







Bahasa
Halaman
Hukum


ORIGINAL PAPER
The bioinvasion of Guam: inferring geographic origin, pace,pattern and process of an invasive lizard (Carlia)in the Pacific using multi-locus genomic data
Christopher C. Austin • Eric N. Rittmeyer •
Lauren A. Oliver • John O. Andermann • George R. Zug •
Gordon H. Rodda • Nathan D. Jackson
Received: 15 October 2010 /Accepted: 27 April 2011 / Published online: 22 May 2011! Springer Science+Business Media B.V. 2011
Abstract Invasive species often have dramatic
negative effects that lead to the deterioration andloss of biodiversity frequently coupled with the
burden of expensive biocontrol programs and sub-
version of socioeconomic stability. The fauna andflora of oceanic islands are particularly susceptible to
invasive species and the increase of global move-ments of humans and their products since WW II has
caused numerous anthropogenic translocations and
increased the ills of human-mediated invasions. Weuse a multi-locus genomic dataset to identify geo-
graphic origin, pace, pattern and historical process of
an invasive scincid lizard (Carlia) that has beeninadvertently introduced to Guam, the Northern
Marianas, and Palau. This lizard is of major impor-
tance as its introduction is thought to have assisted inthe establishment of the invasive brown treesnake
(Boiga irregularis) on Guam by providing a food
resource. Our findings demonstrate multiple waves of
introductions that appear to be concordant withmovements of Allied and Imperial Japanese forces
in the Pacific during World War II.
Keywords Boiga ! Brown Treesnake ! Marianas !Micronesia ! New Guinea ! Palau ! World War II
Introduction
Invasive species affect the ecological co-evolutionary
cohesion of ecosystems with the potential to causedramatic declines in biodiversity over short ecolog-
ical and evolutionary time scales (Rodda et al. 1999).
Invasive species can have extreme negative effects onhuman economic development especially for agricul-
ture and tourism. The economic cost is compounded
by the necessity of establishing and maintainingbiocontrol programs. The seemingly exponential
growth of global movements of humans and theirproducts since World War II has caused numerous
anthropogenic translocations and increased the prob-
lems of human-mediated invasions (Kraus 2009).Species introduced to islands often have a greater
ecological impact than those introduced to mainland
areas (Case and Bolger 1991). Biodiversity lossassociated with invasive species is most easily
studied on islands, and the oceanic islands in the
C. C. Austin (&) ! E. N. Rittmeyer ! L. A. Oliver !J. O. Andermann ! N. D. JacksonDepartment of Biological Sciences, Museum of NaturalScience, 119 Foster Hall, Louisiana State University,Baton Rouge, LA 70803-3216, USAe-mail: [email protected]
G. R. ZugDepartment of Vertebrate Zoology, National Museumof Natural History, Washington, DC 20013, USA
G. H. RoddaUSGS Fort Collins Science Center, 2150 Centre Ave.,Bldg. C, Fort Collins, CO 80526, USA
123
Biol Invasions (2011) 13:1951–1967
DOI 10.1007/s10530-011-0014-y

Pacific offer striking examples of ecosystem altera-tions resulting from biological invasion (Rodda et al.
1991; McKeown 1996; Ota 1999; Urban et al. 2008).
Arguably the best-studied island invasive is thebrown treesnake, Boiga irregularis. This invasive
snake has decimated Guam’s native wildlife and is
responsible for a dramatic decline and extinction ofbird species (Rodda et al. 1999).
Boiga irregularis is assumed to have arrived on
Guam from the Admiralty Islands following WW II(Rodda et al. 1999). Boiga irregularis, however, isnot the only species to arrive on Guam post WW II.
Other presumed post-WWII species colonizing Guamare: the musk shrew (Suncus murinus), the black
drongo (Dicrurus macrocercus), the green anole
(Anolis carolinensis), and the Admiralty brown skink(Carlia ailanpalai) (Fritts and Rodda 1998). This
latter invasive species has received little attention
even though it is hypothesized that C. ailanpalaiindirectly aided the decline and extinction of Guam’s
native birds by providing a stable food source for
juvenile brown treesnakes (Fritts and Rodda 1998). Inaddition, Carlia is invasive on the islands of the
Northern Marianas and the Palau archipelago (Crom-
bie and Pregill 1999).The genus Carlia is composed of more than 40
species with a center of diversity in Australia and New
Guinea (Zug 2004). The Carlia fusca complex is agroup of 18 morphologically similar species found
mainly on New Guinea and surrounding islands (Zug
2004; Zug and Allison 2006; Donnellan et al. 2009).Although the invasive populations of Carlia belong tothe Carlia fusca complex, the morphological unifor-
mity and taxonomic inattention to this group led toconfusion on the specific identity of the invasive
populations (Zug 2004). In 2004, Zug delineated
species boundaries in the fusca complex based ondetailed morphological analysis and assigned species
identity to the invasive populations. He proposed
Carlia ailanpalai (native range, Admiralty Islands) asthe source for the Guam and Northern Marianas
populations and Carlia tutela (native range, Halma-hera) as the source for the Palau populations.
In this study, we use a holotypic and topotypic
multi-locus genomic dataset to identify the species ofintroduced Carlia (Fig. 1; Appendix). These data
reveal the source populations for introduced Carlia ofGuam, the Northern Marianas, and Palau and allowus to examine and correct the taxonomic confusion of
the introduced and native populations (Fig. 1). Inaddition to the pattern of invasion, we address the
mode and timing of translocations and compare the
genetic diversity of various introduced populationswith populations from the native range.
Materials and methods
Sampling
A total of 147 individuals of Carlia were sampled,
including 79 samples from New Guinea and adjacentislands representing potential source populations, and
59 samples from the introduced populations of Guam
and the Northern Marianas Islands (including sam-ples from three major islands: Saipan, Tinian and
Rota), and 9 from the introduced populations from
Palau. Samples were stored in 95% ethanol prior toextraction. Lizards were caught by hand and vou-
chered and deposited in the Texas Natural History
Museum (TNNM), South Australian Museum(SAM), or Louisiana State University Museum of
Zoology (LSUMZ). Outgroup sequences of Carliarhomboidalis, Lygisaurus curtus, Menetia timlowi,and Lampropholis coggeri were selected based on
previous phylogenetic studies involving Carliaskinks (Dolman and Hugall 2008; Donnellan et al.2009), and were obtained from GenBank (Accession
numbers available in Appendix).
DNA isolation, amplification, and sequencing
Whole genomic DNA was extracted from either liveror muscle tissue using an ammonium acetate salt
extraction protocol (Fetzner 1999) or a QiagenDNeasy
Tissue Kit (Qiagen, Inc. Valencia, CA) followingmanufacturer’s instructions. Tissues (*100–200 mg)
were digested overnight with 20 ll proteinase K, andfinal extraction concentrations of DNA were stored inAE Buffer at -20"C prior to amplification.
Two mitochondrial gene regions, including cyto-chrome oxidase b (cyt b) and NADH dehydrogenase
subunit 4 (ND4), and one nuclear locus, Beta-globin
(B-glob), were selected for sequencing based on theutility of these loci in similar groups (primer
sequences and references are available in Table 1).
Target gene regions were amplified by PCR on an MJPTC-200 thermocycler (annealing temperatures
1952 C. C. Austin et al.
123

available in Table 1) using previously published
protocols (Austin et al. 2010b). Amplified PCR
products were purified with 5 units Exonuclease Iand 1.25 units Antarctic Phosphotase (New England
Biolabs, Ipswich, MA) as in Austin et al. (2010a), and
sequenced in both directions using BigDye v. 3.1
Terminator Sequencing Kit (Applied Biosystems,
Foster City, CA) via previously published protocols(Austin et al. 2010b) on an ABI 3100 automated
capillary sequencer.
Fig. 1 Sampling localities of Carlia fusca group skinks inNew Guinea, Palau, and the Marianas Islands. Symbolscorrespond to clades in Fig. 2: open square, C. ailanpalai;closed circle, C. pulla; open triangle, C. mysi, Huon Peninsula;
closed square, C. mysi, Oro, Morobe Provinces; open diamond,C. eothen; closed triangle, C. sp. Northern Milne Bay; opencircle, C. sp. Southern Milne Bay; X, C. sp. Eastern Central;closed diamond, C. luctuosa; ?, C. aenigma
The bioinvasion of Guam 1953
123

Phylogenetic analysis
Sequences were visually verified to check for basedouble-calls or other misreads and complementary
strands were assembled into contigs using Sequencher
v4.7 (Gene Codes Corp., Ann Arbor, MI, USA) andaligned using ClustalX2 (Larkin et al. 2007) under
default settings (Gap opening penalty = 15, Gap
extension penalty = 6.66). All protein-coding regionswere translated to amino acid sequences using Mes-
quite v2.73 (Maddison and Maddison 2010) to verify
that no premature stop codons disrupted the readingframe. Maximum likelihood and Bayesian inference
were used to estimate the phylogeny of Carlia fuscagroup skinks, and to identify the sources of introducedpopulations. Several partitioning schemes, including
partitioning by genome (mitochondrial vs. nuclear),
locus, and codon position, were tested and the optimalpartitioning scheme was selected by comparing the
likelihoods of the resulting topologies using the
Akaike information criterion [AIC] (Akaike 1974).The best fit models of nucleotide substitution for each
partition were selected using the corrected AIC
(Akaike 1974) in jModelTest ver. 0.1.1 (Posada2008). Maximum Likelihood analyses were imple-
mented in using the partition testing version of
GARLI-part v. 0.97 (Zwickl 2006). GARLI analyseswere run under default settings with 50 search
replicates to ensure the maximum likelihood phylog-
eny was identified. Maximum likelihood support forthe optimal partitioning scheme was estimated using
100 bootstrap pseudoreplicates under default settings
with a single search replicate per bootstrap pseudo-replicate. The partitioned Bayesian inference analysis
was implemented in Mr. Bayes v. 3.1.2 (Huelsenbeck
and Ronquist 2001; Ronquist and Huelsenbeck 2003).For all models, the prior nucleotide state frequencies,
transition/transversion ratio, and substitution ratepriors were set as flat Dirichlet distributions, and the
proportion of invariable sites was set as a uniform
(0.0–1.0) prior distribution. Rate priors were set tovariable to allow the nucleotide substitution rates to
vary among partitions. Analyses consisted of two runs,
each of 4 chains with default heating and samplingevery 1,000 generations for 2,50,00,000 generations.
We discarded the first 15,000 topologies as burn-in,
and verified convergence using the program Tracerv1.5 (Rambaut and Drummond 2007) by examining
the posterior probability, log likelihood, and all model
parameters for stationarity and by examining theeffective sample sizes (ESSs), all of which were
substantially greater than 200 at run completion. We
also used Are We There Yet (AWTY) (Nylander et al.2008) to compare the posterior probabilities of all
splits between runs to further verify convergence. At
run completion, the plot comparing posterior proba-bilities of splits between runs was linear, indicating
that the two runs had converged and were sampling the
posterior distribution.To further investigate relationships among intro-
duced Carlia populations and source populations
identified via phylogenetic analyses, we constructedmultilocus haplotype networks for islands containing
introduced populations. Individuals for which we
could not obtain sequence data for all three loci wereexcluded from haplotype network analyses. Genetic
distance matrices were calculated for each locus using
uncorrected p distances and maximum likelihooddistances in Paup* v4.0b10 (Swofford 2003), and
combined to a single genetic distance matrix using
Pofad v.1.03 (Joly and Bruneau 2006). Multilocushaplotype networks were then constructed using the
NeighborNet algorithm (Bryant and Moulton 2004) in
SplitsTree v4.11 (Huson and Bryand 2006).
Table 1 Primers and annealing temperatures used in this study
Locus Primers Primer sequence Temp.("C)
Reference
Cyt B H15149 50-AAA CTG CAG CCC CTC AGA ATG ATA AA-30 48 Kocher et al. (1989)
L14841 50-AAA AAG CTT CCA TCC AAC ATC TCA GC-30 Kocher et al. (1989)
ND4 ND4.light 50-CAC CTA TGA CTA CCA AAA GCT CATGTA GAA GC-30
55 Arevalo et al. (1994)
Leu3.heavy 50-GAA TTA GCA GTT CTT TRT G-30 Stuart-Fox et al. (2002)
b-globin
Bglo1CR 50-GCG AAC TGC ACT GYG ACA AG-30 59 Dolman and Phillips (2004)
Bglo2CR 50-GCT GCC AAG CGG GTG GTG A-30 Dolman and Phillips (2004)
1954 C. C. Austin et al.
123

Genetic diversity
Due to the founder effect associated with introduction,many invasive populations show decreased genetic
diversity relative to native populations. To investigate
this in Carlia, we calculated nucleotide diversity (p)for both native and introduced populations of Carliausing the pegas package ver. 0.3-2 (Paradis 2010) in R
v. 2.11.0. Populations for the calculation of nucleotidediversity were defined as those samples collected with
5 km of one another. Both mitochondrial loci (cyt b
and ND4) were concatenated and samples for whichwe could not obtain sequence for one mitochondrial
locus were excluded; sites with missing data in one or
more samples were also excluded from the calculationof nucleotide diversity.
Results
Phylogenetic analyses
The final aligned dataset was a total of 1603 bp.
Lengths of each partition, numbers of variable sites,and models of nucleotide substitution (selected using
AIC) are provided in Table 2. In both maximum
likelihood and Bayesian inference, a scheme of sevenpartitions was selected as optimal using AIC: B-glob,
cyt b 1st, 2nd, and 3rd codon positions, and ND4 1st,
2nd, and 3rd codon positions. Tracer plots of allparameters, posterior probabilities, and likelihood
were stationary at similar values after the burnin
period for both Bayesian inference analyses, and allESSs were substantially greater than 200 at run
completion. Additionally, AWTY plots comparingthe posterior probabilities of all splits were linear,
indicating that Bayesian inference analyses had
reached convergence.The phylogenetic reconstructions (Fig. 2) strongly
support (Bayesian posterior probability, PP = 1.0,
maximum likelihood bootstrap support, BS = 100)distinct clades that correspond to species. All of the
invasive Carlia populations fall within a clade that
corresponds to C. ailanpalai (holotypic and topotypicmaterial included). Within this C. ailanpalai clade,three major clades are recovered: (A) the Northern
Marianas Islands (Saipan, Tinian and Rota) and theAdmiralty Islands (PP = 0.99, BS = 57), (B) Palau
and New Britain (PP = 1.0, BS = 81), and
(C) Guam, New Ireland, Halmahera and the Admi-ralty Islands (PP = 1.0, BS = 77). These subclades
are all strongly supported in Bayesian analyses
(PP[ 0.99), and, with the exception of subclade A,are strongly supported in maximum likelihood anal-
yses (BS = 57, 81, and 77 for clades A, B, and C,
respectively). Results of the haplotype networks fornative and introduced populations of C. ailanpalaiwere similar between the two methods of genetic
distance calculation, and corroborate the results of thephylogenetic analyses. Samples cluster into three
clades in the haplotype network, corresponding to the
three clades recovered within C. ailanpalai in thephylogenetic analyses (Figs. 2, 3). Genetic diver-
gence among these clades (maximum p distance is
0.0153 for cyt b and 0.0346 for ND4) is lower thanthose among other clades that are currently recong-
nized as good species and indicate at least three
introduction events from three genetically identifiablesource populations for the introduced Pacific popu-
lations of C. ailanpalai: New Britain as the source
population for Palau, the Admiralty Islands as thesource population for the Northern Marianas Islands
(Tinian, Saipan and Rota), and the Admiralty Islands,
New Ireland, or Halmahera as the source populationfor Guam.
Genetic diversity
Nucleotide diversity for the concatenated mitochon-drial dataset is shown in Fig. 4. Despite the larger
sample sizes for the introduced populations relative
to the native populations (Appendix), nucleotidediversity within each of the introduced populations
Table 2 Lengths and number of parsimony informative sitesfor each partition in the optimal partitioning scheme
Partition Length(bp)
Parsimonyinformative sites
Model
cyt b pos. 1 111 25 SYM ? G
cyt b pos. 2 111 7 F81 ? I
cyt b pos. 3 112 92 GTR ? G
ND4 pos. 1 246 79 GTR ? I ? G
ND4 pos. 2 246 34 HKY ? I ? G
ND4 pos. 3 246 206 GTR ? I ? G
B-globin 531 247 GTR ? G
The bioinvasion of Guam 1955
123

is zero, with only single haplotypes recovered for
each sampled population. In contrast, all of the native
populations show expected levels of nucleotidediversity based on previous scincid studies (Smith
et al. 2001; Fuerst and Austin 2004; Austin et al.
2010a; Jackson and Austin 2010). In addition to
nucleotide diversity, nucleotide divergence is alsohigh among native populations and species.
Fig. 2 Maximum likelihood phylogeny of Carlia fusca groupskinks. Numbers on nodes indicate maximum likelihoodbootstrap proportions and Bayesian posterior probabilities,respectively, asterisks indicate bootstrap support of 100 or
posterior probability support of 1.0. The holotype of C.ailanpalai is indicated with an H; paratypes of C. ailanpalaiare indicated with P’s; topotype samples are indicated with T’s
1956 C. C. Austin et al.
123

Discussion
From a broad taxonomic standpoint, some speciesdelimitations of Zug (2004) conflict with the molec-
ular phylogeographic data presented here. Zug (2004)
considered C. ailanpalai as restricted to the Admi-ralty archipelago (Fig. 1); however, our molecular
data demonstrates that C. ailanpalai occurs morewidely including the Bismarck Archipelago, popula-
tions that Zug (2004) assigned to C. mysi, a presumed
widely occurring species distributed along the northcoast of New Guinea. Further, his Halamahera
species, C. tutela, despite being known from pre-
WWII collections (MCZ R-7674, 1907, Patani,Halmahera; RMNH 8659, 3–7 June, 1930 Morotai)
appears to be another invasive population of
C. ailanpalai. Our sampling, however, was limited(n = 2); further sampling from Halmahera and re-
examination of the type series is needed to better
understand the status of the Halmahera population.Our results demonstrate that the uniform morphology
of the C. fusca species group hides considerable
genetic diversity and we are currently investigatingthe apparent decoupling of morphological and
molecular evolution in Carlia.Because of the genetic diversity and divergence
within and among populations of C. ailanpalai acrossits native range, it is possible to trace the geographic
origins of the invasive populations. Interestingly, thethree separate invasions herein identified appear to
track the occupation and movements of Allied and
Imperial Japanese forces in this region during WorldWar II. First, Palau was invaded by Imperial Japanese
forces early in the war as the Japanese forces movedsouth and eastward culminating with their occupation
of northern New Guinea and various Pacific islands.
Fig. 3 Network ofmultilocus data from Carliaailanpalai samplesconstructed using theNeighbornet algorithm inSplitsTree. Clade names(A, B, C) correspond tothose indicated in Fig. 2
Fig. 4 Histogram of intra-population nucleotide diversitycalculated from 1,072 bp of mitochondrial DNA (includingboth cyt b and ND4) for native and introduced populations ofCarlia fusca group skinks. Populations with no bars (includingall introduced populations and some native populations)indicate that a single mitochondrial haplotype was recoveredfrom that population
The bioinvasion of Guam 1957
123

The natural harbor of Rabaul on the northern end ofNew Britain was a major Japanese stronghold, and
the Allies decided to bypass and isolate it in their
campaign to recapture New Guinea and in theirmovement to retake the Philippines (Gailey 2004).
The invasive Palau population is part of a well-
supported clade of samples from New Britainsuggesting that translocation of military materiel
from the Japanese forces on New Britain to Palau
could have resulted in the introduction of C. ailanp-alai from New Britain to Palau. Second, the Guam
populations are part of a well-supported clade from
New Ireland and the Admiralty archipelago, the laterof which was a major base for Allies’ operations and
support the introduction of C. ailanpalai onto Guam
via the transport of Allied military materiel (Morison1953). Finally, the invasive populations from the
Northern Marianas islands of Saipan, Tinian and Rota
are nested in a clade from the Admiralty archipelagoindicating an introduction independent from the
Guam introduction. Thus, the Marianas populations
show the genetic signature of two separate invasions:one invasion of Guam, and a second invasion of the
Northern Marianas Islands of Saipan, Tinian and
Rota.The exact timing of the introductions, however,
remains uncertain. The Bismarck-Palau introduction
presumably occurred during the latter days of theJapanese occupation of Rabaul and their movement
of military materials back to Palau to re-enforce their
Palauan base. The general assumption for the intro-duction of Carlia into the Marianas was post-WWII
removal of military supplies from New Guinea to
permanent bases within the Marianas as there wassubstantial postwar salvage movement of ships and
materiel between Manus and other Pacific islands.
Supporting an hypothesis of WW II era transloca-tions, the first collections of Carlia from these regions
are: Guam 1968 (USNM 192894, CAS 139825-7),
Palau 1968 (MCZ R-104514-6, 104518-24, 111930-73,113491, 114600-40, 118591), and Northern Marianas
(Saipan) 1962 (FMNH 17641-3). Although all thesedates are well past the end of WWII, the initial
propagules may have needed time to become estab-
lished to reach sufficient population densities to attractattention and be collected and deposited in museum
collections.
The broad geographic distribution of C. ailanpalaialong with the corresponding phylogeographic struc-
ture has allowed us to tease out the genetic signature
of the origin, mode and tempo of invasion. Theidentification of the species and specific source
population has the ability to identify potential bio-
control methods such as population-specific parasites.Further, if our hypothesized timeline of WWII
introductions is correct, this suggests that this species,
and possibly other invasive lizards, may require asignificant establishment time (*20–30 years)
before becoming widely abundant. This lag in
establishment, even for species that use secondarygrowth habitats like C. ailanpalai, means that with
appropriate long-term sampling programs there may
be sufficient time to identify, control and eliminateany future island introductions. Vigilant monitoring
and continued museum vouchered collections, there-
fore, are imperative for sound conservation manage-ment. Our data also demonstrates that large
movements of military materiel can transport exotic
invasive species. Given the large scale transport ofmilitary materiel to and from the US and Iraq and
Afghanistan appropriate controls should be imple-
mented to preclude transport of invasive species.
Acknowledgments We thank the people from the manydifferent village communities where we were given theprivilege to conduct fieldwork on their land. We thank B.Roy, V. Kula, and B. Wilmot from the PNG Department ofEnvironment and Conservation, J. Robins from the PNGNational Research Institute. Jim Animiato, Ilaiah Bigilale, andBulisa Iova from the PNG National Museum provided researchassistance in Papua New Guinea. Robert Reed kindly providedsamples from the Marianas and Ben Evans and Iqbal Setiadikindly provided samples from Halmahera. Ken Tighe providedinformation on dates of first collections and discussions withRon Crombie provided valuable information. This manuscriptwas improved from comments from the Austin lab group andLeslie Austin. Fieldwork by GZ in PNG was supported by theSmithsonian Research Awards Program and subsequentmuseum studies of Carlia by the Smithsonian’s ScholarlyStudies Program and Research Opportunities Awards.Research was carried out under LSU IACUC protocol06-071. This research was funded by National ScienceFoundation grants DEB 0445213 and DBI 0400797 to CCA.
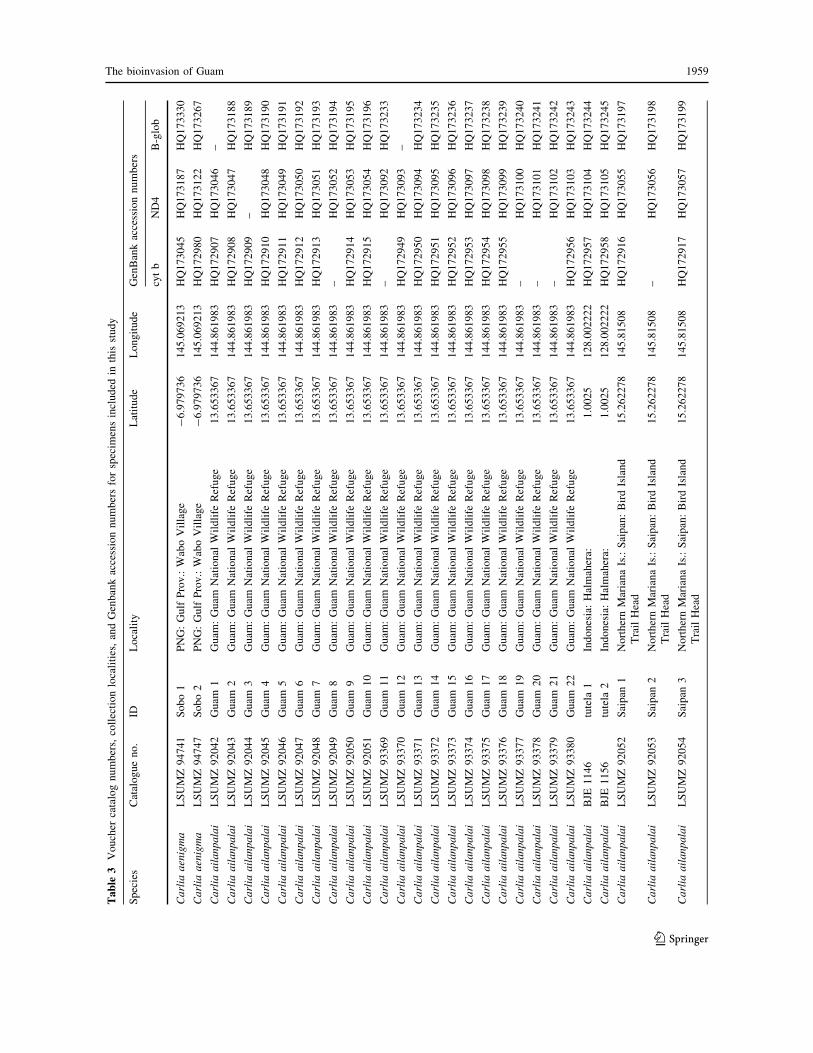
Appendix
See Table 3.
1958 C. C. Austin et al.
123
Tab
le3
Vou
cher
catalognu
mbers,collection
localities,andGenbank
accessionnu
mbers
forspecim
ensinclud
edin
thisstud
y
Species
Catalog
ueno
.ID
Locality
Latitud
eLon
gitude
GenBankaccessionnu
mbers
cytb
ND4
B-glob
Carliaaenigm
aLSUMZ94
741
Sob
o1
PNG:GulfProv.:WaboVillage
-6.97
9736
145.06
9213
HQ17
3045
HQ17
3187
HQ17
3330
Carliaaenigm
aLSUMZ94
747
Sob
o2
PNG:GulfProv.:WaboVillage
-6.97
9736
145.06
9213
HQ17
2980
HQ17
3122
HQ17
3267
Carliaailanp
alai
LSUMZ92
042
Guam
1Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2907
HQ17
3046
–
Carliaailanp
alai
LSUMZ92
043
Guam
2Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2908
HQ17
3047
HQ17
3188
Carliaailanp
alai
LSUMZ92
044
Guam
3Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2909
–HQ17
3189
Carliaailanp
alai
LSUMZ92
045
Guam
4Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2910
HQ17
3048
HQ17
3190
Carliaailanp
alai
LSUMZ92
046
Guam
5Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2911
HQ17
3049
HQ17
3191
Carliaailanp
alai
LSUMZ92
047
Guam
6Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2912
HQ17
3050
HQ17
3192
Carliaailanp
alai
LSUMZ92
048
Guam
7Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2913
HQ17
3051
HQ17
3193
Carliaailanp
alai
LSUMZ92
049
Guam
8Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
–HQ17
3052
HQ17
3194
Carliaailanp
alai
LSUMZ92
050
Guam
9Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2914
HQ17
3053
HQ17
3195
Carliaailanp
alai
LSUMZ92
051
Guam
10Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2915
HQ17
3054
HQ17
3196
Carliaailanp
alai
LSUMZ93
369
Guam
11Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
–HQ17
3092
HQ17
3233
Carliaailanp
alai
LSUMZ93
370
Guam
12Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2949
HQ17
3093
–
Carliaailanp
alai
LSUMZ93
371
Guam
13Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2950
HQ17
3094
HQ17
3234
Carliaailanp
alai
LSUMZ93
372
Guam
14Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2951
HQ17
3095
HQ17
3235
Carliaailanp
alai
LSUMZ93
373
Guam
15Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2952
HQ17
3096
HQ17
3236
Carliaailanp
alai
LSUMZ93
374
Guam
16Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2953
HQ17
3097
HQ17
3237
Carliaailanp
alai
LSUMZ93
375
Guam
17Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2954
HQ17
3098
HQ17
3238
Carliaailanp
alai
LSUMZ93
376
Guam
18Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2955
HQ17
3099
HQ17
3239
Carliaailanp
alai
LSUMZ93
377
Guam
19Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
–HQ17
3100
HQ17
3240
Carliaailanp
alai
LSUMZ93
378
Guam
20Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
–HQ17
3101
HQ17
3241
Carliaailanp
alai
LSUMZ93
379
Guam
21Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
–HQ17
3102
HQ17
3242
Carliaailanp
alai
LSUMZ93
380
Guam
22Guam:Guam
NationalWildlifeRefug
e13
.653
367
144.86
1983
HQ17
2956
HQ17
3103
HQ17
3243
Carliaailanp
alai
BJE
1146
tutela
1Indo
nesia:
Halmahera:
1.00
2512
8.00
2222
HQ17
2957
HQ17
3104
HQ17
3244
Carliaailanp
alai
BJE
1156
tutela
2Indo
nesia:
Halmahera:
1.00
2512
8.00
2222
HQ17
2958
HQ17
3105
HQ17
3245
Carliaailanp
alai
LSUMZ92
052
Saipan1
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2916
HQ17
3055
HQ17
3197
Carliaailanp
alai
LSUMZ92
053
Saipan2
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
–HQ17
3056
HQ17
3198
Carliaailanp
alai
LSUMZ92
054
Saipan3
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2917
HQ17
3057
HQ17
3199
The bioinvasion of Guam 1959
123

Tab
le3
continued
Species
Catalog
ueno
.ID
Locality
Latitud
eLon
gitude
GenBankaccessionnu
mbers
cytb
ND4
B-glob
Carliaailanp
alai
LSUMZ92
055
Saipan4
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
–HQ17
3058
HQ17
3200
Carliaailanp
alai
LSUMZ92
056
Saipan5
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
–HQ17
3059
HQ17
3201
Carliaailanp
alai
LSUMZ92
057
Saipan6
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2918
HQ17
3060
HQ17
3202
Carliaailanp
alai
LSUMZ92
058
Saipan7
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2919
HQ17
3061
HQ17
3203
Carliaailanp
alai
LSUMZ92
059
Saipan8
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2920
HQ17
3062
HQ17
3204
Carliaailanp
alai
LSUMZ92
060
Saipan9
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2921
HQ17
3063
HQ17
3205
Carliaailanp
alai
LSUMZ92
062
Saipan10
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2922
HQ17
3064
–
Carliaailanp
alai
LSUMZ92
064
Saipan11
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2923
HQ17
3065
HQ17
3206
Carliaailanp
alai
LSUMZ92
065
Saipan12
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2924
HQ17
3066
HQ17
3207
Carliaailanp
alai
LSUMZ92
066
Saipan13
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2925
HQ17
3067
HQ17
3208
Carliaailanp
alai
LSUMZ92
067
Saipan14
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2926
HQ17
3068
HQ17
3209
Carliaailanp
alai
LSUMZ92
068
Saipan15
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2927
HQ17
3069
HQ17
3210
Carliaailanp
alai
LSUMZ92
069
Saipan16
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2928
HQ17
3070
HQ17
3211
Carliaailanp
alai
LSUMZ92
070
Saipan17
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2929
HQ17
3071
HQ17
3212
Carliaailanp
alai
LSUMZ92
071
Saipan18
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2930
HQ17
3072
HQ17
3213
Carliaailanp
alai
LSUMZ92
072
Saipan19
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2931
HQ17
3073
HQ17
3214
Carliaailanp
alai
LSUMZ92
073
Saipan20
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2932
HQ17
3074
HQ17
3215
1960 C. C. Austin et al.
123

Tab
le3
continued
Species
Catalog
ueno
.ID
Locality
Latitud
eLon
gitude
GenBankaccessionnu
mbers
cytb
ND4
B-glob
Carliaailanp
alai
LSUMZ92
074
Saipan21
NorthernMariana
Is.:Saipan:
BirdIsland
TrailHead
15.262
278
145.81
508
HQ17
2933
HQ17
3075
HQ17
3216
Carliaailanp
alai
SAMR
4800
2Palau
9Palau:Ang
aur
6.91
6667
134.12
4167
HQ17
2979
HQ17
3121
HQ17
3266
Carliaailanp
alai
SAMR
4770
7Palau
1Palau:Babeldaob
7.39
113
4.51
1433
HQ17
2971
–HQ17
3258
Carliaailanp
alai
SAMR
4770
8Palau
2Palau:Babeldaob
7.39
113
4.51
1433
HQ17
2972
–HQ17
3259
Carliaailanp
alai
SAMR
4775
4Palau
5Palau:Koror
7.33
3333
134.47
HQ17
2975
HQ17
3120
HQ17
3262
Carliaailanp
alai
SAMR
4790
0Palau
6Palau:Koror
7.33
7513
4.47
3333
HQ17
2976
–HQ17
3263
Carliaailanp
alai
SAMR
4790
1Palau
7Palau:Koror
7.33
7513
4.47
3333
HQ17
2977
–HQ17
3264
Carliaailanp
alai
SAMR
4797
5Palau
8Palau:Koror
7.33
7513
4.47
3333
HQ17
2978
–HQ17
3265
Carliaailanp
alai
SAMR
4773
3Palau
4Palau:Malakal
7.32
1833
134.44
1667
HQ17
2974
HQ17
3119
HQ17
3261
Carliaailanp
alai
SAMR
4772
9Palau
3Palau:Ngerm
alik
7.33
6667
134.45
5HQ17
2973
HQ17
3118
HQ17
3260
Carliaailanp
alai
THNM
5138
8New
Britain
1PNG:EastNew
Britain
Prov.:Rabaul
-4.19
5914
152.17
2903
HQ17
2959
HQ17
3106
HQ17
3246
Carliaailanp
alai
THNM
5138
9New
Britain
2PNG:EastNew
Britain
Prov.:Rabaul
-4.19
5914
152.17
2903
HQ17
2960
HQ17
3107
HQ17
3247
Carliaailanp
alai
LSUMZ93
933*
Manus
4PNG:Manus
Prov.:Los
NegrosIs.:Lon
iuVillage
-2.07
0167
147.34
0833
HQ17
2988
HQ17
3130
HQ17
3275
Carliaailanp
alai
LSUMZ93
934*
Manus
5PNG:Manus
Prov.:Los
NegrosIs.:Lon
iuVillage
-2.07
0167
147.34
0833
HQ17
2989
HQ17
3131
HQ17
3276
Carliaailanp
alai
USNM
5600
85**
*Manus
8PNG:Manus
Prov.:Los
NegrosIs.:Peyon
Village
-2.03
2667
147.43
4167
HQ17
2992
HQ17
3134
HQ17
3279
Carliaailanp
alai
USNM
5600
87**
Manus
6PNG:Manus
Prov.:Los
NegrosIs.:Rio
Rio
Village
-2.04
9167
147.41
8667
HQ17
2990
HQ17
3132
HQ17
3277
Carliaailanp
alai
LSUMZ93
941
Manus
7PNG:Manus
Prov.:Los
NegrosIs.:Rio
Rio
Village
-2.04
9167
147.41
8667
HQ17
2991
HQ17
3133
HQ17
3278
Carliaailanp
alai
LSUMZ93
925
Manus
1PNG:Manus
Prov.:Manus
Is.:Kaw
aliap
Village
-2.11
147.06
7667
HQ17
2981
HQ17
3123
HQ17
3268
Carliaailanp
alai
LSUMZ93
926
Manus
2PNG:Manus
Prov.:Manus
Is.:Kaw
aliap
Village
-2.11
147.06
7667
HQ17
2982
HQ17
3124
HQ17
3269
Carliaailanp
alai
LSUMZ93
927
Manus
3PNG:Manus
Prov.:Manus
Is.:Kaw
aliap
Village
-2.11
147.06
7667
HQ17
2983
HQ17
3125
HQ17
3270
Carliaailanp
alai
USNM
5600
88**
Manus
9PNG:Manus
Prov.:Manus
Is.:Tingau
Village
-2.09
614
7.10
55HQ17
2993
HQ17
3135
HQ17
3280
Carliaailanp
alai
USNM
5600
89**
Manus
10PNG:Manus
Prov.:Manus
Is.:Tingau
Village
-2.09
614
7.10
55HQ17
2994
HQ17
3136
HQ17
3281
The bioinvasion of Guam 1961
123

Tab
le3
continued
Species
Catalog
ueno
.ID
Locality
Latitud
eLon
gitude
GenBankaccessionnu
mbers
cytb
ND4
B-glob
Carliaailanp
alai
USNM
5600
90**
Manus
11PNG:Manus
Prov.:Manus
Is.:Tingau’
Village
-2.09
614
7.10
55HQ17
2995
HQ17
3137
HQ17
3282
Carliaailanp
alai
LSUMZ93
942
Manus
12PNG:Manus
Prov.:Ram
butyoIs.:Penchal
Village
-2.34
0514
7.79
45HQ17
2996
HQ17
3138
HQ17
3283
Carliaailanp
alai
USNM
5601
37Manus
13PNG:Manus
Prov.:Ton
gIs.
-2.05
8333
147.75
8333
HQ17
2997
HQ17
3139
HQ17
3284
Carliaailanp
alai
THNM
5146
8New
Ireland3
PNG:New
IrelandProv.:Kavieng
-2.56
6496
150.79
8547
HQ17
2963
HQ17
3110
HQ17
3250
Carliaailanp
alai
THNM
5146
9New
Ireland4
PNG:New
IrelandProv.:Kavieng
-2.56
6496
150.79
8547
HQ17
2964
HQ17
3111
HQ17
3251
Carliaailanp
alai
THNM
5140
6New
Ireland1
PNG:New
IrelandProv.:Lelet
Plateau
-3.43
0352
151.99
8138
HQ17
2961
HQ17
3108
HQ17
3248
Carliaailanp
alai
THNM
5141
1New
Ireland2
PNG:New
IrelandProv.:Lelet
Plateau
-3.43
0352
151.99
8138
HQ17
2962
HQ17
3109
HQ17
3249
Carliaailanp
alai
LSUMZ93
363
Rota1
Rota:
Tenetu
14.137
019
145.15
4436
HQ17
2943
HQ17
3086
HQ17
3227
Carliaailanp
alai
LSUMZ93
364
Rota2
Rota:
Tenetu
14.137
019
145.15
4436
HQ17
2944
HQ17
3087
HQ17
3228
Carliaailanp
alai
LSUMZ93
365
Rota3
Rota:
Veteran’s
Mem
orialPark
14.167
714
145.17
5692
HQ17
2945
HQ17
3088
HQ17
3229
Carliaailanp
alai
LSUMZ93
366
Rota4
Rota:
Veteran’s
Mem
orialPark
14.167
714
145.17
5692
HQ17
2946
HQ17
3089
HQ17
3230
Carliaailanp
alai
LSUMZ93
367
Rota5
Rota:
Veteran’s
Mem
orialPark
14.167
714
145.17
5692
HQ17
2947
HQ17
3090
HQ17
3231
Carliaailanp
alai
LSUMZ93
368
Rota6
Rota:
Veteran’s
Mem
orialPark
14.167
714
145.17
5692
HQ17
2948
HQ17
3091
HQ17
3232
Carliaailanp
alai
LSUMZ92
724
Tinian1
Tinian
14.993
603
145.63
3328
HQ17
2934
HQ17
3076
HQ17
3217
Carliaailanp
alai
LSUMZ92
725
Tinian2
Tinian
14.993
603
145.63
3328
HQ17
2935
HQ17
3077
HQ17
3218
Carliaailanp
alai
LSUMZ92
726
Tinian3
Tinian
14.993
603
145.63
3328
HQ17
2936
HQ17
3078
HQ17
3219
Carliaailanp
alai
LSUMZ92
727
Tinian4
Tinian
14.993
603
145.63
3328
–HQ17
3079
HQ17
3220
Carliaailanp
alai
LSUMZ92
728
Tinian5
Tinian
14.993
603
145.63
3328
HQ17
2937
HQ17
3080
HQ17
3221
Carliaailanp
alai
LSUMZ92
729
Tinian6
Tinian
14.993
603
145.63
3328
HQ17
2938
HQ17
3081
HQ17
3222
Carliaailanp
alai
LSUMZ92
732
Tinian7
Tinian
14.993
603
145.63
3328
HQ17
2939
HQ17
3082
HQ17
3223
Carliaailanp
alai
LSUMZ92
733
Tinian8
Tinian
14.993
603
145.63
3328
HQ17
2940
HQ17
3083
HQ17
3224
Carliaailanp
alai
LSUMZ92
734
Tinian9
Tinian
14.993
603
145.63
3328
HQ17
2941
HQ17
3084
HQ17
3225
Carliaailanp
alai
LSUMZ92
735
Tinian10
Tinian
14.993
603
145.63
3328
HQ17
2942
HQ17
3085
HQ17
3226
Carliaeothen
THNM
5149
6Fergu
sson
1PNG:Milne
Bay
Prov.:D’Entrecasteaux
Arch.:Fergu
sson
Is.
-9.66
4622
150.79
2103
HQ17
2968
HQ17
3115
HQ17
3255
Carliaeothen
THNM
5149
7Fergu
sson
2PNG:Milne
Bay
Prov.:D’Entrecasteaux
Arch.:Fergu
sson
Is.
-9.66
4622
150.79
2103
HQ17
2969
HQ17
3116
HQ17
3256
Carliaeothen
THNM
5150
1Fergu
sson
3PNG:Milne
Bay
Prov.:D’Entrecasteaux
Arch.:Fergu
sson
Is.
-9.66
4622
150.79
2103
HQ17
2970
HQ17
3117
HQ17
3257
1962 C. C. Austin et al.
123

Tab
le3
continued
Species
Catalog
ueno
.ID
Locality
Latitud
eLon
gitude
GenBankaccessionnu
mbers
cytb
ND4
B-glob
Carliaeothen
LSUMZ94
750*
Trobriand
s1
PNG:Milne
Bay
Prov.:Trobriand
Is.:
KiriwinaIs.
-8.54
2208
151.07
9164
HQ17
2984
HQ17
3126
HQ17
3271
Carliaeothen
LSUMZ94
248*
Trobriand
s2
PNG:Milne
Bay
Prov.:Trobriand
Is.:
KiriwinaIs.
-8.54
2208
151.07
9164
HQ17
2985
HQ17
3127
HQ17
3272
Carliaeothen
LSUMZ94
249*
Trobriand
s3
PNG:Milne
Bay
Prov.:Trobriand
Is.:
KiriwinaIs.
-8.54
2208
151.07
9164
HQ17
2986
HQ17
3128
HQ17
3273
Carliaeothen
LSUMZ94
250*
Trobriand
s4
PNG:Milne
Bay
Prov.:Trobriand
Is.:
KiriwinaIs.
-8.54
2208
151.07
9164
HQ17
2987
HQ17
3129
HQ17
3274
Carlialuctuo
saLSUMZ93
564
luctuo
sa1
PNG:Central
Prov.:Kon
eVillage
-8.41
5183
146.95
865
HQ17
3015
HQ17
3157
HQ17
3301
Carlialuctuo
saLSUMZ93
565
luctuo
sa2
PNG:Central
Prov.:Kon
eVillage
-8.41
5183
146.95
865
HQ17
3016
HQ17
3158
HQ17
3302
Carliamysi
THNM
5130
1Huo
n1
PNG:Morob
eProv.:Huo
nPeninsula:near
Oligadu
-6.70
9008
147.57
2558
HQ17
2965
HQ17
3112
HQ17
3252
Carliamysi
THNM
5133
6Huo
n2
PNG:Morob
eProv.:Huo
nPeninsula:near
Oligadu
-6.70
9008
147.57
2558
HQ17
2966
HQ17
3113
HQ17
3253
Carliamysi
THNM
5133
8Huo
n3
PNG:Morob
eProv.:Huo
nPeninsula:near
Oligadu
-6.70
9008
147.57
2558
HQ17
2967
HQ17
3114
HQ17
3254
Carliamysi
LSUMZ93
547
Morob
e1
PNG:Morob
eProv.:Kam
ialiVillage
-7.30
0514
7.13
3667
HQ17
3006
HQ17
3148
HQ17
3292
Carliamysi
LSUMZ93
549
Morob
e2
PNG:Morob
eProv.:Kam
ialiVillage
-7.30
0514
7.13
3667
HQ17
3007
HQ17
3149
HQ17
3293
Carliamysi
LSUMZ93
552
Oro
3PNG:Oro
Prov.:Baseof
Mt.Victory
-9.23
9067
149.05
43HQ17
3009
HQ17
3151
HQ17
3295
Carliamysi
LSUMZ93
553
Oro
4PNG:Oro
Prov.:Baseof
Mt.Victory
-9.23
9067
149.05
43HQ17
3010
HQ17
3152
HQ17
3296
Carliamysi
CCA
2157
Oro
1PNG:Oro
Prov.:KoreafVillage,near
Wanigela
-9.33
7667
149.14
HQ17
2999
HQ17
3141
HQ17
3286
Carliamysi
LSUMZ93
551
Oro
2PNG:Oro
Prov.:KoreafVillage,near
Wanigela
-9.33
7667
149.14
HQ17
3000
HQ17
3142
–
Carliamysi
LSUMZ93
554
Oro
5PNG:Oro
Prov.:Sarad
Mission
,Wanigela
-9.33
914
9.15
8833
HQ17
3011
HQ17
3153
HQ17
3297
Carliapu
lla
LSUMZ93
556
Sandaun
2PNG:Sandaun
Prov.:near
Warom
oVillage
-2.66
6283
141.23
8783
HQ17
3013
HQ17
3155
HQ17
3299
Carliapu
lla
LSUMZ93
557
Sandaun
3PNG:Sandaun
Prov.:near
Warom
oVillage
-2.66
6283
141.23
8783
HQ17
3014
HQ17
3156
HQ17
3300
Carliapu
lla
LSUMZ93
117
Sandaun
4PNG:Sandaun
Prov.:Vanim
o-2.68
4433
141.30
4883
HQ17
3019
HQ17
3161
HQ17
3305
Carliapu
lla
LSUMZ92
712
Sandaun
5PNG:Sandaun
Prov.:Vanim
o-2.68
4433
141.30
4883
HQ17
3020
HQ17
3162
HQ17
3306
Carliapu
lla
LSUMZ92
713
Sandaun
6PNG:Sandaun
Prov.:Vanim
o-2.68
4433
141.30
4883
HQ17
3021
HQ17
3163
HQ17
3307
Carliapu
lla
LSUMZ93
118
Sandaun
7PNG:Sandaun
Prov.:Vanim
o-2.68
4433
141.30
4883
HQ17
3022
HQ17
3164
HQ17
3308
Carliapu
lla
LSUMZ93
555
Sandaun
1PNG:Sandaun
Prov.:Warom
oVillage
-2.66
8214
1.24
42HQ17
3012
HQ17
3154
HQ17
3298
The bioinvasion of Guam 1963
123

Tab
le3
continued
Species
Catalog
ueno
.ID
Locality
Latitud
eLon
gitude
GenBankaccessionnu
mbers
cytb
ND4
B-glob
Carliasp.‘‘Amau
fuscagrou
p’’
LSUMZ94
695
Amau
1PNG:Central
Prov.:Amau
Village
-10
.036
814
8.56
4633
HQ17
3037
HQ17
3179
HQ17
3323
Carliasp.‘‘Amau
fuscagrou
p’’
LSUMZ94
696
Amau
2PNG:Central
Prov.:Amau
Village
-10
.036
814
8.56
4633
HQ17
3038
HQ17
3180
HQ17
3324
Carliasp.‘‘Amau
fuscagrou
p’’
LSUMZ94
697
Amau
3PNG:Central
Prov.:Amau
Village
-10
.036
814
8.56
4633
HQ17
3039
HQ17
3181
HQ17
3325
Carliasp.‘‘Amau
fuscagrou
p’’
LSUMZ94
719
Amau
4PNG:Central
Prov.:Amau
Village
-10
.036
814
8.56
4633
HQ17
3040
HQ17
3182
HQ17
3326
Carliasp.‘‘Milne
Bay
North
Coast’’
LSUMZ93
156
MB
12PNG:Milne
Bay
Prov.:Eastern
Cape
-10
.230
083
150.87
45HQ17
3031
HQ17
3173
HQ17
3317
Carliasp.‘‘Milne
Bay
North
Coast’’
LSUMZ92
880
MB
11PNG:Milne
Bay
Prov.:Huh
unaVillage
-10
.284
817
150.60
105
HQ17
3030
HQ17
3172
HQ17
3316
Carliasp.‘‘Milne
Bay
North
Coast’’
USNM
5601
07MB
1PNG:Milne
Bay
Prov.:Iapo
a#2
Village
-10
.258
667
150.57
35HQ17
3001
HQ17
3143
HQ17
3287
Carliasp.‘‘Milne
Bay
North
Coast’’
USNM
5601
08MB
2PNG:Milne
Bay
Prov.:Iapo
a#2
Village
-10
.258
667
150.57
35HQ17
3002
HQ17
3144
HQ17
3288
Carliasp.‘‘Milne
Bay
North
Coast’’
USNM
5600
99MB
3PNG:Milne
Bay
Prov.:NetuliIs.
-10
.234
333
150.59
7667
HQ17
3003
HQ17
3145
HQ17
3289
Carliasp.‘‘Milne
Bay
North
Coast’’
LSUMZ92
877
MB
10PNG:Milne
Bay
Prov.:Top
uraVillage
-10
.193
315
0.33
705
HQ17
3029
HQ17
3171
HQ17
3315
Carliasp.‘‘Milne
Bay
Sou
th’’
LSUMZ93
128
MB
6PNG:Milne
Bay
Prov.:NapatanaLod
gegrou
nds,Alotau
-10
.305
867
150.43
7183
HQ17
3025
HQ17
3167
HQ17
3311
Carliasp.‘‘Milne
Bay
Sou
th’’
LSUMZ93
129
MB
7PNG:Milne
Bay
Prov.:NapatanaLod
gegrou
nds,Alotau
-10
.305
867
150.43
7183
HQ17
3026
HQ17
3168
HQ17
3312
Carliasp.‘‘Milne
Bay
Sou
th’’
LSUMZ92
874
MB
8PNG:Milne
Bay
Prov.:NapatanaLod
gegrou
nds,Alotau
-10
.305
867
150.43
7183
HQ17
3027
HQ17
3169
HQ17
3313
Carliasp.‘‘Milne
Bay
Sou
th’’
LSUMZ92
875
MB
9PNG:Milne
Bay
Prov.:NapatanaLod
gegrou
nds,Alotau
-10
.305
867
150.43
7183
HQ17
3028
HQ17
3170
HQ17
3314
Carliasp.‘‘Milne
Bay
Sou
th’’
LSUMZ93
102
MB
13PNG:Milne
Bay
Prov.:Oilpalm
plantation
,westof
Alotau
-10
.399
283
150.09
1617
HQ17
3032
HQ17
3174
HQ17
3318
Carliasp.‘‘Milne
Bay
Sou
th’’
LSUMZ93
103
MB
14PNG:Milne
Bay
Prov.:Oilpalm
plantation
,westof
Alotau
-10
.399
283
150.09
1617
HQ17
3033
HQ17
3175
HQ17
3319
Carliasp.‘‘Milne
Bay
Sou
th’’
CCA
4802
MB
16PNG:Milne
Bay
Prov.:PadiPadiVillage
-10
.402
115
0.06
2717
HQ17
3035
HQ17
3177
HQ17
3321
1964 C. C. Austin et al.
123

Tab
le3
continued
Species
Catalog
ueno
.ID
Locality
Latitud
eLon
gitude
GenBankaccessionnu
mbers
cytb
ND4
B-glob
Carliasp.‘‘Milne
Bay
Sou
th’’
LSUMZ92
583
MB
17PNG:Milne
Bay
Prov.:PadiPadiVillage
-10
.402
115
0.06
2717
HQ17
3036
HQ17
3178
HQ17
3322
Carliasp.‘‘Milne
Bay
Sou
th’’
USNM
5601
36MB
4PNG:Milne
Bay
Prov.:SagaAHO
River
-10
.544
167
150.11
5167
HQ17
3004
HQ17
3146
HQ17
3290
Carliasp.‘‘Milne
Bay
Sou
th’’
USNM
5601
01MB
5PNG:Milne
Bay
Prov.:SagaAHO
River
-10
.544
167
150.11
5167
HQ17
3005
HQ17
3147
HQ17
3291
Carliasp.‘‘Milne
Bay
Sou
th’’
LSUMZ95
752
MB
15PNG:Milne
Bay
Prov.:Takwatakwai
Village
-10
.325
617
150.03
765
HQ17
3034
HQ17
3176
HQ17
3320
Outgrou
ps:
Carliabicarina
taLSUMZ93
542
bicarinata
1PNG:NCD:PortMoresby
,National
Museum
grou
nds
-9.42
5565
147.19
0129
HQ17
3008
HQ17
3150
HQ17
3294
Carliabicarina
taLSUMZ93
545
bicarinata
2PNG:NCD:PortMoresby
,2Mile
-9.47
3283
147.17
76HQ17
3017
HQ17
3159
HQ17
3303
Carliabicarina
taLSUMZ93
546
bicarinata
3PNG:NCD:PortMoresby
,2Mile
-9.47
3283
147.17
76HQ17
3018
HQ17
3160
HQ17
3304
Carliabicarina
taLSUMZ92
680
bicarinata
4PNG:Central
Prov.:Boo
tlessBay
-9.50
9333
147.29
0183
HQ17
3023
HQ17
3165
HQ17
3309
Carliabicarina
taLSUMZ92
681
bicarinata
5PNG:Central
Prov.:Boo
tlessBay
-9.50
9333
147.29
0183
HQ17
3024
HQ17
3166
HQ17
3310
Carliasp.‘‘Amau
bicarinate’’
LSUMZ94
724
Amau
bicarinate
1PNG:Central
Prov.:Amau
Village
-10
.036
814
8.56
4633
HQ17
3041
HQ17
3183
HQ17
3327
Carliasp.‘‘Amau
bicarinate’’
LSUMZ94
731
Amau
bicarinate
2PNG:Central
Prov.:Amau
Village
-10
.036
814
8.56
4633
HQ17
3042
HQ17
3184
HQ17
3328
Carliasp.‘‘Amau
bicarinate’’
LSUMZ94
732
Amau
bicarinate
3PNG:Central
Prov.:Amau
Village
-10
.036
814
8.56
4633
HQ17
3043
HQ17
3185
HQ17
3329
Carliasp.‘‘Amau
bicarinate’’
LSUMZ94
526
Amau
bicarinate
4PNG:Central
Prov.:Amau
Village
-10
.036
814
8.56
4633
HQ17
3044
HQ17
3186
–
Carliarhom
boidalis
AJ406
203
DQ35
0070
DQ34
9566
Lygisau
ruscurtus
HQ17
2998
HQ17
3140
HQ17
3285
Menetia
timlowi
–AJ290
552
AY50
8895
Lam
prop
holiscogg
eri
–FJ379
463
AY50
8900
*Indicatestopo
types
**Indicatesparatypes
***Indicatesho
lotypes
The bioinvasion of Guam 1965
123

References
Akaike H (1974) A new look at the statistical model identifi-cation. IEEE Trans Autom Control 19:716–723
Arevalo E, Davis SK, Sites JW Jr (1994) Mitochondrial DNAsequence divergence and phylogenetic relationshipsamong eight chromosome races of the Sceloporus gram-micus complex (Phrynosomatidae) in Central Mexico.Syst Biol 43:387–418
Austin CC, Rittmeyer EN, Richards SJ, et al (2010a) Phylog-eny, historical biogeography and body size evolution inPacific Island Crocodile skinks Tribolonotus (Squamata;Scincidae). Mol Phylogenet Evol 57:227–236
Austin CC, Spataro M, Peterson S, et al (2010b) Conservationgenetics of Boelen’s python (Morelia boeleni) from NewGuinea: reduced genetic diversity and divergence ofcaptive and wild animals. Conserv Genet 11:889–896
Bryant D, Moulton V (2004) Neighbor-Net: an agglomerativemethod for the construction of phylogenetic networks.Mol Biol Evol 21:255–265
Case TJ, Bolger DT (1991) The role of introduced species inshaping the distribution and abundance of island reptiles.Evol Ecol 5:272–290
Crombie RI, Pregill GK (1999) A checklist of the herpetofaunaof the Palau islands (Republic of Belau), Oceania. Her-petol Monogr 13:29–80
Dolman G, Hugall AF (2008) Combined mitochondrial andnuclear data enhance resolution of a rapid radiation ofAustralian rainbow skinks (Scincidae: Carlia). MolPhylogenet Evol 49:782–794
Dolman G, Phillips BP (2004) Single copy nuclear DNAmarkers characterized for comparative phylogeography inAustralian wet tropics rainforest skinks. Mol Ecol Notes4:185–187
Donnellan SC, Couper PJ, Saint KM, et al (2009) Systematicsof the Carlia ‘fusca’ complex (Reptilia: Scincidae) fromnorthern Australia. Zootaxa 2227:1–31
Fetzner FWJ (1999) Extracting high-quality DNA from shedreptile skins: a simplified method. BioTechniques 26:1052–1054
Fritts TH, Rodda GH (1998) The role of introduced species inthe degradation of island ecosystems: a case history ofGuam. Annu Rev Ecol Syst 29:113–140
Fuerst GS, Austin CC (2004) Population genetic structure ofthe prairie Skink (Eumeces septentrionalis): nested cladeanalysis of post Pleistocene populations. J Herpetol38:257–268
Gailey HA (2004) MacArthur’s victory: the war in New Gui-nea, 1943–1944. Presidio Press, New York
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesianinference of phylogeny. Bioinformatics 17:754–755
Huson DH, Bryand D (2006) Application of phylogeneticnetworks in evolutionary studies. Mol Biol Evol23:254–267
Jackson ND, Austin CC (2010) The combined effects of riversand refugia generate extreme cryptic fragmentation withinthe common ground skink (Scincella lateralis). Evolution64:409–428
Joly S, Bruneau A (2006) Incorporating allelic variation forreconstructing the evolutionary history of organisms from
multiple genes: an example from Rosa in North America.Syst Biol 55:623–636
Kocher TD, Thomas WK, Meyer A, et al (1989) Dynamics ofmitochondrial DNA evolution in animals: amplificationand sequencing with conserved primers. Proceedings ofthe National Academy of Sciences of the United States ofAmerica 86:6196–6200
Kraus F (2009) Alien reptiles and amphibians: a scientificcompendium and analysis. Springer Science, New York
Larkin MA, Blackshields G, Brown NP, Chenna R, McGetti-gan PA, McWilliam H, Valentin F, Wallace IM, Wilm A,Lopez R, Thompson JD, Gibson TJ, Higgins DG (2007)Clustal W and Clustal X version 2.0. Bioinformatics23:2947–2948
Maddison WP, Maddison DR (2010) Mesquite: a modular sys-tem for evolutionary analysis. Version 2.73 http://mesquiteproject.org
McKeown S (1996) A field guide to reptiles and amphibians ofthe Hawaiian islands. Diamond Head Publishing, LosOsos
Morison SE (1953) History of United States naval operations inworld war II volume 8: New Guinea and the Marianas,March 1944–August 1944. University of Illinois Press,Urbana
Nylander JAA, Wilgenbusch JC, Warren DL, Swofford DL(2008) AWTY (are we there yet?): a system for graphicalexploration of MCMC convergence in Bayesian phylog-enetics. Bioinformatics 24:581–583
Ota H (1999) Introduced amphibians and reptiles of the Ry-ukyu Archipelago, Japan. In: Rodda GH, Sawai Y, Chis-zar D, Tanaka H (eds) Problem snake management: theHabu an the brown treesnake. Comstock PublishingAssociates, Ithaca, pp 439–452
Paradis E (2010) pegas: an R package for population geneticswith an integrated-modular approach. Bioinformatics26:419–420
Posada D (2008) jModelTest: phylogenetic model averaging.Mol Biol Evol 25:1253–1256
Rambaut A, Drummond AJ (2007) Tracer v1.4. Available fromhttp://beast.bio.ed.ac.uk/Tracer
Rodda GH, Fritts TH, Reichel JD (1991) Distributional pat-terns of reptiles and amphibians in the Mariana Islands.Micronesica 24:195–210
Rodda GH, Sawai Y, Chiszar D, Tanaka H (1999) Problemsnake management: the Habu an the brown treesnake.Comstock Publishing Associates, Ithaca
Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinfor-matics 19:1572–1574
Smith SA, Austin CC, Shine R (2001) A phylogenetic analysisof variation in reproducive mode within an Australianlizard (Saiphos equalis, Scincidae). Biol J Linn Soc74:131–139
Stuart-Fox DM, Hugall AF, Moritz C (2002) A molecularphylogeny of rainbow skinks (Scincidae: Carlia): taxo-nomic and biogeographic implications. Aust J Zool 50:39–51
Swofford DL (2003) PAUP*. Phylogenetic analysis usingparsimony (*and Other Methods). Version 4. SinauerAssociates, Sunderland
1966 C. C. Austin et al.
123

Urban MC, Phillips BL, Skelly DK, Shine R (2008) A toadmore traveled: the heterogeneous invasion dynamics ofcane toads in Australia. Am Nat 171:134–148
Zug GR (2004) Systematics of the Carlia ‘‘fusca’’ lizards ofNew Guinea and nearby islands. Bishop Museum Press,Honolulu
Zug GR, Allison A (2006) New Carlia fusca complex lizards(Reptilia: Squamata: Scincidae) from New Guinea,Papua-Indonesia. Zootaxa 1237:27–44
Zwickl DJ (2006) Genetic algorithm approaches for the phy-logenetic analysis of large biological sequence datasetsunder the maximum likelihood criterion. Ph.D. disserta-tion, The University of Texas, Austin. Available from:http://www.bio.utexas.edu/faculty/antisense/garli/Garli.html
The bioinvasion of Guam 1967
123
Top Related
Copyright © 2022 FDOKUMEN

