







Bahasa
Halaman
Hukum


i
The role of Type VI Secretion System in the
virulence of Klebsiella pneumoniae
Thesis Submitted for the degree of
Doctor of Philosophy
at the University of Leicester
By
David Mwin (B.Sc.)
Department of Infection, Immunity, and Inflammation
College of Medicine, Biological Sciences and Psychology
University of Leicester
October 2017

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
i
Statement of originality
The research work submitted in this thesis for the degree of Doctor of Philosophy PhD entitled “The role
of Type VI secretion system in the virulence of Klebsiella pneumoniae” is based on work conducted by the
author in the Department of Infection, Immunity, and Inflammation (College of Medicine and Biological
Science) of the University of Leicester. during the period between February 2012 and August 2015.
All the work recorded in this thesis is original unless otherwise acknowledged in the text or by references.
No part of this work is submitted elsewhere for another degree in this or any other University.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
ii
Acknowledgement
To God Almighty, I say thank you for Your Wisdom, Protection, and the Gift of life upon me throughout
my studies and beyond. I am highly indebted to my family for their support, patience, and care during my
studies. Thanks to the Government of Ghana for their generous sponsorship and provision of funds for
this research work.
I am very grateful to my Supervisors, Dr Yassine Amrani, Dr Shaun Heaphy, and Dr Kumar Rajakumar,
for all their support and guidance during my studies. I am also very grateful for the support and
constructive criticisms of Prof. Jose Bengoechea and Prof. Peter Andrew. To Dr Hasan Yelsilkaya and
Dr Primerose Fresestone, I am immensely grateful for your mentorship.
The team from the Biotechnology Core was beneficial during confocal and Electron microscopy, and I am
very thankful for their assistance. I am very grateful to Dr Codula Stover and Dr Simon Bath for the
provision of J744 and Acanthamoeba castellani, respectively.
To my loving family whose undivided support and encouragement gave me the wings to saw higher
during and beyond this journey, thank you for being such a pillar in my life. And finally, to all who in one
way or the other supported me in completion of this work, I would like to say God bless you all.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
iii
Dedication
I dedicated this research work/ thesis to my; beautiful family (Mrs Ugomma Lauritta Mwin and our
children), amazing mother (Marcelina Ngmenterebo) and late father, Godfred Ngmenterebo, whose
support and memory inspire me for greater heights.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
iv
Abstract
The role of Type VI secretion system in the virulence of Klebsiella pneumoniae
David Mwin
The enterobacteria, especially the multidrug-resistant strains, pose a serious health threat with evolving
virulence mechanisms against their host. The opportunistic pathogen, K. pneumoniae, is a leading cause of
urinary tract infection, blood and pneumonia in patients at the hospital and remains a common isolate in
community-acquired disease. Many bacteria, including K. pneumoniae, explore a range of factors
/mechanisms such as secreted toxin and antibiotic resistance that may enhance their survival, virulence, and
evasion of the host immune system. Type VI Secretion Systems (T6SSs) is a transmembrane “spring-
loaded” toxin-translocating nanomolecular machinery recently characterized in over 25% of Proteobacteria.
T6SS shares both structural and protein homology with phage tail and is considered to be reminiscent of
the bacteriophage puncturing device. While available data suggest a diverse role of the toxin puncturing
T6SS device in many Gram-negative bacteria, no experimental data have demonstrated the putative T6SS
gene clusters’ role in the virulence of K. pneumoniae as at the time that this research work was conducted.
In silico analysis, using a full range of bioinformatic tools were used to identify and map the T6SS gene
cluster (T6SS1 and T6SS3) in K. pneumoniae. A library of molecular genetic tools was constructed via a
novel strategy and used to disarm the resistance in MDR K. pneumoniae, which enhanced the safety and
genetic manipulation of the strain for T6SS functional studies. Mainly, three T6SS mutants, ∆T1 (T6SS1
mutant), ∆T3 (T6SS3 mutant), and ∆T1∆T3 (T6SS1/T6SS3 mutant) generated via lambda red
recombination allelic exchange and were examined for the role of the T6SS gene clusters in K. pneumoniae.
The data obtained from the various assays and analysis suggest that K. pneumoniae via T6SS mediated
antibacterial virulence against other competing bacteria and played a significant role T4SS-mediated
conjugal transfer of mobile genetic elements. K. pneumoniae used T6SS to resist amoeba phagocytosis and
enhance subsequent survival within the cell. Also, T6SS increased in vivo virulence in Galleria larvae, host
cell invasion, survival, actin filament polymerisation and activation of host pro-inflammatory innate
immunity. Thus, the putative T6SS gene clusters may be mediating a multipurpose virulence against host
organism and other bacteria using pre-emptive contact-dependent strikes and toxin secretion.
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
v
Abbreviations
ABC ATP-binding cassette Mpf Mating pair formation
ATP Adenosine triphosphate MGE Mobile genetic element
ApE A plasmid editor MCS Multiple Cloning Site
Blast Basic Local Alignment Tool SOE-PCR Spliced overlap extension-PCR
bp Base pair EDTA Ethylenediaminetetraacetic acid
CFU Colony-forming unit NEB New England Bio lab
OD Optical Density CRKP Carbapenem-resistant Klebsiella pneumoniae
DNA Deoxyribose nucleic acid Pfam Protein families
dNTP Deoxy-ribonucleotide triphosphate PAI Pathogenicity island
dsDNA Double-stranded DNA PCR Polymerase Chain Reaction
DF Downstream flank PBS Phosphate buffered saline
ddH2O Deionised distilled water RF Right Flank
ESBL extended-spectrum beta-lactamases RNA Ribose nucleic acid
FRT Flippase Recognition Target ST Sequence type
g Gram DR Direct repeat
PO Phenoloxidase SOE Splice overlap extension
GC Guanine and Cytosine T1SS Type I secretion system
GI Genomic island T2SS Type II secretion system
HGT Horizontal gene transfer T3SS Type III secretion system
ICE Integrative and conjugative element T4SS Type IV secretion system
int Integrase T5SS Type V secretion system
IR Inverted repeat T6SS Type VI secretion system
IPTG Isopropyl-β-D-thiogalactopyranoside tRNA Transfer RNA
Kb Kilobase λ Lambda
Kp Klebsiella pneumoniae ⁰ C Degree Celsius
μl Microliter TATOXB Tannic acid-treated oxblood
LA Luria Bertani agar μg Microgram
LAS Luria Bertani agar + sucrose M Micro Molar
LB Luria Bertani broth cDNA Complementary DNA
LB Luria Bertani broth + sucrose CDS Coding domain Sequence
Mpf Mating pair formation DF Conserved downstream flan
GP-RBS Guinea pig red blood cells DMSO Dimethyl sulfoxide
NCBI National Centre for Biotechnology
Information
SDS-PAGE Sodium dodecyl sulphate polyacrylamide gel
electrophoresis
kDa Kilodalton TAE Tris-acetate-EDTA
SOC Super-optimal broth with Catabolite
Repression
qRT-PCR Quantitative real-time polymerase chain
reaction
mM Millimolar pmol Picomole
v/v Volume per volume ratio w/v Weight per volume ratio

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
vi
Table of Contents
Statement of originality .................................................................................................................................. i
Acknowledgement ........................................................................................................................................ ii
Dedication .................................................................................................................................................... iii
Abstract ........................................................................................................................................................ iv
Abbreviations ................................................................................................................................................ v
List of Figures .............................................................................................................................................. xi
List of Tables ............................................................................................................................................. xiv
Publications ................................................................................................................................................. xv
Chapter 1: Introduction ................................................................................................................................. 2
1.1 Klebsiella Species ............................................................................................................................... 2
1.1.1 History and taxonomic structure .................................................................................................. 2
1.1.2 Ecology and Host Range .............................................................................................................. 3
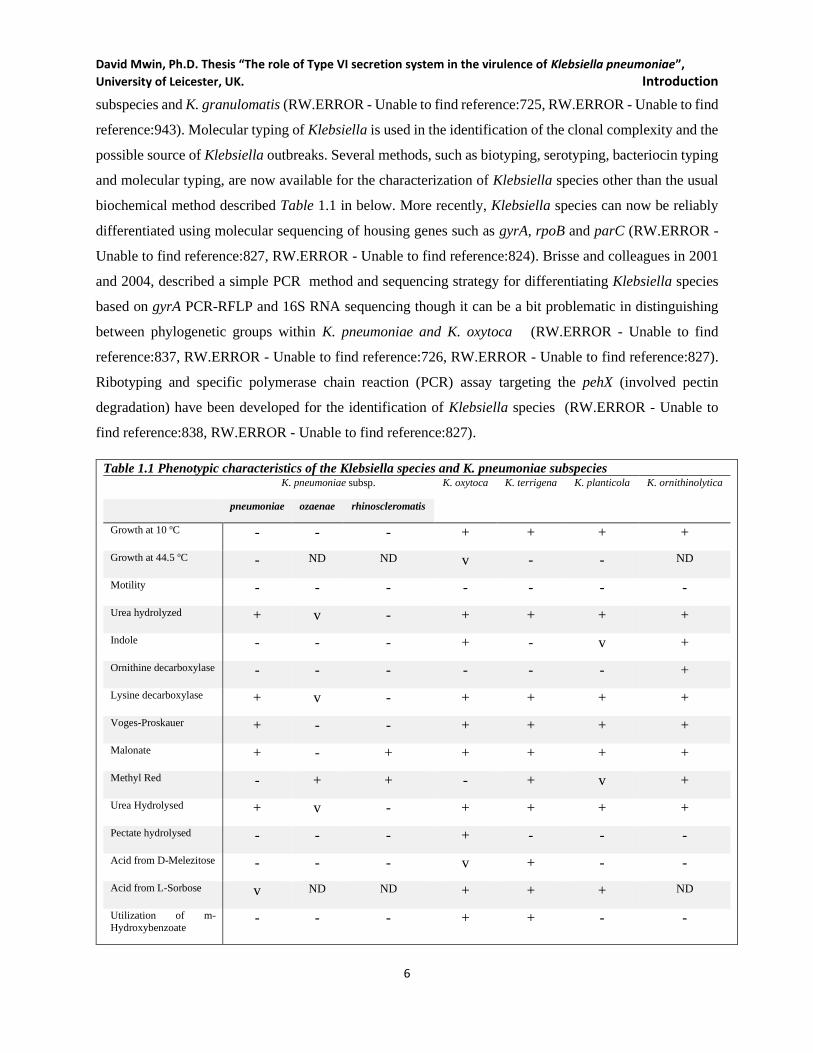
1.1.3 Isolation and Identification .......................................................................................................... 5
1.1.4 Epidemiology ............................................................................................................................... 7
1.1.5 K. pneumoniae ........................................................................................................................... 11
1.1.6 Virulence factors ........................................................................................................................ 11
1.2 Antibiotic resistance and virulence association ................................................................................ 20
1.3 Secretion systems .............................................................................................................................. 25
1.3.1 The Type I Secretion System (T1SS) ........................................................................................ 27
1.3.2 The Type II Secretion System (T2SS) ....................................................................................... 28
1.3.3 Type III secretion system (T3SS) .............................................................................................. 29
1.3.4 Type IV secretion system (T4SS) .............................................................................................. 30
1.3.5 Type V secretion system (T5SS) ............................................................................................... 32
1.3.6 Type VI secretion system (T6SS) .............................................................................................. 34
1.4 Research aims and objectives ........................................................................................................... 47
Chapter 2: Material and Methods................................................................................................................ 51

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
vii
2.1 Bacterial strains, Eukaryotic cells and Plasmids ............................................................................... 51
2.1.1 Media, reagents and solutions .................................................................................................... 57
2.2 DNA-related methods and technique ................................................................................................ 59
2.2.1 Oligonucleotide design, In silico-PCR and synthesis ................................................................ 59
2.2.2 Polymerase chain reaction (PCR) .............................................................................................. 59
2.2.3 Colony PCR ............................................................................................................................... 60
2.2.4 Splice Overlap Extension (SOE)-PCR: Mutant allele construction ........................................... 60
2.2.5 Genomic and plasmid extraction ................................................................................................ 62
2.2.6 Gel electrophoresis, DNA purification and Sequencing ............................................................ 63
2.2.7 Restriction digestion .................................................................................................................. 63
2.2.8 DNA dephosphorylation and ligation ........................................................................................ 63
2.3 RNA-related techniques and methods ............................................................................................... 63
2.3.1 RNA extraction and cDNA library preparation ......................................................................... 63
2.3.2 Transcriptional analysis of T6SS, T4SS and fimbrial gene clusters .......................................... 64
2.3.3 qRT-PCR analysis of T6SS, T4SS, fim and mrk fimbrial clusters. ........................................... 64
2.4 Construction of pDNTOOL plasmids ............................................................................................... 64
2.4.1 Construction of suicide deletion vectors .................................................................................... 65
2.4.2 Variety of Lambda RED plasmids with different antibiotic resistance cassette ........................ 68
2.4.3 FRT-based antibiotic cassettes/ GFP plasmids .......................................................................... 70
2.4.4 Flp recombinase- encoding plasmid construction ...................................................................... 74
2.4.5 Cloning and inducible expression plasmids/ GFP plasmid Construction .................................. 76
2.5 Genetic manipulation ........................................................................................................................ 80
2.5.1 Preparation of Competent bacterial cells ................................................................................... 80
2.5.2 Suicide vector-based allelic exchange ....................................................................................... 81
2.5.3 Lambda RED recombination-based allelic exchange for KPHST6SS mutation ....................... 82
2.5.4 Flp-mediated FRT recombination .............................................................................................. 86
2.5.5 Sucrose counterselection ............................................................................................................ 87

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
viii
2.6 Other methods ................................................................................................................................... 87
2.6.1 Biofilm formation ...................................................................................................................... 87
2.6.2 Growth and cell viability ............................................................................................................ 88
2.6.3 Antibiotic susceptibility testing ................................................................................................. 89
2.6.4 Co-culture experiments .............................................................................................................. 90
2.6.5 Bacterial competition and plasmid mobilization ........................................................................ 90
2.6.6 Agglutination assay .................................................................................................................... 93
2.6.7 G. mellonella larvae In vivo virulence model ............................................................................ 94
2.6.8 Eukaryotic cell-based virulence assays and methods ................................................................. 96
2.6.9 Transmission Electron Microscopy ......................................................................................... 105
2.6.10 Data analysis and Statistical Analysis .................................................................................... 105
2.6.11 Bioinformatic Analysis .......................................................................................................... 105
Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of K. pneumoniae
.................................................................................................................................................................. 110
3.1 Constructed molecular tools and strategic genetic manipulation .................................................... 110
3.1.1 Novel DNA swapping strategy; constructing FRT-flanked antibiotic resistance marker ........ 110
3.1.2 Other constructed genetic tools ................................................................................................ 116
3.2 K. pneumoniae HS11286 mobile genome analysis and disarmament antibiotic resistance ............ 118
3.2.1 The mobile genome of K. pneumoniae HS11286 and analysis ............................................... 118
3.2.2 Resistance genes and the broader contexts of the linked regions ............................................ 120
3.2.3 Genetic disarmament of the resistance of K. pneumoniae HS11286 ....................................... 121
3.3 Disarmament of antibiotic resistance in K. pneumoniae HS11286 enhanced in vitro fitness not in
vivo virulence in G. mellonella. ............................................................................................................ 123
3.4 3.6 K. pneumoniae HSS11286 T6SS clusters are syntenic with similarity in flanking regions to
other K. pneumoniae strains ................................................................................................................. 126
Chapter 4: K. pneumoniae T6SS enhance bacteria-bacteria interaction and biofilm-like formation ........ 142
4.1 Overview ................................................................................................................................... 142

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
ix
4.2 Deletion of T6SS gene clusters (T6SS1 and/ or T6SS3) did not affect the growth of the mutants
relative to the parent strain .................................................................................................................... 144
4.3 T6SS hallmark genes hcp1, hcp2, vgrG1 and vgrG3 were expressed in the K. pneumoniae
HS11286 strain...................................................................................................................................... 145
4.4 Deletion of the K. pneumoniae DN2 T6SS1 and/ or T6SS3 locus results in reduced 'intra-strain'
fitness 146
4.5 K. pneumoniae DN2 inter-species fitness is dependent on which T6SS loci are being deleted 149
4.6 Absence of T6SS1 and T6SS3 enhanced plasmid DNA mobilization via conjugation ............ 152
4.7 T6SS deletion enhanced biofilm formation in K. pneumoniae. ................................................ 153
4.8 Deletion of T6SS1 and T6SS3 clusters enhanced type 1 and type 3 fimbriae expression
respectively ........................................................................................................................................... 157
4.9 K. pneumoniae T6SSs mediate resistance to Acanthamoeba castellanii, complement-mediated
killing and PMN internalisation ............................................................................................................ 161
Chapter 5: K. pneumoniae T6SSs Mediate Virulence and Activation Host Cell Innate Immune Response
.................................................................................................................................................................. 167
5.1 T6SSs in K. pneumoniae promote Galleria killing and in vivo survival of bacteria ...................... 167
5.2 K. pneumoniae T6SSs enhance phagocytic uptake and survival in murine macrophage ............... 173
5.3 K. pneumoniae T6SSs promote attachment, invasion and actin polymerisation of host lungs
epithelial cells ....................................................................................................................................... 176
5.4 T6SSs in K. pneumoniae trigger an inflammatory immune response in both macrophages and
epithelial cell (J774 and A549) ............................................................................................................. 186
5.4.1 Expressed macrophage (J774) inflammatory mediators/ cytokines induced in response to K.
pneumoniae infection ........................................................................................................................ 187
5.4.2 Expressed epithelial (A549) inflammatory mediators/ cytokines stimulated in response to K.
pneumoniae ....................................................................................................................................... 193
5.5 J774 inflammatory mediators induced by hyper mucoid K. pneumoniae NTUH-K2044 T6SS ..... 196
Chapter 6: General Discussion .................................................................................................................. 200
Chapter 7: Conclusion ............................................................................................................................... 220
7.1 General conclusion .......................................................................................................................... 223

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
x
7.2 Future work ..................................................................................................................................... 224
Appendix 1: Primer Table ......................................................................................................................... 282
Appendix 2 ................................................................................................................................................ 289
Appendix 2-1: List of genes deleted from the 26 kb region ................................................................. 289
Appendix 2-2 Antibiotic susceptibility of K. pneumoniae HS11286 and the antibiotic disarmed versions
.............................................................................................................................................................. 290
Appendix 2-3 Resistance genes identified in K. pneumoniae HS11286 that could confer resistance in
Clinical cases ........................................................................................................................................ 291
Appendix 2-4- Distribution of mobile genetic elements in K. pneumoniae HS11286 ......................... 292
Appendix 2-5 Replicons of the K. pneumoniae plasmids ..................................................................... 292
Appendix 2-6 Other GIs identified in K. pneumoniae HS11286 genome ............................................ 292
Appendix 2-7 T6SS gene cluster in K. pneumoniae HS11286 and NTUH-K2044 .............................. 292

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
xi
List of Figures
Figure 1.1 Schematic of phylogeny within the Klebsiella genus .................................................................. 3
Figure 1.2 Factors associated with nosocomial bacterial pneumonia. ...................................................... 10
Figure 1.3 Virulence factors associated with K. pneumoniae. ................................................................... 12
Figure 1.4 Protein secretion systems in Gram-Negative bacteria. ............................................................. 27
Figure 1.5 T4SS Functional representation in most Gram-negative bacteria, including K. pneumoniae. 31
Figure 1.6 Core T6SS gene organisation and Structural resemblance to phage-tail. ................................ 35
Figure 1.7 The assembled structural components of T6SS. ........................................................................ 37
Figure 1.8 Dynamic formations and dissociation T6SS in bacteria. .......................................................... 38
Figure 1.9 T6SS genealogy reported in some commonly encountered bacteria. ........................................ 40
Figure 1.10 T6SS functional versatility. ..................................................................................................... 42
Figure 1.11 Identified T6SS gene clusters in some Klebsiella species. ...................................................... 46
Figure 2.1 Schematic of SOE-PCR for mutant allele creation or joining DNA fragment. ......................... 62
Figure 2.2 Construction of K. pneumoniae NUTH-K2044 T6SS1 deletion vector, pJTOOL-8. ................. 66
Figure 2.3 Construction of K. pneumoniae NUTH-K2044 T6SS3 deletion vector, pDNTOOL-9. ............. 67
Figure 2.4 Construction of pDNTOOL-13 and pDNTOOL-16. .................................................................. 69
Figure 2.5 Construction of pDNTOOL-31 and pDNTOOL-1. .................................................................... 72
Figure 2.6 Construction of pDNTOOL-19 and pDNTOOL-2. .................................................................... 75
Figure 2.7 Construction of pDNTOOL-22 and pDNTOOL-3. .................................................................... 77
Figure 2.8 Construction of pDNTOOL-29 and pDNTOOL-30. .................................................................. 79
Figure 2.9 K. pneumoniae HS11286 T6SS1 mutant allele and knockout. .................................................. 83
Figure 2.10 K. pneumoniae HS11286 T6SS3 mutant allele creation and knockout. .................................. 85
Figure 3.1 Schematic representation of novel in vivo plasmid manipulation. .......................................... 112
Figure 3.2 Construction and verification of pDNTOOL-31. .................................................................... 113
Figure 3.3 Colony PCR to confirm the presences or absence of a target gene, lambda Red and
constructed plasmids. ........................................................................................................................ 114
Figure 3.4 Confirmation of constructed plasmid carrying FRT-Antibiotic resistance marker-FRT. ....... 115
Figure 3.5 The contexts of resistance genes in K. pneumoniae HS11286 (a) pKPHS2 and (b) pKPHS3
plasmids. ........................................................................................................................................... 121
Figure 3.6 Deletion of blaKPC-2 on pKPHS2 and 26 kb MDR region on pKPHS3 in K. pneumoniae
HS11286 background. ....................................................................................................................... 122

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
xii
Figure 3.7 Deletion of the K. pneumoniae HS11286 antibiotic resistance arsenal enhances competitive
fitness but not virulence in Galleria larvae. ..................................................................................... 125
Figure 3.8 T6SS gene clusters identified in K. pneumoniae HS21186. .................................................... 129
Figure 3.9 Arrangement of T6SS1, T6SS2 and T6SS3 gene clusters using genome viewer. .................... 134
Figure 3.10 Schematic procedure for K. pneumoniae HS11286 T6SS1 deletion. .................................... 136
Figure 3.11 Schematic procedure for K. pneumoniae HS11286 T6SS3 deletion. .................................... 137
Figure 3.12 T6SS mutant generation in DN2 (resistance disarmed K. pneumoniae HS11286) background
and T6SS mutant PCR confirmation. ................................................................................................ 139
Figure 4.1 Growth bacteria strains after T6SS gene cluster knockout. .................................................... 145
Figure 4.2 T6SS core genes, hcp1, hcp2, vgrG1 and vgrG3 are expressed in K. pneumoniae. ............... 146
Figure 4.3 K. pneumoniae intra-strain in vitro competition. .................................................................... 148
Figure 4.4 Inter-species in vitro competition. ........................................................................................... 149
Figure 4.5 E. coli DH5α colony reduction due to K. pneumoniae T6SS activity. ..................................... 151
Figure 4.6 K. pneumoniae NTUH-K2044 T6SS1-dependent inhibition of E. coli DH5α. ........................ 152
Figure 4.7 Plasmid mobilization and expression T4SS –related genes.. .................................................. 155
Figure 4.8 Biofilm formation and visualisation. ....................................................................................... 156
Figure 4.9 Expression of mrkD and fimH of K. pneumoniae surface fimbriae. ....................................... 158
Figure 4.10 Transmission electron microscopic (TEM) visualization of K. pneumoniae surface fimbriae.
.......................................................................................................................................................... 160
Figure 4.11 T6SS enhances K. pneumoniae against Acanthamoeba castellanii. ..................................... 162
Figure 4.12 K. pneumoniae T6SS-enhanced resistance to Serum mediated Killing, C3b deposition and
opsonisation.. .................................................................................................................................... 164
Figure 5.1 K. pneumoniae T6SS dependent killing of Galleria larvae. .................................................... 168
Figure 5.2 In vivo competition: DN2 vs T6SS mutants in Galleria larvae. .............................................. 169
Figure 5.3 Hypermucoid K. pneumoniae T6SS enhanced killing of Galleria larvae. .............................. 170
Figure 5.4 Phenoloxidase activity (PO) in infected Galleria larvae. ....................................................... 171
Figure 5.5 Parentage viability cell count in larvae. ................................................................................. 172
Figure 5.6 J7774.A1 phagocytosis uptake and visualization of K. pneumoniae. ..................................... 175
Figure 5.7 K. pneumoniae phagocytic survival in murine macrophage (J774.A1) is T6SS enhanced. ... 176
Figure 5.8 K. pneumoniae T6SS-enhanced attachment and survival in lungs epithelial cells. ................ 177
Figure 5.9 Visualization K. pneumoniae attachment to H292 cells. ........................................................ 180
Figure 5.10 K. pneumoniae T6SS toxicity in H292 cells .......................................................................... 182
Figure 5.11 K. pneumoniae T6SSs mediated A549 invasion and actin polymerisation after 3hrs. .......... 184
Figure 5.12 K. pneumoniae A549 invasion and actin polymerisation after 6hrs. .................................... 185

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
xiii
Figure 5.13 A preliminary detection of cytokines array/ proteome profiler of K. pneumoniae-infected
J774 cell supernatant at 3h post-infection. ....................................................................................... 188
Figure 5.14 ELISA quantification of K. pneumoniae T6SS-dependent induction of inflammatory mediators
in infected J774 supernatant. ............................................................................................................ 192
Figure 5.15 K. pneumoniae T6SS induced secretion of lungs epithelial A549 inflammatory mediators in
the supernatant at 3 post-infection. ................................................................................................... 195
Figure 5.16 Inflammatory mediators induced by hyper mucoid K. pneumoniae NTUH-K2044. ............. 197
Figure 6.1 Schematic representation of K. pneumoniae T6SS used for bacteria competition involved in
DN2 interaction. ............................................................................................................................... 208
Figure 6.2 K. pneumoniae T6SS induces type 1 and 3 fimbria-associated biofilm form. ......................... 211
Figure 6.3 Multipurpose K. pneumoniae T6SS directly interact with target cells in a contact-dependent
manner. ............................................................................................................................................. 217

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
xiv
List of Tables
Table 1.1 Phenotypic characteristics of the Klebsiella species and K. pneumoniae subspecies ________ 6
Table 1.2 Features of bacterial secretion system ___________________________________________ 26
Table 1.3.0 T6SS nomenclatural components and functions __________________________________ 39
Table 1.4.0 Functional diversity of T6SS toward the eukaryotic host. ___________________________ 43
Table 2.1 List of plasmids used in this study _______________________________________________ 52
Table 2.2 List of bacterial strains and Eukaryotic lines used in this study________________________ 55
Table 2.3 Typical PCR cycling conditions ________________________________________________ 60
Table 2.4 Bioinformatic resources used in this study _______________________________________ 105
Table 3.1 K. pneumoniae HS11286 T6SS1 gene cluster: context and content ____________________ 127
Table 3.2 K. pneumoniae HS11286 T6SS3 gene cluster: context and content ____________________ 130
Table 4.1 Agglutination Assay ________________________________________________________ 157

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
xv
Publications
1. BI, D., JIANG, X., SHENG, Z.K., NGMENTEREBO, D., TAI, C., WANG, M., DENG, Z.,
RAJAKUMAR, K. and OU, H.Y., 2015. Mapping the resistance-associated mobilome of a
carbapenem-resistant Klebsiella pneumoniae strain reveals insights into factors shaping these
regions and facilitates the generation of a 'resistance-disarmed' model organism. The Journal of
antimicrobial chemotherapy

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK.
xvi

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
1
CHAPTER ONE
INTRODUCTION

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
2
Chapter 1: Introduction
1.1 Klebsiella Species
In 1882, Austrian microbiologist called Anton von Frisch made the first account of Klebsiella as an
encapsulated bacillus isolated from a chronic rhinoscleroma patient with chronic granulomatous infection
of the upper airway. This bacillus was named in honour of a German Microbiologist, Edwin Klebs, in 1887
for discovering Corynebacterium diphtheria (RW.ERROR - Unable to find reference:758, RW.ERROR -
Unable to find reference:912). Modern microbiology defines Klebsiella as a Gram-negative bacillus that
forms mucoid colonies and non-motile (RW.ERROR - Unable to find reference:873). In the following
sections, the history, taxonomy, ecology and host range, isolation and identification, and epidemiology of
Klebsiella species are described. The next discussion will be the virulence factors of Klebsiella pneumoniae,
including Type 6 secretion systems, which are the focus of this research. Also included in this chapter is
the host immune response to K. pneumoniae, and more importantly, the overall aims and objective of this
research. The Klebsiella species described in the most detail is a clinical isolate K. pneumoniae.
1.1.1 History and taxonomic structure
The genus Klebsiella consists of Gram-negative, non-motile, encapsulated, lactose-fermenting,
facultatively anaerobic, rod-shaped bacteria. The genus Klebsiella belongs to the Enterobacteriaceae
family, named by Trevisan (1885) in honour of Edwin Kleb (RW.ERROR - Unable to find reference:763).
K. pneumoniae subspecies pneumoniae strain C122 is the strain-type for Klebsiella genus and was first
isolated from urinary tract infection (RW.ERROR - Unable to find reference:751, RW.ERROR - Unable to
find reference:749, RW.ERROR - Unable to find reference:757). The capsulated K. rhinoschleromatis,
named by Trevisan (1887) was the first Klebsiella species to be isolated from a rhinoscleroma patient
(RW.ERROR - Unable to find reference:758). Abel (1893) isolated K. ozaenae formally called “Bacillus
mucosus” from the nasal secretion of a patient with ozaena (RW.ERROR - Unable to find reference:761).
In 1982 Friedlander isolated from a pneumonia patient an organism he called “Hyalococcus pneumoniae”
(RW.ERROR - Unable to find reference:762) and K. pneumoniae (RW.ERROR - Unable to find
reference:764). There was considerable confusion about this genus (Klebsiella) for several years as the
organism at the time could not be objectively distinguished from Escherichia (described as “Bacterium
lactis aerogenes” by Escherich, in 1885). However, it was renamed as “Bacillus aerogenes” by Kruse in
1896, to “Aerobacter aerogenes” by Kruse in 1896, and later to Enterobacter aerogenes by Hormaeche and
Edwards in 1960 (RW.ERROR - Unable to find reference:766, RW.ERROR - Unable to find reference:767,
RW.ERROR - Unable to find reference:765).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
3
The widely accepted taxonomic structure of Klebsiella includes K. pneumoniae (K. pneumoniae subsp.
pneumoniae, K. pneumoniae subsp. ozaenae and K. pneumoniae subsp. rhinoscleromatis); K. oxytoca; K.
planticola, K. ornithinolytica, K. terrigena, and K. variicola, as indicated in Figure 1.1. Error! Reference
source not found. (RW.ERROR - Unable to find reference:757, RW.ERROR - Unable to find
reference:768). These Klebsiella species are categorized into three broad clusters; the K. pneumoniae cluster
one (K. pneumoniae subsp. pneumoniae, K. pneumoniae subsp. ozaenae and K. pneumoniae subsp.
rhinoscleramtis), K. oxytoca as cluster three while the other species (K. planticola, K. ornithinolytica, K.
terrigena, and K. variicola) utilize L-sorbose as carbon source and can grow at 10⁰C, classified as cluster
two. K. pneumoniae, K. ozaenae, K. rhinoscleromatis and K. oxytoca are currently the most relevant
clinical species (RW.ERROR - Unable to find reference:757).
Figure 1.1 Schematic of phylogeny within the Klebsiella genus
A phylogenic tree depicting Klebsiella classification Adapted from (RW.ERROR - Unable to find reference:915,
RW.ERROR - Unable to find reference:824, RW.ERROR - Unable to find reference:826, RW.ERROR - Unable to
find reference:757, van Aartsen, 2012)
1.1.2 Ecology and Host Range
Klebsiella is ubiquitously found in diverse environmental habitats such as soil, water and vegetation
(RW.ERROR - Unable to find reference:769, RW.ERROR - Unable to find reference:770, RW.ERROR -

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
4
Unable to find reference:771, RW.ERROR - Unable to find reference:772). It contributes to the biochemical
and geochemical processes of the ecosystem and acting as microflora in nonclinical settings (RW.ERROR
- Unable to find reference:773, RW.ERROR - Unable to find reference:774, RW.ERROR - Unable to find
reference:775, RW.ERROR - Unable to find reference:777, RW.ERROR - Unable to find reference:778).
These bacteria play an essential role in the nitrogen fixation as several Klebsiella species such as K.
pneumoniae, K. oxytoca or K. planticola have been isolated from the plant root surface and nodules
(RW.ERROR - Unable to find reference:779, RW.ERROR - Unable to find reference:780, RW.ERROR -
Unable to find reference:782, RW.ERROR - Unable to find reference:783). The interaction of human and
animals with their immediate environment enhances bacteria colonization to certain parts of the body (gut,
nasopharynx and mucosal membranes) as microflora. Though few environmental Klebsiella strains
occasionally appear in clinical settings, most of them are mostly not as virulent as clinical Klebsiella
(RW.ERROR - Unable to find reference:784, RW.ERROR - Unable to find reference:785).
Klebsiella can exist in a wide range of mammal and insect host. K. oxytica is frequently isolated from
insects, such as cockroaches among others which may act as vectors for the spread of Klebsiella in hospital
settings. The bacteria play a role in the synthesis of pheromone aggregation in insects (RW.ERROR -
Unable to find reference:795, RW.ERROR - Unable to find reference:796, RW.ERROR - Unable to find
reference:797). Klebsiella has been either isolated or implicated in the several animal host diseases, such
metritis, infertility and pyothorax pus accumulation in horses, cattle (bovine mastitis and osteomyelitis
originating from pulmonary lesions), snakes, crocodiles, dogs, Rhesus monkey, guinea pigs, birds, muskrats
and squirrel (RW.ERROR - Unable to find reference:1299). In humans, carriage in the gastrointestinal tract
is a major K. pneumoniae reservoir albeit less frequently isolated from the nasopharynx (RW.ERROR -
Unable to find reference:1361). Human host interactions with Klebsiella ranges from the asymptomatic
carriage in healthy individuals to opportunistic infection in immune-compromised individuals and more
recently in community-acquired diseases (RW.ERROR - Unable to find reference:805, RW.ERROR -
Unable to find reference:806, RW.ERROR - Unable to find reference:812, RW.ERROR - Unable to find
reference:810, RW.ERROR - Unable to find reference:813, RW.ERROR - Unable to find reference:808,
RW.ERROR - Unable to find reference:1361). More recently, Klebsiella is frequently isolated as the
etiologic agent of community-acquired pneumonia (RW.ERROR - Unable to find reference:813,
RW.ERROR - Unable to find reference:814, RW.ERROR - Unable to find reference:815, RW.ERROR -
Unable to find reference:816, RW.ERROR - Unable to find reference:817, RW.ERROR - Unable to find
reference:818).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
5
1.1.3 Isolation and Identification
Klebsiella belongs to the Enterobacteriaceae family and can be further subdivided into a range of species,
including K. granulomatis, K. mobilis, K. ornithinolytica, K. oxytoca, K. planticola, K. pneumoniae, K.
singaporensis, K. terrigena, K. trevisanii and K. variicola. The bacterium K. pneumoniae can be further
subdivided into K. pneumoniae subsp. pneumoniae, K. pneumoniae subsp. ozaenae and K. pneumoniae
subsp. Rhinoscleromatis. K. pneumoniae is closely related to several other genera within the
Enterobacteriaceae family, such as Citrobacter, Escherichia, Enterobacter and Salmonella (RW.ERROR -
Unable to find reference:941, RW.ERROR - Unable to find reference:784, RW.ERROR - Unable to find
reference:942). Being ubiquitous organisms, Klebsiella species grow well on varieties of media such
nutrient agar, tryptic casein soy agar, bro-mocresol purple lactose agar, blood agar, as well as more
differential plating media for Enterobacteriaceae, such as Drigalski agar, MacConkey agar, eosin-
methylene blue agar (EMB), and bromothymol blue agar. K. pneumoniae and K. oxytoca are both lactose
positive with dome-shaped colonies (3-4mm) when grown overnight at 30°C or 37°C and can be mucoid.
K. planticola and K. terrigena are equally lactose positive and dome-shaped colonies but with 1.5- 2.5mm
diameter and weakly mucoid. Almost all Klebsiella strains grow well in minimal medium with ammonium
ions or nitrate as sole nitrogen source and a carbon source without growth factor requirement. Klebsiella
strains can be conserved at room temperature in meat extract semisolid agar, or at –80°C in a broth medium
with 10–50% (v/v) glycerol, or freeze-dried (RW.ERROR - Unable to find reference:821, RW.ERROR -
Unable to find reference:820). Before the use invention of molecular methods, Klebsiella detection was
based on a combination of biochemical tests and microphysical features.
Klebsiella species can be identified based on their differential carbon source utilization alongside other
biochemical tests (RW.ERROR - Unable to find reference:823, RW.ERROR - Unable to find reference:820,
RW.ERROR - Unable to find reference:832, RW.ERROR - Unable to find reference:833). Generally, eight
carbon sources may be sufficient to for the Klebsiella species identification, and about 18 biochemical tests
are often enough for the identification of all Klebsiella species but not K. variicola (RW.ERROR - Unable
to find reference:834, RW.ERROR - Unable to find reference:835). K. pneumoniae is urease and citrate
positive, ferments glucose and lactose, unable to produce indole and lacks ornithine decarboxylase and
motility (RW.ERROR - Unable to find reference:757, RW.ERROR - Unable to find reference:822). Apart
from the inability of the K. pneumoniae subspecies to grow at 10°C or utilise L-sorbose as the sole carbon
source in combination with other biochemical methods (Table 1.1), carbon assimilation, 16S rDNA and β-
subunit of RNA polymerase B (rpoB) sequencing was earlier reported for the taxonomic confirmation of
the genus Klebsiella (RW.ERROR - Unable to find reference:824). In the sequence analysis of 16s rDNA
and rpoB, 98.2% to 99.7% and 99.4% to 100% similarity, respectively, between the three K. pneumoniae
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
6
subspecies and K. granulomatis (RW.ERROR - Unable to find reference:725, RW.ERROR - Unable to find
reference:943). Molecular typing of Klebsiella is used in the identification of the clonal complexity and the
possible source of Klebsiella outbreaks. Several methods, such as biotyping, serotyping, bacteriocin typing
and molecular typing, are now available for the characterization of Klebsiella species other than the usual
biochemical method described Table 1.1 in below. More recently, Klebsiella species can now be reliably
differentiated using molecular sequencing of housing genes such as gyrA, rpoB and parC (RW.ERROR -
Unable to find reference:827, RW.ERROR - Unable to find reference:824). Brisse and colleagues in 2001
and 2004, described a simple PCR method and sequencing strategy for differentiating Klebsiella species
based on gyrA PCR-RFLP and 16S RNA sequencing though it can be a bit problematic in distinguishing
between phylogenetic groups within K. pneumoniae and K. oxytoca (RW.ERROR - Unable to find
reference:837, RW.ERROR - Unable to find reference:726, RW.ERROR - Unable to find reference:827).
Ribotyping and specific polymerase chain reaction (PCR) assay targeting the pehX (involved pectin
degradation) have been developed for the identification of Klebsiella species (RW.ERROR - Unable to
find reference:838, RW.ERROR - Unable to find reference:827).
Table 1.1 Phenotypic characteristics of the Klebsiella species and K. pneumoniae subspecies K. pneumoniae subsp. K. oxytoca K. terrigena K. planticola K. ornithinolytica
pneumoniae ozaenae rhinoscleromatis
Growth at 10 oC - - - + + + +
Growth at 44.5 oC - ND ND v - - ND
Motility - - - - - - -
Urea hydrolyzed + v - + + + +
Indole - - - + - v +
Ornithine decarboxylase - - - - - - +
Lysine decarboxylase + v - + + + +
Voges-Proskauer + - - + + + +
Malonate + - + + + + +
Methyl Red - + + - + v +
Urea Hydrolysed + v - + + + +
Pectate hydrolysed - - - + - - -
Acid from D-Melezitose - - - v + - -
Acid from L-Sorbose v ND ND + + + ND
Utilization of m-
Hydroxybenzoate - - - + + - -
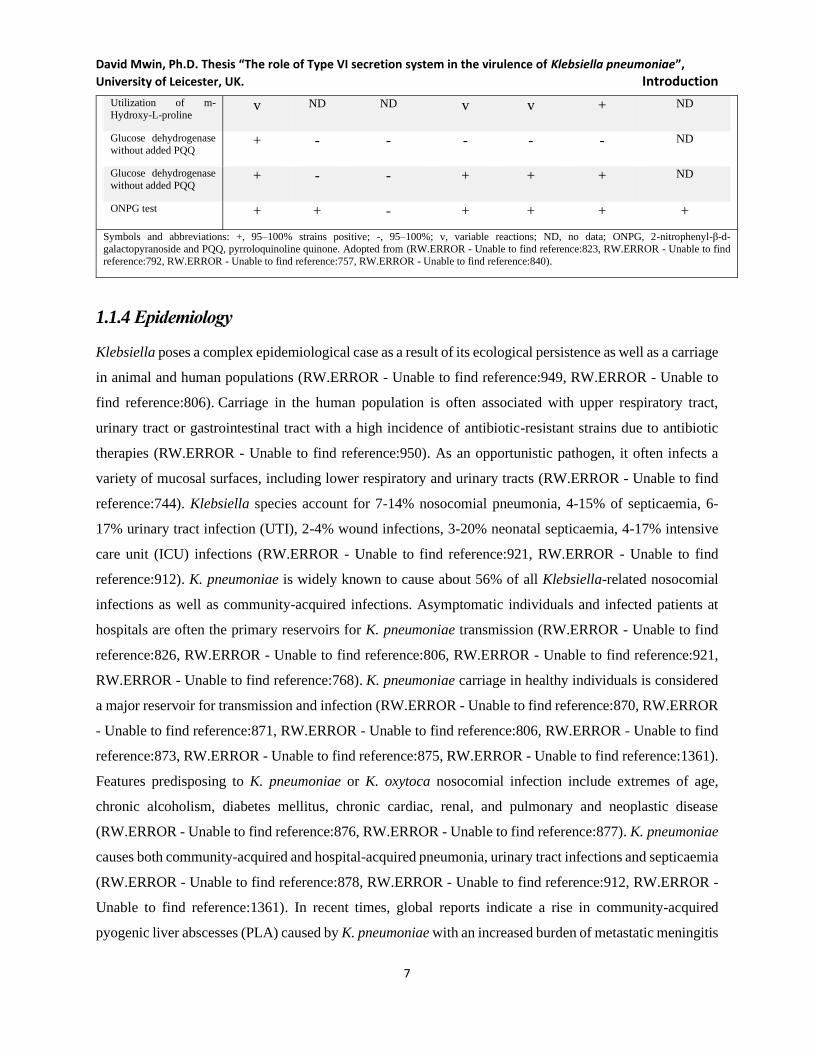
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
7
Utilization of m-
Hydroxy-L-proline v ND ND v v + ND
Glucose dehydrogenase
without added PQQ + - - - - - ND
Glucose dehydrogenase
without added PQQ + - - + + + ND
ONPG test + + - + + + +
Symbols and abbreviations: +, 95–100% strains positive; -, 95–100%; v, variable reactions; ND, no data; ONPG, 2-nitrophenyl-β-d-
galactopyranoside and PQQ, pyrroloquinoline quinone. Adopted from (RW.ERROR - Unable to find reference:823, RW.ERROR - Unable to find
reference:792, RW.ERROR - Unable to find reference:757, RW.ERROR - Unable to find reference:840).
1.1.4 Epidemiology
Klebsiella poses a complex epidemiological case as a result of its ecological persistence as well as a carriage
in animal and human populations (RW.ERROR - Unable to find reference:949, RW.ERROR - Unable to
find reference:806). Carriage in the human population is often associated with upper respiratory tract,
urinary tract or gastrointestinal tract with a high incidence of antibiotic-resistant strains due to antibiotic
therapies (RW.ERROR - Unable to find reference:950). As an opportunistic pathogen, it often infects a
variety of mucosal surfaces, including lower respiratory and urinary tracts (RW.ERROR - Unable to find
reference:744). Klebsiella species account for 7-14% nosocomial pneumonia, 4-15% of septicaemia, 6-
17% urinary tract infection (UTI), 2-4% wound infections, 3-20% neonatal septicaemia, 4-17% intensive
care unit (ICU) infections (RW.ERROR - Unable to find reference:921, RW.ERROR - Unable to find
reference:912). K. pneumoniae is widely known to cause about 56% of all Klebsiella-related nosocomial
infections as well as community-acquired infections. Asymptomatic individuals and infected patients at
hospitals are often the primary reservoirs for K. pneumoniae transmission (RW.ERROR - Unable to find
reference:826, RW.ERROR - Unable to find reference:806, RW.ERROR - Unable to find reference:921,
RW.ERROR - Unable to find reference:768). K. pneumoniae carriage in healthy individuals is considered
a major reservoir for transmission and infection (RW.ERROR - Unable to find reference:870, RW.ERROR
- Unable to find reference:871, RW.ERROR - Unable to find reference:806, RW.ERROR - Unable to find
reference:873, RW.ERROR - Unable to find reference:875, RW.ERROR - Unable to find reference:1361).
Features predisposing to K. pneumoniae or K. oxytoca nosocomial infection include extremes of age,
chronic alcoholism, diabetes mellitus, chronic cardiac, renal, and pulmonary and neoplastic disease
(RW.ERROR - Unable to find reference:876, RW.ERROR - Unable to find reference:877). K. pneumoniae
causes both community-acquired and hospital-acquired pneumonia, urinary tract infections and septicaemia
(RW.ERROR - Unable to find reference:878, RW.ERROR - Unable to find reference:912, RW.ERROR -
Unable to find reference:1361). In recent times, global reports indicate a rise in community-acquired
pyogenic liver abscesses (PLA) caused by K. pneumoniae with an increased burden of metastatic meningitis

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
8
and endophthalmitis (RW.ERROR - Unable to find reference:879, RW.ERROR - Unable to find
reference:880, RW.ERROR - Unable to find reference:857, RW.ERROR - Unable to find reference:882,
RW.ERROR - Unable to find reference:813, RW.ERROR - Unable to find reference:883, RW.ERROR -
Unable to find reference:856).
In the United States, urinary tract infections (UTIs) comprise 30% of nosocomial infections, followed by
pneumonia at 27%, and bloodstream infections at 19%. K. pneumoniae is responsible for about 4-8% of
these cases and continually ranked in the top 5 causative agents in nosocomial and community-acquired
diseases (RW.ERROR - Unable to find reference:884). In Taiwan and USA, K. pneumoniae is one of the
critical emerging pathogen responsible for about 78% and 41% community-acquired liver abscess,
respectively. It remains a problem in most Intensive Care Unit (ICU) acquired infections worldwide
(RW.ERROR - Unable to find reference:907, RW.ERROR - Unable to find reference:906, RW.ERROR -
Unable to find reference:905, RW.ERROR - Unable to find reference:883, RW.ERROR - Unable to find
reference:904, RW.ERROR - Unable to find reference:903). K. pneumoniae related infection have mainly
been problematic in the many Asia nations with an incidence of 1 to 21 per 1000 hospital admissions.
Global point-prevalence studies reported, 6.1% to 15% nosocomial infection rates, though nosocomial
infection in Asian countries range from 4% to 43% of which 45% to 65% are lower respiratory tract
infection (RW.ERROR - Unable to find reference:925). Mortality rate ranged from 25% to 54% has been
reported in the region (RW.ERROR - Unable to find reference:944, RW.ERROR - Unable to find
reference:945, RW.ERROR - Unable to find reference:946). The United States observed more cases of
nosocomial K. pneumoniae-related pneumonia than community pneumonia which accounts for only 1% of
the disease (RW.ERROR - Unable to find reference:806, RW.ERROR - Unable to find reference:951,
RW.ERROR - Unable to find reference:954, RW.ERROR - Unable to find reference:953). The bacteria is
the fifth most prevalent nosocomial bacterial pathogen in the United States, accounting for up to 6% of all
nosocomial bacterial disease (RW.ERROR - Unable to find reference:956).
Understanding nosocomial infection and the associated factors and mechanisms that contribute to the
persistence of K. pneumoniae in hospitals is very paramount in the fight against K. pneumoniae related
pneumonia. A matrix of environmental, bacterial and host-related factors (Figure 1.2) play a huge role the
incidence of K. pneumoniae associated infections (RW.ERROR - Unable to find reference:892,
RW.ERROR - Unable to find reference:923). Many patients, including ICU patients, tend to have low
immunity against most opportunistic pathogens such as K. pneumoniae, Pseudomonas aeruginosa,
Escherichia coli, and other bacteria, thereby making them vulnerable to disease-causing bacteria. These
poor host immune factors in combination to environmental factors predispose patients to K. pneumoniae
found on contaminated hospital floors, surgical tools, invasive devices, hands/gloves of staff, and

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
9
contaminated respiratory therapies (RW.ERROR - Unable to find reference:1364). Typical cases are
observed in patients under antibiotic therapies that often get infected with a deadly multi-drug resistant
(MDR) K. pneumoniae. Treatment of K. pneumoniae respiratory and urinary tract infections with extensive
use of broad-spectrum antibiotics in hospitals sometimes results in the emergence of drugs resistant strains
such as extended-spectrum β-lactamase (ESBL) and carbapenem-resistant strains (RW.ERROR - Unable
to find reference:935, RW.ERROR - Unable to find reference:947). This rapid and easy acquisition of
antimicrobial resistance along with the unmatched development of novel antimicrobials is posing a threat
to the last lines of antimicrobial therapies (RW.ERROR - Unable to find reference:931, RW.ERROR -
Unable to find reference:932, RW.ERROR - Unable to find reference:933, RW.ERROR - Unable to find
reference:934). Thus, the emergence of MDR K. pneumoniae strains is limiting therapeutic options and
contributing to the overall high mortality rates caused by K. pneumoniae (RW.ERROR - Unable to find
reference:928, RW.ERROR - Unable to find reference:938, Bi et al, 2015, RW.ERROR - Unable to find
reference:733, RW.ERROR - Unable to find reference:736). The growth and colonization of the bacterium
are enhanced by a matrix of environmental factors leading to the overwhelming population of the bacteria
in the defenceless host-lungs, mostly through aspiration (oropharynx and gut) or inhalation of contaminated
aerosols. Homogeneous dissemination of the bacteria may also play a role in bacteraemia or the
translocation bacteria from other infected organs to the lungs. The increased imbalance of the bacteria
population in the host lungs overwhelms the compromised host-lung immune systems, hence leading to
pneumonia caused by K. pneumoniae. Though other unknown factors may play a role pneumonia infection,
the below Error! Reference source not found. attempted to summarise the widely reported factors
involved in Klebsiella-related pneumonia.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
10
Figure 1.2 Factors associated with nosocomial bacterial pneumonia.
Factors contributing to the persistence of bacteria nosocomial pneumonia in hospitals (RW.ERROR - Unable to find
reference:892)

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
11
1.1.5 K. pneumoniae
For this research, K. pneumoniae is the organism of interest among the Klebsiella species. K. pneumoniae
has K and O-antigens that are of epidemiological and clinical importance (RW.ERROR - Unable to find
reference:858, RW.ERROR - Unable to find reference:918, RW.ERROR - Unable to find reference:919).
The bacterium has 78 serotypes for capsular polysaccharide (CPS) K-antigen types and 9 serotypes for
lipopolysaccharide (LPS) O-antigen types (RW.ERROR - Unable to find reference:806).
Classification of K. pneumoniae:
Bacteria
Proteobacteria
Gammaproteobacteria
Enterobaceriales
Enterobacteriaceae
K. pneumoniae
K. pneumoniae subsp. pneumoniae
The ubiquitous K. pneumoniae (named after the 19th-century German microbiologist, Edwin Kleb) belongs
to the family Enterobacteriaceae and currently remains the most frequent causative agent of both human
nosocomial and community-acquired infections. It is rod-shaped and measures 2 µm by 0.5 µm. In 1882,
Friedlander C. Uber first discovered Klebsiella to be a pathogen that caused pneumonia (RW.ERROR -
Unable to find reference:841). The bacterium is often found as a commensal resident of the human
gastrointestinal tract (RW.ERROR - Unable to find reference:843).
1.1.6 Virulence factors
In 1890, Robert Koch postulated guidelines for identifying disease-causing organisms. A century later,
Stanley Falkow established the molecular version of Koch’s postulate, thereby allowing the identification
of microbial genes involved in virulence. As part of investigating functional molecular mechanisms,
microbial genes are inactivated, and the appropriate virulence models are then used to determine the
measure of the virulence (RW.ERROR - Unable to find reference:948). K. pneumoniae, as an opportunistic
pathogen has evolved several mechanisms to evade and colonize the host or environment. In the human
host, one or more virulence factors associated with K. pneumoniae may play an essential role in the
pathogenicity of the bacterium. A broad genetic variability of K. pneumoniae plays a role in the dramatic

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
12
differences in the virulence factors and pathogenicity of the bacterium (RW.ERROR - Unable to find
reference:1017). These virulence factors include capsular polysaccharide (CPS), lipopolysaccharide (LPS)
and O-antigen side chains, antibiotic resistance, siderophores and receptors, adhesins/ fimbriae, and
secretion systems (RW.ERROR - Unable to find reference:744, RW.ERROR - Unable to find
reference:951, RW.ERROR - Unable to find reference:730, RW.ERROR - Unable to find reference:731,
RW.ERROR - Unable to find reference:738, RW.ERROR - Unable to find reference:737). indicates some
of the most commonly studied virulence factors (Capsule, Lipopolysaccharide LPS and O-antigens side
chain, siderophore and receptors, adhesins/ fimbriae/ pili, and Secretion system) in K. pneumoniae.
Figure 1.3 Virulence factors associated with K. pneumoniae.
The above figure is a schematic depiction of reported factors (Siderophores, LPS, CPS, secretion systems,
fimbriae and antibiotic resistance) known to mediate virulence in pathogenic K. pneumoniae (adapted from
(RW.ERROR - Unable to find reference:806)
1.1.6.1 capsular polysaccharide
Capsules from a range of pathogenic bacteria are key virulence determinants, so is the capsule of K.
pneumoniae that has been implicated in the virulence of the bacteria (RW.ERROR - Unable to find
reference:958, RW.ERROR - Unable to find reference:960, RW.ERROR - Unable to find reference:965).
As a widely reported virulence factor, K. pneumoniae capsular polysaccharide (CPS) is an acidic
polysaccharide (consisting of 3-6 repeating sugar units) that promote resistance to phagocytosis, serum
killing and in some cases contribute to biofilm through multi-path interaction with surface virulence
factors(RW.ERROR - Unable to find reference:962, RW.ERROR - Unable to find reference:961,
RW.ERROR - Unable to find reference:964). The CPS is synthesized by the Wzy-dependent

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
13
polymerization pathway, consisting of 21-30 kb containing 16-25 gene cluster which encodes the proteins
for the polymerization and assembly of CPS subunits. The 5´ terminal regions of most CPS-expressing gene
clusters are conserved, and consist of galF, orf2, wzi, wza, wzb and wzc while the 3´ end regions contain a
conserved gnd and mostly terminated at the ugd gene (RW.ERROR - Unable to find reference:974,
RW.ERROR - Unable to find reference:975).
CPS synthesis initiates the assembly of individual sugar-repeat units and catalysed by different
glycosyltransferases resulting in long chains of nascent sugar units that are transferred across the membrane
by flippase Wzx for polymerization by Wzy polymerase within the periplasmic space. Subsequent
polymerization and export of the CPS result in the deposition of the CPS on the bacterial surface as a result
of combined action of Wza (an inner membrane tyrosine autokinase), Wzb (a protein tyrosine phosphatase)
and Wzc (an integral outer membrane lipoprotein) (RW.ERROR - Unable to find reference:976,
RW.ERROR - Unable to find reference:977, Li et al, 2014). Capsular magA is a crucial virulent determinant
in K. pneumoniae K1-induced metastatic infections associated with capsular K1-specific Wzy polymerase
but has no role in the synthesis of lipopolysaccharide (LPS) (RW.ERROR - Unable to find reference:978,
RW.ERROR - Unable to find reference:979, RW.ERROR - Unable to find reference:980). Based structural
variability of capsular polysaccharide, Klebsiella characteristically produces 78 capsular (K-antigens)
serotypes covering the bacterial surface though they widely differ in their pathogenicity (RW.ERROR -
Unable to find reference:966, RW.ERROR - Unable to find reference:967, RW.ERROR - Unable to find
reference:968, Li et al, 2014). Epidemiological reports indicate that about 70% of all cases of Klebsiella
bacteremia are caused by 25 different serotypes (RW.ERROR - Unable to find reference:973). Among these
serotypes, K1 and K2 are mostly the predominant serotypes associated pathogenic K. pneumoniae. Several
K-antigen serotypes (including predominantly K1 and K2) have a unique hypermucoviscous
(hypervirulent) phenotype as a result of high production of capsule polysaccharide (CPS), grown
hypermucoviscous colonies of K. pneumoniae are easily identifiable on agar plates (RW.ERROR - Unable
to find reference:969, RW.ERROR - Unable to find reference:970, Li et al, 2014).
CPS is by far the most important virulence factor of K. pneumoniae that plays essential roles in resistance
to phagocytosis and serum killing, suppression of early inflammatory signals, resistance to antimicrobial
peptides and other environmental chemicals, inhibition of dendritic cell maturation and enhancement of
biofilm formation (RW.ERROR - Unable to find reference:964, RW.ERROR - Unable to find
reference:808, Sahly et al, 2004, Li et al, 2014, RW.ERROR - Unable to find reference:981, RW.ERROR
- Unable to find reference:982). The primary contribution of CPS to the virulence of Klebsiella appears to
serve as a protective surface layer enhancing the bacteria to evade host immune systems (RW.ERROR -
Unable to find reference:964, Li et al, 2014). K. pneumoniae capsule is an important virulence factor in the

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
14
UTIs and is reported to inhibit binding to epithelial cells (RW.ERROR - Unable to find reference:1016).
K1 serotype, unlike avirulent counterparts, has a unique capsular nature (extensive pyruvation of glucuronic
acid and acetylation of C2-OH or C3-OH of fucose) helps the bacterium to resist macrophages or escape
neutrophil-mediated intracellular killing while disseminating the bacteria to other host organs during
infection (RW.ERROR - Unable to find reference:985, RW.ERROR - Unable to find reference:984,
RW.ERROR - Unable to find reference:983). K. pneumoniae is considered an extracellular pathogen though
other reports showed that the bacterium could be internalised by epithelial cells (RW.ERROR - Unable to
find reference:992, RW.ERROR - Unable to find reference:993).
It has been demonstrated that K. pneumoniae CPS plays a role in the suppression of inflammatory signals
by inhibiting of IL-8 expression through the inhibition of Toll-Like Receptor 2 (TLR2) and TLR4, and
NOD1-dependent pathways (Li et al, 2014). CPS acts as a surface protective shield against the access of
host-derived antimicrobial peptides, environmental chemical agents, and free release forms of CPS can trap
antimicrobial polypeptides to reduce antimicrobial polypeptides reaching the bacterial surface
(RW.ERROR - Unable to find reference:986, RW.ERROR - Unable to find reference:987). Also, the sub-
lethal concentration of host antimicrobial peptides in airway induces the bacteria CPS production in the
bacterium to protect itself against host airway antimicrobial polypeptides (RW.ERROR - Unable to find
reference:987). K. pneumoniae CPS can impair dendritic cell maturation resulting in the reduction in
dendritic-mediated production of pro-Th1 cytokines (IL-12 and TNF-α). Thus, leading to the functional
abortion of dendritic maturation and reduction in the translocation of dendritic-mediated natural killer cells
during K. pneumoniae antigen presentation (RW.ERROR - Unable to find reference:989). Some K.
pneumoniae strains express a repeated sequence of dimannose or dirhamnose and are recognised by
macrophage receptors for binding, ingestion and digestion (RW.ERROR - Unable to find reference:990).
CPS also affects the expression of K. pneumoniae adhesins which mediate non-phagocytic cell-binding as
well as surface attachment structures (e.g. fimbriae/pili for biofilm formation and exchange of genetic
content) (RW.ERROR - Unable to find reference:964). The above overwhelming data demonstrate the
relevance of CPS in the pathogenicity of K. pneumoniae.
1.1.6.2 Lipopolysaccharide
Lipopolysaccharide (LPS) is one of the significant structural and immunodominant molecules of the outer
membrane (see Figure 1.3) and contains three parts: a highly variable O-antigen as the external part of LPS;
a highly conserved hydrophobic lipid A anchored in the outer membrane; and a core polysaccharide linking
O-antigen with lipid A (Li et al, 2014, RW.ERROR - Unable to find reference:964). At least nine type K.
pneumoniae O-antigens have been reported (O1, O2, O2ac, O3, O4, O5, O7, O8 and O12). Enzymes
encoded by six members wb gene cluster (wzm, wzt, wbbM, glf, wbbN and wbbO) are responsible for the

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
15
biosynthesis of O-antigen. The most prevalent O-antigen often isolated in clinical specimens is O1, which
are usually hypermucoviscous (invasive) strains, unlike the non-tissue invasive strain (Li et al, 2014).
The O-antigen of K. pneumoniae prevents complement components from reaching activators such, porins
and rough LPS, thereby enhancing bacterial resistance to complement-mediated killing (RW.ERROR -
Unable to find reference:994). In clinical settings, there is frequent isolation O1-serotype (known for their
serum resistance) than non-O1-serotype isolates (RW.ERROR - Unable to find reference:918). A strong
virulent role is reported for the O1 antigen of hypermucoviscous K. pneumoniae serotype O1:K1 through
host serum resistance and enhancement of bacterial dissemination and colonization in host organs
(RW.ERROR - Unable to find reference:918). A study has reported that CPS and LPS O- antigens of
hypermucoviscous K. pneumoniae O1: K2 in a murine pneumonia model enhanced bacteria passage in the
blood and sepsis development. CPS but not LPS O-antigen triggered the deposition of C3 on the bacterium
to protects it against human macrophage- phagocytosis in the alveolar (RW.ERROR - Unable to find
reference:995). The core polysaccharide of K. pneumoniae contains type 1 and type 2 core polysaccharides
which are synthesized by two different 13-gene wa gene cluster (RW.ERROR - Unable to find
reference:996, RW.ERROR - Unable to find reference:998, Li et al, 2014). Both wa gene clusters have
similar gene sets (hldD, waaF, waaC, waaL, waaQ, wabG, wabH, orf10, waaA, waaE and coaD). While
type 1 has additional wabI and wabJ which encode 3-deoxy-d-manno-octulosonic acid (Kdo) transferase
and heptosyltransferase, type 2 has wabK and wabM that are involved in the of transfer of the last two outer
core Glc residues (RW.ERROR - Unable to find reference:998).
Mutation of a gene within the K. pneumoniae LPS synthesis gene cluster (waaC, waaF, wabG and wabG)
results in a dramatic attenuation of virulence and urinary tract colonization (RW.ERROR - Unable to find
reference:999). Lipid A is synthesised by a set of constitutive enzymes through ABC transporter MsbA and
hanging onto the outer membrane. K. pneumoniae lipid A modifications are enzymatically catalysed upon
receiving environmental stimuli, resulting in virulence modulation in the pathogen, such change in K.
pneumoniae has attenuated virulence in animal models (RW.ERROR - Unable to find reference:1013,
RW.ERROR - Unable to find reference:1015). Lipid A and core polysaccharide independent of O-antigen
have been reported to be involved in phagocytic resistance against alveolar macrophage and to antibacterial
peptides, resulting in evasion of host immune defence (RW.ERROR - Unable to find reference:1014).
Pathogenic strains of K. pneumoniae serotypes O1:K1 and O1:K2 release an aggregate of an extracellular
toxic complex made of CPS, LPS, and protein that has significant tissue damages as a result of active K.
pneumoniae lobar pneumonia (RW.ERROR - Unable to find reference:999, Li et al, 2014).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
16
1.1.6.3 Adhesins/Fimbriae
K. pneumoniae encodes a vast arsenal of fimbrial and afimbrial adhesins. Fmbrial adhesin was first
described in E. coli in 1950. Pili and fimbriae are interchangeably used to describe nonflagellar
proteinaceous surface appendages on bacteria (RW.ERROR - Unable to find reference:1040, RW.ERROR
- Unable to find reference:1038, RW.ERROR - Unable to find reference:1039). The first study on K.
pneumoniae fimbriae was reported by Duguid in the late 1950s who also used haemagglutination properties
to distinguish between type 1 and type 3 fimbriae (RW.ERROR - Unable to find reference:1008,
RW.ERROR - Unable to find reference:1038). Type 1 fimbriae mediate mannose-sensitive
hemagglutination of guinea pig red blood cells, while type 3 fimbriae mediate mannose-resistant
agglutination of tannic acid-treated ox erythrocytes (RW.ERROR - Unable to find reference:1008,
RW.ERROR - Unable to find reference:1038). However, the advent of whole-genome sequencing has
advanced the distinction among large numbers of phenotypically uncharacterised putative fimbrial gene
clusters (RW.ERROR - Unable to find reference:1041). Four classes of fimbriae have been reported;
conjugative fertility (F) fimbriae, type IV fimbriae, fimbriae assembled by extracellular
nucleation/precipitation pathway and fimbriae assembled by the chaperone/usher (CU) pathway (e.g. Type
1 and type 3 fimbriae) (RW.ERROR - Unable to find reference:1041).
Type 1 fimbriae agglutinate the erythrocytes of fowl or guinea pig in the absence of a-D-mannose
(RW.ERROR - Unable to find reference:1019). Type 1 fimbriae have been characterized in several other
Enterobacteriaceae, including K. pneumoniae (RW.ERROR - Unable to find reference:1023, RW.ERROR
- Unable to find reference:1022, RW.ERROR - Unable to find reference:1021, RW.ERROR - Unable to
find reference:1020). Fimbriae are principal surface appendages on K. pneumoniae that have been well
described to possess a wide range of virulence role in the pathogenicity of Enterobacteriaceae. K.
pneumoniae fimbriae or pili play a massive role in the attachment to abiotic and biotic surfaces. The
capacity to adhere to surfaces is an essential interactive stage for the pathogen to attach, invader and cause
infection in host organisms. At least four types of fimbriae are widely described experimentally; type 1
fimbriae, type 3 fimbriae, Kpc fimbriae and KPF-28 adhesin (RW.ERROR - Unable to find reference:1025,
RW.ERROR - Unable to find reference:1024). Type 1 fimbriae, type 3 fimbriae and Kpc fimbriae but not
KPF-28 have been reported in hypermucoviscous O1:K1 strain, K. pneumoniae NUTH-K2044 with other
uncharacterized fimbrial gene loci (kpa, kpb, kpd, kpe, kpf and kpe) (RW.ERROR - Unable to find
reference:1025). The fimbriae are often encoded by an operonic cluster of genes with fimbrial-related genes
with other putative genes captured within the fimbrial gene cluster.
K. pneumoniae type 1 fimbriae are thin and rigid adhesive thread-like bacterial surface appendages encoded
by fimBEAICDFGHK (RW.ERROR - Unable to find reference:1024). K. pneumoniae, unlike E. coli, has

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
17
a unique transcriptional regulator with an N-terminally located putative DNA-bind region that is involved
in fimA transcription (RW.ERROR - Unable to find reference:1029). FimA is the major fimbria structural
protein at the base of the fimbria, while FimH is the sticky adhesins at the tip of the fimbria (RW.ERROR
- Unable to find reference:1028). A region of inverted repeat called fim switch (fimS) is located upstream
of fimA and is responsible for K. pneumoniae type 1 fimbriae phase-variation (RW.ERROR - Unable to
find reference:1024). Type 1 fimbriae fimH mediates bacterial adherence to mannose-containing structures
on host cell or environment (RW.ERROR - Unable to find reference:1031), hence attachment to surfaces.
Type 1 fimbria of K. pneumoniae has been implicated in the establishment of urinary tract infection but
reported to play less role in intestinal or lung colonization (RW.ERROR - Unable to find reference:1024).
K. pneumoniae type 3 fimbriae encoded by mrkABCDF operon is relatively longer (0.5–2-μm) and thicker
(2–4-nm wide) appendages than type 1 fimbriae. Similar to type 1 fimbriae, MrkA is the main helical
fimbrial shaft. Also, MrkD is at the tip of this fimbriae and forms the adhesive subunit that binds to collagen
molecules. The chaperone, usher and scaffolding proteins responsible for fimbrial assemblage and
stabilization are encoded by mrkB, mrkC and mrkF, respectively (RW.ERROR - Unable to find
reference:1032). Type 3 fimbriae in a mannose resistant fashion are reported to enhance in vitro attachment
to epithelial cells, kidney and lungs tissues (RW.ERROR - Unable to find reference:1033). The primary
function of type 3 fimbria is for biofilm formation, but with little to no role in intestinal and pulmonary
infections (RW.ERROR - Unable to find reference:1024). However, most K. pneumoniae urinary tract
infection is associated with K. pneumoniae biofilm formation on indwelling urinary catheters and other
contaminated medical devices. In a compensative manner, both type 1 and 3 fimbriae enhance biofilm
formation on a urinary catheter. Additionally, type 3 is involved in colonisation role for K. pneumoniae
biofilm-associated urinary infections due to the indwelling of urinary catheters (Stahlhut et al, 2012,
RW.ERROR - Unable to find reference:1035).
Other hyper mucoid K. pneumoniae strains carry Kpc fimbriae encoded by kpcABCD operon (RW.ERROR
- Unable to find reference:1025). Upstream of the gene (kpcA) expressing the primary Kpc fimbria subunit
is a site-specific recombinase encoded by kpcI with a kpcS promoter region, and these two genes together
mediate Kpc fimbria phase variation. The Kpc fimbria in E. coli further confer higher biofilm-forming
activity and believe in playing a similar role of contributing to the enhancement of K. pneumoniae biofilm
(RW.ERROR - Unable to find reference:1025, Li et al, 2014). Apart from the three types of fimbriae
described above, KPF-28 adhesin (a 28-kDa) is expressed by K. pneumoniae. KPF-28 adhesin is a thin and
long flexible major fimbrial subunit encoded by a gene on transferable CAZ-5/SHV-4 β-lactamase encoding
R plasmid (RW.ERROR - Unable to find reference:1025). A previous study indicated that K. pneumoniae
expressing both CAZ-5/SHV-4 β-lactamase and KPF-28 fimbriae were often detected in hospitals in

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
18
Clermont-Ferrand of France (RW.ERROR - Unable to find reference:1037). Adherence to human Caco-2
cell lines was reported for KPF-28 fimbriae expression K. pneumoniae. Thus, these plasmid-mediated
expressed fimbriae play a role in mammalian intestinal colonisation (RW.ERROR - Unable to find
reference:1036, Li et al, 2014). Hence, the K. pneumoniae surface adhesins are vastly involved in biofilm
formation and adherence to host cells, thereby enhancing the pathogenicity of the bacterium cannot be
overemphasised.
1.1.6.4 Iron Acquisition / Siderophores
Bacteria require an iron supply for both in vitro and in vivo growth; hence the growth of bacteria in host
organism is not only limited by host defence system but also the availability of iron to the bacteria
(RW.ERROR - Unable to find reference:1043). Living organisms, including host organisms, require an iron
supply for proper growth (RW.ERROR - Unable to find reference:806). Thus, free iron supply is rarely
available for bacteria in the host, because the irons are bounded to host carrier proteins (haemoglobin,
ferritin, hemosiderin, myoglobin, lactoferrin and transferrin). The iron acts as a redox catalyst in oxygen
and electron transport processes (RW.ERROR - Unable to find reference:1044, RW.ERROR - Unable to
find reference:1045), thus limiting the availability of irons for bacteria. Availability of free irons for bacteria
in the host during specific human pathologies (physiology post-chemotherapy and haemochromatosis)
means many pathogens can take advantage of the iron availability to cause significant morbidity and
mortality on top of the underlying disorder (RW.ERROR - Unable to find reference:1048, RW.ERROR -
Unable to find reference:1047). However, K. pneumoniae, including other bacteria secrete high-affinity,
low-molecular-weight iron chelators called siderophores used in the competitive sequestration of iron from
host molecules (RW.ERROR - Unable to find reference:806, RW.ERROR - Unable to find reference:1050).
Under iron-limiting conditions, K. pneumoniae is reported to produce between four and six outer membrane
proteins, which to play a role in iron acquisition (RW.ERROR - Unable to find reference:1053). K.
pneumoniae secrete siderophores that are specific for ferric iron uptake in bacteria (RW.ERROR - Unable
to find reference:1051). Aerobactin, enterobactin (also known as enterochelin) and yersiniabactin are
common iron chelators secreted by many K. pneumoniae strains that sequester iron from host carrier
molecules due to their high affinity (RW.ERROR - Unable to find reference:1055, RW.ERROR - Unable
to find reference:1054). Cell-specific receptors recognise iron-siderophore complexes and transport them
through TonB- dependent receptors for iron dissociation to be made available for bacteria usage
(RW.ERROR - Unable to find reference:1056). Iron-siderophore complexes are recognised by cell-specific
receptors and are transported into the cell via TonB dependent receptors, where the iron dissociates and is
made available for use by the bacterium (RW.ERROR - Unable to find reference:1056, van Aartsen, 2012).
In addition to yersiniabactin and aerobactin, at least twelve other putative iron uptake systems have been

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
19
described in K. pneumoniae NTUH-K2044 classified into four groups; Fe2+ transporter Feo, ABC
transporter, hemophore-based uptake systems and siderophore-based uptake systems (Li et al, 2014). Of
these twelve, seven are TonB-dependent and shown to be required for the full virulence of K. pneumoniae;
two are ABC transporters (Kfu and Sit), three are siderophore-based systems (Yersinia high-pathogenicity
island), Iuc and IroA. (RW.ERROR - Unable to find reference:1058, RW.ERROR - Unable to find
reference:1052, RW.ERROR - Unable to find reference:1061, RW.ERROR - Unable to find
reference:1060, RW.ERROR - Unable to find reference:1059, Li et al, 2014).
A study demonstrated that when siderophore gene from a K. pneumoniae strain was cloned into a plasmid
and expressed in an avirulent non-siderophore producing K. pneumoniae (strain), it further enhanced the
growth of the bacterium in the host tissues (RW.ERROR - Unable to find reference:1051, RW.ERROR -
Unable to find reference:1052), suggesting the role siderophores play in the growth and survival of K.
pneumoniae in the iron-deficient environment. K. pneumoniae propensity to cause infection was
significantly increased when the guinea pig was supplemented with iron parenterally (RW.ERROR -
Unable to find reference:806). Another study demonstrated the virulent role of hydroxamate-type
aerobactin in K. pneumoniae (RW.ERROR - Unable to find reference:1052). K. pneumoniae NTUH-K2044
tonB mutant reported to have shown attenuated virulence relative to the wild-type, hence reiterating the
importance of siderophores in K. pneumoniae pathogenesis (RW.ERROR - Unable to find reference:1058).
Enterobactin is produced more often by many K. pneumoniae strains and reported to have an additional
advantage of higher affinity for iron than aerobactin. Hence a better candidate for sequestering iron from
host carrier molecules such as transferrin unlike aerobactin iron source sequestered from is host cells
(RW.ERROR - Unable to find reference:806, RW.ERROR - Unable to find reference:1057). However, it
speculated that the production both siderophores, enterobactin and aerobactin, will provide a dual iron
source for K. pneumoniae, thereby enhancing the virulence of the bacterium.
1.1.6.5 Other Virulence factors
In addition to the above-discussed virulence factors, other factors associated with the virulence of K.
pneumoniae include nitrogen source utilization (urease and allantoin metabolism), outer membrane protein
(OmpA, outer membrane porins and efflux), and antibiotic resistance (to beta-lactam, quinolones,
aminoglycosides) are reported to play a considerable role in bacterial virulence (RW.ERROR - Unable to
find reference:1062, RW.ERROR - Unable to find reference:1063, Li et al, 2014). Many gut pathogens,
including K. pneumoniae, can hydrolyze urea to ammonia and carbon dioxide as a nitrogen source for
growth by using cytoplasmic urease encoded by ureDABCEFG operon (RW.ERROR - Unable to find
reference:1065). Allantoin metabolism is noted in hypervirulent K. pneumoniae and is reported to induce
K. pneumoniae associated liver infection such as primary liver abscesses in diabetic patients with increase

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
20
allantoin concentration (RW.ERROR - Unable to find reference:1017). The allantoin utilization phenotype
described in hypervirulent K. pneumoniae elevates its capability to compete for allantoin as a nitrogen
source in mammalian hosts (RW.ERROR - Unable to find reference:1067). However, K. pneumoniae have
a symbiotic association with plant roots where the bacterium can fix nitrogen into the soil for plant use. The
colonization of plant roots is aided by a combination of factor including fimbriae for attachment and
secretion systems for delivery of toxins.
Outer membrane proteins (e,g OmpA) is conserved in Enterobacteriaceae and reported to play a role in the
virulence of K. pneumoniae. A study said that lack of K. pneumoniae CPS is not enough to attenuate airway
epithelial cell-mediated inflammatory response. Rather, K. pneumoniae OmpA is known to be involved in
the inhibition of airways epithelial cell activation by acting on NF-κB-, p38- and p44/42-dependent
pathways, and contributes to resistance to alveolar macrophage phagocytosis (RW.ERROR - Unable to find
reference:1069, RW.ERROR - Unable to find reference:1071, RW.ERROR - Unable to find
reference:1070). The absence of OmpA renders K. pneumoniae susceptible to antimicrobial peptides but
has no effect on the CPS production (RW.ERROR - Unable to find reference:1075). OmpK35 and OmpK36
are two main outer membrane porins of K. pneumoniae through which hydrophilic molecules enter the
bacterium (RW.ERROR - Unable to find reference:1072) even though alternative porins such as KpnO and
OmpK26 are alternative in the absence of OmpK35/36 (RW.ERROR - Unable to find reference:1073,
RW.ERROR - Unable to find reference:1074). Increased resistance to cephalosporins/carbapenems and
attenuated virulence in the mouse model was observed in the absence of OmpK36, KpnO or OmpK26 while
lack of OmpK35 showed no effect on antibiotic resistance and virulence (RW.ERROR - Unable to find
reference:1069, Li et al, 2014, RW.ERROR - Unable to find reference:1072, RW.ERROR - Unable to find
reference:1073, RW.ERROR - Unable to find reference:1074). Absence of OmpK36 affects K. pneumoniae
surface structure altering the bacterium binding ability to phagocytic cells resulting in enhancing
susceptibility to phagocytosis and attenuated virulence (RW.ERROR - Unable to find reference:1069).
Also, efflux pumps (AcrAB) expressed in K. pneumoniae contribute to the export of antimicrobials agents,
hence determinant of K. pneumoniae resistance to host (Li et al, 2014).
1.2 Antibiotic resistance and virulence association
Antibiotic resistance is one of the major problems encountered by most medical facilities globally.
Development of antibiotic resistance by pathogenic bacteria since the discovery of antibiotics has now out-
run the discovery of new antibiotics and antibacterial therapies for solving the problem posed by these
resistant pathogenic bacteria. With a range of antimicrobial resistance and virulence factors in K.
pneumoniae strains with different phylogenetic background, the relationship between resistance and

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
21
virulence is a complex issue than often reported, hence require elucidation. The main aim of this section
of the chapter is to discuss how the association between antimicrobial resistance and virulence in K.
pneumoniae strains play a crucial role in pathogenicity. Despite the different evolutionary timescale,
pathogenic bacteria have competitively evolved through the combination of virulence and antibiotic
resistance with the sole aim of survival and persistence in the host/ environment. Hypervirulent bacteria
pose a significant health problem globally. More so, this is compounded by the emergence of antibiotic
resistance, which is threatening the treatments of many pathogens in many hospitals. Virulence and
antibiotic resistance may play independent or dependent roles during infection. However, these two
determinants share some characteristics; (i) both are necessary for bacteria survival under adverse
conditions, (ii) horizontal gene transfer (HGT) is essential in the transmission of both determinants, but
compensatory/ adaptive mutations may also be involved, (iii) antibiotic resistance is related to virulence
since the earlier is often associated with infection, (iv) both processes often have direct involvement of
efflux pumps, porins, cell wall alterations, and two-component systems that activate or repress the
expression of various resistance and virulence-related genes (RW.ERROR - Unable to find reference:1076,
RW.ERROR - Unable to find reference:1077, RW.ERROR - Unable to find reference:1072, RW.ERROR
- Unable to find reference:1084, RW.ERROR - Unable to find reference:1085).
K. pneumoniae is part of the ESKAPE group (Enterococcus faecium, Staphylococcus aureus, K.
pneumoniae, A. baumannii, P. aeruginosa, and Enterobacter species) that are responsible for most
nosocomial and community-acquired infections (RW.ERROR - Unable to find reference:1080). K.
pneumoniae has a highly plastic genome with ease in losing or gaining genomic segments by lateral gene
transfer (RW.ERROR - Unable to find reference:1077, RW.ERROR - Unable to find reference:1081).
Hypervirulent K. pneumoniae (hvKP) isolates belonging to K1, and K2 serotypes are the two major strains
often involved in infection (RW.ERROR - Unable to find reference:742). The increasing association of
virulence and antibiotic resistance is becoming more beneficial to a lot of pathogenic bacteria, including K.
pneumoniae. Also, antibiotic resistance may come with a fitness cost to K. pneumoniae; this is reported
to reduce the competitive ability of the bacterium in the absence of antibiotics. Deletion or mutations in
chromosomic genes involved in antimicrobial resistance (e.g., porins) lead to a fitness cost. However,
this fitness cost is lower for the deletion of plasmid acquired antibiotic resistance (RW.ERROR - Unable
to find reference:1082, RW.ERROR - Unable to find reference:1081, RW.ERROR - Unable to find
reference:742). In Bejing over a period a decade and a half, multidrug-resistant (MDR), extensively drug-
resistant(XDR), and pan-drug-resistant (PDR) K. pneumoniae isolate have risen to 61.4 %, 22 %, and 1.8
% respectively (RW.ERROR - Unable to find reference:742, RW.ERROR - Unable to find reference:1079).
As discussed above, CPS, LPS, fimbriae, and siderophores and efflux are the main virulence factors of K.
pneumoniae. They seem to be at the centre of classical pathogenesis (RW.ERROR - Unable to find

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
22
reference:806), though other factors such as secretion system as discussed in subsequent sections of this
thesis participate in bacterial virulence as well. K. pneumoniae rapidly acquire resistance to traditional and
new antimicrobial drugs, including resistance to beta-lactams, quinolones and aminoglycosides.
Resistance to beta-lactams: β-Lactam antibiotics are the largest class of antibiotics and are the most widely
used antibiotics. These class of antibiotics usually have a β-lactam ring in their molecular structure
(penicillins, cephalosporins, monobactams, carbapenems, and β-lactamase inhibitors). Expression of β-
lactamases, alteration of permeability and efflux pump are the primary mechanism used by K.
pneumoniae to escape β-lactam antibiotics. At the end of 1980s plasmid encoding, extended-spectrum β
lactamases (ESBL) was noted to hydrolyze oxyimino-cephalosporins (RW.ERROR - Unable to find
reference:1087). But CTX-M changed the game by remaining dominant in ESBLs with CTX-M-15 as
observed in hypervirulent and resistant K. pneumoniae strains. Thus, conferring resistance against
penicillins, first-, second-, and third generation cephalosporins, and aztreonam. ESBLs are inhibited by
β-lactamase inhibitors and remain inactive against cefoxitin and carbapenems (RW.ERROR - Unable to
find reference:742).
It has been reported that K. pneumoniae sometime acquire adhesins or potential virulence regulators
during ESBL acquisition (e.g. adhesin CF29K, aerobactin, and mucoid phenotype, now associated with
RmpA) thereby correlating with increased pathogenicity of the bacterium (Bi et al, 2015, RW.ERROR -
Unable to find reference:1086, RW.ERROR - Unable to find reference:742). Recent studies have
indicated that R-plasmids encoding a non-fimbrial protein (CF29K) and 28-kDa fimbrial protein (KPF-
28) (RW.ERROR - Unable to find reference:1036) aided the adherence of the Caco-2 cells to
siderophores and ESBL expressing K. pneumoniae strains (RW.ERROR - Unable to find reference:1088,
RW.ERROR - Unable to find reference:1089). Another study observed an increase in localized adhesion
pattern, upregulation of type 3 fimbriae expression and invasion process in ESBL-producing (SHV-4 and
SHV-12 β-lactamases) K. pneumoniae, suggesting the interplay of these factors as markers of
pathogenicity (RW.ERROR - Unable to find reference:1091, RW.ERROR - Unable to find
reference:1092). Hypermucoviscosity is more commonly associated with SHV- and TEM-type than with
CTX-M-type ESBLs-producing K. pneumoniae strains but in most cases harmful for RmpA or RmpA2
(RW.ERROR - Unable to find reference:1086, RW.ERROR - Unable to find reference:1094). Plasmids
encoding CTX-M-15 have been demonstrated to end up in strains with different genetic backgrounds,
posing variable virulence though both could cause outbreaks without any fitness cost (RW.ERROR -
Unable to find reference:1095). Strains carrying plasmids encoding CTX-M-15 belong to clonal complex
23 and therefore present worrying concerns as the clone are associated with hypervirulent traits and
multidrug resistance (RW.ERROR - Unable to find reference:742).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
23
Carbapenemases hydrolyze all cephalosporins and aztreonam, OXA-type carbapenemases in K.
pneumoniae are only active against first-generation cephalosporins, cefotaxime, ceftriaxone, and
cefepime, but not ceftazidime and aztreonam (RW.ERROR - Unable to find reference:1101). While
plasmid encoding KPC-2 is the most common, KPC variants (KPC-2 to KPC-12) differ by a few amino
acids changes (RW.ERROR - Unable to find reference:1098). They have rapidly spread due to their
broad-host-range conjugative plasmids. Delhi metallo-beta-lactamase-1 (blaNMD) and blaOXA48 (OXA-48)
have been reported in hypervirulent K. pneumoniae. The explanation for the epidemiological success of
KPC strains remains unclear (RW.ERROR - Unable to find reference:1103, RW.ERROR - Unable to
find reference:1102, RW.ERROR - Unable to find reference:1101). Few studies have examined the
fitness cost of carbapenemase-producing K. pneumoniae strains when the kpc gene is inactivated. A study
of OXA-48 producing K. pneumoniae strains demonstrated no particular trait of virulence (RW.ERROR
- Unable to find reference:1096) while NDM-1-producing strain was virulent and possessed several
virulence factors such as capsular serotype K2, enhanced biofilm formation, and human and murine
serum resistance (RW.ERROR - Unable to find reference:743). Other studies using Caenorhabditis
elegans and Galleria mellonella models demonstrated that KPC isolates showed decreased virulence
compared to non-producing carbapenemase strain (RW.ERROR - Unable to find reference:1098,
RW.ERROR - Unable to find reference:1099). In light of these observations, the success of these strains
does not seem to be linked to their particular virulence.
Apart from K. pneumoniae outer membrane proteins (OmpA, OmpK35, OmpK36) contributing to
phagocytic resistance, Ompk36, unlike OmpA, does not play a role in antimicrobial peptides resistance. In
contrast to OmpK36, OmpA does not play any role in antibiotic resistance (RW.ERROR - Unable to find
reference:1069). In K. pneumoniae strains producing ESBLs, OmpK35 and OmpK36 but the absence of
OmpK36 often result in the strain virulence impairment (RW.ERROR - Unable to find reference:1072,
RW.ERROR - Unable to find reference:1104). Bacteria usually sacrifice one or more protein to survive in
an antibiotic environment resulting in the biological cost of acquiring antibiotic resistance via mutations
(RW.ERROR - Unable to find reference:742). The expression of efflux pumps has been demonstrated to
correlate with virulence. The expression AcrAB multidrug efflux system enhances resistance against lungs
antimicrobial peptides and antibiotics (quinolones, cefoxitin, chloramphenicol, erythromycin, tigecycline)
(RW.ERROR - Unable to find reference:1106, RW.ERROR - Unable to find reference:1107) and OqxAB,
for resistance to nalidixic acid, ciprofloxacin, chloramphenicol, and cefoxitin (RW.ERROR - Unable to
find reference:1108, RW.ERROR - Unable to find reference:1109). K. pneumoniae resistance nodulation–
cell division (RND)-type efflux pump contributes to multidrug resistance still under study (RW.ERROR -
Unable to find reference:1111).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
24
Several mechanisms are available for the resistance of fluoroquinolones, such as plasmid induced
resistance, inhibition of quinolone uptake due to overexpressed efflux, altered permeability due to porin
loss, and mutations in quinolone resistance-determining regions of gyrase and topoisomerase (gyrA and
parC respectively) (RW.ERROR - Unable to find reference:1118). A study reported 3.9% prevalence of
qnr genes in K. pneumoniae isolates from blood specimens in Taiwan (RW.ERROR - Unable to find
reference:1113) while another reported a strong link between fluoroquinolone resistance and fitness
(RW.ERROR - Unable to find reference:1114). Another study suggests that this may be due to efflux
activity rather than amino acid substitutional mutations in quinolones resistance regions (RW.ERROR -
Unable to find reference:742).
Colistin interacts with lipid A resulting in the outer membrane disruption (RW.ERROR - Unable to find
reference:1115) by modifying the LPS following the addition of 4-amino-4-deoxy-L-arabinose to lipid A
associated with pbgPE operon modification (RW.ERROR - Unable to find reference:1116). A study
showed that a hypervirulent K. pneumoniae ST23 resistant to colistin resulted in in vitro fitness defect and
defect in hypermucoviscous, CPS production and serum resistance (RW.ERROR - Unable to find
reference:1117).
In summary, the most commons mechanism of resistance in K. pneumoniae is through enzyme synthesis,
especially β-lactamases (ESBLs), cephalosporinases, and carbapenemases. Resistance against β-lactams is
plasmid-mediated though they often carry some other gene associated with virulence. Consequently, the
acquisition of these plasmids by K. pneumoniae increased its virulence potential. In recent times, plasmid-
mediated CTX-M-type ESBLs and carbapenemases (KPC) have become a primary concern. However, the
plasmid-expressing enzymes do not seem to have any fitness cost, and these K. pneumoniae strains are
usually less virulent. In light of the ongoing discussion, neither antibiotic resistance or virulence in K.
pneumoniae is better over the other. It is, however, very important to state that their existence is of
importance either way and contributes to the survival of the opportunistic pathogen in the human host. In a
nutshell, antibiotic resistance tends to go hand-in-hand with the virulence of K. pneumoniae, and this
ultimately raises the threat level of the bacterium in infections.
As reported in this thesis, the role of the recently discovered bacterial T6SS as a putative virulence factor
in clinical hypervirulent K. pneumoniae HS11286 and NTUH-K2044 was examined to understand better
how T6SS contributed to the overall virulence of K. pneumoniae. But as means to an end, an antibiotic
resistance “disarmed” version of K. pneumoniae HS11286 was first created for safe laboratory use and to
serve as the parent strain for the T6SS virulence role study. Also, reported is the fitness cost of the antibiotic
resistance on the MDR K. pneumoniae HS11286 after deleting the antibiotic resistance genes. Thus, this
research throws more lights on antibiotic resistance and virulence and how the acquisition of these factors

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
25
shape the genome of MDR K. pneumoniae while specifically elucidating the virulent role of putative K.
pneumoniae T6SS gene clusters.
1.3 Secretion systems
Bacterial pathogens utilize diverse ways to survive in abiotic environments competitively or to thwart the
host immune system and to cause damage to host tissue during infection. One of the essential mechanisms
is via bacteria secretion systems used for the secretion of protein and other molecules across phospholipid
membranes. Bacterial secretion systems are dedicated protein secretion systems widely reported to secret
virulence factor from the cytosol of the bacteria into the extracellular milieu (RW.ERROR - Unable to find
reference:1145, RW.ERROR - Unable to find reference:1123). The secreted proteins through these systems
play many roles in promoting bacterial virulence ranging from attachment to abiotic and host cell surfaces
to releasing of toxins into target cells, to functional disruption hosts immune system, to environmental
resource scavenging, promotion of horizontal gene transfer of genetic content known for antibiotic
resistance and virulence (RW.ERROR - Unable to find reference:1123). Bacteria secretion apparatuses
differ types based on their structures, functions, and specificity. Based on the type of protein secretion
system (see Table 1.2 and Figure 1.4), the secreted protein is translocated across a single phospholipid
membrane, two membranes, or even three membranes including the target cell. Understanding of bacterial
protein secretion system creates a huge potential of developing antimicrobials in this era of bacterial
multidrug resistance. Not until 2006, the Gram-negative bacteria secretion systems were characterized into
five types; Type I secretion system (T1SS), Type II secretion system (T2SS), Type III secretion system
(T3SS), Type IV secretion system (T4SS) and Type V secretion system (T5SS). The importance of these
dedicated secretion systems is discussed with regards to the promotion of virulence or survival in the
bacteria niche. Therefore, elucidating protein secretion systems has a significant bearing on the field of
bacterial pathogenesis, especially Enterobacteriaceae.
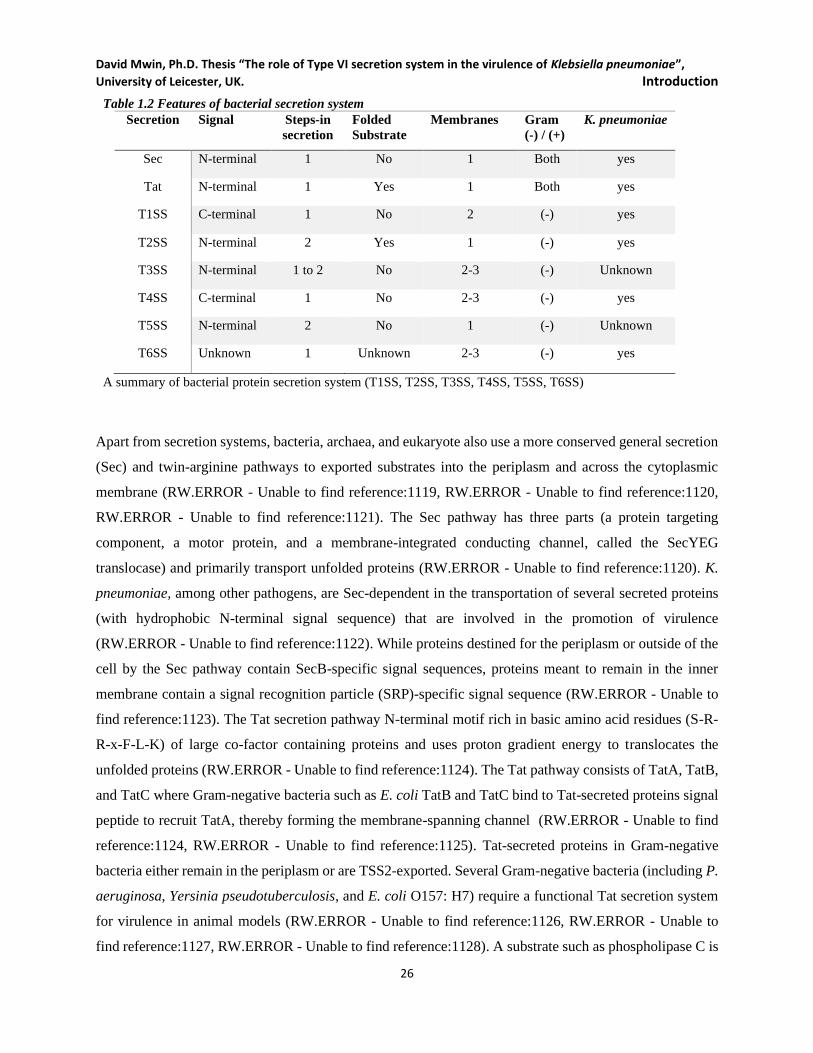
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
26
Table 1.2 Features of bacterial secretion system
Secretion Signal Steps-in
secretion
Folded
Substrate
Membranes Gram
(-) / (+)
K. pneumoniae
Sec N-terminal 1 No 1 Both yes
Tat N-terminal 1 Yes 1 Both yes
T1SS C-terminal 1 No 2 (-) yes
T2SS N-terminal 2 Yes 1 (-) yes
T3SS N-terminal 1 to 2 No 2-3 (-) Unknown
T4SS C-terminal 1 No 2-3 (-) yes
T5SS N-terminal 2 No 1 (-) Unknown
T6SS Unknown 1 Unknown 2-3 (-) yes
A summary of bacterial protein secretion system (T1SS, T2SS, T3SS, T4SS, T5SS, T6SS)
Apart from secretion systems, bacteria, archaea, and eukaryote also use a more conserved general secretion
(Sec) and twin-arginine pathways to exported substrates into the periplasm and across the cytoplasmic
membrane (RW.ERROR - Unable to find reference:1119, RW.ERROR - Unable to find reference:1120,
RW.ERROR - Unable to find reference:1121). The Sec pathway has three parts (a protein targeting
component, a motor protein, and a membrane-integrated conducting channel, called the SecYEG
translocase) and primarily transport unfolded proteins (RW.ERROR - Unable to find reference:1120). K.
pneumoniae, among other pathogens, are Sec-dependent in the transportation of several secreted proteins
(with hydrophobic N-terminal signal sequence) that are involved in the promotion of virulence
(RW.ERROR - Unable to find reference:1122). While proteins destined for the periplasm or outside of the
cell by the Sec pathway contain SecB-specific signal sequences, proteins meant to remain in the inner
membrane contain a signal recognition particle (SRP)-specific signal sequence (RW.ERROR - Unable to
find reference:1123). The Tat secretion pathway N-terminal motif rich in basic amino acid residues (S-R-
R-x-F-L-K) of large co-factor containing proteins and uses proton gradient energy to translocates the
unfolded proteins (RW.ERROR - Unable to find reference:1124). The Tat pathway consists of TatA, TatB,
and TatC where Gram-negative bacteria such as E. coli TatB and TatC bind to Tat-secreted proteins signal
peptide to recruit TatA, thereby forming the membrane-spanning channel (RW.ERROR - Unable to find
reference:1124, RW.ERROR - Unable to find reference:1125). Tat-secreted proteins in Gram-negative
bacteria either remain in the periplasm or are TSS2-exported. Several Gram-negative bacteria (including P.
aeruginosa, Yersinia pseudotuberculosis, and E. coli O157: H7) require a functional Tat secretion system
for virulence in animal models (RW.ERROR - Unable to find reference:1126, RW.ERROR - Unable to
find reference:1127, RW.ERROR - Unable to find reference:1128). A substrate such as phospholipase C is

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
27
one of such virulence proteolytic enzyme secreted by the Tat-pathway of P. aeruginosa (RW.ERROR -
Unable to find reference:1126), Legionella pneumophila (RW.ERROR - Unable to find reference:1129)
and Mycobacterium tuberculosis (RW.ERROR - Unable to find reference:1130) that can lead to the
suppression of immune cell activity and promotion of intracellular survival during infection (RW.ERROR
- Unable to find reference:1131).
Figure 1.4 Protein secretion systems in Gram-Negative bacteria.
Various type of protein secretion systems in Gram-Negative bacteria (T1SS, T2SS, T3SS, T4SS, T5SS and
T6SS).
1.3.1 The Type I Secretion System (T1SS)
T1SSs is a transmembrane Sec-independent system described in many Gram-negative bacteria of plant and
animal origins where exportation of substrates into the external milieu of the cell is achieved in a single
step (Figure 1.4) (RW.ERROR - Unable to find reference:1132). T1SS resemble ATP-binding cassette
transporter and exports unfolded small molecules (such as antibiotics, digestive enzymes and toxins) and
proteins with repeats-in-toxins (RTX) motifs. These substrates play a crucial role in the virulence and
survival of the bacteria (RW.ERROR - Unable to find reference:1133, RW.ERROR - Unable to find
reference:1134). Most of these substrates are without a C-terminal sequence and remain uncleaved even
after being exported (RW.ERROR - Unable to find reference:1123). The three essential structural units of
T1SS include ABC transporter protein in the inner membrane, a membrane fusion protein (MFP) that
crosses the inner membrane and linking the third part, the outer membrane factor (OMF) in the outer
membrane (RW.ERROR - Unable to find reference:1132). ABC transporters generate energy for the
system, MFP links ABC to the OMF across the periplasm as well select substrate using its cytoplasmic-
located N-terminus (RW.ERROR - Unable to find reference:1123) while the OMF forms an outer
membrane pore for substrate release (RW.ERROR - Unable to find reference:1135). T1SS ABC

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
28
transporters may contain C39- peptidase (C39 peptidase domain is Colicin V of E. coli) (RW.ERROR -
Unable to find reference:1136) for recognition and cleavage of N-termini, non-proteolytic C39-like
peptidase domains (CLD) (RW.ERROR - Unable to find reference:1138) and non-N-terminal ABC
transporters with C-terminal that may or may not have Rtx domain (RW.ERROR - Unable to find
reference:1134).
T1SS has been reported in the secreting of V. cholera MARTX toxin (RW.ERROR - Unable to find
reference:1139) and Serratia marcescens hemophore HasA (RW.ERROR - Unable to find reference:1140),
and uropathogenic E. coli HlyA hemolysin protein (RW.ERROR - Unable to find reference:1123). These
secreted toxins rapture host erythrocytes, nucleated eukaryotic cells, and damaging mucosal barriers and
effector immune cells (RW.ERROR - Unable to find reference:1123). Recent studies showed that T1SS is
involved in plant-bacteria association, (RW.ERROR - Unable to find reference:1134) as observed in rice
pathogen Xanthomonas oryzae pv. oryzae that use T1SS complex to express AvrXa21 which is sensed by
plant host (RW.ERROR - Unable to find reference:1141) while other studies reported metalloproteases,
adhesions and glycanases secreted by the T1SS in Agrobacterium tumefaciens, P. syringae pv tomato,
Ralstonia solanacearum, Xanthomonas axonopodis pv. citri and Xylella fastidiosa are avirulent to plants
(RW.ERROR - Unable to find reference:1134, RW.ERROR - Unable to find reference:1142). Putative
T1SS has been identified in K. pneumoniae though its role in the bacterium is unclear (RW.ERROR -
Unable to find reference:1143).
1.3.2 The Type II Secretion System (T2SS)
Type II secretion systems are conserved in most Gram-negative bacteria, where they transport folded
proteins from the periplasm into the extracellular environment. As shown in Figure 1.4, T2SS forms
channel only in the outer membrane, where Sec or Tat-dependent proteins in the periplasm are export across
the outer membrane (RW.ERROR - Unable to find reference:1122, RW.ERROR - Unable to find
reference:1146). T2SS destined substrates must first have cleavable N-terminal signal sequence to enable
it through Sec-Tat transporters before reaching T2SS (RW.ERROR - Unable to find reference:1122). T2SS
folds and export the periplasmic unfolded proteins brought in by the Sec-transporters (RW.ERROR -
Unable to find reference:1147). In some bacteria, T2SS secrete and export single protein or a wide range
of substrates (proteases, lipases, and phosphatases and several carbohydrates-processing proteins) which
contribute to the virulence of pathogens (RW.ERROR - Unable to find reference:1122, RW.ERROR -
Unable to find reference:1148). T2SS apparatus has at least 15 different proteins that are grouped into four
subassemblies; the outer-membrane complex, the inner-membrane platform, the secretion ATPase, and the

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
29
pseudopilus. The outer membrane complex has a multimeric protein called secretin that usually have a long
N-terminus extend across the periplasm to touch the inner membrane (RW.ERROR - Unable to find
reference:1147). The inner membrane platform is made of multiple copies of at least four proteins, is
embedded in the inner membrane and extending across the periplasm to connect with the secretin, thereby
communicating with the secretin, pseudopilus, and the cytoplasmic ATPase to coordinate the export of
substrates (RW.ERROR - Unable to find reference:1122). T2SS inner membrane component is not involved
in substrate transport into the periplasm. Instead, it serves as the anchoring platform for the rest of the
system. T2SS pseudopilus is structurally like T4SS on bacterial cell surfaces suggesting evolutional
relatedness (RW.ERROR - Unable to find reference:1149). Protein substrates interact with periplasmic
secretin domain to stimulate ATPase dependent retraction of T2SS pseudopilus to expel proteins substrates
through the outer membrane secretin channel (RW.ERROR - Unable to find reference:1122, RW.ERROR
- Unable to find reference:1151, RW.ERROR - Unable to find reference:1150). T2SS is deployed by a large
number of pathogens in transporting virulence factors outside the cell as observed in cholera toxin of Vibrio
cholerae (RW.ERROR - Unable to find reference:1153), exotoxin A of P. aeruginosa (RW.ERROR -
Unable to find reference:1160), ADP-ribosylating toxins of enterotoxigenic and enterohemorrhagic E. coli
(RW.ERROR - Unable to find reference:1154, RW.ERROR - Unable to find reference:1155, RW.ERROR
- Unable to find reference:1156), L. pneumophila (RW.ERROR - Unable to find reference:1157), K.
pneumoniae (RW.ERROR - Unable to find reference:1162, RW.ERROR - Unable to find reference:1161),
Aeromonas hydrophila (RW.ERROR - Unable to find reference:1158), and Dickeya dadantii (RW.ERROR
- Unable to find reference:1159).
1.3.3 Type III secretion system (T3SS)
T3SSs are needle-like injectisome/ syringe secreting a wide variety of effector proteins across the
cytoplasmic membranes into target cells in a single step (Figure 1.4) and appears to have a common
evolutionary origin with the flagellum (RW.ERROR - Unable to find reference:1166). The primary function
of the system is to release effector proteins across the bacterial and host membranes into the cytosol of host
cells, modulating a wide range of cell functions (immune and defence responses) (reviewed in (Grant et al.,
2006; Mota and Cornelis, 2005; Lindeberg et al., 2009). The effector proteins carry an embedded N-
terminal secretion signal though they are not usually cleaved. T3SS has nine conserved core proteins and
10 to 20 accessory proteins for the secretion of unfolded effectors that are chaperone-guided to the T3SS
base in an ATP-dependent manner (Abrusci et al., 2014; Burkinshaw and Strynadka, 2014). They share 8
of these proteins with the flagellar apparatus found in many bacteria and are evolutionarily related to
flagellin (Troisfontaines and Cornelis, 2005). Pathogenicity islands in bacterial chromosome or plasmid
may also carry few operons of T3SS encoding structural components. While the T3SS needle forms an

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
30
inner hollow core enough for unfolded effector to traverse (Deane et al., 2006; Demers et al., 2014), the
outer residing T3SS needle tip complex is critical for sensing contact with target cells to insert into the
target cell membrane for the release of secreted effectors (Price et al., 1991; Picking et al., 2005; Holmström
et al., 2001). The translocon does not secrete effectors into the external milieu of the bacterium, instead
provides passage for effectors by forming a pore through the target cell membrane (Price et al., 1991;
Hakansson et al., 1993; Hakansson et al., 1996).
T3SS are acquired typically by horizontal transfer, and evolutionarily distant groups may have similar
T3SSs. Although the genomes similarity between of Shigella and E. coli, the Shigella T3SS is more similar
to the Salmonella T3SS than it is to T3SSs found in the E. coli pathogens EHEC and EPEC (Troisfontaines
and Cornelis, 2005). Most T3SSs are comprised of at least 15 proteins (Abrusci et al., 2014; Burkinshaw
and Strynadka, 2014) grouped into three main components: a base complex or basal body (transmembrane
and cytoplasmic components), the needle component, and the translocon (Abrusci et al., 2014). The basal
complex forms socket-like rings with a central needle rod-like filament extending through the secretin and
into the extracellular space (Kubori et al., 1998). Yersinia, Salmonella, and Shigella T3SSs translocate
effectors into host cells with virulence consequences (Buttner, 2012) such as remodelling of regular host
cellular functions to enable survival in the host cell (Green and Mecsas, 2016). Though the earlier report of
T3SS was in pathogenic bacteria, T3SS is known to be present in rhizobial nitrogen-fixing mutualists of
plants, in the tsetse fly mutualist, Sodalis glossinidius, in the nematode mutualist, Photorhabdus
luminescens and the human commensal Pantoea agglomerans (Tseng, Tyler and Setubal, 2009; Cornelis,
2006). Plant pathogenic bacterial T3SS has two family proteins (Hrp1 and Hrp2). In contrast, the rhizobial
bacteria T3SS possess a third family though other bacteria may harbour multiple T3SS in the same genome
(e.g. SPI-1 and SPI-2 of Salmonella typhimurium) (reviewed in (Tseng, Tyler and Setubal, 2009).
1.3.4 Type IV secretion system (T4SS)
T4SSs are transmembrane secretion system and unique in a sense that, they translocate nucleic acid (DNA)
by conjugation in addition to protein effectors (Figure 1.5) across the cytoplasmic membrane of the target
cell (bacteria, or eukaryotic cell and some archaea) (Christie and Cascales, 2005; Alvarez-Martinez and
Christie, 2009). Found in both Gram-negative and positive bacteria, T4SSs are used to mediate plasmid
conjugation thereby promoting the spread of antibiotic resistance and virulence among bacteria, this leads
to enhance bacterial pathogenicity (Costa et al., 2015; Voth, Broederdorf and Graham, 2012).
Agrobacterium tumefaciens Ti plasmid, and E. coli pKM101 and R388 conjugative plasmids-encoded
T4SSs are some of the well-characterised T4SS (Alvarez-Martinez and Christie, 2009; Fronzes, Christie
and Waksman, 2009). The T4SS consists of 12 proteins (VirB1 to VirB11, and VirD4) (Alvarez-Martinez
and Christie, 2009; Fronzes, Christie and Waksman, 2009). The T4SS scaffold is formed by VirB3, VirB6,

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
31
VirB7, VirB8, VirB9 and VirB10 while VirB2 and VirB5 form the extracellular extending pilus (Trokter
et al., 2014; Christie, Whitaker and González-Rivera, 2014). VirB1 is for lytic transglycosylase-dependent
peptidoglycan degradation and pilus biogenesis while VirB2-VirB11 and VirD4 form the major
components of A. tumefaciens C58 T4SS. Most VirB proteins form the T4SS structure complex of the
secretory machinery under ATP energy hydrolysis (Fronzes, Christie and Waksman, 2009; Costa et al.,
2015).
Figure 1.5 T4SS Functional representation in most Gram-negative bacteria, including K. pneumoniae.
T4SS plays a crucial role in uptake and release of genetic material including antibiotic and virulence factors;
conjugation enables spread of virulence and antibiotic resistance; and the translocation of effector protein/
toxins and T-DNA complexes into the host cell (inhibiting host cell functions) (Fronzes, Christie and Waksman,
2009; Souza et al., 2012).
T4SS operates via two modes; one is the pilus biogenesis in which VirB11 and VirB4 bind together, and
the other mode is substrate-translocation, where VirB11 associates with VirD469 (Costa et al., 2015).
Switching between these two modes is believed to occur when the pilus tip binds to an unknown recipient

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
32
cell receptor thereby triggering the release of VirB11 associated with virB4 to initiate binding with VirD4
(for function substrate translocation)(Costa et al., 2015). T4SS components can be found in both non-
pathogenic bacteria such as Mesorhizobium loti (VirB) (Hubber, Sullivan and Ronson, 2007) and
pathogenic bacteria such as A. tumefaciens C58 (CAG), Helicobacter pylori (ComB), P. aeruginosa
(TraS/TraB), Bordetella pertussis (Ptl), E. coli (Tra) and L. pneumophila (Dot) (Christie and Cascales,
2005). Not all T4SSs contain the same sets of genes (Saier Jr, 2006). T4SS of A. tumefaciens C58 (model
T4SS) export T-DNA complex from Ti plasmid (Christie and Cascales, 2005; Souza et al., 2012). T4SS
injected T-DNA cause cancerous growth or crown gall tumours which results in carbon and energy source
discourse for the pathogen (Tseng, Tyler and Setubal, 2009; Voth, Broederdorf and Graham, 2012; Fronzes,
Christie and Waksman, 2009; Wallden, Rivera-Calzada and Waksman, 2010; Souza et al., 2012).
T4SS plays a role in the virulence of H. pylori, Bordetella pertussis and L. pneumophila by secreting
transforming proteins and effectors that support the intracellular survival of the bacteria (Alvarez-Martinez
and Christie, 2009). The L. pneumophila Dot/Icm T4SS mediate phagosome-lysosome fusion inhibitory
activities and host cell rough ER recruitment to favour the bacterial replication. This enhances the
intracellular survival of the bacterium within the phagosomes (Ensminger and Isberg, 2009). No pilus
structure is associated with B. pertussis T4SS and depend on a VirB2 pilin homologue to release their
substrates into the extracellular milieu rather than injecting them into the target host cell (Craig-Mylius
and Weiss, 1999). The presence of several putative T4SS encoding loci in K. pneumoniae such as MDR K.
pneumoniae HS11286 (Bi et al., 2015) and hyper mucoid K. pneumoniae NUTH-K2044 (Lery et al., 2014;
Bistué et al., 2008) have been demonstrated using genomic analysis. These two strains were mainly used
for the T6SS work presented in the subsequent chapter of this thesis. Pathogenic bacteria T4SSs have
evolved with a variety of functions ranging from the translocation of 50 different effector proteins into host
cells, to horizontal gene transfer-mediated spread of virulence factors and antibiotic resistance among
bacteria (conjugation, uptake and release) (Fronzes, Christie and Waksman, 2009; Voth, Broederdorf and
Graham, 2012; Alvarez-Martinez and Christie, 2009; Wallden, Rivera-Calzada and Waksman, 2010).
1.3.5 Type V secretion system (T5SS)
T5SS is a unique type of secretion systems that depend on Sec-pathway to translocate substrates into
periplasm across the inner and outer membranes of the bacteria, hence exporting the substrates in two steps.
T5SS destined proteins form β-barrel allowing the free passage of the passenger protein domain across the
outer membrane onto the surface of the bacteria. T5SS proteins are grouped into three based on the number
of proteins involved in the mechanistic secretion process; T5aSS (autotransporters, Va), T5bSS (two-
partner system, Vb) and T5cSS (the chaperone-usher system, Vc) (reviewed in (Green and Mecsas, 2016;
Tseng, Tyler and Setubal, 2009)). Autotransporters (T5Ass) are archetypal T5SS proteins that are able to

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
33
secrete themselves with at least three domains; a Sec-cleavable N-terminal passenger domain, effector
domain (linker domain) and the C-terminal β-barrel domain forming the outer membrane channel (Leyton,
Rossiter and Henderson, 2012; van Ulsen et al., 2014; Henderson et al., 2004; Jacob-Dubuisson, Fernandez
and Coutte, 2004). They are called autotransporters because the C-terminal domains form a pore-forming
β-barrel for N-terminal domain delivery(Henderson et al., 2004; Jacob-Dubuisson, Fernandez and Coutte,
2004) even though a recent study suggests otherwise (Bernstein, 2007). The translocator domain forms a β
barrel with the help of accessory components such as chaperone Skp and the Bam complex (Wagner et al,
2009; Ruiz-Perez et al, 2009) while the flexible linker domain leads the passenger domain through the
channel, which remains attached or released by protease cleavage (Leyton, Rossiter and Henderson, 2012).
The Basic structure and biogenesis of T5SS proteins are the same. However, T5aSS are the most common
in Proteobacteria genomes. The periplasmic substrates are usually unfolded, preceded by single N-terminal
signal peptide (for Sec -dependant transport) and followed by effector domain and the C-terminal outer
membrane translocation domain (Leyton, Rossiter and Henderson, 2012b; Pohlner et al., 1987; Dalbey and
Kuhn, 2012).
Few T5SS substrates depend on T5bSS where protein pairs (β-barrel and secreted protein) (Henderson et
al., 2004) are involved in the secretion of virulence proteins such as the filamentous haemagglutinin of B.
pertussis and the high-molecular-weight adhesins HWM1 and HWM2 of Haemophilus influenzae (McCann
and Geme III, Joseph W St, 2014; Lambert‐Buisine et al., 1998). The chaperone-usher secretion of T5SS
recruits the help of β-barrel forming usher protein and chaperone to facilitate the folding of the secreted
proteins before delivery into T5SS the channel (Waksman and Hultgren, 2009). An example of this system
is observed in uropathogenic E. coli for pillin (P-pilus) formation (Waksman and Hultgren, 2009).
Neisseria meningitidis T5SS secretes a multidomain autotransporter protein called NalP as precursor
proteins across the inner membrane into the periplasm via Sec-pathway. The translocator domain of the
protein inserts into the outer membrane for free external localisation of the passenger domain (N-terminal
and effector domain of the protein) (Henderson et al., 2004). IgA proteases are one of the virulence proteins
secreted by N. gonorrhoea and N. meningitidis T4SS for the degradation of host IgA antibodies (Pohlner
et al., 1987). Different translocator protein (TpsB) aid in the secretion of the effector protein (TpsA) through
the outer membrane (ref for this). Most of the T5SS secreted proteins are well characterized as toxin and
adhesins/ receptor-binding protein that significantly contribute to the virulence of pathogens to their host
(Jacob-Dubuisson, Fernandez and Coutte, 2004; Bernstein, 2007; Henderson et al., 2004). The S. flexneri
IcsA protein is for adhesion and actin-based intracellular motility) while Y. enterocolitica YadA promotes
the resistance to host complement system and translocation of T3SS substrates into the host cell (Wagner
et al., 2009; Pohlner, Halter and Meyer, 1988; Mikula, Kolodziejczyk and Goldman, 2014; Brandon et al.,

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
34
2003). Also, many T5SS bacterial adhesins and toxins have reportedly been secreted via T5SS in H. pylori,
Serratia marcescens, Rickettsia sp., Dickeya dadantii (Erwinia chrysanthemii), Proteus mirabilis and
Edwardsiellla tarda (reviewed in (Green and Mecsas, 2016; Tseng, Tyler and Setubal, 2009; Henderson et
al., 2004)). Interestingly, no account of T5SS in K. pneumoniae has been reported despite the presence of
T5SSa in closely related species within the Proteobacteria family.
1.3.6 Type VI secretion system (T6SS)
T6SS is a recently identified Gram-negative bacterial injectisome (Mougous et al., 2006). The role of T6SS
in bacteria is still under exploration for better insights. The focus of this research is on T6SS and is discussed
in great details to lay a fundamental framework for the research work that was conducted. T6SS is a versatile
envelope-spanning spring-loaded nano-machine that resembles an inverted phage-tail spike injectisome
(Figure 1.6B). Many Gram-negative bacteria use T6SS to directly introduce its effector proteins into
prokaryotic and eukaryotic target cells (Mougous et al., 2006; Zoued et al., 2014; Ho, Dong and Mekalanos,
2014; Russell, Peterson and Mougous, 2014; Tseng, Tyler and Setubal, 2009). Although recently
characterised as a functional protein secretion system in 2006 (Pukatzki et al., 2006a), clues to the presence
of T6SSs date back a decade earlier where hemolysin co-regulated protein, (Hcp now a hallmark protein of
T6SS) was reported to be secreted by V. cholera without any signal peptide cleavage as mostly in the case
of all the effector proteins in the other secretion systems (T1SS-T5SS) discussed earlier(Tseng, Tyler and
Setubal, 2009; Green and Mecsas, 2016; Beeckman and Vanrompay, 2010). Similar clues were observed
in R. leguminosarum, P. aeruginosa and L. pneumophila. Hence it was then named IcmF-associated
homologous proteins (IAHP) because this gene cluster encoded a homolog of the IcmF protein associated
with Type IVB secretion in L. pneumophila (Das and Chaudhuri, 2003). In 2006, John Mekalanos of
Harvard Medical School functionally defined The T6SS to be present in V. cholerae (Pukatzki et al., 2006a)
and now reportedly present in a broad range of Proteobacteria (Boyer et al., 2009a). Since then, T6SS is
noted to be conserved among the Proteobacteria and analogous to the T3SS and T4SS machinery even
though a majority of the T6SS proteins are unique (Bingle, Bailey and Pallen, 2008).
1.3.6.1 T6SS components, structure, and mechanism
Different organization of T6SS gene clusters are widely reported to encode T6SSs as indicated in Fig. 1.5,
however, these gene clusters are identifiable by the presence of 13 core components are shown in Figure
1.6 (Boyer et al., 2009a; Zheng and Leung, 2007). These T6SS genes within the clusters have been assigned
TssA to TssM nomenclature (Shalom, Shaw and Thomas, 2007). However, other forms of names have also
widely been used to refer to the T6SS components, such as Hcp, VgrG, ClpV, IcmF (refer to Tab. 1.3) just
to mention a few (Bhatty, Gomez and Christie, 2013; Bingle, Bailey and Pallen, 2008). However, the use

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
35
of Tss or in combination with other specific names is now widely accepted. Apart from the core Tss genes,
other accessory components associated with the T6SS gene clusters, such as fha, ppkA, tagF and sigma-54
dependent transcriptional activator vasH are associated with T6SS (Boyer et al., 2009a). Quite often, there
are other genes that are unique to a specific T6SS cluster or strain-specific (these may be a system-specific
substrate, chaperone or regulatory components). T6SS have been identified in over a quarter of analyzed
sequenced bacterial genomes (especially Proteobacteria but not Epsilon Proteobacteria), and even up to six
T6SS clusters in Burkholderia pseudomallei and Yersinia pestis. Based on the genome analysis, four
families or phylogenetic groups of T6SS, A, B, C and D as shown in Figure 1.9 (Bingle, Bailey and Pallen,
2008; Boyer et al., 2009a; Blondel et al., 2010). The presence of most T6SS clusters on pathogenicity island
together with the lack of correlation of phylogeny of T6SS cluster and bacterial taxonomy points to the
possible acquisition of the T6SS gene via horizontal transfer to fit into the bacterial environment (Boyer et
al, 2009a). Among the core components, Hcp and VgrG are considered hallmark protein and could exist in
multiples within the same clusters or genome (Barret et al., 2011).
Figure 1.6 Core T6SS gene organisation and Structural resemblance to phage-tail.
A. T6SS structure is encoded by at least 13 core/ conserved genes with a cluster or multiple clusters. The T6SS
nanotube is encoded by hcp/tssD, needle-like puncturing device by vgrG/tssI, the baseplate complex by tssE,
tssF, tssG and the sheath complex by tssB, tssB and tssA while tssJ, tssK, tssL and tssM encode the membrane
complex. B. Structural similarity between phage and T6SS. T6SS is often regarded as remnant phage-tail
injectisome. (Zoued et al., 2014a; Basler et al., 2012a; Leiman et al., 2009a; Brunet et al., 2014)
Despite the recent discovery of T6SS, considerable progress has already been made towards understanding
T6SS (Bönemann, Pietrosiuk and Mogk, 2010; Cascales and Cambillau, 2012a; Silverman et al., 2012a).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
36
The structure and function of the T6SS are enabled by 13 core components (TssA-TssM) and are
responsible for the molecular nano-machine that span across the cell envelope of the bacteria to reach the
target cells (see Fig 1.6B and Fig 1.7). Based on comparative genomics, protein remodelling and structural
homology, T6SS is often regarded as a remnant of T4 phage injectisome (Figure 1.6B) with sequence
similarities to some components of T4SS and powered by ATPase activity to contract and release substrates
into their target cell in a single step through a needle-puncturing tip protein (Shrivastava and Mande, 2008;
Leiman et al., 2009b). So far, all the effectors reported have no T6SS substrate-defining signal/ motif and
are translocated in a single step independent of Sec- or Tat pathways. The hexameric Hcp and trimeric
VgrG form the phage-tail like injecting device (with similarity to gp5 tube protein) (Mougous et al., 2006b;
Ballister et al., 2008; Pell et al., 2009). Some core proteins of the T6SS apparatus serve a dual purpose,
both as effector proteins, though other T6SS effector proteins may be secreted alongside such core proteins.
The trimeric Valine-glycine-repeat protein, VgrG (needle-like T6SS protein), forms a spike in the central
baseplate complex (Leiman et al., 2009b; Basler et al., 2012b) and often act both as a structural and effector
protein (Schneider et al., 2013b; Bhatty, Gomez and Christie, 2013). VgrG spikes are nucleation platform
for the T6SS tail tube assemblage, and the tail-tube is made of polymerised Hemolysin-Co-regulated protein
Hcp (a structural homolog of phage tail tube protein) (Brunet et al., 2014; Leiman et al., 2009b). Hcp, just
like the VgrG, served as structural protein as well as a secreted protein of T6SS. In vitro hexameric Hcp
assemble spontaneously assemble into rings with an internal aperture of ~40Å in diameter (Schneider et
al., 2013a; Mougous et al., 2006) unlike in vivo studies, where the Hcp hexamers in a head-to-tail fashion
assemble into a contractile transmembrane tubular structure via which T6SS substrates are translocated
(Pukatzki et al., 2007; Brunet et al., 2014).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
37
Figure 1.7 The assembled structural components of T6SS.
Functional T6SS is made of at least 13 conserved components (TssA-TssM) (Zoued et al., 2014a; Cascales and
Cambillau, 2012a).
The trimeric VgrG (similar gp5-gp27) sits on the Hcp tube and forms the spike-like puncturing device with
other effector domains (Figure 1.7), while the TssB and TssC form the cytoplasmic contractile tail sheath
(Leiman et al., 2009b; Basler and Mekalanos, 2012). TssE share similarity with gp25 of T4 phage, and
forms part of a cytoplasmic baseplate-like structure (Lossi et al., 2011), and TssA, TssF, TssG, TssK are
predicted to be cytoplasmic and involved in the complex basal formation. The T6SS injectisome is anchored
to the cell envelope by complex, long tabular tail sheath structure perpendicular to the membrane, and
extends deep into the bacterial cytoplasm (Basler et al., 2012b). The TssJ (OM protein), TssL (IM protein)
and TssM (lipoprotein) make up the T6SS membrane complex. TssK is a T6SS cytoplasmic baseplate
subunit thought to connect the membrane complex (TssJ–TssL–TssM) to the T6SS tail (Zoued et al., 2014a;
Ma, Lin and Lai, 2009; Felisberto-Rodrigues et al., 2011; Zoued et al., 2013). Aschtgen and colleagues
reported that TssM to be an anchoring protein to the T6SS apparatus via a peptidoglycan-binding domain
or through interaction with an accessory protein, SciZ (Aschtgen, Thomas and Cascales, 2010). Outer
membrane pore protein (analogous to T2SSand T3SS) or shared outer membrane pore protein (analogous
TolC and used by T1SS and other transmembrane systems) has been reported.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
38
Figure 1.8 Dynamic formations and dissociation T6SS in bacteria.
Diagrammatic representation of a mechanistic assemblage of T6SS (from left to right). TssA (red) recruited
at the membrane complex to recruit the baseplate for initiation of Hcp (black rectangles) polymerization to
build Hcp-TssBC (blue rectangles) during the assemblage of the nanotube. But TssA always at the distal end
is release after sheath contraction (Zoued et al., 2016).
During the growing assemblage of the Hcp tube, the heterodimers of TssB–TssC polymerise around the
tube to form the tail sheath (Basler et al., 2012b; Brunet et al., 2014) for the firm stability of Hcp tube
(Figure 1.8). Dynamic formation and disassociation of both constricted and extended Hcp tubular structures
and a tail sheath around the Hcp tube in V. cholera are believed to be ATPase energised by ClpV
dimerization (Basler et al., 2012b; Cascales, 2008). Given the diverse parts, it seems T6SS has functionally
assimilated a wide range of components, (the presence of eukaryotic post-translational regulation systems
(based around PpkA, PppA and Fha proteins).
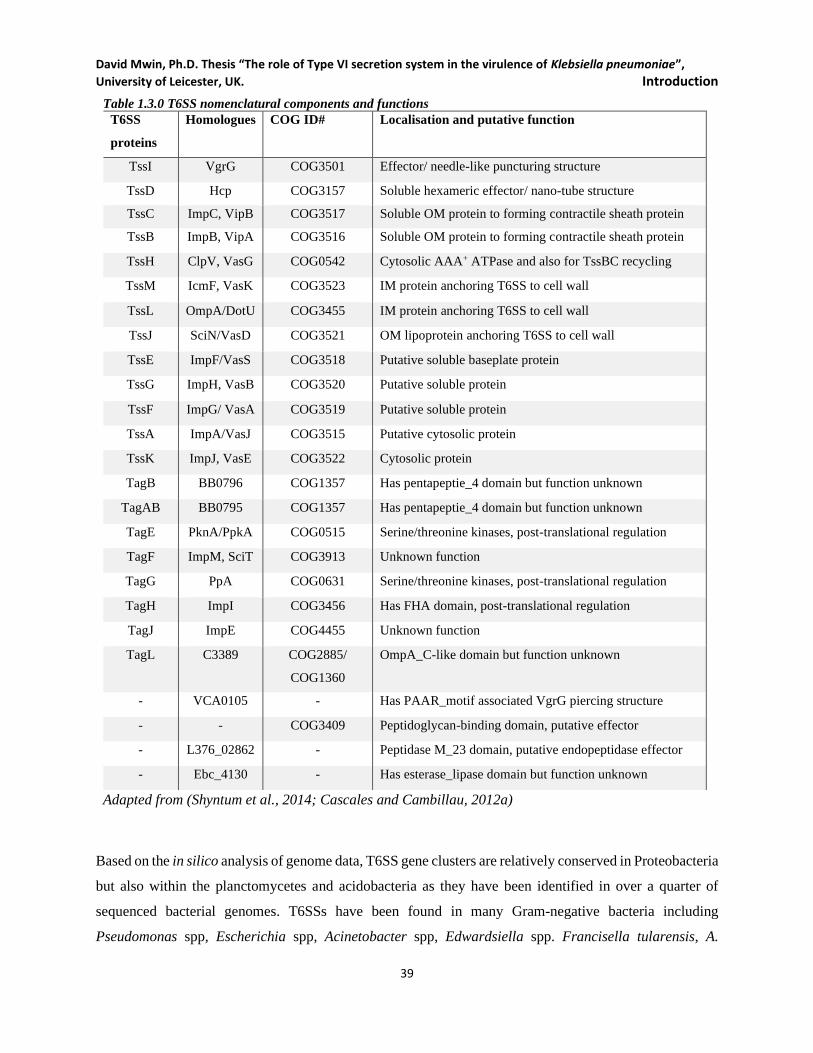
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
39
Table 1.3.0 T6SS nomenclatural components and functions
T6SS
proteins
Homologues COG ID# Localisation and putative function
TssI VgrG COG3501 Effector/ needle-like puncturing structure
TssD Hcp COG3157 Soluble hexameric effector/ nano-tube structure
TssC ImpC, VipB COG3517 Soluble OM protein to forming contractile sheath protein
TssB ImpB, VipA COG3516 Soluble OM protein to forming contractile sheath protein
TssH ClpV, VasG COG0542 Cytosolic AAA+ ATPase and also for TssBC recycling
TssM IcmF, VasK COG3523 IM protein anchoring T6SS to cell wall
TssL OmpA/DotU COG3455 IM protein anchoring T6SS to cell wall
TssJ SciN/VasD COG3521 OM lipoprotein anchoring T6SS to cell wall
TssE ImpF/VasS COG3518 Putative soluble baseplate protein
TssG ImpH, VasB COG3520 Putative soluble protein
TssF ImpG/ VasA COG3519 Putative soluble protein
TssA ImpA/VasJ COG3515 Putative cytosolic protein
TssK ImpJ, VasE COG3522 Cytosolic protein
TagB BB0796 COG1357 Has pentapeptie_4 domain but function unknown
TagAB BB0795 COG1357 Has pentapeptie_4 domain but function unknown
TagE PknA/PpkA COG0515 Serine/threonine kinases, post-translational regulation
TagF ImpM, SciT COG3913 Unknown function
TagG PpA COG0631 Serine/threonine kinases, post-translational regulation
TagH ImpI COG3456 Has FHA domain, post-translational regulation
TagJ ImpE COG4455 Unknown function
TagL C3389 COG2885/
COG1360
OmpA_C-like domain but function unknown
- VCA0105 - Has PAAR_motif associated VgrG piercing structure
- - COG3409 Peptidoglycan-binding domain, putative effector
- L376_02862 - Peptidase M_23 domain, putative endopeptidase effector
- Ebc_4130 - Has esterase_lipase domain but function unknown
Adapted from (Shyntum et al., 2014; Cascales and Cambillau, 2012a)
Based on the in silico analysis of genome data, T6SS gene clusters are relatively conserved in Proteobacteria
but also within the planctomycetes and acidobacteria as they have been identified in over a quarter of
sequenced bacterial genomes. T6SSs have been found in many Gram-negative bacteria including
Pseudomonas spp, Escherichia spp, Acinetobacter spp, Edwardsiella spp. Francisella tularensis, A.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
40
tumefaciens, Pectobacterium atrosepticum, Xanthomonas oryzae Myxococcus xanthus, Dechloromonas
aromatica and Rhodopirellula baltica (Tseng, Tyler and Setubal, 2009; Leo, Grin and Linke, 2012). Based
on T6SSs cluster analysis, T6SS can, therefore, be grouped into four families or phylogenetic groups A, B,
C, D (Figure 1.9). However, complete structures are yet to be provided for the different T6SS groups
(reviewed in (Parizad et al., 2016)). Though the overall identification and functional characterisation of
T6SS effectors poorly understood, some progress has so far been made. VgrG or TssI often carry a variety
of target specific-extensions such as actin-crosslinking domain (ACD), Proline-Alinine-Alinne-arginine
repeat proteins (PAAR), Toxin-like (RtxA), ADP-ribosyl transferase (Vip), peptidases, chitinases, lipases
and adhesins on the C-terminals and as such are collectively called “evolved VgrG’s” (Pukatzki et al., 2007)
For example, VgrG-1 from V. cholera has an actin-cross-linking domain (ACD) at the C-terminal that can
polymerize G-actins monomers (Bhatty, Gomez and Christie, 2013; Parizad et al., 2016). Many of the
effectors secreted by T6SS are of wide forms and function especially against the bacterial cell wall and
membranes further credence for supporting the T6SS interspecies bacterial competition theory (Russell,
Peterson and Mougous, 2014; Russell et al., 2011c). Bacteria carrying T6SSs often have T6SS immune
proteins to protect bacteria from self-intoxication or similar T6SS effectors from other bacteria (Russell,
Peterson and Mougous, 2014).
Figure 1.9 T6SS genealogy reported in some commonly encountered bacteria.
Gene arrangement in different families of T6SS identified in several Gram-negative bacteria (Coulthurst, 2013)

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
41
T6SS contraction and re-assembly cycle is energy-dependent, hence it not surprising that the expression
and assembly of the system are tightly regulated in some kind of transcriptional control, quorum sensing,
iron-depletion, thermoregulation or other environmental stress (reviewed in (Coulthurst, 2013)). In V.
cholera T6SS, VasH is an essential activator of the transcription initiator σ54
(RpoN) though these two are
only noted in control the transcription of two hcp operon but not the primary cluster (Pukatzki et al, 2006a;
Zheng, Ho and Mekalanos, 2011a). Despite the range of T6SS mechanisms and tight control, transcriptional
regulation of T6SS has many limitations as these regulations tend to be strain specific.
1.3.6.2 Role of T6SS; a multipurpose injectisome
The bacterial T6SS was initially reported to translocate substrates directly into the eukaryotic host cells via
needle-like appendages. However, subsequent reports indicate that T6SS is involved in mediating
interbacterial interactions. Although T6SS can, in rare instances, directly influence interactions with higher
organisms, the broader physiological significance of the system is likely to provide defence against simple
eukaryotic cells and other bacteria in the environment. The crucial role of T6SS in bacterial interactions,
along with its presence in many organisms relevant to disease, suggests that it might be a key determinant
in the progression and outcome of certain human polymicrobial infections. T6SS effectors come in two
flavours; one set with a C-terminal extension of VgrG puncturing device for mediating virulence towards
eukaryotic cells and the other set is the free toxins involved in bacterial competition and killing. A recent
study, however, suggested that V. cholerae VgrG-3 C-terminal extension is involved in peptidoglycan
degradation (Brooks et al., 2013). Interestingly, through an unclear mechanism, few emerging data suggest
that T6SS may instead be contributing to a more symbiotic or antivirulence of certain bacteria and
maintaining balance with host microflora. Hence the role of T6SS can broadly be grouped into three (see
Figure 1.10), virulence (mostly towards eukaryotic hosts such as animals and plants), Antivirulence by
inducing a more protective or symbiotic interaction with other host organisms and finally competition
(demonstration of fitness/cooperation/protection against threats of different bacteria, mostly between two
more bacteria in the same ecological niche).
Many in vitro studies have demonstrated T6SS virulence role to host cells (see Table 1.4) range from
cytotoxicity and resistance to epithelial, intracellular growth in macrophage, resistance and activation,
cytoskeletal rearrangement, inflammasome activation; Dictyostelium resistance, adhesion and invasion of
human brain microvascular endothelial cells, and to growth suppression of yeast cells (reviewed in
(Coulthurst, 2013)). Recent studies suggest T6SS functional versatility in the mediation of not just virulence
towards the eukaryotic host. It also functions to enhance commensal or mutualistic relationships between
bacteria and eukaryotes or to mediate cooperative or competitive interactions between bacteria. Pukartzi

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
42
and colleagues demonstrated that VgrG-1 carry actin crosslinking domain (ACD) of RtxA together with
other T6SS components in Vibrio necessary for actin polymerisation in host cells (Pukatzki et al., 2007).
V. cholerae T6SS may mediate an immediate inflammation and in a longer-term contribute to the bacterial
growth within the host and subsequent transmission. T6SS gene clusters in Burkholderia mediate virulence
in acute mouse pneumonia based on a hamster model (Aubert, Flannagan and Valvano, 2008a; Gavrilin et
al., 2012; Schell et al., 2007; French et al., 2011; Burtnick et al., 2010; Burtnick et al., 2011).
Figure 1.10 T6SS functional versatility.
T6SS is a versatile system involved a wide range of function but can be broadly grouped into competition,
antivirulence and virulence. (Hachani, Wood and Filloux, 2016)
Other bacteria mediating T6SS virulence towards the eukaryotic host cell or experimentally demonstrated
using an in vitro and in vivo model includes uropathogenic E. coli, Francisella species, Salmonella species,
Aeromonas hydrophila, Pectobacterium atrosepticum, E. carotovora subsp. atroseptica, just to mention a
few (reviewed in (Coulthurst, 2013)) (reviewed in (Coulthurst, 2013)).
More emerging data are suggesting the involvement of T6SS in bacterial competition other just virulence
towards the eukaryotic host. Also, several bacterial genomes harbour several T6SS-encoding gene clusters
suggesting the possibility of functional diversity of T6SS. Konovalova and colleagues alluded the role of
Myxococcus xanthus T6SS in intraspecies microbial cooperation (Konovalova, Petters and Sogaard-
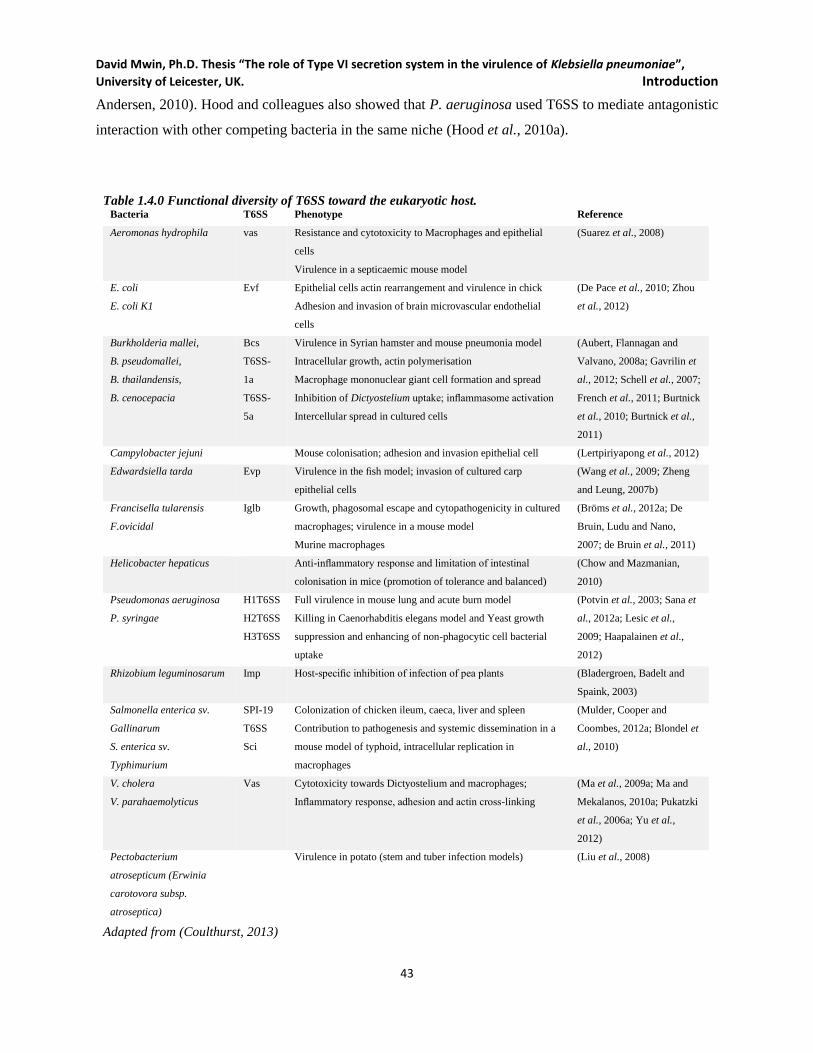
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
43
Andersen, 2010). Hood and colleagues also showed that P. aeruginosa used T6SS to mediate antagonistic
interaction with other competing bacteria in the same niche (Hood et al., 2010a).
Table 1.4.0 Functional diversity of T6SS toward the eukaryotic host. Bacteria T6SS Phenotype Reference
Aeromonas hydrophila vas Resistance and cytotoxicity to Macrophages and epithelial
cells
Virulence in a septicaemic mouse model
(Suarez et al., 2008)
E. coli
E. coli K1
Evf Epithelial cells actin rearrangement and virulence in chick
Adhesion and invasion of brain microvascular endothelial
cells
(De Pace et al., 2010; Zhou
et al., 2012)
Burkholderia mallei,
B. pseudomallei,
B. thailandensis,
B. cenocepacia
Bcs
T6SS-
1a
T6SS-
5a
Virulence in Syrian hamster and mouse pneumonia model
Intracellular growth, actin polymerisation
Macrophage mononuclear giant cell formation and spread
Inhibition of Dictyostelium uptake; inflammasome activation
Intercellular spread in cultured cells
(Aubert, Flannagan and
Valvano, 2008a; Gavrilin et
al., 2012; Schell et al., 2007;
French et al., 2011; Burtnick
et al., 2010; Burtnick et al.,
2011)
Campylobacter jejuni Mouse colonisation; adhesion and invasion epithelial cell (Lertpiriyapong et al., 2012)
Edwardsiella tarda Evp Virulence in the fish model; invasion of cultured carp
epithelial cells
(Wang et al., 2009; Zheng
and Leung, 2007b)
Francisella tularensis
F.ovicidal
Iglb Growth, phagosomal escape and cytopathogenicity in cultured
macrophages; virulence in a mouse model
Murine macrophages
(Bröms et al., 2012a; De
Bruin, Ludu and Nano,
2007; de Bruin et al., 2011)
Helicobacter hepaticus Anti-inflammatory response and limitation of intestinal
colonisation in mice (promotion of tolerance and balanced)
(Chow and Mazmanian,
2010)
Pseudomonas aeruginosa
P. syringae
H1T6SS
H2T6SS
H3T6SS
Full virulence in mouse lung and acute burn model
Killing in Caenorhabditis elegans model and Yeast growth
suppression and enhancing of non-phagocytic cell bacterial
uptake
(Potvin et al., 2003; Sana et
al., 2012a; Lesic et al.,
2009; Haapalainen et al.,
2012)
Rhizobium leguminosarum Imp Host-specific inhibition of infection of pea plants (Bladergroen, Badelt and
Spaink, 2003)
Salmonella enterica sv.
Gallinarum
S. enterica sv.
Typhimurium
SPI-19
T6SS
Sci
Colonization of chicken ileum, caeca, liver and spleen
Contribution to pathogenesis and systemic dissemination in a
mouse model of typhoid, intracellular replication in
macrophages
(Mulder, Cooper and
Coombes, 2012a; Blondel et
al., 2010)
V. cholera
V. parahaemolyticus
Vas Cytotoxicity towards Dictyostelium and macrophages;
Inflammatory response, adhesion and actin cross-linking
(Ma et al., 2009a; Ma and
Mekalanos, 2010a; Pukatzki
et al., 2006a; Yu et al.,
2012)
Pectobacterium
atrosepticum (Erwinia
carotovora subsp.
atroseptica)
Virulence in potato (stem and tuber infection models) (Liu et al., 2008)
Adapted from (Coulthurst, 2013)

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
44
T6SS is essential in the ecological fitness of bacteria living in mixed bacterial communities such as mixed
biofilms (Miyata, Bachmann and Pukatzki, 2013). Some of these bacteria also carry T6SS immunity to
avoid self-killing or resistance the strike of other competing bacteria in the same niche. Many bacteria
secreting effectors often carry cognate protection against the toxins self-intoxication or toxins of other
closely related T6SS clusters of competing bacteria while using their effectors to target the cell wall, cell
membrane and nucleic acid of the competing bacteria (Alcoforado Diniz, Liu and Coulthurst, 2015; Russell,
Peterson and Mougous, 2014). Many T6SS-harbouring bacteria such as P. aeruginosa and B. pseudomallei
possess multiple T6SS clusters. They may persist in both patients and the environment under polymicrobial
setting for many years, suggesting the possibility of functional diversity of these T6SS gene clusters to
enable the bacteria to survive in the stressful ecological niches. P. aeruginosa employs its powerful T6SS
to strike or neutralise the threats from other bacteria and ready to operate on the “tit for tat” principle in
other to survive in the mixed bacterial population (Ho, Basler and Mekalanos, 2013). In addition to targeting
eukaryotic cells, V. cholera uses T6SS to confer toxicity towards other competing bacteria. Thus, T6SS
activity may enhance the survival of the V. cholera in the aquatic ecosystem during and between the
outbreak of cholera (MacIntyre et al., 2010c). Schwarz and colleague (2010) extensively demonstrated the
involvement of Burkholderia T6SS in bacterial ecological fitness, suggesting that T6SS may be the decisive
factor in determining the survival of bacterial species in polymicrobial communities (Schwarz et al., 2010c).
The firing T6SS together with the exuding effectors may indiscriminately kill all susceptible bacteria in the
complex microbial community (Kolenbrander et al., 2010; Peters et al., 2012). A study showed that S.
marcescens T6SS does not contribute to virulence in a non-mammalian model. They instead enhance strain-
specific killing with more effective killing in S. marcescens with a very similar and active T6SS, hence
implicating the role of S. marcescens T6SS in competitiveness (Murdoch et al., 2011a). Similarly, Repizo
and colleagues indicate that A. baumannii used T6SS to prey on E. coli in a contact-dependent manner
(Repizo et al., 2015b). Thus, there are overwhelming data to support the idea that T6SS substrates are
secreted to target other bacterial competitors in a polymicrobial environment to survive and colonize their
niche.
The functional versatility of T6SS does not just end with only virulence and competition towards the
eukaryotic host and bacteria competitor respectively, but controversial antivirulence in the eukaryotic host.
T6SS has been reported to play to the advantage of the host by inducing antivirulence in host models. Few
studies suggest T6SS may be present to limit bacterial replication or virulence in the host and encouraging
a commensal or mutualistic interaction between the two (Jani and Cotter, 2010). For example, imp mutation
in R. leguminosarum promotes mutualistic cooperation by forming functional nitrogen-fixing nodules on

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
45
pea plants (Bladergroen, Badelt and Spaink, 2003) (Bladergroen et al. (2003) Thus, suggesting that T6SS
was involved in secretion of proteins that block infection of pea plants by R. leguminosarum, thus
determining host specificity of the Rhizobium-plant interaction (Jani and Cotter, 2010). Parsons and
Heffron (2005) also reported that a mutant of sciS in S. Typhimurium showed enhanced replication
macrophages and increased virulence in a mouse model suggesting that T6SS via a poorly understood
mechanism is upregulating virulence of S. Typhimurium sciS mutant (Parsons and Heffron, 2005). Also,
Chow and Mazmanian (2010) presented a compelling argument by demonstrating that mutation of T6SS
related genes in the symbiont H. hepaticus can function to limit within-host growth and virulence. The
T6SS mutant had increased intestinal colonization as well as inducing a more robust inflammatory response
in a dysbiosis model (Chow and Mazmanian, 2010). In a nutshell, compelling antivirulence evidence
present here suggest that the role of T6SS maybe be strain-specific and the consideration T6SS as a
virulence factor towards other bacteria and the host should be used with caution. The idea that multiple
T6SS clusters within individual bacterial species could mediate various types of interactions with diverse
hosts, predators, cooperators, or competitors is an enticing piece that demands attention. Also, there is the
need to explore further the role of T6SS alongside other virulence mediating factor such as antibiotic
resistance to further our understanding about how the two may contribute to the pathogenicity of the
bacteria.
1.3.6.3 Putative T6SS gene clusters identified in hyper mucoid and MDR K. pneumoniae
Multidrug-resistant (MDR) K. pneumoniae strains are very problematic and commonly isolated from both
nosocomial and community-acquired infections, thus, stressing the limits therapeutic options for
physicians. The discovery of new antibiotics to counter the evolving antibiotic resistance in bacteria remains
a puzzle and compounded when other virulence factors such as capsular polysaccharides,
lipopolysaccharides, iron acquisition systems (Nassif and Sansonetti, 1986; Cortes et al., 2002b; Shankar-
Sinha et al., 2004a), and protein secretion systems (possibly T6SS) are present in such strains. Based on
comparative in silico mining, Saris and colleagues reported the presence of three syntenic putative T6SS
clusters in four K. pneumoniae strain (three full and one partially sequenced) as well as K. variicola (Figure
1.11) (Sarris et al., 2011b). Little to no experimental data exist for K. pneumoniae T6SS since the
description of these putative T6SS gene clusters in Klebsiella. With all the accumulating literature on T6SS
and the role T6SSs play in bacteria, the role of T6SS in K. pneumoniae is missing and poorly understood.
Here this research examined to confirm the presence of T6SS gene clusters in K. pneumoniae and
demonstrate the what T6SS clusters may be doing in K. pneumoniae.
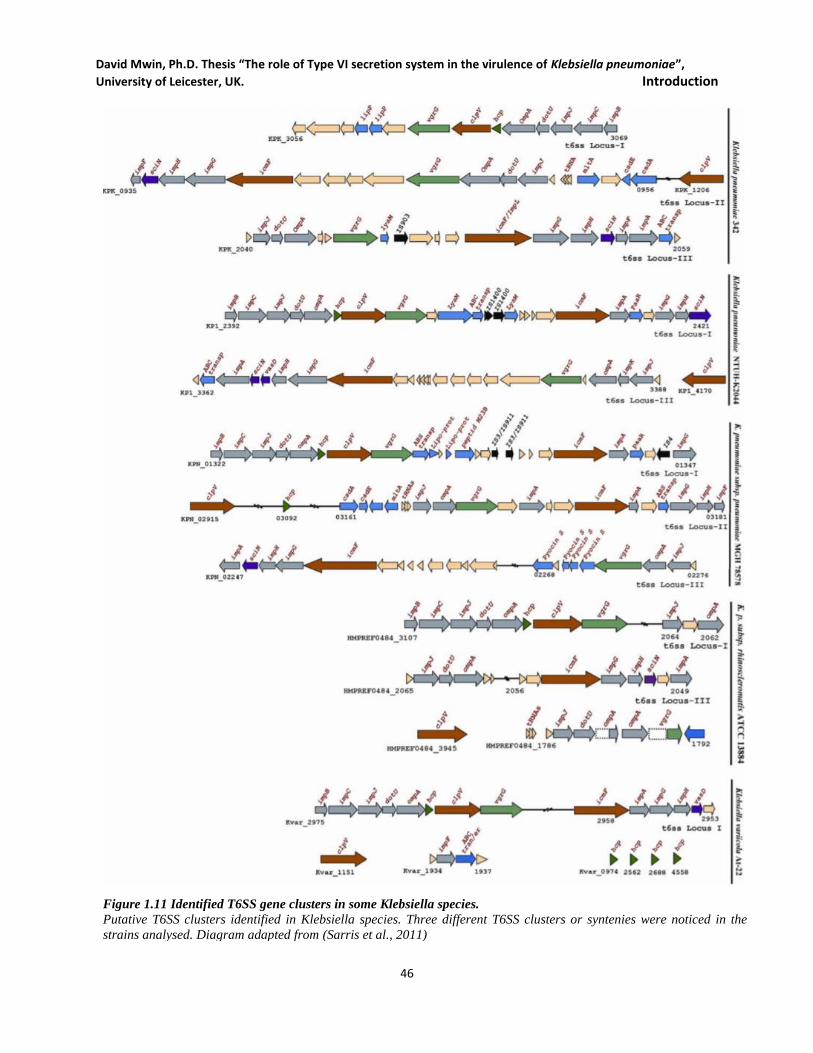
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
46
Figure 1.11 Identified T6SS gene clusters in some Klebsiella species.
Putative T6SS clusters identified in Klebsiella species. Three different T6SS clusters or syntenies were noticed in the
strains analysed. Diagram adapted from (Sarris et al., 2011)

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
47
MDR K. pneumoniae HS11286 is a clinical isolate from Shanghai patient sputum in China. It has a highly
plastic genome with one chromosome and six plasmids and carries two novel integrative and conjugative
elements (ICEKpnHS11286-1 and CEKpnHS11286-2)(Liu et al., 2012a). These plasmids (pKPHS1,
pKPHS2, pKPHS3, pKPHS4, pKPHS5 and pKPHS6) range from 122.8 Kb, 111.2 Kb, 106 Kb, 3.8 Kb, 3.4
Kb to 1.3 Kb respectively. The pKPHS6 is the smallest known K. pneumoniae plasmid. The pKPHS1
encode CTX-M-14 extended-spectrum beta-lactamase while pKPHS2 carries blaTEM-1 and carbapenemase
gene(blaKPC-2). Also, pKPHS3 carry about 13 resistant determinants (such as tetG, cat, sul1, dfra12, aac(3)-
Ia, aph, among others). Other plasmids such as pKPHS4, pKPHS5 and pKPHS6 encode some unknown
protein. At the same time, pKPHS2 and pKPHS3 carry a conjugative transfer gene (tra) that may lead to
the widespread of multidrug resistance antibiotic resistance among different genera (Liu et al., 2012a). K.
pneumoniae NTUH-K2044 is a clinical isolate from a patient with liver abscess and meningitis and
generally susceptible to a lot of antibiotics but very mucoid (high production for capsular polysaccharide)
with demonstrated high virulence in mice (Wu et al., 2009). The T6SS gene clusters in K. pneumoniae
NTUH-K2044 (especially T6SS1) were considered for mutation, but T6SS3 deletion failed. The
microviscosity of the strain and possible presence of some essential genes on T6SS3 posed a huge challenge
in the creation of T6SS3 mutant for functional studies in K. pneumoniae NTUH-K2044. Hence only T6SS1
mutant in K. pneumoniae NTUH-K2044 was analysed in this research. Understanding the antibiotic
resistance and virulence factors (T6SS in this case) associated with K. pneumoniae HS1186 and K.
pneumoniae NTUH-K2044 will not only add to the growing knowledge of antibiotic resistance and
virulence puzzle but also elucidate the role of T6SS in K. pneumoniae pathogenicity.
1.4 Research aims and objectives
Before this research work reported in this thesis, no previous work defining the role of these putative T6SS
gene clusters had been published for K. pneumoniae and more importantly in hyper mucoid or MDR K.
pneumoniae strain such as K. pneumoniae HS11286 and K. pneumoniae NTUH-K2044. However, to allow
multiple uses of various antibiotic cassettes for easy genetic manipulation, safe usage as well as
understanding the resistance repertoire of K. pneumoniae HS11286 (Liu et al., 2012a), the K. pneumoniae
HS11286 multidrug resistance arsenal was disarmed for safe laboratory usage (Bi et al., 2015). The
disarming of this strain provided an antibiotic-susceptible K. pneumoniae model strain for the studies of
putative T6SS and other future works. In a different strain, an attempt was made to study the role of T6SS1
in hyper mucoid K. pneumoniae NTUH-K2044 where T6SS1 was inactivated and examined. The presence

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
48
of T6SS in K. pneumoniae raises concern about how T6SS gene clusters may be shaping the pathogenicity
of hypervirulent K. pneumoniae and other bacteria. Hence with growing data suggesting the functional
versatility of T6SS in bacteria together with the presence of putative T6SS gene clusters in K. pneumoniae,
this research work attempted to elucidate the role T6SS in K. pneumoniae, a common agent for nosocomial
and community-acquired infections,
1. Bioinformatic analysis of K. pneumoniae resistance-mobilome, identification putative K.
pneumoniae T6SS gene clusters and the generation of molecular genetic tools and T6SS mutants
in a resistance-disarmed K. pneumoniae model strain:
a. To in silico analyzes the resistance-mobilome of MDR K. pneumoniae HS11286.
b. To perform T6SS in silico analysis and annotate putative K. pneumoniae T6SS gene
clusters.
c. To generate a library of molecular genetic tool for use in a wide range of Gram-negative
bacteria
d. To create a resistance-disarmed K. pneumoniae HS11286 as a safer model for future
functional studies in K. pneumoniae
e. To generate various K. pneumoniae T6SS gene cluster mutants for T6SS functional studies.
Resistance mobilome and two putative T6SS gene clusters were identified and mapped in K.
pneumoniae based on bioinformatic analysis and subsequently disarmed for functional studies. The
creation of a less resistant version of MDR resistant K. pneumoniae HS11286 did not only provide
safely disarmed model strain with minimized potential health risk but also enhanced the
understanding of antibiotic resistance and acquisition in MDR K. pneumoniae strains. Also, the
disarmed K. pneumoniae HS11286 model strains facilitated easy genetic manipulation for the
successful deletion of the identified putative T6SS gene clusters for functional studies.
2. Investigating the role of the putative K. pneumoniae T6SS in gene clusters in environmental
survival during bacteria-bacteria and bacteria-amoeba interactions:
a. To determine K. pneumoniae T6SS role in antibacterial virulence during bacterial
competition
b. To determine the role T6SS induced expression of other virulent mechanisms such as
fimbria-mediated biofilm formation and T4SS-mediated acquisition of mobile genetic
elements such as plasmids.
c. To determine K. pneumoniae T6SS-mediate resistance to amoeba during environmental
survival

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Introduction
49
After the generation of K. pneumoniae T6SS mutants, the role of T6SS in mediating antibacterial
virulence/ competition was examined. It was realised that K. pneumoniae uses T6SS to for survival
in complex microbial communities by unleashing antibacterial toxins in a contact-dependent
manner. Similarly, K. pneumoniae T6SS mediated antiphagocytic virulence against amoeba or
survival within the amoeba. Also, T6SS played a role in making MDR K. pneumoniae as antibiotic
resistance sink by enhancing the acquisition of foreign DNA such as plasmid and other mobile
genetic elements carrying virulence/ resistance genes. This is the first of such experimental data
that demonstrated that T6SS plays a significant role in the environmental survival of K. pneumoniae
during bacteria-bacteria and bacteria-amoeba interactions.
3. Assessing the role of K. pneumoniae T6SSs in mediating virulence against host and immuno-
stimulation of host innate immune system:
a. To measure the role of T6SS in mediating attachment, invasion and survival within host
cells such as macrophage and epithelial cells
b. To determine the role of T6SS in K. pneumoniae in host cytoskeletal rearrangement and
cell toxicity
c. To measure the K. pneumoniae T6SS innate immuno-stimulation of pro-inflammatory
response based on the induced expression of cytokines/chemokines in host cells.
d. To measure K. pneumoniae T6SS mediated in vivo virulence using Galleria larvae model
The role of K. pneumoniae in mediating anti-phagocytic resistance and survival in the host cell was
examined, and the corresponding pro-inflammatory cytokines and chemokine were measured. As
a survival mechanism within host cells, T6SS enhanced actin filament polymerization and host cell
toxicity. Finally, the T6SS was noted to improved in vivo competition of K. pneumoniae and
increased larval mortality via stimulated phenoloxidase-based melanisation.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
50
Chapter Two
Materials and Methods

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
51
Chapter 2: Material and Methods
2.1 Bacterial strains, Eukaryotic cells and Plasmids
All the bacterial isolates used in this study were obtained from Lab 212 bacterial archives stored at -20℃/-
80℃ in Brain Heart Infusion (BHI) with 30% glycerol. While the E. coli and P. aeruginosa used in this
study were obtained from Lab 212 laboratory bacterial strain collections, the K. pneumoniae strains are
clinical isolates from Shanghai Jia Tong Hospital in China and National Taiwan University Hospital in
Taiwan. Prior to experimental use, these bacterial strains were routinely grown at 37 ℃ overnight using
lysogeny agar (LA) or broth (LB) with shaking at 200 rpm unless otherwise specified. When required, E.
coli growth medium was supplemented with the appropriate antibiotics (100 µg/ml ampicillin, 50 µg/ml
kanamycin, 30 µg/ml apramycin, 25 µg/ml gentamicin, 30 µg/ml chloramphenicol, 50 µg/ml hygromycin
or 50 µg/ml streptomycin). Unless otherwise stated, K. pneumoniae strains were routinely grown in LA or
LB medium and supplemented with antibiotics when required (50 µg/ml apramycin 100 µg/ml ampicillin,
25 µg/ml tetracycline, 50 µg/ml kanamycin, 25 µg/ml gentamicin, 30 µg/ml chloramphenicol, 50 µg/ml,
hygromycin or 50 µg/ml streptomycin).
A. castellanii ATCC 30234 was a gift from Dr Simon Bath’s lab but originally purchased from American
Type Culture Collections. Details of the bacterial strains or plasmids used for this study are listed in Table
2.1 and Table 2.2, respectively. All growth media used for this work were sterilized by autoclaving at 121
℃, 15 psi, for 15 minutes or filtered with 0.45 µm-Merck Millipore filtration system.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
52
Table 2.1 List of plasmids used in this study
Plasmids Alternative designation Relevant Description Reference
pJTOOL-3 Lambda pir dependent suicide vector with sacB negative and cat markers (van Aartsen,
2012)
pKD46 Lambda RED plasmid with ampicillin resistance marker (Datsenko and
Wanner, 2000)
pFLP2 Flipase-FRT excising suicide plasmid with ampicillin marker (Hoang et al.,
1998)
pUC18R6K-mini-tn7t-Gm Source of FRT-aacC1-FRT as a gentamicin marker (Choi et al.,
2005)
pBluescript II KS (+) With lacZa and Ampicillin resistance gene (Short et al.,
1988)
pJTOOL-8 Source FRT-aph-FRT with kanamycin resistance marker Lab 212
pWSK29 Cloning plasmid with ampicillin resistance marker (Wang and
Kushner, 1991)
pKOBEG-Apra Lambda RED plasmid with apramycin resistance marker (Chaveroche,
Ghigo and
d’Enfert, 2000)
pMQ300 Sources of the hygromycin resistance marker (Kalivoda et al.,
2011)
pACYC186-OriT Cloning plasmid with ICE1 oriT and chloramphenicol resistance marker Lab 212
pIM438 Source of green fluorescent protein (GFP) marker (Murin et al.,
2012)
pKPHS6 K. pneumoniae HS11286 plasmid (1.3 kb) backbone with the origin of
replication
(Liu et al.,
2012a)
pDNTOOL-1 pUC18R6K-mini-Tn7t-aac(3)-IV For FRT-aac(3)-IV-FRT with apramycin resistance marker This study
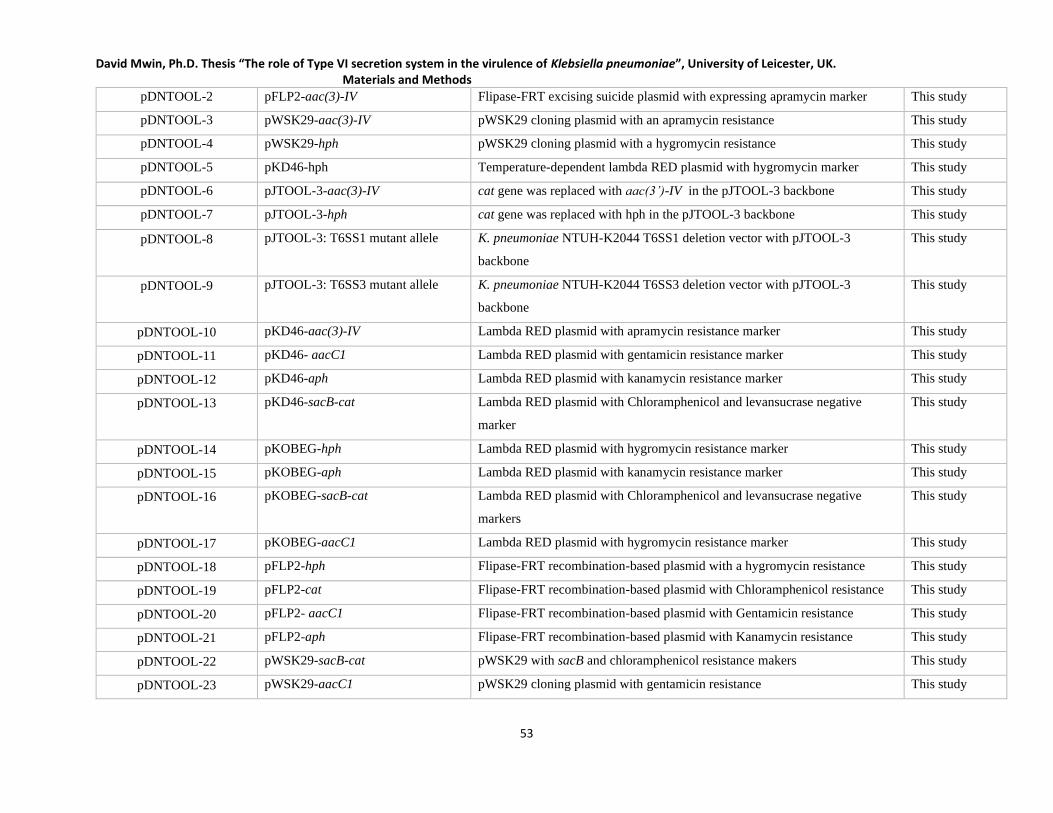
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
53
pDNTOOL-2 pFLP2-aac(3)-IV Flipase-FRT excising suicide plasmid with expressing apramycin marker This study
pDNTOOL-3 pWSK29-aac(3)-IV pWSK29 cloning plasmid with an apramycin resistance This study
pDNTOOL-4 pWSK29-hph pWSK29 cloning plasmid with a hygromycin resistance This study
pDNTOOL-5 pKD46-hph Temperature-dependent lambda RED plasmid with hygromycin marker This study
pDNTOOL-6 pJTOOL-3-aac(3)-IV cat gene was replaced with aac(3’)-IV in the pJTOOL-3 backbone This study
pDNTOOL-7 pJTOOL-3-hph cat gene was replaced with hph in the pJTOOL-3 backbone This study
pDNTOOL-8 pJTOOL-3: T6SS1 mutant allele K. pneumoniae NTUH-K2044 T6SS1 deletion vector with pJTOOL-3
backbone
This study
pDNTOOL-9 pJTOOL-3: T6SS3 mutant allele K. pneumoniae NTUH-K2044 T6SS3 deletion vector with pJTOOL-3
backbone
This study
pDNTOOL-10 pKD46-aac(3)-IV Lambda RED plasmid with apramycin resistance marker This study
pDNTOOL-11 pKD46- aacC1 Lambda RED plasmid with gentamicin resistance marker This study
pDNTOOL-12 pKD46-aph Lambda RED plasmid with kanamycin resistance marker This study
pDNTOOL-13 pKD46-sacB-cat Lambda RED plasmid with Chloramphenicol and levansucrase negative
marker
This study
pDNTOOL-14 pKOBEG-hph Lambda RED plasmid with hygromycin resistance marker This study
pDNTOOL-15 pKOBEG-aph Lambda RED plasmid with kanamycin resistance marker This study
pDNTOOL-16 pKOBEG-sacB-cat Lambda RED plasmid with Chloramphenicol and levansucrase negative
markers
This study
pDNTOOL-17 pKOBEG-aacC1 Lambda RED plasmid with hygromycin resistance marker This study
pDNTOOL-18 pFLP2-hph Flipase-FRT recombination-based plasmid with a hygromycin resistance This study
pDNTOOL-19 pFLP2-cat Flipase-FRT recombination-based plasmid with Chloramphenicol resistance This study
pDNTOOL-20 pFLP2- aacC1 Flipase-FRT recombination-based plasmid with Gentamicin resistance This study
pDNTOOL-21 pFLP2-aph Flipase-FRT recombination-based plasmid with Kanamycin resistance This study
pDNTOOL-22 pWSK29-sacB-cat pWSK29 with sacB and chloramphenicol resistance makers This study
pDNTOOL-23 pWSK29-aacC1 pWSK29 cloning plasmid with gentamicin resistance This study

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
54
pDNTOOL-24 pWSK29-aph pWSK29 cloning plasmid with a kanamycin resistance This study
pDNTOOL-25 pWSK29- aac(3)-IV:gfp pWSK29 cloning plasmid with apramycin resistance and GFP This study
pDNTOOL-26 pWSK29-hph-gfp pWSK29 cloning plasmid with hygromycin resistance and GFP This study
pDNTOOL-27 pWSK29-aacC1-gfp pWSK29 cloning plasmid with gentamicin resistance and GFP This study
pDNTOOL-28 pWSK29-aph-gfp pWSK29 cloning plasmid with kanamycin resistance and GFP This study
pDNTOOL-29 pKPHS6repA:araC:aacC1 Inducible protein expressing vector with hygromycin and gentamicin
resistance
This study
pDNTOOL-30 pKPHS6repA:araC:gfp:aacC1 Inducible protein-gfp expressing vector with hygromycin and gentamicin This study
pDNTOOL-31 pUC18R6K-mini-tn7t-sacB-cat For FRT-sacB-cat-FRT with chloramphenicol and sucrose negative marker This study
pDNTOOL-32 pUC18R6K-mini-tn7t-cat For FRT-cat-FRT with chloramphenicol resistance marker This study
pDNTOOL-33 pUC18R6K-mini-tn7t-hph For FRT-hph-FRT with hygromycin resistance marker This study
pDNTOOL-34 pUC18R6K-mini-tn7t-hph-gfp For FRT-hph-gfp-FRT with hygromycin resistance and GFP markers This study
pDNTOOL-35 pUC18R6K-mini-tn7t- aac(3)-IV -
gfp
For FRT-aac(3’)-IV-gfp-FRT apramycin resistance and GFP markers This study
pDNTOOL-36 pUC18R6K-mini-tn7t-sacB-aacC1 For FRT-sacB-aacC1-FRT with gentamicin and sucrose negative marker This study
pDNTOOL-37 pUC18R6K-mini-tn7t-sacB-hph For FRT-sacB-hph-FRT with hygromycin and sucrose negative markers This study
pDNTOOL-38 pUC18R6K-mini-tn7t-sacB-aac(3)-
IV
For FRT-sacB-aac(3’)-IV-FRT with apramycin and sucrose negative markers This study
pDNTOOL-39 pBluescript II KS (+)-aac(3)-IV aac(3)-IV was used to replace the ampicillin resistance gene This study
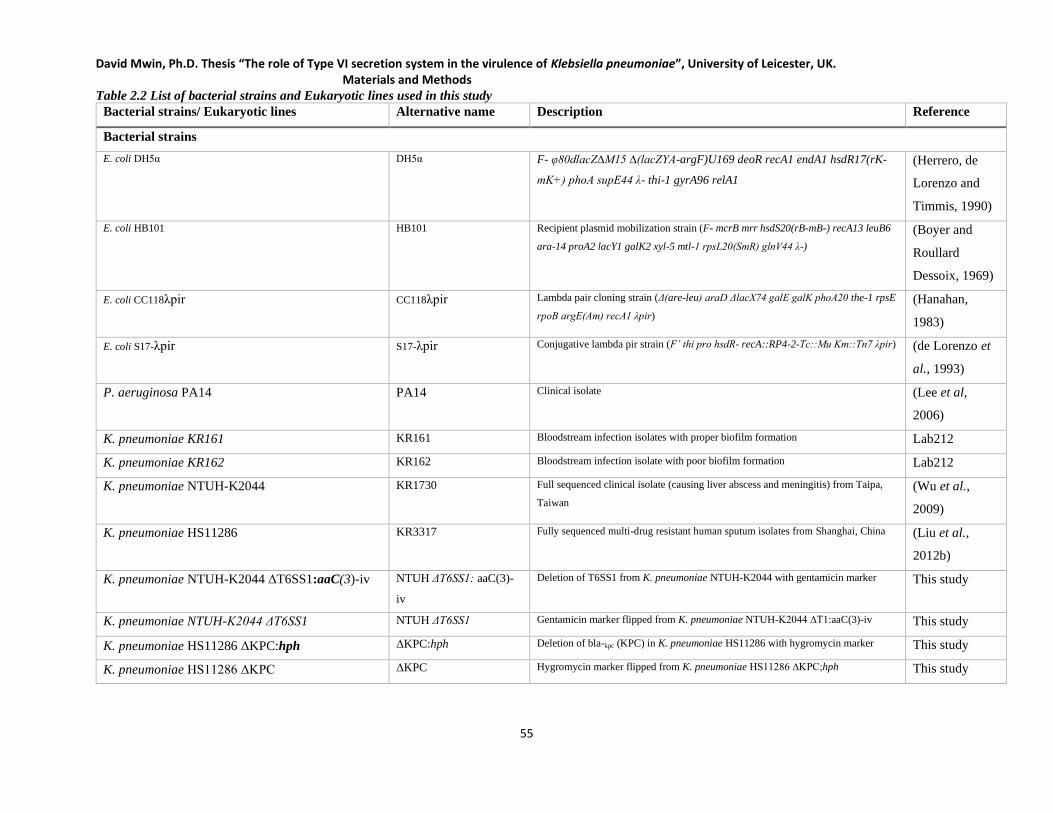
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
55
Table 2.2 List of bacterial strains and Eukaryotic lines used in this study
Bacterial strains/ Eukaryotic lines Alternative name Description Reference
Bacterial strains
E. coli DH5α DH5α F- φ80dlacZ∆M15 ∆(lacZYA-argF)U169 deoR recA1 endA1 hsdR17(rK-
mK+) phoA supE44 λ- thi-1 gyrA96 relA1
(Herrero, de
Lorenzo and
Timmis, 1990)
E. coli HB101 HB101 Recipient plasmid mobilization strain (F- mcrB mrr hsdS20(rB-mB-) recA13 leuB6
ara-14 proA2 lacY1 galK2 xyl-5 mtl-1 rpsL20(SmR) glnV44 λ-)
(Boyer and
Roullard
Dessoix, 1969)
E. coli CC118λpir CC118λpir Lambda pair cloning strain (Δ(are-leu) araD ΔlacX74 galE galK phoA20 the-1 rpsE
rpoB argE(Am) recA1 λpir)
(Hanahan,
1983)
E. coli S17-λpir S17-λpir Conjugative lambda pir strain (F’ thi pro hsdR- recA::RP4-2-Tc::Mu Km::Tn7 λpir) (de Lorenzo et
al., 1993)
P. aeruginosa PA14 PA14 Clinical isolate (Lee et al,
2006)
K. pneumoniae KR161 KR161 Bloodstream infection isolates with proper biofilm formation Lab212
K. pneumoniae KR162 KR162 Bloodstream infection isolate with poor biofilm formation Lab212
K. pneumoniae NTUH-K2044 KR1730 Full sequenced clinical isolate (causing liver abscess and meningitis) from Taipa,
Taiwan
(Wu et al.,
2009)
K. pneumoniae HS11286 KR3317 Fully sequenced multi-drug resistant human sputum isolates from Shanghai, China (Liu et al.,
2012b)
K. pneumoniae NTUH-K2044 ΔT6SS1:aaC(3)-iv NTUH ΔT6SS1: aaC(3)-
iv
Deletion of T6SS1 from K. pneumoniae NTUH-K2044 with gentamicin marker This study
K. pneumoniae NTUH-K2044 ΔT6SS1 NTUH ΔT6SS1 Gentamicin marker flipped from K. pneumoniae NTUH-K2044 ∆T1:aaC(3)-iv This study
K. pneumoniae HS11286 ΔKPC:hph ΔKPC:hph Deletion of bla-kpc (KPC) in K. pneumoniae HS11286 with hygromycin marker This study
K. pneumoniae HS11286 ΔKPC ΔKPC Hygromycin marker flipped from K. pneumoniae HS11286 ∆KPC;hph This study
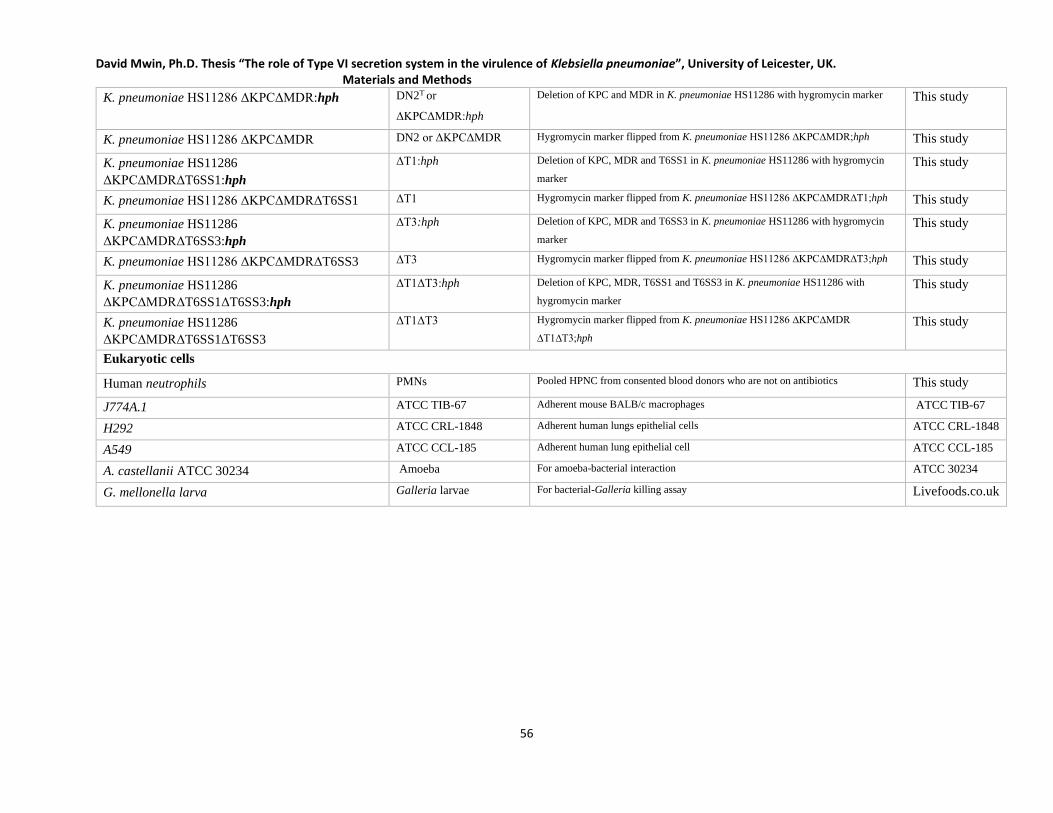
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
56
K. pneumoniae HS11286 ΔKPCΔMDR:hph DN2T or
ΔKPCΔMDR:hph
Deletion of KPC and MDR in K. pneumoniae HS11286 with hygromycin marker This study
K. pneumoniae HS11286 ΔKPCΔMDR DN2 or ΔKPCΔMDR Hygromycin marker flipped from K. pneumoniae HS11286 ∆KPC∆MDR;hph This study
K. pneumoniae HS11286
ΔKPCΔMDRΔT6SS1:hph
ΔT1:hph Deletion of KPC, MDR and T6SS1 in K. pneumoniae HS11286 with hygromycin
marker
This study
K. pneumoniae HS11286 ΔKPCΔMDRΔT6SS1 ΔT1 Hygromycin marker flipped from K. pneumoniae HS11286 ∆KPC∆MDR∆T1;hph This study
K. pneumoniae HS11286
ΔKPCΔMDRΔT6SS3:hph
ΔT3:hph Deletion of KPC, MDR and T6SS3 in K. pneumoniae HS11286 with hygromycin
marker
This study
K. pneumoniae HS11286 ΔKPCΔMDRΔT6SS3 ΔT3 Hygromycin marker flipped from K. pneumoniae HS11286 ∆KPC∆MDR∆T3;hph This study
K. pneumoniae HS11286
ΔKPCΔMDRΔT6SS1ΔT6SS3:hph
ΔT1ΔT3:hph Deletion of KPC, MDR, T6SS1 and T6SS3 in K. pneumoniae HS11286 with
hygromycin marker
This study
K. pneumoniae HS11286
ΔKPCΔMDRΔT6SS1ΔT6SS3
ΔT1ΔT3 Hygromycin marker flipped from K. pneumoniae HS11286 ∆KPC∆MDR
∆T1∆T3;hph
This study
Eukaryotic cells
Human neutrophils PMNs Pooled HPNC from consented blood donors who are not on antibiotics This study
J774A.1 ATCC TIB-67 Adherent mouse BALB/c macrophages ATCC TIB-67
H292 ATCC CRL-1848 Adherent human lungs epithelial cells ATCC CRL-1848
A549 ATCC CCL-185 Adherent human lung epithelial cell ATCC CCL-185
A. castellanii ATCC 30234 Amoeba For amoeba-bacterial interaction ATCC 30234
G. mellonella larva Galleria larvae For bacterial-Galleria killing assay Livefoods.co.uk

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
57
2.1.1 Media, reagents and solutions
Brain Heart Infusion (BHI) broth plus 30 % v/v glycerol: This was prepared by mixing 47 g of brain
heart infusion broth powder (Oxoid) and 30% (v/v) of glycerol in a final volume of 1L and autoclaved
before use.
Lysogeny Broth (LB) and Agar (LA): is also commonly called Luria-Bertani broth or agar. LB or LA
was prepared by mixing 10 g of tryptone, 5 g of yeast extract and 5 g of sodium chloride in distilled water
to a final volume of 1 litre for LB or dissolving 15g of agar base, 10 g of tryptone, 5 g of yeast extract and
5 g of sodium chloride and in distilled water to a final volume of 1 litre for LA.
LA + 5 % sucrose (LAS) or LA + 5 % sucrose + Antibiotics (LASA): LAS and LASA were used for
sucrose-counterselection media during for allelic exchange or plasmid negative selection pressure. LA
media as described above, was supplemented with 5 % sucrose to make LAS or supplemented with 5 %
sucrose and the required antibiotics to make LASA.
MacConkey agar: This was prepared by mixing 52 g of MacConkey agar powder (Oxoid) with 0.5 L of
dH2O stirred with a magnetic stirrer and top-up to 1L with distilled water and autoclaved. MacConkey agar
contains bile salts which inhibit most Gram-positive bacterial growth.
Super Optimal Broth (SOB) or Supper Optimal broth with Catabolite repression (SOC): These media
were generally used for the preparation of competent bacterial cells (SOB) or for immediate bacterial
recovery right after plasmid transformation. An SOB media was prepared by adding 20 g of tryptone, 5 g
of yeast extract and 0.5 g of sodium chloride, 0.2 g of potassium chloride, 1 g of anhydrous magnesium
chloride and 1.2 g of anhydrous magnesium sulphate into 1 litre of distilled water (pH adjusted to 7 when
needed) and autoclave. SOC was similarly prepared just like SOB but with the addition of 3.6 g of sterile
glucose after autoclaving.
M9 minimal salt: This was prepared by adding Na2HPO4 (6.9 g/liter), KH2PO4 (3 g/liter), NaCl (0.5
g/liter), NH4Cl (1 g/liter), CaCl2 (0.1 mM), MgSO4 (2 mM), and 0.5% w/v glycerol as the sole carbon
source in the media. Usually, 1ml of the trace metal mixture solution is added to 1000 ml M9 medium.
Agar base is routinely added when required for M9 minimal plates.
1000X Trace metal mix: This was prepared by adding 49 mg of Co(NO3)2.6H2O, 2860 mg of H3BO3, 222
mg of ZnSO4.7H2O, 1810 mg of MnCl.4H2O, 390 mg of Na2MoO4.2H2O2 and 79 mg of CuSO4.5H2O into
1 litre.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
58
PYG broth: The broth was prepared by mixing 0.75% w/v Difco Proteose peptone, 0.75% w/v Oxoid yeast
extract and 1.5% w/v glucose
Page’s modified Neff’s amoeba saline (PAS): PAS was prepared by mixing 1 Liter of distilled water, 120
mg of NaCl, 4 mg of MgSO4.7H2O, 4 mg of CaCl2.2H2O, 142 mg of Na2HPO4 and 36 mg of KH2PO4
Tris-EDTA (TE) buffer: The buffer was prepared by mixing equal volumes of 10 mM of Tris-HCl (pH
8.0) and 1 mM of EDTA (pH 8.0).
Tris-acetate-EDTA (TAE) buffer: This buffer was prepared by mixing of 2 M Tris-HCl, 2 M Acetic acid.
50 mM EDTA
1 M MgSO4: This was made by dissolving 24.6 g MgSO4·7H2O in 100 ml of distilled water.
0.9% Sodium Chloride (saline): The saline was prepared by dissolving 9 g NaCl (154 mM final
concentration, 0.9% w/v) in 1 litre distilled water and filter-sterilized.
Paraformaldehyde fixation buffer (4%): For a 1l of 4 % paraformaldehyde, 800 ml of 1X PBS and 40 g
of paraformaldehyde powder were mixed in a glass beaker on the heating plate at 60 ℃ under a ventilated
fume hood. The solution was gradually stirred while adding drops of 1 M NaOH to raise the pH until the
solution is clear and completely dissolved. The cooled solution was filtered before adjusting the pH to 6.9
using dilute HCl.
4′,6-Diamidine-2′-phenylindole dihydrochloride (DAPI)
Crystallized DAPI power was dissolved in dH2O to a final concentration of 3 mg/ml as a stock solution
(Store at -20). For the working solution, the stock solution was diluted in methanol to a final concentration
of 1 µg/ml (store at +2 to +8 ℃ for six months).
Rhodamine-Phalloidin working solution from cytoskeleton Inc: The staining reagent was received as
pink solid (MW 1306). The tube containing the reagent was centrifuged briefly and reconstituted with 500
µl of 100 % methanol to create a 14 µM solution stock. Aliquots were stored at -20℃ until needed.
Phosphate buffered saline (PBS): The following items were mixed for 1x PBS and adjusted to pH 7.4 in
500 ml dH2O and sterilised by autoclaving; 8 g NaCl, 0.2 g KCl, 1.15 g Na2HPO4.7H2O, 0.20 g KH2PO4.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
59
2.2 DNA-related methods and technique
2.2.1 Oligonucleotide design, In silico-PCR and synthesis
Oligonucleotides (primers) listed in Appendix 1: Primer Table for this study were all designed using a
combination of NCBI primer blast (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) and primer3/
primer3plus (http://insilico.ehu.es/primer3/ or http://www.bioinformatics.nl/cgi-
bin/primer3plus/primer3plus.cgi) by the setting all the primer design parameters to the highest quality to
increase primer specificity. The primers were then verified with In silco-PCR tool
(http://insilico.ehu.es/PCR/) when required. All primers were synthesized by Sigma Aldrich
(https://www.sigmaaldrich.com/united-kingdom.html) and received as lyophilized in tubes and then
subsequently dissolved in nH2O at 10 pmol/ul and stored at -20℃.
2.2.2 Polymerase chain reaction (PCR)
GoTaq DNA polymerase (Promega)and KOD host start polymerase (Novagen) PCR kits, were used for all
the PCR amplification of various DNA fragments in this work. GoTaq DNA polymerase was used for
regular verification of PCR amplicons sizes in plasmids, mutant alleles and bacterial strains. However, all
high-fidelity amplification of both short and long-range PCR products for cloning, mutant creation or
sequencing was done using KOD Hot Start DNA polymerase. Manufacturer’s instructions were strictly
followed in the used of these DNA enzymes for PCR amplification. Cycling conditions (see Table 2.3)
were set per the primer melting temperature (lowest Tm-2℃) and predicted PCR amplicon size.
Representative PCR cycling conditions for each enzyme are indicated in Table 2.3 below.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
60
Table 2.3 Typical PCR cycling conditions
GoTaq DNA polymerase KOD Hot Start DNA polymerase
Temperature Time Cycles Temp. ℃ Time Cycles
Standard Standard
Initial Denaturation 95℃ 2 min 1 95℃ 10 s 1
Cycle Denaturation 95℃ 30 s
25 -30
95 20 s
30 Annealing (Lowest Tm-2)℃ 30 s (Lowest Tm-2)℃ 10 s
Cycle Extension 72℃ 1 min/ kb 72℃ 20 s/ kb
Final Extension 72℃ 5 min 1 72℃ 5 min 1
Sample Hold 4℃ Until use 4℃ Until use
Touch down Touch down
Initial Denaturation 95℃ 2 min 1 95℃ 10 s 1
Cycle Denaturation 95℃ 30 s
10
95℃ 20 s
10 Annealing Lowest (Tm-2)℃ +10℃,
decrease by1 each cycle 30 s
Lowest (Tm-2)℃ +10℃,
decrease by1℃ each cycle 10 s
Cycle Extension 72℃ 1 min/ kb 68℃ 20 s/ kb
Cycle Denaturation 95℃ 30 s
20
95℃ 20 s
20-25 Annealing (Lowest Tm-2)℃ 30 s (Lowest Tm-2)℃ 10 s
Cycle Extension 72℃ 1 min/ kb 68℃ 20 s/ kb
Final Extension 72℃ 5 min 1 72℃ 5 min 1
Sample Hold 4℃ Until use Until use
Representative standard and touchdown PCR conditions for GoTaq DNA polymerase and KOD Hot Start DNA polymerase.
2.2.3 Colony PCR
A representative colony was resuspended in 100µ l of nH20 and boiled at 100℃ for 10 minutes and gently
vortexed. The cell debris was then pelleted at 12000 ×g for 2 minutes, and 1µl of the clear supernatant used
as the DNA template for that colony.
2.2.4 Splice Overlap Extension (SOE)-PCR: Mutant allele construction
SOE-PCR technique was used to join DNA fragments together for plasmid and mutant allele creation in
this study. A typical mutant allele consisted of a relevant antibiotic marker with or without GFP, and two
homologous flanking arms/fragments all joined together as a single piece of DNA fragment that can be

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
61
cloned into a specified plasmid when required. The size of each homologue flanks (Left flank, LF and Right
Flank, RF) for the target gene to be deleted ranged varied from 300 bp to 1 kb. The LF and RF of the SOE-
PCR product are homologues to the upstream and downstream regions of the target gene. When required
(for cloning into suicide vector), unique restriction sites were added to the extreme ends of mutant alleles
(restrictions sites matching the sites on the multiple cloning site on the suicide vector). The mutant alleles
that were used in the generation of most of the bacterial mutants (derivatives of K. pneumoniae H11286)
for this study via lambda RED recombination did not carry any restriction sites unlike those used in creating
suicide vector-based allelic exchange.
Another unique method of mutant allele creation was used in this study by using two primers (P1 and P2
as shown in Figure 2.1a) where 30 bp – 60 bp sequence of LF and RF were incorporated into both primers
and used to amplify the antibiotic cassette/DNA fragment, resulting in an amplicon with 30- 60 bp LF and
RF. Such unique mutant alleles or SOE-PCR product were less successfully in bacterial mutant creation,
rather the creation of various molecular genetic tools (pDNTOOL plasmids) as described subsequent
sections of this work. Occasionally, two more DNA fragments were joined together by simply design the
primers (P4 and P5 as shown in Figure 2.1b) of both fragments to overlap with complementary sequences.
A typical multi-step SOE-PCR mutant allele creation method (shown in Figure 2.1c) was employed mainly
in the creation of the bacterial mutants. Three amplicons were amplified using primer pair P7/P8 (LF),
P9/P10 (RF) and Ab-F/Ab-R (antibiotic tic cassette with two flanking FRT sites). Note that, P8 /Ab-F
primer pair and P9/Ab-R carry reverse complementary sequence at their 5’ ends. Mutant alleles were
cloned into a suicide vector with the help of restriction-ligation enzymes or used directly as in the case of
lambda RED recombination. During the SOE-PCR process (Figure 2.1a and Figure 2.1b), regular PCR
was run using the variety of overlapping DNA fragments but without primers for 10-15 cycles. The PCR
machine then paused at the extension phase for the addition of the external primers (P3 and P6 in Figure
2.1b and P10 and P11 in Figure 2.1c) and the PCR process was then allowed to continue for another 25-30
cycles.
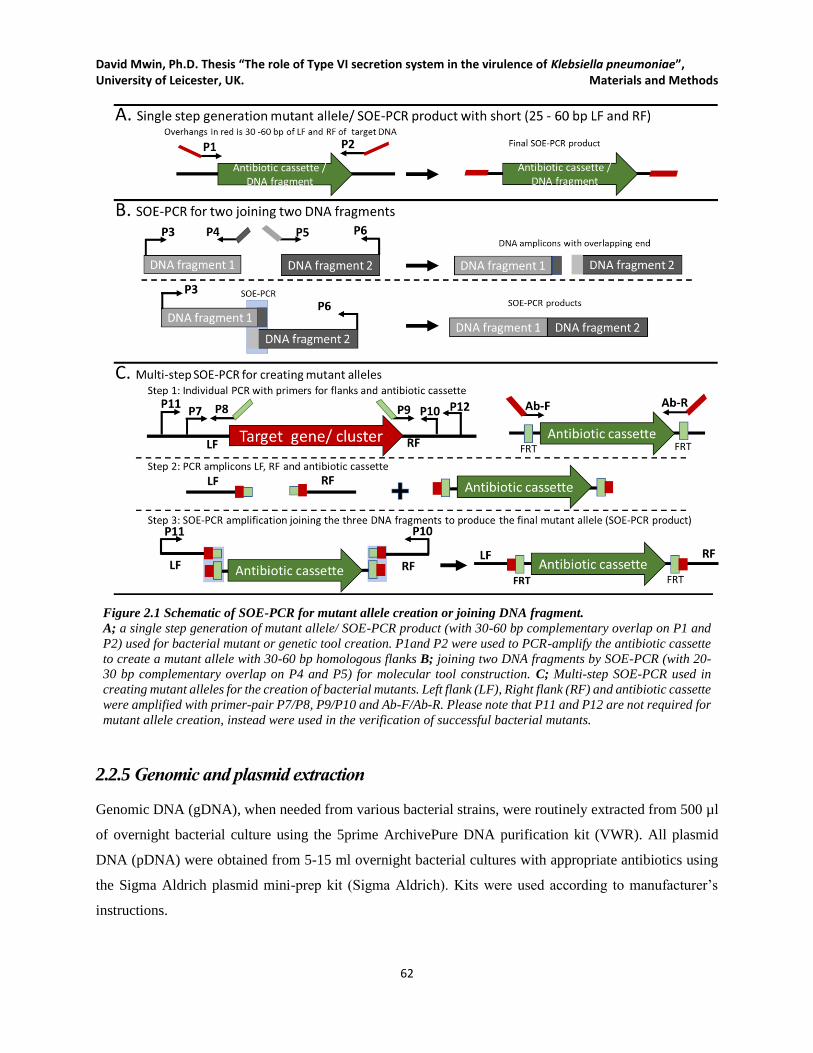
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
62
Figure 2.1 Schematic of SOE-PCR for mutant allele creation or joining DNA fragment.
A; a single step generation of mutant allele/ SOE-PCR product (with 30-60 bp complementary overlap on P1 and
P2) used for bacterial mutant or genetic tool creation. P1and P2 were used to PCR-amplify the antibiotic cassette
to create a mutant allele with 30-60 bp homologous flanks B; joining two DNA fragments by SOE-PCR (with 20-
30 bp complementary overlap on P4 and P5) for molecular tool construction. C; Multi-step SOE-PCR used in
creating mutant alleles for the creation of bacterial mutants. Left flank (LF), Right flank (RF) and antibiotic cassette
were amplified with primer-pair P7/P8, P9/P10 and Ab-F/Ab-R. Please note that P11 and P12 are not required for
mutant allele creation, instead were used in the verification of successful bacterial mutants.
2.2.5 Genomic and plasmid extraction
Genomic DNA (gDNA), when needed from various bacterial strains, were routinely extracted from 500 µl
of overnight bacterial culture using the 5prime ArchivePure DNA purification kit (VWR). All plasmid
DNA (pDNA) were obtained from 5-15 ml overnight bacterial cultures with appropriate antibiotics using
the Sigma Aldrich plasmid mini-prep kit (Sigma Aldrich). Kits were used according to manufacturer’s
instructions.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
63
2.2.6 Gel electrophoresis, DNA purification and Sequencing
Gel electrophoresis for visualization of DNA fragments was routinely performed in 0.8 % to 1.5 % agarose
gels (depending on the DNA fragments) made with agarose powder, 1× TAE buffer and 0.5 µg/ml ethidium
bromide. Quick-Load 2-log DNA Ladder, λ-HindIII, or GeneRulerTM (Thermo Fisher) were used as DNA
standards. DNA fragments were either visualized in the gel using gel documenting system (Bio-Rad) or
excised and subsequently purified from the gel using YORBIO Gel/PCR DNA purification kit (Geneflow)
following manufacturer’s instructions Quantification of purified DNA was done using Nanodrop DNA
quantification system (Thermo Fisher) before being used in any downstream works or sequencing. PCR
amplicons and plasmid inserts were routinely Sanger-sequenced by GATC Biotech.
2.2.7 Restriction digestion
All restriction enzymes were obtained from New England Biolabs, Promega and Roche and sored at -20℃
before usage. 25-50 µl reaction volume was routinely performed according to the manufacturer’s protocol,
chosen enzymes and DNA amount. For complete digestion reaction, the required restriction enzymes were
set up in a compatible buffer at 37℃ for 30 – 60 minutes and then ran on 0.8 -1 % agarose gel. All restriction
digestions meant for downstream application were first heat-inactivated per manufacturer’s protocol.
2.2.8 DNA dephosphorylation and ligation
During ligation, to prevent linearized plasmids from self-ligation after restriction digestion, thermosensitive
alkaline phosphatase (TSAP, Promega) was added to plasmid restriction digestion at 1 U/µg DNA. Once
the reaction was completed, TSAP was inactivated at 74℃ for 15 minutes.
The ligation reaction was performed in a final volume of 10-30 µl containing 3-5 U of T4 DNA ligase
(Promega) and 1-3 µl of 10× ligation buffer incubated at room temperature for one hour followed by 4℃
overnight incubation. The ligation mixture was then transformed into competent E. coli and subsequently
selected with the appropriate antibiotic medium and temperature.
2.3 RNA-related techniques and methods
2.3.1 RNA extraction and cDNA library preparation
Norgen Total RNA purification kit (following manufacturer’s instructions) was used to extract total RNA
from K. pneumoniae grown in LB for 16 h at 37℃ and 200 rpm) unless otherwise mentioned in specific
experiments. Ambion TURBO DNA-free kit was used to remove residual DNA from RNA preparations,

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
64
and cDNA library was synthesized using QuantiTect Reverse Transcription Kit (Qiagen) per
Manufacturer’s instructions Also, an identical reaction without reverse transcriptase was performed as a
control to check for DNA contamination.
2.3.2 Transcriptional analysis of T6SS, T4SS and fimbrial gene clusters
Each cDNA library and its reverse transcriptase negative controls were analyzed for transcripts
corresponding to hcp1 and vgrG1 (T6SS1), hcp2 and vgrG3 (T6SS3), virB4 and mob (T4SS3), fimH (Type
1 fimbriae), and mrkD (Type 3 fimbriae). The regions corresponding to hcp1, vgrG1, hcp2, vgrG3, virB4,
mobB, fimH and mrkD were then PCR-amplified from the cDNA. Hcp and VgrG proteins (Hcp1, VgrG1
Hcp2 and VgrG3) are both structural and secreted proteins of T6SS. VirB4 and MobB are T4SS conjugal
transfer ATPase and T4SS relaxosome-DNA coupling protein for T4SS conjugative machine while FimH
and MrkD are fimbrial adhesins for type 1 and type 3 fimbriae respectively. Primer pair DN135/DN136
(136 bp) were used for hcp1, DN137/DN138 (167 bp) were used for vgrG1, DN139/DN140 (148 bp) were
used for hcp2, DN141/DN142 (173 bp) were used for vgrG3, DN147/DN148 (189 bp) were used for virB4,
DN149/DN150 (165 bp) were used for mobB, DN143/DN144 (187 bp) were used for fimH, DN145/DN146
(198 bp) were used for mrkD.
2.3.3 qRT-PCR analysis of T6SS, T4SS, fim and mrk fimbrial clusters.
For in vitro samples, 2 µl of 10-1 dilutions of cDNA library and its reverse transcriptase negative controls
for each set of experiments were used for quantitative real-time PCR (qRT-PCR). qRT-PCR reactions (20
µl) were set up using SensiMix Plus SYBR (Bioline) according to the manufacturer’s instructions. Also, a
reaction for internal control, rpoD, was set-up using DN151/DN152 primer-pair (164 bp). For
hcp1(DN135/DN136), vgrG1 (DN137/DN138), hcp2 (DN139/DN140), vgrG3 (DN141/DN142), virB4
(DN147/DN148), mob (DN149/DN150), fimH (DN143/DN144), and mrkD (DN145/DN146), 10 pmol of
each primer was added to the reaction mixture. Cycling conditions consisted of 40 cycles of 95ºC for 15
seconds, 60ºC for 15 seconds and 72ºC for 20 seconds. Each qRT-PCR reaction was performed in triplicate
using the Qiagen Rotor Gene 6000. The relative quantification of hcp1, vgrG1, hcp2, vgrG3, virB4, mobB,
fimH, and mrkD transcripts versus rpoD was determined according to the Pfaffl equation (Pfaffl, 2001).
2.4 Construction of pDNTOOL plasmids
Most clinical K. pneumoniae strains are resistant to a broad range of antibiotics (as is the case of K.
pneumoniae HS11286), hence finding the right genetic tools to embark on a successful study of the
bacterium can be problematic. This section describes all the useful plasmid-based tool (pDNTOOLs) that

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
65
were carefully constructed to make available and aid in the genetic manipulation of most Gram-negative
bacteria, including K. pneumoniae and E. coli. The constructed plasmids, pDNTOOLs, were of significant
use in this work and beyond, as was the case with colleague lab members and collaborators. The
pDNTOOLs are categorized into suicide deletion vectors, lambda RED recombination-based plasmids,
necessary antibiotic resistance cassettes with or without GFP flanked by FRTs, flippase-FRT
recombination-based plasmids and expression vectors.
2.4.1 Construction of suicide deletion vectors
Unlike K. pneumoniae HS11286, K. pneumoniae NTUH-K2044 is a hyper-mucoid strain and
transformation with mutant alleles were unsuccessful for mutant creation, as such, a suicide plasmid-based
allelic exchange was employed for the production of T6SS mutants. A lambda pir dependent plasmid,
pJTOOL-3, encoding chloramphenicol resistance (cat gene) to was used as the backbone for the
constructions of K. pneumoniae NTUH-K2044 T6SS1 and T6SS3 deletion vectors (See Figure 2.2 and
Figure 2.3). First, T6SS1 mutant allele (2595 bp) was SOE-PCR constructed using the primer pairs
DN1/DN2 (LF,733 bp), DN3/DN4 (RF, 852 bp) and Gm-F/ Gm-R (gentamicin resistance cassette 1052
bp), while DN7/DN8 (910 bp), DN9/DN10 (845 bp) and Gm-F/ Gm-R were used for T6SS3 mutant allele
(2760 bp). The T6SS mutant alleles were then separately cloned into the spel/ xhol sites of pJTOOL-3 and
transformed into E. coli cc18λpir resulting in pDNTOOL-8 (for T6SS1) and pDNTOOL-9 (for T6SS3).
Plasmids were verified phenotypically for both gentamicin and chloramphenicol resistance and
genotypically by restriction digestion, PCR and sequencing. The plasmids transformed into E. coli cc18λpir
were purified and later transformed into E. coli s17λpir for conjugation with K. pneumoniae NTUH-K2044.
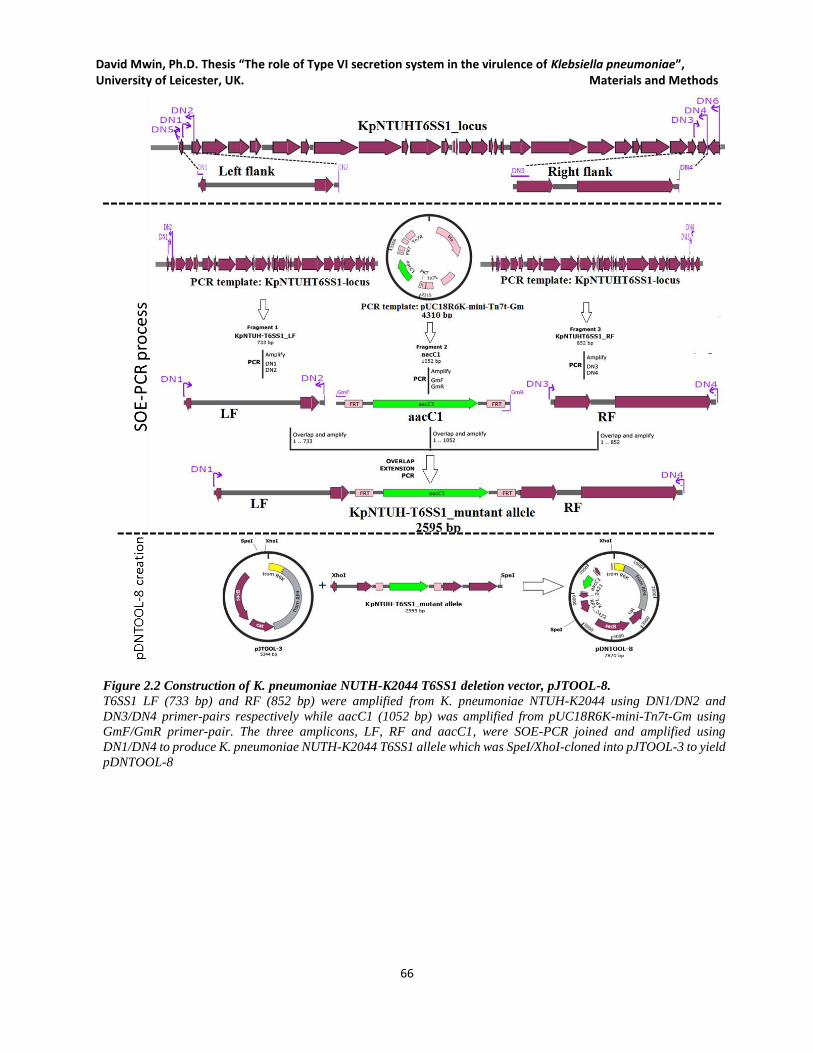
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
66
Figure 2.2 Construction of K. pneumoniae NUTH-K2044 T6SS1 deletion vector, pJTOOL-8.
T6SS1 LF (733 bp) and RF (852 bp) were amplified from K. pneumoniae NTUH-K2044 using DN1/DN2 and
DN3/DN4 primer-pairs respectively while aacC1 (1052 bp) was amplified from pUC18R6K-mini-Tn7t-Gm using
GmF/GmR primer-pair. The three amplicons, LF, RF and aacC1, were SOE-PCR joined and amplified using
DN1/DN4 to produce K. pneumoniae NUTH-K2044 T6SS1 allele which was SpeI/XhoI-cloned into pJTOOL-3 to yield
pDNTOOL-8
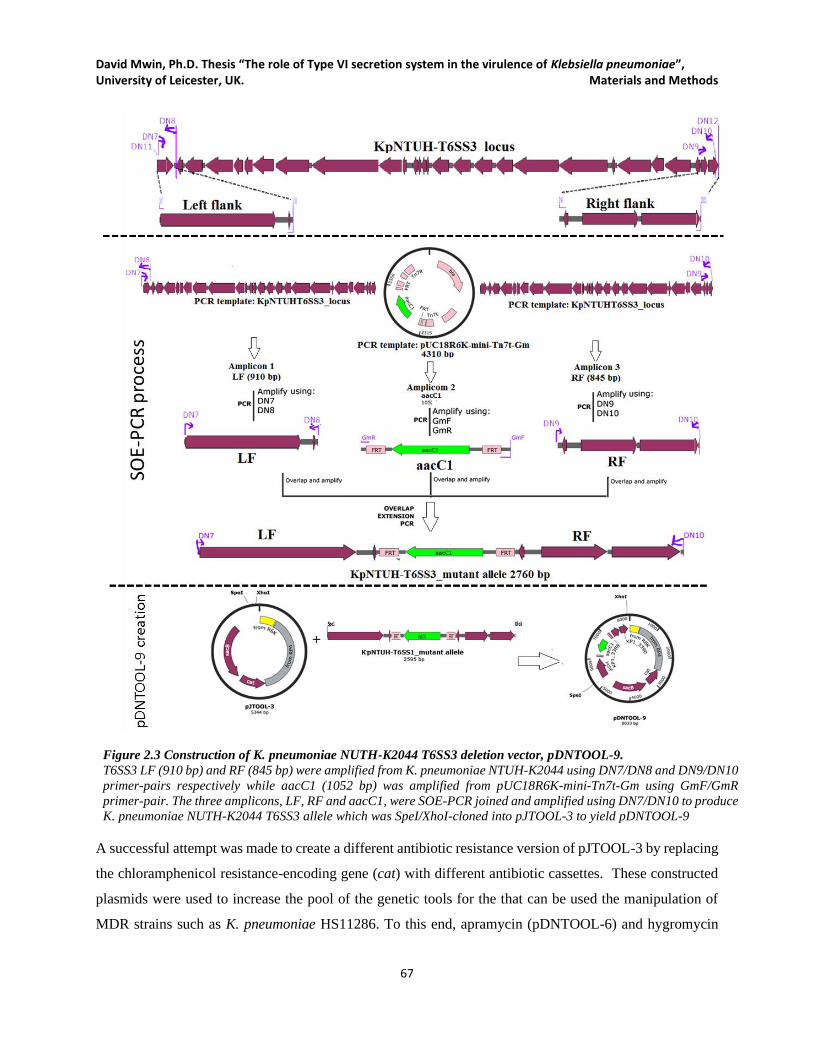
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
67
Figure 2.3 Construction of K. pneumoniae NUTH-K2044 T6SS3 deletion vector, pDNTOOL-9.
T6SS3 LF (910 bp) and RF (845 bp) were amplified from K. pneumoniae NTUH-K2044 using DN7/DN8 and DN9/DN10
primer-pairs respectively while aacC1 (1052 bp) was amplified from pUC18R6K-mini-Tn7t-Gm using GmF/GmR
primer-pair. The three amplicons, LF, RF and aacC1, were SOE-PCR joined and amplified using DN7/DN10 to produce
K. pneumoniae NUTH-K2044 T6SS3 allele which was SpeI/XhoI-cloned into pJTOOL-3 to yield pDNTOOL-9
A successful attempt was made to create a different antibiotic resistance version of pJTOOL-3 by replacing
the chloramphenicol resistance-encoding gene (cat) with different antibiotic cassettes. These constructed
plasmids were used to increase the pool of the genetic tools for the that can be used the manipulation of
MDR strains such as K. pneumoniae HS11286. To this end, apramycin (pDNTOOL-6) and hygromycin

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
68
resistance (pDNTOOL-6) version of pJTOOL-3 were created by using lambda RED recombination-based
allelic exchange.
To create pDNTOOL-6, PCR products of DN13/DN14 (LF, 503 bp) and DN15/DN16 (RF, 520 bp) both
amplified from pJTOOL-3 and DN17/DN17 (apramycin cassette from PKOBEG-apra, 839bp) were joined
by SOE-PCR to produce 1809 bp PCR product that was used as mutant allele in the replacement of cat in
pJTOOL-3. The 1809 bp SOE-PCR product (2-5 µg) was transformed into an 0.2 % arabinose-induced
competent E. coli cc18λpir carrying both the target (pJTOOL-3) and the lambda RED-recombination-based
(pKD46) plasmids and recovered in apramycin plate at 37℃. The grown colonies were then parallel patched
on chloramphenicol apramycin and ampicillin LB agar plates for loss of chloramphenicol and ampicillin.
Similarly, hygromycin version of pJTOOL-3 (pDNTOOL-7) was constructed by using PCR products of
DN13/DN14, DN15/DN16 and DN19/DN20 (hph, 1058 bp) to produce SOE-PCR product of 2022 bp. The
hygromycin SOE-PCR products (2-5 µg) was transformed into 0.2 % arabinose induced competent E. coli
cc18λpir (with pJTOOL-3 and pKD46) and recovered in hygromycin plate at 37℃. Colonies were then
parallel patched on chloramphenicol hygromycin and ampicillin LB agar plates for the loss of
chloramphenicol and ampicillin resistance but not hygromycin.
As usual, all the constructed lambda-pir dependent suicide plasmids (pDNTOOL-6, pDNTOOL-7,
pDNTOOL-8 and pDNTOOL-9) were verified by PCR, restriction digestion and sequencing (when needed)
relative to the parent plasmid, pJTOOOL-3.
2.4.2 Variety of Lambda RED plasmids with different antibiotic resistance cassette
To further enhance the use of lambda RED recombination tools for easy access and usage in a variety of
genetic manipulations in Gram-negative MDR bacteria, pDNTOOOL-10 to pDNTOOL-16, which are
different antibiotic resistance versions of pKD46 and pKOBEG-Apra were constructed.
First, a PCR product of primer-pair DN21/DN22 (sacB-cat, 2199 bp) from pJTOOL-3 was cloned
separately cloned into the inverse-PCR products of DN23/DN24 (10 kb from pKD46) or DN24/DN25 (6
kb from pKOBEG-apra) using PacI and ApaI sites, this gave rise to pDNTOOL-13 (see Figure 2.4a) or
pDNTOOL-16 (see Figure 2.4b) respectively. The individual ligation mixtures were then transformed into
E. coli DH5α and recovered in LA chloramphenicol plate at 30℃. Apramycin and ampicillin resistance-
encoding genes in PKOBEG-apra and PKD46 were successfully replaced with sacB-cat genes encoding
levansucrase for negative selection (only in the presence of sucrose) and chloramphenicol resistance.
Subsequently, these plasmids (pDNTOOL-13 and pDNTOOL-16) were PCR and restriction digest verified.
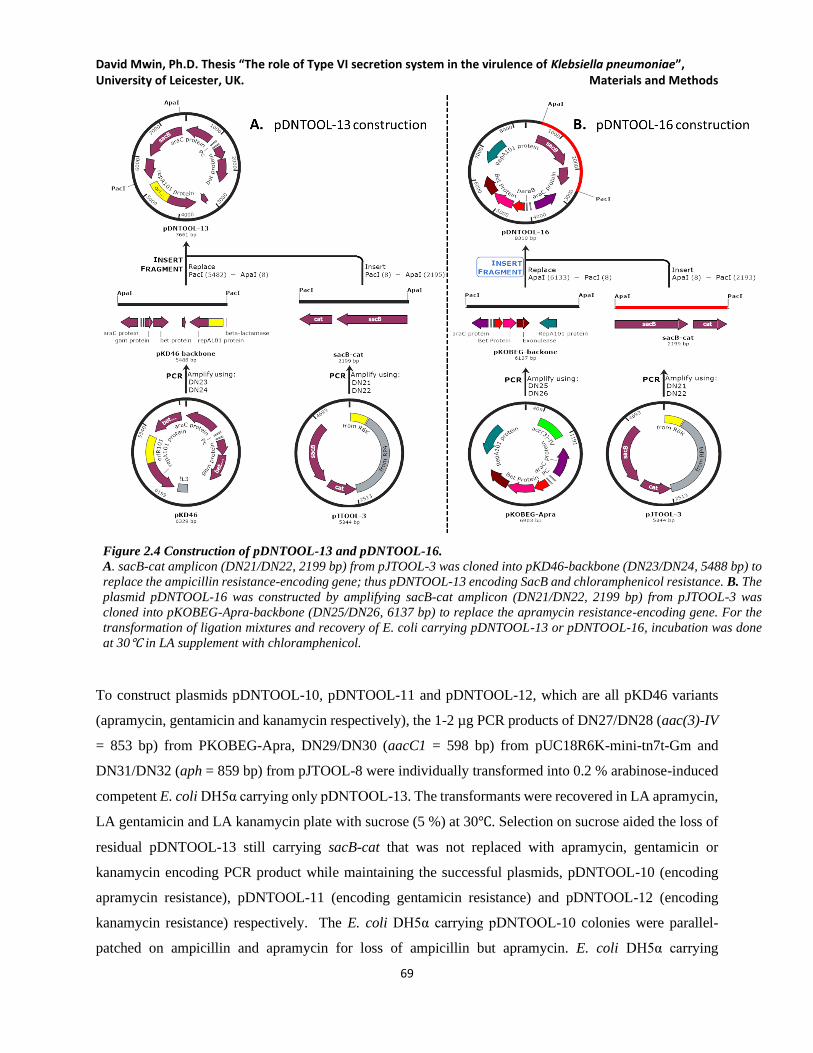
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
69
Figure 2.4 Construction of pDNTOOL-13 and pDNTOOL-16.
A. sacB-cat amplicon (DN21/DN22, 2199 bp) from pJTOOL-3 was cloned into pKD46-backbone (DN23/DN24, 5488 bp) to
replace the ampicillin resistance-encoding gene; thus pDNTOOL-13 encoding SacB and chloramphenicol resistance. B. The
plasmid pDNTOOL-16 was constructed by amplifying sacB-cat amplicon (DN21/DN22, 2199 bp) from pJTOOL-3 was
cloned into pKOBEG-Apra-backbone (DN25/DN26, 6137 bp) to replace the apramycin resistance-encoding gene. For the
transformation of ligation mixtures and recovery of E. coli carrying pDNTOOL-13 or pDNTOOL-16, incubation was done
at 30℃ in LA supplement with chloramphenicol.
To construct plasmids pDNTOOL-10, pDNTOOL-11 and pDNTOOL-12, which are all pKD46 variants
(apramycin, gentamicin and kanamycin respectively), the 1-2 µg PCR products of DN27/DN28 (aac(3)-IV
= 853 bp) from PKOBEG-Apra, DN29/DN30 (aacC1 = 598 bp) from pUC18R6K-mini-tn7t-Gm and
DN31/DN32 (aph = 859 bp) from pJTOOL-8 were individually transformed into 0.2 % arabinose-induced
competent E. coli DH5α carrying only pDNTOOL-13. The transformants were recovered in LA apramycin,
LA gentamicin and LA kanamycin plate with sucrose (5 %) at 30℃. Selection on sucrose aided the loss of
residual pDNTOOL-13 still carrying sacB-cat that was not replaced with apramycin, gentamicin or
kanamycin encoding PCR product while maintaining the successful plasmids, pDNTOOL-10 (encoding
apramycin resistance), pDNTOOL-11 (encoding gentamicin resistance) and pDNTOOL-12 (encoding
kanamycin resistance) respectively. The E. coli DH5α carrying pDNTOOL-10 colonies were parallel-
patched on ampicillin and apramycin for loss of ampicillin but apramycin. E. coli DH5α carrying

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
70
pDNTOOL-11 were parallel-patched on ampicillin and gentamicin for loss of ampicillin but not
gentamicin. Also, E. coli DH5α carrying pDNTOOL-11 were parallel-patched on ampicillin and kanamycin
for loss of ampicillin but not kanamycin.
For the construction of pDNTOOL-14, pDNTOOL-15 and pDNTOOL-17, 1-2 µg PCR products of
DN33/DN34 (hph, 1059 bp), DN35/DN36 (aph, 859 bp) and DN36/DN37 (aacC1, 598 bp) were
transformed into 0.2% arabinose-induced competent E. coli DH5α carrying only pDNTOOL-16. The
transformants were separately recovered in hygromycin (for pDNTOOL-14,), kanamycin (for pDNTOOL-
15) and gentamicin (for pDNTOOL-17) plates with 5 % sucrose at 30℃. Selection on sucrose aided the
loss of residual pDNTOOL-16 whose sacB-cat) was not replaced with hygromycin, kanamycin or
gentamicin while maintaining the successful plasmids (pDNTOOL-14, pDNTOOL-15 and pDNTOOL-17).
E. coli DH5α carrying pDNTOOL-14 colonies were parallel-patched on apramycin and hygromycin for the
loss of apramycin but not hygromycin. E. coli DH5α carrying pDNTOOL-15 were parallel-patched on
apramycin and kanamycin for loss of apramycin resistance, but not kanamycin resistance. Finally, E. coli
DH5α carrying pDNTOOL-17 were parallel-patched on apramycin and gentamicin for the loss of
apramycin but not gentamicin. All lambda RED variant plasmids were verified using restriction digestion
and PCR.
2.4.3 FRT-based antibiotic cassettes/ GFP plasmids
Generally, the selection of antibiotic resistance markers for genetic manipulation in MDR K. pneumoniae
strains (which was the case of K. pneumoniae HS11286) poses a significant difficulty. Hence, a successful
attempt was made to increase the pool of FRT-based antibiotic markers that can be repeatedly used in
creating a clean markerless gene knockout in most Gram-negative bacterial strains.
To create pDNTOOL-32, primer-pair DN39/DN40 (707 bp) was used to amplify cat gene from pJTOOL-
3 and SOE-PCR joined together with LF and RF from pUC18R6K-mini-tn7t-Gm using primer pairs
DN45/DN46 (617 bp) and DN47/DN48 (610 bp), respectively. The 1887 bp LF-cat-RF SOE-PCR product
(1-2 µg) was then transformed into 0.2 % arabinose-induced competent E. coli DH5α carrying target
plasmid pUC18R6K-mini-tn7t-Gm and pDNTOOL-15 (lambda red recombination variant) The
transformants were recovered in LB + chloramphenicol at 37℃. The grown colonies were then parallel-
patched on chloramphenicol, kanamycin and gentamicin LB agar plates for the loss of kanamycin and
gentamicin resistance but not chloramphenicol. The FRT-cat-FRT based plasmid, pDNTOOL-32, was
further verified by restriction digestion and PCR.
The plasmid pDNTOOL-31 (Figure 2.5a) was created by amplifying sacB-cat, (2199 bp) from pJTOOL-3
using primer-pair DN41/DN42 and Mlul/ Pacl-cloning it into an inverse-PCR product of 3796 bp amplicon

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
71
from pUC18R6K-mini-tn7t-Gm (amplified with DN43/DN44). The ligation mixture of sacB-cat and
pUC18R6K-mini-tn7t-Gm backbone which was transformed into competent E. coli DH5α resulted in the
replacement of the aacC1 gene in pUC18R6K-mini-tnt-Gm with sacB-cat genes. This plasmid,
pDNTOOL-31 (carrying FRT-sacB-cat-FRT) was subsequently used as a parent plasmid with negative
(sacB) and positive selection (cat) for the creation of other FRT-based antibiotic cassettes in the
pUC18R6K-mini-tn7t backbone. Plasmid pDNTOOL-31 was now used as parent plasmid in the creation
of pDNTOOL-1 (pUC18R6K-mini-tn7t-aac(3)-IV) pDNTOOL-33 (pUC18R6K-mini-tn7t-hph),
pDNTOOL-34 (pUC18R6K-mini-tn7t-hph-gfp) and pDNTOOL-35 (pUC18R6K-mini-tn7t-aac(3)-IV-
gfp).
To create plasmid pDNTOOL-1 carrying FRT-aac(3)-IV-FRT (Figure 2.5b), the amplicons of DN54/DN55
(aac(3)-IV, 836 bp) from pKOBEG-Apra, DN45/DN46 (LF, 617 bp) and DN47/DN48 (RF, 610 bp) from
pDNTOOL-31 were SOE-PCR joined together to create a mutant allele ( 2016 bp) targeting the replacement
of sacB-cat in pDNTOOL-31. The LF-aac(3)-IV-RF SOE-PCR product (2-5 µg) was transformed into 0.2
% arabinose-induced competent E. coli DH5α carrying target plasmid pDNTOOL-31 (FRT-sacB-cat-FRT
version of pUC18R6K-mini-tn7t) and pDNTOOL-15 (lambda red recombination variant). The
transformants were recovered on apramycin plate with sucrose (5 %) at 37 ℃. Colonies were parallel
patched for loss of chloramphenicol and kanamycin resistance but not apramycin. Restriction digest and
PCR were used to further verify the plasmid, pDNTOOL-1.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
72
Figure 2.5 Construction of pDNTOOL-31 and pDNTOOL-1.
A. Construction of pDNTOOL-31, carrying sacB-cat insert as a negative (sacB in the presence of sucrose) and positive
(chloramphenicol) selection markers during lambda RED mutagenesis. The backbone amplicon of pUC18R6K-mini-tn7t-
Gm without the aacC1 and sacB-cat amplicon were digested with MluI and PacI enzymes, ligated and transformed into E.
coli to give rise to pDNTOOL-31. B. Construction of pDNTOOL-1 was done by transforming a 0.2% arabinose induced
competent E. coli carrying pDNTOOL-31 and lambda RED plasmid pDNTOOL-15 with a 2 kb LF-aac(3)-IV-RF SOE-PCR
product and recovered on apramycin at 37℃. Similarly, pDNTOOL-33, pDNTOOL-34 and pDNTOOL-35 were all
constructed by transforming 0.2% arabinose induced competent E. coli carrying pDNTOOL-31 and pDNTOOL-15 with
SOE-PCR products (LF-hph-RF, LF-hph-gfp-RF and LF- aac(3)-VI-gfp-RF and transformants were recovered on LA plus
respective antibiotics at 37℃. Also, pDNTOOL-36, pDNTOOL-37 and pDNTOOL-38 construction were done by
transforming, the SOE-PCR products (LF-sacB-aacC1-RF, LF-sacB-hph-RF and LF-sacB-aac(3)-IV-RF) into an E. coli
carrying pDNTOOL-32 and pDNTOOL-15 and the transformants were recovered in the gentamicin, hygromycin and
apramycin without sucrose at 37℃.
Like pDNTOOL-1 construction, pDNTOOL-33 was constructed by first SOE-PCR joining the hph
amplicon DN49/DN50 (1046 bp) from pMQ300 to pDNTOOL-31 LF (DN45/DN46, 617 bp) and RF
(DN47/DN48, 610 bp) amplicons. The resultant SOE-PCR product (LF-hph-RF, 2226 bp), was transformed
into 0.2 % arabinose-induced competent E. coli DH5α carrying target plasmid pDNTOOL-31 (carrying
FRT-sacB-cat-FRT) and pDNTOOL-15 (lambda red recombination) and recovered in hygromycin LA
plate with sucrose (10%) at 37℃. Grown colonies were parallel-patched on hygromycin, chloramphenicol

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
73
LA plates. The plasmid was restriction-digest and PCR. In PDNTOOL-33, hph replaced the sacB-cat and
is flanked by two FRTs.
The fluorescent-tagged (GFP) pDNTOOL-34 was constructed from by first joining hph (DN49/DN51, 1043
bp) with gfp (DN52/DN53, 912 bp) amplicons amplified from pMQ300 and pIM438, respectively. The
hph-gfp SOE-PCR product (1935 bp) is further joined together with pDNTOOL-31 LF (DN45/DN46, 617
bp) and RF (DN47/DN48, 610 bp) amplicons to form a mutant allele (LF-hph-gfp-RF, 3115 bp) that was
used for the replacement of sacB-cat in pDNTOOL-31. The LF-hph-gfp-RF mutant allele was transformed
into 0.2 % arabinose-induced competent E. coli DH5α carrying target plasmid pDNTOOL-31 and
pDNTOOL-15 and recovered in hygromycin LA plate with sucrose (5 %) at 37℃. Colonies were verified
phenotypically by parallel-patching and fluorescent microscopy and genotypically by restriction digest and
PCR.
To construct pDNTOOL-35, SOE-PCR was used to join aac(3)-IV (DN54/DN56, 836 bp) and gfp
(DN52/DN53 912 bp) to yield 1725 bp PCR product in the first round. The second round of SOE-PCR was
performed using the 1.7 kb aac(3)-VI-gfp SOE-PCR product together with pDNTOOL-31 LF
(DN45/DN46, 617 bp) and RF (DN47/DN48, 610 bp) amplicons. The LF-aac(3)-IV-gfp- RF SOE-PCR
product (2905 bp) was then transformed into 0.2 % arabinose-induced competent E. coli DH5α carrying
target plasmid pDNTOOL-31 ( with FRT-sacB-cat-FRT) and pDNTOOL-15 (lambda red recombination)
The transformed were recovered in apramycin LA plate with sucrose (10%) at 37℃. Colonies were parallel-
patched on LA apramycin, chloramphenicol and kanamycin to ensure the loss of both kanamycin resistance
(encoded by temperature-sensitive plasmid pDNTOOL-15) and chloramphenicol (encoding by pDNTOOL-
31). Any colony with residual pDNTOOL-31 and pDNTOOL-15 were lost due to the negative sacB
selection and temperature-sensitive origin of replication (37-42℃), respectively
Three more negative-positive antibiotic selections marker plasmids flanked by FRTs (pDNTOOL-36,
pDNTOOL-37 and pDNTOOL-38) were created for other genetic manipulation purposes.
An FRT-sacB-aacC1-FRT based plasmid (pDNTOOL-36) was created by SOE-PCR joining sacB
(DN57/DN58, 1543 bp) and aacC1 (DN59/DN60, 578 bp) amplicons in the first round to give 2100 bp
product. The second round of SOE-PCR joining was doing by SOE-PCR joining LF (DN45/DN46, 617 bp
from pDNTOOL-32) and RF (DN47/DN48, 610 bp from pDNTOOL-32) amplicons with the PCR product
from the first round, this resulted in a final 3280 bp SOE-PCR product (LF-sacB-aacC1-RF). The LF-sacB-
aacC1-RF SOE-PCR product (1-2µg) was transformed into 0.2 % arabinose-induced competent E. coli
DH5α carrying target plasmid pDNTOOL-32 and pDNTOOL-15. The transformants were recovered in
gentamicin LA plates at 37℃ resulting in the lost chloramphenicol resistance as the cat gene was replaced

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
74
with the FRT-sacB-aacC1-FRT and no pDNTOOL-15. The colonies were parallel-patched on gentamicin,
chloramphenicol and kanamycin and the final plasmid, pDNTOOL-36, was verified by restriction digest
and PCR.
Like pDNTOOL-36 construction, FRT-sacB-hph-FRT based plasmid (pDNTOOL-37) and FRT-sacB-
aac(3)-IV-FRT based plasmid (pDNTOOL-38) were created by transforming the respective a 0.2 %
arabinose-induced competent E. coli DH5α (carrying pDNTOOL-32 and pDNTOOL-15) with 1-2µg SOE-
PCR products (LF-sacB-hph-RF and LF-sacB-aac(3)-IV-RF respectively). Four amplicons using primer-
pair DN61/DN62 (to amplify hph from pMQ300, 1043 bp), DN57/DN58 (sacB from pJTOOL-3, 1543 bp),
DN45/DN46 (LF from pDNTOOL-32, 617 bp) and DN47/DN48 (RF from pDNTOOL-32, 610 bp) were
joined together to produce 3745 bp LF-sacB-hph-RF mutant allele that was used for the construction of
pDNTOOL-37. For the construction of pDNTOOL-38, a 3535 bp SOE-PCR product (LF-sacB-aac(3)-IV-
LF) was produced from DN57/DN58, (sacB from pJTOOL-3, 1543 bp), DN63/DN64 (aac(3)-IV amplified
from pKOBEG-Apra, 833 bp ), DN45/DN46 (LF from pDNTOOL-32, 617 bp) and DN47/DN48 (RF from
pDNTOOL-32, 610 bp) for construction of pDNTOOL-38. Transformant colonies were parallel patched
on hygromycin or apramycin, chloramphenicol and kanamycin and the right plasmids were verified by PCR
and restriction digestion.
2.4.4 Flp recombinase- encoding plasmid construction
Flp-recombinase encoding plasmid is very important in the creation of markerless mutant when used in
combination with FRT-based antibiotic cassettes. This ensured the repeated used of a single antibiotic
cassette in the same MDR K. pneumoniae while creating multiple gene knockouts in that strain. To increase
the availability of antibiotic-resistant variants of Flp-recombinase encoding plasmids that mediate FRT
recombination, pDNTOOL-2, pDNTOOL-18, pDNTOOL-19, pDNTOOL-20 and pDNTOOL-21 were
constructed.
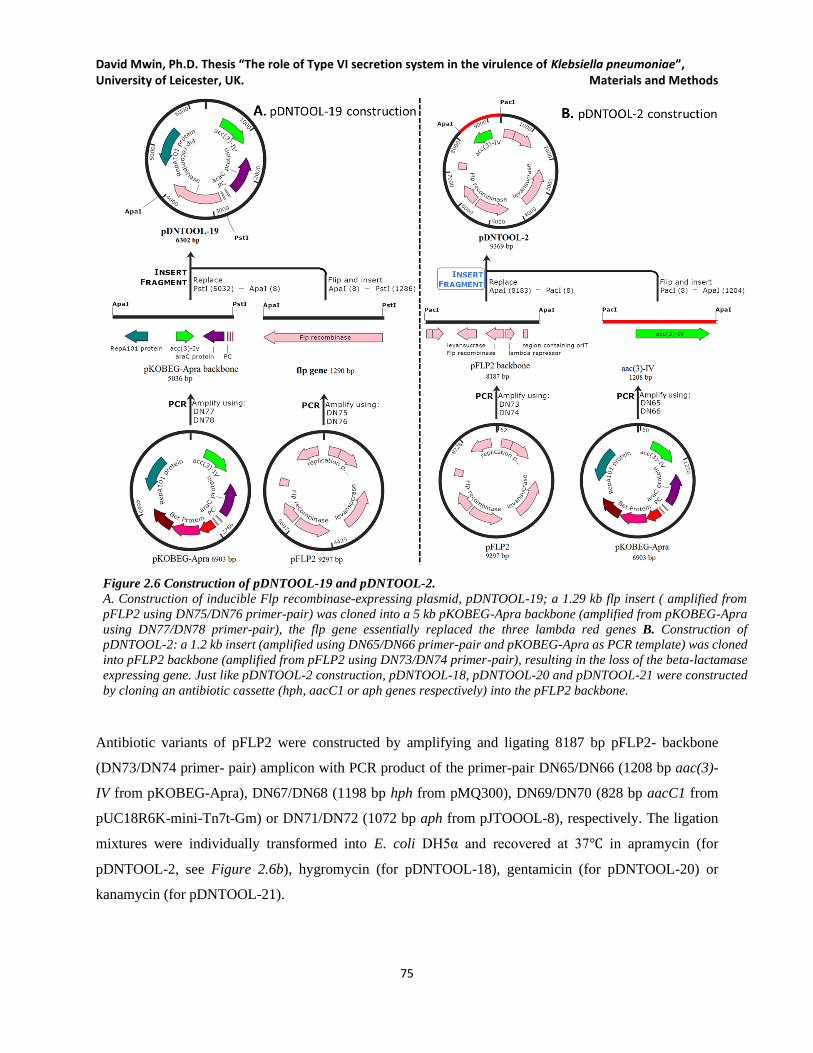
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
75
Figure 2.6 Construction of pDNTOOL-19 and pDNTOOL-2.
A. Construction of inducible Flp recombinase-expressing plasmid, pDNTOOL-19; a 1.29 kb flp insert ( amplified from
pFLP2 using DN75/DN76 primer-pair) was cloned into a 5 kb pKOBEG-Apra backbone (amplified from pKOBEG-Apra
using DN77/DN78 primer-pair), the flp gene essentially replaced the three lambda red genes B. Construction of
pDNTOOL-2: a 1.2 kb insert (amplified using DN65/DN66 primer-pair and pKOBEG-Apra as PCR template) was cloned
into pFLP2 backbone (amplified from pFLP2 using DN73/DN74 primer-pair), resulting in the loss of the beta-lactamase
expressing gene. Just like pDNTOOL-2 construction, pDNTOOL-18, pDNTOOL-20 and pDNTOOL-21 were constructed
by cloning an antibiotic cassette (hph, aacC1 or aph genes respectively) into the pFLP2 backbone.
Antibiotic variants of pFLP2 were constructed by amplifying and ligating 8187 bp pFLP2- backbone
(DN73/DN74 primer- pair) amplicon with PCR product of the primer-pair DN65/DN66 (1208 bp aac(3)-
IV from pKOBEG-Apra), DN67/DN68 (1198 bp hph from pMQ300), DN69/DN70 (828 bp aacC1 from
pUC18R6K-mini-Tn7t-Gm) or DN71/DN72 (1072 bp aph from pJTOOOL-8), respectively. The ligation
mixtures were individually transformed into E. coli DH5α and recovered at 37℃ in apramycin (for
pDNTOOL-2, see Figure 2.6b), hygromycin (for pDNTOOL-18), gentamicin (for pDNTOOL-20) or
kanamycin (for pDNTOOL-21).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
76
Also, arabinose inducing flippase expression plasmid (pDNTOOL-19, see Figure 2.6a) was constructed by
amplifying flippase encoding gene from pFLP2 and cloning it in place of the lambda red recombination
encoding genes in pKOBEG-Apra. Briefly, the PCR product of flp amplicon (1290 bp) using the primer-
pair DN75/DN76 and was PstI/ ApaI digested and ligated to the 5036 bp pKOBEG-backbone amplicon
(using primer-pair DN77/DN78). The ligation mix was transformed into E. coli DH5α and recovered at
30℃ on apramycin LA for pDNTOOL-19.
All the Flp recombinase-expressing plasmids (pDNTOOL-2, pDNTOOL-18, pDNTOOL-19, pDNTOOL-
20 and pDNTOOL-21) were verified by restriction digestion and PCR and the strains carrying the respective
plasmids were subsequently archived in BHI with 30 % glycerol at -80℃.
2.4.5 Cloning and inducible expression plasmids/ GFP plasmid Construction
Several expressing plasmids (from pDNTOOL-3, pDNTOOL-4, and pDNTOOL-22 to pDNTOOL-30)
were constructed using amplicons from the pWSK29 backbone, an inducible promoter from pKOBEG-
Apra, pKPHS6 and different antibiotic markers with or with GFP. First, pDNTOOL-22 was created by PCR
amplifying and ligating pWSK29 backbone (using primer-pair DN81/DN82, 4591 bp) and sacB-cat
amplicons (using primer-pair DN79/DN80, 2225 bp from pJTOOL-3). The ligation mix was then
transformed into E. coli DH5α. The transformants carrying pDNTOOL-22 (see Figure 2.7) were recovered
on chloramphenicol before being parallel-patched on chloramphenicol and ampicillin. Colonies with only
chloramphenicol resistance were considered for molecular confirmation of pDNTOOL-22 (by plasmid
extraction, restriction digestion a PCR)
To construct pDNTOOL-3 (as shown in Figure 2.7), pDNTOOL-4, pDNTOOL-23 and pDNTOOL-24
(apramycin, hygromycin, gentamicin and kanamycin versions of pWSK29), the PCR products of aac(3)-
IV (DN83/DN84, 849 bp from pDNTOOL-1), hph (DN85/DN86, 1059 bp from pMQ300), aacC1
(DN87/DN88, 594 bp from pUC18R6K-mini-tn7t-Gm) and aph (DN89/DN90, 855 bp from pJTOOL-8)
were individually transformed into 0.2% arabinose induced competent E. coli DH5α carrying both
pDNTOOL-22 and pKD46. The transformants were recovered in their respective antibiotic LA plates at
37℃ and patched on chloramphenicol, ampicillin and the associated antibiotic plates (apramycin,
hygromycin, gentamicin or kanamycin). In each case, only colonies that were not resistant to
chloramphenicol and ampicillin but resistant to apramycin (pDNTOOL-3), hygromycin (pDNTOOL-4),
gentamicin (pDNTOOL-23) or kanamycin (pDNTOOL-24) were considered for restriction-digest and PCR
confirmation.
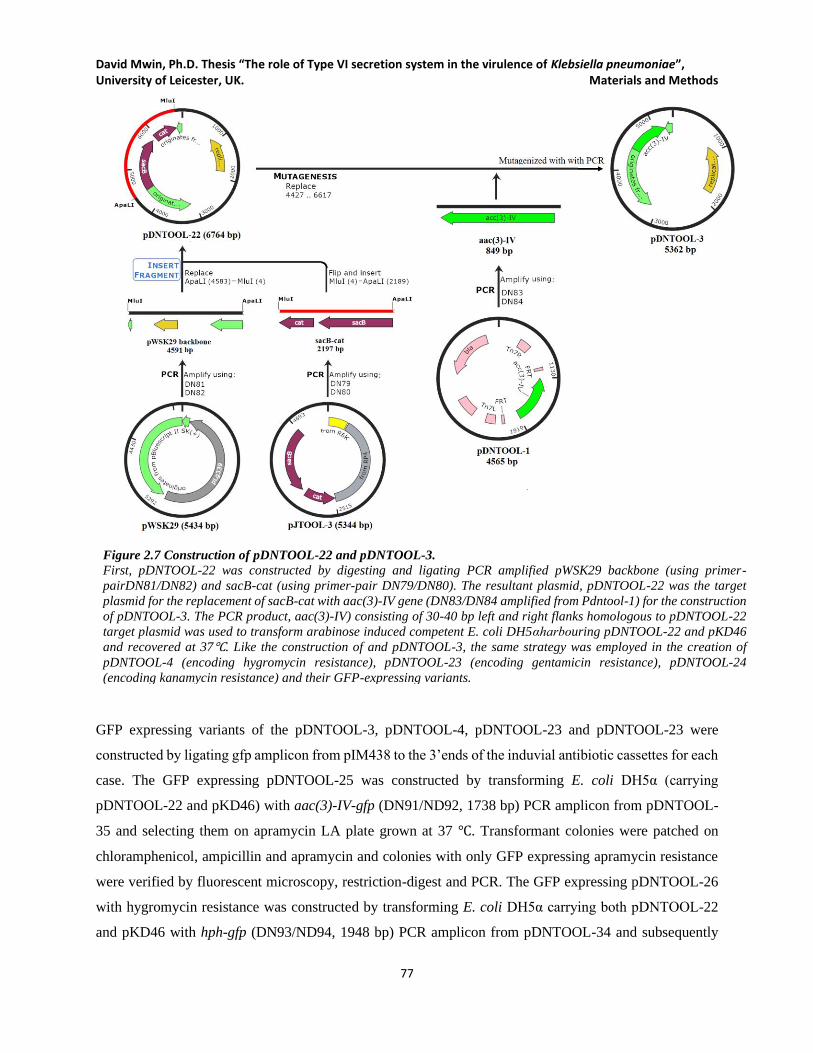
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
77
Figure 2.7 Construction of pDNTOOL-22 and pDNTOOL-3.
First, pDNTOOL-22 was constructed by digesting and ligating PCR amplified pWSK29 backbone (using primer-
pairDN81/DN82) and sacB-cat (using primer-pair DN79/DN80). The resultant plasmid, pDNTOOL-22 was the target
plasmid for the replacement of sacB-cat with aac(3)-IV gene (DN83/DN84 amplified from Pdntool-1) for the construction
of pDNTOOL-3. The PCR product, aac(3)-IV) consisting of 30-40 bp left and right flanks homologous to pDNTOOL-22
target plasmid was used to transform arabinose induced competent E. coli DH5αharbouring pDNTOOL-22 and pKD46
and recovered at 37℃. Like the construction of and pDNTOOL-3, the same strategy was employed in the creation of
pDNTOOL-4 (encoding hygromycin resistance), pDNTOOL-23 (encoding gentamicin resistance), pDNTOOL-24
(encoding kanamycin resistance) and their GFP-expressing variants.
GFP expressing variants of the pDNTOOL-3, pDNTOOL-4, pDNTOOL-23 and pDNTOOL-23 were
constructed by ligating gfp amplicon from pIM438 to the 3’ends of the induvial antibiotic cassettes for each
case. The GFP expressing pDNTOOL-25 was constructed by transforming E. coli DH5α (carrying
pDNTOOL-22 and pKD46) with aac(3)-IV-gfp (DN91/ND92, 1738 bp) PCR amplicon from pDNTOOL-
35 and selecting them on apramycin LA plate grown at 37 ℃. Transformant colonies were patched on
chloramphenicol, ampicillin and apramycin and colonies with only GFP expressing apramycin resistance
were verified by fluorescent microscopy, restriction-digest and PCR. The GFP expressing pDNTOOL-26
with hygromycin resistance was constructed by transforming E. coli DH5α carrying both pDNTOOL-22
and pKD46 with hph-gfp (DN93/ND94, 1948 bp) PCR amplicon from pDNTOOL-34 and subsequently

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
78
selecting the transformants hygromycin LA plate at 37 ℃. Transformant colonies were patched on
chloramphenicol, ampicillin and hygromycin and colonies that were only apramycin resistant and GFP
expressing under fluorescent microscopy were verified by restriction digest and PCR.
The GFP expressing pDNTOOL-27 was constructed by transforming E. coli DH5α carrying both
pDNTOOL-22 and pKD46 with aacC1-gfp SOE-PCR product (1483 bp) of aacC1 (DN95/DN96 583 bp
from pUC18R6K-mini-tn7t-Gm) and gfp amplicon (DN52/DN97, 919 bp from pIM438). Transformant
colonies were patched on chloramphenicol, ampicillin and apramycin and colonies with only apramycin
resistance and GFP expressing under fluorescent microscopy were verified by restriction digest and PCR.
While GFP expressing pDNTOOL-28 was constructed by transforming E. coli DH5α carrying both
pDNTOOL-22 and pKD46 with aph-gfp, (1744 bp) SOE-PCR amplicon. The SOE-PCR product was done
by joining aph amplicon (DN99/DN100, 844 bp from pJTOOL-8) and gfp amplicon (DN52/DN97, 919 bp
from pIM438). The transformant colonies were parallel-patched on chloramphenicol, ampicillin and
apramycin and only GFP expressing kanamycin-resistant colonies were verified using fluorescent
microscopy, restriction digest and PCR.
Finally, two additional protein expression plasmids (pDNTOOL-29 and pDNTOOL-30) with arabinose
inducible promoters with or without GFP at the MCS site, were constructed for future protein expression,
GFP tagging and complementation studies. The PCR amplicons of DN101/DN102 (1334 bp from pKPHS6)
and DN103/DN104 (1287 bp from pKD46) were joined together to produce a 2586 bp SOE-PCR amplicon
(pKPHS6 backbone + arabinose-inducible promoter) with NotI and KpnI sites. A transcriptional terminator
sequence was amplified from pKD46 with DN106/DN107 (308 bp) and then used as a template in second
PCR re-amplification using DN105/DN107 (329 bp). Also, aacC1 was amplified from pUC18R6K-mini-
Tn7t-Gm using DN108/DN109 (854bp). The final transcriptional terminator amplicon (332 bp) and the
aacC1 amplicon were joined together to produce 1140 bp SOE-PCR product with NotI and KpnI sites. The
two SOE-PCR products (2586 bp and 1144 bp) were digested (NotI and KpnI) and ligated and subsequently
transformed into E. coli DH5α at 37 ℃. The transformant carrying pDNTOOL-29 (3.7 kb as shown in
Figure 2.8) were recovered on gentamicin LA plate and verified by restriction digest and PCR.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
79
Figure 2.8 Construction of pDNTOOL-29 and pDNTOOL-30.
The backbone of pKPHS6 was amplified from K. pneumoniae HS11286 was joined by SOE-PCR with araC-ParaB amplicon
from pKD46 to yield a 2.6 kb SOE-PCR product. Also, aacC1 amplicon from Puc18r6k-mini-tn7t-Gm was SOE-PCR joined
with a tL3-mcs amplicon, resulting in a 1140 bp SOE-PCR product. The 2.6 kb amplicon and 1.1 kb amplicon were digested
with NotI and KpnI and ligated to produce a 3.7 kb plasmid. The ligation mixture was transformed into E. coli DH5α. To
confirm the arabinose inducibility of pDNTOOL-29, a gfp amplicon from pIM438 was also cloned inframe into the multiple
cloning site (at PacI/SphI sites) resulting in pDNTOOL-30
To confirm the arabinose inducibility of the promoter for gene expression and as well create protein-GFP
tagging tool, pDNTOOL-30 (see Figure 2.8) was constructed by re-amplifying a 752 bp gfp amplicon
(DN111/DN112, from pIM438) with primer-pair DN110/DN112 (773 bp) and the final gfp amplicon was
digest (SphI/ PacI) and ligated to form pDNTOOL-29. The ligation mix was transformed into E. coli DH5α
at 37 ℃, and the transformants were recovered in gentamicin LA and verified by fluorescent microscopy,
restriction-digestion and PCR.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
80
2.5 Genetic manipulation
2.5.1 Preparation of Competent bacterial cells
Generally, all plasmid and linear DNA were transformed into chemically competent or electrocompetent
bacterial cells. While the transformations of DNA into E. coli were primarily done using both chemically
and electrocompetent cells, K. pneumoniae transformation was only done in electrocompetent cells to
maximize the transformation efficiency.
2.5.1.1 Chemically competent E. coli preparations and transformation
Chemically competent E. coli DH5α, CC118-λpir and S17-λpir were prepared by inoculating 5 ml LB +
appropriate antibiotics /additional supplements (e.g. L-arabinose) if required in a 20 ml tube at 37℃ 200
rpm overnight. E. coli carrying temperature-sensitive origin of replication such pKD46and pKOBEG-Apra
derivatives were grown at 30℃ to avoid loss of the plasmid. The overnight culture was subcultured (1:100
ration) into fresh sterile 1 L LB broth with appropriate supplements or antibiotics if required. The cells were
then grown in the proper temperature (30℃ or 37℃) with shaking at 200 rpm until they reached OD600nm of
0.3 to 0.4. The cells were harvested (3000 ×g 10 minutes at 4℃) and gently washed three times with 50 ml
ice-cold 100 mM MgCl2 (Sigma Aldrich, UK) with intermittent ice incubation for 10 minutes. The final
cell pellet was resuspended in 5 ml ice-cold, sterile 100 mM CaCl2 (Sigma, UK) in 15 % (w/v) glycerol
(Fisher, UK) and 50 µl aliquots were stored in -80℃ until needed.
To transform the chemically competent E. coli using heat shock method, DNA ligation mixture (1-4 µl),
plasmid (10-100 ng) or linear DNA (50 ng– 500 ng) was gently mixed with 50 µl aliquot of thawed
chemically competent E. coli on ice. The DNA-bacterial mixture was ice-incubated for 20 minutes before
placing the tube of the DNA-bacteria mixture in a heat block at 42℃ for 45 seconds and immediately
returned on the ice. After 2 minutes of ice-incubation, 950 µl of Super Optimal broth with catabolite
repression (SOC) broth was added to the DNA-bacteria mixture and incubated at 30 or 37 with shaking
(200 rpm) for 1.5 h. The transformant cells were then recovered in LA plates containing the appropriate
antibiotics and or supplements.
2.5.1.2 Electrocompetent cells preparation and transformation
To prepare electrocompetent E. coli or K. pneumoniae, a single colony of the bacteria was inoculated 5 ml
LB + appropriate antibiotics /additional supplements (e.g. L-arabinose) if required in a 20 ml tube at 30℃

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
81
or 37℃ 200 rpm overnight. E. coli or K. pneumoniae carrying temperature-sensitive origin of replication
such pKD46 and pKOBEG-Apra derivatives were grown at 30℃ to avoid loss of the plasmid. The overnight
bacterial culture was subcultured (1:100 ration) into fresh sterile LB broth with appropriate supplements/
antibiotics if required and grown until cells reached OD600nm of 0.3 to 0.4 The cells were pelleted (3000 ×g
10 minutes at 4℃) and washed three times with successively smaller volumes (100 ml, 50 ml and 25 ml)
of ice-cold 10 % (v/v) glycerol. After the final wash, the cells were resuspended in 4 -10 µl of ice-cold 10
% (v/v) glycerol and aliquoted 50 µl aliquots were stored at -80℃ until needed. EDTA (0.7 mM) was added
to growth medium when higher competency was required for K. pneumoniae.
The bacteria (E. coli or K. pneumoniae) were transformed in 0.2 cm cuvettes using Bio-Rad Gene Pulser
system (Bio-Rad) at the setting of 2.5 kV/cm, 25 µF and 200 Ω. The transformants were recovered in 500-
950 l of SOC immediately after electroporation and grown for 1 h. The electroporated cells were then plated
on LA plates with the appropriate antibiotics/ supplements when required at 30℃ 0r 37℃.
2.5.2 Suicide vector-based allelic exchange
E. coli S17-1λpir was used in conjugation as a donor strain for the mobilization of suicide vectors into K.
pneumoniae NTUH-K2044 for T6SS1 and T6SS3 deletion. E. coli S17-1λpir encoded broad-host-range tra
(transfer) genes of RP4 conjugation system which enable the successful transfer of the mobilization of
vectors with RP4 plasmid mobilization (mob RP4) region. Briefly, E. coli S17-1λpir carrying suicide vector
pDNTOOL-8 or pDNTOOL-9, containing the respective T6SS1 or T6SS3 mutant alleles were grown
overnight in LB plus gentamicin at 37℃, 200 rpm. Also, the recipient K. pneumoniae strain was
simultaneously grown in LB only overnight at 37, rpm. The overnight culture of both bacteria was then
subcultured (1:100) in fresh LB medium with (1:50 for E. coli S17-1λpir carrying pDNTOOL-8 or
pDNTOOL-9) or without (1:100 for K. pneumoniae NTUH-K2044) gentamicin antibiotic and grown to
OD600nm 0.6 - 0.8. A 1:1 ratio (1 ml: 1ml) of the donor to recipient mixture was pelleted and resuspended in
20 µl of 10 mM MgSO4 and aseptically spotted on 2 cm by 2 cm nitrocellulose filter paper (0.45 m pore
size) on a pre-warmed LA plate and incubated at 37℃ overnight. Also, a 1:1 ratio of the donor to donor and
recipient to the recipient were included as necessary controls. The overnight matings on the nitrocellulose
paper were resuspended in 1.5 ml of 10 mM MgSO4, and dilutions were plated on M9 minimal medium
plus gentamicin agar plates and incubated at 37℃ overnight. Single colonies were parallel-patched on LA
only, LA plus gentamicin and LA plus chloramphenicol, to screen for the integration of PDNTOOL-8 or
pDNTOOL-9. The single crossover, SCO (merodiploid) were colony-PCR confirmed for the vector
integration using primer-pair GmF/ DN6 and GmR/DN5 for pDNTOOL-8 integration and primer-pair

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
82
GmF/DN11 and GmR/DN12 for pDNTOOL-9 integration. Also, vector backbone and antibiotic cassette
were confirmed using primer-pair DN21/DN22 and GmF/GmR. A confirmed SCO (merodiploid)colony
was grown in LB and the appropriate dilutions were plated on LA plus 10 % sucrose at 25℃ overnight. The
recovered colonies were then parallel patched onto LA plus gentamicin and LA plus chloramphenicol at
37℃ overnight. Colonies resistant to gentamicin but sensitive to chloramphenicol were selected a potential
T6SS1 or T6SS6 mutants (double crossover, DCOs). Few DCO colonies were chosen for PCR confirmation
using primer-pair DN5/DN6 for ∆T1:aacC1 or DN11/DN12 for ∆T3:aacC1 and the presence of the
gentamicin cassette using GmF/GmR in the mutants. The gentamicin cassette was mapped onto the targeted
locus by using GmF/ DN6 or GmR/DN5 for ∆T1:aacC1, and primer-pair GmF/DN11 or GmR/DN12 for
∆T3:aacC1. Also, PCR to confirm the absence of the vector backbone in the mutants was performed using
DN21/DN22. Only T6SS1 deletion was successful in K. pneumoniae NTUH-K2044 but not T6SS3, and
lambda Red recombination-based allelic exchange in the stain proved to be difficult because it's high
mucoviscosity and low electroporation efficiency.
2.5.3 Lambda RED recombination-based allelic exchange for KPHST6SS mutation
Lambda Red recombination-based allelic exchange uses overexpressed phage λ Red recombinase system
(γ, β, exo) in bacteria to enhance the homologous exchange linear DNA fragments (mostly SOE-PCR
generated mutant alleles with antibiotic cassette) with bacteria chromosome or plasmid ((Davidson and
Szybalski, 1971; Murphy, 1998). All T6SS gene cluster knockout (such as ∆T1:hph ∆T3:hph and
∆T1∆T3:hph) in K. pneumoniae HS11286 were created via lambda Red recombination-based allelic
exchange using T6SS1 and T6SS3 mutant alleles.
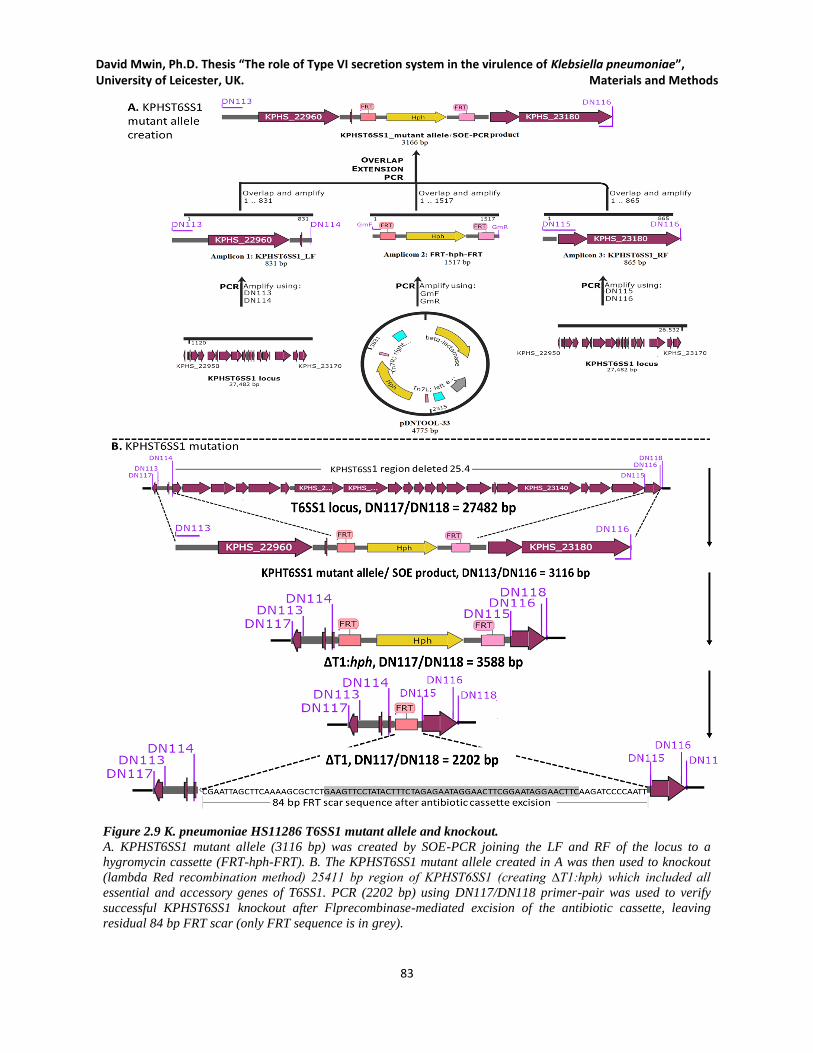
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
83
Figure 2.9 K. pneumoniae HS11286 T6SS1 mutant allele and knockout.
A. KPHST6SS1 mutant allele (3116 bp) was created by SOE-PCR joining the LF and RF of the locus to a
hygromycin cassette (FRT-hph-FRT). B. The KPHST6SS1 mutant allele created in A was then used to knockout
(lambda Red recombination method) 25411 bp region of KPHST6SS1 (creating ∆T1:hph) which included all
essential and accessory genes of T6SS1. PCR (2202 bp) using DN117/DN118 primer-pair was used to verify
successful KPHST6SS1 knockout after Flprecombinase-mediated excision of the antibiotic cassette, leaving
residual 84 bp FRT scar (only FRT sequence is in grey).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
84
Both T6SS single and double knockouts were all done in antibiotic resistance ‘disarmed background’ of
the originally K. pneumoniae HS11286 strain, see published data (Bi et al., 2015), represented in this work
as DN2. To create ∆T1:hph, ∆T3:hph and ∆T1∆T3:hph in DN2, the lambda Red recombination-based
plasmid (pKOBEG-Apra or its derivatives) was initially transformed into in DN2 and recovered on LA
apramycin or the appropriate antibiotics at 30℃. The presence of the pKOBEG-Apra was confirmed by
colony PCR using the primer-pair DN125/DN126 (1949 bp). A single colony of PCR-confirmed DN2
carrying pKOBEG-Apra was inoculated in 5 ml LB with apramycin and grown at 30℃ overnight. A 1:100
subculture of the overnight DN2/pKOBEG-Apra culture was made in fresh LB with antibiotics and 0.2 %
arabinose. The culture was grown to OD600nm 0.8 - 1.0 before being harvested for electrocompetent
DN2/pKOBEG-Apra preparation. T6SS mutant alleles were constructed by SOE-PCR joining T6SS LF,
hygromycin cassette and T6SS RF to produce a 3.5 kb T6SS1 or 3.2 kb T6SS3 mutant alleles (Figure 2.9a
and Figure 2.10a). The 0.5 - 1 µg of the SOE-PCR joined T6SS1, or T6SS3 mutant allele was then
electroporated into the arabinose induced electrocompetent DN2/pKOBEG-Apra K. pneumoniae. The
transformants were recovered in 950 µl of SOC for 1.5 h at 37℃ and then plated onto the LA plus
hygromycin at 37℃.
To create a T6SS1 and T6SS3 double cluster knockout (∆T1∆T3:hph), an arabinose-induced competent
markerless ∆T1 strain carrying pKOBEG-Apra was electroporated with T6SS3 mutant allele (0.5-1µg), and
the transformants were selected at 37℃ overnight on LA hygromycin after a brief recovery in SOC at 37℃
for 1.5 h. Colonies were parallel-patched on LA plus hygromycin and LA plus apramycin overnight grown
at 37℃. Only hygromycin-resistant colonies were PCR verified for the expected knockout (DN117/DN118,
3588 bp for ∆T1 and DN123/124, 3417 bp for ∆T3) and the absence of pKOBEG-Apra (DN125/DN126,
1949 bp).

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
85
Figure 2.10 K. pneumoniae HS11286 T6SS3 mutant allele creation and knockout.
A. KPHST6SS3 mutant allele (3209 bp) was created by SOE-PCR joining the LF and RF of the locus to a hygromycin cassette
(FRT-hph-FRT). B. The KPHST6SS3 mutant allele generated in step A was then used to knockout 30509 bp region of
KPHST6SS3, which included all essential and accessory T6SS3 genes. PCR (1984 bp) using DN123/DN124 primer-pair was
used to verify successful KPHST6SS3 knockout after Flprecombinase-mediated excision of the antibiotic cassette by FRTs
recombining and leaving residual 84 bp FRT scar (only FRT sequence in grey only for FRT) was observed. A markerless
∆T1 was used for KPHST6SS1 and KPHST6SS3 double knockout (∆T1∆T3) by electroporating the arabinose induced strain
(∆T1carrying lambda red plasmid) with KPHST6SS3 mutant allele. Transformants were selected on LA hygromycin plate at
37℃. Just like the excision of the antibiotic marker in the single mutants (∆T1 and ∆T3), the antibiotic cassette in the double
mutant was also excised.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
86
Similarly, lambda Red recombination allelic exchange was also primarily employed in the creation of many
genetic tools as described earlier in this chapter. Briefly, a PCR amplified mutant allele was used to replace
the target gene on a plasmid in E. coli strain also carrying a lambda Red recombination plasmid derivative.
Briefly, E. coli carrying lambda red recombination plasmid and target plasmid were grown in the 5ml LB
and appropriate antibiotics overnight at 30℃ overnight. A 1:50 ratio subculture was made in fresh LB with
appropriate antibiotics supplemented 0.2 % arabinose and grown until OD600nm 0.6-1.0 where they were
harvested for electrocompetent cell preparation. The electrocompetent E. coli was then electroporated either
PCR amplified sacB-cat (negative and positive selection markers) or the appropriate SOE-PCR amplified
linear DNA/ mutant allele. The transformants were selected on the appropriate LA antibiotic plates per
temperature requirements overnight and subsequently parallel-patched on the right antibiotic plates.
2.5.4 Flp-mediated FRT recombination
All the antibiotic cassettes used in the construction of T6SS1 and T6SS3 mutant allele for the T6SS cluster
knock out in K. pneumoniae NTUH-K2044 and K. pneumoniae HS11286, carry two FRT sites flanking the
antibiotic cassette (FRT-antibiotic cassette-FRT). The use of Flp recombinase encoding plasmid to excise
the antibiotic cassette using site-specific recombination of the two FRTs was necessary for the creation of
markerless mutant or double gene knockout for this study.
To excise antibiotic cassette (FRT-aacC1-FRT and FRT-hph-FRT) mediated by Flp-FRT recombination, a
Flp recombinase encoding plasmid, pDNTOOL-2 was electroporated into competent K. pneumoniae T6SS
mutants (K. pneumoniae NTUH-K2044∆T1:aacC1, K. pneumoniae HS11286 ∆T1:hph, K. pneumoniae
HS11286 ∆T3:hph and K. pneumoniae HS11286 ∆T1∆T3:hph) and recovered in LA apramycin at 37℃
overnight. The presence on the pDNTOOL-2 plasmid in the bacteria was PCR-confirmed using
DN129/DN130 (2062 bp). Obtained colonies were phenotypically screened by parallel-patched on LA plus
apramycin, LA plus gentamicin (K. pneumoniae NTUH-K2044∆T1) and LA plus hygromycin (for K.
pneumoniae HS11286 ∆T1:hph, K. pneumoniae HS11286∆T3:hph and K. pneumoniae HS11286
∆T1∆T3:hph) for hph cassette excision. Colonies resistant to apramycin but not gentamicin and hygromycin
(in each case) were considered for PCR verification of loss of aacC1 with one FRT and hph with one FRT
using mutant screening primers and antibiotic cassette primers. The Flp recombinase encoding plasmid was
kicked out by sucrose counterselection by growing bacterial on LA 5% sucrose at 37℃ overnight. Colonies
were parallel-patched, and PCR confirmed (DN) for the loss of the antibiotic cassette and the Flp
recombinase encoding plasmid using T6SS mutant screening primer-pairs, GmF/GmR and DN129/DN130
primer-pair (see Appendix 1). PCR amplicons across the excised region (using mutant screening primers)
was sequenced for residual FRT sequence analysis.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
87
As a result of successful aacC1 or hph excision, an inframe residual FRT scar (Figure 2.9b and Figure
2.10b) encoding a non-functional protein was created in the bacterial mutant chromosome for each
antibiotic cassette excision using an Flp recombinase encoding plasmid (pDNTOOL-2).
2.5.5 Sucrose counterselection
Sucrose counter selection is a negative selection method performed when needed to get rid of plasmids
carrying the sacB gene (encoding for levansucrase in the presence of sucrose which is lethal to the bacteria).
Briefly, after Flp-FRT excision, appropriate dilutions of K. pneumoniae mutants carrying the Flp
recombinase encoding plasmids (pDNTOOL-2) were grown on LA plus 5 % sucrose at 37℃ overnight.
The colonies were then patched on LA and LA plus gentamicin or hygromycin to screen for the loss of the
Flp recombinase encoding plasmid (pDNTOOL-2). Colonies that were gentamicin or hygromycin sensitive
but grew well on LA only were PCR screen for the loss of pDNTOOL-2 using primer-pair DN129/130
(2062 bp) The gentamicin or hygromycin sensitive K. pneumoniae T6SS mutants were grown on LA and
achieved in BHI with 30 glycerol at -80℃.
Also, sucrose counterselection was performed during lambda Red recombination-mediated plasmid
construction. Briefly, arabinose induced competent E. coli strains carrying both the lambda Red plasmids
(pKD46 or other derivatives) and sacB-cat derivative plasmid (pDNTOOL-13, pDNTOOL-16, pDNTOOL-
22 or pDNTOOL-31) were transformed with the appropriate SOE-PCR mutant allele and selected on LA
with 5 % sucrose and antibiotics at 37℃ overnight. This aided the replacement of the sacB-cat region on
the plasmids or loss of residual sacB-cat carrying plasmid and pKD46. To the best of my knowledge, this
is the first time such a novel plasmid construction strategy (without restriction or ligation) is reported. This
strategy was very efficient and reduced the difficulty of restriction-ligation as restriction enzymes sites
selection can be very difficult in certain plasmid construction. This method could be extended to
chromosomal complementation of genes when well optimized.
2.6 Other methods
2.6.1 Biofilm formation
2.6.1.1 Microplate-based biofilm assay
Modified microtitre plate-based method (O'toole and Kolter, 1998) was used to perform biofilm assays.
Briefly, overnight cultures (in LB for 16 h at 37°C, 200 rpm) of K. pneumoniae strains were subcultured
(1:100) into 100 µl LB in 96-well polystyrene (NUNC) microtitre plates and subsequently incubated at
30℃ or 37℃ for 24 h to 72 h without shaking. After the appropriate time, the bacterial cultures inside the

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
88
wells were discarded, and the plate gently washed once with 200µl of dH2O followed by the addition of
125 µl of 0.1 % crystal violet for 10 minutes. The crystal violet was discarded, and the microtitre plate was
washed twice with dH2O (200 µl). After thorough drying, 200 µl of 95 % ethanol was added to each well
and gently vortexed. The absorbance was measured at 595 nm (BioRad Model 680 Microplate reader).
Biofilm data was analyzed from three independent experiments, and each strain had eight well repeats for
each experiment
2.6.1.2 Coverslip based bacterial biofilm-like association
To perform bacterial association with a microscopic coverslip, a 1:100 subculture of overnight culture (in
LB for 16 h at 37°C, 200 rpm) of each K. pneumoniae strains was made into fresh sterile 3ml LB in 6-well
plates containing coverslips. The 6-well plates containing the coverslips and bacterial cultures were
incubated at 30℃ or 37℃ for 24 h or 48 h without shaking. After the set time, coverslips were carefully
washed in a new 6-well plate containing 5ml dH20. The coverslips were stained with 3 ml crystal violet
(0.1 %) for 10 minutes and washed twice with 5 ml dH20. The coverslips were carefully mounted on
microscopic slides and visualize on the light microscope.
2.6.2 Growth and cell viability
2.6.2.1 Bacterial growth curves.
Both bacterial growth measurement in 96 microtitre plate and CFU were employed to assess the growth of
the K. pneumoniae strains. Briefly, 5 ml of LB was inoculated with a colony of K. pneumoniae strains and
incubated 37℃, 200 rpm for overnight. A 1:100 subcultures (37oC at 200rpm) of each strain and grown
until OD600nm 0.6. The bacterial cultures were then normalized to OD600nm 0.06 in 150 µl of LB in 96
microtitre plate (NUNC). The plates with the cultures were immediately placed into a pre-set Varioskan®
flash spectral scanning multimode reader with continuous shaking at 37°C for 24 h. Also, dilutions of each
initial inoculum (at OD600nm 0.06) were plated on LA and incubated at 37℃ overnight to ensure that all the
inoculum gave rise to similar CFUs. The machine measured the absorbance (600 nm) of the cultures in the
microtiter plates every 10 minutes, and the data was retrieved after 24 h for analysis using GraphPad Prism.
Each strain for the growth curves had three biological repeats and three experimental repeats. Also, analysis
of growth fitness of bacterial cell with various plasmid constructs was examined using growth curves, and
inducers such as IPTG were added where required.
Also, growth assessment by CFU was done by making 1:100 subculture of K. pneumoniae overnight
cultures were grown until OD600nm 0.6 and normal to OD600nm 0.06. 100 µl of the normalised bacterial
cultures (OD600nm 0.06) were added to fresh sterile LB (20 ml) in a flask and incubated at 37℃ for 24 h
with shaking (200 rpm). 100 µl of the bacterial cultures were taken from each flask every 2 h, and the

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
89
appropriate dilutions were plated on LA and incubated overnight at 37℃ for colonies. The CFU were
counted and compared to the data obtained from Varioskan® flash spectral scanning multimode reader for
microtiter plate.
2.6.3 Antibiotic susceptibility testing
Before using bacterial strains for experiment or when needed, the antibiotic profile of the K. pneumoniae
strains was established using minimum inhibition concentration (MIC) and E-test strips diffusion methods.
After the disarming of the antibiotic resistance in original K. pneumoniae HS11286, the antibiotic profile
for the mutant was necessary to ensure that the strain was safe to use for downstream experiments such as
T6SS studies. The antibiotic susceptibility data (CLSI and BAC) obtained was used for antibiotic marker
selection for mutant creation or plasmid transformation and selection. Briefly, 2-4 K. pneumoniae colonies
from a fresh overnight LA plate cultures were picked and homogenously resuspended in 0.9 % NaCl. The
turbidity of the bacterial suspensions was adjusted to 0.5 McFarland (0.08-0.13 at OD625nm) containing 1 ×
108 CFU/ml and used with 15 minutes. With the aid of a sterile cotton swab, a semi-confluent lawn of the
bacteria was created on pre-warmed Mueller-Hinton agar (MHA) plates and allowed for 10 minutes to dry.
The E-test strips for various antibiotics were aseptically placed on the lawn, and the plates were then
incubated at 37℃ for 20 h. The results for the E-test strips (µg/ml) were read the following day from the
scale on the Etest strip where the symmetrical inhibition ellipse edge intercepts the E-test strip. BSAC
version 2013 guideline (Wootton 2013) was referred to interpret the MIC and zone diameter breakpoints
for K. pneumoniae.
To perform MIC per CLSI guidelines, increasing dilutions of antibiotic concentrations (0-2000 µg/ml) in
100 µl Mueller-Hinton broth (MHB) were put into in 96-well microtiter plate. Fresh bacterial inoculum
(5×104 CFU/well in 100 µl) made from overnight cultures was carefully mixed with the MHB antibiotics
medium in the microtitre plate and incubated at 37 for 24h or 48 h. To prepare by mixing 50 µl of 0.5
McFarland (1 × 108 CFU/ml) of bacterial suspension with 9.95 ml of MHB and 100 µl of this mixture was
added to each well as the bacterial inoculum (5×104 CFU per well). Note, the final antibiotic concentrations
in the 96-well were adjusted after the addition of the bacterial inoculum, and the appropriate controls were
included. The growth inhibition was observed and recorded in wells with the lowest antibiotic concentration
where the bacteria failed to growth after the incubated time. The MIC experimental data were pooled from
three experimental repeats with triplicate wells for each experiment. Where necessary, antibiotic Etest strips
were used to determine the MIC of the bacteria.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
90
2.6.4 Co-culture experiments
Basically, for the bacteria-bacteria co-cultures (K. pneumoniae - K. pneumoniae, K. pneumoniae- E. coli,
K. pneumoniae - P. aeruginosa) at 1:1 ratio mixture was made to compete for a certain duration before
CFU were enumerated. For bacterial - amoeba co-culture was used for the bacterial association and
phagocytosis assay while bacteria - amoeba zone clearance was done by spotting A. castellanii on a lawn
K. pneumoniae grown in M9 Minimal medium or LA. Also, K. pneumoniae was used in cell line co-culture/
infection assays such as attachment, invasion and survival.
2.6.5 Bacterial competition and plasmid mobilization
In this study, bacteria-bacteria contact competition on agar surface or in the broth was used to access the
impact of T6SS in bacterial competition in a polymicrobial environment. The bacterial competition
included K. pneumoniae head-to-head intra-strain competition, Head-to-head K. pneumoniae - E. coli inter-
competition and Head-to-head K. pneumoniae - P. aeruginosa inter-species competition.
2.6.5.1 Intra-strain competition
An antibiotic disarmed version of K. pneumoniae (Bi et al., 2015) was competed head-to-head against its
T6SS1 mutant, T6SS3 mutant, and T6SS1 and T6SS3 double knockout mutant in both LB broth and
nitrocellulose filter paper (0.45 µm pore size) placed on LA. Subculture (1:100) of K. pneumoniae HS11286
antibiotic disarmed version (DN2T) and its associated T6SS mutants were made from overnight cultures
(37℃, 200rpm for 16 h). All the K. pneumoniae T6SS mutants (∆T1, ∆T3 and ∆T1∆T3) previously show
similar growth pattern to their respective parent strains (DN2 or DN2T). The subculture of each strain in
LB was grown to OD600nm 0.3 – 0.4 at 37℃, 200rpm.
For intra K. pneumoniae strain LA plate competition, 300 µl of each subculture was pelleted and
resuspended in 300 µl of PBS (Sigma Aldrich). A 1:1 ratio mixture of DN2T (2 ×108 CFU in 100 µl PBS )
to T6SS mutant (2 ×108 CFU in 100 µl PBS of ∆T1, ∆T3 or ∆T1∆T3) was made and 20 µl of the mixtures
(DN2T + ∆T1, DN2T + ∆T3, or DN2T + ∆T1∆T3) were aseptically spotted on a pre-warmed filter paper
placed on LA. Individual strains (20 µl the of DN2T only, ∆T1 only, ∆T3 only and ∆T1∆T3 only) were
also spotted for comparative CFU data. The bacterial mixtures were incubated for 2 - 4 days and the CFU
estimated by resuspending the filter paper with the bacteria in 1000 µl of PBS and appropriate dilutions
plated on LA only and LA plus hygromycin and incubated at 37℃ overnight. Note that, K. pneumoniae
(DN2T) has hygromycin resistance-encoding gene, this enabled the differential CFU count on LA plus
antibiotics and LA only.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
91
For intra-K. pneumoniae strain competition in Liquid medium (LB), the inoculum was prepared just as
before, 300 µl of fresh subculture (grown to OD600nm 0.3 – 0.4 at 37℃. 200 rpm) of K. pneumoniae (DN2T)
was mixed with 300 µl of a new subculture of T6SS mutants (∆T1, ∆T3 or ∆T1∆T3). Then, 100 µl of each
1:1 ratio mixture (DN2T + ∆T1, DN2T + ∆T3, DN2T + ∆T1∆T3) or individual strains (DN2T only, ∆T1 only,
∆T3 only or ∆T1∆T3 only) were aseptically added to 100 ml LB in labelled 500 ml flasks. The individual
K. pneumoniae strains (DN2T, ∆T1, ∆T3 or ∆T1∆T3) were necessary to enable the appropriate comparative
CFU estimation. The culture in the flask was grown at 37℃ with shaking (200 rpm) for seven days. The
CFU from each flask was estimated daily by sampling 100 µl of the bacterial culture and plating the
appropriate dilution on LA only and LA plus hygromycin and incubated at 37℃ overnight.
All intra-strain competition data was pooled from three experimental repeats and used to analyze the
competitive fitness of each K. pneumoniae strains.
2.6.5.2 Head-to-head inter-bacterial species
Like intra-K. pneumoniae strain competition described above, the inter-species competition of K.
pneumoniae strains against E. coli, or P. aeruginosa was also performed on both agar surface (LA) and
liquid medium (LB) to access the effect of T6SS deletion on K. pneumoniae interaction with other bacteria.
For the agar-based inter-bacterial competition, 1:100 subculture of overnight K. pneumoniae, E. coli and P.
aeruginosa were made in fresh sterile LB and grown to OD600nm 0.3 - 0.5 at 37℃ with shaking (200rpm).
A volume (300 µl) of each bacteria subculture was pelleted (3000 ×g) and resuspended in 300 µl of PBS.
A 1:1 ratio of K. pneumoniae (2 ×108 CFU in 100 µl PBS) to E. coli (2 ×108 CFU in 100 µl PBS ) or P.
aeruginosa (2 ×108 CFU in 100 µl PBS ) mixtures were made, and 20 µl of the K. pneumoniae - E. coli
mixtures (DN2T + E. coli, ∆T1T + E. coli, ∆T3T + E. coli, and ∆T1∆T3T + E .coli) or K. pneumoniae – P.
aeruginosa mixtures (DN2T + PA14, ∆T1T + PA14, ∆T3T + PA14, and ∆T1∆T3T + PA14) were spotted on
sterile filter paper placed on a pre-warmed LA. Also, 20 µl of individual bacterial strains (DN2T only, ∆T1T,
∆T3T, ∆T1∆T3 T, E. coli and PA14) were spotted on a filter paper placed on LA plates. After incubating the
inoculated plates for 1 - 4 days at 37℃, the CFUs (K. pneumoniae, E. coli and P. aeruginosa) were estimated
by resuspending the individual or mix bacterial cultures on the filter paper in 1000 µl of PBS and the
appropriate dilutions were plated on LA only, LA + hygromycin, LA + streptomycin, LA + apramycin, LA
+ streptomycin & apramycin. IPTG/ X-gal was added to the recovery media when required. All plates were
incubated at 37℃ overnight for bacterial CFU enumeration.
Unless otherwise stated, when E. coli DH5α (carrying pDNTOOL-39) or E. coli HB101 (carrying
pDNTOOL-39) was used in the competition assay, it was recovered in LA + apramycin supplemented with
IPTG/ X-gal, or LA + apramycin & streptomycin supplemented with IPTG/ X-gal for the E. coli HB101.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
92
For head-to-head inter-bacterial competition in liquid medium (LB) 300 µl of K. pneumoniae was mixed
with 300 µl of the other competing bacteria (E. coli or P. aeruginosa) to make a 1:1 ratio mixture. The
mixtures were pelleted and resuspended in 600 µl of sterile LB, and 100 µl of K. pneumoniae - E. coli
mixtures (DN2T + E. coli, ∆T1T + E. coli, ∆T3T + E. coli, and ∆T1∆T3T + E .coli) or K. pneumoniae – P.
aeruginosa mixtures (DN2T + PA14, ∆T1T + PA14, ∆T3T + PA14, and ∆T1∆T3T + PA14) containing 2
×108 CFU was added to 100 ml of LB in labelled flasks. Also, 100 µl (2 ×108 CFU in LB) of the individual
strains (DN2T only, ∆T1T, ∆T3T, ∆T1∆T3 T, E. coli and PA14) were added to 100 ml of LB in separately
labelled flasks for comparative CFU estimation. The cultures were incubated at 37℃ with shaking (200
rpm) for four days. The CFU K. pneumoniae, E. coli and P. aeruginosa from the individual or mix cultures
were estimated by plating on LA only, LA + hygromycin, LA + streptomycin, LA + apramycin, LA +
streptomycin & apramycin and IPTG/ X-gal were added to the recovery media when required. All plates
were incubated at 37℃ overnight for bacterial CFU enumeration.
All inter-species competition data was pooled from three experimental repeats and used to analyze the
competitive fitness of each K. pneumoniae relative E. coli and P. aeruginosa strains.
2.6.5.3 T4SS conjugative plasmid mobilization assay.
Being a multi-drug resistant strains with many integrative conjugative elements (ICEs), K. pneumoniae
HS11286 has been purported to be a bacterial antibiotic sink. Hence K. pneumoniae HS11286 with active
T6SS could potentially use independent DNA uptake mechanisms such as T4SS to scavenge for other
genetic elements/ virulence genes or work together with other secretory systems such as T6SS enhance its
take- (DNA)-and kill (bacteria) method of environmental survival. T4SS-mediated conjugative
mobilization of pACYC184-oriT-ICEKP-1 into E. coli HB101 was used assessed the effect of T6SS
deletion on MDR K. pneumoniae plasmid mobilization.
ICE1 marker conjugative plasmid (pACYC184-oriT-ICEKP-1) was constructed to ensure that K.
pneumoniae HS11286 has a functional T4SS. The plasmid pACYC184-oriT-ICEKp-1 was constructed by
cloning a 1760 bp oriT amplicon of ICEKp-1 from K. pneumoniae NTUH-K2044 into HindII/ SalI sites in
pACYC184 (a non-conjugative plasmid) as this plasmid has been previously used in similar studies
(Schubert et al., 2004; Lin et al., 2008). Filter mating conjugation was performed between K. pneumoniae
strains carrying pACYC184-oriT-ICEKp-1 and streptomycin-resistant E. coli HB101. The frequency of
mobilization of pACYC184-oriT-ICEKp-1 from either K. pneumoniae DN2 or TSS mutants (donor) to E.
coli HB101 (recipient) was assessed from the bacterial mixtures mated overnight on a membrane filter. The
transconjugants E. coli carrying three plasmids were enumerated in the antibiotic medium.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
93
Briefly, pACYC184-oriT-ICEKp-1 was electroporated into K. pneumoniae strains (DN2, ∆T1, ∆T3 or
∆T1∆T3) and were recovered in LA chloramphenicol. They were colony-PCR verified for the presence of
the plasmid (pACYC184-oriT-ICEKp-1) using DN/DN. Single colonies of each K. pneumoniae strain
carrying pACYC184-oriT-ICEKp-1 was put into 5 ml LB plus chloramphenicol for overnight cultures at
37℃, 200 rpm. Also, a colony of streptomycin-resistant E. coli HB101 was put into 5 ml of LB with
streptomycin and incubated at 37℃ overnight. A 1:100 of a subculture of E. coli HB101 and K. pneumoniae
were made and grow to exponential phase, ~4.5 hr and ~4 hr with shaking at 37℃ respectively. K.
pneumoniae (DN2, ∆T1, ∆T3 and ∆T1∆T3) carrying pACYC184-oriT-ICEKp-1 were considered as the
donor while E. coli HB101 as the recipient.
Bacterial cells (1ml each) from the subcultures were harvest and pelleted (3000 ×g for 10 min) and washed
twice with 1 ml of PBS to remove residual antibiotics media. A 10:1 ratio (10 or 20 µl) of the donor (K.
pneumoniae) to recipient (E. coli HB101) mixture was spotted on 2 cm x 2 cm 0.45 µm membrane filter
(Thermo Fisher) placed on a pre-warmed LA plate. Also, individual strains only were included as controls
for comparative CFU counts. The inoculated plates were incubated at 37 for 20 h before the membranes
were resuspended in 1 ml PBS and vortexed. The appropriate dilutions from the mixtures or neat
suspensions were plated on LA antibiotics plates for bacteria CFU.
The CFU of the output K. pneumoniae donor or E. coli HB101 recipient were enumerated by plating the
mixtures or the individual cultures on LA only, LA + streptomycin, and LA + ampicillin, LA+
chloramphenicol and LA + chloramphenicol & streptomycin. The plates were incubated at 37℃ overnight,
and only E. coli HB101 transconjugants carrying the mobilized plasmid (pACYC184-oriT-ICEKp-1) grew
on LA with chloramphenicol & streptomycin plates. Transconjugants (E. coli HB101 carrying pACYC184-
oriT-ICEKp-1) were PCR confirmed with ICEKp-1 oriT specific primers and E. coli specific primers (see
Appendix 1). Data from three experimental repeats were analyzed for the K. pneumoniae conjugation
efficiency.
The conjugation efficiency was estimated using the below formula;
Conjugation efficiency= (# of Transconjugants) / (# of recipients recovered) CFU/ml
% Conjugation efficiency = [= (# of Transconjugants) / (# of recipients recovered) CFU/ml] x 100
2.6.6 Agglutination assay
Agglutination assays were performed using guinea pig red blood cells (GP-RBCs). GP-RBCs were stored
at 4℃ as 20 % solutions in Alsever’s (Matrix Biologicals). When required, a 5% suspension of GP-RBCs
was freshly prepared by dilution the 20% stock solution 1 in 4 ml PBS. Briefly, agglutination assays were

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
94
performed by mixing 10 µl of overnight K. pneumoniae cultures (37°C, 200 rpm) separately with 10 µl of
the 5 % guinea pig red blood cells suspension on a microscopic slide. The microscope slide was tipped back
and forth and observed for signs of agglutination for up to 15 minutes. To test for mannose-sensitivity, 20
μl of 10 % (w/v) mannose was added to the agglutination reaction. Also, to test for type 3 fimbria expression
by the various K. pneumoniae strains, 10 µl of overnight K. pneumoniae strain each was mixed with 10 µl
of 5% tannic acid-treated Oxblood with or without 5% mannose. The mixture was gently rocked on a
microscope slide for 10-15 for visible agglutination.
2.6.7 G. mellonella larvae In vivo virulence model
The unpigmented wax moth (G. mellonella) larvae were all purchased from Livefood UK Ltd and stored in
darkness at 15℃ and used with 2-3 days upon arrival as previous studies have reported that long-time food
deprivation may lead to a reduction in cellular and immune response (Banville, Browne and Kavanagh,
2012; Browne et al., 2015). The use of G. mellonella as a K. pneumoniae virulence model has been widely
described (Insua et al., 2013b; Wand et al., 2013a). To investigate K. pneumoniae T6SS-induced virulence
in Galleria larvae and the interplay between K. pneumoniae (DN2, ∆T1, ∆T3 and ∆T1∆T3) and Galleria
larva innate immune system, the larvae were infected with either 2 × 108 CFU or 2 × 106 CFU of K.
pneumoniae for the examination of for Galleria mortality, K. pneumoniae recovery, phenoloxidase activity,
and viable hemocyte counts.
2.6.7.1 Galleria killing assay
To prepare bacterial inoculums for Galleria larvae infection, a single colony of each K. pneumoniae strain
was inoculated in 5ml LB for overnight at 37℃, 200 rpm. A 1:100 subculture of each strain was made in
fresh 5 ml LB to grow for approximately 4 h at 37℃ 200 rpm. The 5 ml subcultures were pelleted (3000
×g, 10 min, room temperature) and washed twice in 10 ml of 10 mM MgSO4 for OD600nm adjustment to 0.2.
Prior to the injection of the larvae with 10 µl of the bacterial inoculum (2 × 108 CFU) into the last right
proleg, the larvae were swabbed with 70% (v/v) ethanol. Random selection of 30 larvae for each group
(untouched, MgSO4 only, DN2 only, ∆T1 only, ∆T3 only, ∆T1∆T3 only, KR1730 only and NTUH∆T1) in
labelled Petri dishes. After injection of larvae with 10 µl of bacterial inoculum containing 2 × 108 CFU
(using a BD micro-fine 1 ml syringe affixed on a repetitive pipette from StepperTM series), the larvae were
incubated at 37℃ for seven days. Dilutions of the inoculums were also made and plated on LA to enumerate
inoculum CFUs. No larva from the control groups (untouched and MgSO4) died until the experiments were
terminated at day 7. A larva was considered dead when it fails to elicit a response to physical stimuli and
the larva mortality from each group was recorded daily post-infection. The progression of K. pneumoniae

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
95
infection in Galleria larva was observed to be associated with increased pigmentation. Statistical analysis
was performed analyzed from triplicate experiments.
2.6.7.2 K. pneumoniae 50% Lethal dose (LD50) determination
To enable bacterial recovery and prevent the larvae from dying during the experimental time, 50% lethal
dose (LD50) experiments was determined by inoculating larvae with 10 µl K. pneumoniae inoculum from a
series of 10-fold dilutions (2 × 104 to 2 × 108 CFU) prepared in 10 mM MgSO4. Twenty larvae were used
for each 10-fold dilution, and the larvae were incubated at 37℃ for seven days. The inoculum was also
plated on LA to confirm the bacterial numbers injected into the worms. The inoculum, with about 50 %
mortality by the end of day seven, was used as LD50. Data from three independent experiments were pooled
and used for the calculation of LD50 (2 × 106 CFU) according to the formula of Reed and Muench (Thakur
and Fezio, 1981).
2.6.7.3 In vivo bacterial recovery from Galleria larva
For bacterial recovery, 25 larvae per group were infected with 10 µl of LD50 (2 × 106 CFU) of K. pneumoniae
individual strains only (DN2T, ∆T1, ∆T3, ∆T1∆T3, KR1730 and NTUH∆T1::aacC1 only) or mixture ( 1:1
ratio of DN2T + ∆T1, DN2T + ∆T3, DN2T + ∆T1∆T3, and KR1730 + NTUH∆T1::aacC1) and incubated
for 4 days post-infection. Also, the necessary controls (untouched and injected with 10 mM MgSO4) were
included. The inoculum was also plated on LA to confirm the bacterial numbers injected into the worms.
No larva from the control group died until the experiments were terminated. To recover bacteria, five larvae
per group were randomly selected and homogenized in 5 ml of 0.5% Triton X-100/PBS lysis buffer
(containing phenylthiourea). The appropriate dilutions of the larva homogenates/lysates were a plate on LA
only, LA + ampicillin, LA + ampicillin + hygromycin or LA + ampicillin + gentamicin and incubated at
37℃ overnight to enumerate the CFU of bacterial from individual inoculation or the mixtures culture
inoculation. No bacterial growth was recovered from the untouched or MgSO4 injected larvae grown on
LA agar supplemented with ampicillin at 37℃ overnight. At least three independent experiments were
performed, and the data pooled were subjected to statistical analysis.
2.6.7.4 Phenoloxidase activity (PO) from Galleria larvae
PO activity was performed as previously described (Insua et al., 2013b). Fifteen larvae per group were
injected 10 µl LD50 of K. pneumoniae (DN2 only, ∆T1 only, ∆T3 only, ∆T1∆T3 only, KR1730 and
NTUH∆T1only). Five larvae from each group on day two and day four post-infection were surface
sterilized with 70% ethanol. The hemolymphs (15 µl each) were collected and diluted in 10 µl of ice-cold

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
96
saturated N-phenylthiourea solution (diluted in dH20) in microcentrifuge tube to prevent melanization.
Samples for each group were pooled together and centrifuge (4,000 × g for 10 min at 4℃) to eliminate cell
debris. The supernatants (hemolymph plasma) were diluted at 3:1 ratio (v/v) in 50 mM PBS (pH 6.5). The
phenoloxidase activity was measured using previously describe microplate enzyme assay (Eleftherianos,
Millichap and Reynolds, 2006). Briefly, a reaction mixture was set up containing 10 µl of diluted
hemolymph plasma, 115 µl of 50 mM PBS (pH 6.5), 2 µl of E. coli lipopolysaccharide (LPS, 5 mg/ml,
Sigma Aldrich) at 25℃. The reaction mixture was left to stand for 60 minutes to activate the enzyme before
adding 25 µl of 20 mM 4-methyl catechol (Sigma Aldrich) to initiate the reaction. The final reaction volume
was adjusted with dH2O to 200 µl. The durational change in absorbance (490 nm) was measured every 5
minutes using a pre-set Varioskan® flash spectral scanning multimode reader with continuous shaking for
1 h at 25 ℃. The experiment was performed in triplicates for three independent experimental repeats, and
the PO was expressed as mean OD490 nm/minute for each of the treatment groups.
2.6.7.5 Hemocytes quantification and viability staining from infected Galleria larva
For viable hemocyte count, 15 larvae per group were injected 10 µl LD50 of K. pneumoniae (DN2 only,
∆T1 only, ∆T3 only, ∆T1∆T3 only, KR1730 and NTUH∆T1only). Larva hemocytes were extracted as
previously described (Banville, Browne and Kavanagh, 2012; Insua et al., 2013b). Briefly, five larvae from
each infected group at day two and day four were surface sterilized with 70% ethanol and dissected for the
hemolymph extraction. The pooled hemolymph samples (10 µl) were mixed immediately with 10 μl of
phenylthiourea (Sigma Aldrich) in distilled water in a microcentrifuge tube containing and hemocytes were
recovered by centrifugation at 1,500 × g for 3 min. The hemocytes suspension were immediately mixed
with 20 μl of 0.4 % trypan blue and incubated for 5 minutes before counting total hemocyte and viable
hemocytes using a hemocytometer. The data from three experimental repeats in triplicates were expressed
as relative percentage viability of hemocytes.
2.6.8 Eukaryotic cell-based virulence assays and methods
To further investigate the K. pneumoniae T6SS virulence, mouse and human cell lines (J774A.1, H292 and
A549), human polymorphonuclear cells (neutrophils) and Acanthamoeba castellani were used in various
assays to measure the virulence-related effect of T6SS.
2.6.8.1 Cells and culture conditions
Three cell lines were used in this study, a mouse macrophage (J774A.1) and two human epithelial
carcinoma cells (H292 and A549) in various assays to evaluate the T6SS dependent virulence in K.
pneumoniae. While J774A.l cell line was kindly provided by Dr Coudla Stover’s Lab (University of

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
97
Leicester), H292 and A549 were both provided Dr Yassine Amrani’s Lab (University of Leicester), all cell
lines were originally obtained from American Type culture collection.
Mouse macrophage, J774A.1 (ATCC TIB-67) from BalB/cN strain were cultured in Dulbecco’s modified
Eagle’s medium (DMEM) with 2 mM L-glutamine (Gibco) containing 10% heat-inactivated fetal bovine
serum (FBS, Gibco) and 1% penicillin/streptomycin (Gibco). The cell lines were routinely maintained in
complete growth medium at 37℃ in 5% CO2 in 75 cm3 flask for 80-90 % confluence before passaging
them. For long term culture, growth medium was changed every 2-3 days. When needed, the appropriate
cells were seeded in 6, 24 or 96 well plates with DMEM + 10 % FBS growth medium and incubated for 2-
3 h before use or overnight at 37℃ in 5% CO2 overnight depending on the experiment. The viability (≥
97%) of the various cell line were periodically tested using trypan blue exclusion method before being used
for any experimental assays.
Human bronchial epithelial cells, NCI-H292 (ATCC CLR-1848), were routinely cultured and maintained
in 75 cm3 flask containing RPMI-1640 with L-glutamine (Gibco) supplemented with 10% FBS (Gibco) and
1% penicillin-streptomycin (Gibco). The cell line was routinely maintained in complete growth medium at
37℃ in 5% CO2 in 75 cm3 flask for 80-90 % confluence before passaging them. For long term culture,
growth medium was changed every 2-3 days. When experimental required, the appropriate cell density was
seeded in 6, 24 or 96 well plates with RPMI -1640 with 10 % FBS only and incubated at 37℃ in 5% CO2
overnight. Prior to experiments, the viability of the cells (≥ 97%) was usually checked.
A549 cells (ATCC CCL-185) were routinely cultured in 75 cm3 flask in DMEM with 2 mM L-glutamine
(Gibco) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Gibco) and 1%
penicillin/streptomycin (Gibco) at 37℃ with 5% CO2. When required, the appropriate cell density was
seeded in 6, 24 or 96 well plates with DMEM + 10 % FBS growth medium and incubated at 37℃ with 5%
CO2 overnight. Cell viability (≥ 97%) was usually checked before being used for any downstream
experiments.
2.6.8.2 Macrophage uptake assay and visualization
Murine macrophage, J774A.1 was (1×105/ well) in DMEM with L-glutamine supplemented with only heat-
inactivated 10% FBS were seeded in 24-well plates and incubated at 37℃ in 5% CO2 15 h before the
experiment. Prior to infection, the spent medium was replaced with complete medium without antibiotics
or FBS. Fresh exponential phase K. pneumoniae culture (DN2, ∆T1, ∆T3 and ∆T1∆T3) was PBS-washed
three-time and resuspended in DMEM without antibiotics and FBS. The bacteria suspension was then used
to infect the monolayer of J774A.1 at MOI 10:1 for 1h at 37℃ in 5% CO2. After 1 h post-infection, each
well was wash twice with PBS and incubated with 100 g/ml gentamicin to the well for 20 minutes.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
98
Gentamicin concentration per time used to kill extracellular bacteria was empirically determined. The cells
were then washed three times with PBS and lysed with 0.5 ml of 0.5 % Triton X-100 in PBS. The lysates
were diluted and plated on LA and incubated at 37℃ overnight for CFU estimation. The analyzed data were
obtained from three independent experiments with triplicate wells in each case.
Also, a monolayer of J774A.1 established on sterile coverslips placed inside 6- well plates were infected
with K. pneumoniae (carrying GFP plasmid, pIM438) at MOI of 10:1 for 1 h at 37℃ in 5% CO2. Similarly,
the monolayer on the coverslips was washed twice with PBS and gentamicin was added to kill the non-
phagocytosed bacteria. The coverslips were washed twice with PBS and then covered with 1 ml of 4%
paraformaldehyde for 15 minutes. After few intermittent washes with PBS, the monolayer on the coverslips
was permeabilized with 0.2 % Triton X-100 for 5 minutes and nuclei-stained with DAPI and mounted for
confocal microscopy. All assays were performed in triplicate experiments.
2.6.8.3 Macrophage survival assay
To assess the ability of K. pneumoniae to survive in J774A.1, the cell density of 1×104 per well were seeded
in 96-well plates 15 h before infection. Using exponential phase K. pneumoniae, each strain (DN2, ∆T1,
∆T3 and ∆T1∆T3) was used to infect five wells at an MOI of 100:1 for 1h before washing and addition of
gentamicin to kill extracellular bacteria. The antibiotics were then maintained in the medium for 2, 4, and
6 h at 37℃ in 5% CO2 and then the antibiotics washed from the cells with PBS. The cells were lysed with
0.5 ml of 0.5 % Triton X-100 in PBS and dilutions of the lysate were plated on LA for CFU estimations.
CFUs from three experimental repeats were used for statistical analysis. Data were analyzed from triplicate
experiments.
2.6.8.4 K. pneumoniae attachment, invasion and replication in lungs epithelial cells
Human lungs epithelial cells, A549 and H292, were challenged with K. pneumoniae strains to assess the
effect of T6SS on the attachment, invasion and replication of the bacteria.
2.6.8.4.1 K. pneumoniae H292 and A549 attachment assays
H292 cells (1×105) were seeded in a 96-well plate 20 h before infection with K. pneumoniae (DN2, ∆T1,
∆T3 and ∆T1∆T3). Also, a monolayer of H292 was seeded on coverslips a day earlier. The H292 cells in
the 96-well plates or on the coverslip were then infected with K. pneumoniae (50:1) for 30 minutes at 37 in
5% CO2. The cells in 96-well plates were then washed 3 times with PBS and lysed with 0.5% Triton X-
100. The lysates were diluted and plated on LA for enumeration of bacteria CFUs. The coverslip monolayer
washed three times with PBS and fixed with 4% paraformaldehyde for 15 minutes. With intermittent
washes with PBS, the coverslips were stained with rhodamine-phalloidin and DAPI for confocal
microscopy. All the experiments were done in triplicates.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
99
For A549 attachment assay, 1×105 cells per well were seeded 20 h in advance of K. pneumoniae infection.
The cells were infected with exponential phase bacteria at MOI of 10:1 and incubated at 37 in 5% for 30
minutes. At the end of the time-point, the cells were washed three times with PBS and lysed with 0.5%
Triton X-100. The lysates were plated on LA plated and incubated at 37℃ for CFU enumerations.
2.6.8.4.2 K. pneumoniae H292 and A549 invasion and survival assays
The cells (1×105) were in 96 well plates and incubated at 37℃ in 5% CO2 20 h before K. pneumoniae
infection. Exponential phase K. pneumoniae at MOI of 100:1 was used to infect the cells for 1 h before
using antibiotic (gentamicin100 µg/ml for 20 minutes) to kill all extracellular bacteria. The infected H292
and A549 cells were maintained in the gentamicin medium for 2, 4, and 6 h at 37℃ in 5% CO2 and then the
antibiotics washed away twice with PBS. The cells were then lysed with 0.5 % Triton X-100/PBS, and the
appropriate dilutions were plated on LA and incubated at 37℃ overnight for CFU enumerations.
2.6.8.5 Lactate dehydrogenase (LDH) assay
Seeded A549 cells (2 × 105/well) in 24 well plate at 37℃ in 5% CO2 was infected with K. pneumoniae
(DN2, ∆T1, ∆T3 and ∆T1∆T3) at MOI of 100:1 for 1 h. Each strain was used to infect triplicate wells, and
the infected cells culture supernatant (500 µl) for each strain was centrifuged to eliminate cell debris at
3000 ×g for 5 minutes. An LDH assay kit (Thermo Fisher) following manufacturer’s recommendation was
used to quantify the amount of LDH in each supernatant. This experiment was repeated three times to obtain
enough data for statistical analysis.
2.6.8.6 H292 ethidium bromide uptake assay
Ethidium bromide (EtBr) assay was used to assess the effect of K. pneumoniae T6SS on H292 membrane
integrity and nuclear staining. EtBr (Sigma Aldrich UK) has very low permeability to live cell membrane
unlike DAPI, but once the membrane of the cell is compromised, the accumulation EtBr increases and
subsequently stains the nuclei of the cells. DAPI (Sigma Aldrich UK) with more specificity and sensitivity
to DNA, used a counterstained for any unstained nuclei after fixation.
First, a monolayer of H292 (37℃ in 5% CO2) was established on coverslips 20 h prior to infection. The
monolayer was infected with exponential phase K. pneumoniae (non-GFP tagged DN2, ∆T1, ∆T3 and
∆T1∆T3) at MOI of 100:1 for 1.5h and washed twice with PBS. The coverslip monolayers were incubated
with antibiotics (gentamicin) in growth medium for 20 minutes and subsequently washed three times with
PBS. ErBr in growth medium (100 µg/ml) was added the monolayers and incubated for 30 minutes at 37℃
in 5% CO2. The coverslips were PBS-washed three times and fixed with 4% paraformaldehyde for 15

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
100
minutes. With intermittent PBS washes, the cells were then counterstained with DAPI and mounted on
microscopic slides for fluorescent microscopy.
2.6.8.7 K. pneumoniae survival in normal Human Serum and polymorphonuclear cells
Venous blood was obtained from 5 consented healthy normal donors who were not on medications or
alcohol a month before donation. One portion of the blood was collected in anticoagulant tubes and the
other portion collected in tubes without anticoagulants.
Serum isolation: The portion of the blood collected in vacutainer (Becton Dickson) without any
anticoagulants was incubated upright at room temperature for 30 minutes until clotting was achieved. The
clotted blood was then centrifuged at 2000 RCF for 15 minutes, and the supernatant (serum) was carefully
transferred into tubes (NUNC) and stored at -20℃ until needed.
Polymorphonuclear Neutrophil (PMN) isolation: The second portion of the donated blood collected in
anticoagulant vacutainer tubes (Becton Dickson) was used for the extraction of polymorphonuclear
neutrophils by standard density gradient separation method as previously describe (Oh, Siano and
Diamond, 2008; Kuhns et al., 2015). Briefly, 7 ml of neutrophil isolation media called lympholyte-poly
solution (a mixture of Sodium metrizoate and Dextran-500, Cedarlane Laboratories) in a 15 ml falcon tube
was carefully over layered with an equal volume of the anticoagulated blood (7 ml). The tube was
centrifuged (500 RCF at 25℃ for 35 minutes) to separate the mixture into six distinct bands (from top to
down; plasma, monocytes, isolation media, neutrophil, more isolation media and red blood cells, RBCs).
With the help of Pasteur pipette, the first three layers (plasma, monocytes and isolation media) were
discarded while the neutrophil and the below isolation media layers (4th and 5th bands respectively) were
collected into a new centrifuge tube. The collected neutrophil containing suspension was diluted with
Hank’s Balanced Salt Solution without Ca2+ & Mg2+ (HBSS-, Thermo Fisher) to a final volume of 10 ml
while inverting the tube a few times ensure adequate mixing. The suspension was centrifuged (350 RCF at
25℃ for 10 minutes) to obtain a red pellet containing neutrophil and residual RBCs and supernatant was
discarded. The residual RBCs in the pellet were lysed by resuspending the pellet in RBCs lysis buffer (2ml,
Thermo Fisher) and slowly pulse-vortexed to mix before centrifugation (250 RCF for 5 minutes at 25℃)
for a neutrophil pellet. To ensure the elimination of RBCs, the RBCs lysis step above was repeated, and the
pellet was resuspended in 500 µl of HBSS- (pulse-vortexed a few times and dilute with HBSS- to 10 ml).
The neutrophil suspension in HBSS- was centrifuged and the final pellet was resuspended in 250 µl of
HBSS+ (Ca2+ & Mg2+, Thermo Fisher) with 2% human serum albumin (Sigma Aldrich). After counting,
more than ≥95.5% of cells were neutrophil (5×106 cells/ml, and ≥96.7% viability), and the desired
concentrations were adjusted accordingly for downstream application requirements.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
101
2.6.8.7.1 K. pneumoniae serum resistance assay
To assess the ability of K. pneumoniae (DN2, ∆T1, ∆T3 and ∆T1∆T3) to survive in serum and cause
infection, a serum resistance assay as previously conducted reported (Lai, Peng and Chang, 2003; Sahly et
al., 2004) was conducted. Briefly, 1×108 CFU of bacteria in 50 µl of PBS was mixed with 50 µl of pooled
normal human Serum and incubated for 1, 2, & 3 h. Also, the same amount of bacterial inoculum was
incubated in heat-inactivated of pooled normal human Serum (56℃ for 30 minutes) for 1, 2, & 3 h. At the
end of each time point, appropriate dilutions from the incubation mixtures were plated on LA and incubated
at 37℃ overnight for the enumeration of bacterial CFU. The percentage K. pneumoniae survival was
calculated as;
[(normal serum CFU)/ (heat-inactivated serum CFU)] x 100%
2.6.8.7.2 C3b deposition on K. pneumoniae
The deposition of C3 complement on K. pneumoniae assay was carried out as previously reported (de
Astorza et al., 2004). Briefly, K. pneumoniae (1×106 CFU per well) was opsonized in 75% normal serum
for 20 minutes and 2 hrs. The opsonized bacteria were washed three times in PBS (without ions), and the
deposited C3b fragments were dislodged by resuspending the bacteria in 50 nM carbonate-bicarbonate
buffer (pH 9.0) containing 1 M NH4OH for 2h at 37℃. The suspension was then centrifuged, and the
supernatant containing the dislodged C3b fragments were collected and processed for ELISA. The C3b in
the suspension were detected following the manufacturer’s recommendation using Abcam C3b kit
catalogue (Ab195461).
2.6.8.7.3 Polymorphonuclear (PMN) leucocytes Internalization of K. pneumoniae
Human Polymorphonuclear neutrophils (PMN) are the most abundant nucleated cells circulating in the
blood with antimicrobial polypeptides in their characteristic cytoplasmic granules and well as possible
antimicrobial reactive oxygen species (Kuhns et al., 2015; Zarember and Kuhns, 2011; Kohanski et al.,
2007; Ganz, 1999). To assess the ability of K. pneumoniae to survive in neutrophils, 1×104 isolated PMN
cells per well were seeded in 96-well plate and incubated at 37℃ for 2h. The cells were then infected with
opsonized K. pneumoniae (1×106 CFU bacteria per well) and incubated at 37℃ with 5% CO2 for 30
minutes. The cells were washed twice with PBS and incubated with gentamicin to eliminate extracellular
bacteria and lysed with 0.5% Triton X-100/PBS at 1 and 2 h post-infection. K. pneumoniae CFUs from the
lysates were enumerated by plating on LA and incubating at 37℃ overnight. Final data were expressed as
CFU/ml from three experimental repeats, with triplicates well for each strain.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
102
2.6.8.8 K. pneumoniae interaction with Acanthamoeba castellani
2.6.8.8.1 The culture of A. castellanii
The A. castellanii ATCC 30234 was axenically grown in 75 cm3 or 150 cm3 tissue flask in sterile PYG
broth at 27℃ to attain at least 95% for trophozoites formation, and the medium changed every 17-20 h.
Trophozoites were harvested by tapping flask and subsequently washing trophozoites three times with
Page’s modified Neff’s amoeba saline (PAS) or PBS for downstream application. Trophozoite viability (≥
97%) were checked using trypan blue staining exclusion and hemocytometer under the microscope.
2.6.8.8.2 A. castellanii zone of clearance
An overnight culture of K. pneumoniae was subcultured at 1:100 in LB and grown for 4 h. The bacteria
were pelleted (3000 ×g for 10 minutes) and resuspended in PBS and the OD600nm adjusted to 0.2. Using a
sterile cotton swab, a lawn of K. pneumoniae was made on Mueller Hinton agar (MHA) or M9 agar, and
the moisture was allowed to dry. A. castellanii trophozoites (1 ×106) resuspended in 20 µl of PBS was then
added to the centre of the bacterial lawn and allowed to dry before incubation. The bacterial lawn together
with A. castellanii was incubated at 37℃ for 24 h or 72 h for the zone of clearance caused by the A.
castellanii. The zone of clearance was measured, and representative images were also taken. After the
incubation, the centre of each zone of clearance was stabbed with a pipette tip and resuspended in PBS.
Trypan blue exclusion was used to count the viable of the A. castellanii in the resuspension for the
estimation of A. castellani viability.
2.6.8.8.3 K. pneumoniae – A. castellanii association
To study the effect of deleting T6SS on the interaction of K. pneumoniae with A. castellanii, an association
assay was performed as previously described (Alsam et al., 2006). Briefly, 2 ×105 cells of A. castellanii
trophozoites resuspended in 100 µl PAS-containing 10% RPMI (PAS-RMPI) were seeded in 96-well plate
and incubated for 2h at 27℃ to allow the cells to adhere to the plate. Exponential phase K. pneumoniae
(MOI 100:1) harvested and resuspended in 100 µl of PAS-RPMI containing 2×107 CFU was added to the
respective wells of A. castellanii trophozoites. Following 1 h co-incubation of K. pneumoniae and A.
castellanii co-culture at 30℃ for 1 h, the mixture was subsequently washed three times with PBS. Using
hemocytometer and trypan blue, A. castellanii viability count at the end of the association time point. To
enumerate the CFU on LA plates, the A. castellanii was lysed in 0.5% Triton X-100/PBS, and the
appropriate dilutions were plated and incubated at 37℃ overnight. The data analyzed was obtained from
three experimental repeats, triplicate in each case. The percentage of K. pneumoniae associated with A.
castellanii was calculated as;
(number of enumerated bacteria/ numbers of enumerated amoeba) x 100%.

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
103
2.6.8.8.4 K. pneumoniae intracellular survival assay
K. pneumoniae intracellular survival was performed as previously reported (Alsam et al., 2006) 2006)).
Briefly, A. castellanii (1 ×106 cells/per well) in 400 µl of PAS-RPMI were seeded in 24-well plate and
incubated in 27℃ for the cell to adhere to the plate. K. pneumoniae (5 ×107 CFU) at MOI of 50:1 in 100 µl
PAS-RPMI was used to infect the cells and incubated for 1 h. Following the incubation of K. pneumoniae
– Acanthamoeba co-culture, the cells were washed twice with PBS and maintained in 500 µl of PAS-RPMI
with gentamicin (100 µg/ml) for 2 and 4 h post-infection at 30℃. At each time-point, A. castellanii counts
were determined using trypan blue and hemocytometer. The infected A. castellanii lysates (0.5% Triton X-
100/PBS) was plated on LA and incubated at 37℃ overnight for CFU were enumerated. The data were
analyzed from three experimental repeats, triplicate wells for each strain infection. The percentage of K.
pneumoniae survival in A. castellanii was calculated as follows;
(number of enumerated bacteria/ numbers of enumerated amoeba) x 100%.
2.6.8.9 K. pneumoniae induced A549 actin filament polymerization
To assess the effect of T6SS deletion on cytoskeletal rearrangement such as actin filaments, a monolayer
of A549 was established on sterile coverslips placed in 6-well plates and incubated at 37℃ in 5% CO2. The
A549 monolayer of the cell was then infected with K. pneumoniae inoculum (5 × 106 CFU) at MOI of 50:1
and incubated for 1h. Gentamicin was maintained in the medium for 3 and 6 h before they washed three
times with PBS. The infected A549 monolayer was then fixed with 4% paraformaldehyde for 15 minutes.
After gently washing the coverslips twice with 20 mM glycine and PBS, they were stained with rhodamine-
phalloidin (Cytoskeleton Inc) and DAPI (Sigma Aldrich) for 5 minutes and mounted on microscopic slides
for confocal microscope.
2.6.8.10 Staining and Confocal microscopy
To visualize GFP tagged K. pneumoniae, DAPI, EtBr or rhodamine phalloidin-stained samples, Olympus
F1000 confocal microscope at The Center for Core Biotechnology service (University of Leicester) was
employed. Basically, after washing the samples with PBS, the samples were fixed with 4%
paraformaldehyde (Thermo Fisher) for 15 minutes. After fixation, the free aldehyde groups in the sample
were blocked by incubating the samples in 20 mM glycine solution (Sigma Aldrich) for 10 minutes and
washing twice with PBS. Triton X-100/PBS (0.5%) were used to permeabilized the samples for 5 minutes,
and the samples were then washed twice with PBS. When required, samples were stained with rhodamine-
phalloidin (200 µl of 100 nM, Cytoskeleton Inc) for 30 minutes and washed twice with PBS. DAPI (200 µl
of 100 nM, Sigma Aldrich) as a counterstain for 5 minutes and washed twice in PBS. GFP (green),

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
104
rhodamine-phalloidin (red) and DAPI (blue) were used to tagged K. pneumoniae, stain actin filaments and
nuclei of the cells respectively. To obtain good quality and reproducible results, all washes, fixation and
permeabilization were done at room temperature (23℃-24℃) while staining-incubations were carried out
in the dark at room temperature. With a drop of vectashield antifade mountain medium (Vector
Laboratories), the samples were mounted on microscopic slides and confocally imaged. Slides were usually
stored in the Dark until required for imaging. The excitation and emission of 385 nm /505 nm (GFP), 540
nm/540 nm (rhodamine-phalloidin) and 365nm/ 461 nm (DAPI) was used for the visualization of the
samples.
2.6.8.11 Proteomic profiling of infected A549 and J774
To explore the innate immune response to K. pneumoniae, various type of cytokines expressed by A549
and J774A.1 cells when infected with the different K. pneumoniae strain (DN2, ∆T1, ∆T3, ∆T1∆T3,
KR1730, NTUH∆T1) is imperative in understanding how T6SS may influence the host immune response
during infection. A preliminary semi-quantitative method was used to assess the proteomic profile of mouse
cytokine array expressed in response to K. pneumoniae infection of A549 and J774A.1 by using the K.
pneumoniae - A549 or J774A.1 co-cultured supernatants or cell lysates. Briefly, cell line (A549 or J774A.1)
was infected with bacteria in 24-well plate at an MOI of 10:1 or 20:1 and allowed for various time point
post-infection. The culture supernatant was collected while the cells were lysed with RIPA lysis buffer
(Thermo Fisher). The samples (cell supernatants or lysates) were processed for their cytokine profiling in
each case using R&D proteomic profiler human cytokine kit (A549 samples) or proteomic profiler mouse
cytokine kit (J774A.1) following manufacturer’s recommendation The proteomic profiler cytokine array
kit is a membrane-based sandwich immunoassay where samples are first mixed with a cocktail of
biotinylated detection antibiotics as and then incubated with the membrane (carrying the duplicate dot-spots
of cytokine capture antibodies arrays). Captured protein was visualized using chemiluminescent detection
reagent, and the signal (pixel intensity) was proportional to the amount of the cytokine in the analytes/
samples. ImageJ software package 6.0 was used to acquire and analyzed the pixel intensities of the captured
cytokines in the analytes/samples
2.6.8.12 ELISA cytokine/chemokine quantification assay: A549 and J774
Following the use of the semi-quantitative profile cytokine kits, a more specific and fully quantitative
traditional sandwich ELISA was performed using various ELISA kits obtained from R&D. Briefly, the
supernatant of infected J774A.1 or A549 (infected with K. pneumoniae at MOI 10:1 or 20:1) were collected,
centrifuged and filtered with 0.45m filter (Millipore) and stored at -80℃ until required. Abcam or R&D

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
105
Elisa kits for mouse (J774A.1) and human (A549) were used to quantify the amount TNF-α, IL-8,
RANTES, IL-1a, IP-10, MIP-1a, MIP-b and MCP-1 in the infected J774A.1 culture supernatant, and IL-6,
IL-8, MIF, GRoa, and TNF-α from infected A549 culture supernatant per manufacturer’s recommendations.
2.6.9 Transmission Electron Microscopy
Briefly, a 5 μl volume of K. pneumoniae samples (DN2, ∆T1, ∆T3, ∆T1∆T3, KR1730, NTUH∆T1) were
applied to a hydrophilic Formvar-carbon coated copper grid (Agar Scientific). The samples were allowed
to adsorb for 5 minutes, and after wicking the excess liquid, the grid was washed once using distilled
deionized water and then stained for 15 seconds with 5 μl of 1 % uranyl acetate (pH 4.5). All the
Transmission Electron Microscopy were performed at the Centre for Core Biotechnology Services
(University of Leicester) with the help of Stefan Hyman using JEOL JEM-1400 microscope at 80 kV.
2.6.10 Data analysis and Statistical Analysis
Various programs/ software packages were used to analyze the data as listed in the bioinformatics section
below. The statistical analysis (with the help of Graph pad prism and excel analysis) in growth curves,
biofilm assay, survival curves in killing assay, cell association and invasion, bacterial competition, Elisa,
and bacteria recovery in Galleria, was performed using of the following; One or two way ANOVA, multiple
t-test using Holm-Sidak method, student t-test, Mantel-cox test, non-parametric Kruskal-Wallis test,
Dunn’s posthoc analysis, and non-parametric Mann-Whitney U test. The p values less than 0.05 were
considered as statistically significant.
2.6.11 Bioinformatic Analysis
A variety of stand-alone or online/web-based programs (listed in the Table 2.4 below) were used in the
bioinformatic analysis for this study.
Table 2.4 Bioinformatic resources used in this study
Name Purpose Reference
PrimerBlast Designing and blasting primers (Jian et al, 2012)
Primer3 Designing primers (Koressaar and Remm, 2007; Untergasser et
al, 2012)
Nebcutter For restriction enzyme selection and digestion (Vincze, Posfai and Roberts, 2003)
ORF Finder (NCBI) For identifying open reading frames of
unknown genes
(Wheeler et al, 2003)
https://www.ncbi.nlm.nih.gov/orffinder/
BPROM Identifying the bacterial gene promoters (Salamov and Solovyevand, 2011)
MobilomeFinder Bacterial genomic island identification (Ou et al, 2007)
David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
106
APE plasmid editor Linear or circular DNA sequence
manipulation and analysis
http://jorgensen.biology.utah.edu/wayned/ap
e/
Snapgene Manipulation of line or circular DNA, in silico
PCR and restriction digestion
SnapGene® software (from GSL Biotech;
available at snapgene.com)
Neural Network promoter
prediction
Bacteria promoter analysis (Reese, 2001; Reese, 2000)
BlasTn Searching NCBI nucleotide database using
nucleotide sequence query
https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAG
E_TYPE=BlastSearch
BlasTp Searching NCBI protein database using
protein sequence query
(Altschul et al, 1997)
https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAG
E=Proteins
BlasTx Using translated nucleotide to search NCBI
protein database
https://blast.ncbi.nlm.nih.gov/Blast.cgi?LIN
K_LOC=blasthome&PAGE_TYPE=BlastSe
arch&PROGRAM=blastx
tBlasTx Using translated nucleotide to search NCBI
translated nucleotide database
https://blast.ncbi.nlm.nih.gov/Blast.cgi?PRO
GRAM=tblastx&PAGE_TYPE=BlastSearch
&BLAST_SPEC=&LINK_LOC=blasttab
CDD /RPS-Blast Conserved protein domain/ function search (Marchler-Bauer et al, 2017)
https://www.ncbi.nlm.nih.gov/Structure/cdd/
wrpsb.cgi
Pfam Conserved protein domain/ function search (Finn et al, 2016) http://pfam.xfam.org/
PROSITE https://prosite.expasy.org/
GeneMark.hmm For gene predictions and analysis http://exon.gatech.edu/GeneMark/gmhmmp.
cgi
ClustaX
In silico PCR Online/web-based PCR analysis with primers (Bikandi et al, 2004; San Millan et al, 2013)
Oligo Calc For primer property calculation (such as Tm) (Kibbe, 2007)
Phylogeny Fr For phylogenetic tree analysis (Dereeper et al, 2008; Dereeper et al, 2010)
MUSCLE Protein/nucleotide sequence alignment https://www.ebi.ac.uk/Tools/msa/muscle/
MAFFT Protein/nucleotide sequence alignment https://www.ebi.ac.uk/Tools/msa/mafft/
Clusta Omega Protein/nucleotide sequence alignment https://www.ebi.ac.uk/Tools/msa/clustalo/
BAGET Bacterial and Archaeal gene exploration tool (Oberto, 2008)
Absynte Archaeal and Bacterial Synteny explorer (Despalins, Marsit and Oberto, 2011)
SyntTax Prokaryotic synteny and taxonomy explorer (Oberto, 2013)
MGcV Microbial genomic context viewer (Overmars et al, 2013)
Genearks Microbial gene prediction and trimming (Besemer, Lomsadze and Borodovsky, 2001)

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
107
ARDB Antibiotic resistance database for predictions (Liu and Pop, 2008)
CARD Comprehensive antibiotic resistance database (McArthur et al, 2013)

David Mwin, Ph.D. Thesis “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Materials and Methods
108

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
109
Chapter Three
Generation of molecular tools, in silico analysis and
genetic manipulation of K. pneumoniae

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
110
Chapter 3: Generation of molecular tools, in silico analysis and
genetic manipulation of K. pneumoniae
3.1 Constructed molecular tools and strategic genetic manipulation
To help advance T6SS works in K. pneumoniae and ease the genetic manipulation of multidrug-resistant
Gram-negative bacterial strains, various molecular tools were constructed and either used in this work or
elsewhere by colleagues in Lab 212 or collaborators. Recently, many bacterial species, including the K.
pneumoniae, have become resistant to a broad range of antibiotics, thereby evading clinical therapies. K.
pneumoniae HS11286 used in this work is clinical isolate with multidrug-resistant properties that was
isolated from the sputum of a patient at Huashan Hospital, Shanghai-China (Liu et al., 2012a). The strain
is also resistant to carbapenems, which are often considered the “last-resort” antibiotics in Clinical settings.
Many of such resistant Gram-negative bacteria such as K. pneumoniae also double their arsenal potency
with other virulence factors (e,g T6SS) that may have been acquired via horizontal gene transfer. Also, at
the beginning of these research work, there very limited molecular tools that could be used for the genetic
manipulation of MDR K. pneumoniae genomes, several molecular tools were constructed to cover for this
need. Using allelic exchanged approach via lambda red mutagenesis, a novel strategy was developed to
swap genes in plasmids in E. coli background with 100% efficiency without the use of restriction-ligation.
3.1.1 Novel DNA swapping strategy; constructing FRT-flanked antibiotic resistance
marker
To create variants of a plasmid, a novel approach was developed by first constructing a negative selection
and antibiotic markers (sacB-cat) version of that plasmid before subsequently swapping the negative and
antibiotic markers with the required gene/marker in the presence of sucrose. This approach is easy, fast and
efficient in constructing varieties of a plasmid by eliminating the difficulty of looking for compatible
restriction enzymes sites for cloning purposes. A well-established negative and positive selection marker
(sacB-cat) was amplified from pJTOOL-3 and used to replace the target gene in the plasmid of interest
(Figure 3.1). This intermediate plasmid (the sac-cat bearing plasmid) was then transformed into an E. coli
carrying a lambda red recombination plasmid and recovered at 30℃ with the appropriate antibiotics. The
Examples of such constructed intermediate plasmids carrying sacB-cat negative-antibiotic marker include
pDNTOOL-13, pDNTOOL-16, pDNTOOL-22 and pDNTOOL-31. A mutant allele constructed by joining
the antibiotic marker /gene of interest to LF and RF of the target gene was used to transform arabinose

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
111
induced E. coli carrying both the intermediate plasmid and the lambda red recombination plasmid. The
transformants were then recovered at 37 on LA supplemented with 5% sucrose and the appropriate
antibiotics. The recovery of the transformants on that appropriate antibiotics LA plate supplemented with
5% sucrose at 30℃ enabled the successful replacement of the sacB-cat on the intermediate plasmid with
gene B (Figure 3.1) while losing both the temperature-sensitive lambda Red recombination plasmid and
any residual intermediate sacB-cat bearing plasmid. The negative selection pressure in the E. coli is because
of the lethality levansucrase expressed by sacB in the presence of sucrose while at a higher temperature
(≥37℃) the temperature-sensitive lambda Red recombination plasmid is lost from the E. coli. With this
technique, a wide range of plasmid variants (with the same backbone) can be constructed via lambda Red
recombination-based mutagenesis using various linear mutant alleles (LF-Antibiotic maker-RF) is a single
electroporation event.
The E. coli carrying both the sacB-cat-intermediate plasmid and the lambda Red recombination plasmid
was then repeatedly electroporated with mutant alleles resulting in plasmid variant construction. Hence,
after transformation with the mutant alleles, only transformants carrying the right constructed plasmid were
recovered on the plate at 37℃ and subsequently confirmed by PCR and restriction digestion. This approach
was so efficient (Figure 3.3) that with little to no optimization, it could be extrapolated to making marker-
less chromosomal complementation.
Based on this novel strategy of gene swapping strategy, several molecular genetic tools have been
constructed and used to facilitate the genetic manipulation of various bacteria for this work or elsewhere.
Some of the tools constructed include FRT-flanked antibiotic resistance marker, lambda Red variant
plasmids, Flprecombinase-expressing variants, and expressing/cloning plasmids.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
112
Figure 3.1 Schematic representation of novel in vivo plasmid manipulation.
A negative selection-antibiotic cassette marker, sacB-cat (1a) amplicon from pJTOOL-3 and an inverse-PCR amplicon
of the plasmid to be manipulated (1b) were restriction digested and ligated to form the intermediate plasmid (2), which
becomes the secondary target plasmid. Gene B was indirectly used to replace gene A via lambda mutagenesis. The
intermediate plasmid was then transformed into a competent E. coli strain carrying a lambda Red plasmid
recombination plasmid and recovered on LA with chloramphenicol and any other appropriate antibiotic at 30℃. A
mutant allele consisting of the antibiotic cassette, LF and RF of the target gene was transformed into a 0.2% arabinose-
induced E. coli strain carrying both plasmids (intermediate sacB-cat plasmid and lambda Red recombination plasmid),
The transformants were recovered in LA supplemented with 5% sucrose and the appropriate antibiotic at 37℃
overnight. In the presence of sucrose, sacB produces levansucrase which lethal to E. coli. Hence any residual plasmid
with sacB-cat is kicked out of bacteria, and only colonies with the appropriate antibiotic marker are retrieved.
To investigate the feasibility of this in vivo plasmid cloning approach using E. coli as a host, the construction
of FRT flanked antibiotic marker variants in pUC18R6K-mint-tn7t backbone was used to validate this novel
strategy while testing for its versatility and efficiency. An FRT-aacC1-FRT based plasmid (pUC18R6K-
mint-tn7t-Gm) was used as the PCR template for the LF and RF of the target gene (aacC1) and as primary
target plasmid with the target gene to be replaced. A 2.2 kb amplicon of sacB-cat (using primer-pair
DN41/DN42) from pJTOOL-3 was ligated with a 3.8 kb pUC18R6K-mini-Tn7t backbone (using
DN43/DN44), and the ligation mixture was transformed into E. coli and recovered on LA +
chloramphenicol at 37℃ overnight. To verify the functionality of the sacB-cat on the pDNTOOL-31, a
verified transformant colony of E. coli carrying the 6 kb intermediate pDNTOOL-31 was grown in 5ml LB
+ 5% sucrose overnight at 37℃ with shaking and an appropriate dilution was plated on LA and LA+
chloramphenicol. The plates examine for colonies, but no colonies were noted on LA+ chloramphenicol,
unlike LA, indicating that the E. coli had loss the pDNTOOL-31 carrying the sacB-cat in the presence of
sucrose due to the lethality of expressed levansucrase. A negative colony PCR results from 10 randomly
selected colonies were used a PCR authenticate the loss of sacB-cat bearing pDNTOOL-31.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
113
Figure 3.2 Construction and verification of pDNTOOL-31.
The intermediate plasmid (secondary target plasmid), pDNTOOL-31, was constructed by ligating sacB-cat amplicon
(DN41/DN42, 2199 bp) with pUC18R6K-mini-Tn7t backbone amplicon (DN43/DN44, 3796 bp) and transforming the
ligation mixture into E. coli carrying lambda Red plasmid (pDNTOOL-15). The transformants were recovered at 30℃
on LA plates supplemented with sucrose and the appropriate antibiotics. To construct various antibiotic cassettes
flanked by FRTs, arabinose induced E. coli carrying both pDNTOOL-15 and pDNTOOL-31 was transformed with the
mutant allele (LF-antibiotic cassette-RF), this resulted in the replacement of the sacB-cat region with the mutant allele
by allelic exchange.
The 6 kb intermediate pDNTOOL-31 was extracted and verified by both PCR and restriction digestion
(Figure 3.2) before it was transformed into an electrocompetent E. coli carrying pDNTOOL-15 and
recovered at 30℃ on LA supplemented with antibiotics. Transformants colonies were parallel patched on
LB + chloramphenicol, LA + kanamycin and LB + chloramphenicol and kanamycin at 30℃ overnight. An
overnight culture (5 ml in LB + chloramphenicol and kanamycin) from a verified E. coli carrying
pDNTOOL-31 & pDNTOOL-15 and grown at 30℃ with shaking. A 1:100 subculture of E. coli (carrying
pDNTOOL-31 & pDNTOOL-15) in 100ml of LB + chloramphenicol and kanamycin at 30℃ with shaking
to grow to OD600nm 0.2 before the addition of 0.2% arabinose. The culture was harvested for

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
114
electrocompetent cells preparation when the OD600nm of cells was between 0.6-0.8. Aliquots of the
arabinose-induced electrocompetent of E. coli (carrying pDNTOOL-31 & pDNTOOL-15) were separately
transformed with an array of mutant alleles to give rise to various pDNTOOL plasmids as indicated in
Figure 3.4.
Figure 3.3 Colony PCR to confirm the presences or absence of a target gene, lambda Red and constructed plasmids.
To test for the efficiency of this in vivo cloning approach, ten E. coli tranformant colonies originally carrying both
pDNTOOL-31 & pDNTOOL-15 were randomly selected for PCR confirmation for the presence or absence of
pDNTOOL-31, pDNTOOL-15 and constructed plasmid before and after transformation. Before transformation with the
mutant alleles, colony PCR amplification of sacB-cat from pDNTOOL-31, beg region (bet, exo, gam) from pDNTOOL-
15, and mutant alleles indicated that only pDNTOOL-31 & pDNTOOL-15 were present in the E. coli. However, after
transformation of arabinose induced E. coli carrying the two plasmids (pDNTOOL-31 & pDNTOOL-15) with the
various mutant allele, a transformant colony PCR was performed for the presence of sacB-cat bearing-pDNTOOL-31
(DN41/DN42, 2.2 kb), beg-bearing pDNTOOL-15 (DN125/DN126, 1.9 kb) and constructed pDNTOOL-bearing the
mutant alleles [DN54/DN55 (aac(3)-IV, 836 bp), DN49/DN50 (hph, 1046 bp), DN49/ DN53(hph-gfp 1935 bp),
DN54/DN53 (aac(3)-VI-gfp 1725 bp)]. The efficiency index of the in vivo cloning strategy was calculated as; # of
colonies PCR-positive for the plasmid divided by the # of colonies randomly selected, all multiplied by 100%. Unlike
before transformation, all the transformant colonies carried only the mutant allele in the constructed plasmid but not
pDNTOOL-31 & pDNTOOL-15 after transformation.
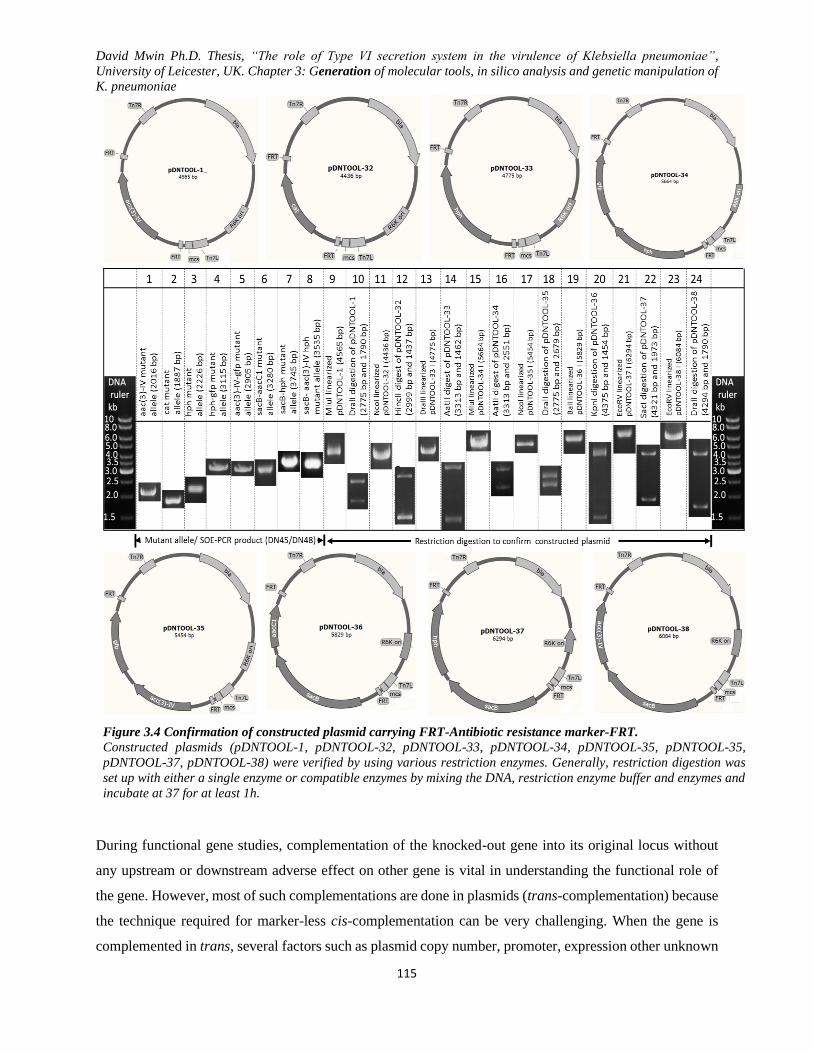
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
115
Figure 3.4 Confirmation of constructed plasmid carrying FRT-Antibiotic resistance marker-FRT.
Constructed plasmids (pDNTOOL-1, pDNTOOL-32, pDNTOOL-33, pDNTOOL-34, pDNTOOL-35, pDNTOOL-35,
pDNTOOL-37, pDNTOOL-38) were verified by using various restriction enzymes. Generally, restriction digestion was
set up with either a single enzyme or compatible enzymes by mixing the DNA, restriction enzyme buffer and enzymes and
incubate at 37 for at least 1h.
During functional gene studies, complementation of the knocked-out gene into its original locus without
any upstream or downstream adverse effect on other gene is vital in understanding the functional role of
the gene. However, most of such complementations are done in plasmids (trans-complementation) because
the technique required for marker-less cis-complementation can be very challenging. When the gene is
complemented in trans, several factors such as plasmid copy number, promoter, expression other unknown

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
116
proteins in the plasmid backbone and plasmid compatibility among many others affect the expression of
the complemented gene and other neighbouring genes. As a result, the gene could be over-expressed, under-
expressed or even not expressed at all, thus affecting the overall complementation studies. Unlike trans-
complementation studies, this novel approach of gene swapping can easily be extrapolated for cis-
complementation studies. With careful optimization, the gene to be complemented can be precisely put
back into its original locus, hence minimizing or eliminating any deleterious effect of trans-
complementation.
3.1.2 Other constructed genetic tools
Using the above-described approach, together with standard cloning techniques, other important molecular
tools were constructed. These included lambda Red recombination-based variant plasmids, Flp-
recombinase encoding plasmids and cloning/expression plasmids. These tools provided the flexibility of
antibiotic marker selection during the mutagenesis stage, post-mutation antibiotic resistance marker
excision and cloning/ expression of genes for functional studies. In the case of the T6SS study, some of
these tools provided the means further to explore the role of T6SS in K. pneumoniae.
3.1.1.1 Lambda Red recombination plasmid derivatives
For mutagenesis, variants of lambda red recombination-based plasmids (pKOBEG-Apra and pKD46) were
mainly constructed using the in vivo cloning as described earlier in this section. These plasmids include
pDNTOOL-5, pDNTOOL-10, pDNTOOL-11, pDNTOOL-12, pDNTOOL-13, pDNTOOL-14,
pDNTOOL-15, pDNTOOL-16 and pDNTOOL-17. The lambda red plasmid, pKD46 encoding ampicillin
resistance was not selectable during the mutagenesis of MDR K. pneumoniae. Hence, the plasmid was used
as the parent plasmid in the construction of hygromycin, apramycin, gentamicin, kanamycin and
chloramphenicol variants (pDNTOOL-5, pDNTOOL-10, pDNTOOL-11, pDNTOOL-12, pDNTOOL-13
respectively) of pKD46 were constructed. Though pKOBEG-Apra encoding apramycin resistance was
selectable in MDR K. pneumoniae HS11286 during mutagenesis and the availability and diversity of such
plasmids are essential in the genetic manipulation of many MDR Gram-negative bacteria. Thus, pKOBEG-
Apra was also used as a parent plasmid for the construction of variants including pDNTOOL-14,
pDNTOOL-15, pDNTOOL-16 and pDNTOOL-17, encoding hygromycin, kanamycin, chloramphenicol
and gentamicin resistance, respectively. All the lambda red recombination-based plasmid variants were
verified by phenotypic- selection for that associated antibiotic resistance and genotypically by restriction
digestion and PCR. Plasmid and primer sequences are in Table 2.1 and Appendix 1: Primer Table,
respectively. Some of these constructed lambda red plasmid variants (pDNTOOL-5, pDNTOOL-10,
pDNTOOL-14 and pDNTOOL-17) were used in the constructing various gram-negative bacterial mutants

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
117
including K. pneumoniae, A. baumannii and E. coli. The constructed lambda red recombination-based
plasmid carried a temperature-sensitive origin of replication (30℃) and an arabinose-inducible promoter
which responded to maximal expression with the addition of 0.2% of L-arabinose.
3.1.1.2 Flp-recombinase encoding plasmids
A flippase recombinase encoding plasmid pFL2, carrying ampicillin marker was used for the construction
of different antibiotic resistance encoding versions for mutant antibiotic marker excision to generate
markerless mutants. The constructed Flp-recombinase encoding plasmids include pDNTOOL-2,
pDNTOOL-18, pDNTOOL-19, pDNTOOL-20 and pDNTOOL-21 respectively encoding apramycin,
hygromycin, chloramphenicol, gentamicin and kanamycin resistance. The plasmids were mainly
constructed by electroporating E. coli carrying the lambda red plasmid and the pFLP2 with an SOE-PCR
product (LF-antibiotic marker-RF). This resulted in the successful replacement of the ampicillin gene in
the original pFLP2 with the donor antibiotic resistance marker. All the Flp recombinase-based plasmid
variants constitutively expressed flippase recombinase which recombines the two FRT sites flanking a
mutant antibiotic marker thereby excising the antibiotic marker and making the mutant markerless. Also,
these plasmids carry a sacB gene which expresses lethal levansucrase in the presence of sucrose, thus
causing the Flp-recombinase plasmid to be negatively selected against by the markerless mutant. With the
construction of the Flp-recombinase plasmid variants, it provided the flexibility of antibiotic selection and
an efficient and safe way of making MDR bacterial mutants. The Flp recombinase plasmids were verified
by the antibiotic selection, restriction digestion and PCR. While the plasmids were widely used by
colleagues and collaborators, they were particularly important in the generation of ∆T1, ∆T3 and ∆T1∆T3
for the study of T6SSs in K. pneumoniae.
3.1.1.3 Cloning/expression plasmids
For the purposes of gene expression and cloning for functional studies, several plasmids were constructed
for used in this work as well as made available to colleagues and collaborators. The cloning/expressing
constructed included pDNTOOL-3, pDNTOOL-4, pDNTOOL-22 to pDNTOOL-30 and pDNTOOL-39.
These plasmids encoded a variety of antibiotic resistance, promoters and replication origin to diversify the
in MDR Gram-negative bacteria. Some of the plasmids (pDNTOOL-26, pDNTOOL-27, pDNTOOL-28
and pDNTOOL-30) expressing GFP can be used for tagging bacteria for confocal/ fluorescence
microscopy.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
118
3.2 K. pneumoniae HS11286 mobile genome analysis and disarmament antibiotic resistance
3.2.1 The mobile genome of K. pneumoniae HS11286 and analysis
MDR and hyper mucoid K. pneumoniae strains are currently a global threat in many hospitals and rendering
most the available antibiotic therapies ineffectual, particularly some last-line antibiotic options such as
carbapenem and polymyxins. This chapter specifically focuses on the exploration and mapping of the
landscape of ST11 Carbapenem-Resistant K. pneumoniae HS11286 (CRKP) mobile genome. The
bacterium is a fully sequenced (GenBank accession number CP003200) and was isolated from the sputum
of a patient at Huashan Hospital, Shanghai-China (Liu et al., 2012a). K. pneumoniae HSS11286 is part of
dominant ST11 clone in China and closely related to world-wide dominant ST258 clone (Qi et al., 2010;
Munoz-Price et al., 2013; Chen et al., 2014b). This makes K. pneumoniae HS11286 a very important model
organism to explore its resistance or virulence-associated mechanisms by annotating its resistance
mobilome and creating an attenuated model strain. Certain mobile genetic elements (plasmids, integrons,
transposons, ISs, prophages, ICEs and GIs) are crucial in the acquisition and spread of virulence or
resistance genes, thus playing a role in the bacteria evolution (Frost et al., 2005a; Partridge, 2011a;
Partridge, Zong and Iredell, 2011).
K. pneumoniae HSS11286 has one chromosome and six plasmids (pKPHS1, pKPHS2, pKPHS3, pKPHS4,
pKPHS5 and pKPHS6), three of which are large plasmids (pKPHS1=122,799 bp, pKPHS2=111,195 bp,
pKPHS3=105, 974 bp) believed to be involved in multidrug resistance including carbapenemase producers
while the rest are small plasmids (pKPHS4= 3,751 bp, pKPHS5= 3,353 bp and pKPHS6= 1,308 bp) that
code for unknown proteins. With a similar backbone to K. pneumoniae plasmid pKP048 (Jiang et al., 2010),
pKPHS2 has blaTEM-1 and carbapenemase gene blaKPC-2 genes. Also, pKPHS3 has similarity with Yersinia
pestis plasmid pIPI202 (Galimand et al., 1997) with essential resistance genes such as tetG, cat, sulI, dfraI2,
aac(3) and aph.
Both pKPHS2 and pKPHS3 has a conjugative transfer (tra) and is thought to play a role in multidrug
resistance of the strain. K. pneumoniae HS11286 has a very plastic genome, hence suggesting the role of
the mobile genetic elements in the bacteria evolution and the spread of antibiotic resistance/ virulence-
associated genes. The K. pneumoniae HS11286 genome has two novel integrative and conjugative
elements; a 63 kb asn tRNA gene associated ICEPKpnHS11286-1 and 56 kb phe tRNA gene associated
ICEKpnHS11286-6 (Bi et al., 2012; Liu et al., 2012a).
As reported in Bi et al. 2015, various mobile genetic elements including 49 ISs, nine transposons, two
separate InC2-like integron remnants, seven prophages, 2 ICEs and four genomic islands were identified
(Appendix 2-4- Distribution of mobile genetic elements in K. pneumoniae HS11286) and annotated. The

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
119
replicon IncFIB (pKPHS1), IncFIIK/ IncR (pKPHS2), IncA/C2 (pKPHS3), ColE2-like (pKPHS4), ColE1-
like (pKPHS5) and Col (KPHS6) were identified (Appendix 2-5 Replicons of the K. pneumoniae plasmids)
to be associated with the plasmids in K. pneumoniae HS11286. Also, pKPHS2 shared similarity to pKP048
and has F-like type IV secretion system (T4SS) gene cluster, an oriT and a relaxase gene traI while pKPHS4
may be mobilizable as it carries relaxase and cognate oriT but not T4SS.plasmid. Among the ISs identified,
30 were in the chromosome, 2 in pKPHS1, 9 in pKPHS2 and 8 in pKPHS3. Plasmid pKPHS1 has one
transposon while pKPHS2 or pKPHS3 each carried four transposons. Seven prophages, two ICEs and four
other GIs were identified on the chromosome of K. pneumoniae HS11286 (Appendix 2-4- Distribution of
mobile genetic elements in K. pneumoniae HS11286). Also, presented in Appendix 2-3 Resistance genes
identified in K. pneumoniae HS11286 that could confer resistance in Clinical cases are genes predicted to
play a role in antibiotic resistance in clinical practice that was identified in the K. pneumoniae HS11286.
The antibiotic resistance regions/genetic elements on pKPHS2 and pKPHS3 were mapped to fine detail
(Figure 3.5) (Bi et al., 2015) and subsequently postulated as the regions responsible for the multidrug
resistance of the K. pneumoniae HS11286 including carbapenem. The region includes blaKPC-2 on pKPHS2
and 12 other clustered antibiotic resistance genes on a 26 kb region on pKPHS3. The strain has many mobile
genetic elements including six plasmids, 49 ISs, nine transposons, two In2-like integron remnants, two ICEs
and seven prophages. Out of 16 identified plasmid-born resistance genes, 14 of them were associated with
Tn1721-, Tn3-, Tn5393-, In2-, ISCR2- and ISCR3-derived elements.
Also, the results from resistance mobilome mapping and disarmament in K. pneumoniae HS11286 provided
insights and facilitated the easy generation of “resistance-disarmed” model organism (Bi et al., 2015). The
antibiotic resistance regions were identified and mapped in K. pneumoniae HS11286, and the knowledge
was used to successfully created a safer and easy-to-manipulate resistance disarmed version for further
work on T6SS (Bi et al., 2015). First, blaKPC-2 was deleted, then followed by the deletion of 26 kb region
(carrying 12 clustered resistance genes) resulting in a significant decrease in the antibiotic resistance
spectrum demonstrated by the resultant mutant, DN2 (Bi et al., 2015). The “resistant disarmed” version
was created by deleting blaKPC-2, and 26 kb region (12 clustered antibiotic resistance genes and one biocides
gene) and the MICs of the mutant was determined to ensure the loss the related antibiotic resistance.
The results re-emphasized plasmid’s role in the acquisition and spread of virulence/ antibiotic resistance in
K. pneumoniae and how ISs, transposons and integrons shape the resistance-coding regions in this clinically
important strain, K. pneumoniae HS11286 (Bi et al., 2015). Hence, the “resistant-disarmed” K. pneumoniae
ST11 strain will offer an easier and modifiable model organism referred to in subsequent chapters of this
thesis as DN2 or DN2T strain for T6SS works.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
120
The mapping out of the resistance region provided an insight into the content and context of the antibiotic
resistance region on the two plasmids, and this facilitated the successfully knocking out of the desired
regions in K. pneumoniae HS11286. This suggests that acquisition and maintenance of as many as six
plasmids in the K. pneumoniae HS11286 through horizontal transfer was critical in mediating plasmid-
based antibiotic resistance. Notably, pKPHS2 and pKPHS3 play a significant role in mediating antibiotic
resistance, supporting the notion that plasmid is the principal bearer of the resistance genes in K.
pneumoniae.
3.2.2 Resistance genes and the broader contexts of the linked regions
In the context of antibiotic resistance, eighteen genes relevant in clinical antibiotic resistance were identified
(Appendix 2-3 Resistance genes identified in K. pneumoniae HS11286 that could confer resistance in
Clinical cases). The blaKPC-2 located on pKPHS2 in the context of Tn1721 transposon (truncated by IS26)
region and highly syntenic to pKP048 (Jioan et al., 2010). The similarity of pKP048 to blaKPC-2 bearing
pKPHS2 plasmid which lacks a 35.3 kb central region with almost perfect identical flanking regions suggest
that pKPHS2 may have originated from a pKP048-bearing plasmid with IS26 insertion into Tn 1712
element and subsequent IS26 recombination. This possibly caused the loss of 35.3 kb central region
(Partridge, 2011a; Cain and Hall, 2012; Partridge, Zong and Iredell, 2011; Harmer, Moran and Hall, 2014).
Regarding antibiotic resistance, more resistance genes were associated with pKPHS3 (Figure 3.5b) than
the other plasmids and were clustered within a 26 kb region that partly shares similarity with an MDR
region on pSRC27-H (Cain and Hall, 2012; Bi et al., 2015). Located within In2-derived class 1 integron
remnant are aadA2, dfrA12-like gene, sul1and qacED1 and In2-derived integron and pKPHS2 integron are
related to Tn21 (Liebert, Hall and Summers, 1999). While tetAR and floR genes may have been acquired
through ISCR3-mediated co-transposition, the three copies of blaTEM-1 carried on varying degrees of Tn3
transposons are consistent with IS26-blaTEM or pSRC27-H configuration (Partridge, Zong and Iredell, 2011;
Bailey et al., 2011; Cain and Hall, 2012). Also, two copies of blaCTX-14 within Tn721-derived element
were identified with the MDR region on the pKHPS3 (Bi et al., 2015). The strA, strB and sul2 may have
been acquired similarly to RSF1010 plasmid (Patridge, 2011) and rmtB, aacC2, fosA and blaSHV-11 could
rarely be shown to be related to mobile genetic elements though rmtB and blaSHV genes are possibly related
to ISCR3 and K. pneumoniae (Bi et al., 2015; Partridge, 2011a; Toleman and Walsh, 2008).
K. pneumoniae HS1128 (dominant CRKP ST11) and other ST11 related strains tend to carry most of the
resistance genes on their plasmids (Bi et al., 2015), which aid the acquisition and spread of such antibiotic
resistance genes or virulent factor within K. pneumoniae species and other species. In a broader context,

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
121
most of these acquired resistance genes have undergone some form of evolution mostly mediated by mobile
genetic elements such as ISs, transposons and integrons. Over time, these evolutionary processes gradually
accumulate virulent factor or resistance genes within the bacterial genome, which become crucial factors
in the bacteria survival and cause disease potential (Bi et al., 2015).
Figure 3.5 The contexts of resistance genes in K. pneumoniae HS11286 (a) pKPHS2 and (b) pKPHS3 plasmids.
The relevant parts of pKP048 and pSRC27-H are also shown to highlight the syntenic regions. The horizontal lines
shown above or below the schematics, with one or both ends demarcated by solid triangles to indicated inverted
repeats, represent intact, interrupted or truncated ISs, transposons or integrons as appropriate. Interruptions in the
above structures are indicated as dashed lines. Regions of synteny between adjacent schematics are indicated by the
shaded areas; the matching percentage nucleotide sequence identity for each such region is indicated. Associated
flanking direct repeat sequences are shown above matching finer lines linked to the solid triangles. GenBank accession
numbers for pKP048 and pSRC27-H are FJ628167 and HQ840942, respectively. The genetic context of the pKPHS3-
borne blaCTX-M-14 is not shown here. Schematic: (Bi et al., 2015)
3.2.3 Genetic disarmament of the resistance of K. pneumoniae HS11286
The clinical K. pneumoniae HS11286 strain poses a significant health risk and difficult to genetically
manipulate due to the acquired multidrug resistance, thus there was the need to create a safer strain that is
easier to manipulate genetically. This was done by sequentially knocking out blaKPC-2 on pKPHS2 and the
26 kb MDR region on pKPHS3. The single (∆KPC) and double knockout (DN2 or ∆KPC∆MDR) were

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
122
employed in antimicrobial susceptibility testing (Appendix 2-2 Antibiotic susceptibility of K. pneumoniae
HS11286 and the antibiotic disarmed versions) against a wide range of antibiotics belonging to 19 classes.
The ∆KPC mutant was sensitive to cefoxitin and ceftazidime, and dose-dependent susceptibility to
cefepime, this indicates that cephalosporin resistance in K. pneumoniae HS11286 most likely due to blaKPC-
2, unlike other b-lactamase encoding genes. As expected, while the wild-type KR3317 was only sensitive
to levofloxacin, colistin, polymyxin B and tigecycline, the mutants (especially DN2) were susceptible to a
wide range of antibiotics. This suggested that plasmid play a key role in the acquisition and dissemination
of resistance genes.
Figure 3.6 Deletion of blaKPC-2 on pKPHS2 and 26 kb MDR region on pKPHS3 in K. pneumoniae
HS11286 background.
A: Disarming the blaKPC-2 (KPHS_p200360) located on pKPHS2 from the original parent strain (K.
pneumoniae HS11286), this resulted in ∆KPC which was then used for the next stage of double knockout. B:
Knocking out the 26-kb multidrug resistance region (KPHS_p300510-KPHS_p300880) located on pKPHS3 in
a ∆KPC background.
The pKPHS2 blaKPC-2 deleted by first constructing an SOE-PCR product (UF-hph-DF) using FRT flanked
hph and ~500 bp upstream and downstream flanks amplified using GmF/GmR, DN157/ DN158 and
DN159/ DN160 respectively. Using lambda Red (pKOBEG-Apra) recombination-based allelic exchange,
blaKPC-2 was then swapped with the SOE-PCR (UF-hph-DF) and the mutants ∆KPC:hph were selected on
LA hygromycin at 37℃. Using a flippase expressing plasmid, the hph cassette in ∆KPC:hph was excised
based on flp-FRT recombination to create a marker-less ∆KPC (Figure 3.6). To further knockout the

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
123
pKPHS3 26 kb MDR region (KPHS_p300510-KPHS_p300880) in ∆KPC background, the upstream
(DN165/DN166) and downstream flanks (DN167/DN168) and hph cassette (GmF/GmR) were used to form
a mutant allele/ SOE-PCR product. Again, the MDR mutant allele was then used to replace the 26 kb MDR
region using lambda recombination to create ∆KPC∆MDR:hph (DN2T) at 37℃. Then, the hph cassette in
∆KPC∆MDR:hph was again flipped out using flippase expressing plasmid to induce flp-FRT
recombination, resulting in the final attenuated DN2 or ∆KPC∆MDR (Figure 3.6).
After the deletion of blaKPC-2 and 26 kb MDR region that carries genes transposases, regulators and
hypothetical proteins with vast resistance arsenal, the final mutant DN2 (∆KPC∆MDR) was susceptible to
a wider group of antibiotics (Appendix 2-2 Antibiotic susceptibility of K. pneumoniae HS11286 and the
antibiotic disarmed versions) but not certain b-lactam (due to the residual blaTEM-1, blaCTX-M-14 and blaSHV-
11). The MIC data of DN2 mutant also showed attenuated resistance to certain b-lactam, suggesting the loss
of the blaTEM-1 and blaKPC-2 contribute to some form of b-lactam resistance. Additionally, both mutants
(∆KPC and DN2) showed no reduction in Fosfomycin (due to fosA), nalidixic acid and subtle reduction to
ciprofloxacin probably due to due Ser83 Ile mutation in gyrA and the Ser80 Ile mutation in parC (Deguchi
et al., 1997; Fu et al., 2008) (Ddeguchi et al., 1997; Fu et al., 2008). Overall, the final mutant ∆KCP∆MDR
(DN2) was safer to genetically manipulate and expands its use as K. pneumoniae model organism for future
studies such T6SS virulence studies as presented in this report.
To create the attenuated antibiotic-resistant version, blaKPC-2 on pKPHS2 was first replaced with FRT-
flanked hph cassette via lambda red recombination and the antibiotic marker cassette was excised through
flippase mediated FRT recombination to create a marker-less ∆KPC (blaKPC-2 mutant). Similarly, a 26 kb
multi-antibiotic resistant region on PKPHS3 was further deleted from ∆KPC by replacing the 26 kb region
with FRT-flanked hph using lambda red recombination. The antibiotic cassette (hph) was then excised by
the recombination of FRT mediated by flippase, resulting in the final attenuated model strain DN2
(∆KPC∆MDR). To confirm the associated attenuation in antibiotic resistance, antibiotic susceptibility test
(Appendix 2-2 Antibiotic susceptibility of K. pneumoniae HS11286 and the antibiotic disarmed versions)
for ∆KPC and DN2 was performed relative to the wild-type strain KR3317.
3.3 Disarmament of antibiotic resistance in K. pneumoniae HS11286 enhanced in vitro fitness not in
vivo virulence in G. mellonella.
The original K. pneumoniae HS11286 strain was difficult to genetically manipulate and pose a significant
health hazard in the event of acc4idental infection during routine laboratory experiments. As such in order
to be able to investigate T6SSs in K. pneumoniae HS11286, a disarmed antibiotic-resistant version of the

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
124
strain (DN2) was created using lambda red recombination-based allelic exchange. This involved the
knockout of blaKPC-2 and a 26 kb antibiotic resistance region (consisting of 12 antibiotic resistance genes
and one biocide resistance gene, Figure 3.5, Figure 3.6 and Appendix 2). To perform the in vitro
competition assay, a 1:1 ratio of the original wild-type strain K. pneumoniae HS11286 (KR3317) and its
antibiotic resistance disarmed version (DN2) were mixed and grown in LB for 24 h at 37℃. Additionally,
both strains were also separately cultured. A 1:100 subculture of the bacterial mixture was made into a fresh
LB and grown for another 24h. At each time-point (day 1 and 2), a sample from the growing mixed culture
was then plated onto LA and LA plus antibiotics and the CFU were enumerated. Also, Galleria mellonella
larvae killing assay was then used to measure whether there was reduced virulence in DN2 after the
disarming of the antibiotic resistance region relative to its parent strain, KR3317. Briefly, 5×106 CFU of
either KR3317 and ‘resistance-disarmed’ DN2 strain were used to inoculate the larvae of Galleria
mellonella (at least 30 in each group), and the survival of these larvae was measured of 6 days period. Death
of Galleria larvae was characterised by lack of sensitivity or response, immobility of the larva and
melanisation between 24 h interval inspection post-infection with the various bacteria strains.
The data (Figure 3.7A) obtained from the in vitro culture revealed that more DN2 were recovered from the
mixture more than KR3317, suggesting that the resistance disarmed DN2 was much fitter and outcompeted
its parent strain. The deletion of the blaKPC-2 and the 26 kb MDR region significantly enhanced the fitness
of the DN2 relative to the parent strain, which encountered a huge physiological fitness cost just by bearing
more antibiotic resistance regions. This was not very surprising as earlier studies have reported that a large
acquisition of antibiotic resistance region poses a high cost to that bacteria growth. Also, the antibiotic
susceptibility data (see Appendix 2-2 Antibiotic susceptibility of K. pneumoniae HS11286 and the
antibiotic disarmed versions) revealed that the resistance-disarmed strain (DN2) became very susceptible
to many antibiotics including ceftazidime, Ceftazidime, imipenem, meropenem, gentamicin, tobramycin,
amikacin, streptomycin, kanamycin, levofloxacin, trimethoprim-sulfamethoxazole, trimethoprim and
chloramphenicol. The predicted genes with the 26 kb antibiotic region thought to be responsible for the
antibiotic resistance in KR3317 are tabulated in Appendix 2-1: List of genes deleted from the 26 kb region.
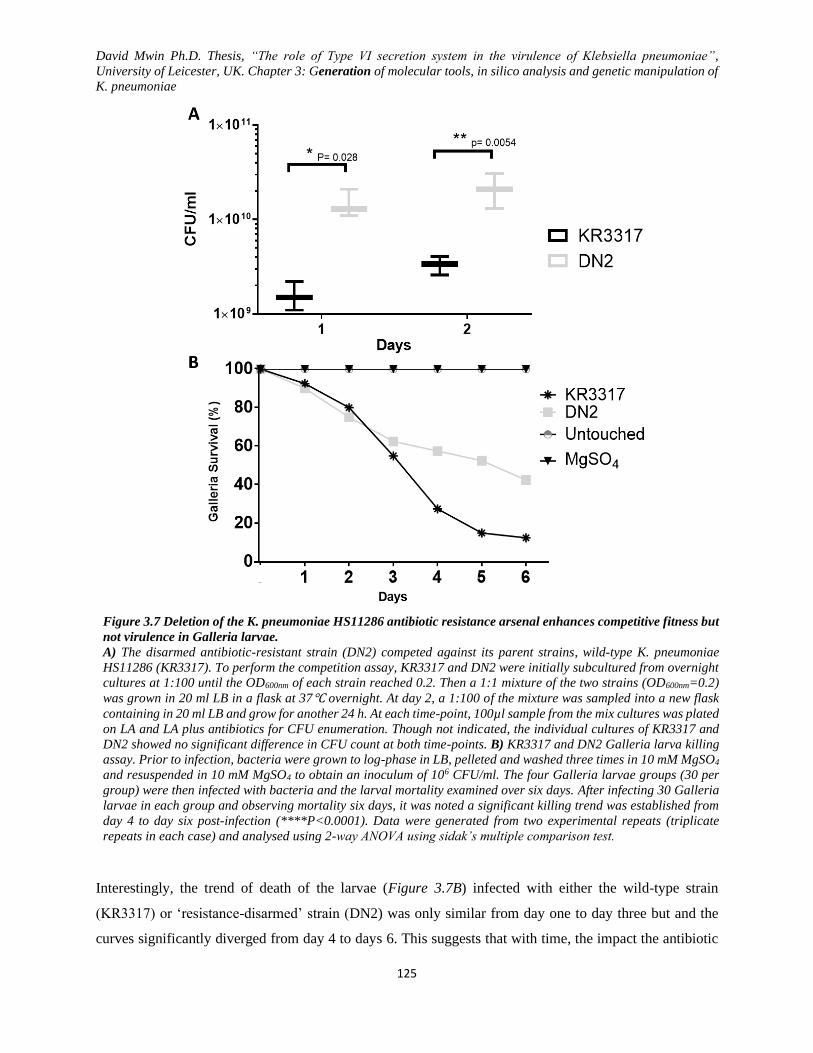
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
125
Figure 3.7 Deletion of the K. pneumoniae HS11286 antibiotic resistance arsenal enhances competitive fitness but
not virulence in Galleria larvae.
A) The disarmed antibiotic-resistant strain (DN2) competed against its parent strains, wild-type K. pneumoniae
HS11286 (KR3317). To perform the competition assay, KR3317 and DN2 were initially subcultured from overnight
cultures at 1:100 until the OD600nm of each strain reached 0.2. Then a 1:1 mixture of the two strains (OD600nm=0.2)
was grown in 20 ml LB in a flask at 37℃ overnight. At day 2, a 1:100 of the mixture was sampled into a new flask
containing in 20 ml LB and grow for another 24 h. At each time-point, 100µl sample from the mix cultures was plated
on LA and LA plus antibiotics for CFU enumeration. Though not indicated, the individual cultures of KR3317 and
DN2 showed no significant difference in CFU count at both time-points. B) KR3317 and DN2 Galleria larva killing
assay. Prior to infection, bacteria were grown to log-phase in LB, pelleted and washed three times in 10 mM MgSO4
and resuspended in 10 mM MgSO4 to obtain an inoculum of 106 CFU/ml. The four Galleria larvae groups (30 per
group) were then infected with bacteria and the larval mortality examined over six days. After infecting 30 Galleria
larvae in each group and observing mortality six days, it was noted a significant killing trend was established from
day 4 to day six post-infection (****P<0.0001). Data were generated from two experimental repeats (triplicate
repeats in each case) and analysed using 2-way ANOVA using sidak’s multiple comparison test.
Interestingly, the trend of death of the larvae (Figure 3.7B) infected with either the wild-type strain
(KR3317) or ‘resistance-disarmed’ strain (DN2) was only similar from day one to day three but and the
curves significantly diverged from day 4 to days 6. This suggests that with time, the impact the antibiotic

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
126
resistance genes deletion may be more observable given enough time. DN2 strain killed the least larvae,
unlike KR3317 strain that nearly wiped out the entire infected larvae group after day 6. Just as the deletion
of antibiotic resistance made the strain susceptible to a wide range of antibiotics (see Appendix 2-2
Antibiotic susceptibility of K. pneumoniae HS11286 and the antibiotic disarmed versions), so did the
antibiotic resistance deletion through an unknown mechanism enhance DN2 strain susceptibility to the
Galleria larva immune system even though DN2 strain showed an enhanced growth fitness relative to the
KR3317 in the in vitro growth assay.
Many studies have linked antibiotic resistance with virulence (Kidd et al, 2017; Padilla et al, 2010b; Kidd
et al, 2016) which was reaffirmed by the data obtained from the in vitro and Galleria larvae killing assays.
The data suggest that K. pneumoniae HS11286 antibiotic resistance region posed a significant physiological
burden to maintain though it enhanced the virulence of KR3317 towards the moth larvae. Though DN2 was
safer to work with as a model due to the disarmed resistance, its ability to kill the larvae was stunted relative
to the parent, KR3317. Hence, acquisition of antibiotic resistance contributes to the virulence of the
bacterial strain enable it to evade the host immune system while inducing a massive physiologically costly
in the absence of selection pressure.
3.4 3.6 K. pneumoniae HSS11286 T6SS clusters are syntenic with similarity in flanking regions to
other K. pneumoniae strains
Sarris and colleagues (2011) identified several T6SS gene clusters in K. pneumoniae based on in silico data
(Sarris et al., 2010). Here, in silico analysis was performed using several databases but most importantly,
NCBI to obtained deposited sequence (reference genome data). Also, plasmid editor and snapgene tools
were employed in this analysis to further the course of this research piece. A comparative genomic analysis
of 14 fully sequenced genomes of K. pneumoniae was analysed (including K. pneumoniae HS11286) with
a particular focus on the identified T6SS gene clusters. Core T6SS members’ DNA and protein sequence
similarities and syntenies were also performed (data not shown here) to elucidate the intra- and the inter-
strain relationship between the putative T6SS gene clusters.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
127
Table 3.1 K. pneumoniae HS11286 T6SS1 gene cluster: context and content
S/No Protein Locus Tag Strand Location Length (bp) Description
1 KPHS_22930 + 2322412..2322621 210 hypothetical protein
2 KPHS_22940 + 2323071..2324558 1488 putative reverse transcriptase/maturase
3 KPHS_22950 - 2325143..2325391 249 DinI family protein
4 KPHS_22960 + 2325956..2326078 123 hypothetical protein
5 ImpB/ TssB KPHS_22970 + 2326237..2326728 492 hypothetical protein
6 ImpC/ TssC KPHS_22980 + 2326771..2328315 1545 EvpB/family type VI secretion protein
7 ImpJ/ TssK KPHS_22990 + 2328325..2329668 1344 hypothetical protein
8 DotU/ TssL KPHS_23000 + 2329665..2330354 690 hypothetical protein
9 OmpA KPHS_23010 + 2330351..2332057 1707 OmpA/MotB domain-containing protein
10 Hcp/ TssD KPHS_23020 + 2332062..2332553 492 hypothetical protein
11 ClpV/ TssH KPHS_23030 + 2332818..2335472 2655 putative protease
12 VgrG/ TssI KPHS_23040 + 2335474..2337843 2370 Rhs element Vgr protein
13 KPHS_23050 + 2337844..2338623 780 hypothetical protein
14 KPHS_23060 + 2338687..2339217 531 putative lipoprotein
15 KPHS_23070 + 2339286..2339816 531 putative lipoprotein
16 KPHS_23080 + 2339884..2340414 531 putative lipoprotein
17 KPHS_23090 + 2340482..2341012 531 putative lipoprotein
18 KPHS_23100 + 2341000..2342034 1035 hypothetical protein
19 KPHS_23110 + 2342120..2343418 1299 hypothetical protein
20 KPHS_23120 + 2343463..2343720 258 PaaR repeat-containing protein
21 KPHS_23130 + 2343717..2344856 1140 hypothetical protein
22 IcmF/TssM KPHS_23140 + 2344840..2348265 3426 ImcF domain-containing protein
23 ImpA/ TssA KPHS_23150 + 2348265..2348672 408 type VI secretion-associated protein
24 TssA-like KPHS_23160 + 2348724..2349857 1134 type VI secretion-associated protein
25 ImpG/ TssF KPHS_23170 + 2349937..2351691 1755 hypothetical protein
26 ImpH/ TssG KPHS_23180 + 2351655..2352740 1086 hypothetical protein
27 VasD/ TssJ KPHS_23190 + 2352718..2353260 543 hypothetical protein
28 KPHS_23200 - 2353231..2353359 129 hypothetical protein
29 KPHS_23210 + 2353389..2353883 495 hypothetical protein
30 KPHS_23220 + 2353950..2354663 714 Oxidoreductase
31 KPHS_23230 + 2354737..2355330 594 putative regulatory protein TetR
32 KPHS_23240 - 2355335..2356090 756 Putative dehydrogenase/reductase oxidoreductase
Note: T6SS1 gene cluster is shaded grey with core T6SS proteins boldened and the flanking upstream (1-4) and
downstream (28-32) flanking region/genes are unshaded.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
128
The result obtained from these analyses suggest that K. pneumoniae HS11286 T6SS gene clusters possess
similar upwards flanking (UF) sequences just like many other K. pneumoniae strains as well as similar
downwards flanking (DF) regions, hence a possible evolutionary relatedness. T6SS1 and T6SS3 are flanked
by conserved regions both upwards and downwards unlike T6SS2. The reserved regions extend over 10 kb
for the UF and nearly 5 kb for the DF. While T6SS1 is complete and may be functionally independent of
the other clusters, T6SS3 lack some core genes such as clpV and hcp. The result suggests that T6SS2 in K.
pneumoniae HS11286 is a degenerated or incomplete cluster and is possibly carrying the T6SS3 missing
set of genes such as clpV and hcp. The insights from these basic bioinformatics analyses indicate that K.
pneumoniae HS11286 T6SS2 may only be evolutionarily displaced but still complements T6SS3 for
functional reasons. Thus, the bacterium has only two sets of functional T6SS gene clusters which are T6SS1
and T6SS3 (the two clusters consider for T6SS mutant generation). Within each cluster, several other genes
whose function is unknown or unrelated to T6SSs were found to be present in both T6SS1 and T6SS3. A
complete deletion of T6SS1 and T6SS3 were using allelic exchange method was used, this created the non-
polar mutants (∆T1, ∆T3 ∆T1∆T3) that were used for K. pneumoniae virulence assays.
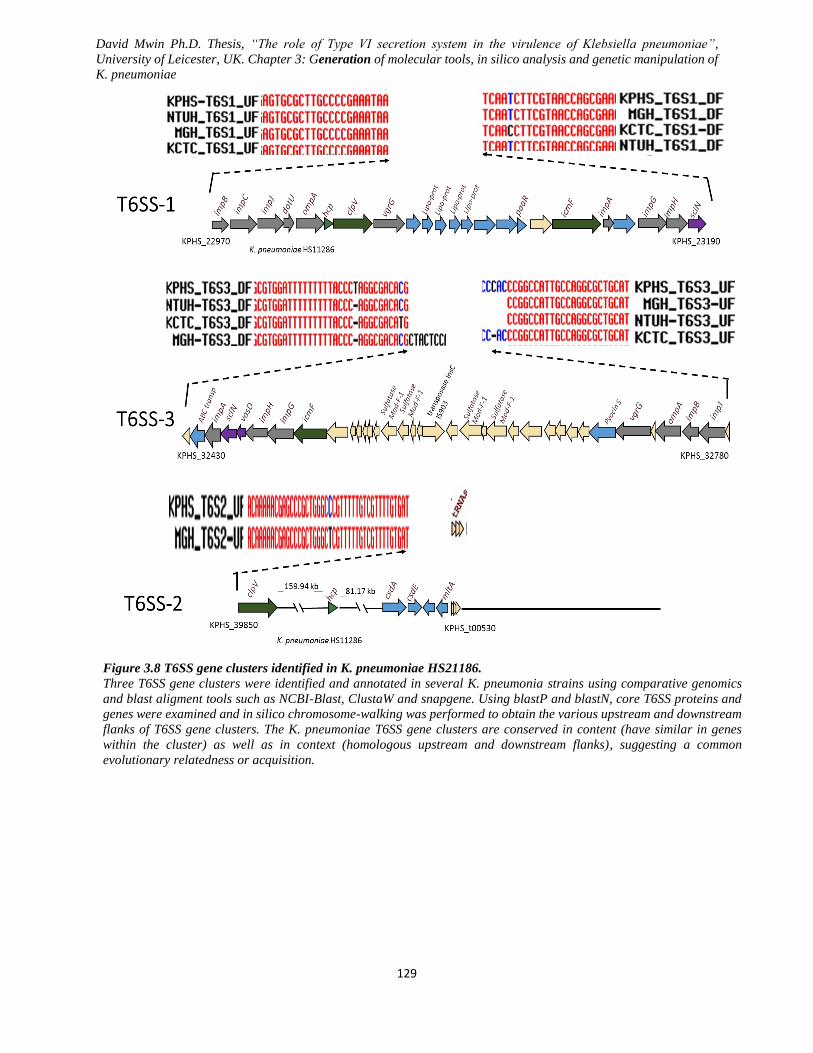
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
129
Figure 3.8 T6SS gene clusters identified in K. pneumoniae HS21186.
Three T6SS gene clusters were identified and annotated in several K. pneumonia strains using comparative genomics
and blast aligment tools such as NCBI-Blast, ClustaW and snapgene. Using blastP and blastN, core T6SS proteins and
genes were examined and in silico chromosome-walking was performed to obtain the various upstream and downstream
flanks of T6SS gene clusters. The K. pneumoniae T6SS gene clusters are conserved in content (have similar in genes
within the cluster) as well as in context (homologous upstream and downstream flanks), suggesting a common
evolutionary relatedness or acquisition.
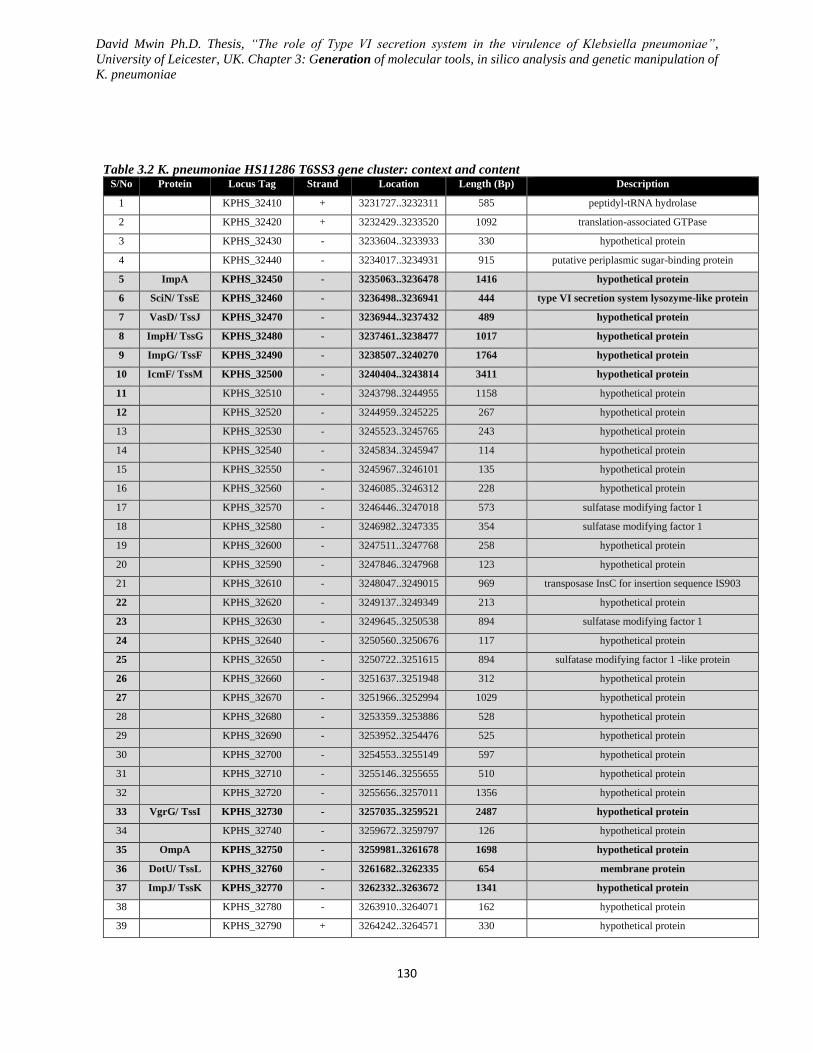
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
130
Table 3.2 K. pneumoniae HS11286 T6SS3 gene cluster: context and content S/No Protein Locus Tag Strand Location Length (Bp) Description
1 KPHS_32410 + 3231727..3232311 585 peptidyl-tRNA hydrolase
2 KPHS_32420 + 3232429..3233520 1092 translation-associated GTPase
3 KPHS_32430 - 3233604..3233933 330 hypothetical protein
4 KPHS_32440 - 3234017..3234931 915 putative periplasmic sugar-binding protein
5 ImpA KPHS_32450 - 3235063..3236478 1416 hypothetical protein
6 SciN/ TssE KPHS_32460 - 3236498..3236941 444 type VI secretion system lysozyme-like protein
7 VasD/ TssJ KPHS_32470 - 3236944..3237432 489 hypothetical protein
8 ImpH/ TssG KPHS_32480 - 3237461..3238477 1017 hypothetical protein
9 ImpG/ TssF KPHS_32490 - 3238507..3240270 1764 hypothetical protein
10 IcmF/ TssM KPHS_32500 - 3240404..3243814 3411 hypothetical protein
11 KPHS_32510 - 3243798..3244955 1158 hypothetical protein
12 KPHS_32520 - 3244959..3245225 267 hypothetical protein
13 KPHS_32530 - 3245523..3245765 243 hypothetical protein
14 KPHS_32540 - 3245834..3245947 114 hypothetical protein
15 KPHS_32550 - 3245967..3246101 135 hypothetical protein
16 KPHS_32560 - 3246085..3246312 228 hypothetical protein
17 KPHS_32570 - 3246446..3247018 573 sulfatase modifying factor 1
18 KPHS_32580 - 3246982..3247335 354 sulfatase modifying factor 1
19 KPHS_32600 - 3247511..3247768 258 hypothetical protein
20 KPHS_32590 - 3247846..3247968 123 hypothetical protein
21 KPHS_32610 - 3248047..3249015 969 transposase InsC for insertion sequence IS903
22 KPHS_32620 - 3249137..3249349 213 hypothetical protein
23 KPHS_32630 - 3249645..3250538 894 sulfatase modifying factor 1
24 KPHS_32640 - 3250560..3250676 117 hypothetical protein
25 KPHS_32650 - 3250722..3251615 894 sulfatase modifying factor 1 -like protein
26 KPHS_32660 - 3251637..3251948 312 hypothetical protein
27 KPHS_32670 - 3251966..3252994 1029 hypothetical protein
28 KPHS_32680 - 3253359..3253886 528 hypothetical protein
29 KPHS_32690 - 3253952..3254476 525 hypothetical protein
30 KPHS_32700 - 3254553..3255149 597 hypothetical protein
31 KPHS_32710 - 3255146..3255655 510 hypothetical protein
32 KPHS_32720 - 3255656..3257011 1356 hypothetical protein
33 VgrG/ TssI KPHS_32730 - 3257035..3259521 2487 hypothetical protein
34 KPHS_32740 - 3259672..3259797 126 hypothetical protein
35 OmpA KPHS_32750 - 3259981..3261678 1698 hypothetical protein
36 DotU/ TssL KPHS_32760 - 3261682..3262335 654 membrane protein
37 ImpJ/ TssK KPHS_32770 - 3262332..3263672 1341 hypothetical protein
38 KPHS_32780 - 3263910..3264071 162 hypothetical protein
39 KPHS_32790 + 3264242..3264571 330 hypothetical protein

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
131
40 KPHS_32800 + 3264686..3265225 540 nucleoprotein/polynucleotide-associated enzyme
41 KPHS_32810 - 3265251..3266042 792 ribosomal large subunit pseudouridine synthase A
T6SS3 gene cluster (shaded grey) with core T6SS proteins boldened and flanking upstream (1-4) and downstream (38-41) flanking region/genes

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
132
Genome sequence viewer was used to view the genomic location of the T6SS1, T6SS3 and purported TSS2
from an aerial view, and it was stunningly shocking to note the exact similarities in the pattern and location
the various T6SS clusters with respect to the origin. The location and arrangement of T6SS gene clusters
within K. pneumoniae genomes were observed to appear to have a unique arrangement pattern (T6SS1
coded grey, T6SS3 coded yellow and T6SS2 coded light brown). Based on the 14 analysed K. pneumoniae
genomes, the arrangement of these T6SS gene clusters was grouped ton sub into three subgroups. Hence,
the results obtained from the genomic sequence viewing suggest syntenic and similarities in the distribution
of T6SS gene clusters and flanking regions as well as relatedness even at arrangement/ pattern of the T6SS
gene cluster from an aerial viewpoint.
Group A: K. pneumoniae JM45, K. pneumoniae 342, K. pneumoniae 234-12, K. pneumoniae 1084 showing a similar
pattern or arrangement of T6SS1, T6SS2 and T6SS3
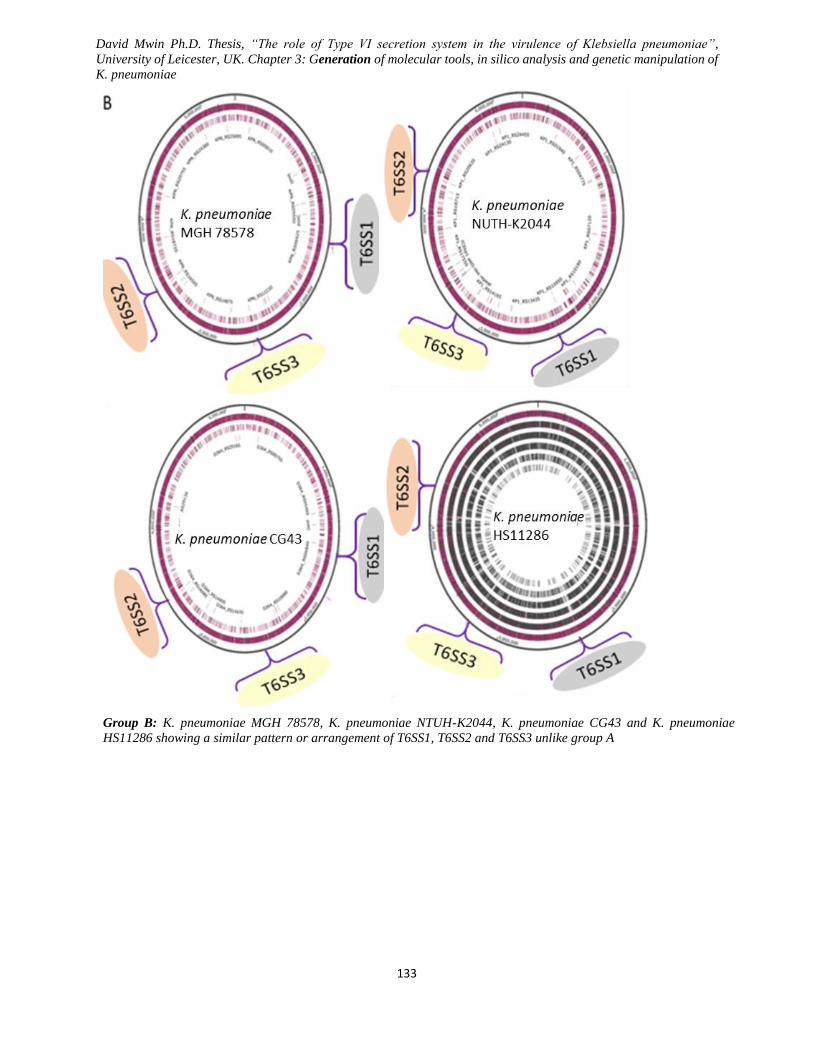
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
133
Group B: K. pneumoniae MGH 78578, K. pneumoniae NTUH-K2044, K. pneumoniae CG43 and K. pneumoniae
HS11286 showing a similar pattern or arrangement of T6SS1, T6SS2 and T6SS3 unlike group A

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
134
Group A: K. pneumoniae INH27, K. pneumoniae IH31, K. pneumoniae subsp. Rhinoscleromatis SB3432, K.
pneumoniae ATTC BAA-2146, K. pneumoniae 52-145 and K. pneumoniae KCTC2242 showing a similar pattern or
arrangement of T6SS1, T6SS2 and T6SS3.
Figure 3.9 Arrangement of T6SS1, T6SS2 and T6SS3 gene clusters using genome viewer.
Using snapgene genome viewing tool in combination with universally designed K. pneumoniae T6SS primers, T6SS1,
T6SS3 (both with functional gene clusters/genes) and T6SS2 (unfunctional gene cluster) in about 14 completely
sequenced K. pneumoniae strains were identified and grouped. Conserved T6SS genes were largely present in K.
pneumoniae, and the three T6SS gene clusters can be probably grouped in similar cluster context and content relative
to the genome. From the above figures, A had slightly different T6SS cluster arrangement pattern compared to B and C
despite the overall similarity in gene content and context.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
135
3.8 Deleting mapped T6SS clusters in resistance-disarmed K. pneumoniae HS11286 for further T6SS
studies
Majority of resistance determinants in K. pneumoniae HS11286, a representative of the dominant CRKP
ST11 lineage, are plasmid-borne. The wider genetic contexts associated with resistance genes may have
undergone complex evolutionary events, driven at least in part by ISs, transposons and integrons, which
have resulted in a gradual accumulation of resistance genes and a potential structural, mobility-linked and
expression-associated optimization of these hazardous DNA assemblages. The multi-drug resistant K.
pneumoniae strain was successfully disarmed of most of the antibiotic-resistant gene resulting in a safer
and antibiotic-susceptible strain, DN2, but still exhibiting some level of virulence level when test in
Galleria Killing model relative parent strain. Thus, the deletion of these antibiotic regions did not have any
impact on the virulence, suggesting that there are other virulent factors expressed by the strain other than
antibiotic resistance, these results support the notion that antibiotic resistance contributes to the bacterial
virulence and the overall survival and disease-causing potential of the bacterium.
Using defined nucleotide and protein sequence alignments of upward flanks (UF) and downward flanks
(DF) of three K, pneumoniae (NTUH-K2044, KCTC AND MGH 78578) T6SS gene clusters from online
database and as defined in Sarris et al., (2011), T6SS1, T6SS2 and T6SS3 were captured. All the UFs of
T6SS clusters in these strains showed near-perfect 10 kb alignment, and similarly, 5 kb DF of the T6SS
clusters was conserved in T6SS1 and T6SS3 but not T6SS2. The lack of conserved DF of T6SS2 was not
surprising since this cluster only had two core genes (hcp and clpV) which were distantly apart. Even though
three T6SS gene clusters were identified and mapped out, further analysis suggested that the genome of the
two K. pneumoniae strains (HS11286 and NTUH-K2044) only had two complete/ functional set of T6SS
gene clusters (T6SS1 and T6SS3). T6SS2 was disintegrated and only had two core gene (hcp and clpV)
which surprisingly are the only core genes missing from the T6SS3 gene cluster, suggesting that T6SS2
genes (hcp and clpV) could be complementing T6SS3, unlike T6SS1 that had a complete set functional of
T6SS core genes. From the T6SS in silico analysis using a wide range of bioinformatic tools, a step-by-step
concept of capturing, identifying and successfully knocking out the entire T6SS clusters or individual genes
in clinical strains was employed based on the T6SS annotation in the genome of fully sequenced K.
pneumoniae strains. Also, the knowledge obtained from these bioinformatic analyses could be extended to
other Gram-negative bacterial strain carrying T6SS gene clusters since most of the T6SS genes tend to
share high similarity across various bacterial species. The conserved upstream (UF) and downstream (DF)
flanks suggest that T6SS may not be native to the bacteria but may have been acquired via horizontal gene
transfer and later evolved.
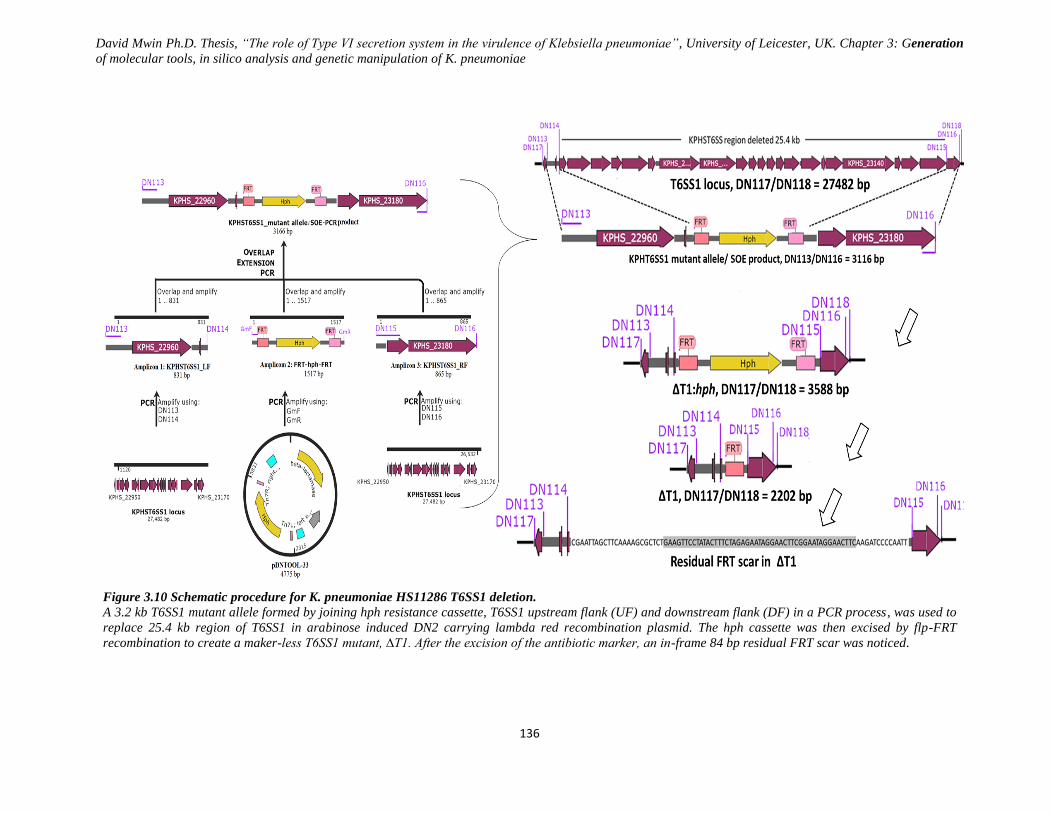
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Chapter 3: Generation
of molecular tools, in silico analysis and genetic manipulation of K. pneumoniae
136
Figure 3.10 Schematic procedure for K. pneumoniae HS11286 T6SS1 deletion.
A 3.2 kb T6SS1 mutant allele formed by joining hph resistance cassette, T6SS1 upstream flank (UF) and downstream flank (DF) in a PCR process, was used to
replace 25.4 kb region of T6SS1 in arabinose induced DN2 carrying lambda red recombination plasmid. The hph cassette was then excised by flp-FRT
recombination to create a maker-less T6SS1 mutant, ∆T1. After the excision of the antibiotic marker, an in-frame 84 bp residual FRT scar was noticed.
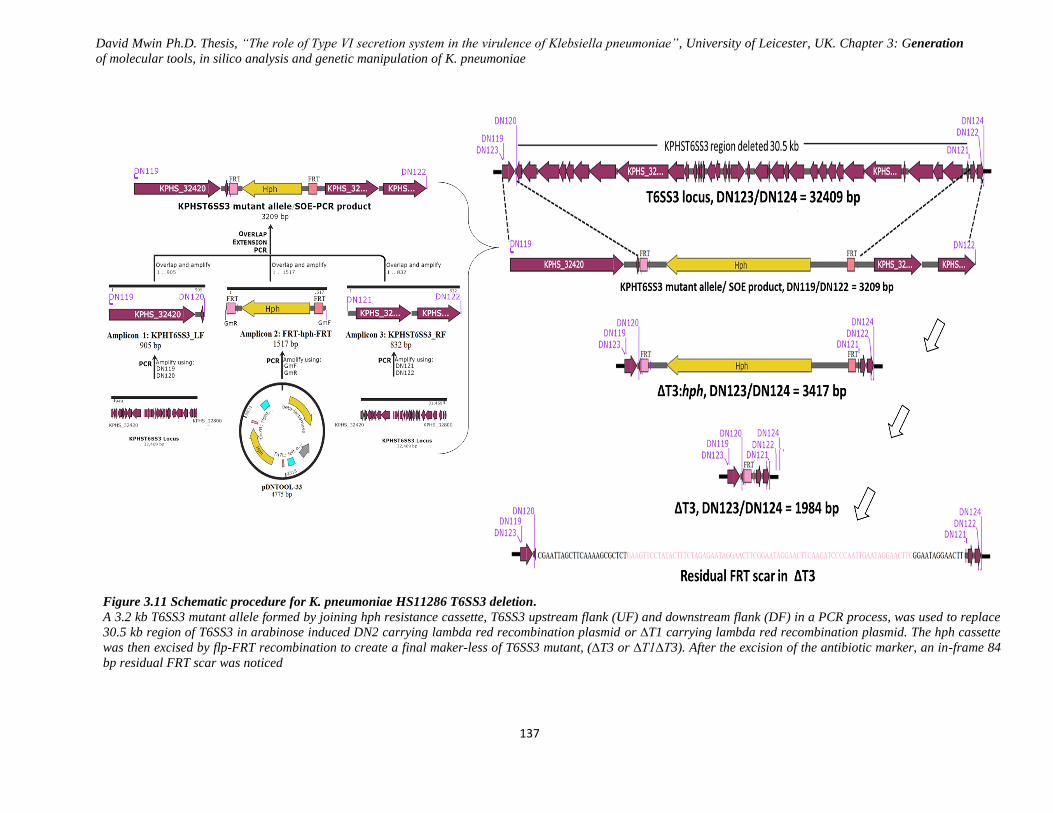
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”, University of Leicester, UK. Chapter 3: Generation
of molecular tools, in silico analysis and genetic manipulation of K. pneumoniae
137
Figure 3.11 Schematic procedure for K. pneumoniae HS11286 T6SS3 deletion.
A 3.2 kb T6SS3 mutant allele formed by joining hph resistance cassette, T6SS3 upstream flank (UF) and downstream flank (DF) in a PCR process, was used to replace
30.5 kb region of T6SS3 in arabinose induced DN2 carrying lambda red recombination plasmid or ∆T1 carrying lambda red recombination plasmid. The hph cassette
was then excised by flp-FRT recombination to create a final maker-less of T6SS3 mutant, (∆T3 or ∆T1∆T3). After the excision of the antibiotic marker, an in-frame 84
bp residual FRT scar was noticed

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
138
The two main T6SS gene clusters were deletion from the DN2 background via lambda red recombination-
based allelic exchange where a mutant allelic carrying the antibiotic cassette mostly (hph or aacC1) was
used to replace the T6SS1 or T6SS3 gene cluster. A flippase expressing plasmid was then introduced into
the T6SS mutant carrying the antibiotic marker to induce flp-FRT recombination to excise the antibiotic
marker. All the T6SS mutants were PCR verified, and amplicons of the mutated T6SS regions were
sequenced. Briefly, the UF and DF of the T6SS1 of T6SS3 were amplified, and SOE-PCR joined together
with FRT-flanked hph antibiotic marker to produce a mutant allele ( see Figure 3.10, Figure 3.11 and Figure
3.12). The T6SS mutant alleles were electroporated into arabinose induced DN2 carrying lambda red
recombination plasmid (for T6SS1 or T6SS3 single knockout) or ∆T1 carrying lambda red recombination
plasmid (for T6SS1 and T6SS3 double knockout). After each knockout, the temperature-sensitive lambada
red recombination plasmid (usually maintained at (25-30℃) was eliminated by growing the mutants at 37℃.
Also, the flippase expressing plasmid (pDNTOOL-2) was eliminated by growing the mutant on LA plus 5-
10% sucrose; the sacB gene in the presence of sucrose express levansucrase, which is lethal to the bacteria.
As a result, the bacteria kid out the plasmid to avoid expressing the lethal levansucrase.
Unlike the deletion of the T6SS gene clusters in K. pneumoniae HS11286, T6SS1 deletion in K. pneumoniae
NTUH-K2044 was doing using a constructed suicide deletion vector (pDNTOOL-8). Briefly, the K.
pneumoniae T6SS1 mutant allele cloned into pJTOOL-3 to form pDNTOOL-8. The deletion vector was
then electroporated into E. coli cc118 λpir and later into E. coli s17. Using the filter mating technique, the
E. coli S17-λpir carrying the plasmid was then conjugated with hyper-mucoid K. pneumoniae NTUH-
K2044. Single crossover mutants were selected, and PCR verified before induced double crossover by
growing the bacteria in LA plus 5% sucrose. The final double-crossover mutant (NTUH∆T1) was then PCR
confirmed and sequenced. Similarly, deletion of K. pneumoniae NTUH-K2044 T6SS3 gene cluster was
attempted, and only a single crossover was obtained, but the double crossover failed. The failure to obtain
the double crossover mutants of K. pneumoniae NTUH-K2044 T6SS3 gene cluster maybe because of some
essential genes within that gene cluster.
Using the above-described method, T6SS1 mutant (∆T1), T6SS3 mutant (∆T3) and T6SS1 and T6SS3
double mutant (∆T1∆T3) all in the resistance disarmed K. pneumoniae HS11286 background (Figure 3.12)
and NTUH∆T1 in K. pneumoniae NTUH-K2044 background were created and verified by PCR and
sequencing. Based on the sequence analysis of the mutated region, only one in-frame 84 bp FRT scar was
left behind after T6SS deletion and subsequent antibiotic marker excision.
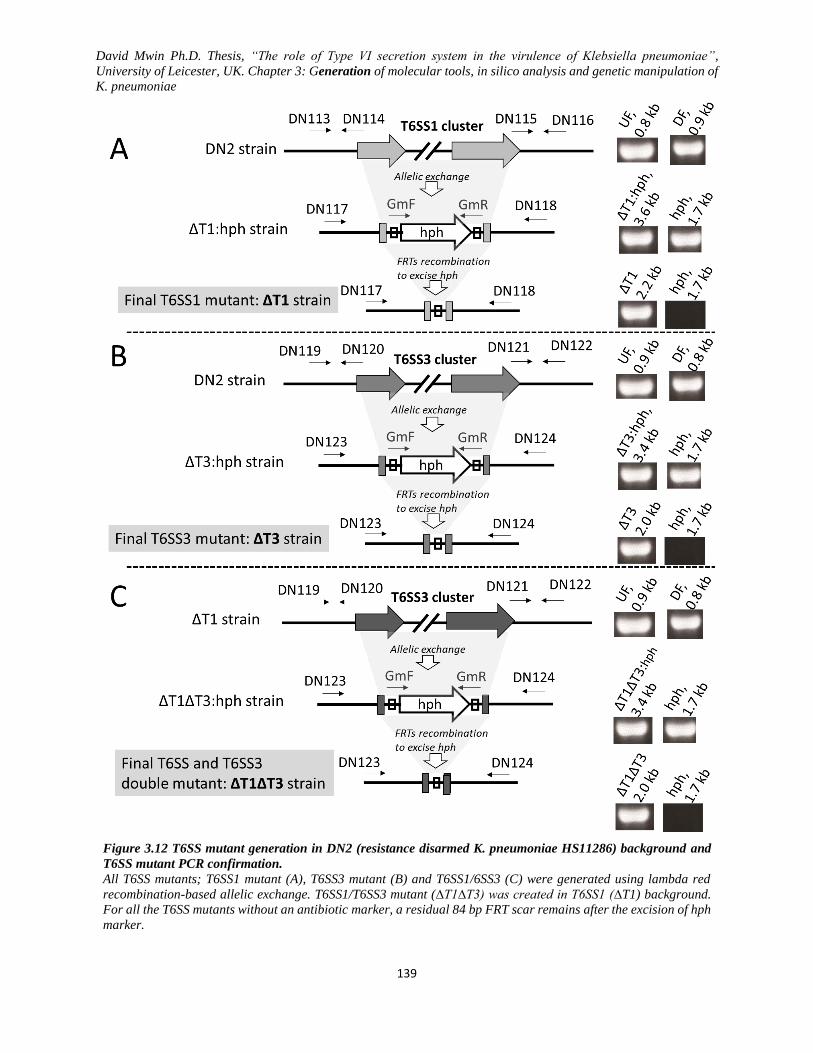
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
139
Figure 3.12 T6SS mutant generation in DN2 (resistance disarmed K. pneumoniae HS11286) background and
T6SS mutant PCR confirmation.
All T6SS mutants; T6SS1 mutant (A), T6SS3 mutant (B) and T6SS1/6SS3 (C) were generated using lambda red
recombination-based allelic exchange. T6SS1/T6SS3 mutant (∆T1∆T3) was created in T6SS1 (∆T1) background.
For all the T6SS mutants without an antibiotic marker, a residual 84 bp FRT scar remains after the excision of hph
marker.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 3: Generation of molecular tools, in silico analysis and genetic manipulation of
K. pneumoniae
140

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
141
Chapter Four
K. pneumoniae T6SS enhances bacterial fitness,
fimbria-mediated biofilm formation and resistance
and survival in amoeba

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
142
Chapter 4: K. pneumoniae T6SS enhance bacteria-bacteria
interaction and biofilm-like formation
4.1 Overview
Presented in this chapter is the role the putative T6SSs gene clusters play in K. pneumoniae bacteria-bacteria
interactions, biofilm formation, surface fimbriation and amoeba phagocytic resistance. The Intra-strain
competitive fitness of K. pneumoniae was assessed by competing DN2 (parent strain) against its isogenic
T6SS mutants (∆T1, ∆T3 and ∆T1∆T3) and recovering the bacteria growth from bacteria mixed cultures
over the examined duration. The T6SS mutants lost the competition relative to DN2 over the examined
duration, with ∆T1∆T3, ∆1T1 and ∆T3 showing increasing order of viable bacteria recovery, respectively,
relative to DN2. In an inter-species competition, K. pneumoniae DN2 inhibited the growth of Escherichia
coli (Non-T6SS prey) in a T6SS-dependent manner whilst the T6SS mutants hardly inhibited the prey.
Also, Pseudomonas aeruginosa (T6SS competitor) competed against DN2 and the T6SS mutants, unlike
DN2, the mutants easily lost the competition to P. aeruginosa when the bacteria growths were assessed
from the mixed culture. This antagonistic property of T6SS affects how K. pneumoniae interacts with other
bacteria in a complex ecosystem where other isogenic T6SS mutants (intra-strain competition) or other
T6SS and non-T6SS harbouring bacteria such as P. aeruginosa and E. coli (inter-species competition)
respectively, will face the defensive-attaching wrath of K. pneumoniae T6SSs. The data suggest that T6SS
is offensive and defensive nano-arsenal and is a critical factor for K. pneumoniae survival and subsequent
colonisation of the polymicrobial environment with limiting resources. K. pneumoniae in polymicrobial
infection may deploy it T6SS to dominant or resist attack from other T6SS harbouring bacteria. The data
also suggest K. pneumoniae T6SSs may play a role theT4SS-dependent horizontal transfer of antibiotic
resistance and other virulence factors.
Furthermore, deletion of T6SS affected the ability of K. pneumoniae to form biofilm on an abiotic surface
(polystyrene and glass coverslip) and surface fimbriae expression. When T6SS1 or T6SS3 was knocked
out, ∆T1 and ∆T3 showed enhanced type 1 and type 3 fimbriae-dependent biofilm formation with ∆T3 is
being the most biofilm former of all. Just like DN2, ∆T1∆T3 surprisingly showed little to no difference in
the amount of biofilm formation relative to DN2. Thus, the TEM visualisation of the bacterial fimbriae
agrees with their respective biofilm phenotype. T6SS-dependent phagocytic resistance was noted in K.
pneumoniae against A. castellanii. Finally, the ability for K. pneumoniae to combined T6SS antagonistic-
defensive behaviour, T4SS-dependent dissemination of resistance, biofilm formation and amoebal
phagocytic resistance, will potentially enhance the pathogenicity and environmental persistence of K.
pneumoniae.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
143
Bacteria are ubiquitous in nature and evolve to adapt to their complex ecological niche where they often
compete for the limiting resources, of which. K. pneumoniae is not an exception. With over 25% sequenced
Gram-negative bacteria harbouring T6SS gene clusters, T6SSs are widely implicated in host-pathogen
interaction and bacterial antagonism (Boyer et al., 2009b; Zheng and Leung, 2007b). T6SS, as a puncturing
and toxin delivery organelle, that contributes to the bacteria social interaction in a complex microbial
community. Though available data describes T6SS in Vibrio, Pseudomonas, Edwardsiella, Bordetella,
Bulkodaria, Escherichia, Acinetobacter, Serratia and many other bacteria species (Decoin et al., 2014b;
Murdoch et al., 2011a; Sanchez, 2011; Unterweger et al., 2014b; MacIntyre et al., 2010a; Zheng, Ho and
Mekalanos, 2011b; Chen et al., 2015; Chieng, Mohamed and Nathan, 2015; Schwarz et al., 2010a; Schwarz
et al., 2014), virtually nothing is known about K. pneumoniae, a clinically relevant bacterium in nosocomial
infections. Saris and colleagues reported the presence of putative T6SS clusters in some K. pneumoniae
strain based on bioinformatics data (Sarris et al., 2011b) though until now, nothing is known about the role
of these putative T6SS gene clusters in this important opportunistic pathogen, K. pneumoniae.
T6SS has been implicated in a range of functions, noticeably important bacterial fitness (Murdoch et al.,
2011a; Unterweger et al., 2014b; Schwarz et al., 2010a; Hood et al., 2010c). T6SS has been reported to be
involved in a myriad of bacterial activities such as biofilm formation, horizontal gene transfers, fitness
quorum sensing regulation, and in both the enhancement and attenuation of virulence (Hood et al., 2010a;
Aschtgen et al., 2008; De Pace et al., 2010; Enos-Berlage et al., 2005; Borgeaud et al., 2015a; Das et al.,
2002; Weber et al., 2009; Zheng et al., 2010; Filloux, 2009). There seems to be a wide range of mechanisms
underpinning how the conserved T6SS is able to exert it defence-deleterious effect on other cells, one of
such mechanisms using the C-terminal extension on VgrG. T6SS is co-expressed with other bacterial
virulent factors). Using eukaryotic cell and whole animal model, T6SS has been shown to exhibit a virulent
role while showing antibacterial or inhibitory role towards other non-immune bacteria (Schwarz et al.,
2010a). Competitive fitness is a necessary natural strategy used by many bacteria to enable them to obtain
the crucial resources for their growth in polymicrobial niches. Many pathogenic gram-negative bacteria
employ the phage-tail-like T6SS nano-syringe as an arsenal against other non-immune competing bacteria.
Based on Insilco data, Sarris and colleagues previously reported the presence of putative T6SS gene clusters
in Klebsiella Species in K. pneumoniae (Sarris et al., 2011b) even though there is not enough experimental
data demonstrating the role of these putative gene clusters in these clinically significant species. Being
opportunistic pathogen and ubiquitous, K. pneumoniae is faced with both biological and environmental
factors in its quest to colonise a niche. K. pneumoniae is posed with intra-species and interspecies fitness
challenge for it to between it and other species but with the same species and strain to survive in

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
144
polymicrobial environment or infection. Hence, this research unravelled how K. pneumoniae may deploy
T6SSs in as a virulence and survival organelle.
After the successful creation of the T6SS gene cluster mutants, bacterial curves and RNA expression for
important T6SS-related gene were performed. The absorbance (600 nm) and qRT-PCR were analysed to
be sure that the T6SS gene clusters were functional and even after knocking them out and did not impact
the bacterial growth in any significant way. Also, intra-strain competitions (T6SS mutant against vrs DN2)
in both shaking and standing medium and inter-species competitions (K. pneumoniae vs E. coli or P.
aeruginosa) was measured to determine the role of T6SS in the fitness of the K. pneumoniae. For all fitness
assay, 1:1 ratio of both competing strains were mixed and allowed to compete for a set time, and the bacteria
were then recovered. To measure the effect of T6SS deletion on the T4SS conjugal transfer, a plasmid
(pACYC184-oriTICE1) was introduced into the K. pneumoniae donors strain and conjugated with the E. coli
HB101 recipient strain on agar surface for overnight. The transconjugants were distinguished on antibiotic
selection media which were later PCR-confirmed. Also, T4SS vriB4 and mobB genes expression were
determined by measuring the RNA expression using qRT-PCR. One in hundred overnight culture of the K.
pneumoniae strains were inoculated in 96-well polystyrene, and 6-well tissue plates (with immersed
coverslip) were used to determine and visualise biofilm formation at 37 oC. RNA expression of fim and mrk
related genes (fimH and mrkD) and later confirmed with Transmission Electron microscopy (TEM) was
used visualise the presence of different surface fimbriae on various the T6SS mutants. To demonstrate the
role of T6SS in aiding K. pneumoniae to thrive in the environment and resist amoeba uptake, a lawn of the
bacterial was spotted with amoeba, and the zone of clearance was observed. Acanthamoeba monolayer
inoculated with bacteria (T6SS mutants and DN2) and on one hand, the percentage association without
antibiotic treatment was determined. On the other hand, the internalised bacteria were protected with
antibiotics and subsequently lysed to enumerate the amoeba internalised K. pneumoniae. All these data
obtained suggest that K. pneumoniae uses T6SSS to survive in a complex polymicrobial environment by
defending and attacking other bacteria, forming biofilm and enhancing its amoeba phagocytic resistance
while spreading resistance and virulence factors.
4.2 Deletion of T6SS gene clusters (T6SS1 and/ or T6SS3) did not affect the growth of the
mutants relative to the parent strain
To determine if the T6SS gene cluster knock introduced any polar effect on the bacterial growth of the
mutants relative to their parent strain, the optical density (at 600 nm) as well as the CFU of the bacteria was
measured at 2 h time-interval for 16 hours. The growth of all the mutant was in synchrony with the parent
strain DN2 suggesting no polar effect of T6SS was invoked, and T6SS may not be any fitness cost on the
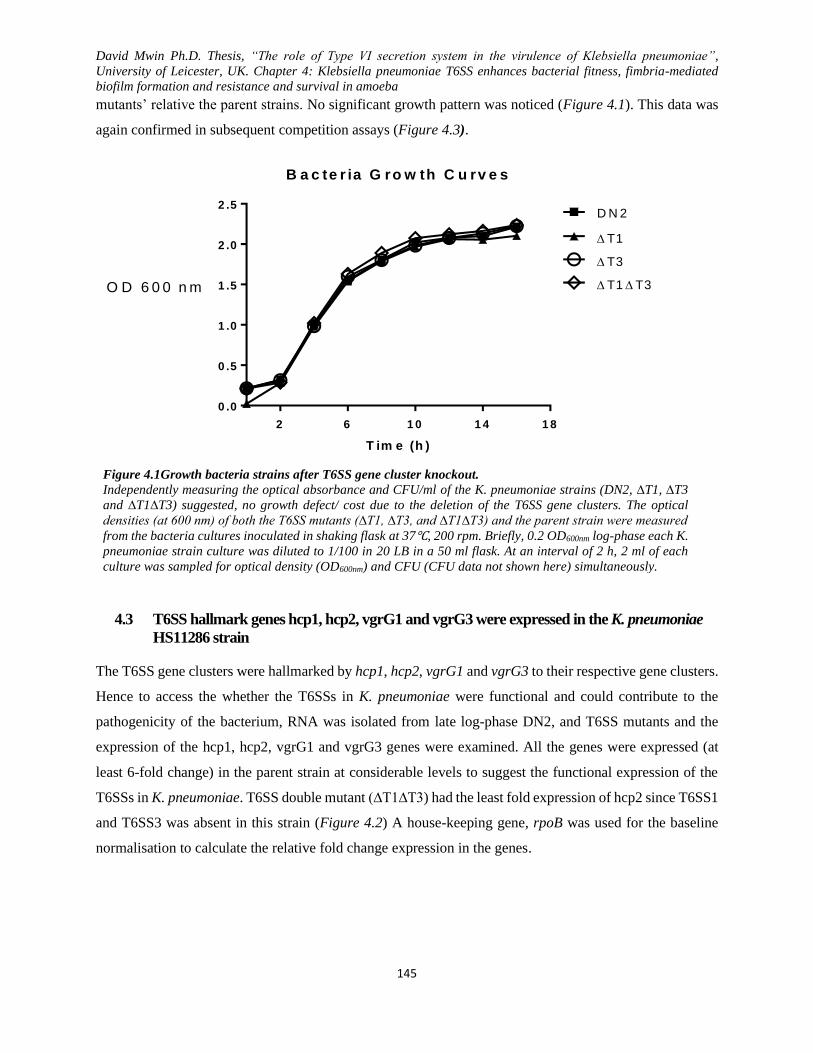
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
145
mutants’ relative the parent strains. No significant growth pattern was noticed (Figure 4.1). This data was
again confirmed in subsequent competition assays (Figure 4.3).
Figure 4.1Growth bacteria strains after T6SS gene cluster knockout.
Independently measuring the optical absorbance and CFU/ml of the K. pneumoniae strains (DN2, ∆T1, ∆T3
and ∆T1∆T3) suggested, no growth defect/ cost due to the deletion of the T6SS gene clusters. The optical
densities (at 600 nm) of both the T6SS mutants (∆T1, ∆T3, and ∆T1∆T3) and the parent strain were measured
from the bacteria cultures inoculated in shaking flask at 37℃, 200 rpm. Briefly, 0.2 OD600nm log-phase each K.
pneumoniae strain culture was diluted to 1/100 in 20 LB in a 50 ml flask. At an interval of 2 h, 2 ml of each
culture was sampled for optical density (OD600nm) and CFU (CFU data not shown here) simultaneously.
4.3 T6SS hallmark genes hcp1, hcp2, vgrG1 and vgrG3 were expressed in the K. pneumoniae
HS11286 strain
The T6SS gene clusters were hallmarked by hcp1, hcp2, vgrG1 and vgrG3 to their respective gene clusters.
Hence to access the whether the T6SSs in K. pneumoniae were functional and could contribute to the
pathogenicity of the bacterium, RNA was isolated from late log-phase DN2, and T6SS mutants and the
expression of the hcp1, hcp2, vgrG1 and vgrG3 genes were examined. All the genes were expressed (at
least 6-fold change) in the parent strain at considerable levels to suggest the functional expression of the
T6SSs in K. pneumoniae. T6SS double mutant (∆T1∆T3) had the least fold expression of hcp2 since T6SS1
and T6SS3 was absent in this strain (Figure 4.2) A house-keeping gene, rpoB was used for the baseline
normalisation to calculate the relative fold change expression in the genes.
2 6 1 0 1 4 1 8
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
B a c te r ia G ro w th C u rv e s
T im e (h )
D N 2
T1
T3
T1 T3O D 6 0 0 n m
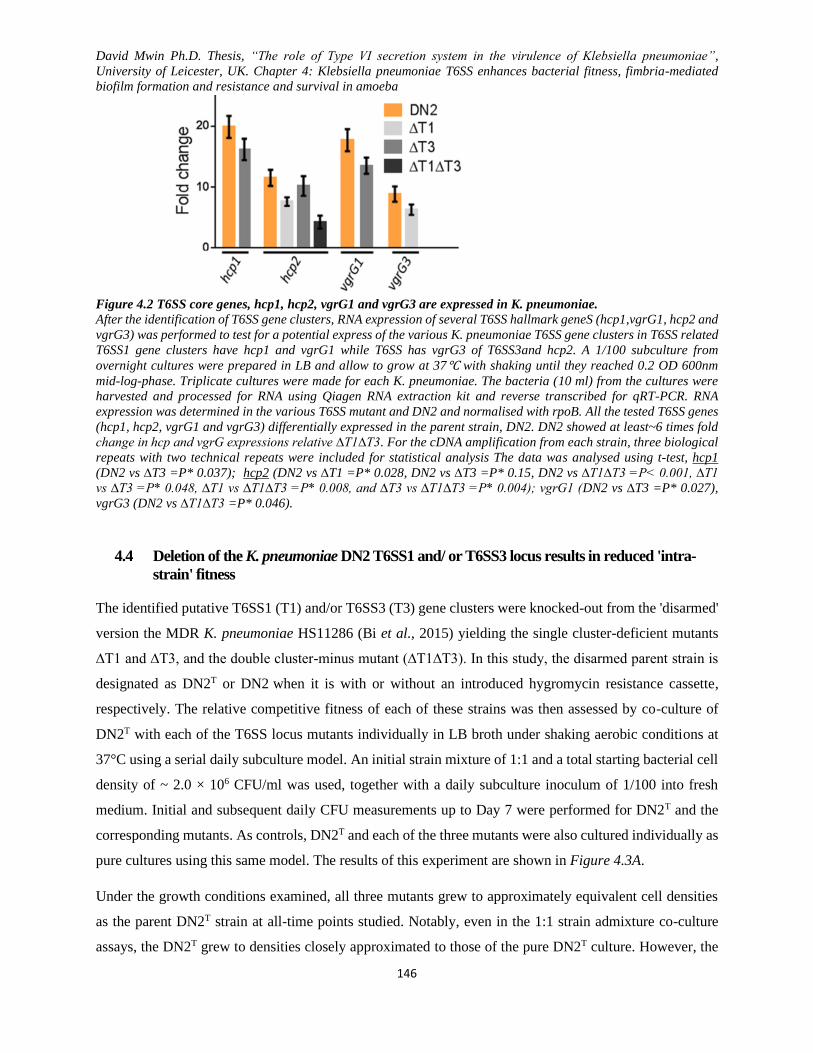
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
146
Figure 4.2 T6SS core genes, hcp1, hcp2, vgrG1 and vgrG3 are expressed in K. pneumoniae.
After the identification of T6SS gene clusters, RNA expression of several T6SS hallmark geneS (hcp1,vgrG1, hcp2 and
vgrG3) was performed to test for a potential express of the various K. pneumoniae T6SS gene clusters in T6SS related
T6SS1 gene clusters have hcp1 and vgrG1 while T6SS has vgrG3 of T6SS3and hcp2. A 1/100 subculture from
overnight cultures were prepared in LB and allow to grow at 37℃ with shaking until they reached 0.2 OD 600nm
mid-log-phase. Triplicate cultures were made for each K. pneumoniae. The bacteria (10 ml) from the cultures were
harvested and processed for RNA using Qiagen RNA extraction kit and reverse transcribed for qRT-PCR. RNA
expression was determined in the various T6SS mutant and DN2 and normalised with rpoB. All the tested T6SS genes
(hcp1, hcp2, vgrG1 and vgrG3) differentially expressed in the parent strain, DN2. DN2 showed at least~6 times fold
change in hcp and vgrG expressions relative ∆T1∆T3. For the cDNA amplification from each strain, three biological
repeats with two technical repeats were included for statistical analysis The data was analysed using t-test, hcp1
(DN2 vs ∆T3 =P* 0.037); hcp2 (DN2 vs ∆T1 =P* 0.028, DN2 vs ∆T3 =P* 0.15, DN2 vs ∆T1∆T3 =P< 0.001, ∆T1
vs ∆T3 =P* 0.048, ∆T1 vs ∆T1∆T3 =P* 0.008, and ∆T3 vs ∆T1∆T3 =P* 0.004); vgrG1 (DN2 vs ∆T3 =P* 0.027),
vgrG3 (DN2 vs ∆T1∆T3 =P* 0.046).
4.4 Deletion of the K. pneumoniae DN2 T6SS1 and/ or T6SS3 locus results in reduced 'intra-
strain' fitness
The identified putative T6SS1 (T1) and/or T6SS3 (T3) gene clusters were knocked-out from the 'disarmed'
version the MDR K. pneumoniae HS11286 (Bi et al., 2015) yielding the single cluster-deficient mutants
∆T1 and ∆T3, and the double cluster-minus mutant (∆T1∆T3). In this study, the disarmed parent strain is
designated as DN2T or DN2 when it is with or without an introduced hygromycin resistance cassette,
respectively. The relative competitive fitness of each of these strains was then assessed by co-culture of
DN2T with each of the T6SS locus mutants individually in LB broth under shaking aerobic conditions at
37°C using a serial daily subculture model. An initial strain mixture of 1:1 and a total starting bacterial cell
density of ~ 2.0 × 106 CFU/ml was used, together with a daily subculture inoculum of 1/100 into fresh
medium. Initial and subsequent daily CFU measurements up to Day 7 were performed for DN2T and the
corresponding mutants. As controls, DN2T and each of the three mutants were also cultured individually as
pure cultures using this same model. The results of this experiment are shown in Figure 4.3A.
Under the growth conditions examined, all three mutants grew to approximately equivalent cell densities
as the parent DN2T strain at all-time points studied. Notably, even in the 1:1 strain admixture co-culture
assays, the DN2T grew to densities closely approximated to those of the pure DN2T culture. However, the

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
147
concentration as determined by CFU counts of each of the three mutants in the co-culture assays were
significantly reduced compared to that of DN2T, with the discordance becoming progressively more
pronounced from day 2 to Day 7. Furthermore, the data revealed a pattern of descending competitive fitness
relative to DN2T, with the ∆T1∆T3 double-mutant being the least fit, ∆T1 being the next most impaired,
and even ∆T3, the least handicapped, still exhibited significant reduced 'intra-strain' fitness in competition
with DN2T. Indeed, the DN2T: mutant ratios measured at day 7 for co-culture mixtures with ∆T1∆T3, ∆T1
and ∆T3 were 1:0.04, 1:0.09 and 1:0.4, respectively.
Just as described above, a similar inoculum mixture of DNT and each mutant was spotted on a filter-
membrane placed on a warm LB agar surface, and each bacterial strain was then subsequently recovered
from the filter-membrane at day 2 and 4. Data from mixed co-cultures (DNT and ∆T1, DNT and ∆T3, and
DNT and ∆T1∆T3) retrieved from the filter-membrane are presented here. As shown in Figure 4.3B,
∆T1∆T3 and ∆T1 (P< 0.05 each) began losing the competition unlike ∆T3 (P> 0.05) at day 2 when
compared to DNT. Additionally, even though all the mutants were less fit at day four relative to DNT,
∆T1∆T3 and ∆T1 (P<0.01 each) again proved to be the least fit at this time-point.
Interestingly, not much difference in the recovery of the mutants was noted at day 2 of the shaking LB broth
aerobic co-culture. However, recoveries made from the filter-membrane surface cultures at day 2 were very
apparent with a lot of dramatic differences on day 4. More recovery ratios of DNT to ∆T1 or ∆T3 or ∆T1∆T3
were made from shaking aerobic LB broth (1:0.90, 1:10 and 1:0.71 respectively) than those made when
mixtures were grown on a filter membrane (1:0.22, 1:0.34, and 1:0.10) at day 2.
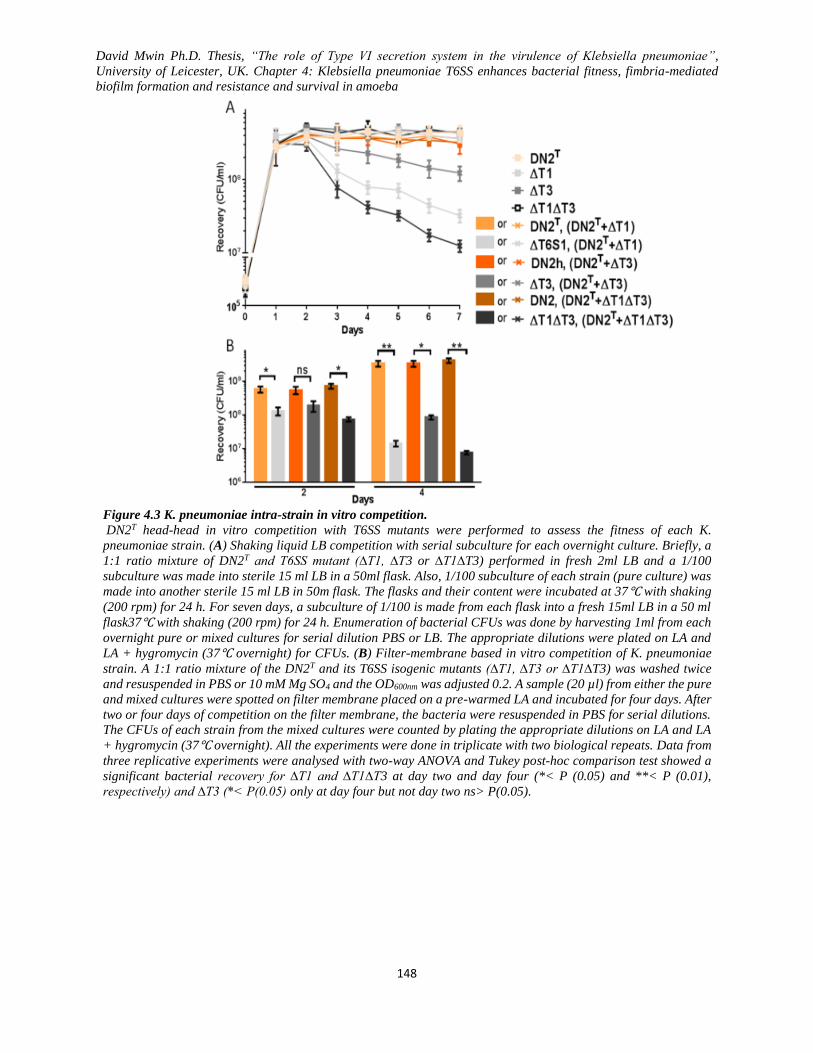
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
148
Figure 4.3 K. pneumoniae intra-strain in vitro competition.
DN2T head-head in vitro competition with T6SS mutants were performed to assess the fitness of each K.
pneumoniae strain. (A) Shaking liquid LB competition with serial subculture for each overnight culture. Briefly, a
1:1 ratio mixture of DN2T and T6SS mutant (∆T1, ∆T3 or ∆T1∆T3) performed in fresh 2ml LB and a 1/100
subculture was made into sterile 15 ml LB in a 50ml flask. Also, 1/100 subculture of each strain (pure culture) was
made into another sterile 15 ml LB in 50m flask. The flasks and their content were incubated at 37℃ with shaking
(200 rpm) for 24 h. For seven days, a subculture of 1/100 is made from each flask into a fresh 15ml LB in a 50 ml
flask37℃ with shaking (200 rpm) for 24 h. Enumeration of bacterial CFUs was done by harvesting 1ml from each
overnight pure or mixed cultures for serial dilution PBS or LB. The appropriate dilutions were plated on LA and
LA + hygromycin (37℃ overnight) for CFUs. (B) Filter-membrane based in vitro competition of K. pneumoniae
strain. A 1:1 ratio mixture of the DN2T and its T6SS isogenic mutants (∆T1, ∆T3 or ∆T1∆T3) was washed twice
and resuspended in PBS or 10 mM Mg SO4 and the OD600nm was adjusted 0.2. A sample (20 µl) from either the pure
and mixed cultures were spotted on filter membrane placed on a pre-warmed LA and incubated for four days. After
two or four days of competition on the filter membrane, the bacteria were resuspended in PBS for serial dilutions.
The CFUs of each strain from the mixed cultures were counted by plating the appropriate dilutions on LA and LA
+ hygromycin (37℃ overnight). All the experiments were done in triplicate with two biological repeats. Data from
three replicative experiments were analysed with two-way ANOVA and Tukey post-hoc comparison test showed a
significant bacterial recovery for ∆T1 and ∆T1∆T3 at day two and day four (*< P (0.05) and **< P (0.01),
respectively) and ∆T3 (*< P(0.05) only at day four but not day two ns> P(0.05).

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
149
4.5 K. pneumoniae DN2 inter-species fitness is dependent on which T6SS loci are being deleted
Bacteria fitness is employed by various bacterial species to outwit their competitors in fight for the limiting
resources in a mixed bacterial environment (Hibbing et al., 2010). The previous report suggests T6SS may
influence the competitive fitness of various gram-negative bacteria by inhibiting their prey (Alcoforado
Diniz and Coulthurst, 2015; Borgeaud et al., 2015b) in their quest to survive and colonize their biological
niche.
Figure 4.4 Inter-species in vitro competition.
The fitness of K. pneumonia strains (DN2T, ∆T1, ∆T3 and ∆T1∆T3) was assessed in a head-to-head- in vitro
competition against other bacteria species (E. coli and P. aeruginosa) on a filter membrane placed on antibiotic-
free LA for 24 h. The mixtures include; K. pneumoniae vs. K. pneumoniae, K. pneumoniae vs. E. coli, and K.
pneumoniae vs. P. aeruginosa. A and B indicate overnight in vitro bacteria recovery when of K. pneumoniae was
competed for heads-head against a T6SS- E. coli HB101 in shaking liquid LB and on filter-membrane respectively.
C; in vitro competition of K. pneumoniae and T6SS+ P. aeruginosa PA14 on Filter-membrane overnight. MC
stands for strains from the same mixed culture. Briefly, a 1:1 ratio mixture of K. pneumoniae and E. cole or P.
aeruginosa was from fresh mid-log-phage cultures was washed twice and resuspended in PBS or 10 mM Mg SO4
and the OD600nm was adjusted 0.2. A sample (20 µl) from either the pure (K. pneumoniae only, E. col only or P.
aeruginosa only) and mixed cultures (K. pneumoniae + E. coli or K. pneumonia + P. aeruginosa) were spotted
on filter membrane placed on a pre-warmed LA and incubated 37℃ overnight. Also, another sample (1/100) of
the pure or mixed cultures were subcultured into 15 ml LB at 37℃ with shaking (200 rpm) for 24 h. After 24 h of
competition on the filter membrane or LB, the bacteria were resuspended in PBS for serial dilutions or sampled
from the LB competition cultures for CFUs enumeration. The CFUs of each bacterial strain from the mixed
cultures were counted by plating the appropriate dilutions on LA and LA + ampicillin (37℃ overnight). Note that
data from K. pneumoniae + P. aeruginosa liquid LB culture competition is not presented. All the above data are
from three experimental replicates, and one-way ANOVA and Tukey post-hoc test were used to analyse the data
statistically, ns (P> 0.05), *< P (0.05), **> P(0.01) and ***< P (0.001). K. pneumoniae recovery from different
mixtures is shown as solid colour bars while the shaded pattern colour bars represent E. coli or P. aeruginosa
PA14 from the mix cultures are shaded/pattern bars..

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
150
In this study, the competitive fitness of E. coli and K. pneumoniae, and P. aeruginosa and K. pneumoniae
from mixed cultures was measured. To measure the ability of K. pneumoniae to inhibit E. coli, a 1:1 of E.
coli and K. pneumoniae were co-cultured in either shaking liquid LB broth or on filter membrane at 37⁰C
and the individual bacteria were subsequently recovered after 24 hours of competition (). While no
significant observable difference in bacteria recoveries was noted when E. coli HB101 and K. pneumoniae
DN2 or T6SS mutants were co-cultured in shaking LB broth, a dramatic reduction in the recovered E. coli
was evident when the bacteria mixture was co-cultured on a filter-membrane placed on a pre-ward LB agar
plate. From the Filter membrane co-culture, the E. coli HB101 growths in the admixtures were as
significantly inhibited by K. pneumoniae DN2T and ∆T3 but not ∆T1 and ∆T1∆T3. As predicted, K.
pneumoniae DN2T caused a 10.56 or 1.31 or 15.39-fold more reduction in the E. coli HB101 recovered
relative to that of ∆T1 or ∆T3 or ∆T1∆T3, respectively. Additionally, a 8.034 and 11.71 fold increase in
the E. coli HB101 recoveries were made when it was co-cultured with ∆T1 and ∆T1∆T3 respectively,
relative to the E. coli HB101 recovery from the admixture with ∆T3. Though no statistically significant
difference in E. coli HB101 recoveries from the admixture with ∆T1 or ∆T1∆T3 was obtained, a 1.46-fold
increase in E. coli HB101 was observed from ∆T1∆T3 admixture.
The killing behaviour that was observed during the plasmid mobilization prompted for a more visual way
of assessing T6SS killing impact of K. pneumoniae of E. coli. An E. coli DH5α carrying PDNTOOL-39
encoding apramycin resistance and blue colour phenotype on X-gal/IPTG media was mixed at a 1:1 ratio
with the various K. pneumoniae strains and allowing to compete overnight. The mixture was then spotted
on apramycin media with X-gal/IPTG, killing all the K. pneumoniae and allowing the blue forming E. coli
colonies to grow. As can be seen in Figure 4.5, DN2 and ∆T3 yielded fewer colonies of E. coli DH5α
colonies compared to the ones recovered from ∆T1 and ∆T1∆T3 mixture with E. coli DH5α. This data
presented in Figure 4.4A, Figure 4.4B, and Figure 4.5 suggest that K. pneumoniae HSS11286 T6SS was
actively involved in the killing of E. coli numbers as means to out-competing other bacteria in the mix-
culture environment. Similarly, the only successful K. pneumoniae NTUH-K2044 T6SS mutant
(NTUH∆T1) or its wildtype was mixed with GFP tagged E. col DH5α and grown for 20 hr, the recovered
E. coli DH5α by both colony counting and fluorescence imaging (Figure 4.6) revealed that the T6SS1
mutant could barely inhibit the growth of the E. coli DH5α unlike its parent strain (KR1730).
P. aeruginosa has previously been reported to secrete an abundance of Hcp even under laboratory
conditions (Jiang et al., 2014; Hood et al., 2010a; Bleves et al., 2010; Pukatzki et al., 2006b; Bernard et
al., 2011; Mougous et al., 2006b). Hence, understanding how K. pneumoniae T6SS may influence its
survival in a polymicrobial community with other T6SS positive bacteria such as P. aeruginosa may
improve the overall understanding of the interaction among T6SS positive bacteria of different species.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
151
Having noted that, the K. pneumoniae T6SS direct contact and killing effect on E. coli HB101 was further
enhanced when the bacteria admixtures were co-cultured on a filter-membrane surface, another K.
pneumoniae competition assay as similarly described above was performed. A 1:1 ratio mixture of K.
pneumoniae DN2 or T6SS mutants and P. aeruginosa were co-cultured on filter-membrane for 24 hours
and the bacteria CFU subsequently recovered. As presented in Figure 4.4C, the P. aeruginosa being T6SS
positive bacteria species had a significant huge inhibitory growth effect on the K. pneumoniae T6SS
mutants but not DN2T. Unlike the absence of noticeable P. aeruginosa inhibitory effect on K. pneumoniae
DNT, the T6SS mutants’ growth (∆T1, ∆T3 and ∆T1∆T3) were significantly reduced by 4.40, 2.90 and
8.65 folds respectively, relative the DN2T recovered. Among the T6SS mutants, ∆T3 was the least inhibited
by P. aeruginosa while ∆T1∆T3 was the most handicapped in its recovery. Interestingly, the P. aeruginosa
recoveries from the admixture with K. pneumoniae did not vary significantly.
Figure 4.5 E. coli DH5α colony reduction due to K. pneumoniae T6SS activity.
K. pneumoniae preyed upon E. coli in a T6SS dependent manner as revealed by spotting growing a mixture of K.
pneumoniae (predator) and E. coli DH5α (prey) together. The E. coli DH5α used in this assay carry pDNTOOL-39
which express blue colonies on LA (X-gal + IPTG) supplemented with apramycin. All the K. pneumoniae strain are
sensitive to apramycin. Briefly, 20µl of a 1:1 mixture of log-phase K. pneumoniae and E. coli DH5α was spotted on a
nitrocellous membrane on an antibiotic-free agar at 37℃ for 24 h. The bacteria spot mixtures on the membranes were
recovered and resuspended in PBS, and 10µl from the appropriate dilution of each mixture was spotted in duplicates
on LA+apramycin and X-gal/IPTG and incubated at 37℃ for 16 h. Only E. coli colonies carrying the resistance
plasmid were recovered from the mixtures since all the K. pneumoniae strains are susceptible to apramycin.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
152
Figure 4.6 K. pneumoniae NTUH-K2044 T6SS1-dependent inhibition of E. coli DH5α.
Bacteria recovery and fluorescent imaging were performed to examine the survival of the E. coli DH5α after co-
culture with either K. pneumoniae NTUH-K2044 wildtype (KR1730) or its associated T6SS1 mutant (NTUH∆T1)
for 20 hours. E. coli DH5α carried pIM438 that encoded GFP, Kanamycin and hygromycin resistance for the
colony selection, unlike the K. pneumoniae strains which were sensitive to both kanamycin and hygromycin. A
1:1 mixture of log-phase K. pneumoniae and E. coli DH5α were grown on nitrocellous membrane placed on an
antibiotic-free agar at 37℃ overnight. The spot mixtures on the membranes were recovered, and the appropriate
dilutions were plated on LA and LA+kanamycin+hygromycin for enumeration. Also, 20µl of the undiluted
mixtures were on a thin layer of media (LA+kanamycin+hygromycin) in a plate or coated on a sterile slide. The
plates and/ or slides were incubated 37℃ for 20 h. The percentage recovery was estimated as several E. coli
DH5α colonies recovered from the mixed culture divided by the number of E. coli DH5α colonies from E. coli
DH5α pure culture multiplied by 100%. The data was analysed using student t-test, and P-value for the difference
in percentage recovery of E. coli was 0.002.
4.6 Absence of T6SS1 and T6SS3 enhanced plasmid DNA mobilization via conjugation
In the natural environment, bacteria spread resistance and virulence through horizontal gene transfer to
improve their subsequent survival in host or environment. Previous studies suggest an interplay between
T6SS and horizontal gene transfer (Blokesch, 2015a; Borgeaud et al., 2015a; Das et al., 2002; Ho, Basler
and Mekalanos, 2013).
In this section, pACYC184-oriTICE plasmid mobilisation frequency from either K. pneumoniae DN2 or TSS
mutants (∆T1, ∆T3 and ∆T1∆T3) to E. coli HB101 (recipient) was assessed. The bacterial mixtures were
mated overnight on a membrane filter. The transconjugants E. coli carrying three plasmids were enumerated
in an antibiotic medium that killed all the K. pneumoniae donor cells. The transconjugants were then PCR-
confirmed for the presence of the plasmid. The deletion of either T6SS1 or T6SS3 resulted in about 6 or 2–
fold increase in the frequency of plasmid mobilization respectively, from K. pneumoniae to E. coli HB101
via conjugation. Surprisingly, ∆T1∆T3 relative to DN2 showed no difference in the plasmid mobilization
frequency. This was quite unexpected as the deletion of both T6SS1, and T6SS3 (∆T1∆T3) resulted in more
E. coli HB101 recoveries. Among the T6SS mutants, ∆T1 was the most efficient in the conjugal

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
153
mobilization of the plasmid, and next was the ∆T3 while ∆T1∆T3 was the least efficient. To have observed
an increased transconjugant frequency in ∆T1 and ∆T3 but not ∆T61∆T3 was quite surprising (Figure
4.7A). One would have thought that the deletion of both clusters will then result in a higher E. coli HB101
recovery (this was later answered with TEM and virB4 and mobB expression data).
But interestingly, ∆T1∆T3 expectedly yielded the most E. coli recovery (Figure 4.4 and Figure 4.5), but
with the lowest transconjugants (Figure 4.7A), predicting that the different levels of expression of the T4SS
conjugative machinery could affect the frequencies of the plasmid mobilization. The expression of key
genes (virB4 and mobB) known to be involved in the expression of the T4SS conjugative module was
analysed by qRT-PCR. K. pneumoniae house-keeping gene (rpoB) was used for all normalization purposes.
The expression data revealed that ∆T1∆T3 had its mobB and virbB4 at least two folds down-regulated
relative to its DN2 while that of ∆T3 had at least two-fold increase (Figure 4.7B). There was no significant
difference in the virB4 and mob expression for ∆T1 and that of the parent strain DN2. The expression data
support the conjugation frequency data all together reinforcing the hypothesis that, T6SS-dependent killing
of E. coli and the co-regulation with T4SS conjugative system synergistically accounts for K. pneumoniae-
E. coli predatory interaction and horizontal gene transfer of resistance between the two bacteria (target-kill-
and-prey-for-for colonization or cooperate in spreading resistance).
4.7 T6SS deletion enhanced biofilm formation in K. pneumoniae.
T6SS has previously been linked to biofilm formation of Vibrio, P. aeruginosa, E. coli (Aschtgen et al.,
2008; De Pace et al., 2010; Enos-Berlage et al., 2005). Biofilm formation in K. pneumoniae is a critical
mechanism in nosocomial infection by evading the host immune system and antibiotic treatment (Fierer,
2012; Boddicker et al., 2006; Huang et al., 2014; Olson et al., 2012). Previous studies suggest that some
virulent factors are possibly tightly co-regulated or induced by specific cues their co-expression (Hood et
al., 2010c; Bernard et al., 2011; Russell et al., 2011a). Thus, understanding how the presence or absence of
T6SSs could impact biofilm formation is vital in studying the virulence mechanism of K. pneumoniae.
T6SS, with its diverse role, has now been implicated in fimbriae and biofilm formation (Aschtgen et al.,
2008; De Pace et al., 2010; Enos-Berlage et al., 2005). Biofilm formation at 37ᴼC was quantified by
measuring the absorbance (595 nm) after staining with crystal violet and biofilm formed on coverslips were
imaged after overnight incubation where coverslips were submerged in the bacterial cultures at 37ᴼC. Note
that two KR161 and KR162 are positive and negative biofilm former previously validated (van Aartsen,
2012). Biofilm was also formed at 30 ᴼC, and it showed similar results (data not included). Thus,
understanding how the presence or absence of T6SSs could impact biofilm formation is critical in studying

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
154
the virulence mechanism of K. pneumoniae. To test the impact of T6SS deletion on the K. pneumoniae
biofilm phenotype, the strains were grown at 1/100 subculture from overnight cultures for biofilm formation
in 9-well plate and 6-well tissue culture plates with an immersed coverslip. And the data collected at the at
various time-points.
All the T6SS mutants have either formed more biofilm or almost equivalent to that of the parent strain
(DN2) at all the time-points. Generally, all the strains (DN2, ∆T1, ∆T3, and ∆T1∆T3) formed biofilm at
24, 48, and 72 h with relatively no significant difference between time points. From the biofilm data (Figure
4.8A), deletion of either T6SS1 or T6SS3 in K. pneumoniae further enhanced their ability to form biofilm
but not when both T6SS1 and T6SS3 clusters are deleted as it results in no significant biofilm formation
relative to the parent strains, DN2. Interestingly, while ∆T3 had the most biofilm formation relative to DN2
and the other mutants, ∆T1∆T3 was the least biofilm former though there was no significant difference
(P>0.05) for day 48 and 72 h except 24h. The loss of both T6SS clusters (T6SS1 and T6SS3). The visualized
images of all the biofilm formed (Figure 4.8B) support the quantitative biofilm data in Figure 4.8A. The
imaged biofilm of ∆T3 shows bacterial aggregates or clusters, unlike ∆T1 and DN2. Biofilm formation at
30ᴼC for all the strains was significantly enhanced (data are not shown) than those obtained from 37ᴼC
suggesting the how temperature difference could influence T6SS-dependent biofilm formation. The
quantitative Biofilm data are from three experimental replicates while the imaged biofilm data are
representative of data form only overnight biofilm formation.
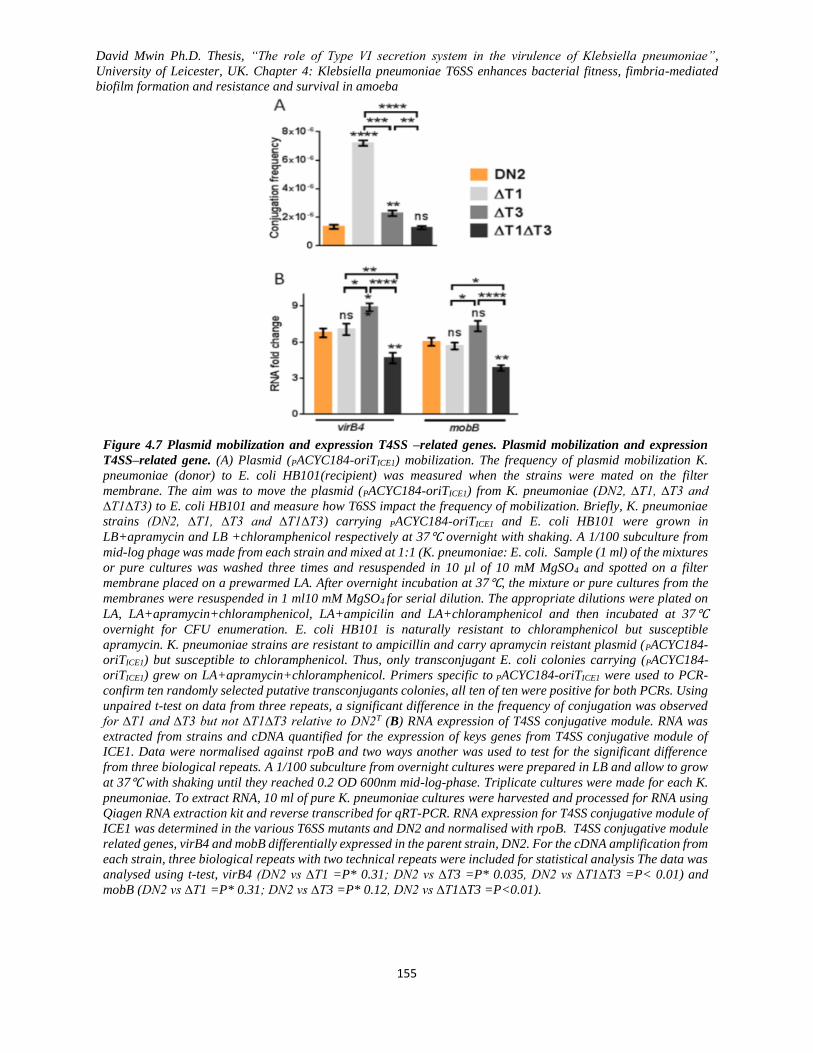
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
155
Figure 4.7 Plasmid mobilization and expression T4SS –related genes. Plasmid mobilization and expression
T4SS–related gene. (A) Plasmid (PACYC184-oriTICE1) mobilization. The frequency of plasmid mobilization K.
pneumoniae (donor) to E. coli HB101(recipient) was measured when the strains were mated on the filter
membrane. The aim was to move the plasmid (PACYC184-oriTICE1) from K. pneumoniae (DN2, ∆T1, ∆T3 and
∆T1∆T3) to E. coli HB101 and measure how T6SS impact the frequency of mobilization. Briefly, K. pneumoniae
strains (DN2, ∆T1, ∆T3 and ∆T1∆T3) carrying PACYC184-oriTICE1 and E. coli HB101 were grown in
LB+apramycin and LB +chloramphenicol respectively at 37℃ overnight with shaking. A 1/100 subculture from
mid-log phage was made from each strain and mixed at 1:1 (K. pneumoniae: E. coli. Sample (1 ml) of the mixtures
or pure cultures was washed three times and resuspended in 10 µl of 10 mM MgSO4 and spotted on a filter
membrane placed on a prewarmed LA. After overnight incubation at 37℃, the mixture or pure cultures from the
membranes were resuspended in 1 ml10 mM MgSO4 for serial dilution. The appropriate dilutions were plated on
LA, LA+apramycin+chloramphenicol, LA+ampicilin and LA+chloramphenicol and then incubated at 37℃
overnight for CFU enumeration. E. coli HB101 is naturally resistant to chloramphenicol but susceptible
apramycin. K. pneumoniae strains are resistant to ampicillin and carry apramycin reistant plasmid (PACYC184-
oriTICE1) but susceptible to chloramphenicol. Thus, only transconjugant E. coli colonies carrying (PACYC184-
oriTICE1) grew on LA+apramycin+chloramphenicol. Primers specific to PACYC184-oriTICE1 were used to PCR-
confirm ten randomly selected putative transconjugants colonies, all ten of ten were positive for both PCRs. Using
unpaired t-test on data from three repeats, a significant difference in the frequency of conjugation was observed
for ∆T1 and ∆T3 but not ∆T1∆T3 relative to DN2T (B) RNA expression of T4SS conjugative module. RNA was
extracted from strains and cDNA quantified for the expression of keys genes from T4SS conjugative module of
ICE1. Data were normalised against rpoB and two ways another was used to test for the significant difference
from three biological repeats. A 1/100 subculture from overnight cultures were prepared in LB and allow to grow
at 37℃ with shaking until they reached 0.2 OD 600nm mid-log-phase. Triplicate cultures were made for each K.
pneumoniae. To extract RNA, 10 ml of pure K. pneumoniae cultures were harvested and processed for RNA using
Qiagen RNA extraction kit and reverse transcribed for qRT-PCR. RNA expression for T4SS conjugative module of
ICE1 was determined in the various T6SS mutants and DN2 and normalised with rpoB. T4SS conjugative module
related genes, virB4 and mobB differentially expressed in the parent strain, DN2. For the cDNA amplification from
each strain, three biological repeats with two technical repeats were included for statistical analysis The data was
analysed using t-test, virB4 (DN2 vs ∆T1 =P* 0.31; DN2 vs ∆T3 =P* 0.035, DN2 vs ∆T1∆T3 =P< 0.01) and
mobB (DN2 vs ∆T1 =P* 0.31; DN2 vs ∆T3 =P* 0.12, DN2 vs ∆T1∆T3 =P<0.01).

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
156
Figure 4.8 Biofilm formation and visualisation.
(A) Biofilm formation 96-well polystyrene plate and (B) biofilm formation and visualization on submerged cover-
slips. Briefly, K. pneumoniae strains (DN2, ∆T1, ∆T3 and ∆T1∆T3) were grown in LB overnight and subcultured
(1/100) in 96-well polystyrene plate at 37℃ for 24, 48 and 72 h. For visualization of formed biofilms on coverslips,
a 1/100 subculture was made into 6- well tissue culture plates containing submerged coverslips in sterile LB and
grown for 24 h at 37℃. After the formation of biofilm, the cultures were gently discared, washed three times with
distilled water and stained with crystal violet. Residual crystal violet was washed away, and the plate air-dried
and solubilized in 95% ethanol for the measurement of OD595nm. The coverslips were mounted on slides for light
microscopy. Data is a representation of three experimental repeats, and the statistical difference was tested using
two-way ANOVA with Tukey post-hoc test. P-values; *<0.05, **<0.01, ***<0.001 and ****<0.0001 (B). The
scale indicated on all the images is 10 µm.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
157
4.8 Deletion of T6SS1 and T6SS3 clusters enhanced type 1 and type 3 fimbriae expression
respectively
Transmembrane bacterial surface appendages such as pili and fimbriae are major organelles used in
bacteria-bacteria interaction or bacteria-abiotic surface interactions (Alcántar-Curiel et al., 2013; Huang et
al., 2014), indicating their crucial role in colonization, dissemination of antibiotic resistance and pathogen-
host interaction. Previous studies have suggested that certain pathogenic bacteria may explore an interplay
between T6SS and fimbriae/ biofilm/ conjugation (De Pace et al., 2010; Blokesch, 2015b; Ho, Basler and
Mekalanos, 2013) as a virulence mechanism towards the host. Having observed unique T6SS-dependent
pattern in conjugation and biofilm, thus an investigation of the potential surface structures such as fimbriae
was performed. The deletion of T6SS clusters was predicted to significantly altered the type and pattern of
pili and fimbriae on the bacterial surface. The examination of expressed fimbriae using agglutination assay,
qRT-PCR and Transmission Electron Microscopic (TEM) was conducted.
Agglutination assay was performed to test for the presence of type 1 and type 3 fimbriae which were
expressed by each K. pneumoniae strain and how these fimbriae may play a role in the biofilm formation.
Briefly, overnight K. pneumoniae cultures were either mixed with guinea pig red blood cells or 5% tannic
acid-treated Oxblood (TATOXB) with or without mannose. From Table 4.1, while all the K. pneumoniae
strains agglutinated GP-RBCs with 15 minutes with mannose sensitivity, suggesting the presence of type 1
fimbria, only ∆T3 showed mannose-resistant agglutination of tannic acid-treated ox blood unlike DN2, ∆T1
and ∆T1∆T3. The results from GP-RBCs and TATOXB agglutination assay suggested that DN2, ∆T1 and
∆T1∆T3 probably expressed only type 1 fimbria while ∆T3 expressed both type 1 and type 3 fimbriae. The
deletion of either T6SS cluster the K. pneumoniae strain affected the fimbriation of the bacterium and its
ability to form biofilm formation as presented in Table 4.1 and Figure 4.8. The possible expression of type
two fimbriae by ∆T3 could be linked to the enhanced biofilm formation as observed Figure 4.8.
Table 4.1 Agglutination Assay
GP-RBC TATOXB
DN2 + / S -
∆T1 + / S -
∆T3 + / S + / R
∆T1∆T3 + / S -
+; agglutinate within 15 minutes, -; no agglutinate within 15 minutes, S;
mannose sensitive and R; mannose resistant
Agglutination assay was by mixing each overnight K. pneumoniae cultures (109 CFU/ml) independently with 5% GP-
RBCs or 5% TATOXB with or with 5% mannose on the slide for visible agglutination within 15 minutes

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
158
To confirm the presence of type 1 and type 3 fimbriae on the various K. pneumoniae, RNA was extracted
from log-phase culture, and revere transcribed into cDNA which was subsequently used for RT-PCR
analysis. The level of expression of FimH an MrkD in the cDNA was used as a measure of the expression
of type 1 and type 3 fimbriae, respectively. From the cDNA analysis, FimH was noted to have been
significantly expressed by ∆T1 by almost 2-fold increase relative to DNA but was attenuated in ∆T1∆T3
by an almost 1-fold decrease. The expression of fimH in ∆T3 showed no significant difference relative to
DN2 but not to ∆T1∆T3. For the expression of type 3 fimbria associated mrkD, ∆T3 significantly expressed
MrkD relative to the other K. pneumoniae strains. Just like the fimH expression, mrkD was least expressed
by ∆T1∆T3 with at least a fold decrease relative to DN2. The RT-qPCR data suggest that K. pneumoniae
express type 1 and type 3 fimbriae which are widely reported to be associated with biofilm formation. ∆T1
and ∆T3 exhibited higher levels of fimH (type 1 fimbria related) and MrkD (type 3 fimbria related) unlike
DN2 and ∆T1∆T3.
Figure 4.9 Expression of mrkD and fimH of K. pneumoniae surface fimbriae.
Quantitative qRT-PCR of RNA of mrkD and fimH extracted from K. pneumoniae and T6SS mutants was
assessed. RNA was extracted from the strains and cDNA quantified for the expression of type 1 and type 3
fimbriae. A 1/100 subculture from overnight cultures were prepared in LB and allow to grow at 37℃ with
shaking until they reached 0.2 OD 600nm mid-log-phase. Triplicate cultures were made for each K.
pneumoniae. To extract RNA, 10 ml of pure K. pneumoniae cultures were harvested and processed for RNA
using Qiagen RNA extraction kit and reverse transcribed for qRT-PCR. RNA expression for type 1 and type3
was determined in the various T6SS mutants and DN2 and normalised with rpoB. Type 1 and type 3 related
genes, mrkD and FimH were differentially expressed in the parent strain the K, pneumoniae strains. For the
cDNA amplification from each strain, three biological repeats with two technical repeats were included for
statistical analysis The data were analysed using one-way ANOVA with Tukey post-hoc test t-test, mrkDvirB4
(DN2 vs ∆T1 =P> 0.05; DN2 vs ∆T3 =P< 0.001, DN2 vs ∆T1∆T3 =P< 0.05) and fimH (DN2 vs ∆T1
=P<0.05; DN2 vs ∆T3 =P>0.05 0.12, DN2 vs ∆T1∆T3 =P<0.05).

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
159
The RNA expression data (Figure 4.9) for ∆T3 indicated about two or more folds increase in the expression
of mrKD relative to DN2 or T6SS mutant, respectively. Two or more-fold increase in the expression of
fimH was observed in ∆T1 when compared to the other strain (DN2. ∆T3, and ∆T1∆T3). While ∆T1∆T3
showed the least fimH and mrkD expression supporting its visualised fimbriae-like structures, no significant
difference in the expression of fimH was observed between DN2, and ∆T3 neither was there any difference
between DN2 and ∆T6S1 for the expression of mrkD. All RT-qPCR data were from three biological repeats
and normalised the house-keeping gene, rpoB.
Having confirmed the presence expression of type 1 and type 3 fimbriae using agglutination and RT-qPCR,
Transmission Electron Microscopic (TEM) imaging was used to visualise the expressed surface
appendages. The data obtained from TEM imaging supports the biofilm, agglutination and RNA expression
data; highly expressed fimbriae with different types were noted on ∆T3 but not the other mutants or DN2.
The mutation of T6SS clusters in the K. pneumoniae HS11286 consequentially induced the expression of
different surface fimbriae-like structures (Figure 4.10) on ∆T3 and ∆T1.
The TEM images revealed three types of fimbriae; long-thick fimbriae (1.4 ± 0.4 µm long and 9.50 ± 3 nm
wide), long-thin fimbriae (1.2 ± 0.2 µm long and 7.2 ± 0.3 nm wide) and short-thin fimbriae (400 ± 100 nm
long and 5.2 ± 0.5 nm wide). All fimbriae were digitally measured based on un-stretched fimbriae length
and width (Figure 4.10). T6SS3 mutant (∆T3) was highly fimbriated with an abundance of surface
appendages, most of which are long thick and bundled fimbriae covering the whole surface of the bacterial
cell (1.2 ± 0.4 µm long 7.2 ± 0.3 nm width) and few thin long and short fimbriae. While ∆T3 revealed
numerous long-thick and short-thin fimbriae, ∆T1 expressed both showing long-thin and short-thin
fimbriae, and ∆T1∆T3 only had fewer long-thin fimbriae just like DN2.
The T6SS1 mutant (∆T1) equally expressed a good amount of long thin fimbriae that appear to span across
the surface of the bacterium. Unlike ∆T1 and ∆T3, the parent strain (DN2) and ∆T1∆T3 barely expressed
any surface fimbriae, and this was not surprising as biofilm, agglutination and RNA fimbriae expression
data corroborated with the TEM data. Unlike ∆T3, few long thin fimbriae-like structures were visualized
on the surface of ∆T1 and DN2 while much fewer long thin fimbriae-like structures were also seen on the
surface of ∆T1∆T3.
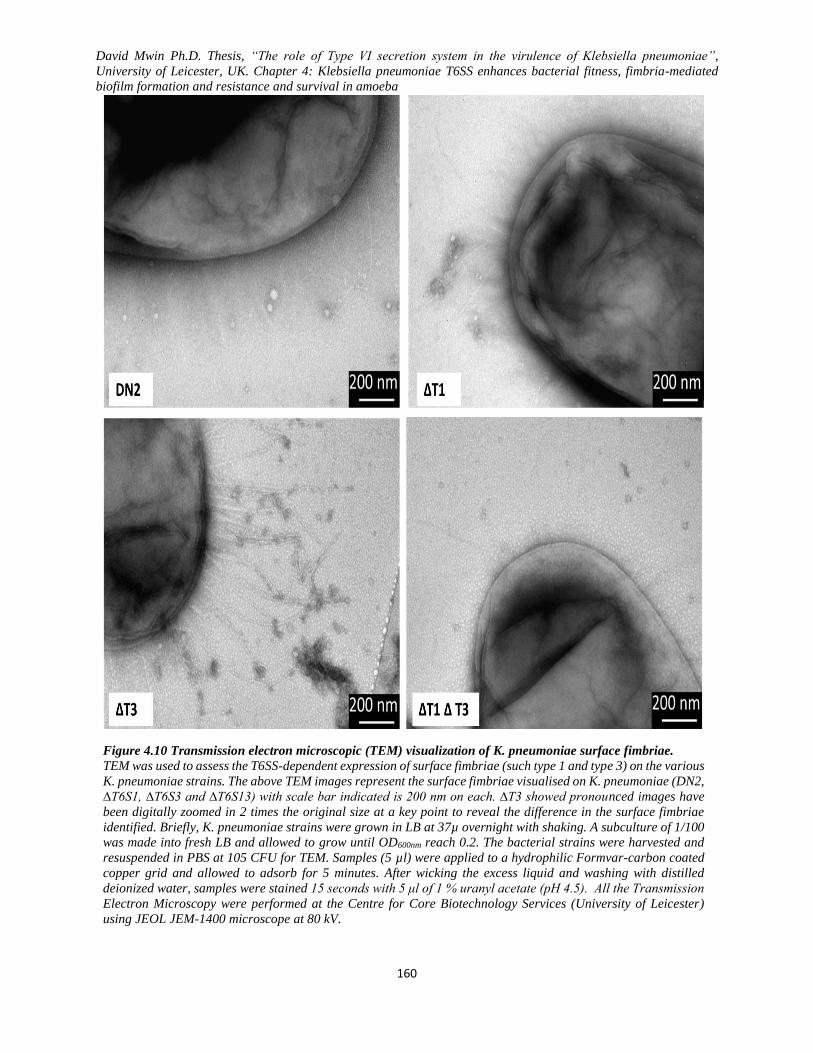
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
160
Figure 4.10 Transmission electron microscopic (TEM) visualization of K. pneumoniae surface fimbriae.
TEM was used to assess the T6SS-dependent expression of surface fimbriae (such type 1 and type 3) on the various
K. pneumoniae strains. The above TEM images represent the surface fimbriae visualised on K. pneumoniae (DN2,
∆T6S1, ∆T6S3 and ∆T6S13) with scale bar indicated is 200 nm on each. ∆T3 showed pronounced images have
been digitally zoomed in 2 times the original size at a key point to reveal the difference in the surface fimbriae
identified. Briefly, K. pneumoniae strains were grown in LB at 37µ overnight with shaking. A subculture of 1/100
was made into fresh LB and allowed to grow until OD600nm reach 0.2. The bacterial strains were harvested and
resuspended in PBS at 105 CFU for TEM. Samples (5 µl) were applied to a hydrophilic Formvar-carbon coated
copper grid and allowed to adsorb for 5 minutes. After wicking the excess liquid and washing with distilled
deionized water, samples were stained 15 seconds with 5 μl of 1 % uranyl acetate (pH 4.5). All the Transmission
Electron Microscopy were performed at the Centre for Core Biotechnology Services (University of Leicester)
using JEOL JEM-1400 microscope at 80 kV.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
161
4.9 K. pneumoniae T6SSs mediate resistance to Acanthamoeba castellanii, complement-
mediated killing and PMN internalisation
K. pneumoniae, in its natural habitat or host, faces phagocytic pressure and hence, the organism may have
evolved new mechanisms to evading phagosome or possible strike back when under attack. To survive in
both the host and environment, K. pneumoniae is taking no chances but T6SS deploy its T6SS as a full
virulence factor. To demonstrate that K. pneumoniae requires T6SSs to avoid phagocytic protozoans such
as environmental amoeba and persist in the environment, A. castellanii-K. pneumoniae interaction put to
the test relative T6SS mutagenesis in K. pneumoniae. Briefly, a monolayer of A. castellanii was infected
with the various K. pneumoniae strains at an MOI of 100. A blank media without Aa castellanii was
inoculated with the different strains and counted at the end of the time point to serve as the denominator in
the calculation of the percentage. Percentage survival is equal to CFU/ml from the sample (with amoeba)
divided by the CFU/ml of bacterial strains from the blank medium multiply by 100%. For association assay,
the A. castellanii monolayer was washed three times with PBS after infection and lysed to determine the
bacterial CFU. In the case of bacterial internalisation assay, 100µg/ml kanamycin was used to kill the
extracellular bacteria for 20 minutes after post-infection, and the bacterial CFU were then determined after
three repeated washes with PBS. To perform A. castellanii zone of clearance, thin lawns of the various
bacterial strains were on a solid agar surface. A. castellanii plaque formation was observed after ~ 2 ₓ 105
was spotted on the bacterial lawn and incubated at 37ᴼC overnight for the zone of clearance. Similarly, just
like the plaque formation, the percentage viability of the A. castellanii spotted on the agar surface was to
determine using trypan blue staining after 2 h post-infection and the viability of the amoeba estimated.
For the first time, I shown K. pneumoniae resist A. castellanii phagocytosis through a T6SS-dependent
puncturing mechanism, rendering the inviable with short space of time. From the data collected, the parent
strain (DN2) was able to comfortably resist the amoeba both in the association and the invasion assays
compared to the other strains, while both ∆T1 and ∆T3 showed show 20-30 fold change in the association
assay (Figure 4.11A). ∆T1∆T3 was not able to attach to the amoeba well even though it did not have active
T6SS to have resisted. This was not surprising since it was demonstrated earlier where ∆T1∆T3 showed

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
162
the poor formation of biofilm and surface fimbriae (Figure 4.8, Figure 4.9 and Figure 4.10) though it was
the most internalised of all the mutants relative to DN2. As shown in Figure 4.11A about 5-times, more of
∆T1 was internalised compared to ∆T3. In the amoeba plaque formation (Figure 4.11B), ∆T1∆T3 lawn
showed the widest and most visible zone of clearance by the A. castellanii with about 91% of the initial A.
castellanii still viable surviving after the 2-hr endpoint. Once again, DN2 just like the phenotypes shown
in both the association and internalisation assays was able to resist the clearance by the amoeba and lowed
the viability of the amoeba to a damming 78% (Figure 4.11). ∆T3 was the next most internalised with after
DN2 while ∆T1 shown arguably wide zone of clearance after ∆T1∆T3.
Figure 4.11 T6SS enhances K. pneumoniae against Acanthamoeba castellanii.
(A) At an MOI of 100, the bacterial strains association or internalisation with A. castellanii after 2 h post-infection.
For association assay, the A. castellanii monolayer was infected with K pneumoniae (MOI 1:100) FOR 1 h at 30℃
and subsequently washed three times with PBS. K. pneumoniae CFUs were enumerated by lyzing the A.castellanii
trophozoites and plating the diluted lysates on LA and LA+ ampicillin. In the case of bacterial internalisation assay,
100µg/ml kanamycin was used to kill the extracellular bacteria for 20 minutes post-infection, and the bacterial CFU
were then determined as decribed above. Plates were incubated at 37 overnight for K. pneumoniae CFUs. Two-way
ANOVA with Tukey post-hoc for multiple comparisons, the data indicate the means from three experimental
replicates and experiments (B) A. castellanii plaque formation were observed after ~ 2 ₓ 105 was spotted on the
bacterial lawn and incubated at 37ᴼC overnight. The percentage viability of the A. castellanii was determined after
2 h post-infection using trypan blue staining. Using trypan blue and hemocytometer, A.castellanii trophozoites
viability was determined by collecting the spotted A.castellanii trophozoites on the bacterial lawns. Data are shown
as mean + SD from three experimental replicates.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
163
Host infection, K. pneumoniae do not only battle with cell uptake but are faced with the full wrath of the
human immune system, including the antimicrobial properties of the serum and cells. Unfortunately, the
same cannot be said for immunocompromised individuals. The serum has a compliment. Complement-
mediated killing is in serums is a significant barrier against K. pneumoniae infection. Complement-
mediated killing largely dependents on bacteria surface receptor- binding could affect bacterial clearance
in the blood should these surface receptors go imbalance. Here I tested how the deletion T6SS1 and/or
T6SS3 could affect the bacterial ability to resist serum-mediated killing and C3b deposition on to the
bacterial surface. Briefly, log-phase bacteria (DN2, ∆T1, ∆T3 and ∆T1∆T3) were resuspended in 75% of
pool normal human serum (PNHS) and incubated 37 oC for the various time-points. Heat-inactivated PNHS
was equally inoculated with the bacteria and used as control and use as the denominator for the calculation
of the bacteria percentage survival. For the C3 deposition, the bacteria were opsonised for 20 minutes in
PNHS and the C3b was determine by Elisa method. Also, opsonised bacteria used to inoculate a monolayer
of polymorphonuclear (PMN) leukocytes A. castellanii monolayer and the internalised bacteria was
determined just as in the previous case.
This is the first demonstration of T6SS involvement in the deposition of C3b onto the bacterial surface and
resistance to the complement-mediated killing of K. pneumoniae (Figure 4.12). From the date, ∆T1∆T3
was the lest recovered, and less C3b deposition since over 75% of ∆T1 was killed at the endpoint of the
experiment. Over 75% of DN2 showed survived the serum killing but had less C3b deposition within the
first 20 minutes. ∆T3 had the highest C3b deposition probably accounting for it diminishing survival in the
serum after ∆T1 and ∆T1∆T3 C3b deposition. Since the DN2 had a very poor association and
internalisation, I decided to enhance the bacteria (DN2, ∆T1, ∆T3 and ∆T1∆T3) uptake by A. castellanii
opso-phagocytosis after opsonising the bacteria with PNHS for 20 minutes. The data showed that ∆T1 and
∆T3 were the most internalised after the first 1h. This was not surprising as these bacteria expressed high
surface attachments and C3b deposition even though only ∆T1 and ∆T1∆T3 were the most digested after 3
h. DN2 was the most internalised at the end of the 3h, maybe because they can survive within the amoeba.
All these data suggest that T6SS is an essential virulence factor in K. pneumoniae, especially in MDR
strains. During K. pneumoniae infection, T6SS may be deployed to enhanced biofilm formation in
intravenous devices, fimbriae expression for attachment and resistance to phagocytosis, thereby evading
the host immune system. In the environment, K. pneumoniae may be good at using T6SS-dependent killing
and defence to outcompete and colonise polymicrobial niche. My data also suggest that T6SS may be
involved in resistance to complement-mediated killing and evasion of environmental amoeba phagocytosis.
For the first with overwhelming evidence from my data, I suggest that the putative T6SS is a crucial factor
in the pathogenicity of K. pneumoniae
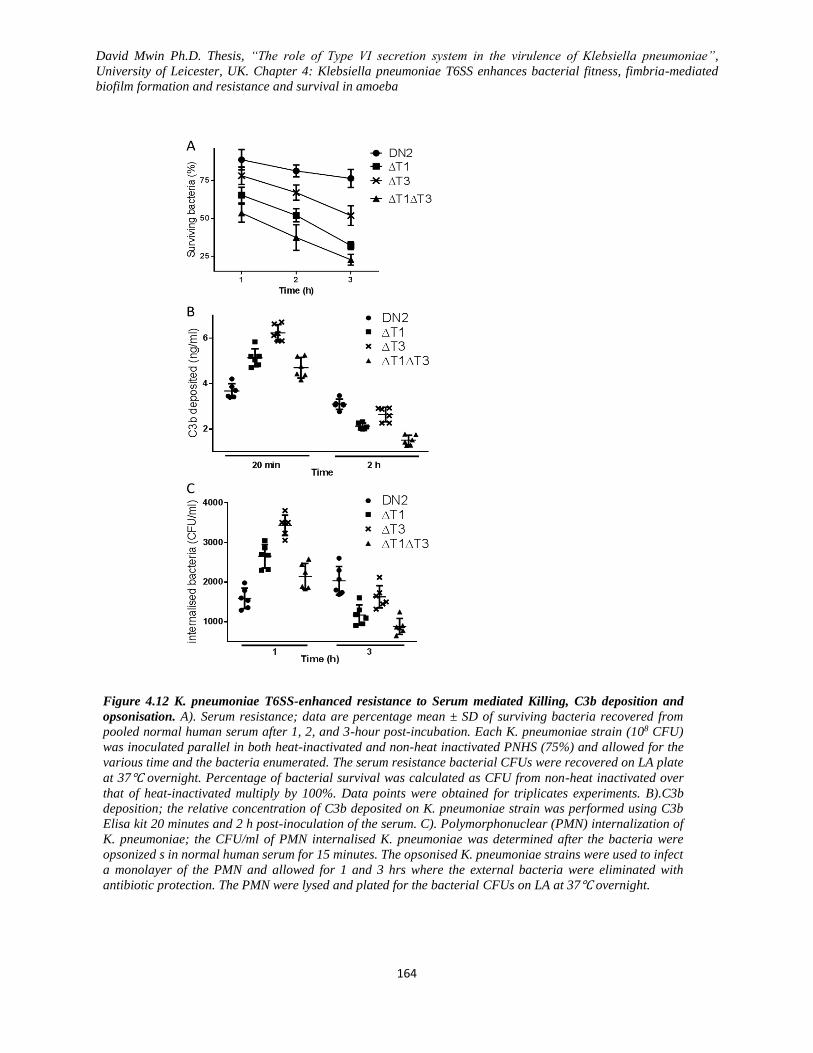
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
164
Figure 4.12 K. pneumoniae T6SS-enhanced resistance to Serum mediated Killing, C3b deposition and
opsonisation. A). Serum resistance; data are percentage mean ± SD of surviving bacteria recovered from
pooled normal human serum after 1, 2, and 3-hour post-incubation. Each K. pneumoniae strain (108 CFU)
was inoculated parallel in both heat-inactivated and non-heat inactivated PNHS (75%) and allowed for the
various time and the bacteria enumerated. The serum resistance bacterial CFUs were recovered on LA plate
at 37℃ overnight. Percentage of bacterial survival was calculated as CFU from non-heat inactivated over
that of heat-inactivated multiply by 100%. Data points were obtained for triplicates experiments. B).C3b
deposition; the relative concentration of C3b deposited on K. pneumoniae strain was performed using C3b
Elisa kit 20 minutes and 2 h post-inoculation of the serum. C). Polymorphonuclear (PMN) internalization of
K. pneumoniae; the CFU/ml of PMN internalised K. pneumoniae was determined after the bacteria were
opsonized s in normal human serum for 15 minutes. The opsonised K. pneumoniae strains were used to infect
a monolayer of the PMN and allowed for 1 and 3 hrs where the external bacteria were eliminated with
antibiotic protection. The PMN were lysed and plated for the bacterial CFUs on LA at 37℃ overnight.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 4: Klebsiella pneumoniae T6SS enhances bacterial fitness, fimbria-mediated
biofilm formation and resistance and survival in amoeba
165

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
166
Chapter Five
K.pneumoniae T6SS mediate virulence and activates
a pro-inflammatory innate immune response in the
host.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
167
Chapter 5: K. pneumoniae T6SSs Mediate Virulence and
Activation Host Cell Innate Immune Response
5.1 T6SSs in K. pneumoniae promote Galleria killing and in vivo survival of bacteria
Galleria mellonella larvae are widely adopted as an in vivo model in the study of bacterial virulence, and
this model has been used for determining the virulence of K. pneumoniae strains(Wand et al., 2013a; Wand
et al., 2013b). This in vivo model has been well established in the laboratory for several years and has
shown many prospects in studying the virulence of many of the laboratory strains. T6SS has been widely
reported to pose not only antagonism towards bacteria but also eukaryotic host cell. To determine the role
virulence of K. pneumoniae T6SS exhibit towards larvae, an inoculum of 20µl containing ~108 CFU of
each strain (DN2, ∆T1, ∆T3 or ∆T1∆T3) resuspended in 10 mM MgSO4 was injected into twenty larvae for
each group and the motility of the larvae was observed and enumerated of time (day 1 to day 6). Two
control group (one group untouched and the other group injected with 20µl bMgSO4) were used to assess
the quality of each experiment. The percentage motility of the larvae is represented in a graph as shown
below. All the larvae were weighed (~250g) to ensure consistency of weight within the various groups and
the data points are means of three independent experiments. For the in vivo bacterial survival assay (single
strain survival or competition mixtures), the larvae were injected with a sub-lethal dose (2×106 CFU) of
either a single bacterial strain (DN, ∆T1, ∆T3 or ∆T1∆T3) or a 1:1 ratio mixtures of DN2 and the mutants
(DN2+∆T1, DN2+∆T3 or DN2+∆T1∆T3) to ensure that all the larvae survived by the end of the
experiment. The CFU/ml of each bacterial strain either from the single strain inoculation samples or
competition mixtures at each time point were enumerated from the 20µl hemocoel extracted from each
larva within the group treatments. The virulence of T6SS in hyper mucoid K. pneumoniae NTUH-K2044
(also known as KR1730) was tested relative to its T6SS mutant (NTUH∆T1) as shown in the in Figure 5.3.
From the Galleria larvae mortality data, it was noticed that DN2 had had high mortality effect on the
population of the larvae by reducing the infected larvae to about 50% within day 2, steadily to 15% by day
6, and finally to at least 85% mortality rate by the end of the experiment. Similarly, the larvae group that
was infected with ∆T3 showed a lower survival (at least 30% by day 6) relative to that of ∆T1 (at least 60%
by 6) and ∆T1∆T3 (at least 75% by 6) but not DN2 (at least 15% by day 6). At the end of the experiment,
the survival rate of the group treated with ∆T1∆T3 was almost equal to the mortality rate of the group
treated with DN. Likewise, ∆T1-treated larva group had a survival rate similar to the mortality rate of the
∆T3-treated larva group. The group treatment with ∆T1∆T3 survived better among all the groups while the
group treated with DN2 showed the least survival. The graph shown in Figure 5.1 is from two experimental
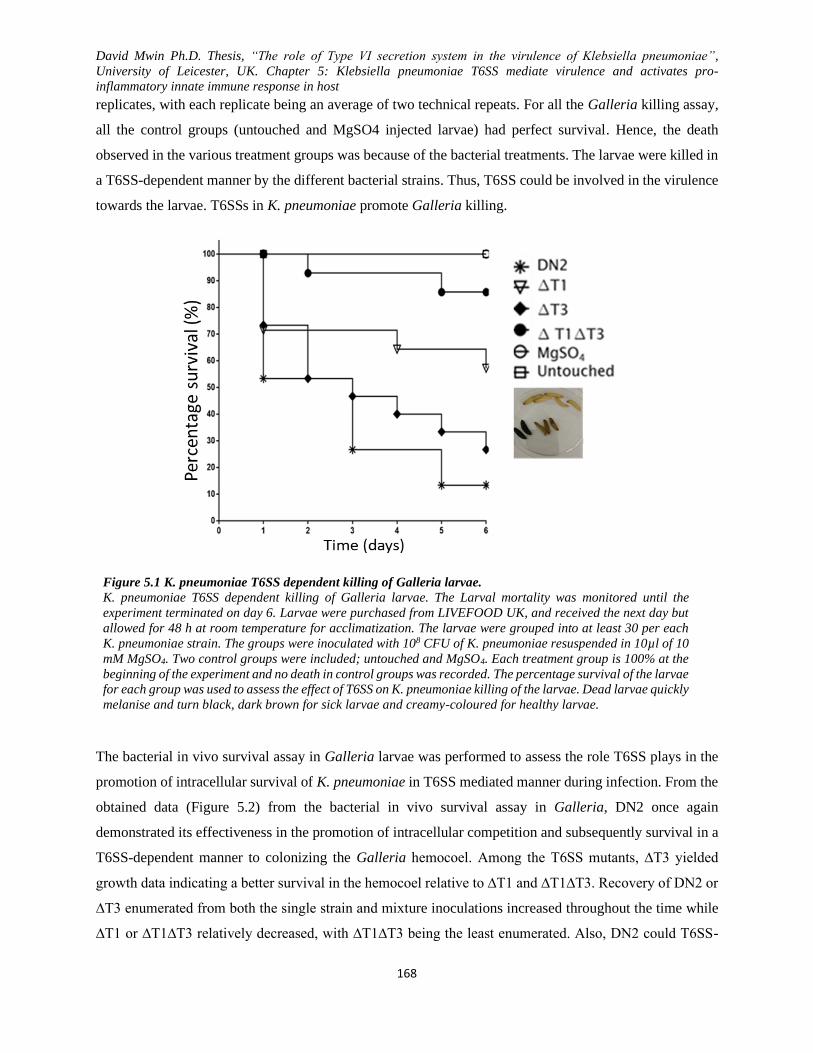
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
168
replicates, with each replicate being an average of two technical repeats. For all the Galleria killing assay,
all the control groups (untouched and MgSO4 injected larvae) had perfect survival. Hence, the death
observed in the various treatment groups was because of the bacterial treatments. The larvae were killed in
a T6SS-dependent manner by the different bacterial strains. Thus, T6SS could be involved in the virulence
towards the larvae. T6SSs in K. pneumoniae promote Galleria killing.
Figure 5.1 K. pneumoniae T6SS dependent killing of Galleria larvae.
K. pneumoniae T6SS dependent killing of Galleria larvae. The Larval mortality was monitored until the
experiment terminated on day 6. Larvae were purchased from LIVEFOOD UK, and received the next day but
allowed for 48 h at room temperature for acclimatization. The larvae were grouped into at least 30 per each
K. pneumoniae strain. The groups were inoculated with 108 CFU of K. pneumoniae resuspended in 10µl of 10
mM MgSO4. Two control groups were included; untouched and MgSO4. Each treatment group is 100% at the
beginning of the experiment and no death in control groups was recorded. The percentage survival of the larvae
for each group was used to assess the effect of T6SS on K. pneumoniae killing of the larvae. Dead larvae quickly
melanise and turn black, dark brown for sick larvae and creamy-coloured for healthy larvae.
The bacterial in vivo survival assay in Galleria larvae was performed to assess the role T6SS plays in the
promotion of intracellular survival of K. pneumoniae in T6SS mediated manner during infection. From the
obtained data (Figure 5.2) from the bacterial in vivo survival assay in Galleria, DN2 once again
demonstrated its effectiveness in the promotion of intracellular competition and subsequently survival in a
T6SS-dependent manner to colonizing the Galleria hemocoel. Among the T6SS mutants, ∆T3 yielded
growth data indicating a better survival in the hemocoel relative to ∆T1 and ∆T1∆T3. Recovery of DN2 or
∆T3 enumerated from both the single strain and mixture inoculations increased throughout the time while
∆T1 or ∆T1∆T3 relatively decreased, with ∆T1∆T3 being the least enumerated. Also, DN2 could T6SS-
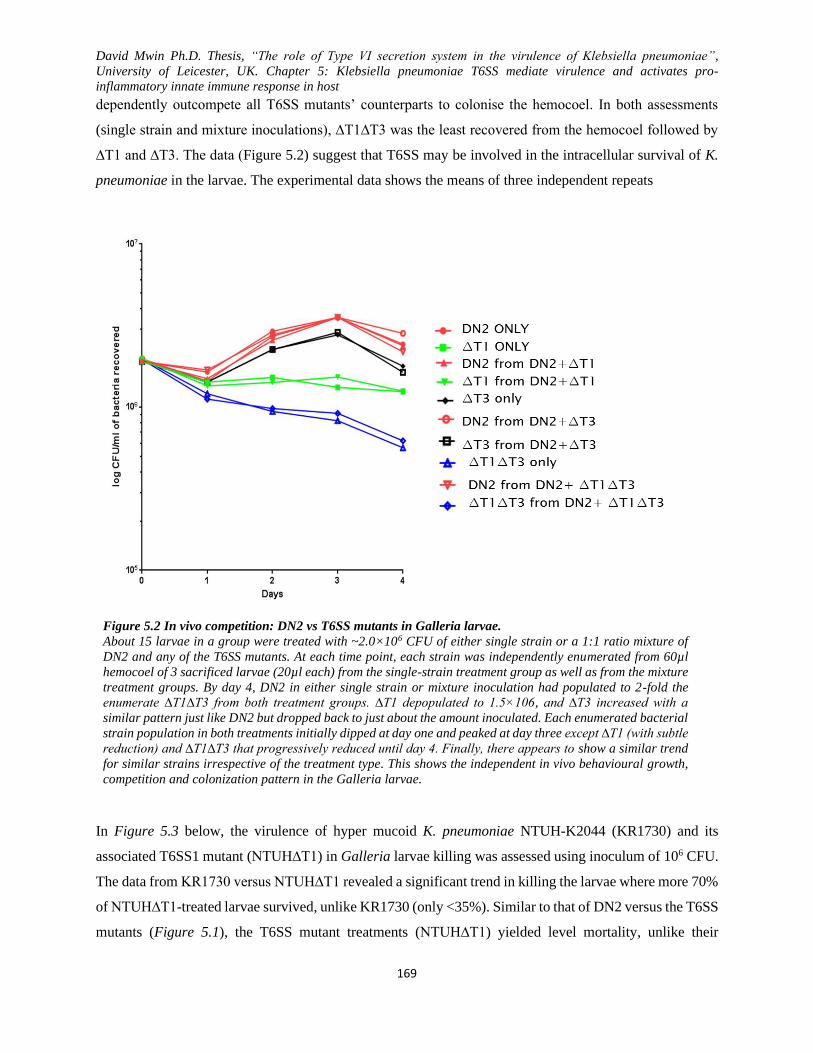
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
169
dependently outcompete all T6SS mutants’ counterparts to colonise the hemocoel. In both assessments
(single strain and mixture inoculations), ∆T1∆T3 was the least recovered from the hemocoel followed by
∆T1 and ∆T3. The data (Figure 5.2) suggest that T6SS may be involved in the intracellular survival of K.
pneumoniae in the larvae. The experimental data shows the means of three independent repeats
Figure 5.2 In vivo competition: DN2 vs T6SS mutants in Galleria larvae.
About 15 larvae in a group were treated with ~2.0×106 CFU of either single strain or a 1:1 ratio mixture of
DN2 and any of the T6SS mutants. At each time point, each strain was independently enumerated from 60µl
hemocoel of 3 sacrificed larvae (20µl each) from the single-strain treatment group as well as from the mixture
treatment groups. By day 4, DN2 in either single strain or mixture inoculation had populated to 2-fold the
enumerate ∆T1∆T3 from both treatment groups. ∆T1 depopulated to 1.5×106, and ∆T3 increased with a
similar pattern just like DN2 but dropped back to just about the amount inoculated. Each enumerated bacterial
strain population in both treatments initially dipped at day one and peaked at day three except ∆T1 (with subtle
reduction) and ∆T1∆T3 that progressively reduced until day 4. Finally, there appears to show a similar trend
for similar strains irrespective of the treatment type. This shows the independent in vivo behavioural growth,
competition and colonization pattern in the Galleria larvae.
In Figure 5.3 below, the virulence of hyper mucoid K. pneumoniae NTUH-K2044 (KR1730) and its
associated T6SS1 mutant (NTUH∆T1) in Galleria larvae killing was assessed using inoculum of 106 CFU.
The data from KR1730 versus NTUH∆T1 revealed a significant trend in killing the larvae where more 70%
of NTUH∆T1-treated larvae survived, unlike KR1730 (only <35%). Similar to that of DN2 versus the T6SS
mutants (Figure 5.1), the T6SS mutant treatments (NTUH∆T1) yielded level mortality, unlike their
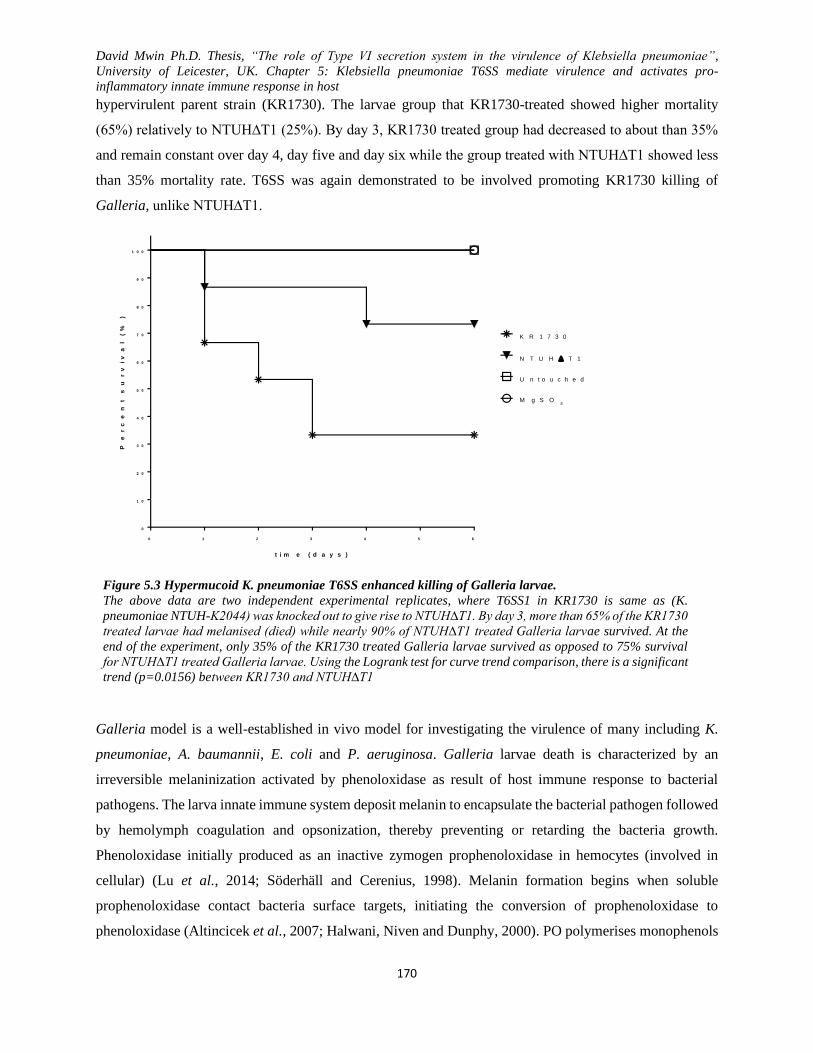
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
170
hypervirulent parent strain (KR1730). The larvae group that KR1730-treated showed higher mortality
(65%) relatively to NTUH∆T1 (25%). By day 3, KR1730 treated group had decreased to about than 35%
and remain constant over day 4, day five and day six while the group treated with NTUH∆T1 showed less
than 35% mortality rate. T6SS was again demonstrated to be involved promoting KR1730 killing of
Galleria, unlike NTUH∆T1.
Figure 5.3 Hypermucoid K. pneumoniae T6SS enhanced killing of Galleria larvae.
The above data are two independent experimental replicates, where T6SS1 in KR1730 is same as (K.
pneumoniae NTUH-K2044) was knocked out to give rise to NTUH∆T1. By day 3, more than 65% of the KR1730
treated larvae had melanised (died) while nearly 90% of NTUH∆T1 treated Galleria larvae survived. At the
end of the experiment, only 35% of the KR1730 treated Galleria larvae survived as opposed to 75% survival
for NTUH∆T1 treated Galleria larvae. Using the Logrank test for curve trend comparison, there is a significant
trend (p=0.0156) between KR1730 and NTUH∆T1
Galleria model is a well-established in vivo model for investigating the virulence of many including K.
pneumoniae, A. baumannii, E. coli and P. aeruginosa. Galleria larvae death is characterized by an
irreversible melaninization activated by phenoloxidase as result of host immune response to bacterial
pathogens. The larva innate immune system deposit melanin to encapsulate the bacterial pathogen followed
by hemolymph coagulation and opsonization, thereby preventing or retarding the bacteria growth.
Phenoloxidase initially produced as an inactive zymogen prophenoloxidase in hemocytes (involved in
cellular) (Lu et al., 2014; Söderhäll and Cerenius, 1998). Melanin formation begins when soluble
prophenoloxidase contact bacteria surface targets, initiating the conversion of prophenoloxidase to
phenoloxidase (Altincicek et al., 2007; Halwani, Niven and Dunphy, 2000). PO polymerises monophenols
0 1 2 3 4 5 6
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
2 : G a l l e r i a K i l l i n g a s s a y ; K R 1 7 3 0 v s i t T 6 S S 1 m u t a n t E x p 2
t i m e ( d a y s )
Pe
rc
en
t
su
rv
iv
al
(
%)
K R 1 7 3 0
N T U H T 1
U n t o u c h e d
M g S O4
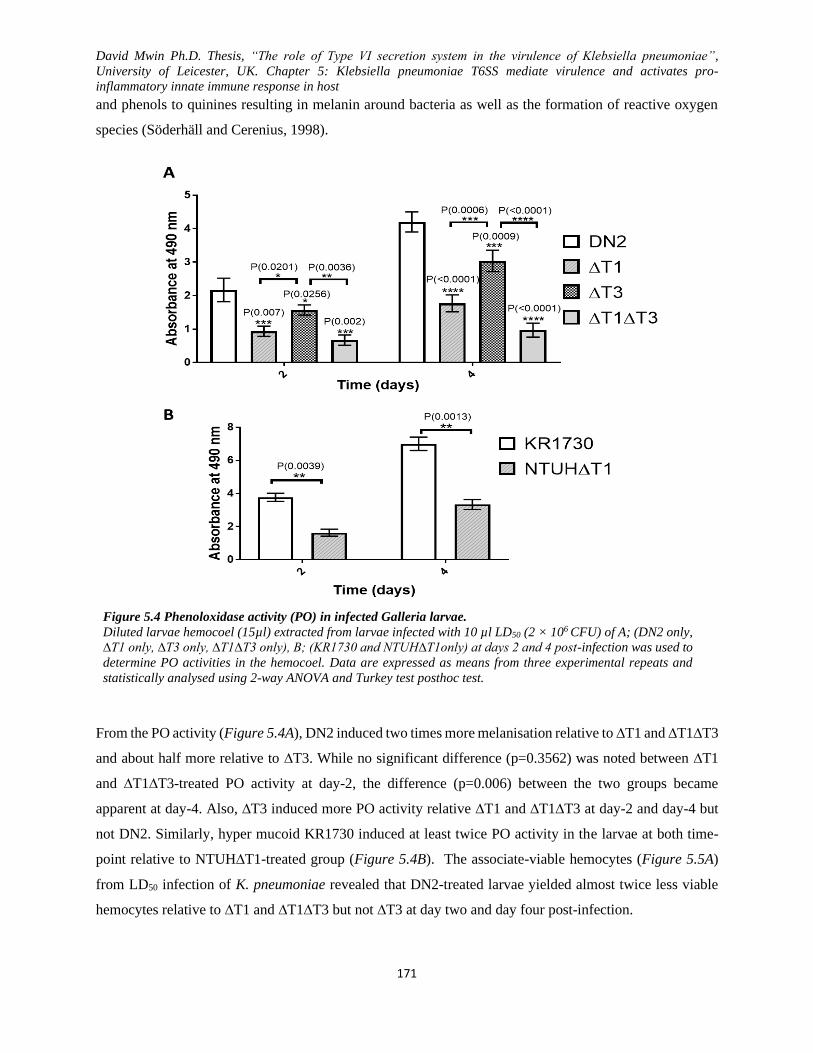
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
171
and phenols to quinines resulting in melanin around bacteria as well as the formation of reactive oxygen
species (Söderhäll and Cerenius, 1998).
Figure 5.4 Phenoloxidase activity (PO) in infected Galleria larvae.
Diluted larvae hemocoel (15µl) extracted from larvae infected with 10 µl LD50 (2 × 106 CFU) of A; (DN2 only,
∆T1 only, ∆T3 only, ∆T1∆T3 only), B; (KR1730 and NTUH∆T1only) at days 2 and 4 post-infection was used to
determine PO activities in the hemocoel. Data are expressed as means from three experimental repeats and
statistically analysed using 2-way ANOVA and Turkey test posthoc test.
From the PO activity (Figure 5.4A), DN2 induced two times more melanisation relative to ∆T1 and ∆T1∆T3
and about half more relative to ∆T3. While no significant difference (p=0.3562) was noted between ∆T1
and ∆T1∆T3-treated PO activity at day-2, the difference (p=0.006) between the two groups became
apparent at day-4. Also, ∆T3 induced more PO activity relative ∆T1 and ∆T1∆T3 at day-2 and day-4 but
not DN2. Similarly, hyper mucoid KR1730 induced at least twice PO activity in the larvae at both time-
point relative to NTUH∆T1-treated group (Figure 5.4B). The associate-viable hemocytes (Figure 5.5A)
from LD50 infection of K. pneumoniae revealed that DN2-treated larvae yielded almost twice less viable
hemocytes relative to ∆T1 and ∆T1∆T3 but not ∆T3 at day two and day four post-infection.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
172
Figure 5.5 Parentage viability cell count in larvae.
Haemocytes viability in larvae infected with 10 µl LD50 (2 × 106 CFU) of A) DN2, ∆T1, ∆T3 and ∆T1∆T3 and
B) KR1730 and NTUH∆T1 at 2- and 4-days post-infection. Data are expressed as means from three
experimental repeats and statistically analysed using 2-way ANOVA and Turkey test posthoc test
Generally, there was a decrease in the viable hemocytes at days-4 relative to day-2 though ∆T1∆T3 yielded
the most viable cells followed by ∆T1 and ∆T3 and DN2, respectively. No significant difference was noted
between ∆T1 and ∆T1∆T3 at days-2 (p=0.15) but not at day-4 (p=0.0072) post-infection. NTUH∆T1
(p<0.0001) significantly yielded more viable hemocytes relative to its parent strain, KR1730 at both day-2
and day-4 (Figure 5.5B). The ability of K. pneumoniae to induce melanin formation (Figure 5.4) and
reduction in hemocytes viability (Figure 5.5) in a virulent T6SS-dependent-manner was consistent with the
larva killing assay (Figure 5.1 and Figure 5.3), suggesting that the killing of the larvae is due to the
virulence of the K. pneumoniae strains. The deletion of T6SS1 and/ or T6SS3 clusters in K. pneumoniae
significantly attenuate the virulence in G. mellonella larvae.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
173
5.2 K. pneumoniae T6SSs enhance phagocytic uptake and survival in murine macrophage
T6SS has been reported to be involved in the mediation of virulence and evasion of host innate immune
system (Tan et al., 2010; Suarez et al., 2010b; Ma and Mekalanos, 2010b; Jayamani and Mylonakis, 2014).
Phagocytosis is a necessary process used by many host cells such as macrophages to attach and digest
pathogens, including bacteria. Briefly, a monolayer of J774.A1 was infected at MOI 10:1 for 1h post-
infection, and gentamicin was used to kill the extracellular bacteria, and the bacteria were enumerated on
LA at 37 ℃. For visualization of phagocytosed GFP-tagged K. pneumoniae, a J774.A1 monolayer on
coverslip was infected for 1h and treated with gentamicin and subsequently fixed with 4%
paraformaldehyde and DAPI-stained. Finally, to investigate how K. pneumoniae T6SSs promote
phagocytotic uptake and survival, J774.A1 cell line was infected (107 CFU) with DN2, ∆T1, ∆T3 and
∆T1∆T3. After 1hr post-infection, antibiotic was added to the media and maintained for 2, 4, and 6 and
subsequently washed three times with PBS before Triton-X lysis for bacteria CFU enumeration.
Presented in Figure 5.6A, the J774.A1 uptake data suggest that the recovered K. pneumoniae was
phagocytosed in a manner that is consistent with the presence or absence of T6SSs. J774.A1 phagocytosed
more DN2 (p<0.0001) relative to the T6SS mutants (∆T1, ∆T3 and ∆T1∆T1) for the 1 h post-infection.
Additionally, ∆T3-treated J774.A1 significantly yielded more bacteria recovery relative to ∆T1 (p=0.0051)
and ∆T1∆T3 (p<0.0001) while ∆T1∆T3-treated J774.A1 yielded the least number of bacteria. The data
demonstrate that murine macrophages can actively phagocytose K. pneumoniae in a T6SS-dependent
manner where the deletion of T6SSs in K. pneumoniae attenuates bacteria recovery from the J774.A1
phagocytosis assay.
To ascertain the K. pneumoniae phagocytosis by J774.A1, the GFP-tagged bacteria were visualized using
confocal microscopy. The confocal microscopy data (Figure 5.6B) was consistent with the bacteria
recovery data, suggesting that more loads of DN2 unlike ∆T1 or ∆T1∆T3 that were taken- up by the
macrophage could easily be seen around the nuclei of the macrophages. More DN2 were visualized to be
present in the macrophage more than those of ∆T1, ∆T3 and ∆T1∆T3. Next, a relatively good amount of
∆T3 was noted in the macrophages relative to ∆T1 and ∆T1∆T3. The confocal microscopy data revealed
increased intracellular presence of DN2 in J774.A1 when compared to the T6SS mutant (∆T1 and ∆T1∆T3)
but not ∆T3, which showed few invaded bacteria. This strongly suggests that K. pneumoniae in T6SS-
dependent fashion can be phagocytosed and survive in murine macrophages. K. pneumoniae may be
actively using T6SS to engage the macrophage during the uptake and survival within the macrophage. Also,
fewer bacteria recovery was observed for K. pneumoniae T6SS mutants, particularly ∆T1 and ∆T1∆T3,
which were noted to have reduced it intracellular virulence due to the deletion of T6SSs, thus resulting in
lesser intracellular survival even have J774.A1 uptake.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
174
Having noticed the phagocytotic uptake of the K. pneumoniae at 1 h, to better assess the ability of K.
pneumoniae to survive in macrophages, the bacteria recovered from the infected cell line was done at the
various time point (2, 4, & 6 h), Figure 5.7. Generally, while all the T6SS mutants yield a reducing bacteria
recovery over the 6 h period, their parent strain (DN2) showed an increasing trend in the number of bacteria
that were recovered. Among the T6SS mutants, T6SS double knockout (∆T1∆T3) produced the least
amount of bacteria while ∆T3 was recovered more than ∆T1. The deletion of T6SSs impaired the ability of
the F6SS mutants to survive in the macrophage intracellularly.
DN2 with both active T6SS1 and T6SS3 was largely by phagocytosed by the J774.A1 at all the time points,
unlike the T6SS mutants. The results suggest that the presence or absence of T6SS influence the phagocytic
uptake of K. pneumoniae and subsequent survival in J774.A1. Though K. pneumoniae is largely considered
not to be an intracellular bacterium, more DN2 invaded and survived better in the macrophages and possibly
replicated over the examined time-points compared to the T6SS mutants (∆T1, ∆T3 and ∆T1∆T3). Unlike
the recovered DN2, T6SS mutants initially invaded the macrophages and over time and subsequently
decreased in population/survival in the J774.A1. The recovered population for the highly fimbriated ∆T3
from the macrophages was significantly higher than that of ∆T1 and ∆T1∆T3 but not DN2 suggesting that,
though the bacterium was taken up by the macrophage, it subsequently struggled to survive in the
macrophage, unlike DN2. After 6 hours post-infection of the macrophage monolayer, while DN2 invaded
and survived well in the macrophages, ∆T1, ∆T3 and ∆T1∆T3 that had either one or both T6SS clusters
deleted could not progressively invade and survive in the macrophages. These data, as shown in Figure 5.6
and Figure 5.7 suggest that T6SS in mediate virulence towards macrophages by enhancing phagocytic
uptake and intracellular survival within the macrophages. The involvement of K. pneumoniae T6SS in
macrophage invasion and survival unlike resistance of macrophages reported in other studies is yet another
virulence decoy employed by a K. pneumoniae evade host immune system while unleashing its virulence.
This data has there informed me to hypothesizes that the putative T6SS gene clusters (T6SS1 and T6SS3)
may play a role in the expression of T6SS -dependent surface antigen/ receptors for binding and surviving
in host cells, such macrophages. This would not be surprising since the deletion of T6SSs enhanced surface
fimbriae expression and biofilm. This data is the first to demonstrate the virulent of K. pneumoniae T6SSs
towards murine macrophages (J774.A1), which in this case, enhanced invasion and intracellular survival
within the macrophages.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
175
Figure 5.6 J7774.A1 phagocytosis uptake and visualization of K. pneumoniae.
K. pneumoniae phagocytic uptake and survival in murine macrophage (J774.A1) is T6SS enhanced. A) Invasion
and intracellular survival in J774.A1 at 1 hr post-infection; the infected bacteria strains were recovered on LA
at 37 ℃ after gentamicin treatment and several washes. The data is a mean of triplicate experiment with
duplicate repeats and statistically analysed using one-way ANOVA with Turkey’s multiple comparisons test.
B) J744.A1 infected by GFP-tagged K. pneumoniae strains for 1 h and the extracellular bacteria were killed
by gentamicin. The nuclei of the infected monolayer on coverslips were DAPI-stained and visualised with
confocal microscopy.
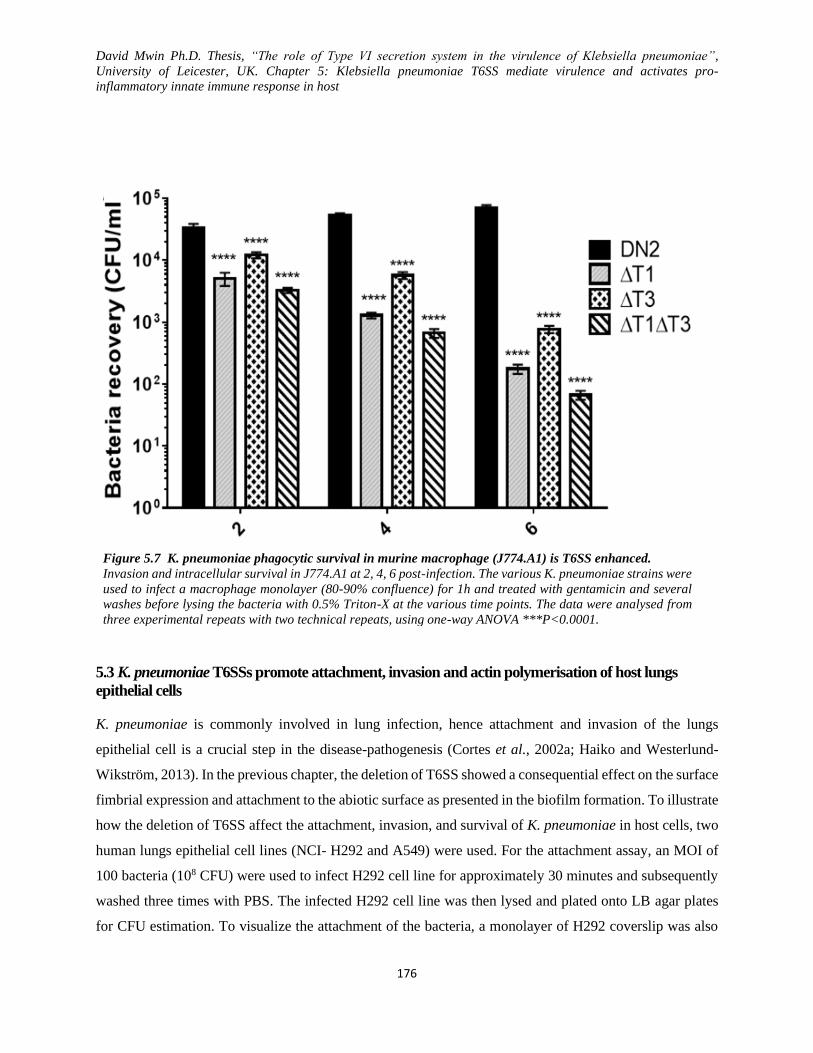
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
176
Figure 5.7 K. pneumoniae phagocytic survival in murine macrophage (J774.A1) is T6SS enhanced.
Invasion and intracellular survival in J774.A1 at 2, 4, 6 post-infection. The various K. pneumoniae strains were
used to infect a macrophage monolayer (80-90% confluence) for 1h and treated with gentamicin and several
washes before lysing the bacteria with 0.5% Triton-X at the various time points. The data were analysed from
three experimental repeats with two technical repeats, using one-way ANOVA ***P<0.0001.
5.3 K. pneumoniae T6SSs promote attachment, invasion and actin polymerisation of host lungs
epithelial cells
K. pneumoniae is commonly involved in lung infection, hence attachment and invasion of the lungs
epithelial cell is a crucial step in the disease-pathogenesis (Cortes et al., 2002a; Haiko and Westerlund-
Wikström, 2013). In the previous chapter, the deletion of T6SS showed a consequential effect on the surface
fimbrial expression and attachment to the abiotic surface as presented in the biofilm formation. To illustrate
how the deletion of T6SS affect the attachment, invasion, and survival of K. pneumoniae in host cells, two
human lungs epithelial cell lines (NCI- H292 and A549) were used. For the attachment assay, an MOI of
100 bacteria (108 CFU) were used to infect H292 cell line for approximately 30 minutes and subsequently
washed three times with PBS. The infected H292 cell line was then lysed and plated onto LB agar plates
for CFU estimation. To visualize the attachment of the bacteria, a monolayer of H292 coverslip was also
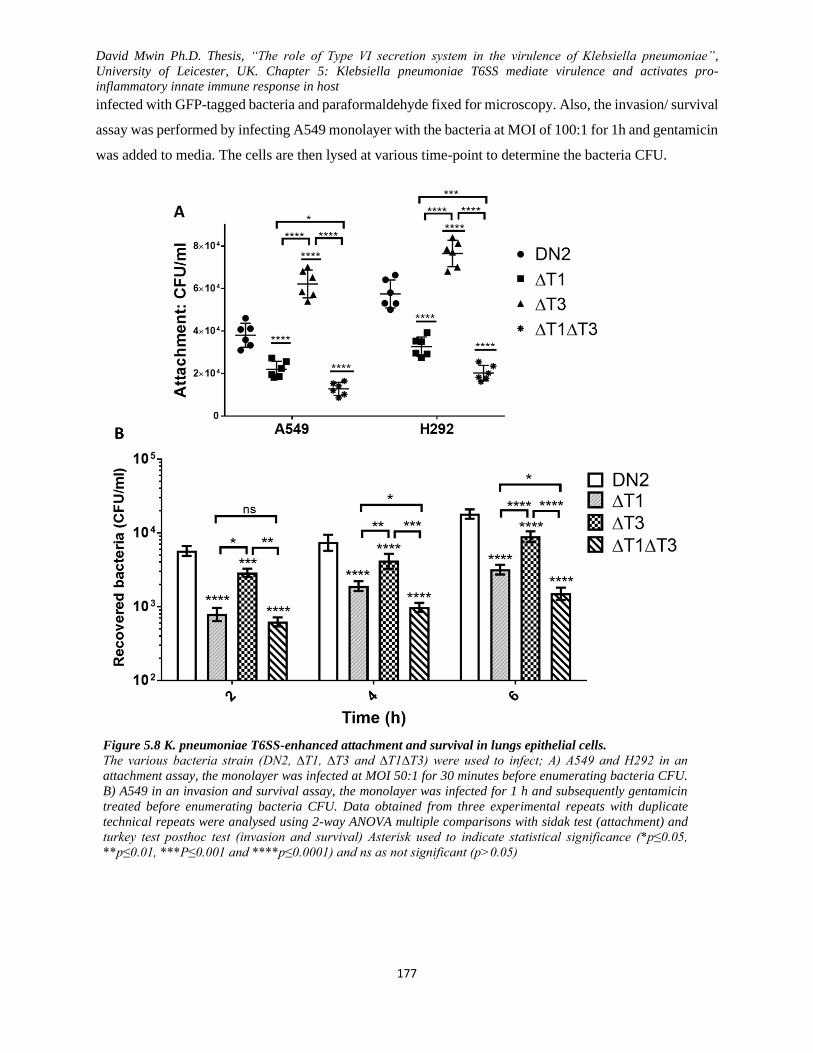
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
177
infected with GFP-tagged bacteria and paraformaldehyde fixed for microscopy. Also, the invasion/ survival
assay was performed by infecting A549 monolayer with the bacteria at MOI of 100:1 for 1h and gentamicin
was added to media. The cells are then lysed at various time-point to determine the bacteria CFU.
Figure 5.8 K. pneumoniae T6SS-enhanced attachment and survival in lungs epithelial cells.
The various bacteria strain (DN2, ∆T1, ∆T3 and ∆T1∆T3) were used to infect; A) A549 and H292 in an
attachment assay, the monolayer was infected at MOI 50:1 for 30 minutes before enumerating bacteria CFU.
B) A549 in an invasion and survival assay, the monolayer was infected for 1 h and subsequently gentamicin
treated before enumerating bacteria CFU. Data obtained from three experimental repeats with duplicate
technical repeats were analysed using 2-way ANOVA multiple comparisons with sidak test (attachment) and
turkey test posthoc test (invasion and survival) Asterisk used to indicate statistical significance (*p≤0.05,
**p≤0.01, ***P≤0.001 and ****p≤0.0001) and ns as not significant (p>0.05)

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
178
The attachment of the bacteria strain to A549 and H292 monolayer (Figure 5.8A) revealed that ∆T3 yielded
the highest bacterial attachment/ association with the cells relative to DN2 ∆T1 and ∆T1∆T3, possibly
because of the increased surface appendages/ fimbrial expression, unlike the other strain. It was surprising
that DN2 that previously showed more phagocytic uptake in macrophage had reduced attachment to the
epithelial cells relative to ∆T3. Though T6SS1-related mutants (∆T1 and ∆T1∆T3) were the least attached
bacteria to the cells, ∆T1 made a significant attachment more the that of ∆T1∆T3. The slight fimbria
expression in ∆T1 could also be aiding more attachment of the strain to the epithelial cells, unlike ∆T1∆T3.
To further investigate the trend of the bacterial attachment to epithelial cells, a monolayer of H292 was
infected with GFP-tagged bacteria and visualized using confocal microscopy. The microscopy data (Figure
5.9) further corroborated the quantitative attachment data, revealing that more ∆T3 attached to the cells
followed by DN2, then ∆T1 and ∆T1∆T3. Also, even though more ∆T3 attached to H292 cells, DN2-treated
cell showed a significant actin filament destruction more than all the T6SS mutants, however, ∆T1∆T3-
and ∆T1 treated cells showed little to no filament destruction. The confocal microscopy of the infected cells
was reliable confirmatory data that agreed with the attachment quantification data. Once again, the imaged
∆T3 attached to the cells out rated all the other T6SS mutants and the parent strain, DN2. Despite the
overwhelming attachment ∆T3 to the epithelial cells but many of the bacteria were not internalised, rather
on the surface of the cells yielding little cytoskeletal rearrangement relative to the other T6SS mutant-
treated cells (∆T1 and ∆T1∆T3) and DN2.
Interestingly, ∆T1-treated cells also yielded little bacteria invasion when compared to ∆T3 but not invaded
∆T31∆T3, hence minimal actin destruction in the cells. Objectively, no significant difference in actin
polymerisation was noted between ∆T1 and ∆T3, although ∆T3 showed more attachment and possibly
invaded more quickly relative to ∆T1. This observation was similar to the EtBr assays, noticeably, where
there was no difference in the nuclei uptake of EtBr between ∆T1 and ∆T3 but not the same for ∆T1∆T3,
all relative to DN2. These observations alongside the quantitative data have led to a hypothesis that, T6SS
is used by K. pneumoniae to attach, invade, toxicate the cell as well as mediate actin filament collapse. A
limitation in the use of these NCI-H292 was noted, the cells did not have unique actin filament across the
cytoplasm.
To understand the mechanistic virulent role of T6SS in K. pneumoniae during infection, T6SS-dependent
promotion of the host epithelial cells invasion and survival of K. pneumoniae was examined by infecting
A549 monolayer (MOI= 100) with the various K. pneumoniae strains for 1h and recovering the invaded
bacteria (2, 4, and 6 h post-infection) after gentamicin treatment to eliminate extracellular bacteria. Having

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
179
confirmed that T6SS affect the ability of K. pneumoniae HS11286 to attach to human epithelial cells, the
ability of the various strain to invade and survive in the host epithelial cells made a fascinating revelation
Figure 5.8B. From Figure 5.8B, the T6SS mutants’ invasion and survival in A549 at all the investigated
time-points were hindered but not their parent strain, DN2. The A549 invasion data revealed that, even
though the bacteria invasion of the cell was time-course dependent, DN2 invasion was more pronounced
relative to T6SS mutants. Unsurprisingly, the double T6SS mutants ∆T1∆T3, yielded the least recovered
bacteria though recovery numbers increased over the time-course. Similarly, ∆T1 and ∆T3 recoveries were
attenuated relative to their parent stain DN2 but more than ∆T1∆T3. It was however interesting to note that,
despite the high attachment of ∆T3 to A549, it could not invade and survive in the cell when compared to
DN2 at all the three time-points. Though ∆T3 invasion and survival was attenuated relative to DN2, ∆T3
recovery revealed that it could invade and survive better than the other two T6SS mutants (∆T1 and
∆T1∆T3).
Inferring from the attachment and invasion data Figure 5.8, ∆T3 yielded more attachment but less invasion
because of highly expressed surface adhesins/fimbriae (mostly type 3 fimbria) which enabled the
attachment but the associated reduced virulence due to T6SS3 deletion affected the ability of the bacteria
to survive in the A549 cells. The expressed type 3 fimbria on ∆T3 does not only enhance biofilm formation
but mediate binding to cell surface receptors on lung epithelial cell, A549, which is consistent with
previously reported studies have(Murphy et al., 2013; Stahlhut, Struve and Krogfelt, 2012; Schroll et al.,
2010; Johnson et al., 2011). Hence, the mutation of T6SS affected the surface fimbrial expression, which
in turn affected the ability of the bacteria to attach to surface and receptor. No statistically significant
difference was noted between ∆T1 and ∆T1∆T3 at 2h, but the difference became apparent with time,
particularly at 6h. ∆T1∆T3 invasion/ survival in A549 was highly attenuated followed by ∆T1 and ∆T3 in
decreasing level of attenuation, all relative to the parent strain, DN2. Both ∆T1 and ∆T1∆T3 exhibited less
invasion and survival in the A549 cell, which corroborates the attachment data from Figure 5.8A.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
180
Figure 5.9 Visualization K. pneumoniae attachment to H292 cells.
K. pneumoniae attachment to NCI-H292. An H292 monolayer on coverslips was infected with the various K.
pneumoniae strains (DN2, ∆T1, ∆T3 and ∆T1∆T3) at MOI 100 for 30 minutes and subsequently washed three times
with PBS (no antibiotics). NCI-H292 actin filament was stained with rhodamine-phalloidin, the nuclei stained with
DAPI while the bacteria were tagged green with GFP plasmid. More attachment was observed in the ∆T3 -infected
cells relative to DN2-treated cells; the reverse was true for ∆T1∆T3-treated cells. NCI-H292 experienced more
actin destruction in cell treated with DN2 followed by ∆T3 and arguably ∆T1. The above figure is representative
images taken with a scale bar of 10µm indicate at the lower right corner of each image

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
181
K. pneumoniae HS11286 has three categorised T6SS clusters, but only two of these clusters (T6SS1 and
T6SS3) are showed functional potential due to the gene content of these clusters (they contained almost all
the known functional T6SS components). But the K. pneumoniae T6SS3 lacks one of the essential
components of T6SS (hcp) although an hcp homologue found elsewhere in the genome as truncate T6SS2
may be complementing the T6SS3. Unlike T6SS3, T6SS1 has all the functional T6SS components and
contributed significantly to the overall virulence T6SS in K. pneumoniae than T6SS3 cluster. The absence
of T6SS1 gene cluster (∆T1) resulted in a significant virulence attenuation, unlike T6SS3 mutant (∆T3)
that showed subtle virulence attenuation in attachment and invasion data. The deletion of T6SS3 enhances
fimbrial expression that promotes biofilm formation, attachment and invasion to host lungs epithelial cell.
Deletion of T6SS1 or T6SS3 increased cell attachment, but the deletion of both clusters terminated virulent
attachment and invasion of K. pneumoniae to the host epithelial cells. The invasion and survival assay was
also confirmed by confocal microscopy at of infected A549 cell at two-time points (3 and 6 h) as shown in
Figure 5.11 and Figure 5.12, suggesting that T6SS is involved the promotion of attachment, invasion and
survival. Hence these data indicated that K. pneumoniae uses T6SSs to promote attachment, invasion and
survival within host lung epithelial cells and there exist a functional difference in the various T6SS clusters
in the genome.
Also, quick ethidium bromide (EtBr) assay was used to assess the viability of the H292. Briefly, the infected
monolayer was fixed and first stained with DAPI and then counter-stained with EtBr for confocal
microscopy for the presence of DAPI and/ or EtBr. In the presence of EtBr, compromised cell membrane
allows EtBr uptake, which may stain the nuclei (replacing lower molecular weight DAPI) unlike healthy
viable cells with the uncompromised cell membrane. Consequentially, higher EtBr nuclei fluorescence
indicated more EtBr uptake was attained through compromised cell membranes, while DAPI-stained nuclei
indicate viable cells. Interestingly, all the K. pneumoniae (DN2, ∆T1 and ∆T3) treated H292 cells except
that of ∆T1∆T3 showed the varying level of EtBr fluorescence. From the data, DN2 had much damaging
effect on the cells resulting in more EtBr uptake that stained the nuclei of the cells. Though ∆T1 and ∆T3
treated cells show some level of EtBr nuclei fluorescence but not as much as that of DN2-treated cells. The
EtBr uptake was attenuated in ∆T1∆T3 treated cells suggesting that ∆T1∆T3 had the least damaging effect
on the H292 cells. This was however expected as have been demonstrated extensively in the previous
chapters that, the absence of both T6SS clusters in K. pneumoniae (∆T1∆T3) resulted in T6SS1 and T6SS3
synergistic virulence attenuation towards the cells.
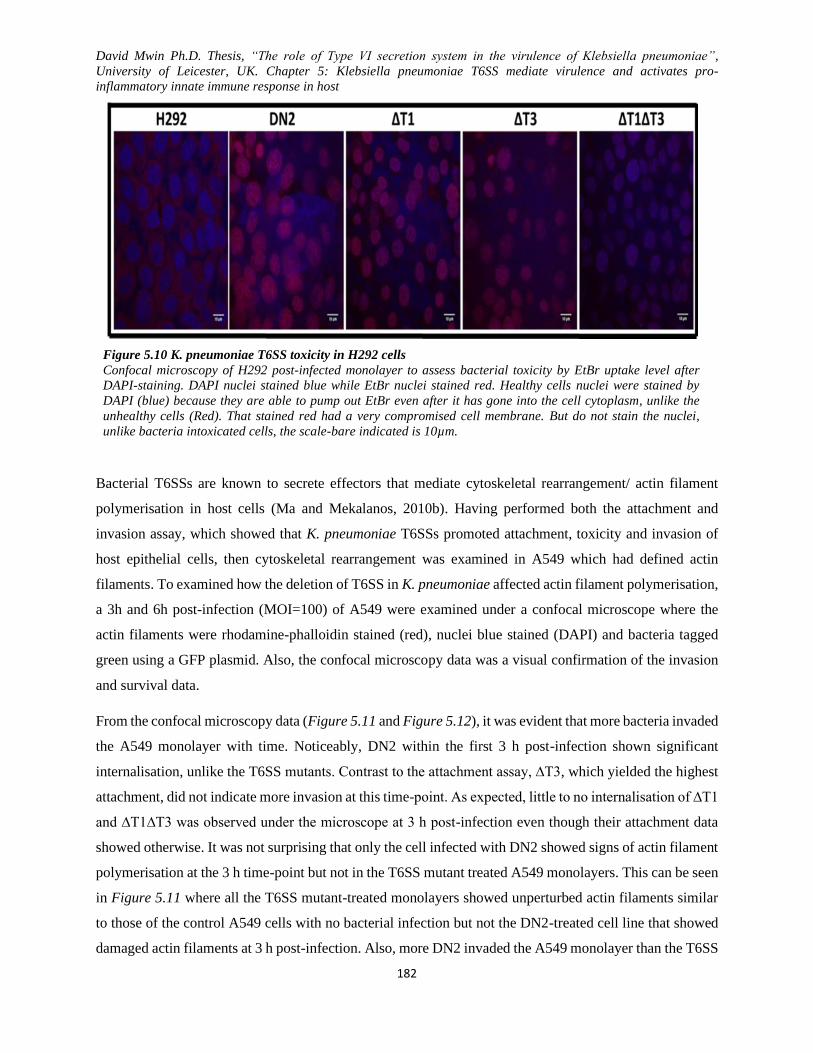
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
182
Figure 5.10 K. pneumoniae T6SS toxicity in H292 cells
Confocal microscopy of H292 post-infected monolayer to assess bacterial toxicity by EtBr uptake level after
DAPI-staining. DAPI nuclei stained blue while EtBr nuclei stained red. Healthy cells nuclei were stained by
DAPI (blue) because they are able to pump out EtBr even after it has gone into the cell cytoplasm, unlike the
unhealthy cells (Red). That stained red had a very compromised cell membrane. But do not stain the nuclei,
unlike bacteria intoxicated cells, the scale-bare indicated is 10µm.
Bacterial T6SSs are known to secrete effectors that mediate cytoskeletal rearrangement/ actin filament
polymerisation in host cells (Ma and Mekalanos, 2010b). Having performed both the attachment and
invasion assay, which showed that K. pneumoniae T6SSs promoted attachment, toxicity and invasion of
host epithelial cells, then cytoskeletal rearrangement was examined in A549 which had defined actin
filaments. To examined how the deletion of T6SS in K. pneumoniae affected actin filament polymerisation,
a 3h and 6h post-infection (MOI=100) of A549 were examined under a confocal microscope where the
actin filaments were rhodamine-phalloidin stained (red), nuclei blue stained (DAPI) and bacteria tagged
green using a GFP plasmid. Also, the confocal microscopy data was a visual confirmation of the invasion
and survival data.
From the confocal microscopy data (Figure 5.11 and Figure 5.12), it was evident that more bacteria invaded
the A549 monolayer with time. Noticeably, DN2 within the first 3 h post-infection shown significant
internalisation, unlike the T6SS mutants. Contrast to the attachment assay, ∆T3, which yielded the highest
attachment, did not indicate more invasion at this time-point. As expected, little to no internalisation of ∆T1
and ∆T1∆T3 was observed under the microscope at 3 h post-infection even though their attachment data
showed otherwise. It was not surprising that only the cell infected with DN2 showed signs of actin filament
polymerisation at the 3 h time-point but not in the T6SS mutant treated A549 monolayers. This can be seen
in Figure 5.11 where all the T6SS mutant-treated monolayers showed unperturbed actin filaments similar
to those of the control A549 cells with no bacterial infection but not the DN2-treated cell line that showed
damaged actin filaments at 3 h post-infection. Also, more DN2 invaded the A549 monolayer than the T6SS

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
183
mutants despite its lower attachment relative to ∆T3. More DN2 was noted in the epithelial cells imaged at
6 h showed better within the epithelial cells unlike The representative images obtained at 6 post-infection
were equally confirmatory of T6SS active role in the invasion and survival of K. pneumoniae in the lungs
epithelial cell relative to the T6SS mutants derivatives. Also, produced at the 6 h time point was more
damaging and disintegration of the A549 actin filaments and rounding-up of the DN2-infected cells.
Surprisingly, the nuclei that were easily stained and imaged at 3 h were poorly seen 6 h in the DN2-infected
A549 cells. Apart from the DN2 deleterious effect on the cells at 6 h, it was also noted that ∆T3-treated
cells also began rounding-up and showed collapsed actin filament as well as more invasion of the bacteria
when compared to the other T6SS mutants (∆T1 and ∆T1∆T3) treated A549 cell line. Arguably, though
few invaded ∆T1 were observed, the actin filaments were relatively conserved but not as much the
conserved actin filaments in the ∆T1∆T3-treated cells. Little to no damage was done to the ∆T1- and
∆T1∆T3-infected cells when compared to the control cells (A549 cells only) unlike the DN2-infected cells,
and ∆T3 infected cells. Comparing the images at 3 h to those of the 6 h, more bacteria invaded and
internalised at the latter time point and consequently, more cell toxicity, actin filament destruction and cell
rounding.
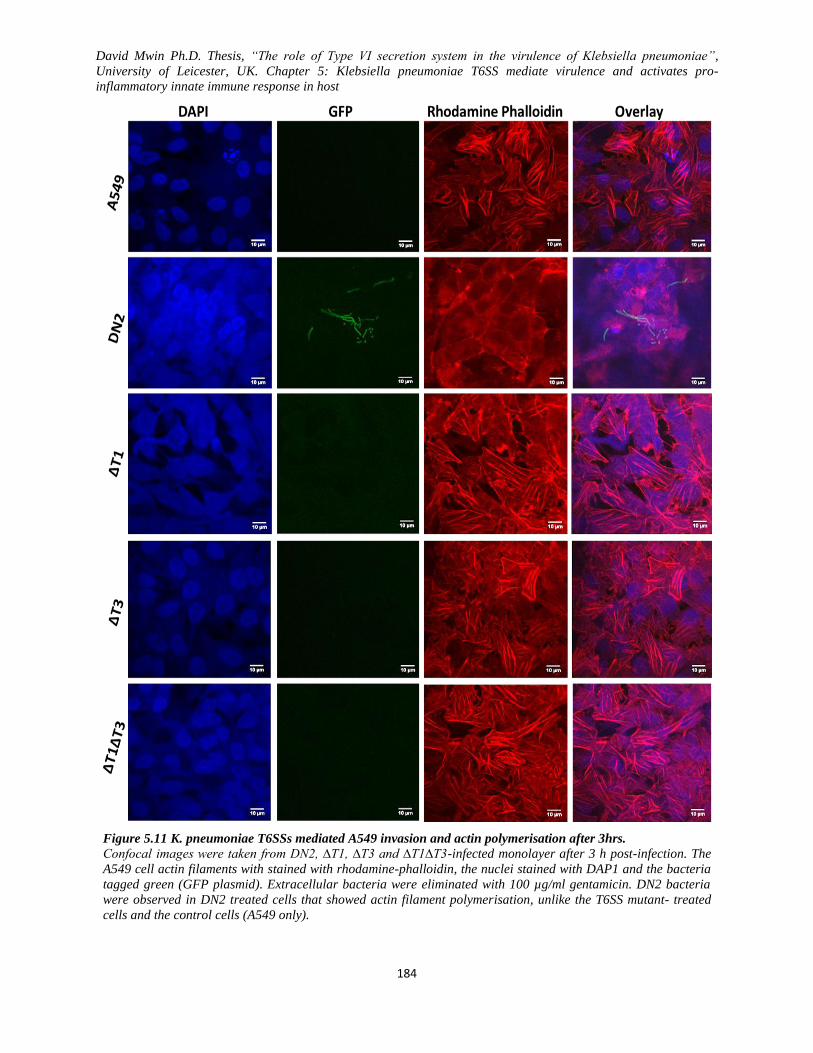
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
184
Figure 5.11 K. pneumoniae T6SSs mediated A549 invasion and actin polymerisation after 3hrs.
Confocal images were taken from DN2, ∆T1, ∆T3 and ∆T1∆T3-infected monolayer after 3 h post-infection. The
A549 cell actin filaments with stained with rhodamine-phalloidin, the nuclei stained with DAP1 and the bacteria
tagged green (GFP plasmid). Extracellular bacteria were eliminated with 100 µg/ml gentamicin. DN2 bacteria
were observed in DN2 treated cells that showed actin filament polymerisation, unlike the T6SS mutant- treated
cells and the control cells (A549 only).

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
185
Figure 5.12 K. pneumoniae A549 invasion and actin polymerisation after 6hrs.
K. pneumoniae T6SSs mediated A549 invasion and actin polymerisation after 6hrs. Confocal images were taken
from DN2, ∆T1, ∆T3 and ∆T1∆T3-infected monolayer after 6 h post-infection. The A549 cell actin filaments with
stained with rhodamine-phalloidin, the nuclei stained with DAP1 and the bacteria tagged green (GFP plasmid).
Extracellular bacteria were eliminated with 100 µg/ml gentamicin. DN2 bacteria were observed in DN2, and
∆T3 treated cells that showed even more actin filament polymerisation unlike the ∆T1 and ∆T1∆T3 iT6SS mutant-
treated cells and the control cells (A549 only).

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
186
5.4 T6SSs in K. pneumoniae trigger an inflammatory immune response in both macrophages and
epithelial cell (J774 and A549)
Many hosts (including humans) use several evolved mechanisms as a response to the presence of pathogenic
microbes, thereby preventing such pathogens from causing infections. One such common mechanism
employed by many hosts is the innate immune system, which is the primary immune system that deals with
infection. During bacterial infections, immune cells respond by secretion or induction of cytokines and
chemokines and activating pathways involved in the clearance of the pathogen. Having looked at several
assays that measured the interactive impact of the K. pneumoniae T6SS on abiotic and biotic surfaces
(attachment and invasion), a hypothesis that the innate immune system would respond differently to the
various T6SS mutants was considered. Here, two forms of binding immunoassays were used, preliminary
cytokine array/ proteome profiler to examine the crude cytokine/proteins in the cell line-bacteria induced
supernatants, and the other was the traditional ELISA. Supernatants from macrophages (J774) and epithelial
(A549) cells were taken at different time points and examined for their cytokine and chemokine response.
Briefly, R&D human/ mouse cytokine array/proteome profiler kit was initially used for the preliminary
detection of the various cytokine expression induced by K. pneumoniae T6SS, and the traditional ELISA
was later used as a confirmatory test for some of the cytokines detected earlier. The human/ mouse cytokine
array/proteome profiler kits are membrane-based sandwich immunoassay where the membranes were
incubated in a mixture of samples and cocktail of biotinylated detection antibodies for the
chemiluminescent visualisation for the captured proteins. The intensity of the signals (as duplicate dots)
produced by each cytokine/chemokine on the membrane was proportional to the amount of the
cytokine/chemokine present in the analytes. All supernatants used were filtered to get rid of all bacteria and
cells debris. Captured cytokines on the membranes were observed as duplicates and the final images
obtained were analysed with ImageJ software for the average pixel intensities represented in graphs (Figure
5.13 and Figure 5.15A). Filtered supernatants meant for ELISA were stored in -80OC until required for the
experiment.
To perform the traditional Sandwich ELISA, 96-well plates were pre-coated with the captured antibody
for overnight at room temperature. The plates were then washed, blocked and rewashed before adding the
samples/ standards to immobilize the cytokines while the unbound materials were washed away. The
primary antibodies were added to bind the cytokines, and then an enzyme-linked secondary/ HRP-
conjugated antibodies were applied for detection. The substrates for colour development was added, and
the absorbance of the wells was measured after being stopped.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
187
5.4.1 Expressed macrophage (J774) inflammatory mediators/ cytokines induced in
response to K. pneumoniae infection
Macrophages are an essential part of the innate immune cells that act as the first line of defence against
pathogens though it plays other roles as well, including homeostasis, coordinate adaptive immune response,
inflammation and repair innate immune system (Adams and Hamilton, 1984; Wynn, Chawla and Pollard,
2013; Burke and Lewis, 2002). Macrophage defends the body against invading microbes (bacteria, viruses,
fungi and protozoa) by phagocytosis, pathogen destruction, antigen presentation and secretion of cytokine/
chemokine, enzymes, complement, enzyme inhibitors, chemokine the invading pathogen (Burke and Lewis,
2002).
Interestingly, both data obtained from the semi-quantitative mouse cytokine array/ proteome profiler kit
(Figure 5.13) and the traditional ELISA kits (Figure 5.14) detected relatively similar J774 cytokines
expressed in the culture supernatants. The mouse cytokine array/ proteome profiler kit was used to
determine a broad range of cytokines/ transduction signal mediators that were secreted into the J774 culture
supernatant upon exposure to the various K. pneumoniae strains (DN2, ∆T1, ∆T3 and ∆T1∆T3). Twelve
different cytokine/ chemokine (IL-1a, IL-1ra, MCP-1, MIP-1a, MIP-1b, MIP-2, IP-10, GM-CSF, M-CSF,
TNF-a, RANTES and SDF-1), mostly inflammatory mediators were noted to be induced in the infected
J774 supernatant. However, for confirmatory purposes, the traditional ELISA was used to measure TNF-a,
IL-6, RANTES, IL1a, IP-10, MIP-1a, MIP-1b and MCP-1 in bacteria-treated J774 cell supernatant.
From the preliminary cytokine array data in Figure 5.13, almost the same amount of-of IL-1a, IL-1ra, MIP-
1a, and RANTES was induced by DN2-infected macrophages relative to those expressed by the T6SS
mutant treated cell supernatant. Additionally, DN2-infected macrophages secreted a relatively low amount
of IP-10, MCP-1, MIP-1b and MIP-2 relative to the T6SS mutant-infected macrophages. In contrast, much
lower levels of GM-CSF, M-CSF, SDF-1 and TNF-a were expressed by ∆T1∆T3-infected macrophages
relative to the other treatment groups. Generally, variable amounts of T6SSS-dependent induced
inflammatory mediators were preliminarily detected in the supernatant of the infected macrophages using
the mouse cytokine array/proteome profile.
Granulocyte-macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor
(M-CSF) are monomeric glycoprotein-based cytokine secreted by macrophages, T cells, mast cells, NK
cells, endothelial cells and fibroblast (Francisco-Cruz et al., 2014; Metcalf and Nicola, 1995; Ushach and
Zlotnik, 2016). They play an essential role including; acting on WBC and stimulate stem cells to produce
granulocytes and monocytes, involved in inflammatory cascade by activating and recruiting cells,
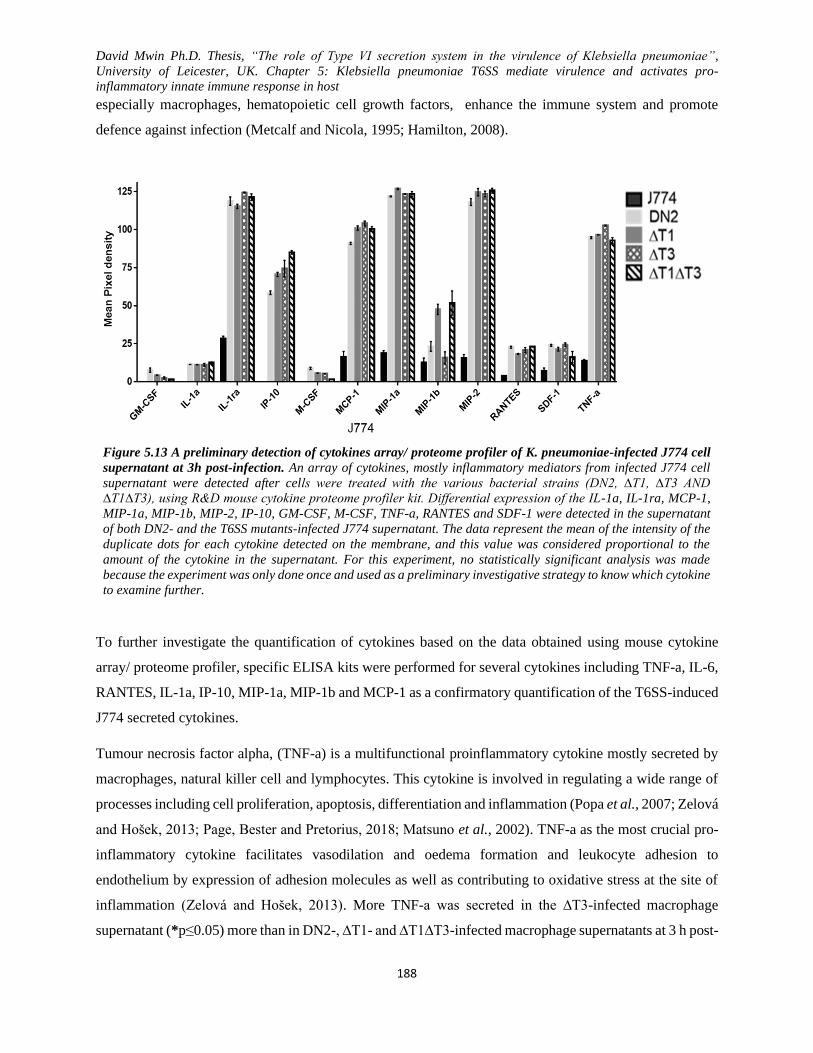
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
188
especially macrophages, hematopoietic cell growth factors, enhance the immune system and promote
defence against infection (Metcalf and Nicola, 1995; Hamilton, 2008).
Figure 5.13 A preliminary detection of cytokines array/ proteome profiler of K. pneumoniae-infected J774 cell
supernatant at 3h post-infection. An array of cytokines, mostly inflammatory mediators from infected J774 cell
supernatant were detected after cells were treated with the various bacterial strains (DN2, ∆T1, ∆T3 AND
∆T1∆T3), using R&D mouse cytokine proteome profiler kit. Differential expression of the IL-1a, IL-1ra, MCP-1,
MIP-1a, MIP-1b, MIP-2, IP-10, GM-CSF, M-CSF, TNF-a, RANTES and SDF-1 were detected in the supernatant
of both DN2- and the T6SS mutants-infected J774 supernatant. The data represent the mean of the intensity of the
duplicate dots for each cytokine detected on the membrane, and this value was considered proportional to the
amount of the cytokine in the supernatant. For this experiment, no statistically significant analysis was made
because the experiment was only done once and used as a preliminary investigative strategy to know which cytokine
to examine further.
To further investigate the quantification of cytokines based on the data obtained using mouse cytokine
array/ proteome profiler, specific ELISA kits were performed for several cytokines including TNF-a, IL-6,
RANTES, IL-1a, IP-10, MIP-1a, MIP-1b and MCP-1 as a confirmatory quantification of the T6SS-induced
J774 secreted cytokines.
Tumour necrosis factor alpha, (TNF-a) is a multifunctional proinflammatory cytokine mostly secreted by
macrophages, natural killer cell and lymphocytes. This cytokine is involved in regulating a wide range of
processes including cell proliferation, apoptosis, differentiation and inflammation (Popa et al., 2007; Zelová
and Hošek, 2013; Page, Bester and Pretorius, 2018; Matsuno et al., 2002). TNF-a as the most crucial pro-
inflammatory cytokine facilitates vasodilation and oedema formation and leukocyte adhesion to
endothelium by expression of adhesion molecules as well as contributing to oxidative stress at the site of
inflammation (Zelová and Hošek, 2013). More TNF-a was secreted in the ∆T3-infected macrophage
supernatant (*p≤0.05) more than in DN2-, ∆T1- and ∆T1∆T3-infected macrophage supernatants at 3 h post-

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
189
infection but not at 6 h, (Figure 5.14A). However, at 6h, DN2 strain induced more macrophage secretion
of TNF-a than induced by ∆T1 (****p≤0.0001) and ∆T1∆T3 (****p≤0.0001) but not ∆T3 (ns p>0.05) even
though ∆T3 (**p≤0.01) induction of TNF-a at 6h was significantly more than that of ∆T1 and ∆T1∆T3
(Figure 5.14A).
Interleukin-6 (IL-6), is a soluble mediator with pleiotropic effect on inflammatory, immunity response and
haematopoiesis (Akira, Taga and Kishimoto, 1993). IL-6 is often produced in response to bacterial infection
and injuries to the host immune defence and pose a dual effect; as pro-inflammatory and anti-inflammatory
role depending on the condition (acts as a defence mechanism and proinflammatory effect in chronic
inflammation) (Gabay, 2006; Scheller et al., 2011; Tanaka, Narazaki and Kishimoto, 2014). For IL-6
(Figure 5.14B), significantly more amount was detected at in ∆T3-infected macrophage supernatant
(*p≤0.05) relative to DN2 at 3h and even more IL-6 was induced at 6h by ∆T3 and DN2 unlike ∆T1 and
∆T1∆T3.
Regulated (increased production) upon Activation, expression of Normal T- cElls (RANTES/ CCL5) is a
hematopoietic and non-hematopoietic cell type chemokine that plays a vital role in homing and migration
of effector and memory T cells during acute infection (Alam et al., 1993; Hopp, 1994; Levy, 2009).
RANTES previous thought to be specific to T cells is now reported to be secreted by many cell types (such
as macrophages, platelets, eosinophil, fibroblast, endothelial, epithelial and endometrial cells). RANTES
has been reported to exhibit direct antimicrobial activity and beneficial chemotactic activity of bringing
immune cells (T cells, dendritic cells, eosinophils, NK cells, mast cell and basophils) to the site of
inflammation and infection (Appay and Rowland-Jones, 2001; Levy, 2009; Villalta et al., 1998). It can,
however, exhibit deleterious effect when it recruits immune cells that enhance the inflammatory process in
arthritis, nephritis, colitis, arteriosclerosis, asthma and endometriosis among other disorders (Appay and
Rowland-Jones, 2001; Levy, 2009; Rossi and Zlotnik, 2000). RANTES (Figure 5.14C) was significantly
induced at both 3, and 6-h postinfection in DN2-treated macrophage supernatant relative to all the T6SS
mutant treated macrophage supernatant (*p≤0.05). No significant difference in RANTES (ns p>0.05) was
detected among the T6SS mutant infected macrophage supernatants at 6h but not at 3h (where ∆T3 induced
more RANTES relative to ∆T1).
ELISA for other cytokines, IL-1a, IP-10, MIP1a, MIP-1b and MCP-1, was only performed for the infected
macrophage supernatant at 3h post-infection only.
Interleukin 1 alpha (L-1a) is a known cytokine mainly produced by activated macrophages, epithelial
neutrophil and endothelial cells responsible for the production of inflammation, fever and sepsis (Di Paolo
and Shayakhmetov, 2016; Dube et al., 2001; Dinarello, 2011). It’s on the same pathway that activates the

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
190
TNF-a and been reported to induced IL-8 during infection (Brito et al., 1999; Cheng et al., 2008; Di Paolo
et al., 2015; Oppenheim et al., 1989). IL-1a is constitutively expressed by epithelial cell and in epidermal
keratinocyte, playing an essential role in maintaining the skin barrier fidelity to prevent the entry of
pathogens. While no significant difference in IL-1a was noted between DN2- and ∆T3-infected macrophage
supernatant at 3h, DN2 or ∆T3 significantly induced macrophage expression of IL-1a relative to ∆T1 and
∆T1∆T3 or ∆T1∆T3 infected macrophage supernatants at 3h, respectively (Figure 5.14D).
Interferon-gamma-induced protein 10 (IP-10/ CXCL10) is a chemokine is secreted by a variety of cell types
(monocytes, fibroblast and endothelial cells) in response to interferon (IFN-y) and inducing chemoattraction
of monocytes/macrophages, T cell, NK cells and dendritic cells, facilitating the attachment
of T cell to endothelial cells, among many others (Dufour et al., 2002; Liu et al., 2011; Gotsch et al., 2007;
Angiolillo et al., 1995). Also, all the T6SS mutants (∆T1, ∆T3 and ∆T1∆T3) significantly induced more
IP-10 in the infected macrophage supernatant than induced by their parent strain, DN2 (**p≤0.01) at 3h
(Figure 5.14D). Additionally, ∆T1∆T3 expressed more IP-10 more than ∆T1 (**p≤0.01) but not ∆T3 (ns
p>0.05) at 3h.
Macrophage inflammatory protein (MIP-1) is known chemokine secreted by macrophages when they are
stimulated by bacterial endotoxins (Sherry et al., 1988). MIP-1 exist as MIP-1a and MIP-1b and are
involved in the inflammatory process of the innate immunity, and the recruitment and activation of
polymorphonuclear leukocytes, NK cells, monocytes and other immune cells (Bystry et al., 2001; Wolpe
et al., 1988). MIP-1a and MIP-1b are a significant immune response to infection and inflammation as well
as synthesizing and release of other pro-inflammatory cytokines (IL-1, IL-6 and TNF) from fibroblast and
macrophages (Ren et al., 2010; Maurer and Von Stebut, 2004). At the 3h post-infection, ∆T1 and ∆T1∆T3
induced high expression of MIP-1a and MIP-1b (Figure 5.14E) relative to ∆T3 and DN2 infected
macrophage supernatant at 3h. While no significant difference between DN2 and ∆T3 was observed for the
induced expression of MIP-1a, DN2 induced the substantial expression of MIP-1b than induced by ∆T3.
No difference was noted between ∆T1 and ∆T1∆T3 induced expression of MIP-1a and MIP-1b at 3h (see
Figure 5.14E).
Monocytes chemoattractant protein 1 and 2 (MCP-1 and MCP-2 ) are cytokines involved in the initiation,
regulation and recruitment of monocytes, granulocytes, NK cell, memory T cells and dendritic cells to the
inflammatory sites during injury/ infection (Carr et al., 1994; Xu et al., 1996; Proost, Wuyts and Van
Damme, 1996; Gong et al., 1998). Finally, the infected macrophage was induced to express more MCP-1
(Figure 5.14E) in their supernatant by ∆T3 (***P≤0.001) than induced DN2, ∆T1, and ∆T1∆T3 at 3h. Also,

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
191
DN2 induce the least amount of macrophage-expressed MCP-1 in the culture supernatant relative to ∆T1
(****p≤0.0001), and ∆T1∆T3 (****p≤0.0001) at 3h.
In summary; T6SS mutants induced more expression of IP-10, MIP-1a, MIP-1b and MCP-1 except for IL-
1a, DN2- and ∆T1∆T3 -infected macrophage showed the highest and least expressions of IL-a respectively
and ∆T3 induced more expression of IL-1a than induced by ∆T1. Also, it was not surprising that ∆T1- and
∆T1∆T3- treated cells yielded the most expression of MIP-1a and MIP-b compared to DN2 and ∆T3 -
treated cells. However, ∆T3 -treated cell expressed the least MIP-1a and MIP-1b compared to the rest of
the treatment. Increased expression of IP-10 was induced T6SS mutants (∆T1∆T3, ∆T3 and ∆T1 in
decreasing manner) was observed when compared to that the parent strain DN2. More distinctively, ∆T3 -
treated cell expressed the highest level of MCP-1 in the supernatant than DN2-, ∆T1- and ∆T1∆T3- treated
macrophage supernatants. DN2 significantly inhibited the expression of MCP-1 compared to ∆T1, ∆T3 and
∆T1∆T3 -treated cells. Notably, DN2 carrying both active T6SS clusters induced more expression of TNF-
a, IL-6, RANTES and IL-1a (at 6h) than induced by the T6SS mutants. Also, while ∆T3 induced more
expression of MCP-1, ∆T1 and ∆T1∆T3 induced more expression of MIP-1a and MIP-1b; and macrophage
IP-10 was expressed more in all the T6SS mutants infected supernatant.
Generally, K. pneumoniae, in a T6SS-dependent manner, trigger a variable expression of inflammatory
signal transduction mediators in macrophages (Figure 5.14). On a broad scale, DN2 treatment inhibited of
increased expression of the inflammatory cytokines through a yet-to-be-explored mechanism as an evasive
strategy to prevent the recruitment of immune cells and factors to the area of infection.
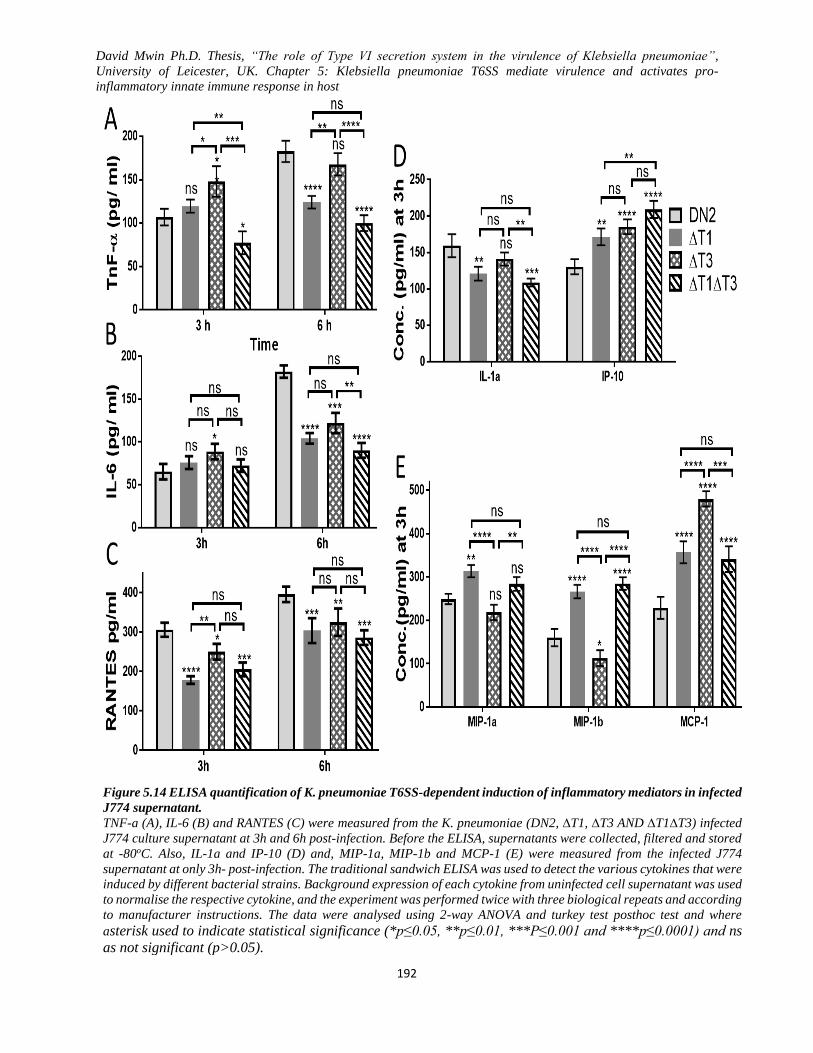
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence and activates pro-
inflammatory innate immune response in host
192
Figure 5.14 ELISA quantification of K. pneumoniae T6SS-dependent induction of inflammatory mediators in infected
J774 supernatant.
TNF-a (A), IL-6 (B) and RANTES (C) were measured from the K. pneumoniae (DN2, ∆T1, ∆T3 AND ∆T1∆T3) infected
J774 culture supernatant at 3h and 6h post-infection. Before the ELISA, supernatants were collected, filtered and stored
at -80oC. Also, IL-1a and IP-10 (D) and, MIP-1a, MIP-1b and MCP-1 (E) were measured from the infected J774
supernatant at only 3h- post-infection. The traditional sandwich ELISA was used to detect the various cytokines that were
induced by different bacterial strains. Background expression of each cytokine from uninfected cell supernatant was used
to normalise the respective cytokine, and the experiment was performed twice with three biological repeats and according
to manufacturer instructions. The data were analysed using 2-way ANOVA and turkey test posthoc test and where
asterisk used to indicate statistical significance (*p≤0.05, **p≤0.01, ***P≤0.001 and ****p≤0.0001) and ns
as not significant (p>0.05).

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence in host and activates host pro-
inflammatory innate immune response
193
5.4.2 Expressed epithelial (A549) inflammatory mediators/ cytokines stimulated in
response to K. pneumoniae
Human cytokine array/ proteome profile was used to preliminarily detected the various A549 cytokines
expressed in the supernatant upon induction by K. pneumoniae. Briefly, the membrane with the duplicates
captured cytokines antibodies were incubated with the supernatants of DN2, ∆T1, ∆T3 and ∆T1∆T3
infected epithelial cells at 3 h post-infection. Using the detection antibodies, the cytokines were detected at
duplicate black dots on the membrane, and their average pixel intensities were obtained using ImageJ. Five
different inflammatory cytokines (IL-8, MIF, PAI-1, sICAM-1, and GROa) were detected using the human
cytokine array/ proteome profiler. The data (Figure 5.15A) revealed that while DN2 stimulated the highest
expression of GROa and MIF, ∆T1∆T3 induced the least expression of GROa and MIF in A549 cells.
Additionally, A549 cells that were infected with ∆T3 showed increased expression of IL-8 and PAI-1
relative to DN2, ∆T1 and ∆T1∆T3. Similarly, ∆T1 and ∆T3 infected epithelial cells showed increased
expression of sCAM-1 relative to DN2 and ∆T1∆T3. Among five induced A549 cytokines, ∆T1∆T3-treated
A549 cell showed the least expression of IL-8, MIF, PAI-1, sICAM-1, and GROa while DN2-treated cell
supernatant expressed higher in MIF and GROa but not PAI-1. This experiment was only performed once
as a preliminary screening strategy for target ELISA quantification. Thus, no statistical analysis was
performed to determine the significant difference among the various cytokines.
Soluble intracellular adhesion molecule-1 (sICAM-1) are cell adhesion molecules representing the
circulating form of ICAM constitutively expressed or inducible on many cell lines surface as an important
inflammatory biomarker for activation and destruction of the endothelium(Mendez et al., 2011; Witkowska
and Borawska, 2004). It serves as a counter-receptor for lymphocytes function-associated antigen (LFA-1)
and the interaction between endothelial ICAM-1 and LFA-1 promote leukocyte attachment and migration
through the endothelium (Witkowska and Borawska, 2004; Meyer, Dustin and Carron, 1995; Abe et al.,
1998). DN2- and ∆T1∆T3-treated cells supernatant yielded lesser sICAM-1 when compared to ∆T1- and
∆T3-treated cell supernatant
To quantify the elicited cytokines induced in the epithelial cell line, ELISA was performed for some of the
A549 inflammatory mediators (IL-8, GROa and MIF) induced by K. pneumoniae. DN2- treated cell
expressed more GROa and MIF compared to the other treatments but not IL-8. ∆T1∆T3-treated A549 cells
secreted the least amount the assessed inflammatory mediators (IL-8, GROa and MIF) followed by ∆T1 for
IL-8 and GROa mediators but not MIF. Unlike in the quantified of MIF mediator quantification in A549,
∆T3 induced more IL-8 and GROa production compared to the other T6SS mutants. Among the T6SS

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence in host and activates host pro-
inflammatory innate immune response
194
mutants, ∆T3 induced higher expression of IL-8 and GROa but not MIF, which showed increased
expression in ∆T1-treated cells compared to the other T6SS mutants.
IL-8 (also known as CXCL8) is an essential inflammatory chemokine of the innate immune response known
to mediate chemotaxis of neutrophils and other granulocytes to migrate to the area of infection as well as
stimulating phagocytosis (Harada et al., 1994; Köhidai and Csaba, 1998). It is widely secreted by
macrophages and epithelial. After examining the supernatant for IL-8 using ELISA kit, it was realised that
∆T3-infected A549 cells expressed more IL-8 in the supernatant than induced by DN2 (*p=0.019), ∆T1
(***p=0.0002) and ∆T1∆T3 (***p≤0.0001) infection of d epithelial cell. While ∆T1 and ∆T1∆T3
significantly induced different amounts of IL-8 (*p=0.0239), no difference was noted between DN2 and
∆T1 induced expression of IL-8 (ns p=0.2528). The drastic induced expression of IL-8 by ∆T3 maybe
because of the high expression of surface fimbriae and combination with the bacteria native virulence
factors
Growth-regulated alpha protein (GROa), has chemotactic neutrophils and plays a role in inflammation and
exerts its effect on endothelial cells (Son et al., 2007; Cubitt, Lausch and Oakes, 1997). More GROa was
secreted by DN2-treated A549 cells and followed by ∆T3-treated A549. The least amount of GROa was
secreted by ∆T1∆T3-treated A549 relative to all the other bacterial strain.
Macrophage migration inhibitory factor (MIF) is an important inflammatory cytokine that is involved in
the regulation of innate immunity by binding to CD70 on other immune cells to trigger an acute immune
response (Calandra and Roger, 2003; Günther et al., 2018). It is mostly realised into the blood when
bacterial antigen stimulates leukocytes. It is involved in cell mediated-immunity and inflammation as well
as suppress the anti-inflammatory effect of glucocorticoids to regulate macrophage function in the host
(Larson and Horak, 2006; Lue et al., 2002). The preliminary data indicated high expression of MIF for
DN2-treated cell supernatant when compared to the T6SS mutant. Among the T6SS mutant treated cell
supernatant, ∆T1, ∆T3 and ∆T1∆T3 expressed a decreasing amount of MIF, respectively.
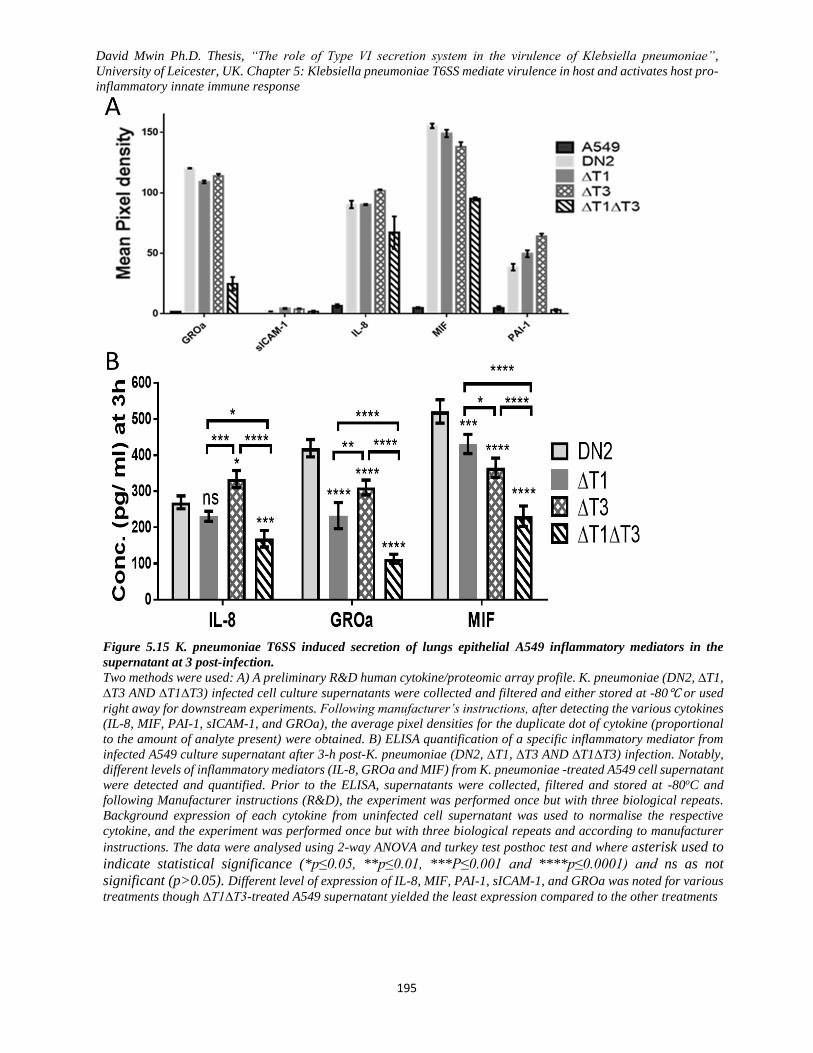
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence in host and activates host pro-
inflammatory innate immune response
195
Figure 5.15 K. pneumoniae T6SS induced secretion of lungs epithelial A549 inflammatory mediators in the
supernatant at 3 post-infection.
Two methods were used: A) A preliminary R&D human cytokine/proteomic array profile. K. pneumoniae (DN2, ∆T1,
∆T3 AND ∆T1∆T3) infected cell culture supernatants were collected and filtered and either stored at -80℃ or used
right away for downstream experiments. Following manufacturer’s instructions, after detecting the various cytokines
(IL-8, MIF, PAI-1, sICAM-1, and GROa), the average pixel densities for the duplicate dot of cytokine (proportional
to the amount of analyte present) were obtained. B) ELISA quantification of a specific inflammatory mediator from
infected A549 culture supernatant after 3-h post-K. pneumoniae (DN2, ∆T1, ∆T3 AND ∆T1∆T3) infection. Notably,
different levels of inflammatory mediators (IL-8, GROa and MIF) from K. pneumoniae -treated A549 cell supernatant
were detected and quantified. Prior to the ELISA, supernatants were collected, filtered and stored at -80oC and
following Manufacturer instructions (R&D), the experiment was performed once but with three biological repeats.
Background expression of each cytokine from uninfected cell supernatant was used to normalise the respective
cytokine, and the experiment was performed once but with three biological repeats and according to manufacturer
instructions. The data were analysed using 2-way ANOVA and turkey test posthoc test and where asterisk used to
indicate statistical significance (*p≤0.05, **p≤0.01, ***P≤0.001 and ****p≤0.0001) and ns as not
significant (p>0.05). Different level of expression of IL-8, MIF, PAI-1, sICAM-1, and GROa was noted for various
treatments though ∆T1∆T3-treated A549 supernatant yielded the least expression compared to the other treatments

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence in host and activates host pro-
inflammatory innate immune response
196
The result obtained from these experiments suggests that K. pneumoniae was able to induce an innate
inflammatory response in both host A549 human epithelial cell and J774 mouse cells in a T6SS-dependent
manner. The data suggest that K. pneumoniae T6SS via an unknown mechanism, stimulates an
inflammatory response in the host cells. More so, the K. pneumoniae parent strain, DN2 and sometimes
∆T3 was found to induced higher expression of the inflammatory immune proteins (TNF-a, IL-6, RANTES
and IL-1a) unlike ∆T1 and ∆T1∆T3 (IP-10, MCP-1 and MIP-1) infected host macrophages. Also, the
epithelial cells infected with DN2 expressed more IL-8, GROa and MIF than induced by ∆T1∆T3 or ∆T1.
Except for IP-10 from infected macrophage, and IL-8, GROa and MIF from infected A549, there was no
significant difference between ∆T1 or ∆T1∆T3 treatment groups was observed. T6SS3 mutants (∆T3),
though induced more cytokines expression than ∆T1 and ∆T1∆T3, attenuated cytokine expression was
induced by ∆T3 relative DN2.
5.5 J774 inflammatory mediators induced by hyper mucoid K. pneumoniae NTUH-K2044 T6SS
Despite the inability to create the T6SS3 mutant in K. pneumoniae NTUH-K2044, the T6SS1 mutant
(NTUH∆T1) and its parent strain (KR1730) were used in the innate immune response mediator
immunoassay was performed using the parent strain (KR1730) and its T6SS1 mutant in J774 cell line
infection. Both the supernatant and the cell lysate were analysed for the expression of cytokines. However,
the results obtained were interestingly similar to the J774 inflammatory mediators induced by DN2 and its
T6SS derivatives. Generally, all the NTUH∆T1 induced inflammatory mediators were highly expressed in
the cell supernatant, while the KR1730 induced inflammatory mediators were significantly in the cell lysate.
The J774 induced inflammatory mediators from the cell supernatant included IL-ra, IP-10, MCP-1, MIP1a,
MIP2, SDF-1 and TNF-a while that from the cell lysate included sICAM-1, IL-a, IL-ra, MCP-1, MIP1a,
MIP2, SDF-1 and TNF-a.
No significant difference was noted for the induced IL-1a and TNF-a from the cell lysate but not from the
cell supernatant. NTUH∆T1 induced cell supernatant showed the presence of MCP-1, MIP-1a, SDF-1 and
TNF-a just like IL-area that was only present in KR1730 induced cell lysate. Comparatively, the little
expression of IL-1ra, IP-10 and MIP-2 were detected in KR1730 induced cell supernatant unlike NTUH∆T1
induced cell supernatant. Also, sICAM-1, MIP-1a, MIP-2 and SDF-1.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence in host and activates host pro-
inflammatory innate immune response
197
Figure 5.16 Inflammatory mediators induced by hyper mucoid K. pneumoniae NTUH-K2044.
A) K. pneumoniae (KR1730 and NTUH∆T1) induced J774 innate proinflammatory mediators in; A) culture
supernatant and B) cell lysate after infection in the supernatant at 3h post-infection. K. pneumoniae (KR1730
and NTUH∆T1) infected cell culture supernatants were collected and filtered and either stored at -80℃ for
downstream experiments or used right away mouse cytokine/proteomic array profile. Following
manufacturer’s instructions, after detecting the various cytokines (IL-1ra, IP-10, MCP-1, MIP-1a MIP-2, SDF-
1 & TNF-a for supernatant and, sCAM-1, IL-1a, IL-1ra, MIP-1a MIP-2, SDF-1 & TNF-a FOR cell lysates),
the average pixel densities for the duplicate dot of cytokine (proportional to the amount of analyte present)
were obtained.
From the hyper mucoid K. pneumoniae NTUH-K2044 induction, NTUH∆T1 induced higher expression of
inflammatory mediators in the cell supernatant but not in the cell lysate while KR1730 induced
inflammatory mediators were more expressed in the cell lysate unlike in the cell supernatant. DN2 and its
mutants distinctly demonstrated variable inflammatory mediator expressions in both hosts epithelial cell
(human lungs A549) and macrophages (mouse J774). The presence or absence of T6SS in K. pneumoniae
influenced the host cell immune response. Hence these results suggest that T6SSs plays a significant role
in the activation of host innate immune systems

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 5: Klebsiella pneumoniae T6SS mediate virulence in host and activates host pro-
inflammatory innate immune response
198

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
199
Chapter Six
General Discussion

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
200
Chapter 6: General Discussion
The natural environment, plant and animal hosts provide a home to diverse pathogenic and non-pathogenic
microbial species (Fuqua et al., 2010). Not only are resources limited in such ecological important niche
but many of the microbes are faced with unfavourable environmental factors where only the fittest or those
that are well adapted survive (Fuqua et al., 2010; Feldman et al., 2002; Wloch-Salamon et al., 2008). Most
well study populations are either driven by a transition (competitive forces) or a balanced population. K.
pneumoniae survival and subsequent colonization in a mixed microbial niche depend on how successful it
favourably competes with the diverse normal flora and other pathogenic bacteria species for the niche’s
limited resources in the face of all the opposing forces posed by other competing bacteria species.
Competition within K. pneumoniae (intraspecies competition) and among diverse bacterial species (inter-
species competition) including those with or without T6SS immunity is key to microbial survival and
colonization.
T6SS is a transmembrane spring-loaded nano-molecular dagger expressed by a quarter of Gram-negative
bacteria for diverse use ranging from competition and antimicrobial to virulence towards host cells (Dong
et al., 2013a; Schwarz et al., 2010b; Hood et al., 2010b; Miyata et al., 2011; Mulder, Cooper and Coombes,
2012b; Bönemann, Pietrosiuk and Mogk, 2010; MacIntyre et al., 2010b). Earlier study bioinformatically
identified putative T6SS clusters in K. pneumoniae by comparative sequenced genome analysis. However,
the role of these putative T6SS clusters in K. pneumoniae were unknown. In the natural environment, K.
pneumoniae is faced with many environmental or host obliterating factor if it must survive to colonize or
cooperate in a polymicrobial community. With the recent discovery of T6SS in most Gram-negative
bacteria and the rise of carbapenem-resistant K. pneumoniae (CRK) related infection threatening last
medical antibiotic resort, the opportunistic pathogens keep evolving in many virulent factors which are yet
to be understood.
Hence, the purpose of this research was to explore the role of these putative T6SS clusters in K. pneumoniae
using less antibiotic-resistant K. pneumoniae NTUH-K2044 and a multidrug-resistant K. pneumoniae
HS11286 as model strains. Unfortunately, as at the time of this research, the lab had limited molecular tools
that could be used to explore the MDR K. pneumoniae HS11286 hence new molecular tools were built to
facilitate the easy genetic manipulation of the Gram-negative multidrug-resistant strains especially, K.
pneumoniae, E. coli and A. baumannii. MDR strains. Briefly, chapter three of this research work describe
the various constructed molecular tools, and the novel gene swapping technique that was used in the
construction of the molecular tools, and the role of T6SS in biofilm, fimbriae and competition was

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
201
demonstrated in chapter four while chapter five elucidated the role of T6SS in posing virulence towards
host cells.
Presented in chapter three is a novel DNA swapping technique (Figure 3.1) that was used mainly in the
rapid and efficient construction of a library of unique molecular tools (pDNTOOL) for exploring T6SS
virulent role in K. pneumoniae. This strategy involved the use of a sucrose counterselection marker, sacB
together with an antibiotic resistance gene (such as cat for chloramphenicol resistance) as a gene knockout
selection marker to first replace the target region via lambda red recombination or traditional cloning
method. Then the intermediate sacB-cat plasmid-bearing strain was then selected, and PCR verified. A
library of plasmids with different resistance markers or GFP were then created by transforming an
arabinose-induced E. coli strain carrying both sacB-cat intermediate plasmid and a lambda red plasmid with
different SOE-PCR products and simultaneously selecting on LA sucrose plate with or without antibiotics
as required. During this allelic exchange, the lambda red recombination used 30-100 bp as flanking up and
downstream target sequence to replace sacB-cat with the desired SOE-PCR product. As in most cases, the
intermediate sacB-cat bearing plasmid exists in multiple copies in the E. coli, and not all these copies are
mutated. To eliminate these residual/ unmutated sacB-cat bearing plasmids, the transformants were selected
on LA sucrose and antibiotics at 37-42℃, this caused sacB to express lethal levansucrase as a counter-
selection marker resulting in the loss of the all the sacB-cat bearing plasmids. Also, by growing
transformant at 37-42℃ resulted in the loss of the temperature-sensitive lambda red plasmid. Thus, the final
desired constructed plasmid was maintained in the E. coli with antibiotic selection while both the
temperature-sensitive lambda red plasmid and residual sacB-cat bearing plasmid were lost simultaneously.
The uniqueness of this novel strategy was not only in the creation of various tools that were used for T6SS
related downstream application but how rapid and efficient this method facilitated the construction of the
pNDTOOOLs without any false-positive results (Figure 3.2 and Figure 3.3). Surprisingly, after the
transformation of SOE-PCR product into the E. coli strain carrying both the intermediate sacB-cat and
lambda red plasmids (on LA sucrose plus antibiotic selection), only the desired plasmid was obtained
without any residual sacB-cat or lambda, indicating that was near This suggested that this method is near
perfect in the generation of genetic tool library. More importantly, this method avoided the cost associated
oligo primers and restriction enzymes, technical issue and time associated with standard cloning technique.
Generating a genetic tool library using standard cloning technique is not only time consuming but very
costly and technically challenging. Hence this novel strategy offers a new approach of rapidly generating a
diverse library of molecular genetic tools while mitigating all the issues associated with traditional cloning
method. Based on the demonstrated efficiency and results of this novel DNA swapping method, the
technique could be further optimized for marker-less chromosomal gene complementation.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
202
Using the novel DNA swapping strategy and standard cloning method (occasionally), an array of genetic
tools was constructed for downstream use or increase the antibiotic selection option of various necessary
plasmid available in Lab 212 archive. The MDR K. pneumoniae HS11286 genetic manipulation offered the
use of limited antibiotic-resistant markers. Hence the diversification and increase of antibiotic-resistant
variants of such suitable plasmids were necessary to aid gene knockout and DNA manipulation in
multidrug-resistant Gram-negative bacteria strains including K. pneumoniae HS11286. Notably, several
antibiotic resistance derivatives of lambda red plasmids (pDNTOOL-5, pDNTOOL-10, pDNTOOL-11,
pDNTOOL-12, pDNTOOL-13, pDNTOOL-14, pDNTOOL-15, pDNTOOL-16 and pDNTOOL-17) were
constructed using pKOBEG-Apra (expressing apramycin resistance) and pKD46 (ampicillin resistance) the
backbone plasmid. Several FRT-flanked antibiotic-resistant markers (pDNTOOL-1, pDNTOOL-32,
pDNTOOL-33, pDNTOOL-34, pDNTOOL-35, pDNTOOL-35, pDNTOOL-37, pDNTOOL-38) were
constructed to provided reusable makers during gene knockout. Another important group of plasmid
derivatives that were created included flippase-encoding plasmids which were used in excising antibiotic-
resistant marker after gene knockout. These plasmids include pDNTOOL-2, pDNTOOL-18, pDNTOOL-
19, pDNTOOL-20 and pDNTOOL-21. Also cloning/ expression plasmids derivatives (pDNTOOL-3,
pDNTOOL-4, pDNTOOL-22 to pDNTOOL-30, pDNTOOL-39, pDNTOOL-26, pDNTOOL-27,
pDNTOOL-28 and pDNTOOL-30) were constructed for gene expression and GFP tagging. This wide array
of plasmids was constructed using the novel DNA swapping strategy or traditional cloning method and
were subsequently use used for exploring the role of T6SS in K. pneumoniae. These plasmids also increase
the availability of diverse antibiotic-resistant derivatives of various plasmids that could potentially be used
for other research purposes.
Also, in chapter three, it was demonstrated that the resistance repertoire of the MDR K. pneumoniae
HS11286 strain could be mapped out and conveniently disarmed to create a model K. pneumoniae safer to
work with for T6SS functional studies and future routine laboratory usage. K. pneumoniae HS11286 is
carbapenem-resistant strain isolated from the sputum of a patient at Huashan Hospital, Shanghai-China (Liu
et al., 2012a). The strain has a plastic genome and six plasmids (pKPHS1, pKPHS2, pKPHS3, pKPHS4,
pKPHS5 and pKPHS6) but more importantly, pKPHS2 and pKPHS3 played a significant role in the
antibiotic resistance of the strain. Multidrug resistance of the strain was primarily mediated via its plasmids
(Appendix 2-3 Resistance genes identified in K. pneumoniae HS11286 that could confer resistance in
Clinical cases). While pKPHS2 (similar backbone to K. pneumoniae plasmid pKP048) has blaTEM-1 and
carbapenemase gene blaKPC-2 genes and, pKPHS3 showed significant similarity to Yersinia pestis plasmid
pIPI202 and possessed several resistant genes (genes such as tetG, cat, sulI, dfraI2, aac(3) and aph)
contributing to the multidrug resistance of K. pneumoniae HS11286. The blaKPC-2 gene on pKPHS2 and a
26 kb MDR region containing 12 clustered antibiotic resistance genes and one biocide resistance gene on

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
203
pKPHS3 (Appendix 2-1: List of genes deleted from the 26 kb region) were mapped out and successfully
deleted for safer use of the strain for downstream T6SS related work. After the deletion of the antibiotic
resistance genes, the MIC data (Appendix 2-2 Antibiotic susceptibility of K. pneumoniae HS11286 and the
antibiotic disarmed versions) of the antibiotic disarmed strain (DN2) relative to the parent strain (KR3317)
showed an attenuated resistance to a wide range of antibiotics including cefoxitin, ceftazidime, imipenem,
meropenem, gentamicin, tobramycin, amikacin, streptomycin, kanamycin, levofloxacin, trimethoprim-
sulfamethoxazole, trimethoprim, chloramphenicol, tetracycline, spectinomycin, colistin, polymyxin B and
tigecycline.
The deletion of only blaKPC-2 on pKPHS2 resulted in a significant loss of carbapenem (imipenem and
meropenem), cefoxitin and ceftazidime which remained unchanged even after the deletion of the 26 kb
MDR region on pKPHS3. After the deletion of the 26 kb MDR region, the attenuation aminoglycoside
resistance was most likely due to the loss of aacC2, aacA2, rmtB and strAB while the loss of aacA2, the
dfrA12-like gene, floR, and tetAR resulted in susceptibility to spectinomycin, trimethoprim,
chloramphenicol and tetracycline respectively. Likewise, loss of dfrA12-like, sul1 and sul2 most likely
resulted in the loss of resistance to trimethoprim-sulfamethoxazole. The disarmament of both the blaKPC-2
and 26 kb MDR region on the K. pneumoniae HS11286, therefore, resulted in a safer generation of a more
harmless and antibiotic-susceptible strain (DN2) which was easily genetically manipulated as model K.
pneumoniae.
Additionally, the antibiotic susceptible strain, DN2, showed a significant increase in competition relative
to the parent strain in an in-vitro culture but not in vivo Galleria larvae model, suggesting that the deleted
resistance repertoire posed a physiological and energetic cost to K. pneumoniae HS11286 while enhancing
virulence in the strain. This was not surprising as other studies have reported similar observations where
enhanced antibiotic resistance acquisition increased physiological burden on the bacteria but provide a
mechanism of subduing and evading host defence (Kidd et al., 2017; Campos et al., 2004b; Padilla et al.,
2010a; Sahly et al., 2008; Beceiro, Tomas and Bou, 2013; Beyrouthy et al., 2014; Bialek-Davenet et al.,
2015; Cannatelli et al., 2013; Chen et al., 2010; da Silva and Mendonça, 2012; El Fertas-Aissani et al.,
2013; Lavigne et al., 2013; McLaughlin et al., 2014; Ramirez et al., 2014).
The acquisition and spread of resistance-associated genes and evolution of bacteria are
significantly influenced by certain types of mobile genetic elements including plasmids, ISs,
transposons, integrons and associated gene cassettes, prophages, integrative and conjugative
elements (ICEs) and genomic islands (GIs) (Frost et al., 2005b; Frost et al., 2005c; Kottara et al.,
2017; Partridge, 2011b; Thomas and Nielsen, 2005). The mapping and disarmament of resistance

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
204
genes on pKPHS2 and pKPHS3 (blaKPC-2 and the 26 kb MDR region, respectively) reiterated
the role of plasmids as one of the most important mobile genetic elements that bear many resistance
genes in K. pneumoniae for dissemination. The resistance-disarmed K. pneumoniae HS11286
(∆kpc∆MDR or DN2) generated in this study offered a more suitable, safer and genetically
modifiable K. pneumoniae model strain that was later used for T6SS functional studies and future
works.
An in silico analysis showed the presence of different types of T6SS gene clusters (T6SS1, T6SS2 and
T6SS3) of in K. pneumoniae (Sarris et al., 2011a). However, no experimental data investigating the
contributory role of such putative T6SS gene clusters in K. pneumoniae was presented. Also, the in silico
analysis that was done then was only limited to few strains of K. pneumoniae, including K. pneumoniae
NTUH-K2044, K. pneumoniae MGH78578 and K. pneumoniae 342. But MDR K. pneumoniae HS11286
sequenced data was only published a year later (Liu et al., 2012a) and being dominant KPC-producing
ST11clone in China and closely related to CRKP clone ST258 (dominant Worldwide), it was imperative to
assess the virulence of this strain and the role of T6SS virulence in it pathogenicity. Comparative genomic
analysis revealed that K. pneumoniae carried two putative T6SS clusters (T6SS1 and T6SS3) and a
disintegrated T6SS2 (notably carrying hcp and clpV genes).
Interestingly, the T6SS1 was not only similar in content (genes within the cluster) but in context (upstream
and downstream flanking regions) to those predicted earlier by Saris et al., (2011). T6SS1 had all the
important genes considered to be necessary for assemblage and functioning of T6SS in bacteria and was
flanked by at least 5 kb conserved upstream and downstream sequences (Figure 3.8). Similarly, T6SS3
had almost all the hallmark genes for a functional T6SS except hcp (which encodes both structural and
effector proteins) and ATPase energizing clpV genes. Also, T6SS had unique conserved upstream and
downstream sequences flanking T6SS3. However, the disintegrated T6SS2 notably had hcp and clpV as the
only T6SS hallmark genes among other genes. Based on this surprising observation, it was postulated the
hcp and clpV in T6SS2 were functionally complementing the T6SS3 while T6SS1 can independently
function on its own. Inferring from these in silico analyses, it seems that K. pneumoniae possess two
functional T6SS gene clusters, T6SS1 and T6SS3.
Based on the similarity of the T6SS genes and conserved upstream and downstream flanking sequences,
unique probing primers were designed and used to identify T6SS1, T6SS2 and T6SS3 in the fourteen
sequenced K. pneumoniae strains with high specificity (Figure 3.9). These strains included K. pneumoniae
JM45, K. pneumoniae 342, K. pneumoniae 234-12, K. pneumoniae 1084, K. pneumoniae MGH 78578, K.
pneumoniae NTUH-K2044, K. pneumoniae HS11286, K. pneumoniae CG43, K. pneumoniae INH27, K.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
205
pneumoniae IH31, K. pneumoniae subsp. Rhinoscleromatis SB3432, K. pneumoniae ATTC BAA-2146, K.
pneumoniae 52-145 and K. pneumoniae KCTC2242. The identified T6SS1, T6SS2 and T6SS3 in the
fourteen K. pneumoniae strains showed different parent of arrangement in the strains. Majority of these
strain only had two functional T6SS gene clusters as it is the case of K. pneumoniae HS11286 and K.
pneumoniae NTUH-K2044. The in silico analysis provides a means of identifying and broadening the list
of sequenced K. pneumoniae strains with putative T6SS gene clusters. This also suggests that T6SS in K.
pneumoniae T6SS gene clusters may have evolved through horizontal gene transfer, supporting the notion
that T6SS is an ancestral/rudimentary feature of phage tail.
Having successfully identified the various T6SS gene clusters in K. pneumoniae, the required molecular
tools from the constructed genetic tool library were employed to generate multiple T6SS mutants in K.
pneumoniae NTUH-K2044 and resistance disarmed K. pneumoniae HS11286. Initially, the aim was to
create T6SS1 and T6SS3 single and double mutants in both strains but unfortunately, for some unknown
reason, knocking out T6SS3 in K. pneumoniae NTUH-K2044 proved unsuccessful. However, while T6SS1
and T6SS3 single or double mutants were created in K. pneumoniae HS11286, only T6SS1 mutant was
successfully created in K. pneumoniae NTUH-K2044. The susceptibility of DN2 to a wide range of
antibiotic and the availability of constructed molecular tools together with the knowledge obtained from
the in silico T6SS analysis significantly aided the generation of T6SS1 and T6SS3 single or double mutants
in K. pneumoniae HS11286 and T6SS1 mutant in K. pneumoniae NTUH-K2044. These mutants, together
with their parent strains were then employed in downstream assays to assess the virulent contributory role
of T6SS in K. pneumoniae.
In a nutshell, chapter one of this report presented a novel DNA swapping technique that was developed and
used to create a library of molecular genetic tools. Also, the antibiotic resistance landscape of K.
pneumoniae was mapped and knocked out to create a safer model strain for future K. pneumoniae studies.
Additionally, the T6SS in silico analysis provided an insight into the various type of putative T6SS gene
clusters and their conserved upstream and downstream regions present in fourteen sequenced K.
pneumoniae strains. The constructed plasmids and resistance-disarmed DN2 were used to generate various
T6SS mutants in the K. pneumoniae for functional studies based on the knowledge obtained from the T6SS
in silico analysis.
Presented in chapter four is the role of the putative K. pneumoniae T6SS gene clusters in competitive fitness,
biofilm, fimbriation and amoeba phagocytic resistance. Both intra (K. pneumoniae-K. pneumoniae
interaction) and interspecies (K. pneumoniae-E. coli or K. pneumoniae-P. aeruginosa interaction)

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
206
competition were employed to study how T6SS mediated antibacterial/competitive interaction in K.
pneumoniae. Biofilm formation in 96-well polyvinyl plate both at 30℃ and 37℃ was used to assess the
role of T6SS in enhancing biofilm formation in K. pneumoniae. TEM visualisation of the surface fimbriae
was then used to confirm the expressed fimbrial type that was involved in the biofilm formation. T6SS
mediation of antiphagocytic resistance to environmental amoeba was assessed using A. castellanii. This
provides insight as to how K. pneumoniae may survive in its natural environment.
After the deletion of T6SS gene clusters from DN2, both the mutant and parent strains were grown
separately, and the bacterial OD600nm or CFU was assessed to determine the bacteria growth curve. It was
observed that the deletion of the T6SS gene clusters did not significantly alter the growth curves of the
strains over an 18 h period, suggesting that the deleted T6SS gene clusters. Additionally, to assess whether
the T6SS gene clusters were functional or not, RNA expression of several T6SS core gene (hcp and vgrG)
were used the determinant of the functionality of the two gene clusters in K. pneumoniae. Hcp and VgrG
are two important components of T6SS that both act as a structural and effector proteins (Silverman et al.,
2012b; Cascales and Cambillau, 2012b; Zoued et al., 2014b), the RNA expression of hcp1 and vgrG1 for
T6SS1, hcp2 for T6SS2 and vgrG3 for T6SS3. Interestingly, hcp1 and vgrG1 expressed at least 5-fold more
when compared to hcp2 and vgrG3 in the wild-type K. pneumoniae (DN2) relative to the T6SS mutants.
Similarly, hcp2 which was deleted expressed more in the wild-type parent K. pneumoniae followed by
T6SS1 and T6SS3 single mutants. It was noted that hcp1 was expressed in ∆T3 despite T6SS1 deletion.
Likewise, vgrG3 was expressed in ∆T1 even after T6SS3 deletion, suggesting that the T6SS1 gene cluster
did not significantly affect T6SS3 gene cluster and the vice versa. Studies have demonstrated that different
T6SS gene clusters in the same bacteria may contribute differently to the overall role of T6SS (Zoued et
al., 2014c; Repizo et al., 2015c). Collectively, the RNA expression data (Figure 4.2) indicates that T6SS1-
related genes were more expressed in K. pneumoniae relative T6SS3, suggesting that both T6SS1 and
T6SS3 were both functional and contributed to the overall role played by K. pneumoniae T6SS.
Bacterial fitness and survival are essential in their colonization and subsequent disease-causation in the
polymicrobial environment (Sanchez, 2011; Fuqua et al., 2010; Stubbendieck, Vargas-Bautista and
Straight, 2016; Sistrunk et al., 2016; Messer et al., 2017). Since the discovery of T6SS in bacteria, many
studies have reported that T6SS is used by the bacteria in intra and interspecies bacteria competition and
fitness and evasion of amoeba (Alcoforado Diniz and Coulthurst, 2015; Alteri et al., 2013; Bartonickova et
al., 2013; Borgeaud et al., 2015c; Brunet et al., 2013a; Carruthers et al., 2013; Decoin et al., 2015; Decoin
et al., 2014a; Haapalainen et al., 2012; Hachani, Lossi and Filloux, 2013; MacIntyre et al., 2010c; Pukatzki
et al., 2006b; Repizo et al., 2015d; Russell et al., 2013; Sanchez, 2011; Unterweger et al., 2014a;
Unterweger et al., 2012; Zheng, Ho and Mekalanos, 2011a). From the bacteria-bacteria interaction data

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
207
reported in chapter four, it was noted that when T6SS mutants competed against their parent strain (DN2),
a significant attenuation in competitive fitness was observed in the T6SS mutants relative to the parent
strains. Similarly, T6SS mediated killing of E. coli was abrogated in the mutants but not the parent strain.
Among the mutants, T6SS mutant-mediated more competitive fitness or killing when it was competed head-
to-head with its parent strains or E. coli, respectively. The T6SS1 and T6SS3 double mutant (∆T1∆T3)
could barely keep up relative to the other mutants in competition or E. coli killing assay. The absence of
both T6SS gene clusters significantly lowered the competitive fitness or antibacterial potential of the K.
pneumoniae. Also, when K. pneumoniae competed against P. aeruginosa with active T6SS, it was observed
that more bacterial recovery was obtained for the DN2 but not the T6SS mutants which were significantly
inhibited P. aeruginosa. Cell contact-mediated T6SS action of T6SS bearing bacteria (Brunet et al, 2013b;
Wösten et al, 2013) is used to deliver t6ss toxins to other bacteria. A direct contact-mediated antibacterial
action of T6SSof K. pneumoniae T6SS was observed as a critical factor in the killing of E. coli. When the
bacterial mixtures were co-cultured in liquid media, K. pneumoniae could barely inhibit the growth of the
E. coli unlike when bacterial mixtures were cultured on the static surface (filter membrane). The effect of
K. pneumoniae T6SS antibacterial killing of E. coli was evident in the static bacteria mixed cultures on the
filter paper. The antibacterial action of K. pneumoniae T6SS mediated and enhanced via direct contact
delivery of effectors onto its prey, E. coli.
This was not surprising as many studies have reported T6SS immune system mostly located within the
T6SS cluster employed in protecting themselves against other bacteria T6SS (Dong et al., 2013b; Hood et
al., 2010b; MacIntyre et al., 2010a; Ishikawa et al., 2012; Murdoch et al., 2011b). Both T6SS gene clusters
individually or synergistically enhanced K. pneumoniae competitive fitness, antibacterial potential and
immunity against other bacterial T6SS action. K. pneumoniae in its native environment may employ the
use of T6SS to mediate survival and colonization by the release of T6SS effectors into the environment or
directly into competitively inhibiting the growth of other competing bacteria in the same biological niche.
These data were in concordance with earlier reports, supporting the hypothesis that K. pneumoniae uses its
T6SS to mediates antibacterial competition in the polymicrobial environment, thus, suggesting how K.
pneumoniae may persist in many habitats such as human gut and nasopharynx, water and soil.
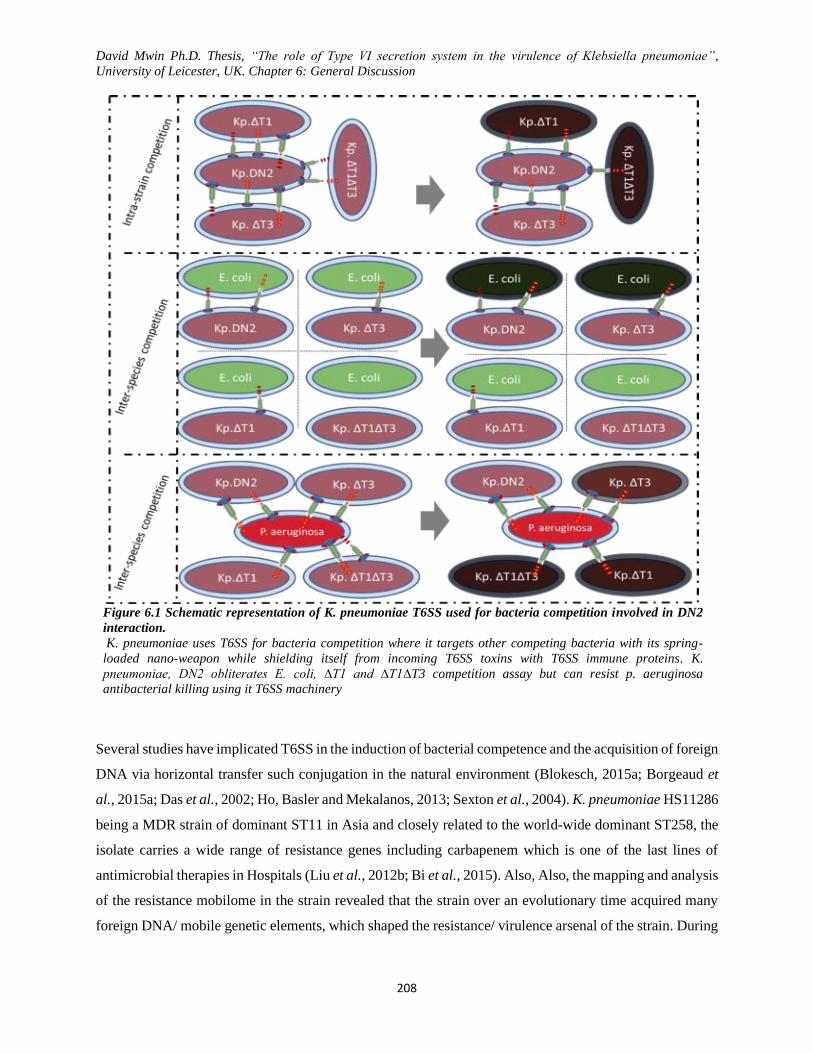
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
208
Figure 6.1 Schematic representation of K. pneumoniae T6SS used for bacteria competition involved in DN2
interaction.
K. pneumoniae uses T6SS for bacteria competition where it targets other competing bacteria with its spring-
loaded nano-weapon while shielding itself from incoming T6SS toxins with T6SS immune proteins. K.
pneumoniae, DN2 obliterates E. coli, ∆T1 and ∆T1∆T3 competition assay but can resist p. aeruginosa
antibacterial killing using it T6SS machinery
Several studies have implicated T6SS in the induction of bacterial competence and the acquisition of foreign
DNA via horizontal transfer such conjugation in the natural environment (Blokesch, 2015a; Borgeaud et
al., 2015a; Das et al., 2002; Ho, Basler and Mekalanos, 2013; Sexton et al., 2004). K. pneumoniae HS11286
being a MDR strain of dominant ST11 in Asia and closely related to the world-wide dominant ST258, the
isolate carries a wide range of resistance genes including carbapenem which is one of the last lines of
antimicrobial therapies in Hospitals (Liu et al., 2012b; Bi et al., 2015). Also, Also, the mapping and analysis
of the resistance mobilome in the strain revealed that the strain over an evolutionary time acquired many
foreign DNA/ mobile genetic elements, which shaped the resistance/ virulence arsenal of the strain. During

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
209
plasmid transfer via conjugation, T6SS lethal response may be triggered by mating pair formation (Mpf)
encoded by the recipient bacterial T4SS conjugation system (Ho, Basler and Mekalanos, 2013). Thus, it
was postulated that K. pneumoniae presence or absence of T6SS could significantly affect the ability of K.
pneumoniae to acquire mobile genetic elements such as plasmids via T4SS conjugative transfer to enhance
resistance and survival.
Have demonstrated that K. pneumoniae T6SS was able to induce a direct antibacterial activity on E. coli, a
plasmid mobilization assay was used to examine the how K. pneumoniae T6SS could affect the frequency
of plasmid transfer to an E. coli donor. K. pneumoniae strains carrying a pACYC184-oriTICE were separately
mated with streptomycin-resistant E. coli HB101, and E. coli HB101 transconjugants were subsequently
selected on LA plus streptomycin or LA plus streptomycin and chloramphenicol on a filter paper.
Surprisingly, only E. coli HB101 transconjugants recovered from a mixture of with T6SS1 mutant and E.
coli HB101 showed an elevated conjugal transfer of the plasmid but not T6SS3 mutant or T6SS1/T6SS3
double mutant. A reduced plasmid mobilization frequency was induced by both the parent K. pneumoniae
HS11286 strain (DN2) and ∆T1∆T3 (T6SS1/T6SS3 double mutant) relative to T6SS1 and T6SS3 single
mutants. However, the CFU of the E. coli recovered from ∆T1∆T3 + E. coli HB101 or ∆T1 + E. coli HB101
was significantly higher than that of the DN2. Thus, though ∆T1∆T3 showed a lower plasmid mobilization
frequency just like DN2 strain. The antibacterial killing of E. coli was significantly diminished just like
∆T1 strain but not DN2. Also, ∆T3 induced a significant plasmid mobilization relative to DN2, and ∆T1∆T3
strains even though the E. coli recovered from the mixture not substantial relative to DN2 strains. The RNA
expression of two crucial T4SS genes (virB4 and mobB) was assessed to understand further why
T6SS1/T6SS3 double mutant (∆T1∆T3) did not yield higher frequency as anticipated. The RNA expression
data revealed the both virB4 and mobB were highly expressed in DN2, ∆T1 and ∆T3 strain but not ∆T1∆T3
strain. The lower expression of the T4SS conjugative machinery in ∆T1∆T3 accounted for its lower induced
plasmid mobilization frequency despite its abrogated E. coli killing ability. The results suggest that the
deletion of T6SS1 in K. pneumoniae may induce an enhanced conjugative plasmid mobilization or mobile
genetic elements but reduced antibacterial killing potential. On the other hand, though the deletion of T6SS3
in K. pneumoniae may relatively enhance T4SS conjugal transfer of genetic elements, its T6SS antibacterial
activity was not abolished since T6SS1 may still be active in the background. These findings suggest that
T6SS and T4SS may be working together to attract other bacteria via conjugal mating pair formation, kill
them and subsequently some genetic material when required to survive. Thus, T6SS in K. pneumoniae
exhibits direct antibacterial and anti-genetic material transfer against other bacteria. This agrees with other
studies suggesting that T6SS may be triggered by the mating pair formation T4SS conjugal machinery and
actively used as an antibacterial and genetic material sequestrating arsenal in partnership with other virulent

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
210
factors to out-compete other bacteria to survive in the same environment (Ho, Basler and Mekalanos, 2013;
Blokesch, 2015a; Borgeaud et al., 2015a; Das et al., 2002).
Biofilm formation associated with bacterial fimbriae and capsule remains one of the most explored virulent
related area of bacteria investigated for their role in antibiotic resistance and bacterial pathogenicity(Dos et
al., 2014; Fierer, 2012; Hancock, Witsø and Klemm, 2011; Klausen et al., 2003; Stahlhut et al., 2012; Olson
et al., 2012; Schembri et al., 2005). T6SS has been reported to play a significant role in biofilm formation
and fimbriae expression (De Pace et al., 2010; Aubert, Flannagan and Valvano, 2008b; Gallique et al.,
2017; Khajanchi et al., 2009; Linares et al., 2016; Tian et al., 2015; Vaysse et al., 2009). It was hypothesized
that K. pneumoniae HS11286 T6SS contributes to biofilm formation and the expression of surface
appendages such as fimbriae. The biofilm data revealed that the deletion of T6SS1 (∆T1) or T6SS3 (∆T3)
significantly enhanced the biofilm formation at all the time-points and temperatures relative to the parent
strain, DN2 and ∆T1∆T3. The T6SS double mutant (∆T1∆T3) showed lesser biofilm formation phenotype
at 48h and 72 h but not 24 h post-inoculation. A microscopic visualisation the biofilm formation on
coverslips confirmed the data obtained from the biofilm formation in the polyvinyl 96 well plates; many
attached ∆T1 films were almost evenly distributed on the coverslips unlike many massive clumps/ clusters
of ∆T3 films relative to the biofilm associated with the parent strain, DN2. Also, few scattered clumps of
∆T1∆T3 films were observed but did not amount to the biofilm-associated with ∆T1 and ∆T3. An
agglutination test confirmed the expression of different fimbrial types induced by T6SS. The ∆T3 strain
agglutinated GP-RBCs and TATOXB but was mannose-sensitive or mannose- resistant respectively. The
agglutination data suggested T6SS-induced differential expression of various type of fimbriae on K.
pneumoniae. A further investigation using, a fimH (type 1 fimbria) and mrkD (type 3 fimbria) RNA
expression and Transmission Electron Microscopy (TEM) data revealed that the deletion of T6SS1 (∆T1)
resulted in an enhanced expression of type 1 fimbriae while the deletion of T6SS3 (∆T3) resulted in a much-
enhanced type 3 fimbria expression relative to the other K. pneumoniae HS11286 strains (DN2 and
∆T1∆T3). The observed poor biofilm formation in DN2 and ∆T1∆T3 strains could be associated with low
expression of type 1 and type 3 fimbriae. Once again, this data suggests that T6SS is not only co-expressed
with T4SSS but are also involved with the expression of K. pneumoniae fimbrial appendages (type 1 and
3) and associated biofilm formation, this has been reported in other bacteria (De Pace et al., 2010; Gallique
et al., 2017; Linares et al., 2016; Tian et al., 2015).
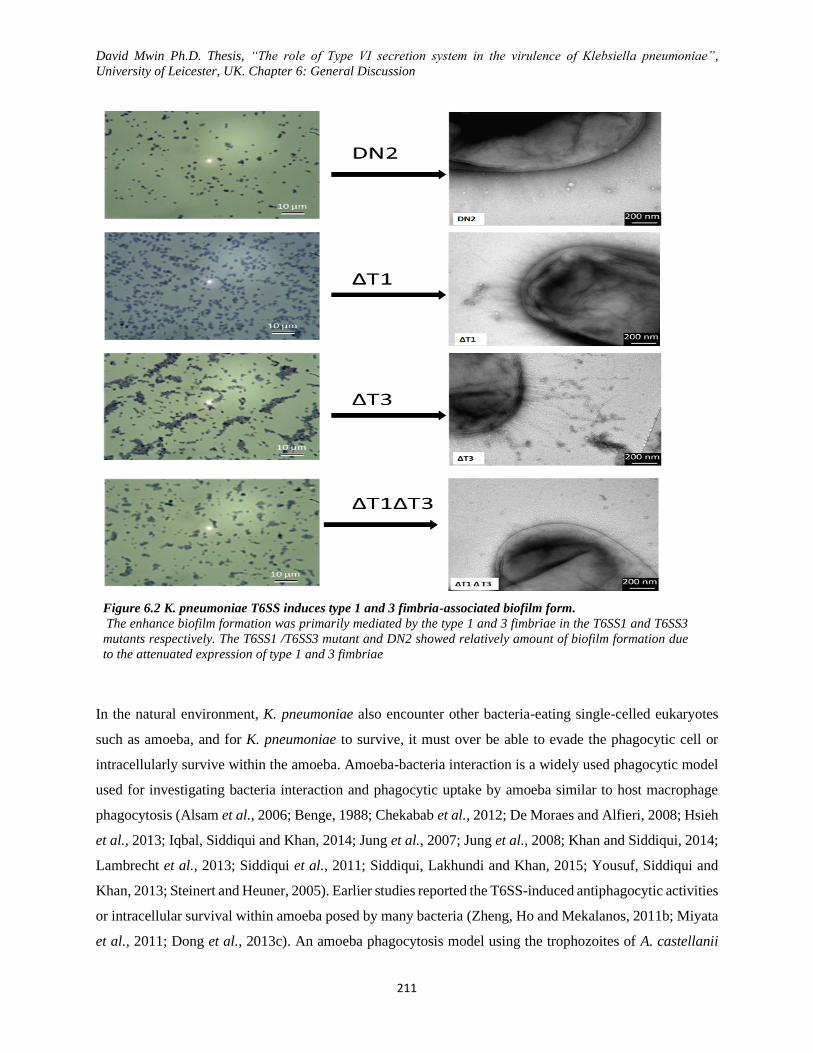
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
211
Figure 6.2 K. pneumoniae T6SS induces type 1 and 3 fimbria-associated biofilm form.
The enhance biofilm formation was primarily mediated by the type 1 and 3 fimbriae in the T6SS1 and T6SS3
mutants respectively. The T6SS1 /T6SS3 mutant and DN2 showed relatively amount of biofilm formation due
to the attenuated expression of type 1 and 3 fimbriae
In the natural environment, K. pneumoniae also encounter other bacteria-eating single-celled eukaryotes
such as amoeba, and for K. pneumoniae to survive, it must over be able to evade the phagocytic cell or
intracellularly survive within the amoeba. Amoeba-bacteria interaction is a widely used phagocytic model
used for investigating bacteria interaction and phagocytic uptake by amoeba similar to host macrophage
phagocytosis (Alsam et al., 2006; Benge, 1988; Chekabab et al., 2012; De Moraes and Alfieri, 2008; Hsieh
et al., 2013; Iqbal, Siddiqui and Khan, 2014; Jung et al., 2007; Jung et al., 2008; Khan and Siddiqui, 2014;
Lambrecht et al., 2013; Siddiqui et al., 2011; Siddiqui, Lakhundi and Khan, 2015; Yousuf, Siddiqui and
Khan, 2013; Steinert and Heuner, 2005). Earlier studies reported the T6SS-induced antiphagocytic activities
or intracellular survival within amoeba posed by many bacteria (Zheng, Ho and Mekalanos, 2011b; Miyata
et al., 2011; Dong et al., 2013c). An amoeba phagocytosis model using the trophozoites of A. castellanii

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
212
was used to investigate the role of T6SS in amoeba phagocytosis of K. pneumoniae HS11286. Data from
the bacterial association and internalization assay revealed that the deletion of T6SS1 or T6SS3 enhanced
K. pneumoniae HS11286 association with A. castellanii however the deletion of T6SS1 (∆T1) or
T6SS1/T6SS3 (∆T1∆T3) increased K. pneumoniae uptake and internalization within A. castellanii relative
to the parent K. pneumoniae HS11286, DN2 strain. The enhanced bacterial association and internalization,
viable amoeba recovery and amoeba clearance of the K. pneumoniae T6SS mutant could be associated with
the attenuation in virulence-associated with T6SS deletion. Expectedly, the deletion of both T6SS1 and
T6SS3 gene clusters significantly impaired the antiphagocytic activity of K. pneumoniae HS11286 and
enhanced the ability of A. castellanii to phagocytose and digest K. pneumoniae suggesting that T6SS was
prominent in the survival of K. pneumoniae HS11286 in an aquatic environment when interacting with the
amoeba. Also, the ability for K. pneumoniae to combined T6SS antagonistic- defensive behaviour, T4SS-
dependent acquisition of mobile genetic elements, biofilm formation and amoebal phagocytic resistance
adds to K. pneumoniae pathogenicity in the host and environmental persistence.
Host blood serum with abundance complements present a significant immune bactericidal barrier to
invading bacterial pathogens and results in the elimination of the pathogen. However, when pathogens
evade the bactericidal activities of serum, they persist, spread and pose serious virulence to the host. Many
K. pneumoniae strains via various mechanisms such as capsule, lipopolysaccharide among others has been
reported to resist serum bactericidal activities posing significant diseases potential (Sharma, Fatma and
Thukral, 1999; Shankar-Sinha et al, 2004b; Sahly et al, 2004; Kumar et al, 2011a; Hsieh et al, 2012b;
Doorduijn et al, 2016; de Astorza et al, 2004; Cox et al, 2015). MDR K. pneumoniae HS11286 exhibit a
broad range of antibiotic resistance even though its serum bactericidal resistance was unknown. Having
observed the varying impact of T6SS deletion on the K. pneumoniae HS1186 phenotype, resistance to pool
normal human serum and C3b deposition was used to assess T6SS role in serum bactericidal resistance.
C3b through the various downstream processes of serum factor binding and cleavage is converted to form
C3 convertase which cleaves C3 (C3a and C3b) leading C5 convertase. The C5 convertase then cleaves C5
into C5a and C5b where the latter bind to C6, C7 and C8 in succession to form C5b-8 complex which gets
polymerise after forming a complex with C9, leading to membrane attack complex formation on the bacteria
membrane. The data revealed that T6SS mutants were impaired in survival, suggesting reduced resistance
to the pooled normal human serum relative to the parent strains due to the high deposition of C3b.
Noticeably, though the deletion of T6SS3 mutant showed the highest C3b deposition, more T6SS3 mutant
was recovered from the serum when compared to T6SS1 (∆T1), and T6SS1/T6SS3 (∆T1∆T3), however the
parent K. pneumoniae HS11286 strain (DN2) showed the most serum resistance and but low C3b
deposition. Also, the deletion of T6SS gene clusters enhanced opso-phagocytosis by PMN at 1h post-
infection a 1h but not at 3 h. Thus, the low C3b deposition and opso-phagocytosis in T6SS1 (∆T1) a and

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
213
T6SS1/T6SS3 (∆T1∆T3) resulted in attenuated survival of T6SS1 (∆T1), and T6SS1/T6SS3 (∆T1∆T3)
relative to DN2 and T6SS3 mutant at 2 and 3h post-inoculation. The T6SS3 mutant surprisingly exhibited
more C3b deposition, serums survival and internalization within neutrophils when compared to the other
T6SS mutant at 3 h post-inoculation/infection. This may be because of the residual functional T6SS1 and
increased fimbrial expression in ∆T3 mutant. These data revealed that K. pneumoniae HS11286 T6SS via
an unknown mechanism promotes resistance to opso-phagocytosis and serum bactericidal resistance.
Many studies have demonstrated that T6SS is involved in a host in vivo virulence among various model
(Weyrich et al., 2012; Ma and Mekalanos, 2010a; Schell et al., 2007; Brodmann et al., 2017; Lennings,
West and Schwarz, 2018; Ungureanu et al., 2019; Repizo et al., 2015a). G. mellonella larvae killing is a
well-established and widely used bacterial in vivo model used to assess the virulence of bacteria. The larvae
hemocoel carry specialized phagocytic cells called hemocytes as part of the larva innate immunity resemble
mammalian phagocytes (Mylonakis, Casadevall and Ausubel, 2007). These cells kill pathogen by releasing
antimicrobial properties during phagocytosis and reactive oxygen species (human neutrophil to NADPH
oxidase homologue) (Bergin et al., 2005). Few studies have reported the use of Galleria larvae to assess
the T6SS in vivo virulence in bacteria and remains a cost-effective in vivo model for virulence exploration.
Mortality of Galleria larvae whose phagocytic antimicrobial innate immunity share similarity with the
phagocytic mechanism used by host phagocytes, was used to assess the in vivo virulence role of T6SS in
K. pneumoniae HS11286. Melanisation of larvae which often leads to death, is consistent with the virulence
of the infecting strains, this was measured using phenoloxidase activities. I postulated that K. pneumoniae
T6SS enhances the bacterial in vivo virulence in the host organism. As expected, the deletion of T6SS
related gene clusters significantly mitigated the T6SS-associated in vivo virulence evidenced by the reduced
killing of Galleria larvae, low phenoloxidase activities and inferior competitive edge. The attenuation of
T6SS mutant virulence in the larvae ultimately enhanced more hemocyte viability (expressing more
antimicrobial properties), reduced melanisation rate and low mortality. Also, the T6SS promoted in vivo
competing in the larvae hemocoel where more parent DN2 strains were significantly recovered, unlike the
T6SS mutants that were least recovered from parent + T6SS mutant mixture. Among T6SS mutants, the
deletion of both T6SS gene clusters (∆T1∆T3) did not only attenuate in vivo virulence but also impaired K.
pneumoniae HS11286 in vivo competition potential against the other T6SS mutants ((∆T1 and ∆T3) strains
or parent strains, DN2. DN2 strain (carrying both T6SS gene clusters) induced early melanisation of
infected larvae leading to high larval mortality which was corroborated by the increased phenoloxidase
activities stimulated by DN2 larval infection. Thus, these obtained data proved that Galleria larvae is very
good in vivo virulence model for opportunistic pathogens and was used to confirm the hypothesis that K.
pneumoniae T6SS does not only enhance antibacterial competition but promotes in vivo virulence in the
host organism.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
214
Host immunity system has evolved via perpetual interaction of pathogens and multicellular host under
acting bidirectional selective pressure leading to the diverse mechanism of attack- and counter-attack
relationship between the pathogen and host, respectively. Host innate immune system is populated with a
myriad of immune cells (macrophages, dendritic cells, neutrophils, epithelial cells, etc.) which acts as a line
of defence against invading pathogens via of mechanisms (Chaplin, 2006; Chaplin, 2010; Parkin and
Cohen, 2001). Host macrophage is on one of the first cells to response at the site of infection by initiating
phagocytosis and subsequent release of inflammatory cytokines and other immune products to orchestrate
cellular defence in the host organisms. Some of the immune products released include IL-12 for activation
of NK cells, neutrophil chemoattractant LTB4 via IL-8, harmful oxygen radicals, T-cell stimulatory and
pro-inflammatory cytokines (TNF-a, IL-1a) and antimicrobial lactoferrin and lysozyme among many others
(Adams and Hamilton, 1984; Burke and Lewis, 2002; Gordon and Read, 2002; Parameswaran and Patial,
2010; Ross and Auger, Wynn, Chawla and Pollard, 2013). Similarly, epithelial cells play an important role
in initiating and regulating both innate and adaptive immunity. Host epithelial cells are involved
mucociliary clearance, production of enzymes and complements, permeabilizing peptides, inhibitors,
reactive oxygen species and stimulatory cytokines among other products as a means of defence against
pathogens (Bals and Hiemstra, 2004; Schleimer et al, 2007; Stanke, 2015). Bacterial virulence via various
mechanism towards host immune cell as a strategy to evade the host immune system to enhance its
pathogenicity presents a significant danger to the host. Many reports have suggested that T6SS induced
virulence toward host by targeting and picturing host cells with the loaded toxin in a contact-dependent
manner (Sana et al., 2012b; Bröms et al., 2012b; Burtnick et al., 2010; Ma and Mekalanos, 2010b; De
Ceuleneer, Vanhoucke and Beyaert, 2015; Wan et al., 2017; Weyrich et al., 2012; Zhang et al., 2012;
Russell et al., 2011b) and some accompanying phenotypic immune cell response such as actin cross-
linking/cytoskeletal rearrangement and cytokine induction (Satchell, 2009; Ma et al., 2009b; Dutta et al.,
2017; Durand et al., 2012; Suarez et al., 2010a; Suarez et al., 2010b; Ma and Mekalanos, 2010b)
Data obtained from macrophage infection revealed that the presence of T6SS gene clusters in the parent
strain, DN2 promotes the antiphagocytic resistance and subsequent survival of K. pneumoniae within the
murine macrophage. However, the deletion of T6SS gene clusters resulted in attenuated antiphagocytic
resistance and reduced survival within the macrophage. Interestingly, the T6SS3 mutant resisted and
survived better in the macrophages relative to the T6SS1 and T6SS1/T6SS3 mutants. Relatively in order
increasing attenuation, T6SS1 mutant (∆T1) and T6S T6SS1/T6SS3 mutants (∆T1∆T3) were the least
recovered or visualized surviving bacteria within the macrophages and could hardly keep up with the other
K. pneumoniae strains. Among the T6SS mutants, more T6SS3 mutants were visualized or recovered from
the macrophage relative to the other T6SS mutants, which maybe because of the presence of the functional
T6SS1 gene cluster in the T6SS3 mutant background. Importantly, while the presence of T6SS in DN2

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
215
gradually promoted increased bacterial CFU recovery from the macrophages, the absence of T6SS -related
gene clusters in theT6SS mutants significantly inhibited the recovery of the T6SS mutants. The low
visualization or bacterial CFU recovery for the T6SS mutant could be associated with the abolished
virulence in the T6SS mutants but not in the parent K. pneumoniae strains.
It was observed that K. pneumoniae T6SS enhanced the attachment, invasion and survival of epithelial cells
(A549 and H292) and caused polymerization of the actin filaments of the cells. Additionally, the deletion
of T6SS3 gene cluster significantly enhanced the attachment of the K. pneumoniae to the epithelial cells
when compared to the attachment of DN2 and the other T6SS mutants. However, T6SS3 mutants could not
invade and survive within the epithelial cell as much as the parent strains DN2. The deletion of T6SS1 and
T6SS1/T6SS3 abrogated the attachment, invasion and survival of K. pneumoniae in the epithelial cell
relative to the T6SS mutant or DN2 parent strain. The enhanced attachment of T6SS3 mutants is possibly
imitated by the abundant expression of surface fimbriae, mostly type 3 fimbriae. Confocal microscopy data
confirmed that more T6SS3 mutants followed by DN2 parent strain attached to the epithelial cells but lesser
T6SS3 mutant were recovered from the epithelial cell invasion and survival assay relative to DN2 strain.
Furthermore, the staining of the actin filament with rhodamine-phalloidin revealed that T6SS in K.
pneumoniae stimulated rearrangement and polymerization of the epithelial cell actin filament. The parent
strains DN2 enhanced cell membrane permeability and toxicity and completely obliterated the epithelial
cell actin filament relative to the T6SS mutant. Among the mutants, while actin filament and cell membrane
fidelity were almost preserved in epithelial cells infected by T6SS1 and T6SS1/T6SS3. T6SS3 mutant
showed a relative level of actin-cross linking polymerization because of its increased invasion and survival
in the epithelial cell relative to the other T6SS mutants. The data suggest that increased K. pneumoniae
invasion, survival and actin-cross linking polymerization of the epithelial cells was facilitated by T6SS.
The induction of cell actin-cross linking/ actin filament polymerization by T6SS associated toxins was
reported in several studies (Ma and Mekalanos, 2010b; Dutta et al., 2017; Durand et al., 2012; Satchell,
2009). While K. pneumoniae is mainly extracellular opportunistic pathogens, is capable of intracellular
survival within host cells. Opportunistic pathogens such as K. pneumoniae via the release of effector
proteins such as those associated with T6SS exploit actin filament/cytoskeletal rearrangement to enable
anti-phagocytosis, attachment, invasion, movement and survival within host cells, thereby facilitating
infection.
While K. pneumoniae T6SS may differentially stimulate J774 to express a wide range of cytokines as
demonstrated with the mouse cytokine array proteome profiler kit, several inflammatory cytokines (such
as TNF-a, IL-6, RANTES, IL-1a, IP-10, MIP-1a and MCP-1) were considered for quantitative ELISA.
Elevated expression of TNF-a, IL-6 and RANTES in the infected J774 supernatant was initially stimulated

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
216
by T6SS3 mutant (at 3 h) but was later more expressed my DN2-infected macrophages at 6 h. The T6SS
mutants failed to stimulate the expression of TNF-a, IL-6 and RANTES in the macrophages when compared
to that of the parent K. pneumoniae strain, DN2. Mainly, the T6SS1/T6SS3 mutant (∆T1∆T3) stimulated
the least J774 induced expression of TNF-a, IL-6 and RANTES followed by T6SS1 mutant (∆T1).
Additionally, more macrophage-induced expression of IP-10 and MCP-1 were stimulated T6SS mutants,
unlike the parent strain where T6SS1/T6SS3 mutant T6SS3 mutant stimulated the highest IP-10 and MCP-
1 respectively. Also, T6SS mutants (∆T1 and ∆T1∆T3) stimulated more expression of macrophage MIP-1
(a and b subunits) and IL-1a when compared to DN2- and ∆T3- induced macrophages expression of MIP-
1 and IL-1a. Similarly, it was also observed K. pneumoniae stimulated the expression of several
inflammatory cytokines such as IL-8, GROa and MIF in lungs epithelial cell, A549 supernatant. T6SS3
mutant stimulated more expression of IL-8, unlike T6SS1/T6SS3 mutant which stimulated the least IL-8,
GROa and MIF. More expression of GROa and MIF were significantly stimulated by DN2 more than T6SS
mutants, T6SS1, T6SS3 or T6SS1/T6SS3 mutants. T6SS mutants (∆T1, ∆T3 and ∆T1∆T3 in decreasing
order) stimulation of A549 to expressed MIF was noticeably attenuated relative to their parent strain DN2.
The T6SS1/T6SS3 mutant (∆T1∆T3) among all the T6SS mutants stimulated the least amount of IL-8,
GROa and MIF. The K. pneumoniae stimulated macrophage, and epithelial induced cytokine expression
data suggest that T6SS is actively involved in triggering an elicited inflammatory cytokine immune
response where different K. pneumoniae T6SS gene cluster could potentially stimulate specific
inflammatory cytokine in host organisms, thereby recruiting inflammatory-associated cellular infiltration
to the site of infection.
Generally, K. pneumoniae T6SS facilitated an enhanced host cell attachment, invasion, survival and the
collapse of the actin filament cytoskeletal structure. The various T6SS gene clusters mediated varying level
of virulence towards the target competing bacterial or host cells. This was not surprising as the in silico
mapping and comparative T6SS gene cluster analysis in K. pneumoniae HS11286 revealed that T6SS1
VgrG1 might be conically sharpened with pAAR (proline-alanine-alanine-arginine) repeat protein which
tends to carry different cell targeting effector domain proteins unlike the T6SS3 VgrG3 with a C-terminal
S-type pyocin that share similarity with that of P. aeruginosa suggesting a novel antibacterial mechanism.
While K. pneumoniae T6SS generally promoted competition and virulence, deletion of T6SS1 and/ or
T6SS3 revealed different phenotypic interaction with bacteria or host cells. The distribution of pAAR
related VgrG1 and s-type pyocin related VgrG3 for T6SS1 and T6SS3 respectively was widely distributed
among the analysed K. pneumoniae genome. Thus, the unique phenotype exhibited by T6SS1 and T6SS3
gene cluster may be due to the specific effectors expressed by each T6SS gene cluster for different
requirements. The different effector proteins possibly translocated by T6SS1 and T6SS3 onto target cells
may account for the displayed difference in virulence towards bacterial and host cell targets and the wide
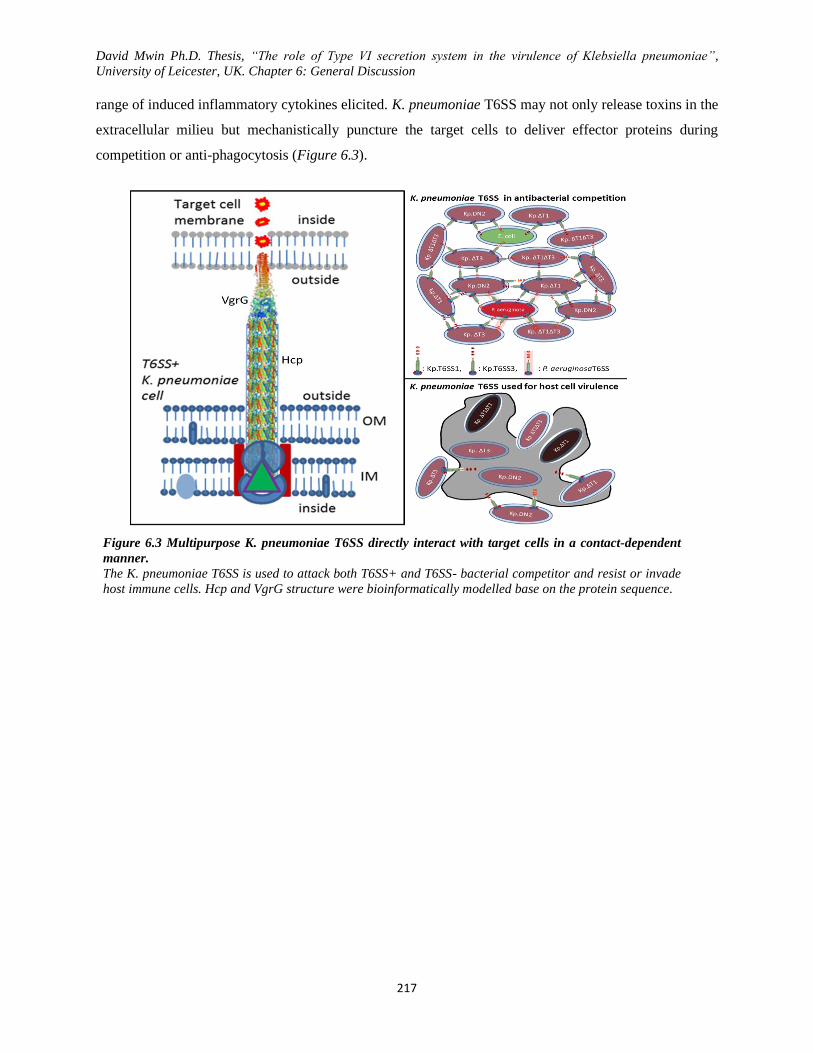
David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
217
range of induced inflammatory cytokines elicited. K. pneumoniae T6SS may not only release toxins in the
extracellular milieu but mechanistically puncture the target cells to deliver effector proteins during
competition or anti-phagocytosis (Figure 6.3).
Figure 6.3 Multipurpose K. pneumoniae T6SS directly interact with target cells in a contact-dependent
manner.
The K. pneumoniae T6SS is used to attack both T6SS+ and T6SS- bacterial competitor and resist or invade
host immune cells. Hcp and VgrG structure were bioinformatically modelled base on the protein sequence.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 6: General Discussion
218

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 7: Conclusion
219
Chapter Seven
Conclusion

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 7: Conclusion
220
Chapter 7: Conclusion
Genetic manipulation in hyper mucoid and multidrug-resistant K. pneumoniae can be very daunting,
especially when the needed genetic tools are limited or unavailable. The generation of genetic tool library
required to investigate the virulence and functional studies of K. pneumoniae and other Gram-negative
bacteria can be very laborious and costly. Thus a need for a more efficient strategy of gene swapping and
DNA manipulating during plasmid construction cannot be overemphasized. In this report, a novel gene
swapping and manipulating strategy were established and used to generate a library of genetic tools
efficiently. This technique used a negative selection marker (sacB) together with an antibiotic selection
marker and flanking upstream and downstream homologues regions to first swab the gene via lambda red
recombination to create an intermediate plasmid in E. coli background. The desired library of SOE-PCR
products/ mutant alleles with matching upstream and downstream homologues regions was used to swap
the sacB + antibiotic marker on the intermediate plasmid via lambda red recombination. The E. coli
transformants were first selected on LA + required antibiotic before subsequently selection them on LA
sucrose + required antibiotic to eliminate residual intermediate plasmid (carrying sacB) while maintaining
the desired constructed plasmid in the E. coli. This strategy was easier and very efficient (100%) in the
generation of various genetic tools (pDNTOOOL plasmids) of interest with and little or use of restriction
enzymes and fewer oligonucleotide primers (with 30-100 bp homologous upstream and downstream
flanks). Some of these generated tools were used employed in the genetic manipulation of MDR K.
pneumoniae for T6SS functional studies. This gene/DNA swapping technique with little optimization could
be potentially extended to generate markerless chromosomal gene complementation for functional gene
studies in most Gram-negative bacteria. This novel gene/DNA swapping strategy proved to be a very
efficient method. It aided the generation of a wide range of plasmids that were desperately needed for
exploring K. pneumoniae.
The antibiotic resistance mobilome of K. pneumoniae HS11286 was successfully identified, mapped and
disarmed to produce a susceptible K. pneumoniae HS11286 strains that were subsequently used for T6SS
functional studies. The antibiotic resistance of K. pneumoniae HS11286 was mediated mainly by plasmids
carrying the resistance genes and other mobile genetic elements acquired via horizontal gene transfer. It
may have undergone several evolutionary events to shape the resistance mobilome of the bacterium. Using

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 7: Conclusion
221
lambda red recombination strategies, blaKPC-2 (on pKPHS2) and a 26 kb antibiotic resistance region (on
pKPHS2) were sequentially deleted from K. pneumoniae HS11286 to produced resistance disarmed and
safer K. pneumoniae model strain. The deletion of only blaKPC-2 resulted in carbapenem (imipenem and
meropenem), cefoxitin and ceftazidime resistance attenuation. The deletion of the 26 kb resistance region
further abrogated aminoglycoside spectinomycin, trimethoprim, trimethoprim-sulfamethoxazole,
chloramphenicol and tetracycline resistance. Antibiotic susceptibility testing of the resistance disarmed K.
pneumoniae strain revealed that the strain was susceptible to a wide range of antibiotics, including
meropenem. The resistance regions were easily identified, mapped and knocked out with the use of some
of the constructed genetic tools. The mapping and disarmament of blaKPC-2 and the 26 kb MDR region on
pKPHS2 and pKPHS3 respectively and subsequent antibiotic susceptibility testing indeed confirmed
associated-multidrug resistance observed in the original K. pneumoniae HS11286 isolate while highlighting
the role of plasmids as the main bearer of these resistance genes in K. pneumoniae HS11286.
Additionally, the deletion of the resistance regions in K. pneumoniae HS11286 enhanced in vitro
competition but showed dwindled virulence in Galleria larvae relative to the parent strain. This agrees with
other studies and confirms the associated burden of physiological and energetic cost accompanying the
acquisition of antibiotic resistance to enhance bacterial virulence. The resistance-disarmed K.
pneumoniae HS11286 (∆kpc∆MDR or DN2) provided a more suitable, safer and genetically
modifiable K. pneumoniae model strain for T6SS functional studies and other future research.
Based on in silico analysis, only two major T6SS gene clusters, T6SS1 and T6SS3 were identified
(T6SS1 and T6SS3) and noted to be widely distributed among K. pneumoniae genomes. T6SS1
gene cluster carried all the known hallmark genes of T6SS within the same cluster, unlike T6SS3
missing ClpV and Hcp genes within the cluster.
The hcp2 and ClpV2 distantly located away from T6SS3 gene cluster is believed to be a
complementing the missing Hcp and ClpV genes in T6SS3 Per the analysed K. pneumoniae
genomes, T6SS1 and T6SS3 have unique conserved upstream and downstream flanking regions
across the K. pneumoniae genome, suggesting the common ancestral evolution of the T6SS gene
clusters. Also, carefully analysing each K. pneumoniae T6SS cluster, the VgrG terminal shaping
protein for each cluster may be different; a pAAR protein that can carry other effector domains
was associated with VgrG1 of the T6SS1 while S-type pyocin protein was found to be associated
with VgrG3 of the T6SS3. The knowledge obtained from T6SS in silico analysis together
resistance disarming of MDR K. pneumoniae and the constructed library of molecular genetic tools
were used to successfully generate was T6SS1 mutant, T6SS3 mutant and TSS1/T6SS3 double

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 7: Conclusion
222
mutant all in resistance-disarmed K. pneumoniae HS11286 background and T6SS1 in hyper mucoid
K. pneumoniae NTUH-K2044 though knocking out T6SS3 in K. pneumoniae NTUH-K2044 was
unsuccessful. These T6SS mutants were then used in various downstream virulence determining assay to
assess the role of K. pneumoniae T6SS.
The data obtained from the bacteria-bacteria suggest that T6SS in K. pneumoniae facilitates antibacterial
competition bacteria by targeting both T6SS negative and T6SS positive Gram-negative competing bacteria
with toxins in a T6SS contact-dependent manner. The deletion of T6SS1 and/or T6SS3 significantly
attenuated the antibacterial virulence of K. pneumoniae even though deletion of only T6SS3 still showed a
significant antibacterial virulence when compared to that of T6SS1 and T6SS1/T6SS3 mutants. Inferring
from this, K. pneumoniae T6SS gene clusters (T6SS1 and T6SS3) exhibit different levels of antibacterial
virulence and possibly play a different role since each carry’s different effector proteins. Furthermore, the
frequency plasmid mobilization from the T6SS mutant (T6SS1 and T6SS3 but not T6SS1/ T6SS3 mutant)
to donor E. coli was enhanced compared to the parent strain. The low plasmid mobilization frequency and
attenuated antibacterial virulence induced by T6SS1/T6SS3 was because of the reduced expression of T4SS
conjugal machinery and T6SS inactivation, respectively. This suggests that MDR K. pneumoniae HS11286
may be acting as an antibiotic resistance sink by actively using T6SS to target and kill other bacteria while
acquiring foreign DNA/ mobile genetic elements via horizontal gene transfer. This T6SS antibacterial
mechanisms and acquisition of mobile genetic elements are probably explored by K. pneumoniae to survive
in a complex microbial community in the natural environment or host.
The deletion K. pneumoniae T6SS gene cluster differentially induced significant type 1 and 3 fimbriae
expression which is responsible for the upregulated biofilm formation in the T6SS mutants (T6SS1 or
T6SS3) but not T6SS1/T6SS3 double mutants. Confirmed with GP-RBS and TATOXB agglutination teste,
RNA expression and electron microscopy, T6SS1 mutant mainly expressed type 1 fimbria while type 3
fimbriae expressed by T6SS3 mutants. The expression T6SS1 and T6SS3 gene clusters may be tightly
regulated with virulence gene such as biofilm forming type1 and type 3 fimbriae. Additionally, K.
pneumoniae T6SS promotes resistance and survival in amoeba and macrophage and serum complement-
mediated bactericidal resistance, confirming the T6SS virulence mechanism toward host cells. The various
K. pneumoniae T6SS gene cluster exports different effector protein to mediate varying level of
antiphagocytic virulence to amoeba and host macrophages. The attenuated serum complement-mediated
bactericidal resistance was because of the increased deposition of complements (C3b) on the T6SS mutants.
K. pneumoniae T6SS also mediate enhanced invasion and intracellular survival of K. pneumoniae within
the host epithelial and PMN cells while activating actin filament cross-linking/ cytoskeletal rearrangement.
Mutation of K. pneumoniae T6SS1 or T6SS1/T6SS3 significantly attenuated the actin filament

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 7: Conclusion
223
polymerization and cell toxicity relative to T6SS3 mutant and the parent strain. This indicates that K.
pneumoniae T6SS gene clusters (particularly T6SS1) may potentially secrete actin filament targeting
effectors as VgrG1 (T6SS1) and VgrG3 (T6SS3) carry different effector domain. Host cell cytokine
expression analysis revealed that K. pneumoniae T6SS sufficiently triggered the host innate immune system
by stimulating differential expression of pro-inflammatory cytokines/ chemokines such IL-1, MIP-1 MCP-
1, IL-6, TNF-a, RANTES, IP-10, IL-8, GROa and MIF These cytokines and chemokines may eventually
enhance the inflammation reaction or chemokines induce the chemoattraction of immune cell to the
infection site in the host, respectively.
Finally, K. pneumoniae T6SS induced in vivo virulence to Galleria larvae by resisting the bactericidal
clearance activities of the larvae innate immunity, particularly the haemocytes and stimulating
phenoloxidase-mediated larval melanisation and mortality. The diminished mortality of T6SS infected
larvae was because of the attenuated in vivo virulence to the larvae. K. pneumoniae T6SS did not only
promote in vitro antibacterial coemption but enhanced in vivo competition, survival and virulence in
Galleria larvae.
7.1 General conclusion
In summary, the in silico and bioinformatic analysis aided the identification and mapping of the resistance
mobilome and the T6SS gene clusters (T6SS1 and T6SS3) in K. pneumoniae to further enhanced insight
on how these virulence-associated regions shape the genome and pathogenicity of K. pneumoniae. Notably,
the content and context of T6SS gene clusters in K. pneumoniae are highly conserved and suggest a similar
common evolutionary event. The various tools within the genetic library constructed using a novel DNA
swapping technique provided a flexible and applicable range of molecular genetic tools for the deletion of
the mapped resistance mobilome or T6SS gene clusters in K. pneumoniae.
K. pneumoniae T6SS promoted antibacterial competitive fitness against other bacteria, horizontal
mobilization of mobile genetic elements such as plasmids, type 1 and 3 fimbria-mediate biofilm formation,
phagocytic and serum bactericidal resistance. The T6SS mediated anti-phagocytic virulence is probably
enable K. pneumoniae survival in the environment (soil and water) and host.
K. pneumoniae T6SS induces invasion, survival and actin filament / cytoskeletal rearrangement in host
cells. The various T6SS gene cluster stimulated differential expression pro-inflammatory innate immune
response in target host cells. Loss of T6SS gene clusters inhibited K. pneumoniae T6SS-induced
melanisation, in vivo bacterial competition and survival, and mortality in Galleria larva, thereby signifying
attenuated in vivo virulence stimulated by the T6SS mutants.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 7: Conclusion
224
The differential plasmid mobilization frequencies, antibacterial, in vitro and in vivo virulence mediated by
T6SS1 and T6SS3 gene clusters suggest that the various K. pneumoniae T6SS gene clusters within the same
genome may independently secrete different effectors for a different function. Also, mutation of T6SS gene
clusters that significantly impacted the expression of T4SS, type 1 and 3 fimbriae emphasizing the potential
tight co-regulation of other virulent factors and T6SS expression in K. pneumoniae.
7.2 Future work
As it is the case for most research work, many aspects of this research were limited by several factors such
as time, resources and technical expertise. These factors, together with other bacterial strain intrinsic
problem and limited knowledge on various aspects of this research, impeded the progress of the project.
Thus, the following aspects of the project will be investigated in the future to advance the understanding of
this research;
1. To optimize the novel DNA/gene swapping technique for maker-less chromosomal gene
complementation.
2. To design a novel single-step markerless deletion of the large pathogenic island (e,g T6SS) as
well as capture such large pathogenic islands in plasmid for T6SS mobilization and functional
studies.
3. To perform in-depth T6SS bioinformatic analysis and experimental assays to elucidate the
mechanism responsible for the co-regulation of how T6SS gene clusters and other virulence
factors.
4. To investigate the mechanism underpinning specific antibiotic resistance virulence in K.
pneumoniae and other bacteria.
5. To generate express Hcp1, VgrG1 and Hcp2 and VgrG3 protein of T6SS1 and T6SS3 clusters at
under various conditions.
6. To generate hcp1 vgrG1, hcp2 and vgrG3 mutant and assess their specific role in antibacterial and
in vivo virulence.
7. To tag expressing T6SS1 and T6SS3 with a separate fluorescent protein and capture T6SS in
action during interaction with other bacteria, amoeba and host cells.
8. To determine the various T6SS secreted effectors that are translocated with VgrG during
virulence-targeting of host and bacteria.
9. To design new assays to measure virulence mediated by T6SS and other virulent factors in
bacteria.

David Mwin Ph.D. Thesis, “The role of Type VI secretion system in the virulence of Klebsiella pneumoniae”,
University of Leicester, UK. Chapter 7: Conclusion
225
10. To measure T6SS mediated antibacterial virulence in gut, nasopharyngeal and urinary tract
complex microbial communities.
11. To assess bacterial T6SS mediated resistance to viral genomic integration and predation
12. Re-examine T6SS gene cluster in K. pneumoniae NTUH-K2044 to create T6SS3 and
T6SS1/T6SS3 mutants and perform functional studies in the hyper mucoid strain.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
226
Bibliography
Abbott S. (ed.) (2007) Klebsiella, Enterobacter, Citrobacter, and Serratia. In, Murray PR (Ed), Manual
of Clinical Microbiology
. 9th Ed edn. Washington DC: ASM Press.
Abbott, S. (2003) '44. Klebsiella, Enterobacter, Citrobacter, Serratia, Plesiomonas, and other
Enterobacteriaceae. W: Manual of clinical microbiology, Murray PR i in', ASM Press, Washington,
DC, 1, pp. 684-700.
Abe, Y., El-Masri, B., Kimball, K.T., Pownall, H., Reilly, C.F., Osmundsen, K., Smith, C.W. and
Ballantyne, C.M. (1998) 'Soluble cell adhesion molecules in hypertriglyceridemia and potential
significance on monocyte adhesion', Arteriosclerosis, Thrombosis, and Vascular Biology, 18(5), pp.
723-731.
Abrusci, P., McDowell, M.A., Lea, S.M. and Johnson, S. (2014) 'Building a secreting nanomachine: a
structural overview of the T3SS', Current opinion in structural biology, 25, pp. 111-117.
Adams, D.O. and Hamilton, T.A. (1984) 'The cell biology of macrophage activation', Annual Review of
Immunology, 2(1), pp. 283-318.
Ah, Y., Kim, A. and Lee, J. (2014) 'Colistin resistance in Klebsiella pneumoniae', International journal of
antimicrobial agents, 44(1), pp. 8-15.
Akira, S., Taga, T. and Kishimoto, T. (1993) 'Interleukin-6 in biology and medicine', in Anonymous
Advances in immunology. Elsevier, pp. 1-78.
Alam, R., Stafford, S., Forsythe, P., Harrison, R., Faubion, D., Lett-Brown, M.A. and Grant, J.A. (1993)
'RANTES is a chemotactic and activating factor for human eosinophils', Journal of immunology
(Baltimore, Md.: 1950), 150(8 Pt 1), pp. 3442-3448.
Alcántar-Curiel, M.D., Blackburn, D., Saldaña, Z., Gayosso-Vázquez, C., Iovine, N.M., De la Cruz, M.A.
and Girón, J.A. (2013) 'Multi-functional analysis of Klebsiella pneumoniae fimbrial types in
adherence and biofilm formation', Virulence, 4(2), pp. 129-138.
Alcoforado Diniz, J., Liu, Y. and Coulthurst, S.J. (2015) 'Molecular weaponry: diverse effectors delivered
by the Type VI secretion system', Cellular microbiology, 17(12), pp. 1742-1751.
Alcoforado Diniz, J. and Coulthurst, S.J. (2015) 'Intraspecies Competition in Serratia marcescens Is
Mediated by Type VI-Secreted Rhs Effectors and a Conserved Effector-Associated Accessory
Protein', Journal of Bacteriology, 197(14), pp. 2350-2360.
Al-Husseiny, K.R. and Nakkash, A.F. (2008) 'Study Some Virulence Factors of Klebsiella pneumoniae
Isolated From Clinical Sources', J.Thi-Qar Univ, 4(3), pp. 45-50.
Alsam, S., Jeong, S.R., Sissons, J., Dudley, R., Kim, K.S. and Khan, N.A. (2006) 'Escherichia coli
interactions with Acanthamoeba: a symbiosis with environmental and clinical implications', Journal
of medical microbiology, 55(6), pp. 689-694.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
227
Alteri, C.J., Himpsl, S.D., Pickens, S.R., Lindner, J.R., Zora, J.S., Miller, J.E., Arno, P.D., Straight, S.W.
and Mobley, H.L. (2013) 'Multicellular bacteria deploy the type VI secretion system to preemptively
strike neighboring cells', PLoS pathogens, 9(9), pp. e1003608.
Altincicek, B., Linder, M., Linder, D., Preissner, K.T. and Vilcinskas, A. (2007) 'Microbial
metalloproteinases mediate sensing of invading pathogens and activate innate immune responses in
the lepidopteran model host Galleria mellonella', Infection and immunity, 75(1), pp. 175-183.
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J., Zhang, Z., Miller, W. and Lipman, D.J. (1997)
'Gapped BLAST and PSI-BLAST: a new generation of protein database search programs', Nucleic
acids research, 25(17), pp. 3389-3402.
Alvarez-Martinez, C.E. and Christie, P.J. (2009) 'Biological diversity of prokaryotic type IV secretion
systems', Microbiology and molecular biology reviews : MMBR, 73(4), pp. 775-808.
Angiolillo, A.L., Sgadari, C., Taub, D.D., Liao, F., Farber, J.M., Maheshwari, S., Kleinman, H.K.,
Reaman, G.H. and Tosato, G. (1995) 'Human interferon-inducible protein 10 is a potent inhibitor of
angiogenesis in vivo', The Journal of experimental medicine, 182(1), pp. 155-162.
Appay, V. and Rowland-Jones, S.L. (2001) 'RANTES: a versatile and controversial chemokine', Trends
in immunology, 22(2), pp. 83-87.
Aschtgen, M., Thomas, M.S. and Cascales, E. (2010) 'Anchoring the type VI secretion system to the
peptidoglycan TssL, TagL, TagP,... what else?', Virulence, 1(6), pp. 535-540.
Aschtgen, M.S., Bernard, C.S., De Bentzmann, S., Lloubes, R. and Cascales, E. (2008) 'SciN is an outer
membrane lipoprotein required for type VI secretion in enteroaggregative Escherichia coli', Journal
of Bacteriology, 190(22), pp. 7523-7531.
Aubert, D.F., Flannagan, R.S. and Valvano, M.A. (2008a) 'A novel sensor kinase-response regulator
hybrid controls biofilm formation and type VI secretion system activity in Burkholderia
cenocepacia', Infection and immunity, 76(5), pp. 1979-1991.
BAER, H. and EHRENWORTH, L. (1956) 'The pathogenicity of Klebsiella pneumoniae for mice: the
relationship to the quantity and rate of production of type-specific capsular polysaccharide', Journal
of Bacteriology, 72(5), pp. 713-717.
Bagley, S.T. (1985) 'Habitat association of Klebsiella species', Infection control, 6(02), pp. 52-58.
Bailey, J.K., Pinyon, J.L., Anantham, S. and Hall, R.M. (2011) 'Distribution of the bla TEM gene and bla
TEM-containing transposons in commensal Escherichia coli', Journal of antimicrobial
chemotherapy, 66(4), pp. 745-751.
Balakrishnan, L., Hughes, C. and Koronakis, V. (2001) 'Substrate-triggered recruitment of the TolC
channel-tunnel during type I export of hemolysin by Escherichia coli', Journal of Molecular
Biology, 313(3), pp. 501-510.
Baldi, D.L., Higginson, E.E., Hocking, D.M., Praszkier, J., Cavaliere, R., James, C.E., Bennett-Wood, V.,
Azzopardi, K.I., Turnbull, L., Lithgow, T., Robins-Browne, R.M., Whitchurch, C.B. and Tauschek,

David Mwin Ph.D. Thesis, University of Leicester, UK. References
228
M. (2012) 'The type II secretion system and its ubiquitous lipoprotein substrate, SslE, are required
for biofilm formation and virulence of enteropathogenic Escherichia coli', Infection and immunity,
80(6), pp. 2042-2052.
Ballister, E.R., Lai, A.H., Zuckermann, R.N., Cheng, Y. and Mougous, J.D. (2008) 'In vitro self-assembly
of tailorable nanotubes from a simple protein building block', Proceedings of the National Academy
of Sciences of the United States of America, 105(10), pp. 3733-3738.
Ballows, A., Truper, H., Dworkin, M., Harder, W. and Schleifer, K. (1992) 'The prokaryotes, a handbook
on the biology of bacteria: Ecophysiology, isolation, identification, applications', .
Bals, R. and Hiemstra, P.S. (2004) 'Innate immunity in the lung: how epithelial cells fight against
respiratory pathogens', The European respiratory journal, 23(2), pp. 327-333.
Banville, N., Browne, N. and Kavanagh, K. (2012) 'Effect of nutrient deprivation on the susceptibility of
Galleria mellonella larvae to infection', Virulence, 3(6), pp. 497-503.
Barret, M., Egan, F., Fargier, E., Morrissey, J.P. and O'Gara, F. (2011) 'Genomic analysis of the type VI
secretion systems in Pseudomonas spp.: Novel clusters and putative effectors uncovered',
Microbiology, 157(6), pp. 1726-1739.
Bartonickova, L., Sterzenbach, T., Nell, S., Kops, F., Schulze, J., Venzke, A., Brenneke, B., Bader, S.,
Gruber, A.D., Suerbaum, S. and Josenhans, C. (2013) 'Hcp and VgrG1 are secreted components of
the Helicobacter hepaticus type VI secretion system and VgrG1 increases the bacterial colitogenic
potential', Cellular microbiology, 15(6), pp. 992-1011.
Basler, á, Pilhofer, M., Henderson, G., Jensen, G. and Mekalanos, J. (2012a) 'Type VI secretion requires a
dynamic contractile phage tail-like structure', Nature, 483(7388), pp. 182-186.
Basler, M. and Mekalanos, J.J. (2012) 'Type 6 secretion dynamics within and between bacterial cells',
Science (New York, N.Y.), 337(6096), pp. 815.
Basler, M., Pilhofer, M., Henderson, G.P., Jensen, G.J. and Mekalanos, J.J. (2012b) 'Type VI secretion
requires a dynamic contractile phage tail-like structure', Nature, 483(7388), pp. 182.
Beceiro, A., Tomas, M. and Bou, G. (2013) 'Antimicrobial resistance and virulence: a successful or
deleterious association in the bacterial world?', Clinical microbiology reviews, 26(2), pp. 185-230.
Beeckman, D. and Vanrompay, D. (2010) 'Bacterial secretion systems with an emphasis on the
chlamydial Type III secretion system', Curr Issues Mol Biol, 12(1), pp. 17-41.
Benge, G.R. (1988) 'Bactericidal activity of human serum against strains of Klebsiella from different
sources', Journal of medical microbiology, 27(1), pp. 11.
Bergey, D.H., Harrison F.C., Breed, R.S., Hammer, B.W. and Huntoon F.M. (1925) Bergey’s Manual of
Determinative Bacteriology. 1st edn. Baltimore, MD: W illiams and Wilkins.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
229
Bergin, D., Reeves, E.P., Renwick, J., Wientjes, F.B. and Kavanagh, K. (2005) 'Superoxide production in
Galleria mellonella hemocytes: identification of proteins homologous to the NADPH oxidase
complex of human neutrophils', Infection and immunity, 73(7), pp. 4161-4170.
Bernard, C.S., Brunet, Y.R., Gavioli, M., Lloubès, R. and Cascales, E. (2011) 'Regulation of type VI
Secretion gene clusters by σ 54 and cognate enhancer binding proteins', Journal of Bacteriology,
193(9), pp. 2158-2167.
Bernstein, H.D. (2007) 'Are bacterial ‘autotransporters’ really transporters?', Trends in microbiology,
15(10), pp. 441-447.
Besemer, J., Lomsadze, A. and Borodovsky, M. (2001) 'GeneMarkS: a self-training method for prediction
of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions',
Nucleic acids research, 29(12), pp. 2607.
Beyrouthy, R., Robin, F., Dabboussi, F., Mallat, H., Hamze, M. and Bonnet, R. (2014) 'Carbapenemase
and virulence factors of Enterobacteriaceae in North Lebanon between 2008 and 2012: evolution via
endemic spread of OXA-48', The Journal of antimicrobial chemotherapy, 69(10), pp. 2699-2705.
Bhatty, M., Gomez, J.A.L. and Christie, P.J. (2013) 'The expanding bacterial type IV secretion lexicon',
Research in microbiology, 164(6), pp. 620-639.
Bi, D., Jiang, X., Sheng, Z.K., Ngmenterebo, D., Tai, C., Wang, M., Deng, Z., Rajakumar, K. and Ou,
H.Y. (2015) 'Mapping the resistance-associated mobilome of a carbapenem-resistant Klebsiella
pneumoniae strain reveals insights into factors shaping these regions and facilitates generation of a
'resistance-disarmed' model organism', The Journal of antimicrobial chemotherapy, .
Bi, D., Xu, Z., Harrison, E.M., Tai, C., Wei, Y., He, X., Jia, S., Deng, Z., Rajakumar, K. and Ou, H.Y.
(2012) 'ICEberg: a web-based resource for integrative and conjugative elements found in Bacteria',
Nucleic acids research, 40(Database issue), pp. D621-6.
Bialek-Davenet, S., Lavigne, J.P., Guyot, K., Mayer, N., Tournebize, R., Brisse, S., Leflon-Guibout, V.
and Nicolas-Chanoine, M.H. (2015) 'Differential contribution of AcrAB and OqxAB efflux pumps
to multidrug resistance and virulence in Klebsiella pneumoniae', The Journal of antimicrobial
chemotherapy, 70(1), pp. 81-88.
Bikandi, J., San Millán, R., Rementeria, A. and Garaizar, J. (2004) 'In silico analysis of complete bacterial
genomes: PCR, AFLP-PCR and endonuclease restriction', Bioinformatics (Oxford, England), 20(5),
pp. 798.
Bingle, L.E., Bailey, C.M. and Pallen, M.J. (2008) 'Type VI secretion: a beginner's guide', Current
opinion in microbiology, 11(1), pp. 3-8.
Bistué, A.J.S., Birshan, D., Tomaras, A.P., Dandekar, M., Tran, T., Newmark, J., Bui, D., Gupta, N.,
Hernandez, K. and Sarno, R. (2008) 'Klebsiella pneumoniae multiresistance plasmid pMET1:
similarity with the Yersinia pestis plasmid pCRY and integrative conjugative elements', PloS one,
3(3), pp. e1800.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
230
Bladergroen, M., Badelt, K. and Spaink, H. (2003) 'Infection-blocking genes of a symbiotic Rhizobium
leguminosarum strain that are involved in temperature-dependent protein secretion', Molecular
Plant-Microbe Interactions, 16(1), pp. 53-64.
Bleves, S., Viarre, V., Salacha, R., Michel, G.P.F., Filloux, A. and Voulhoux, R. (2010) 'Protein secretion
systems in Pseudomonas aeruginosa: A wealth of pathogenic weapons', International Journal of
Medical Microbiology, 300(8), pp. 534-543.
Blokesch, M. (2015a) 'Competence‐induced type VI secretion might foster intestinal colonization by
Vibrio cholerae', BioEssays, 37(11), pp. 1163-1168.
Blokesch, M. (2015b) 'Competence-induced type VI secretion might foster intestinal colonization by
Vibrio cholerae: Intestinal interbacterial killing by competence-induced V. cholerae', BioEssays :
news and reviews in molecular, cellular and developmental biology, 37(11), pp. 1163.
Blondel, C.J., Yang, H.-., Castro, B., Chiang, S., Toro, C.S., Zaldïvar, M., Contreras, I., Andrews-
Polymenis, H.L. and Santiviago, C.A. (2010) 'Contribution of the type VI secretion system encoded
in SPI-19 to chicken colonization by Salmonella enterica serotypes Gallinarum and enteritidis',
PLoS ONE, 5(7).
Boddicker, J.D., Anderson, R.A., Jagnow, J. and Clegg, S. (2006) 'Signature-Tagged Mutagenesis of
Klebsiella pneumoniae To Identify Genes That Influence Biofilm Formation on Extracellular Matrix
Material', Infection and immunity, 74(8), pp. 4590-4597.
Bönemann, G., Pietrosiuk, A. and Mogk, A. (2010) 'Tubules and donuts: A type VI secretion story:
MicroReview', Molecular microbiology, 76(4), pp. 815-821.
Borgeaud, S., Metzger, L.C., Scrignari, T. and Blokesch, M. (2015a) 'The type VI secretion system of
Vibrio cholerae fosters horizontal gene transfer', Science, 347(6217), pp. 63-67.
Borgeaud, S., Metzger, L.C., Scrignari, T. and Blokesch, M. (2015b) 'The type VI secretion system of
Vibrio cholerae fosters horizontal gene transfer', Science, 347(6217), pp. 63-67.
Borgeaud, S., Metzger, L.C., Scrignari, T. and Blokesch, M. (2015c) 'The type VI secretion system of
Vibrio cholerae fosters horizontal gene transfer', Science, 347(6217), pp. 63-67.
Boucher, H.W., Talbot, G.H., Bradley, J.S., Edwards, J.E., Gilbert, D., Rice, L.B., Scheld, M., Spellberg,
B. and Bartlett, J. (2009) 'Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases
Society of America', Clinical infectious diseases : an official publication of the Infectious Diseases
Society of America, 48(1), pp. 1-12.
Boye, K. and Hansen, D.S. (2003) 'Sequencing of 16S rDNA of Klebsiella: taxonomic relations within
the genus and to other Enterobacteriaceae', International journal of medical microbiology, 292(7),
pp. 495-503.
Boyer, F., Fichant, G., Berthod, J., Vandenbrouck, Y. and Attree, I. (2009a) 'Dissecting the bacterial type
VI secretion system by a genome wide in silico analysis: what can be learned from available
microbial genomic resources?', BMC genomics, 10, pp. 104-2164-10-104.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
231
Boyer, F., Fichant, G., Berthod, J., Vandenbrouck, Y. and Attree, I. (2009b) 'Dissecting the bacterial type
VI secretion system by a genome wide in silico analysis: what can be learned from available
microbial genomic resources?', BMC genomics, 10, pp. 104-2164-10-104.
Boyer, W.H. and Roullard Dessoix, D. (1969) Journal of Molecular Biology, 41, pp. 459-472.
Braiteh, F. and Golden, M.P. (2007) 'Cryptogenic invasive Klebsiella pneumoniae liver abscess
syndrome', International journal of infectious diseases, 11(1), pp. 16-22.
Brandon, L.D., Goehring, N., Janakiraman, A., Yan, A.W., Wu, T., Beckwith, J. and Goldberg, M.B.
(2003) 'IcsA, a polarly localized autotransporter with an atypical signal peptide, uses the Sec
apparatus for secretion, although the Sec apparatus is circumferentially distributed', Molecular
microbiology, 50(1), pp. 45-60.
Brisse, S., Grimont, F. and Grimont, P.A. (2006) 'The Genus Klebsiella', in Anonymous The Prokaryotes.
Springer, pp. 159-196.
Brisse, S. and Verhoef, J. (2001a) 'Phylogenetic diversity of Klebsiella pneumoniae and Klebsiella
oxytoca clinical isolates revealed by randomly amplified polymorphic DNA, gyrA and parC genes
sequencing and automated ribotyping.', International Journal of Systematic and Evolutionary
Microbiology, 51(3), pp. 915-924.
Brisse, S. and Verhoef, J. (2001b) 'Phylogenetic diversity of Klebsiella pneumoniae and Klebsiella
oxytoca clinical isolates revealed by randomly amplified polymorphic DNA, gyrA and parC genes
sequencing and automated ribotyping.', International Journal of Systematic and Evolutionary
Microbiology, 51(3), pp. 915-924.
Brisse, S., Issenhuth-Jeanjean, S. and Grimont, P.A.D. (2004) 'Molecular Serotyping of Klebsiella
Species Isolates by Restriction of the Amplified Capsular Antigen Gene Cluster', Journal of clinical
microbiology, 42(8), pp. 3388.
Brito, B.E., O’Rourke, L.M., Pan, Y., Anglin, J., Planck, S.R. and Rosenbaum, J.T. (1999) 'IL-1 and TNF
receptor–deficient mice show decreased inflammation in an immune complex model of uveitis',
Investigative ophthalmology & visual science, 40(11), pp. 2583-2589.
Brock, J.H., Williams, P.H., Liceaga, J. and Wooldridge, K.G. (1991) 'Relative availability of transferrin-
bound iron and cell-derived iron to aerobactin-producing and enterochelin-producing strains of
Escherichia coli and to other microorganisms', Infection and immunity, 59(9), pp. 3185-3190.
Brodmann, M., Dreier, R.F., Broz, P. and Basler, M. (2017) 'Francisella requires dynamic type VI
secretion system and ClpB to deliver effectors for phagosomal escape', Nature communications, 8,
pp. 15853.
Bröms, J.E., Meyer, L., Lavander, M., Larsson, P. and Sjöstedt, A. (2012a) 'DotU and VgrG, core
components of type VI secretion systems, are essential for Francisella LVS pathogenicity', PLoS
ONE, 7(4).

David Mwin Ph.D. Thesis, University of Leicester, UK. References
232
Bröms, J.E., Meyer, L., Sun, K., Lavander, M. and Sjöstedt, A. (2012b) 'Unique Substrates Secreted by
the Type VI Secretion System of Francisella tularensis during Intramacrophage Infection', PLoS
ONE, 7(11).
Brooks, T.M., Unterweger, D., Bachmann, V., Kostiuk, B. and Pukatzki, S. (2013) 'Lytic activity of the
Vibrio cholerae type VI secretion toxin VgrG-3 is inhibited by the antitoxin TsaB', The Journal of
biological chemistry, 288(11), pp. 7618-7625.
Broug-Holub, E., Toews, G.B., van Iwaarden, J.F., Strieter, R.M., Kunkel, S.L., Paine, R.,3rd and
Standiford, T.J. (1997) 'Alveolar macrophages are required for protective pulmonary defenses in
murine Klebsiella pneumoniae: elimination of alveolar macrophages increases neutrophil
recruitment but decreases bacterial clearance and survival', Infection and immunity, 65(4), pp. 1139-
1146.
Brown, C. and Seidler, R.J. (1973) 'Potential pathogens in the environment: Klebsiella pneumoniae, a
taxonomic and ecological enigma', Applied Microbiology, 25(6), pp. 900-904.
Browne, N., Surlis, C., Maher, A., Gallagher, C., Carolan, J.C., Clynes, M. and Kavanagh, K. (2015)
'Prolonged pre-incubation increases the susceptibility of Galleria mellonella larvae to bacterial and
fungal infection', Virulence, 6(5), pp. 458-465.
Brunet, Y.R., Henin, J., Celia, H. and Cascales, E. (2014) 'Type VI secretion and bacteriophage tail tubes
share a common assembly pathway', EMBO reports, 15(3), pp. 315-321.
Brunet, Y.R., Espinosa, L., Harchouni, S., Mignot, T. and Cascales, E. (2013a) 'Imaging type VI
secretion-mediated bacterial killing', Cell reports, 3(1), pp. 36-41.
Brunet, Y.R., Espinosa, L., Harchouni, S., Mignot, T. and Cascales, E. (2013b) 'Imaging type VI
secretion-mediated bacterial killing', Cell reports, 3(1), pp. 36-41.
Bullen, J., Rogers, H.J. and Griffiths, E. (1978) 'Role of iron in bacterial infection', in Anonymous
Current topics in microbiology and immunology. Springer, pp. 1-35.
Bullen, J.J., Rogers, H.J., Spalding, P.B. and Ward, C.G. (2006) 'Natural resistance, iron and infection: a
challenge for clinical medicine', Journal of medical microbiology, 55(3), pp. 251-258.
Bullen, J., Griffiths, E., Rogers, H. and Ward, G. (2000) 'Sepsis: the critical role of iron', Microbes and
Infection, 2(4), pp. 409-415.
Buré, A., Legrand, P., Arlet, G., Jarlier, V., Paul, G. and Philippon, A. (1988) 'Dissemination in five
French hospitals of Klebsiella pneumoniae serotype K25 harbouring a new transferable enzymatic
resistance to third generation cephalosporins and aztreonam', European Journal of Clinical
Microbiology and Infectious Diseases, 7(6), pp. 780-782.
Burke, B. and Lewis, C.E. (2002) 'The macrophage', .
Burkinshaw, B.J. and Strynadka, N.C. (2014) 'Assembly and structure of the T3SS', Biochimica et
Biophysica Acta (BBA)-Molecular Cell Research, 1843(8), pp. 1649-1663.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
233
Burtnick, M.N., Brett, P.J., Harding, S.V., Ngugi, S.A., Ribot, W.J., Chantratita, N., Scorpio, A., Milne,
T.S., Dean, R.E., Fritz, D.L., Peacock, S.J., Prior, J.L., Atkins, T.P. and DeShazer, D. (2011) 'The
cluster 1 type VI secretion system is a major virulence determinant in Burkholderia pseudomallei',
Infection and immunity, 79(4), pp. 1512-1525.
Burtnick, M.N., DeShazer, D., Nair, V., Gherardini, F.C. and Brett, P.J. (2010) 'Burkholderia mallei
cluster 1 type VI secretion mutants exhibit growth and actin polymerization defects in RAW 264.7
murine macrophages', Infection and immunity, 78(1), pp. 88-99.
Bush, K. (2012) 'Improving known classes of antibiotics: an optimistic approach for the future', Current
opinion in pharmacology, 12(5), pp. 527-534.
Buttner, D. (2012) 'Protein export according to schedule: architecture, assembly, and regulation of type III
secretion systems from plant- and animal-pathogenic bacteria', Microbiology and molecular biology
reviews : MMBR, 76(2), pp. 262-310.
Bystry, R.S., Aluvihare, V., Welch, K.A., Kallikourdis, M. and Betz, A.G. (2001) 'B cells and
professional APCs recruit regulatory T cells via CCL4', Nature immunology, 2(12), pp. 1126.
Cain, A.K. and Hall, R.M. (2012) 'Evolution of a multiple antibiotic resistance region in IncHI1 plasmids:
reshaping resistance regions in situ', Journal of antimicrobial chemotherapy, 67(12), pp. 2848-2853.
Calandra, T. and Roger, T. (2003) 'Macrophage migration inhibitory factor: a regulator of innate
immunity', Nature reviews immunology, 3(10), pp. 791.
Calhau, Vera Mónica Tavares Vinhas (2015) 'Virulence factors associated with antimicrobial resistance
determinants among Escherichia coli and Klebsiella spp', .
Campos, M.A., Vargas, M.A., Regueiro, V., Llompart, C.M., Alberti, S. and Bengoechea, J.A. (2004a)
'Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides', Infection and
immunity, 72(12), pp. 7107-7114.
Campos, M.A., Vargas, M.A., Regueiro, V., Llompart, C.M., Alberti, S. and Bengoechea, J.A. (2004b)
'Capsule Polysaccharide Mediates Bacterial Resistance to Antimicrobial Peptides', Infection and
immunity, 72(12), pp. 7107.
Cannatelli, A., D'Andrea, M.M., Giani, T., Di Pilato, V., Arena, F., Ambretti, S., Gaibani, P. and
Rossolini, G.M. (2013) 'In vivo emergence of colistin resistance in Klebsiella pneumoniae producing
KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator',
Antimicrobial Agents and Chemotherapy, 57(11), pp. 5521-5526.
Caplenas, N.R., Kanarek, M.S. and Dufour, A.P. (1981) 'Source and extent of Klebsiella pneumoniae in
the paper industry', Applied and Environmental Microbiology, 42(5), pp. 779-785.
Carpenter, J.L. (1990) 'Klebsiella pulmonary infections: occurrence at one medical center and review',
Review of Infectious Diseases, 12(4), pp. 672-682.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
234
Carr, M.W., Roth, S.J., Luther, E., Rose, S.S. and Springer, T.A. (1994) 'Monocyte chemoattractant
protein 1 acts as a T-lymphocyte chemoattractant', Proceedings of the National Academy of Sciences
of the United States of America, 91(9), pp. 3652-3656.
Carruthers, M.D., Nicholson, P.A., Tracy, E.N. and Munson, J.,Robert S. (2013) 'Acinetobacter
baumannii utilizes a type VI secretion system for bacterial competition', PloS one, 8(3), pp. e59388.
Cascales, E. (2008) 'The type VI secretion toolkit', EMBO reports, 9(8), pp. 735-741.
Cascales, E. and Cambillau, C. (2012a) 'Structural biology of type VI secretion systems', Philosophical
transactions of the Royal Society of London.Series B, Biological sciences, 367(1592), pp. 1102-
1111.
Cascales, E. and Cambillau, C. (2012b) 'Structural biology of type VI secretion systems', Philosophical
Transactions of the Royal Society B: Biological Sciences, 367(1592), pp. 1102-1111.
Chang, S., Fang, C., Hsueh, P., Chen, Y. and Luh, K. (2000) 'Klebsiella pneumoniae isolates causing liver
abscess in Taiwan', Diagnostic microbiology and infectious disease, 37(4), pp. 279-284.
Chang, F.Y. and Chou, M.Y. (1995) 'Comparison of pyogenic liver abscesses caused by Klebsiella
pneumoniae and non-K. pneumoniae pathogens', Journal of the Formosan Medical Association =
Taiwan yi zhi, 94(5), pp. 232-237.
Chaplin, D.D. (2010) 'Overview of the immune response', Journal of Allergy and Clinical Immunology,
125(2), pp. S3-S23.
Chaplin, D.D. (2006) '1. Overview of the human immune response', Journal of allergy and clinical
immunology, 117(2), pp. S430-S435.
Chaveroche, M., Ghigo, J. and d’Enfert, C. (2000) 'A rapid method for efficient gene replacement in the
filamentous fungus Aspergillus nidulans', Nucleic acids research, 28(22), pp. e97-e97.
Chawla, R. (2008) 'Epidemiology, etiology, and diagnosis of hospital-acquired pneumonia and ventilator-
associated pneumonia in Asian countries', American Journal of Infection Control, 36(4), pp. S93-
S100.
Chekabab, S.M., Daigle, F., Charette, S.J., Dozois, C.M. and Harel, J. (2012) 'Survival of
enterohemorrhagic Escherichia coli in the presence of Acanthamoeba castellanii and its dependence
on Pho regulon', MicrobiologyOpen, 1(4), pp. 427-437.
Chen, Z., Liu, M., Cui, Y., Wang, L., Zhang, Y., Qiu, J., Yang, R., Liu, C. and Zhou, D. (2014a) 'A novel
PCR-based genotyping scheme for clinical Klebsiella pneumoniae', Future microbiology, 9(1), pp.
21-32.
Chen, J.H., Siu, L.K., Fung, C.P., Lin, J.C., Yeh, K.M., Chen, T.L., Tsai, Y.K. and Chang, F.Y. (2010)
'Contribution of outer membrane protein K36 to antimicrobial resistance and virulence in Klebsiella
pneumoniae', The Journal of antimicrobial chemotherapy, 65(5), pp. 986-990.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
235
Chen, L., Mathema, B., Pitout, J.D., DeLeo, F.R. and Kreiswirth, B.N. (2014b) 'Epidemic Klebsiella
pneumoniae ST258 is a hybrid strain', mBio, 5(3), pp. e01355-14.
Chen, L., Zou, Y., She, P. and Wu, Y. (2015) 'Composition, function, and regulation of T6SS in
Pseudomonas aeruginosa', Microbiological research, 172, pp. 19.
Cheng, W., Shivshankar, P., Zhong, Y., Chen, D., Li, Z. and Zhong, G. (2008) 'Intracellular interleukin-
1alpha mediates interleukin-8 production induced by Chlamydia trachomatis infection via a
mechanism independent of type I interleukin-1 receptor', Infection and immunity, 76(3), pp. 942-951.
Chieng, S., Mohamed, R. and Nathan, S. (2015) 'Transcriptome analysis of Burkholderia pseudomallei
T6SS identifies Hcp1 as a potential serodiagnostic marker', Microbial pathogenesis, 79, pp. 47-56.
Chiu, C., Lin, D. and Liaw, Y. (1988) 'Metastatic septic endophthalmitis in pyogenic liver abscess.',
Journal of clinical gastroenterology, 10(5), pp. 524-527.
Choi, K.H., Gaynor, J.B., White, K.G., Lopez, C., Bosio, C.M., Karkhoff-Schweizer, R.R. and Schweizer,
H.P. (2005) 'A Tn7-based broad-range bacterial cloning and expression system', Nature methods,
2(6), pp. 443-448.
Choi, M.J. and Ko, K.S. (2015) 'Loss of hypermucoviscosity and increased fitness cost in colistin-
resistant Klebsiella pneumoniae sequence type 23 strains', Antimicrobial Agents and Chemotherapy,
59(11), pp. 6763-6773.
Chou, H.C., Lee, C.Z., Ma, L.C., Fang, C.T., Chang, S.C. and Wang, J.T. (2004) 'Isolation of a
chromosomal region of Klebsiella pneumoniae associated with allantoin metabolism and liver
infection', Infection and immunity, 72(7), pp. 3783-3792.
Chow, J. and Mazmanian, S.K. (2010) 'A pathobiont of the microbiota balances host colonization and
intestinal inflammation', Cell Host and Microbe, 7(4), pp. 265-276.
Christie, P.J. and Cascales, E. (2005) 'Structural and dynamic properties of bacterial type IV secretion
systems (review)', Molecular membrane biology, 22(1-2), pp. 51-61.
Christie, P.J., Whitaker, N. and González-Rivera, C. (2014) 'Mechanism and structure of the bacterial
type IV secretion systems', Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 1843(8),
pp. 1578-1591.
Chung, D., Lee, S., Lee, H., Kim, H., Choi, H., Eom, J., Kim, J., Choi, Y., Lee, J. and Chung, M. (2007)
'Emerging invasive liver abscess caused by K1 serotype Klebsiella pneumoniae in Korea', Journal of
Infection, 54(6), pp. 578-583.
Cianciotto, N.P. (2013) 'Type II secretion and Legionella virulence', in Anonymous Molecular
Mechanisms in Legionella Pathogenesis. Springer, pp. 81-102.
Cianciotto, N.P. (2005) 'Type II secretion: a protein secretion system for all seasons', Trends in
microbiology, 13(12), pp. 581-588.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
236
Clements, A., Tull, D., Jenney, A.W., Farn, J.L., Kim, S.H., Bishop, R.E., McPhee, J.B., Hancock, R.E.,
Hartland, E.L., Pearse, M.J., Wijburg, O.L., Jackson, D.C., McConville, M.J. and Strugnell, R.A.
(2007) 'Secondary acylation of Klebsiella pneumoniae lipopolysaccharide contributes to sensitivity
to antibacterial peptides', The Journal of biological chemistry, 282(21), pp. 15569-15577.
Cornelis, G.R. (2006) 'The type III secretion injectisome', Nature Reviews Microbiology, 4(11), pp. 811-
825.
Cortes, G., Alvarez, D., Saus, C. and Alberti, S. (2002a) 'Role of lung epithelial cells in defense against
Klebsiella pneumoniae pneumonia', Infection and immunity, 70(3), pp. 1075-1080.
Cortes, G., Borrell, N., de Astorza, B., Gomez, C., Sauleda, J. and Alberti, S. (2002b) 'Molecular analysis
of the contribution of the capsular polysaccharide and the lipopolysaccharide O side chain to the
virulence of Klebsiella pneumoniae in a murine model of pneumonia', Infection and immunity, 70(5),
pp. 2583-2590.
Costa, T.R., Felisberto-Rodrigues, C., Meir, A., Prevost, M.S., Redzej, A., Trokter, M. and Waksman, G.
(2015) 'Secretion systems in Gram-negative bacteria: structural and mechanistic insights', Nature
reviews.Microbiology, 13(6), pp. 343-359.
Cotton, M., Wasserman, E., Pieper, C., Theron, D., Van Tubbergh, D., Campbell, G., Fang, F. and
Barnes, J. (2000) 'Invasive disease due to extended spectrum beta-lactamase-producing Klebsiella
pneumoniae in a neonatal unit: the possible role of cockroaches', Journal of Hospital Infection,
44(1), pp. 13-17.
Coulthurst, S.J. (2013) 'The Type VI secretion system - a widespread and versatile cell targeting system',
Research in microbiology, 164(6), pp. 640.
Cox, B.L., Schiffer, H., Dagget Jr, G., Beierschmitt, A., Sithole, F., Lee, E., Revan, F., Halliday-
Simmonds, I., Beeler-Marfisi, J. and Palmour, R. (2015) 'Resistance of Klebsiella pneumoniae to the
innate immune system of African green monkeys', Veterinary microbiology, 176(1-2), pp. 134-142.
Craig-Mylius, K.A. and Weiss, A.A. (1999) 'Mutants in the ptlA-H genes of Bordetella pertussis are
deficient for pertussis toxin secretion', FEMS microbiology letters, 179(2), pp. 479-484.
Cryz, S.J.,Jr, Furer, F. and Germanier, R. (1984) 'Experimental Klebsiella pneumoniae burn wound
sepsis: role of capsular polysaccharide', Infection and immunity, 43(1), pp. 440-441.
Cryz, S.J.,Jr, Mortimer, P.M., Mansfield, V. and Germanier, R. (1986) 'Seroepidemiology of Klebsiella
bacteremic isolates and implications for vaccine development', Journal of clinical microbiology,
23(4), pp. 687-690.
Cubitt, C.L., Lausch, R.N. and Oakes, J.E. (1997) 'Differential induction of GRO alpha gene expression
in human corneal epithelial cells and keratocytes exposed to proinflammatory cytokines.',
Investigative ophthalmology & visual science, 38(6), pp. 1149-1158.
da Silva, F.G., Shen, Y., Dardick, C., Burdman, S., Yadav, R.C., de Leon, A.L. and Ronald, P.C. (2004)
'Bacterial genes involved in type I secretion and sulfation are required to elicit the rice Xa21-
mediated innate immune response', Molecular Plant-Microbe Interactions, 17(6), pp. 593-601.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
237
da Silva, G.J. and Mendonça, N. (2012) 'Association between antimicrobial resistance and virulence in
Escherichia coli', Virulence, 3(1), pp. 18-28.
Dalbey, R.E. and Kuhn, A. (2012) 'Protein traffic in Gram-negative bacteria--how exported and secreted
proteins find their way', FEMS microbiology reviews, 36(6), pp. 1023-1045.
Darfeuille-Michaud, A., Jallat, C., Aubel, D., Sirot, D., Rich, C., Sirot, J. and Joly, B. (1992) 'R-plasmid-
encoded adhesive factor in Klebsiella pneumoniae strains responsible for human nosocomial
infections', Infection and immunity, 60(1), pp. 44-55.
Das, S. and Chaudhuri, K. (2003) 'Identification of a unique IAHP (IcmF associated homologous
proteins) cluster in Vibrio cholerae and other proteobacteria through in silico analysis', In silico
biology, 3(3), pp. 287-300.
Das, S., Chakrabortty, A., Banerjee, R. and Chaudhuri, K. (2002) 'Involvement of in vivo induced icmF
gene of Vibrio cholerae in motility, adherence to epithelial cells, and conjugation frequency',
Biochemical and biophysical research communications, 295(4), pp. 922-928.
Datsenko, K.A. and Wanner, B.L. (2000) 'One-step inactivation of chromosomal genes in Escherichia
coli K-12 using PCR products', Proceedings of the National Academy of Sciences of the United
States of America, 97(12), pp. 6640.
Davidson, N. and Szybalski, W. (1971) 'In The Bacteriophage Lambda (Hershey, AD, ed.)', Cold Spring
Harbor Laboratory, New York.DNA CONDENSATION WITH POLYAMINES.II, 337, pp. 45-82.
Davis, G. and Price, L. (2016) 'Recent Research Examining Links Among Klebsiella pneumoniae from
Food, Food Animals, and Human Extraintestinal Infections', Current Environmental Health Reports,
3(2), pp. 128-135.
de Astorza, B., Cortes, G., Crespi, C., Saus, C., Rojo, J.M. and Alberti, S. (2004) 'C3 promotes clearance
of Klebsiella pneumoniae by A549 epithelial cells', Infection and immunity, 72(3), pp. 1767-1774.
de Bruin, O.M., Duplantis, B.N., Ludu, J.S., Hare, R.F., Nix, E.B., Schmerk, C.L., Robb, C.S., Boraston,
A.B., Hueffer, K. and Nano, F.E. (2011) 'The biochemical properties of the Francisella
pathogenicity island (FPI)-encoded proteins IglA, IglB, IglC, PdpB and DotU suggest roles in type
VI secretion', Microbiology, 157(12), pp. 3483-3491.
De Bruin, O.M., Ludu, J.S. and Nano, F.E. (2007) 'The Francisella pathogenicity island protein IglA
localizes to the bacterial cytoplasm and is needed for intracellular growth', BMC microbiology, 7(1),
pp. 1.
De Ceuleneer, M., Vanhoucke, M. and Beyaert, R. (2015) 'Role of the bacterial type VI secretion system
in the modulation of mammalian host cell immunity', Current medicinal chemistry, 22(14), pp.
1734-1744.
De Jesus, M.B., Ehlers, M.M., Dos Santos, R.F. and Kock, M.M. (2015) 'Review-Understanding β-
lactamase Producing Klebsiella pneumoniae', .

David Mwin Ph.D. Thesis, University of Leicester, UK. References
238
de Lorenzo, V., Cases, I., Herrero, M. and Timmis, K.N. (1993) 'Early and late responses of TOL
promoters to pathway inducers: identification of postexponential promoters in Pseudomonas putida
with lacZ-tet bicistronic reporters', Journal of Bacteriology, 175(21), pp. 6902-6907.
De Moraes, J. and Alfieri, S.C. (2008) 'Growth, encystment and survival of Acanthamoeba castellanii
grazing on different bacteria', FEMS microbiology ecology, 66(2), pp. 221-229.
De Pace, F., Nakazato, G., Pacheco, A., De Paiva, J.B., Sperandio, V. and Da Silveira, W.D. (2010) 'The
type VI secretion system plays a role in type 1 fimbria expression and pathogenesis of an avian
pathogenic Escherichia coli strain', Infection and immunity, 78(12), pp. 4990-4998.
Deane, J.E., Cordes, F.S., Roversi, P., Johnson, S., Kenjale, R., Picking, W.D., Picking, W.L., Lea, S.M.
and Blocker, A. (2006) 'Expression, purification, crystallization and preliminary crystallographic
analysis of MxiH, a subunit of the Shigella flexneri type III secretion system needle', Acta
Crystallographica Section F: Structural Biology and Crystallization Communications, 62(3), pp.
302-305.
Decoin, V., Gallique, M., Barbey, C., Le Mauff, F., Poc, C.D., Feuilloley, M.G., Orange, N. and Merieau,
A. (2015) 'A Pseudomonas fluorescens type 6 secretion system is related to mucoidy, motility and
bacterial competition', BMC microbiology, 15, pp. 72-015-0405-9.
Decoin, V., Barbey, C., Bergeau, D., Latour, X., Feuilloley, M.G.J., Orange, N. and Merieau, A. (2014a)
'A type VI secretion system is involved in Pseudomonas fluorescens bacterial competition', PloS
one, 9(2), pp. e89411.
Decoin, V., Barbey, C., Bergeau, D., Latour, X., Marc G J Feuilloley, Orange, N. and Merieau, A.
(2014b) 'A Type VI Secretion System Is Involved in Pseudomonas fluorescens Bacterial
Competition: e89411', PLoS One, 9(2).
Deguchi, T., Fukuoka, A., Yasuda, M., Nakano, M., Ozeki, S., Kanematsu, E., Nishino, Y., Ishihara, S.,
Ban, Y. and Kawada, Y. (1997) 'Alterations in the GyrA subunit of DNA gyrase and the ParC
subunit of topoisomerase IV in quinolone-resistant clinical isolates of Klebsiella pneumoniae',
Antimicrobial Agents and Chemotherapy, 41(3), pp. 699-701.
Delepelaire, P. (2004) 'Type I secretion in gram-negative bacteria', Biochimica et Biophysica Acta (BBA)-
Molecular Cell Research, 1694(1), pp. 149-161.
Demers, J., Habenstein, B., Loquet, A., Vasa, S.K., Giller, K., Becker, S., Baker, D., Lange, A. and
Sgourakis, N.G. (2014) 'High-resolution structure of the Shigella type-III secretion needle by solid-
state NMR and cryo-electron microscopy', Nature communications, 5.
Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., Dufayard, J., Guindon, S.,
Lefort, V., Lescot, M., Claverie, J. and Gascuel, O. (2008) 'Phylogeny.fr: robust phylogenetic
analysis for the non-specialist', Nucleic acids research, 36, pp. W465.
Dereeper, A., Audic, S., Claverie, J. and Blanc, G. (2010) 'BLAST-EXPLORER helps you building
datasets for phylogenetic analysis', BMC Evolutionary Biology, 10(1), pp. 8-8.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
239
Despalins, A., Marsit, S. and Oberto, J. (2011) 'Absynte: a web tool to analyze the evolution of
orthologous archaeal and bacterial gene clusters', Bioinformatics (Oxford, England), 27(20), pp.
2905.
Di Martino, P., Bertin, Y., Girardeau, J.P., Livrelli, V., Joly, B. and Darfeuille-Michaud, A. (1995)
'Molecular characterization and adhesive properties of CF29K, an adhesin of Klebsiella pneumoniae
strains involved in nosocomial infections', Infection and immunity, 63(11), pp. 4336-4344.
Di Martino, P., Livrelli, V., Sirot, D., Joly, B. and Darfeuille-Michaud, A. (1996) 'A new fimbrial antigen
harbored by CAZ-5/SHV-4-producing Klebsiella pneumoniae strains involved in nosocomial
infections', Infection and immunity, 64(6), pp. 2266-2273.
Di Paolo, N.C., Shafiani, S., Day, T., Papayannopoulou, T., Russell, D.W., Iwakura, Y., Sherman, D.,
Urdahl, K. and Shayakhmetov, D.M. (2015) 'Interdependence between interleukin-1 and tumor
necrosis factor regulates TNF-dependent control of Mycobacterium tuberculosis infection',
Immunity, 43(6), pp. 1125-1136.
Di Paolo, N.C. and Shayakhmetov, D.M. (2016) 'Interleukin 1α and the inflammatory process', Nature
immunology, 17(8), pp. 906.
Diancourt, L., Passet, V., Verhoef, J., Grimont, P.A.D. and Brisse, S. (2005) 'Multilocus Sequence Typing
of Klebsiella pneumoniae Nosocomial Isolates', Journal of clinical microbiology, 43(8), pp. 4178.
Dillon, R., Vennard, C. and Charnley, A. (2002) 'A note: gut bacteria produce components of a locust
cohesion pheromone', Journal of applied microbiology, 92(4), pp. 759-763.
Dinarello, C.A. (2011) 'Interleukin-1 in the pathogenesis and treatment of inflammatory diseases', Blood,
117(14), pp. 3720-3732.
Dolores, J.S., Agarwal, S., Egerer, M. and Satchell, K.J. (2015) 'Vibrio cholerae MARTX toxin
heterologous translocation of beta‐lactamase and roles of individual effector domains on
cytoskeleton dynamics', Molecular microbiology, 95(4), pp. 590-604.
Domenico, P., Schwartz, S. and Cunha, B.A. (1989) 'Reduction of capsular polysaccharide production in
Klebsiella pneumoniae by sodium salicylate', Infection and immunity, 57(12), pp. 3778-3782.
Dong, Y., Iniguez, A.L. and Triplett, E.W. (2003) 'Quantitative assessments of the host range and strain
specificity of endophytic colonization by Klebsiella pneumoniae 342', Plant and Soil, 257(1), pp. 49-
59.
Dong, T.G., Ho, B.T., Yoder-Himes, D.R. and Mekalanos, J.J. (2013a) 'Identification of T6SS-dependent
effector and immunity proteins by Tn-seq in Vibrio cholerae', Proceedings of the National Academy
of Sciences of the United States of America, 110(7), pp. 2623-2628.
Dong, T.G., Ho, B.T., Yoder-Himes, D.R. and Mekalanos, J.J. (2013b) 'Identification of T6SS-dependent
effector and immunity proteins by Tn-seq in Vibrio cholerae', Proceedings of the National Academy
of Sciences, 110(7), pp. 2623-2628.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
240
Dong, T.G., Ho, B.T., Yoder-Himes, D.R. and Mekalanos, J.J. (2013c) 'Identification of T6SS-dependent
effector and immunity proteins by Tn-seq in Vibrio cholerae', Proceedings of the National Academy
of Sciences, 110(7), pp. 2623-2628.
Doorduijn, D.J., Rooijakkers, S.H., van Schaik, W. and Bardoel, B.W. (2016) 'Complement resistance
mechanisms of Klebsiella pneumoniae', Immunobiology, 221(10), pp. 1102-1109.
Dos, S.G., Delattre, C., Balestrino, D., Charbonnel, N., Elboutachfaiti, R., Wadouachi, A., Badel, S.,
Bernardi, T., Michaud, P. and Forestier, C. (2014) 'Anti-Biofilm Activity: A Function of Klebsiella
pneumoniae Capsular Polysaccharide.(Research Article)', PLoS ONE, 9(6).
Drancourt, M., Bollet, C., Carta, A. and Rousselier, P. (2001) 'Phylogenetic analyses of Klebsiella species
delineate Klebsiella and Raoultella gen. nov., with description of Raoultella ornithinolytica comb.
nov., Raoultella terrigena comb. nov. and Raoultella planticola comb. nov.', International Journal
of Systematic and Evolutionary Microbiology, 51(3), pp. 925-932.
Dube, P.H., Revell, P.A., Chaplin, D.D., Lorenz, R.G. and Miller, V.L. (2001) 'A role for IL-1 alpha in
inducing pathologic inflammation during bacterial infection', Proceedings of the National Academy
of Sciences of the United States of America, 98(19), pp. 10880-10885.
Dufour, J.H., Dziejman, M., Liu, M.T., Leung, J.H., Lane, T.E. and Luster, A.D. (2002) 'IFN-gamma-
inducible protein 10 (IP-10; CXCL10)-deficient mice reveal a role for IP-10 in effector T cell
generation and trafficking', Journal of immunology (Baltimore, Md.: 1950), 168(7), pp. 3195-3204.
Duguid, J. (1959) 'Fimbriae and adhesive properties in Klebsiella strains', Microbiology, 21(1), pp. 271-
286.
Duguid, J., Smith, I.W., Dempster, G. and Edmunds, P. (1955) 'Non‐flagellar filamentous appendages
(“fimbriæ”) and hæmagglutinating activity in Bacterium coli', The Journal of pathology and
bacteriology, 70(2), pp. 335-348.
Duncan, D.W. and Razzell, W.E. (1972) 'Klebsiella biotypes among coliforms isolated from forest
environments and farm produce', Applied Microbiology, 24(6), pp. 933-938.
Durand, E., Derrez, E., Audoly, G., Spinelli, S., Ortiz-Lombardia, M., Raoult, D., Cascales, E. and
Cambillau, C. (2012) 'Crystal structure of the VgrG1 actin cross-linking domain of the Vibrio
cholerae type VI secretion system', The Journal of biological chemistry, 287(45), pp. 38190-38199.
Dutta, P., Jijumon, A., Mazumder, M., Dileep, D., Gourinath, S., Mukhopadhyay, A.K. and Maiti, S.
(2017) 'Presence of WH2 like domain in VgrG-1 toxin of Vibrio cholerae reveals the molecular
mechanism of actin cross-linking', BioRxiv, , pp. 226605.
El Fertas-Aissani, R., Messai, Y., Alouache, S. and Bakour, R. (2013) 'Virulence profiles and antibiotic
susceptibility patterns of Klebsiella pneumoniae strains isolated from different clinical specimens',
Pathologie Biologie, 61(5), pp. 209-216.
Eleftherianos, I., Millichap, P.J. and Reynolds, S.E. (2006) 'RNAi suppression of recognition protein
mediated immune responses in the tobacco hornworm Manduca sexta causes increased susceptibility

David Mwin Ph.D. Thesis, University of Leicester, UK. References
241
to the insect pathogen Photorhabdus', Developmental & Comparative Immunology, 30(12), pp. 1099-
1107.
Elhani, D., Bakir, L., Aouni, M., Passet, V., Arlet, G., Brisse, S. and Weill, F. (2010) 'Molecular
epidemiology of extended‐spectrum β‐lactamase‐producing Klebsiella pneumoniae strains in a
university hospital in Tunis, Tunisia, 1999–2005', Clinical Microbiology and Infection, 16(2), pp.
157-164.
Enos-Berlage, J.L., Guvener, Z.T., Keenan, C.E. and McCarter, L.L. (2005) 'Genetic determinants of
biofilm development of opaque and translucent Vibrio parahaemolyticus', Molecular microbiology,
55(4), pp. 1160-1182.
Ensminger, A.W. and Isberg, R.R. (2009) 'Legionella pneumophila Dot/Icm translocated substrates: a
sum of parts', Current opinion in microbiology, 12(1), pp. 67-73.
Escherich, T. (1885) 'D ie darmbakterien des neugeborenen und saunglings. . 3:515–522.', Fortsch.
Deutsch. Med, 3, pp. 515-522.
EUZéBY, J.P. (1997) 'List of Bacterial Names with Standing in Nomenclature: a folder available on the
Internet', International Journal of Systematic and Evolutionary Microbiology, 47(2), pp. 590-592.
Evrard, B., Balestrino, D., Dosgilbert, A., Bouya-Gachancard, J.L., Charbonnel, N., Forestier, C. and
Tridon, A. (2010) 'Roles of capsule and lipopolysaccharide O antigen in interactions of human
monocyte-derived dendritic cells and Klebsiella pneumoniae', Infection and immunity, 78(1), pp.
210-219.
Fader, R.C., Duffy, L.K., Davis, C.P. and Kurosky, A. (1982) 'Purification and chemical characterization
of type 1 pili isolated from Klebsiella pneumoniae', The Journal of biological chemistry, 257(6), pp.
3301-3305.
Fang, G., Fine, M., Orloff, J., Arisumi, D., Victor, L.Y., Kapoor, W., Grayston, J.T., Wang, S.P., Kohler,
R. and Muder, R.R. (1990) 'New and emerging etiologies for community-acquired pneumonia with
implications for therapy: a prospective multicenter study of 359 cases.', Medicine, 69(5), pp. 307-
316.
Fang, C.T., Chuang, Y.P., Shun, C.T., Chang, S.C. and Wang, J.T. (2004) 'A novel virulence gene in
Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic complications',
The Journal of experimental medicine, 199(5), pp. 697-705.
Fang, C.T., Lai, S.Y., Yi, W.C., Hsueh, P.R., Liu, K.L. and Chang, S.C. (2007) 'Klebsiella pneumoniae
genotype K1: an emerging pathogen that causes septic ocular or central nervous system
complications from pyogenic liver abscess', Clinical infectious diseases : an official publication of
the Infectious Diseases Society of America, 45(3), pp. 284-293.
Fang, F.C., Sandler, N. and Libby, S.J. (2005) 'Liver abscess caused by magA+ Klebsiella pneumoniae in
North America', Journal of clinical microbiology, 43(2), pp. 991-992.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
242
Farmer, J. and Hickman-Brenner, F. (1992) 'The prokaryotes. A handbook on the biology of bacteria:
ecophysiology, isolation, identification, applications', Edited by A.Balows.New York: Springer, , pp.
2952-3011.
Feldman, C., Ross, S., Mahomed, A.G., Omar, J. and Smith, C. (1995) 'The aetiology of severe
community-acquired pneumonia and its impact on initial, empiric, antimicrobial chemotherapy',
Respiratory medicine, 89(3), pp. 187-192.
Feldman, C., Smith, C., Levy, H., Ginsburg, P., Miller, S. and Koornhof, H. (1990) 'Klebsiella
pneumoniae bacteraemia at an urban general hospital', Journal of Infection, 20(1), pp. 21-31.
Feldman, M.W., Riley, M.A., Kerr, B. and Bohannan, B.J.M. (2002) 'Local dispersal promotes
biodiversity in a real-life game of rock-paper-scissors', Nature, 418(6894), pp. 171-174.
Felisberto-Rodrigues, C., Durand, E., Aschtgen, M., Blangy, S., Ortiz-Lombardia, M., Douzi, B.,
Cambillau, C. and Cascales, E. (2011) 'Towards a structural comprehension of bacterial type VI
secretion systems: characterization of the TssJ-TssM complex of an Escherichia coli pathovar', PLoS
Pathog, 7(11), pp. e1002386.
Fierer, J. (2012) 'Biofilm formation and Klebsiella pneumoniae liver abscess: True, true and unrelated?',
Virulence, 3(3), pp. 241-242.
Filloux, A. (2009) 'The type VI secretion system: a tubular story', The EMBO journal, 28(4), pp. 309-310.
Finn, R.D., Coggill, P., Eberhardt, R.Y., Eddy, S.R., Mistry, J., Mitchell, A.L., Potter, S.C., Punta, M.,
Qureshi, M., Sangrador-Vegas, A., Salazar, G.A., Tate, J. and Bateman, A. (2016) 'The Pfam protein
families database: towards a more sustainable future', Nucleic acids research, 44, pp. D279.
Fodah, R.A., Scott, J.B., Tam, H., Yan, P., Pfeffer, T.L., Bundschuh, R. and Warawa, J.M. (2014)
'Correlation of Klebsiella pneumoniae comparative genetic analyses with virulence profiles in a
murine respiratory disease model', PloS one, 9(9), pp. e107394.
Francisco-Cruz, A., Aguilar-Santelises, M., Ramos-Espinosa, O., Mata-Espinosa, D., Marquina-Castillo,
B., Barrios-Payan, J. and Hernandez-Pando, R. (2014) 'Granulocyte–macrophage colony-stimulating
factor: not just another haematopoietic growth factor', Medical oncology, 31(1), pp. 774.
French, C.T., Toesca, I.J., Wu, T.-., Teslaa, T., Beaty, S.M., Wong, W., Liu, M., Schröder, I., Chiou, P.-.,
Teitell, M.A. and Miller, J.F. (2011) 'Dissection of the Burkholderia intracellular life cycle using a
photothermal nanoblade', Proceedings of the National Academy of Sciences of the United States of
America, 108(29), pp. 12095-12100.
Fresno, S., Jimenez, N., Canals, R., Merino, S., Corsaro, M.M., Lanzetta, R., Parrilli, M., Pieretti, G.,
Regue, M. and Tomas, J.M. (2007) 'A second galacturonic acid transferase is required for core
lipopolysaccharide biosynthesis and complete capsule association with the cell surface in Klebsiella
pneumoniae', Journal of Bacteriology, 189(3), pp. 1128-1137.
Friedländer, C. (1882) 'Über die Schizomyceten bei der acuten fibrösen Pneumonie', Virchows Archiv,
87(2), pp. 319-324.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
243
Fronzes, R., Christie, P.J. and Waksman, G. (2009) 'The structural biology of type IV secretion systems',
Nature Reviews Microbiology, 7(10), pp. 703-714.
Frost, L.S., Leplae, R., Summers, A.O. and Toussaint, A. (2005a) 'Mobile genetic elements: the agents of
open source evolution', Nature Reviews Microbiology, 3(9), pp. 722.
Frost, L.S., Leplae, R., Summers, A.O. and Toussaint, A. (2005b) 'Mobile genetic elements: the agents of
open source evolution', Nature Reviews Microbiology, 3(9), pp. 722.
Frost, L.S., Leplae, R., Summers, A.O. and Toussaint, A. (2005c) 'Mobile genetic elements: the agents of
open source evolution', Nature Reviews Microbiology, 3(9), pp. 722.
Fu, Y., Guo, L., Xu, Y., Zhang, W., Gu, J., Xu, J., Chen, X., Zhao, Y., Ma, J., Liu, X. and Zhang, F.
(2008) 'Alteration of GyrA amino acid required for ciprofloxacin resistance in Klebsiella
pneumoniae isolates in China', Antimicrobial Agents and Chemotherapy, 52(8), pp. 2980-2983.
Fumagalli, O., Tall, B.D., Schipper, C. and Oelschlaeger, T.A. (1997) 'N-glycosylated proteins are
involved in efficient internalization of Klebsiella pneumoniae by cultured human epithelial cells',
Infection and immunity, 65(11), pp. 4445-4451.
Fuqua, C., Hibbing, M.E., Peterson, S.B. and Parsek, M.R. (2010) 'Bacterial competition: surviving and
thriving in the microbial jungle', Nature Reviews Microbiology, 8(1), pp. 15-25.
Fuursted, K., Scholer, L., Hansen, F., Dam, K., Bojer, M.S., Hammerum, A.M., Dagnaes-Hansen, F.,
Olsen, A., Jasemian, Y. and Struve, C. (2012) 'Virulence of a Klebsiella pneumoniae strain carrying
the New Delhi metallo-beta-lactamase-1 (NDM-1)', Microbes and infection / Institut Pasteur, 14(2),
pp. 155-158.
Gabay, C. (2006) 'Interleukin-6 and chronic inflammation', Arthritis research & therapy, 8(2), pp. S3.
Galimand, M., Guiyoule, A., Gerbaud, G., Rasoamanana, B., Chanteau, S., Carniel, E. and Courvalin, P.
(1997) 'Multidrug resistance in Yersinia pestis mediated by a transferable plasmid', New England
Journal of Medicine, 337(10), pp. 677-681.
Gallique, M., Decoin, V., Barbey, C., Rosay, T., Feuilloley, M.G., Orange, N. and Merieau, A. (2017)
'Contribution of the Pseudomonas fluorescens MFE01 type VI secretion system to biofilm
formation', PLoS One, 12(1), pp. e0170770.
Ganz, T. (1999) 'Defensins and host defense', Science (New York, N.Y.), 286(5439), pp. 420-421.
Garcia-Sureda, L., Domenech-Sanchez, A., Barbier, M., Juan, C., Gasco, J. and Alberti, S. (2011)
'OmpK26, a novel porin associated with carbapenem resistance in Klebsiella pneumoniae',
Antimicrobial Agents and Chemotherapy, 55(10), pp. 4742-4747.
Gasink, L.B., Edelstein, P.H., Lautenbach, E., Synnestvedt, M. and Fishman, N.O. (2009) 'Risk factors
and clinical impact of Klebsiella pneumoniae carbapenemase-producing K. pneumoniae', Infection
Control & Hospital Epidemiology, 30(12), pp. 1180-1185.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
244
Gavrilin, M.A., Abdelaziz, D.H.A., Mostafa, M., Abdulrahman, B.A., Grandhi, J., Akhter, A., Khweek,
A.A., Aubert, D.F., Valvano, M.A., Wewers, M.D. and Amer, A.O. (2012) 'Activation of the pyrin
inflammasome by intracellular Burkholderia cenocepacia', Journal of Immunology, 188(7), pp.
3469-3477.
Gerlach, G.F., Clegg, S. and Allen, B.L. (1989) 'Identification and characterization of the genes encoding
the type 3 and type 1 fimbrial adhesins of Klebsiella pneumoniae', Journal of Bacteriology, 171(3),
pp. 1262-1270.
Giamarellou, H. (2010) 'Multidrug-resistant Gram-negative bacteria: how to treat and for how long',
International journal of antimicrobial agents, 36, pp. S50-S54.
Giske, C.G., Monnet, D.L., Cars, O., Carmeli, Y. and ReAct-Action on Antibiotic Resistance (2008)
'Clinical and economic impact of common multidrug-resistant gram-negative bacilli', Antimicrobial
Agents and Chemotherapy, 52(3), pp. 813-821.
Golia, P. and Sadler, M. (2006) 'Pyogenic liver abscess: Klebsiella as an emerging pathogen', Emergency
radiology, 13(2), pp. 87-88.
Gong, W., Howard, O.M., Turpin, J.A., Grimm, M.C., Ueda, H., Gray, P.W., Raport, C.J., Oppenheim,
J.J. and Wang, J.M. (1998) 'Monocyte chemotactic protein-2 activates CCR5 and blocks
CD4/CCR5-mediated HIV-1 entry/replication', The Journal of biological chemistry, 273(8), pp.
4289-4292.
Gordon, D.M. and FitzGibbon, F. (1999) 'The distribution of enteric bacteria from Australian mammals:
host and geographical effects', Microbiology, 145(10), pp. 2663-2671.
Gordon, S. and Read, R. (2002) 'Macrophage defences against respiratory tract infections: The
immunology of childhood respiratory infections', British medical bulletin, 61(1), pp. 45-61.
Gorrie, C.L., Mirceta, M., Wick, R.R., Edwards, D.J., Thomson, N.R., Strugnell, R.A., Pratt, N.F.,
Garlick, J.S., Watson, K.M., McGloughlin, D.V.P.S.A., Spelman, D.W., Jenney, A.W.J. and Holt,
K.E. (2017) 'Gastrointestinal Carriage Is a Major Reservoir of Klebsiella pneumoniae Infection in
Intensive Care Patients.(Report)', Clinical Infectious Diseases, 65(2), pp. 208.
Gotsch, F., Romero, R., Friel, L., Kusanovic, J.P., Espinoza, J., Erez, O., Than, N.G., Mittal, P., Edwin, S.
and Yoon, B.H. (2007) 'CXCL10/IP-10: a missing link between inflammation and anti-angiogenesis
in preeclampsia?', The Journal of Maternal-Fetal & Neonatal Medicine, 20(11), pp. 777-792.
Grant, S.R., Fisher, E.J., Chang, J.H., Mole, B.M. and Dangl, J.L. (2006) 'Subterfuge and manipulation:
type III effector proteins of phytopathogenic bacteria', Annu.Rev.Microbiol., 60, pp. 425-449.
Green, E.R. and Mecsas, J. (2016) 'Bacterial Secretion Systems: An Overview', Microbiology spectrum,
4(1), pp. 10.1128/microbiolspec.VMBF-0012-2015.
Griffith, E. (1987) 'The iron-uptake systems of pathogenic bacteria', Iron and Infection: Molecular,
Physiological and Clinical Aspects, , pp. 69-137.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
245
Griffiths, E. (1988) 'High-affinity iron uptake systems and bacterial virulence', Virulence mechanisms of
bacterial pathogens, , pp. 121-137.
Grimont F., Grimont P.A.D. and Richard C. (1992) 'The genus Klebsiella', in Balows A, , T.H.G.,
Dworkin M., Harder W. and Schleifer K. H. (eds.) The Prokaryotes. III edn. New York: Springer-
Verlag, pp. 2775-2795.
Günther, S., Fagone, P., Jalce, G., Atanasov, A., Guignabert, C. and Nicoletti, F. (2018) 'Role of MIF and
D-DT in immune-inflammatory, autoimmune, and chronic respiratory diseases: From pathogenic
factors to therapeutic targets', Drug discovery today, .
Haapalainen, M., Mosorin, H., Dorati, F., Wu, R.-., Roine, E., Taira, S., Nissinen, R., Mattinen, L.,
Jackson, R., Pirhonen, M. and Lin, N.-. (2012) 'Hcp2, a secreted protein of the phytopathogen
Pseudomonas syringae pv. tomato DC3000, is required for fitness for competition against bacteria
and yeasts', Journal of Bacteriology, 194(18), pp. 4810-4822.
Hachani, A., Wood, T.E. and Filloux, A. (2016) 'Type VI secretion and anti-host effectors', Current
opinion in microbiology, 29, pp. 81-93.
Hachani, A., Lossi, N.S. and Filloux, A. (2013) 'A visual assay to monitor T6SS-mediated bacterial
competition', Journal of visualized experiments : JoVE, (73), pp. e50103.
Haiko, J. and Westerlund-Wikström, B. (2013) 'The role of the bacterial flagellum in adhesion and
virulence', Biology, 2(4), pp. 1242-1267.
Hakansson, S., Bergman, T., Vanooteghem, J.C., Cornelis, G. and Wolf-Watz, H. (1993) 'YopB and
YopD constitute a novel class of Yersinia Yop proteins', Infection and immunity, 61(1), pp. 71-80.
Hakansson, S., Schesser, K., Persson, C., Galyov, E.E., Rosqvist, R., Homble, F. and Wolf-Watz, H.
(1996) 'The YopB protein of Yersinia pseudotuberculosis is essential for the translocation of Yop
effector proteins across the target cell plasma membrane and displays a contact-dependent
membrane disrupting activity', The EMBO journal, 15(21), pp. 5812-5823.
Halwani, A., Niven, D. and Dunphy, G. (2000) 'Apolipophorin-III and the interactions of lipoteichoic
acids with the immediate immune responses of Galleria mellonella', Journal of invertebrate
pathology, 76(4), pp. 233-241.
Hamilton, J.A. (2008) 'Colony-stimulating factors in inflammation and autoimmunity', Nature Reviews
Immunology, 8(7), pp. 533.
Hanahan, D. (1983) 'Studies on transformation of Escherichia coli with plasmids', Journal of Molecular
Biology, 166(4), pp. 557-580.
Hancock, V., Witsø, I.L. and Klemm, P. (2011) 'Biofilm formation as a function of adhesin, growth
medium, substratum and strain type', International Journal of Medical Microbiology, 301(7), pp.
570-576.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
246
Hansen, D.S., Gottschau, A. and Kolmos, H.J. (1998) 'Epidemiology of Klebsiella bacteraemia: a case
control study using Escherichia coli bacteraemia as control', Journal of Hospital Infection, 38(2), pp.
119-132.
Hansen, D., Skov, R., Benedi, J., Sperling, V. and Kolmos, H. (2002) 'Klebsiella typing: pulsed‐field gel
electrophoresis (PFGE) in comparison with O: K‐serotyping', Clinical microbiology and infection,
8(7), pp. 397-404.
Hansen, D.S., Aucken, H.M., Abiola, T. and Podschun, R. (2004) 'Recommended test panel for
differentiation of Klebsiella species on the basis of a trilateral interlaboratory evaluation of 18
biochemical tests', Journal of clinical microbiology, 42(8), pp. 3665-3669.
Harada, A., Sekido, N., Akahoshi, T., Wada, T., Mukaida, N. and Matsushima, K. (1994) 'Essential
involvement of interleukin‐8 (IL‐8) in acute inflammation', Journal of leukocyte biology, 56(5), pp.
559-564.
Harmer, C.J., Moran, R.A. and Hall, R.M. (2014) 'Movement of IS26-associated antibiotic resistance
genes occurs via a translocatable unit that includes a single IS26 and preferentially inserts adjacent
to another IS26', mBio, 5(5), pp. e01801-14.
He, S.Y., Lindeberg, M., Chatterjee, A.K. and Collmer, A. (1991) 'Cloned Erwinia chrysanthemi out
genes enable Escherichia coli to selectively secrete a diverse family of heterologous proteins to its
milieu', Proceedings of the National Academy of Sciences of the United States of America, 88(3), pp.
1079-1083.
Henderson, I.R., Navarro-Garcia, F., Desvaux, M., Fernandez, R.C. and Ala'Aldeen, D. (2004) 'Type V
protein secretion pathway: the autotransporter story', Microbiology and molecular biology reviews :
MMBR, 68(4), pp. 692-744.
Hennequin, C. and Robin, F. (2016) 'Correlation between antimicrobial resistance and virulence in
Klebsiella pneumoniae', European journal of clinical microbiology & infectious diseases : official
publication of the European Society of Clinical Microbiology, 35(3), pp. 333-341.
Herrero, M., de Lorenzo, V. and Timmis, K.N. (1990) 'Transposon vectors containing non-antibiotic
resistance selection markers for cloning and stable chromosomal insertion of foreign genes in gram-
negative bacteria', Journal of Bacteriology, 172(11), pp. 6557-6567.
Hibbing, M.E., Fuqua, C., Parsek, M.R. and Peterson, S.B. (2010) 'Bacterial competition: surviving and
thriving in the microbial jungle', Nature reviews.Microbiology, 8(1), pp. 15-25.
Highsmith, A.K. and Jarvis, W.R. (1985) 'Klebsiella pneumoniae: selected virulence factors that
contribute to pathogenicity', Infection control : IC, 6(2), pp. 75-77.
Ho, B.T., Dong, T.G. and Mekalanos, J.J. (2014) 'A view to a kill: the bacterial type VI secretion system',
Cell host & microbe, 15(1), pp. 9-21.
Ho, B.T., Basler, M. and Mekalanos, J.J. (2013) 'Type 6 secretion system-mediated immunity to type 4
secretion system-mediated gene transfer', Science (New York, N.Y.), 342(6155), pp. 250-253.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
247
Hoang, T.T., Karkhoff-Schweizer, R.R., Kutchma, A.J. and Schweizer, H.P. (1998) 'A broad-host-range
Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences:
application for isolation of unmarked Pseudomonas aeruginosa mutants', Gene, 212(1), pp. 77-86.
Hobbs, M. and Mattick, J.S. (1993) 'Common components in the assembly of type 4 fimbriae, DNA
transfer systems, filamentous phage and protein‐secretion apparatus: a general system for the
formation of surface‐associated protein complexes', Molecular microbiology, 10(2), pp. 233-243.
Holmström, A., Olsson, J., Cherepanov, P., Maier, E., Nordfelth, R., Pettersson, J., Benz, R., Wolf‐Watz,
H. and Forsberg, Å (2001) 'LcrV is a channel size‐determining component of the Yop effector
translocon of Yersinia', Molecular microbiology, 39(3), pp. 620-632.
Hood, R.D., Singh, P., Hsu, F., Güvener, T., Carl, M.A., Trinidad, R.R.S., Silverman, J.M., Ohlson, B.B.,
Hicks, K.G., Plemel, R.L., Li, M., Schwarz, S., Wang, W.Y., Merz, A.J., Goodlett, D.R. and
Mougous, J.D. (2010a) 'A Type VI Secretion System of Pseudomonas aeruginosa Targets a Toxin to
Bacteria', Cell Host and Microbe, 7(1), pp. 25-37.
Hood, R.D., Singh, P., Hsu, F., Güvener, T., Carl, M.A., Trinidad, R.R.S., Silverman, J.M., Ohlson, B.B.,
Hicks, K.G., Plemel, R.L., Li, M., Schwarz, S., Wang, W.Y., Merz, A.J., Goodlett, D.R. and
Mougous, J.D. (2010b) 'A Type VI Secretion System of Pseudomonas aeruginosa Targets a Toxin
to Bacteria', Cell Host and Microbe, 7(1), pp. 25-37.
Hood, R.D., Singh, P., Hsu, F., Güvener, T., Carl, M.A., Trinidad, R.R.S., Silverman, J.M., Ohlson, B.B.,
Hicks, K.G., Plemel, R.L., Li, M., Schwarz, S., Wang, W.Y., Merz, A.J., Goodlett, D.R. and
Mougous, J.D. (2010c) 'A Type VI Secretion System of Pseudomonas aeruginosa Targets a Toxin to
Bacteria', Cell Host & Microbe, 7(1), pp. 25-37.
Hopp, R.J. (1994) 'RANTES is a chemotactic and activating factor for human eosinophils', Pediatrics,
94(2), pp. 259-260.
Hormaeche, E. and Edwards P. R. (1960) 'A proposal genus Enterobacter', 10, pp. 71-74.
Hou, S.R., Xu, X.J. and Yi, P. (1992) 'Nosocomial pneumonia: a report of 372 cases', Zhonghua nei ke za
zhi, 31(6), pp. 338-40, 380.
Houwink, A. and Van Iterson, W. (1950) 'Electron microscopical observations on bacterial cytology II. A
study of flagellation', Biochimica et biophysica acta, 5, pp. 10-44.
Hsieh, P., Lin, T., Yang, F., Wu, M., Pan, Y., Wu, S. and Wang, J. (2012a) 'Lipopolysaccharide O1
antigen contributes to the virulence in Klebsiella pneumoniae causing pyogenic liver abscess', PloS
one, 7(3), pp. e33155.
Hsieh, P., Lin, T., Yang, F., Wu, M., Pan, Y., Wu, S. and Wang, J. (2012b) 'Lipopolysaccharide O1
antigen contributes to the virulence in Klebsiella pneumoniae causing pyogenic liver abscess', PloS
one, 7(3), pp. e33155.
Hsieh, P.F., Lin, T.L., Lee, C.Z., Tsai, S.F. and Wang, J.T. (2008) 'Serum-induced iron-acquisition
systems and TonB contribute to virulence in Klebsiella pneumoniae causing primary pyogenic liver
abscess', The Journal of infectious diseases, 197(12), pp. 1717-1727.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
248
Hsieh, P., Liu, J., Pan, Y., Wu, M., Lin, T., Huang, Y. and Wang, J. (2013) 'Klebsiella pneumoniae
peptidoglycan-associated lipoprotein and murein lipoprotein contribute to serum resistance,
antiphagocytosis, and proinflammatory cytokine stimulation', The Journal of infectious diseases,
208(10), pp. 1580.
Hsu, C., Lin, T., Pan, Y., Hsieh, P. and Wang, J. (2013) 'Isolation of a bacteriophage specific for a new
capsular type of Klebsiella pneumoniae and characterization of its polysaccharide depolymerase',
PLoS One, 8(8), pp. e70092.
Huang, T., Lam, I., Chang, H., Tsai, S., Palsson, B.O. and Charusanti, P. (2014) 'Capsule deletion via a
[lambda]-Red knockout system perturbs biofilm formation and fimbriae expression in Klebsiella
pneumoniae MGH 78578', BMC Research Notes, 7, pp. 13.
Hubber, A.M., Sullivan, J.T. and Ronson, C.W. (2007) 'Symbiosis-induced cascade regulation of the
Mesorhizobium loti R7A VirB/D4 type IV secretion system', Molecular Plant-Microbe Interactions,
20(3), pp. 255-261.
Hunt, J.J., Wang, J. and Callegan, M.C. (2011) 'Contribution of mucoviscosity-associated gene A (magA)
to virulence in experimental Klebsiella pneumoniae endophthalmitis', Investigative ophthalmology &
visual science, 52(9), pp. 6860-6866.
Huntley, B., Jones, A. and Cabelli, V.J. (1976) 'Klebsiella densities in waters receiving wood pulp
effluents', Journal (Water Pollution Control Federation), , pp. 1766-1771.
Insua, J.L., Llobet, E., Moranta, D., Perez-Gutierrez, C., Tomas, A., Garmendia, J. and Bengoechea, J.A.
(2013a) 'Modeling Klebsiella pneumoniae pathogenesis by infection of the wax moth Galleria
mellonella', Infection and immunity, 81(10), pp. 3552-3565.
Insua, J.L., Llobet, E., Moranta, D., Pérez-Gutiérrez, C., Tomás, A., Garmendia, J. and Bengoechea, J.A.
(2013b) 'Modeling Klebsiella pneumoniae pathogenesis by infection of the wax moth Galleria
mellonella', Infection and immunity, 81(10), pp. 3552.
Iqbal, J., Siddiqui, R. and Khan, N.A. (2014) 'Acanthamoeba and bacteria produce antimicrobials to target
their counterpart', Parasites & vectors, 7(1), pp. 56-56.
Ishida, T., Hashimoto, T., Arita, M., Ito, I. and Osawa, M. (1998) 'Etiology of community-acquired
pneumonia in hospitalized patients: a 3-year prospective study in Japan', Chest, 114(6), pp. 1588-
1593.
Ishikawa, T., Sabharwal, D., Bröms, J., Milton, D.L., Sjöstedt, A., Uhlin, B.E. and Wai, S.N. (2012)
'Pathoadaptive conditional regulation of the type VI secretion system in Vibrio cholerae O1 strains',
Infection and immunity, 80(2), pp. 575-584.
Jacob-Dubuisson, F., Fernandez, R. and Coutte, L. (2004) 'Protein secretion through autotransporter and
two-partner pathways', Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 1694(1), pp.
235-257.
Janda, J. and Abbott, S. (2006) 'The Genera Klebsiella and Raoultella', The Enterobacteria, , pp. 115-129.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
249
Jani, A.J. and Cotter, P.A. (2010) 'Type VI Secretion: Not just for pathogenesis anymore', Cell Host and
Microbe, 8(1), pp. 2-6.
Jayamani, E. and Mylonakis, E. (2014) 'Effector triggered manipulation of host immune response elicited
by different pathotypes of Escherichia coli', Virulence, 5(7), pp. 733-739.
Jean, S.S., Lee, W.S., Lam, C., Hsu, C.W., Chen, R.J. and Hsueh, P.R. (2015) 'Carbapenemase-producing
Gram-negative bacteria: current epidemics, antimicrobial susceptibility and treatment options',
Future microbiology, 10(3), pp. 407-425.
Jian, Y., George, C., Irena, Z., Ioana, C., Steve, R. and Madden Thomas, L. (2012) 'Primer-BLAST: A
tool to design target-specific primers for polymerase chain reaction', BMC Bioinformatics, 13(1), pp.
134.
Jiang, B. and Howard, S.P. (1992) 'The Aeromonas hydrophila exeE gene, required both for protein
secretion and normal outer membrane biogenesis, is a member of a general secretion pathway',
Molecular microbiology, 6(10), pp. 1351-1361.
Jiang, F., Waterfield, N.R., Yang, J., Yang, G. and Jin, Q. (2014) 'A Pseudomonas aeruginosa type VI
secretion phospholipase D effector targets both prokaryotic and eukaryotic cells', Cell host &
microbe, 15(5), pp. 600-610.
Jiang, Y., Yu, D., Wei, Z., Shen, P., Zhou, Z. and Yu, Y. (2010) 'Complete nucleotide sequence of
Klebsiella pneumoniae multidrug resistance plasmid pKP048, carrying blaKPC-2, blaDHA-1,
qnrB4, and armA', Antimicrobial Agents and Chemotherapy, 54(9), pp. 3967-3969.
Johnson, J.G., Murphy, C.N., Sippy, J., Johnson, T.J. and Clegg, S. (2011) 'Type 3 Fimbriae and Biofilm
Formation Are Regulated by the Transcriptional Regulators MrkHI in Klebsiella pneumoniae',
Journal of Bacteriology, 193(14), pp. 3453-3460.
Jonas, D., Spitzmüller, B., Daschner, F.D., Verhoef, J. and Brisse, S. (2004) 'Discrimination of Klebsiella
pneumoniae and Klebsiella oxytoca phylogenetic groups and other Klebsiella species by use of
amplified fragment length polymorphism', Research in microbiology, 155(1), pp. 17-23.
Jong, G., Hsiue, T., Chen, C., Chang, H. and Chen, C. (1995) 'Rapidly fatal outcome of bacteremic
Klebsiella pneumoniae pneumonia in alcoholics', CHEST Journal, 107(1), pp. 214-217.
Jung, S., Alsam, S., Kim, K.S. and Khan, N.A. (2008) 'Pathogen–pathogen interactions: a comparative
study of Escherichia coli interactions with the clinical and environmental isolates of Acanthamoeba',
World Journal of Microbiology and Biotechnology, 24(10), pp. 2339-2348.
Jung, S., Matin, A., Kim, K.S. and Khan, N.A. (2007) 'The capsule plays an important role in Escherichia
coli K1 interactions with Acanthamoeba', International journal for parasitology, 37(3), pp. 417-423.
Kabha, K., Nissimov, L., Athamna, A., Keisari, Y., Parolis, H., Parolis, L.A., Grue, R.M., Schlepper-
Schafer, J., Ezekowitz, A.R. and Ohman, D.E. (1995) 'Relationships among capsular structure,
phagocytosis, and mouse virulence in Klebsiella pneumoniae', Infection and immunity, 63(3), pp.
847-852.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
250
Kalivoda, E.J., Horzempa, J., Stella, N.A., Sadaf, A., Kowalski, R.P., Nau, G.J. and Shanks, R.M. (2011)
'New vector tools with a hygromycin resistance marker for use with opportunistic pathogens',
Molecular biotechnology, 48(1), pp. 7-14.
Kang, C.I., Kim, S.H., Bang, J.W., Kim, H.B., Kim, N.J., Kim, E.C., Oh, M.D. and Choe, K.W. (2006)
'Community-acquired versus nosocomial Klebsiella pneumoniae bacteremia: clinical features,
treatment outcomes, and clinical implication of antimicrobial resistance', Journal of Korean medical
science, 21(5), pp. 816-822.
Kang, Y., Tian, P. and Tan, T. (2015) 'Research advances in the virulence factors of Klebsiella
pneumoniae--A review', Wei sheng wu xue bao = Acta microbiologica Sinica, 55(10), pp. 1245-
1252.
Kesteman, A.S., Perrin-Guyomard, A., Laurentie, M., Sanders, P., Toutain, P.L. and Bousquet-Melou, A.
(2010) 'Emergence of resistant Klebsiella pneumoniae in the intestinal tract during successful
treatment of Klebsiella pneumoniae lung infection in rats', Antimicrobial Agents and Chemotherapy,
54(7), pp. 2960-2964.
Khajanchi, B.K., Sha, J., Kozlova, E.V., Erova, T.E., Suarez, G., Sierra, J.C., Popov, V.L., Horneman,
A.J. and Chopra, A.K. (2009) 'N-acylhomoserine lactones involved in quorum sensing control the
type VI secretion system, biofilm formation, protease production, and in vivo virulence in a clinical
isolate of Aeromonas hydrophila', Microbiology (Reading, England), 155(Pt 11), pp. 3518-3531.
Khan, N.A. and Siddiqui, R. (2014) 'Predator vs aliens: bacteria interactions with Acanthamoeba',
Parasitology, 141(7), pp. 869.
Kibbe, W.A. (2007) 'OligoCalc: an online oligonucleotide properties calculator', Nucleic acids research,
35, pp. W43-W46.
Kidd, T.J., Mills, G., Brennan, I., Dumigan, A., Hobley, L., Insua, J.L., Sá-Pessoa, J., Roets, L. and
Bengoechea, J.A. (2016) 'Molecular mechanisms and virulence of colistin-resistant klebsiella
pneumoniae', European Respiratory Journal, 48, pp. PA2625.
Kidd, T.J., Mills, G., Sá‐Pessoa, J., Dumigan, A., Frank, C.G., Insua, J.L., Ingram, R., Hobley, L. and
Bengoechea, J.A. (2017) 'A Klebsiella pneumoniae antibiotic resistance mechanism that subdues
host defences and promotes virulence', EMBO Molecular Medicine, 9(4), pp. 430-447.
Kim, J.M., Park, E.S., Jeong, J.S., Kim, K.M., Kim, J.M., Oh, H.S., Yoon, S.W., Chang, H.S., Chang,
K.H. and Lee, S.I. (2000) 'Multicenter surveillance study for nosocomial infections in major
hospitals in Korea', American Journal of Infection Control, 28(6), pp. 454-458.
Klausen, M., Heydorn, A., Ragas, P., Lambertsen, L., Aaes‐Jørgensen, A., Molin, S. and Tolker‐Nielsen,
T. (2003) 'Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili
mutants', Molecular microbiology, 48(6), pp. 1511-1524.
Kline, K.A., Dodson, K.W., Caparon, M.G. and Hultgren, S.J. (2010) 'A tale of two pili: assembly and
function of pili in bacteria', Trends in microbiology, 18(5), pp. 224-232.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
251
Knittel, M.D., Seidler, R.J., Eby, C. and Cabe, L.M. (1977) 'Colonization of the botanical environment by
Klebsiella isolates of pathogenic origin', Applied and Environmental Microbiology, 34(5), pp. 557-
563.
Ko, W., Paterson, D.L., Sagnimeni, A.J., Hansen, D.S., Von Gottberg, A., Mohapatra, S., Casellas, J.M.,
Goossens, H., Mulazimoglu, L. and Trenholme, G. (2002) 'Community-acquired Klebsiella
pneumoniae bacteremia: global differences in clinical patterns', Emerging infectious diseases, 8(2),
pp. 160-166.
Koczura, R. and Kaznowski, A. (2003) 'Occurrence of the Yersinia high-pathogenicity island and iron
uptake systems in clinical isolates of Klebsiella pneumoniae', Microbial pathogenesis, 35(5), pp.
197-202.
Kohanski, M.A., Dwyer, D.J., Hayete, B., Lawrence, C.A. and Collins, J.J. (2007) 'A common
mechanism of cellular death induced by bactericidal antibiotics', Cell, 130(5), pp. 797-810.
Köhidai, L. and Csaba, G. (1998) 'Chemotaxis and chemotactic selection induced with cytokines (IL-8,
Rantes and TNF-α) in the unicellular Tetrahymena pyriformis', Cytokine, 10(7), pp. 481-486.
Kohno, K., Yamamoto, T., Kuroiwa, A. and Amako, K. (1984) 'Purification and characterization of
Serratia marcescens US5 pili', Infection and immunity, 46(2), pp. 295-300.
Kolenbrander, P.E., Palmer, R.J., Periasamy, S. and Jakubovics, N.S. (2010) 'Oral multispecies biofilm
development and the key role of cell–cell distance', Nature Reviews Microbiology, 8(7), pp. 471-
480.
Konovalova, A., Petters, T. and Sogaard-Andersen, L. (2010) 'Extracellular biology of Myxococcus
xanthus', FEMS microbiology reviews, 34(2), pp. 89-106.
Koressaar, T. and Remm, M. (2007) 'Enhancements and modifications of primer design program
Primer3', 23(10), pp. 1289-1291.
Korhonen, T.K., Lounatmaa, K., Ranta, H. and Kuusi, N. (1980) 'Characterization of type 1 pili of
Salmonella typhimurium LT2', Journal of Bacteriology, 144(2), pp. 800-805.
Korotkov, K.V., Gonen, T. and Hol, W.G. (2011) 'Secretins: dynamic channels for protein transport
across membranes', Trends in biochemical sciences, 36(8), pp. 433-443.
Korotkov, K.V., Sandkvist, M. and Hol, W.G. (2012) 'The type II secretion system: biogenesis, molecular
architecture and mechanism', Nature Reviews Microbiology, 10(5), pp. 336-351.
Kottara, A., Hall, J.P., Harrison, E. and Brockhurst, M.A. (2017) 'Variable plasmid fitness effects and
mobile genetic element dynamics across Pseudomonas species', FEMS microbiology ecology, 94(1),
pp. fix172.
Kovtunovych, G., Lytvynenko, T., Negrutska, V., Lar, O., Brisse, S. and Kozyrovska, N. (2003)
'Identification of Klebsiella oxytoca using a specific PCR assay targeting the polygalacturonase
pehX gene', Research in microbiology, 154(8), pp. 587-592.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
252
Kruse, W. (ed.) (1896) Systematik der Streptothricheen and Bakterien.
Kubori, T., Matsushima, Y., Nakamura, D., Uralil, J., Lara-Tejero, M., Sukhan, A., Galan, J.E. and
Aizawa, S.I. (1998) 'Supramolecular structure of the Salmonella typhimurium type III protein
secretion system', Science (New York, N.Y.), 280(5363), pp. 602-605.
Kuhns, D.B., Priel, D.A.L., Chu, J. and Zarember, K.A. (2015) 'Isolation and functional analysis of
human neutrophils', Current protocols in immunology, 111(1), pp. 7.23. 1-7.23. 16.
Kulkarni, R., Dhakal, B.K., Slechta, E.S., Kurtz, Z., Mulvey, M.A. and Thanassi, D.G. (2009) 'Roles of
putative type II secretion and type IV pilus systems in the virulence of uropathogenic Escherichia
coli', PLoS One, 4(3), pp. e4752.
Kumar, A., Chakraborti, S., Joshi, P., Chakrabarti, P. and Chakraborty, R. (2011a) 'A multiple antibiotic
and serum resistant oligotrophic strain, Klebsiella pneumoniae MB45 having novel dfrA30, is
sensitive to ZnO QDs', Annals of clinical microbiology and antimicrobials, 10(1), pp. 19-19.
Kumar, V., Sun, P., Vamathevan, J., Li, Y., Ingraham, K., Palmer, L., Huang, J. and Brown, J.R. (2011b)
'Comparative genomics of Klebsiella pneumoniae strains with different antibiotic resistance profiles',
Antimicrobial Agents and Chemotherapy, 55(9), pp. 4267-4276.
Ladha, J., Barraquio, W. and Watanabe, I. (1983) 'Isolation and identification of nitrogen-fixing
Enterobacter cloacae and Klebsiella planticola associated with rice plants', Canadian journal of
microbiology, 29(10), pp. 1301-1308.
Lai, Y.C., Peng, H.L. and Chang, H.Y. (2003) 'RmpA2, an activator of capsule biosynthesis in Klebsiella
pneumoniae CG43, regulates K2 cps gene expression at the transcriptional level', Journal of
Bacteriology, 185(3), pp. 788-800.
Lambert‐Buisine, C., Willery, E., Locht, C. and Jacob‐Dubuisson, F. (1998) 'N‐terminal characterization
of the Bordetella pertussis filamentous haemagglutinin', Molecular microbiology, 28(6), pp. 1283-
1293.
Lambrecht, E., Baré, J., Damme, I.V., Bert, W., Sabbe, K. and Houf, K. (2013) 'Behavior of Yersinia
enterocolitica in the Presence of the Bacterivorous Acanthamoeba castellanii', Applied and
Environmental Microbiology, 79(20), pp. 6407-6413.
Larson, D.F. and Horak, K. (2006) 'Macrophage migration inhibitory factor: controller of systemic
inflammation', Critical Care, 10(2), pp. 138.
Lathem, W.W., Grys, T.E., Witowski, S.E., Torres, A.G., Kaper, J.B., Tarr, P.I. and Welch, R.A. (2002)
'StcE, a metalloprotease secreted by Escherichia coli O157: H7, specifically cleaves C1 esterase
inhibitor', Molecular microbiology, 45(2), pp. 277-288.
Lavander, M., Ericsson, S.K., Broms, J.E. and Forsberg, A. (2006) 'The twin arginine translocation
system is essential for virulence of Yersinia pseudotuberculosis', Infection and immunity, 74(3), pp.
1768-1776.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
253
Lavigne, J., Cuzon, G., Combescure, C., Bourg, G., Sotto, A. and Nordmann, P. (2013) 'Virulence of
Klebsiella pneumoniae isolates harboring bla KPC-2 carbapenemase gene in a Caenorhabditis
elegans model', PloS one, 8(7), pp. e67847.
Lawlor, M.S., O'connor, C. and Miller, V.L. (2007) 'Yersiniabactin is a virulence factor for Klebsiella
pneumoniae during pulmonary infection', Infection and immunity, 75(3), pp. 1463-1472.
Lecher, J., Schwarz, C.K., Stoldt, M., Smits, S.H., Willbold, D. and Schmitt, L. (2012) 'An RTX
transporter tethers its unfolded substrate during secretion via a unique N-terminal domain', Structure,
20(10), pp. 1778-1787.
Lederman, E.R. and Crum, N.F. (2005) 'Pyogenic liver abscess with a focus on Klebsiella pneumoniae as
a primary pathogen: an emerging disease with unique clinical characteristics', The American Journal
of Gastroenterology, 100(2), pp. 322-331.
Lee, D.G., Urbach, J.M., Wu, G., Liberati, N.T., Feinbaum, R.L., Miyata, S., Diggins, L.T., He, J.,
Saucier, M. and Déziel, E. (2006) 'Genomic analysis reveals that Pseudomonas aeruginosa virulence
is combinatorial', Genome biology, 7(10), pp. R90.
Leiman, P.G., Basler, M., Ramagopal, U.A., Bonanno, J.B., Sauder, J.M., Pukatzki, S., Burley, S.K.,
Almo, S.C. and Mekalanos, J.J. (2009a) 'Type VI secretion apparatus and phage tail-associated
protein complexes share a common evolutionary origin', Proceedings of the National Academy of
Sciences of the United States of America, 106(11), pp. 4154-4159.
Leiman, P.G., Basler, M., Ramagopal, U.A., Bonanno, J.B., Sauder, J.M., Pukatzki, S., Burley, S.K.,
Almo, S.C. and Mekalanos, J.J. (2009b) 'Type VI Secretion Apparatus and Phage Tail-Associated
Protein Complexes Share a Common Evolutionary Origin', Proceedings of the National Academy of
Sciences of the United States of America, 106(11), pp. 4154-4159.
Lennings, J., West, T.E. and Schwarz, S. (2018) 'The Burkholderia type VI secretion system 5:
composition, regulation and role in virulence', Frontiers in microbiology, 9.
Leo, J.C., Grin, I. and Linke, D. (2012) 'Type V secretion: mechanism(s) of autotransport through the
bacterial outer membrane', Philosophical transactions of the Royal Society of London.Series B,
Biological sciences, 367(1592), pp. 1088-1101.
Lertpiriyapong, K., Gamazon, E.R., Feng, Y., Park, D.S., Pang, J., Botka, G., Graffam, M.E., Ge, Z. and
Fox, J.G. (2012) 'Campylobacter jejuni type VI secretion system: roles in adaptation to deoxycholic
acid, host cell adherence, invasion, and in vivo colonization', PLoS One, 7(8), pp. e42842.
Lery, L.M., Frangeul, L., Tomas, A., Passet, V., Almeida, A.S., Bialek-Davenet, S., Barbe, V.,
Bengoechea, J.A., Sansonetti, P. and Brisse, S. (2014) 'Comparative analysis of Klebsiella
pneumoniae genomes identifies a phospholipase D family protein as a novel virulence factor', BMC
biology, 12(1), pp. 1.
Lesic, á, Starkey, á, He, J., Hazan, R. and Rahme, L. (2009) 'Quorum sensing differentially regulates
Pseudomonas aeruginosa type VI secretion locus I and homologous loci II and III, which are
required for pathogenesis', Microbiology, 155(9), pp. 2845-2855.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
254
Letoffe, S., Delepelaire, P. and Wandersman, C. (1996) 'Protein secretion in gram-negative bacteria:
assembly of the three components of ABC protein-mediated exporters is ordered and promoted by
substrate binding', The EMBO journal, 15(21), pp. 5804-5811.
Leverstein-Van Hall, M.A., Stuart, J.C., Voets, G.M., Versteeg, D., Tersmette, T. and Fluit, A.C. (2010)
'Global spread of New Delhi metallo-beta-lactamase 1', The Lancet.Infectious diseases, 10(12), pp.
830-831.
Levy, J.A. (2009) 'The unexpected pleiotropic activities of RANTES', Journal of immunology (Baltimore,
Md.: 1950), 182(7), pp. 3945-3946.
Leyton, D.L., Rossiter, A.E. and Henderson, I.R. (2012a) 'From self sufficiency to dependence:
mechanisms and factors important for autotransporter biogenesis', Nature Reviews Microbiology,
10(3), pp. 213-225.
Leyton, D.L., Rossiter, A.E. and Henderson, I.R. (2012b) 'From self sufficiency to dependence:
mechanisms and factors important for autotransporter biogenesis', Nature Reviews Microbiology,
10(3), pp. 213-225.
Li, B., Yi, Y., Wang, Q., Woo, P.C., Tan, L., Jing, H., Gao, G.F. and Liu, C.H. (2012) 'Analysis of drug
resistance determinants in Klebsiella pneumoniae isolates from a tertiary-care hospital in Beijing,
China', PLoS One, 7(7), pp. e42280.
Li, B., Zhao, Y., Liu, C., Chen, Z. and Zhou, D. (2014a) 'Molecular pathogenesis of Klebsiella
pneumoniae', Future Microbiology, 9(9), pp. 1071.
Li, W., Sun, G., Yu, Y., Li, N., Chen, M., Jin, R., Jiao, Y. and Wu, H. (2014b) 'Increasing occurrence of
antimicrobial-resistant hypervirulent (hypermucoviscous) Klebsiella pneumoniae isolates in China',
Clinical infectious diseases : an official publication of the Infectious Diseases Society of America,
58(2), pp. 225-232.
Liam, C., Lim, K. and Wong, C.M. (2001) 'Community‐acquired pneumonia in patients requiring
hospitalization', Respirology, 6(3), pp. 259-264.
Liao, C.H., Hsueh, P.R., Jacoby, G.A. and Hooper, D.C. (2013) 'Risk factors and clinical characteristics
of patients with qnr-positive Klebsiella pneumoniae bacteraemia', The Journal of antimicrobial
chemotherapy, 68(12), pp. 2907-2914.
Liebert, C.A., Hall, R.M. and Summers, A.O. (1999) 'Transposon Tn21, flagship of the floating genome',
Microbiology and molecular biology reviews : MMBR, 63(3), pp. 507-522.
Lin, T., Yang, F., Yang, A., Peng, H., Li, T., Tsai, M., Wu, S. and Wang, J. (2012) 'Amino acid
substitutions of MagA in Klebsiella pneumoniae affect the biosynthesis of the capsular
polysaccharide', PloS one, 7(10), pp. e46783.
Lin, T.L., Lee, C.Z., Hsieh, P.F., Tsai, S.F. and Wang, J.T. (2008) 'Characterization of integrative and
conjugative element ICEKp1-associated genomic heterogeneity in a Klebsiella pneumoniae strain
isolated from a primary liver abscess', Journal of Bacteriology, 190(2), pp. 515-526.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
255
Linares, D., Jean, N., Van Overtvelt, P., Ouidir, T., Hardouin, J., Blache, Y. and Molmeret, M. (2016)
'The marine bacteria Shewanella frigidimarina NCIMB400 upregulates the type VI secretion system
during early biofilm formation', Environmental microbiology reports, 8(1), pp. 110-121.
Lindeberg, M., Biehl, B.S., Glasner, J.D., Perna, N.T., Collmer, A. and Collmer, C.W. (2009) 'Gene
Ontology annotation highlights shared and divergent pathogenic strategies of type III effector
proteins deployed by the plant pathogen Pseudomonas syringae pv tomato DC3000 and animal
pathogenic Escherichia coli strains', BMC microbiology, 9(1), pp. 1.
Liu, B. and Pop, M. (2008) 'ARDB—antibiotic resistance genes database', Nucleic acids research,
37(suppl_1), pp. D443-D447.
Liu, H., Coulthurst, S.J., Pritchard, L., Hedley, P.E., Ravensdale, M., Humphris, S., Burr, T., Takle, G.,
Brurberg, M. and Birch, P.R. (2008) 'Quorum sensing coordinates brute force and stealth modes of
infection in the plant pathogen Pectobacterium atrosepticum', PLoS Pathog, 4(6), pp. e1000093.
Liu, M., Guo, S., Hibbert, J.M., Jain, V., Singh, N., Wilson, N.O. and Stiles, J.K. (2011) 'CXCL10/IP-10
in infectious diseases pathogenesis and potential therapeutic implications', Cytokine & growth factor
reviews, 22(3), pp. 121-130.
Liu, P., Li, P., Jiang, X., Bi, D., Xie, Y., Tai, C., Deng, Z., Rajakumar, K. and Ou, H.Y. (2012a)
'Complete genome sequence of Klebsiella pneumoniae subsp. pneumoniae HS11286, a multidrug-
resistant strain isolated from human sputum', Journal of Bacteriology, 194(7), pp. 1841-1842.
Liu, P., Li, P., Jiang, X., Bi, D., Xie, Y., Tai, C., Deng, Z., Rajakumar, K. and Ou, H.Y. (2012b)
'Complete genome sequence of Klebsiella pneumoniae subsp. pneumoniae HS11286, a multidrug-
resistant strain isolated from human sputum', Journal of Bacteriology, 194(7), pp. 1841-1842.
Livrelli, V., De Champs, C., Di Martino, P., Darfeuille-Michaud, A., Forestier, C. and Joly, B. (1996)
'Adhesive properties and antibiotic resistance of Klebsiella, Enterobacter, and Serratia clinical
isolates involved in nosocomial infections', Journal of clinical microbiology, 34(8), pp. 1963-1969.
Llobet, E., Tomas, J.M. and Bengoechea, J.A. (2008) 'Capsule polysaccharide is a bacterial decoy for
antimicrobial peptides', Microbiology, 154(12), pp. 3877-3886.
Llobet, E., Campos, M.A., Gimenez, P., Moranta, D. and Bengoechea, J.A. (2011) 'Analysis of the
networks controlling the antimicrobial-peptide-dependent induction of Klebsiella pneumoniae
virulence factors', Infection and immunity, 79(9), pp. 3718-3732.
Llobet, E., March, C., Gimenez, P. and Bengoechea, J.A. (2009) 'Klebsiella pneumoniae OmpA confers
resistance to antimicrobial peptides', Antimicrobial Agents and Chemotherapy, 53(1), pp. 298-302.
Lossi, N.S., Dajani, R., Freemont, P. and Filloux, A. (2011) 'Structure–function analysis of HsiF, a gp25-
like component of the type VI secretion system, in Pseudomonas aeruginosa', Microbiology,
157(12), pp. 3292-3305.
Lu, A., Zhang, Q., Zhang, J., Yang, B., Wu, K., Xie, W., Luan, Y. and Ling, E. (2014) 'Insect
prophenoloxidase: the view beyond immunity', Frontiers in physiology, 5, pp. 252.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
256
Lu, H.M. and Lory, S. (1996) 'A specific targeting domain in mature exotoxin A is required for its
extracellular secretion from Pseudomonas aeruginosa', The EMBO journal, 15(2), pp. 429-436.
Lue, H., Kleemann, R., Calandra, T., Roger, T. and Bernhagen, J. (2002) 'Macrophage migration
inhibitory factor (MIF): mechanisms of action and role in disease', Microbes and Infection, 4(4), pp.
449-460.
Ma, A.T., McAuley, S., Pukatzki, S. and Mekalanos, J.J. (2009a) 'Translocation of a Vibrio cholerae type
VI secretion effector requires bacterial endocytosis by host cells', Cell host & microbe, 5(3), pp. 234-
243.
Ma, A.T., McAuley, S., Pukatzki, S. and Mekalanos, J.J. (2009b) 'Translocation of a Vibrio cholerae type
VI secretion effector requires bacterial endocytosis by host cells', Cell host & microbe, 5(3), pp. 234-
243.
Ma, A.T. and Mekalanos, J.J. (2010a) 'In vivo actin cross-linking induced by Vibrio cholerae type VI
secretion system is associated with intestinal inflammation', Proceedings of the National Academy of
Sciences of the United States of America, 107(9), pp. 4365-4370.
Ma, A.T. and Mekalanos, J.J. (2010b) 'In vivo Actin Cross-Linking Induced by Vibrio cholerae Type VI
Secretion System Is Associated with Intestinal Inflammation', Proceedings of the National Academy
of Sciences of the United States of America, 107(9), pp. 4365-4370.
Ma, L.C., Fang, C.T., Lee, C.Z., Shun, C.T. and Wang, J.T. (2005) 'Genomic heterogeneity in Klebsiella
pneumoniae strains is associated with primary pyogenic liver abscess and metastatic infection', The
Journal of infectious diseases, 192(1), pp. 117-128.
Ma, L.S., Lin, J.S. and Lai, E.M. (2009) 'An IcmF family protein, ImpLM, is an integral inner membrane
protein interacting with ImpKL, and its walker a motif is required for type VI secretion system-
mediated Hcp secretion in Agrobacterium tumefaciens', Journal of Bacteriology, 191(13), pp. 4316-
4329.
MacIntyre, D.L., Miyata, S.T., Kitaoka, M. and Pukatzki, S. (2010a) 'The Vibrio cholerae type VI
secretion system displays antimicrobial properties', Proceedings of the National Academy of
Sciences of the United States of America, 107(45), pp. 19520-19524.
MacIntyre, D.L., Miyata, S.T., Kitaoka, M. and Pukatzki, S. (2010b) 'The Vibrio cholerae type VI
secretion system displays antimicrobial properties', Proceedings of the National Academy of
Sciences of the United States of America, 107(45), pp. 19520-19524.
MacIntyre, D.L., Miyata, S.T., Kitaoka, M., Pukatzki, S. and Mekalanos, J.J. (2010c) 'The Vibrio
cholerae type VI secretion system displays antimicrobial properties', Proceedings of the National
Academy of Sciences of the United States of America, 107(45), pp. 19520-19524.
MAHL, M.C., WILSON, P.W., FIFE, M.A. and EWING, W.H. (1965) 'Nitrogen Fixation by Members of
the Tribe Klebsielleae', Journal of Bacteriology, 89, pp. 1482-1487.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
257
March, C., Cano, V., Moranta, D., Llobet, E., Pérez-Gutiérrez, C., Tomás, J.M., Suárez, T., Garmendia, J.
and Bengoechea, J.A. (2013) 'Role of bacterial surface structures on the interaction of Klebsiella
pneumoniae with phagocytes', PLoS One, 8(2), pp. e56847.
March, C., Moranta, D., Regueiro, V., Llobet, E., Tomas, A., Garmendia, J. and Bengoechea, J.A. (2011)
'Klebsiella pneumoniae outer membrane protein A is required to prevent the activation of airway
epithelial cells', The Journal of biological chemistry, 286(12), pp. 9956-9967.
Marchler-Bauer, A., Bo, Y., Han, L., He, J., Lanczycki, C.J., Lu, S., Chitsaz, F., Derbyshire, M.K., Geer,
R.C., Gonzales, N.R., Gwadz, M., Hurwitz, D.I., Lu, F., Marchler, G.H., Song, J.S., Thanki, N.,
Wang, Z., Yamashita, R.A., Zhang, D., Zheng, C., Geer, L.Y. and Bryant, S.H. (2017)
'CDD/SPARCLE: functional classification of proteins via subfamily domain architectures', Nucleic
acids research, 45, pp. D200.
Maroncle, N.M., Sivick, K.E., Brady, R., Stokes, F.E. and Mobley, H.L. (2006) 'Protease activity,
secretion, cell entry, cytotoxicity, and cellular targets of secreted autotransporter toxin of
uropathogenic Escherichia coli', Infection and immunity, 74(11), pp. 6124-6134.
Marston, B.J., Plouffe, J.F., File, T.M., Hackman, B.A., Salstrom, S., Lipman, H.B., Kolczak, M.S. and
Breiman, R.F. (1997) 'Incidence of community-acquired pneumonia requiring hospitalization: results
of a population-based active surveillance study in Ohio', Archives of Internal Medicine, 157(15), pp.
1709-1718.
Matsuno, H., Yudoh, K., Katayama, R., Nakazawa, F., Uzuki, M., Sawai, T., Yonezawa, T., Saeki, Y.,
Panayi, G. and Pitzalis, C. (2002) 'The role of TNF‐α in the pathogenesis of inflammation and joint
destruction in rheumatoid arthritis (RA): a study using a human RA/SCID mouse chimera',
Rheumatology, 41(3), pp. 329-337.
Maurer, M. and Von Stebut, E. (2004) 'Macrophage inflammatory protein-1', The international journal of
biochemistry & cell biology, 36(10), pp. 1882-1886.
Mazzariol, A., Zuliani, J., Cornaglia, G., Rossolini, G.M. and Fontana, R. (2002) 'AcrAB Efflux System:
Expression and Contribution to Fluoroquinolone Resistance in Klebsiella spp', Antimicrobial Agents
and Chemotherapy, 46(12), pp. 3984-3986.
McArthur, A.G., Waglechner, N., Nizam, F., Yan, A., Azad, M.A., Baylay, A.J., Bhullar, K., Canova,
M.J., De Pascale, G., Ejim, L., Kalan, L., King, A.M., Koteva, K., Morar, M., Mulvey, M.R.,
O'Brien, J.S., Pawlowski, A.C., Piddock, L.J., Spanogiannopoulos, P., Sutherland, A.D., Tang, I.,
Taylor, P.L., Thaker, M., Wang, W., Yan, M., Yu, T. and Wright, G.D. (2013) 'The comprehensive
antibiotic resistance database', Antimicrobial Agents and Chemotherapy, 57(7), pp. 3348-3357.
McCann, J.R. and Geme III, Joseph W St (2014) 'The HMW1C-like glycosyltransferases—an enzyme
family with a sweet tooth for simple sugars', PLoS Pathog, 10(4), pp. e1003977.
McDonough, J.A., McCann, J.R., Tekippe, E.M., Silverman, J.S., Rigel, N.W. and Braunstein, M. (2008)
'Identification of functional Tat signal sequences in Mycobacterium tuberculosis proteins', Journal of
Bacteriology, 190(19), pp. 6428-6438.
McLaughlin, M.M., Advincula, M.R., Malczynski, M., Barajas, G., Qi, C. and Scheetz, M.H. (2014)
'Quantifying the clinical virulence of Klebsiella pneumoniae producing carbapenemase Klebsiella

David Mwin Ph.D. Thesis, University of Leicester, UK. References
258
pneumoniae with a Galleria mellonella model and a pilot study to translate to patient outcomes',
BMC infectious diseases, 14(1), pp. 1.
Mendez, M.P., Monroy, Y.K., Du, M., Preston, A.M., Tolle, L., Lin, Y., VanDussen, K.L., Samuelson,
L.C., Standiford, T.J. and Curtis, J.L. (2011) 'Overexpression of sICAM-1 in the alveolar epithelial
space results in an exaggerated inflammatory response and early death in Gram negative pneumonia',
Respiratory research, 12(1), pp. 12.
Merino, S., Altarriba, M., Izquierdo, L., Nogueras, M.M., Regue, M. and Tomas, J.M. (2000) 'Cloning
and sequencing of the Klebsiella pneumoniae O5 wb gene cluster and its role in pathogenesis',
Infection and immunity, 68(5), pp. 2435-2440.
Messer, J.S., Liechty, E.R., Vogel, O.A. and Chang, E.B. (2017) 'Evolutionary and ecological forces that
shape the bacterial communities of the human gut', Mucosal immunology, 10(3), pp. 567.
Metcalf, D. and Nicola, N. (1995) The hemopoietic colony-stimulating factors: from biology to clinical
applications. Cambridge University Press.
Meyer, D.M., Dustin, M.L. and Carron, C.P. (1995) 'Characterization of intercellular adhesion molecule-1
ectodomain (sICAM-1) as an inhibitor of lymphocyte function-associated molecule-1 interaction
with ICAM-1', Journal of immunology (Baltimore, Md.: 1950), 155(7), pp. 3578-3584.
Miethke, M. and Marahiel, M.A. (2007) 'Siderophore-based iron acquisition and pathogen control',
Microbiology and molecular biology reviews : MMBR, 71(3), pp. 413-451.
Mikula, K.M., Kolodziejczyk, R. and Goldman, A. (2014) 'Yersinia infection tools—characterization of
structure and function of adhesins', The pathogenic Yersiniae–advances in the understanding of
physiology and virulence, , pp. 7.
Mirza, M.S., Ahmad, W., Latif, F., Haurat, J., Bally, R., Normand, P. and Malik, K.A. (2001) 'Isolation,
partial characterization, and the effect of plant growth-promoting bacteria (PGPB) on micro-
propagated sugarcane in vitro', Plant and Soil, 237(1), pp. 47-54.
Miyata, S.T., Kitaoka, M., Brooks, T.M., McAuley, S.B. and Pukatzki, S. (2011) 'Vibrio cholerae
requires the type VI secretion system virulence factor vasx to kill Dictyostelium discoideum',
Infection and immunity, 79(7), pp. 2941-2949.
Miyata, S.T., Bachmann, V. and Pukatzki, S. (2013) 'Type VI secretion system regulation as a
consequence of evolutionary pressure', Journal of medical microbiology, 62(Pt 5), pp. 663.
Mobley, H.L., Island, M.D. and Hausinger, R.P. (1995) 'Molecular biology of microbial ureases',
Microbiological reviews, 59(3), pp. 451-480.
Monnet, D., Freney, J., Brun, Y., Boeufgras, J. and Fleurette, J. (1991) 'Difficulties in identifying
Klebsiella strains of clinical origin', Zentralblatt für Bakteriologie, 274(4), pp. 456-464.
Monnet, D. and Freney, J. (1994) 'Method for differentiating Klebsiella planticola and Klebsiella
terrigena from other Klebsiella species', Journal of clinical microbiology, 32(4), pp. 1121-1122.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
259
Montgomerie, J.Z. (1979) 'Epidemiology of Klebsiella and hospital-associated infections', Reviews of
infectious diseases, , pp. 736-753.
Mota, L.J. and Cornelis, G.R. (2005) 'The bacterial injection kit: type III secretion systems', Annals of
Medicine, 37(4), pp. 234-249.
Mougous, J.D., Cuff, M.E., Raunser, S., Shen, A., Zhou, M., Gifford, C.A., Goodman, A.L., Joachimiak,
G., Ordonez, C.L., Lory, S., Walz, T., Joachimiak, A. and Mekalanos, J.J. (2006a) 'A virulence locus
of Pseudomonas aeruginosa encodes a protein secretion apparatus', Science (New York, N.Y.),
312(5779), pp. 1526-1530.
Mougous, J.D., Cuff, M.E., Raunser, S., Shen, A., Zhou, M., Gifford, C.A., Goodman, A.L., Joachimiak,
G., Ordonez, C.L., Lory, S., Walz, T., Joachimiak, A. and Mekalanos, J.J. (2006b) 'A virulence locus
of Pseudomonas aeruginosa encodes a protein secretion apparatus', Science (New York, N.Y.),
312(5779), pp. 1526-1530.
Moya, B., Juan, C., Alberti, S., Perez, J.L. and Oliver, A. (2008) 'Benefit of having multiple ampD genes
for acquiring beta-lactam resistance without losing fitness and virulence in Pseudomonas
aeruginosa', Antimicrobial Agents and Chemotherapy, 52(10), pp. 3694-3700.
Mulder, D.T., Cooper, C.A. and Coombes, B.K. (2012a) 'Type VI secretion system-associated gene
clusters contribute to pathogenesis of Salmonella enterica serovar typhimurium', Infection and
immunity, 80(6), pp. 1996-2007.
Mulder, D.T., Cooper, C.A. and Coombes, B.K. (2012b) 'Type VI secretion system-associated gene
clusters contribute to pathogenesis of Salmonella enterica serovar typhimurium', Infection and
immunity, 80(6), pp. 1996-2007.
Müller, M. (2005) 'Twin-arginine-specific protein export in Escherichia coli', Research in microbiology,
156(2), pp. 131-136.
Munoz-Price, L.S., Poirel, L., Bonomo, R.A., Schwaber, M.J., Daikos, G.L., Cormican, M., Cornaglia,
G., Garau, J., Gniadkowski, M. and Hayden, M.K. (2013) 'Clinical epidemiology of the global
expansion of Klebsiella pneumoniae carbapenemases', The Lancet infectious diseases, 13(9), pp.
785-796.
Murdoch, S.L., Trunk, K., English, G., Fritsch, M.J., Pourkarimi, E. and Coulthurst, S.J. (2011a) 'The
opportunistic pathogen Serratia marcescens utilizes type VI secretion to target bacterial
competitors', Journal of Bacteriology, 193(21), pp. 6057-6069.
Murdoch, S.L., Trunk, K., English, G., Fritsch, M.J., Pourkarimi, E. and Coulthurst, S.J. (2011b) 'The
opportunistic pathogen Serratia marcescens utilizes type VI secretion to target bacterial
competitors', Journal of Bacteriology, 193(21), pp. 6057-6069.
Murin, C.D., Segal, K., Bryksin, A. and Matsumura, I. (2012) 'Expression vectors for Acinetobacter
baylyi ADP1', Appl.Environ.Microbiol., 78(1), pp. 280-283.
Murphy, C.N. and Clegg, S. (2012) 'Klebsiella pneumoniae and type 3 fimbriae: nosocomial infection,
regulation and biofilm formation', Future microbiology, 7(8), pp. 991-1002.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
260
Murphy, C.N., Mortensen, M.S., Krogfelt, K.A. and Clegg, S. (2013) 'Role of Klebsiella pneumoniae type
1 and type 3 fimbriae in colonizing silicone tubes implanted into the bladders of mice as a model of
catheter-associated urinary tract infections', Infection and immunity, 81(8), pp. 3009-3017.
Murphy, K.C. (1998) 'Use of bacteriophage lambda recombination functions to promote gene replacement
in Escherichia coli', Journal of Bacteriology, 180(8), pp. 2063-2071.
Mylonakis, E., Casadevall, A. and Ausubel, F.M. (2007) 'Exploiting amoeboid and non-vertebrate animal
model systems to study the virulence of human pathogenic fungi', PLoS pathogens, 3(7), pp. e101.
Nassif, X. and Sansonetti, P.J. (1986) 'Correlation of the virulence of Klebsiella pneumoniae K1 and K2
with the presence of a plasmid encoding aerobactin', Infection and immunity, 54(3), pp. 603-608.
Natale, P., Brüser, T. and Driessen, A.J. (2008) 'Sec-and Tat-mediated protein secretion across the
bacterial cytoplasmic membrane—distinct translocases and mechanisms', Biochimica et Biophysica
Acta (BBA)-Biomembranes, 1778(9), pp. 1735-1756.
Niemela, S.I. and Vaatanen, P. (1982) 'Survival in lake water of Klebsiella pneumoniae discharged by a
paper mill', Applied and Environmental Microbiology, 44(2), pp. 264-269.
Nuccio, S.P. and Baumler, A.J. (2007) 'Evolution of the chaperone/usher assembly pathway: fimbrial
classification goes Greek', Microbiology and molecular biology reviews : MMBR, 71(4), pp. 551-
575.
Oberto, J. (2013) 'SyntTax: a web server linking synteny to prokaryotic taxonomy.(Report)', BMC
Bioinformatics, 14(1).
Oberto, J. (2008) 'BAGET: a web server for the effortless retrieval of prokaryotic gene context and
sequence', Bioinformatics (Oxford, England), 24(3), pp. 424.
Ochsner, U.A., Snyder, A., Vasil, A.I. and Vasil, M.L. (2002) 'Effects of the twin-arginine translocase on
secretion of virulence factors, stress response, and pathogenesis', Proceedings of the National
Academy of Sciences of the United States of America, 99(12), pp. 8312-8317.
Oelschlaeger, T.A. and Tall, B.D. (1997) 'Invasion of cultured human epithelial cells by Klebsiella
pneumoniae isolated from the urinary tract', Infection and immunity, 65(7), pp. 2950-2958.
Ofek, I., Goldhar, J., Keisari, Y. and Sharon, N. (1995) 'Nonopsonic phagocytosis of microorganisms',
Annual Reviews in Microbiology, 49(1), pp. 239-276.
Ogawa, W., Onishi, M., Ni, R., Tsuchiya, T. and Kuroda, T. (2012) 'Functional study of the novel
multidrug efflux pump KexD from Klebsiella pneumoniae', Gene, 498(2), pp. 177-182.
Oh, H., Siano, B. and Diamond, S. (2008) 'Neutrophil isolation protocol', Journal of visualized
experiments : JoVE, (17). pii: 745. doi(17), pp. 10.3791/745.
Okano, H., Shiraki, K., Inoue, H., Kawakita, T., Yamamoto, N., Deguchi, M., Sugimoto, K., Sakai, T.,
Ohmori, S. and Murata, K. (2002) 'Clinicopathological analysis of liver abscess in Japan',
International journal of molecular medicine, 10, pp. 627-630.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
261
Old, D. (1972) 'Inhibition of the interaction between fimbrial haemagglutinins and erythrocytes by D-
mannose and other carbohydrates', Microbiology, 71(1), pp. 149-157.
Olson, R., MacDonald, U., Pomakov, A.O., Shon, A.S., Kong, Q., Metzger, D.J., Russo, T.A. and
Beanan, J.M. (2012) 'Biofilm formed by a hypervirulent (hypermucoviscous) variant of Klebsiella
pneumoniae does not enhance serum resistance or survival in an in vivo abscess model', Virulence,
3(3), pp. 309-318.
Oppenheim, J., Matsushima, K., Yoshimura, T., Leonard, E. and Neta, R. (1989) 'Relationship between
interleukin 1 (IL1), tumor necrosis factor (TNF) and a neutrophil attracting peptide (NAP-1)', Agents
and Actions, 26(1-2), pp. 134-140.
Ørskov, I. (1984) 'Genus v. Klebsiella', Bergey's manual of systematic bacteriology, 1, pp. 461-465.
Ørskov, I. and Ørskov, F. (1984) '4 Serotyping of Klebsiella', Methods in microbiology, 14, pp. 143-164.
O'toole, G.A. and Kolter, R. (1998) 'Initiation of biofilm formation in Pseudomonas fluorescens WCS365
proceeds via multiple, convergent signalling pathways: a genetic analysis', Molecular microbiology,
28(3), pp. 449-461.
Ou, H., He, X., Harrison, E.M., Kulasekara, B.R., Thani, A.B., Kadioglu, A., Lory, S., Hinton, J.C.D.,
Barer, M.R., Deng, Z. and Rajakumar, K. (2007) 'MobilomeFINDER: web-based tools for in silico
and experimental discovery of bacterial genomic islands', Nucleic acids research, 35, pp. W97.
Overmars, L., Kerkhoven, R., Siezen, R.J. and Francke, C. (2013) 'MGcV: the microbial genomic context
viewer for comparative genome analysis', BMC Genomics, 14(1), pp. 209-209.
Padilla, E., Llobet, E., Domenech-Sanchez, A., Martinez-Martinez, L., Bengoechea, J.A. and Alberti, S.
(2010a) 'Klebsiella pneumoniae AcrAB efflux pump contributes to antimicrobial resistance and
virulence', Antimicrobial Agents and Chemotherapy, 54(1), pp. 177-183.
Padilla, E., Llobet, E., Domenech-Sanchez, A., Martinez-Martinez, L., Bengoechea, J.A. and Alberti, S.
(2010b) 'Klebsiella pneumoniae AcrAB Efflux Pump Contributes to Antimicrobial Resistance and
Virulence', Antimicrobial Agents and Chemotherapy, 54(1), pp. 177.
Page, M.J., Bester, J. and Pretorius, E. (2018) 'The inflammatory effects of TNF-α and complement
component 3 on coagulation', Scientific reports, 8(1), pp. 1812.
Pan, Y., Lin, T., Chen, Y., Hsu, C., Hsieh, P., Wu, M. and Wang, J. (2013) 'Capsular types of Klebsiella
pneumoniae revisited by wzc sequencing', PLoS One, 8(12), pp. e80670.
Pan, P.C., Chen, H.W., Wu, P.K., Wu, Y.Y., Lin, C.H. and Wu, J.H. (2011) 'Mutation in fucose synthesis
gene of Klebsiella pneumoniae affects capsule composition and virulence in mice', Experimental
biology and medicine (Maywood, N.J.), 236(2), pp. 219-226.
Pan, Y.J., Fang, H.C., Yang, H.C., Lin, T.L., Hsieh, P.F., Tsai, F.C., Keynan, Y. and Wang, J.T. (2008)
'Capsular polysaccharide synthesis regions in Klebsiella pneumoniae serotype K57 and a new
capsular serotype', Journal of clinical microbiology, 46(7), pp. 2231-2240.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
262
Papanikou, E., Karamanou, S. and Economou, A. (2007) 'Bacterial protein secretion through the
translocase nanomachine', Nature Reviews Microbiology, 5(11), pp. 839-851.
Parameswaran, N. and Patial, S. (2010) 'Tumor necrosis factor-α signaling in macrophages', Critical
Reviews™ in Eukaryotic Gene Expression, 20(2).
Parizad, E.G., Parizad, E.G., Pakzad, I. and Valizadeh, A. (2016) 'A Review of Secretion Systems in
Pathogenic and Non-Pathogenic Bacteria', Biosciences Biotechnology Research Asia, 13(1), pp. 135-
145.
Parkin, J. and Cohen, B. (2001) 'An overview of the immune system', The Lancet, 357(9270), pp. 1777-
1789.
Parsons, D.A. and Heffron, F. (2005) 'sciS, an icmF homolog in Salmonella enterica serovar
Typhimurium, limits intracellular replication and decreases virulence', Infection and immunity, 73(7),
pp. 4338-4345.
Partridge, S.R. (2011a) 'Analysis of antibiotic resistance regions in Gram-negative bacteria', FEMS
microbiology reviews, 35(5), pp. 820-855.
Partridge, S.R. (2011b) 'Analysis of antibiotic resistance regions in Gram-negative bacteria', FEMS
microbiology reviews, 35(5), pp. 820-855.
Partridge, S.R., Zong, Z. and Iredell, J.R. (2011) 'Recombination in IS26 and Tn2 in the evolution of
multiresistance regions carrying blaCTX-M-15 on conjugative IncF plasmids from Escherichia coli',
Antimicrobial Agents and Chemotherapy, 55(11), pp. 4971-4978.
Paterson, D.L. and Bonomo, R.A. (2005) 'Extended-spectrum beta-lactamases: a clinical update', Clinical
microbiology reviews, 18(4), pp. 657-686.
Pedersen, W.L., Chakrabarty, K., Klucas, R.V. and Vidaver, A.K. (1978) 'Nitrogen fixation (acetylene
reduction) associated with roots of winter wheat and sorghum in Nebraska', Applied and
Environmental Microbiology, 35(1), pp. 129-135.
Pell, L.G., Kanelis, V., Donaldson, L.W., Howell, P.L. and Davidson, A.R. (2009) 'The phage lambda
major tail protein structure reveals a common evolution for long-tailed phages and the type VI
bacterial secretion system', Proceedings of the National Academy of Sciences of the United States of
America, 106(11), pp. 4160-4165.
Perry, R.D. and San Clemente, C.L. (1979) 'Siderophore synthesis in Klebsiella pneumoniae and Shigella
sonnei during iron deficiency', Journal of Bacteriology, 140(3), pp. 1129-1132.
Peters, B.M., Jabra-Rizk, M.A., O'May, G.A., Costerton, J.W. and Shirtliff, M.E. (2012) 'Polymicrobial
interactions: impact on pathogenesis and human disease', Clinical microbiology reviews, 25(1), pp.
193-213.
Pfaffl, M.W. (2001) 'A new mathematical model for relative quantification in real-time RT-PCR', Nucleic
acids research, 29(9), pp. e45.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
263
Picking, W.L., Nishioka, H., Hearn, P.D., Baxter, M.A., Harrington, A.T., Blocker, A. and Picking, W.D.
(2005) 'IpaD of Shigella flexneri is independently required for regulation of Ipa protein secretion and
efficient insertion of IpaB and IpaC into host membranes', Infection and immunity, 73(3), pp. 1432-
1440.
Piddock, L.J. (2006) 'Multidrug-resistance efflux pumps? not just for resistance', Nature Reviews
Microbiology, 4(8), pp. 629-636.
Podschun, R., Acktun, H., Okpara, J., Linderkamp, O., Ullmann, U. and Borneff-Lipp, M. (1998)
'Isolation of Klebsiella planticola from newborns in a neonatal ward', Journal of clinical
microbiology, 36(8), pp. 2331-2332.
Podschun, R., Pietsch, S., Holler, C. and Ullmann, U. (2001) 'Incidence of Klebsiella species in surface
waters and their expression of virulence factors', Applied and Environmental Microbiology, 67(7),
pp. 3325-3327.
Podschun, R. and Ullmann, U. (1998) 'Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy,
typing methods, and pathogenicity factors', Clinical microbiology reviews, 11(4), pp. 589-603.
Pohlner, J., Halter, R. and Meyer, T. (1988) 'Neisseria gonorrhoeae IgA protease. Secretion and
implications for pathogenesis', in Anonymous Gonococci and Meningococci. Springer, pp. 427-432.
Pohlner, J., Halter, R., Beyreuther, K. and Meyer, T.F. (1987) 'Gene structure and extracellular secretion
of Neisseria gonorrhoeae IgA protease', Nature, 325(6103), pp. 458-462.
Poirel, L., Heritier, C., Tolun, V. and Nordmann, P. (2004) 'Emergence of oxacillinase-mediated
resistance to imipenem in Klebsiella pneumoniae', Antimicrobial Agents and Chemotherapy, 48(1),
pp. 15-22.
Pollack, M., Nieman, R., Reinhardt, J., Charache, P., Jett, M. and Hardy, P. (1972) 'Factors influencing
colonisation and antibiotic-resistance patterns of gram-negative bacteria in hospital patients', The
Lancet, 300(7779), pp. 668-671.
Popa, C., Netea, M.G., van Riel, P.L., van der Meer, J.W. and Stalenhoef, A.F. (2007) 'The role of TNF-
alpha in chronic inflammatory conditions, intermediary metabolism, and cardiovascular risk',
Journal of lipid research, 48(4), pp. 751-762.
Potgieter, P.D. and Hammond, J. (1992) 'Etiology and diagnosis of pneumonia requiring ICU admission.',
CHEST Journal, 101(1), pp. 199-203.
Potvin, E., Lehoux, D.E., Kukavica‐Ibrulj, I., Richard, K.L., Sanschagrin, F., Lau, G.W. and Levesque,
R.C. (2003) 'In vivo functional genomics of Pseudomonas aeruginosa for high‐throughput screening
of new virulence factors and antibacterial targets', Environmental microbiology, 5(12), pp. 1294-
1308.
Pradel, N., Ye, C., Livrelli, V., Xu, J., Joly, B. and Wu, L.F. (2003) 'Contribution of the twin arginine
translocation system to the virulence of enterohemorrhagic Escherichia coli O157:H7', Infection and
immunity, 71(9), pp. 4908-4916.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
264
Price, S.B., Cowan, C., Perry, R.D. and Straley, S.C. (1991) 'The Yersinia pestis V antigen is a regulatory
protein necessary for Ca2(+)-dependent growth and maximal expression of low-Ca2+ response
virulence genes', Journal of Bacteriology, 173(8), pp. 2649-2657.
Proost, P., Wuyts, A. and Van Damme, J. (1996) 'Human monocyte chemotactic proteins‐2 and‐3:
structural and functional comparison with MCP‐1', Journal of leukocyte biology, 59(1), pp. 67-74.
Pugsley, A.P., Chapon, C. and Schwartz, M. (1986) 'Extracellular pullulanase of Klebsiella pneumoniae is
a lipoprotein', Journal of Bacteriology, 166(3), pp. 1083-1088.
Pukatzki, S., Ma, A.T., Sturtevant, D., Krastins, B., Sarracino, D., Nelson, W.C., Heidelberg, J.F. and
Mekalanos, J.J. (2006a) 'Identification of a conserved bacterial protein secretion system in Vibrio
cholerae using the Dictyostelium host model system', Proceedings of the National Academy of
Sciences of the United States of America, 103(5), pp. 1528-1533.
Pukatzki, S., Ma, A.T., Revel, A.T., Sturtevant, D. and Mekalanos, J.J. (2007) 'Type VI Secretion System
Translocates a Phage Tail Spike-Like Protein into Target Cells Where It Cross-Links Actin',
Proceedings of the National Academy of Sciences of the United States of America, 104(39), pp.
15508-15513.
Pukatzki, S., Ma, A.T., Sturtevant, D., Krastins, B., Sarracino, D., Nelson, W.C., Heidelberg, J.F. and
Mekalanos, J.J. (2006b) 'Identification of a Conserved Bacterial Protein Secretion System in Vibrio
cholerae Using the Dictyostelium Host Model System', Proceedings of the National Academy of
Sciences of the United States of America, 103(5), pp. 1528-1533.
Qi, Y., Wei, Z., Ji, S., Du, X., Shen, P. and Yu, Y. (2010) 'ST11, the dominant clone of KPC-producing
Klebsiella pneumoniae in China', Journal of Antimicrobial Chemotherapy, 66(2), pp. 307-312.
Rahn, A., Drummelsmith, J. and Whitfield, C. (1999) 'Conserved organization in the cps gene clusters for
expression of Escherichia coli group 1 K antigens: relationship to the colanic acid biosynthesis locus
and the cps genes from Klebsiella pneumoniae', Journal of Bacteriology, 181(7), pp. 2307-2313.
Ramirez, M.S., Traglia, G.M., Lin, D.L., Tran, T. and Tolmasky, M.E. (2014) 'Plasmid-Mediated
Antibiotic Resistance and Virulence in Gram-negatives: the Klebsiella pneumoniae Paradigm',
Microbiology spectrum, 2(5), pp. 1-15.
Ramos, P.I.P., Picão, R.C., Vespero, E.C., Pelisson, M., Zuleta, L.F.G., Almeida, L.G.P., Gerber, A.L.,
Vasconcelos, A.T.R., Gales, A.C. and Nicolás, M.F. (2012) 'Pyrosequencing-based analysis reveals
a novel capsular gene cluster in a KPC-producing Klebsiella pneumoniae clinical isolate identified in
Brazil', BMC microbiology, 12(1), pp. 1.
Reddy, J.D., Reddy, S.L., Hopkins, D.L. and Gabriel, D.W. (2007) 'TolC is required for pathogenicity of
Xylella fastidiosa in Vitis vinifera grapevines', Molecular Plant-Microbe Interactions, 20(4), pp.
403-410.
Reese, M.G. (2001) 'Application of a time-delay neural network to promoter annotation in the Drosophila
melanogaster genome', Computers & chemistry, 26(1), pp. 51-56.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
265
Reese, M.G. (2000) PhD thesis. University of California, Berkeley, CA/University of Hohenheim,
Stuttgart, Germany.
Regue, M., Izquierdo, L., Fresno, S., Pique, N., Corsaro, M.M., Naldi, T., De Castro, C., Waidelich, D.,
Merino, S. and Tomas, J.M. (2005) 'A second outer-core region in Klebsiella pneumoniae
lipopolysaccharide', Journal of Bacteriology, 187(12), pp. 4198-4206.
Regueiro, V., Moranta, D., Frank, C.G., Larrarte, E., Margareto, J., March, C., Garmendia, J. and
Bengoechea, J.A. (2011) 'Klebsiella pneumoniae subverts the activation of inflammatory responses
in a NOD1‐dependent manner', Cellular microbiology, 13(1), pp. 135-153.
Ren, M., Guo, Q., Guo, L., Lenz, M., Qian, F., Koenen, R.R., Xu, H., Schilling, A.B., Weber, C., Ye,
R.D., Dinner, A.R. and Tang, W.J. (2010) 'Polymerization of MIP-1 chemokine (CCL3 and CCL4)
and clearance of MIP-1 by insulin-degrading enzyme', The EMBO journal, 29(23), pp. 3952-3966.
Repizo, G.D., Gagné, S., Foucault-Grunenwald, M., Borges, V., Charpentier, X., Limansky, A.S., Gomes,
J.P., Viale, A.M. and Salcedo, S.P. (2015a) 'Differential Role of the T6SS in Acinetobacter
baumannii Virulence', PloS one, 10(9), pp. e0138265.
Repizo, G.D., Gagné, S., Foucault-Grunenwald, M., Borges, V., Charpentier, X., Limansky, A.S., Gomes,
J.P., Viale, A.M. and Salcedo, S.P. (2015b) 'Differential Role of the T6SS in Acinetobacter
baumannii Virulence: e0138265', PLoS One, 10(9).
Repizo, G.D., Gagné, S., Foucault-Grunenwald, M., Borges, V., Charpentier, X., Limansky, A.S., Gomes,
J.P., Viale, A.M. and Salcedo, S.P. (2015c) 'Differential Role of the T6SS in Acinetobacter
baumannii Virulence: e0138265', PLoS One, 10(9).
Repizo, G.D., Gagné, S., Foucault-Grunenwald, M., Borges, V., Charpentier, X., Limansky, A.S., Gomes,
J.P., Viale, A.M. and Salcedo, S.P. (2015d) 'Differential Role of the T6SS in Acinetobacter
baumannii Virulence: e0138265', PLoS One, 10(9).
Richards, M.J., Edwards, J.R., Culver, D.H. and Gaynes, R.P. (2000) 'Nosocomial infections in combined
medical-surgical intensive care units in the United States', Infection Control & Hospital
Epidemiology, 21(08), pp. 510-515.
Richards, M.J., Edwards, J.R., Culver, D.H. and Gaynes, R.P. (1999) 'Nosocomial infections in medical
intensive care units in the United States. National Nosocomial Infections Surveillance System',
Critical Care Medicine, 27(5), pp. 887-892.
Robinson, C. and Bolhuis, A. (2004) 'Tat-dependent protein targeting in prokaryotes and chloroplasts',
Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 1694(1), pp. 135-147.
Rose, H. and Schreier, J. (1968) 'The effect of hospitalization and antibiotic therapy on the gram-negative
fecal flora.', The American Journal of the Medical Sciences, 255(4), pp. 228-236.
Rosen, D.A., Pinkner, J.S., Walker, J.N., Elam, J.S., Jones, J.M. and Hultgren, S.J. (2008) 'Molecular
variations in Klebsiella pneumoniae and Escherichia coli FimH affect function and pathogenesis in
the urinary tract', Infection and immunity, 76(7), pp. 3346-3356.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
266
Rosenblueth, M., Martínez, L., Silva, J. and Martínez-Romero, E. (2004) 'Klebsiella variicola, a novel
species with clinical and plant-associated isolates', Systematic and applied microbiology, 27(1), pp.
27.
Ross, J. and Auger, M.'The biology of the macrophage. Burke B, Lewis CE, editors', .
Rossi, D. and Zlotnik, A. (2000) 'The biology of chemokines and their receptors', Annual Review of
Immunology, 18(1), pp. 217-242.
Rossier, O. and Cianciotto, N.P. (2005) 'The Legionella pneumophila tatB gene facilitates secretion of
phospholipase C, growth under iron-limiting conditions, and intracellular infection', Infection and
immunity, 73(4), pp. 2020-2032.
Ruiz-Perez, F., Henderson, I.R., Leyton, D.L., Rossiter, A.E., Zhang, Y. and Nataro, J.P. (2009) 'Roles of
periplasmic chaperone proteins in the biogenesis of serine protease autotransporters of
Enterobacteriaceae', Journal of Bacteriology, 191(21), pp. 6571-6583.
Russell, A.B., Peterson, S.B. and Mougous, J.D. (2014) 'Type VI secretion system effectors: poisons with
a purpose', Nature Reviews Microbiology, 12(2), pp. 137-148.
Russell, A.B., Hood, R.D., Bui, N.K., LeRoux, M., Vollmer, W. and Mougous, J.D. (2011a) 'Type VI
secretion delivers bacteriolytic effectors to target cells', Nature, 475(7356), pp. 343-347.
Russell, A.B., Hood, R.D., Bui, N.K., LeRoux, M., Vollmer, W. and Mougous, J.D. (2011b) 'Type VI
secretion delivers bacteriolytic effectors to target cells', Nature, 475(7356), pp. 343-347.
Russell, A.B., Hood, R.D., Bui, N.K., LeRoux, M., Vollmer, W. and Mougous, J.D. (2011c) 'Type VI
secretion delivers bacteriolytic effectors to target cells', Nature, 475(7356), pp. 343-347.
Russell, A.B., LeRoux, M., Hathazi, K., Agnello, D.M., Ishikawa, T., Wiggins, P.A., NyuntWai, S.,
Mougous, J.D., Molekylär Infektionsmedicin, S.(., Institutionen för molekylärbiologi (Medicinska
fakulteten), Medicinska fakulteten and Umeå universitet (2013) 'Diverse type VI secretion
phospholipases are functionally plastic antibacterial effectors', Nature, 496(7446), pp. 508-512.
Russotto, V., Cortegiani, A., Raineri, S.M. and Giarratano, A. (2015) 'Bacterial contamination of
inanimate surfaces and equipment in the intensive care unit', Journal of Intensive Care, 3(54).
Saenghirunvattana, S., Charoenpan, P., Kiatboonsri, S. and Aeursudkij, B. (1991) 'Nosocomial
pneumonias in Thailand.', Group, .
Sahly, H., Podschun, R. and Ullmann, U. (2002) 'Klebsiella infections in the immunocompromised host',
in Anonymous The Biology and Pathology of Innate Immunity Mechanisms. Springer, pp. 237-249.
Sahly, H., Aucken, H., Benedí, V.J., Forestier, C., Fussing, V., Hansen, D.S., Ofek, I., Podschun, R.,
Sirot, D., Tomás, J.M., Sandvang, D. and Ullmann, U. (2004) 'Increased Serum Resistance in
Klebsiella pneumoniae Strains Producing Extended-Spectrum β-Lactamases', Antimicrobial Agents
and Chemotherapy, 48(9), pp. 3477-3482.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
267
Sahly, H., Navon-Venezia, S., Roesler, L., Hay, A., Carmeli, Y., Podschun, R., Hennequin, C., Forestier,
C. and Ofek, I. (2008) 'Extended-spectrum beta-lactamase production is associated with an increase
in cell invasion and expression of fimbrial adhesins in Klebsiella pneumoniae', Antimicrobial Agents
and Chemotherapy, 52(9), pp. 3029-3034.
Sahly, H., Podschun, R., Oelschlaeger, T.A., Greiwe, M., Parolis, H., Hasty, D., Kekow, J., Ullmann, U.,
Ofek, I. and Sela, S. (2000) 'Capsule impedes adhesion to and invasion of epithelial cells by
Klebsiella pneumoniae', Infection and immunity, 68(12), pp. 6744-6749.
Saier Jr, M.H. (2006) 'Protein Secretion and Membrane Insertion Systems in Gram-Negative Bacteria',
Journal of Membrane Biology, 214(1), pp. 75-90.
Salamov, V.S.A. and Solovyevand, A. (2011) 'Automatic annotation of microbial genomes and
metagenomic sequences', Metagenomics and its applications in agriculture.Nova Science
Publishers, Hauppauge, , pp. 61-78.
Salit, I.E. and Gotschlich, E.C. (1977) 'Hemagglutination by purified type I Escherichia coli pili', The
Journal of experimental medicine, 146(5), pp. 1169-1181.
San Millan, R.M., Martinez-Ballesteros, I., Rementeria, A., Garaizar, J. and Bikandi, J. (2013) 'Online
exercise for the design and simulation of PCR and PCR-RFLP experiments.(Report)', BMC
Research Notes, 6(1).
Sana, T.G., Hachani, A., Bucior, I., Soscia, C., Garvis, S., Termine, E., Engel, J., Filloux, A. and Bleves,
S. (2012a) 'The second type VI secretion system of Pseudomonas aeruginosa strain PAO1 is
regulated by quorum sensing and fur and modulates internalization in epithelial cells', Journal of
Biological Chemistry, 287(32), pp. 27095-27105.
Sana, T.G., Hachani, A., Bucior, I., Soscia, C., Garvis, S., Termine, E., Engel, J., Filloux, A. and Bleves,
S. (2012b) 'The second type VI secretion system of Pseudomonas aeruginosa strain PAO1 is
regulated by quorum sensing and Fur and modulates internalization in epithelial cells', The Journal
of biological chemistry, 287(32), pp. 27095-27105.
Sanchez, C. (2011) 'Vibrio cholerae beats the competition', Nature Reviews Microbiology, 9(1), pp. 3.
Sandkvist, M., Michel, L.O., Hough, L.P., Morales, V.M., Bagdasarian, M., Koomey, M., DiRita, V.J.
and Bagdasarian, M. (1997) 'General secretion pathway (eps) genes required for toxin secretion and
outer membrane biogenesis in Vibrio cholerae', Journal of Bacteriology, 179(22), pp. 6994-7003.
Sargent, F., Stanley, N.R., Berks, B.C. and Palmer, T. (1999) 'Sec-independent protein translocation in
Escherichia coli. A distinct and pivotal role for the TatB protein', The Journal of biological
chemistry, 274(51), pp. 36073-36082.
Sarris, P.F., Skandalis, N., Kokkinidis, M. and Panopoulos, N.J. (2010) 'In silico analysis reveals multiple
putative type VI secretion systems and effector proteins in Pseudomonas syringae pathovars',
Molecular Plant Pathology, 11(6), pp. 795-804.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
268
Sarris, P.F., Zoumadakis, C., Panopoulos, N.J. and Scoulica, E.V. (2011a) 'Distribution of the putative
type VI secretion system core genes in Klebsiella spp', Infection, genetics and evolution : journal of
molecular epidemiology and evolutionary genetics in infectious diseases, 11(1), pp. 157-166.
Sarris, P.F., Zoumadakis, C., Panopoulos, N.J. and Scoulica, E.V. (2011b) 'Distribution of the putative
type VI secretion system core genes in Klebsiella spp.', Infection, Genetics and Evolution, 11(1), pp.
157-166.
Satchell, K.J. (2009) 'Actin crosslinking toxins of Gram-negative bacteria', Toxins, 1(2), pp. 123-133.
Sauvonnet, N., Vignon, G., Pugsley, A.P. and Gounon, P. (2000) 'Pilus formation and protein secretion by
the same machinery in Escherichia coli', The EMBO journal, 19(10), pp. 2221-2228.
Schell, M.A., Ulrich, R.L., Ribot, W.J., Brueggemann, E.E., Hines, H.B., Chen, D., Lipscomb, L., Kim,
H.S., Mrázek, J., Nierman, W.C. and DeShazer, D. (2007) 'Type VI secretion is a major virulence
determinant in Burkholderia mallei', Molecular microbiology, 64(6), pp. 1466-1485.
Scheller, J., Chalaris, A., Schmidt-Arras, D. and Rose-John, S. (2011) 'The pro-and anti-inflammatory
properties of the cytokine interleukin-6', Biochimica et Biophysica Acta (BBA)-Molecular Cell
Research, 1813(5), pp. 878-888.
Schembri, M.A., Blom, J., Krogfelt, K.A. and Klemm, P. (2005) 'Capsule and fimbria interaction in
Klebsiella pneumoniae', Infection and immunity, 73(8), pp. 4626-4633.
Schleimer, R.P., Kato, A., Kern, R., Kuperman, D. and Avila, P.C. (2007) 'Epithelium: at the interface of
innate and adaptive immune responses', The Journal of allergy and clinical immunology, 120(6), pp.
1279-1284.
Schroeter, J. (ed.) (1889) Kriptogamen Flora von Schlesien.
Schroeter, J. (1886) 'In: F. Cohn, Kriptogamen Flora von Schlesien.', 3, pp. 131-256.
Schroll, C., Barken, K.B., Krogfelt, K.A. and Struve, C. (2010) 'Role of type 1 and type 3 fimbriae in
Klebsiella pneumoniae biofilm formation', BMC microbiology, 10(1), pp. 179-179.
Schubert, D., Lechtenberg, B., Forsbach, A., Gils, M., Bahadur, S. and Schmidt, R. (2004) 'Silencing in
Arabidopsis T-DNA transformants: the predominant role of a gene-specific RNA sensing
mechanism versus position effects', The Plant Cell, 16(10), pp. 2561-2572.
Schwarz, S., Singh, P., Robertson, J.D., LeRoux, M., Skerrett, S.J., Goodlett, D.R., West, T.E. and
Mougous, J.D. (2014) 'VgrG-5 is a Burkholderia type VI secretion system-exported protein required
for multinucleated giant cell formation and virulence', Infection and immunity, 82(4), pp. 1445-1452.
Schwarz, S., West, T.E., Boyer, F., Chiang, W., Carl, M.A., Hood, R.D., Rohmer, L., Tolker-Nielsen, T.,
Skerrett, S.J. and Mougous, J.D. (2010a) 'Burkholderia type vi secretion systems have distinct roles
in eukaryotic and bacterial cell interactions', PLoS Pathogens, 6(8), pp. 77-78.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
269
Schwarz, S., West, T.E., Boyer, F., Chiang, W., Carl, M.A., Hood, R.D., Rohmer, L., Tolker-Nielsen, T.,
Skerrett, S.J. and Mougous, J.D. (2010b) 'Burkholderia type vi secretion systems have distinct roles
in eukaryotic and bacterial cell interactions', PLoS Pathogens, 6(8), pp. 77-78.
Schwarz, S., West, T.E., Boyer, F., Chiang, W., Carl, M.A., Hood, R.D., Rohmer, L., Tolker-Nielsen, T.,
Skerrett, S.J. and Mougous, J.D. (2010c) 'Burkholderia type VI secretion systems have distinct roles
in eukaryotic and bacterial cell interactions', PLoS pathogens, 6(8), pp. e1001068.
Seaton, D. (2000) 'Pneumonia', in Seaton, D., Seaton and Leitch A.G. (eds.) Crofton and Douglas's
respiratory diseases
. 1st edn. Malden, MA.: Blackwell Science, pp. 406-407.
Seidler, R.J. (1981) 'The genus Klebsiella (nonmedical aspects)', The Prokaryotes: a handbook on
habitats, isolation, and identification of bacteria.Berlin: Springer-Verlag, , pp. 1166-1172.
SELDEN, R., Lee, S., WANG, W.L.L., BENNETT, J.V. and EICKHOFF, T.C. (1971) 'Nosocomial
Klebsiella infections: intestinal colonization as a reservoir', Annals of Internal Medicine, 74(5), pp.
657-664.
Sexton, J.A., Miller, J.L., Yoneda, A., Kehl-Fie, T.E. and Vogel, J.P. (2004) 'Legionella pneumophila
DotU and IcmF are required for stability of the Dot/Icm complex', Infection and immunity, 72(10),
pp. 5983-5992.
Shalom, G., Shaw, J.G. and Thomas, M.S. (2007) 'In vivo expression technology identifies a type VI
secretion system locus in Burkholderia pseudomallei that is induced upon invasion of macrophages',
Microbiology, 153(8), pp. 2689-2699.
Shankar-Sinha, S., Valencia, G.A., Janes, B.K., Rosenberg, J.K., Whitfield, C., Bender, R.A., Standiford,
T.J. and Younger, J.G. (2004a) 'The Klebsiella pneumoniae O antigen contributes to bacteremia and
lethality during murine pneumonia', Infection and immunity, 72(3), pp. 1423-1430.
Shankar-Sinha, S., Valencia, G.A., Janes, B.K., Rosenberg, J.K., Whitfield, C., Bender, R.A., Standiford,
T.J. and Younger, J.G. (2004b) 'The Klebsiella pneumoniae O antigen contributes to bacteremia and
lethality during murine pneumonia', Infection and immunity, 72(3), pp. 1423-1430.
Sharma, S.K., Fatma, T. and Thukral, S.S. (1999) 'A simple and rapid serum bactericidal assay and its
evaluation in clinical isolates of Klebsiella pneumoniae', Journal of microbiological methods, 39(1),
pp. 45-48.
Sherry, B., Tekamp-Olson, P., Gallegos, C., Bauer, D., Davatelis, G., Wolpe, S.D., Masiarz, F., Coit, D.
and Cerami, A. (1988) 'Resolution of the two components of macrophage inflammatory protein 1,
and cloning and characterization of one of those components, macrophage inflammatory protein 1
beta', The Journal of experimental medicine, 168(6), pp. 2251-2259.
Shevchik, V.E., Robert-Baudouy, J. and Condemine, G. (1997) 'Specific interaction between OutD, an
Erwinia chrysanthemi outer membrane protein of the general secretory pathway, and secreted
proteins', The EMBO journal, 16(11), pp. 3007-3016.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
270
Shin, J. and Soo Ko, K. (2014) 'Single origin of three plasmids bearing blaCTX-M-15 from different
Klebsiella pneumoniae clones', The Journal of antimicrobial chemotherapy, 69(4), pp. 969-972.
Shneider, M.M., Buth, S.A., Ho, B.T., Basler, M., Mekalanos, J.J. and Leiman, P.G. (2013a) 'PAAR-
repeat proteins sharpen and diversify the type VI secretion system spike', Nature, 500(7462), pp.
350-353.
Shneider, M.M., Buth, S.A., Ho, B.T., Basler, M., Mekalanos, J.J. and Leiman, P.G. (2013b) 'PAAR-
repeat proteins sharpen and diversify the type VI secretion system spike', Nature, 500(7462), pp.
350-353.
Shon, A.S., Bajwa, R.P. and Russo, T.A. (2013) 'Hypervirulent (hypermucoviscous) Klebsiella
pneumoniae: a new and dangerous breed', Virulence, 4(2), pp. 107-118.
Shon, A.S. and Russo, T.A. (2012) 'Hypervirulent Klebsiella pneumoniae: the next superbug?', Future
microbiology, 7(6), pp. 669-671.
Short, J.M., Fernandez, J.M., Sorge, J.A. and Huse, W.D. (1988) 'λ ZAP: a bacteriophage λ expression
vector with in vivo excision properties', Nucleic acids research, 16(15), pp. 7583-7600.
Shrivastav, A., Lee, J., Kim, H.Y. and Kim, Y.R. (2013) 'Recent insights in the removal of Klebseilla
pathogenicity factors for the industrial production of 2,3-butanediol', Journal of microbiology and
biotechnology, 23(7), pp. 885-896.
Shrivastava, S. and Mande, S.S. (2008) 'Identification and functional characterization of gene components
of Type VI Secretion system in bacterial genomes', PloS one, 3(8), pp. e2955.
Shu, H., Fung, C., Liu, Y., Wu, K., Chen, Y., Li, L., Liu, T., Kirby, R. and Tsai, S. (2009) 'Genetic
diversity of capsular polysaccharide biosynthesis in Klebsiella pneumoniae clinical isolates',
Microbiology, 155(12), pp. 4170-4183.
Shyntum, D.Y., Venter, S.N., Moleleki, L.N., Toth, I. and Coutinho, T.A. (2014) 'Comparative genomics
of type VI secretion systems in strains of Pantoea ananatis from different environments', BMC
genomics, 15(1), pp. 1.
Siddiqui, R., Lakhundi, S. and Khan, N.A. (2015) 'Interactions of Pseudomonas aeruginosa and
Corynebacterium spp. with non-phagocytic brain microvascular endothelial cells and phagocytic
Acanthamoeba castellanii', Parasitology research, 114(6), pp. 2349-2356.
Siddiqui, R., Malik, H., Sagheer, M., Jung, S. and Khan, N.A. (2011) 'The type III secretion system is
involved in Escherichia coli K1 interactions with Acanthamoeba', Experimental parasitology,
128(4), pp. 409-413.
Silverman, J.M., Brunet, Y.R., Cascales, E. and Mougous, J.D. (2012a) 'Structure and regulation of the
type VI secretion system', Annual Review of Microbiology, 66, pp. 453-472.
Silverman, J.M., Brunet, Y.R., Cascales, E. and Mougous, J.D. (2012b) 'Structure and regulation of the
type VI secretion system', Annual Review of Microbiology, 66, pp. 453-472.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
271
Sirot, D., Sirot, J., Labia, R., Morand, A., Courvalin, P., Darfeuille-Michaud, A., Perroux, R. and Cluzel,
R. (1987) 'Transferable resistance to third-generation cephalosporins in clinical isolates of Klebsiella
pneumoniae: identification of CTX-1, a novel beta-lactamase', The Journal of antimicrobial
chemotherapy, 20(3), pp. 323-334.
Sistrunk, J.R., Nickerson, K.P., Chanin, R.B., Rasko, D.A. and Faherty, C.S. (2016) 'Survival of the
Fittest: How Bacterial Pathogens Utilize Bile To Enhance Infection', Clinical microbiology reviews,
29(4), pp. 819-836.
Sjogren, R.E. and Gibson, M.J. (1981) 'Bacterial survival in a dilute environment', Applied and
Environmental Microbiology, 41(6), pp. 1331-1336.
Skerman, V.B.D., McGowan, V. and Sneath, P.H.A. (1989) Approve Lists of Bacterial Names. Amended
edition edn. Washington: American Society for Microbiology.
Söderhäll, K. and Cerenius, L. (1998) 'Role of the prophenoloxidase-activating system in invertebrate
immunity', Current opinion in immunology, 10(1), pp. 23-28.
Son, D., Parl, A.K., Montgomery Rice, V. and Khabele, D. (2007) 'Keratinocyte chemoattractant
(KC)/human growth-regulated oncogene (GRO) chemokines and pro-inflammatory chemokine
networks in mouse and human ovarian epithelial cancer cells', Cancer biology & therapy, 6(8), pp.
1308-1318.
Songer, J.G. (1997) 'Bacterial phospholipases and their role in virulence', Trends in microbiology, 5(4),
pp. 156-161.
Souza, R.C., Saji, Guadalupe del Rosario Quispe, Costa, M.O., Netto, D.S., Lima, N.C., Klein, C.C.,
Vasconcelos, A.T.R. and Nicolás, M.F. (2012) 'AtlasT4SS: a curated database for type IV secretion
systems', BMC microbiology, 12(1), pp. 1.
Srinivasan, V.B., Venkataramaiah, M., Mondal, A., Vaidyanathan, V., Govil, T. and Rajamohan, G.
(2012) 'Functional characterization of a novel outer membrane porin KpnO, regulated by PhoBR
two-component system in Klebsiella pneumoniae NTUH-K2044', PloS one, 7(7), pp. e41505.
Stahlhut, S.G., Struve, C. and Krogfelt, K.A. (2012) 'Klebsiella pneumoniae type 3 fimbriae agglutinate
yeast in a mannose-resistant manner', Journal of medical microbiology, 61(3), pp. 317-322.
Stahlhut, S.G., Struve, C., Krogfelt, K.A. and Reisner, A. (2012) 'Biofilm formation of Klebsiella
pneumoniae on urethral catheters requires either type 1 or type 3 fimbriae', FEMS Immunology &
Medical Microbiology, 65(2), pp. 350-359.
Stanke, F. (2015) 'The Contribution of the Airway Epithelial Cell to Host Defense', Mediators of
inflammation, 2015, pp. 463016.
Steinert, M. and Heuner, K. (2005) 'Dictyostelium as host model for pathogenesis', Cellular microbiology,
7(3), pp. 307-314.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
272
Struve, C., Bojer, M. and Krogfelt, K.A. (2009) 'Identification of a conserved chromosomal region
encoding Klebsiella pneumoniae type 1 and type 3 fimbriae and assessment of the role of fimbriae in
pathogenicity', Infection and immunity, 77(11), pp. 5016-5024.
Struve, C. and Krogfelt, K.A. (2003) 'Role of capsule in Klebsiella pneumoniae virulence: lack of
correlation between in vitro and in vivo studies', FEMS microbiology letters, 218(1), pp. 149-154.
Struve, C., Roe, C.C., Stegger, M., Stahlhut, S.G., Hansen, D.S., Engelthaler, D.M., Andersen, P.S.,
Driebe, E.M., Keim, P. and Krogfelt, K.A. (2015) 'Mapping the Evolution of Hypervirulent
Klebsiella pneumoniae', mBio, 6(4), pp. e00630-15.
Stubbendieck, R.M., Vargas-Bautista, C. and Straight, P.D. (2016) 'Bacterial communities: interactions to
scale', Frontiers in microbiology, 7, pp. 1234.
Suarez, G., Sierra, J.C., Sha, J., Wang, S., Erova, T.E., Fadl, A.A., Foltz, S.M., Horneman, A.J. and
Chopra, A.K. (2008) 'Molecular characterization of a functional type VI secretion system from a
clinical isolate of Aeromonas hydrophila', Microbial pathogenesis, 44(4), pp. 344-361.
Suarez, G., Sierra, J.C., Erova, T.E., Sha, J., Horneman, A.J. and Chopra, A.K. (2010a) 'A type VI
secretion system effector protein, VgrG1, from Aeromonas hydrophila that induces host cell toxicity
by ADP ribosylation of actin', Journal of Bacteriology, 192(1), pp. 155-168.
Suarez, G., Sierra, J.C., Kirtley, M.L. and Chopra, A.K. (2010b) 'Role of Hcp, a type 6 secretion system
effector, of Aeromonas hydrophila in modulating activation of host immune cells', Microbiology,
156(12), pp. 3678-3688.
Sun, W.W., Syu, W., Ho, W., Lin, C., Tsai, S. and Wang, S. (2014) 'SitA contributes to the virulence of
Klebsiella pneumoniae in a mouse infection model', Microbes and Infection, 16(2), pp. 161-170.
Symmons, M.F., Bokma, E., Koronakis, E., Hughes, C. and Koronakis, V. (2009) 'The assembled
structure of a complete tripartite bacterial multidrug efflux pump', Proceedings of the National
Academy of Sciences of the United States of America, 106(17), pp. 7173-7178.
Tablan, O.C., Anderson, L.J., Arden, N.H., Breiman, R.F., Butler, J.C., McNeil, M.M. and Pearson, M.L.
(1997) 'Guidelines for prevention of nosocomial pneumonia', Morbidity and Mortality Weekly
Report: Recommendations and Reports, , pp. i-79.
Tablan, O.C., Anderson, L.J., Arden, N.H., Breiman, R.P., Butler, J.C. and McNeil, M.M. (1994)
'Guideline for prevention of nosocomial pneumonia', Infection Control & Hospital Epidemiology,
15(09), pp. 588-627.
Tan, K.S., Chen, Y., Lim, Y.C., Tan, G.Y., Liu, Y., Lim, Y.T., Macary, P. and Gan, Y.H. (2010)
'Suppression of host innate immune response by Burkholderia pseudomallei through the virulence
factor TssM', Journal of immunology (Baltimore, Md.: 1950), 184(9), pp. 5160-5171.
Tanaka, T., Narazaki, M. and Kishimoto, T. (2014) 'IL-6 in inflammation, immunity, and disease', Cold
Spring Harbor perspectives in biology, 6(10), pp. a016295.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
273
Tauschek, M., Gorrell, R.J., Strugnell, R.A. and Robins-Browne, R.M. (2002) 'Identification of a protein
secretory pathway for the secretion of heat-labile enterotoxin by an enterotoxigenic strain of
Escherichia coli', Proceedings of the National Academy of Sciences of the United States of America,
99(10), pp. 7066-7071.
Thakur, A.K. and Fezio, W.L. (1981) 'A computer program for estimating LD50 and its confidence limits
using modified Behrens-Reed-Muench cumulant method', Drug and chemical toxicology, 4(3), pp.
297-305.
Thomas, C.M. and Nielsen, K.M. (2005) 'Mechanisms of, and barriers to, horizontal gene transfer
between bacteria', Nature reviews microbiology, 3(9), pp. 711.
Thomas, S., Holland, I.B. and Schmitt, L. (2014) 'The type 1 secretion pathway—the hemolysin system
and beyond', Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 1843(8), pp. 1629-
1641.
Tian, Y., Zhao, Y., Wu, X., Liu, F., Hu, B. and Walcott, R.R. (2015) 'The type VI protein secretion
system contributes to biofilm formation and seed‐to‐seedling transmission of A Cidovorax citrulli on
melon', Molecular plant pathology, 16(1), pp. 38-47.
Toleman, M.A. and Walsh, T.R. (2008) 'Evolution of the ISCR3 group of ISCR elements', Antimicrobial
Agents and Chemotherapy, 52(10), pp. 3789-3791.
Tóth, A., Kocsis, B., Damjanova, I., Kristóf, K., Jánvári, L., Pászti, J., Csercsik, R., Topf, J., Szabó, D.
and Hamar, P. (2014) 'Fitness cost associated with resistance to fluoroquinolones is diverse across
clones of Klebsiella pneumoniae and may select for CTX-M-15 type extended-spectrum β-
lactamase', European journal of clinical microbiology & infectious diseases, 33(5), pp. 837-843.
Trevisan, V. (1887) 'Sul micrococco della rabia e sulla possibilità di r iconoscere durante il periodo
d’incubazione, dall’esame del sangue della persona morsicata, se ha contratta l’infezione rabbica. R.
C', Ist. Lombardo, 20(ser. II), pp. 88-105.
Trevisan, V. (1885) 'Caratteri di alcuni nuovi generi di Batteriaceae', Atti. Accad. fis.- med-stat. M ilano
Ser, 4(3), pp. 92-106.
Troisfontaines, P. and Cornelis, G.R. (2005) 'Type III secretion: more systems than you think', Physiology
(Bethesda, Md.), 20, pp. 326-339.
Trokter, M., Felisberto-Rodrigues, C., Christie, P.J. and Waksman, G. (2014) 'Recent advances in the
structural and molecular biology of type IV secretion systems', Current opinion in structural
biology, 27, pp. 16-23.
Tsai, F., Huang, Y., Chang, L. and Wang, J. (2008) 'Pyogenic liver abscess as endemic disease, Taiwan',
Emerg Infect Dis, 14(10), pp. 1592-1600.
Tsai, Y.K., Fung, C.P., Lin, J.C., Chen, J.H., Chang, F.Y., Chen, T.L. and Siu, L.K. (2011) 'Klebsiella
pneumoniae outer membrane porins OmpK35 and OmpK36 play roles in both antimicrobial
resistance and virulence', Antimicrobial Agents and Chemotherapy, 55(4), pp. 1485-1493.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
274
Tseng, T., Tyler, B.M. and Setubal, J.C. (2009) 'Protein secretion systems in bacterial-host associations,
and their description in the Gene Ontology', BMC microbiology, 9(1), pp. 1.
Tugal, D., Lynch, M., Hujer, A.M., Rudin, S., Perez, F. and Bonomo, R.A. (2015) 'Multi-drug-resistant
Klebsiella pneumoniae pancreatitis: a new challenge in a serious surgical infection', Surgical
infections, 16(2), pp. 188-193.
Ungureanu, V.A., Stratakos, A.C., Gundogdu, O., Stef, L., Pet, I., Pet, E., Pacala, N. and Corcionivoschi,
N. (2019) 'Virulence of a T6SS Campylobacter jejuni chicken isolate from North Romania', BMC
Research Notes, 12(1), pp. 180.
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B.C., Remm, M. and Rozen, S.G. (2012)
'Primer3--new capabilities and interfaces', Nucleic acids research, 40(15), pp. e115.
Unterweger, D., Miyata, S.T., Bachmann, V., Brooks, T.M., Mullins, T., Kostiuk, B., Provenzano, D. and
Pukatzki, S. (2014a) 'The Vibrio cholerae type VI secretion system employs diverse effector
modules for intraspecific competition', Nature communications, 5, pp. 3549.
Unterweger, D., Kitaoka, M., Miyata, S.T., Bachmann, V., Brooks, T.M., Moloney, J., Sosa, O., Silva, D.,
Duran-Gonzalez, J., Provenzano, D. and Pukatzki, S. (2012) 'Constitutive Type VI Secretion System
Expression Gives Vibrio cholerae Intra- and Interspecific Competitive Advantages: e48320', PLoS
One, 7(10).
Unterweger, D., Miyata, S.T., Bachmann, V., Brooks, T.M., Mullins, T., Kostiuk, B., Provenzano, D. and
Pukatzki, S. (2014b) 'The Vibrio cholerae type VI secretion system employs diverse effector
modules for intraspecific competition', Nature communications, 5, pp. 3549.
Ushach, I. and Zlotnik, A. (2016) 'Biological role of granulocyte macrophage colony‐stimulating factor
(GM‐CSF) and macrophage colony‐stimulating factor (M‐CSF) on cells of the myeloid lineage',
Journal of leukocyte biology, 100(3), pp. 481-489.
van Aartsen, J.J. (2012) Genetic and Functional Characterisation of a Novel Klebsiella pneumoniae
Genomic Island Harbouring an Accessory Chaperone/Usher Fimbrial Operon. University of
Leicester.
van Ulsen, P., ur Rahman, S., Jong, W.S., Daleke-Schermerhorn, M.H. and Luirink, J. (2014) 'Type V
secretion: from biogenesis to biotechnology', Biochimica et Biophysica Acta (BBA)-Molecular Cell
Research, 1843(8), pp. 1592-1611.
Vasconcelos, G.J. and Swartz, R.G. (1976) 'Survival of bacteria in seawater using a diffusion chamber
apparatus in situ', Applied and Environmental Microbiology, 31(6), pp. 913-920.
Vaysse, P., Prat, L., Mangenot, S., Cruveiller, S., Goulas, P. and Grimaud, R. (2009) 'Proteomic analysis
of Marinobacter hydrocarbonoclasticus SP17 biofilm formation at the alkane-water interface reveals
novel proteins and cellular processes involved in hexadecane assimilation', Research in
microbiology, 160(10), pp. 829-837.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
275
Veleba, M., Higgins, P.G., Gonzalez, G., Seifert, H. and Schneiders, T. (2012) 'Characterization of RarA,
a novel AraC family multidrug resistance regulator in Klebsiella pneumoniae', Antimicrobial Agents
and Chemotherapy, 56(8), pp. 4450-4458.
Viale, P., Giannella, M., Lewis, R., Trecarichi, E.M., Petrosillo, N. and Tumbarello, M. (2013) 'Predictors
of mortality in multidrug-resistant Klebsiella pneumoniae bloodstream infections', Expert review of
anti-infective therapy, 11(10), pp. 1053-1063.
Victor, L.Y. (2007) 'Virulence characteristics of Klebsiella and clinical manifestations of K. pneumoniae
bloodstream infections', Emerging Infectious Disease, 13(7), pp. 123-128.
Villalta, F., Zhang, Y., Bibb, K.E., Kappes, J.C. and Lima, M.F. (1998) 'The cysteine-cysteine family of
chemokines RANTES, MIP-1alpha, and MIP-1beta induce trypanocidal activity in human
macrophages via nitric oxide', Infection and immunity, 66(10), pp. 4690-4695.
Vincze, T., Posfai, J. and Roberts, R.J. (2003) 'NEBcutter: A program to cleave DNA with restriction
enzymes', Nucleic acids research, 31(13), pp. 3688.
Vogwill, T. and MacLean, R.C. (2015) 'The genetic basis of the fitness costs of antimicrobial resistance: a
meta‐analysis approach', Evolutionary applications, 8(3), pp. 284-295.
Von Frisch, A. (1882) 'Zur aetiologie des rhinoskleroms', Wien. Med. Wschr. Jahrg, (32), pp. 969-972.
Voth, D.E., Broederdorf, L.J. and Graham, J.G. (2012) 'Bacterial Type IV secretion systems: Versatile
virulence machines', Future Microbiology, 7(2), pp. 241-257.
Voulgari, E., Zarkotou, O., Ranellou, K., Karageorgopoulos, D.E., Vrioni, G., Mamali, V., Themeli-
Digalaki, K. and Tsakris, A. (2013) 'Outbreak of OXA-48 carbapenemase-producing Klebsiella
pneumoniae in Greece involving an ST11 clone', The Journal of antimicrobial chemotherapy, 68(1),
pp. 84-88.
Voulhoux, R., Ball, G., Ize, B., Vasil, M.L., Lazdunski, A., Wu, L.F. and Filloux, A. (2001) 'Involvement
of the twin-arginine translocation system in protein secretion via the type II pathway', The EMBO
journal, 20(23), pp. 6735-6741.
Wagner, J.K., Heindl, J.E., Gray, A.N., Jain, S. and Goldberg, M.B. (2009) 'Contribution of the
periplasmic chaperone Skp to efficient presentation of the autotransporter IcsA on the surface of
Shigella flexneri', Journal of Bacteriology, 191(3), pp. 815-821.
Waksman, G. and Hultgren, S.J. (2009) 'Structural biology of the chaperone–usher pathway of pilus
biogenesis', Nature Reviews Microbiology, 7(11), pp. 765-774.
Wallden, K., Rivera-Calzada, A. and Waksman, G. (2010) 'Type IV secretion systems: versatility and
diversity in function', Cellular microbiology, 12(9), pp. 1203-1212.
Wan, B., Zhang, Q., Ni, J., Li, S., Wen, D., Li, J., Xiao, H., He, P., Ou, H. and Tao, J. (2017) 'Type VI
secretion system contributes to Enterohemorrhagic Escherichia coli virulence by secreting catalase
against host reactive oxygen species (ROS)', PLoS pathogens, 13(3), pp. e1006246.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
276
Wand, M.E., McCowen, J.W., Nugent, P.G. and Sutton, J.M. (2013a) 'Complex interactions of Klebsiella
pneumoniae with the host immune system in a Galleria mellonella infection model', Journal of
medical microbiology, 62(12), pp. 1790-1798.
Wand, M., McCowen, J., Nugent, P. and Sutton, J. (2013b) 'Use of the Galleria mellonella infection
model to study the opportunistic pathogen Klebsiella pneumoniae supports the presence of a
multifaceted interaction with the host immune response', J Med Microbiol, 62(Pt 12), pp. 1790-1798.
Wang, R.F. and Kushner, S.R. (1991) 'Construction of versatile low-copy-number vectors for cloning,
sequencing and gene expression in Escherichia coli', Gene, 100, pp. 195-199.
Wang, X., Wang, Q., Xiao, J., Liu, Q., Wu, H., Xu, L. and Zhang, Y. (2009) 'Edwardsiella tarda T6SS
component evpP is regulated by esrB and iron, and plays essential roles in the invasion of fish', Fish
& shellfish immunology, 27(3), pp. 469-477.
Wang, Z., Huang, C., Huang, Y., Wu, C. and Peng, H. (2013) 'FimK regulation on the expression of type
1 fimbriae in Klebsiella pneumoniae CG43S3', Microbiology, 159(7), pp. 1402-1415.
Wang, J.H., Liu, Y.C., Lee, S.S., Yen, M.Y., Chen, Y.S., Wang, J.H., Wann, S.R. and Lin, H.H. (1998)
'Primary liver abscess due to Klebsiella pneumoniae in Taiwan', Clinical infectious diseases : an
official publication of the Infectious Diseases Society of America, 26(6), pp. 1434-1438.
Weber, B., Hasic, M., Chen, C., Wai, S.N. and Milton, D.L. (2009) 'Type VI secretion modulates quorum
sensing and stress response in Vibrio anguillarum', Environmental microbiology, 11(12), pp. 3018-
3028.
Weinberg, E.D. (1978) 'Iron and infection', Microbiological reviews, 42(1), pp. 45-66.
Weller, T., MacKenzie, F. and Forbes, K. (1997) 'Molecular epidemiology of a large outbreak of
multiresistant Klebsiella pneumoniae', Journal of medical microbiology, 46(11), pp. 921-926.
Weyrich, L.S., Rolin, O.Y., Muse, S.J., Park, J., Spidale, N., Kennett, M.J., Hester, S.E., Chen, C.,
Dudley, E.G. and Harvill, E.T. (2012) 'A Type VI Secretion System Encoding Locus Is Required for
Bordetella bronchiseptica Immunomodulation and Persistence In Vivo', PLoS ONE, 7(10).
Wheeler, D.L., Church, D.M., Federhen, S., Lash, A.E., Madden, T.L., Pontius, J.U., Schuler, G.D.,
Schriml, L.M., Sequeira, E., Tatusova, T.A. and Wagner, L. (2003) 'Database resources of the
National Center for Biotechnology', Nucleic acids research, 31(1), pp. 28.
Whitfield, C. (2006) 'Biosynthesis and assembly of capsular polysaccharides in Escherichia coli',
Annu.Rev.Biochem., 75, pp. 39-68.
Whitfield, C., Richards, J.C., Perry, M.B., Clarke, B.R. and MacLean, L.L. (1991) 'Expression of two
structurally distinct D-galactan O antigens in the lipopolysaccharide of Klebsiella pneumoniae
serotype O1', Journal of Bacteriology, 173(4), pp. 1420-1431.
Williams, P., Chart, H., Griffiths, E. and Stevenson, P. (1987) 'Expression of high affinity iron uptake
systems by clinical isolates of Klebsiella', FEMS microbiology letters, 44(3), pp. 407-412.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
277
Williams, P., Lambert, P.A., Brown, M.R. and Jones, R.J. (1983) 'The role of the O and K antigens in
determining the resistance of Klebsiella aerogenes to serum killing and phagocytosis', Microbiology,
129(7), pp. 2181-2191.
Witkowska, A.M. and Borawska, M.H. (2004) 'Soluble intercellular adhesion molecule‐1 (sICAM‐1): an
overview', European cytokine network, 15(2), pp. 91-98.
Wloch-Salamon, D.M., Gerla, D., Hoekstra, R.F. and J. Arjan G.M de Visser (2008) 'Effect of dispersal
and nutrient availability on the competitive ability of toxin-producing yeast', Proceedings of the
Royal Society B: Biological Sciences, 275(1634), pp. 535-541.
Wolpe, S.D., Davatelis, G., Sherry, B., Beutler, B., Hesse, D.G., Nguyen, H.T., Moldawer, L.L., Nathan,
C.F., Lowry, S.F. and Cerami, A. (1988) 'Macrophages secrete a novel heparin-binding protein with
inflammatory and neutrophil chemokinetic properties', The Journal of experimental medicine,
167(2), pp. 570-581.
Wood, L., Klucas, R. and Shearman, R. (1981) 'Nitrogen fixation (acetylene reduction) by Klebsiella
pneumoniae in association with'Park'Kentucky bluegrass (Poa pratensis L.)', Canadian journal of
microbiology, 27(1), pp. 52-56.
Wösten, M.M.S.M., Bleumink-Pluym, N.M.C., van Alphen, L.B., Bouwman, L.I. and van Putten, J.P.M.
(2013) 'Identification of a functional type VI secretion system in Campylobacter jejuni conferring
capsule polysaccharide sensitive cytotoxicity', PLoS Pathogens, 9(5), pp. e1003393.
Wu, C., Huang, Y., Fung, C. and Peng, H. (2010) 'Regulation of the Klebsiella pneumoniae Kpc fimbriae
by the site-specific recombinase KpcI', Microbiology, 156(7), pp. 1983-1992.
Wu, J.H., Wu, A.M., Tsai, C.G., Chang, X.Y., Tsai, S.F. and Wu, T.S. (2008) 'Contribution of fucose-
containing capsules in Klebsiella pneumoniae to bacterial virulence in mice', Experimental biology
and medicine (Maywood, N.J.), 233(1), pp. 64-70.
Wu, K.H. and Tai, P.C. (2004) 'Cys32 and His105 are the critical residues for the calcium-dependent
cysteine proteolytic activity of CvaB, an ATP-binding cassette transporter', The Journal of
biological chemistry, 279(2), pp. 901-909.
Wu, K.M., Li, L.H., Yan, J.J., Tsao, N., Liao, T.L., Tsai, H.C., Fung, C.P., Chen, H.J., Liu, Y.M., Wang,
J.T., Fang, C.T., Chang, S.C., Shu, H.Y., Liu, T.T., Chen, Y.T., Shiau, Y.R., Lauderdale, T.L., Su,
I.J., Kirby, R. and Tsai, S.F. (2009) 'Genome sequencing and comparative analysis of Klebsiella
pneumoniae NTUH-K2044, a strain causing liver abscess and meningitis', Journal of Bacteriology,
191(14), pp. 4492-4501.
Wynn, T.A., Chawla, A. and Pollard, J.W. (2013) 'Macrophage biology in development, homeostasis and
disease', Nature, 496(7446), pp. 445.
Xu, L.L., Warren, M.K., Rose, W.L., Gong, W. and Wang, J.M. (1996) 'Human recombinant monocyte
chemotactic protein and other C‐C chemokines bind and induce directional migration of dendritic
cells in vitro', Journal of leukocyte biology, 60(3), pp. 365-371.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
278
Yang, F.L., Yang, Y.L., Liao, P.C., Chou, J.C., Tsai, K.C., Yang, A.S., Sheu, F., Lin, T.L., Hsieh, P.F.,
Wang, J.T., Hua, K.F. and Wu, S.H. (2011) 'Structure and immunological characterization of the
capsular polysaccharide of a pyrogenic liver abscess caused by Klebsiella pneumoniae: activation of
macrophages through Toll-like receptor 4', The Journal of biological chemistry, 286(24), pp. 21041-
21051.
Yeh, K.M., Lin, J.C., Yin, F.Y., Fung, C.P., Hung, H.C., Siu, L.K. and Chang, F.Y. (2010) 'Revisiting the
importance of virulence determinant magA and its surrounding genes in Klebsiella pneumoniae
causing pyogenic liver abscesses: exact role in serotype K1 capsule formation', The Journal of
infectious diseases, 201(8), pp. 1259-1267.
Yeung, A.T., Bains, M. and Hancock, R.E. (2011) 'The sensor kinase CbrA is a global regulator that
modulates metabolism, virulence, and antibiotic resistance in Pseudomonas aeruginosa', Journal of
Bacteriology, 193(4), pp. 918-931.
Yousuf, F.A., Siddiqui, R. and Khan, N.A. (2013) 'Acanthamoeba castellanii of the T4 genotype is a
potential environmental host for Enterobacter aerogenes and Aeromonas hydrophila', Parasites &
vectors, 6(1), pp. 169-169.
Yu, Y., Yang, H., Li, J., Zhang, P., Wu, B., Zhu, B., Zhang, Y. and Fang, W. (2012) 'Putative type VI
secretion systems of Vibrio parahaemolyticus contribute to adhesion to cultured cell monolayers',
Archives of Microbiology, 194(10), pp. 827-835.
Yu, W.L., Chuang, Y.C. and Walther-Rasmussen, J. (2006) 'Extended-spectrum beta-lactamases in
Taiwan: epidemiology, detection, treatment and infection control', Journal of microbiology,
immunology, and infection = Wei mian yu gan ran za zhi, 39(4), pp. 264-277.
Zarember, K.A. and Kuhns, D.B. (2011) 'Will the real neutrophil please stand up?', Journal of leukocyte
biology, 90(6), pp. 1039-1041.
Zelová, H. and Hošek, J. (2013) 'TNF-α signalling and inflammation: interactions between old
acquaintances', Inflammation Research, 62(7), pp. 641-651.
Zhang, W., Wang, Y., Song, Y., Wang, T., Xu, S., Peng, Z., Lin, X., Zhang, L. and Shen, X. (2012) 'A
type VI secretion system regulated by OmpR in Yersinia pseudotuberculosis functions to maintain
intracellular pH homeostasis', Environmental microbiology, .
Zheng, J. and Leung, K.Y. (2007a) 'Dissection of a type VI secretion system in Edwardsiella tarda',
Molecular microbiology, 66(5), pp. 1192-1206.
Zheng, J., Ho, B. and Mekalanos, J.J. (2011a) 'Genetic analysis of anti-amoebae and anti-bacterial
activities of the type vi secretion system in vibrio cholerae', PLoS ONE, 6(8).
Zheng, J., Ho, B. and Mekalanos, J.J. (2011b) 'Genetic analysis of anti-amoebae and anti-bacterial
activities of the type vi secretion system in vibrio cholerae', PLoS ONE, 6(8).
Zheng, J. and Leung, K.Y. (2007b) 'Dissection of a type VI secretion system in Edwardsiella tarda',
Molecular microbiology, 66(5), pp. 1192-1206.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
279
Zheng, J., Shin, O.S., Cameron, D.E. and Mekalanos, J.J. (2010) 'Quorum sensing and a global regulator
TsrA control expression of type VI secretion and virulence in Vibrio cholerae', Proceedings of the
National Academy of Sciences of the United States of America, 107(49), pp. 21128-21133.
Zhou, Y., Tao, J., Yu, H., Ni, J., Zeng, L., Teng, Q., Kim, K.S., Zhao, G.-., Guo, X. and Yao, Y. (2012)
'Hcp family proteins secreted via the type VI secretion system coordinately regulate Escherichia coli
K1 interaction with human brain microvascular endothelial cells', Infection and immunity, 80(3), pp.
1243-1251.
Zoued, A., Durand, E., Brunet, Y.R., Spinelli, S., Douzi, B., Guzzo, M., Flaugnatti, N., Legrand, P.,
Journet, L. and Fronzes, R. (2016) 'Priming and polymerization of a bacterial contractile tail
structure', Nature, 531(7592), pp. 59-63.
Zoued, A., Durand, E., Bebeacua, C., Brunet, Y.R., Douzi, B., Cambillau, C., Cascales, E. and Journet, L.
(2013) 'TssK is a trimeric cytoplasmic protein interacting with components of both phage-like and
membrane anchoring complexes of the type VI secretion system', The Journal of biological
chemistry, 288(38), pp. 27031-27041.
Zoued, A., Brunet, Y.R., Durand, E., Aschtgen, M., Logger, L., Douzi, B., Journet, L., Cambillau, C. and
Cascales, E. (2014a) 'Architecture and assembly of the Type VI secretion system', Biochimica et
Biophysica Acta (BBA) - Molecular Cell Research, 1843(8), pp. 1664-1673.
Zoued, A., Brunet, Y.R., Durand, E., Aschtgen, M., Logger, L., Douzi, B., Journet, L., Cambillau, C. and
Cascales, E. (2014b) 'Architecture and assembly of the Type VI secretion system', Biochimica et
Biophysica Acta (BBA) - Molecular Cell Research, 1843(8), pp. 1664-1673.
Zoued, A., Brunet, Y.R., Durand, E., Aschtgen, M., Logger, L., Douzi, B., Journet, L., Cambillau, C. and
Cascales, E. (2014c) 'Architecture and assembly of the Type VI secretion system', Biochimica et
Biophysica Acta (BBA) - Molecular Cell Research, 1843(8), pp. 1664-1673.

David Mwin Ph.D. Thesis, University of Leicester, UK. References
280

David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
281
Appendices
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
282
Appendix 1: Primer Table
Primer Sequence (5’→3’) Feature Template Purpose
GmF CGAATTAGCTTCAAAAGCGCTCTGA - pUC18R6K-mini-tn7-Gm Amplification of
antibiotic cassettes GmR AATTGGGGATCTTGAAGTTCCT - pUC18R6K-mini-tn7-Gm
DN1 ATACTCGAGGCCGACCAGCATTAGGATT XhoI site K. pneumoniae NTUH K2044
T6SS1 mutant allele
(2595 bp) in pDNTOOL-8
DN2 agagcgcttttgaagctaattcgTGGGGACTTCATTCTGGA with overlap K. pneumoniae NTUH K2044
DN3 aacttcaagatccccaattAGTCTGAATATGCCGCTGG with overlap K. pneumoniae NTUH K2044
DN4 ATAACTAGTTCGCCTGTCAGGGATCTT SpeI site K. pneumoniae NTUH K2044
DN5 CTAGCGCATCGTAGTTCTCG - Kp. NTUH-K2044 ∆T1 T6SS1 mutation
verification primers DN6 GTCGGTGGTGTTTAGCTATTCC - Kp. NTUH-K2044 ∆T1
DN7 TTATACTAGTGTTCGCTGCTTTGAAAACG SpeI site K. pneumoniae NTUH K2044
T6SS3 mutant allele in
pDNTOOL-9
DN8 aggAacttcaagatccccaattATATTGTGGAGTAGCGTGTC with overlap K. pneumoniae NTUH K2044
DN9 tcagagcgcttttgaagctaattcgAGATACCCGGAGGAGTGC with overlap K. pneumoniae NTUH K2044
DN10 ATGTCTCGAGACCGTAATGCGGTTCATCTC XhoI site K. pneumoniae NTUH K2044
DN11 TCCAAAGGTGAAGGTCTTGG - Kp. NTUH-K2044 ∆T3 T6SS3 mutant verification
primers DN12 TCACCTTCGGGCTTGTTATC - Kp. NTUH-K2044 ∆T3
DN13 TGAAATTGAACGCGCGAACG - pJTOOL-3 LF of pJTOOL-3
DN14 TTTAGCTTCCTTAGCTCCTGCC - pJTOOL-3
DN15 TTCCCGGGTCATGGCTGCG - pJTOOL-3 RF of pJTOOL-3
DN16 ACATCAAGGCCAAGCCCGCC - pJTOOL-3
DN17 ccatagggcaggagctaaggaagctaaaATGGTGCAATACGAATGGCGAAAAG with overlap pKOBEG-Apra aac(3)-IV for PNDTOOL-6
DN18 cgggtgtcggggcgcagccatgacccgggaaTCATGAGCTCAGCCAATCG with overlap pKOBEG-Apra
DN19 ccatagggcaggagctaaggaagctaaaATGACACAAGAATCCCTGTT with overlap pMQ300 hph for PNDTOOL-7
DN20 cgggtgtcggggcgcagccatgacccgggaaTCAGGCGCCGGGGGCGGT with overlap pMQ300
DN21 tctGGGCCCATGAACATCAAAAAGTTTGCAAAAC ApaI site pJTOOL-3 sacB-cat for pDNTOOL-13
and pDNTOOL-16 DN22 ctcTTAATTAATTACGCCCCGCCCTGCCAC PacI site pJTOOL-3
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
283
DN23 tctGGGCCCACTCTTCCTTTTTCAATATTATTG ApaI site pKD46 For pDNTOOL-13
DN24 ctcTTAATTAACTGTCAGACCAAGTTTACTCATATATAC PacI site pKD46
DN25 tctGGGCCCGACATTGCACTCCACCGCTG ApaI site pKOBEG-Apra For pDNTOOL-16
DN26 ctcTTAATTAAGCGGAGAACGAGATGACGTTGGAG PacI site pKOBEG-Apra
DN27 aaatgcttcaataatattgaaaaaggaagagtATGGTGCAATACGAATGG with overlap pKOBEG-Apra aac(3)-IV for PNDTOOL-
10 DN28 taaagtatatatgagtaaacttggtctgacagTCATGAGCTCAGCCAATC with overlap pKOBEG-Apra
DN29 aaatgcttcaataatattgaaaaaggaagagtATGTTACGCAGCAGCAAC with overlap pUC18R6K-mini-tn7-Gm aacC1 for PNDTOOL-11
DN30 taaagtatatatgagtaaacttggtctgacagTTAGGTGGCGGTACTTGG with overlap pUC18R6K-mini-tn7-Gm
DN31 aaatgcttcaataatattgaaaaaggaagagtATGATTGAACAAGATGG with overlap pJTOOL-8 aph for pNDTOOL-12
DN32 taaagtatatatgagtaaacttggtctgacagTCAGAAGAACTCGTCAAG with overlap pJTOOL-8
DN33 ctgatgtcatcagcggtggagtgcaatgtcATGACACAAGAATCCCTG with overlap pMQ300 hph for for pNDTOOL-14
DN34 ttgcccctccaacgtcatctcgttctccgcTCAGGCGCCGGGGGCGGT with overlap pMQ300
DN35 gactgatgtcatcagcggtggagtgcaatgtcATGATTGAACAAGATGGA with overlap pJTOOL-8 aph for for pNDTOOL-15
DN36 ccttgcccctccaacgtcatctcgttctccgcTCAGAAGAACTCGTCAAG with overlap pJTOOL-8
DN37 gactgatgtcatcagcggtggagtgcaatgtcATGTTACGCAGCAGCAAC with overlap pUC18R6K-mini-tn7-Gm aacC1 for pNDTOOL-17
DN38 ccttgcccctccaacgtcatctcgttctccgcTTAGGTGGCGGTACTTGG with overlap pUC18R6K-mini-tn7-Gm
DN39 gtttgatgttatggagcagcaacgATGGAGAAAAAAATCACTGGATATAC with overlap pJTOOL-3 cat for pNDTOOL-32
DN40 ccgatctcggcttgaacgaattgTTACGCCCCGCCCTGCCACT with overlap pJTOOL-3
DN41 acgACGCGTatgaacatcaaaaagtttgc MluIsite pJTOOL-3 sacB-cat for pNDTOOL-31
DN42 ttgTTAATTAAttacgccccgccctgccact PacI site pJTOOL-3
DN43 catACGCGTcgttgctgctccataacatcaaac MluI site pUC18R6K-mini-tn7-Gm pUC18R6K-mini-tn7-Gm
for pNDTOOL-31 DN44 ccgTTAATTAAcaattcgttcaagccgagatcg PacI site pUC18R6K-mini-tn7-Gm
DN45 GCGCCCAAACATAACAGGAAG - pUC18R6K-mini-tn7-Gm LF of pUC18R6K-mini-
tn7-Gm DN46 CGTTGCTGCTCCATAACATCAAAC - pUC18R6K-mini-tn7-Gm
DN47 CAATTCGTTCAAGCCGAGATCGG - pUC18R6K-mini-tn7-Gm RF of pUC18R6K-mini-
tn7R-Gm DN48 GCAAACTCTTCATTTTCTGAAGTGC - pUC18R6K-mini-tn7-Gm
DN49 gtttgatgttatggagcagcaacgATGACACAAGAATCCCTGTTAC with overlap pMQ300
DN50 ccgatctcggcttgaacgaattgTCAGGCGCCGGGGGCGGTGT with overlap pMQ300
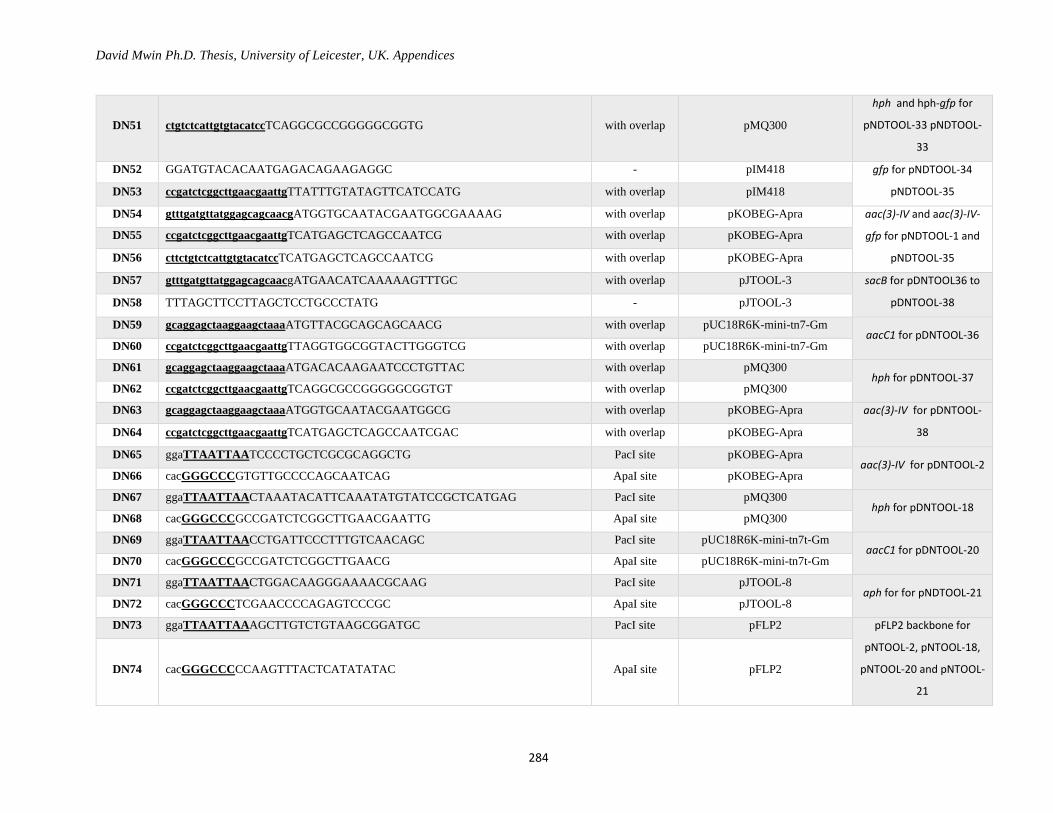
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
284
DN51 ctgtctcattgtgtacatccTCAGGCGCCGGGGGCGGTG with overlap pMQ300
hph and hph-gfp for
pNDTOOL-33 pNDTOOL-
33
DN52 GGATGTACACAATGAGACAGAAGAGGC - pIM418 gfp for pNDTOOL-34
pNDTOOL-35 DN53 ccgatctcggcttgaacgaattgTTATTTGTATAGTTCATCCATG with overlap pIM418
DN54 gtttgatgttatggagcagcaacgATGGTGCAATACGAATGGCGAAAAG with overlap pKOBEG-Apra aac(3)-IV and aac(3)-IV-
gfp for pNDTOOL-1 and
pNDTOOL-35
DN55 ccgatctcggcttgaacgaattgTCATGAGCTCAGCCAATCG with overlap pKOBEG-Apra
DN56 cttctgtctcattgtgtacatccTCATGAGCTCAGCCAATCG with overlap pKOBEG-Apra
DN57 gtttgatgttatggagcagcaacgATGAACATCAAAAAGTTTGC with overlap pJTOOL-3 sacB for pDNTOOL36 to
pDNTOOL-38 DN58 TTTAGCTTCCTTAGCTCCTGCCCTATG - pJTOOL-3
DN59 gcaggagctaaggaagctaaaATGTTACGCAGCAGCAACG with overlap pUC18R6K-mini-tn7-Gm aacC1 for pDNTOOL-36
DN60 ccgatctcggcttgaacgaattgTTAGGTGGCGGTACTTGGGTCG with overlap pUC18R6K-mini-tn7-Gm
DN61 gcaggagctaaggaagctaaaATGACACAAGAATCCCTGTTAC with overlap pMQ300 hph for pDNTOOL-37
DN62 ccgatctcggcttgaacgaattgTCAGGCGCCGGGGGCGGTGT with overlap pMQ300
DN63 gcaggagctaaggaagctaaaATGGTGCAATACGAATGGCG with overlap pKOBEG-Apra aac(3)-IV for pDNTOOL-
38 DN64 ccgatctcggcttgaacgaattgTCATGAGCTCAGCCAATCGAC with overlap pKOBEG-Apra
DN65 ggaTTAATTAATCCCCTGCTCGCGCAGGCTG PacI site pKOBEG-Apra aac(3)-IV for pDNTOOL-2
DN66 cacGGGCCCGTGTTGCCCCAGCAATCAG ApaI site pKOBEG-Apra
DN67 ggaTTAATTAACTAAATACATTCAAATATGTATCCGCTCATGAG PacI site pMQ300 hph for pDNTOOL-18
DN68 cacGGGCCCGCCGATCTCGGCTTGAACGAATTG ApaI site pMQ300
DN69 ggaTTAATTAACCTGATTCCCTTTGTCAACAGC PacI site pUC18R6K-mini-tn7t-Gm aacC1 for pDNTOOL-20
DN70 cacGGGCCCGCCGATCTCGGCTTGAACG ApaI site pUC18R6K-mini-tn7t-Gm
DN71 ggaTTAATTAACTGGACAAGGGAAAACGCAAG PacI site pJTOOL-8 aph for for pNDTOOL-21
DN72 cacGGGCCCTCGAACCCCAGAGTCCCGC ApaI site pJTOOL-8
DN73 ggaTTAATTAAAGCTTGTCTGTAAGCGGATGC PacI site pFLP2 pFLP2 backbone for
pNTOOL-2, pNTOOL-18,
pNTOOL-20 and pNTOOL-
21
DN74 cacGGGCCCCCAAGTTTACTCATATATAC ApaI site pFLP2
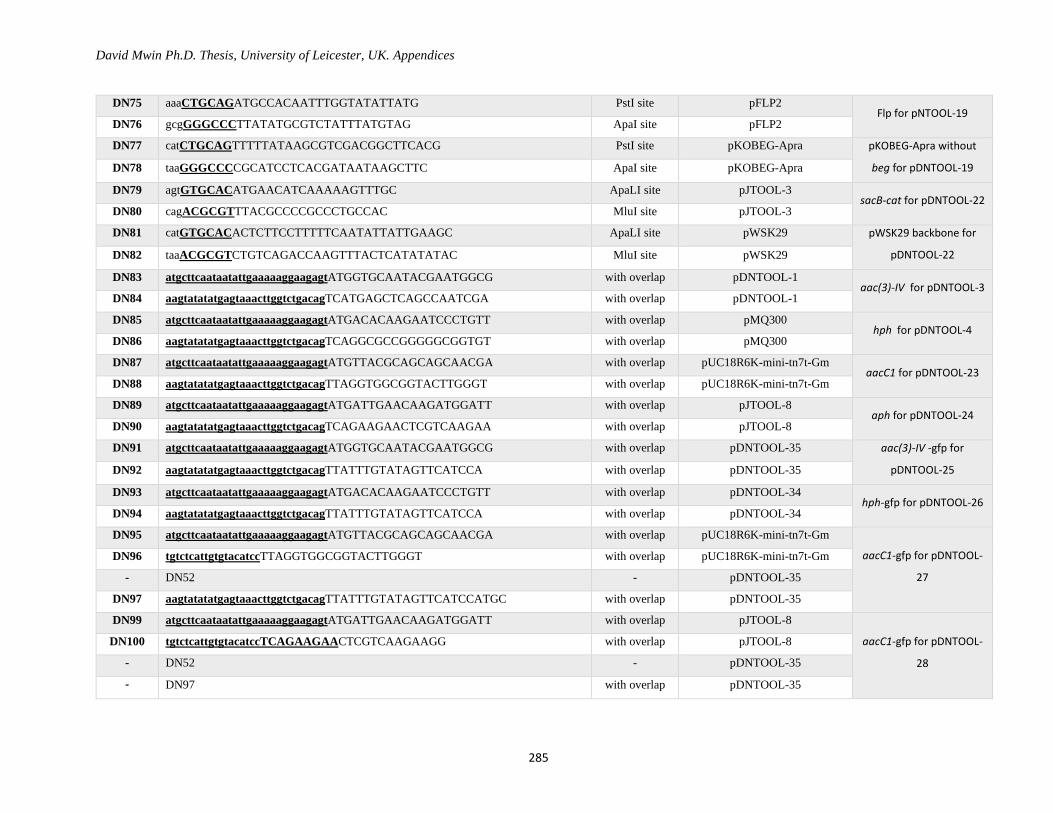
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
285
DN75 aaaCTGCAGATGCCACAATTTGGTATATTATG PstI site pFLP2 Flp for pNTOOL-19
DN76 gcgGGGCCCTTATATGCGTCTATTTATGTAG ApaI site pFLP2
DN77 catCTGCAGTTTTTATAAGCGTCGACGGCTTCACG PstI site pKOBEG-Apra pKOBEG-Apra without
beg for pDNTOOL-19 DN78 taaGGGCCCCGCATCCTCACGATAATAAGCTTC ApaI site pKOBEG-Apra
DN79 agtGTGCACATGAACATCAAAAAGTTTGC ApaLI site pJTOOL-3 sacB-cat for pDNTOOL-22
DN80 cagACGCGTTTACGCCCCGCCCTGCCAC MluI site pJTOOL-3
DN81 catGTGCACACTCTTCCTTTTTCAATATTATTGAAGC ApaLI site pWSK29 pWSK29 backbone for
pDNTOOL-22 DN82 taaACGCGTCTGTCAGACCAAGTTTACTCATATATAC MluI site pWSK29
DN83 atgcttcaataatattgaaaaaggaagagtATGGTGCAATACGAATGGCG with overlap pDNTOOL-1 aac(3)-IV for pDNTOOL-3
DN84 aagtatatatgagtaaacttggtctgacagTCATGAGCTCAGCCAATCGA with overlap pDNTOOL-1
DN85 atgcttcaataatattgaaaaaggaagagtATGACACAAGAATCCCTGTT with overlap pMQ300 hph for pDNTOOL-4
DN86 aagtatatatgagtaaacttggtctgacagTCAGGCGCCGGGGGCGGTGT with overlap pMQ300
DN87 atgcttcaataatattgaaaaaggaagagtATGTTACGCAGCAGCAACGA with overlap pUC18R6K-mini-tn7t-Gm aacC1 for pDNTOOL-23
DN88 aagtatatatgagtaaacttggtctgacagTTAGGTGGCGGTACTTGGGT with overlap pUC18R6K-mini-tn7t-Gm
DN89 atgcttcaataatattgaaaaaggaagagtATGATTGAACAAGATGGATT with overlap pJTOOL-8 aph for pDNTOOL-24
DN90 aagtatatatgagtaaacttggtctgacagTCAGAAGAACTCGTCAAGAA with overlap pJTOOL-8
DN91 atgcttcaataatattgaaaaaggaagagtATGGTGCAATACGAATGGCG with overlap pDNTOOL-35 aac(3)-IV -gfp for
pDNTOOL-25 DN92 aagtatatatgagtaaacttggtctgacagTTATTTGTATAGTTCATCCA with overlap pDNTOOL-35
DN93 atgcttcaataatattgaaaaaggaagagtATGACACAAGAATCCCTGTT with overlap pDNTOOL-34 hph-gfp for pDNTOOL-26
DN94 aagtatatatgagtaaacttggtctgacagTTATTTGTATAGTTCATCCA with overlap pDNTOOL-34
DN95 atgcttcaataatattgaaaaaggaagagtATGTTACGCAGCAGCAACGA with overlap pUC18R6K-mini-tn7t-Gm
aacC1-gfp for pDNTOOL-
27
DN96 tgtctcattgtgtacatccTTAGGTGGCGGTACTTGGGT with overlap pUC18R6K-mini-tn7t-Gm
- DN52 - pDNTOOL-35
DN97 aagtatatatgagtaaacttggtctgacagTTATTTGTATAGTTCATCCATGC with overlap pDNTOOL-35
DN99 atgcttcaataatattgaaaaaggaagagtATGATTGAACAAGATGGATT with overlap pJTOOL-8
aacC1-gfp for pDNTOOL-
28
DN100 tgtctcattgtgtacatccTCAGAAGAACTCGTCAAGAAGG with overlap pJTOOL-8
- DN52 - pDNTOOL-35
- DN97 with overlap pDNTOOL-35

David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
286
DN101 cctGCGGCCGCATTTTTTTGTTGCCAACGCACCG NotI site pKPHS6 pKPHS6 backbone for
pDNTOOL-29 DN102 gccgtcaagttgtcataataaatcgCGGAACCCCTGAAGGGGCCC with overlap pKPHS6
DN103 gggccccttcaggggttccgCGATTTATTATGACAACTTGACGGC with overlap pKD46 Arabinose inducible
promoter for pDNTOOL-
29 DN104 cggGGTACCGTGCACCTGCAGAAGCTTTTTTTATAACCTCCTTAGAGCTC with overlap pKD46
DN105 attGGTACCCCGCGGCCCGGGTCTAGAACTAGTCCATGGGCATGCTT with overlap Multiple cloning site (MCS) MCS
DN106 AGAACTAGTCCATGGGCATGCTTAATTAACGCATCCTCACGATAATATCC with overlap pKD46 Terminator sequence
from pKD46 DN107 gttcaagccgagatcggctATTCTTCGTCTGTTTCTACTGGTATTG with overlap pKD46
DN108 caataccagtagaaacagacgaagaatAGCCGATCTCGGCTTGAACG with overlap pUC18R6K-mini-tn7t-Gm aacC1 for pDNTOOL-29
DN109 aatGCGGCCGCAAGATCCCCTGATTCCCTTTG NotI site pUC18R6K-mini-tn7t-Gm
DN110 tggGCATGCGCAGAAGCAGCTGCAAAGGAGGCTGCTGCAAAGGCA SphI site - MCS-polylinker sequence
DN111 gcaaaggaggctgctgcaaaggcaATGCGTAAAGGAGAAGAACTTTTC with overlap pMQ300 gfp for pDNTOOL-230
DN112 gcgTTAATTAATTATTTGTATAGTTCATCCATGCC PacI site pMQ300
DN113 GAGATGATTACTAGTCAAATGGATTG - K. pneumoniae HS11286
3166 bp KPHST6SS1
mutant allele
DN114 tcagagcgcttttgaagctaattcgCATAGCAGTTCCTTTCCTTAAGG with overlap K. pneumoniae HS11286
DN115 aggAacttcaagatccccaattTGGAATGGGAAGAGAAGCACAGC with overlap K. pneumoniae HS11286
DN116 AGCAGGATGAGAAAGTCCC - K. pneumoniae HS11286
DN117 CCGCGAATGATTATATAGACCGATC - K. pneumoniae HS11286 ∆T1 For KPHS∆T1
verification (3588 bp) DN118 TTCATACCGAGCCAGAACGG - K. pneumoniae HS11286 ∆T1
DN119 GTGGTTCGCTGCTTTGAAAACG - K. pneumoniae HS11286
3209 bp KPHST6SS1
mutant allele
DN120 aggAacttcaagatccccaattATATTGTGGAGTAGCGTGTCG with overlap K. pneumoniae HS11286
DN121 agagcgcttttgaagctaattcgAGATGCCCGGAGGAGCGCC with overlap K. pneumoniae HS11286
DN122 CCGTAATGCGGTTCATCTCG - K. pneumoniae HS11286
DN123 TCCAAAGGTGAAGGTCTTGGC - K. pneumoniae HS11286 ∆T3 For KPHS∆T1
verification (3417 bp) DN124 TCACCTTCGGGCTTGTTATCC - K. pneumoniae HS11286 ∆T3
DN125 CTAGCGAAAAGATGTTTCGTGAAGC -- pKOBEG-Apra For pKOBEG-Apra
verification (1949 bp) DN126 GCTTATTATCGTGAGGATGCGTCA - pKOBEG-Apra & derivatives
ND127 GGGAATTCGAGCTCTAAGGAGG - pKD46 and derivatives
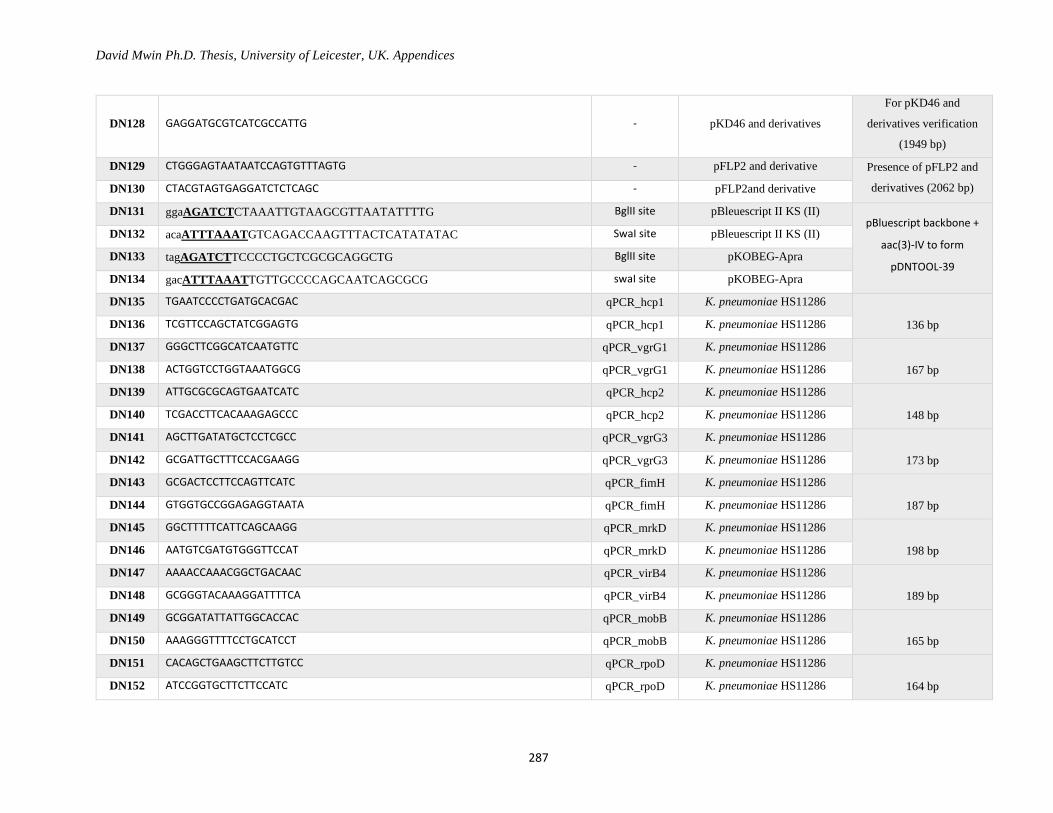
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
287
DN128 GAGGATGCGTCATCGCCATTG - pKD46 and derivatives
For pKD46 and
derivatives verification
(1949 bp)
DN129 CTGGGAGTAATAATCCAGTGTTTAGTG - pFLP2 and derivative Presence of pFLP2 and
derivatives (2062 bp) DN130 CTACGTAGTGAGGATCTCTCAGC - pFLP2and derivative
DN131 ggaAGATCTCTAAATTGTAAGCGTTAATATTTTG BglII site pBleuescript II KS (II) pBluescript backbone +
aac(3)-IV to form
pDNTOOL-39
DN132 acaATTTAAATGTCAGACCAAGTTTACTCATATATAC SwaI site pBleuescript II KS (II)
DN133 tagAGATCTTCCCCTGCTCGCGCAGGCTG BglII site pKOBEG-Apra
DN134 gacATTTAAATTGTTGCCCCAGCAATCAGCGCG swaI site pKOBEG-Apra
DN135 TGAATCCCCTGATGCACGAC qPCR_hcp1 K. pneumoniae HS11286
136 bp DN136 TCGTTCCAGCTATCGGAGTG qPCR_hcp1 K. pneumoniae HS11286
DN137 GGGCTTCGGCATCAATGTTC qPCR_vgrG1 K. pneumoniae HS11286
167 bp DN138 ACTGGTCCTGGTAAATGGCG qPCR_vgrG1 K. pneumoniae HS11286
DN139 ATTGCGCGCAGTGAATCATC qPCR_hcp2 K. pneumoniae HS11286
148 bp DN140 TCGACCTTCACAAAGAGCCC qPCR_hcp2 K. pneumoniae HS11286
DN141 AGCTTGATATGCTCCTCGCC qPCR_vgrG3 K. pneumoniae HS11286
173 bp DN142 GCGATTGCTTTCCACGAAGG qPCR_vgrG3 K. pneumoniae HS11286
DN143 GCGACTCCTTCCAGTTCATC qPCR_fimH K. pneumoniae HS11286
187 bp DN144 GTGGTGCCGGAGAGGTAATA qPCR_fimH K. pneumoniae HS11286
DN145 GGCTTTTTCATTCAGCAAGG qPCR_mrkD K. pneumoniae HS11286
198 bp DN146 AATGTCGATGTGGGTTCCAT qPCR_mrkD K. pneumoniae HS11286
DN147 AAAACCAAACGGCTGACAAC qPCR_virB4 K. pneumoniae HS11286
189 bp DN148 GCGGGTACAAAGGATTTTCA qPCR_virB4 K. pneumoniae HS11286
DN149 GCGGATATTATTGGCACCAC qPCR_mobB K. pneumoniae HS11286
165 bp DN150 AAAGGGTTTTCCTGCATCCT qPCR_mobB K. pneumoniae HS11286
DN151 CACAGCTGAAGCTTCTTGTCC qPCR_rpoD K. pneumoniae HS11286
164 bp DN152 ATCCGGTGCTTCTTCCATC qPCR_rpoD K. pneumoniae HS11286
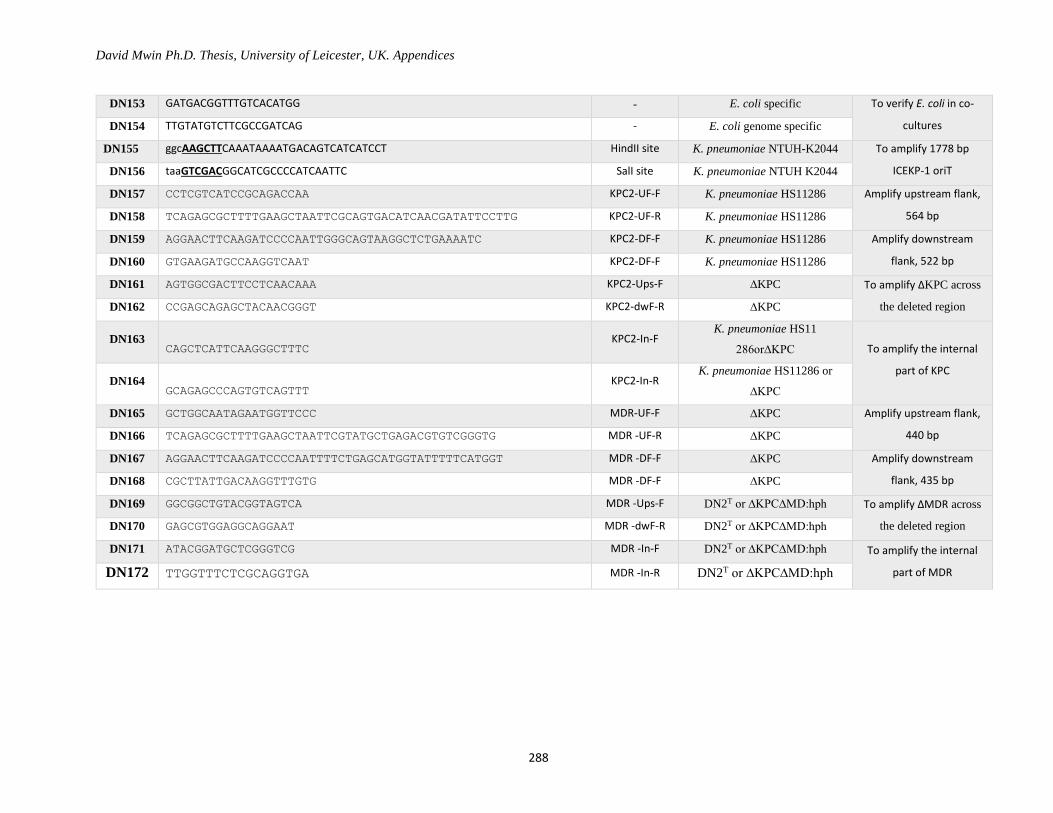
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
288
DN153 GATGACGGTTTGTCACATGG - E. coli specific To verify E. coli in co-
cultures DN154 TTGTATGTCTTCGCCGATCAG - E. coli genome specific
DN155 ggcAAGCTTCAAATAAAATGACAGTCATCATCCT HindII site K. pneumoniae NTUH-K2044 To amplify 1778 bp
ICEKP-1 oriT DN156 taaGTCGACGGCATCGCCCCATCAATTC SalI site K. pneumoniae NTUH K2044
DN157 CCTCGTCATCCGCAGACCAA KPC2-UF-F K. pneumoniae HS11286 Amplify upstream flank,
564 bp DN158 TCAGAGCGCTTTTGAAGCTAATTCGCAGTGACATCAACGATATTCCTTG KPC2-UF-R K. pneumoniae HS11286
DN159 AGGAACTTCAAGATCCCCAATTGGGCAGTAAGGCTCTGAAAATC KPC2-DF-F K. pneumoniae HS11286 Amplify downstream
flank, 522 bp DN160 GTGAAGATGCCAAGGTCAAT KPC2-DF-F K. pneumoniae HS11286
DN161 AGTGGCGACTTCCTCAACAAA KPC2-Ups-F ∆KPC To amplify ∆KPC across
the deleted region DN162 CCGAGCAGAGCTACAACGGGT KPC2-dwF-R ∆KPC
DN163 CAGCTCATTCAAGGGCTTTC
KPC2-In-F K. pneumoniae HS11
286or∆KPC To amplify the internal
part of KPC DN164
GCAGAGCCCAGTGTCAGTTT KPC2-In-R
K. pneumoniae HS11286 or
∆KPC
DN165 GCTGGCAATAGAATGGTTCCC MDR-UF-F ∆KPC Amplify upstream flank,
440 bp DN166 TCAGAGCGCTTTTGAAGCTAATTCGTATGCTGAGACGTGTCGGGTG MDR -UF-R ∆KPC
DN167 AGGAACTTCAAGATCCCCAATTTTCTGAGCATGGTATTTTTCATGGT MDR -DF-F ∆KPC Amplify downstream
flank, 435 bp DN168 CGCTTATTGACAAGGTTTGTG MDR -DF-F ∆KPC
DN169 GGCGGCTGTACGGTAGTCA MDR -Ups-F DN2T or ∆KPC∆MD:hph To amplify ∆MDR across
the deleted region DN170 GAGCGTGGAGGCAGGAAT MDR -dwF-R DN2T or ∆KPC∆MD:hph
DN171 ATACGGATGCTCGGGTCG MDR -In-F DN2T or ∆KPC∆MD:hph To amplify the internal
part of MDR DN172 TTGGTTTCTCGCAGGTGA MDR -In-R DN2T or ∆KPC∆MD:hph
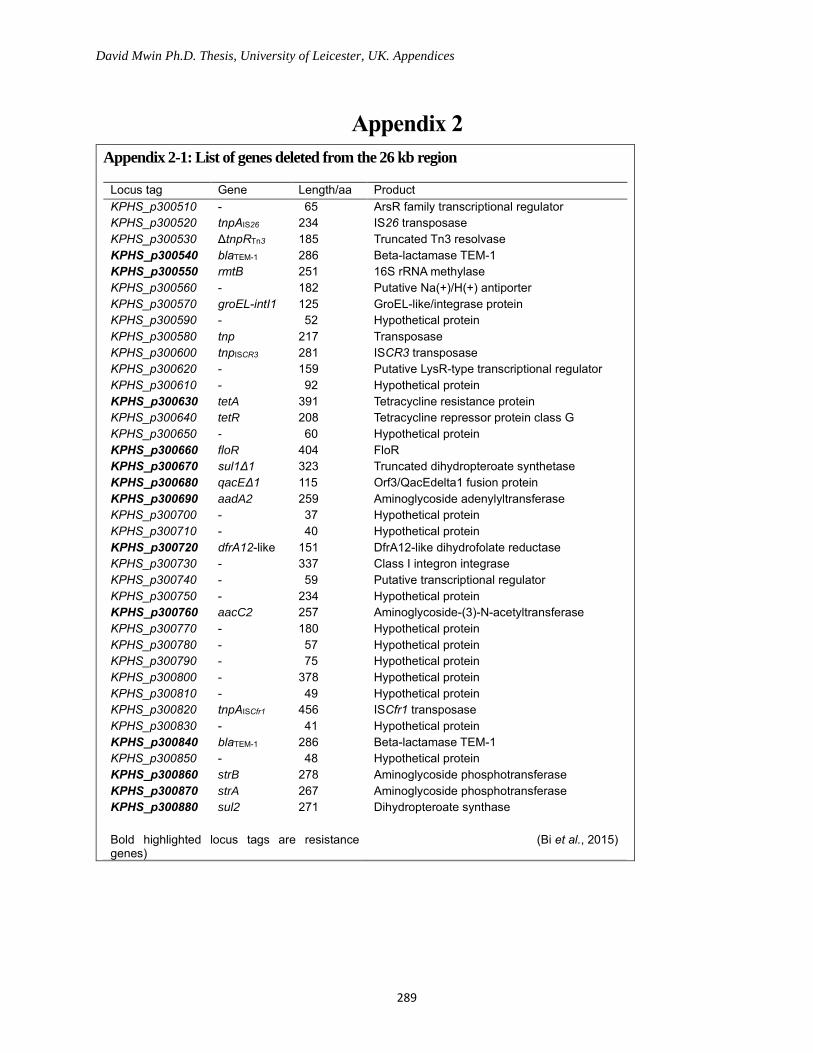
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
289
Appendix 2
Appendix 2-1: List of genes deleted from the 26 kb region
Locus tag Gene Length/aa Product
KPHS_p300510 - 65 ArsR family transcriptional regulator
KPHS_p300520 tnpAIS26 234 IS26 transposase
KPHS_p300530 ΔtnpRTn3 185 Truncated Tn3 resolvase
KPHS_p300540 blaTEM-1 286 Beta-lactamase TEM-1
KPHS_p300550 rmtB 251 16S rRNA methylase
KPHS_p300560 - 182 Putative Na(+)/H(+) antiporter
KPHS_p300570 groEL-intI1 125 GroEL-like/integrase protein
KPHS_p300590 - 52 Hypothetical protein
KPHS_p300580 tnp 217 Transposase
KPHS_p300600 tnpISCR3 281 ISCR3 transposase
KPHS_p300620 - 159 Putative LysR-type transcriptional regulator
KPHS_p300610 - 92 Hypothetical protein
KPHS_p300630 tetA 391 Tetracycline resistance protein
KPHS_p300640 tetR 208 Tetracycline repressor protein class G
KPHS_p300650 - 60 Hypothetical protein
KPHS_p300660 floR 404 FloR
KPHS_p300670 sul1Δ1 323 Truncated dihydropteroate synthetase
KPHS_p300680 qacEΔ1 115 Orf3/QacEdelta1 fusion protein
KPHS_p300690 aadA2 259 Aminoglycoside adenylyltransferase
KPHS_p300700 - 37 Hypothetical protein
KPHS_p300710 - 40 Hypothetical protein
KPHS_p300720 dfrA12-like 151 DfrA12-like dihydrofolate reductase
KPHS_p300730 - 337 Class I integron integrase
KPHS_p300740 - 59 Putative transcriptional regulator
KPHS_p300750 - 234 Hypothetical protein
KPHS_p300760 aacC2 257 Aminoglycoside-(3)-N-acetyltransferase
KPHS_p300770 - 180 Hypothetical protein
KPHS_p300780 - 57 Hypothetical protein
KPHS_p300790 - 75 Hypothetical protein
KPHS_p300800 - 378 Hypothetical protein
KPHS_p300810 - 49 Hypothetical protein
KPHS_p300820 tnpAISCfr1 456 ISCfr1 transposase
KPHS_p300830 - 41 Hypothetical protein
KPHS_p300840 blaTEM-1 286 Beta-lactamase TEM-1
KPHS_p300850 - 48 Hypothetical protein
KPHS_p300860 strB 278 Aminoglycoside phosphotransferase
KPHS_p300870 strA 267 Aminoglycoside phosphotransferase
KPHS_p300880 sul2 271 Dihydropteroate synthase
Bold highlighted locus tags are resistance genes)
(Bi et al., 2015)
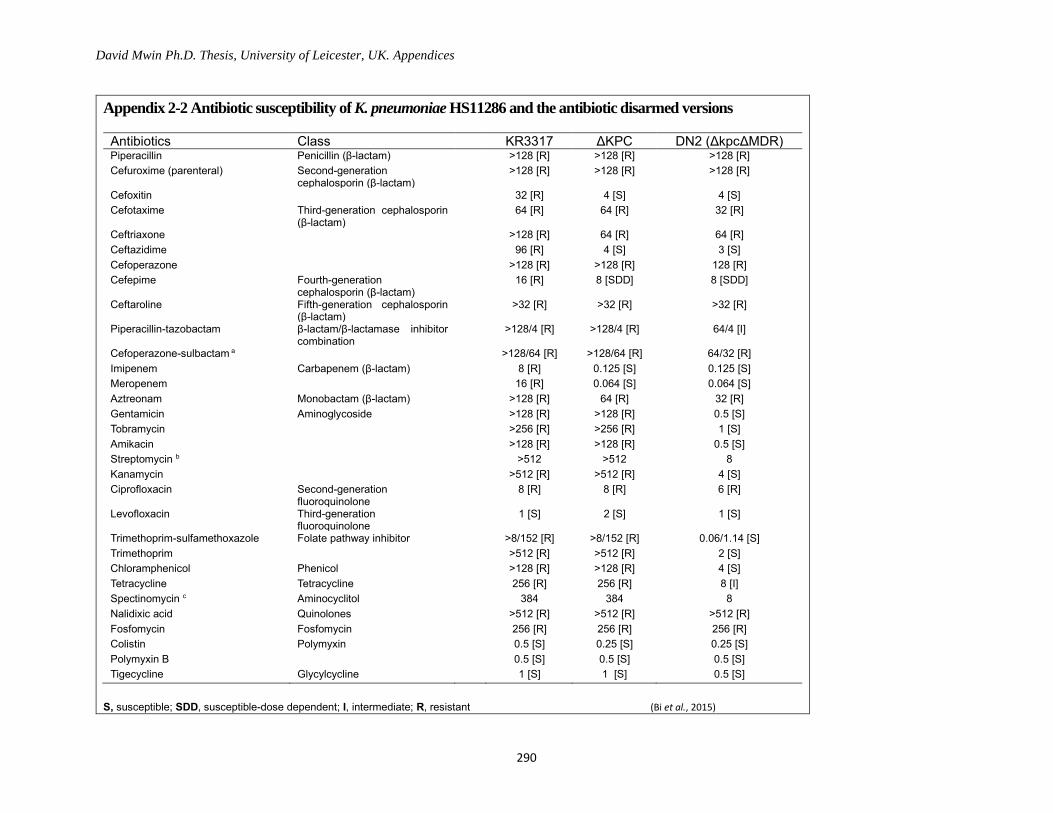
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
290
Appendix 2-2 Antibiotic susceptibility of K. pneumoniae HS11286 and the antibiotic disarmed versions
Antibiotics Class KR3317 ΔKPC DN2 (ΔkpcΔMDR) Piperacillin Penicillin (β-lactam) >128 [R] >128 [R] >128 [R]
Cefuroxime (parenteral) Second-generation cephalosporin (β-lactam)
>128 [R] >128 [R] >128 [R]
Cefoxitin 32 [R] 4 [S] 4 [S]
Cefotaxime Third-generation cephalosporin (β-lactam)
64 [R] 64 [R] 32 [R]
Ceftriaxone >128 [R] 64 [R] 64 [R]
Ceftazidime 96 [R] 4 [S] 3 [S]
Cefoperazone >128 [R] >128 [R] 128 [R]
Cefepime Fourth-generation cephalosporin (β-lactam)
16 [R] 8 [SDD] 8 [SDD]
Ceftaroline Fifth-generation cephalosporin (β-lactam)
>32 [R] >32 [R] >32 [R]
Piperacillin-tazobactam β-lactam/β-lactamase inhibitor combination
>128/4 [R] >128/4 [R] 64/4 [I]
Cefoperazone-sulbactam a >128/64 [R] >128/64 [R] 64/32 [R]
Imipenem Carbapenem (β-lactam) 8 [R] 0.125 [S] 0.125 [S]
Meropenem 16 [R] 0.064 [S] 0.064 [S]
Aztreonam Monobactam (β-lactam) >128 [R] 64 [R] 32 [R]
Gentamicin Aminoglycoside >128 [R] >128 [R] 0.5 [S]
Tobramycin >256 [R] >256 [R] 1 [S]
Amikacin >128 [R] >128 [R] 0.5 [S]
Streptomycin b >512 >512 8
Kanamycin >512 [R] >512 [R] 4 [S]
Ciprofloxacin Second-generation fluoroquinolone
8 [R] 8 [R] 6 [R]
Levofloxacin Third-generation fluoroquinolone
1 [S] 2 [S] 1 [S]
Trimethoprim-sulfamethoxazole Folate pathway inhibitor >8/152 [R] >8/152 [R] 0.06/1.14 [S]
Trimethoprim >512 [R] >512 [R] 2 [S]
Chloramphenicol Phenicol >128 [R] >128 [R] 4 [S]
Tetracycline Tetracycline 256 [R] 256 [R] 8 [I]
Spectinomycin c Aminocyclitol 384 384 8
Nalidixic acid Quinolones >512 [R] >512 [R] >512 [R]
Fosfomycin Fosfomycin 256 [R] 256 [R] 256 [R]
Colistin Polymyxin 0.5 [S] 0.25 [S] 0.25 [S]
Polymyxin B 0.5 [S] 0.5 [S] 0.5 [S]
Tigecycline Glycylcycline 1 [S] 1 [S] 0.5 [S]
S, susceptible; SDD, susceptible-dose dependent; I, intermediate; R, resistant (Bi et al., 2015)

David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
291
Appendix 2-3 Resistance genes identified in K. pneumoniae HS11286 that could confer resistance in Clinical cases
Replicon Gene Locus tag Product Antibiotics to which resistance is known to be conferred
pKPHS1 blaCTX-M-14 KPHS_p100340 Beta-lactamase CTX-M-14 Cefotaxime, ceftriaxone, and aztreonam
pKPHS2 blaKPC-2 KPHS_p200360 KPC-2 Doripenem, ertapenem, imipenem, and meropenem
blaTEM-1 KPHS_p200640 Beta-lactamase TEM-1 Penicillin antibiotics and 1st generation cephalosporin
pKPHS3 blaTEM-1 KPHS_p300540 Beta-lactamase TEM-1 Penicillin antibiotics and 1st generation cephalosporin
rmtB KPHS_p300550 16S rRNA methylase Amikacin, gentamicin, tobramycin, and kanamycin
tetA (G) KPHS_p300630 Tetracycline resistance protein Tetracycline
floR KPHS_p300660 FloR Florfenicol and chloramphenicol
sul1Δ1 KPHS_p300670 Truncated dihydropteroate synthetase
Sulfonamide antibiotics
qacEΔ1 KPHS_p300680 QacEdelta1 fusion protein Intercalating dyes and quaternary ammonium compounds
aadA2 KPHS_p300690 Aminoglycoside adenylyltransferase Streptomycin and spectinomycin.
dfrA12-like KPHS_p300720 Dihydrofolate reductase Trimethoprim
aacC2 KPHS_p300760 Aminoglycoside-(3)-N-acetyltransferase
Gentamicin and tobramycin
blaTEM-1 KPHS_p300840 Beta-lactamase TEM-1 Penicillin antibiotics and 1st generation cephalosporin
strB KPHS_p300860 Aminoglycoside phosphotransferase
Streptomycin
strA KPHS_p300870 Aminoglycoside phosphotransferase
Streptomycin
sul2 KPHS_p300880 Dihydropteroate synthase Sulfonamide antibiotics
blaCTX-M-14 KPHS_p301310 Beta-lactamase CTX-M-14 Cefotaxime, ceftriaxone, and aztreonam
Chromosome blaSHV-11 KPHS_25220 Beta-lactamase SHV-11 Oxacillin, cloxacillin, cefotaxime and ceftriaxon
fosA KPHS_06030 Glutathione transferase Fosfomycin
(Bi et al., 2015)
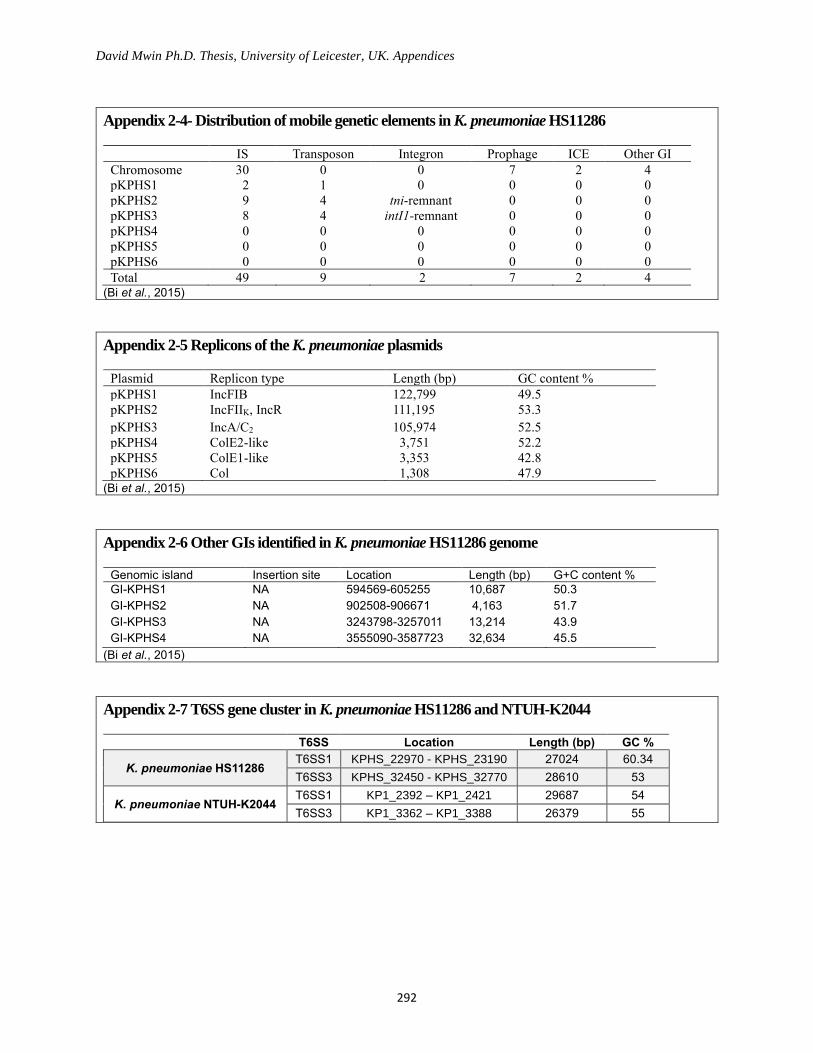
David Mwin Ph.D. Thesis, University of Leicester, UK. Appendices
292
Appendix 2-4- Distribution of mobile genetic elements in K. pneumoniae HS11286
IS Transposon Integron Prophage ICE Other GI
Chromosome 30 0 0 7 2 4
pKPHS1 2 1 0 0 0 0
pKPHS2 9 4 tni-remnant 0 0 0
pKPHS3 8 4 intI1-remnant 0 0 0
pKPHS4 0 0 0 0 0 0
pKPHS5 0 0 0 0 0 0
pKPHS6 0 0 0 0 0 0
Total 49 9 2 7 2 4 (Bi et al., 2015)
Appendix 2-5 Replicons of the K. pneumoniae plasmids
Plasmid Replicon type Length (bp) GC content %
pKPHS1 IncFIB 122,799 49.5
pKPHS2 IncFIIK, IncR 111,195 53.3
pKPHS3 IncA/C2 105,974 52.5
pKPHS4 ColE2-like 3,751 52.2
pKPHS5 ColE1-like 3,353 42.8
pKPHS6 Col 1,308 47.9 (Bi et al., 2015)
Appendix 2-6 Other GIs identified in K. pneumoniae HS11286 genome
Genomic island Insertion site Location Length (bp) G+C content %
GI-KPHS1 NA 594569-605255 10,687 50.3
GI-KPHS2 NA 902508-906671 4,163 51.7
GI-KPHS3 NA 3243798-3257011 13,214 43.9
GI-KPHS4 NA 3555090-3587723 32,634 45.5
(Bi et al., 2015)
Appendix 2-7 T6SS gene cluster in K. pneumoniae HS11286 and NTUH-K2044
T6SS Location Length (bp) GC %
K. pneumoniae HS11286 T6SS1 KPHS_22970 - KPHS_23190 27024 60.34
T6SS3 KPHS_32450 - KPHS_32770 28610 53
K. pneumoniae NTUH-K2044 T6SS1 KP1_2392 – KP1_2421 29687 54
T6SS3 KP1_3362 – KP1_3388 26379 55
Top Related
Copyright © 2022 FDOKUMEN

