
BAB II KAJIAN PUSTAKA - sinta.unud.ac.id filefokal simtomatik 12% dan fokal kriptogenik 37% dari 613...
59
BAB II KAJIAN PUSTAKA 2.1 Epilepsi Kejang merupakan manifestasi klinis akibat adanya lompatan muatan listrik berlebihan pada neuron-neuron di otak yang terganggu fungsinya. Epilepsi adalah kejang berulang tanpa pencetus (provokasi) ≥2 dengan interval >24 jam antara kejang pertama dan berikutnya. Manifestasi klinis epilepsi dapat berupa gangguan kesadaran, motorik, sensoris, autonom, atau psikis (Swaiman dan Ashwal, 2012). Insiden epilepsi tertinggi pada golongan usia dini, menurun pada usia dewasa muda, dan meningkat pada usia lanjut (Hauser dan Nelson, 2013). Sebanyak 25% dari seluruh kasus epilepsi terjadi pada anak umur kurang lima tahun (Yilmas dkk., 2013). Sebuah penelitian melaporkan bahwa insiden epilepsi pada umur 0-14 tahun sebesar 82,2 kasus/100.000 populasi/tahun (Khatria dkk., 2003). Klasifikasi epilepsi dibuat berdasarkan tipe kejang, penyebab, dan sindroma epilepsi. International leage againts epilepsy (ILAE) pada tahun 2010 menetapkan klasifikasi epilepsi berdasarkan tipe kejang. Tipe kejang dibuat berdasarkan semiologi kejang dan gambaran EEG, dibedakan menjadi epilepsi fokal (parsial) dan epilepsi umum. Epilepsi fokal adalah kejang dimulai dari fokus tertentu yang terlokalisir di otak dan kejang muncul pada satu sisi tubuh saja. Epilepsi fokal dapat sekunder menjadi umum jika terjadi propagasi listrik otak ke hemispher kontralateral. Epilepsi umum adalah kejang pada daerah lebih luas di
Transcript of BAB II KAJIAN PUSTAKA - sinta.unud.ac.id filefokal simtomatik 12% dan fokal kriptogenik 37% dari 613...

BAB II
KAJIAN PUSTAKA
2.1 Epilepsi
Kejang merupakan manifestasi klinis akibat adanya lompatan muatan
listrik berlebihan pada neuron-neuron di otak yang terganggu fungsinya. Epilepsi
adalah kejang berulang tanpa pencetus (provokasi) ≥2 dengan interval >24 jam
antara kejang pertama dan berikutnya. Manifestasi klinis epilepsi dapat berupa
gangguan kesadaran, motorik, sensoris, autonom, atau psikis (Swaiman dan
Ashwal, 2012). Insiden epilepsi tertinggi pada golongan usia dini, menurun pada
usia dewasa muda, dan meningkat pada usia lanjut (Hauser dan Nelson, 2013).
Sebanyak 25% dari seluruh kasus epilepsi terjadi pada anak umur kurang lima
tahun (Yilmas dkk., 2013). Sebuah penelitian melaporkan bahwa insiden epilepsi
pada umur 0-14 tahun sebesar 82,2 kasus/100.000 populasi/tahun (Khatria dkk.,
2003).
Klasifikasi epilepsi dibuat berdasarkan tipe kejang, penyebab, dan
sindroma epilepsi. International leage againts epilepsy (ILAE) pada tahun 2010
menetapkan klasifikasi epilepsi berdasarkan tipe kejang. Tipe kejang dibuat
berdasarkan semiologi kejang dan gambaran EEG, dibedakan menjadi epilepsi
fokal (parsial) dan epilepsi umum. Epilepsi fokal adalah kejang dimulai dari fokus
tertentu yang terlokalisir di otak dan kejang muncul pada satu sisi tubuh saja.
Epilepsi fokal dapat sekunder menjadi umum jika terjadi propagasi listrik otak ke
hemispher kontralateral. Epilepsi umum adalah kejang pada daerah lebih luas di

kedua hemispher otak dan manifestasi kejang pada kedua sisi tubuh (Berg dkk.,
2012). Pada anak lebih dari 60% epilepsi tergolong epilepsi fokal (Rauchenzauner
dan Luef, 2010). Penelitian terdahulu mendapatkan epilepsi fokal idiopatik 10%,
fokal simtomatik 12% dan fokal kriptogenik 37% dari 613 kasus epilepsi umur
kurang 16 tahun (Berg dkk., 2012).
Klasifikasi epilepsi berdasarkan penyebabnya, digolongkan menjadi tiga
jenis yaitu idiopatik, simtomatik, dan kriptogenik. Epilepsi idiopatik adalah
epilepsi yang tidak jelas ditemukan penyebabnya dan sering dihubungkan dengan
faktor genetik. Epilepsi simtomatik jika penyakit yang mendasari jelas ditemukan,
sedangkan epilepsi kriptogenik, diduga ada penyebab yang mendasari tetapi
belum bisa dibuktikan (Kwan dkk., 2011). Sekitar 60% epilepsi tergolong epilepsi
didapat, biasanya disebabkan cedera otak seperti infeksi, malformasi otak,
masalah perinatal, masalah metabolik, kejang demam lama, status epileptikus,
trauma kepala dan stroke. Penyebab epilepsi pada bayi dan anak di Finlandia
adalah idiopatik (64%), masalah prenatal (15%), perinatal (9%), dan post natal
(12%) (Sillanpaa dan Schmidt, 2011).
International leage againts epilepsy (ILAE) juga membuat klasifikasi
sindroma epilepsi pada tahun 1989. Sindroma epilepsi adalah epilepsi unik
dengan gejala dan tanda yang kompleks dengan penyebab berbeda-beda (Berg
dkk., 2012). Sindroma epilepsi umum yang sering pada bayi dan anak adalah
sindrom Ohtahara, sindroma West, sindroma Lenox-Gestaut, epilepsi mioklonik
juvenile, sedangkan sindroma epilepsi fokal antara lain epilepsi Rolandic, epilepsi
lobus temporal, dan epilepsi oksipital benigna (Stafstorm dkk., 2011).

2.2 Patofisiologi Epilepsi atau Epileptogenesis
Penyebab yang berperan pada onset dan progresivitas epilepsi ada tiga
faktor utama yaitu: 1) faktor genetik; 2) gangguan perkembangan dan atau
malformasi susunan saraf pusat; 3) cedera otak atau interaksi dari ketiga faktor
tersebut (Jefferys, 2010). Perubahan seluler dan molekuler yang terjadi akibat
cedera otak dan menyebabkan kejang berulang tanpa provokasi dinamakan
epileptogenesis. Kematian neuron di otak dipercaya sebagai faktor propagasi yang
menyebabkan epilepsi dan banyak bukti menunjukkan bahwa kematian neuron di
otak menyebabkan kejang dan sebaliknya kejang menimbulkan kematian neuron.
Kematian neuron terlibat dalam eksitoksisitas dimana kelebihan glutamat
menyebabkan stimulasi berlebihan terhadap reseptor N-methil-D-aspartat
(NMDA), dan menyebabkan akumulasi kalsium di dalam sel dan diikuti dengan
cedera dan kematian neuron. Proses seluler lain yang terlibat pada epileptogenesis
adalah neurogenesis, gliosis, axonal sprouting, cedera axon, remodeling dendrit,
angiogenesis, stres oksidatif, inflamasi, perubahan matriks ekraseluler, dan
perubahan saluran ion (Pitkanen dan Lukasiuk, 2011;Reddy, 2013).
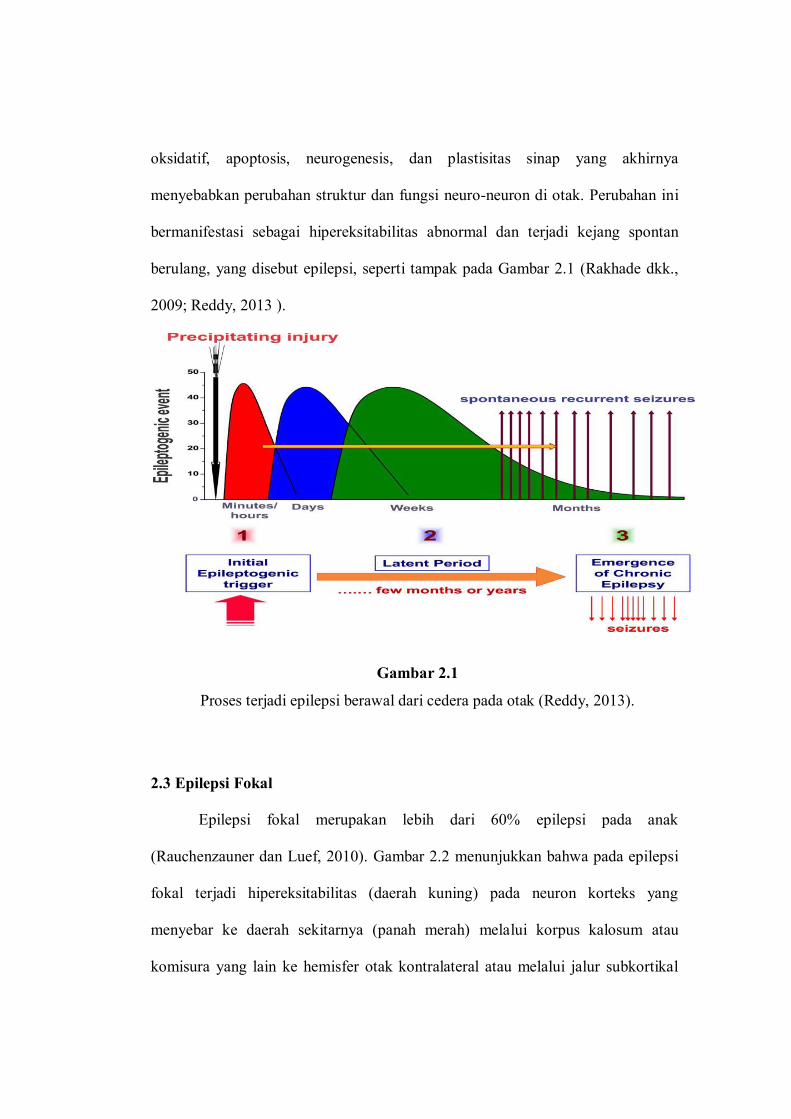
Proses epileptogenesis dapat dibagi menjadi tiga fase yakni: 1) cedera otak
awal; 2) fase laten, tanpa aktivitas kejang berlangsung beberapa bulan sampai
beberapa tahun; 3) fase kronis, dengan kejang spontan berulang. Walaupun
mekanisme yang mendasari terjadinya epilepsi belum sepenuhnya dipahami,
tetapi dipercaya bahwa terjadi interaksi akut dan kronis secara anatomis,
molekuler, dan fisiologis yang kompleks dan bersifat multifaktor. Faktor-faktor
pencetus awal mengaktivasi berbagai jalur sinyal, seperti inflamasi, stres
oksidatif, apoptosis, neurogenesis, dan plastisitas sinap yang akhirnya
menyebabkan perubahan struktur dan fungsi neuro-neuron di otak. Perubahan ini
bermanifestasi sebagai hipereksitabilitas abnormal dan terjadi kejang spontan
berulang, yang disebut epilepsi, seperti tampak pada Gambar 2.1 (Rakhade dkk.,
2009; Reddy, 2013 ).
Gambar 2.1
Proses terjadi epilepsi berawal dari cedera pada otak (Reddy, 2013).
2.3 Epilepsi Fokal
Epilepsi fokal merupakan lebih dari 60% epilepsi pada anak
(Rauchenzauner dan Luef, 2010). Gambar 2.2 menunjukkan bahwa pada epilepsi
fokal terjadi hipereksitabilitas (daerah kuning) pada neuron korteks yang
menyebar ke daerah sekitarnya (panah merah) melalui korpus kalosum atau
komisura yang lain ke hemisfer otak kontralateral atau melalui jalur subkortikal

(thalamus, batang otak) pada kejang fokal yang berkembang menjadi kejang
umum (Gambar 2.2.A). Pada epilepsi umum, kejang mulai pada satu area tertentu
di otak, kemudian secara serentak menyebar ke hemisfer otak kiri dan kanan
(Gambar 2.2.B)(Stafstorm, 1998).
Manifestasi klinis kejang fokal pada anak dapat muncul dalam bentuk
aura, automatisme, versive, kepala menengok (nonversive head turning), deviasi
mulut, disfasia pascakejang, iktal speech, dan automatisme unilateral. Manifestasi
klinis epilepsi fokal tergantung dari lobus mana fokus epileptik berasal, setiap
lobus memberikan manifestasi yang khas (Berg dkk., 2012). Kejang yang berasal
dari lobus frontalis merupakan 30% dari seluruh pasien epilepsi fokal, dan
merupakan fokus tersering ke dua setelah lobus temporalis. Kejang fokal tanpa
penurunan kesadaran disebut epilepsi fokal sederhana, jika disertai penurunan
kesadaran disebut epilepsi fokal kompleks (Swaiman dan Ashwal, 2012).
Kejang fokal kompleks hanya sedikit yang berasal dari lobus parietalis.
Manifestasi kejang fokal dengan penurunan kesadaran terjadi bila melibatkan
lobus temporalis, manifestasi klinis dapat berupa perubahan perilaku, halusinasi
visual, panik, atau bengong. Kejang yang berasal dari lobus oksipitalis berupa
kejang fokal kompleks: a) aura visual, berupa cahaya warna putih atau berwarna;
b) buta saat serangan (ictal blindness); c) gerakan kepala dan mata ke arah
kontralateral atau; d) kedip-kedip mata yang cepat. Gejala lain dapat terjadi bila
melibatkan lobus temporalis atau parietalis (Niedermeyer, 2005; Schaefer-
Foldvary, 2005).

Gambar 2.2
A. Kejang fokal dan B. Kejang umum (Stafstorm, 1998).
2.4 Diagnosis Epilepsi
Epilepsi adalah diagnosis klinis, ditegakkan berdasarkan anamnesis dan
pemeriksaan fisik. Pemeriksaan penunjang EEG untuk konfirmasi diagnosis,
melihat sindroma epilepsi tertentu dan pencitraan kepala yaitu computed
tomography scan (CT scan) atau magnetic resonance imaging (MRI) untuk
menemukan penyebabnya. Pemeriksaan MRI kepala dapat memberikan informasi
lebih baik dibandingkan CT scan kepala (Kuzniecky, 2005).
2.4.1 Anamnesis
Anamnesis harus dilakukan secara cermat, rinci, dan menyeluruh karena
pemeriksa atau dokter jarang menyaksikan langsung serangan kejang yang
dialami penderita. Penjelasan segala sesuatu yang terjadi sebelum, selama, dan

sesudah kejang merupakan informasi yang sangat berarti dan menjadi kunci
diagnosis epilepsi (Sillanpaa dan Schmidt, 2011).
Anamnesis penderita epilepsi harus meliputi: tipe kejang, lama kejang,
gejala sebelum, selama dan setelah kejang, frekuensi kejang, dan adanya penyakit
penyerta. Umur saat kejang pertama sangat penting dicatat, riwayat kehamilan,
persalinan dan tumbuh-kembang, riwayat penyakit, penyebab, pengobatan
sebelumnya, respon awal pengobatan, dan riwayat penyakit epilepsi dalam
keluarga (Maria, 2009; Swaiman dan Aswal, 2012). Pemeriksa harus dapat
memastikan apakah kejadian itu memang kejang atau bukan kejang. Rekaman
video kejadian kejang di rumah, bisa sangat membantu dokter. Jika belum jelas,
sebaiknya ditunggu sampai bisa dipastikan bahwa kejadian itu memang kejang
atau bukan kejang. Kejang harus dipastikan berulang ≥ 2 kali dengan interval >24
jam. Kejang yang berulang serial dalam rentang waktu 24 jam dianggap kejang
episode tunggal dan diagnosis epilepsi belum bisa ditegakkan (Berg dkk., 2012).
2.4.2 Pemeriksaan fisik umum dan neurologis
Tanda-tanda gangguan yang berhubungan dengan epilepsi harus
diidentifikasi seperti trauma kepala, kelainan kongenital dan gangguan
neurologik. Pemeriksa harus memastikan bahwa kejang tidak ada pencetus yang
jelas, seperti demam, gangguan elektrolit dan gangguan metabolik lainnya.
Adanya keterlambatan perkembangan, organomegali, asimetri ukuran anggota
tubuh dapat menunjukkan awal gangguan pertumbuhan otak. Gambaran dismorfik
pada muka, tanda-tanda tertentu pada bagian tubuh seperti hemangioma, nodul,

makula warna pucat, dan sebagainya untuk melihat kemungkinan sindroma
epilepsi tertentu (Hauser dan Nelson, 2013).
2.4.3 Pemeriksaan penunjang
2.4.3.1 Elektroensefalografi (EEG)
Pemeriksaan EEG sebaiknya dilakukan pada semua penderita epilepsi jika
fasilitas tersedia. Rekaman elektroensefalogram (EEGram) memperlihatkan
aktivitas listrik otak yang direkam menggunakan elektroda yang diletakkan pada
tulang kepala dengan tatacara tertentu. Elektroda kemudian merefleksikan
gabungan eksitasi dan inhibisi potensial post-sinaptik pada ujung apikal dendrit
sel piramidal. Elektroensefalografi berperan dalam konfirmasi diagnosis epilepsi,
menentukan tipe kejang dan sindrom epilepsi, pemilihan OAE, serta menentukan
prognosis. Rekaman EEG pertama menunjukkan abnormalitas sekitar 37-40%
kasus (So, 2011).
Metodelogi rekaman EEG dilakukan sesuai standarisasi American Clinical
Neurophysiology Society (ACNS). Prosedur aktivasi standar adalah stimulasi fotik,
hiperventilasi, rekaman mengantuk sampai tidur stadium non-REM, serta sleep
deprivasi. Penderita sebaiknya tidur alami pada saat direkam, premedikasi
dengan kloralhidrat dosis minimal diberikan kalau sangat diperlukan, karena dapat
menekan aktivitas gelombang epileptik (Sillanpaa dan Shinnar, 2010).
Rekaman EEG hanya bisa merekam 20 -70% gelombang paku yang ada
pada otak penderita. Pada orang normal sekitar 10% mempunyai abnormalitas
tidak khas pada rekaman EEG, sedangkan kurang lebih 1% pasien mempunyai

kelainan paroksismal EEG berupa epileptiform tanpa adanya kejang secara klinis,
pada anak-anak mencapai 2-4%. Penilaian abnormalitas EEG ditentukan
berdasarkan gelombang irama dasar dan adanya gelombang epileptik (Chabolla
dan Cascino, 2005; Wirrell dkk., 2012).
2.4.3.1.1 Gelombang irama dasar
Gelombang irama dasar menunjukkan perbedaan sesuai usia anak. Adanya
perkembangan otak ditunjukkan dengan perubahan gelombang irama dasar mulai
dari 3-4 siklus/detik pada usia 4 bulan, 5 siklus/detik usia 6 bulan, 6-7 siklus/detik
usia 9-18 bulan, 7-8 siklus/detik usia 2 tahun, 9 siklus/detik usia 7 tahun, dan 10-
11 siklus/detik pada usia 10-15 tahun (Niedermeyer, 2005; Chabolla dan Cascino,
2005).
2.4.3.1.2 Gelombang epileptik pada epilepsi fokal
Kerentanan beberapa area di otak berbeda terhadap epilepsi. Lobus
temporalis mempunyai nilai ambang kejang sangat rendah diikuti daerah motorik
Rolandik dan lobus frontalis sehingga lebih mudah mengalami epilepsi. Lobus
oksipitalis dan parietalis merupakan daerah terendah derajat epileptogenitasnya
sehingga kejadian epilepsinya lebih jarang (Chabolla dan Cascino, 2005).
Gelombang epileptik fokal dapat terjadi pada setiap daerah otak
tergantung pada usia penderita. Penderita epilepsi lobus temporalis sebagian besar
(90%) memperlihatkan gelombang paku pada saat tidur. Sebagian besar pasien
dengan gelombang paku bilateral bisinkron pada lobus temporalis memberikan

gejala epilepsi parsial lobus temporalis. Pasien dengan epilepsi parsial kompleks
yang berasal dari lobus temporalis anterior, memberikan gambaran fokal,
lateralisasi atau umum pada gelombang iktal EEG(Niedermeyer, 2005; Chabolla
dan Cascino, 2005).
Area otak kedua yang sensitif terhadap epilepsi fokal adalah lobus
frontalis. Pada area ini sangat sulit menemukan daerah epileptik dengan rekaman
EEG interiktal. Rekaman EEG interiktal tidak spesifik dan tidak sensitif pada
epilepsi lobus frontalis, tidak seperti pada epilepsi lobus temporalis. Daerah
epileptogenik pada lobus frontalis tidak berhubungan dengan aktivitas interiktal
pada rekaman EEG yang diulang atau diperpanjang. Penambahan elektroda
supraorbita dapat meningkatkan sensitifitas dan spesifisitas rekaman EEG pada
pasien epilepsi lobus frontalis yang berasal dari daerah orbitofrontal
(Niedermeyer, 2005).
Gelombang paku oksipital lebih jarang ditemukan dibandingkan lobus
temporalis dan frontalis. Penderita yang mempunyai gelombang paku oksipital
40-50% menunjukkan gejala kejang klinis (Ghofrani dan Akhondian, 2010).
2.4.3.2 Pencitraan (Neuroimaging)
Pemeriksaan MRI kepala merupakan pencitraan pilihan terbaik pada
epilepsi. MRI kepala dengan atau tanpa kontras dapat menemukan etiologi
epilepsi seperti neoplasma otak, ensefalitis autoimun, leukomalasia serebral, dan
sebagainya. Pada keadaan fasilitas MRI tidak tersedia, pemeriksaan CT scan

kepala tanpa atau dengan kontras dapat dilakukan, meskipun memberikan hasil
tidak sebaik MRI kepala (Kuzniecky, 2005).
2.5 Epilepsi Resisten Obat
Resistensi obat merupakan tantangan yang besar pada penatalaksanaan
epilepsi. Istilah epilepsi resisten obat (ERO) mempunyai pengertian serupa
dengan epilepsi refrakter, epilepsi intractabl, dan epilepsi pharmaco-resistenace
(Patil dkk., 2009). Definisi ERO sampai saat ini masih diperdebatkan
(Alexopoulos, 2013 ), ada definisi sangat ketat dan ada lebih longgar, para ahli
menggunakan sesuai dengan kepentingan klinis masing-masing (Kwan dkk.,
2011; Ramos-Lizana dkk., 2012).
Pada konsep ERO terdapat tiga komponen yang perlu diperhatikan yaitu
jumlah OAE yang dikonsumsi sebelumnya, frekuensi kejang, dan durasi kejang
yang tidak terkontrol (Beleza,2009; So, 2011). Definisi ERO, merupakan suatu
konsep relatif (Kwan dkk.,2011). Sifat relatif konsep ERO memerlukan beberapa
persyaratan yang perlu dinilai agar definisi ini lebih mudah dipahami sebelum
mempertimbangkan terapi non-farmakologik misalnya pembedahan, stimulasi
saraf vagus, diet ketogenik, atau opsi lain (Caraballo dkk., 2011; Hao dan Kwan,
2011).
2.5.1 Mekanisme resistensi obat antiepilepsi (OAE)
Patomekanisme mengenai farmako-resisten pada epilepsi ada beberapa
hipotesis yang diajukan oleh para ahli antara lain 1) hipotesis target, 2) hipotesis

transporter obat, 3) hipotesis genetik atau mutasi, 4) hipotesis mekanisme yang
berhubungan dengan penyakit(disease-related mechanisms), 5) hipotesis
mekanisme yang berhubungan dengan obat (drug-related mechanisms)( Schmidt
dan Löscher, 2005; Viteva, 2014). Dua hipotesis utama yang banyak dibahas saat
ini adalah “hipotesis target” dan “hipotesis transporter obat”. Hipotesis target
menyatakan bahwa kegagalan OAE untuk berefek disebabkan oleh adanya
perubahan biomelokuler pada target. Hipotesis transporter obat menyatakan
bahwa kegagalan OAE untuk mencapai targetnya dalam konsentrasi yang cukup
disebabkan oleh transporter obat yang terlalu aktif bekerja pada sawar darah otak
(SDO).
“Hipotesis target” menyatakan bahwa sasaran utama OAE adalah gerbang
listrik saluran ion dan reseptor neurotransmiter. Beberapa OAE terutama bekerja
pada saluran natrium dan OAE lain bekerja pada saluran kalsium, meskipun OAE
juga bekerja pada saluran ion yang lain. Beberapa OAE juga bekerja pada efek
modulasi neurotransmiter inhibisi GABA atau menekan neurotransmiter eksitasi
glutamat (Beleza, 2009). Hipotesis target menunjukkan bahwa perubahan
biomolekuler pada target OAE, apakah bersifat intrinsik seperti polymorphisme,
atau didapat oleh karena proses penyakit itu sendiri, akibat kejang dan obat-
obatan, menyebabkan penurunan sensitivitas target terhadap OAE. Pada epilepsi,
kejang yang berulang menyebabkan terjadinya stres oksidatif dan inflamasi
kronis. Reactive oxygen space (ROS) dan berbagai sitokin proinflamasi terutama
IL-β1, IL-6 dan TNF-α menimbulkan kerusakan dan perubahan biomolukuler
pada reseptor-reseptor tersebut melalui beberapa mekanisme seperti menurunkan

reuptake dan meningkatkan pelepasan glutamat, perubahan fungsi klasik
neurotransmiter melalui remodeling N-methyl-D-aspartate (NMDA) reseptor,
meningkatkan permeabilitas vaskuler dan angiogenesis, meningkatkan kematian
sel, neurogenesis, dan menurunkan nilai ambang kejang, sehingga terjadi
penurunan sensitivitasnya terhadap OAE (Schmidt dan Löscher, 2005; Viteva,
2014), seperti tampak pada Gambar 2.3.
Gambar 2.3
Skema hipotesis target dan hipotesis transporter obat (Schmidt dan Löscher,
2005).
“Hipotesis transporter obat” menyatakan bahwa resistensi OAE
disebabkan oleh aktivitas berlebih transporter yang menghambat OAE masuk ke
dalam otak dalam konsentrasi yang cukup. Transporter obat yang ada pada SDO

adalah kelompok protein ATP-binding cassette (ABC). Transporter ini pada
keadaan normal mencegah masuknya berbagai racun lipophilik atau xenobiotik
dari darah ke otak dengan mekanisme tertentu. Pada saat penderita epilepsi
memerlukan obat untuk masuk ke dalam otak, transporter ini juga bekerja aktif sehingga
menghambat OAE mencapai otak. Transporter obat utama adalah: P-glycoprotein
(P-gp) dan multidrug resistance related proteins (Mrps) (Viteva, 2014).
Transporter ini diregulasi pada suatu kompleks dan proses berbeda, yang mana
pada penderita epilepsi sangat dipengaruhi oleh stres oksidatif dan inflamasi.
Stress oksidatif, inflamasi, dan kejang merupakan mekanisme yang saling
mempengaruhi (Vezzani dkk., 2013). ROS dan berbagai sitokin proinflamasi
terutama IL-β1, IL-6 dan TNF-α menyebabkan overekpresi gen yang mengkode
beberapa transporter obat tersebut melalui aksis glutamat, N-methyl-D-Aspartate
(NMDA) reseptor, dan sinyal cycloxygenase-2 (COX-2). Mekanisme ini
menyebabkan aktivitas transpoter meningkat dan menghambat OAE masuk ke
otak dengan konsentrasi yang cukup (Miller, 2010).
Para ahli, dalam dekade terakhir ini menitikberatkan perhatian pada
beberapa faktor utama yang berperan penting pada patomekanisme epilepsi dan
ERO yakni stres oksidatif dan inflamasi kronis (Vezzani dkk., 2012). Dua faktor
ini merupakan risiko yang sangat mungkin bisa dikoreksi sehingga merupakan
harapan baru pada tatalaksana ERO (Aguiar dkk., 2012).
Penderita ERO sering disertai penyakit lain seperti palsi serebral,
gangguan menelan, dan mengunyah sehingga menyebabkan malnutrisi. Beberapa
OAE seperti topiramat juga mempunyai efek menurunkan nafsu makan. Keadaan

malnutrisi menyebabkan penurunan nilai ambang kejang dan perubahan fungsi
neurotransmiter sehingga memperberat epilepsi (Crepin dkk., 2011).
2.5.2 Resistensi obat antiepilepsi dan inflamasi
Berbagai penelitian pada hewan uji dalam satu dekade terakhir
membuktikan bahwa mediator inflamasi dilepaskan dalam jumlah besar pada saat
kejang dan eksaserbasinya. Sitokin TNF-α dilepaskan dalam jangka waktu cukup
lama setelah kejang ( > 4 jam), menyebabkan peningkatan transporter P-gp, dan
setelah 6 jam paparan oleh P-gp akan meningkatkan TNF-α lebih dua kali lipat.
Interleukin-6 juga ditemukan meningkat akibat peningkatan ekspresi P-gp
(Vezzani dkk., 2011).
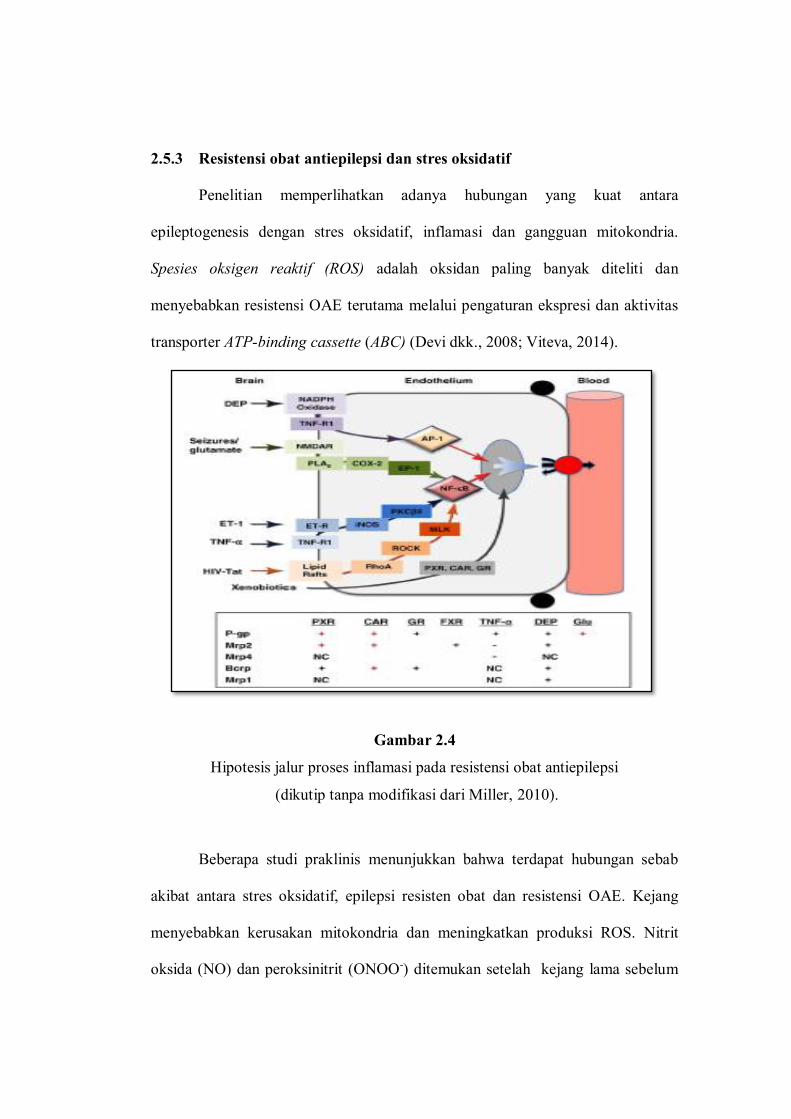
Resistensi OAE melalui jalur inflamasi dapat dibuktikan pada percobaan
binatang dimana terjadi over-ekspresi transporter obat yang melibatkan ATP, dan
mengganggu kerja protein-protein seperti P-gp, Mrp-1, Mrp-2, dan Bcrp (breast
cancer-related protein). Isolasi pada kapiler otak memperlihatkan bahwa pada
saat kejang terjadi peningkatan neurotransmiter glutamat dan neurotransmitter ini
melepaskan sinyal melalui reseptor NMDA, COX-2, prostaglandin E-2 dan NF-
kB, kemudian menyebabkan peningkatan ekspresi P-gp. Sebaliknya, kejang dapat
dihentikan pada percobaan tikus yang diberikan inhibitor COX-2, antagonis
reseptor NMDA, dan antagonis reseptor prostaglandin E2, seperti tampak pada
Gambar 2.4. Meskipun pada hewan uji menunjukkan hasil yang memuaskan,
peranan P-gp dalam resistensi OAE pada manusia masih terus diteliti (Miller,
2010).
2.5.3 Resistensi obat antiepilepsi dan stres oksidatif
Penelitian memperlihatkan adanya hubungan yang kuat antara
epileptogenesis dengan stres oksidatif, inflamasi dan gangguan mitokondria.
Spesies oksigen reaktif (ROS) adalah oksidan paling banyak diteliti dan
menyebabkan resistensi OAE terutama melalui pengaturan ekspresi dan aktivitas
transporter ATP-binding cassette (ABC) (Devi dkk., 2008; Viteva, 2014).
Gambar 2.4
Hipotesis jalur proses inflamasi pada resistensi obat antiepilepsi
(dikutip tanpa modifikasi dari Miller, 2010).
Beberapa studi praklinis menunjukkan bahwa terdapat hubungan sebab
akibat antara stres oksidatif, epilepsi resisten obat dan resistensi OAE. Kejang
menyebabkan kerusakan mitokondria dan meningkatkan produksi ROS. Nitrit
oksida (NO) dan peroksinitrit (ONOO-) ditemukan setelah kejang lama sebelum

terjadinya kematian neuron. Transporter P-gp ditemukan meningkat 4 jam setelah
paparan ROS, dan tetap tinggi hingga 48 jam kemudian dan kembali normal
setelah 48 jam dengan pemberian oksigenasi yang baik (Devi dkk., 2008).
Hidrogenperoksida (H2O2) dan deplesi partikel glutathion juga ditemukan
menyebabkan reaksi yang serupa pada hewan uji. Sebaliknya, kejang dapat
diinduksi oleh peningkatan radikal bebas. Peningkatan produksi anion
superoksida (O2-) dalam mitokondria kemudian menyebabkan peningkatan
aktivasi reseptor NMDA merupakan komponen penting terjadinya resistensi OAE
(Waldbaum dan Patel, 2010).
Mekanisme stres oksidatif menyebabkan resistensi OAE yang sudah
diketahui adalah terjadi melalui proses kerusakan DNA mitokondria (mt-DNA),
terbentuknya H2O2, penurunan GSH/GSSG, CoA reduksi (CoASH) dan GSH
disulfide (CoASSG) dalam mitokondria (Waldbaum dan Patel, 2010).
Peranan stres oksidatif dan inflamasi yang terbukti menyebabkan resistensi
OAE dan epilepsi resisten obat, menunjukkan adanya potensi terapi epilepsi
menggunakan antiinflamasi dan antioksidan di masa depan. Meskipun terbukti
ada hubungan kuat antara inflamasi dan stres oksidatif pada kejadian resistensi
OAE, belum ada penjelasan tuntas mengenai cara kerjanya, sehingga diperlukan
penelitian untuk memahami cara kerja pada tingkat seluler (Miller, 2010;
Stafstorm dkk., 2011).
2.6 Stres Oksidatif dan Epilepsi

2.6.1 Stres oksidatif dan pertahanan antioksidan
Sel tubuh manusia secara terus-menerus menghasilkan radikal bebas
berupa ROS melalui rantai respirasi selama metabolisme aerob untuk membentuk
energi (Aguiar dkk., 2012). Tubuh juga dapat memproduksi radikal bebas lain,
yakni nitrogen yang dikelompokkan dalam reactive nitrogen species (RNS).
Keberadaan ROS dan RNS pada konsentrasi rendah atau sedang berperan sangat
penting pada fisiologi normal tubuh seperti mengatur tranduksi sinyal,
menginduksi respon mitogenik, melawan infeksi, dan sebagainya (Rodriguez
dkk., 2013). Jenis ROS antara lain anion superoksida (O2-), radikal hidroksil (OH-
), dan hidrogen peroksida (H2O2) yang non-radikal. Beberapa RNS yang terlibat
dalam kematian sel karena apoptosis dan nekrosis antara lain nitric oxide (NO),
anion nitroksil (NO-), kation nitrosonium (NO+), dan peroksinitrit (ONOO-)
(Aldy, 2010;Rodriques dkk., 2013).
Radikal bebas dan antioksidan tubuh dalam keadaan normal berada dalam
keseimbangan dinamis, tetapi jika keseimbangan itu terganggu menimbulkan
keadaan stres oksidatif, seperti tampak pada Gambar 2.5. Radikal bebas
berlebihan dapat menembus membran plasma sel, menimbulkan kerusakan DNA,
lipid, dan protein pada struktur sel dan mengakibatkan modifikasi sinyal, intrupsi
transkripsi, mutasi gen, dan kematian sel termasuk neuron-neuron di otak.
Kerusakan sel jangka panjang menimbulkan berbagai penyakit seperti kanker,
arterosklerosis, arthritis, dan beberapa penyakit neurodegeneratif termasuk
epilepsi (Li dkk, 2013).

Reactive oxygen species dihasilkan di mitokondria, retikulum endoplasma,
plasma membran, sitoplasma, phagosit, peroxisome, dan enzim-enzim
cytochrome P450. Mitokondria merupakan tempat utama pembentukan ROS
karena mitokondria paling tinggi mengkonsumsi O2. Dalam keadaaan normal, 1-
2% elektron bocor dari rantai transport elektron di mitokondria dan membentuk
anion superoksida (O2-).
Gambar 2.5
Ketidakseimbangan antara radikal bebas dan antioksidan (Aldy, 2010).
Pembentukan O2- pada rantai respirasi mitokondria terjadi pada 2 tempat yaitu
pada NADH-ubiquinone oxidoreductase (kompleks I) dan ubiquinol-cytochrome c
oxidoreductase (kompleks III), sedangkan di dalam sitosol xantine oxidase (XO)
merupakan proses enzimatik lain sebagai sumber O2- dan H2O2. Anion superoksid
(O2-) juga dibentuk secara non-enzimatik melalui mentransfer elektron tunggal

kepada O2 oleh reduksi co-enzim, kelompok prosthetik yaitu flavin autoiron
sulfur cluster atau reduksi xenobiotik (Li dkk., 2013).
Anion superoksida (O2-) merupakan prekursor sebagian besar ROS dan
sebagai mediator pada rantai reaksi oksidatif. Anion superoksid secara spontan
diubah (dikatalisasi) oleh antioksidan superoksida dismutase (SOD) menjadi H2O2
dan O2, kemudian sebagian H2O2 direduksi menjadi radikal hidroksil (OH-)
dengan adanya Fe2+ melalui reaksi Fenton. Radikal hidroksil (OH-) juga dapat
dihasilkan melalui reaksi katalisasi logam (reaksi Heber-Weiss) (Maio, 2011; Li
dkk., 2013).
Nitric oxide (NO) dihasilkan dari L-arginine oleh enzim nitric oxide
synthases (NOS), termasuk neuronal NOS (nNOS), endothelial NOS (eNOS) dan
inducible NOS (iNOS), yang semuanya berlokasi di dalam sitosol. Hal sebaliknya,
mitokondrial NOS (mtNOS) suatu α-isoform dari nNOS, berlokasi di mitokondria,
yang mana merupakan co-existence dari NO dan O2- membentuk peroksinitrit
(ONOO-). Pada dasarnya NO- dihasilkan dari rekasi NO dengan Heme-Fe2+, dan
NO+ berasal dari reaksi NO dengan Heme-Fe3+ (Aguiar, 2012; Li dkk., 2013).
Sel tubuh melindungi diri terhadap kerusakan oksidatif melalui beberapa
cara yaitu 1) dua sistem buffer redoks yakni gluthation (GSH) dan thioredoksin
(TRX), 2) antioksidan enzimatik yaitu SOD, CAT, TR, GPx, serta 3) antioksidan
non-enzimatik, terdiri dari molekul endogen yaitu glutathion, asam urat, dan
nicotinamide adenine dinucleotida phosphate (NADPH) serta molekul eksogen
seperti vitamin C, vitamin E, asam α-lipoik, dan flavonoid (Vries dkk., 2011;
Ross, 2012).

Mekanisme sistem buffer GSH, GPx mengubah H2O2 menjadi H2O dan
O2, dan pada konversi ini mengubah GSH menjadi bentuk teroksidasi disulfida
(GSSG). GSSH kemudian direduksi oleh gluthation reduktase (GR) menghasilkan
GSH untuk digunakan kembali. Pada sistem buffer TRX, TRX dalam status
tereduksi (TRXR) dioksidasi menjadi thioredoxin teroksidasi (TRXo) selama
degradasi H2O2 dan kemudian direduksi oleh thioredoksin reduktase (TR). Selain
ROS, NO- secara enzimatik dibentuk oleh NOS, bereaksi dengan O2- untuk
membentuk peroksinitrit (ONOO-), seperti tampak pada Gambar 2.6 (Li dkk.,
2013).
Flavonoid merupakan antioksidan non-enzimatik eksogen yang bersumber
dari buah-buhan, umbi-umbian, dan sayur-sayuran. Efek antioksidan flavonoid
melalui beberapa mekanisme yang berbeda-beda. Antioksidan non-enzimatik
lainnya di dalam tubuh antara lain selenium, karotenoid, asam lipoik, coenzyme-
Q, melatonin, dan sebagainya (Delanty dan Dichter, 2000; Li dkk., 2013).
Gambar 2.6
Produksi ROS dan sistem antioksidan dalam sel tubuh (Li dkk., 2013).
2.6.2 Peranan radikal bebas pada epilepsi

Kejang berulang terus-menerus pada epilepsi terutama ERO,
meningkatkan kebutuhan energi sehingga konsumsi oksigen dan pembentukan
radikal bebas juga meningkat, menimbulkan keadaan stres oksidatif. Kejang dan
stres oksidatif menimbulkan inflamasi dan sebaliknya stres oksidatif dan inflamasi
menyebabkan hipereksitasi membran neuron dan kematian neuron serta memicu
kejang berikutnya, terbentuk sebuah lingkaran setan (Aguiar dkk., 2012; Vezzani
dkk., 2013).
Para ahli, saat ini percaya bahwa radikal bebas berhubungan dengan
epilepsi terutama ERO, baik sebagai penyebab kejang dan sebaliknya radikal
bebas timbul akibat kejang (Shin dkk., 2011). Radikal bebas menimbulkan
kerusakan lipid, protein dan DNA pada struktur sel sehingga menimbulkan
gangguan fungsi bahkan kematian sel termasuk neuron-neuron di otak (Hooper
dkk., 2011; Tov dan Gadoth, 2011).
2.6.2.1 Kerusakan sel akibat radikal bebas
Kerusakan oksidatif pada sel terjadi pada beberapa komponen sel seperti
asam nukleat (DNA/RNA), lipid, protein, dan karbohidrat (Pandey dkk., 2012).
2.6.2.1.1 Kerusakan asam nukleat (DNA)
Radikal bebas dapat menimbulkan pecahnya untaian rantai DNA atau
secara langsung memodifikasi basa purine dan pyridine, menyebabkan dilesi atau
mutasi yang mengakibatkan penyimpangan ekspresi gen dan kematian sel. DNA
mitokondria (mtDNA) merupakan target utama kerusakan oksidatif karena

terbatasnya mekanisme perbaikan dan kurangnya histones, menunjukkan tingkat
mutasi 10 kali lebih besar dibandingkan DNA pada inti sel (nDNA). Dirupsi
mtDNA menyebabkan gangguan fungsi mitokondria yang mengakibatkan
gangguan fungsi sel pada tingkat yang berbeda-beda, termasuk kegagalan rantai
transport elektron sehingga semakin menambah produksi ROS, yang mana
menciptakan lingkaran setan stres oksidatif. Biomarkers kerusakan oksidatif
terhadap DNA yang paling sering diperiksa: 8-hidroksi-2-deoksiguanosin (8-
OHdG) dan 8-hidroksiguanosin (8-OHG). Mekanisme yang terjadi adalah
oksidasi pada salah satu basa penyusun DNA yakni guanosin, dimana guanosin
yang teroksidasi membentuk 8-OHdG (Aldy, 2010).
Ribo Nucleac Acid (RNA) diketahui lebih rentan terhadap kerusakan
oksidatif dibandingkan DNA, karena RNA sebagian besar rantai tunggal, tidak
diproteksi oleh ikatan hidrogen dan kurang dilindungi oleh protein. Kerusakan
RNA dapat menyebabkan kesalahan dalam ekspresi protein atau disregulasi
ekspresi gen (Tov dan Gadoth, 2011).
2.6.2.1.2 Kerusakan lipid
Lipid polyunsaturated dalam lipoprotein dan membran sel sangat peka
terhadap kerusakan oksidatif, yang mengakibatkan peroksidasi lipid. Peroksidasi
lipid dapat menghasilkan aldehid reaktif yang disebut malondialdehid (MDA).
Aldehid reaktif dapat berikatan dengan protein secara kovalen, selanjutnya
mengganggu fungsinya dan menginduksi kerusakan sel. Produk peroksidasi
(radikal peroksil) lebih stabil dibandingkan ROS dan dapat berdifusi menyeberang

sel dan bereaksi dengan konstituen sel lain menyebabkan kerusakan sel secara
difus. Produk akhir peroksidasi lipid yang dapat dijadikan biomarker yakni:
MDA, hexanoyllysine adduct (HEL), acrolein-lysine adduct (ACR) dan 4-hydroxy
nonenal (4-HNE). Neuron di otak sangat sensitif terhadap kerusakan oksidatif
karena membran neuron sebagian besar dibentuk oleh fosfolipid (Waldbaum dan
Patel, 2010).
2.6.2.1.3 Kerusakan protein
Radikal bebas dapat mengoksidasi baik rantai utama atau rantai samping
protein. Modifikasi rantai utama menyebabkan pembentukan radikal peroksil
dengan sebuah rantai reaksi yang menyebabkan protein cross-linking dan atau
pembelahan ikatan peptida. Residu oksidasi rantai samping menyebabkan
gangguan fungsi enzim, reseptor, neurotransmiter, dan struktur protein. Inaktivasi
enzim aconitase digunakan sebagai marker kerusakan oksidatif protein.
Keberadaan iron yang labil di dalam iron sulfur center aconitase membuatnya
sensitif terhadap radikal bebas, khususnya anion superoksida dan peroksinitrit.
Modifikasi protein oleh peroksinitrit terutama terjadi pada residu tirosin dan
triptofan. Kadar 3-nitrotirosin, produk utama nitrasi protein merupakan marker
penting kerusakan protein akibat stres oksidatif (Aldy, 2010).
2.6.2.2 Pembentukan radikal bebas sebagai akibat kejang epileptik
Kadar radikal bebas meningkat setelah kejang dibuktikan pada banyak
percobaan hewan uji dengan metoda pemeriksaan berbeda, termasuk pengukuran

produksi radikal bebas secara tidak langsung, marker kerusakan oksidatif DNA,
lipid, dan protein serta evaluasi kadar antioksidan tubuh (Tov dan Gadoth, 2011;
Rodriguez dkk., 2013). Bukti-bukti tersebut antara lain: peningkatan kadar radikal
bebas pada cairan cerebrospinal otak tikus yang diinduksi kejang dengan
bicuculline. Peterson dkk (2006), melaporkan peningkatan produksi anion
superoksida dengan pengecatan hidroetidium pada otak tikus yang mengalami
status epileptikus karena induksi litium-pilokarpin. Radikal hidroksil meningkat
pada tikus yang diinduksi kejang dengan pentilenetetrazol (PTZ). Peningkatan
kadar 8-OHdG pada tikus yang diinduksi kejang dengan asam kainat. Erakovic
dkk (2007), penurunan kadar antioksidan SOD dan GPx pada otak tikus yang
mengalami syok elektrokonvulsif (Tov dan Gadoth, 2011).
2.6.2.3 Radikal bebas menyebabkan kejang epileptik
Cedera otak seperti trauma kepala, hipoksia, infeksi, dan sebagainya
bersifat epileptogenik, hal ini berhubungan dengan stres oksidatif (Aldy, 2010).
Cedera otak menyebabkan perubahan kaskade struktur, fisiologi, dan biokimia
pada otak seperti perubahan aliran darah otak (cerebral bloob flow) dan
vasoregulasi, dirupsi SDO, peningkatan tekanan intrakranial, perdarahan
fokal/difuse, inflamasi, stres oksidatif, nekrosis, dan dirupsi serabut saraf dan
pembuluh darah (Ogino dan Wang, 2007).
Peningkatan produksi ROS menginduksi perubahan seluler dan molekuler
yang menyebabkan hipereksitabilitas atau kematian neuron. Salah satu akibatnya
adalah kerusakan aktivitas Na+/K+-ATP-ase, yang dalam kondisi normal

memelihara gradient ionic membran neuron. Penurunan aktivitas Na+/K+-ATP-
ase, dapat menurunkan nilai ambang kejang. Cedera terhadap membran neuron
juga dapat menyebabkan peningkatan pelepasan neurotransmiter eksitasi utama
seperti glutamat dan aspartat atau menurunkan neurotransmiter inhibisi seperti
GABA. Jadi pelepasan berlebihan asam amino yang bersifat eksitasi dapat
mencetuskan kejang, peningkatan produksi NO dan ROS, serta radikal bebas ini
menyebabkan kerusakan sel dan memicu kejang berulang selanjutnya (Rodriguez
dkk., 2013).
Hubungan radikal bebas dan epileptogenesis didukung oleh penelitian
hewan uji dan penelitian klinis. Gangguan fungsi mitokondria yang menimbulkan
stres oksidatif ditemukan pada fokus kejang penderita epilepsi lobus temporal dan
hewan model epilepsi.Pada penderita epilepsi mioklonik progresif, aktivitas
SOD1 (cytosolic SOD) rendah pada serum. Peneliti lain melaporkan SOD2
(mitokondria manganese SOD) regulasinya menurun pada korteks serebri
penderita epilepsi dibandingkan bukan epilepsi. Aktivitas antioksidan GPx dan
selenium yang berperan penting pada aktivitas GPx rendah pada anak epilepsi
(Tov dan Gadoth, 2011; Frietas, 2012).
2.7 Epilepsi dan Inflamasi
Peranan inflamasi pada patofisiologi kejang banyak diteliti dalam dekade
terakhir, dimana terdapat banyak bukti menunjukkan adanya hubungan antara
epilepsi dan sistem imun. Sitokin dan proses inflamasi yang terjadi, tampaknya
memiliki peranan penting dalam patogenesis epilepsi (Li dkk., 2011). Pada

epilepsi resisten obat ditemukan gliosis reaktif bermakna yang terlibat dalam
epileptogenesisnya. Patogenesis lain yang diduga terlibat pada epilepsi resisten
obat adalah gangguan pada saluran membran astrosit, reseptor, dan transporternya
(Vezzani, 2011). Penelitian klinis dan patologis memperlihatkan bahwa inflamasi
dapat menjadi pemain tunggal dalam kejadian kejang tanpa disertai infeksi
ataupun adanya proses lain (Vezzani dkk., 2012; Shimada dkk., 2014). Kejang
tidak hanya menimbulkan induksi ekspresi sitokin di otak, tetapi mengakibatkan
juga perubahan sitokin di perifer (Li dkk., 2011). Proses inflamasi di otak dapat
disebabkan oleh kejang, dengan onset yang cepat (< 30 menit) dan berlangsung
beberapa hari walaupun kejang sudah berhenti, dimana mekanisme inflamasi
endogen juga turut terlibat secara tidak efisien (Vezzani dkk., 2012).
Kerusakan permeabilitas sawar darah otak (SDO) merupakan salah satu
jalur yang memudahkan terjadinya kejang pada epilepsi. Inflamasi intrinsik
menyebabkan induksi inflamasi pada keadaan kejang yang kemudian
menyebabkan kerusakan SDO (Li dkk., 2011). Adanya kerusakan SDO
memungkinkan sitokin menginduksi kejadian kejang berikutnya, seperti tampak
pada Gambar 2.7 (Shimada dkk., 2014).
2.7.1 Peranan inflamasi pada eksitabilitas otak
Cedera neuron akibat kejang, selanjutnya mengakibatkan sitokin,
prostaglandin, dan berbagai molekul inflamasi lainnya bersama reseptornya
mengalami induksi di dalam neuron dan mengaktifkan sel glia dan juga sel

endotel pada SDO di area otak yang mengalami cedera (Vezzani, 2011). Sitokin
dan berbagai
Gambar 2.7
Mekanisme inflamasi yang terlibat dalam epileptogenesis (dikutip tanpa
modifikasi dari Shimada dkk., 2014).
mediator ini merupakan bagian dari aktivasi imunitas innate dan adaptif di perifer,
namun akhir-akhir ini diketahui juga berperan dalam fungsi neuromodulator non
konvensional dalam susunan saraf pusat yang bekerja secara langsung dan tidak
langsung yang akhirnya mempengaruhi nilai ambang kejang pada tingkat selular
(Vezzani dkk., 2012).
IL-1ß
HMGB-1
IL-6 TNF-α

2.7.2 Interleukin-6
Interleukin-6 (IL-6) merupakan sitokin multifungsi yang mengatur
berbagai respon imun dan inflamasi. Dalam sel target, sinyal IL-6 ini
ditransmisikan melalui kompleks reseptor IL-6 (IL-6R) dan dua molekul gp130.
Baik IL-6 dan sIL-6R (bentuk IL-6R yang terlarut) dapat memicu terbentuknya
sinyal gp130 tanpa harus melalui ikatan membran dengan Il-6R (Li dkk., 2011;
Erta dkk., 2012).
Interleukin-6 dengan cepat terbentuk di daerah hipokampus, korteks, girus
dentata, amigdala, dan meningen setelah terjadinya kejang, namun mRNA
reseptor IL-6 tampaknya hanya terbentuk pada hipokampus (Vezzani, 2011).
Interleukin-6 meningkat sangat cepat pada sel glia, namun sifatnya juga transien
dan menghilang dengan cepat, maksimal 6 jam setelah terjadinya status
epileptikus, hal ini mungkin disebabkan oleh depolarisasi membran yang
menyebabkan terakumulasinya protein IL-6 dan mRNA IL-6 dalam neuron
(Vezzani dkk.,2013). Pada sisi lain mRNA IL-6 meningkat pada thalamus,
korteks, amigdala, dan hipokampus dalam 10-16 jam setelah terjadinya kejang
dan menghilang setelah lima hari, sementara mRNA gp130 meningkat di lokasi
yang serupa dalam waktu yang lambat (Erta dkk., 2012; Vezzani dkk.,2013),
seperti tampak pada Gambar 2.8 di bawah ini.

Gambar 2.8
Mekanisme kerja IL-6 di dalam neuron (Erta dkk., 2012).
Penelitian mengenai keterlibatan IL-6 dalam kejadian kejang memberikan
hasil beragam dan masih terus diperdebatkan (Erta dkk., 2012). Pemberian IL-6
intranasal pada tikus dapat memperpanjang latensi dan memperpendek durasi
kejang yang diinduksi oleh demam, sementara pada penelitian lain, IL-6 intranasal
pada tikus dewasa menyebabkan latensi memendek, tingkat keparahan lebih
besar, dan mortalitas lebih tinggi pada status epileptikus. Interleukin-6 pada tikus
yang mengalami overekspresi IL-6 akan menyebabkan kekambuhan kejang tonik-
klonik, yang kemungkinan melalui reduksi inhibisi GABA (Li dkk., 2011; Erta
dkk., 2012).
Beberapa penelitian menunjukkan bahwa seperti sitokin lain, IL-6
menunjukkan efek ganda, seperti tampak pada Gambar 2.9, yang mungkin
dibutuhkan dalam perkembangan normal sistem saraf dan neuroplastisitas, namun
penjelasan mengenai bagaimana efek ini dapat saling melengkapi membutuhkan
penelitian lebih lanjut (Li dkk., 2011; Erta dkk., 2012).

Peningkatan IL-6 dalam plasma dan CSS terjadi secara bermakna dalam
24 jam setelah kejang epilepsi dan kejang demam. Kadar IL-6 pada CSS jauh
lebih tinggi dibandingkan kadarnya dalam plasma, sehingga diduga awalnya IL-6
terbentuk pada neuron di otak (Erta dkk., 2012).
Gambar 2.9
Peranan IL-6 pada cedera otak (Erta dkk., 2012).
Kejang dapat memengaruhi otak dan menginduksi pelepasan katekolamin.
Katekolamin yang dilepaskan dapat mempengaruhi sel mononuklear di dalam
darah perifer, yang akan memicu respon IL-6 di perifer selain prosesnya di neuron
otak. Kadar IL-6 pada darah hingga kini belum diketahui apakah memberikan
hasil sebanding dengan kadarnya dalam otak, walaupun dalam kedua sampel
didapatkan peningkatan kadar yang bermakna (Li dkk., 2011; Youn, dkk., 2013).
Pada akhirnya dapat disimpulkan bahwa, terdapat hubungan timbal balik
seperti sebuah lingkaran setan antara cedera otak, stres oksidatif, inflammasi,
kejang, dan kematian neuron. Masing-masing proses tersebut saling
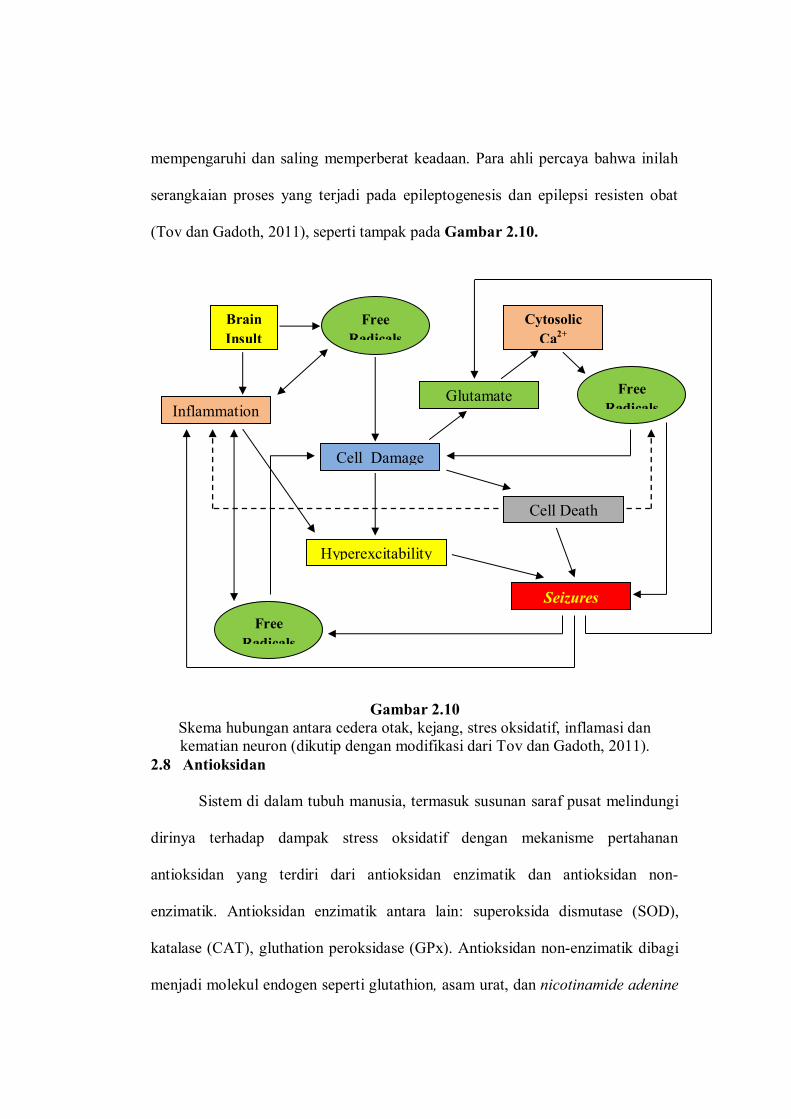
mempengaruhi dan saling memperberat keadaan. Para ahli percaya bahwa inilah
serangkaian proses yang terjadi pada epileptogenesis dan epilepsi resisten obat
(Tov dan Gadoth, 2011), seperti tampak pada Gambar 2.10.
Gambar 2.10 Skema hubungan antara cedera otak, kejang, stres oksidatif, inflamasi dan kematian neuron (dikutip dengan modifikasi dari Tov dan Gadoth, 2011).
2.8 Antioksidan
Sistem di dalam tubuh manusia, termasuk susunan saraf pusat melindungi
dirinya terhadap dampak stress oksidatif dengan mekanisme pertahanan
antioksidan yang terdiri dari antioksidan enzimatik dan antioksidan non-
enzimatik. Antioksidan enzimatik antara lain: superoksida dismutase (SOD),
katalase (CAT), gluthation peroksidase (GPx). Antioksidan non-enzimatik dibagi
menjadi molekul endogen seperti glutathion, asam urat, dan nicotinamide adenine
Hyperexcitability
Cell Damage
Cell Death
Glutamate Inflammation
Free Radicals
Seizures
Brain Insult
Cytosolic Ca2+
Free Radicals
Free Radicals

dinucleotida phosphate (NADPH) serta molekul eksogen seperti vitamin C,
vitamin E, asam α-lipoik, dan flavonoid (Vries dkk., 2011; Ross, 2012).
Keadaan stres oksidatif mengaktifkan mekanisme adaptif tubuh yang
bertujuan melindungi sel terhadap kerusakan oksidatif dan menjaga keseimbangan
reaksi reduksi-oksidasi di jaringan. Respon ini diatur pada tingkat transkripsional
dengan transkripsi faktor nuclear factor-Erythroid 2-related factor-2 (Nrf2).
Dalam kondisi fisiologis, Nrf2 terikat pada actin-bound Kelch-like ECH yang
berhubungan dengan protein Kelch-like ECH-associated molecule-1 (Keap-1)
yang berlokasi di dalam sitoplasma. Akibat paparan ROS,Nrf2 berdisosiasi dari
Keap-1 dan berpindah tempat (translokasi) ke inti sel (nukleus), kemudian
berikatan dengan antioxidant response element (ARE) di dalam gen yang
mengkode enzim antioksidan. Sampai saat ini diketahui lebih dari 200 Nrf2-ARE
gen yang terlibat pada detoksifikasi dan pertahanan antioksidan yang telah
diidentifikasi dengan baik yakni: katalase, SOD, GPx, peroxiredoksin(Prxs), heme
oxygenases, NAD(P)H:quinineoxireductase-1(NQO1), dan NHR:quinone
oxidoreductase-2 (NQO-2). Antioksidan enzimatik terpenting di dalam tubuh
manusia adalah SOD, CAT dan GPx (Vries dkk., 2011).
2.8.1 Antioksidan enzimatik
2.8.1.1 Superoksida dismutase
Anion superoksida (O2-) merupakan salah satu radikal bebas utama di
dalam sel, sehingga antioksidan SOD memegang peranan penting pada
detoksifikasi O2-. Superoksida dismutase tergolong metalloenzym yang

mengkatalisa perubahan O2- menjadi H2O2 dan O2, yang mana H2O2 merupakan
senyawa yang kurang toksik dibandingkan O2- (Vries dkk., 2011).
Sitosol sel mengandung copper-zinc SOD (CuZnSOD/SOD1), matriks
mitokondria memiliki manganese SOD (MnSOD/SOD2) yang mengeliminasi
superoksida di dalam matriks atau pada bagian dalam membran dalam
mitokondria, disamping itu juga terdapat ekstraseluler CuZnSOD (SOD3). Di
dalam otak banyak ditemukan SOD1 dan SOD2, SOD1 terutama diekspresi oleh
astrosit, sedangkan SOD2 oleh neuron. Aktivitas SOD1 dan SOD2 relatif rendah
di dalam mikroglia dan oligodendrosit. Ada bukti kuat keterlibatan SOD pada
penyakit neurodegeneratif dan neuroinflamasi termasuk epilepsi (Delanty dan
Dichter, 2000; Vries dkk., 2011).
2.8.1.2 Katalase
Anion superoksida dikonversi oleh SOD menghasilkan hidrogen peroksida
(H2O2) dan O2 yang dapat berdifusi menembus membran sel. Keberadaan metal
iron atau copper, H2O2 dapat berubah menjadi radikal hidroksil (OH-) yang sangat
reaktif, sedangkan pertemuan H2O2 dengan NO membentuk anion peroksinitrit
(ONOO-) yang juga bersifat reaktif. Upaya menyingkirkan H2O2, sel dilindungi
oleh beberapa hydrogen peroxide–removing enzyme seperti katalase, gluthation
peroksidase (GPx), dan peroksiredoksin (Prxs). Katalase terutama dihasilkan di
peroksisom yang berfungi mengkatalisa konversi H2O2 menjadi H2O dan O2
(Vries dkk., 2011).

2.8.1.3 Gluthation peroksidase
Glutathion peroksidase yang berlokasi di dalam sitosol dan kompartemen
mitokondria, merupakan kelompok enam enzim yang mengandung selenium yang
mendetoksifikasi organik peroksida dan hidrogen peroksida. Aktivitas Glutathion
peroksidase-1 pada umumnya lebih tinggi dibandingkan katalase, menandakan
bahwa di dalam otak peranan glutathion peroksidase lebih penting dibandingkan
katalase untuk menghilangkan H2O2. Glutathion peroksidase juga mempunyai
peranan protektif pada neuroinflammasi (Hooper dkk., 2011; Vries dkk., 2011).
2.8.2 Antioksidan non-enzimatik
Antioksidan non-enzimatik dapat dibagi menjadi molekul endogen seperti
glutathion, asam urat, dan nicotinamide adenine dinucleotida phosphate
(NADPH) serta antioksidan eksogen seperti vitamin C, vitamin E, asam α-lipoik,
dan flavonoid (Park dan Hallerm, 2011; Frietas, 2012).
2.8.2.1 Vitamin C dan vitamin E
Vitamin C merupakan antioksidan larut dalam air yang dapat mereduksi
radikal bebas dari berbagai sumber. Vitamin C juga berperan pada daur ulang
radikal vitamin E. Suatu hal yang menarik, vitamin C juga berfungsi sebagai pro-
oksidan dalam kondisi tertentu. Vitamin E adalah antioksidan larut dalam lemak
yang memainkan peranan penting melindungi membran sel terhadap kerusakan
oksidatif. Aktivitas primer vitamin E adalah menghancurkan radikal peroksil di
dalam membran sel (Vries dkk., 2011; Ogunro dkk., 2013).

2.8.2.2 Glutathion
Glutathion dipertahankan sebagai thiol oleh aksi glutathion reduktase,
sebuah enzim yang mereduksi oxidized glutathione (GSSG) menjadi glutathion
(GSH). Glutathion berperan penting sebagai antioksidan di matriks mitokondria.
Dalam hal berpartisipasi sebagai substrat di dalam reaksi GPx dan GPHx, GSH
juga dapat secara langsung menangkap radikal bebas dan oksidan lainnya seperti
OH-, O2-, H2O2 dan ONOO-. GSH juga mempunyai target utama peroksinitrit
(ONOO-) (Frietas, 2012; Ogunro dkk., 2013).
2.9 Pengobatan Epilepsi
Pengobatan epilepsi bertujuan untuk mencapai remisi (“sembuh”)
sehingga dampak buruk terhadap tumbuh kembang anak dapat diminimalkan.
Beberapa pilihan pengobatan yang telah dilakukan antara lain: obat antiepilepsi
(OAE), pembedahan, stimulasi saraf vagus, dan diet ketogenik (Kwan dkk., 2011;
Berg dkk., 2012).
2.9.1 Pengobatan farmakologis menggunakan OAE
Sekitar 20 macam OAE telah dikenal sampai saat ini. Pada anak ada empat
macam OAE lini pertama yang direkomendasi penggunaannya secara rutin yaitu
fenobarbital, asam valproat, karbamazepin, dan fenitoin (Berg dkk., 2012). Prinsip
pengobatan epilepsi adalah dimulai dengan monoterapi I, bila kejang tidak
terkontrol, dilanjutkan monoterapi II dan jika kejang tidak juga terkontrol dapat

diberikan kombinasi (duoterapi/politerapi) dengan dua atau tiga jenis OAE lini
pertama.
Pada epilepsi umum pilihan monoterapi I adalah asam valproat, sedangkan
monoterapi II dapat dipilih salah satu dari OAE lini pertama yang lain. Pada
epilepsi fokal, pilihan monoterapi I adalah karbamazepin, sedangkan monoterapi
II dapat dipilih salah satu dari OAE lini pertama yang lain. Jika dengan
monoterapi I dan II kejang tidak terkontrol, maka dapat dilakukan terapi
kombinasi (duoterapi/politerapi) dengan dua atau tiga OAE lini pertama
(selengkapnya dapat dilihat pada lampiran 1).
Penggunaan OAE mencapai remisi (“sembuh”) 60-70% kasus, dengan
rincian OAE monoterapi I menghasilkan remisi 47%, dan OAE monoterapi II dan
kombinasi 2 OAE (duoterapi) menambah angka remisi 13% kasus, serta
pemberian OAE ke-3 atau ke-4 menambah remisi hanya sekitar 4% kasus
sehingga sekitar 30-40% menjadi epilepsi resisten obat (Kwan dkk, 2011; Berg
dkk., 2012).
Beberapa macam OAE lini kedua atau OAE generasi baru seperti
topiramat, levitirasetam, vigabatrin, gabapentin, oksarbazepin, dan sebagainya
belum digunakan secara rutin, hanya direkomendasi untuk pengobatan add-on
pada beberapa jenis sindroma epilepsi bayi dan anak (Sillanpaa dan Schmidt,
2011).
Epilepsi dinyatakan remisi atau“sembuh” jika dengan OAE tercapai
keadaan bebas kejang selama dua sampai tiga tahun dengan gambaran EEG
normal, meskipun ada epilepsi yang membutuhkan pengobatan OAE seumur

hidup yakni epilepsi mioklonik juvenile (Berg dkk., 2012). Epilepsi yang sudah
remisi, kemungkinan mengalami kumat lagi sekitar 15-20% (Kwan dkk, 2011).
2.9.2 Terapi pembedahan pada epilepsi
Pembedahan yang dilakukan adalah menghilangkan fokus epileptogenik
atau area kecil pada otak yang dicurigai merupakan fokus kejang dengan tindakan
lobektomi, lesionectomy atau corpus callosumectomy. Tindakan bedah hanya
tepat untuk epilepsi fokal yang berasal dari satu fokus yang jelas di otak, seperti
epilepsi lobus temporalis dengan tingkat keberhasilan beragam (Kelly dan Chung,
2011). Tindakan pembedahan epilepsi di Indonesia sampai saat ini belum banyak
dikerjakan.
2.9.3 Stimulasi saraf vagus
Stimulasi saraf vagus adalah modalitas pengobatan non-farmakologi yang
merupakan alternatif atas kegagalan OAE untuk mengontrol kejang. Pengobatan
ini memberi keuntungan pada sindrom Lennox-Gastaut dan epilepsi absanse yang
resisten obat. Saraf vagus berhubungan dengan nukleus traktus solitarius pada
batang otak dan pada tempat tersebut akan terjadi efek yang diharapkan. Area
yang dapat diaktifkan oleh saraf vagus antara lain amygdala, hipokampus, girus
cingulate, dan korteks somatosensori kontralateral. Stimulasi pada area-area
tersebut berdampak sebagai inhibitor serangan kejang pada epilepsi (Min dkk.,
2011; Patil dkk., 2011). Stimulasi saraf vagus untuk epilepsi di Indonesia belum
banyak dikerjakan.

2.9.4 Diet ketogenik
Beberapa studi membuktikan bahwa anak epilepsi mengkonsumsi banyak
lemak, sedikit karbohidrat dan protein dapat mengalami kejang lebih jarang. Diet
ini disebut diet ketogenik yang menyebabkan tubuh berada dalam kondisi ketosis.
Bagaimana keadaan ketosis dapat menghambat terjadinya kejang, belum dipahami
dengan baik. Satu penelitian menunjukkan bahwa produk keadaan ketosis yang
dinamakan beta-hydroxybutyrate (BHB) dapat menghambat kejang pada binatang
model (Neal dkk., 2010). Pemberian secara oral sering sulit ditoleransi, tetapi
keberhasilan diet ketogenik jika diberikan secara parenteral selama beberapa
bulan dan penderita harus rawat inap (Caraballo dkk., 2011). Peneliti pernah
mencoba diet ketogenik secara oral pada beberapa pasien ERO di RSUP Sanglah,
tetapi hasilnya tidak memuaskan karena kurang dapat ditoleransi oleh penderita.
Obat antiepilepsi bersifat simtomatik saja, hanya dapat menekan aktivitas
kejang tanpa dapat memperbaiki proses patologis yang mendasari. Para ahli terus
berusaha menemukan formula pengobatan yang dapat menekan progresivitas
epilepsi melalui patofisiologi yang mendasari antara lain dengan antioksidan dan
antiinflamasi karena stres oksidatif dan inflamasi diyakini sangat berperan pada
epileptogenesis dan epilepsi resisten obat (Rodriguez dkk., 2013; Vezzani dkk.,
2013).
2.10 Peranan Antiinflamasi pada Pengobatan Epilepsi Resisten Obat

Bukti adanya peranan antiinflamasi pada epilepsi adalah penggunaan
ACTH, kortikosteroid, plasmapharesis, dan immunoglobulin untuk pengobatan
kejang dan beberapa macam epilepsi memberikan keberhasilan beragam.
Mekanisme kerja antiinflamasi adalah menekan inflamasi atau secara langsung
pada eksitabilitas neuron otak dan menekan agen proinflamasi endogen otak
(Vezzani, 2011; Vezzani dkk., 2013). Prednison dan ACTH telah digunakan
dengan hasil yang cukup memuaskan pada sindrom epilepsi spasme infantil
(Stafstorm dkk., 2011).
2.11 Peranan Antioksidan pada Pengobatan Epilepsi Resisten Obat
Efek antioksidan terhadap otak sangat tergantung pada kemampuan
senyawa tersebut melewati SDO, senyawa larut dalam lemak memberikan efek
lebih baik dibandingkan senyawa larut dalam air. Inflamasi pada otak
berhubungan dengan gangguan integritas SDO yang membuat senyawa-senyawa
tersebut lebih mudah menembusnya (Delanty dan Dichter, 2000).
Pada beberapa penelitian, terbukti stres oksidatif terlibat dalam
patogenesis dan progresivitas epilepsi (Surekha dan Melinkeri, 2010; Miguel,
2011). Efek pemberian antioksidan pada kejang yang diinduksi pilokarpin,
menunjukkan penurunan kadar pro-oksidan nitrit pada hipokampus tikus. Efek
biologi radikal bebas dikontrol secara in vivo oleh antioksidan seperti vitamin E,
vitamin C, vitamin A, dan glutathion (Freitas, 2012).
Bukti pertama manfaat antioksidan pada epilepsi dilaporkan oleh Ben-
Menachem, dkk (1995), menemukan kadar dan aktivitas antioksidan GPx, CAT,

dan SOD rendah pada epilepsi mioklonik progresif resisten obat dibandingkan
kontrol sehat. Pemberian antioksidan N-acetylcystein (NAC) dengan dosis tinggi
(6 gram/hari) dapat memperbaiki kondisi penderita (Rodriques dkk., 2013).
Pemberian vitamin E dapat mencegah berkembangnya aktivitas gelombang
epileptik yang diinduksi Fe pada otak tikus. Vitamin E juga dilaporkan nyata
dapat menghambat onset kejang elektrografik yang diinduksi dengan injeksi
ferrous klorida intraserebral. Pemberian vitamin E dan vitamin C dapat
menurunkan produksi marker stress oksidatif dan memperlambat atau menekan
gelombang epileptik pada tikus model epilepsi. Pada tikus yang diinduksi kejang
dengan Fe, pemberian antioksidan melatonin menghambat aktivitas gelombang
kejang elektrografik dengan menekan peroksidasi (Freitas, 2012).
Antioksidan eksogen yang mulai banyak digunakan pada beberapa macam
penyakit neurodegeneratif termasuk epilepsi adalah golongan flavonoid.
Flavonoid adalah kelompok senyawa polifenol yang bersumer dari berbagai buah-
buahan, umbi-umbian, dan sayur-sayuran, mempunyai efek antioksidan yang
poten, jauh lebih poten dibandingkan antioksidan klasik vitamin E. Flavonoid
yang telah diidentifikasi sampai saat ini lebih dari 8000 jenis. Flavonoid bisa
tampil sebagai glycosides yang terikat dengan sebuah molekul gula atau sebagai
aglycones yang tidak terikat molekul gula (Rytter, 2011).
Salah satu bentuk flavonoid yang mempunyai efek antioksidan sangat
poten dan efek antiinflamasi adalah anthosianin (Han dkk., 2007; Montilla dkk.,
2010), disamping itu anthosianin juga memiliki efek antikejang melalui potensiasi
reseptor GABA (Katzung, 2011 ). Anthosianin banyak terkadung pada sayur,

buah-buahan, dan umbi-umbian termasuk umbi ubi jalar ungu (Suprapta dkk.,
2004).
2.12 Ubi Jalar Ungu
Ubi jalar, ketela rambat atau sweet potato berasal dari benua Amerika,
mulai menyebar ke seluruh dunia terutama negara-negara beriklim tropis pada
abad ke 16, dibawa oleh orang-orang Spanyol ke kawasan Asia, terutama Filipina,
Jepang, dan Indonesia. Ubi jalar ada yang berwarna putih, kuning, oranye, dan
ungu. Ubi jalar ungu tergolong Plantae, divisi Magnoliophyta, kelas
Magnoliopsida, ordo Solanales, famili Convolvulaceae, genus Ipomoea, dan
spesies Ipomoea batatas. Umbi ubi jalar ungu banyak mengandung flavonoid
terutama anthosianin, disamping zat-zat lainnya (Yusuf dkk., 2008).
Ubi jalar ungu varietas baru dengan kandungan anthosianin (flavonoid)
lebih tinggi banyak dikembangkan di Jepang, Korea, dan New Zaeland. Kultivar
paling penting adalah varietas Ayamurasaki dan Yamagamamurasaki dengan
beberapa macam klonnya. Berdasarkan rasio peonidin dan cyanidin (peo/cy ratio),
klon ubi jalar ungu dibedakan dua kelompok yaitu tipe cyanidin (peo/cy < 1,0)
dengan dominan warna ungu dan tipe peonidin (peo/cy > 1,0) dengan dominan
warna merah.
Beberapa varietas ubi jalar ungu ditemukan di Indonesia, termasuk di Bali
yang dikenal dengan nama lokal sela bojog atau ubi bojog yang mempunyai
kandungan anthosianin cukup tinggi dibandingkan ubi jalar ungu di daerah lain
(Suprapta, 2004). Sejak diperkenalkan dua varietas ubi jalar ungu dari Jepang

yaitu Ayamurasaki dan Yamagamamurasaki, budidayanya semakin luas di
Indonesia. Selain itu Balitkabi juga memiliki tiga klon ubi jalar ungu yaitu MSU
01022-12, MSU 01008-16 dabn MSU 01016-19, seperti tampak pada Gambar
2.11 (Yusuf dkk., 2003; Montilla dkk., 2010). Ubi jalar ungu banyak diminati
konsumen karena umbinya mengandung gizi yang baik dan mempunyai fungsi
fisiologis tertentu untuk kesehatan termasuk untuk otak (Jeong dkk., 2013).
Kandungan anthosianin bervariasi pada beberapa kultivar ubi jalar ungu.
Di Jepang, varietas Chiran murasaki mengandung anthosianin 29,1 mg/100 gram,
Naka murasaki 6,5 mg/100 gram, Purple sweet 12,4 mg/100 gram, dan
Tanegashima murasaki 9,3 mg/100gram (Montilla dkk., 2010). Umbi ubi jalar
ungu yang tumbuh di Bali mengandung anthosianin berkisar 110-210 mg/100
gram (Suprapta, 2004), sedangkan umbi ubi jalar ungu di daerah lain berbeda-
beda seperti: umbi ubi jalar ungu Malang mengandung anthosianin 511,70
mg/100 gram, Bone 530,06/100 gram, sumedang 508,45 mg/100 gram, Salatiga
79,47 mg/100 gram, Sukabumi 606,08 mg/100 gram, Bangkok 50,68 mg/100
gram, Jambi 69,37 mg/100 gram, Yangyang 65,16 mg/100 gram, dan selo
Banyuwangi 76 mg/100 gram (Widiati, 2010).
Anthosianin berasal dari kata Yunani Antos berarti bunga dan cyanos
berarti biru. Anthosianin pada umbi ubi jalar ungu memiliki stabilitas tinggi
dibandingkan anthosianin sumber lain dan memiliki kasiat antioksidan 2,5-3,2
kali lebih tinggi dibandingkan blueberry (Ebrahimi dan Schluesener, 2012 ). Umbi
ubi jalar ungu juga sumber karbohidrat dan kalori, mengandung protein, lemak,
dan sejumlah kecil vitamin dan mineral (Dreiseitel, 2011).

Varietas ubi jalar ungu di Bali Beberapa varietas ubi jalar ungu di (Sela Bojog) Jepang
Gambar 2.11 Tanaman dan umbi ubijalar ungu (Suprapta, 2004; Jusuf dkk., 2011).
2.12.1 Flavonoid
Flavonoid tergolong senyawa polifenol, dengan struktur dasar adalah inti
flavan mengandung 15 kerangka carbon yang disusun dalam dua cincin phenyl
terikat oleh tiga jembatan carbon yang umumnya dikelilingi oksigen, kemudian
membentuk tiga cincin dinamakan cincin A,B,C, seperti tampak pada Gambar
2.12. Variasi klas flavonoid berbeda pada tingkat oksidasi dan perubahan pada
cincin C, sedangkan kandungan senyawa dalam satu klas berbeda pada cincin A
dan B. Beberapa klas flavonoid utama yaitu flavonols, flavones, flavanones,
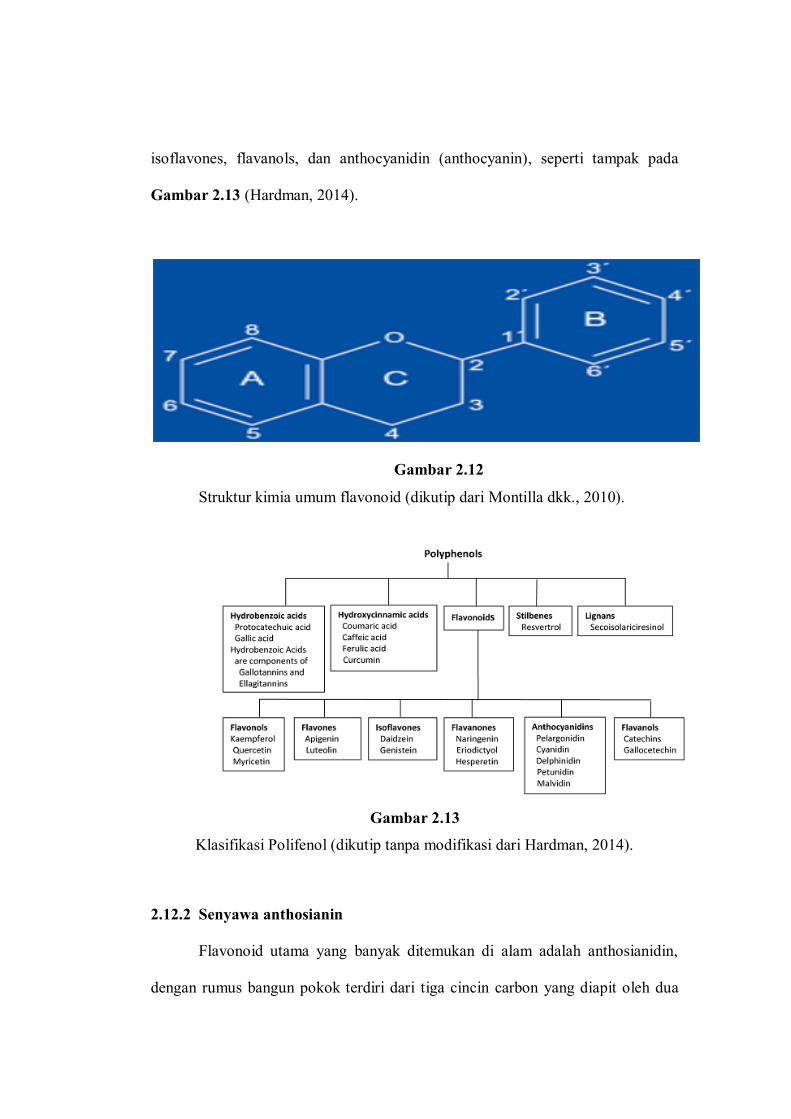
isoflavones, flavanols, dan anthocyanidin (anthocyanin), seperti tampak pada
Gambar 2.13 (Hardman, 2014).
Gambar 2.12
Struktur kimia umum flavonoid (dikutip dari Montilla dkk., 2010).
Gambar 2.13
Klasifikasi Polifenol (dikutip tanpa modifikasi dari Hardman, 2014).
2.12.2 Senyawa anthosianin
Flavonoid utama yang banyak ditemukan di alam adalah anthosianidin,
dengan rumus bangun pokok terdiri dari tiga cincin carbon yang diapit oleh dua

cincin benzene. Bentuk 3 atau 3,5-glikosida dari anthosianidin disebut anthosianin
(Dreiseitel, 2011). Sebanyak 17 macam anthosianidin telah diketahui sampai
tahun 2003, tetapi hanya enam yang terpenting dengan distribusinya masing-
masing yakni: cyanidin (cy) 50%, peonidin (pn) 12%, pelargonidin (pg) 12%,
delphinidin (dp) 12%, malvidin (mv) 7%, dan petunidin (pt) 7% (Dreiseitel,
2011). Perbedaan struktur kimia enam anthosianidin/anthosianin terletak pada
posisi cincin 3 dan 5, dengan struktur kimia umum seperti tampak pada Gambar
2.14 (Montilla dkk., 2010).
Anthosianin dalam bentuk glikosida semua larut dalam air dan tidak larut
dalam pelarut organik. Perbedaan individual antara beberapa bentuk anthosianidin
terletak pada tingkat hidroksilasi cincin aromatik. Perbedaan individual antara
senyawa anthosianin terletak pada posisi gugus glikosil (residu gula) di dalam
molekulnya dan perbedaan ini sangat menentukan bioaktivitas dan efek
farmakologisnya masing-masing anthosianin (Kovacsova, dkk., 2010).
Gambar 2.14
Struktur kimia umum anthosianin (anthosianidin) (Montilla dkk., 2010).
Anthocyanins (Anthocyanidin)

2.12.3 Farmakokinetik anthosianin
Efek farmakologis anthosianin setelah dikonsumsi sangat bervariasi
tergantung pada bioavailabilitasnya. Bioavailabilitas anthosianin secara oral
cukup rendah karena agak sulit diserap pada saluran cerna, sedangkan beberapa
jenis senyawa polifenol yakni isoflavon dan asam fenolik lebih mudah diserap
(Han dkk., 2007). Anthosianin larut dalam air, sulit larut dalam lemak,
penyerapannya di saluran cerna mencapai 13-22% (Dreiseitel, 2011). Konsentrasi
maksimal pada plasma dicapai 2 jam setelah konsumsi, menurun setelah 4 jam,
dan mencapai kadar terendah setelah 16 jam konsumsi (SutirtaYasa dan Jawi,
2014). Sekitar 68% anthosianin yang diabsorpsi, dimetabolisme secara cepat dan
sebagian besar diekskresi melalui feces dan urin (Shipp dan Abdoel, 2010).
Pemberian anthosianin secara oral, sebelum diabsorpsi anthosianin
mengalami deglikosilasi oleh enzim lactase phloridzin hydrolase (LPH) di lumen
usus atau enzim cytosolic ß-glukosidase(CBG) pada enterosit. Setelah bentuk
aglikon diabsorpsi, kemudian dikonyugasi secara methilasi, sulphatisasi, atau
glukoronidasi. Bentuk aglikon dan konyugasinya masuk ke dalam sirkulasi
sistemik dan dapat melewati sawar darah otak (SDO) (Jager dan Saaby, 2011;
Viteva, 2014).
Banyak faktor dapat memengaruhi penyerapan anthosianin yaitu pH usus,
mikroflora usus, adanya makanan, dan bentuk sediaan (Han dkk., 2007).
Penelitian in vitro menunjukkan bahwa penyerapan anthosianin terjadi paling baik
pada jejenum, dan sedikit di duodenum, hampir tidak diabsorpsi di ileum dan

kolon. Bioavilabilitas cukup rendah akibat metabolismenya cepat dan diekskresi
dalam bentuk terkonyugasi glucuronidated, methylated, dan glycosylated melalui
urin dan feces. Metabolisme anthosianin diketahui juga terjadi di hati, yakni
proses metilasi yang dibantu enzim catechol-O-methyltransferase (COMT),
menghasilkan bentuk metilasi anthosianin, diekskresi lewat empedu, seperti
tampak pada Gambar 2.15 (Dreiseitel, 2011).
Konsentrasi anthosianin utuh yang dideteksi di dalam jaringan otak setelah
pemberian secara oral cukup rendah di bawah konsentrasi yang mempunyai efek
farmakologis dan neuroprotektif bermakna yang dibuktikan in vitro. Metabolit
anthosianin dipercaya mempunyai efek farmakologis nyata in vivo dan
bertanggung jawab pada tingginya bioaktivitas, walaupun bioavailabilitasnya
rendah (Montilla dkk., 2010). Metabolit anthosianin lebih stabil daripada
anthosianin utuh, sehingga menjadi gugus yang bioaktif di jaringan target,
termasuk bentuk sulfat, methylated, glucuronidated, dan glycosylated (Bhullar
dan Rupasinghe, 2013).

Gambar 2.15 Mekanisme penyerapan anthosianin (dikutip tanpa modifikasi dari Dreiseitel,
2011).
Otak dilindungi oleh sawar darah otak (SDO) dengan integritas yang
sangat baik sehingga sangat sulit dilewati oleh senyawa-senyawa dari luar otak.
Tingkat kemampuan anthosianin (flavonoid) melewati SDO tergantung pada
tingkat lipofilisitasnya. Metabolit derivat O-methylated mempunyai kemampuan
lebih besar melewati SDO dan masuk ke dalam otak dibandingkan metabolit
derivat glukuronid, tetapi dilaporkan juga bahwa bentuk glukuronid tertentu dapat
melewati SDO dengan mekanisme spesifik in vivo (Jager dan Saaby, 2011).
Kemampuan memasuki otak juga dimodulasi oleh interaksinya dengan specific
efflux transporters yang diekspresi di dalam SDO seperti P-glikoprotein (Viteva,
2014).

2.12.4 Efek farmakologis anthosianin
Beberapa efek farmakologis anthosianin pada susunan saraf pusat terutama
otak sangat bermanfaat dan memegang peranan penting melawan beberapa
penyakit degeneratif kronis seperti penyakit Parkinson, Alzheimer, stroke, dan
epilepsi (Ogino dan Wang, 2007). Efek farmakologis terpenting anthosianin
adalah: 1) antioksidan; 2) antiinflamasi; dan 3) antikejang (Han dkk., 2007;
Montilla dkk., 2010).
2.12.4.1 Peranan anthosianin mengatasi stres oksidatif
Produksi ROS dan RNS yang berlebihan melampaui kemampuan
antioksidan tubuh menimbulkan keadaan stres oksidatif (Han dkk., 2007; Ross,
2012). Keberadaan ROS dan RNS menginisiasi serta merangsang kematian
neuron melalui kerusakan oksidatif terhadap DNA, protein, dan lipid, diikuti oleh
inflamasi dan keadaan ini akan mempermudah terjadinya kejang berulang. Stres
oksidatif merupakan mekanisme yang menonjol pada perkembangan dan
progresivitas epilepsi dan beberapa penyakit neurodegeneratif lainnya. Neuron
dan glia di otak sangat rentan terhadap kerusakan oksidatif terutama otak anak
yang sedang berkembang (Farooqui, 2012).
Aktivitas antioksidan anthosianin tergantung pada struktur dan substituen
dari cincin heterocyclic dan cincin B, khususnya keberadaan struktur O-di-OH
pada cincin B, 2-3 double bond dalam konyugasi dengan fungsi 4-oxo dan
tambahan adanya grup 3- dan 5-OH pada cincin heterocyclic (Montilla dkk.,
2010). Konfigurasi dan jumlah grup hidroksil anthosianin secara substansial

memengaruhi mekanisme aktivitas antioksidan. Konfigurasi hidroksil cincin B
paling signifikan menentukan aktivitas ROS scavenging, sedangkan substitusi
cincin A dan C mempunyai sedikit pengaruh pada efek superoxide anion
scavenging (Farooqui, 2012; Ross, 2012).
Aktivitas antioksidan anthosianin terjadi secara langsung maupun secara
tidak langsung. Secara umum mekanisme kerja anthosianin sebagai antioksidan
adalah: 1) mencegah pembentukan radikal bebas dengan cara menghambat enzim
xanthine oksidase dan khelasi metal transisi; 2) menghambat radikal bebas
merusak sel target (mendonorkan elektron dan penangkap radikal bebas); 3)
menghambat propagasi reaksi oksidatif (chain breaking antioxidant); 4)
menguatkan (reinforce) kapasitas antioksidan sel (sparing effect) dan
menginduksi ekspresi antioksidan endogen (Han dkk., 2007; Ross, 2012). Efek
antioksidan anthosianin jauh lebih kuat dibandingkan antioksidan klasik seperti
butil hidroksi anilin (BHA), butil hidroksi toluen (BHT), vitamin E, cathecin, dan
quercetin (Shipp dan Abdoel, 2010).
Efek antioksidan anthosianin meningkatkan antioksidan endogen melalui
aktivasi jalur Nrf2 dan meningkatkan ikatan Nrf2 dengan ARE pada inti sel
(Parooqui, 2012). Peningkatan transkripsi Nrf2 dapat meningkatkan ekspresi
enzim-enzim detoksifikasi seperti Glutathion S-Transferase A2 (GSTA2),
NADPH: Quinone Oxidoreductase 1(NQO-1), Hem Oxigenase-1 (HO-1), γ-
Glutamylcysteine synthetase (γGCS), dan Glutamate Cysteine Ligase (GCL)
sehingga terjadi peningkatan antioksidan endogen terutama SOD, dan antioksidan
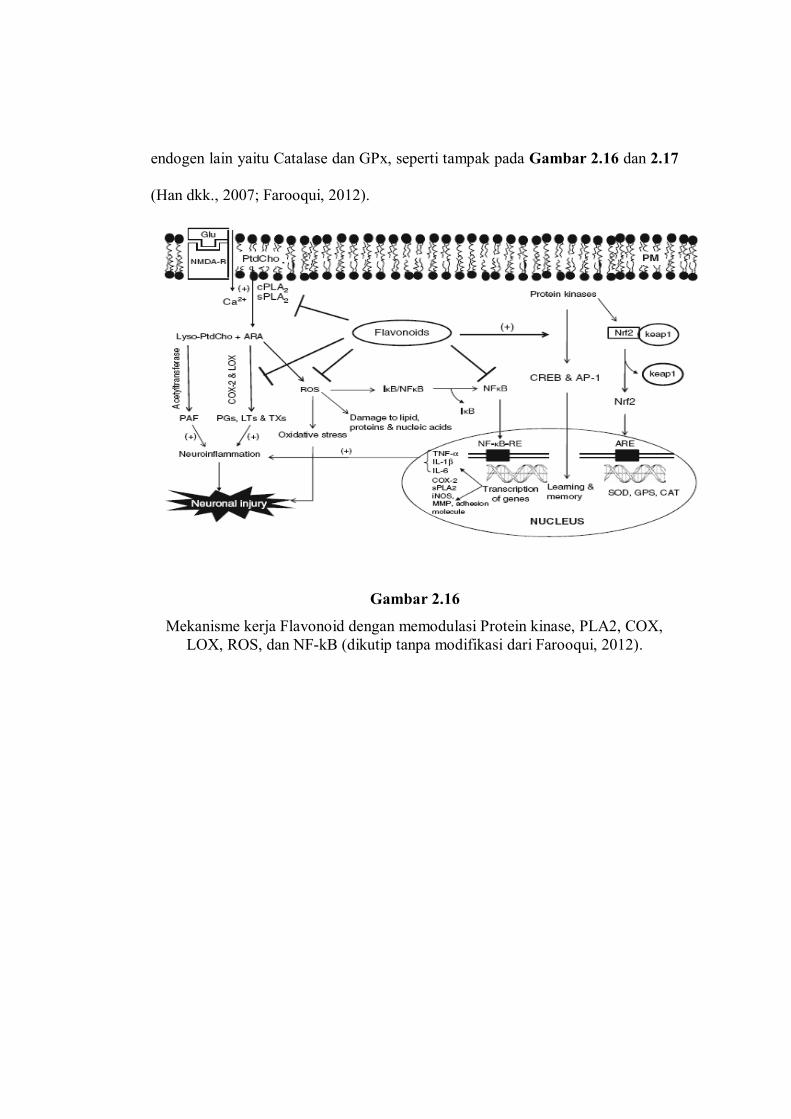
endogen lain yaitu Catalase dan GPx, seperti tampak pada Gambar 2.16 dan 2.17
(Han dkk., 2007; Farooqui, 2012).
Gambar 2.16
Mekanisme kerja Flavonoid dengan memodulasi Protein kinase, PLA2, COX, LOX, ROS, dan NF-kB (dikutip tanpa modifikasi dari Farooqui, 2012).
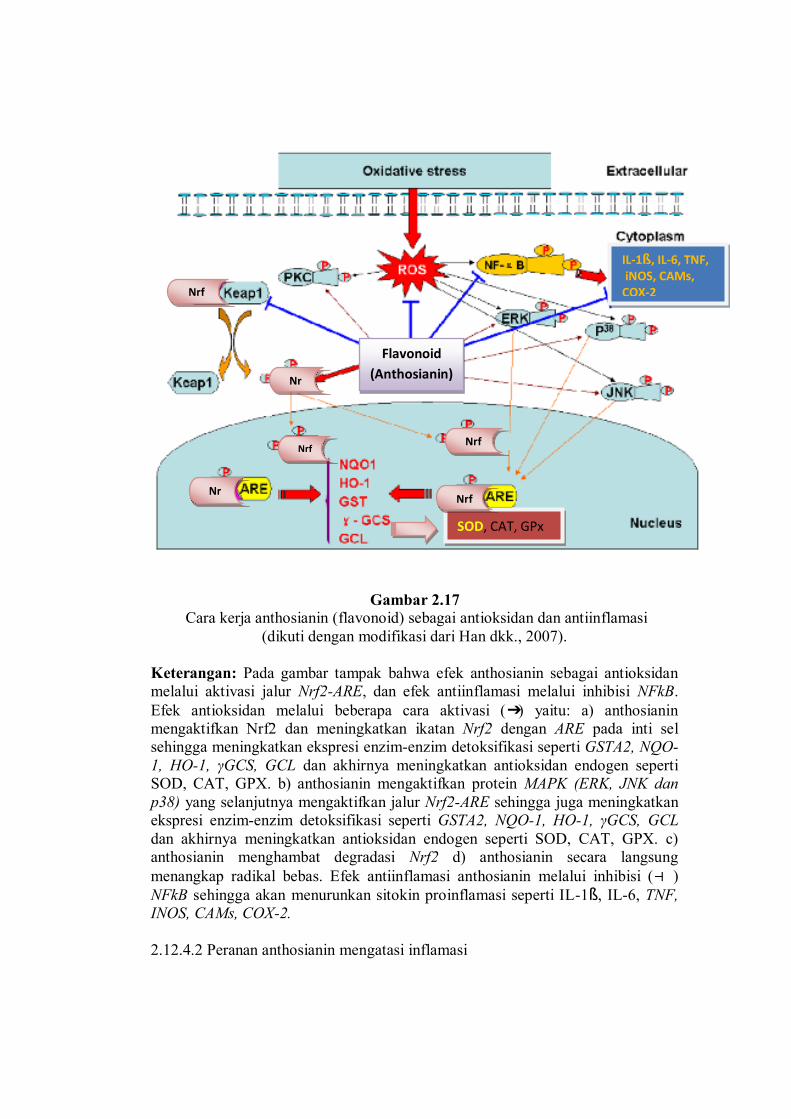
Gambar 2.17 Cara kerja anthosianin (flavonoid) sebagai antioksidan dan antiinflamasi
(dikuti dengan modifikasi dari Han dkk., 2007).
Keterangan: Pada gambar tampak bahwa efek anthosianin sebagai antioksidan melalui aktivasi jalur Nrf2-ARE, dan efek antiinflamasi melalui inhibisi NFkB. Efek antioksidan melalui beberapa cara aktivasi (➔) yaitu: a) anthosianin mengaktifkan Nrf2 dan meningkatkan ikatan Nrf2 dengan ARE pada inti sel sehingga meningkatkan ekspresi enzim-enzim detoksifikasi seperti GSTA2, NQO-1, HO-1, γGCS, GCL dan akhirnya meningkatkan antioksidan endogen seperti SOD, CAT, GPX. b) anthosianin mengaktifkan protein MAPK (ERK, JNK dan p38) yang selanjutnya mengaktifkan jalur Nrf2-ARE sehingga juga meningkatkan ekspresi enzim-enzim detoksifikasi seperti GSTA2, NQO-1, HO-1, γGCS, GCL dan akhirnya meningkatkan antioksidan endogen seperti SOD, CAT, GPX. c) anthosianin menghambat degradasi Nrf2 d) anthosianin secara langsung menangkap radikal bebas. Efek antiinflamasi anthosianin melalui inhibisi (⊣ ) NFkB sehingga akan menurunkan sitokin proinflamasi seperti IL-1ß, IL-6, TNF, INOS, CAMs, COX-2. 2.12.4.2 Peranan anthosianin mengatasi inflamasi
Flavonoid
(Anthosianin)
n)
SOD, CAT, GPx
IL-1ß, IL-6, TNF, iNOS, CAMs, COX-2
Nrf
2
Nrf
2
Nr
f2
Nrf
22
Nr
f2
Nrf
2

Efek antiinflamasi anthosianin secara in vitro terutama melalui inhibisi
sinyal NF-kB dan menekan ekspresi marker proinflamasi seperti IL-1ß, IL-6 dan
TNF-α.
Kapasitas anthosianin menghambat marker proinflamasi tergantung pada
gambaran struktur spesifiknya. Substitusi 5,7 hidroksil pada cincin A dan 2,3
double bond serta
4-keto group pada cincin C merupakan susunan struktur yang sangat penting.
Sebaliknya, substitusi hidroksil dari cincin B dan C, tetapi bukan cincin A, adalah
esensial untuk aktivitas antioksidan (Montilla dkk., 2010).
Aktivasi mikroglia dan astrosit pada penyakit denegeratif di otak
diketahui dapat memproduksi nitric oxide dan merangsang ekspresi sitokin pro-
inflamasi. Proses ini melibatkan sinyal jalur mitogen-activated protein kinase
(MAPK) dan kaskade sinyal NF-kB. Mitogen-activated protein kinase terdiri dari
Extracelluler signal-regulated kinase (ERK 1/2), c-Jun N-terminal kinase (JNK
1/2/3), dan p38 kinase (p38abcd), yang berhubungan dengan transfer signal dari
luar ke dalam sel dan respon sel. Selama proses trandusi, aktivasi kinase
phosphorylate dan cytosolic transcription factors (seperti STAT-1/2/3, NF-kB,
CREB), menyebabkan terjadinya nuclear translocation yang pada akhirnya
menghasilkan ekspresi gen untuk pembentukan sitokin proinflamasi (Parooqui,
2012), seperti tampak pada Gambar 2.16 dan 2.17.
Anthosianin menunjukkan efek antiinflamasi yang nyata, melalui
penekanan aktivasi NFkB, yang bertanggung jawab pada ekspresi beberapa gen
proinflamasi (Han dkk., 2007), menghambat MAPKs, JNK, dan ERK, yang

bertanggungjawab pada ekspresi sejumlah sitokin proinflamasi dan juga
menghambat cyclooxigenase-2 (COX-2),yang merupakan enzim proinflamasi
penting (Han dkk., 2007; Ross, 2012).
Eksperimen pada sel mikroglia Murine BV2 menunjukkan bahwa
anthosianin secara nyata menghambat mediator proinflamasi yang diinduksi LPS
seperti nitric oxide (NO), dan prostaglandin E2, serta sitokin proinflamasi seperti
TNF-α, IL-6 dan IL-1ß, tanpa efek toksis (Ross, 2012).
2.12.4.3 Peranan anthosianin sebagai antikejang
Glutamat dan GABA secara kuantitatif merupakan neurotransmiter
eksitasi dan neurotransmiter inhibisi terpenting pada otak mamalia.
Ketidakseimbangan sistem neurotransmiter ini sangat berhubungan dengan
epilepsi. Stimulasi berlebihan glutamat dan NMDA reseptor serta defisiensi
GABA pada neuron menyebabkan kejang sehingga dua neurotransmiter ini
menjadi target penting obat antiepilepsi (Katzung, 2011).
GABA reseptor distribusinya luas pada otak mamalia, dan merupakan
target untuk obat antiepilepsi. GABA-A reseptor merupakan heteromeric GABA-
gated chloride channel dan saluran transmembran ion dibuka oleh stimulus yang
ditimbulkan oleh GABA, serta diikuti influx ion klorida, dan menghasilkan
penurunan efek depolarisasi dan mendepresi eksitabilitas neuron. GABA-A
reseptor mengandung tempat ikatan (binding sites) untuk senyawa-senyawa yang
secara allosterik memodifikasi saluran klorida seperti benzodiazepin dan

barbiturat, yang dapat menimbulkan efek sedatif dan antikonvulsan (Parooqui,
2012).
GABA merupakan neurotransmiter inhibisi utama yang berperan pada
epilepsi, dan diperankan melalui ikatan dengan reseptor GABA. Reseptor GABA-A
ada di post sinap, permeabel terhadap ion klorida. Saluran ion membran terbuka
karena stimulasi oleh GABA, diikuti aktivasi klorida influx untuk menimbulkan
penurunan efek depolarisasi (efek kejang) dan terjadi keadaan hiperpolarisasi
sehingga menghambat potensial aksi serta akhirnya menekan aktivitas kejang
(Jager dan Saaby, 2011).
Bukti eksperimental jelas menunjukkan bahwa anthosianin memiliki
struktur yang menyerupai benzodiazepin sehingga dapat terikat pada binding site
reseptor GABA sehingga dapat menimbulkan efek antikonvulsan, seperti tampak
pada Gambar 2.18 (Katzung, 2011).
Gambar 2.18 Mekanisme aksi benzodiazepin dan ikatan anthosianin (flavonoid) pada
benzodiazepin binding site (dikutip dengan modifikasi dari Katzung, 2011).
Anthosianin (FLavonoid)

Pada reseptor GABA, disamping tempat ikatan untuk neurotransmitter
(neurotransmitter binding site), ada juga tempat ikatan modulasi (modulatory
binding site). Anthosianin dapat terikat pada kedua tempat ikatan tersebut,
menimbulkan modulasi reseptor GABA untuk lebih sensitif terhadap GABA
sehingga meningkatkan efek inhibisi kejang. Sebuah penelitian melaporkan
bahwa flavone ororylin yang diisolasi dari Lamiaceae di China, menunjukkan
efek antikejang in vivo, dengan memodulasi reseptor GABA spesifik (Jager dan
Saaby, 2011).
2.12.5 Asupan dan toksisitas anthosianin
Anthosianin (E 163), merupakan makanan tambahan yang resmi diakui di
Eropa dan telah dievaluasi oleh SCF 1975 dan JECFA pada tahun 1982. Asupan
harian (Acceptive daily intake) anthosianin 2,5 mg/kgbb/hari (Aguilar dkk., 2013).
Penelitian pada babi dan anjing, pemberian oral anthosianin ekstrak kulit
anggur sebanyak 3 gram/kg dan pada tikus 6 gram/hari, tidak ada efek samping.
Pemberian anthosianin ekstrak kulit anggur pada diet sukarelawan dengan dosis
1780 mg/kgbb/hari pada laki-laki dan 2150 mg/kgbb/hari pada perempuan selama
90 hari, tidak menimbulkan efek samping atau efek toksis (Aguilar dkk., 2013).
Asupan anthosianin pada diet rutin sangat rendah, rata-rata 0.1
mg/kgbb/hari pada dewasa dan 0,3 mg/kgbb/hari pada anak-anak. Rekomendasi
EFSA (European Food Safety Authoritry), konsumsi anthosianin disesuaikan
dengan umur seperti tampak pada Tabel 2.1.

Tabel 2.1
Asupan regular anthosianin yang dianjurkan (mg/kgbb/hari)(Aguilar dkk., 2013)
__________________________________________________________________
__
Toddlers anak-anak remaja dewasa orang tua
(12-35 bln) (3-9 th) (10-17 th) (18-64 th) (>65 th)
____________________________________________________________________ Rerata 1,5-4,0 1,5-4,7 1,0-2,5 0,7-1,9 0,5-1,1
Level tinggi 3,2-6,9 2,7-7,8 1,6-3,9 1,1-3,4 0,9-2,3 ___________________________________________________________________ Toksisitas pemberian anthosianin oral telah diteliti pada tikus dan kelinci.
Anthosianin dari ekstraks currant, blueberries dan elderberries, nilai LD50
ditentukan 25.000 mg/kgbb pada tikus, dan tidak ditemukan efek genotoksisitas,
toksisitas kronis maupun karsinogenik (Aguilar dkk., 2013), juga tidak ditemukan
toksisitas terhadap reproduksi dan pertumbuhan-perkembangan anak (Jeong dkk.,
2013).
Indikasi bahwa anthosianin ekstrak currant, blueberries atau elderberries
menginduksi genotoksis, toksisitas pertumbuhan-perkembangan, dan
karsinogenesis pada tikus atau rabbit dengan dosis tinggi 9 gram/kgbb/hari tidak
ditemukan (Aguilar dkk., 2013). Pemberian ekstrak air umbi ubi jalar ungu dosis
tunggal 4 ml, nyata meningkatkan kadar anthosianin dengan profil farmakokinetik
baik pada tikus sehat dan tikus yang diinduksi menjadi diabetes (Sutirta-Yasa dan
Jawi, 2014).

Jadi sudah dibuktikan bahwa anthosianin ekstrak air umbi ubi jalar ungu
yang merupakan salah satu bentuk flavonoid, mempunyai efek antioksidan kuat
dengan menghambat stres oksidatif melalui proses mendonorkan elektron,
menangkap radikal bebas, memutus rantai reaksi oksidatif, dan meningkatkan
ekspresi gen antioksidan endogen melalui aktivasi jalur Nrf2-ARE. Anthosianin
juga mempunyai efek antiinflamasi melalui penghambatan NF-kB dan COX-2;
dan efek antikejang melalui modulasi efek neurotransmiter GABA pada
reseptornya.
Berdasarkan hal tersebut di atas, pemberian ekstrak air umbi ubi jalar ungu
diasumsikan bahwa dapat menurunkan biomarker stres oksidatif seperti 8-OHdG,
meningkatkan antioksidan endogen SOD dan menurunkan biomarker inflamasi
seperti IL-6. Stres oksidatif dan inflamasi berperan penting pada patogenesis dan
progresivitas ERO,sehingga dengan menghambat stres oksidatif dan proses
inflamasi, meningkatkan antioksidan endogen tubuh serta menekan aktivitas
kejang pada neuron melalui modulasi GABA, secara rasional akan terjadi
perbaikan gambaran EEG dan menurunkan frekuensi kejang pada anak dengan
EFRO, yang dibuktikan pada penelitian ini.




















