PUSTAKA
Chung GF 2011 Management of Ganoderma diseases in oil palm plantations Planter 87(1022)325ndash339
Cooper RM Flood J Rees R 2011 Ganoderma boninense in oil palm plantations current thinking on epidemiology resistance and pathology Planter 87(1024)515ndash526
Durand-Gasselin T Asmady H Flori A Jacquemard Jc Hayun Z Breton F De Franqueville H 2005 Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq) to basal stem rot caused by Ganoderma boninensemdashprospects for future breeding Mycopathologia 159(1)93ndash100 DOI httpdxdoiorg10 1007s11046-004-4429-1
Flood J Keenan L Wayne S Hasan Y 2005 Studies on oil palm trunks as sources of infection in the field Mycopathologia
159(1)101ndash107 DOI httpdxdoiorg 101007s11046-004-4430-8
Hasan Y Turner P 1998 The comparative importance of different oil palm tissues as infection sources for basal stem rot in replantings Planter 74(864)119ndash135
Ho CT Hashim K 1997 Usefulness of soil mounding treatments in prolonging productivity of prime-aged Ganoderma infected palms Planter 73(854)239ndash244
Idris AS Kushairi A Ismail S Ariffin D 2004 Selection for partial tolerance in oil palm progenies to Ganoderma basal stem rot J Oil Palm Res 16(2)12ndash18
Kartika E Yahya S Wilarso S 2006 Isolasi karakterisasi dan pemurnian cendawan mikoriza arbuskular dari dua lokasi perkebunan kelapa sawit (bekas hutan dan bekas kebun karet) J Penelitian Kelapa Sawit 14(3)145ndash155
Marshall R Hunt R Pilotti C 2004 Low cost control for basal stem rotmdasha Poliamba initiative Planter 80(936)173ndash176
Naher L Yusuf UK Tan SG Ismail A 2013 Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq) Aus Sci 7(11)1723ndash1727
Paterson RRM 2007 Ganoderma disease of oil palmmdasha white rot perspective necessary for integrated control Crop Protect 26(2007)1369ndash1376 DOI httpdxdoiorg101016jcropro200611009
Priwiratama H Susanto A 2014 Utilization of fungi for the biological control of insect pests and Ganoderma disease in the Indonesian oil palm industry Agr Sci Tech A 4(2014)103ndash111
Susanto A Prasetyo AE Wening S 2013 Laju infeksi Ganoderma pada empat kelas tekstur tanah J Fitopatol Indones 9(2)39-46 DOI httpdxdoiorg1014692jfi9239
Virdiana I Flood J Sitepu B Hasan Y Aditya R Nelson S 2012 Integrated disease management to reduce future Ganoderma infection during oil palm replanting Planter 88(1305)383ndash393
7
Volume 10 Nomor 1 Februari 2014Halaman 8ndash16
DOI 1014692jfi1018
Pengelolaan Penyakit Kuning pada Tanaman Lada oleh Petani di Wilayah Bangka
Management of Yellow Diseases on Pepper by Farmers in Bangka Region
Abdul Munif Ita SulistiawatiInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit kuning pada tanaman lada yang disebabkan oleh nematoda parasit tumbuhan hingga kini masih menjadi masalah utama pada pertanaman lada di wilayah Bangka Kehilangan hasil berupa penurunan produksi lada yang diakibatkan oleh penyakit kuning cukup besar Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan Penelitian ini meliputi dua kegiatan yaitu survei tingkat kejadian penyakit kuning pada pertanaman lada di lapangan dan wawancara dengan petani lada Hasil survei menunjukkan bahwa tingkat kejadian penyakit kuning sudah merata dan menyerang hampir semua lahan yang diamati Hampir semua petani responden menyatakan bahwa penyakit kuning merupakan penyakit utama yang sangat merusak pertanaman lada Tingkat pemahaman petani terhadap penyebab gejala dan ekologi penyakit kuning dan cara-cara pengendaliannya masih sangat terbatas Tindakan pengendalian yang dilakukan oleh petani terhadap penyakit kuning ialah pencabutan dan pembakaran tanaman terserang dan pemberian kapur Beberapa petani tidak melakukan tindakan pengendalian namun sebagian besar petani menggunakan pestisida Penggunaan pestisida untuk pengendalian penyakit kuning oleh petani belum sepenuhnya mengikuti prinsip pengendalian hama terpadu Masih rendahnya pemahaman petani terhadap penyakit kuning mengakibatkan semakin sulit bagi petani untuk melakukan pengendaliannya
Kata kunci nematoda pengendalian penyakit pestisida Piper nigrum
ABSTRACT
Yellow disease caused by plant-parasitic nematodes are still a major problem on pepper production area in Bangka region Significant yield losses occurred due to yellow disease The objective of this research was to survey the yellow disease incidence and to study farmerrsquos knowledge attitude and practices on the management of yellow disease of black pepper in Bangka district Central Bangka and South Bangka This study consisted of a survey of the incidence of yellow disease in the field and interviews to the farmers The result showed that the yellow disease had infected nearly all of black pepper fields in the areas with incidence of as much as 20ndash60 Almost all of the respondents reported that the yellow disease was the major disease of black pepper in Bangka region Farmersrsquo level of knowledge about the causal of disease symptoms ecology and control methods of the yellow disease are still very limited Various control measures have been done by farmers to combat the disease especially by removing and burning the infected plants or by application of lime A few farmers did not even controlling the disease Pesticides have been used by a few farmers to control the yellow disease but the application of pesticide does not in line with integrated pest management principles
Key words disease control nematode pesticide Piper nigrumAlamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor
Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel abdulmunifipbacid
8
ISSN 2339-2479
J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-
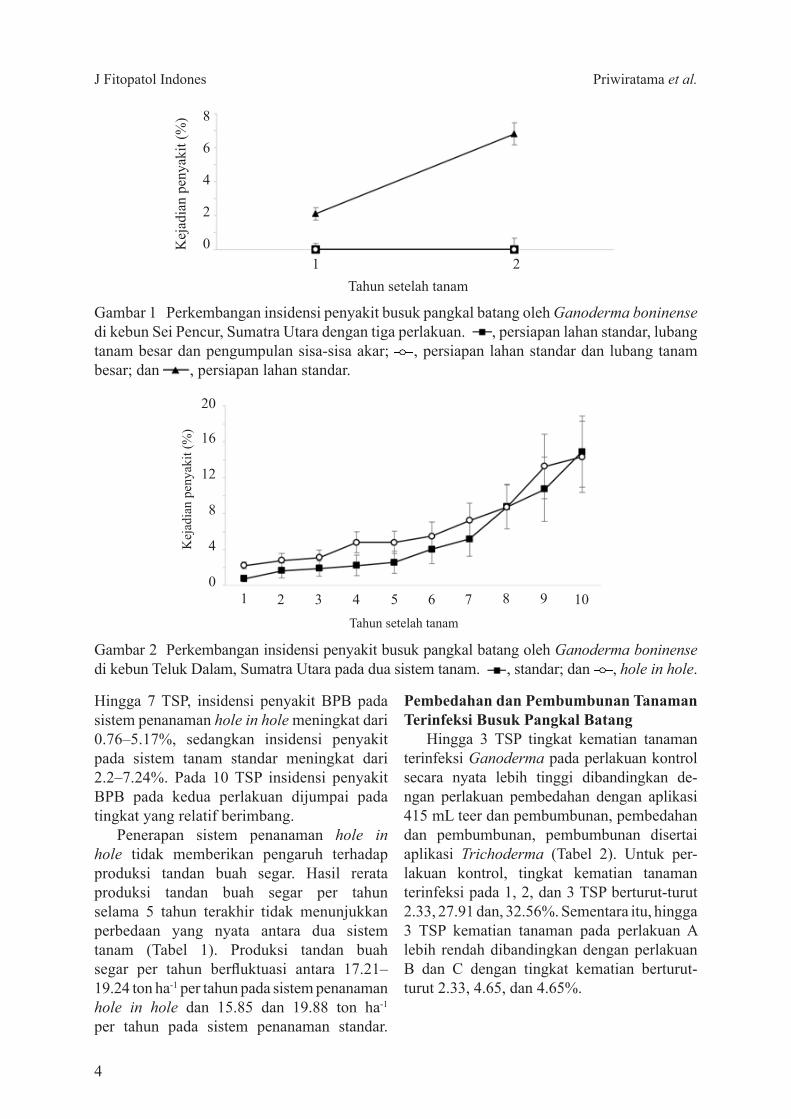
J Fitopatol Indones Priwiratama et al
Hingga 7 TSP insidensi penyakit BPB pada sistem penanaman hole in hole meningkat dari 076ndash517 sedangkan insidensi penyakit pada sistem tanam standar meningkat dari 22ndash724 Pada 10 TSP insidensi penyakit BPB pada kedua perlakuan dijumpai pada tingkat yang relatif berimbang
Penerapan sistem penanaman hole in hole tidak memberikan pengaruh terhadap produksi tandan buah segar Hasil rerata produksi tandan buah segar per tahun selama 5 tahun terakhir tidak menunjukkan perbedaan yang nyata antara dua sistem tanam (Tabel 1) Produksi tandan buah segar per tahun berfluktuasi antara 1721ndash1924 ton ha-1 per tahun pada sistem penanaman hole in hole dan 1585 dan 1988 ton ha-1 per tahun pada sistem penanaman standar
Pembedahan dan Pembumbunan Tanaman Terinfeksi Busuk Pangkal Batang
Hingga 3 TSP tingkat kematian tanaman terinfeksi Ganoderma pada perlakuan kontrol secara nyata lebih tinggi dibandingkan de-ngan perlakuan pembedahan dengan aplikasi 415 mL teer dan pembumbunan pembedahan dan pembumbunan pembumbunan disertai aplikasi Trichoderma (Tabel 2) Untuk per-lakuan kontrol tingkat kematian tanaman terinfeksi pada 1 2 dan 3 TSP berturut-turut 233 2791 dan 3256 Sementara itu hingga 3 TSP kematian tanaman pada perlakuan A lebih rendah dibandingkan dengan perlakuan B dan C dengan tingkat kematian berturut-turut 233 465 dan 465
4
Gambar 1 Perkembangan insidensi penyakit busuk pangkal batang oleh Ganoderma boninense di kebun Sei Pencur Sumatra Utara dengan tiga perlakuan persiapan lahan standar lubang tanam besar dan pengumpulan sisa-sisa akar persiapan lahan standar dan lubang tanam besar dan persiapan lahan standar
Kej
adia
n pe
nyak
it (
) 8
6
4
2
01 2
Tahun setelah tanam
Gambar 2 Perkembangan insidensi penyakit busuk pangkal batang oleh Ganoderma boninense di kebun Teluk Dalam Sumatra Utara pada dua sistem tanam standar dan hole in hole
Kej
adia
n pe
nyak
it (
)
20
16
12
8
4
01 2 3 4 5 6 7 8 9 10
Tahun setelah tanam
J Fitopatol Indones Priwiratama et al
Parit IsolasiSecara umum insidensi penyakit BPB pada
perlakuan kontrol lebih tinggi dibandingkan dengan perlakuan parit isolasi (Tabel 3) Insidensi penyakit BPB pada perlakuan kontrol sudah teramati sejak 1 TSP (052) dan terus meningkat hingga 182 pada 3 TSP Sebaliknya pada perlakuan parit isolasi insidensi penyakit BPB baru teramati pada 3 TSP dengan tingkat yang lebih rendah (026)
PEMBAHASAN
Sanitasi sisa-sisa akar kelapa sawit saat tanam ulang memiliki peran penting untuk mencegah penularan dini G boninense Pada
sistem tanam ulang standar kelapa sawit memiliki risiko terinfeksi G boninense lebih tinggi sejak tahun pertama setelah penanaman yang dapat dilihat dari insidensi penyakit mencapai 2 Hal ini terjadi karena tingginya peluang terjadi kontak akar antara perakaran kelapa sawit sehat dan sisa-sisa akar terinfeksi yang lebih cepat pada sistem tanam ulang standar dibandingkan dengan sistem yang diujikan Virdiana et al (2012) melaporkan bahwa penanaman bibit kelapa sawit dengan jarak sekurang-kurangnya 2 m dari lubang tanam lama pada saat tanam ulang dapat menunda proses infeksi penyakit melalui kontak akar dan menekan tingkat insidensi penyakit secara nyata hingga 11 tahun setelah tanam Sebelumnya Flood et al (2005) juga telah melaporkan bahwa penanaman bibit kelapa sawit jauh dari tumpukan batang dan bonggol saat tanam ulang dapat menunda infeksi Ganoderma
Upaya menekan laju penyakit BPB dapat juga dilakukan melalui penggunaan sistem tanam hole in hole Sistem tanam ini bertujuan mengurangi sumber inokulum Ganoderma hingga seminimal mungkin pada titik pe-nanaman bibit kelapa sawit Berdasarkan percobaan jangka panjang yang dilakukan di daerah endemik penyakit BPB di kebun
5
Tabel 1 Rerata produksi tandan buah segar di kebun Teluk Dalam Sumatra Utara pada sistem tanam standar dan hole in hole
Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Perlakuan
Lubang tanam standarHole in hole
20102009 2011Rerata produksi TBS (tonhatahun)
201320121585 a1721 a
1810 a1927 a
1988 a1974 a
1924 a1853 a
1880 a1752 a
Tabel 2 Perkembangan kematian tanaman terinfeksi Ganoderma pada perlakuan pembedahan dan pembumbunan
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
PerlakuanPembedahan dengan aplikasi teer dan pembumbunanPembedahan dan pembumbunanPembumbunanKontrol
2 TSP1 TSP 3 TSPMortalitas tanaman ()
000 a000 a000 a233 a
233 a 000 a 000 a2791 b
233 a 465 a 465 a3256 b
Tabel 3 Insidensi penyakit busuk pangkal batang di kebun Sampit Kalimantan Tengah pada sistem parit isolasi
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Sistem Tanam
Parit isolasiKontrol
2 TSP1 TSP 3 TSPInsidensi penyakit ()
000 a052 a
000 a104 b
026 a182 b
J Fitopatol Indones Priwiratama et al
Teluk Dalam Sumatera Utara sistem tanam hole in hole dapat mengurangi tingkat G boninense pada masa awal penanaman kelapa sawit di lapangan Sistem penanaman hole in hole mampu menekan tingkat insidensi penyakit BPB hingga 7 TSP dibandingkan dengan sistem penanaman dengan lubang tanam standar Hal ini dapat disebabkan oleh perbedaan pola sebaran dan jangkauan akar kelapa sawit yang berbeda dengan pada kedua sistem penanaman Perakaran kelapa sawit berumur 3 tahun pada lubang tanam standar dapat menjangkau area sepanjang 467 m dengan sebaran merata sehingga meningkatkan potensi kontak dengan inokulum G boninense di lapangan Pada sistem hole in hole jangkauan akar hanya mencapai 301 m dengan pola sebaran akar terpusat di dalam lubang tanam besar sehingga memperkecil peluang terjadinya kontak akar dengan sumber inokulum Namun demikian setelah 7 TSP sistem tanam hole in hole tidak mampu menghambat laju perkembangan penyakit BPB Pada tahap ini perakaran kelapa sawit telah tumbuh berkembang melebihi area lubang sanitasi sehingga peluang terjadinya kontak dengan sumber inokulum G boninense di luar area hole in hole menjadi semakin tinggi
Penggunaan sistem hole in hole dianggap memberikan dampak negatif terhadap per-kembangan dan produktivitas kelapa sawit karena sebagian besar lapisan tanah bagian atas yang memiliki kandungan hara lebih tinggi terbuang dan hanya menyisakan lapisan tanah dalam yang miskin unsur hara Namun berdasarkan pengamatan di lapangan sistem tanam ini tidak memberikan pengaruh terhadap keragaan tanaman dan produktivitas kelapa sawit Data produksi TBS selama lima tahun terakhir (Tabel 1) memperlihatkan produktivitas kelapa sawit pada kedua sistem tanam relatif berimbang Dalam praktiknya penambahan bahan organik berupa tandan kosong kelapa sawit pada penerapan sistem hole in hole merupakan salah satu faktor penting agar asupan nutrisi kelapa sawit selama tahap awal perkembangannya dapat terpenuhi
sehingga keragaan dan produktivitasnya tidak tertinggal dari sistem tanam standar
Penerapan teknik pembedahan dan pem-bumbunan untuk mengendalikan penyakit BPB menunjukkan hasil yang cukup menjanjikan Meskipun tidak dapat menghindarkan tanaman dari infeksi G boninense pembedahan jaringan sakit yang disertai pembumbunan mampu memperpanjang masa hidup tanaman terinfeksi sekurang-kurangnya hingga 3 TSP dibandingkan dengan tanaman tanpa perlakuan Sedangkan pada tanaman dengan hanya pembumbunan mampu bertahan hingga 2 TSP Hal ini cukup penting mengingat dalam kurun waktu 2ndash3 tahun tanaman sakit masih terus memproduksi TBS Pembedahan dilaporkan dapat memperpanjang masa hidup kelapa sawit hingga 2ndash3 tahun (Ho dan Hashim 1997) Pembedahan di perkebunan rakyat mampu memperpanjang masa hidup kelapa sawit di lapangan (Marshall et al 2004 Cooper et al 2011) Berdasarkan pengamatan di lapangan tingkat keberhasilan teknik pembedahan ini sangat bergantung pada ketepatan jaringan sakit yang dibuang Bila jaringan sakit tidak seluruhnya dibuang umumnya kelapa sawit hanya akan bertahan selama satu tahun karena sisa jaringan sakit tersebut berperan sebagai sumber inokulum yang akan terus berkembang hingga menyebabkan pembusukan pada pangkal batang tanaman yang dibumbun
Teknik pengendalian kultur teknis lain yang dapat diterapkan di lapangan ialah pembuatan parit isolasi yang bertujuan mencegah terjadinya kontak akar Hasil pe-ngamatan menunjukkan bahwa parit isolasi mampu mencegah terjadinya infeksi pada tanaman tetangga di luar parit hingga 2 TSP Keberadaan parit isolasi akan mencegah terjadinya kontak akar sehingga tanaman sehat terhindar dari infeksi G boninense dari jaringan tanaman terinfeksi dalam area isolasi Meskipun cukup sulit ketepatan waktu dan titik pembuatan parit isolasi sangat mempengaruhi keefektifan pencegahan pe-nyebaran G boninense di lapangan Dengan akurasi waktu pembuatan yang baik parit isolasi berukuran 2 m times 2 m dilaporkan mampu
6
J Fitopatol Indones Priwiratama et al
mencegah penularan penyakit BPB hingga 14 tahun (Hasan dan Turner 1998 Chung 2011)
Dari uraian di atas dapat diketahui bahwa tujuan pengendalian penyakit BPB secara kultur teknis dititikberatkan untuk menekan adanya sumber inokulum potensial sehingga dapat mencegah terjadinya infeksi G boninense di lapangan Metode pengendalian kultur teknis yang digunakan dapat disesuaikan dengan tahap perkembangan kelapa sawit Di areal pengembangan atau penanaman baru pengendalian dilakukan sejak tahap awal penanaman melalui sanitasi sumber inokulum terutama sisa-sisa perakaran kelapa sawit dan penggunaan lubang tanam besar Sistem penanaman hole in hole dapat diterapkan untuk melakukan penyisipan tanaman baru terutama di areal endemik G boninense Sementara itu untuk tanaman terserang dapat dilakukan tindakan pembedahan diiringi pembumbunan dan bila sudah tidak memungkinkan dapat dilakukan isolasi tanaman terserang untuk mencegah penularan penyakit pada tanaman sehat di sekitarnya Beberapa penelitian di atas akan terus dilanjutkan untuk melihat pengaruh jangka panjang dari teknik-teknik yang diterapkan
DAFTAR PUSTAKA
Chung GF 2011 Management of Ganoderma diseases in oil palm plantations Planter 87(1022)325ndash339
Cooper RM Flood J Rees R 2011 Ganoderma boninense in oil palm plantations current thinking on epidemiology resistance and pathology Planter 87(1024)515ndash526
Durand-Gasselin T Asmady H Flori A Jacquemard Jc Hayun Z Breton F De Franqueville H 2005 Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq) to basal stem rot caused by Ganoderma boninensemdashprospects for future breeding Mycopathologia 159(1)93ndash100 DOI httpdxdoiorg10 1007s11046-004-4429-1
Flood J Keenan L Wayne S Hasan Y 2005 Studies on oil palm trunks as sources of infection in the field Mycopathologia
159(1)101ndash107 DOI httpdxdoiorg 101007s11046-004-4430-8
Hasan Y Turner P 1998 The comparative importance of different oil palm tissues as infection sources for basal stem rot in replantings Planter 74(864)119ndash135
Ho CT Hashim K 1997 Usefulness of soil mounding treatments in prolonging productivity of prime-aged Ganoderma infected palms Planter 73(854)239ndash244
Idris AS Kushairi A Ismail S Ariffin D 2004 Selection for partial tolerance in oil palm progenies to Ganoderma basal stem rot J Oil Palm Res 16(2)12ndash18
Kartika E Yahya S Wilarso S 2006 Isolasi karakterisasi dan pemurnian cendawan mikoriza arbuskular dari dua lokasi perkebunan kelapa sawit (bekas hutan dan bekas kebun karet) J Penelitian Kelapa Sawit 14(3)145ndash155
Marshall R Hunt R Pilotti C 2004 Low cost control for basal stem rotmdasha Poliamba initiative Planter 80(936)173ndash176
Naher L Yusuf UK Tan SG Ismail A 2013 Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq) Aus Sci 7(11)1723ndash1727
Paterson RRM 2007 Ganoderma disease of oil palmmdasha white rot perspective necessary for integrated control Crop Protect 26(2007)1369ndash1376 DOI httpdxdoiorg101016jcropro200611009
Priwiratama H Susanto A 2014 Utilization of fungi for the biological control of insect pests and Ganoderma disease in the Indonesian oil palm industry Agr Sci Tech A 4(2014)103ndash111
Susanto A Prasetyo AE Wening S 2013 Laju infeksi Ganoderma pada empat kelas tekstur tanah J Fitopatol Indones 9(2)39-46 DOI httpdxdoiorg1014692jfi9239
Virdiana I Flood J Sitepu B Hasan Y Aditya R Nelson S 2012 Integrated disease management to reduce future Ganoderma infection during oil palm replanting Planter 88(1305)383ndash393
7
Volume 10 Nomor 1 Februari 2014Halaman 8ndash16
DOI 1014692jfi1018
Pengelolaan Penyakit Kuning pada Tanaman Lada oleh Petani di Wilayah Bangka
Management of Yellow Diseases on Pepper by Farmers in Bangka Region
Abdul Munif Ita SulistiawatiInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit kuning pada tanaman lada yang disebabkan oleh nematoda parasit tumbuhan hingga kini masih menjadi masalah utama pada pertanaman lada di wilayah Bangka Kehilangan hasil berupa penurunan produksi lada yang diakibatkan oleh penyakit kuning cukup besar Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan Penelitian ini meliputi dua kegiatan yaitu survei tingkat kejadian penyakit kuning pada pertanaman lada di lapangan dan wawancara dengan petani lada Hasil survei menunjukkan bahwa tingkat kejadian penyakit kuning sudah merata dan menyerang hampir semua lahan yang diamati Hampir semua petani responden menyatakan bahwa penyakit kuning merupakan penyakit utama yang sangat merusak pertanaman lada Tingkat pemahaman petani terhadap penyebab gejala dan ekologi penyakit kuning dan cara-cara pengendaliannya masih sangat terbatas Tindakan pengendalian yang dilakukan oleh petani terhadap penyakit kuning ialah pencabutan dan pembakaran tanaman terserang dan pemberian kapur Beberapa petani tidak melakukan tindakan pengendalian namun sebagian besar petani menggunakan pestisida Penggunaan pestisida untuk pengendalian penyakit kuning oleh petani belum sepenuhnya mengikuti prinsip pengendalian hama terpadu Masih rendahnya pemahaman petani terhadap penyakit kuning mengakibatkan semakin sulit bagi petani untuk melakukan pengendaliannya
Kata kunci nematoda pengendalian penyakit pestisida Piper nigrum
ABSTRACT
Yellow disease caused by plant-parasitic nematodes are still a major problem on pepper production area in Bangka region Significant yield losses occurred due to yellow disease The objective of this research was to survey the yellow disease incidence and to study farmerrsquos knowledge attitude and practices on the management of yellow disease of black pepper in Bangka district Central Bangka and South Bangka This study consisted of a survey of the incidence of yellow disease in the field and interviews to the farmers The result showed that the yellow disease had infected nearly all of black pepper fields in the areas with incidence of as much as 20ndash60 Almost all of the respondents reported that the yellow disease was the major disease of black pepper in Bangka region Farmersrsquo level of knowledge about the causal of disease symptoms ecology and control methods of the yellow disease are still very limited Various control measures have been done by farmers to combat the disease especially by removing and burning the infected plants or by application of lime A few farmers did not even controlling the disease Pesticides have been used by a few farmers to control the yellow disease but the application of pesticide does not in line with integrated pest management principles
Key words disease control nematode pesticide Piper nigrumAlamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor
Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel abdulmunifipbacid
8
ISSN 2339-2479
J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-
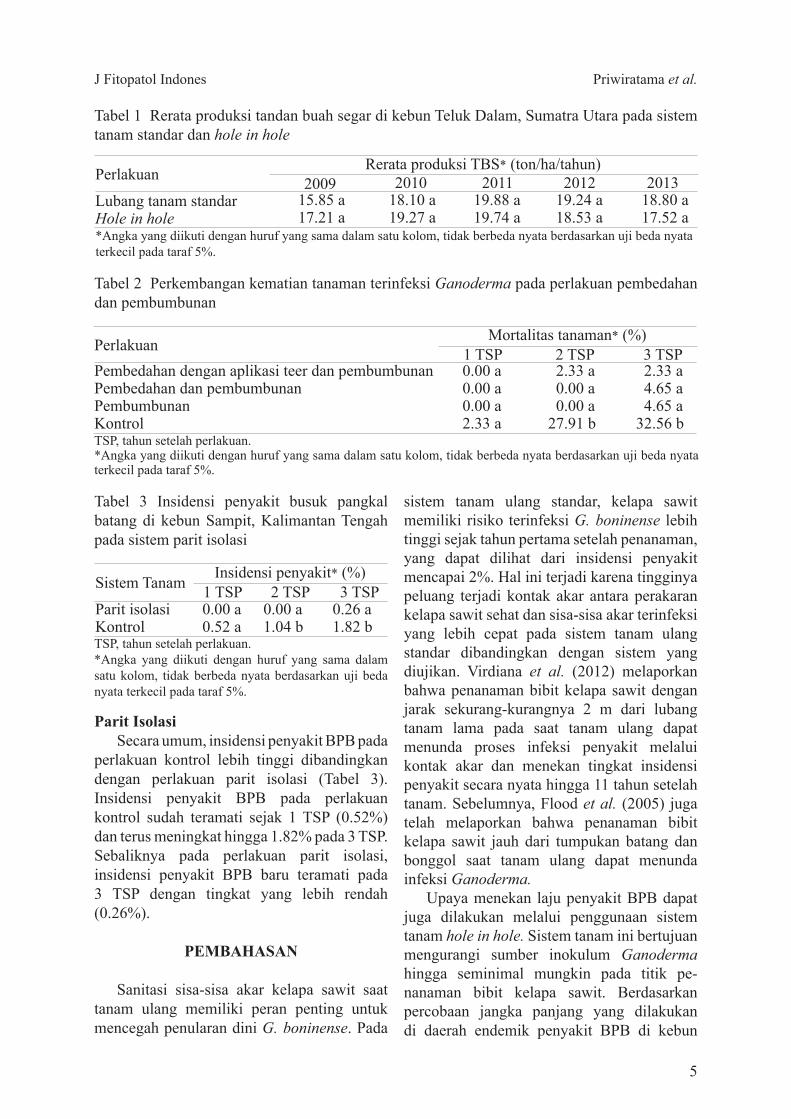
J Fitopatol Indones Priwiratama et al
Parit IsolasiSecara umum insidensi penyakit BPB pada
perlakuan kontrol lebih tinggi dibandingkan dengan perlakuan parit isolasi (Tabel 3) Insidensi penyakit BPB pada perlakuan kontrol sudah teramati sejak 1 TSP (052) dan terus meningkat hingga 182 pada 3 TSP Sebaliknya pada perlakuan parit isolasi insidensi penyakit BPB baru teramati pada 3 TSP dengan tingkat yang lebih rendah (026)
PEMBAHASAN
Sanitasi sisa-sisa akar kelapa sawit saat tanam ulang memiliki peran penting untuk mencegah penularan dini G boninense Pada
sistem tanam ulang standar kelapa sawit memiliki risiko terinfeksi G boninense lebih tinggi sejak tahun pertama setelah penanaman yang dapat dilihat dari insidensi penyakit mencapai 2 Hal ini terjadi karena tingginya peluang terjadi kontak akar antara perakaran kelapa sawit sehat dan sisa-sisa akar terinfeksi yang lebih cepat pada sistem tanam ulang standar dibandingkan dengan sistem yang diujikan Virdiana et al (2012) melaporkan bahwa penanaman bibit kelapa sawit dengan jarak sekurang-kurangnya 2 m dari lubang tanam lama pada saat tanam ulang dapat menunda proses infeksi penyakit melalui kontak akar dan menekan tingkat insidensi penyakit secara nyata hingga 11 tahun setelah tanam Sebelumnya Flood et al (2005) juga telah melaporkan bahwa penanaman bibit kelapa sawit jauh dari tumpukan batang dan bonggol saat tanam ulang dapat menunda infeksi Ganoderma
Upaya menekan laju penyakit BPB dapat juga dilakukan melalui penggunaan sistem tanam hole in hole Sistem tanam ini bertujuan mengurangi sumber inokulum Ganoderma hingga seminimal mungkin pada titik pe-nanaman bibit kelapa sawit Berdasarkan percobaan jangka panjang yang dilakukan di daerah endemik penyakit BPB di kebun
5
Tabel 1 Rerata produksi tandan buah segar di kebun Teluk Dalam Sumatra Utara pada sistem tanam standar dan hole in hole
Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Perlakuan
Lubang tanam standarHole in hole
20102009 2011Rerata produksi TBS (tonhatahun)
201320121585 a1721 a
1810 a1927 a
1988 a1974 a
1924 a1853 a
1880 a1752 a
Tabel 2 Perkembangan kematian tanaman terinfeksi Ganoderma pada perlakuan pembedahan dan pembumbunan
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
PerlakuanPembedahan dengan aplikasi teer dan pembumbunanPembedahan dan pembumbunanPembumbunanKontrol
2 TSP1 TSP 3 TSPMortalitas tanaman ()
000 a000 a000 a233 a
233 a 000 a 000 a2791 b
233 a 465 a 465 a3256 b
Tabel 3 Insidensi penyakit busuk pangkal batang di kebun Sampit Kalimantan Tengah pada sistem parit isolasi
TSP tahun setelah perlakuan Angka yang diikuti dengan huruf yang sama dalam satu kolom tidak berbeda nyata berdasarkan uji beda nyata terkecil pada taraf 5
Sistem Tanam
Parit isolasiKontrol
2 TSP1 TSP 3 TSPInsidensi penyakit ()
000 a052 a
000 a104 b
026 a182 b
J Fitopatol Indones Priwiratama et al
Teluk Dalam Sumatera Utara sistem tanam hole in hole dapat mengurangi tingkat G boninense pada masa awal penanaman kelapa sawit di lapangan Sistem penanaman hole in hole mampu menekan tingkat insidensi penyakit BPB hingga 7 TSP dibandingkan dengan sistem penanaman dengan lubang tanam standar Hal ini dapat disebabkan oleh perbedaan pola sebaran dan jangkauan akar kelapa sawit yang berbeda dengan pada kedua sistem penanaman Perakaran kelapa sawit berumur 3 tahun pada lubang tanam standar dapat menjangkau area sepanjang 467 m dengan sebaran merata sehingga meningkatkan potensi kontak dengan inokulum G boninense di lapangan Pada sistem hole in hole jangkauan akar hanya mencapai 301 m dengan pola sebaran akar terpusat di dalam lubang tanam besar sehingga memperkecil peluang terjadinya kontak akar dengan sumber inokulum Namun demikian setelah 7 TSP sistem tanam hole in hole tidak mampu menghambat laju perkembangan penyakit BPB Pada tahap ini perakaran kelapa sawit telah tumbuh berkembang melebihi area lubang sanitasi sehingga peluang terjadinya kontak dengan sumber inokulum G boninense di luar area hole in hole menjadi semakin tinggi
Penggunaan sistem hole in hole dianggap memberikan dampak negatif terhadap per-kembangan dan produktivitas kelapa sawit karena sebagian besar lapisan tanah bagian atas yang memiliki kandungan hara lebih tinggi terbuang dan hanya menyisakan lapisan tanah dalam yang miskin unsur hara Namun berdasarkan pengamatan di lapangan sistem tanam ini tidak memberikan pengaruh terhadap keragaan tanaman dan produktivitas kelapa sawit Data produksi TBS selama lima tahun terakhir (Tabel 1) memperlihatkan produktivitas kelapa sawit pada kedua sistem tanam relatif berimbang Dalam praktiknya penambahan bahan organik berupa tandan kosong kelapa sawit pada penerapan sistem hole in hole merupakan salah satu faktor penting agar asupan nutrisi kelapa sawit selama tahap awal perkembangannya dapat terpenuhi
sehingga keragaan dan produktivitasnya tidak tertinggal dari sistem tanam standar
Penerapan teknik pembedahan dan pem-bumbunan untuk mengendalikan penyakit BPB menunjukkan hasil yang cukup menjanjikan Meskipun tidak dapat menghindarkan tanaman dari infeksi G boninense pembedahan jaringan sakit yang disertai pembumbunan mampu memperpanjang masa hidup tanaman terinfeksi sekurang-kurangnya hingga 3 TSP dibandingkan dengan tanaman tanpa perlakuan Sedangkan pada tanaman dengan hanya pembumbunan mampu bertahan hingga 2 TSP Hal ini cukup penting mengingat dalam kurun waktu 2ndash3 tahun tanaman sakit masih terus memproduksi TBS Pembedahan dilaporkan dapat memperpanjang masa hidup kelapa sawit hingga 2ndash3 tahun (Ho dan Hashim 1997) Pembedahan di perkebunan rakyat mampu memperpanjang masa hidup kelapa sawit di lapangan (Marshall et al 2004 Cooper et al 2011) Berdasarkan pengamatan di lapangan tingkat keberhasilan teknik pembedahan ini sangat bergantung pada ketepatan jaringan sakit yang dibuang Bila jaringan sakit tidak seluruhnya dibuang umumnya kelapa sawit hanya akan bertahan selama satu tahun karena sisa jaringan sakit tersebut berperan sebagai sumber inokulum yang akan terus berkembang hingga menyebabkan pembusukan pada pangkal batang tanaman yang dibumbun
Teknik pengendalian kultur teknis lain yang dapat diterapkan di lapangan ialah pembuatan parit isolasi yang bertujuan mencegah terjadinya kontak akar Hasil pe-ngamatan menunjukkan bahwa parit isolasi mampu mencegah terjadinya infeksi pada tanaman tetangga di luar parit hingga 2 TSP Keberadaan parit isolasi akan mencegah terjadinya kontak akar sehingga tanaman sehat terhindar dari infeksi G boninense dari jaringan tanaman terinfeksi dalam area isolasi Meskipun cukup sulit ketepatan waktu dan titik pembuatan parit isolasi sangat mempengaruhi keefektifan pencegahan pe-nyebaran G boninense di lapangan Dengan akurasi waktu pembuatan yang baik parit isolasi berukuran 2 m times 2 m dilaporkan mampu
6
J Fitopatol Indones Priwiratama et al
mencegah penularan penyakit BPB hingga 14 tahun (Hasan dan Turner 1998 Chung 2011)
Dari uraian di atas dapat diketahui bahwa tujuan pengendalian penyakit BPB secara kultur teknis dititikberatkan untuk menekan adanya sumber inokulum potensial sehingga dapat mencegah terjadinya infeksi G boninense di lapangan Metode pengendalian kultur teknis yang digunakan dapat disesuaikan dengan tahap perkembangan kelapa sawit Di areal pengembangan atau penanaman baru pengendalian dilakukan sejak tahap awal penanaman melalui sanitasi sumber inokulum terutama sisa-sisa perakaran kelapa sawit dan penggunaan lubang tanam besar Sistem penanaman hole in hole dapat diterapkan untuk melakukan penyisipan tanaman baru terutama di areal endemik G boninense Sementara itu untuk tanaman terserang dapat dilakukan tindakan pembedahan diiringi pembumbunan dan bila sudah tidak memungkinkan dapat dilakukan isolasi tanaman terserang untuk mencegah penularan penyakit pada tanaman sehat di sekitarnya Beberapa penelitian di atas akan terus dilanjutkan untuk melihat pengaruh jangka panjang dari teknik-teknik yang diterapkan
DAFTAR PUSTAKA
Chung GF 2011 Management of Ganoderma diseases in oil palm plantations Planter 87(1022)325ndash339
Cooper RM Flood J Rees R 2011 Ganoderma boninense in oil palm plantations current thinking on epidemiology resistance and pathology Planter 87(1024)515ndash526
Durand-Gasselin T Asmady H Flori A Jacquemard Jc Hayun Z Breton F De Franqueville H 2005 Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq) to basal stem rot caused by Ganoderma boninensemdashprospects for future breeding Mycopathologia 159(1)93ndash100 DOI httpdxdoiorg10 1007s11046-004-4429-1
Flood J Keenan L Wayne S Hasan Y 2005 Studies on oil palm trunks as sources of infection in the field Mycopathologia
159(1)101ndash107 DOI httpdxdoiorg 101007s11046-004-4430-8
Hasan Y Turner P 1998 The comparative importance of different oil palm tissues as infection sources for basal stem rot in replantings Planter 74(864)119ndash135
Ho CT Hashim K 1997 Usefulness of soil mounding treatments in prolonging productivity of prime-aged Ganoderma infected palms Planter 73(854)239ndash244
Idris AS Kushairi A Ismail S Ariffin D 2004 Selection for partial tolerance in oil palm progenies to Ganoderma basal stem rot J Oil Palm Res 16(2)12ndash18
Kartika E Yahya S Wilarso S 2006 Isolasi karakterisasi dan pemurnian cendawan mikoriza arbuskular dari dua lokasi perkebunan kelapa sawit (bekas hutan dan bekas kebun karet) J Penelitian Kelapa Sawit 14(3)145ndash155
Marshall R Hunt R Pilotti C 2004 Low cost control for basal stem rotmdasha Poliamba initiative Planter 80(936)173ndash176
Naher L Yusuf UK Tan SG Ismail A 2013 Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq) Aus Sci 7(11)1723ndash1727
Paterson RRM 2007 Ganoderma disease of oil palmmdasha white rot perspective necessary for integrated control Crop Protect 26(2007)1369ndash1376 DOI httpdxdoiorg101016jcropro200611009
Priwiratama H Susanto A 2014 Utilization of fungi for the biological control of insect pests and Ganoderma disease in the Indonesian oil palm industry Agr Sci Tech A 4(2014)103ndash111
Susanto A Prasetyo AE Wening S 2013 Laju infeksi Ganoderma pada empat kelas tekstur tanah J Fitopatol Indones 9(2)39-46 DOI httpdxdoiorg1014692jfi9239
Virdiana I Flood J Sitepu B Hasan Y Aditya R Nelson S 2012 Integrated disease management to reduce future Ganoderma infection during oil palm replanting Planter 88(1305)383ndash393
7
Volume 10 Nomor 1 Februari 2014Halaman 8ndash16
DOI 1014692jfi1018
Pengelolaan Penyakit Kuning pada Tanaman Lada oleh Petani di Wilayah Bangka
Management of Yellow Diseases on Pepper by Farmers in Bangka Region
Abdul Munif Ita SulistiawatiInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit kuning pada tanaman lada yang disebabkan oleh nematoda parasit tumbuhan hingga kini masih menjadi masalah utama pada pertanaman lada di wilayah Bangka Kehilangan hasil berupa penurunan produksi lada yang diakibatkan oleh penyakit kuning cukup besar Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan Penelitian ini meliputi dua kegiatan yaitu survei tingkat kejadian penyakit kuning pada pertanaman lada di lapangan dan wawancara dengan petani lada Hasil survei menunjukkan bahwa tingkat kejadian penyakit kuning sudah merata dan menyerang hampir semua lahan yang diamati Hampir semua petani responden menyatakan bahwa penyakit kuning merupakan penyakit utama yang sangat merusak pertanaman lada Tingkat pemahaman petani terhadap penyebab gejala dan ekologi penyakit kuning dan cara-cara pengendaliannya masih sangat terbatas Tindakan pengendalian yang dilakukan oleh petani terhadap penyakit kuning ialah pencabutan dan pembakaran tanaman terserang dan pemberian kapur Beberapa petani tidak melakukan tindakan pengendalian namun sebagian besar petani menggunakan pestisida Penggunaan pestisida untuk pengendalian penyakit kuning oleh petani belum sepenuhnya mengikuti prinsip pengendalian hama terpadu Masih rendahnya pemahaman petani terhadap penyakit kuning mengakibatkan semakin sulit bagi petani untuk melakukan pengendaliannya
Kata kunci nematoda pengendalian penyakit pestisida Piper nigrum
ABSTRACT
Yellow disease caused by plant-parasitic nematodes are still a major problem on pepper production area in Bangka region Significant yield losses occurred due to yellow disease The objective of this research was to survey the yellow disease incidence and to study farmerrsquos knowledge attitude and practices on the management of yellow disease of black pepper in Bangka district Central Bangka and South Bangka This study consisted of a survey of the incidence of yellow disease in the field and interviews to the farmers The result showed that the yellow disease had infected nearly all of black pepper fields in the areas with incidence of as much as 20ndash60 Almost all of the respondents reported that the yellow disease was the major disease of black pepper in Bangka region Farmersrsquo level of knowledge about the causal of disease symptoms ecology and control methods of the yellow disease are still very limited Various control measures have been done by farmers to combat the disease especially by removing and burning the infected plants or by application of lime A few farmers did not even controlling the disease Pesticides have been used by a few farmers to control the yellow disease but the application of pesticide does not in line with integrated pest management principles
Key words disease control nematode pesticide Piper nigrumAlamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor
Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel abdulmunifipbacid
8
ISSN 2339-2479
J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Priwiratama et al
Teluk Dalam Sumatera Utara sistem tanam hole in hole dapat mengurangi tingkat G boninense pada masa awal penanaman kelapa sawit di lapangan Sistem penanaman hole in hole mampu menekan tingkat insidensi penyakit BPB hingga 7 TSP dibandingkan dengan sistem penanaman dengan lubang tanam standar Hal ini dapat disebabkan oleh perbedaan pola sebaran dan jangkauan akar kelapa sawit yang berbeda dengan pada kedua sistem penanaman Perakaran kelapa sawit berumur 3 tahun pada lubang tanam standar dapat menjangkau area sepanjang 467 m dengan sebaran merata sehingga meningkatkan potensi kontak dengan inokulum G boninense di lapangan Pada sistem hole in hole jangkauan akar hanya mencapai 301 m dengan pola sebaran akar terpusat di dalam lubang tanam besar sehingga memperkecil peluang terjadinya kontak akar dengan sumber inokulum Namun demikian setelah 7 TSP sistem tanam hole in hole tidak mampu menghambat laju perkembangan penyakit BPB Pada tahap ini perakaran kelapa sawit telah tumbuh berkembang melebihi area lubang sanitasi sehingga peluang terjadinya kontak dengan sumber inokulum G boninense di luar area hole in hole menjadi semakin tinggi
Penggunaan sistem hole in hole dianggap memberikan dampak negatif terhadap per-kembangan dan produktivitas kelapa sawit karena sebagian besar lapisan tanah bagian atas yang memiliki kandungan hara lebih tinggi terbuang dan hanya menyisakan lapisan tanah dalam yang miskin unsur hara Namun berdasarkan pengamatan di lapangan sistem tanam ini tidak memberikan pengaruh terhadap keragaan tanaman dan produktivitas kelapa sawit Data produksi TBS selama lima tahun terakhir (Tabel 1) memperlihatkan produktivitas kelapa sawit pada kedua sistem tanam relatif berimbang Dalam praktiknya penambahan bahan organik berupa tandan kosong kelapa sawit pada penerapan sistem hole in hole merupakan salah satu faktor penting agar asupan nutrisi kelapa sawit selama tahap awal perkembangannya dapat terpenuhi
sehingga keragaan dan produktivitasnya tidak tertinggal dari sistem tanam standar
Penerapan teknik pembedahan dan pem-bumbunan untuk mengendalikan penyakit BPB menunjukkan hasil yang cukup menjanjikan Meskipun tidak dapat menghindarkan tanaman dari infeksi G boninense pembedahan jaringan sakit yang disertai pembumbunan mampu memperpanjang masa hidup tanaman terinfeksi sekurang-kurangnya hingga 3 TSP dibandingkan dengan tanaman tanpa perlakuan Sedangkan pada tanaman dengan hanya pembumbunan mampu bertahan hingga 2 TSP Hal ini cukup penting mengingat dalam kurun waktu 2ndash3 tahun tanaman sakit masih terus memproduksi TBS Pembedahan dilaporkan dapat memperpanjang masa hidup kelapa sawit hingga 2ndash3 tahun (Ho dan Hashim 1997) Pembedahan di perkebunan rakyat mampu memperpanjang masa hidup kelapa sawit di lapangan (Marshall et al 2004 Cooper et al 2011) Berdasarkan pengamatan di lapangan tingkat keberhasilan teknik pembedahan ini sangat bergantung pada ketepatan jaringan sakit yang dibuang Bila jaringan sakit tidak seluruhnya dibuang umumnya kelapa sawit hanya akan bertahan selama satu tahun karena sisa jaringan sakit tersebut berperan sebagai sumber inokulum yang akan terus berkembang hingga menyebabkan pembusukan pada pangkal batang tanaman yang dibumbun
Teknik pengendalian kultur teknis lain yang dapat diterapkan di lapangan ialah pembuatan parit isolasi yang bertujuan mencegah terjadinya kontak akar Hasil pe-ngamatan menunjukkan bahwa parit isolasi mampu mencegah terjadinya infeksi pada tanaman tetangga di luar parit hingga 2 TSP Keberadaan parit isolasi akan mencegah terjadinya kontak akar sehingga tanaman sehat terhindar dari infeksi G boninense dari jaringan tanaman terinfeksi dalam area isolasi Meskipun cukup sulit ketepatan waktu dan titik pembuatan parit isolasi sangat mempengaruhi keefektifan pencegahan pe-nyebaran G boninense di lapangan Dengan akurasi waktu pembuatan yang baik parit isolasi berukuran 2 m times 2 m dilaporkan mampu
6
J Fitopatol Indones Priwiratama et al
mencegah penularan penyakit BPB hingga 14 tahun (Hasan dan Turner 1998 Chung 2011)
Dari uraian di atas dapat diketahui bahwa tujuan pengendalian penyakit BPB secara kultur teknis dititikberatkan untuk menekan adanya sumber inokulum potensial sehingga dapat mencegah terjadinya infeksi G boninense di lapangan Metode pengendalian kultur teknis yang digunakan dapat disesuaikan dengan tahap perkembangan kelapa sawit Di areal pengembangan atau penanaman baru pengendalian dilakukan sejak tahap awal penanaman melalui sanitasi sumber inokulum terutama sisa-sisa perakaran kelapa sawit dan penggunaan lubang tanam besar Sistem penanaman hole in hole dapat diterapkan untuk melakukan penyisipan tanaman baru terutama di areal endemik G boninense Sementara itu untuk tanaman terserang dapat dilakukan tindakan pembedahan diiringi pembumbunan dan bila sudah tidak memungkinkan dapat dilakukan isolasi tanaman terserang untuk mencegah penularan penyakit pada tanaman sehat di sekitarnya Beberapa penelitian di atas akan terus dilanjutkan untuk melihat pengaruh jangka panjang dari teknik-teknik yang diterapkan
DAFTAR PUSTAKA
Chung GF 2011 Management of Ganoderma diseases in oil palm plantations Planter 87(1022)325ndash339
Cooper RM Flood J Rees R 2011 Ganoderma boninense in oil palm plantations current thinking on epidemiology resistance and pathology Planter 87(1024)515ndash526
Durand-Gasselin T Asmady H Flori A Jacquemard Jc Hayun Z Breton F De Franqueville H 2005 Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq) to basal stem rot caused by Ganoderma boninensemdashprospects for future breeding Mycopathologia 159(1)93ndash100 DOI httpdxdoiorg10 1007s11046-004-4429-1
Flood J Keenan L Wayne S Hasan Y 2005 Studies on oil palm trunks as sources of infection in the field Mycopathologia
159(1)101ndash107 DOI httpdxdoiorg 101007s11046-004-4430-8
Hasan Y Turner P 1998 The comparative importance of different oil palm tissues as infection sources for basal stem rot in replantings Planter 74(864)119ndash135
Ho CT Hashim K 1997 Usefulness of soil mounding treatments in prolonging productivity of prime-aged Ganoderma infected palms Planter 73(854)239ndash244
Idris AS Kushairi A Ismail S Ariffin D 2004 Selection for partial tolerance in oil palm progenies to Ganoderma basal stem rot J Oil Palm Res 16(2)12ndash18
Kartika E Yahya S Wilarso S 2006 Isolasi karakterisasi dan pemurnian cendawan mikoriza arbuskular dari dua lokasi perkebunan kelapa sawit (bekas hutan dan bekas kebun karet) J Penelitian Kelapa Sawit 14(3)145ndash155
Marshall R Hunt R Pilotti C 2004 Low cost control for basal stem rotmdasha Poliamba initiative Planter 80(936)173ndash176
Naher L Yusuf UK Tan SG Ismail A 2013 Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq) Aus Sci 7(11)1723ndash1727
Paterson RRM 2007 Ganoderma disease of oil palmmdasha white rot perspective necessary for integrated control Crop Protect 26(2007)1369ndash1376 DOI httpdxdoiorg101016jcropro200611009
Priwiratama H Susanto A 2014 Utilization of fungi for the biological control of insect pests and Ganoderma disease in the Indonesian oil palm industry Agr Sci Tech A 4(2014)103ndash111
Susanto A Prasetyo AE Wening S 2013 Laju infeksi Ganoderma pada empat kelas tekstur tanah J Fitopatol Indones 9(2)39-46 DOI httpdxdoiorg1014692jfi9239
Virdiana I Flood J Sitepu B Hasan Y Aditya R Nelson S 2012 Integrated disease management to reduce future Ganoderma infection during oil palm replanting Planter 88(1305)383ndash393
7
Volume 10 Nomor 1 Februari 2014Halaman 8ndash16
DOI 1014692jfi1018
Pengelolaan Penyakit Kuning pada Tanaman Lada oleh Petani di Wilayah Bangka
Management of Yellow Diseases on Pepper by Farmers in Bangka Region
Abdul Munif Ita SulistiawatiInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit kuning pada tanaman lada yang disebabkan oleh nematoda parasit tumbuhan hingga kini masih menjadi masalah utama pada pertanaman lada di wilayah Bangka Kehilangan hasil berupa penurunan produksi lada yang diakibatkan oleh penyakit kuning cukup besar Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan Penelitian ini meliputi dua kegiatan yaitu survei tingkat kejadian penyakit kuning pada pertanaman lada di lapangan dan wawancara dengan petani lada Hasil survei menunjukkan bahwa tingkat kejadian penyakit kuning sudah merata dan menyerang hampir semua lahan yang diamati Hampir semua petani responden menyatakan bahwa penyakit kuning merupakan penyakit utama yang sangat merusak pertanaman lada Tingkat pemahaman petani terhadap penyebab gejala dan ekologi penyakit kuning dan cara-cara pengendaliannya masih sangat terbatas Tindakan pengendalian yang dilakukan oleh petani terhadap penyakit kuning ialah pencabutan dan pembakaran tanaman terserang dan pemberian kapur Beberapa petani tidak melakukan tindakan pengendalian namun sebagian besar petani menggunakan pestisida Penggunaan pestisida untuk pengendalian penyakit kuning oleh petani belum sepenuhnya mengikuti prinsip pengendalian hama terpadu Masih rendahnya pemahaman petani terhadap penyakit kuning mengakibatkan semakin sulit bagi petani untuk melakukan pengendaliannya
Kata kunci nematoda pengendalian penyakit pestisida Piper nigrum
ABSTRACT
Yellow disease caused by plant-parasitic nematodes are still a major problem on pepper production area in Bangka region Significant yield losses occurred due to yellow disease The objective of this research was to survey the yellow disease incidence and to study farmerrsquos knowledge attitude and practices on the management of yellow disease of black pepper in Bangka district Central Bangka and South Bangka This study consisted of a survey of the incidence of yellow disease in the field and interviews to the farmers The result showed that the yellow disease had infected nearly all of black pepper fields in the areas with incidence of as much as 20ndash60 Almost all of the respondents reported that the yellow disease was the major disease of black pepper in Bangka region Farmersrsquo level of knowledge about the causal of disease symptoms ecology and control methods of the yellow disease are still very limited Various control measures have been done by farmers to combat the disease especially by removing and burning the infected plants or by application of lime A few farmers did not even controlling the disease Pesticides have been used by a few farmers to control the yellow disease but the application of pesticide does not in line with integrated pest management principles
Key words disease control nematode pesticide Piper nigrumAlamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor
Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel abdulmunifipbacid
8
ISSN 2339-2479
J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Priwiratama et al
mencegah penularan penyakit BPB hingga 14 tahun (Hasan dan Turner 1998 Chung 2011)
Dari uraian di atas dapat diketahui bahwa tujuan pengendalian penyakit BPB secara kultur teknis dititikberatkan untuk menekan adanya sumber inokulum potensial sehingga dapat mencegah terjadinya infeksi G boninense di lapangan Metode pengendalian kultur teknis yang digunakan dapat disesuaikan dengan tahap perkembangan kelapa sawit Di areal pengembangan atau penanaman baru pengendalian dilakukan sejak tahap awal penanaman melalui sanitasi sumber inokulum terutama sisa-sisa perakaran kelapa sawit dan penggunaan lubang tanam besar Sistem penanaman hole in hole dapat diterapkan untuk melakukan penyisipan tanaman baru terutama di areal endemik G boninense Sementara itu untuk tanaman terserang dapat dilakukan tindakan pembedahan diiringi pembumbunan dan bila sudah tidak memungkinkan dapat dilakukan isolasi tanaman terserang untuk mencegah penularan penyakit pada tanaman sehat di sekitarnya Beberapa penelitian di atas akan terus dilanjutkan untuk melihat pengaruh jangka panjang dari teknik-teknik yang diterapkan
DAFTAR PUSTAKA
Chung GF 2011 Management of Ganoderma diseases in oil palm plantations Planter 87(1022)325ndash339
Cooper RM Flood J Rees R 2011 Ganoderma boninense in oil palm plantations current thinking on epidemiology resistance and pathology Planter 87(1024)515ndash526
Durand-Gasselin T Asmady H Flori A Jacquemard Jc Hayun Z Breton F De Franqueville H 2005 Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq) to basal stem rot caused by Ganoderma boninensemdashprospects for future breeding Mycopathologia 159(1)93ndash100 DOI httpdxdoiorg10 1007s11046-004-4429-1
Flood J Keenan L Wayne S Hasan Y 2005 Studies on oil palm trunks as sources of infection in the field Mycopathologia
159(1)101ndash107 DOI httpdxdoiorg 101007s11046-004-4430-8
Hasan Y Turner P 1998 The comparative importance of different oil palm tissues as infection sources for basal stem rot in replantings Planter 74(864)119ndash135
Ho CT Hashim K 1997 Usefulness of soil mounding treatments in prolonging productivity of prime-aged Ganoderma infected palms Planter 73(854)239ndash244
Idris AS Kushairi A Ismail S Ariffin D 2004 Selection for partial tolerance in oil palm progenies to Ganoderma basal stem rot J Oil Palm Res 16(2)12ndash18
Kartika E Yahya S Wilarso S 2006 Isolasi karakterisasi dan pemurnian cendawan mikoriza arbuskular dari dua lokasi perkebunan kelapa sawit (bekas hutan dan bekas kebun karet) J Penelitian Kelapa Sawit 14(3)145ndash155
Marshall R Hunt R Pilotti C 2004 Low cost control for basal stem rotmdasha Poliamba initiative Planter 80(936)173ndash176
Naher L Yusuf UK Tan SG Ismail A 2013 Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq) Aus Sci 7(11)1723ndash1727
Paterson RRM 2007 Ganoderma disease of oil palmmdasha white rot perspective necessary for integrated control Crop Protect 26(2007)1369ndash1376 DOI httpdxdoiorg101016jcropro200611009
Priwiratama H Susanto A 2014 Utilization of fungi for the biological control of insect pests and Ganoderma disease in the Indonesian oil palm industry Agr Sci Tech A 4(2014)103ndash111
Susanto A Prasetyo AE Wening S 2013 Laju infeksi Ganoderma pada empat kelas tekstur tanah J Fitopatol Indones 9(2)39-46 DOI httpdxdoiorg1014692jfi9239
Virdiana I Flood J Sitepu B Hasan Y Aditya R Nelson S 2012 Integrated disease management to reduce future Ganoderma infection during oil palm replanting Planter 88(1305)383ndash393
7
Volume 10 Nomor 1 Februari 2014Halaman 8ndash16
DOI 1014692jfi1018
Pengelolaan Penyakit Kuning pada Tanaman Lada oleh Petani di Wilayah Bangka
Management of Yellow Diseases on Pepper by Farmers in Bangka Region
Abdul Munif Ita SulistiawatiInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit kuning pada tanaman lada yang disebabkan oleh nematoda parasit tumbuhan hingga kini masih menjadi masalah utama pada pertanaman lada di wilayah Bangka Kehilangan hasil berupa penurunan produksi lada yang diakibatkan oleh penyakit kuning cukup besar Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan Penelitian ini meliputi dua kegiatan yaitu survei tingkat kejadian penyakit kuning pada pertanaman lada di lapangan dan wawancara dengan petani lada Hasil survei menunjukkan bahwa tingkat kejadian penyakit kuning sudah merata dan menyerang hampir semua lahan yang diamati Hampir semua petani responden menyatakan bahwa penyakit kuning merupakan penyakit utama yang sangat merusak pertanaman lada Tingkat pemahaman petani terhadap penyebab gejala dan ekologi penyakit kuning dan cara-cara pengendaliannya masih sangat terbatas Tindakan pengendalian yang dilakukan oleh petani terhadap penyakit kuning ialah pencabutan dan pembakaran tanaman terserang dan pemberian kapur Beberapa petani tidak melakukan tindakan pengendalian namun sebagian besar petani menggunakan pestisida Penggunaan pestisida untuk pengendalian penyakit kuning oleh petani belum sepenuhnya mengikuti prinsip pengendalian hama terpadu Masih rendahnya pemahaman petani terhadap penyakit kuning mengakibatkan semakin sulit bagi petani untuk melakukan pengendaliannya
Kata kunci nematoda pengendalian penyakit pestisida Piper nigrum
ABSTRACT
Yellow disease caused by plant-parasitic nematodes are still a major problem on pepper production area in Bangka region Significant yield losses occurred due to yellow disease The objective of this research was to survey the yellow disease incidence and to study farmerrsquos knowledge attitude and practices on the management of yellow disease of black pepper in Bangka district Central Bangka and South Bangka This study consisted of a survey of the incidence of yellow disease in the field and interviews to the farmers The result showed that the yellow disease had infected nearly all of black pepper fields in the areas with incidence of as much as 20ndash60 Almost all of the respondents reported that the yellow disease was the major disease of black pepper in Bangka region Farmersrsquo level of knowledge about the causal of disease symptoms ecology and control methods of the yellow disease are still very limited Various control measures have been done by farmers to combat the disease especially by removing and burning the infected plants or by application of lime A few farmers did not even controlling the disease Pesticides have been used by a few farmers to control the yellow disease but the application of pesticide does not in line with integrated pest management principles
Key words disease control nematode pesticide Piper nigrumAlamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor
Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel abdulmunifipbacid
8
ISSN 2339-2479
J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

Volume 10 Nomor 1 Februari 2014Halaman 8ndash16
DOI 1014692jfi1018
Pengelolaan Penyakit Kuning pada Tanaman Lada oleh Petani di Wilayah Bangka
Management of Yellow Diseases on Pepper by Farmers in Bangka Region
Abdul Munif Ita SulistiawatiInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit kuning pada tanaman lada yang disebabkan oleh nematoda parasit tumbuhan hingga kini masih menjadi masalah utama pada pertanaman lada di wilayah Bangka Kehilangan hasil berupa penurunan produksi lada yang diakibatkan oleh penyakit kuning cukup besar Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan Penelitian ini meliputi dua kegiatan yaitu survei tingkat kejadian penyakit kuning pada pertanaman lada di lapangan dan wawancara dengan petani lada Hasil survei menunjukkan bahwa tingkat kejadian penyakit kuning sudah merata dan menyerang hampir semua lahan yang diamati Hampir semua petani responden menyatakan bahwa penyakit kuning merupakan penyakit utama yang sangat merusak pertanaman lada Tingkat pemahaman petani terhadap penyebab gejala dan ekologi penyakit kuning dan cara-cara pengendaliannya masih sangat terbatas Tindakan pengendalian yang dilakukan oleh petani terhadap penyakit kuning ialah pencabutan dan pembakaran tanaman terserang dan pemberian kapur Beberapa petani tidak melakukan tindakan pengendalian namun sebagian besar petani menggunakan pestisida Penggunaan pestisida untuk pengendalian penyakit kuning oleh petani belum sepenuhnya mengikuti prinsip pengendalian hama terpadu Masih rendahnya pemahaman petani terhadap penyakit kuning mengakibatkan semakin sulit bagi petani untuk melakukan pengendaliannya
Kata kunci nematoda pengendalian penyakit pestisida Piper nigrum
ABSTRACT
Yellow disease caused by plant-parasitic nematodes are still a major problem on pepper production area in Bangka region Significant yield losses occurred due to yellow disease The objective of this research was to survey the yellow disease incidence and to study farmerrsquos knowledge attitude and practices on the management of yellow disease of black pepper in Bangka district Central Bangka and South Bangka This study consisted of a survey of the incidence of yellow disease in the field and interviews to the farmers The result showed that the yellow disease had infected nearly all of black pepper fields in the areas with incidence of as much as 20ndash60 Almost all of the respondents reported that the yellow disease was the major disease of black pepper in Bangka region Farmersrsquo level of knowledge about the causal of disease symptoms ecology and control methods of the yellow disease are still very limited Various control measures have been done by farmers to combat the disease especially by removing and burning the infected plants or by application of lime A few farmers did not even controlling the disease Pesticides have been used by a few farmers to control the yellow disease but the application of pesticide does not in line with integrated pest management principles
Key words disease control nematode pesticide Piper nigrumAlamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor
Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel abdulmunifipbacid
8
ISSN 2339-2479
J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Munif dan Sulistiawati
PENDAHULUAN
Lada (Piper nigrum) merupakan jenis rempah yang paling sering digunakan di Eropa dibandingkan dengan rempah-rempah lainnya Daerah utama penghasil lada di Indonesia ialah daerah Lampung yang terkenal sebagai penghasil lada hitam atau lampung black pepper dan Bangka yang terkenal sebagai penghasil lada putih atau muntok white pepper Daras dan Pranowo (2009) melaporkan bahwa Indonesia masih menjadi salah satu produsen lada yang diperhitungkan di pasar dunia namun produktivitas lada nasional terus menurun dalam sepuluh tahun terakhir Faktor yang mempengaruhi produksi lada ialah gangguan penyakit antara lain penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici dan penyakit kuning yang disebabkan oleh nematoda (Manohara et al 2006)
Penyakit kuning merupakan salah satu penyakit penting pada pertanaman lada di wilayah Bangka Penyakit kuning sangat merugikan karena dapat menyebabkan tanaman lada berhenti berkembang sehingga menurunkan hasil panen Penyakit ini dilaporkan merusak pertanaman lada wilayah Bangka hingga 32 Penyebab penyakit tersebut adalah nematoda Meloidogyne incognita dan Radopholus similis (Mustika 1990) Deteksi awal penyakit ini relatif sulit biasanya tanaman diketahui sakit setelah menampakkan gejala menguning pada daun Perkembangan penyakit tanaman sangatbergantung pada banyak faktor baik ling-kungan tanaman maupun teknik budi daya yang digunakan Teknik atau sistem budi daya yang digunakan petani diduga berpengaruh terhadap perkembangan penyakit kuning pada pertanaman lada
Penggunaan bahan kimia dalam pertanian di Indonesia terutama pestisida untuk tujuanpengendalian hama dan penyakit tanaman masih merupakan cara yang paling disukai oleh petani Pada komoditas tertentu pe-ngeluaran petani untuk membeli pestisida dapat mencapai 40 dari total biaya produksi keseluruhan Penggunaan pestisida yang terlalu intensif di lapang dapat berakibat tidak
baik seperti kerancunan kontaminasi racun pestisida pada air sumur bahan makanan dan kolam ikan serta munculnya hama dan patogen yang resisten terhadap suatu pestisida Alternatif pengendalian hama dan penyakit tanaman yang ramah lingkungan untuk mendukung kehidupan yang lebih sehat perlu terus dikembangkan sejalan dengan konsep pengendalian hama terpadu (PHT)
Penelitian ini bertujuan memperoleh informasi mengenai pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada dan pengelolaan penyakit khususnya penyakit kuning di wilayah Bangka
BAHAN DAN METODE
Penelitian dilaksanakan di Desa Petaling Kemuja dan Cengkong Abang Kecamatan Mendo Barat Kabupaten Bangka Desa Sungkap Celuak dan Teru Kecamatan Simpang Katis Kabupaten Bangka Tengah dan Desa Delas Nyelanding dan Air Gegas Kecamatan Air Gegas Kabupaten Bangka Selatan Penelitian dilaksanakan dari bulan April sampai Juni 2010 dan bulan Agustus sampai Desember 2011 Metode Penelitian
Penelitian ini terdiri atas dua kegiatan yaitu survei kejadian penyakit kuning di lapangan dan wawancara dengan petani lada Kegiatan survei lapangan bertujuan menentu-kan tingkat kejadian dan luas serangan penyakit kuning pada lada di lahan petani sedangkan wawancara dengan petani dimaksudkan untuk memperoleh informasi sistem pengelolaan atau pengendalian penyakit kuning oleh petani
Survei Penyakit KuningPengamatan penyakit kuning dilakukan
pada lahan tanaman lada milik petani di tiga kabupaten yaitu Kabupaten Bangka Bangka Tengah dan Bangka Selatan Setiap kabuapten diambil satu kecamatan dan dari setiap kecamatan dipilih 3 desa Setiap desa dipilih 4 kebun lada yang ditentukan secara purposif dengan kriteria kebun mempunyai
9
J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Munif dan Sulistiawati
tanaman lada paling sedikit 2000 tanaman dan umur lada antara 1ndash2 tahun Penentuan umur tanaman 1ndash2 tahun dengan alasan gejala awal penyakit kuning sudah terlihat saat tanaman berumur 1 tahun setelah di lapangan Pada setiap lahan diamati 25 tanaman contoh yang ditentukan secara sistematis Penilaian gejala penyakit kuning pada lada mengacu pada gejala umum di lapangan Gejala awal penyakit kuning yaitu daun tanaman lada berwarna kuning atau pucat dan daun menggulung ke arah batang sedangkan gejala lanjut berupa tanaman menguning kerdil tunas mati dan akhirnya seluruh daun gugur (Mustika 1990)
Tanaman lada yang menunjukkan gejala penyakit kuning baik gejala masih awal atau gejala lanjut dianggap tanaman sudah terserang secara keseluruhan Selanjutnya jumlah tanaman contoh yang terserang dicatat untuk menentukan kejadian penyakit kuning pada lahan tersebut Kejadian penyakit di-hitung dengan rumus
n jumlah tanaman yang terserang penyakit kuning N jumlah tanaman yang diamati
Wawancara dengan PetaniWawancara dengan petani dimaksudkan
untuk mendapatkan gambaran tentang pe-ngetahuan dan tindakan pengendalian ter-hadap penyakit kuning pada tanaman lada yang dilakukan petani responden Wawancara secara langsung dilakukan terhadap petani lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan dengan menggunakan kuisioner Dari setiap kabupaten ditentukan 40 responden yang menyebar di tiga desa sehingga jumlah total responden dari ketiga kabupaten adalah 120 petani responden Petani responden dari tiap desa ditentukan secara purposif dengan kriteria petani tersebut menanam lada dalam tiga tahun terakhir dan memiliki tanaman lada saat wawancara Data yang dikumpulkan meliputi karakteristik petani responden (tingkat pendidikan umur kepemilikan lahan luas lahan garapan pengalaman bertani lada) dan pengetahuan sikap dan tindakan petani dalam pengelolaan tanaman lada (pemilihan bibit pengolahan
lahan pengetahuan terhadap penyebab dan gejala penyakit kuning teknik-teknik pengendalian penyakit kuning yang sudah digunakan dan berikut alasannya serta kendala yang dihadapi petani dalam pengendalian penyakit kuning selama ini) Data yang diperoleh dianalisis menggunakan program Microsoft Excel
HASIL
Hasil survei menunjukkan rata-rata ke-jadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan berkisar antara 33 dan 41 (Tabel 1) Persentase kejadian penyakit kuning paling tinggi ditemukan 40 di Desa Petaling Kabupaten Bangka 60 di Desa Sungkap Kabupaten Bangka Tengah dan 48 di Desa Nyelanding Kabupaten Bangka Selatan Kejadian penyakit ini tergolong tinggi diduga berkaitan dengan umur tanaman lada yang diamati relatif masih muda yaitu baru berumur 2 tahun dan belum berproduksi Contoh akar dari tanaman lada yang terinfeksi penyakit kuning menunjukkan lada yang membengkak yang merupakan gejala serangan nematoda Meloidogyne spp dan sebagian lagi akar membusuk yang merupakan gejala serangan nematoda R similis yang sudah lanjut
Hasil survei menunjukkan bahwa lada masih merupakan komoditas yang banyak diusahakan oleh petani di wilayah Bangka Umumnya petani menanam lada pada lahan milik sendiri atau milik keluarga dan sebagian lagi dengan menyewa lahan milik petani lain Rata-rata luasan lahan tanaman lada antara 1ndash2 ha walaupun sebagian besar terutama di wilayah Bangka Selatan 50 petani responden memiliki lahan tanaman lada lebih dari 2 ha Sebagian besar lada ditanam dengan pola tumpang sari dengan tanaman karet atau kelapa sawit namun akhir-akhir ini petani kembali menanam dengan pola monokultur (Tabel 2) Lebih dari 70 varietas lada yang ditanam oleh petani di wilayah Bangka adalah varietas Lampung Daun Lebar (LDL) dan sisanya adalah varietas Merapin dan Jambi
Hampir semua petani responden menyatakan bahwa gangguan hama dan
10
Kejadian penyakit = times 100nN dengan
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-
J Fitopatol Indones Munif dan Sulistiawati
penyakit merupakan masalah utama dalam budi daya lada di Bangka bahkan lebih penting dari modal (Tabel 3) Menurut petani penyakit kuning merupakan penyakit yang paling merusak tanaman lada dan merugikan petani Namun ternyata pengetahuan petani tentang penyakit kuning masih tergolong sangat kurang Beberapa petani menduga bahwa penyebab penyakit kuning adalah cendawan pupuk asal bibit air dan tanah yang tidak subur namun sebagian besar petani tidak mengetahui penyebab pasti penyakit ini (Tabel 3)
Petani responden pada umumnya mengetahui gejala penyakit kuning yaitu berupa perubahan warna tanaman menjadi kuning namun belum mengetahui secara pasti tahapan perkembangan gejala penyakit ini Beberapa petani mengira bahwa gejala menguning pada tanaman merupakan hal yang biasa terjadi di lapangan sehingga beberapa petani membiarkannya saja tetapi sebagian melakukan tindakan pengendalian (Tabel 4) Pengendalian penyakit kuning yang banyak dilakukan oleh petani tidak hanya dengan
11
Tabel 1 Rata-rata persentase kejadian penyakit kuning pada tanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
Kabupaten
Bangka
Bangka Tengah
Bangka Selatan
Desa pengamatanRata-rata kejadian
penyakit ()Persentase kejadian
penyakit ()3333
3067
4133
482428602016443248
PetalingKemujaCengkong AbangSungkapCeluakTeruAir GegasDelasNyelanding
Tabel 2 Karakteristik budi daya lada oleh petani responden di Kabupaten Bangka Bangka Tengah dan Bangka Selatan
PersenPeubah
Bangka
Luasan tanaman lada lt1 ha 1-2 ha gt2 haPola tanaman lada Monokultur TumpangsariVarietas lada yang ditanam LDL (Lampung) Merapin JambiPenggunaan pupuk organik Ya TidakPenggunaan pupuk sintetik NPK Urea TSP KCl
Jumlah petani
Bangka Selatan
Persen Jumlah petani Persen
Jumlah petani
Bangka Tengah
9274
733
3055
1525
402000
225 675 100
175 825
750 120 120
375 625
100 50 0 0
61915
832
3190
2317
401700
150475375
200800
77532500
575425
1004250000
41620
2020
30100
2020
4013126
100400500
500500
75025000
500500
100325300150
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-
J Fitopatol Indones Munif dan Sulistiawati
12
Tabel 3 Gangguan hama dan penyakit pada pertanaman lada di Kabupaten Bangka Bangka Tengah dan Bangka Selatan menurut petani responden
Peubah
Permasalahan utama usaha tani lada Hama dan penyakit Modal usahaPenyakit utama lada yang paling merugikan Penyakit kuning Penyakit Busuk pangkal batang Penyakit oleh virusPenyebab penyakit kuning Tidak tahu Asal dari bibit Asal dari air Asal dari pupuk Asal dari tanah Jenis jamur
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
346
3721
3010333
850150
9255025
75025
0757575
382
3899
2546050
95050
950225225
62510015000
12500
364
404
11
2840080
900100
100100275
7001000000
20000
Tabel 4 Beberapa teknik pengendalian terhadap penyakit kuning pada lada yang dilakukan oleh petani responden
Petani responden bisa memilih atau memberikan jawaban lebih dari satu
Peubah
Sikap petani terhadap penyakit kuning Dibiarkan DikendalikanCara pengendalian terhadap penyakit kuning yang pernah dilakukan Dicabut dan diganti Pemberian kapur Pestisida sintetik Tanah bekas penyakit dibakar Pemberian tanah atau bahan bekas pembakaran Aplikasi vapam Aplikasi bahan organik
BangkaJumlah petani
Bangka Selatan
Jumlah petani Jumlah
petani
Bangka Tengah
1030
1210302
8
03
250750
30025075050
200
0075
832
020320
4
03
200800
0050080000
100
0075
436
4123400
2
200
150900
10030085000
50
5000
menggunakan pestisida sintetik namun juga beberapa cara pengendalian lainnya seperti penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar aplikasi bahan organik sampai
aplikasi jenis fungisida vapam (Tabel 4) Cara pengendalian penyakit kuning yang paling banyak dilakukan oleh petani responden ialah menggunakan pestisida sintetik (Tabel 5)
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-
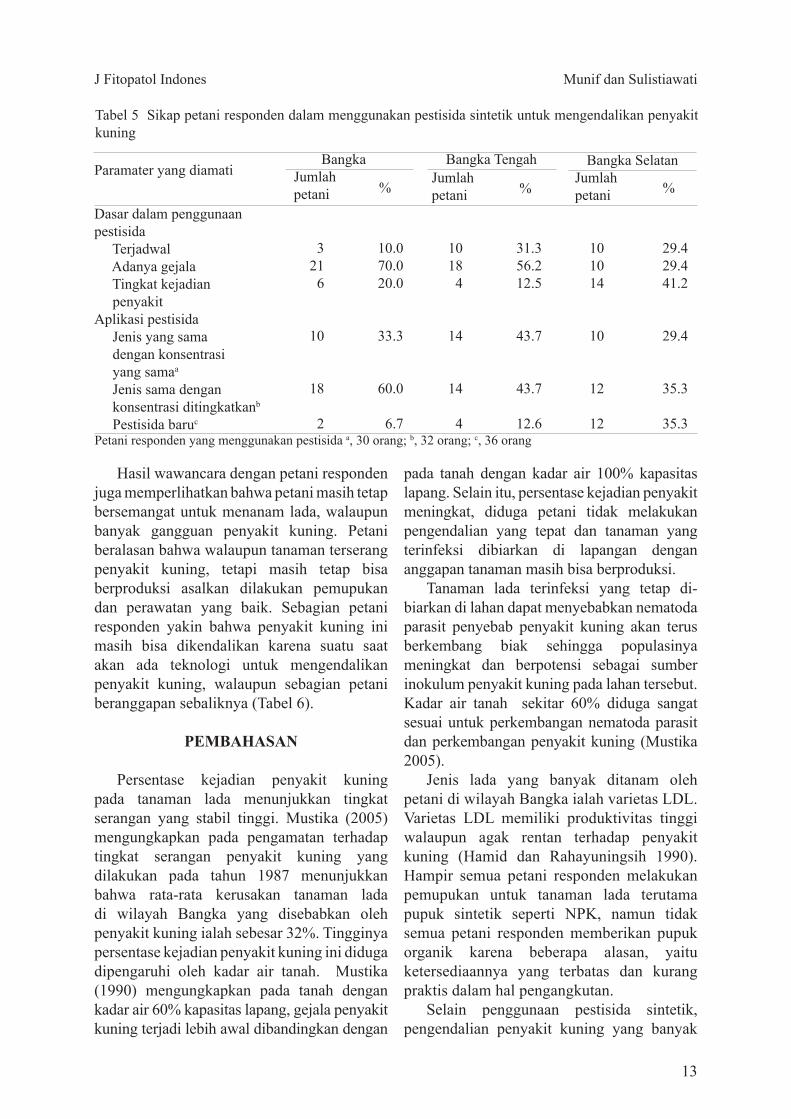
J Fitopatol Indones Munif dan Sulistiawati
13
Tabel 5 Sikap petani responden dalam menggunakan pestisida sintetik untuk mengendalikan penyakit kuning
Petani responden yang menggunakan pestisida a 30 orang b 32 orang c 36 orang
Paramater yang diamati
Dasar dalam penggunaan pestisida Terjadwal Adanya gejala Tingkat kejadian penyakitAplikasi pestisida Jenis yang sama dengan konsentrasi yang samaa
Jenis sama dengan konsentrasi ditingkatkanb
Pestisida baruc
BangkaJumlah petani
Bangka Selatan
Jumlah petani
Jumlah petani
Bangka Tengah
3216
10
18
2
100700200
333
600
67
10184
14
14
4
313562125
437
437
126
101014
10
12
12
294294412
294
353
353
Hasil wawancara dengan petani responden juga memperlihatkan bahwa petani masih tetap bersemangat untuk menanam lada walaupun banyak gangguan penyakit kuning Petani beralasan bahwa walaupun tanaman terserang penyakit kuning tetapi masih tetap bisa berproduksi asalkan dilakukan pemupukan dan perawatan yang baik Sebagian petani responden yakin bahwa penyakit kuning ini masih bisa dikendalikan karena suatu saat akan ada teknologi untuk mengendalikan penyakit kuning walaupun sebagian petani beranggapan sebaliknya (Tabel 6)
PEMBAHASAN
Persentase kejadian penyakit kuning pada tanaman lada menunjukkan tingkat serangan yang stabil tinggi Mustika (2005) mengungkapkan pada pengamatan terhadap tingkat serangan penyakit kuning yang dilakukan pada tahun 1987 menunjukkan bahwa rata-rata kerusakan tanaman lada di wilayah Bangka yang disebabkan oleh penyakit kuning ialah sebesar 32 Tingginya persentase kejadian penyakit kuning ini diduga dipengaruhi oleh kadar air tanah Mustika (1990) mengungkapkan pada tanah dengan kadar air 60 kapasitas lapang gejala penyakit kuning terjadi lebih awal dibandingkan dengan
pada tanah dengan kadar air 100 kapasitas lapang Selain itu persentase kejadian penyakit meningkat diduga petani tidak melakukan pengendalian yang tepat dan tanaman yang terinfeksi dibiarkan di lapangan dengan anggapan tanaman masih bisa berproduksi
Tanaman lada terinfeksi yang tetap di-biarkan di lahan dapat menyebabkan nematoda parasit penyebab penyakit kuning akan terus berkembang biak sehingga populasinya meningkat dan berpotensi sebagai sumber inokulum penyakit kuning pada lahan tersebutKadar air tanah sekitar 60 diduga sangat sesuai untuk perkembangan nematoda parasit dan perkembangan penyakit kuning (Mustika 2005)
Jenis lada yang banyak ditanam oleh petani di wilayah Bangka ialah varietas LDL Varietas LDL memiliki produktivitas tinggi walaupun agak rentan terhadap penyakit kuning (Hamid dan Rahayuningsih 1990) Hampir semua petani responden melakukan pemupukan untuk tanaman lada terutama pupuk sintetik seperti NPK namun tidak semua petani responden memberikan pupuk organik karena beberapa alasan yaitu ketersediaannya yang terbatas dan kurang praktis dalam hal pengangkutan
Selain penggunaan pestisida sintetik pengendalian penyakit kuning yang banyak
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-
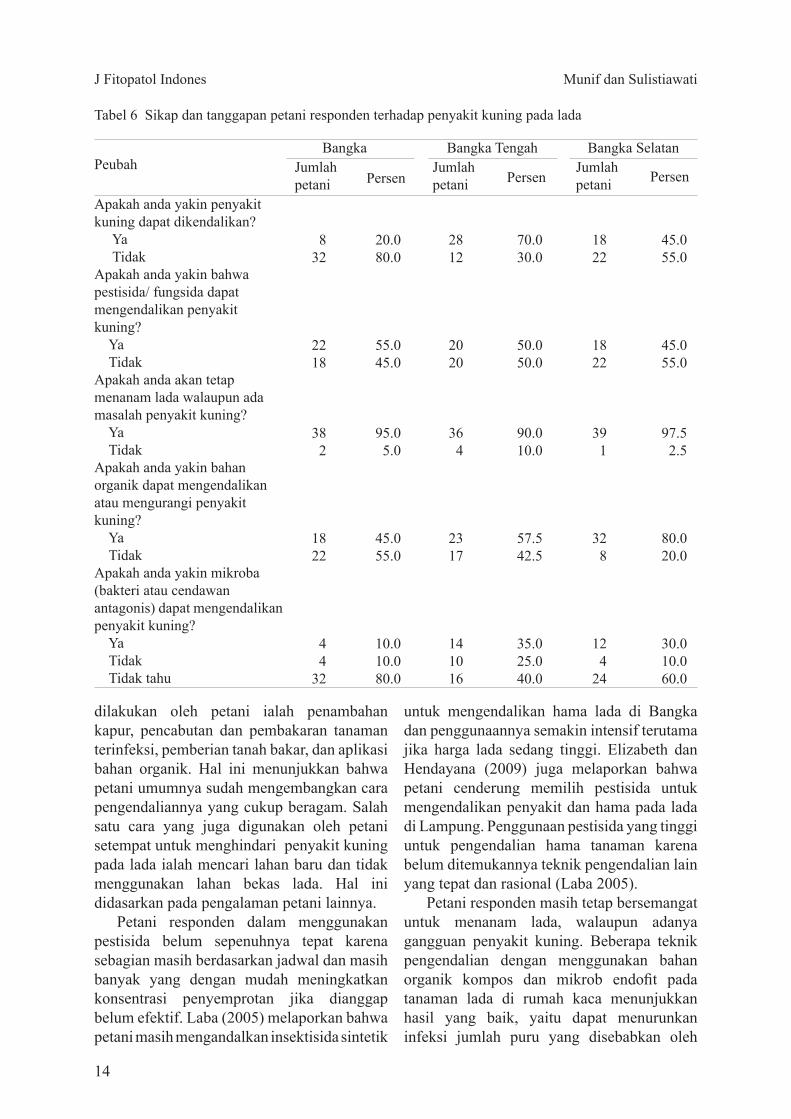
J Fitopatol Indones Munif dan Sulistiawati
dilakukan oleh petani ialah penambahan kapur pencabutan dan pembakaran tanaman terinfeksi pemberian tanah bakar dan aplikasi bahan organik Hal ini menunjukkan bahwa petani umumnya sudah mengembangkan cara pengendaliannya yang cukup beragam Salah satu cara yang juga digunakan oleh petani setempat untuk menghindari penyakit kuning pada lada ialah mencari lahan baru dan tidak menggunakan lahan bekas lada Hal ini didasarkan pada pengalaman petani lainnya
Petani responden dalam menggunakan pestisida belum sepenuhnya tepat karena sebagian masih berdasarkan jadwal dan masih banyak yang dengan mudah meningkatkan konsentrasi penyemprotan jika dianggap belum efektif Laba (2005) melaporkan bahwa petani masih mengandalkan insektisida sintetik
untuk mengendalikan hama lada di Bangka dan penggunaannya semakin intensif terutama jika harga lada sedang tinggi Elizabeth dan Hendayana (2009) juga melaporkan bahwa petani cenderung memilih pestisida untuk mengendalikan penyakit dan hama pada lada di Lampung Penggunaan pestisida yang tinggi untuk pengendalian hama tanaman karena belum ditemukannya teknik pengendalian lain yang tepat dan rasional (Laba 2005)
Petani responden masih tetap bersemangat untuk menanam lada walaupun adanya gangguan penyakit kuning Beberapa teknik pengendalian dengan menggunakan bahan organik kompos dan mikrob endofit pada tanaman lada di rumah kaca menunjukkan hasil yang baik yaitu dapat menurunkan infeksi jumlah puru yang disebabkan oleh
14
Peubah
Apakah anda yakin penyakit kuning dapat dikendalikan Ya TidakApakah anda yakin bahwa pestisida fungsida dapat mengendalikan penyakit kuning Ya TidakApakah anda akan tetap menanam lada walaupun ada masalah penyakit kuning Ya TidakApakah anda yakin bahan organik dapat mengendalikan atau mengurangi penyakit kuning Ya TidakApakah anda yakin mikroba (bakteri atau cendawan antagonis) dapat mengendalikan penyakit kuning Ya Tidak Tidak tahu
Persen
BangkaJumlah petani
Bangka Selatan
Persen Jumlah petani Persen Jumlah
petani
Bangka Tengah
832
2218
382
1822
44
32
200800
550450
95050
450550
100100800
2812
2020
364
2317
141016
700300
500500
900100
575425
350250400
1822
1822
391
328
124
24
450550
450550
97525
800200
300100600
Tabel 6 Sikap dan tanggapan petani responden terhadap penyakit kuning pada lada
J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Munif dan Sulistiawati
nematoda Meloidogyne spp dibandingkan dengan kontrol (Munif dan Harni 2012) Manohara dan Wahyuno (2009) juga melapor-kan bahwa penggunaan bahan organik bungkil jarak dapat mengendalikan penyakit kuning dan penyakit busuk pangkal batang Pengembangan pengendalian nematoda parasit berbasis pada komponen biologi dengan pemanfaatan bahan organik dan mikrob antagonis adalah teknik yang tepat karena lebih murah dan ramah lingkungan (Sikora 1992 Munif dan Kristina 2012)
Upaya untuk mengembangkan teknologi pengendalian penyakit kuning pada lada menghidupkan kelompok tani dan program sekolah lapang pengendalian hama terpadu (Elizabeth dan Hendayana 2009) Jumlah petani yang pernah mengikuti SLPHT lada masih sangat terbatas Sebagian besar petani responden sangat mengharapkan adanya kegiatan penyuluhan dan pelatihan mengenai penyakit kuning dan cara pengendaliannya secara teratur maupun cara budi daya lada yang lebih baik daripada pihak pemerintah Sebagian besar masih cukup aktif dalam kegiatan kelompok tani terutama karena sangat diperlukan untuk memperoleh pembagian pupuk subsidi
Penyakit kuning merupakan permasalahan utama petani dalam budi daya lada di wilayah Bangka Terbatasnya informasi yang dimiliki oleh petani mengenai penyakit kuning menyebabkan perkembangan penyakit ini masih tetap tinggi Sampai saat ini belum ditemukan strategi atau cara pengendalian penyakit kuning yang efektif Evaluasi dan pengembangan teknologi pengendalian penyakit kuning yang efektif sangat diperlu-kan dalam rangka meningkatkan produktivitas lada khususnya di wilayah Bangka
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Penelitian dan Pengembangan Kementerian Pertanian atas dukungan dana penelitian melalui program KKP3T
DAFTAR PUSTAKA
Daras U Pranowo D 2009 Kondisi kritis lada putih Bangka Belitung dan alternatif pemulihannya J Litbang Pert 28(1)1ndash6
Elizabeth R Hendayana R 2010 Peran dan peluang SL-PHT komoditi lada mempengaruhi kognitif petani perkebunan rakyat (Studi kasus Propinsi Lampung) Bogor (ID) Pusat Penelitian Sosial Ekonomi Pertanian
Hamid A Rahayuningsih ST 1990 Identifikasi dan pengenalan empat varietas utama tanaman lada Di dalam Simposium I Hasil Penelitian dan Pengembangan Tanaman Industri Bogor (ID) Pusat Penelitian dan Pengembangan Tanaman Industri hlm 586ndash590
Laba IW 2005 Kepik Renda lada Doconocoris hewetti (Dist) (Hemiptera Tingidae) Biologi kelimpahan populasi dan pengaruhnya terhadap kehilangan hasil [disertasi] Bogor (ID) Institut Pertanian Bogor
Manohara D Wahid P Wahyono D Nuryani Y Mustika I Laba IW Yuhono Rivai AM Saefudin 2006 Status Teknologi Tanaman Lada Di dalam Prosiding Status Teknologi Tanaman Rempah dan Aneka Tanaman Industri 2006 26 Sep Parungkuda-Sukabumi (ID) Pusat Penelitian Tanaman Perkebunan Deptan hlm 1ndash54
Manohara D Wahyuno D 2009 Kontroversi penggunaan bungkil jarak (Ricinus communis) pada penyakit busuk pangkal batang dan penyakit kuning tanaman lada Warta Penel Pengembangan Tan Indust15(2)1ndash3
Munif A Harni R 2012 Keefektifan bakteri endofit untuk mengendalikan nematoda parasit Meloidogyne incognita pada tanaman lada Bul Riset Tan Rempah Aneka Indust 2(3)377ndash382
Munif A Kristiana 2012 Hubungan bakteri endofit dan nematoda parasit penyebab penyakit kuning pada tanaman lada di
15
J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Munif dan Sulistiawati
provinsi Bangka Belitung Bul RISTRI 3(1)71ndash78
Mustika I 1990 Studies on the interaction of Meloidogyne incognita Radopholus similis and Fusarium solani on black pepper (Piper nigrum L) [disertasi] Netherland (NL) Wageningen AgricUniv
Mustika I 2005 Penyakit kuning pada tanaman lada dan cara pengendaliannya Di dalam Materi Pelatihan Teknologi Imunisasi Silang untuk Pengendalian
OPT Vanili Lada dan Jambu Mete 2005 Agu 22ndash26 Bogor (ID) Balai Penelitian Tanaman Rempah dan Obat dan Direktorat Perlindungan Perkebunan Ditjen Perkebunan Deptan hlm 28
Sikora RA 1992 Management of the antagonistic potential in agriculture system for the biological control of plant parasitic nematodes Annu Rev Phytopathol 30245ndash270 DOI httpdxdoiorg101146annu revpy30090192001333
16
Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

Volume 10 Nomor 1 Februari 2014Halaman 17ndash22
DOI 1014692jfi10117
Sensitivitas Metode Serologi dan Polymerase Chain Reactionuntuk Mendeteksi Bean Common Mosaic Potyvirus
pada Kacang Panjang
Sensitifity of Serology and Polymerase Chain Reaction Methods for Detection of Bean Common Mosaic Potyvirus in Yard Long Bean
Sherli Anggraini Sri Hendrastuti HidayatInstitut Pertanian Bogor Bogor 16680
ABSTRAK
Penyakit mosaik kacang panjang yang disebabkan Bean common mosaic potyvirus (BCMV) merupakan penyakit penting yang berpengaruh pada produksi tanaman Metode umum untuk mendeteksinya ialah dengan serologi dan polymerase chain reaction (PCR) Penelitian ini bertujuan mengevaluasi tiga metode deteksi yaitu indirect enzym-linked immunosorbent assay (I-ELISA) dot immunobinding assay (DIBA) dan reverse transcription (RT)-PCR yang sensitif sebagai metode diagnosis BCMV pada kacang panjang Sampel yang digunakan adalah isolat BCMV asal Cirebon yang dipelihara pada tanaman kacang panjang di rumah kaca Batas sensitivitas masing-masing metode diuji melalui pengenceran cairan perasan tanaman dan antiserum untuk I-ELISA dan DIBA serta cDNA sebagai template pada RT-PCR Batas sensitivitas masing-masing metode berbeda-beda metode I-ELISA ialah pada tingkat pengenceran cairan perasan 10-3 dan pengenceran antiserum 10-2 metode DIBA ialah pada tingkat pengenceran cairan perasan 10-5 dan metode RT-PCR pada pengenceran cDNA 10-4
Kata kunci enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
ABSTRACT
Mosaic disease in yard long bean is caused by Bean common mosaic potyvirus (BCMV) and has been reported to affect yield Common method to detect infection of BCMV involves serological assay and polymerase chain reaction (PCR) The aims of this research is to assess the sensitivity of three methods ie Indirect Enzym-Linked Immunosorbent Assay (I-ELISA) Dot Immunobinding Assay (DIBA) and reverse transcription (RT)-PCR as detection method for BCMV infection in yard long bean Sensitivity level of the methods was evaluated by diluting plant extract and antisera for I-ELISA and DIBA and cDNA as template in RT-PCR Virus isolate from Cirebon was maintained in yard long bean in screenhouse and used for the assessment Absorbance value of ELISA showed that dilution end point for I-ELISA was reached at 10-3 and 10-2 of plant extract and antisera dilution respectively Positive infection was still detected using DIBA when the plant extract was diluted up to 10-5 based on development of color intensity on nitrocellulose membrane Specific viral DNA fragment was still amplified when cDNA was diluted up to 10-4 indicated higher sensitivity level of RT-PCR method
Key words enzym-linked immunosorbent assay dot immunobinding assay reverse transcription-polymerase chain reaction
Alamat penulis korespondensi Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper Kampus Darmaga Bogor 16680Tel 0251-8629364 Faks 0251-8629362 Surel srihendrastutihidayatgmailcom
17
ISSN 2339-2479
J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Anggraini dan Hidayat
PENDAHULUAN
Bean common mosaic potyvirus (BCMV) termasuk dalam famili Potyviridae kelompok virus tumbuhan yang terbesar Virus ini terbawa benih yang dapat ditularkan secara mekanis melalui cairan perasan tanaman dan ditularkan secara nonpersisten oleh beberapa jenis kutudaun termasuk Aphis craccivora (Nalini et al 2006) Penyakit mosaik yang disebabkan oleh BCMV merupakan penyakit penting pada tanaman kacang-kacangan karena dapat menyebabkan kehilangan hasil hingga 98 (Verma dan Gupta 2010) Sejak tahun 2008 persentase insidensi penyakit mosaik kacang panjang dilaporkan di beberapa daerah di Jawa Barat Jawa Tengah dan Banten meningkat Damayanti et al (2009) melaporkan bahwa salah satu virus yang berasosiasi dengan penyakit mosaik tersebut ialah Bean common mosaic potyvirus
Metode yang umum dilakukan untuk mendeteksi infeksi virus pada tanaman adalah uji serologi yang didasarkan pada reaksi antara antigen (virus) dan antibodi seperti metode enzyme-linked immunosorbent assay (ELISA) (Nalini et al 2006 Verma dan Gupta 2010) metode gel double-diffusion test dot immunobinding assay (DIBA) immuno-blotting atau western blotting (Xie et al 2013) Metode deteksi lain yang sering digunakan dewasa ini ialah metode deteksi molekuler polymerase chain reaction (PCR) yang memanfatkan sifat spesifik urutan nukleotida virus Pemilihan metode deteksi yang tepat sangat menentukan hasil deteksi karena metode yang kurang baik akan menyebabkan kesalahan pada identifikasi penyebab penyakit
Penelitian bertujuan mengevaluasi tiga metode deteksi virus yaitu ELISA DIBA dan RT-PCR sebagai metode yang sensitif untuk mendeteksi infeksi BCMV pada kacang panjang
BAHAN DAN METODE
Sumber Inokulum VirusIsolat BCMV yang digunakan berasal dari
Cirebon yang merupakan koleksi Laboratorium
Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian Institut Pertanian Bogor Isolat tersebut diperbanyak pada tanaman kacang panjang varietas Parade melalui penularan secara mekanis
Metode Indirect Enzyme-Linked Immunosorbent Assay
Sampel daun tanaman terinfeksi BCMV digerus dalam buffer coating dengan perbandingan 1100 (bv) Cairan perasan tanaman hasil gerusan sebanyak 100 microL dimasukkan ke dalam masing-masing sumur pada plat mikrotiter ELISA dan diinkubasi pada suhu 4 degC selama 24 jam Sap tanaman dibuang dan sumur dicuci dengan phosphate buffer saline tween-20 (PBST) sebanyak 7 kali Anti serum BCMV dilarutkan dalam bufer ECL (2 g bovine serum albumin 20 g PVP 02 g NaN3) dengan perbandingan 1200 (bv) selanjutnya sebanyak 100 microL antiserum BCMV dimasukkan ke dalam sumur plat mikrotiter ELISA dan diinkubasi pada suhu 37 degC selama 2 jam Sumur dicuci kembali dan kemudian diisi dengan 100 microL konjugat yang dilarutkan dalam bufer ECL dengan perbandingan 11000 (bv) dan diinkubasi pada suhu ruang selama 2 jam Sumuran dicuci kembali sebanyak 7 kali dengan PBST selanjutnya diisi dengan 100 microL PNP yang dilarutkan dalam bufer PNP Setelah diinkubasi pada suhu ruang selama 30 menit pengamatan dilakukan secara kuantitatif menggunakan ELISA reader (Bio-RAD 550) pada panjang gelombang 405 nm Reaksi dihentikan dengan cara menambahkan larutan NaOH 3 M dalam masing-masing sumur Pada setiap pengujian disertakan kontrol negatif yaitu tanaman sehat dan bufer
Evaluasi metode I-ELISA dilakukan untuk menentukan batas sensitivitas yaitu dengan mengencerkan sap tanaman dan antiserum Sap tanaman diencerkan secara berseri sampai 10-6 dan antiserum diencerkan hingga 10-5
Metode Dot Immunobinding Assay Metode DIBA dilakukan berdasarkan
metode Mahmood et al (1997) Membran nitroselulosa direndam dalam metanol 100
18
J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Anggraini dan Hidayat
selama 10 detik dan dikeringanginkan Jaringan daun tanaman terinfeksi BCMV digerus dalam tris buffer saline (TBSTris-HCl 002 M dan NaCl 015 M pH 75) dengan perbandingan 110 (bv) Sebanyak 10 μL cairan perasan tanaman diteteskan di atas membran nitroselulosa Setelah tetesan sampel kering membran direndam di dalam 10 mL larutan blocking non-fat milk 2 dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2 Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam dengan menggunakan EYELA multi shaker MMS Membran kemudian dicuci 5 kali dengan dH2O setiap pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm Membran selanjutnya direndam dalam 5 mL TBS yang mengandung antibodi 5 μL ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi semalam pada suhu kamar sambil digoyang dengan kecepatan 50 rpm Membran kemudian dicuci sebanyak 5 kali dengan Tween 005 dalam TBS (TBST) Membran selanjutnya direndam dalam 5 mL TBS yang mengandung 5 μL konjugat (goat anti rabbit-IgG Sigma USA) ditambah non-fat milk dengan konsentrasi akhir 2 dan kemudian membran diinkubasi selama 60 menit sambil digoyang dengan kecepatan 50 rpm Membran selanjutnya dicuci kembali dengan TBST dan direndam selama 5 menit dalam 10 mL bufer substrat (Tris-HCl 01 M NaCl 01 M dan MgCl2 5 mM) yang mengandung 66 μL nitro blue tetrazolium (NBT) dan 30 μL bromo chloro indolil phosphate (BCIP) Bila reaksi positif akan terjadi perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dapat dihentikan dengan merendam membran dalam dH2O
Evalusi metode DIBA dilakukan dengan perlakuan pengenceran sap tanaman hingga 10-6 dan penggunaan dua membran yang berbeda yaitu Amersham HybondTM-P dan Nitropure GE untuk menentukan sensitivitas metode deteksi
Metode Reverse Transcription-Polymerase Chain Reaction
Tahapan metode deteksi BCMV meng-gunakan metode RT-PCR terdiri atas tahapan ekstraksi RNA total transkripsi balik dan amplifikasi cDNA Metode ekstraksi RNA dilakukan menggunakan NucleoSpin RNA Plant mini kit dan hasil ekstrak disimpan pada suhu -80 degC sampai akan digunakan
Reaksi transkripsi balik (10 μL) terdiri atas 22 μL water free nuclease 2 μL bufer RT 10x 035 μL dNTP 10 mM 075 μL oligo d(T) 10 μM 035 μL RNAse Inhibitor 035 μL MmULV RT dan 2 μL RNA Reaksi transkripsi balik dilakukan menggunakan thermal cycler (Gene Amp PCR System 9700) dengan kondisi yang telah diatur sebagai berikut 25 degC selama 5 menit 37 degC selama 90 menit dan 70 degC selama 15 menit Hasil akhir reaksi transkripsi balik adalah produk cDNA yang akan digunakan pada reaksi selanjutnya yaitu amplifikasi
Amplifikasi dilakukan menggunakan pasangan primer BIC-cpF (5rsquo-TCA GGA ACT GGG CAG CCG CAA C-3rsquo) dan BIC-cpR (5rsquo-CTG CGG GGA ACC CAT GCC AAG-3rsquo) (Damayanti et al 2009) Reaksi amplifikasi (25 μL) terdiri atas 163 μL water free nuclease 25 μL PCR buffer 02 μL dNTP 25 mM 25 μL sucrose creasol masing-masing 1μL primer BIC-F dan BIC-R (masing-masing 10 microM) 1 μL taq polymerase 1 μL cDNA hasil RT Semua bahan tersebut diisi ke dalam tabung mikro selanjutnya tabung mikro tersebut dimasukkan ke dalam thermal cycler (Gene Amp PCR System 9700) Amplifikasi BCMV dilakukan sebanyak 35 siklus mengikuti metode Udayashankar et al (2010) melalui beberapa tahapan pemisahan utas DNA pada suhu 94 degC selama 2 menit penempelan primer pada DNA 68 degC selama 1 menit dan sintesis DNA pada suhu 72 degC selama 1 menit Khusus untuk siklus terakhir ditambah tahapan sintesis selama 10 menit kemudian siklus berakhir dengan suhu 4 degC
Evaluasi metode RT-PCR dilakukan dengan perlakuan pengenceran cDNA hingga 10-5 untuk menentukan batas cDNA yang dapat digunakan untuk deteksi BCMV
19
J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Anggraini dan Hidayat
HASIL
Deteksi BCMV Pengamatan pada metode ELISA
secara visual terhadap perubahan warna yang terjadi pada plat mikrotiter ELISA menunjukkan perbedaan intensitas warna kuning Warna kuning terlihat sangat jelas pada pengenceran sap dengan konsentrasi 10-2 dan warna kuning semakin berkurang dengan semakin tingginya pengenceran sap tanaman Demikian pula dengan pengenceran antiserum semakin tinggi pengenceran maka intensitas warna kuning akan semakin berkurang Nilai absorbansi ELISA (NAE) pada 405 nm untuk pengenceran sap dengan konsentrasi 10-2 dapat mencapai lebih dari 6 kali NAE kontrol negatif bila konsentrasi antiserum yang digunakan ialah 100 dan 10-1 (Tabel 1) Nilai absorbansi tersebut menurun mengikuti faktor pengenceran antiserum Nilai absorbansi ELISA pada pengenceran sap dengan konsentrasi 10-3 dapat mencapai lebih dari 2 kali NAE kontrol negatif untuk konsentrasi antiserum 100 dan 10-1 dan NAE akan menurun mengikuti faktor pengenceran antiserum Batas sensitivitas pengujian dengan metode I-ELISA ialah pada tingkat peng-enceran sap tanaman dengn konsentrasi 10-3 dengan pengenceran antiserum 10-2
Metode DIBA menggunakan membran HybondTM-P Amersham dan Nitropure science GE memberikan reaksi positif yang ditandai dengan adanya perubahan warna ungu pada membran (Gambar 1) Reaksi positif yang ditunjukkan oleh warna ungu yang sangat tebal terlihat pada tingkat pengenceran sap dengan
konsentrasi 10-2 namun semakin melemah pada tingkat pengenceran berikutnya Reaksi positif pada membran HybondTM-P Amersham dapat terlihat sampai tingkat pengenceran sap 10-5 sementara pada membran Nitropure science GE reaksi positif terlihat sampai tingkat pengenceran sap 10-4
Amplifikasi RT-PCR menggunakan pasangan primer BIC-cpFBIC-cpR meng-hasilkan produk yang berukuran sekitar 850 pb (Gambar 2) Pengenceran bertingkat terhadap
20
Gambar 1 Reaksi perubahan warna pada metode DIBA dengan berbagai tingkat pengenceran cairan perasan tanaman sakit a menggunakan membran HybondTM-P Amersham b Nitropure science GE Sampel pada masing-masing membran terdiri atas 1 bufer 2 daun tanaman sehat 3 daun tanaman terinfeksi 4 sampai 8 daun tanaman bergejala berturut-turut dengan konsentrasi 10-2 10-3 10-4 10-5 dan 10-6
1
2
3
4
5
6
7
8a b
Tabel 1 Deteksi BCMV menggunakan metode I-ELISA pada berbagai konsentrasi cairan perasan tanaman dan antiserum
- absorban lt 2 kali kontrol negatif + nilai absorban 2 kali kontrol negatif ++ nilai absorban gt 2 ndash 4 kali kontrol negatif +++ nilai absorban gt 4 - 6 kali kontrol negatif ++++ nilai absorban gt 6 kali kontrol negatif
10-3100 10-1Konsentrasi cairan perasan tanaman terinfeksi
Konsentrasi antiserum
10-2
10-3
10-4
10-5
10-6
++++++---
10-410-2 10-5
-----
++++++---
++----
-----
-----
J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Anggraini dan Hidayat
cDNA yang digunakan sebagai DNA cetakan pada tahap amplifikasi menunjukkan bahwa semakin banyak cDNA yang digunakan maka hasil amplifikasi akan semakin jelas Produk amplifikasi DNA berhubungan dengan konsentrasi cDNA Pita DNA masih terbentuk pada pengenceran cDNA 10-4 dan pita tidak terbentuk lagi pada pengenceran 10-5 10-6 dan 10-7
PEMBAHASAN
Deteksi dan identifikasi virus dapat dilakukan berdasarkan karakter biologi dan molekuler Deteksi virus berdasarkan karakter biologi dapat dilakukan dengan pengujian tanaman indikator penularan dan bentuk partikel Deteksi dan identifikasi menggunakan karakter molekuler umumnya dilakukan berdasarkan sifat protein misalnya dengan uji serologi dan sifat asam nukleat dengan hibridisasi DNA dan PCRRT-PCR Metode ELISA dan DIBA merupakan metode serologi yang banyak digunakan untuk mendeteksi dan mengidentifikasi virus tumbuhan Dalam metode ini antiserum dikonjugasikan dengan enzim sehingga bila ditambahkan substrat enzim maka kompleks antigen-antibodi dalam jumlah yang sedikit saja dapat tervisualisasiAnalisis hasil ELISA dapat dilakukan secara kuantitatif dengan spektrofotometer (ELISA reader) sementara hasil DIBA hanya dapat dianalisis secara kualitatif Keuntungan metode ELISA ialah sensitivitas yang sangat
tinggi sehingga dapat mendeteksi virus pada konsentrasi rendah (1ndash10 ng mL-1) dapat menguji sampel dalam jumlah banyak secara cepat menggunakan antiserum sedikit memperoleh data secara kualitatif dan kuantitatif dan tahapan pengujian yang mudah
Deteksi serologi dengan metode I-ELISA memberi hasil positif untuk BCMV sampai pengenceran sap 10-3 dan pengenceran antiserum 10-3 Tingkat pengujian yang lebih sensitif diperoleh dengan metode DIBA yang mampu memberikan hasil positif sampai pengenceran sap 10-5 Batas sensitifitas sap DIBA lebih tinggi dibandingkan dengan metode I-ELISA dan hanya memerlukan antiserum yang lebih sedikit dibandingkan dengan I-ELISA untuk pengujian dengan sampel yang sama
Saat ini metode deteksi yang banyak digunakan ialah metode deteksi berdasarkan sifat molekuler virus Metode ini lebih sensitif akurat dan lebih efisien dalam peng-gunaan waktu dan tenaga Hasil visualisasi elektroforesis menunjukkan bahwa metode RT-PCR berhasil mengamplifikasi BCMV dengan konsentrasi template cDNA yang rendah Hal tersebut menunjukkan keunggulan teknik PCR yang mampu mendeteksi virus dari jaringan tanaman dengan konsentrasi virus yang rendah
Metode deteksi I-ELISA DIBA dan PCR memiliki kelebihan dan kekurangan (Tabel 2) Ditinjau dari segi kemudahan pelaksanaan
1000 pb
850 pb500 pb
M 1 2 3 4 5 6 7 8 9
Gambar 2 Hasil amplifikasi RT-PCR dari tanaman kacang panjang terinfeksi BCMV dengan cDNA yang diencerkan bertingkat menggunakan primer BIC-F dan BIC-R M penanda DNA 1 kb 1 tanaman sehat 2ndash9 sampel cDNA yang diencerkan bertingkat berturut-turut 100 10-1 10-2 10-3 10-4 10-5 10-6 10-7
21
J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Anggraini dan Hidayat
pengujian metode I-ELISA dan DIBA dalam menentukan hasil termasuk metode yang mudah dilakukan dan tidak memerlukan alat yang canggih Hal ini disebabkan hasil I-ELISA dan DIBA dapat ditentukan dengan membandingkan warna substrat antarsampel kontrol positif kontrol negatif dan larutan penyangga Sebaliknya RT-PCR tidak dapat dilakukan pada kondisi lapangan karena memerlukan peralatan yang canggih Teknik RT-PCR memiliki keunggulan pada aspek kecepatan diagnosis Waktu yang dibutuhkan dalam teknik RT-PCR ialah sekitar 65 jam sedangkan untuk teknik I-ELISA dan DIBA sekitar 10ndash15 jam Teknik RT-PCR memiliki tingkat kesulitan yang lebih tinggi daripada teknik I-ELISA dan untuk mendapatkan hasil yang akurat dibutuhkan keterampilan yang tinggi Berdasarkan kebutuhan biaya pengujian I-ELISA dan DIBA memerlukan biaya lebih rendah dibandingkan RT-PCR karena bahan-bahan untuk I-ELISA dan DIBA relatif lebih murah (Kartiningtyas et al 2006)
Ketersediaan metode deteksi virus yang sangat beragam perlu disikapi dengan tepat dan bijaksana terutama untuk menetapkan metode yang akan digunakan Kriteria yang perlu diperhatikan untuk memilih metode deteksi yang akan digunakan ialah sensitivitas spesifikasi waktu pengerjaan dan biaya (Figueira et al 1997)
DAFTAR PUSTAKA
Damayanti TA Alibi OJ Naidu RA Rauf A 2009 Severe outbreak of a yellow mosaic disease on the yard long bean in Bogor West Java Hayati J Biosci16 78ndash82 DOI httpdxdoiorg104308hjb16278
Figueira AR Domier LL DrsquoArcy CJ 1997 Comparison of Technique for Detection of Barley yellow dwarf virus-PAV-IL Plant Dis 81(11)1236ndash1246 DOI 101094PDIS 199781111236
Kartiningtyas Hidayat SH 2006 Deteksi Turnip mosaic virus (TuMV) pada benih dan jaringan daun J HPT Tropica 632ndash40
Mahmood T Hein GL French RC 1997 Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite Plant Dis 81250ndash253 DOI httpdxdoiorg101094PDIS1997813250
Nalini MS Shylaja MD Prakash HS Shetty HS 2006 Production of polyclonal antibody to Bean common mosaic virus and its application in seed health testing Indian J Microbiol 4697ndash108
Udayashankar AC Nayaka SC Kumar HB Mortensen CN Shetty HS Prakash HS 2010 Establishing inoculum threshold levels for Bean common mosaic virus strain blackeye cowpea mosaic infection in cowpea seed African J Biotech 9(53) 8958ndash8969 DOI 105897AJB091066
Verma P Gupta UP 2010 Immunological detection of Bean common mosaic virus in French bean (Phaseolus vulgaris L) Indian J Microbiol 50 263ndash265 DOI httpdxdoiorg101007s12088-010-0019-8
Xie Y Jiao X Zhou X Liu H Ni Y Wa J 2013 Highly sensitive serological methods for detecting Tomato yellow leaf curl virus in tomato plants and whiteflies Virol J 10142 DOI httpdxdoiorg1011861743-422X-10-142
22
Tabel 2 Pembandingan metode I-ELISA DIBA dan RT-PCR untuk mendeteksi BCMV pada kacang panjang
0001 Sedang-tinggi
10-30 45-90
Rendah -Sedang
00002 Tinggi3-65
96 Tinggi
RT-PCRI-ELISA DIBAFaktor Pembanding Metode
Bobot sampel daun (g)Akurasi Waktu (jam)Kapasitas (sampel)Biaya
001 Sedang-tinggi
10-30 45-90
Sedang-tinggi
Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

Penapisan dan Identifikasi Bakteri Agens Biokontrol Penyakit Layu Fusarium Hasil Isolasi dari Rizosfer Pisang
Screening and Identification of Bacteria as Biological Control Agents for Fusarium Wilt Disease from Banana Rhizosphere
Dwi Agustiyani Achirul Nditasari Nur Laili Sarjiya AntoniusLembaga Ilmu Pengetahuan Indonesia Cibinong 16911
ABSTRAK
Penyakit layu Fusarium pada tanaman pisang yang diakibatkan oleh Fusarium oxysporum f sp cubense merupakan salah satu penyakit yang sangat merusak tanaman pisang di daerah tropik Pengendalian penyakit layu secara biologi menjadi salah satu solusi Penelitian ini bertujuan mendapatkan agens pengendali F oxysporum f sp cubense yang potensial Bakteri dan aktinomiset diisolasi dari sampel tanah perakaran tanaman pisang di Lampung dan Cianjur Sebanyak 64 isolat aktinomiset dan 142 isolat bakteri diperoleh dari lokasi tersebut Uji antagonis terhadap F oxysporum f sp cubense dari isolat tersebut menunjukkan bahwa 10 isolat aktinomiset dan 21 isolat bakteri positif memiliki daya hambat Isolat aktinomiset memperlihatkan kemampuan menghambat F oxysporum f sp cubense relatif lebih tinggi dibandingkan dengan bakteri Isolat-isolat yang bersifat antagonis terhadap F oxysporum f sp cubense diuji aktivitas enzim protease kitinase dan selulase secara kualitatif pada medium spesifik Semua isolat aktinomiset yang diuji mempunyai aktivitas enzim kitinase tetapi hanya 5 isolat bakteri mempunyai aktivitas enzim tersebut Sebanyak 13 isolat bakteri memiliki aktivitas enzim protease dan hanya 1 isolat aktinomiset yang mempunyai aktivitas protease Dua isolat bakteri (LII4ND dan LAI-5DW) dan 3 isolat aktinomiset (LAIDW L31DW dan CiIA5DW) mampu menghambat F oxysporum f sp cubense cukup tinggi dan mempunyai aktivitas enzim lisis Isolat memiliki homologi 99 dengan Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) dan Streptomyces sp (CiIA5DW)
Kata kunci antagonis kitinase protease selulase
ABSTRACT
Banana wilt disease caused by Fusarium oxysporum fspcubense is considered as one of the most destructive diseases on banana plants in the tropical region Biological control agents (BCArsquos) have become a promising solution to overcome this disease The objective of this study was to find potential BCArsquos for wilt disease of banana plants Bacteria and actinomycetes were isolated from bananarsquos rhizosphere in Lampung and Cianjur As much as 64 actinomycetes and 142 bacteria isolates were obtained Antagonistic test against F oxysporum f sp cubense of those isolates showed that 21 bacteria and 10 actinomycete isolates have abilities to inhibit F oxysporum f sp cubense Actinomycetes showed relatively higher inhibition against F oxysporum f sp cubense compared to bacteria Isolates which have positive antagonistic activities against F oxysporum f sp cubense were then tested for their protease chitinase and selulase activities qualitatively on specific medium All actinomycetes which were tested had chitinase enzyme activities while only 5 bacterial isolates had chitinase activities On the other hand 13 bacterial isolates showed protease activities and only 1 actinomycete showed
Alamat penulis korespondensi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Jalan Raya Bogor-Jakarta Km 46 Cibinong 16911Tel 021-8765066 Faks 021-8765062 Surel titinagustinyahoocom
Volume 10 Nomor 1 Februari 2014Halaman 23ndash30
DOI 1014692jfi10123ISSN 2339-2479
23
J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Agustiyani et al
protease activity Two bacterial isolates (LII4ND and LAI-5DW) and 3 actinomycetes (LAIDW L31DW and CiIA5DW) which showed high inhibition against F oxysporum f sp cubense and lyses enzymes activities were identified based on 16S rRNA genes Analysis based on GenBank data those isolates have 99 homology to Klebsiella pneumonia (LII4ND) Burkholderia sp (LAI-5DW) Streptomyces sp (LAIDW) Streptomyces sp (L31DW) and Streptomyces sp (CiIA5DW)
Key words antagonistic cellulase khitinase protease
PENDAHULUAN
Penyakit layu Fusarium atau lebih dikenal dengan Panama disease merupakan penyakit mematikan paling penting pada tanaman pisang (Ploetz 2006) Penyakit ini disebabkan oleh Fusarium oxysporum f sp cubense Fusarium menginfeksi akar tanaman kemudian men-jalar ke rizom dan batang semu sehingga menyebabkan kematian pada jaringan atau seluruh bagian tanaman F oxysporum f sp cubense sangat sulit dikendalikan secara tuntas karena hidup sebagai saprob Patogen ini juga dapat bertahan sangat lama di tanah hingga mendapatkan atau mengenali tanaman yang dapat diinfeksi Penyakit layu kemungkinan berasal dari Asia Tenggara tetapi penyakit ini pertama kali dilaporkan di Australia pada tahun 1876 Penyakit ini ditemukan di semua area yang memproduksi pisang kecuali Mediterrenean Melanesia Somalia dan beberapa pulau di Pasifik Selatan (Ploetz 2006) Penelitian mengenai upaya pengendalian F oxysporum f sp cubense telah banyak dilakukan termasuk upaya pengendalian secara kimia dengan menggunakan fungisida (Nel et al 2007) pengendalian secara fisika rotasi tanaman (Huang et al 2012) pengelolaan nutrisi tanah dan varietas pisang yang resisten (Hwang dan Ko 2004) maupun secara biologi menggunakan agens pengendali hayati (Sivamani et al 1988) Pengendalian secara biologi merupakan salah satu alternatif yang menjanjikan karena selain aman secara ekologi dan spesifik terhadap patogen agens pengendali biologi (APB) mampu hidup di tanah dalam jangka waktu lama Pada dekade terakhir perhatian publik kembali tercurah pada penelitian dasar dan pemanfaatan mikrob endofit dan rizosfer sebagai APB Pemanfaatan komunitas mikrob rizosfer sama
pentingnya dengan mikrob endofit karena sistem perakaran merupakan daerah penting penyerapan nutrisi bagi tanaman dan juga daerah yang rentan bagi masuknya penyebab penyakit tanaman yang berasal dari tanah
Bakteri rizosfer seperti Arthrobacter Bacillus Pseudomonas Serratia dan Streptomyces merupakan bakteri agens biokontrol untuk patogen tanaman (Kloepper et al 2004 de Vasconcellos dan Cardoso 2009) Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol dilakukan melalui mekanisme kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glucanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Bacillus sp dan Streptomyces sp diketahui mampu mensekresikan protease dan kitinase yang berpotensi dalam melisiskan dinding sel F oxysporum f sp cubense dan menghambat pertumbuhannya (Kloepper et al 2004 de Azaredo et al 2004 Rodas-Junco et al 2009)
Pada penelitian ini dilakukan penapisan sejumlah isolat bakteri dan aktinomiset yang diisolasi dari tanah perakaran tanaman pisang sebagai agens biokontrol terhadap F oxysporum f sp cubense
BAHAN DAN METODE
Isolat Bakteri dan Aktinomiset Bakteri dan aktinomiset diisolasi dari
rizosfer tanaman pisang yang dikoleksi dari perkebunan pisang di Lampung dan Cugenang-Cianjur Tanah rizosfer perkebunan pisang di Lampung ada tiga kategori yaitu tanah rizosfer dengan tingkat serangan F oxysporum f sp cubense tinggi (gt 20) sedang (6)
24
J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Agustiyani et al
dan rendah (2) Sampel lainnya ialah tanah rizosfer pisang SJ20 yang cukup resisten terhadap serangan F oxysporum f sp cubense dan tanah rizosfer pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense Sampel dari perkebunan pisang di Cianjur ialah tanah rizosfer pisang yang relatif tidak terserang F oxysporum f sp cubense terserang F oxysporum f sp cubense dan selama 5 tahun tidak panen juga tanah rizosfer pisang yang paling tinggi dengan serangan F oxysporum f sp cubense dan serangan penyakit lainnya
Bakteri dan aktinomiset diisolasi dari sampel tanah menggunakan metode sebar dengan pengenceran berseri pada medium yeast soluble agar (YSA) (2 g ekstrak khamir 10 g soluble starch 15 g agar-agar 1 L akuades pH 72) Isolasi bakteri menggunakan medium agar-agar nutrien (10 g ekstrak kaldu 10 g pepton 5 g NaCl 15 g agar-agar dan 1 L akuades) Cawan petri diinkubasi pada suhu 28 degC selama 2‒7 hari
Koloni bakteriaktinomiset dimurnikan pada medium YSA atau agar-agar nutrien Biakan murni bakteri disimpan di tabung reaksi pada medium agar-agar nutrien dan isolat aktinomiset pada medium agar-agar international streptomyces project (ISP2) (4 g ekstrak khamir 10 g ektrak malt 4 g dekstrosa 16 g agar-agar 1 L akuades pH 72) Isolat tersebut diinkubasi pada suhu 28 degC sampai terjadi sporulasi kemudian disimpan pada suhu 4 degC Uji Aktivitas Antagonisme Bakteri dan Aktinomiset terhadap F oxysporum f sp cubense secara in Vitro
Fusarium oxysporum f sp cubense yang digunakan sebagai patogen uji berasal dari Lampung (koleksi Loekas Soesanto Unsoed Purwokerto) Uji aktivitas biokontrol terhadap patogen uji menggunakan cross-plug dengan dua ulangan Satu ose biakan bakteri atau aktinomiset ditumbuhkan di tengah cawan petri yang berisi medium PDA dan diinkubasi pada suhu 28 degC selama 2 hari Masing-masing 1 blok agar-agar patogen uji berdiameter 5 mm ditumbuhkan pada medium PDA pada
kedua sisi cawan dengan jarak 2 cm dari bakteri atau aktinomiset Untuk perlakuan kontrol 1 blok agar-agar patogen uji ditumbuhkan pada medium PDA di kedua sisi cawan petri yang tidak diinokulasi dengan bakteri atau aktinomiset
Setelah 5 hari inkubasi zona penghambatan yang terbentuk dari aktivitas bakteri atau aktinomiset diamati Persentase penghambatan dihitung mengikuti rumus (Nourozian et al 2006)
Penghambatan = ((C-T)C) times 100 dengan
C diameter F oxysporum f sp cubense kontrol T diameter F oxysporum f sp cubense dengan perlakuan agens pengendali hayati
Uji Kualitatif Aktivitas ProteaseUji aktivitas protease secara kualitatif
ditentukan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium nutrient broth (NB) dan di-inkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose biakan bakteri atau aktinomiset diletakkan di bagian tengah cawan petri yang berisi medium protease-skim milk agar 1 pH 70 dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Zona bening yang terbentuk di sekelilingkoloni bakteriaktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan ber-dasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas KitinaseUji aktivitas kitinase secara kualitatif
ditentukan dengan metode disc blank (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48ndash72 jam Selanjutnya 1 ose bakteri atau aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium agar-agar kitin 05 (5 g kitin 3 g glukosa 1 g polipepton
25
J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Agustiyani et al
1 g KH2PO4 05 g MgSO47H2O 20 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC
Pengamatan zona bening yang terbentuk di sekeliling koloni bakteri atau aktinomiset diukur diameternya Aktivitas secara kualitatif ditentukan berdasarkan nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Uji Kualitatif Aktivitas Selulase Uji aktivitas selulase secara kualitatif
ditentukan dengan menggunakan metode disc blank yang telah dimodifikasi (Basha dan Ulaganathan 2002) Isolat bakteri diinokulasi pada medium NB dan diinkubasi pada rotary shaker selama 24 jam sedangkan isolat aktinomiset diinokulasi pada medium ISP2 dan diinkubasi pada rotary shaker selama 48‒72 jam Selanjutnya 1 ose bakteri aktinomiset diletakkan pada bagian tengah cawan petri yang berisi medium carboxymethyl cellulose (CMC) (10 g CMC 02 g MgSO47H2O 075 g KNO3 05 g K2HPO4 002 g FeSO47H2O 004 g CaCl22H2O 2 g ekstrak khamir 1 g glukosa 15 g agar-agar dan 1 L akuades pH 68) dengan 2 ulangan dan diinkubasi pada suhu 28 degC Koloni yang tumbuh pada cawan petri kemudian ditetesi congo red 1 kemudian diamati terbentuknya zona bening di sekelilingnya Aktivitas secara kualitatif ditentukan berdasarkan pada nilai indeks yaitu diameter total zona dibagi diameter koloni bakteri
Identifikasi Bakteri dan Aktinomiset Ekstraksi DNA bakteri dilakukan ber-
dasarkan metode GES Sebanyak plusmn1 ose sel bakteri disuspensikan dalam ependorf (15 mL) yang berisi 1 mL bufer TE dihomogenkan dengan vorteks kemudian disentrifugasi dengan kecepatan 10 000 rpm selama 15 menit pada suhu ruang Supernatan dibuang dan sel disuspensikan dalam 50 microL lisozim dengan konsentrasi 0001 g 15 mL (yang terlebih dahulu dipanaskan pada suhu 37 degC selama 10 menit) dan diinkubasi pada suhu 37 degC selama 30 menit selanjutnya ditambahkan 250 microL reagen GES (guanidium thyocyanat
5 mol L-1 EDTA 100 mmol L-1 dan sarkosyl 05) dihomogenkan secara hati-hati meng-gunakan pipet mikro dan didiamkan selama 10 menit Selanjutnya 125 microL larutan natrium asetat 75 M ditambahkan dan diinkubasi di dalam es selama 10 menit Sebanyak 500 microL kloroform ditambahkan dan dihomogenkan dengan dibolak-balik sebanyak 50 kali dan disentrifugasi dengan kecepatan 10 000 rpm selama 10 menit untuk memisahkan 2 lapisan yang terbentuk Lapisan paling atas ditransfer ke dalam ependorf baru dan ditambah alkohol absolut sebanyak setengah bagian dari volume larutan yang dipindahkan tersebut Ependorf dibolak-balik dengan hati-hati sampai ter-bentuk benang-benang DNA Benang-benang DNA dipindahkan ke ependorf baru dan dicuci dengan etanol 70 dan disentrifugasi Supernatan dibuang DNA dikeringanginkan selama plusmn10 menit dan kemudian disuspensikan dengan 100 microL 02 x TE
Ekstraksi DNA aktinomiset dilakukan menggunakan PowerSoilreg DNA Isolation Kit sesuai petunjuk dari produsen Amplifikasi PCR menggunakan Primer 20F 5rsquoGATTTTGATCCTGGCTCAG3rsquo dan Primer 520R 5rsquoGTGCCAGCAGCCGCGG3rsquo Hasil amplifikasi PCR dari DNA isolat bakteri dan aktinomiset dirunut urutan kode DNAnya dalam format FASTA secara on-line di Genbank (wwwncbinlmnihgov) dengan memasukkan data sikuen ke dalam program BLAST
HASIL
Bakteri dan Aktinomiset dari Tanah Perakaran Tanaman Pisang
Bakteri dan aktinomiset yang berhasil diisolasi dari tanah perakaran pisang per-kebunan pisang di Lampung dan kebun pisang lokal di Cianjur ialah 206 isolat Isolat bakteri terdiri atas 151 jumlahnya lebih banyak dibandingkan dengan aktinomiset yang hanya 55 isolat (Tabel 1)
Aktivitas Penghambatan F oxysporum f sp cubense oleh Isolat Bakteri dan Aktinomiset
Hasil uji antibiosis atau antagonis bakteri dan aktinomiset terhadap patogen uji
26
J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Agustiyani et al
F oxysporum f sp cubense menunjukkan 21 isolat bakteri dan 10 isolat aktinomiset dapat menghambat patogen uji Secara umum isolat aktinomiset memiliki persentase peng-hambatan yang relatif lebih tinggi dibanding-kan dengan bakteri Aktinomiset LAIDW L31DW dan CiIA5DW memperlihatkan tingkat hambatan yang cukup tinggi (45ndash60) dibandingkan dengan isolat bakteri yang hanya mencapai 15ndash45 Isolat bakteri yang memperlihatkan kemampuan penghambatan F oxysporum f sp cubense cukup tinggi ialah bakteri LII4ND dan LAI-5DW
Aktivitas Enzim Protease Kitinase dan Selulase
Aktivitas enzim secara kualitatif ditentukan berdasarkan pada terbentuknya zona bening di sekililing koloni bakteri dan aktinomiset (Tabel 2 Tabel 3) Isolat bakteri yang mempunyai aktivitas enzim protease ialah sebanyak 13 isolat 3 isolat bakteri (LAI-22DW LAI-5DW dan LAII-18DW) mem-punyai aktivitas protease cukup tinggi yang
diindikasikan dari indeks zona bening 5 isolat bakteri (LI2ND LII 4ND LAI-5DW LAII-18DW dan LPII-19DW) mempunyai aktivitas kitinase yang relatif rendah (Tabel 2)
Sebanyak 10 isolat aktinomiset mampu menghambat F oxysporum f sp cubense (Tabel 3) Semua isolat mempunyai aktivitas kitinase dan 7 isolat mempunyai aktivitas selulase Berbeda dengan isolat bakteri yang sebagian besar mempunyai aktivitas protease hanya 2 isolat aktinomiset yang mempunyai aktivitas protease yaitu aktinomiset LPI-5bDW dan L88DW
Dua bakteri diidentifikasi sebagai Klebsiella pneumonia (LII4ND) dan Burkholderia galur LAI-5DW (Tabel 2) serta tiga aktinomiset diidentifikasi semuannya sebagai Streptomyces yaitu galur L31DW LAIDW dan CiIA5DW (Tabel 3)
PEMBAHASAN
Secara umum isolat aktinomiset lebih mampu menghambat F oxysporum f sp cubense dibandingkan dengan bakteri Nourozian et al (2006) juga mengemukakan penghambatan patogen oleh Streptomyces sp lebih tinggi dibandingkan dengan Bacillus subtilis Ada berbagai macam mekanisme penghambatan patogen secara langsung salah satunya ialah melalui mekanisme parasitisme atau pengerusakan dinding sel patogen meng-gunakan enzim hidrolase Enzim hidrolase yang terlibat dalam mekanisme hidrolisis polimer penyusun dinding sel patogen antara lain ialah kitinase protease dan selulase (Ramesh dan Mathivanan 2009 Ara et al 2012) Selain mempunyai hidrolase aktinomiset juga menghasilkan antibiotik dan metabolit sekunder (Adegboye dan Babalola 2012)
Sampai sekarang aktinomiset masih menjadi sumber utama senyawa bioaktif potensial dalam berbagai bidang industri termasuk pertanian Metabolit sekunder dari aktinomiset ini telah terbukti dapat berperan sebagai agens biokontrol (Doumbou et al 2001) Isolat Streptomyces angustmyceticus L31 juga menghasilkan metabolit sekunder
Daerah Tingkat serangan pada perakaran
Jumlah
Lampung Tinggi (gt 20) 29Sedang (6) 32Rendah (2) 17
Rejang Cianjur
Relatif resisten 18
Cukup resisten 25Cianjur Relatif tidak
terserang31
Terserang selama 5 tahun dan tidak dapat panen
28
Serangan paling tinggi dan serangan penyakit lainnya
26
Total 206
Tabel 1 Isolat bakteri dan aktinomiset dari tanah perakaran tanaman pisang dengan tingkat serangan Fusarium oxysporum f sp cubense di Lampung dan Cianjur
Perakaran pisang Rejang Perakaran pisang SJ 20
27
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-
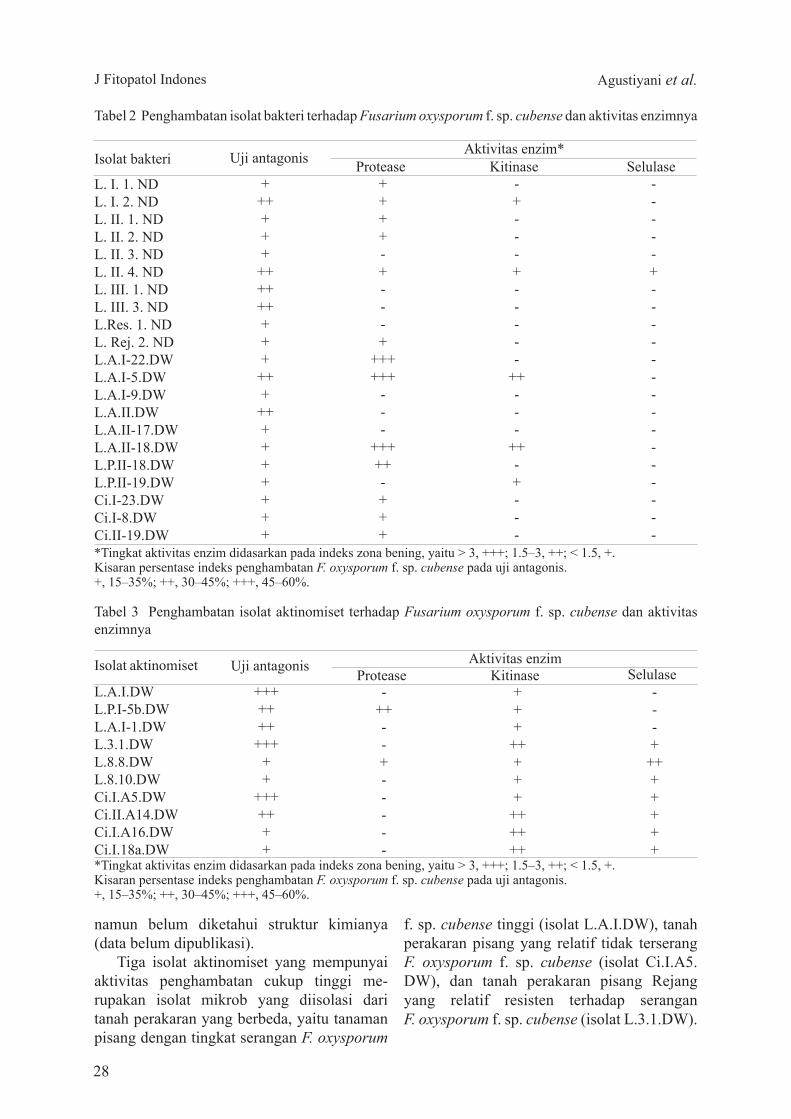
J Fitopatol Indones Agustiyani et al
namun belum diketahui struktur kimianya (data belum dipublikasi)
Tiga isolat aktinomiset yang mempunyai aktivitas penghambatan cukup tinggi me-rupakan isolat mikrob yang diisolasi dari tanah perakaran yang berbeda yaitu tanaman pisang dengan tingkat serangan F oxysporum
f sp cubense tinggi (isolat LAIDW) tanah perakaran pisang yang relatif tidak terserang F oxysporum f sp cubense (isolat CiIA5DW) dan tanah perakaran pisang Rejang yang relatif resisten terhadap serangan F oxysporum f sp cubense (isolat L31DW)
Tabel 2 Penghambatan isolat bakteri terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
Isolat bakteri
L I 1 NDL I 2 NDL II 1 NDL II 2 NDL II 3 NDL II 4 NDL III 1 NDL III 3 NDLRes 1 NDL Rej 2 NDLAI-22DWLAI-5DWLAI-9DWLAIIDWLAII-17DWLAII-18DWLPII-18DWLPII-19DWCiI-23DWCiI-8DWCiII-19DW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
++++++
+++++++++
+++
+++++++++
++++-+---+
++++++
---
+++++-+++
-+---+-----
++---
++-+---
-----+---------------
Tabel 3 Penghambatan isolat aktinomiset terhadap Fusarium oxysporum f sp cubense dan aktivitas enzimnya
Isolat aktinomiset
LAIDW LPI-5bDWLAI-1DW L31DWL88DWL810DWCiIA5DWCiIIA14DWCiIA16DWCiI18aDW
ProteaseAktivitas enzimUji antagonis SelulaseKitinase
+++++++
+++++
+++++++
-++--+-----
+++
+++++
++++++
---+
+++++++
28
Tingkat aktivitas enzim didasarkan pada indeks zona bening yaitu gt 3 +++ 15ndash3 ++ lt 15 + Kisaran persentase indeks penghambatan F oxysporum f sp cubense pada uji antagonis + 15ndash35 ++ 30ndash45 +++ 45ndash60
J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Agustiyani et al
Data ini menunjukkan bahwa tidak ada korelasi antara sumber isolat (jenis tanah perakaran) dan tingkat kemampuan isolat dalam menghambat F oxysporum f sp cubense
Semua isolat aktinomiset mempunyai aktivitas kitinase sedangkan sebagian besar isolat bakteri mempunyai aktivitas protease Apabila dihubungkan antara aktivitas enzim dan kemampuan penghambatan F oxysporum f sp cubense tampak tidak ada korelasi yang nyata Isolat bakteri yang mempunyai aktivitas protease tinggi (LAII-18DW LAI-22DW dan LAI-5DW) ternyata hanya memperlihatkan tingkat hambatan F oxysporum f sp cubense yang relatif rendah (+ atau ++) Sebaliknya isolat aktinomiset yang mempunyai tingkat hambatan terhadap patogen uji cukup tinggi (LAIDW L31DW dan CiIA5DW) ternyata tidak memiliki aktivitas enzim protease dan hanya memiliki aktivitas enzim kitinase dan selulase yang relatif rendah (+ atau ++) Kemampuan penghambatan terhadap F oxysporum f sp cubense tidak hanya disebabkan oleh adanya aktivitas enzim tetapi kemungkinan besar diduga juga oleh adanya metabolit sekunder (antibiotik) yang dihasilkan oleh isolat bakteri atau aktinomiset Penghambatan pertumbuhan F oxysporum f sp cubense oleh bakteri biokontrol telah dilaporkan kompetisi nutrisi oksigen dan habitat sintesis enzim hidrolase seperti kitinase protease dan β-1-3 glukanase serta sekresi senyawa antibiotik atau senyawa anticendawan lainnya (Benyagoub et al 1998 Getha dan Vikineswary 2002 de Azaredo et al 2004 Nourozian et al 2006 Rodas-Junco et al 2009) Jadi kemungkinan kemampuan penghambatan F oxysporum f sp cubense dari isolat-isolat uji disebabkan oleh kombinasi aktivitas enzim hidrolase dan sekresi senyawa antibiotik yang dihasilkan
Lima isolat uji yang menunjukkan hasil potensial ialah bakteri Klebsiella pneumonia (LII4ND) dan Burkholderia sp (LAI-5DW) serta Streptomyces sp L31DW dan dua spesies Streptomycessp LAIDW dan Streptomyces sp CiIA5DW Genus Streptomyces merupakan kelompok bakteri
gram positif yang telah banyak dilaporkan sebagai mikroba pendorong pertumbuhan tanaman Sebuah penelitian melaporkan kemampuan Streptomyces sp sebagai agens biokontrol terhadap F oxysporum f sp ciceri pada tanaman polong (Gopalakrishnan et al 2013) Sedangkan Klebsiella pneumonia dan Burkholderia sp merupakan bakteri penambat nitrogen yang sangat penting bagi pertumbuhan tanaman Dalam beberapa studi kedua bakteri tersebut juga dapat menaikkan imunitas tanaman terhadap pathogen (Liu et al 2011 Suarez-Moreno et al 2008)
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Loekas Soetanto atas pemberian isolat Fusarium oxysporum f sp cubense ras 4 dan proyek Dipa Kompetitif-LIPI atas dana penelitian ini
DAFTAR PUSTAKA
Adegboye MF Babalola OO 2012 Taxonomy and ecology of antibiotic producing actinomycetes Afr J Agric Res 7(15)2255ndash2261 DOI httpdxdoiorg105897AJARX11071
Ara I Bukhari NA Wijayanti DR Bakir MA 2012 Proteolytic activity of alkaliphilic salt-tolerant actinomycetes from various region in Saudi Arabia Afr J Biotechnol 11(16)3894ndash3857
Basha S Ulaganathan K 2002 Antagonism of Bacillus spesies (strain BC121) towards Curvularia lunata Curr Sci 82(12)1457ndash1463
Benyagoub M Benhamou N Carisse O 1998 Cytochemical investigation of the antagonistic interaction beetween a Microsphaeropsis sp (isolate P130A) and Venturia inaequalis Phytopathology 88(7)605ndash613 DOI httpdxdoiorg10 1094PHYTO1998887605
de Azaredo LAI Freire DMG Soares RMA Leite SGF Coelho RRR 2004 Production and partial characterization of thermophilic proteases from Streptomyces
29
J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Agustiyani et al
sp isolated from Brazilian cerrado soil Enzyme Microb Technol 34(3)354ndash358 DOI httpdxdoiorg101016jenzmictec200311015
de Vasconcellos RLF Cardoso EJBN 2009 Rhizospheric streptomycetes as potential biocontrol agents of Fusarium and Armillaria pine rot and as PGPR for Pinus taeda Biocontrol 54(6)807ndash816 DOI httpdxdoiorg101007s10526-009-9226-9
DoumbouCL Hamby Salove MK Crawford DL Beaulieu C 2001 Actinomycetes promising tools to control plant disease and to promote plant growth Phytoprotection 82(3) 85-102 DOI httpdxdoiorg10 7202706219ar
Getha K Vikineswary S 2002 Antagonistic effect of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f sp cubense race 4 Indirect evidence for the role of antibiosis in the antagonistic process J Ind Microbiol Biotechnol 28(6)303ndash310 DOI httpdxdoiorg10 1038sjjim7000247
Gopalakrishnan S Srinivas V Sree Vidya M Rathore A 2013 Plant growth-promoting activities of Streptomyces spp in sorghum and rice Springer Plus 2(1) 574 DOI httpdxdoiorg1011862193-1801-2-574
Huang YH Wang RC Lia CH Zuo CW Wei YR Zhang L Yi GJ 2012 Control of Fusarium wilt in banana with Chinese leek Eur J Plant Pathol 134(1) 87ndash95 DOI httpdxdoiorg101007s10658-012-0024-3
Hwang SC Ko WH 2004 Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in Taiwan Am Phytopathol Soc 88(6)580ndash588
Kloepper JW Ryu CM Zhang S 2004 Induced systemic resistance and promotion of plant growth by Bacillus spp Phytopathology 94(11) 1259ndash1266 DOI httpdxdoiorg101094PHYTO200494111259
Liu Y Wang H Sun X Yang H Wang Y Song W 2011 Study on mechanisms of colonization of nitrogen-fixing PGPB
Klebsiella pneuminiae NG14 on the root surface of rice and the formation of biofilm Curr Microbiol 62(4) 1113ndash1122 DOI httpdxdoiorg101007s00284-010-9835-7
Nourozian J Etebarian HR Khodakaramian G 2006 Biological control of Fusarium graminearum on wheat by antagonistic bacteria Songklanakarin J Sci Technol 28(1)29ndash38
Nel B Steinberg C Labuschange N Viljoen A 2007 Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana Crop Protection 26(4)697ndash705 DOI httpdxdoiorg101016jcropro200606008
Ploetz RC 2006 Fusarium wilt of bananan is caused by several pathogens referred to as Fusarium oxysporum f sp cubense Phytopathology 96(6)653ndash656 DOI httpdxdoiorg101094PHYTO-96-0653
Ramesh S Mathiavanan N 2009 Screening of marine actinomycetes isolated from the Bay of Bengal India for antimicrobial activity and industrial enzyme World J Microbiol Biotechnol 25(12)2103ndash2111 DOI httpdxdoiorg101007s11274-009-0113-4
Rodas-Junco BA Magana-Sevilla HF Tun-Suarez JM Reyes-Ramirez A 2009 Antifungal activity in vitro of native Bacillus sp strain against Macrophomina phaseolina (Tassi) Goid Res J Biol Sci 4(9)985ndash989
Sivamani E Gnanamanickam SS 1988 Biological control of Fusarium oxysporum f sp cubense in banana by inoculation with Pseudomonas fluorescens Plant Soil 107(1)3ndash9 DOI httpdxdoiorg10 1007BF02371537
Suarez-Moreno ZR Caballero-Mellado J Venturi V 2008 The new group of non-pathogenic plant-associated nitrogen fixing Burkholderia spp shared conserved quorum-sensing system which is tightly regulated by the RsaL repressor Microbiol 154(7)2048-2059 DOI httpdxdoiorg101099mic02008017780-0
30
Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

Volume 10 Nomor 1 Februari 2014Halaman 31ndash36
DOI 1014692jfi10131
Trichoderma spp dan Penicillium spp dari Tanah Rizosfer Lahan Rawa Lebak dalam Menginduksi Ketahanan Tanaman Cabai
Terhadap Serangan Penyakit Rebah Kecambah
Induced Resistance to Damping-off Disease in Chili Using Trichoderma spp and Penicillium spp Isolated from Soil Rhizosphere
in Lowland Swampy Area
Ahmad Muslim Syahri Harman Hamidson Abdullah SalimUniversitas Sriwijaya Ogan Ilir 30662
ABSTRAK
Mikroba yang berasosiasi dengan rizosfer sangat penting bagi pertumbuhan tanaman dan dapat melindungi tanaman dari penyakit Penelitian dilakukan untuk menentukan kemampuan isolat cendawan Trichoderma spp dan Penicillium spp yang diisolasi dari rizosfer di lahan rawa lebak dalam mengendalikan penyakit rebah kecambah yang disebabkan oleh Rhizoctonia solani pada tanaman cabai Trichoderma spp dan Penicillium spp diaplikasikan dalam bentuk inokulum dengan bahan pembawa dedak jagung dan merang padi Aplikasi kedua isolat cendawan pada pembibitan tanaman cabai selama 2 minggu mampu menginduksi ketahanan tanaman cabai terhadap infeksi R solani Perlakuan Trichoderma spp dan Penicillium spp dapat menekan insidensi penyakit berturut-turut sebesar 615ndash 100 dan 462ndash100 dan keparahan penyakit berturut-turut sebesar 50ndash100 dan 30ndash959 Hasil tersebut menunjukkan potensi Trichoderma spp dan Penicillium spp sebagai agens biokontrol untuk mengendalikan penyakit rebah kecambah pada tanaman cabai
Kata kunci induksi ketahanan insidensi penyakit keparahan penyakit Rhizoctonia solani
ABSTRACT
Soil microbes associated with rhizosphere are important for promoting plant growth and inducing resistance to diseases The research was conducted to study the ability of Trichoderma spp and Penicillium spp isolated from rhizosphere in lowland swampy area for controlling damping-off disease caused by Rhizoctonia solani Khun Trichoderma spp and Penicillium spp were cultured in bran corn meal and rice straw containing media and applied as inoculum to 2-weeks old seedlings Application of two fungi isolates effectively induced resistance of chili plants to damping-off disease Trichoderma spp and Penicillium spp were significantly reduced disease incidence by 615ndash100 to 462ndash100 respectively and disease severity by 50ndash100 and 30ndash959 respectively This experiment showed the potential of Trichoderma spp and Penicillium spp as biocontrol agents to control damping-off disease on chili
Key words chili disease incidence disease severity induced resistance Rhizoctonia solani
Alamat penulis korespondensi Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas SriwijayaJalan Raya Palembang-Prabumulih Km 32 Indralaya Ogan Ilir 30662 Tel 0711-580663 Faks0711-580059 Surel limpal2003yahoocom
31
ISSN 2339-2479
J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Muslim et al
PENDAHULUAN
Daerah rizosfer merupakan daerah yang sangat penting bagi pertumbuhan tanaman dan juga daerah pertahanan yang paling penting dari serangan patogen Mikroorganisme yang berasosiasi dan hidup di sekitar akar (rizosfer dan rizoplen) sangat penting bagi pertumbuhan tanaman dan juga dalam ekosistem tanah Shivana et al (1996) melaporkan bahwa cendawan steril Phoma Trichoderma Fusarium dan Penicillium merupakan cendawan rizosfer yang efektif dalam menekan berbagai penyakit rebah kecambah dan layu fusarium pada tanaman mentimun di samping itu cendawan tersebut juga dapat memacu pertumbuhan tanaman sehingga digolongkan sebagai cendawan pemacu pertumbuhan tanaman (PPT) Merra et al (1994) membuktikan bahwa cendawan tersebut mempunyai kemampuan mengolonisasi akar mentimun dengan baik dan mampu menginduksi ketahanan tanaman terhadap penyakit antraknosa
Eksplorasi cendawan rizosfer dari daerah rawa lebak yang memiliki potensi sebagai penginduksi ketahanan tanaman belum banyak dilaporkan Penelitian mengenai cendawan PPT yang memiliki kemampuan melindungi tanaman dari penyakit baik sebagai agens biokontrol ataupun penginduksi ketahanan tanaman akan sangat berguna dalam mendukung pengembangan pengendalian ataupun pengelolaan penyakit secara terpadu dan berkelanjutan khususnya di daerah rawa lebak Penelitian ini bertujuan menentukan kemampuan cendawan Trichoderma dan Penicillium dalam menginduksi ketahanan tanaman cabai terhadap penyakit rebah kecambah yang merupakan penyakit penting pada tanaman cabai di lahan rawa lebak
BAHAN DAN METODE
Inokulum Cendawan Rizosfer dan PatogenInokulum cendawan Trichoderma spp
dan Penicillium spp yang digunakan diisolasi dari tanah rizosfer lahan rawa lebak di Kabupaten Ogan Ilir Sumatera Selatan yang
sudah diuji kemampuannya sebagai cendawan PPT (Muslim et al 2006) Isolat patogen R solani diisolasi dari akar tanaman cabai yang terserang penyakit rebah kecambah Masing-masing cendawan Trichoderma spp Penicillium spp dan Rhizoctonia solani disiapkan dengan cara yang sama dalam bentuk inokulum yang menggunakan substrat campuran dedak jagung dan merang padi kering dengan perbandingan masing-masing volume yang sama Masing-masing isolat cendawan ditumbuhkan pada medium agar-agar dekstrosa kentang (ADK) selama 3 hari pada suhu ruangan Sebanyak 5 koloni miselium (oslash 5 mm) dari masing-masing biakan dipotong dari ujung pertumbuhan yang termuda dan diletakkan ke dalam 80 g substrat campuran dedak jagung merang padi basah (dicampur air distilasi dengan perbandingan 108 bv campuran dedak jagung dan merang padi keringair distilasi) yang sudah disterilkan dengan autoklaf di dalam botol erlemneyer (500 mL) Masing-masing biakan diinkubasikan selama 10-14 hari pada suhu ruangan Biakan digoyang setiap hari supaya kolonisasi cendawan pada subsrat merata Substrat yang terkolonisasi oleh masing-masing cendawan dikering anginkan selama 7 hari dan disimpan pada suhu 4 oC sebelum digunakan (Muslim 2003)
Uji Induksi Ketahanan Prinsip induksi ketahanan ialah
perlindungan tanaman secara tidak langsung tanpa kontak langsung antara agens hayati dan patogen Uji induksi ketahanan pada tanaman cabai terhadap penyakit rebah kecambah ini dilakukan mengikuti Cardosa dan Echandi (1987) yang dimodifikasi Benih tanaman cabai ditanam pada wadah yang diisi tanah yang telah diinfestasi dengan Trichoderma spp dan Penicillium spp dengan konsentrasi 2 (bb) Tanah sebagai media tumbuh berasal dari lahan rawa lebak Bibit dibiarkan tumbuh selama 2 minggu kemudian bibit dicabut dengan hati-hatiuntuk menghindari akar tanaman patah atau rusak Trichoderma spp dan Penicillium spp yang mengolonisasi akar tersebut dimatikan dengan
32
J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Muslim et al
merendam akar bibit dalam larutan NaOCl 1 selama 1 menit lalu alkohol 70 selama 1 menit Bibit tersebut ditanam kembali pada tanah yang telah diinfestasi R solani dengan konsentrasi 1 (bb) dari bobot medium tanam Untuk memastikan Trichoderma spp dan Penicillium spp sudah tidak berasosiasi dengan akar tanaman akar yang telah disterilisasi permukaannya dipotong-potong sepanjang 1 cm kemudian potongan akar diletakkan pada medium agar-agar air atau medium ADK dan diamati pertumbuhan agens hayati
Peubah yang diamati pada uji induksi ketahanan ialah insidensi penyakit dan persentase keparahan penyakit Insidensi dan keparahan penyakit dihitung dengan rumus M insidensi penyakit n jumlah bibit mati N jumlah seluruh bibit yang disemai
I keparahan penyakit n jumlah bibit yang terserang Z harga numerik dari nilai kategori tertinggi N jumlah benih yang disemai v harga numerik dari setiap nilai kategori (0-5) 0 tidak ada penyakit 1 lesio muncul pada leher akar sepanjang 1 mm 2 lesio cokelat sampai cokelat gelap sepanjang 2-10 mm mengelilingi akar 3 lesio cokelat gelap sepanjang 10-25 mm ketika miselium mengolonisasi koleoptil 4 gt25 mm area akar menjadi hitam dan busuk pada koleoptil 5 bibit busuk secara menyeluruh atau bibit mati
Persentase penghambatan terhadap insidensi dan keparahan penyakit dihitung berdasarkan rumus sebagai berikut
K nilai insidensi penyakit atau keparahan penyakit pada perlakuan kontrol P nilai insidensi penyakit atau keparahan penyakit pada perlakuan cendawan agens biokontrol
Rancangan Percobaan Penelitian disusun dalam rancangan acak
lengkap dengan 13 perlakuan yang terdiri
atas kontrol 7 perlakuan isolat Trichoderma spp (galur T1 T2 T5 T6 T9 T10 dan T11) dan 5 perlakuan isolat Penicillium spp (galur P1 P2 P3 P8 dan P11) Masing-masing perlakuan diulang 4 kali dengan 5 tanaman untuk masing-masing ulangan
Data yang diperoleh dianalisis menggunakan analysis of variance dan uji lanjut dengan uji Beda Nyata Jujur menggunakan SAS 90
HASIL
Semua isolat Trichoderma spp dan Penicillium spp yang diuji mempunyai kemampuan sebagai penginduksi ketahanan terhadap penyakit rebah kecambah pada tanaman cabai Kemampuan isolat-isolat tersebut menginduksi ketahanan ditunjukkan dengan nilai insidensi dan keparahan penyakit pada masing-masing perlakuan (Tabel 1 dan 2) Insidensi penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash25 dan 0ndash35 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 65 Penghambatan insidensi penyakit oleh isolat-isolat Trichoderma spp mencapai gt846 bahkan isolat-isolat T5 T6 T9 T10 dan T11 mampu menghambat penyakit sebesar 100 Perlakuan Penicillium spp mampu menekan insidensi penyakit sebesar 462ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan P11
Demikian pula dengan pengamatan keparahan penyakit menunjukkan bahwa perlakuan Trichoderma spp dan Penicillium spp sangat efektif menekan penyakit Keparahan penyakit pada perlakuan Trichoderma spp dan Penicillium spp berturut-turut berkisar 0ndash37 dan 3ndash515 jauh lebih rendah dibandingkan perlakuan kontrol yaitu 74 Penghambatan keparahan penyakit pada perlakuan Trichoderma spp Berkisar 50ndash100 dengan penghambatan tertinggi (100) terjadi pada perlakuan T11 Penghambatan keparahan penyakit pada perlakuan Penicillium spp berkisar 304ndash959 dengan penghambatan tertinggi (9511) terjadi pada perlakuan P11
M = n
N x 100 dengan
00100
)(times
times
times= sum
NZvn
I dengan
Persentase penghambatan= K - PK
X 100 dengan
33
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-
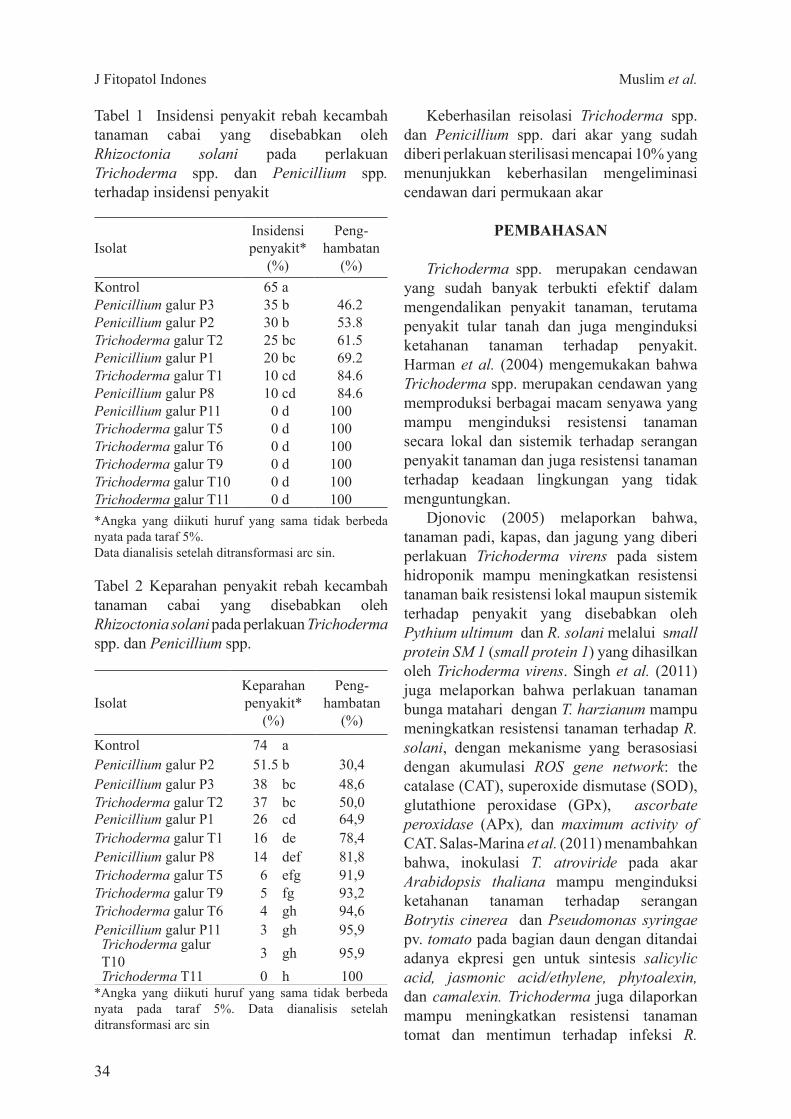
J Fitopatol Indones Muslim et al
Keberhasilan reisolasi Trichoderma spp dan Penicillium spp dari akar yang sudah diberi perlakuan sterilisasi mencapai 10 yang menunjukkan keberhasilan mengeliminasi cendawan dari permukaan akar
PEMBAHASAN
Trichoderma spp merupakan cendawan yang sudah banyak terbukti efektif dalam mengendalikan penyakit tanaman terutama penyakit tular tanah dan juga menginduksi ketahanan tanaman terhadap penyakit Harman et al (2004) mengemukakan bahwa Trichoderma spp merupakan cendawan yang memproduksi berbagai macam senyawa yang mampu menginduksi resistensi tanaman secara lokal dan sistemik terhadap serangan penyakit tanaman dan juga resistensi tanaman terhadap keadaan lingkungan yang tidak menguntungkan
Djonovic (2005) melaporkan bahwa tanaman padi kapas dan jagung yang diberi perlakuan Trichoderma virens pada sistem hidroponik mampu meningkatkan resistensi tanaman baik resistensi lokal maupun sistemik terhadap penyakit yang disebabkan oleh Pythium ultimum dan R solani melalui small protein SM 1 (small protein 1) yang dihasilkan oleh Trichoderma virens Singh et al (2011) juga melaporkan bahwa perlakuan tanaman bunga matahari dengan T harzianum mampu meningkatkan resistensi tanaman terhadap R solani dengan mekanisme yang berasosiasi dengan akumulasi ROS gene network the catalase (CAT) superoxide dismutase (SOD) glutathione peroxidase (GPx) ascorbate peroxidase (APx) dan maximum activity of CAT Salas-Marina et al (2011) menambahkan bahwa inokulasi T atroviride pada akar Arabidopsis thaliana mampu menginduksi ketahanan tanaman terhadap serangan Botrytis cinerea dan Pseudomonas syringae pv tomato pada bagian daun dengan ditandai adanya ekpresi gen untuk sintesis salicylic acid jasmonic acidethylene phytoalexin dan camalexin Trichoderma juga dilaporkan mampu meningkatkan resistensi tanaman tomat dan mentimun terhadap infeksi R
IsolatInsidensi penyakit
()
Peng-hambatan
()Kontrol 65 aPenicillium galur P3 35 b 462Penicillium galur P2 30 b 538Trichoderma galur T2 25 bc 615Penicillium galur P1 20 bc 692Trichoderma galur T1 10 cd 846Penicillium galur P8 10 cd 846Penicillium galur P11 0 d 100Trichoderma galur T5 0 d 100Trichoderma galur T6 0 d 100Trichoderma galur T9 0 d 100Trichoderma galur T10 0 d 100Trichoderma galur T11 0 d 100
Tabel 1 Insidensi penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp terhadap insidensi penyakit
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
34
Tabel 2 Keparahan penyakit rebah kecambah tanaman cabai yang disebabkan oleh Rhizoctonia solani pada perlakuan Trichoderma spp dan Penicillium spp
Angka yang diikuti huruf yang sama tidak berbeda nyata pada taraf 5 Data dianalisis setelah ditransformasi arc sin
Isolat Keparahan penyakit
()
Peng-hambatan
()
Kontrol 74 aPenicillium galur P2 515 b 304Penicillium galur P3 38 bc 486Trichoderma galur T2 37 bc 500Penicillium galur P1 26 cd 649Trichoderma galur T1 16 de 784Penicillium galur P8 14 def 818Trichoderma galur T5 6 efg 919Trichoderma galur T9 5 fg 932Trichoderma galur T6 4 gh 946Penicillium galur P11 3 gh 959Trichoderma galur T10 3 gh 959Trichoderma T11 0 h 100
J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Muslim et al
solani melalui peningkatan hydrogen peroxide (H2O2) lipid peroxidation dan aktivitas enzim phenylalanine ammonia lyase (PAL) guaiacol (POG) dan ascorbate peroxidases (APX) (Nawrocka et al 2011)
Koike et al (2001) melaporkan bahwa isolat cendawan PPT seperti Trichoderma Fusarium Penicillium Phoma dan cendawan steril yang diaplikasikan pada akar tanaman mentimun dalam bentuk inokulum biji gandum miselium dan filtrat biakan ternyata efektif dalam menginduksi ketahanan secara sistemik pada tanaman terhadap penyakit angural leaf spot yang disebabkan oleh bakteri Pseudomonas syringae pv lachrymans dan juga layu fusarium oleh cendawan Fusarium oxysporum fsp cucumerinum Perlakuan dengan cendawan PPT ini meningkatkan sistem pertahanan tanaman melalui lignifikasi superoxide generation dan aktivitas chemiluminescence
De Cal et al (2000) melaporkan bahwa penurunan penyakit layu fusarium pada tanaman tomat berhubungan dengan induksi ketahanan melalui aktivitas kambium dan pembentukan tambahan xilem sekunder pada tanaman tomat yang diberi perlakuan P oxalicum Hossain et al (2008) melaporkan bahwa tanaman Arabidopsis thaliana yang diberi perlakuan biakan sel dan filtrat Penicillium isolat GP 16-2 mampu menginduksi ketahanan tanaman secara sistemik melalui peningkatan asam jasmonat asam salisianat etilena dan non-expressor of PR genes 1 (NPR1)
Perlakuan Trichoderma spp dan Penicillium spp pada bibit tanaman cabai sangat efektif dalam menekan insidensi dan keparahan penyakit rebah kecambah Penelitian perlu dilanjutkan untuk mempelajari mekasnisme induksi ketahanan yaitu secara biokimia histologi atau genetik
UCAPAN TERIMA KASIH
Ucapan terima kasih penulis sampaikan kepada Direktorat Pembinaan Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional yang telah membiayai penelitian ini dengan nomor kontrak 026SP2HPPDP2MIII2007
DAFTAR PUSTAKA
De Cal A Garcia-Lepe R Melgarejo P 2000 Induced resistance by Penicillium oxalicum against Fusarium oxysporum f sp lycopersici histological studies of infected and induced tomato stems Phytopathology 90260-268 DOI httpdxdoiorg10 1094PHYTO2000903260
Djonovic S 2005 Role of two secreted proteins from trichoderma virens in mycoparasitism and induction of plant resistance [disertasi] Texas (US) UMI
Harman GE Howell ChR Viterbo A Chet I Lorito M 2004 Trichoderma Species Opportunistic Avirulent Plant Symbionts Nat Rev 243-56
Hossain MdM Sultana F Kubota M Hyakumachi M 2008 Differential inducible defense mechanisms against bacterial speck pathogen in Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp GP16-2 and its cell free filtrate Plant Soil 304227-239 DOI httpdxdoiorg101007s11104-008-9542-3
Koike N Hyakumachi M Kageyama K Tsuyumu S Doke N 2001 Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi lignification and superoxide generation Eur J Plant Pathol 107523-533 DOI httpdxdoiorg101023A1011203826805
Merra MS Shivanna MB Kageyama K Hyakumachi M 1994 Plant growth-promoting fungi from zoyusia grass rhizosophere as potential inducer of systemic resistance in cucumbers Phytopathology 841399-1406 DOI httpdx doiorg101094Phyto-84-1399
Muslim A Horinouchi H Hyakumachi M 2003 Control of fusarium crown and root rot of tomato with hypovirulent binucleate Rhizioctonia in soil and rock wool systems
35
J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-

J Fitopatol Indones Muslim et al
Plant Dis 87739-747 DOI httpdxdoiorg101094PDIS2003876739
Nawrocka J Snochowska M Gajewska E Pietrowska E Szczech M 2011 Activation of defense responses in cucumber and tomato plants by selected polish Trichoderma strains Veget Crops Res Bull 75105-116 DOI httpdxdoiorg102478v10032-011-0022-6
Salas-marina MA Silva-Flores MA Uresti-Rivera EE Castro-Longoria E Herrera-Estrella A 2011 Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acidethylene and salicylic acid pathways Eur J Plant Pathol 13115-26
DOI httpdxdoiorg101007s10658-011-9782-6
Shivanna MB Merra MS and Hyakumachi M 1996 Role of root colonization ability of plant growth promoting fungi in the suppression of take-all and common root rot of wheat Crop Proec 15497-504 DOI httpdxdoiorg1010160261-2194 (96)00004-X
Singh BN Singh A Singh SP Singh HB 2011 Trichoderma harzianum - mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani Eur J Plant Pathol 131121ndash134 DOI httpdxdoiorg101007s10658-011-9792-4
36
- 8516-23984-1-PBpdf (p1-7)
- 8517-23985-1-PBpdf (p8-16)
- 8518-23986-1-PBpdf (p17-22)
- 8519-23987-1-PBpdf (p23-30)
- 8520-23988-1-PBpdf (p31-36)
-





![[Kelapa Sawit] Profil Komoditi Kelapa Sawit](https://static.fdokumen.com/doc/165x107/5571fa8a4979599169927503/kelapa-sawit-profil-komoditi-kelapa-sawit.jpg)














