






Bahasa
Halaman
Hukum


Physical interaction between a novel domain of thereceptor Notch and the transcription factor RBP-JK/Su(H)
Kumiko Tamura, Yoshihito Taniguchi, Shigeru Minoguchi, Takashi Sakai,Tin Tun, Takahisa Furukawa and Tasuku Honjo
Department of Medical Chemistry, Kyoto University Faculty of Medicine, Yoshida, Sakyo-ku, Kyoto 606, Japan
Background: The mammalian transcription factor RBP-JK binds to the DNA sequence motif CGTGGGAA andis involved in the regulation of gene expression; forexample, it plays a part in the transactivation of viral andcellular genes by Epstein-Barr virus nuclear antigen-2.The Drosophila homologue of RBP-JK is the product ofthe Suppressor of Hairless (Su(H)) gene. Su(H) is a neuro-genic gene that acts downstream of Notch, which encodesa cell-surface receptor. Furthermore, in the mouse, thephenotypes of homozygous mutant Notch l embryos arevery similar to those of homozygous mutant RBP-JKembryos. Recent studies, using the yeast two-hybrid sys-tem, have led to the suggestion that the CDC10/ankyrin-like repeats of the Drosophila Notch protein interact withthe Su(H) protein.Results: We searched for proteins that interact withmouse RBP-JK using the yeast two-hybrid system, and inthis way identified a short intracellular region (mRAM23)
of the mouse Notchl protein that lacks any knownsequence motif. In vitro interaction studies, using proteinsfused to glutathione-S-transferase, showed that RBP-JKand Su(H) bind directly to the RAM23 regions of mouseNotchl and Drosophila Notch, respectively. Immuno-precipitation analysis showed that RBP-JK and themRAM23 region of mouse Notchl also interact in vivo.Further studies, including site-directed mutagenesisexperiments, narrowed down the region of mouseNotchl that interacts with RBP-JK. The results indicatethat this region is less than 50 amino-acid residues inlength, and lies immediately downstream of the trans-membrane region.Conclusions: We show that the transcription factorRBP-JK/Su(H) interacts directly with a novel intracellulardomain of the cell-surface receptor Notch. RBP-JK/Su(H) does not appear to interact with Notch via theCDC10/ankyrin repeats implicated in previous studies.
Current Biology 1995, 5:1416-1423
Background
Studies of a large number of Drosophila Notch (Notch)mutants have shown that Notch is involved in cell-fatedetermination in various lineages, including neuronalprecursors, myoblasts and Malpighian tubules [1,2]. TheNotch protein is a cell-surface receptor with several dis-tinct domains. The extracellular domain contains 36 tan-dem repeats of a 40 amino-acid, cysteine-rich motifhomologous to epidermal growth factor (EGF), and threecopies of another cysteine-rich region motif called theLNR motif. The intracellular domain of Notch containssix copies of the 33-residue CDC10/ankyrin-like repeat.The general structure of the Notch receptor, includingthese three motifs, is highly conserved through evolution.
A series of mutations of residues in the EGF-like repeatregion of Notch have been shown to affect binding ofthe Notch receptor to the ligand encoded by the Deltagene. The precise physiological role of the LNR motif isnot clear. Although CDC10/ankyrin-like repeats areknown to be involved in the protein-protein interactionsof cytoskeletal proteins and transcription factors [3,4], theexact role of the CDC10/ankyrin-like motif of Notchremains to be investigated, as the signal transductionpathway downstream of Notch has not been elucidated.
Drosophila Notch is a member of a family of neurogenicgenes, which include Delta, Enhancer of split, master mind,Hairless and Suppressor of Hairless (Su(H)). Genetic crosseshave indicated that Su(H) has a close genetic interactionwith Notch [1,2]. In most Schneider (S2) cells overex-pressing both Notch and Su(H), Su(H) was found in thecytoplasm, but when Su(H) was expressed alone, itwas nuclear in most cells [5]. Deletion of the CDC10/ankyrin-like repeats from Notch reduced the frequencywith which Su(H) was cytoplasmically localized incotransfected S2 cells. These studies suggested that Notchand Su(H) interact through the CDC10/ankyrin-likerepeats of Notch, although no direct evidence for theirphysical interaction has been provided.
We have previously isolated a 60 kDa nuclear protein,known as RBP-JK, from a nuclear extract of a mouse pre-B cell line [6]. RBP-JK recognizes the DNA sequencemotif CGTGGGAA [7], and is highly conserved throughevolution from Drosophila to humans [8,9]. Studies ofmutant RBP-JK proteins have shown that the two regionsimmediately upstream and downstream of the integrase-related motif in the middle of the protein are criticalfor its DNA-binding activity, and that RBP-JK bindsto DNA as a monomer [10]. To understand the functionof RBP-JK, we and others have isolated its Drosophila
Correspondence to: Tasuku Honjo. E-mail address: [email protected]
© Current Biology 1995, Vol 5 No 121416

Interaction between a novel domain of Notch and RBP-JK/Su(H) Tamura et al.
homologue, and shown that the Drosophila RBP-JK geneis identical to Su(H) [11,12]. RBP-JK is involved in thetranscriptional regulation of a number of cellular and viralgenes: for example, it mediates the repression of the ade-novirus pIX gene promoter and of other promoters [13].RBP-JK is also essential for the transactivation of viral andcellular genes by Epstein-Barr virus nuclear antigen-2(EBNA-2), which plays a crucial role in human B-cellimmortalization by the virus. Unlike most transcriptionfactors, EBNA-2 does not bind directly to its cis-respon-sive DNA element but binds to RBP-JK, which doesbind directly to DNA [14-18]. Su(H) is also involved intranscriptional regulation in Drosophila [19].
Three mouse homologues of Notch - mNotch 1, mNotch2and mNotch3 - have already been cloned [20-22]. Thehuman Notch I gene (Tan- ; [23]) and the mouse int-3gene [24] were cloned as oncogenes. These studies indi-cated that the deregulated expression of the cytoplasmicdomain of the truncated Notch protein contributes to thetumorigenic phenotype [23,24]. Homozygous mutantmNotch 1 (mNotch 1-/-) mice showed growth retardation ataround 9.5 days post-coitum (dpc), and died before11.5 dpc with widespread cell death [25,26]; the micealso showed defects both in somite development and inneural tube formation. Intriguingly, these phenotypes arevery similar to those found in homozygous mutant RBP-JK (RBP-JK-/ -) mice, although the onset of the pheno-type and the lethality phase of RBP-JK-/ - mice areslightly earlier than in mNotch 1-/ - mice [27]. The pheno-typic similarity between RBP-JK-/- and mNotch 1-/ - micesuggests that both genes may be involved in the same,conserved developmental pathway.
To explore the molecular mechanisms whereby RBP-JKregulates mouse development, we searched for proteinsthat interact with RBP-JK using the yeast to-hybridsystem. As we report here, in this way we isolated a
DNA sequence encoding a short intracellular region ofmNotchl, which did not contain the CDC10/ankyrin-like repeat. Further studies indicated that a small region,of about 50 amino-acid residues, immediately down-stream of the transmembrane region of mNotch, isresponsible for its interaction with RBP-JK.
Results and discussion
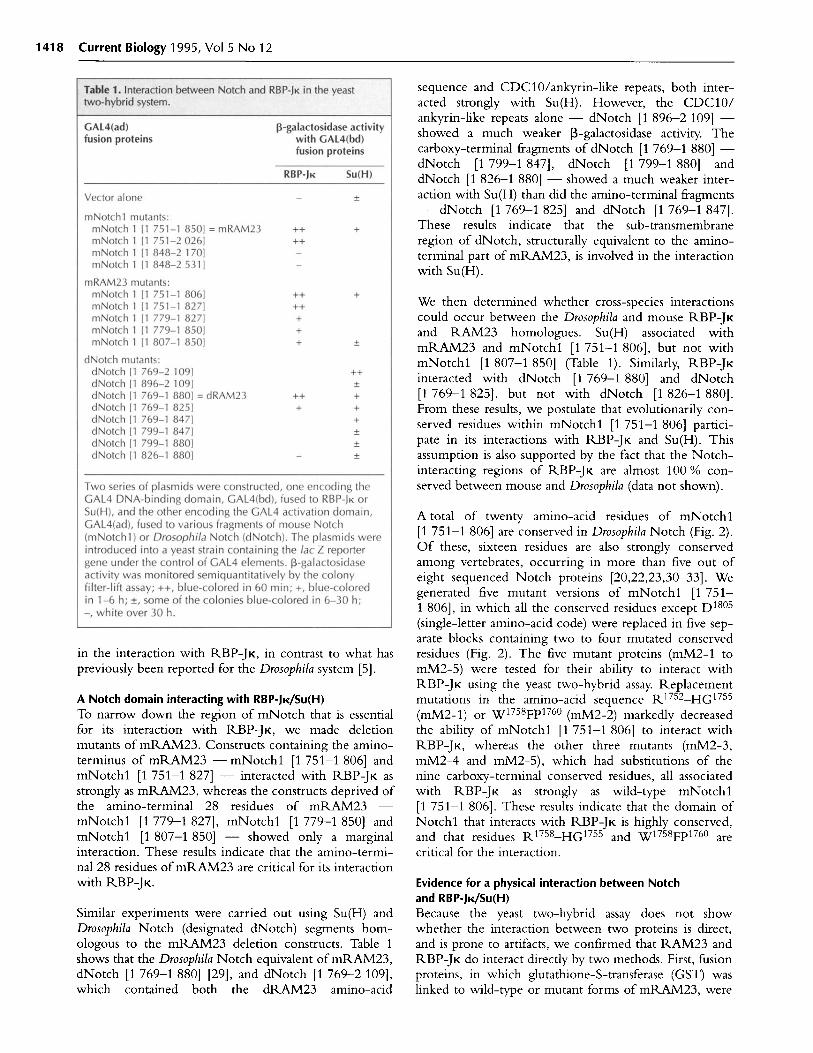
RBP-JK and mNotchl interact specifically in the yeasttwo-hybrid systemWe used the yeast two-hybrid system to screen for pro-teins that interact with RBP-JK. The 'bait' plasmid,which was constructed by fusing the DNA sequenceencoding RBP-JK to that encoding the DNA-bindingprotein LexA, was used to screen a cDNA library, pre-pared from a 9.5-day-old mouse embryo, which directsthe synthesis of activator-tagged mouse proteins in yeast[28]. After screening one million transformants by theP-galactosidase colony filter-lift assay, we isolated fiveplasmids, four of which encoded previously uncharacter-ized proteins and will be described elsewhere. One of thecDNA clones we isolated turned out to encode a seg-ment of mNotchl, residues 1 751-1 850, just down-stream of the transmembrane region, which we havedesignated mRAM23 (Fig. la) [20]. It should be notedthat mRAM23 does not contain any CDC10/ankyrin-like repeats.
We tested whether other truncated mNotchl proteins(Fig. la) could interact with RBP-JK in the yeast two-hybrid system. Table 1 shows that RBP-JK interactedwith mNotchl [1 751-1 850] and mNotchl [1 751-2 026], but not with mNotchl [1 848-2 170] ormNotchl [1 848-2 531], neither of which containedmRAM23 residues. These results suggest that theCDC10/ankyrin-like repeat of mNotchl is not involved
Fig. 1. (a) Schematic representation ofthe carboxy-terminal region of Notch.Numbers represent amino-acid positionsof mouse Notch (mNotchl), and those ofDrosophila Notch (dNotch) are shown inparentheses. Bars represent regions ofmNotchl and dNotch deletion mutantswhich are used in two-hybrid and/orGST assays. (b) COS 7 cell expressionconstructs of mNotchl mutants. Myc,Myc-epitope tag; mNotchl-RAMIC,mNotchl [1 747-2 531]; mNotchl-IC,mNotchl [1 810-2 531].
RESEARCH PAPER 1417
1418 Current Biology 1995, Vol 5 No 12
sequence and CDC10/ankyrin-like repeats, both inter-acted strongly with Su(H). However, the CDC10/ankyrin-like repeats alone - dNotch [1 896-2 109] -showed a much weaker -galactosidase activity. Thecarboxy-terminal fragments of dNotch [1 769-1 880] -dNotch [1 799-1 847], dNotch [1 799-1 880] anddNotch [1 826-1 880] - showed a much weaker inter-action with Su(H) than did the amino-terminal fragments- dNotch [1 769-1 825] and dNotch [1 769-1 847].These results indicate that the sub-transmembraneregion of dNotch, structurally equivalent to the amino-terminal part of mRAM23, is involved in the interactionwith Su(H).
We then determined whether cross-species interactionscould occur between the Drosophila and mouse RBP-JKand RAM23 homologues. Su(H) associated withmRAM23 and mNotchl [1 751-1 806], but not withmNotchl [1 807-1 850] (Table 1). Similarly, RBP-JKinteracted with dNotch [1 769-1 880] and dNotch[1 769-1 825], but not with dNotch [1 826-1 880].From these results, we postulate that evolutionarily con-served residues within mNotchl [1 751-1 806] partici-pate in its interactions with RBP-JK and Su(H). Thisassumption is also supported by the fact that the Notch-interacting regions of RBP-JK are almost 100 % con-served between mouse and Drosophila (data not shown).
in the interaction with RBP-JK, in contrast to what haspreviously been reported for the Drosophila system [5].
A Notch domain interacting with RBP-JK/Su(H)To narrow down the region of mNotch that is essentialfor its interaction with RBP-JK, we made deletionmutants of mRAM23. Constructs containing the amino-terminus of mRAM23 - mNotchl [1 751-1 806] andmNotchl [1 751-1 827] - interacted with RBP-JK asstrongly as mRAM23, whereas the constructs deprived ofthe amino-terminal 28 residues of mRAM23 -mNotchl [1 779-1 827], mNotchl [1 779-1 850] andmNotchl [1 807-1 850] - showed only a marginalinteraction. These results indicate that the amino-termi-nal 28 residues of mRAM23 are critical for its interactionwith RBP-JK.
Similar experiments were carried out using Su(H) andDrosophila Notch (designated dNotch) segments hom-ologous to the mRAM23 deletion constructs. Table 1shows that the Drosophila Notch equivalent of mRAM23,dNotch [1 769-1 880] [29], and dNotch [1 769-2 109],which contained both the dRAM23 amino-acid
A total of twenty amino-acid residues of mNotchl[1 751-1 806] are conserved in Drosophila Notch (Fig. 2).Of these, sixteen residues are also strongly conservedamong vertebrates, occurring in more than five out ofeight sequenced Notch proteins [20,22,23,30-33]. Wegenerated five mutant versions of mNotchl [1 751-1 806], in which all the conserved residues except D180 5
(single-letter amino-acid code) were replaced in five sep-arate blocks containing two to four mutated conservedresidues (Fig. 2). The five mutant proteins (mM2-1 tomM2-5) were tested for their ability to interact withRBP-JK using the yeast two-hybrid assay. Replacementmutations in the amino-acid sequence R17 52-HG17 5 5
(mM2-1) or W17 58 Fp 7 6 0 (mM2-2) markedly decreasedthe ability of mNotchl [1 751-1 806] to interact withRBP-JK, whereas the other three mutants (mM2-3,mM2-4 and mM2-5), which had substitutions of thenine carboxy-terminal conserved residues, all associatedwith RBP-JK as strongly as wild-type mNotchl[1 751-1 806]. These results indicate that the domain ofNotchl that interacts with RBP-JK is highly conserved,and that residues R1 75 8-HG 17 5 5 and W17 58Fp 17 60 arecritical for the interaction.
Evidence for a physical interaction between Notchand RBP-JK/Su(H)Because the yeast two-hybrid assay does not showwhether the interaction between two proteins is direct,and is prone to artifacts, we confirmed that RAM23 andRBP-JK do interact directly by two methods. First, fusionproteins, in which glutathione-S-transferase (GST) waslinked to wild-type or mutant forms of mRAM23, were

Interaction between a novel domain of Notch and RBP-JK/Su(H) Tamura et al.
Fig. 2. The effect of replacement mutations on the interaction between mNotchl and RBP-JK in the yeast two-hybrid system. Theamino-acid sequence of mNotchl [1 751-1 8061 and other characterized Notch sequences [20-23,29-33] are aligned. Strongly con-served amino acids are shadowed. The amino acids that the conserved residues are changed to in each mutant are shown above thealigned sequences. The altered amino acids, shown in bold, followed by the corresponding DNA sequence for each mutant residue, areas follows: mM2-1, A(GCG)/Q/D(GAT)/C(TGC); mM2-2, L(TTG)/A(GCC)/A(GCT); mM2-3, A(GCG)/A(GCT)/A(GCA); mM2-4,A(GCG)/A(GCA)/E/A(GCC)/S(TCG)/V(GTC); mM2-5, A(GCC)/A(GCG)/P/L/K/N/T(ACC). The results of interaction between RBP-JK andthe mNotchl replacement mutants are shown above (highlighted in yellow). Construction of the plasmids encoding the GAL4 fusionproteins and the criteria for the assessment of 3-galactosidase activity are as described in Table 1. The numbers indicate amino-acidpositions in the mNotchl and Drosophila Notch sequences.
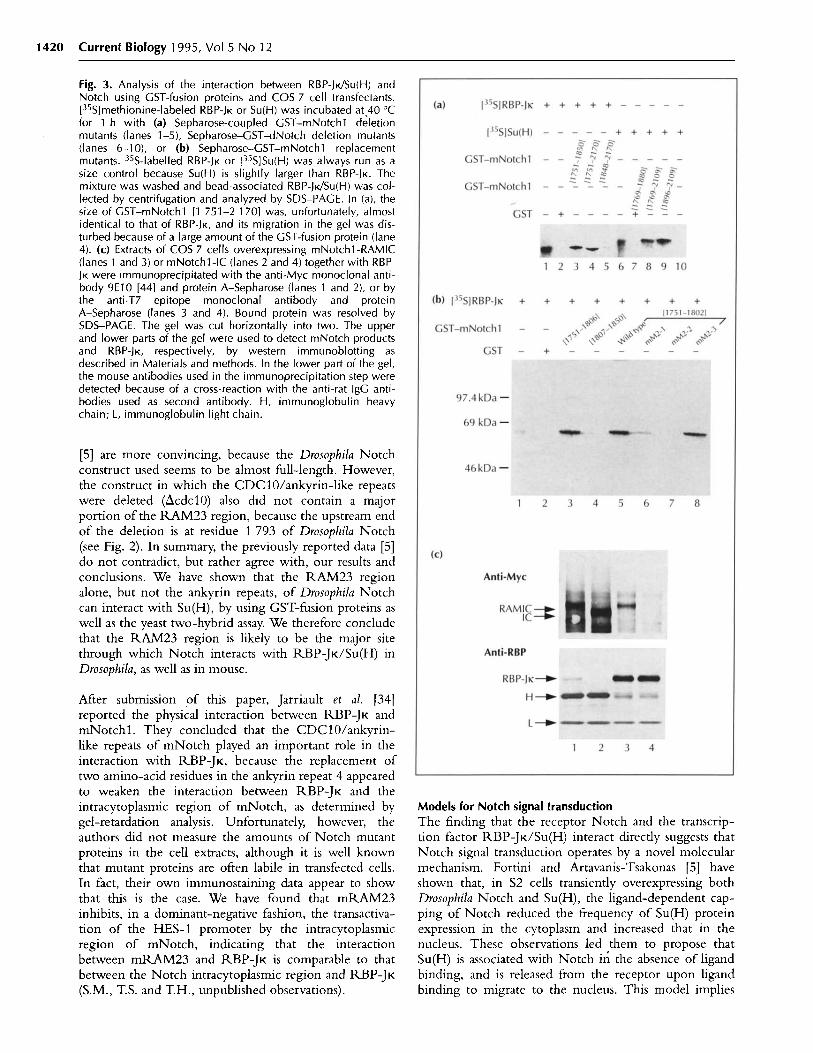
synthesized in bacteria. In order to remove 5' endsequences from the mRAM23 clone that was originallyisolated by the yeast two-hybrid assay, the mRAM23cDNA and its mutant versions were synthesized by thepolymerase chain reaction (PCR) and inserted into aGST-fusion vector. GST-fusion proteins were immobi-lized on glutathione-Sepharose beads and tested for theirability to bind to 35 S-labeled RBP-JK (Fig. 3a, lanes 1-5).
In agreement with the results from the yeast two-hybridassay, RBP-JK was specifically retained on beads coupledto GST-mRAM23, GST-mNotchl [1 751-1 850] andGST-mNotchl [1 751-2 170], but not on beads linkedto GST alone or GST-mNotchl [1 848-2 170]. Wedetected a marginal association between RBP-JK andGST-mNotchl [1 807-1 850]. As a negative' control,35 S-labeled luciferase was not retained on beads coupledto GST-mRAM23 (data not shown). Results with GST-fusion proteins containing either wild-type mNotchl[1 751-1 806], or the mutant forms mM2-1, mM2-2 ormM2-3, also supported the conclusions from the yeasttwo-hybrid assay (Fig. 3b). The mM2-2 mutant com-pletely failed to bind to RBP-JK, and mM2-1 interactedonly weakly with RBP-JK; mM2-3, however, interactedwith RBP-JK as strongly as wild-type mRAM23.The carboxy-terminal region of mRAM23 (mNotchl[1 807-1 850]) was inactive in the binding assay.
We further confirmed that the Drosophila homologues,dRAM23 and Su(H), also interact directly by using GST-fusion proteins (Fig. 3a; lanes 6-10). GST-dRAM23 andGST-dNotch [1 769-2 109] interacted strongly withSu(H). However, dNotch [1 896-2 109], which con-tained the CDC10/ankyrin-like repeats but not thedRAM23 region, did not interact with Su(H) at all, inagreement with the yeast two-hybrid assay.
Finally, we tested whether the intracellular region ofmNotchl can interact with RBP-JK in mammalian
cells. An expression plasmid encoding either the entirecytoplasmic region of mNotchl (mNotchl-RAMIC)or its mRAM23-truncated construct (mNotchl-IC),fused to six copies of a Myc-epitope tag (Fig. lb), wascoexpressed in COS 7 cells with RBP-JK tagged withthe bacteriophage T7 gene 10 product. Cell extracts wereincubated with a monoclonal antibody directed againstthe T7- or the Myc-epitope tag. The anti-Myc antibodyco-immunoprecipitated RBP-JK and the mNotchl-RAMIC protein (Fig. 3c). Conversely, the anti-T7 anti-body co-immunoprecipitated mNotchl-RAMIC andRBP-JK. In contrast, mNotchl-IC did not interact withRBP-JK at all. The results clearly indicate that theCDC10/ankyrin-like repeats of mNotchl are unable tointeract with RBP-JK in COS 7 cells. Furthermore, insitu chemical crosslinking was found to augment complexformation between mNotch and RBP-JK in the samecotransfected COS 7 cells (data not shown), indicatingthat the interaction did take place in the cell. Theseresults, taken together the yeast two-hybrid and GST-fusion protein data, indicate that RBP-JK interacts withthe amino-terminal region of mRAM23, but not withthe CDC10/ankyrin-like repeat of mNotchl.
Is Drosophila Notch different from mouse Notch?The CDC10/ankyrin-like repeats of Drosophila Notchwere previously reported to interact with Su(H), on thebasis of experiments using CDC10-deletion constructs inthe yeast two-hybrid assay and in S2 cell transfectants [5].However, the construct used as a positive control in theyeast assay did not contain the RAM23 region. Deletionof the sequence encoding the CDC10/ankyrin-likerepeats from this construct did reduce its interaction withSu(H), but the low level of interaction detected with thepositive control could have been a result of non-specificweak interactions between the CDC10/ankyrin-likerepeats and yeast proteins. In fact, the CDC10/ankyrin-like repeats alone did not interact with Su(H) in theseexperiments [5]. The S2 cell cotransfection experiments
RESEARCH PAPER 1419
1420 Current Biology 1995, Vol 5 No 12
Fig. 3. Analysis of the interaction between RBP-JK/Su(H) andNotch using GST-fusion proteins and COS 7 cell transfectants.[35S]methionine-labeled RBP-JK or Su(H) was incubated at 40 °Cfor 1 h with (a) Sepharose-coupled GST-mNotchl deletionmutants (lanes 1-5), Sepharose-GST-dNotch deletion mutants(lanes 6-10), or (b) Sepharose-GST-mNotchl replacementmutants. 35S-labelled RBP-JK or [35S]Su(H) was always run as asize control because Su(H) is slightly larger than RBP-JK. Themixture was washed and bead-associated RBP-JK/Su(H) was col-lected by centrifugation and analyzed by SDS-PAGE. In (a), thesize of GST-mNotchl [1 751-2 170] was, unfortunately, almostidentical to that of RBP-JK, and its migration in the gel was dis-turbed because of a large amount of the GST-fusion protein (lane4). (c) Extracts of COS 7 cells overexpressing mNotchl-RAMIC(lanes 1 and 3) or mNotchl -IC (lanes 2 and 4) together with RBP-JK were immunoprecipitated with the anti-Myc monoclonal anti-body 9E10 [44] and protein A-Sepharose (lanes 1 and 2), or bythe anti-T7 epitope monoclonal antibody and proteinA-Sepharose (lanes 3 and 4). Bound protein was resolved bySDS-PAGE. The gel was cut horizontally into two. The upperand lower parts of the gel were used to detect mNotch productsand RBP-JK, respectively, by western immunoblotting asdescribed in Materials and methods. In the lower part of the gel,the mouse antibodies used in the immunoprecipitation step weredetected because of a cross-reaction with the anti-rat IgG anti-bodies used as second antibody. H, immunoglobulin heavychain; L, immunoglobulin light chain.
[5] are more convincing, because the Drosophila Notchconstruct used seems to be almost full-length. However,the construct in which the CDC10/ankyrin-like repeatswere deleted (AcdclO) also did not contain a majorportion of the RAM23 region, because the upstream endof the deletion is at residue 1 793 of Drosophila Notch(see Fig. 2). In summary, the previously reported data [5]do not contradict, but rather agree with, our results andconclusions. We have shown that the RAM23 regionalone, but not the ankyrin repeats, of Drosophila Notchcan interact with Su(H), by using GST-fusion proteins aswell as the yeast two-hybrid assay. We therefore concludethat the RAM23 region is likely to be the major sitethrough which Notch interacts with RBP-JK/Su(H) inDrosophila, as well as in mouse.
After submission of this paper, Jarriault et al. [34]reported the physical interaction between RBP-JK andmNotchl. They concluded that the CDC10/ankyrin-like repeats of mNotch played an important role in theinteraction with RBP-JK, because the replacement oftwo amino-acid residues in the ankyrin repeat 4 appearedto weaken the interaction between RBP-JK and theintracytoplasmic region of mNotch, as determined bygel-retardation analysis. Unfortunately, however, theauthors did not measure the amounts of Notch mutantproteins in the cell extracts, although it is well knownthat mutant proteins are often labile in transfected cells.In fact, their own immunostaining data appear to showthat this is the case. We have found that mRAM23inhibits, in a dominant-negative fashion, the transactiva-tion of the HES-1 promoter by the intracytoplasmicregion of mNotch, indicating that the interactionbetween mRAM23 and RBP-JK is comparable to thatbetween the Notch intracytoplasmic region and RBP-JK(S.M., T.S. and T.H., unpublished observations).
Models for Notch signal transductionThe finding that the receptor Notch and the transcrip-tion factor RBP-JK/Su(H) interact directly suggests thatNotch signal transduction operates by a novel molecularmechanism. Fortini and Artavanis-Tsakonas [5] haveshown that, in S2 cells transiently overexpressing bothDrosophila Notch and Su(H), the ligand-dependent cap-ping of Notch reduced the frequency of Su(H) proteinexpression in the cytoplasm and increased that in thenucleus. These observations led them to propose thatSu(H) is associated with Notch in the absence of ligandbinding, and is released from the receptor upon ligandbinding to migrate to the nucleus. This model implies

Interaction between a novel domain of Notch and RBP-JK/Su(H) Tamura et al.
Fig. 4. Three models for Notch signaltransduction. The RBP-JK signal trans-ducer model (a), the receptor-cleavagemodel (b) and the combined model (c)are schematically presented to explainthe direct interaction between the cell-surface receptor Notch and the tran-scription factor RBP-JK/Su(H) that wehave detected. Notch is assumed toform a dimer upon interaction with theligand (Delta), which is expressed onthe, surface of other cells. Horizontalarrows indicate proteolytic cleavage ofthe intracellular portion of Notch.
that Su(H) serves not only as a transcription factor butalso as a signal transducer (Fig. 4a).
A complicating factor, however, is that RBP-JK is foundalmost exclusively in the nuclei of differentiated as well asundifferentiated mammalian cells expressing Notch,although a small fraction of RBP-JK may be present inthe cytoplasm [35,36]. Our experiments, in which RBP-JK and full-length mNotch were coexpressed in COS 7cells, showed that RBP-JK was not located in the cyto-plasm (unpublished data). In addition, there are a numberof reports indicating that the intracytoplasmic region ofNotch is localized in the nuclei of Drosophila and mam-malian cells ([37]; reviewed in [1,38]). Taken together,these results lead to another model (Fig. 4b), which sug-gests that ligand binding to Notch induces proteolyticcleavage of the receptor at a membrane-proximal site onthe intracytoplasmic region. This cleavage results in therelease of almost the entire intracytoplasmic region,which then moves to the nucleus to interact with RBP-JK/Su(H), and possibly other proteins; the Notch cyto-plasmic region has, in fact, been shown to contain nuclearlocalization signals [39]. Although the observation thatSu(H) is predominantly cytoplasmic in cells coexpressingSu(H) and dNotch [5] appears to contradict this model,actually Su(H) was also predominantly cytoplasmic in asmany as 21 % of cells expressing Su(H) in the absence ofNotch [5]. These observations suggest that the intracellu-lar distribution of Su(H) may be influenced by several un-known factors, such as the amount of Su(H) produced bytransfection, and it is therefore difficult to draw definitiveconclusions from these data. In mammalian cells, RBP-JKis not predominantly expressed in the cytoplasm [36].
A third possibility is a combination of the two mecha-nisms described above (Fig. 4c). Namely, RBP-JK/Su(H)
associates with Notch at the cell surface, and ligand bind-ing induces the proteolytic cleavage of Notch, resultingin the migration of a complex composed of the Notchintracytoplasmic region and RBP-JK/Su(H) to thenucleus. At this stage, none of the available evidencedefinitively rules out any of the three models describedabove. Finally, it is formally possible that mouse andDrosophila use slightly different strategies for Notch signaltransduction, although this seems unlikely. In any case,the fact that Notch and RBP-JK/Su(H) interact directlyimplies that Notch signal transduction involves a novelmolecular mechanism.
Conclusions
The results we have reported in this paper show that anovel domain, RAM23, of the cell-surface receptorNotch physically interacts with the RBP-JK/Su(H)transcription factor. The direct interaction betweenRBP-JK/Su(H) and Notch implies that Notch signaltransduction employs a novel molecular mechanism.
Materials and methods
Plasmid constructionMolecular manipulations were conducted essentially accordingto standard protocols [40]. Constructs were usually generatedby inserting fragments prepared by restriction enzyme digestionor PCR amplification into appropriate vectors and sites. Toconstruct GAL4(ad)-mNotchl [1 751-1 806] replacementmutants, two-step PCR mutagenesis [41] was carried out. Thesequences of the PCR products were verified by dideoxysequencing. Plasmids encoding the sequence of GST fused tothe sequences encoding mRAM23, dRAM23, dNotch andtheir derivatives were constructed by direct cloning of PCR
RESEARCH PAPER 1421

1422 Current Biology 1995, Vol 5 No 12
amplified fragments into the pGEX-4T-1 or pGEX-4T-3 vec-tor (Pharmacia). Plasmids encoding mNotchl and RBP-JKfused to GST were generated by recloning cDNAs obtainedfrom Gridley [20] and Matsunami [6], respectively. The plas-mid encoding mNotchl-IC was obtained from R. Kopan [42]and contains six Myc-epitope tags at the amino-terminus ofmNotchl [1 810-2 531]. The plasmid encoding mNotchl-RAMIC was generated by inserting a PCR-amplified RAMregion (residues 1 747-1 809) between the Myc-epitope tagand the mNotchl sequences of mNotchl-IC. Oligonu-cleotides encoding the bacteriophage T7 gene 10 epitope(Novagen) were synthesized and appended to the sequenceencoding the carboxy-terminus of RBP-JK. DNA encodingthe mNotchl constructs (IC and RAMIC) and T7-taggedRBP-JK were recloned into the expression vectors pEF-BOS[43] and pMKITNeo (a gift from K. Maruyama), respectively.
Yeast two-hybrid screening and interaction assaysFor the yeast two-hybrid screening, the mouse RBP-JK cDNA(encoding amino-acid residues 1-526) [6] was inserted into thepBTM116 vector as a fusion with the sequence encodingLexA. The resultant plasmid, pLRBP, was cotransformed withthe 9.5-day-old mouse embryo cDNA library in the pVP16vector into the L40 strain and 106 transformants were analyzedas described [28]. Library plasmids were rescued in Escherichiacoli strain HB101 and were classified according to insertlengths. Individual interprotein interactions were tested usingstrains similar to those described above. Yeast manipulationsand 3-galactosidase assays were carried out on individual trans-formations according to standard procedures [40]. Levels ofGAL4(bd) fusion protein expression in yeast were monitoredby western immunoblotting, using an anti-GAL4(bd) antibody(Upstate Biotechnology Incorporated), especially in caseswhere a protein-protein interaction was not detected.
In vitro interactions of GST-fusion proteinsE. coli DH5ot was transformed with either pGEX4T-1 or anappropriate expression plasmid, and GST-fusion proteins wereextracted as described by the manufacturer (Pharmacia). Thesupernatant of extracts was mixed with 50 I1 gluta-thione-Sepharose 4B beads (Pharmacia), and washed with 1 mlof phosphate buffer containing 1% Triton X-100, followed by1 ml RIPA buffer; beads were stored as a 50:50 slurry withRIPA. 35S-labeled RBP-JK or Su(H) was synthesized in vitrofrom pBluescript II vectors containing the RBP-JK or Su(H)sequences, using the TNT-coupled rabbit reticulocyte lysatesystem (Promega). 3 5S-labeled RBP-JK or Su(H) was bound tothe preincubated immobilized GST proteins. Fusion proteinswere used after washing six times in RIPA buffer. Theamounts of GST-fusion proteins were monitored bySDS-PAGE of the Sepharose-GST-fusion complex andequimolar amounts were used for each set of experiments;bound protein was released by boiling in gel sample buffer.
Immunoprecipitation experimentsCOS 7 cells (7.5 x 105) were cotransfected with expressionvectors encoding T7-epitope-tagged RBP-JK and eithermNotchl-IC or mNotchl-RAMIC by lipofection. After 24 h,transfected cells were harvested and lysed in lysis buffer(50 mM Tris-HCI (pH 8.0), 0.5 % NP40, 150 mM NaCl,1 mM EDTA, 0.4 mM PMSF, 10 .g m1-1 aprotinin and1 mM DTT). The suspension was rotated at 4 °C for 1 h, andinsoluble material was removed by centrifugation. The super-natant was mixed with either a mouse monoclonal antibodydirected against the T7-epitope tag (Novagen) or the mousemonoclonal antibody, 9E10, specific for human c-Myc [44],
rotated at 4 C for h, and then mixed with 20 lIl of a 50 %suspension of protein A-Sepharose. After incubation for 1 h,the beads were washed eight times with lysis buffer. Boundproteins were released by boiling in a gel sample buffer andanalyzed by SDS-PAGE. The gel was cut into two and ana-lyzed by western immunoblotting to monitor Notch proteinexpression, using 9E10 and anti-mouse IgG antibodies (upperpart), and to monitor RBP-JK, using T6709/T6719 anti-RBP-JK monoclonal antibodies [36] and anti-rat gG antibodies(lower part).
Acknowledgements: We thank S. Hollenberg, A. Kakizuka, P. Barteland S. Fields for supplying components of the yeast two-hybridsystem. We are grateful to T. Gridley and R. Kopan for plasmidsencoding mNotchl and to P. Simpson for the plasmid encodingdNotch. This investigation was supported by grants from the Min-istry of Education, Science, Sports and Culture ofJapan.
References
1. Artavanis-Tsakonas S, Matsuno K, Fortini ME: Notch signaling. Sci-ence 1995, 268:225-232.
2. Muskavitch MAT: Delta-Notch signaling and Drosophila cell fatechoice. Dev Biol 1994, 166:415-430.
3. Blank V, Kourilsky P, Israel A: NF-KB and related proteins: Rel/dor-sal homologies meet ankyrin-like repeats. Trends Biochem Sci1992,17:135-140.
4. Nolan GP, Baltimore D: The inhibitory ankyrin and activator Relproteins. Curr Opin Gen Dev 1992, 2:211-220.
5. Fortini ME, Artavanis-Tsakonas S: The Suppressor of Hairless pro-tein participates in Notch receptor signaling. Cell 1994,79:273-282.
6. Matsunami N, Hamaguchi Y, Yamamoto Y, Kuze K, Kangawa K,Matsuo H, et al.: A protein binding to the JK recombinationsequence of immunoglobulin genes contains a sequence related tothe integrase motif. Nature 1989, 342:934-937.
7. Tun T, Hamaguchi Y, Matsunami N, Furukawa T Honjo T, KawaichiM: Recognition sequence of a highly conserved DNA binding pro-tein RBP-JK. Nucleic Acids Res 1994, 22:965-971.
8. Furukawa T, Kawaichi M, Matsunami N, Ryo H, Nishida Y, Honjo T:The Drosophila RBP-JK gene encodes the binding protein for theimmunoglobulin K recombination signal sequence. Biol Chem1991, 266:23334-23340.
9. Amakawa R, Wu J, Ozawa K, Matsunami N, Hamaguchi Y, MatsudaF, et al.: Human K recombination signal binding protein gene(IGKJRB): Comparison with its mouse homologue. Genomics 1993,17:306-315.
10. Chung C-N, Hamaguchi Y, Honjo T, Kawaichi M: Site-directedmutagenesis study on DNA binding regions of the mouse homo-logue of Suppressor of Hairless, RBP-JK. Nucleic Acids Res 1994,22:2938-2944.
11. Furukawa T, Maruyama S, Kawaichi M, Honjo T: The Drosophilahomolog of the immunoglobulin recombination signal-binding pro-tein regulates peripheral nervous system development. Cell 1992,69:1191-1197.
12. Schweisguth F, Posakony JW: Suppressor of Hairless, theDrosophila homolog of the mouse recombination signal-bindingprotein gene, controls sensory organ cell fates. Cell 1992,69:1199-1212.
13. Dou S, Zeng X, Cortes P, Erdjument-Bromage H, Tempst P, Honjo T,et al.: The recombination signal sequence-binding protein RBP-2Nfunctions as a transcriptional repressor. Mol Cell Biol 1994,14:3310-3319.
14. Grossman SR, ohannsen E, Tong X, Yalamanchili R, Kieff E: TheEpstein-Barr virus nuclear antigen 2 transactivator is directed toresponse elements by the Jk recombination signal binding protein.Proc Natl Acad Sci USA 1994, 91:7568-7572.
15. Henkel T, Ling PD, Hayward SD, Peterson MG: Mediation ofEpstein-Barr virus EBNA2 transactivation by recombination signal-binding protein K. Science 1994, 265:92-95.
16. Laux G, Adam B, Strobl L, Moreau-Gachelin F: The Spi-1/PU.1 andSpi-B ets family transcription factors and the recombination signalbinding protein RBP-JK interact with an Epstein-Barr virus nuclearantigen 2 responsive cis-element. EMBOJ 1994, 13:5624-5632.
17. Zimber-Strobl U, Strobl L, Meitinger C, Hinrichs R, Sakai T,Furukawa T, et al.: Epstein-Barr virus nuclear antigen 2 exerts its

Interaction between a novel domain of Notch and RBP-JK/Su(H) Tamura et al.
transactivating function through interaction with recombinationsignal binding protein RBP-JK, the homologue of Drosophila Sup-pressor of Hairless. EMBO J 1994, 13:4973-4982.
18. Waltzer L, Logeat F, Brou C, Israel A, Sergeant A, Manet E: Thehuman JK recombination signal sequence binding protein (RBP-JK)targets the Epstein-Barr virus EBNA2 protein to its DNA responsiveelements. EMBOJ 1994, 13:5633-5638.
19. Furukawa T, Kobayakawa Y, Tamura K, Kimura K, Kawaichi M, Tan-imura T et al.: Suppressor of Hairless, the Drosophila homologue ofRBP-JK, transactivates the neurogenic gene E(spl)m8. Jpn J Genet1995, 70:505-524.
20. Franco del Amo F, Gendron-Maguire M, Swiatek P, Jenkins NA,Copeland NG, Gridley T: Cloning, analysis, and chromosomallocalization of Notch-i, a mouse homolog of Drosophila Notch.Genomics 1993, 15:259-264.
21. Lardelli M, Lendahl U: Motch A and Motch B - Two mouse Notchhomologues coexpressed in a wide variety of tissues. Exp Cell Res1993, 204:364-372.
22. Lardelli M, Dahlstrand J, Lendahl U: The novel Notch homologuemouse Notch 3 lacks specific epidermal growth factor-repeats andis expressed in proliferating neuroepithelium. Mech Dev 1994,46:123-136.
23. Ellisen LW, Bird J, West DC, Soreng AL, Reynolds TC, Smith SD, etal.: TAN-I, the human homolog of the Drosophila Notch gene, isbroken by chromosomal translocations in T lymphoblastic neo-plasms. Cell 1991, 66:649-661.
24. Robbins , Blondel B, Callahan D, Callahan R: Mouse mammarytumor gene int-3: A member of the notch gene family transformsmammary epithelial cells. I Virol 1992, 66:2594-2599.
25. Swiatek PJ, Lindsell CE, Franco del Amo F, Weinmaster G, GridleyT: Notch is essential for postimplantation development in mice.Genes Dev 1994, 8:707-719.
26. Conlon RA, Reaume AG, Rossant J: Notch 1 is required for the coor-dinate sementation of somites. Development 1995, 121:1553-1545.
27. Oka C, Nakano T, Wakeham A, Luis de la Pompa J, Mori C, Sakai T,et al.: Disruption of the mouse RBP-JK gene results in early embry-onic death. Development 1995, 121:3291-3301.
28. Vojtek AB, Hollenberg SM, Cooper JA: Mammalian Ras interactsdirectly with the serine/threonine kinase Raf. Cell 1993,74:205-214.
29. Kidd S, Kelley MR, Young MW: Sequence of the Notch locus ofDrosophila melanogaster: Relationship of the encoded protein tomammalian clotting and growth factors. Mol Cell Biol 1986,6:3094-3108.
30. Coffman C, Harris W, Kintner C: Xotch, the Xenopus homolog ofDrosophila Notch. Science 1990, 249:1438-1441.
31. Weinmaster GC, Roberts VJ, Lemke G: A homolog of DrosophilaNotch expressed during mammalian development. Development1991, 113:199-105.
32. Weinmaster G, Roberts VJ, Lemke G: Notch2: a second mammalianNotch gene. Development 1992, 116:931-941.
33. Bierkamp C, Campos-Ortega JA: A zebrafish homologue of theDrosophila neurogenic gene Notch and its pattern of transcriptionduring early embryogenesis. Mech Dev 1993, 43:87-100.
34. Jarriault S, Brou C, Logeat F, Schroeter EH, Kopan R, Israel A: Sig-nalling downstream of activated mammalian Notch. Nature 1995,377:355-358.
35. Hamaguchi Y, Yamamoto Y, Iwanari H, Maruyama S, Furukawa T,Matsunami N, et al.: Biochemical and immunological characteriza-tion of the DNA binding protein (RBP-JK) to mouse JK recombina-tion signal sequence. J Biochem 1992, 112:314-320.
36. Sakai T, Furukawa T, Iwanari H, Oka C, Nakano T, Kawaichi M etal.: Loss of immunostaining of the RBP-JK transcription factor uponF9 cell differentiation induced by retinoic acid. Biochem 1995,118:621-628.
37. Zagouras P, Stifani S, Blaumueller CM, Carcangiu ML, Artavanis-Tsakonas 5: Alterations in Notch signaling in neoplastic lesions ofthe human cervix. Proc Natl Acad Sci USA 1995, 92:6414-6418.
38. Greenwald I: Structure/function studies of lin-12/Notch proteins.Curr Opin Gen Dev 1994, 4:556-562.
39. Lieber T, Kidd S, Alcamo E, Corbin V, Young MW: Antineurogenicphenotypes induced by truncated Notch proteins indicate a role insignal transduction and may point to a novel function for Notch innuclei. Genes Dev 1993, 7:1949-1965.
40. Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, SmithIA, Struhl K: Current Protocols in Molecular Biology. New York:John Wiley and Sons; 1987-1995.
41. Kammann M, Laufs J, Schell J, Gronenborn B: Rapid insertionalmutagenesis of DNA by polymerase chain reaction (PCR). NucleicAcids Res 1989, 17:5404.
42. Kopan R, Nye JS, Weintraub H: The intracellular domain of mouseNotch: a constitutively activated repressor of myogenesis directedat the basic helix-loop-helix region of MyoD. Development 1994,120:2385-2396.
43. Mizushima S, Nagata S: pEF-BOS, a powerful mammalian expres-sion vector. Nucleic Acids Res 1990, 18:5322.
44. Evan GI, Lewis GK, Ramsay G, Bishop JM: Isolation of monoclonalantibodies specific for human c-myc proto-oncogene product. MolCell Biol 1985, 5:3610-3616.
Received: 26 June 1995; revised: 29 September 1995.Accepted: 23 October 1995.
RESEARCH PAPER 1423
Top Related
Copyright © 2022 FDOKUMEN

