







Bahasa
Halaman
Hukum


www.theriojournal.com
Theriogenology 68 (2007) 223–236
Respiration rates correlate with mRNA expression of G6PD and
GLUT1 genes in individual bovine in vitro-produced blastocysts
A.S. Lopes a,b,*, C. Wrenzycki c, N.B. Ramsing d, D. Herrmann c,H. Niemann c, P. Løvendahl a, T. Greve b, H. Callesen a
a Department of Genetics and Biotechnology, Faculty of Agricultural Sciences, University of Aahrus, DK-8830 Tjele, Denmarkb Department of Large Animal Sciences, Faculty of Life Sciences, University of Copenhagen, DK-1870 Frederiksberg C, Denmark
c Department of Biotechnology, Institute for Animal Breeding, Federal Agricultural Research Centre (FAL), Mariensee, 31535 Neustadt, Germanyd Unisense FertiliTech A/S, DK-8200 Skejby, Denmark
Received 2 February 2007; received in revised form 23 April 2007; accepted 25 April 2007
Abstract
Quantification of embryo respiration is a promising procedure to assess embryonic metabolism and possibly select viable
embryos. At the blastocyst stage, ATP is produced by glycolysis and oxidative phosphorylation, processes that require uptake of
oxygen and glucose, which is regulated by the expression of GLUT1 and G6PD. The purpose of the present study was to investigate
the relationship between respiration rates and relative abundances of G6PD and GLUT1 transcripts in individual bovine blastocysts
produced in vitro. Respiration rates of 104 bovine in vitro-produced blastocysts were measured individually using the
nanorespirometer technology. Real-time RT-PCR was employed to determine the relative abundance of G6PD and GLUT1
mRNA in individual embryos. The mean respiration rates were similar for male and female blastocysts of the same developmental
stage, but the sex ratio was skewed towards males. GLUT1 expression was down-regulated in female versus male embryos. In
contrast, a �1.8-fold increase in the expression of G6PD mRNA was observed in female blastocysts when compared to male
blastocysts, indicating that dosage compensation for this gene had not yet occurred. Both GLUT1 and G6PD expression levels were
affected by morphological quality and stage of development. Expression of GLUT1 and G6PD mRNAs was correlated with
respiration rates, indicating that, in metabolically active blastocysts, uptake of oxygen and glucose are jointly increased. These
findings suggest that expression of genes for oxidative phosphorylation and glycolysis are both involved in oxygen demanding ATP
production.
# 2007 Elsevier Inc. All rights reserved.
Keywords: Respiration; GLUT1; G6PD; Oxygen; Bovine embryo
1. Introduction
Assessment of embryo viability is crucial for the
successful establishment and maintenance of pregnancy
* Corresponding author. Current address: Research Centre for
Reproductive Health, Department of Obstetrics and Gynaecology,
The University of Adelaide, 2nd Floor, Medical School South, Ade-
laide 5005, SA, Australia. Tel.: +61 8 83038188; fax: +61 8 83034099.
E-mail address: [email protected] (A.S. Lopes).
0093-691X/$ – see front matter # 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.theriogenology.2007.04.055
following embryo transfer, both in human and livestock
species [1–4]. Usually, single embryos are transferred in
cattle breeding and commercial herds, and elective
single embryo transfer is increasingly applied also in
human IVF clinics [5]. Thus, it is essential to select and
transfer only good quality embryos in order to increase
in vivo development.
Morphological evaluation based on light micro-
scopic observation is still the most commonly used
method to assess embryo quality [2,6–8]. However, this

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236224
type of quality evaluation is subjective and requires
specialized training [2,9,10]. To overcome these limita-
tions, several approaches for embryo quality evaluation
have been proposed in the past few years including
analysis of the kinetics of embryonic development [11–
13], gene expression profiling [14–16] and determination
of embryo metabolism [17,18].
The energy metabolism of the embryo has been
subjected to conspicuous evaluation, and considerable
evidence has accumulated that embryonic metabolic
activity may be used to predict embryonic develop-
mental potential [9,19–27]. At the blastocyst stage, ATP
is generated by glycolysis and in particular oxidative
phosphorylation [28–30], processes that ultimately
require oxygen [31]. The oxygen consumption of
murine [23,25,27,32,33], bovine [28,34–36] and por-
cine [37] embryos have been previously assessed and a
relationship between embryonic oxygen consumption
and viability following transfer has been earlier
described [36,38–40].
Oxygen consumption of individual embryos increases
significantly around compaction and blastulation [25,28,
33,41,42], and is associated with a higher ATP demand to
accommodate increased protein synthesis and activity of
the Na+/K+-dependent ATPase [28,43]. Embryonic
oxygen consumption is significantly affected by culture
conditions, specifically by the glucose [20], serum [44],
amino acids and protein content in culture medium
[45,46] or by oxygen incubation conditions [47]. Oxygen
consumption is higher among embryos of superior
quality [32,36,48–50], but large variations in respiration
rates within each class of morphological quality have
been reported [34,35,50]. Embryonic diameter is some-
what related to respiration as bovine embryos of greater
size have higher respiration rates [35,50]. The stage of the
bovine blastocyst has also been associated with oxygen
consumption, with embryos of more advanced stages
showing higher oxygen consumption [37,51–52]. Over-
all, embryonic sex does not seem to influence respiration
rates [32,35] but the oxygen consumption of good quality
female bovine embryos has been reported to be higher
than that of good quality male embryos. [32].
Glucose is an important energy substrate for the
development of bovine embryos, particularly during
the post compaction period [53]. After entering the
embryonic cells via facilitative glucose transporters
(GLUT), glucose can be metabolized through the
glycolytic pathway or may enter the pentose phosphate
pathway (PPP). The metabolism of glucose through
glycolysis generates pyruvate, which can either be
transformed to lactate or to acetyl CoA that in turn can
be further metabolized by the tricarboxylic acid cycle
and the oxidative phosphorylation pathway. Concomi-
tant with compaction and blastulation, glucose uptake
and metabolism are substantially increased [22,28,
33,41,54]. Despite this increase, the glycolytic pathway
contributes little to ATP production as most of the
glucose is primarely converted to lactate, which is an
ineffective source of ATP production. In fact, glycolysis
only contributes 12, 15 and 28% to the total ATP
produced by mouse, bovine and human blastocysts,
respectively [29]. Glucose uptake tends to be higher
in female as compared to male embryos [19,55].
Furthermore, glucose uptake was shown to be higher in
morphologically normal than in degenerated embryos
[56] and in blastocysts with an increased viability
following transfer to a recipient animal [19,55,57].
Glucose-6-phosphate dehydrogenase gene (G6PD)
maps to the X-chromosome [58] and encodes the rate-
limiting enzyme of the PPP, which is crucially involved
in the metabolism of glucose-6-phosphate. The PPP is
activated during development and generates nicotina-
mide adenine dinucleotide phosphate (NADPH) for
biosynthesis of steroids and fatty acids. It also provides
cells with ribose-5-phosphate for the synthesis of
nucleotides and nucleic acids [59]. Through the
NADPH generation in the PPP, G6PD also participates
in the detoxification of oxygen radicals [60,61] and
therefore has been considered a cytoprotective enzyme
for oxidative stress and DNA damage [61]. The relative
abundance (RA) of G6PD has shown to be nearly two-
fold higher in female pre-implantation bovine embryos
produced in vitro as compared to their male counter-
parts [62–64]. This phenomenon has been associated
with alterations in dosage compensation under in vitro
conditions, likely induced by problems or delay in X-
chromosome inactivation. The high levels of G6PD
were proposed to be responsible for the delayed
development of in vitro female embryos by mediating
NADPH production and thus modulating radical levels
[62,65].
GLUT1 belongs to the family of facilitative glucose
transporters and is expressed after in vitro maturation
of the oocyte [16]. Expression levels of GLUT1 are
significantly higher in the trophectoderm cells than in
the inner cell mass cells of bovine embryos [66], and
within the trophectoderm cells GLUT1 is specifically
found in the lateral membranes. GLUT1 expression
increases with increasing glucose uptake by the embryo
[67,68]. In vitro culture conditions, such as the oxygen
tension [42,69], the presence or absence of serum
[67,70] and the levels of glucose in the culture medium
[71] were shown to affect the expression of GLUT1.
This transcript is down-regulated in in vitro-produced

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236 225
bovine embryos as compared to their in vivo counter-
parts [70,72,73].
The present study investigated the relationship
between individual respiration rates of day 7 bovine
in vitro-produced blastocysts and the relative abundance
of GLUT1 and G6PD mRNA transcripts. To gain a
better understanding on whether genetic patterns can
be reflected at the phenotypic level, we evaluated the
effects of morphological quality, stage of embryonic
development, sex and embryo diameter on the GLUT1
and G6PD transcriptional activity. Preliminary results
of this work have been presented earlier [74].
2. Materials and methods
2.1. Experimental design
A total of 750 bovine oocytes were used in this study
(50 oocytes per replicate with 15 independent repli-
cates). Oocytes were submitted to in vitro production
(IVP) and from the resulting embryos, 5 or 10
blastocysts were randomly selected for subsequent
assessment of oxygen consumption. On each day of
measurement, the respiration rate of 5 (one rosette/day)
or 10 (two rosettes/day) day 7 embryos produced in
vitro were measured in duplicate. In total, the
respiratory activity of 104 embryos was assessed
during 15 experimental days. The measured embryos
were classified in six classes, according to the stage of
embryonic development: not expanded (n = 16), early
expanded (n = 20), expanded (n = 35), late expanded
(n = 22), collapsed (n = 5) or hatching (n = 6) blas-
tocysts. Sex diagnosis and gene expression analysis
were performed on 81 embryos. The RA of GLUT1 and
G6PD transcripts was not considered for 3 embryos due
to insufficient amount of DNA of the samples. Oxygen
measurements on 4 embryos were regarded as non-
consistent and these results were not included in the
analysis. Furthermore, 3 embryos with extreme values
for RA of G6PD or respiration rate were considered as
outliers and thus not included in the analysis. Thus, only
71 embryos were included in the full statistical analysis
to evaluate the relationship between respiration rate,
embryo morphology, sex, stage of development and
abundance of GLUT1 and G6PD transcripts.
2.2. In vitro embryo production (IVP)
The method used for IVP has been reported
previously [75]. Briefly, bovine immature cumulus-
oocyte complexes (COCs) were aspirated from slaugh-
terhouse ovaries, selected (i.e. excluding denuded
COCs and COCs with clear degenerative signs) and
matured for 24 h in four-well dishes (Nunc, Roskilde,
Denmark). Each well contained 400 ml of bicarbonate
buffered TCM-199 medium (Gibco BRL, Paisley, UK)
supplemented with 15% cattle serum (CS; Danish Food
and Veterinary Institute, Frederiksberg, Denmark),
10 IU/ml eCG and 5 IU/ml hCG (Suigonan Vet; Intervet
Scandinavia, Skovlunde, Denmark). The oocytes were
matured under mineral oil at 38.5 8C in 5% CO2 in
humidified air. Fertilization was performed in modified
Tyrode’s medium [76] using frozen–thawed, Percoll-
selected sperm. After 22 h, cumulus cells were removed
by vortexing and presumptive zygotes were transferred
to 400 ml of culture medium composed of synthetic
oviduct fluid (SOF) medium containing amino acids,
citrate and inositol (SOFaaci [75]) supplemented with
antibiotics (gentamycin sulphate, 50 mg/ml) and 5%
CS, and subsequently incubated at 38.5 8C in 5% CO2,
5% O2, 90% N2 atmosphere with maximum humidity.
2.3. In vitro embryo selection and evaluation
On each day of experiment, 5–10 embryos at the
blastocyst stage (day 7) were randomly selected to
represent all classes of morphological quality, and
individually transferred into one well of a four-well dish
containing culture medium. Blastocysts were evaluated
according to the stage of embryonic development (see
above) and classified as quality I, II, III or IV based on
microscopic observation of the morphological quality
(excellent, good, fair or poor blastocysts, respectively).
In a previous work using the same IVP system the mean
cell numbers of day 7 expanded or haching blastocysts,
early expanded blastocysts and not expanded blasto-
cysts were 134 � 33, 95 � 26 and 76 � 22, respectively
[75]. The diameter of each blastocyst (i.e. from the outer
periphery of the zona pellucida) was measured under a
stereomicroscope (SMZ800; Nikon, Tokyo, Japan)
using an ocular micrometer eyepiece. Digital images
of each embryo were acquired using a digital still
camera (GC-X3E; JVL, Yokohama, Japan) mounted on
an inverted optical microscope (TDM, Nikon) with a
thermal control microscope stage (CO 102; Linkam
Scientific Instruments Ltd., Tadworth, Surrey, UK).
2.4. Measurement of oxygen consumption
The nanorespirometer system (Unisense A/S, Aar-
hus, Denmark) used to measure the oxygen consump-
tion of single embryos has recently been described [35].
Briefly, each of the embryos was transferred from the
four-well dish into the bottom of each glass capillary in

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236226
the rosette (measuring unit composed of seven fused
glass capillaries). A total of five blastocysts were
loaded into each rosette while two glass capillaries
were left empty to serve as negative references
without respiratory activity (=control). The rosette
was subsequently placed into the rosette disc holder
and submerged into the culture medium in the beaker
(80 ml), which was maintained in a semi-closed
system at 38.5 8C and under constant flow of
humidified air (21% oxygen) with 5% CO2. The
system was left undisturbed for one h, until a steady-
state linear oxygen gradient was established inside the
glass capillaries of the rosette. Subsequently, oxygen
concentration gradients generated by the respiration
of the embryo were determined by measuring the
oxygen concentration at consecutive measurement
points inside the capillary with an oxygen micro-
sensor. Both embryo-containing capillaries and empty
capillaries were measured in duplicate. Calculation of
the mean respiration rate was based on the two oxygen
concentration profiles obtained from each embryo,
after subtraction of the background noise measured in
the empty reference capillaries.
2.5. Embryo processing after the measurements and
freezing procedure
With the purpose of preserving the genetic material
for sexing and gene expression analysis, day 7
embryos produced in vitro were individually trans-
ferred from the rosette into a well of a four-well dish
containing culture medium. Subsequently, individual
embryos were transferred to a four-well dish contain-
ing 500 ml Ca2+-free and Mg2+-free phosphate-
buffered saline (Biowhittaker, Walkersville, MD)
supplemented with 4 mg/ml of polyvinylpyrrolidone
and washed four times using a glass pipette. Finally,
embryos were individually transferred under the
stereomicroscope in a minimum volume of medium
(5 ml or less) into 1.5 ml eppendorf tubes, which were
immediately snap-frozen in liquid nitrogen and stored
at �80 8C.
2.6. Sex determination
DNA from each individual embryo was prepared
using the Microcon YM-100 (42412; Millipore,
Eschborn, Germany) columns according to the instruc-
tions of the manufacturer, except for minor modifica-
tions. The supernatant from the RNA extraction was
diluted with 200 ml H2O and centrifuged for 10 min.
Another 200 ml were then added to the column followed
by centrifugation for 8 min. Subsequently, the column
was inverted and centrifuged at 350 � g for 2 min. One
half of the supernatant (�10 ml) was used for PCR.
PCR analysis was performed using bovine-specific
(50-AGGTCGCGAGATTGGTCGCT-AGGTC ATGC
A-30 and 30-AAGACCTCGAGAGACCCTCTTCAA-
CACGT-50; accession number PCT WO 86/07095) and
Y-chromosome-specific primers (50-CCTCCCCTTGT-
TCAAACGCCCGGAATCATT-30 and 30-TGCTTGA-
CTGCAGGGACCGAGAGGTTTGGG-50; accession
number PCT WO 86/07095) as previously described
[61]. The PCR products were subjected to electrophor-
esis on a 2% agarose gel in 1� TBE buffer (90 mM Tris,
90 mM borate, 2 mM EDTA, pH 8.3) containing
0.2 mg/ml ethidium bromide. The image of each gel
was recorded using a CCD camera (Quantix; Photo-
metrics, Munchen, Germany). Sex determination was
performed in duplicate for each individual embryo.
2.7. Determination of the relative abundance of
GLUT1 and G6PD transcripts in individual
embryos by real-time reverse transcription-
polymerase chain reaction (RT-PCR)
Poly(A)+ RNA was isolated from single blastocysts
as previously described [67] and immediately used for
reverse transcription (RT), which was carried out in a
total volume of 20 ml using 2.5 mM random hexamers
(Applied Biosystems, Vaterstetten, Germany). Prior to
RNA isolation, 1 pg of rabbit globin RNA (Invitrogen,
Karlsruhe, Germany) was added as an internal standard
and was analyzed simultaneously. The rabbit globin
mRNA represents a mixture of the a- and b-chains
derived from polyribosomes of reticulocytes and was
purified via oligo-dT cellulose chromatography. The
reaction mixture consisted of 1� RT buffer (50 mM
KCl, 10 mM Tris–HCl, pH 8.3, Invitrogen), 5 mM
MgCl2, 1 mM of each dNTP (Amersham, Brunswick,
Germany), 2.5 mM random hexamers (Applied Biosys-
tems), 20 IU RNase inhibitor (Applied Biosystems) and
50 IU MuLV reverse transcriptase (Applied Biosys-
tems). The RT reaction was carried out in a PTC-200
thermocycler (MJ Research, Watertown, MA, USA) for
10 min at 25 8C, 60 min at 42 8C, followed by a
denaturation step for 5 min at 99 8C and flash cooling on
ice.
Immediately after RT, the samples were subjected to
real-time RT-PCR, which was performed with 0.05
embryo equivalents (percentage of the volume from
the RT reaction employing one embryo in a defined
volume). The PCR primers for each gene were designed
using the OLIGOTM program (Plymouth, MN, USA).

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236 227
The sequences of the primer pairs to detect globin,
GLUT1 and G6PD transcripts were respectively:
50-GCAGCCACGGTGGCGAGTAT-30 (position 241–
260) and 30-GTGGGACAGGAGCTT-GAAAT-50 (posi-
tion 555–657), 50-CAGGAGATGAAGGAGGAGAGC-
30 (position 894–915) and 30-CACAAATAGCGACAC-
GACAGT-50 (position 1131–1152), 50-GTTCTTCAA-
CCC-CGAGGAGTC-30 (position 748–769) and 30-ATGTGGTGGAGCAGTGGAGTG-50 (position 910–
931). The corresponding accession numbers were
X04751, M60448.1 and XM_583628.1, respectively.
A standard curve consisting of dilutions of globin
cDNA originating from 1 pg globin RNA was included
as reference in each run. The standard curve consisted
of five standards of the globin cDNA diluted from 50 to
0.08 fg. This standard curve was used for calculation of
the concentration of each target gene as the amplifica-
tion efficiency of the target genes (GLUT1, G6PD)
and the reference gene (globin) is comparable. The
amplification efficiency of GLUT1, G6PD and globin
was compared by preparing serial dilutions for all the
genes from cDNA samples. After real-time amplifica-
tion, the threshold cycle (CT) values obtained were
used for the standard curve construction. The ampli-
fication efficiency (E) was calculated according to the
following equation: E = 10(�1/S) � 1 (S: slope of the
standard curve). Differences in the CT values between
the target genes and the reference gene were then
plotted against the logarithm of the template amount
(slope of the resulting straight line <0.1; data not
shown).
Real-time RT-PCR was carried out in a Lightcycler
apparatus using the LightCyclerFastStart DNA Mas-
ter + SYBR Green I Kit (Roche Diagnostik, Man-
nheim, Germany) according to the instructions of
the manufacturer. The PCR reaction consisted of a
cDNA equivalent to 0.05 blastocysts, 0.5 mM of each
primer, 4 ml of the mix containing dNTP’s, MgCl2,
FastStart Taq DNA polymerase and SYBR Green I,
in a total volume of 20 ml. The cycling profile
included the denaturation of the sample and activation
of the FastStart Taq DNA polymerase at 95 8Cfor 15 min, and 44 PCR cycles (denaturation at
96 8C for 10 s, annealing at 60 8C for 10 s and
elongation at 72 8C for 10 s) followed by a melting
curve. The LightCycler Software Version 3.5 (Roche
Diagnostik) was used to quantify and analyze the
results. Purity and specificity of both GLUT1 and
G6PD amplicons were verified by analysis of the
melting curve and the correct size of the products
was confirmed by electrophoresis on a 2% agarose
gel.
2.8. Statistical analysis
Sex ratios of the embryos were compared with
an expected 50:50 ratio by a corrected chi square
procedure (Freq Procedure [77]; Yate’s adjustment
[78]).
The background noise of the nanorespirometer,
measured by assessing the apparent oxygen consump-
tion within the empty reference capillaries (zero-value),
was evaluated by a two-tailed Student’s t-test. Linear
regression (Reg Procedure, [77]) was used to determine
the association between the first and second oxygen
measurements of each embryo. Normalization of the
results obtained for the individual genes of each
blastocyst by Real-Time RT-PCR was performed by
calculating the ratio of abundance of mRNA for the
target genes to globin mRNA.
Respiration rates and the RA of GLUT1 and G6PD in
in vitro embryos were tested for the distributional
properties and deviation from normal distribution was
significant (Univariate procedure [77]). Data was
subsequently square root transformed, as this was
effective in stabilizing variance and obtaining approx-
imate normal distribution of data and residuals.
Simple relationships between respiration rates and
RA of GLUT1 and of G6PD were firstly investigated as
correlations and through linear regression of respiration
rate on RA of GLUT1 and G6PD.
The mRNA expression of GLUT1 and G6PD were
analyzed independently using linear mixed models
(Mixed Procedure, [77]). For GLUT1, the model
included sex, diameter, time point of the measurement
in relation to in vitro fertilization, square root of
the RA of G6PD as well as interaction between
morphological quality and stage of embryonic
development as fixed effects. For G6PD, the model
included sex, diameter, time point of the measurement
in relation to in vitro fertilization, square root of the
RA of GLUT1 expression as well as interaction
between morphological quality and stage of embryo-
nic development as fixed effects. Random effects
were not included in these models as the effects of
rosette and date of measurement were non-significant
in relation to the variable in question. Least square
means (LSM) were produced for the factors in the
final model.
Factors affecting embryo diameter were assessed
using a linear mixed model (Mixed Procedure [77]).
The model included sex, time point of the measurement
in relation to in vitro fertilization, morphological
quality, stage of embryonic development, RA of GLUT1
and G6PD and the interaction of morphological quality
A.S. Lopes et al. / Theriogenology 68 (2007) 223–236228
and stage of development as fixed effects. Date of
measurement was tested as random effect but it was
regarded as non-significant and consequently excluded
from the model. LSM were produced by the final model
after removal of both interactions, which were shown to
be non-significant.
Respiration rates of day 7 embryos were subse-
quently analyzed using a linear mixed model (Mixed
Procedure [77]), including time points of the mea-
surement in relation to in vitro fertilization, diameter,
stage of embryonic development, morphological
quality, sex, square root of the RA of GLUT1 and
G6PD transcripts and interaction between morpho-
logical quality and stage of embryonic development
as fixed effects. The random part of the model
contained an interaction between rosette and date of
measurement. LSM were produced by the previous
model after removal of non-significant effects. The
interaction of date of measurement and rosette was
tested by a linear mixed model (Asycov, Covtest [77])
before being introduced in the random part of the
model.
All statistical procedures were performed using the
computational software of SAS [77]. Data was
presented as LSM � S.E. and the level of significance
was P < 0.05, unless otherwise indicated.
3. Results
3.1. Sex ratio of embryos selected for oxygen
measurements
Among the 81 blastocysts evaluated for sex and
morphology (Table 1), a total of 51 were male and thus
the average ratio of male to female blastocysts was
63:37, which deviated significantly from the expected
50:50 ratio in all morphological quality categories
(P < 0.05).
Table 1
Morphological quality and sex ratio of day 7 bovine in vitro-produced
blastocysts
Morphological
quality
Number of
embryos
Male:female
number/(ratio)
I 19 13:6/(68:32a)
II 27 17:10/(63:37a)
III 27 16:11/(59:41a)
IV 8 5:3/(62:38a)
Total 81 51:30/(63:37a)
a Significantly different from anticipated 50:50 ratio of males to
females (P < 0.05).
3.2. Consistency and accuracy of the oxygen
measurements
The background noise of the nanorespirometer
system was very small, although significantly different
from zero, with the empty reference capillaries showing
an apparent oxygen consumption of 0.016 � 0.0047 nl/
h (n = 35; P < 0.01). The results of the first and second
oxygen measurements of the same embryos were highly
correlated (R2 = 0.985; n = 85; P < 0.0001) and the
slope of the regression line was not significantly
different from one (y2 = 0.981 (�0.014) y1 + 0.025
(�0.023); P > 0.05).
3.3. Relative abundance of GLUT1 and G6PD
transcripts in in vitro-produced embryos
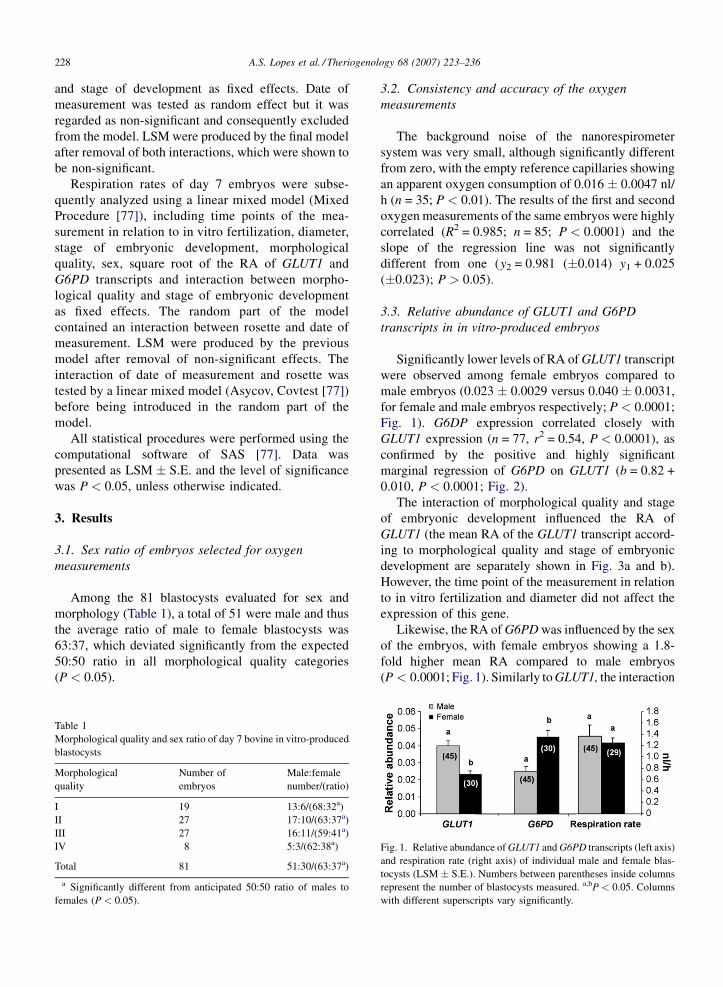
Significantly lower levels of RA of GLUT1 transcript
were observed among female embryos compared to
male embryos (0.023 � 0.0029 versus 0.040 � 0.0031,
for female and male embryos respectively; P < 0.0001;
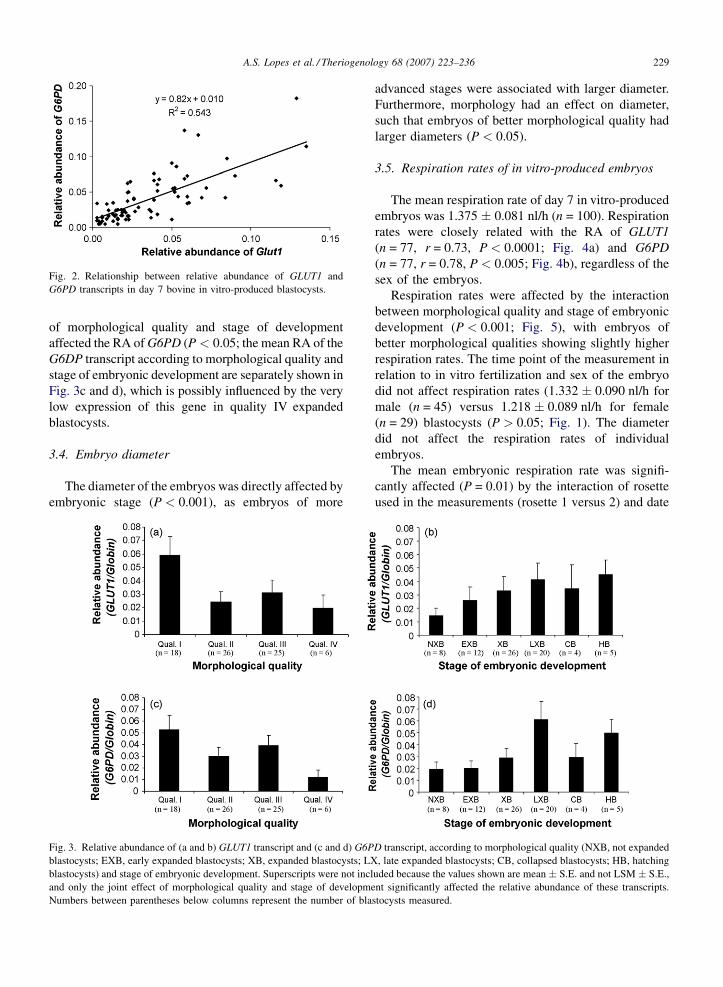
Fig. 1). G6DP expression correlated closely with
GLUT1 expression (n = 77, r2 = 0.54, P < 0.0001), as
confirmed by the positive and highly significant
marginal regression of G6PD on GLUT1 (b = 0.82 +
0.010, P < 0.0001; Fig. 2).
The interaction of morphological quality and stage
of embryonic development influenced the RA of
GLUT1 (the mean RA of the GLUT1 transcript accord-
ing to morphological quality and stage of embryonic
development are separately shown in Fig. 3a and b).
However, the time point of the measurement in relation
to in vitro fertilization and diameter did not affect the
expression of this gene.
Likewise, the RA of G6PD was influenced by the sex
of the embryos, with female embryos showing a 1.8-
fold higher mean RA compared to male embryos
(P < 0.0001; Fig. 1). Similarly to GLUT1, the interaction
Fig. 1. Relative abundance of GLUT1 and G6PD transcripts (left axis)
and respiration rate (right axis) of individual male and female blas-
tocysts (LSM � S.E.). Numbers between parentheses inside columns
represent the number of blastocysts measured. a,bP < 0.05. Columns
with different superscripts vary significantly.
A.S. Lopes et al. / Theriogenology 68 (2007) 223–236 229
Fig. 2. Relationship between relative abundance of GLUT1 and
G6PD transcripts in day 7 bovine in vitro-produced blastocysts.
of morphological quality and stage of development
affected the RA of G6PD (P < 0.05; the mean RA of the
G6DP transcript according to morphological quality and
stage of embryonic development are separately shown in
Fig. 3c and d), which is possibly influenced by the very
low expression of this gene in quality IV expanded
blastocysts.
3.4. Embryo diameter
The diameter of the embryos was directly affected by
embryonic stage (P < 0.001), as embryos of more
Fig. 3. Relative abundance of (a and b) GLUT1 transcript and (c and d) G6P
blastocysts; EXB, early expanded blastocysts; XB, expanded blastocysts; LX
blastocysts) and stage of embryonic development. Superscripts were not incl
and only the joint effect of morphological quality and stage of developme
Numbers between parentheses below columns represent the number of bla
advanced stages were associated with larger diameter.
Furthermore, morphology had an effect on diameter,
such that embryos of better morphological quality had
larger diameters (P < 0.05).
3.5. Respiration rates of in vitro-produced embryos
The mean respiration rate of day 7 in vitro-produced
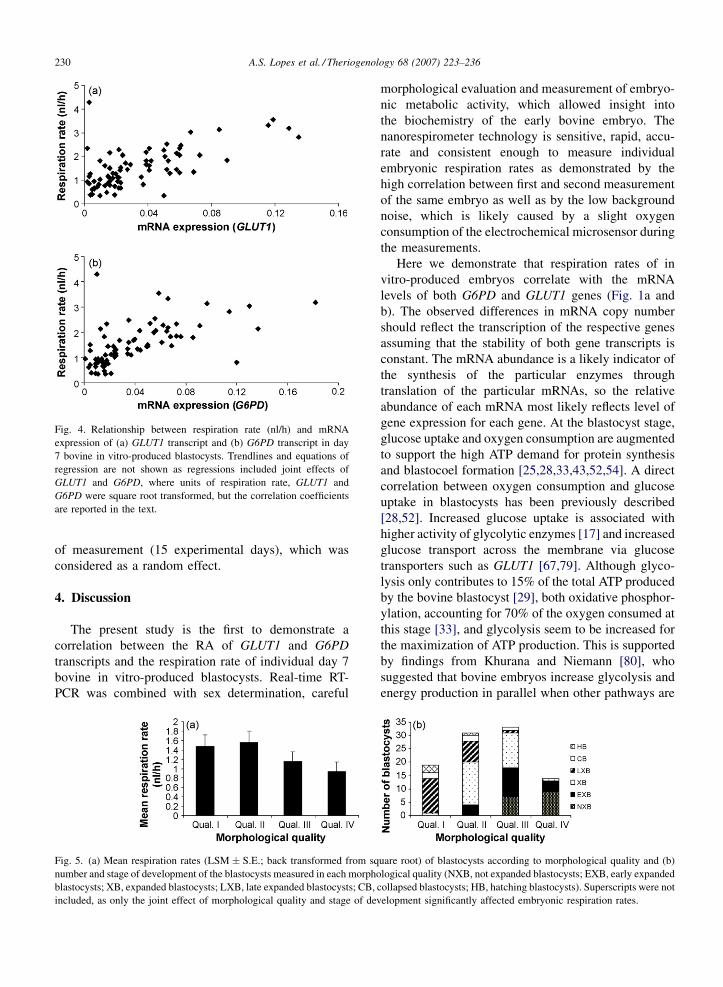
embryos was 1.375 � 0.081 nl/h (n = 100). Respiration
rates were closely related with the RA of GLUT1
(n = 77, r = 0.73, P < 0.0001; Fig. 4a) and G6PD
(n = 77, r = 0.78, P < 0.005; Fig. 4b), regardless of the
sex of the embryos.
Respiration rates were affected by the interaction
between morphological quality and stage of embryonic
development (P < 0.001; Fig. 5), with embryos of
better morphological qualities showing slightly higher
respiration rates. The time point of the measurement in
relation to in vitro fertilization and sex of the embryo
did not affect respiration rates (1.332 � 0.090 nl/h for
male (n = 45) versus 1.218 � 0.089 nl/h for female
(n = 29) blastocysts (P > 0.05; Fig. 1). The diameter
did not affect the respiration rates of individual
embryos.
The mean embryonic respiration rate was signifi-
cantly affected (P = 0.01) by the interaction of rosette
used in the measurements (rosette 1 versus 2) and date
D transcript, according to morphological quality (NXB, not expanded
, late expanded blastocysts; CB, collapsed blastocysts; HB, hatching
uded because the values shown are mean � S.E. and not LSM � S.E.,
nt significantly affected the relative abundance of these transcripts.
stocysts measured.
A.S. Lopes et al. / Theriogenology 68 (2007) 223–236230
Fig. 4. Relationship between respiration rate (nl/h) and mRNA
expression of (a) GLUT1 transcript and (b) G6PD transcript in day
7 bovine in vitro-produced blastocysts. Trendlines and equations of
regression are not shown as regressions included joint effects of
GLUT1 and G6PD, where units of respiration rate, GLUT1 and
G6PD were square root transformed, but the correlation coefficients
are reported in the text.
of measurement (15 experimental days), which was
considered as a random effect.
4. Discussion
The present study is the first to demonstrate a
correlation between the RA of GLUT1 and G6PD
transcripts and the respiration rate of individual day 7
bovine in vitro-produced blastocysts. Real-time RT-
PCR was combined with sex determination, careful
Fig. 5. (a) Mean respiration rates (LSM � S.E.; back transformed from sq
number and stage of development of the blastocysts measured in each morpho
blastocysts; XB, expanded blastocysts; LXB, late expanded blastocysts; CB,
included, as only the joint effect of morphological quality and stage of dev
morphological evaluation and measurement of embryo-
nic metabolic activity, which allowed insight into
the biochemistry of the early bovine embryo. The
nanorespirometer technology is sensitive, rapid, accu-
rate and consistent enough to measure individual
embryonic respiration rates as demonstrated by the
high correlation between first and second measurement
of the same embryo as well as by the low background
noise, which is likely caused by a slight oxygen
consumption of the electrochemical microsensor during
the measurements.
Here we demonstrate that respiration rates of in
vitro-produced embryos correlate with the mRNA
levels of both G6PD and GLUT1 genes (Fig. 1a and
b). The observed differences in mRNA copy number
should reflect the transcription of the respective genes
assuming that the stability of both gene transcripts is
constant. The mRNA abundance is a likely indicator of
the synthesis of the particular enzymes through
translation of the particular mRNAs, so the relative
abundance of each mRNA most likely reflects level of
gene expression for each gene. At the blastocyst stage,
glucose uptake and oxygen consumption are augmented
to support the high ATP demand for protein synthesis
and blastocoel formation [25,28,33,43,52,54]. A direct
correlation between oxygen consumption and glucose
uptake in blastocysts has been previously described
[28,52]. Increased glucose uptake is associated with
higher activity of glycolytic enzymes [17] and increased
glucose transport across the membrane via glucose
transporters such as GLUT1 [67,79]. Although glyco-
lysis only contributes to 15% of the total ATP produced
by the bovine blastocyst [29], both oxidative phosphor-
ylation, accounting for 70% of the oxygen consumed at
this stage [33], and glycolysis seem to be increased for
the maximization of ATP production. This is supported
by findings from Khurana and Niemann [80], who
suggested that bovine embryos increase glycolysis and
energy production in parallel when other pathways are
uare root) of blastocysts according to morphological quality and (b)
logical quality (NXB, not expanded blastocysts; EXB, early expanded
collapsed blastocysts; HB, hatching blastocysts). Superscripts were not
elopment significantly affected embryonic respiration rates.

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236 231
already operating at a maximum rate. Furthermore,
glycolysis is affected by oxygen availability in each
blastomere via the hypoxia-inducing factor 1 (HIF-1),
which is regulated by the oxygen sensor, that itself
accounts for part of the oxidative phosphorylation-
independent oxygen consumed by the blastocyst [41].
The observed increase in GLUT1 expression among
embryos with higher respiration rates, and hence with
higher metabolic activity, suggests an important role of
this glucose transporter in the metabolism of the pre-
implantation bovine embryo. However, it has been
suggested that GLUT3 instead of GLUT1 is the main
transporter of glucose from the external environment
and that GLUT1 is primarily responsible for mediating
the glucose efflux into the blastocoel cavity and the
uptake by the inner cell mass cells [79,81]. It would thus
be interesting to investigate if the expression of GLUT3
is equally increased in embryos with higher respiration
rates.
Similar to GLUT1, mRNA expression of G6PD was
up-regulated in embryos with a higher level of
respiratory activity (Fig. 4b). The finding that G6PD
expression was correlated with GLUT1 expression
(Fig. 2) suggests that higher levels of GLUT1 increase
the uptake of glucose by the cells, which in turn leads to
higher amounts of glucose metabolized through the PPP
in the form of glucose-6-phosphate. The high influx of
glucose is likely to be associated with an allosteric
increase of the activity of G6PD, which in turn
stimulates glucose cycling through the PPP. It thus
seems that higher ATP demands are responsible for
higher respiration rates and glucose uptake by the
embryos, leading to enhanced glycolytic and pentose
phosphate pathways and consequently to increased
GLUT1 and G6PD expression. Alternatively, the
increased G6PD expression associated with the higher
oxygen uptake by the embryos could be caused by the
presence of reactive oxygen species (ROS). A higher
oxygen consumption causes an enhanced ATP produc-
tion by mitochondrial oxidative phosphorylation, but
also produces ROS [41]. G6PD is critically involved in
the production of NADPH, which detoxifies ROS.
Furthermore, G6PD is the only NADPH-producing
enzyme activated in response to oxidative stress [82]. It
is therefore possible that the presence of higher levels of
ROS generated by the increased oxygen consumption
triggers G6PD activation. To further unravel this
intriguing hypothesis, the expression pattern of genes
related to stress response, such as the CU/ZN-SOD and
in particular the HSP70.1, should be investigated. This
would help clarifying whether the higher levels of
respiratory activity and G6PD are indicative of embryo
viability or are merely a reaction to an increased stress
response. Moreover, assessment of cell number in each
individual blastocyst would be useful for understanding
if the increased expression of GLUT1 and G6PD in
more advances stages is or not merely a reflection of a
higher cell number.
Individual respiration rates were correlated with the
RA of GLUT1 and G6PD transcripts. Furthermore, the
interaction between morphological quality and stage of
embryonic development also influenced respiration
rates (Fig. 5), but the limited number of observations
within each stage of development and morphological
quality made results interpretation difficult. The fact
that collapsing embryos might not necessarily have
been at a more advanced stage than expanding embryos
could have also been a confounding factor. Differences
in respiration rates between embryos of different
morphological qualities have shown that oxygen
consumption is higher among embryos of superior
quality [32,35,38,49,50]. However the partial correla-
tion between morphological quality and respiration rate
of individual embryos observed in the present and
recent studies [35,41] also supports the concept that
morphological differences are to a certain extent
reflected at the physiological level. Thus, a combination
of oxygen consumption measurements and morpholo-
gical assessment of quality will be instrumental to
improve embryo selection. However, it should be noted
that high respiration rates could reflect an overproduc-
tion of ROS, and thus very high respiration rates could
represent a manifestation of metabolic stress instead of
superior quality.
Changes in morphology at the blastocyst stage
require the expression of key genes from the embryonic
genome. To our knowledge, differences in GLUT1 and
G6PD expression in relation to the morphological
quality of the embryos have not been reported
previously. This is likely explained by the fact that
transcriptional profiles are normally analyzed solely in
good quality embryos. Patterns of gene expression in
relation to morphological quality have been analyzed by
comparing gene expression of in vitro-produced
embryos with that of their in vivo counterparts
[16,42,70], which are considered to be of superior
quality. Blastocysts of the best morphological quality
and at the hatching/hatched stage expressed the highest
amount of GLUT1 mRNA among all analyzed embryos
(Fig. 3a and b). In contrast, GLUT1 expression was
relatively low in embryos of inferior morphological
quality (IV) (Fig. 3a). In agreement with the present
study, Wrenzycki et al. [70] and Harvey et al. [42]
reported that GLUT1 expression was reduced in in

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236232
vitro-produced embryos as compared to embryos
produced in vivo and suggested that reduced amounts
of GLUT1 were indicative of compromised develop-
mental potential (poor bovine embryo quality). Based
on these results, it is plausible to speculate that a higher
expression of the GLUT1 transcript is observed in better
quality embryos, but a larger study including a higher
number of embryos representative of all morphological
qualities and stages of development is required to fully
clarify this. Furthermore, quantification of the total cell
number of each blastocyst will help elucidating if the
abundance of transcripts represents increased quality or
is merely a result of the number of cells in each
blastocyst, which is not related to embryo quality as
evaluated following embryo transfer [3].
Quality I embryos seemed to show the highest G6PD
mRNA levels (Fig. 3c). Similar to GLUT1 expression,
mRNA expression of G6PD was relatively low in
quality IV embryos. Previous reports have shown that
G6PD expression was significantly higher in in vitro-
produced blastocysts than in their in vivo counterparts
[64]. However, these differences probably reflect
incomplete X-chromosome inactivation of in vitro-
produced blastomeres rather than differences in embryo
quality.
Embryonic diameter was not correlated with
respiration rates or expression of GLUT1 and G6PD.
However, diameter was affected by the embryonic
stage, which might explain the lack of influence of
diameter on G6PD and GLUT1 expression levels as
well as on respiration, when both parameters were
simultaneously analyzed. Embryonic diameter had
previously been related to respiration, with significantly
larger embryos associated with higher respiration rates
[35,50], but not with glucose metabolism [80]. In the
present study, the number of cells of each blastocyst
was not determined. The number of cells of individual
embryos appears to be correlated with oxygen consum-
ption [83] but effects of cell number on the mRNA
expression of GLUT1 and G6PD have only been
indirectly described [42,66].
We observed that the majority of the analyzed
blastocysts were males. Similar skewed sex ratios have
been reported in previous studies [35,64,84–88]. The
higher number of male blastocysts in culture at defined
points has been attributed to their faster development in
glucose-containing culture media [86–89]. According
to Gutierrez-Adan et al. [90], the presence of serum in
the culture media also affects the sex and kinetics of the
blastocysts, favoring male development. The culture
media (SOFaaci) employed in this study were not
supplemented with glucose but included 5% bovine
serum, which contains non-defined levels of amino
acids, proteins, vitamins, growth factors and energy
substrates such as glucose [91,92]. Thus, the differential
response of the embryos to serum components might
have been responsible for the observed sex ratio.
Irrespectively of the skewed sex ratio observed for all
morphological qualities, the respiration rates were
similar for male and female blastocysts (Fig. 1). Inte-
restingly, Agung et al. [32] also found no differences in
the oxygen consumption between male and female
embryos of excellent quality, but observed that the
oxygen consumption of good quality female embryos
was higher than that of male embryos. Thus, it seems
that sexual dimorphism does not exist at the level of
oxidative phosphorylation, and oxygen consumption is
mostly associated with quality rather than with the sex
of the embryos.
On the contrary, expression of GLUT1 and G6PD
was influenced by the sex of embryos (Fig. 1). In the
present study, the DNA used for the sexing procedure
was extracted from the supernatant of the RNA isolation
buffer and no biopsy had to be performed, ruling out the
possibility of alterations in gene expression patterns
caused by the manipulation procedure. The lower levels
of GLUT1 in day 7 female blastocysts (Fig. 1) have not
previously been reported. This observation conflicts
with other reports, in which female embryos showed
higher glucose uptake [19,55]. Later in development,
Bertolini et al. [72] also observed that day 16 male
bovine in vivo-produced embryos had higher levels of
GLUT1 and GLUT3 expression than female in vivo-
produced embryos. Moreover, Morton et al. [93] found
that the levels of GLUT3 were lower in female embryos
as compared to male embryos. Potentially, the increased
level of expression of the GLUT1 gene in males is
associated with the faster development of male embryos
under in vitro conditions [84,85]. A faster development
could lead to an increase in glucose uptake, mediated by
GLUT1 expression. However, the increased metabolism
of faster growing embryos (males) was not significantly
reflected at the respiratory level, even though the
respiration rate of male embryos was 9% higher than
that of female embryos. In fact, if GLUT1 contributes to
glucose uptake and the mRNA level corresponds to
GLUT1 activity then we would only expect a 15%
difference in respiration rate between the sexes
(considering that glycolysis only contributes in 15%
for the overall oxygen consumption), even though the
GLUT1 transcript level is twice as large for males. Even
so, the contribution of GLUT1 to the glucose transport
from the external environment requires further inves-
tigation.

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236 233
The nearly 1.8-fold higher levels of G6PD observed
among female embryos produced in vitro (Fig. 1) was
expected and confirmed the delayed dosage com-
pensation of G6PD reported by others [62–64,94].
By inhibiting G6PD with dehydroepiandrosterone
(DHEA), Perez-Crespo et al. [65] observed similar
mRNA expression of G6PD between the two sexes and
did not detect a skewed sex ratio for in vitro-produced
embryos. Thus, the sexual dimorphism regarding the
G6PD transcript could also be responsible for the
skewed sex ratio observed in our study. It is likely that
the higher expression of G6PD leads to higher PPP
activity in females [95] and thus higher production of
NADPH and higher antioxidative capacity by female
embryos. This leads to a reduction in the amount of free
radical levels, which have a growth stimulating effect
[59] and thus conditions the subsequent development of
female embryos. Even so, it seems that the delayed
dosage compensation of G6PD and the higher activity
of the PPP in female embryos did not affect oxygen
consumption and thus oxidative phosphorylation.
In conclusion, the present study showed sexual
dimorphism in regards to the expression of both GLUT1
and G6PD transcripts, which was not observed in the
levels of oxygen consumed by individual bovine
embryos produced in vitro. Furthermore, the RA of
GLUT1 and G6PD transcripts correlate with each other
and with individual respiration rates, suggesting that
more metabolically active embryos may have higher
developmental competence. However, the present study
only investigated two gene transcripts, and thus only
provides a limited view of the involved complex
metabolic pathways. It is important to stress that
changes in transcriptional levels do not necessarily
reflect cellular and physiological alterations and there-
fore caution should be applied when interpreting
transcriptional profiles. Combining assessment of
embryonic respiration with genetic investigations using
microarray technology, which is capable of analyzing
the expression patterns of thousands of genes simulta-
neously, and with quantification of protein abundance
may allow for a better understanding of the involvement
of key genes in specific metabolic processes.
Acknowledgements
The authors thank Anette Pedersen, Klaus Villemoes
and Ruth Kristensen for their technical assistance. Novo
Nordisk Foundation financially supported the acquisi-
tion of the nanorespirometer technology and the Danish
Directorate for Food, Fisheries and Agri Business
(TFH03-8) the embryo work. Ana Sofia Lopes is
supported by a grant from Foundation for Science and
Technology (FCT), Ministry of Science and Technol-
ogy, Portugal (SFRH/BD/8486/2002). Dr. Per Horn is
thanked for critically revising the manuscript.
References
[1] Ziebe S, Petersen K, Lindenberg S, Andersen AG, Gabrielsen A,
Nyboe Andersen A. Embryo morphology or cleavage stage: how
to select the best embryos for transfer after in-vitro fertilization.
Hum Reprod 1997;12:1545–9.
[2] Merton S. Morphological evaluation of embryos in domestic
animals. In: Van Soom A, Boerjan M, editors. Assessment of
mammalian embryo quality: invasive and non-invasive techni-
ques. 1st ed, Dordrecht, The Netherlands: Kluwer Academic
Publishers; 2002. p. 31–55.
[3] Donnay I. Metabolic markers of embryo viability. In: Van Soom
A, Boerjan M, editors. Assessment of mammalian embryo
quality: invasive and non-invasive techniques. 1st ed, Dordrecht,
The Netherlands: Kluwer Academic Publishers; 2002. p. 57–94.
[4] Scott L. The biological basis of non-invasive strategies for
selection of human oocytes and embryos. Hum Reprod Update
2003;9:237–49.
[5] Bergh C, Aittomaki K, Loft A, Hazekamp J, Nygren KG.
Attitudes to single embryo transfer among Nordic IVF doctors.
Hum Reprod 2005;20:54. Abstract.
[6] Van Royen E, Mangelschots K, De Neubourg D, Laureys I,
Ryckaert G, Gerris J. Calculating the implantation potential of
day 3 embryos in women younger than 38 years of age: a new
model. Hum Reprod 2001;16:326–32.
[7] Boiso I, Veiga A, Edwards RG. Fundamentals of human embryo-
nic growth in vitro and the selection of high-quality embryos for
transfer. Reprod Biomed Online 2002;5:328–50.
[8] Corcoran D, Fair T, Lonergan P. Predicting embryo quality:
mRNA expression and the preimplantation embryo. Reprod
Biomed Online 2005;11:340–8.
[9] Overstrom EW. In vitro assessment of embryo viability. Ther-
iogenology 1996;45:3–16.
[10] Farin PW, Slenning BD, Britt JH. Estimates of pregnancy out-
comes based on selection of bovine embryos produced in vivo or
in vitro. Theriogenology 1999;52:659–70.
[11] Holm P, Shukri NN, Vajta G, Booth P, Bendixen C, Callesen H.
Developmental kinetics of the first cell cycles of bovine in vitro
produced embryos in relation to their in vitro viability and sex.
Theriogenology 1998;50:1285–99.
[12] Sakkas D, Shoukir Y, Chardonnens D, Bianchi PG, Campana A.
Early cleavage of human embryos to the two-cell stage after
intracytoplasmic sperm injection as an indicator of embryo
viability. Hum Reprod 1998;13:182–7.
[13] Lonergan P, Khatir H, Piumi F, Rieger D, Humblot P, Boland MP.
Effect of time interval from insemination to first cleavage on the
developmental characteristics, sex ratio and pregnancy rate after
transfer of bovine embryos. J Reprod Fertil 1999;117:159–67.
[14] Munne S, Wells D. Preimplantation genetic diagnosis. Curr Opin
Obstet Gynecol 2002;14:239–44.
[15] Gianaroli L, Magli M, Ferraretti A, Munne S. Preimplantation
diagnosis for aneuploidies in patients undergoing in vitro ferti-
lization with a poor prognosis: identification of the categories for
which it should be proposed. Fertil Steril 1999;72:837–44.
[16] Wrenzycki C, Herrmann D, Lucas-Hahn A, Korsawe K, Lemme
E, Niemann H. Messenger RNA expression patterns in bovine

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236234
embryos derived from in vitro procedures and their implications
for development. Reprod Fertil Dev 2005;17:23–35.
[17] Houghton FD, Hawkhead JA, Humpherson PG, Hogg JE, Balen
AH, Rutherford AJ, et al. Non-invasive amino acid turnover
predicts human embryo developmental capacity. Hum Reprod
2002;17:999–1005.
[18] Brison DR, Houghton F, Falconer D, Roberts SA, Hawkhead J,
Humpherson PG, et al. Identification of viable embryos in IVF
by non-invasive measurement of amino acid turnover. Hum
Reprod 2004;19:2319–24.
[19] Lane M, Gardner DK. Selection of viable mouse blastocysts
prior to transfer using a metabolic criterion. Hum Reprod
1996;11:1975–8.
[20] Donnay I, Feugang JM, Bernard S, Marchandise J, Pampfer S,
Moens A, et al. Impact of adding 5.5 mM glucose to SOF
medium on the development, metabolism and quality of in vitro
produced bovine embryos from the morula to the blastocyst
stage. Zygote 2002;10:189–99.
[21] Leese HJ. Non-invasive amino acid turnover predicts human
embryo developmental capacity. Hum Reprod 2002;17:999–
1005.
[22] Thompson JG. In vitro culture and embryo metabolism of cattle
and sheep embryos—a decade of achievement. Anim Reprod Sci
2000;60/61:263–75.
[23] Trimarchi JR, Liu L, Porterfield DM, Smith PJ, Keefe DL. A
non-invasive method for measuring preimplantation embryo
physiology. Zygote 2000;8:15–24.
[24] Rieger D, Loskutoff NM. Changes in the metabolism of glucose,
pyruvate, glutamine and glycine during maturation of cattle
oocytes in vitro. J Reprod Fertil 1994;100:257–62.
[25] Houghton FD, Thompson JG, Kennedy CJ, Leese HJ. Oxygen
consumption and energy metabolism of the early mouse embryo.
Mol Reprod Dev 1996;44:476–85.
[26] Houghton FD, Humpherson PG, Hawkhead JA, Hall CJ, Leese
HJ. Na+, K+, ATPase activity in the human and bovine pre-
implantation embryo. Dev Biol 2003;263:360–6.
[27] Houghton FD, Leese HJ. Metabolism and developmental com-
petence of the preimplantation embryo. Eur J Obstet Gynecol
Reprod Biol 2004;115:92–6.
[28] Thompson JG, Partridge RJ, Houghton FD, Cox CI, Leese HJ.
Oxygen uptake and carbohydrate metabolism by in vitro derived
bovine embryos. J Reprod Fertil 1996;106:299–306.
[29] Leese H. What does an embryo need? Hum Fertil 2003;6:180–5.
[30] Houghton FD. Energy metabolism of the inner cell mass and
trophectoderm of the mouse blastocyst. Differentiation 2006;74:
11–8.
[31] Benos DJ, Baladan RS. Energy metabolism of preimplantation
mammalian blastocysts. Am J Physol 1983;245:40–5.
[32] Agung B, Otoi T, Abe H, Hoshi H, Murakami M, Karja NW,
et al. Relationship between oxygen consumption and sex of
bovine in vitro fertilized embryos. Reprod Domest Anim
2005;40:51–6.
[33] Trimarchi JR, Liu L, Porterfield DM, Smith PJ, Keefe DL.
Oxidative phosphorylation-dependent and -independent oxygen
consumption by individual preimplantation mouse embryos.
Biol Reprod 2000;62:1866–74.
[34] Overstrom EW. Manipulation of early embryonic development.
Anim Reprod Sci 1992;28:277–85.
[35] Lopes AS, Larsen LH, Ramsing N, Løvendahl P, Raty M, Peippo
J, et al. Respiration rates of individual bovine in vitro-produced
embryos measured with a novel, non-invasive and rapid micro-
sensor system. Reproduction 2005;130:669–79.
[36] Lopes AS, Madsen SE, Ramsing N, Løvendahl P, Greve T,
Callesen H. Respiration of individual bovine in vivo and in
vitro-produced embryos: correlation with viability following
transfer. Hum Reprod 2006;22:558–66.
[37] Sturmey RG, Leese HJ. Energy metabolism in pig oocytes and
early embryos. Reproduction 2003;126:197–204.
[38] Abe H, Shiku H, Yokoo M, Aoyagi S, Moriyasu S, Minamihashi
A, et al. Evaluating the quality of individual embryos with a non-
invasive and highly sensitive measurement of oxygen consump-
tion by scanning electrochemical microscopy. J Reprod Dev
2006;52:55–64.
[39] Moriyasu S, Hirayama H, Sawai K, Kageyama S, Aoyagi S,
Shiku H, et al. Relationship between respiratory activity and the
pregnancy rate of bisected bovine embryos in vivo. Reprod Fertil
Dev 2007;19:219. Abstract.
[40] Sakagami N, Akiyama K, Nakazawa Y. The relationship
between oxygen consumption rate and pregnancy rate of bovine
embryos. Reprod Fertil Dev 2007;19:225. Abstract.
[41] Harvey AJ, Kind KL, Thompson JG. REDOX regulation of early
embryo development. Reproduction 2002;123:479–86.
[42] Harvey AJ, Kind KL, Pantaleon M, Armstrong DT, Thompson
JG. Oxygen-regulated gene expression in bovine blastocysts.
Biol Reprod 2004;71:1108–19.
[43] Leese HJ. Metabolism of the preimplantation mammalian
embryo. Oxf Rev Reprod Biol 1991;13:35–72.
[44] Abe H, Shiku H, Aoyagi S, Matsue T, Hoshi H. Respiration
activity of bovine embryos cultured in serum-free and serum-
containing media. Reprod Fertil Dev 2005;17:215. Abstract.
[45] Eckert J, Pugh PA, Thompson JG, Niemann H, Tervit HR.
Exogenous protein affects development competence and meta-
bolic activity of bovine pre-implantation embryos in vitro.
Reprod Fertil Dev 1998;10:327–32.
[46] Manser RC, Leese HJ, Houghton FD. Effect of inhibiting nitric
oxide production on mouse preimplantation embryo develop-
ment and metabolism. Biol Reprod 2004;71:528–33.
[47] Hooper K, Lane M, Gardner DK. Reduced oxygen concentration
increases mouse embryo development and oxidative metabo-
lism. Theriogenology 2001;55:334. Abstract.
[48] Abe H, Hoshi H. Evaluation of bovine embryos produced in high
performance serum-free media. J Reprod Dev 2003;49:193–202.
[49] Magnusson C, Hillensjo T, Hamberger L, Nilsson L. Oxygen
consumption by human oocytes and blastocysts grown in vitro.
Hum Reprod 1986;1:183–4.
[50] Shiku H, Shiraishi T, Ohya H, Matsue T, Abe H, Hoshi H, et al.
Oxygen consumption of single bovine embryos probed by
scanning electrochemical microscopy. Anal Chem 2001;73:
3751–8.
[51] Thompson JG, Partridge RJ, Houghton FD, Kennedy CJ, Pullar
D, Wrathall AE, et al. Preliminary observations of the uptake of
oxygen by day 7 bovine blastocysts. Theriogenology 1995;43:
337. Abstract.
[52] Donnay I, Leese HJ. Embryo metabolism during the expansion
of the bovine blastocyst. Mol Reprod Dev 1999;53:171–8.
[53] Leese HJ. Metabolic control during preimplantation mammalian
development. Hum Reprod Update 1995;1:63–72.
[54] Rieger D, Loskutoff NM, Betteridge KJ. Developmentally
related changes in the metabolism of glucose and glutamine
by cattle embryos produced and co-cultured in vitro. J Reprod
Fertil 1992;95:585–95.
[55] Gardner DK, Leese HJ. Assessment of embryo viability prior to
transfer by the noninvasive measurement of glucose uptake. J
Exp Zool 1987;242:103–5.

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236 235
[56] Rieger D. The measurement of metabolic activity as an approach
to evaluating viability and diagnosing sex in early embryos.
Theriogenology 1984;21:138–49.
[57] Renard JP, Philippon A, Menezo Y. J In-vitro uptake of glucose
by bovine blastocysts. J Reprod Fertil 1980;58:161–4.
[58] Shimizu N, Shimizu Y, Kondo I, Woods C, Wegner T. The bovine
genes for phosphoglycerate kinase, glucose-6-phosphate dehy-
drogenase, alpha-galactosidase, and hypoxanthine phosphoribo-
syltransferase are linked to the X chromosome in cattle-mouse
cell hybrids. Cytogenet Cell Genet 1981;29:26–31.
[59] Rieger D. Relationships between energy metabolism and devel-
opment of early mammalian embryos. Theriogenology 1992;37:
75–93.
[60] Peippo J, Bredbacka P. Sex-related growth rate differences in
mouse preimplantation embryos in vivo and in vitro. Mol Reprod
Dev 1995;40:56–61.
[61] Nicol CJ, Zielenski J, Tsui LC, Wells PG. An embryoprotective
role for glucose-6-phosphate dehydrogenase in developmental
oxidative stress and chemical teratogenesis. FASEB J 2000;14:
111–27.
[62] Gutierrez-Adan A, Oter M, Martinez-Madrid B, Pintado B, De
La Fuente J. Differential expression of two genes located on the
X chromosome between male and female in vitro-produced
bovine embryos at the blastocyst stage. Mol Reprod Dev
2000;55:146–51.
[63] Peippo J, Farazmand A, Kurkilahti M, Markkula M, Basrur PK,
King WA. Sex-chromosome linked gene expression in in-vitro
produced bovine embryos. Mol Reprod Dev 2002;8:923–9.
[64] Wrenzycki C, Lucas-Hahn A, Herrmann D, Lemme E, Korsawe
K, Niemann H. In vitro production and nuclear transfer affect
dosage compensation of the X-linked gene transcripts G6PD,
PGK, and Xist in preimplantation bovine embryos. Biol Reprod
2002;66:127–34.
[65] Perez-Crespo M, Ramirez MA, Fernandez-Gonzalez R, Rizos D,
Lonergan P, Pintado B, et al. Differential sensitivity of male and
female mouse embryos to oxidative induced heat-stress is
mediated by glucose-6-phosphate dehydrogenase gene expres-
sion. Mol Reprod Dev 2005;72:502–10.
[66] Wrenzycki C, Herrmann D, Niemann H. Timing of blastocyst
expansion affects spatial messenger RNA expression patterns of
genes in bovine blastocysts produced in vitro. Biol Reprod
2003;68:2073–80.
[67] Wrenzycki C, Herrmann D, Carnwath JW, Niemann H. Altera-
tions in the relative abundance of gene transcripts in preimplan-
tation bovine embryos cultured in medium supplemented with
either serum or PVA. Mol Reprod Dev 1999;53:8–18.
[68] Leppens-Luisier G, Urner F, Sakkas D. Facilitated glucose
transporters play a crucial role throughout mouse preimplanta-
tion embryo development. Hum Reprod 2001;16:1229–36.
[69] Kind KL, Collett RA, Harvey AJ, Thompson JG. Oxygen-
regulated expression of GLUT-1, GLUT-3, and VEGF in the
mouse blastocyst. Mol Reprod Dev 2004;70:37–44.
[70] Wrenzycki C, Herrmann D, Keskintepe L, Martins Jr A, Sir-
isathien S, Brackett B, et al. Effects of culture system and protein
supplementation on mRNA expression in pre-implantation
bovine embryos. Hum Reprod 2001;16:893–901.
[71] Kimura K, Spate LD, Green MP, Roberts RM. Effects of D-
glucose concentration, D-fructose, and inhibitors of enzymes of
the pentose phosphate pathway on the development and sex ratio
of bovine blastocysts. Mol Reprod Dev 2005;72:201–7.
[72] Bertolini M, Beam SW, Shim H, Bertolini LR, Moyer AL,
Famula TR, et al. Growth, development, and gene expression
by in vivo- and in vitro-produced day 7 and 16 bovine embryos.
Mol Reprod Dev 2002;63:318–28.
[73] Knijn HM, Wrenzycki C, Hendriksen PJ, Vos PL, Herrmann D,
van der Weijden GC, et al. Effects of oocyte maturation regimen
on the relative abundance of gene transcripts in bovine blas-
tocysts derived in vitro or in vivo. Reproduction 2002;124:365–
75.
[74] Lopes AS, Wrenzycki C, Ramsing NB, Herrmann D, Niemann
H, Greve T, et al. Relationship between respiration rates and
mRNA expression of G6PD and Glut-1 genes in individual
blastocysts produced in vitro. Hum Reprod 2006;21:48–9.
Abstract.
[75] Holm P, Booth PJ, Schmidt MH, Greve T, Callesen H. High
bovine blastocyst development in a static in vitro production
system using SOFaa medium supplemented with sodium citrate
and myo-inositol with or without serum-proteins. Theriogenol-
ogy 1999;52:683–700.
[76] Parrish JJ, Susko-Parrish JL, Liebfried-Rutledge ML, Critser ES,
Eyestn WH, First NL. Bovine in vitro fertilization with frozen–
thawed semen. Theriogenology 1986;25:591–600.
[77] SAS Institute. SAS/STAT user’s guide (release 8.2). Cary, NC:
Statistical Analysis System Institute Inc.; 1999.
[78] Box G, Hunter WG, Hunter JS. Statistics for experimenters, 1st
ed, New York: John Wiley & Sons; 1978. p. 150.
[79] Pantaleon M, Kaye PL. Glucose transporters in preimplantation
development. Rev Reprod 1998;3:77–81.
[80] Khurana NK, Niemann H. Energy metabolism in preimplanta-
tion bovine embryos derived in vitro or in vivo. Biol Reprod
2000;62:847–56.
[81] Augustin R, Pocar P, Navarrete-Santos A, Wrenzycki C, Gan-
dolfi F, Niemann H, et al. Glucose transporter expression is
developmentally regulated in in vitro derived bovine preimplan-
tation embryos. Mol Reprod Dev 2001;60:370–6.
[82] Filosa S, Fico A, Paglialunga F, Balestrieri M, Crooke A, Verde
P, et al. Failure to increase glucose consumption through the
pentose-phosphate pathway results in the death of glucose-6-
phosphate dehydrogenase gene-deleted mouse embryonic stem
cells subjected to oxidative stress. Biochem J 2003;370:935–43.
[83] Kaidi S, Bernard S, Lambert P, Massip A, Dessy F, Donnay I.
Effect of conventional controlled-rate freezing and vitrification
on morphology and metabolism of bovine blastocysts produced
in vitro. Biol Reprod 2001;65:1127–34.
[84] Avery B, Madison V, Greve T. Sex and development in bovine in-
vitro fertilized embryos. Theriogenology 1991;35:953–63.
[85] Xu KP, Yadav BR, King WA, Betteridge KJ. Sex-related differ-
ences in developmental rates of bovine embryos produced and
cultured in vitro. Mol Reprod Dev 1992;31:249–52.
[86] Bredbacka K, Bredbacka P. Glucose controls sex-related growth
rate differences of bovine embryos produced in vitro. J Reprod
Fertil 1996;106:169–72.
[87] Gutierrez-Adan A, Granados J, Pintado B, De La Fuente J.
Influence of glucose on the sex ratio of bovine IVM-IVF
embryos cultured in vitro. In: Proceedings of the 14th scientific
meeting, AETE; 1998. p. 166. Abstract.
[88] Larson MA, Kimura K, Kubisch HM, Roberts RM. Sexual
dimorphism among bovine embryos in their ability to make
the transition to expanded blastocyst and in the expression of the
signaling molecule IFN-tau. Proc Natl Acad Sci USA
2001;98:9677–82.
[89] Gutierrez-Adan A, Granados J, Pintado B, De La Fuente J.
Influence of glucose on the sex ratio of bovine IVM/IVF
embryos cultured in vitro. Reprod Fertil Dev 2001;13:361–5.

A.S. Lopes et al. / Theriogenology 68 (2007) 223–236236
[90] Gutierrez-Adan A, Lonergan P, Rizos D, Ward FA, Boland MP,
Pintado B, et al. Effect of the in vitro culture system on the
kinetics of blastocyst development and sex ratio of bovine
embryos. Theriogenology 2001;55:1117–26.
[91] Kane MT. A low molecular weight extract of bovine serum
albumin stimulates rabbit blastocyst cell division and expansion
in vitro. J Reprod Fertil 1985;73:147–50.
[92] Maurer HR. Towards serum-free, chemically defined media for
mammalian cell culture. In: Freshney RI, editor. Animal cell
culture: a practical approach. 2nd ed, Oxford, UK: Oxford
University Press; 1992. p. 15–46.
[93] Morton KM, Herrmann D, Sieg B, Struckmann C, Maxwell
WMC, Rath D, et al. Altered mRNA expression patterns in
bovine blastocysts after fertilization in vitro using flow-cyto-
metrically sex-sorted sperm. Mol Reprod Dev 2007;74:931–
40.
[94] Lonergan P, Gutierrez-Adan A, Pintado B, Fair T, Ward F,
Fuente JD, et al. Relationship between time of first cleavage
and the expression of IGF-I growth factor, its receptor, and
two housekeeping genes in bovine two-cell embryos and
blastocysts produced in vitro. Mol Reprod Dev 2000;57:
146–52.
[95] Tiffin GJ, Rieger D, Betteridge KJ, Yadav BR, King WA.
Glucose and glutamine metabolism in pre-attachment cattle
embryos in relation to sex and stage of development. J Reprod
Fertil 1991;93:125–32.
Top Related
Copyright © 2022 FDOKUMEN

