







Bahasa
Halaman
Hukum


PAPERS
Department of Geology
University of Queensland
J. B. WATERHOUSE
World Correlations for Permian Marine Faunas
\ NJ�mber 2

f)(E
I�� sqq
(). � .
V- 1 .m.o �
'1
j�

World Correlations for Permian Marine Faunas

World Correlations for Permian Marine Faunas
by
J. B . WATERHOUSE
Department of Geology and Mineralogy, University of Queensland, Brisbane
UNIVERSITY OF QUEENSLAND PAPERS DEPARTMENT OF GEOLOGY
Volume 7 Number 2

© Geology Department, University of Queensland , St. Lucia, 1 976
This book is copyright. Apart from any fair dealing for the purposes of private study, research , criticism, or review, as permitted under the Copyright Act, no part may be reproduced by any process without written permission. Enquiries should be made to the publisher.
Typeset, printed and bound by Academy Press Pty Ltd, Brisbane

Contents
Abstract xiIntroduction x111Transliteration of Russian names xvii Acknowledgments xviii
Chapter 1 . Correlation Methods for Marine Pennian Invertebrate Faunas
Communities and biozones 4
CORRELATION OF BIOZONES 5
Succession 5 Units 5 Subzones and superzones, substages 7 Key species 11 Recurrent and alternating species and genera 11 Faunal signature 11 Limits of correlation techniques for brachiopods 14 Use of statistical techniques 16
Chapter 2. Review of Chronologie and Biostratigraphic Subdivisions of the Marine Pennian 18
RELATIVE VALUE OF CERTAIN MARINE FOSSIL GROUPS 20
The primary of Brachiopoda 20 Fusulinacea and nomenclatural problems 20 The value and limitation of Ammonoidea 22 Correlation potential 24 Correlation value 24 Risk of error 25 Conodont studies 25
DISCUSSION OF CORRELATION SCHEMES 29
Correlations proposed by Cooper & Grant ( 1973) 29 Ammonoid scheme of Furnish ( 1973) 31 Literature review by Anderson ( 1971) 34 Correlation scheme by Ustritsky (1971) 34
TYPE REGIONS 34
Type sections and stage names 37 Substages 37 Major subdivisions of the period 39
Chapter 3. The Base of the Pennian 40
A. Significant marine biota 40 Fusulinacea 40 Ammonoidea 40

vi Contents
Brachiopoda 44 B. Regions significant for placement of the boundary 44 Pamirs, Austria 44 West Texas, United States 45 Australia 45 Canada 46 Verchoyan, Orulgania, north-east Siberia 46 Options over the Carboniferous-Permian boundary 48
Chapter 4. Biochronology for the Marine Permian 53 ASSELIAN STAGE 53 Urals 53 Donetz Basin, north Russia, Siberia 55 Karakorum Range, Tibet 6 1 Mongolia, China 6 1 South-east Asia, Japan 64 Turkey, Austria, Spain 66 Spitsbergen, Canada, United State!� 69 Bolivia, Peru 7 5 Pakistan, India, Himalayas, Afghanistan 75 South Africa, Argentina, Australia 77 SAKMARIAN STAGE 78 Urals 78 North Siberia, Pamirs 79 Europe 82 Armenia, Iran, Afghanistan, Karakorum, Chitral 82 China, Japan 85 Spitsbergen, Canada, Alaska 85 United States 87 Bolivia, Madagascar, India 92 Australia, New Zealand 92 BAIGENDZINIAN STAGE 95 Urals 95 North Siberia, Pamirs, Afghanistan, Armenia 96 China, Japan 97 Canada, United States, Mexico 97 India, Australia, New Zealand 99 KUNGURIAN STAGE 1 02 Russian Platform, Urals 1 02 Kanin Peninsula, Pai Hoi, Kolyma, Sikhote Alin 1 05 Pamirs, Afghanistan, Iran 1 06 China, Japan, south-east Asia, Pakistan 1 09 North-west Europe 1 1 0 Spitsbergen, Greenland 1 1 3 Canada 1 1 6 United States, Guatemala 1 1 8 Sumatra, Timor 1 22 Australia 1 24 New Zealand 1 27

KAZANIAN STAGE 1 27
Russian Platform, Urals, North Russia 1 27Northern Siberia, Caucasus, Crimea, Afghanistan 1 29 China, Japan, south-east Asia 1 3 1 Pakistan, Iran, Mediterranean 1 34 Spitsbergen, Greenland, North America 1 3 5 Timor, Australia, New Zealand 1 37
PUNJABIAN STAGE 1 4 1
Pakistan 140 Himalayas 141 Karakorum, Afghanistan 1 45South-east Asia, China, Japan 146 South Primoyre, Kolyma River 1 49 Mongolia, Pamirs, Armenia, Iran 149 United States, Mexico 1 52Madagascar, Timor, Australia, New Zealand 1 53
DJULFIAN STAGE 1 57
Armenia, Iran, Caucasus Pakistan 1 6 1Pamirs, Mongolia, China Japan, south-east Asia United States, Mexico New Zealand 1 66
DORASHAMIAN STAGE
1 57
1 63 1 65 1 66
1 68
Vedian and Ogbinan Substages 1 68 Armenia, Iran 1 68Europe 1 68China, Japan 1 68Madagascar, New Zealand 1 69 Griesbachian Substage 1 72
Contents vii
Chapter S. Summary of Pennian Stages and Substages: Problems and Proposals 176
Asselian Stage 1 76 Sakmarian Stage 1 76 Baigendzinian Stage 1 79 Kungurian Stage 1 79 Kazanian Stage 1 8 1 Punjabian Stage 1 8 1 Djulfian Stage 1 8 1 Dorashamian Stage 1 83
References 1 84
Geographic Index 207Stratigraphic and Biostratigraphic Index 2 1 2
Systematic Index for Gene�a and Species 222

Tables
1 . Subdivisions of the marine Permian System xiv 2a,b. World correlations for the most significant marine Permian sequences
3. Faulty standard sections 6
4. Early Permian occurrences of Attenuatella
5 . Alternating chonetids in the mid-Permian 6. Alternative correlations for New Zealand 7. Conodont zonation 27
1 2 1 3 1 7
8. Range of key conodont species 28
9 . Scheme for world correlation proposed b y Grant and Cooper { 1 973)
10. World correlations proposed by Furnish { 1973) 33
1 1 . World correlations proposed by Ustritsky ( 1971 ) 3 5
1 2. Upper Palaeozoic successions of the Russian Platform and Urals 13 . Middle and Late Carboniferous correlations for north and north-east
Siberia 47
XV
30
4 1
14. Permian stratigraphic succession of Pai Hoi, north Russia 56 1 5 . Correlation of Siberian sequences from Taimyr Peninsula, Lena River-Verchoyan
16 . 17 . 18 . 19 . 20. 21 . 22. 23. 24. 25. 26. 27.
28 . 29. 30. 3 1 . 32. 33 . 34. 35 . 36. 37. 38. 39. 40. 41 . 42. 43 . 44.
Massif, and Kolyma River-Omolon Massif 58 Siberian correlations, after Menner et a/. 1970 59 Permian of Darvas, central Asia 60 Permian sequence of south China 62 Permian correlations for south-east Asia Permian correlations for south-west Japan Auernig sequence, Carnian Alps 70
65 67
Permian correlations for Arctic Canada 73 Permian succession in Fergana, c�ntral Asia 8 1Late Palaeozoic succession in Carnic Alps, Austria 83 Late Palaeozoic successions of Spitsbergen and Bj¢rn�ya 86 Permian formations and correlations of west Texas, United States 88 Correlation of McCloud fusulinid zones of California with other lower Permian sequences of United States, following Wilde ( 1 97 1 , fig. 8) 9 1Permian sequence of Salt Range, Pakistan 1 00 Permian correlations for Western Australia 1 0 1 Nomenclature of early Middle Permian Units 1 04 Permian faunal succession of Elburz Mountains, North Iran Permian succession for northern Japan 1 1 1Middle and Late Permian rocks of the Zechstein Sea Phosphoria rock complex, north-west United States Permian sequence at Las Delicias, Mexico 1 23Permian sequences of Queensland 1 25Permian brachiopod zonal succession in New Zealand Permian succession in central Afghanistan 130 Permian sequence at Sisophon, Cambodia 1 33
1 1 4 1 20
1 28
Permian correlations for Australia and New Zealand 1 3 8Permian correlations for Queensland, proposed by Dear ( 1971 )Permian sequence in Indo-China, according to Deprat ( 1 9 1 3)Permian sequence of Sikhote Alin, west Siberia 1 50Permian faunas of Timor 1 54
1 08
139 1 47

45. Pennian sequence in Armenia 1 58 46. Correlation of Annenia sequence, proposed by Furnish ( I 973) 1 59 47. Faunal sequence in Jisu Honguer Formation, Mongolia 1 64 48. Late Pennian sequences in Japan 1 7049. Significant Early Permian genera and species 1 77 50. Significant Early Permian genera and species 1 78 5 1 . Significant Middle Pennian genera and species 1 80 52. Significant Middle and Late Permian genera and species 1 82
Tables ix

Text Figures
1 . World locality map showing places with significant marine Permian outcrops xii
2. Brachiopod biozones for the Permian of Yukon Territory, Canada 2 3. Biozones for the Permian of New Zealand 3 4. Biozones for the Permian of central Afghanistan 8 5. Faunal signature 1 0 6. Evolution of genus Spiriferella 1 5 7. World marine Permian sequences, showing number ef biozones with
Fusulinacea, Brachiopoda, and Ammonoidea 2 1 . 8 . Evolution of Cyclolobidae, according to Furnish ( 1 973) 32 9. Evolution of Cyclolobidae, according to Waterhouse ( 1 972b) 32
10. Hypothetical climatic change for Permian Period 38 1 1 . Evidence from spores on the base of the Permian System 43 1 2. Option 1 for the base of the Permian System 50 1 3 . Option 2 for the base of the Permian System 50 14. Option 3 for the base of the Permian System 5 1 1 5 . Preferred option 4 for the base o f the Permain System 5 1 16 . Map of Permian of Soviet Union 54 1 7. Geographic map of China and surrounding regions 63 1 8. Map of Permian of Spitsbergen 68 19. Map of Permian and Carboniferous sediments in northern Yukon
Territory , Arctic Canada 7 120. Permian basins in South America 74 2 1 . Permian basins in Australia 76 22. Palaeogeographic setting of Permian in west Texas 89 23 . Geology of the north Bowen Basin, Queensland 93 24. Permian rock in New Zealand 94 25. Zechstein Sea of northwest Europe 1 1 2 26. Section of Middle and Late Permian in Greenland 1 1 5 27. Permian formations in Canadian Arctic Archipelago 1 1 7 28. Section of Permian at El Antimonio, north Mexico 1 2 1 29. Permian form�tions in part of the Glass Mountains, west Texas 136 30. Stratigraphic column for Permian-Triassic of Nepal 1 42 3 1 . Permian of Southland, New Zealand 1 5 5 32. Top of Permia� sequence in Salt Range, Pakistan 1 60 33. Alternative interpretations of Permian-Triassic contact in Salt Range ,
Pakistan 1 62 34. Distribution of late Middle and Late Permian Fusulinacea I-67 35. Distribution of Otoceras, key to Late Permian 1 7 136. Late Permian in Guryul Ravine, Kashmir 1 7 1 37. Permian-Triassic contact in New Zealand 1 73 38. Late Permian (Griesbachian) in Canadian Arctic 1 73

Abstract
The marine Permian faunas of the world are subdivided into eight stages and nineteen substages, the Asselian Stage, with the Surenan, Uskalikian, and Kurmaian Substages; the Sakmarian Stage� with the Tastubian, Sterlitamakian, and Aktastinian Substages; the Baigeildzinian Stage, with the Sarginian and Krasnoufimian Substages; the Kungurian Stage, with the Filippovian and Irenian Substages, the latter including the Nevolin, Elkin, and Ufimian horizons; the Kazanian Stage with the Kalinovian and Sosnovian Substages; the Punjabian Stage with the Kalabaghian and Chhidruan Substages; the Djulfian Stage with the Urushtenian and Baisalian Substages, and the Dorashamian Stage with the Vedian and Ogbinan Substages and perhaps Griesbachian Substage or modification thereof. This offers the most extensive and refined scheme of correlation �vailable for the Permian System, in which brachiopod correlations agree well with those proposed for the less abundant Permian ammonoids, and palaeotropical Fusulinacea. Each stage is discussed In terms of its type region and fauna based chiefly on Soviet or Pakistan outcrops,. and then traced over the world in various palaeogeographic settings. The base of the Permian Period as understood in some regions possibly overlaps with Late Carboniferous, and the. top of the marine Permian pr�bably needs to be elevated to incorporate part or all of the Griesbachian 'Stage', generally classed as basal Triassic. Such questions should be resolved by international discussion.

��"nuaoa:�
r::P
a
UCIRNOYA
,
KAZA
KHDAN
�
'-J·�
w-
c::::::::::, ..,o,
,. �
..... �
"'
�..tj
-�-j
NIW
CAUDO
NIA
wtm
iH
�
�-
;wn)•-AIMftAUA
-��
• YuNI,-
Gya ..
... a.
l�elrw
lnll
WAU
l -I.
NOIIl
ll ..
._,. "'AUCICLAN
D
_,..'i!l
-�.v
ICIUniLAN
D -
-
�·
�
-
11 _
i I
/ ....._')
r
7/
Ji;;JC AR
CTIC
CIRCL
E
------TR
OPIC
OF CA
NCER
----------f------
-'<=:;--------EOOATO
R
--r«----TROPI
C OF CA
PRICO
RN
-------7fh/-
----
·-----
ANTA
RCTIC C
III:CLE
Fig.
1.
Me
rc
at
or
pr
oje
ct
ion
, sh
ow
ing
so
me
of
th
e p
lac
e n
am
es
sig
nif
ica
nt
fo
r m
arin
e P
er
mia
n c
orr
lea
tio
ns
.

Introduction
The Permian System, proposed on the basis of rocks and faunas of the Ural Mountains .an� Russian �latform in the Soviet Union by Murchison ( 184 1), was one ofthe more s1gmficant penods of earth history. From a scientific viewpoint the period ended with the greatest of life catastrophes (McAlester, 1973). At its base p;culiarly distributed tillites helped suggest the theory of continental drift to Wegener ( 1929), and its rocks have provided some of the most convincing evidence for continental movement (Irving & Parry 1963). Economically the system has proved to be of high value. Indeed, the copper deposits in the.middle Permian Kupferschiefer of Thuringia and Saxony gave tl:e impetus to studies by one of the fathers of geology, Abraham Gottlieb Werner, who taught at the mining school in Freiberg, east Germany. The rich salt, copper, uranium, phosphate, and other mineral deposits of sedimentary and igneous origin, found widely over the world, are reinforced by many oil and gas fields, and enormous coal resources, notably in the fragments of Gondwana and the Tunguskian and other basins of the Soviet Union.
But any full understanding of Permian events and causes in the evolution of life, climatic change, and development of continents and mineral wealth depends on firm control of sequence, or relative age, and this has been markedly lacking for the period. In the first place, in Europe and European Russia, where stratigraphy and palaeontology were established during the nineteenth century, much of the upper half of the Permian System is terrestrial, without marine fossils. This meant that studies elsewhere were done in a vacuum, because no good European section was available for comparison. Secondly, many of the best studies on Permian fossils have been concentrated on Fusulinacea and Ammonoidea. But Fusulinacea are absent from extensive Permian outcrops, including large tracts of the Arctic, the entire continent of Australia, and many other important regions. Ammonoidea are also restricted in distribution, and are seldom numerous enough to establish even local zonation. Both are superbly useful where found, but neither group was common enough and widespread enough to permit detailed and world wide zonation for Permian faunas. We are forced to use ordinary benthos, such as Brachiopoda and to less extent Bivalvia and Gastropoda, and these are so numerous and diverse that it has taken more than a century of study to master the distribution and range of useful species and genera.
It is now timely to incorporate all these studies into a detailed correlation of marine faunas in the Permian System (Table 1 , 2). The following outline summarizes the stratigraphic and faunal content of the Russian-based stages and zones, and then traces the faunas over the globe, through similar and different palaeogeographic realms. The study is based on fnur interrelated• procedures by the writer: 1 . Detailed studies of faunas and stratigraphy in western and Arctic Canada, Nepal,
New Zea:land, Thailand, New Caledonia, and Australia, with more limited studiesor assessments of faunas from Iran, Burma, India, and England.
2. Study at institutions and museums of faunas described from Mexico, United States,Bolivia, Peru, Spitsbergen, Siberia, Urals, Russian Platform, Tunisia, E.ngland, Germany, Sicily, Armenia, Madagascar, Salt Range, Himalayas, �ambod1a, and La�s.Faunas from Japan, China, Bellerophonkalk of Austna and Yugoslavia,
Karakorum and Greenland have not been seen at first hand.3. Field observations in United States, Mexico, Bolivia, Brazil, Alaska, East G�r
many, Carnian Alps (Austria and Italy), Salt Range, Him�layas .• and Armema.4. Perusal of literature on Permian stratigraphy and faunas, m wh1ch faunas were

Stage Substage Horizons P Number
Series & Symbol
Permian or Diener ian Triassic
GriesbachJ.an 19 p� Upper
Dorashamian Ogbinan 18 PDo Permian
Vedian 17 PDv Dj ulfian Baisalian 16 PJb
Urushtenian 15 PJu Punj ab ian Chhidruan 14 PPc
Kalaba51:hian 13 PPk Middle Kazan ian Sosnovian 12 PZs Permian
Kalinovian 11 PZk Kungurian Ufimian 10 c
Irenian Elkin 10 b PKl Nevolin 10 a
Filippovian 9 PKf Baigendzinian Krasnoufimian 8 PBk
S ar51:inian 7 PBs Sakmarian Aktastinian 6 PSa
Lower Sterl itamakian 6 PSs Permian
Tastubian 4 PSt Asselian Kurmaian 3 PAk
Uskalikian 2 PAU Sur en an 1 PAs
Upper Orenbur2ian 0 Carboniferous
Gshelian 0
Kasimovian 0
Table 1 . Subdivis ions of the marine Permian System

Introduction xv
reassessed, reidentified, and aged, it must be stressed, independently from the view expressed in the work, against a total world framework of zones and ages. As far as possible, studies are complete up to the end of 1 973.A companion work to this study is the paper 'Global distributions and character of
Permian biomes based on brachiopod assemblages' by Waterhouse & Bonham-Carter (1975) which provides a compact data bank for the distribution of marine brachiopod faunas, substage by substage, with occurrences of brachiopod families and cluster analyses of their associations. The paper by Waterhouse ( 1 975) sets out new taxa for use in the present work. A forthcoming paper will summarize radiometric values for the period, in the light of new stratigraphic advances.
TRANSLITERA TION OF R USSIAN NAMES
In the transliteration of Russian names it has been decided to use proper nouns rather than adjectival derivatives for formational and other stratigraphic terms, thus eliding the 'sk' from many words. In view of the fact that English is not phonetic and not consistent, it is impossible to provide a phonetic rendition of foreign words although various pseudophonetic and somewhat comical renditions are available. Therefore the chief concern has been to trea:t the words with consistent spelling, and to slightly anglicize them, as must be done if they are to be acceptable, and in some instances, perhaps to enter common usage amongst English-speaking geologists.

Stage
Dorashamian
Djulfian
Punjab ian
Kazanian
Kungurian
Baigendzinian
Sakmarian
Asselian
Tab le 2 a .
Subs tage Permian P la t form Sa l t Range Armenia Ura l s
Griesbachian Kutuluk I
Ogb inan Pa:zoatizooUtes z Come Zicania z +
Vedian Ma lokinel ArawiZevis z � Codonofusiella Baisalian ..... � Z • Abadeh
Urushtenian � Bolshkinel Kathwai M
Chhidruan Chhidru FIII + Hachik H
Kalabagh ian Sok Kalabagh M + Gnishik. H
Sosnovian Sosnovian + Middle
Armik H Ka linovian Ka linovian + Warga l
Fm Ufimian H +
lrenian Elkin H + Lower Nevolin H +
F i lippovian Fi lippovian + Asni H
Krasnoufimian Saranin (+) Amb / Sarginian Sarginian + F
Aktas tinian Akta s tinian + Sardi Fm Dava lin
S ter 1i tamakian Sterli tamakian + Warchha Fm H
Tastubian Tastub ian + Conutaztia Beds
Kurmaian Kurmaian + Eur:ydesma beds Talchir
Uska likian Uskalikian + / Surenan Surenan +
Intercorrelations of the mos t signi fi�ant marine s equences in the world ,
for the Permian Period .
M Member , Fm- Formation , H Horizon , Z - Zone , f - fauna,
+ type for sub s tage .

r.. .. � 'tl a
,>1. � E:<
r.. ,>1. Gl Gl II u Gl ,..j tJI § ..,
Arctic canada USA Texas New Zealand Queensland China Australia
Griesbachian +
Ochoan ? Rewan Fm Chang sing
Aperi spi rifer Baralaba CM ? Fml!J nelsonensis z
Gp ·O"iIll J?urvi lleoceras z +' ·rl Wuchiaping Hegler M � Spinomartini a z Tamaree Fm? Fm .:I
a "' I Plekonella Upper South +' multicostata z Curra Lmst ·rl Cherry � l!J u Canyon M
Martiniopsis woodi .!( Cll z Cll Appel Ranch M H Terrakea brachg- Pelican u
"' thaerum z Ck f ::s Maokou
Cancrinelloides Willis Ranch M +' g Echinalosia ovalis
z � � Notospirifer Z Scottville f Fm
e � "' H ? � J..i.ssochonetes
China Tank M
Pseudosgrinx z Road Canyon Fm Terrakea Gebbie IIIc exmoorensis z G IIIb
III a Sowerbina Z Cathedral Echinalosia z
Mountain Fm l!J Antiquatoni a z Martinia z
Jakutoproductus Lenox Notostrophi a sirius Shale Chihsia z � z Hills Fm ·O"i Tiverton Fm Fm
Tornqui stia z .... ·O"i � Lizzie Ck Fm
Yakovlevi a z E-< Tomiopsis z Mourlonia
Neal Ranch ? Burnett Fm Orthoti chia z .Fm Maping
Fm Kochiproductus � Gap tank Atomodesma Joe Joe Fm
Table 2b . Intercorrelations of the most siqnlficant marine sequences in the
world for the Permian Period .
CM - Coal Measures ; Fm Formation ; f - fauna ; Gp - Group ;
H - Horizon ; L - Limestone; M - Member ; Z - Zone

Acknowledgements
A number of individuals have contributed greatly to this present study, amongst whom I would especially mention my Canadian colleagues Drs E.W. Bamber, W.W. Nassichuk, and E.T. Tozer, Geological Survey of Canada; and American colleagues Drs G.A. Cooper and R.E. Grant of the Smithsonian Institution, Washington, D.C. Drs Nassichuk and Grant have been especially helpful in reading the manuscript. Dr D.J. Gobbett, Solihull, England; and Dr R. Douglass, U.S. Geological Survey, Washington D.C. have also read parts of the manuscript, and helped correct Fusuline names. Dr H. Kozur, Meiningen, East Germany has advised me on conodont correlations. Dr T.G. Sarytcheva, Palaeontological Institute, Moscow, provided copies of much of the Russian literature now so hard to obtain, and Dr M.V. Kulikov, VSEGEI, Leningrad, has also been most helpful in many ways. Many colleagues have guided me in the field, or assisted in examining collections, including Mr M. Banks, University of Tasmania; Dr K.S.W. Campbell, Australian National University; and Dr J.M. Dickins, Bureau of Mineral Resources, Australia; Dr G. Fuchs, Geological Bundesanstalt, Austria; Dr H. Jordan, East Germany; Dr E.G. Malkhaisian, Armenia; Mr A.N. Fatmi, Geological Survey of Pakistan; Mr A. Khan, Geological Survey of Nepal; Mr F. Branisa, Geological Survey of Bolivia; Mr K. Pitakpaivan, Geological Survey of Thailand, with many others. I also wish to underline the great helpfulness from staff of the University of Toronto Library, under Mr R.H. Blackburn. The study has been financed almost entirely by the National Research Council of Canada, from grant A4263, from 1967-1 975.

1 . Correlation Methods for Marine PermianInvertebrate Faunas
Permian marine faunas and floras are abundant and diverse, and it will probably be fou.nd that a nu�ber of groups are useful for correlation, when sufficiently studied.Brachtopoda are wtdespread, moderately numerous, and have been studied for over a ce�tury so that these have to �ssume the main burden of correlation in many regions(Ftg. 1 ). Where present, Fusuhnacea and Ammonoidea are of high value. All three groups were subject to facies and climatic interference. Fusulinacea were least tolerant, and showed strong provinciality (Ross, 1 967b). Ammonoidea were a little more tolerant, but their species and genera were not as wide ranging geographically, nor as tolerant of so many facies as a number of brachiopod species, in spite of what has been written about free floating short lived ammonoid shells (cf. Ustritsky, 1 974). Thus ammonoids are absent from most Permian outcrops. Brachiopods varied in their distribution and control by lithofacies, but a number were highly tolerant, widely dispersed, and very short lived. They are the most common macro-invertebrate marine fossil in many Permian deposits.
A number of other groups have high potential. Bivalvia and Gastropoda were rather rare in many parts of the world during the Permian, but ranged more widely than Fusulinacea or Ammonoidea, and locally are valuable. Bryozoa are particularly widespread and numerous, and will probably prove to be as useful as Brachiopoda, but have been somewhat neglected. Studies of microfossils are beginning to show high promise. Small Foraminifera and ostracods have been used successfully for subdividing the Kungurian and Kazanian stages in the Soviet Union and conodonts appear to have excellent potential. Admittedly Grant & Cooper ( 1 973) devastatingly criticized the work by Sweet ( 1 970) on the Permian-Triassic conodonts as being inconsistent, and 'too deferential': but such flaws may not lie in the conodonts themselves. However it still has not been established that they occur abundantly in high palaeolatitudes or in rock with glacial sediment. Plant spores and acritarchs have been studied widely in recent years (Balme, 1 970; Hart, 1 970; Visscher, 1 973), but floral divisions are extremely broad at the moment.
Because of controversy over correlations for the Permian Period, some writers have stressed that each fossil group obeyed its own laws, with evolution proceeding at various rates, so that species of one group were unlikely to have lived and died in phase with species of other groups. This has been most absurdly, yet seriously epitomised in the remark 'Ammonoids don't speak to Fusulines and Fusulines don't speak to Brachiopods'. Although it would be possible to express this concept in some sort of evolutionary model, there is little evidence to support such a view. The present controversies over correlation have resulted from differing interpretations of the same fossils. Errors of correlation have been made, and these may not be excused by allegations of widely disparate behaviour· of different fossil groups. The present work will show how, over the entire globe, Fusulinacea, Brachiopoda, and Ammonoidea, and from what is known the less studied groups, all behaved in harmony. Already it is clear that the Ammonoidea and Fusulinacea are in close agreement for the Early Permian, andbrachiopod evidence agrees perfectly with world al_llmonoi� subdivisions for the Ear�yand most of the Middle Permian, indeed for the enttre Permtan but for one sequence, m the Salt Range. A study by Waterhouse ( 1 973a) on.
faunas in south-east Asia s�owedthat Fusulinacean zones conformed well with brachiOpod zones of the same reg10n. It appears likely that for the duration of a zone various life forms bel�nged to a �losely. interlinked plexus of contemporaneous communities, governed by chmate and hthofactes,

2 Correlation Methods
I 10m B RA C H I O P O D G R OUP
ZON E 1 2 3 4 5 6 7 8
! Gc
I I ! ! I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I Fl I I I I I I I I I I I I I I I I I I I I I I I I I ! I j I I I I I I I l I I Fps I I I I I Fs I I I ; Fa
! ! ! ! ! ! ! ! ! ! ! ! I I i ! I I I i i Ej I ! I
i I i I i I i i i j I i i i i
Et s ! s s s s s s s s s s � I ! ! ! s � s: I I !
Ea I s s s: s s s � s 7 s I I . I I I I
! ! ! ! ! ! ! ; ! ! ! ! IIII I
Ey I I I I I
il I i i i i i i I i i Eta I I I I I I J I I I I
! ! ! ! ! ! ! ! ! ! ! ! ! ! i I I ! I ! I I ! ! ! I i i I i ! ! i I I I i ! ! i ! I I Eog i i ! i I ! i I i I i I I i ' I I i i I i i I I ! I i I i i i ! ! ! i Eka l I I I I
Fig. 2. Range chart of brachiopod species in Permian beds of the north Yukon Territory, Arctic Canada. Species that commence in a zone are assigned a distinct kind of line, and many are replaced from zone to zone, though a few persist. The species are grouped as follows: 1 . Inarticulata (zone Ea) ; Orthida; 2. Davidsoniacea; 3. Chonetidina; 4. Strophalosicacea; 5 . Productacea; 6. Rhynchonellida; 7. Atrypida; 8. Spiriferida; 9. Terebratulida. Zones match the type stages and sub-stages, based also on biozones, of the Urals as follows: GC - Lower Kazanian or Kalinovian; Fl - Nevolin ; Fps -Filippovian; Fs - Krasnoufimian; Fa - Sarginian; Ej - Aktastinian; Et and Ea - Sterlitamakian; Ey - Tastubian; Eta - Kurmaian; Eok - Uskalikian; Eka - Surenan. The symbol S links species absent from an intervening zone. Thickness approximate, exaggerated for narrow zones.
9
I
!
I
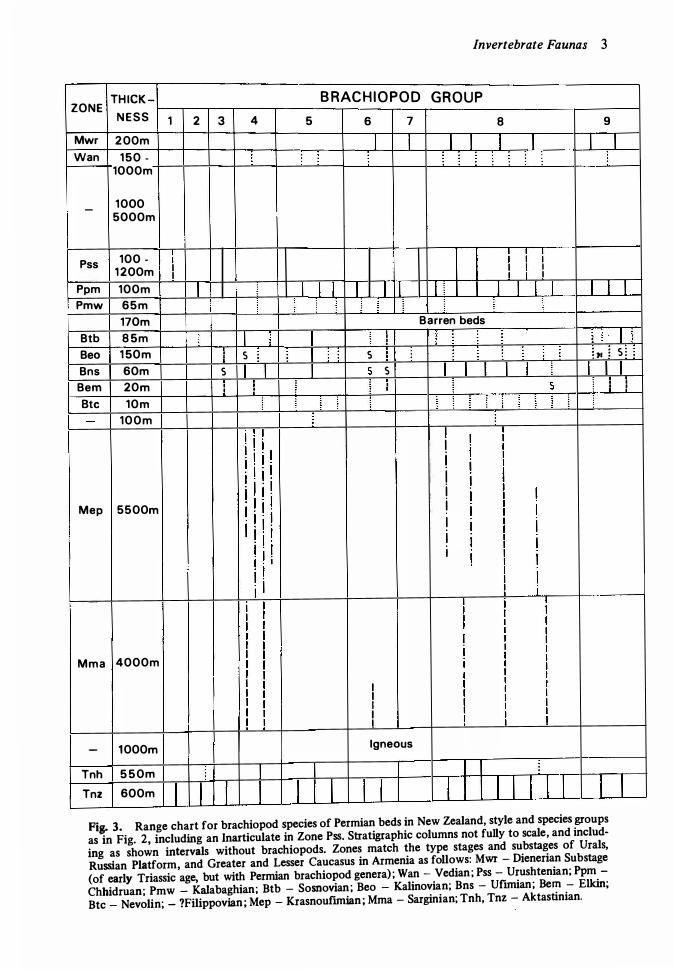
Invertebrate Faunas 3
ZONE TH ICK - B RAC H I O PO D GROUP
N E S S 1 2 3 4 5 6 7 8 9 Mwr 2 00m I I I I I I I Wan 1 50 -
1000m : : :
1 000 -5000m
Pss 100 - : : : : 1 200m I I I : I I I
Ppm 100m I I I I I I . I I ! I I I I I I I Pmw 6 5 m
170m Barren beds
Btb 8 5m I : I I I �
Beo 1 50m I s s I ;, : I s :
Bns 60m s I I s I I I I I I I I Bern 20m I I I I I I
Btc 10m
1 0 0 m
! I ! I iI i
Mep 5500m i I
i I
I I
M m a 4000m
I I I I i i I
- 1 000m Igneous
Tnh 5 5 0m
Tnz 600m I I I I 1 I I I 1 I I I I I Fig. 3. Range chart for brachiopod species of Permian beds in New Zealand, style and species groups
as in Fig. 2, including an Inarticulate in Zone Pss. Stratigraphic columns not fully to scale, and includ
ing as shown intervals without brachiopods. Zones match the type stages and substages of Urals,
Russian Platform, and Greater and Lesser Caucasus in Armenia as follows: Mwr - Dienerian Substage
(of early Triassic age, but with Permian brachiopod genera) ; Wan - Vedian; Pss - Urushtenian; Ppm
Chhidruan; Pmw - Kalabaghian; Btb - Sosnovian; Beo - Kalinovian; Bns - Ufunian; Bern - Elkin;
Btc - Nevolin; - ?Filippovian; Mep - Krasnoufunian; Mma - Sarginian; Tnh, Tnz - Aktastinian.
I I I I I
I

4 Correlation Methods
under prevailing trophic and geochemical conditions. Any evolutionary change to one segment would have had drastic effects on co-habiting groups, at least at the same trophic level, which would then affect life at other trophic levels. Evidence is convincing that Fusulinacea and Brachiopoda (and one would suspect, Bryozoa and Anthozoa) were closely interlinked. Certainly some ammonoid species behaved with greater toleration by living for a longer time, but generally they still show, within their limits ·of distribution, new incomings at the same time as the rest of the faunas.
Communities and Biozones Permian marine fauna and flora were organised in communities, for which studies
are just commencing (Waterhouse, 1 973c). Communities are characterised by their numerically preponderant and large-sized species-perhaps one, to three or four-and may be objectively established by counting individuals in collections after assessing the extent of interference by transport, postburial solution, and extraction. Within the marine environment, a number of different communities generally developed in any one region, each governed by different primary species that were favoured, in the case of brachiopods, by bottom sediment especially, and also influenced by depth, and other biota. As today, a large measure of background control was exerted by salinity, and by temperature, affecting geochemical conditions and trophic resources, so that diversity increased towards the palaeoequator. Over the world, a succession of fossil biomes or major associations of communities were organised, each occupying 1 5°-30° latitude, in reflection of climate and trophic resources.
The relationship between communities and biozones is particularly critical for biostratigraphy and chronostratigraphy. In practice, palaeontologists have included a number of correlative communities in a single biozone, because the communities shared a sufficient number of species to make up a 'faunal assemblage zone', or each had individuals of a given key species, sufficient to designate a 'range zone', or all shared an important species that made a significant 'first entry', often of high correlative value. Individual communities, though treated as zones in some texts, occupied too small a part of the sedimentary column to be readily mapped as single zones, for one low bluff may expose two to five communities or even more, recurrently. Thus, biozones embrace various fossil communities which were distinguished by their differing response to different environmental parameters, and are correlated because they share significant species. These major assemblages of contemporaneous communities strongly approach the modern 'biome' (Krassilov, 1 974; Waterhouse 1 974a; Waterhouse & BonhamCarter, 1 975), and are essentially identical with biotic provinces.
Few species within zones of Permian age show morphological clines, anobservation confirmed by Eldredge & Gould ( 1972) who stated that the geological record seldom reveals what they called 'phyletic gradualism'. Specific and general changes, and as a result, zonal and community boundaries, were abrupt, greatly aiding correlation. This is an important observation, against current fashions in evolutionary theory, but cannot be denied for at least a number of Permian successions that have been discussed in modern terms. Two examples are shown for the brachiopod faunules of Yukon Terr�tory and New Zealand (Fig.2,3). A further example may be drawn from the descriptiOn of faunas and floras from central Afghanistan (Fig.4) by Termier et al. ( 1 974),where a moderately complete succession displays marked and episodic changes in the faunas, changes which appear to coincide with those of the Canadian and New Zealand s�ccessions. Termier et al. ( 1974) concluded that the faunal changes were to constdera�le extent climatically induced, as also hypothesised for the New Zealand and Cana�tan chang�s (� �ter�ouse, 1 963 ; Bamber & Waterhouse, 1 97 1 ); Once again,there ts a sugges�lVe stmtla�tty between biozones which appear to have been climaticallycontrolled� and btomes, whic? are definitely climatically controlled, (Krassilov, 1 974), or more parttcularly, geo_graphtc subdivisions of a biome, that is, provinces.

Correlation of Biozones 5
CORRELA TION OF BIOZONES
Succession
. The abrupt changes in the fossil record which provide boundaries for biozones remfo�ced by th� recurrent nat�re of major faunal assemblages (Waterhouse, 1 9 73c): estabh�h �he bas�s for �orld wide correlation. In correlating strata and faunas primeval�e hes m stratigraphic succession, as also emphasized by Cooper & Grant { 1973) in their discussion of the Permian in west Texas. Degrees of freedom, and chance or error are greater in fitt�ng a single faunule into a standard sequence, than in matching a compl�te sequence With the standard. Let the standard sequence S1 contain the sequential umts:
a., b., c., d., e., and f • .From . faunal anal�sis, �1 differs from b1 , and from stratigraphic {and rarelyevoluttonary) analysis, a1 IS older than b1• Let the sequence S2 be correlated contain the sequential units :
�. b2, c2, d2, e2. An established correlation, i.e. faunal similarity between b1 and b2 severely restricts possibilities for remaining correlations, for � must now be either older than a1, as old as a1, or perhaps as old as b2. For correlating c2, the choices are narrowed to six (b1 ; c1; d1; e1; f1 ; or younger). If each faunule differs, and has upper and lower age limits, the choice is further restricted. And if b1 = b2, and d1 = d2, it becomes easy to correlate all five units.
It is likely in many sequential correlations that some faunas may be more positively correlated than others. Some horizons that do not share key species, but share a similar relationship to horizons that do share key species, are deemed correlative. In following this practice, it is advisable to assess where possible a number of communities, for one outstanding community may have been distinguished from contemporaneous communities only by its ecologic rather than temporal parameters.
From inspection it is clear that there are two prime requisites of the standard sequence : 1 . that it be complete, 2. that it be in stratigraphic order.
The reason that Permian correlations have been so long delayed lies in the fact that, for more than a century, the world standard has not been complete, and that for the last decade, the sequence has been disordered because the Cyclolobus ammonoid zone has been placed too high in the column. As shown in Table 3 the Upper Permian has long been based on the Tatarian red beds, virtually useless for world correlation. The Tatarian equivalents in west Texas-New Mexico, mainly in the so-called "Ochoan Stage, are of little more use. An attempt to replace the gap with marine based sequences by ammonoid workers (Miller & Furnish, 1 940; Glenister & Furnish, 1 96 1 ; Furnish, 1 973) recognised three stages called Chhidruan, Amarassian, and Capitanian, for units that overlap to considerable degree, being each based on different and miscorrelated sections in the Salt Range, Timor, and New Mexico. Moreover the Chhidruan Stage was incorrectly placed above the so-called Araksian Stage instead of below it (Waterhouse, 1 972a, b; Stepanov, 1 973 ; Grant & Cooper, 1 973 ; Taraz, 1973; Termier et a/., 1 974; Kozur, 1 974).
Units
A fundamental questiQn lies in the nature of the units that we seek to correlate. It is widely understood that stages, as defined in Hedberg ( 1 972), are the basic chronostratigraphic unit, but in fact, stages are more or less arbitrary, convenient or historically based bundles of zones-both vertically and laterally. If we are to search for

6 Correlation Methods
Tab l e 3 .
Trea t i s e ,
" Europe" *
Ta tar ian (non-marine)
Kazanian
Kungurian
Ar t inskian
Sakmarian
Sources of difficulty a r i s ing from use of faul ty s tandard sec tions
Moore e t a l . 1965 . Furnish 1973 Present S tandard
"North American" + Anunonoidea Stage Sub stage
Gr iesbachian
Dora shamian Ogb inan
Changh s ingian Vedian Ochoan Chhidruan (No few marine ( out of order)
Bai.sa lian fos s i l s ) D j u l f ian
Araksian Urushtenian
Amarass ian (•Chhi druan) Chhidruan
Capi tanian Punjab ian (=Chhidruan Ka labagh ian) Kalabaghian
Sosnovian Wordian Kazanian Ka s inov ian
Guada lupian Kungurian Irenian
Road ian Filippovian
Leonardian -Leonardian Baigendz inian Kra snoufimian
Aktas tinian
Sterli tamakiar:
Wo lfcampian Sakmarian
Ta s t ubian
*
+
As s e l ian
As s e l ian
Europe , meaning wor ld s tandard
North America , meaning Texa s , and certainly not Canada or Ala ska
Sarginian
Aktas tinian
S ter li tamakian
Tas tubian -
Kc::.naian
Uska likian
Surenan

Co"elation of Biozones 7
m�re objec�ive means of e!fecting correlation, we look in the first place at biozones(Ft�.2,3). B10zon�s s.ummanse the faunal ranges, acmes, entries, and assemblages, all ofwhtch largely cm�ctde at least for the Permian Period. It is biozones that carry the burden of correlation. Stages are further removed from actual outcrops and fossils, and may serve as a convenient simplification and summary of the evidence based on zones. They are much less 'realistic' or 'actualistic' than zones because they often contain widely disparate zones. For instance the Asselian Stage includes three biozonal horizons referred herein to the Surenan, Uskalikian and Kurmaian Substages. The upper and lower faunas are rather similar, the intervening faunas very different. In the Kungurian Stage, the Filippovian Substage is impoverished, the following Nevolin horizon of the Irenian Substage is very richly fossiliferous and different, the following Elkin and Ufimian horizons are somewhat like the Filippovian. The groupings are somewhat arbitrary and have to be, because each zone differs so widely from preceding and succeeding zones.
Perhaps the main usefulness of stages is that they: 1, group disparate zones into a few units, readily understood by non-specialists, and readily mapped for geological surveys; and 2, incorporate barren rock, that often may be at least roughly correlated by its lithologic content, or by radiometry. In a very approximate way, zones scale off the days, and stages scale off the weeks for a period, which matches the month. This analogy expresses well the comparative accuracy.
Subzones and superzones, substages
There are some problems in the use of biozones, even leaving aside the question of exact relationship between communities and zones, discussed previously, with attendant difficulties offered by barren or faunally impoverished beds, and areas with deeply weathered or no outcrop. The prime difficulty centres around scale, or duration of a biozone. Although many sequences display a succession of assemblage zones, that could equally well be treated as range zones on the basis of a number of species, the pattern is frequently overprinted by more or less short-lived acme zones or even range zones of usually one or a very few species that often invade a number of communities simultaneously. Evidently a species has suddenly enjoyed a highly successful burst, and thus marks, at least locally, a most useful datum place. Now how are these 'bursts' to be treated? They correspond in most respects with zones. But the fact that it is sometimes hard to match these zones with comparable zones elsewhere would suggest that they were in some cases very short lived, and possibly of local extent only. Therefore such phenomena were classed as subzones, or 'zone within a zone' by Bamber & Waterhouse ( 1 97 1 ). This appears to be a realistic and pragmatic method of sustaining, instead of concealing the evidence. Because the subzone may be upgraded if correlative subzones are found to be extensive and significant, the terminology allows flexibility.
A second term that is mentioned widely in literature, yet which has found no formal definition or consistent application is 'superzone'. It is here suggested that superzone should be restricted chiefly to laterally contiguous and probably correlative biozones, characterised by different assemblages, acme zones and range zones, but linked by certain key species, which do not necessarily range for the full extent of the zone or superzone. This in other words is a step towards the abstract stage concept, still governed by faunal data, but going beyond actual species presence and absence to incorporate species and biozones believed to be contemporaneous. It is close especially to substage, which differs by incorporating barren rock, non-marine rock, and igneous rock. In the past superzones have sometimes been used as a term for very long ranging species but this has seldom been very useful for correlation, and is the same as the range zone. In the present study nineteen substages are recognised and widely correlated around the globe, corresponding approximately with the number of brachiopod super-

8 Correlation Methods
... Q) a. a. ::l
z � ! ID � 'C
'C (!j a: :::E ::l :::E
... G) � 0 ..J
z � lJi:: (/) z 1-a:
�
.!! 'C 'C z :::E�
a: � :::E lJi:: � ... (/) Q)
� 0 ..J
z � ::::i w (/) (/) �
1
I I I I I I I
I I I I I I I I I I
F O S S I L G R O U P
2
I I II I I I I I I I I I I 1 1 1 1 1 1 1 1 1 1 1 1 1 1I I I 1 I 1 1 1 1 I I 1 I I1 1 1 1 1 1 1 1 1 1 1 1 1 11 1 1 1 1 1 1 1 1 1 1 1 1 1I I I I I I I I I I I I I II I I I I 1 I I I I I
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 J I I I I I 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1
3
? ? I I
I l l I l l I l l 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1
4 5 6 7
I I I I I I I I I I I I I I
I l l
I l l
Fig. 4. Chart of species ranges for Permian at Wardak, central Afghanistan, compiled from Termier et al. (1 974). Note repeated influx of short-lived new or different species, and predominance of brachiopoda, with limited occurrences of Cephalopoda and Fusulinacea, as is usual for most Permian successions. 1. Algae ; 2. Foraminifera; 3. Bryozoa; 4. Scyphozoa; 5. Annelids; 6. Bivalvia; 7. Cephalopoda; 8. Gastropoda; 9. Brachiopoda; 10. Arthropoda. In addition some sixteen species of Anthozoa occur in the lower Murgabian (Kungurian approximately). lntercorrelations are somewhat more complicated than for Yukon and New Zealand, so that this chart is not completely correct. It is

8
... Q) Q. Q.
::J
z < - Q) ID -« , , " a: � ::J �
... G)
� 0 ..J
z I< � rJ) z
I
1-a: <
Q) , , z � <
a: < � � < ... rJ) Q)
� 0 ..J
z <
..J w rJ) rJ) <
Correlation of Biozones 9
F O S S I L G R O U P
ITTl l I I I I I 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 I I I I I I I I I 1 1 1 1 1 1 1 1 1
II I I I I I I I I I I I I I I I I I I I I I I I I I I
9
I I I I I I I I
I ' I I I I I I I I I I I I I I I I I I I I I IT 1 I I I I
I I I I I I I I I I I I I I 1 I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I
I I I I I I I I I I I I I I I I I I I I I I I I I
I ' I I I I I I I I I I I I I I I I I I I I I I I I . I I I I I I I I I I I I I I I I I I I I I I I I I
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I ! I l I I I I I I 1 I I I I I I I I I
1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 !
I I I I
i . . . I I I I I . . . . . I I I I I . . . . . ! ! ! ! ! I I I I I . . . . . I ! ! I !
10
I I I I I I I I
based solely on Termier et al. (1974). The ·Murgabian is probably ?Fillippovian, ?lrenian and Kazanian.
Fusulinacea and ammonoidea are more abundant nearby in the sections of Tezak (see Termier et al. ,
1972).

1 0 Correlation Methods
Kazan ian Stage
Kungurian Stage
0
Sosnovian
., o .•• ·
·.. Kali . • ..:o nova'� • · • · • · • · ·
· · · · · · · · · • ·
JJ;. o • • • • • • • •
Ufimian
I -... .. Ji. o. • • • Elkin
· · · · · · • · · · · · · · · . . . . . . . '\()
� · · • · · · · · · · · · · · · ·
Newlin
Iii. 0 · · · · · · · · · · · · ·
. . . Fili ppovian
10 20 30 40 Number of brachiopod species
Fig. 5. Diversity graph or "Faunal Signature" of brachiopod species from early Middle Permian sequences of Russian Platform - Urals (world standard) - dotted line - and New Zealand - solid line - showing similar and correlative signatures. Russian data based on studies summarised in Branson (1948), Likharev (1966), Chalishev (1966) and Zolotova et al. (1966). New Zealand data summarised from Waterhouse (1 973c). Correlations are supported by entry of key species, and succession.

Co"elation of Biozones I I
zones . . There are li�el� to be a few more still to be unravelled, perhaps between theKazaman and PunJabian Stages and within the Chhidruan Substage. Definition of the P�rmian superzones lies beyond the scope of the present study, for it is easier to dealWith substages, but the succession is generalized in Tables 49-52,and the concept underpins the correlations set out in this study.
Key species
The entry of key species that suddenly became prolific and enjoyed a short lived burst offers a most practical method for correlation. There are many such instances at generic and specific level amongst brachiopods. The entry (or re-entry) of the genera Kochiproductus and A ttenuatella at the base of the Permian Period, the incoming of certain species of Streptorhynchus, Cleiothyridina, Spiriferellina, and Dielasma in the Kungurian Stage, and of Cancrinelloides, Licharewia, and Permospirifer at the base of the Kazanian Stage enable extensive correlation. All of course are open to revision. The species that are useful for zonal correlation, to the level of substage, are wide-ranging tolerant forms, found in many communities, as either dominants or subordinates, or both. Others were briefly so preponderant numerically that they were prime species over a wide range of lithofacies.
Recurrent and alternating species and genera
On a pragmatic basis, it has been found that some genera appear widely for brief recurrent intervals-such as Attenuatella (Table 4). Studies are not yet advanced enough to decide whether one or several species were involved at any one) horizon, but the presence and absence of this genus provide several datum planes, especially for the Asselian and Sakmarian Stages. It may be doubted that this genus would everywhere mark exactly the same time, and it is assumed that the recurrent entry of the genus was probably limited to certain palaeogeographic realms, apparently of palaeotropical and especially palaeotemperate character, although this cannot be finally established until the occurrences in Australia come under good control.
In some instances, genera alternated for the part of a succession. For instance throughout the early Middle Permian, the finely costate genus Neochonetes alternated with more diversely ornamented allies Lissochonetes, Dyoros, and Chonetina (Table 5). The principal exception is offered by the Magnesian Limestone of England, which has Neochonetes and has been regarded as of Nevolin age. Perhaps high salinity affected the fauna, or perhaps the age is really Ufimian, as proposed by Likharev ( 1 959).
Faunal Signature
Throughout a succession zones vary in the number of taxa present, and by plotting the numbers a zig-zag curve is derived, (cf. Miloradovich, I 940) which could be called a faunal signature (Fig.5). This is clearly in part controlled by facies and sampling, but the relative diversity in different collections may be assessed by the formula :
no. of species present-uo. of shared species no. of shared species
As a control for collections from a longer time interval, the number of shared species may be replaced by the number in each collection of the ten most common genera present in the entire succession. Clearly lithofacies and other environmental parameters may have a strong influence on faunal signature. But environment itself changed through time, and thus recorded time.
The concept of faunal signature is in its infancy, but if it works, then departures from the typical signature should reflect local interference to diversity, caused ultimately by unsuitable facies, or other environmental parameters than can be investi.gated aft�radequate sampling. It is to be expected that the curves of the faunal signature m

Gla
ss
Yu
ko
n
an
d
Ko
lyma
R
. S
pi
tsb
er
gen
K
az
ha
kst
an
M
ong
ol
ia
T
ibe
t
Ca
rn
ian
A
us
tr
a-
Br
az
il
- I'-)
S
tan
da
rd
S
ub
st
ag
es
Hu
ec
o
Alp
s
lia
M
ts.
� S
ar
gin
ian
� �
S"
... c:;· :::s
Ak
tas
tin
ian
�
�
:; c �
St
er
li
tam
ak
ian
A
A
?A
?A
Ta
stu
bi
an
?
-
Ku
nna
ian
A
?
-A
A
A
Usk
al
ikia
n
?-
Su
re
na
n
A
?-
?A
A
A
A
Ta
bl
e 4
. S
umma
ry
of
occu
rren
ce o
f At
tenu
a te1
1a i
n a
numbe
r of
cor
rela
tive
hor
izon
s (A
)
and
abse
nce
from
int
erven
ing
hori
zons
. Oc
curr
ence
poo
rly
cont
roll
ed a
s ye
t
in E
ast
Aust
rali
a.
Braz
il.
and
Ruec
o an
d G1
ass
Mts.

?S
osn
ov
ian
Ka
lin
ov
ian
Ufi
mian
El
ki
n
Ne
vo
li
n
Fi
lip
pov
ian
Ru
ss
ian
Ko
lym
a
(0)
Gr
ee
nl
and
(G
) C
ana
di
an
We
st
C
al
if
orn
ia
S
ikh
ot
e
Sa
lt
R
ang
e
Pl
at
form
P
et
ch
or
a
No
va
ya
A
rc
tic
(A)
Al
ller
ic
as
(N
C)
Ali
n
(R)
Zem
lya
(N
Z)
Arc
hipe
lag
o
Eu
ro
pe
T
aim
yr
(T
) Sum
at
ra
Y
uko
n
(Y)
Liss
ocho
n-Li
s so-
Chon
eti
na
etes
clw
nete
s R
0
T
Neo
chon
-To
mqui
stia
0
Ne
ochon
ete6
Ne
ochon
etes
et
es
NZ
AY
?Neo
chon
-et
es
Chon
et-
Liss
ocho
n-in
eU
a
etes
Chon
et-
Liss
ocho
n-Ch
onet
ina
Liss
ocho
n-M
ea
de
P
eak
Li
ssoc
hon-
Chon
etin
a Ch
onet
ina
ine
l.'La
et
es
G
etes
AY
et
es
Liss
ocho
n-Ch
onet
ina
To:m
quis
tia
Liss
ocho
n-Ch
onet
in-
Liss
ochon
-Ch
onet
in-
etes
--
----
----
-et
es
eZ.Z.
a et
es
eZ.Z.
a N
C
?Dyo
xoos
G
NZ
A
Axoct
ocho
nete
s N
eoch
onet
es
To:m
quis
tia
Liss
ochon
-et
es
Chon
etin
a Ne
ocho
nete
s
Neoc
hon-
?Neo
chon
etes
N
eoch
cmete
s et
es
AY
Cho
ca
l -
--
-T
abl
e
5.
Co
rr
el
at
ion
s o
f e
ar
ly
Mid
dle
P
ermi
an
str
at
a w
ith
a
lte
rna
tin
g
oc
cur
re
nc
es
of
Neo
chon
etes
wi
tr
ot
he
r
chon
et
ids
.
Qu
ee
ns
lan
d
(Q)
New
Z
ea
lan
d
Neoc
honet
es
Neoc
hone
tes
Liss
ochon
e tea
Q
g � l:r
... �· ;::s � b::l c:;· �
0
;::s � ......
w

14 Correlation Methods
reflecting diversity may also convey data about biomass, and so help calibrate environment and time by reflecting changes in trophic resources. Little use of the concept has been made in the present study, other than to use a sudden change in diversity as a suggestion for more refined correlation, to be tested by other means.
Limits of correlation techniques for brachiopods
Some correlation techniques will not work for brachiopods. Few genera were so short-lived that their ranges can be reliably used in the way that ammonoid genera are used, so that correlation must be at a specific rather than generic level. There are some exceptions to this rule, though it will probably transpire that such short-lived genera were rather rare, and sometimes of limited distribution, including some palaeotropical brachiopods. Amongst examples of short-lived genera, Timaniella Barkhatova, member of the Spiriferellinae, is found in early Middle Permian beds of Timan, northern Can ada, Kamchatka, and perhaps northern Manchuria and central British Columbia.
First entries may also be unreliable. For instance, Cooper ( 1 957) correlated a sizable brachiopod faunule from central Oregon with the Wordian (Mid-Permian), largely, it would seem, on the basis of the presence of Productacean genus Muirwoodia Likharev, which entered the Texan sequences of the Glass Mountains in the Wordian. But Muirwoodia itself is found widely in the early Permian of the Soviet Union and Canada, and a number of specific brachiopod links strongly suggested that the Oregon faunas are Early Permian (Bamber & Waterhouse, 1 97 1 ), as confirmed by Fusulinacea (Wilde, 197 1). Campbell ( 1 965) stated that Maorielasma Waterhouse, a Terebratulid, was restricted to early Middle Permian faunas (chiefly Kungurian-Kazanian), but it is found in late Sakmarian faunas in New Zealand. Armstrong ( 1968) asserted that the Ambocoeliid genus Attenuatel/a Stehli was not known below the Artinskian. Yet the type species is Sakmarian, and the genus is most widespread and numerous in rocks of Asselian age.
The nature of error in these three examples differed in each instance. Cooper ( 1 957) was well aware that Muirwoodia ranged into older rocks in the Soviet Union, but preferred to make correlations with nearby sequences in North America. Subsequent studies have shown that the faunas belonged to different palaeogeographic realms. Armstrong ( 1 968), on the other hand, appears to have overlooked or misinterpreted various references, something that is easily excused in these days of burgeoning literature and inadequate tabulation of data. Campbell ( 1 965) thoroughly studied world literature, and his statement was correct on the basis of what was then known. Any statement on age carried the risk of being upset by later discoveries.
The supposed upper limit of a genus also has dangers. Grant ( 1 970, p. l i 1) placed correlative value on the disappearance of Waagenoconcha after the early Guadalupian (Wordian) in the Glass Mountains, but correlations by Grant & Cooper ( 1 973) clearly revised the concept and extended the upper range. On the other hand, as Grant ( 1970) noted, the Rhynchotetradidae, represented by Septacamera, does not appear to be known from any faunas younger than Kungurian. �erhaps we need to be cautious here, for the genus, which is easily confused with Stenoscismatids, may yet be found in younger rocks. Bamber & Waterhouse ( 1 9 7 1 ) and Waterhouse ( 1 972b) noted that the Overtoniidae largely disappeared at the end of the Sakmarian Stage in widely disparate geographic realms of North America from the Canadian Arctic Islands, Yukon, Canadian Rockies, to Texas. Yet a few younger occurrences were noted in that region (Waterhouse, 1972b), and the family appears to have persisted in the Urals and North Russia into the Baigendzinian Stage, with a few younger occurrences. The change was geographic i.e. provincial, rather than climatic or biomal. The Rhipidomellidae also disappeared or more accurately, diminished, early (Dagis & Ustritsky 1 973).
Faunal assemblages are easily misjudged without firm control of species. It is all

S T A G E
DORAS H AMIAN
DJULFIAN
PUNJAB IAN
K A Z A N I AN
K U NG U R I A N
BAIG E N DZINIAN
SA K MA R I A N
A S S E L I A N
ORENBUR GIAN K A S I MOV I A N
MOSCO V I A N
B A S H K I R I A N
Correlation of Biozones 1 5
S P I R I F E R E L L A E L I V I N A T I M A�I E L L A I ELtVA
L I N E A G E s ' Ill l . A B c D E
. I •P
mOnrJO 1cal ! inte:p/icata I J pars 1 ranae semiOvalis mex1cana
I '
IT1"cla
JJ cambodgiansis 1 kupangensis : : rajah ..._ I vthtana
• • . . I I . . I keilhavii I I I
austra IISICB . sco�ma "I votniskll: l '-1 ;I I scobm
polaris • -� OldB< I talbBIC '\ .
pseudo - ! waagani I tibet ana I I .I
pseudod'"
T"'
i i
. I
lsuppl- tovem anta
'I vasrovskti
sa ranee
I digna
salta" pseudosaranae I I : � ploskaiae I
mica J petit I' a:taica I I
BSIBIICB I I I
yukonensis I pnmoeva ll
. . . •
I .
llevtpltce l ....I I .
I PBJC�OICB
I . . I . . fbasc�kirica
' '"'+·. . . . 9/Bii&nSIS I I I I
te;unus
I Soviet sp
I
l ••ngo o l spec i e s
• ....... .
gydar . 80$/$ . . . . I I I I
I I I
ordinaria .I .I
0 1tbeta;jl • tenu1-• • sulcal a
,, ' I rbucharens1s _ 1 sulc,Jer . T"l, I
: monguye�sis
I
I herkeri : I testa 1
Icordiformis
Tocciden ta/is
ne,mengens,s
tschernyscttaw' II mtrJnon
lyra
Fig. 6. Range chart and interrelationships for the genera Spiriferella, Timaniella, and Elivina, based on examination of material and wide survey of literature, by Mrs J. Waddington and the writer in a study on the genera for the Geological Survey of Canada.

1 6 Correlation Methods
too easy to go on incorporating fauna after fauna into one huge or portmanteau assemblage zone on the basis of a few shared, perhaps long ranging species, or even to use different shared species in progressing up the column. It is the uncontrolled use of this technique that explains the enormous units recognised by Runnegar ( 1 969b) in the east Australia Permian, in which virtually the entire Permian has been subdivided into a mere five divisions. Runnegar ( 1969b) suggested that several New Zealand biozones in a Permian succession 4,000 to 5,000 m thick were of Kazanian age (Table 6) because some Kazanian-like species were present. To the writer the entries of many new genera and species, some specifically close to Chhidruan species, suggest a post-Kazanian age, confirmed by the stratigraphic position over Lepidolina and Yabeina of post-Kazanian (Kalabaghian) age. A still younger Permian fauna from New Zealand was included by Runnegar ( 1969b) in the same Kazanian Stage. It is indeed of Kazanian aspect generically, but the species differ, and, with stratigraphic position, demand a Late Permian or Vedian age. Allied faunas in Australia were included in the same all-embracing 'Kazanian'Stage (Runnegar & Ferguson, 1 969), incorporating faunas and rocks much younger than the type Kazanian of the world standard.
The difficulties and possibilities inherent in brachiopods may be assessed from the species range and evolutionary chart of Fig. 6. The genus Spiriferella has many species in at least five distinct lineages which have to be disentangled before any evolutionary pattern can be assembled, not an easy task because there are so many species, and still many gaps in the record, although these are being reduced rapidly by numerous Russian studies. Species lived through one to often three or four zones, a relatively long time for brachiopods, and prime correlation is better based on Productida. Moreover the genus was not world-wide, being rare in palaeotropical regions, and completely absent from the south polar waters of east Australia, the highly saline Zechstein Sea of Europe, and rare or absent from the Phosphoria Sea of north-west United States. Even so, this genus was diverse, and tolerated a wider range of lithotopes and temperature than any known Fusulinacean or ammonoid genus of Late Palaeozoic age. It is clear that the real difficulty in using brachiopod species lies not in the short or lengthy ranges or restricted distribution, but in the very large number of taxa to be resolved before identification can be ascertained. The huge number of brachiopod species have held up advances in systematics and correlation, in contrast to Ammonoidea and Fusulinacea, where progress has been aided by the small number of taxa involved, enabling ready mastery of morphological and chronological differences, through brilliant research. A further barrier to the use of brachiopods lies in the need for revision of much of the literature, in which specimens have been misidentified, or not described in modern terms. This is one of the chief facets underlying the temporary and tentative nature of the scheme here proposed for subdividing the Permian, because too many of the zones and stages are based on faunai assemblages and presence or absence of genera rather than on species ranges. But there is little alternative, pending revision of species from Pakistan, Himalayas, south east Asia, and the Urals.
Use of statistical techniques
So far no statistical treatment has proved satisfactory for making correlations, probably because species have not been used from restricted zones. Grant & Cooper ( 1 973) used cluster analyses of brachiopod genera from a few regions to support their correlations, but their scheme is unconvincing, because the clusters appear to indicate palaeogeographic affinities rather than chronologie control. Several of their clusters link faunas clearly in sequence from one region, rather than link correlative faunas from differen.t regions. Williams ( 1 973) more appropriately clustered genera to indicate faunalprovmces, and Waterhouse & Bonham-Carter ( 1 972) clustered families to indicate major assemblages, controlled principally by climatic rather than temporal factors, and probably corresponding with fossil biomes.

Rurmegar
Kazan ian
Stage
Correlation of Biozones 1 7
( 1969b) New Zealand Sequence Waterhouse 1963 , l97 3c , herein
Subs tage Stage
Lower Stephens Fm Aperispirifer ne lsonensis Zone , l50-2 00m Dorashamian
Waiua Fm Vedian 700m, barren
Grevi11e Fm Dur>vi Ueoaeras woodmani Baisa1ian Zone , SO Om
Djulfian Little Ben Ss etc .
Spinomartinia spinosa Urushtenian Zone , 30-200m
Spirifere lla zone Tramway Ss , Wooded Peak Chhidrua �
Lmst
Plekone l la multiaostata Zone , 20-2 00m
Punj abian
Lower Ag4 Lmst
Martiniopsis woodi Ka1abaghian
Zone , 20-100m
barren ? ? Upper Mangarewa Fm 0-1 70m
Middle Mangarewa Fm Terrakea braahythaerum Sosnovian Kazan ian
Zone , BSm
Lower Mangarewa Fm Eahinalosia ova Us Kalinovian
Zone , 150 m
Table 6 . Alternative correlations for New Zealand
Permian , by Runnegar ( l969b) and Waterhouse ( 19 6 3 ; 197 3c) .
Fm - Formation ; Lmst - Limestone ; Ss - S andstone

2. Review of Chronologie and Biostratigraphic Sub-divisions of the Marine Permian
The Permian Period as proposed by Murchison ( 1 84 1 ) embraced rocks and faunas in the Urals Mountains and Russian Platform, now chiefly referred to the Middle Permian; followed by terrestrial beds of the Tatarian Stage. Karpinsky ( 1 874) showed that the underlying 'Artian' or 'Artinskian' beds were of post-Carboniferous age, and in 1889 included them as well in the Permian System. During the twentieth century, attention focussed on the pre-Artinskian Schwagerina and Pseudofusulina beds, more recently called the Sakmarian Stage, (see Dunbar, 1 940) and these have come to be incorporated in the Permian System, although a number of Fusulinacean experts have maintained, in both Russia and China, that the Schwagerina horizon is Carboniferous.
In recent years palaeontologists of the Soviet Union have done much to clarify their own Permian sequence, by re-examining their successions and faunas in detail. The Sakmarian has been subdivided into a number of units, by Rauser-Chernossova and colleagues working on Fusulinacea, and by Ruzencev, studying Cephalopoda. The local Russian subdivisions, recognised by fauna and lithology, offer a standard for wideranging correlation, and it is a prime contention of this study that the subdivisions can be recognised anywhere in the world through detailed faunal analysis. The Middle Permian has received attention from brachiopod workers such as Gerassimov ( 1 952a, b, 1 956) and Grigorieva ( 1 962) and stratigraphers such as Zolotova et a/. ( 1 966), with fine studies on Ostracoda and Foraminifera. Both Early and Middle Permian faunas have been extensively and well studied in Siberia by many palaeontologists. Because the Kungurian Stage has few Fusulinacea, or Ammonoidea, and the Kazanian Stage has none, a number of North American authorities have tried to transfer the world standard to west Texas and New Mexico, and made very grave miscorrelations as a consequence. Understandably, Australian palaeontologists such as Dickins ( 1956) and Campbell ( 1 953, 1 965) who have had to rely on Brachiopoda and Bivalvia have correlated their faunas more successfully. In truth, the Kungurian and Kazanian Stages have had their faunas fairly fully described, and there should be little difficulty in understanding and correlating the middle part of the Permian Period.
The Late Permian Period has been studied principally in Armenia, and across the border in Iran (Ruzencev & Sarytcheva, 1 965 ; Stepanov et a/. 1 969). However the Late Permian has been a considerable source of difficulty and controversy during the last thirty years. Much of the reason lies in the terrestrial nature of the standard Tatarian Stage, making it very difficult to correlate with marine sequences. An alternative standard, used in the United States, has fossils slightly higher in the column than in the Russian Platform, but also is unsatisfactory. During the late 1 930's and early 1 940's, a quite erroneous view of the Late Permian prevailed, in which a single stage was recognised, on the basis of the Fusulinacean Yabeina (Thompson, 1 946) and correlative ammonoid Cyclolobus (Miller & Furnish, 1 940). Fusulinacean experts soon recognised that Yabeina-Lepidolina beds were not of Late Permian age, ao; especially well demonstrated by Chinese sequences (Sheng, 1 964). But one school of thought has campaigned to retain Cyclolobus as a key to the top of the Permian (Ruzencev & Sarytcheva, 1 965 ; Stepanov et a/., 1 969 ; Glenister & Furnish, 1 96 1 ; Furnish, 1966, 1 97 3 ; Kummel & Teichert, 1 964, 1 970; Nakazawa et a/., 1 970), even though Spath ( 1 934, p.24) and Schenk et a/. ( 1 94 1 , p.2 1 97) had clearly indicated that beds and faunas of the �arne age as Cyclo/obus were overlain by younger pre-Triassic beds at Djulfa in Armema. The alleged young age for Cyclolobus was based primarily on the supposed

Relative Value of Certain Marine Fossil Groups 1 9
evolutionary developmen� o f an ammonoid lineage from Waagenoceras to Timorites to Cyclo/obus. This is largely theoretical, because the three have never been found in any one sequence, except at Timor. where Timorites co-exists with Cyclolobus. Grant ( 1 968), Grant & Cooper ( 1 973), and Waterhouse ( 1966, 1 972a, b) have contended that Cyclolobus was essentially correlative with Timorites, and that neither were of Late Permian age. Glenister & Furnish ( 1970) and Furnish ( 1970, 1 973) even appear to have inverted the sequence, to retain Cyc/olobus as the key to the Late Permian. Grant & Cooper ( 1973) criticised the scheme in detail for the Salt Range and west Texas, and Waterhouse ( 1 972a) demonstrated that Cyclolobus and Timorites could have evolved from different lineages at the same time, and occupied different palaeogeographic realms in the Late Middle Permian. Other authors have also not accepted the inversion of the sequence (e.g. Stepanov, 1973 ; Dickins in Grant & Cooper, 1 973, p.579; Taraz, 1 973 ; Termier et al. 1 974; Kozur, 1 974).
A second related problem concerned the top of the Permian System. Ruzencev & Sarytcheva ( 1 965) referred Permian-type ammonoids and brachiopods from Armenia to the 'Eotriassic', and Stepanov et a/. ( 1 969) even included as Eotriassic an underlying zone with Comelicania and Janiceps, clearly correlative with the Late Permian Bellerophon Limestone of the Carnian Alps in Austria and Italy. This view was opposed by Waterhouse ( I 967a, 1 972a, b, etc.) on the basis of brachiopods. Although Grant ( 1 970) at first accepted the Eotriassic age, it was put aside by Grant & Cooper ( 1 973). The conclusions based on brachiopods were anticipated by Yakovlev ( 1 93 1 ) and by Spath ( I 934). Rostovtsev & Azaryan ( 1 973) demonstrated that the controversial 'Eotriassic' beds were overlain by beds with the 'Early Triassic' ammonoids Ophiceras and Gyronites, and agreed that the underlying beds were Late Permian in age.
Further confusion concerns the top of the Permian System. Diener ( 1 909) defined the basal Triassic faunas primarily by the absence of the Productida so characteristic of the Permian Period. The first Triassic faunas, he believed, were typified by the ammonoids Otoceras and Ophiceras, with the bivalve Claraia, and supposed Norella, a Triassic brachiopod. However Norella is very like the mid-Permian genus Boloria Grunt in appearance and the writer found Norella-like forms in ?Smithian, not Griesbachian, faunas of Nepal. Nevertheless Diener's definition if not his application was sound, for Chao ( 1 966), Tozer ( 1 969, 1 973) afiel Waterhouse ( l967a, 1 972a, b) used the presence of Permian-type brachiopods in the so-called Eotriassic of Armenia to date those rocks as Permian, and Grant & Cooper ( 1973) followed a similar approach in fixing the Permian-Triassic boundary in Kashmir. It has now been established that Permian-type brachiopt>ds, including Productida, occur higher than realised by Diener ( 1 909), certainly with Otoceras, and possibly with Ophiceras (Waterhouse, 1 973b). These brachiopods have not been reworked, but lived in place with Otoceras ·and accompanying 'Triassic' ammonoids. Moreover, as similar faunas are found all over the globe, there is no question of them being pockets of late survivors that somehow survived an earlier major catastrophe that wiped out most forms of Permian life, an unsubstantiated thesis reiterated by Kummel and Teichert since 1964. The 'final catastrophe', if there was one, post-dated the Griesbachian Stage or Substage. Does this mean we have to redraw the boundary, or change the definition? Kozur ( 1973a, b), Newell ( 1973) and Waterhouse ( 1973b) have argued for placing the Permian-Triassic boundary within or above the Griesbachian, and international discussion is required to decide whether the boundary should be moved, or retained.

20 Biostratigraphic Subdivisions
RELATIVE VAL UE OF CERTAIN MARINE FOSSIL GROUPS
The primacy of Brachiopoda
The responsibility of correlation for Permian marine faunas has fallen in t�e past principally on Fusulinacea and Ammonoidea. But the present study allots prime value to Brachiopoda, and this shift in emphasis needs to be explained. The overriding reason for using Brachiopoda is because they were the predominant marine macrofossil of the Permian Period, and therefore can be used to correlate more rocks, on a wider basis. Against an ill-informed but widely reiterated misapprehension, brachiopod species and genera, during the Permian Period, were less latitudinally or facies-controlled than Fusulinacea or Ammonoidea, and just as short-lived. Severe problems remain. They were so diverse a group that mastery on a world-wide basis is difficult, if not impossible provided we persist in our present techniques, because systematic, communal and zonal (i.e. ecologic and temporal) studies are all required before their potential can be realised. At present the most critical short-coming lies in the failure of palaeontologists to reexamine the Early Permian brachiopod faunules of the Urals. If the standard sections are to be in the Urals, as argued herein, it is the duty of the host country to provide adequate and up modern studies of Brachiopoda. Until that is done, some questions of correlation remain insoluble.
Fusulinacea and nomenclatural problems
In Permian carbonates of relatively low palaeolatitudes, Fusulinacea are more numerous, and more useful than even Brachiopoda, but they are severely restricted in distribution, with none known from vast tracts of Permian marine rocks, including those of the entire continent of Australia. It is therefore impossible to use them for world-wide correlation. Moreover, in many regions, such as Arctic Canada, Afghanistan, New Zealand, to mention a few, they are present at only a few intervals. Additional problems that beset the stratigrapher lie in the lack of knowledge of communal associations, and the response of species to various ecologic and biologic parameters has scarcely been assessed. Consequently we understand little of the apparent wide differences of species ranges in different parts of the world, and the marked provincialism set out by Ross ( 1 967b).
A frustrating nomenclatural tangle bedevils our appreciation of key Fusuline genera of the Late Carboniferous and Early Permian, with baffling effects on correlation and boundary problems. This concerns the three genera Schwagerina Moeller 1877, Pseudoschwagerina Dunbar & Skinner 1 936, and Pseudofusulina Dunbar & Skinner 193 1 . According to Kahler & Kahler ( 1 966), Schwagerina was initially based on shells mistakenly identified as Borelia princeps Ehrenburg. Russian authorities have granted primacy to the intention of Moeller, and retained the definition, by renaming Moeller's specimens as the type species Schwagerina moelleri Rauser-Chernossova. American authorities have stressed the citation of the type species, and centred their understanding of Schwagerina on Borelia princeps Ehrenburg. Pseudoschwagerina Dunbar and Skinner 1936, type species Schwagerina uddeni Beede & Kniker was erected for shells long considered by the Russians to be true Schwagerina. Now the two schools persist side by side, with the ridiculous situation that a Russian Schwagerina is not an American Schwagerina. Throughout this text Schwagerina as used by the Russians is accompanied by an asterisk, following Kahler & Kahler ( 1 966).
Pseudofusulina is just as confusing. Kahler & Kahler have discriminated a number
.of �pecies groups, those of uncertain standing, those as understood by the Soviet school, mdtcated throughout the text as Pseudofusulina*, a group as used by the Russians, and by Thompson ( 1948), shown herein as !Pseudofusulina* and the group Pseudofusulina

/
"' 40°
_._
_ I
20°-
----
--+--
I '
0" ·-
---
· A/
at
e
T A
• Fu
1ulln
ac-
41•
5%
a
• B
rach
iopoda
90
7. C
• A
mm
on
oidea
39'/.
T
• To
tal MI
LES
0 20
00
0 30
00
KILO
MET
RES
.....-16
5° 12
0°
I I --
t--
--
1 I I -
--
--t
- 1 I
60°
0°
so•
''·"'-,
OhJ/2•1 Oj 'l!
2 o " 12
•1
i
!ot:t2
II
---�/
,-
-;:? /' -
40°
�
//
/15
/
/
/
1800
60
" _1
65°
Fig
. 7.
P
rovi
sio
nal
est
imat
es o
f n
um
ber
of
sub
stag
es r
epre
sen
ted
fau
nal
ly i
n v
ario
us
regi
on
s, w
ith
est
imat
es o
f n
um
ber
of
sub
stage
s b
eari
ng
Fu
suli
nac
ea,
Bra
chio
p
od
a, a
nd
A
mm
on
oid
ea.
Th
e ch
art
om
its
seve
ral
sign
ifican
t fu
ll se
qu
ence
s o
f F
usu
lin
acea
, su
ch a
s th
at o
f th
e M
cLo
ud
Lim
esto
ne
of
Cal
ifor
nia
, an
d m
ay u
nd
er
esti
mat
e th
e n
um
ber
of
Fu
suli
nac
ean
zo
nes
fo
r A
rcti
c C
anad
a,
Pam
irs,
Ch
ina
and
Jap
an.
It a
lso
fai
ls t
o s
ho
w t
hat
Fu
suli
nac
ea w
her
e p
rese
nt
wer
e ge
ner
ally
ab
un
dan
t, a
nd
so
pro
vid
e a
goo
d i
nd
icat
ion
of
age.
Bra
chio
po
da
clear
ly w
ere
of
maj
or
sign
ific
ance
in
Per
mia
n s
equ
ence
s as
sh
ow
n b
y t
his
ch
art,
an
d t
he
nu
mb
er
is u
nd
eres
tim
ated
fo
r se
vera
l se
qu
ence
s w
her
e th
ey h
ave
bee
n p
rese
nt,
or
pro
bab
ly p
rese
nt,
bu
t n
ot
des
crib
ed, a
s in
Arc
tic
Can
ada,
Arm
enia
, P
amir
s an
d J
apan
, an
d
pro
bab
ly C
hin
a.
Am
mo
no
idea
are
so
mew
hat
ex
agge
rate
d i
n s
ign
ifica
nce
, b
ecau
se t
hey
wer
e co
un
ted
eve
n w
her
e ra
re -
and
gai
n b
ecau
se t
he
Gri
esb
ach
ian
-b
asal
D
ien
eria
n i
s in
clu
ded
, wh
en a
mm
on
oid
s fo
r th
e fu
st t
ime
bec
ame
wid
esp
read
en
ou
gh t
o a
llo
w w
orl
d-w
ide
corr
elat
ion
. N
um
ber
of
sub
stag
es w
ith
Fu
suli
nac
ea 4
1.5%
; nu
mb
er o
f su
bst
ages
wit
h B
rach
iop
od
a 90
%; n
um
ber
of
sub
stag
es wi
th A
mm
on
oid
ea 3
9%.
To
tal
nu
mbe
r o
f su
bst
ages
with
marin
e in
vert
ebra
te f
aun
as:
265.
�
� S" ... �- �
� E" � � � ... � s· � ... s· � � t: ::::.: C':l � 1:: �
... IV .....

22 Biostratigraphic Subdivisions
as understood by Thompson ( 1 948). Throughout this text the nomenclature of Fusulinacea is matched where possible to
Kahler & Kahler ( 1 966).
The value and limitation of Ammonoidea
Students of ammonoids have long shared the lead in subdividing the Permian Period on a world-wide basis. Of recently proposed classifications one of the most outstanding in applicability and fineness of control is that proposed by Furnish (1973), building on the excellent studies by a number of American and Russian experts. But ammonoids cannot be allotted primacy of place as an aid for correlation, because they are too rare in too many sequences. This comparative rarity may be objectively established from basic data provided by Furnish ( 1973, fig. 1 , reproduced herein as Table 10), which shows how the twelve so-called ammonoid stages are based on only fifteen regional sequences around the entire world. Fifteen would hardly seem enough, but indeed, to reach even that modest score, the Glass Mountains and adjoining Delaware Basin were counted separately, and the nearby mid continental states separately again. Timor was included even though it offers no stratigraphic sequence. Even sequences in the Basin and Range Province (with only two scattered stages), western Canada (two), Mediterranean (three widely scattered), and the huge region of eastern Australia (three, widely scattered) were counted as offering serious evidence. One ammonoid succession was tabulated in the 'Himalayan System', perhaps referring to the Salt Range, where ammonoids of pre-Kalabagh age are scarce, and not significant. The Himalayas themselves include one or perhaps two ammonoid zones, with a further Griesbachian horizon. · Published literature on 'Arctic America' shows a modest sequence of ammonoid zones that depend heavily on other fossils for local intercorrelation : few stratigraphic sections show a succession of ammonoid zones, so far as is known. Thus the 'valuable regions' showing a mere three or more ammonoid faunules in some sort of stratigraphic sequence are about eight in number (Mexico, United States, Armenia, Pamirs, Western Australia, Urals, Verchoyan Mts., China), and perhaps the Canadian Arctic Archipelago should now be included. They do indeed provide a most valuable scheme, in spite of the scarcity of ammonoids, thanks to the excellent ammonoid studies. By contrast, as outlined in the present work, the following areas contain three or more brachiopod faunas in sequence (see Fig. 7).
Americas, Arctic: Glass Mts., and Delaware Basin, Midcontinental U.S.A., Coahuila, El Antimonio, Canadian Arctic Archipelago, western Canada (eastern Rockies), Yukon, Alaska, Alaska Panhandle, Spitsbergen-Bear Island.
USSR: Urals, Russian Platform, Petchora, Pai Hoi, Timan, Taimyr Peninsula, Verchoyan Mountains, Kolyma River-Omolon Plateau, Fergana-Pamirs, Donetz Basin, Greater Caucasus, Crimea, Armenia-North Iran.
Asia: . Ce.ntral Iran, Elburz Mountains, Salt Range (Pakistan), Kashmir, Thailand,Cambodia-VIetnam-Laos, South China, Central China, North China, Japan.
Ausfralasia: Western Australia (Carnarvon, Bonaparte Gulf, Perth, Canning Bas�ns), Queen�land (�owen, Yarrol, Gympie Basins), New South Wales (SydneyBasm), Tasmama, south and north South Island, New Zealand.
Regions such �s Novaya Zemlya, Nepal, Baikal, Ussuriland, Mongolia, and Karakorun;t are om1tted because although sections are known, they have not been resolved, e1ther. because work !s in preparation, or was done so long ago that it has dated badly. T1mor, though w1th large brachiopod faunas, should not be counted.

Relative Value of Certain Marine Fossil Groups 23
Regi�ns of equal s�ze, say 1 5° latitude and 15° longitude (at the equator), provide t?� followmg comp�ra�tve number of sequences with three or more abundantly fossthferous substages m mdisputable sequence.
Brachiopoda Mexico - United States Rocky Mts Alaska - Yukon Canadian Arctic Archipelago Spitsbergen, North Greenland, Petchora
Novaya Zemlya Peru - Bolivia Urals Europe (Austria, Italy, Yugoslavia) Armenia - Iran Pamirs, Salt Range, Kashmir Nepal, Burma Taimyr Lena and Kolyma Rivers Japan, Korea, Ussuriland Lake Baikal, Mongolia China South-east Asia Timor, northwest Australia Southwest Australia Queensland Tasmania, New South Wales New Zealand Total : 22 regions
Ammonoidea Mexico - United States
Canadian Arctic Archipelago
Urals
Armenia - Iran Pamirs - Salt Range
Lena River
China
Timor, Western Australia
8 regions
Of these major regions, only central North America has more than six ammonoid zones in sequence, in one compact area, compared with more than twenty of the regions with five or more brachiopod zones in sequence. Moreover in each region, sections that have ammonoids are extremely few. Even in the Urals, richest of all Permian areas in terms of numbers of ammonoids, the material is found in scattered outcrops that often had to be correlated by means of Fusulinacea (Barkhatova, 1964). As Furnish noted, 'At most localities, in America at least, Cephalopods are generally too rare to provide a satisfactory guide' ( 1 973, p.528). The same holds true for most of the world.
Coahuila, north-west Mexico, is one of a few areas with ammonoids scattered through several zones in section. In the Salt Range, ammonoids are extremely rare (Grant & Cooper, 1 973), and cannot be used for even local correlation. In the closely studied region of west Texas, unusually rich in ammonoids, Dr R.E. Grant, (pers. comm.) has estimated that some 5,000 ammonoids have been recovered from 97 localities, compared with some 3,000,000 brachiopods from about 800 localities. In Yukon Territory, Arctic Canada, some 1 ,500 localities have been collected from 50 sections, by staff of the Geological Survey of Canada and the writer with his associates. Ammonoids have been found in five of those localities-that is 0.3%-scattered in two or three zones out of the eleven zones present. In New Zealand, five areas have been mapped in examining some fourteen brachiopod zones. Ammonoids have been found in only two areas, in one zone, at about 10 localities. In east Australia, Permian rocks with more than ten brachiopod zones are exposed in numerous sections, in several huge

24 Biostratigraphic Subdivisions
basins. Widely scattered few ammonoids are found within only three zones in some ten localities. A serious problem lies in the sparse occurrence of ammonoids in any one section. There are so many intervals without ammonoids, let alone diagnostic ammonoids, that only fragments of the section can be accurately dated, or zoned by ammonoids. The bulk of sections lies between zones, and boundaries are impossible to draw with accuracy. Interestingly enough, Furnish ( 1 973) commented that it would be possible to record some 300 occurrences of ammonoid families in various localities. In a massive compilation of Permian brachiopod families, we have just compressed the data from more than 50,000 occurrences (Waterhouse & Bonham-Carter, 1 975).
The results of using for prime correlauon a group ot tossils that are so rare may be illustrated for the Late Palaeozoic Permian successions of Queensland, eastern Australia. We could erect a sequence of some 9- 1 0 zones based on brachiopods, these clearly being the prime fossil for at least local correlation, as established for example by Maxwell ( 1954), Campbell ( 1 96 1 ), and Dear ( 1 972), •in contrast to the long ranges ascribed to some of the bivalves by Runnegar ( 1969a). If world correlation is to be based on ammonoids, only two horizons can be ascribed an international stage or substage, and of course these lack any supporting evidence, so that there can be no possibility of falsifying or verifying the assumptions. (It may be added that the ages ascribed to the ammonoids by Armstrong et al., 1 967 are almost certainly false, when other evidence is considered). But by refusing to consider other evidence, some degree of superficial consistency is attained. The procedure of relying solely on Ammonoidea amounts basically to an unproved and unadmitted assertion that world-wide correlation by using benthonic fossils is intrinsically impossible, except perhaps at the scale of a period.
Correlation potential
The relative usefulness of ammonoids and brachiopods may be assessed by ratio of correlation potential (cp).
Zi cp = -s
where Zi = number of zones in sequence i with given group of fossils; and S = total number of fossiliferous zones in a succession.
As summarized in Fig. 7, the world correlation potential for Fusulinacea is only 0.4 1 ; and for Ammonoidea only 0.39 ; and for Brachiopoda, 0.9. For local sequences, the correlation potential for Ammonoidea is generally very much lower, insofar as ammonoids may be found in only a few localities, compared with hundreds of localities with Fusulinacea or Brachiopoda.
For instance, in the Permian zones of the Ogilvie Mountains, Yukon Territory, as set out previously, Ammonoidea and Fusulinacea are found in two of eleven zones, yielding a correlation potential of 0. 1 8, compared with a potential of 1 .0 for Brachiopoda. If we consider the number of macrofossil localities for the entire northern Yukon Territory, the correlation potential for ammonoids falls to 0.003, compared with a ratio of 0.99 for Brachiopoda.
Correlation value
The correlation value (cv), may be assessed as a percentage from the formula:
cv

Relative Value of Certain Marine Fossil Groups 25
where �z = total number �f Permian zones, and Ri = range of given fossil group. The correlation value of brachiopod genera is relatively low : say _ [0 9 x ( 1 9)] .
1 9 19 X 100 = 4.7%
Whereas the correlation for an ammonoid genus may in favourable instances be :
r 39 X ( 1 9)] t" 1 9
3 X I 00 = 13%
But the average brachiopod species ranges from one to four zones with an average correlation potential of ' 40-50%and an optimum value up to 90% Compare this with an average correlation potential for ammonoid species of some : 20% and optimum value for short-lived ammonoid species of 39%
In most sequences, the correlation value for ammonoids is well below 10% com-pared with well over 50% for Brachiopoda. ' Risk of error
If correlation potential (cp) is low the chance of error. increases, as expressed by the concept 'risk of error' (E)
E = ( 1 - cp) x 100I
If, for example, we have as in Queensland, Australia, only two ammonoid biozones in a sequence of at least eleven biozones, the risk of error
E = (1 - _1) X IOO = 82%
1 1 I This appears to be too high to be acceptable under normal standards of correlation accuracy.
With such a low correlation potential and high risk of error, it must be asked why ammonoids have proved so useful in the past for correlation. The answer probably lies in the brilliance of research by ammonoid experts, and especially in their willingness to undertake world-wide correlation, as opposed to more regional correlations attempted by other palaeontologists.
Conodont studies
Conodont studies such as those by Sweet ( 1 970a, b, 1973), Kozur ( 1973a), and Clark & Behnken ( 1 97 1) have on the whole conformed to the correlation scheme outlined by Glenister & Furnish (e.g. 196 1), and so have been somewhat superseded by the more refined scheme offered by Furnish ( 1 973). As emphasised by Clark & Behnken ( 197 1 , p.4 1 7), 'the Permian System represents the last frontier for conodont research' and this modest appraisal would appear to be well justified in view of the critical reassessments of Late Permian and Early Triassic conodont correlations of Sweet ( 1 970a, b, 1 973) by Kozur ( 1 973a, b, 1 974), Newell ( 1973), and Grant & Cooper (1 973). The chief problem appears to lie in the newness of the work. So few sequences have been available for cross-comparison that data have naturally been adjusted to what was understood to be the current world classification of the Permian Period.

26 Biostratigraphic Subdivisions
The conodont zones as set out by Clark & Behnken ( 1 97 1 ) are reproduced in Table7, and may be discussed in turn. .
The zone placed at the start of the Permtan by Clark & Behnken (1971), withIdiognathodus ellisoni, is now considered to be Carboniferous (Dr H. Kozur, Freiberg, pers. comm.). The overlying Sweetognathodus whitei zone was too broadly defined, and includes three species, of which whitei is restricted to the Asselian. Younger forms not yet investigated occur in the Sakmarian Stage. (Dr H. Kozur, pers. comm.). According to Dr Kozur (in litt. I May, 1 975), Neostreptognathodus sulcoplicatus is in fact restricted to the Roadian (which I assume includes Meade Peak, that is, Nevolin rather than Filippovian) but the genus Neostreptognathodus occurs in the Leonardian (Baigendzinian) and in the Roadian, as well as the 'Artinskian' of the Cis-Ural. Gondolella occurs in a different facies, and G. idahoensis is found in both 'Leonardian' and 'Roadian'.
The supposedly following-zone of Neogondolella serrata is reported from the Bone Springs Formation, Leonardian according to Clark & Ethington ( 1962), 'basal Wordian or Guadalupian' according to Clark & Behnken ( 197 1 ), and Sweet (1973), presumably implying a roughly Roadian (Filippovian) age as confirmed to me by DrR.E. Grant. It thus almost certainly commenced at the same time as the Gondolella idahoensis Zone, and the sequence appears to have been reversed by Clark & Behnken ( 1 97 1). Sweet ( I 973, p.64 1) found Neogondolella serrata (Clark & Ethington) throughout the Word, and into the Capitan as high as the mid-Capitan Rader Limestone in the type Guadalupian, but Dr Kozur (in litt. May 1 97 5) considers this may be a distinctive subspecies. Contemporaneous faunas of western United States had different species that failed to enter the Glass Mountains and Delaware Basin (Sweet, 1973).
The name species of the supposedly overlying Merrillina arcucristatus Zone occurs in the Plympton and lower Gerster beds of Nevada and Utah. Although correlated with the high Word and lower Capitan by Clark & Behnken ( 1 97 1 , p.427), these beds are probably very little younger than the Meade Peak Shale. Kozur ( 1975) has synonymised arcucristatus with galeatus Bender & Stoppel from the Kazanian of Sicily. It is clearly coeval with serrata s.l.
Fortunately the following zones occur in a sequence of faunas, although even in these zones, ranges of critical species overlap considerably, including even the name species. The supposed Capitan succession of conodonts commences with the Gondolella n.sp. previously of the 'Gondole/la rosenkrantzi' Assemblage Zqne, found not in the type Capitan of New Mexico but in the upper Gerster Formation of Nevada, with a socalled Punctospirifer pulchra (Meek) fauna. Leaving aside dubious or unsubstantiated reports, this fauna is found in the upper Phosphoria beds and Ervay and Franson Members of the Park City Formation of Idaho, Utah, and Wyoming, just above a Filippovian-Nevolin ammonoid-brachiopod assemblage, and is assessed as being Nevolin to Kazanian age, in other words, Wordian. Dr G.A. Cooper has correlated the Gerster brachiopods with those of the Appel Ranch Member of the type Word (i.e. Sosnovian) in the Glass Mountains, Texas (Cooper in Bissel, 1 973, p.324). But the conodont workers have assumed a lower Capitanian age. Other worJd (aunas with some of the sam� conodonts are also Kungurian-Kazanian, i.e. Wordian, coming from the lower Zechstem and Greenland, (both Kungurian as here defined, to include Ufimian) and Sosio beds of Sicily (Kazanian). Disturbingly, the supposed key species G. rozenkrantzi Bender & Stoppel is known from the underlying Neospathodus arcucristatus Clark & Behnken (Merrillina galeatus) Fauna as well, so that the zones are ill-defined, to say the least. However, according to Dr Kozur, various so-called rozenkrantzi need to be reassigned .
. M_errillina divergens (Bender & Stoppel) characterises the succeeding zone, but
agam, IS stated to overlap the underlying zone. It is ascribed, with no independent sup-

Wo
rld
st
an
da
rd
Ch
hi
dr
ua
n
I
Ca
pit
ani
an
=K
aza
n i
an
Ar
tin
skian
Sak
mar
ian
---
-
Te
xa
n
sta
nd
ar
d
Ch
hid
ru
an
� H llo
Cap
ita
nian
� � 0
Wo
rd
ian
{..'
Le
on
ar
dian
Wo
lfc
amp
ian
Cla
rk
&
Beh
nk
en
, 1
97
1
Co
no
do
nt
z
on
es
Anc
hi gn
at
ho
du
s F
aun
a
�-
--
--
--
--
--
--
--
--
--
--
--
-M
err
il
lin
a d
ive
rg
en
s F
au
na
f--
--
--
--
--
(ov
er
lap
)--
--
--
--
--
-G
on
do
le
ll
a n
. sp
. =
so
-a
all
ed
"r
ose
nk
ra
nt
zi
...
Ass
emb
lag
e
Zon
e
f--
--
--
--
--
--
--
--
--
--
--
--
-M
er
ri
lli
na
ar
cu
cr
ist
at
us
=
ga
lea
tu
s F
aun
a
--
--
--
--
--
--
--
--
--
--
--
--
Ne
ogo
nd
ol
el
la
ser
ra
ta
Fa
una
f--
--
--
--
--
(ov
er
lap
)--
--
--
--
--
-
Go
nd
ol
el
la
id
ah
oe
ns
is
Fau
na
--
--
--
--
-Co
ver
la
p) -
--
--
--
--
--
Ne
or
epto
gn
at
ho
du
s su
lcop
lic
at
us
Unit
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
--
Go
nd
ol
el
la
bi
ss
el
li
-s
we
et
ogn
at
ho
du
s w
hit
ei
Ass
emb
lag
e
Zon
e
�-
--
--
--
--
--
--
--
--
--
--
--
-Id
iogn
at
ho
du
s e
ll
iso
ni
Ass
emb
lag
e
Zon
e
�-
--
--
--
--
--
--
--
--
--
--
--
-
---
----
--L....--
Su
gg
est
ed
co
rr
el
at
ion
Gr
ie
sba
ch
ian
-
Ch
hid
ru
an
? K
az
ani
an
Ka
zan
ia
n
Ka
za
nia
n-K
ung
ur
ian
Ka
zan
ian
t
o
Ba
ig
en
dzi
ni
an
Up
pe
r
Ba
ige
nd
zin
ian
?N
ev
oli
n
Ne
vo
lin
Sak
mar
ian
As
se
lia
n
Ca
rb
on
if
er
ou
s
Tab
le
7.
C
on
od
on
t
zon
at
ion
, b
ase
d
pa
rt
ly
on
Cl
ark
&
Be
hn
ke
n
(19
71
, f
ig
. 4
),
eme
nd
ed
t
o
fo
ll
ow
the
ir
tex
t
(p.
42
7)
, a
nd
Sw
ee
t
et
a
l.
(19
71
, p
. 4
44
) •
i
�
11) S'
... �- 11) � � � g ...
�
;;· � ... ;;· 11) � ::::;
c;) � 1:
: � N
-.1
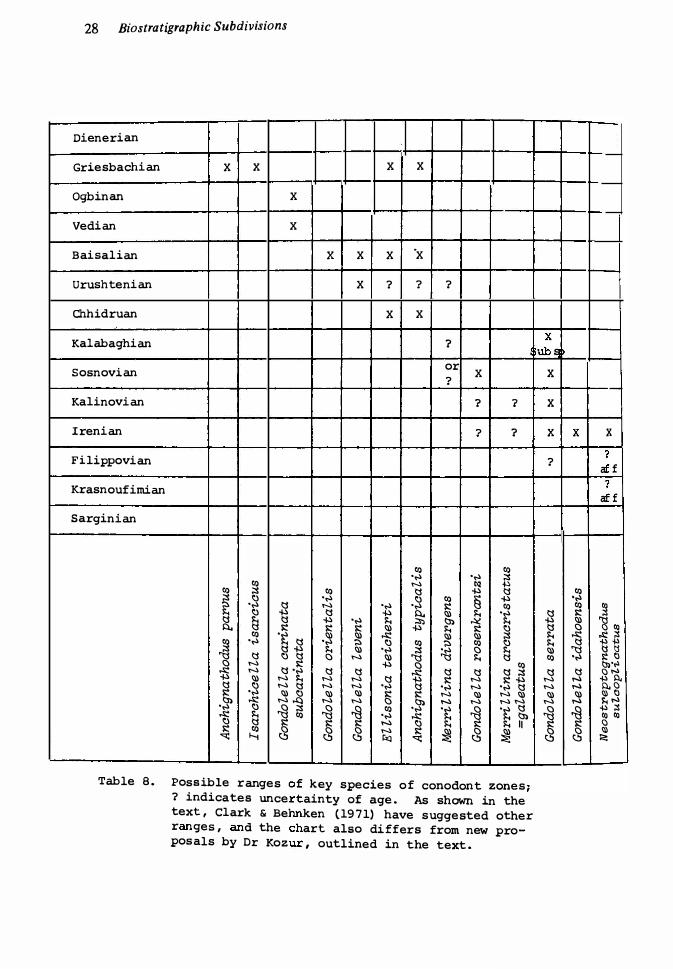
t-3 g. 1-'
CD
(X)
.
'011
rtoV
't!
0
Ill CD
0
Il
l p
� ..
... Ill
lll
o.Ort
::lll
l 1-'
CD
,
p. 1-'
· Il
l Il
l ..
... tr
,()
01-'
tr
1-'
Il
l CD
'<
Il
l Il
l rt
P
11
CD
11
�
o.:o�· m
g rt
R'>§ o.O
:;><:
::r
CD
0
CD
ll:J 0
Il
l N
CD
CD
�
0::1'
110
11
P'P
rtH1
'
Ill
�Il
l 11
CDI-'
·�
O
rtP
PCD
�
rt
'<
rt
Ill
...... '<
1-'
1-'1-'
Il
l 1-'
·ll
li.O
O'O
::
:10
-..J
H!CD
CD
1-'
0
p.
p.�
IIJI-'·
1-'
· o.Q
CD
1-'
·Hl
::TCD
Ill
p H1
Il
l •
CD
<
0
rt
11
CD
H1
::r
Ill
!!:' CD
Il
l Il
l 0
H1
�
0
rt
l1o.O
IIl::l
CD
0
o.Q
::T
0
>C!:I
CD�
P.
rt
Ill
0
• p
rt
::s
CD
CD
rt
� p.
.....
'0
0
p �
11rt
rt::S
0
::1'
::1' CD
I
CD
CD
Ill
11
An
chig
na
thod
us
pa
rvu
s
Isa
rc
hic
eZZ
a i
sar
cic
us
Gon
do
ZeZ
Za c
ar
ina
ta
sub
ca
rin
ata
Gon
do
ZeZ
Za o
Pie
nta
Zis
Gon
dbZe
ZZa
Ze
ven
i
EZ
Z.is
onia
t
ei
ch
ePti
An
ch
ign
ath
odu
s ty
pic
aZi
s
Me
PP
iZ.
Zin
a d
iveP
gen
s
Gon
do
ZeZ
Za P
ose
nk
Pan
tzi
Me
PP
iZ
Zin
a a
rcu
cPi
sta
tus
=g
aZe
atu
s
Gon
do
ZeZ
Za
se
PP
ata
Gon
db Z.
e Z.
Z.a i
da
ho
ensi
s
Neos
��ep
�ogn
a�ho
dus
suZ.c
opti.
cat;u
s
til ::><:
t'lj H
::><:
Ill
11
.....
11
Ill
11
Ill
1-'
CD
1-'
o.Q
Ill
.....
p..
... ..
... p
:g ..
... p
p 0
�
0
.....
�
0
<
� H1
<
..
... ..
... ..
... �
�-g
�
'"
""
""
""
'"
:>< :><
:<
PI
PI
.... ""
.... ""
:<
..
.. ..
..
til ::><:
9 c
b:l <:
0
Ill
11
Ill
CD
Ill
1-'
::r
�
.....
p.
p g.
.....
Ill
Ill
.....
0
p.
::r
Ill
� <
Il
l 11
rt
1-'
..
... '§.
�
CD
.....
� �
p �
.....
.....
� �
:><
:><
:>< :><
:>< '"
:><
:>< ' "
:><'
' "
0
""
'"
11
:>< :>< 6-
:-:
Ill
0 G)
'§. 11
.....
.....
CD
p Il
l g
tr
Ill g. ..... � :>< :><
:><
:>< :><
t:l
.....
CD p CD
11
.....
Ill p
I
N
00 t::l;, c;· "'
..
. i:! ....
�-
1:::! ::s- ;::;· C'J
I:: <:::!"
l::l.. � o;: · c;· ;:
:s "'

Discussion of Correlation Schemes 29
P.orting evid�n�e, to the l�te Capitanian by Clark & Behnken (1971) and Dr Kozur conSiders that 1t 1s Abadeh1an (Urushtenian). �ecent work by Kozur ( 1974) has greatly enhanced our understanding of laterPerm1an conodont zones : Conodont Assemblage Zone Ammonoid Zone Stage Anchignathodus parvus /sarcicella isarcicus Gondo/ella carinata
subcarinata Gondolella orienta/is Gondolella /eveni
Merrillina divergens
upper Ophiceras commune lower Ophiceras commune Phisonites-Paratiro/ites
Vedioceras ventroplanum up to top of Araxoceras
Ia tum
"Griesbachian" "Griesbachian" Dorashamian
Baisalian Baisalian
Urushtenian
Most conodont studies (e.g. Sweet, 1973 ; Kozur, 1 973a, b) have accepted and sought to establish a Late Permian age for the ammonoid Cyclolobus, and this erroneous assumption has probably undercut their preferred correlations. However new and refmed work by Kozur ( 1974) has reversed earlier conclusions, and now adds strong support for the position of Cyclolobus as adduced from brachiopod studies by Grant ( 1 970; Grant & Cooper, 1 973) and Waterhouse ( 1966, 1 972a). Some aspects of Kozur's present scheme still differ from the correlations discussed here, for he correlates the Chhidruan Substage and the lower Zechstein with the Urushtenian (Abadehian) Substage; the Kazanian Stage with the Capitanian; and the Sosio fauna with the Kungurian Stage. These are substantial differences, but discrepancies} will probably be overcome now that the true position of Cyclolobus has been recognised. Moreover, Kozur and his colleagues are now revising and subdividing long-ranging guide species, and beginning to recognise the occurrence of contemporary but different guide species and communities. Nonetheless, it appears doubtful that conodonts will ever be able to provide world-wide correlations for the marine Permian. None have yet been found in the cold-water Permian faunas, despite intensive search in east Australia (Nicol, 1 975).
DISCUSSION OF CORRELA TION SCHEMES
Correlations proposed by Grant & Cooper (1973)
A number of tentative schemes for correlating Permian marine faunas have been offered in recent years, and some examples may be chosen to illustrate the difficulties involved. That of Grant & Cooper ( 1973) is reproduced in Table 9. These authors placed high value on a number of different groups, including conodonts, sponges, Fusulinacea, Brachiopoda, and Ammonoidea. They were highly critical of conodont work by Sweet ( 1 970) and ammonoid studies by Furnish & Glenister ( 1970) but successfully reinterpreted the basic data, especially for the Salt Range, Pakistan. Grant & Cooper ( 1 973, p.578) deprecated the use of first appearances for correlation, one of the chief criteria used by many authorities. The present writer would regard first appearances as a prime method of correlation, and fully concurs with Wilde ( 1968, p. 12) when he 'prefers basing major time stratigraphic breaks on the first appearance of new faunas'. Grant & Cooper ( 1 973, p.578) 'preferred as complete analysis as possible of the total fauna'. Why this contradicts Wilde's view may not be clear, but elsewhere they emphasized a preference for relying on faunal assemblage (Cooper & Grant, 1 973). As a natural consequence value was placed on cluster analyses of genera for correlation (Grant & Cooper, 1 973, p.588, figs. 5 ,6). Their graphs confirmed that the Basleo and Kalabagh-Chhidru faunas have a high coefficient of similarity at a generic level. But so

�ort
h
Ame
ri
ca
r S
ou
th
H
ima
la
ya
E
. G
re
en
la
nd
I
Eu
ro
pe
an
Se
ri
es
P
ro
vi
nc
ia
l
Wo
rl
d
St
ag
e
Sa
lt
Ra
ng
e
Ch
in
a
an
d
Ax
el
H
ei
be
rg
I
s.
Se
ri
es
K
as
luni
r
Lo
we
r
Sc
yt
hi
an
(G
ri
es
ba
ch
ia
n)
Tr
ia
s
1::
Ch
an
gh
-C
ha
ng
h-
Ch
an
gh
-
ns
si
ng
ia
n
si
ng
ia
n
si
ng
ia
n
Dz
hu
lf
ia
n?
·.-I
0>
�
1:
: T
at
ar
ian
O
ch
oan
.-i
Ar
ak
si
an
·.-I
B
ra
ch
io
po
ds
::l
.9
Wu
ch
ia
pi
ng
1:
: ..c:
ns
N
?
?
?
·.-I0
Ch
hi
dr
u
= P
ro
du
ct
us
,.
.. S
va
rt
ev
ae
g
Cll
p..
Ka
la
ba
gh
,...
I Cll
Ca
pi
ta
ni
an
-
Sh
al
e
=
0.
Ka
za
n i
an
G
ua
da
lu
pi
an
Fo
ld
vi
k
Cr
ee
k
0. W
ar
ga
l
(ab
se
nt
a
t
0
Wo
rd
ia
n
Ma
ok
ou
G
ur
yu
l
Ra
vi
ne
)
?
?
?
Uf
im
ian
R
oa
d i
an
Amb
Ch
ih
Si
a
Kun
gu
ri
an
L
eo
na
rd
ian
B
ai
ge
nd
zi
ni
an
Ze
wa
n
Se
ri
es
1:
: a
nd
ns
·.-I
L
av
en
de
r
= Ar
ti
ns
ki
an
Ak.t
as
ti
ni
an
Gang
amop
te:ro
is
,...
Cl
ay
?
Cll
p..
,...
Ya
ng
si
n
Lo
we
r
Go
nd
wa
na
Cl
l S
pe
ck
le
d
� ..:I S
ak
ma
ri
an
W
ol
fc
am
pi
an
s
an
ds
to
ne
GZo
ss
opte
:l'is
As
se
li
an
Ta
lc
hi
r
Ma
pi
ng
j S
er
ie
s
Table
9.
Su
gg
es
te
d
co
rr
el
at
io
ns
o
f
Pe
rm
ia
n
se
ct
io
ns
b
y
Gr
ant
&
Co
op
er
{1
973,
f
ig
. 4
, p
.
586)
.
w
0
� c· ""
� � i:l � � ;:;· V:l
;:: c::r
t:).. � <::; · c· ::
:s ""

Discussion of Co"elation Schemes 3 1
do the Word and Capitan faunas, according to their graphs, and these faunas are not correlative, but in sequence. The only other close lateral pairing is shown between Greenland (one brachiopod zone) and Spitsbergen (at least three zones and two stages). The so-called Guadalupian and Djulfian Stages are also paired, and again, we know that they are in sequence, not correlative. Clearly, the clusters demonstrate ecologic rather than temporal links--especially pronounced at generic levels, as well shown by Williams ( 1 973) in his cluster analyses of Ordovician genera. There is time significance as well of course, but this would be enhanced by analysis of species rather than genera.
The correlation table offered by Grant & Cooper ( 1 973) does not depend entirely on cluster analyses, although it is difficult to be sure, because little documentation is provided. Miscorrelations between the Texas-New Mexico standard, and the world or so called European standard are particularly severe. The Kazanian Stage is supposed to match the entire Guadalupian Stage for instance, and the Roadian Stage is supposed to match the Ufimian. The Chhidruan is correlated with the lower Djulfian. The Svartevaeg fauna of Axel Heiberg Island, and the Foldvik Creek fauna of east Greenland are correlated with the Kazanian and entire Guadalupian. (They appear to be Kungurian.) So one may go on. The reason for such errors probably lies in the likelihood that Grant & Cooper ( 1 973) did not follow their own advice, and carefully analyse faunal content, including brachiopods, for many of the faunas. Instead, they used assessments from the literature, often based on a few ammonoids. The world correlation table of Grant & Cooper ( 1 973) contrasts in accuracy with the interpretation of the richly fossiliferous and facially complex Permian of west Texas and New Mexico by Cooper & Grant ( 1 973). Here their correlations are indeed based on species ranges and faunal assemblages, and provide overwhelming proof of the value of their procedures, and degree of sophisticated correlation that can be achieved through mastery of brachiopod faunas.
Ammonoid scheme of Furnish '( I 973) (Table 10) The scheme outlined by Furnish ( 1 973) provides world correlations in good agree
ment with the Lower Permian and much of the Middle Permian subdivisions used in the present scheme. Furnish relied solely on ammonoid data, and therefore severely restricted possibilities of correlation. Concentrating on only one group must deprive any scheme of checks and balances from other groups, and ammonoids are not a good class on which to place sole reliance for they are too rare to resolve problems of correlation for many parts of many columns in most parts of the world. They have to be used so broadly in international correlation that verification or falsification is seldom possible. In many instances, the most ammonoids can achieve is correlation of one or two parts of the column, leaving gaps so huge that it is impossible to map boundaries. Yet ammonoids, because they are few, and have been well studied, are highly useful in making pioneer studies and initial correlations, to precede studies of the benthos. For a full two thirds of the Permian Period to the 'Amarassian Stage', agreement between the studies· by Furnish ( 1 973) and the larger number of sequences in the present work is excellent. The younger Permian differs. between the two models, because the Chhidruan Stage of the Salt Range appears to have been miscorrelated with younger faunas of China and Armenia (see Waterhouse, 1 972a, b). This is not just a question of ammonoids versus brachiopods. It reflects two different understandings of the ammonoid Cyclolobus. According to Furnish ( 1 973), one lineage was present (Fig. 8), that evolved orthogenetically from Waagenoceras into Timorites, into Cyclo/obus. Cyclolobus characterised one Permian stage, with primitive Cyclolobus at the base ( =Godthaabites Frebold) and Changhsingoceras Chao at the top. According to Waterhouse ( 1 972a, b), Waagenoceras evolved into Timorites, at the same time as Cyclolobus evolved from Godthaabites, and Changhsingoceras was a much later derivation from Cyclolobus

32 Biostratigraphic Subdivisions
S T A G E
Changhsingian J CMnghsi"ffiC'r•
Chhidruan }J� .C Ill Araksian Q. Q.
Amarassian :::) •
Capitan ian • ? I -- -
Wordian WU(ItlfiOCtlfW J ,_� M•xicoceras I
Road ian Gl.,.octJras r·· I SMngoc.,. I Kuftmgocer•.
Fig. 8. The evolution of the Cyclolobidae, summarised from Furnish (1973). Note the linear development from Glassoceras through to Cyclolobus, and probably on to the poorly known Changhsingoceras. The family commenced in the "Lower Permian" i.e. Roadian Substage, according to Furnish (1973), and typified the Upper Permian in his sen� of the period.
S U B S T A G E
a: Griesbach ian Ill Q. Q. Ogbinan
:::) Vedian
Baisalian
Urushtenian
Chhidruan Ill ...
Kalabaghian Cl Cl - Sosnovian MuicOCtlras :I
Kalinovian
Iranian ? --Filippovion Gl.,.oet�r.s
Krasnoufimian
r-WNgenoet�ras·
--{ ?
I�
1 Ch.nghrillf/O(:tnS ? r·----�
• •
u . . . . • . • •
Cyclolobus
1 J GodrhHbit:ft
r;;�
-Fig. 9. Evolutionary-development of the Cyclolobidae, following Waterhouse (1972a). There is some uncertainty over some ages, indicated by queries. Note the gaps in the record, not unreasonable in view of the delicacy of the shell. The family i$ regarded principally as a bilinear Middle Permian group.

Region < Cf.l ., QJ ::. c:: c:: 00
'rl 'rl c:: ..... .. ., � s:: ..... Cll
Sy stem QJ s:: pCI s:: ::I "'0
'rl 0 QJ Cll c:: ..... lE: ... .... Cll s:: Series .. 'rl 0 IIJ � ::I s:: (,) CD ..c 'rl "'0 .. .... .. CD
Stage 'rl .... Ql 0 Cll lE: C) � u pCI
Triassic X
s:: Changhs ingian Cll
'rl ""' .... Chhidruan
· s:: ::I .. ..c
'rl N
e � Araksian Ql
Po. l ... Amaras sian X X
Ql � .... � .. Capi tanian X X X
::. "'0 �
C) Wordian X X X
]" 'rl
Road ian X X X X X
.!i: CD
s:: s:: Leonardian X X X X
'rl Cll .....
'rl � e Aktas tinian X X X
Ql Po.
s:: S terl'itamakian X X X X ... .. Ql 'rl
3 ...
J Tas tubian X X X
.. Cf.l As selian X X X
Pennsy lvanian X X
Discussion of Co"elation Schemes 33
� u m ....
.. 'rl ., ID "'0 ..... c:: s:: ::I .....
..
.. u Cll 'rl ., ...
Ill ..... s:: � QJ .. Cll � � CD Cll s:: ..... u � u .. s:: ::I 'rl
s:: ... ::I Cll .. ..c s:: Cll ... . o u >. u s:: ... u QJ lE: I .. QJ
... 'rl ..... CD .... ..c ... Ql ..... ... 'rl .... s:: Cll ..... 0 ..... CD Ql "'0 QJ �
Cll .. a ::I a CD
:3:: � � � 'rl 0 'rl Ql :r: Cf.l £:-< :3::
X X X X X X X
X X
X X X
X X X
X X
X X
X X X X X
X X X X
X X X X X X X
X X X
X X X
X X X \
X
X X
Tab le 10. Char t i l lustrat ing some of the areas with important Permian ammonoid
fauna s in sequenc e , from Furni sh ( 1973) .
� .... .. ... ..... CD
� s:: ... QJ ..... CD Cll rzl
X
X
X
X

34 Biostratigraphic Subdivisions
(Fig. 9). Waterhouse ( 1 972b) adduced much support from correlations based independently on Fusulinacea and Brachiopoda and stratigraphic sequence. Furnish ( 1973) based his argument on internal evidence of sutural development.
Furnish ( 1973) does not discuss methodology for his correlations, but the reader may infer that first appearances are allotted some significance, for instance in the acceptance of Otoceras as marking the base of the Triassic. Ammonoid sutural evolution played a critical role in Furnish's understanding of the Late Permian, though this procedure has been attacked by Spath ( 1 934), Cooper & Grant ( 1973) and Grant &Cooper ( 1973). Interestingly, it would appear that succession also played a vital role in Furnish's scheme, because many of the correlations are not in fact based on shared species, or in some instances, even shared genera. Correlations are stated ex cathedra. It is believed that they are correct, showing that high value is to be attached to the concepts of succession and ammonoid evolution held by Dr Furnish and his colleagues, provided that these are adequately checked against other evidence.
Literature review by Anderson (1 973)
A number of large charts and map-summaries of the Permian Period have been presented by Anderson ( 1 973) from compilations of the literature, without critical assessment of evidence. In striving to allot primacy to ammonoids Anderson was of course confronted by the scarcity of ammonoids in so many of the significant sections and his columns reveal many miscorrelations, and non-existent unconformities. Virtually no attention was paid to the type world Permian of Russia and extensive sequences of Siberia.
Correlation scheme of Ustritsky (1971) (Table 1 1 ) The Permian correlations offered by U stritsky ( 1 9 7 1 ) are of considerable interest
because they most closely approximate the arrangement used some decades ago, before recent advances by Fusuline, brachiopod and ammonoid studies especially in Armenia and south China. The scheme is thus rather close to that used in the Treatise volumes (Moore, 1 965), or in Australia by Runnegar ( 1 969b). Thus Cyclolobus is shown as latest Permian, as the linear descendant of Timorites and Waagenoceras. As a consequence, the significance of the Tatarian Stage or its marine correlatives is not conveyed, being reduced to a sliver in one column and omitted entirely in favour of the Kazanian in another column. The entire Middle and Late Permian is severely condensed. Moreover, the Kazanian Stage is shown as Late Permian, post-Yabeina and postTimorites in age, a totally unwarranted correlation. The Ufimian is exaggerated in significance, and shown as equivalent to Timorites and Waagenoceras, as well as Yabeina and Verbeekina. The use of a mere four major subdivisions does not appear to have saved the scheme from serious error. The scheme of Furnish ( 1 973) is not only more refined, but certainly an enormous ,improvement in accuracy. There is little guarantee that broad accuracy will be achieved by using 'safe' and coarse units of correlation.
TYPE REGIONS
There are several regions contending for consideration as world standard sections for stages of the Permian Period. Although the Permian System was first conceived from rocks and faunas of the Urals and Russian Platform, as confirmed by the very name Permian, based on the Russian city of Perm, several workers from the United States have attempted to have much or even all of the world standard transferred to west Texas and New Mexico. For example Furnish (1 973) based 'World Ammonoid Stages' on the Permian of Texas-New Mexico for the so-called Leonardian, Roadian,

Ur
al
s
St
ag
es
Ta
ta
ri
an
Ka
zan
ia
n
Uf
im
ian
S
ui
te
or
St
ag
e
Ku
ng
ur
ian
Ar
ti
ns
ki
an
Sak
ma
ri
an
Ass
el
ian
--
--
--
Mi
kh
luk
o-
Ma
ck
la
y
Ammon
oi
d
Zo
ne
s
St
ag
es
Ka
zan
ian
P
ami
ran
Cy
cto
tob
us
Zo
ne
Pa
ik
ho
ian
M
ur
gah
ia
n
Timo
ri
tes
Waa
gen
ocer
as
Ar
ti
ns
ki
an
Pe
T'r
ini
tes
Da
rva
si
an
As
se
li
an
K
ar
ac
ha
ti
ri
an
Prop
err
init
es
-··-
-·-
--
--
Tab
le
1
1.
Co
rr
el
at
io
n
sc
he
me
o
f
Us
tr
it
sk
y
(19
71
, T
abl
e
9)
.
Fu
su
li
na
ce
a
Zo
ne
s
Sm
al
l
Fo
ram
in
if
er
s
Yab
ein
a
Ver
be
eki
na
Par
af
usu
tin
a
Pse
ud
osch
wag
erin
a
�--
--
·---
I I
� � !:1::1 � c· ;:
:r ..
.
w
VI

36 Biostratigraphic Subdivisions
Wordian, and Capitanian Stages. Younger �ermian standards were erected from scattered regions in Timor, Salt R�ge, and c.hina. Grant f!L C:ooper. ( 1973) asserted th�tthe world standard for the Roadtan, Wordtan, and Cap1tan1an umts are to be found m Texas, and also used the Changhsingian Stage of China. The basal Permian (Asselian and Sakmarian) was left blank, implying at least some doubt about the use of AsselianSakmarian as a world standard. Of course the designation of world standards should properly be left to an international commission. In the meantime it is proposed that, wherever possible and conveniept, the standard sections should be somewhere in the Soviet Union for the following reasons : 1 . Historically, the Permian System was first conceived and proposed on the basis of
Russian rocks and faunas (Murchison, 1 84 1}. 2. The Soviet Union has an abundance of rocks and faunas providing adequate stan
dards in most instances. 3. The volume of work by Soviet palaeontologists on Permian problems exceeds that
from any other country. They have attained fine control over many regions, and their work may be readily interpreted and extended elsewhere. There is good emphasis on stratigraphic and faunal succession for a number of fossil groups.
The Asselian and Sakmarian rocks and faunas are all diverse and well known, though revision is required for the brachiopods. The suggestion that the Leonard of west Texas replace the Baigendzinian is indefensible, as Baigendzinian faunas are virtually as diverse, historically long established, and much more extensive geographically. Moreover Baigendzinian zonation is more refined, whereas the Leonardian (=Cathedral Mountain) has not been subdivided. Faunally, the Kungurian and Kazanian Stages are so well entrenched in the literature that it would seem impossible, certainly most undesirable, to replace them. They are finely divided, in contrast to the crude double division of the Texas sequence (Roadian, Wordian). Indeed it may be noted that Cooper ( 1957) and Grant ( 197 1) miscorrelated even other North American faunas, from Oregon . and Arctic Canada, with faunas of west Texas, to demonstrate that the Glass Mountains standard does not guarantee successful correlation.
The next segment oftime offers more difficulty. The Capitanian Stage of New Mexico, reinforced by the equivalent faunas of Las Delicias at Coahuila, Mexico, offers diverse though geographically restricted and somewhat unusual faunas, sufficient for an excellent standard. However, the Kalabaghian, Amarassian, and Chhidruan Stages from the Salt Range, Pakistan and Timor, Indonesia, have also been proposed. Waterhouse ( 1972b) and Grant & Cooper ( 1973) pointed out that the absence of any sequence must rule out the Amarassian Stage. Furnish ( 1973) disagreed and commented that 'the stratotype as a reference serves secondarily to the. faunal content in time-stratigraphic definitions. This thought is not in accord with the Stratigraphic Code' . . . (Furnish, 1973, p.523). The Kalabaghian and Chhidruan Stages, used and defined by Waterhouse ( 1972a, b) and here treated as substages of the Punjabian Stage (Schenk et a/. 194 1), are based on rocks and faunas in the Salt Range, Pakistan, and are underlain and overlain by faunas in sequence. Although lacking Fusulinacea, they have highly diverse brachiopods, numerous bivalves and other invertebrates and a few ammonoids, sufficient to act as standards. Perhaps there will be some preference for replacing them with Soviet sequences. The Murgabian Stage of central Asia appears to h�ve �iverse Fusulinacea, and more brachiopods are being described (Grunt &Dtmetnev, 1 973). Alternatively, the Gnishik and Hachik faunas described by Ruzencev & Sarytcheva ( 1965} and Stepanov et al. ( 1969), as evaluated by Waterhouse ( 1972a), may suffice .. Ammonoidea as usual are sparse or missing in these faunas, but selectionof these umts would help provide the standard for many substages in one region. Moreoyer, such a selection might obviate political difficulties, as similar sequencesoccur m Iran.
The Late Permian is well displayed by sequences in Armenia. Faunal units have

Type Regions 37
been formally proposed by Waterhouse ( 1 972a), based on geographic localities, with designated type sections, specific zones, and faunas described by Ruzencev & Sarytcheva ( 1965). Rostovtsev & Azaryan ( 1973) have further proposed a comprehensive name for the upper two faunas from the same region, so that the two works combine to give a logical, and one would hope, widely acceptable subdivision. Glenister & Furnish ( 1970) proposed an undefined Araksian Stage which Furnish (1973) equated with the zones equivalent to the lower and middle Baisalian fauna of Waterhouse (1972c). Fal,lnally the Araksian Stage is inadequate, for the real boundaries lie with the Baisalian Substage, as also shown by Kozur ( 1974), not Araksian Stage. The Baisalian unit no.t only was defined earlier (of small consequence) buf is more realistic. The base of the Baisalian Substage presents some problems, Meagre faunas in the area, as discussed by Waterhouse ( 1 972b ), point to a related but slightly older substage, called the Abadehian in central Iran by Taraz (1971, 1973)� Furnish (1973) suggested that tlris interval matches the Amarassian · Stage Qf Timor, and the uppermost Coahuila beds. ofMexico and the writer agrees, at least with the Mexican correlation. Kozur
· (1974)
matched the Abadehian with the bulk of the Chhidru Formation. The same Abadehian interval has had very rich faunas described from the Urushtenian Horizon of the Greater Caucasus. I provisionally use the name Urushtenian Substage, long established through Fusulinacean and brachiopod work, and with much richer faunas than the Abadehian of central Iran, but leave the Russians to designate a type area where they wish.
Grant & Cooper ( 1973) and Furnish (i973) referred Late Permian beds to the Changsingian (Changhsingian) Stage of south-west China, based on superb sections and rich faunas described in part by Huang (1933), Sheng (1964), and Chao (1966). But Rostovtsev & Azaryan (1973, p.94) pointed out that the stratigraphic position of the Changhsing Limestone is not well established, and that it is not latest Permian, and that its faunas are not well known. Kozur (1974) judged the Changhsing interval to be late Baisalian and early Dorashamian.
Finally there is the question of the Griesbachian or its equivalents. This stage, of debated Permian or Triassic age, has been carefully defined and its ·ammonoids described by Tozer ( 1967). Brachiopods are rare, and some may wish to transfer the standard section to Armenia. Waterhouse ( 1973b) argued for retention of the stage as it 1ow stands, subject to international adjudication, but it is true that it has the disadvantage of commencing above an unconformity.
Type sections and stage names
An international subcommission is required to resolve the replication of available names for stages, and designation of various type sections, especially for the younger Permian rocks and faunas. In such decisions it is to be hoped that the decision be based primarily not on the priority of a proposal, but especially on the usefulness and relevance to the rest of the world, and to lesser degree on the proper observation of the rules of stratigraphic nomenclature (Hedberg, 1972).
Finally, it must be observed that some obligation lies with the country responsiblefor the standard sections. Access should be readily granted to overseas specialists, and faunal elements should be provided to at least some insti.t�tiO!J§. itt _Q!her countries. In this regard, Drs G.A. Cooper and R.E. Grant have provided a model of generosity and helpfulness. They have not only guided numerous. specialists in the �eld thro�gh the Glass Mountains, west Texas, but have donated limestone blocks wtth magmficently silicified brachiopods to many institutions.
Substages
The earlier subdivisions treated as stages in Waterhouse ( 19�2a, 1973a, b) are

Serie
s
Stag
es
140
'f
40
'C
E ;) � Q. 0
LOW
ER P
ERM
IAN
I As
selia
n I
Sakm
arian
I Baig
endz
inian
Kun
guria
n
4
5 9
MID
DLE
PER
MIA
N
UPP
ER
PERM
IAN
Kaza
nian
Pu
njabi
an
Djul
fian
Dora
sham
ian
J2'f
o·c�
�-
--
--
--
-_j
Fig
. 10
. H
yp
oth
ecat
ed w
orl
d t
emp
erat
ure
ch
ange
s an
d t
hei
r re
lati
on
ship
to
Per
mia
n s
erie
s an
d s
tage
s, w
ith
su
bst
ages
nu
mb
ered
as
in
Tab
le
1,
p.
3. B
ased
on
dis
trib
uti
on
of
till
ites
, co
ral r
eefs
, sal
t d
epo
sits
, an
d c
oal
dep
osi
ts,
and
flu
ctu
atin
g fa
un
al d
iver
siti
es.
Red
raw
n
fro
m W
ater
ho
use
(19
73
d,
p.
46
2,
fig.
10
).
w
00
t:l::l B· "'
... i:l �· i:l '1::3 ;:::-
;:;- VJ
1::: (;)- � �· t:; · B· ::s
"'

Type Regions 39
perhaps too ambitious, and therefore a number have been reduced herein to substages. This makes the scheme simple and easy to remember, retains a fair degree of consistency with previous work, and enables broad correlation for inadequately understood
· faunas that cannot be matched precisely with a zone.
Major subdivisions of the period
To some extent stages can only be arbitrarily grouped, but for the Permian Period a natural grouping is suggested by major faunal changes that occurred in response to major climatic changes (Fig. ! 0). It is clear from the stratigraphic record of east Australia, New Zealand and north-east Siberia that three major glacial episodes may be recognised, in the Surenan Substage, Filippovian Substage, and Vedian Substage. The first two were followed by several further glacial episodes and world-wide refrigeration, and then by world-wide climatic amelioration. Thus a three-fold division seems natural. The first, and longest, early Permian subdivision retained many Carboniferous genera. Dillgnostic forms included Pseudoschwagerina, numerous brachiopods, and Perrinitid ammonoids. The Middle Permian saw the significant entry of Neoschwagerina, and Cyclolobid ammonoids. The Late Permian saw the entry of Phisonites , an ammonoid, and brachiopods Comelicania and Janiceps.
A two-fold division for the system is widespread in the literature, although earlywork in the United States, Japan, and· China recognised a three-fold division. A two-folddivision was warranted for rocks of the Urals and Russian Platform, because the upper third or half was terrestrial, and too poor to justify more than two divisions. Now that a fully marine sequence is established, it is clear that a three-fold division is justified, especially as the Russians for a time referred the upper third, not to just a discrete section of the Permian, but an entirely different system.
A second and minor question concerns the start of the Middle Permian, placed by some authorities at the base of the Kungurian Stage, and now by some American workers at the start of the correlative Road Canyon Formation in the Glass Mountains of west Texas. Others have favoured a higher boundary, at the top of the Kungurian Stage, or above the Road Canyon Formation, by stressing the survival in the Kungurian Stage of earlier forms. Dubious statistical support was claimed from a cluster analysis that slightly discriminates the Road Canyon faunas from Wordian and Capitanian faunas, with no data on the relationship to the Cathedral Mountain faunas. Other workers, such as Wilde ( 1968) and Waterhouse ( I 972a, 1973a), have stressed the significance of incoming rather than outgoing forms. There can be no question that a great faunal change occurred at the start of the Kungurian Stage, even though many species survived from older faunas.

3. The Base of the Permian
In recent years the base of the Permian System has been placed at the base of the Asselian deposits and faunas of the Urals and Russian Platform. It is widely agreed that to some extent the boundary is fixed arbitrarily, because successive faunas show incremental changes to the extent that various options have been supported at different times, ranging recently from the base of the Orenburgian, (see Table 12) to the base of the Sakmarian, the latter boundary still favoured by Chinese geologists.
A. SIGNIFICANT MARINE BIOTA
Fusulinacea
The Asselian Stage has been primarily distinguished by its Schwageriilids. Pseudoschwagerina and in particular Schwagerina are characteristic genera, although they do not always provide consistent time planes, and Barkhatova (1964) stated that neither the start nor end of Schwagerina• (in the Russian sense) can be relied on for dating. In the Russian Platform and Urals, the Asselian Stage is underlain by the Orenburgian Stage with Pseudofusulina• in the Russian sense (e.g. Nalivkin, 1972). Dr R.A. Douglass and Dr G. Wilde have informed me that Pseudofusulina (in the American sense) is a reliable key to the Permian Period. Certainly, in North America, and over much of Asia, Pseudofusulina appears to be restricted to the Permian, though beds with 'Pseudofusulina' from the Pamirs and indeed Austria are generally assigned to the Late Carboniferous as discussed subsequently. Judged from brachiopods, the Austrian faunas could be Permian but the faunas of south Fergana are likely to be Carboniferous. It would appear that the communities of Fusulinacea should be studied as well as zones, and until this is done, and until Pseudofusu/ina is clearly delimited, . it remains difficult to decide whether or not Pseudofusulina commenced in pre-Permian time. And until this is decided, it is obviously difficult to draw the boundary on Fusulinacean evidence.
Ammonoidea
Ruzencev ( 1952} has done a great deal �o clarify and establish the Asselian Stage from ammonoid studies in the Urals. It is he who has most clearly enunciated reasons for fixing the boundary at ·the base of the Asselian Stage.
'The position of the upper boundary of the Carboniferous System has long been under discussion, a few stratigraphers drawing it at the base of the Orenburgian Stage, but the great majority placing it at the base of the Asselian Stage and a minority at the top of that stage. Such diversity of opinion is probably largely due to the fact tha� the upper boundary of the system is not clearly so sharply expressed as the lower boundary, even in terms of ammonoid changes ... The Permian ammonoid assemblage as a whole is peculiar, but it formed very gradually. Of the eight highly characteristic Permian families, the Medlicottiidae and Shumarditidae appeared in the Moscovian Stage, the Marathonitidae in the Zhigulian (=.Jigulevian), the Adrianitidae and Vidrioceratidae in the Orenburgian, the Paragastrioceratidae, Metalegoceratidae and Popanoceratidae in the Asselian. It is interesting that in the Sakmarian Stage no new families appeared. This constitutes definite evidence against the third version of the boundary, in which it is placed at the top of the Asselian Stage .. . A count shows that the Zhigulian (=Jigulevian)

Significant Marine Biota 41
system Staqe Kotelnich River Sakmara
Upper Variegated sandstones and clays Red sandstones , siltstones Tatarian with beds of marl ( 162m) and marls (200-1500m)
Marls and clays with beds of Lower dolomite ( 113m)
Tatarian Variegated sandstones and clays ( 153m)
Upper Gypsiferous , muddy limestones Above , red beds ; below, lilae-Kazanian (28m) stones & grey clays (200m)
Lower Grey limestones . Marls and clays :or; .
Kazanian at the .bottom; and a fauna ( 79m) .c H Ufian (•Ufimian) Red, sands and clay ( 29m) Red beds (200m) :£ � Kungurian Gypsum and anhydrite with beds Gypsum with sequences of clays 1&1 of dolomite ( 175m) and sandstones towards the � bottom ( 1800m)
Artinskian Conglomerates & sandstones (450m)
Clays with bands of sandstones ( 700m)
Sakmarian Interbedded dolomites and Ammonite-bearing argillites with anhydrites ( 56111) beds of sandstone and marl
(100-1200m)
Asselian Grey and dark grey gypsiferous Argillites , marls and sand-dolomites ( 17m) stones. Fusulinids are
present ( 500m)
Orenburgian Grey dolomites and limestones Marls·, argillites with beds and with beds of gypsum ( 128m) sandstone & limestone ( 200-:-300111)
Gshelian
Cll 0 Kasimovian Grey limestones , dolomites and Flysch-like alternating sequence
0 gypsum ( 75111) of argillites and sandstones
� with boulder conglomerates ( lOOm)
1&1 limestones and r.. Mayachkovian Interbedded pale
(Moscovian) H � suite dolomites (113111)
z H ( 52m) 0 8 Podol ian ste . Grey limestones & dolomites
11:1 � �hirian S te . Above , limestones ; be low , sand-� stones & clays (46111) .c Verelian S te . Re d clays & sandstones with beds u of limestone (40111)
Dolomitized limestones {12� Namurian dolomites
Table 12 . Upper Palaeozoic successions in the Russian Platform and Urals slightly emended
from Nalivkin ( 19 7 3 , tables 12 , 30) .

42 The Base of the Permian
and Orenburgian assemblages have 2 1 genera in common, the Orenburgian and Asselian 1 6, and the Asselian and Sakmarian 2 1 . Six new genera appeared in the Orenburgian age, 1 1 in the Asselian and 7 in the Sakmarian. These data are sufficient evidence in favour of the second version of the boundary, which places it at the base of the Asselian. But the arithmetical count is not the whole story.
'In the Orenburgian assemblage such Carboniferous genera as Uddenoceras, Daixites, Neodimorphoceras, Aristoceras, Gonioloboceras, Glaphyrites, Paraschistoceras, Schistoceras, and Shumardites are clearly predominant. Ammonoids of Permian appearance play a completely subordinate part here. There are accordingly no sufficient grounds for assigning the Orenburgian Stage to the Permian System ... In the Asselian assemblage all the principal families of the Permian ammonoids are already represented together with such characteristic Permian genera as Sakmarites, Akmilleria, Artinskia, Paragastrioceras, Juresanites, Properrftdtes, Kargalites (subgenus Kargalites), Marathonites (subgenus A /mites), Prostacheoceras, Protopopanoceras and others. The number of ammonoids of Carboniferous appearance in this complex is quite insignificant. It must be emphasized that this peculiar fauna developed at the beginning of the Asselian or 'Schwagerinan' age; therefore the ammonoid evolution offers no grounds whatsoever for assigning the Asselian Stage to the Carboniferous System. The boundary between the Carboniferous and the Permian must therefore be drawn at the base of the Asselian.' Quoted from Ruzencev, 1 965, slightly emended from translation in International Geological Review, volume 8, No. 1 , pp. 57,58.
However, few informed readers would be willing to accept Ruzencev's conclusions from the data he has presented. After all, the Orenburgian is clearly close to the Asselian in terms of ammonoids, underlining the fact that the decision is arbitrary. There is however, a more critical point to be evaluated. Ruzencev ( 1 965) stated that the Asselian fauna developed at the beginning of the Asselian or 'Schwagerina' Stage, a point crucial for placing the boundary at the start of the Asselian. But the statement is not sustained by his more thorough discussion of Asselian ammonoids (R uzencev, 1 9 52). According to this treatise, three ammonoid zones are recognised in the Asselian, and the basal zone is characterised by the loss of a number of Orenburgian genera including Uddenites and Prouddenites, and by the proliferation of Glaphyrites, without the appearance of a single new genus (italics mine, directly translated from Russian). If this is correct, and it has never been countermanded in the literature, it suggests that the start of the Asselian was diagnosed simply by means of a depauperized ammonoid fraction in animal communities. Even the proliferation of Glaphyrites only repeated a phenomenon that occurred also in Moscovian times, and the genus ranged from Mississippian to Sakmarian (Dr W.W. Nassichuk, pers. comm.) The basal Asselian may mark a significant event, but then again, it may mark simply the final dwindling of an essentially Orenburgian faunule. It must be asked, was the early Asselian really Orenburgian? Barkhatova ( 1 964) asserted that Ruzencev ( 1 952) had in practice alloted primacy to Fusulinacea, even though he strongly criticised their value :
'In the type section of the subdivision described by Ruzencev in Bashkiria, along the Suren and Uskalik rivers . . . ammonoids are absent. At a number of places their position within the section cannot be accurately located in relationship to the base and top of the given subdivision' . . . (Barkhatova, 1 964, p.274). It can thus be said that the basal Asselian was characterised by early Asselian Fusulinacea, and by impoverished, late Orenburgian ammonoids. It might then be argued that Pseudofusulina and Pseudoschwagerina, or shells so-called, dominated different but contemporaneous communities, to suggest that the Orenburgian and Asselian belong together in one major unit.

� N.W. EUROPE
z W. GERMANY; SAAR c LOW BASIN -I COU NTRIES,
� FRANCE. Ill AUTUNIAN
KUSELER
a. SCHICHTEN
"' ::) 2 - - - - - c
m Ill a:: STEPHANIAN.. c( - u z .
� 0 z WEST-a:l c( PHALIAN � ;;; c
c w .....
u ;;;
Significant Marine Biota 43
._.,_R USSIA .-.- u sA ...
MOSCON BASIN
---------PSEUOOFUSULINE
HORIZON
GZH E L HORIZON KASIMOV HORIZ.- - - -
MYACHKOIIIAN HORIZON
PODOL IAN HORIZON
KASHIRIAN HORIZON
L VEREYAN HORIZON
M I D -CONTINENT REGION
WOLF CAM P
-· � .. -- - - - - ·
_ V I RG I L
! z c( MISSOURI - �- - -_ ..... z � ;.., >- z "' OESMOI N ES z c( z .....
� w 0 A.
Fig. 1 1 . Late Carboniferous correlations suggested by spores, according to Anderson ( 1 973, p. 69, table 3), based on Helby ( 1 969, pp. 69-7 1) .

44 The Base of the Permian,
Brachiopoda
Clearly what is needed is· a third source of evidence, from the brachiopods, which by virtue of being less facies controlled and far more widespread, co1,1ld prove decisive. But there are few modern studies on the Uralian brachiopods, and Asselian,and youn�er brachiopods from the Urdl.s and Russian Platform require re-examination before they can be accurately fitted into a modern stratigraphic framework. More attention has been paid to Late Carboniferous brachiopods, summarised by Miloradovich (1949), Stepanov ( 195 1), Likharev (1939), Ivanov ( 1935), Mironova ( 1967), Ivanova & Ivanova ( 1955), and Sarytcheva & Sokolskaia ( 1952), but the attempted division of faunas is not as fine as for Fusulinacea or Ammonoidea. Generally the Late Carboniferous Period is subdivided, if at all, into only two .units, Kasimovian and Gshelian, which apparently incorporates the Orenburgian. In. other works, Kasimovian and Gshelian are combined to make the Jigulevian, below the Orenburgian. From very imperfect stratigraphic control, it is judged that the Kasimovian, and Gshelian form one major faunal assemblage, of about stage rank, as distinct from the Asselian Stage. But where the Orenburgian brachiopod affmities lie is far from clear. There can be little confidence in the boundary until the brachiopods are studied or reassessed zone by zone.
REGIONS SIGNIFICANT FOR PLACEMENT OF THE BOUNDARY
Pamirs, A ustria
Several other regions are significant in assessing where the bouodary should be drawn, Rich faunas. are known in the Pamirs, where extensive Fusulinacean zones, rich brachiopod faunas (Volgin, 1 960), and a number of ammonoid specimens are found in the Sakmarian and Baigendzinian Stages. Russian fusulinacean experts have placed the Permian-Carboniferous boundary at the base of the Karachatirian Suite with Pseudoschwagerina, above the Dastar Horizon with Pseudofusulina, and the Uchbulak Horizon with Triticites and Pseudofusulina, regarded. as Gshelian, including Oten-burgian, and Kasimovian (Mikhluko-Maklay, 1 958). . .
The tectonically complicated Austrian sequences in 'the Carnian Alps include Fusulinacea and Brachiopoda, with no significant Ammonoidea. Pseudoschwagerina enters the lower Pseudoschwagerina limestone at the base of the Rattendorfer beds, and is taken to indicate the base of the Permian by Kahler & Pray·( l963). Pseudofusulina is found in underlying faunas in the upper Auernig beds. The Auernig beds also contain brachiopod faunas of general Late Carboniferous and Early Permian appearance, with Attenuatellafrechi (Schellwien), member of a genus which is widespread in Early Permian deposits. Perhaps, like Pseudofusulina, Attenuatella entered pre-Permian faunas in some regions. But the general correlations in this region could accommodate a Permian age, provided that cryptic Carboniferous or Early Permian unconformities are present. It is interesting to note the presence of rolled blocks of sediment in the Auernig beds, ·suggesting the possibility of sharply lowered (glacio-eustatic?) sea-level. A fmal decisionrequires close analysis of the fusuline and brachiopod species. Floras from these bedsare generally assigned to the Stephanian, which opens the further question of adjudicating a boundary between Carboniferous typified by non-marine beds in Europe,and Permian based on marine beds in Russia. The possibility of offiap, with a sizablegap, proposed by Helby ( 1969), or overlap is high. Helby ( 1969) suggested that Stephanian C matched the Kasimovian horizon of Russia (Fig. 1 1). But Wagner & Prinz( 1970) recorded plants and brachiopods from Spain that may not support Helby'shypothesis. They assigned the plants from Spain to Stephanian A, and the brachiopodsto the 'Kasimovian', chiefly on the basis of direct correlation with the Middle Kalkarme

Regions Significant for Placement of the Boundary 45
beds of Austria. If the Spanish and Austrian faunas are really basal Permian it seems that much of the younger Stephanian may prove to be Permian.
West Texas, United States
The Early Permian faunas of west Texas, ranging into north Texas, Kansas, etc., are virtually as diverse as those of the Urals. In the standard Wolfcamp sequence primitive Schwagerinids and Pseudofusulina, according to American defmitions, and various brachiopods indicate a basal Permian age for the Uddenites Shale in the Gaptank beds, as summarized by Cooper & Grant ( 1973). Strong objection to this is voiced by Furnish ( 1973), who argued that the boundary should be retained for the sake of stability and because ammonoids from this shale include several with Carboniferous links, including Uddenites, Prouddenites, and Uddenoceras, listed also from the Jigulevian (Kasimovian, Gshelian) and Orenburgian of the Late Carboniferous in the Urals by Ruzencev ( 1962, p.352). Yochelson ( 19S4) pointed out that the gastropod Omphalotrochus normally typified Permian faunas, including the Gaptank beds, but was found in the earlier (apparently Gshelian) Omphalotrochus Zone, well below the Asselian Stage.
It is noteworthy that Girty in King (1937) rec�rded 'Ambocoelia', from the Uddenites fauna. This might · prove to be Crurithyris or Attenuatella which widely characterized Early Permian faunas. Cooper & Grant ( 1973, p.366) in their discussion of the problem noted the presence of the following allegedly Permian brachiopods in the Uddenites shale: Scacchinella, Waagenoconcha•, Kochiproductus, Limbella, J'.?guliferina, Orthotetella, Echinauris, Fimbrinia•, Martinia•, Dip/anus, Tropidelasma, Parenteletes, Spirifer of S. condor type•. Of these, Kochiproductus is in my view the most significant, and several of the other genera have scarcely any other than local significance, those genera asterisked being particularly common in Carboniferous deposits, with Teguliferina typifying early Gshelian-Kasimovian faunas in the Moscow Basin. Similar brachiopods, including Waagenoconcha, Martinia, and Spirifer of the condor type are found in the Cisco beds of the mid-continent (i.e. North Texas), with an Uddenites ammonoid fauna, and Teguliferina, Limbella, and Scacchinella occur in the Marathon Basin with Virgilian Fusulinacea. Cooper & Grant ( 1973) included these faunas also in the basal Permian. The Pennsylvanian would close in their view with the Missourian Stage, which may be approximately Kasimovian according to the Russian standard. If so, this would appear to lower the Permian boundary to include the Gshelian faunas of Russia.
A ustralia
East Australia has impoverished Late Carboniferous and Early Permian faunas, that are significant for Gondwana correlations. As in Argentina, there has been considerable discussion over the start of the period, which is usually taken as indicated by the plant Glossopteris. The Late Carboniferous rocks of Australia appear to be extremely condensed, and no sizable marine post-Moscovian fauna of Carboniferou� age �s known from east Australia. Just above the ?Moscovian faunas are found glacial sediments, which have been dated as Carboniferous, but could be early Permian, with some support from rare Eurydesma found close to this horizon in New South Wales. However Gonzalez ( 1973) has recorded Eurydesma from the Argentina with Levipustula, a genus especially typical of the early Moscovian, although perhaps persisting into younger faunas.
Helby ( 1 969) suggested that the Stephanian to Permian floral change was represented in east Australia as the replaceme�t of the GrandisfJ!'ra fl.ora by the Potonieisporites flora in the basal part of the glacial Seaham Formation (Ftg. 1 1), correlated with the Kasimovian Stage. He showed the Dalwood Group to be as old as

46 The Base of the Permian
Gshelian, with Glossopteris entering a little later, and replacing Rhacopteris. Thus even plants, and the significant bivalve Eurydesma, would appear to have been of Permian aspect by supposed Gshelian time, suggesting that the boundary could be lowered. However, none of Helby's Late Carboniferous ages for Australian beds are well established.
Yukon Territory, Canada
The Yukon Territory, especially in the north Ogilvie Mountains, offers superb, well exposed, and continuously fossiliferous sequences, especially rich in brachiopods. Fusulinacea have not been fully studied, and are missing from the Early Permian segment. Ammonoids are too few or localized to provide any sort of succession. But the brachiopods are closely allied to those of the Urals and Russian Platform, and reveal links with the Glass Mountains.
In the Ettrain Formation (Bamber & Waterhouse, 1 97 1) the early zones are approximately early Moscovian-late Bashkiri<.n, judged from small Foraminifera. They are followed by three consecutive brachiopod zones called the Buxtonia Zone, the Gemmulicosta, Praehorridonia, and Reticulatia zones, all correlative, and the PurdonellaGibbospirifer zones which may approximately match the Kashirian, Podolian, and Mayachkovian suites respectively of the Moscovian Stage (Table I I). The overlying faunas in the Yukon Territory, chiefly in a new formation yet to be named, are characterized by an influx of brachiopod genera, including species of Orthotichia, schuchertellids, Tubersulculus, Krotovia, Kozlowskia, Chaoiella, Crurithyris, Atriboniidae, Brachythyris ufensis, broad ribbed Choristites, and the distinctive genus Duartia. These faunas include the Ck and Dos (Kozlowskia and OrthotichiaSeptospirifer) zones in sequence of Bamber & Waterhouse ( 1 97 1 ). In a general way these faunas are Kasimovian-Gshelian, and perhaps Orenburgian, in terms of the Russian succession.
They are followed by a suite of zones, characterized by many different brachiopod genera, including Tomiopsis, Attenuatella, Kochiproductus, and many productids that clearly mark a major faunal change, in the E faunas of Bamber & Waterhouse ( 1971). Fusulinacea and Ammonoidea are too rare to be highly significant. It is clear that the major faunal boundary falls at the base of the E faunas, or Jungle Creek Formation, but this does not necessarily mean that the boundary has been placed at that horizon in other countries.
Verchoyan, Orulgania, North-east Siberia
Thick, chiefly clastic sequences with abundant brachiopods and frequent intervals with Ammonoidea are found in the Verchoyan region near the Lena River of north-east Siberia. In north Verchoyan, and Orulgania, as outlined by Menner et a/. ( 1970, p.44fl), the Tiksin Suite appears to be of mid-Carboniferous age, correlated with the Makarov Horizon of Taimyr Peninsula (Table 1 3). It is overlain by the Soybol Suite with early Jakutoproductus and various Cephalopoda, including Yakutoceras and Parajakutoceras. In Orulgania on the east flank of the Verchoyan anticline, many brachiopod species are found in the Yupenchin Suite, including genera Semicostella, Antiquatonia, Plictotorynifer, and others that suggest a Carboniferous age. Younger beds in this region, assigned to the Suyorgan Suite, include Fimbrinia, Jakutoproductus, Cancrinella, Linoproductus, Brachythyris, Paeckelmanella, Settedabania, Martinia and Attenuatella. Menner et a/. ( 1 970, p.47) correlated the faunas with those of the Ekachan Suite of Set-Davan, and upper Makarov beds of Taimyr Peninsula. Ammonoids include Owenoceras orulganense Popov, Agathiceras uralicum (Karp.) and Eoshumardites artigensis Popov, leaning towards a Carboniferous age. Plant horizons of the Suyorgan beds include Noeggerathiopsis, often regarded as exclusively of Per-
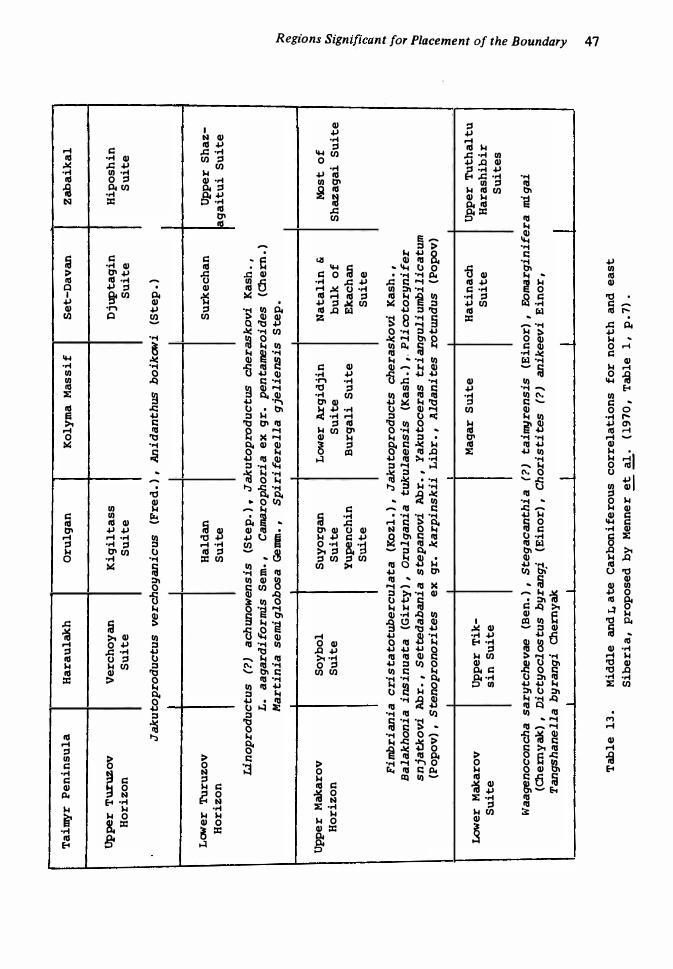
Ta
imy
r
Pe
ni
ns
ul
a
Uppe
r T
uruzo
v
Ho
ri
zo
n
Ha
ra
ul
akh
Ve
rc
ho
yan
S
ui
te
Or
ul
qa
n
Ki
qi
lt
as
s
Su
it
e
Ko
ly
ma
M
as
si
f
Se
t-
ca
van
Dj
upt
aq
in
S
ui
te
Jak
uto
pr
od
uc
tus
v
er
cho
yan
ic
us
(Fr
ed
.)
, Jln
id
an
thus
b
oik
owi
(St
ep
.)
Low
er
Tu
ru
zo
v
Ho
ri
zo
n
Ha
ld
an
Su
it
e
Su
rke
ch
an
Lin
opr
od
uc
tu
s
(?)
ach
uno
wen
si
s
(St
ep
.)
� J
aku
top
rod
uc
tus
ch
er
ask
ovi
K
as
h.
,
L. a
ag
ard
ifo
rmi
s
Se
m.
, C
ama.
ro
ph
or
ia
e
x
qr
. p
en
tame
ro
id
es
(Ch
em
.)
M
ar
tin
ia
se
mig
lo
bos
a Ge
DIIl.,
S
pi
ri
fer
el
la
g
jel
ie
nsi
s
St
ep
.
Upp
er
Mak
ar
ov
H
or
iz
on
S
oy
bo
l
Su
it
e
Su
yo
rq
an
Su
it
e
Yu
pe
nc
hi
n
Sui
te
Low
er
Ar
qi
dji
n
Su
it
e
Bu
rq
al
i
Su
it
e
Na
ta
li
n
& b
ul
k
of
E
ka
ch
an
Su
it
e
Fi
mbr
ian
ia
c
ris
tato
tube
rc
ul
at
a
(Koz
l.
) ,
Jak
uto
pr
od
uc
ts
che
ra
sk
ovi
K
as
h.
, B
al
ak
ho
ni
a
ins
in
ua
ta
(G
ir
ty
),
Or
ul
ga
ni
a
tuku
lae
ns
is
(Ka
sh
.)
,. P
li
coto
ryn
if
er
s
nj
at.k
ovi
Ab
r.
, S
et
ted
ab
ani
a
ste
pa
no
vi
Abr
.,
Ya
ku
toc
er
as
tr
ian
gul
iumb
il
ic
at
um
(Po
po
v)
, S
ten
op
ro
no
ri
tes
e
x
qr
. k
arp
in
sk
ii
Li
br
.,
Ald
an
ite
s
rotun
du
s
(Po
pov
)
Lowe
r
Mak
ar
ov
S
ui
te
U
pp
er
T
ik
s
in
S
ui
te
Ma
ga
r
Su
it
e
Ha
ti
na
ch
Su
it
e
Zab
ai
ka
l
Hi
po
sh
in
S
ui
te
Upp
er
S
ha
z
aq
ai
tu
i
Su
it
e
Mos
t
of
S
haz
aq
ai
S
ui
te
Up
pe
r T
ut
ha
l t
u
Ha
ra
sh
ib
ir
S
ui
te
s
Wa
age
no
co
nch
a
sa
ry
tc
hev
ae
(Be
n.
),
Ste
ga
can
thi
a
(?)
ta
irm)
re
ns
is
(Ei
no
r)
, Eo
ma.r
gi
nif
er
a
mi.g
ai
(Che
my
ak)
, D
ic
ty
ocl
ostu
s b
yr
ang_i
(E
in
or
),
Cho
ri
sti
tes
(?
) an
ike
ev
i E
in
or
, T
angs
ha
ne
ll
a b
yr
ang
i Ch
em
yak
I I
Tab
le
1
3.
I I
Mi
dd
le
an
d L
at
e
Ca
rb
on
if
er
ou
s
co
rr
el
at
io
ns
f
or
n
or
th
a
nd
e
as
t
Si
be
ri
a,
pr
op
os
ed
b
y
Me
nn
er
e
t
�·
(19
70
, T
abl
e
1,
p.7
).
I
� � <:;· ::s
"'
IJ:l
oq·
::s .;; ff ::s
..
.. 'C' ... "'o
i:i
(")
"' :i "'
::s
.... � ....
::s- "' t:x:l
<;:)
1:::
::s f} � �

48 The Base of the Permian
mian age. The supposedly La�e Carboniferous Ha!dan Suite incl�des �rac�i�pods thatsuggest either an Early Permian or Late Carbomferous age, With Fzmbrzma, Tubersulculus Jakutoproductus, Levicamera, Stenoscisma. A mbocoelia, and Orulgania. ByCanadi;n standards they appear rather like those of the Late Carboniferous D faunas,if Ambocoelia is correctly identified. Overlying faunas of the Verchoyan Suite includeKochiproductus porrectus (Kut.), possibly indicative of an Asselian age, withNeoglaphyrites, Agathiceras uralicum (Karp.), Uraloceras simense Ruzh., Neopronorites skvorzovi (Chern.) and Paragastrioceras verneuili Ruzh.
Clearly Verchoyan offers valuable sequences, probably comparable with those of Canada, with Ammonoidea more common, and Fusulinacea apparently absent. A number of inconsistencies arise from the correlations preferred by Menner et al. ( 1970), especially amongst the Ammonoidea, compared with ranges in the Urals. Of genera found in the Verchoyan Suite, Paragastrioceras characterises midAsselian faunas of the Urals, Neoglaphyrites disappeared at the end of the Asselian, but Uraloceras did not appear until the Tastubian (Ruzencev, 1 952), yet all are found together in north-east Siberia. Kochiproductus characterised Asselian beds, but A ttenuatella is recorded well below in the ?Middle Carboniferous Suyorgan Suite, with Noeggerathiopsis that is also usually regarded as no older than Permian. Admittedly this material has not been described, and could have been misidentified. But it must raise the possibility that Attenuatella did enter Carboniferous successions, :as is possibly the case also for Spain and Austria. Equally it must raise the possibility that the Verchoyan sequences have been miscorrelated, Asselian. rocks and faunas having been referred by Menner et a/. to the Late Carboniferous. Certainly, in this regard, many of Menner•s Lower Permian faunas are Sakmarian rather than Asselian, which would allow a basal Permian rather than Late Carboniferous age for some of their pre-Sakmarian suites.
Options over the Carboniferous-Permian boundary
From the foregoing review, it would appear that we face several difficulties in adjudicating a boundary. Overall, post-Moscovian faunas are fairly similar until the end of the Sakmarian Stage, and there is, apparently, no one indisputable key, plant or animal, that offers a widespread guide to the base of the Permian System. Fusulinacea have assumed the prime position in the Ural Mountains. But they were not particularly widespread, formed several contemporaneous provinces, and the nomenclature for the key genera are tangled in a bewildering maze at the moment. Ammonoidea would appear to be only slightly less provincial, and there is a critical apparent gap in the sequence in the Urals for the basal Asselian. Perhaps we should ignore Fusuline evidence, and allow the base of the Permian to coincide with the entry of 'basal Permian ammonoid genera' in the mid-Asselian. But we know of other instances where ammonoid genera are missing or belonged to a local and unusual community, and cannot judge yet the significance of the absence of Asselian ammonoids from the basal Asselian. Clearly Ruzencev ( 1 952, 1 965) considered that the absence was not significant, and gave greater weight to the Fusulinacea. The Brachiopoda at this stage are no more useful. Although they are much more widespread, and therefore less affected by environmental paramete�s, they are poorly known for the critical sequences of the Urals, especially for the Assehan . and Sakmarian Stages. It is thus impossible to finally correlate betterknown brachiopod sequences. But enough is known to offer several alternative models for correlation, pending the overdue revision of the brachiopod faunules.
Option 1 , Fig. 1 2 In Option 1 , reliance is placed on the ranges of brachiopods as in Cooper & Grant
( 1 973), Wit? a� attempt to synchronise the incomings of such genera as Kochiproductus and Teguliferma, renewal of Tomiopsis and renewal or incoming of Attenuatella as

Regions Significant for Placement of the Boundary 49
�arking the base of the Permian Period. This yields an impressive array of first beginnmgs, for the Fusulinacea show similar significant incomings with Schwagerinids (in the Gaptan�) and . 'Pseudofusulina' (as variously understood, including the Carnian Alpsand Soviet Umon), and the gastropod Omphalotrochus, and bivalve Eurydesma (except f�r a report of an earlier form in Argentina by Gonzalez, 1973), and probably the bivalve Atomodesma. The overall picture is reasonable, because it suggests that the Permian Period commenced with glaciation, recorded by the Seaham deposits in New South Wales, Australia, which coincided with the widespread introduction or reinvigoration of 'cool-water' genera such as Eurydesma, and the brachiopods Kochiproductus, Tomiopsis; and Attenuatella, and the evolution in palaeotropics of many new Fusulinacea and Brachiopoda. Correlation is sustained between the Gaptank and Orenburgian on the basis of ammonoids, but the Suren fauna, with its indistinctive ammonoids, is relegated to an insignificant part of the basal Asselian. As a variation, the Gaptank might be solely Orenburgian, not Suren, and the range of Uddenites and Prouddenites should be reduced accordingly. The Fusulinacea suggest that the Surenan Substage is more significant than this, but they may have been of only local importance. 'Pseudofusulina' s.l. assumes · high value as a Permian index, though there is evidence, discussed previously, that at least some species perhaps mistakenly ascribed to that genus entered the Pamirs earlier. If the Seaham flora of Australia were regarded as Stephanian C rather than A as in Helby ( 1 969), and were basal Permian in age, we would have a complete floral succession.
Although we need not discuss the Siberian faunas, it appears probable under this scheme that several of the faunas assigned to the Late Carboniferous by Likharev ( 1 966) would also be Permian in age.
There are some objections, but none are firm. My overall impression of at least the Gshelian brachiopods is that they are very close to those of the underlying Kasimovian faunules, whereas the Surenan brachiopods· in the Urals see the introduction of numerous Juresania (though not limited to this horizon), and Tomiopsis. A further objection lies in the treatment of Pseudofusulina. It appears very difficult to enforce a Permian age for this genus or at least species so named in the Pamirs, south Fergana, where associated brachiopods appear to be very close to late Moscovian species (Volgin, 1960), but perhaps the species concerned should be reassigned.
The range of Duartea deserves more study. This distinctive genus disappeared at the start of the E faunas in Canada, and is replaced by Yakovlevia. Duartea could berepresented in Gshelian faunas by 'Muirwoodia' pseudoartiensis.
Option 2, Fig. 1 3 High value is placed on ammonoid distribution as interpreted by Ruzencev ( 1965)
and Furnish ( 1 973) with the boundary placed at the base of the Asselian, and above the upper Gaptank fauna. The Canadian Eka fauna is lowered, with its supposedly correlative faunas of Spitsbergen, Kazakhstan, and Austria to match the Orenburgian, and retain links with the Gaptank (chiefly through the incoming of Kochiproductus). This disperses a number of entries that coincided in the previous model, and makes Schwagerina (s.l.) and Pseudofusu/ina (s.l.) pre-Permian, and opens up a gap .in the record of characteristic ammonoids. Brachiopods from Kazakhstan and Austna and Spitsbergen do show a number of Orenburgia� affiniti�s, bu� canno� be assessed as �iscriminating between correlative, or sequential relationships untd the Orenburgian brachiopods are revised. If the correlation scheme �ere correct, it would appear that there is much to be gained from lowering the Permtan boundary, so that it matches significant Fusulinacean and brachiopod incomings, instead of being represented by an ammonoid 'gap'. From the account in C�oper & Grant ( 1 973).i� appears u�likely that agap is really present in the Glass Mountams column at the position shown m the figure.

a .5 "5 1ft � ... 0 -a � • l.
50 The Base of the Permian
1ft
I I � "' a .t:
-a ..2 a u ·� E 0 i c .. 't -a 'i: 1ft .. Gl 0 Gl 0 a Gl -a a lSI .. � ... 0 1ft 1 A. a ·- .t: Gl :c c E .. Gl ·s: � 0 A. ·;: u .. lSI E 0 .. • � .. a u > c 0 "' � c 1ft 0 •
� .. :. � ""'
a Ill ;: ! .. c a � • Ci -a -a
:a
Ura l s Te xas Yu kon A ustria Australia substage formations brachiopod member formations
zones
Kurmain Neal
Eta Allandale
Ranch U skalik Eo Lochinvar
Suren
Orenburg Mid.
Gaptank E ka zone Kalkarme Sea ham
c 1ft • .. c • -a -a � 0 .. A.
Fig. 12. One optional scheme for correlation of basal Permian sequences.
1ft 1ft 1ft � a I � .. � .t: a • .. ..2 u u u 0 E 1ft 0 Ill � .!! 'i .. 1ft ! "t • "t -a .. Gl .. .. 0 .. "t • 0 > a -a ·;: 1ft at ... . � a 0 a a .!- > a a c .t: E 1ft at E U rals � .t: 0 • A. • 2 .t: c; � :: E 0 .. � U S S R u � :, , c .. 1ft � l a
"' 0 c • ""' "' .. Ill ·c:
I
• • Kurmaln .. ·c: -a -a • �
a -a 0 Uskalik ; -a .. .. :a A. a �
S uren Ci
Orenburg
Gshel
Kasimov
Fig. 1 3 . Alternate option for basal Permian correlations.
Texas Yukon Austria
Pseudosch • E ta
Kalk Neal - ? Ranch Up.
?Eo Kalkarme
Up. --Kalkreiche
M. Gaptank E kct Kalkarme
D

Regions Significant for Placement of the Boundary 5 1
Cl 5 :.. :II "' :II ... 0 , :II • Ill A.
"' "' :II , .. ·- u .! c :II 'i: 1 · j .! & .! :II J j :i
c l t l ..��� u X c �lit :Ill:
Cl E "' "' • • ..
, "2 0 a E ! 0 c :II .., "' I •
"' .. • 1: = e , II i i l t
'V ::�� , II � ; 6
.. J! Cl .. c u � u 0 "' 'i: • .. .!: "' Cl ..
Cl Cll E Ura ls Texas Cl .. .Ill l. JlI Kurmaln Neal
Uskallk Ranch - - -
Suren
Orenburg Gaptank
Gshel .. • Kaslmov .. i. J: A. II ij
Fig. 14. Alternate option for basal· Permian correlations.
! ..
I· i :! t .. .5 -= .s l • 2 ' i f 2 1• u .Ill l. JC � '
.! .. .. II &
"' .! .! ! � .. .. ; 1i .! , 5 II 0 i 1 Ef E ! c 0 c "'
• "' !: • c:
!: • c: :aj l
Ura ls
Kurmaln
Uskallk
Suren
Oshel
Texas
Neal aanch
Gaptank
Yukon
Eta
Eo
Ek
Yukon
Eta
Eo
Eka
Fig. 15. Preferred option for correlation of basal Permian sequences.
Austria
u. Kalkarme
u. Kalkrelche
M . Kalkarme
Austria
Pseudosch Kalk
U.l
�- South Wales
Allandale
Lochinvar
s-ham
�South Wales
Allandale
Lochlnvar
M. Kalkarme Seaham

52 The Base of the Permian
Option 3, Fig. 14 In the third scherne, the Canadian sequence i s matched more closely with the
Soviet sequence on the basis of brachiopods, the Eka zone with the Surenan Substage on the basis of Tomiopsis, and the D faunas with the ?Orenburgian to Kasimovian brachiopods. But the Gaptank and Orenburgian are intercorrelated by means of ammonoids. This appears to leave the brachiopod and Fusulinacean incomings in disarray and the positive links of the Orenburgian ammonoids are not reinforced by any notable entries of new forms at the start of the Permian. As a variation from previous charts, the traditional correlations of the Austrian and Australian sequences are shown as Late Carboniferous, i.e. Kasimovian. The scheme may well prove to be the right alternative, for it has essentially extended the column. This would accommodate well a Carboniferous age for Attenuatella in Verchoyan (Menner et al., 1 970). But as a result, lengthy unconformities have opened up in the Glass Mountains and Australia, and no confirmation is yet known of three rather than two cold episodes in the Late Carboniferous and Early Permian. Nonetheless, I incline to this view.
Option 4, Fig. 1 5 The fourth alternative will be the one adopted herein. It shows the Gaptank as cor
relative with the Surenan Substage, and delineates a fairly well defined CarboniferousPermian boundary, marked by the incoming of Schwagerinids (but not perhaps Pseudofusulina), with the incoming brachiopod Kochiproductus, renewal or incoming of Attenuatella, and the renewal of Tomiopsis. Further support from the brachiopods appears to be provided from the underlying faunas, which through their characteristic suite of Duartea, Kozlowskia, Brachythyris, Choristites, Krotovia etc., form a general Late Carboniferous suite, involving the Missourian faunas of United States, the Kasimovian-Gshelian faunas of Russia and D faunas of Canada. As in other interpretations, the position of the Seaham beds in Australia is difficult to resolve, and not entirely relevant to the problem. They could be fitted into the Early Permian, at the entry of Eurydesma in Australia, and shortly before the entry of Glossopteris. And part of the debated Austrian succession, though shown as Permian, could . be Late Carboniferous without critically affecting other correlations.
This scheme seems to fit best with most evidence, and of course requires least change in world correlations, or the Permian-Carboniferous boundary, except for the United States, where the Virgilian Stage may need to be treated as Permian, following Cooper & Grant ( 1973). But the Orenburgian could still be Permian.
The proposed solution is of course tentative, and it must be ·emphasized, needs verification from modern studies of brachiopods from the Urals. The data agree well with those adduced by Russian experts, including Ruzencev ( 1 952), and also agrees with early conclusions by Miller ( 1 93 1 , p.384, 385) in which he accepted a Permian age for the Uddenites shale, and allowed that nearby Prouddenites faunas could be slightly older, as Late Carboniferous. But an intriguing paper by Miloradovich ( 1940) underlines the need for caution. He showed that Brachiopoda, Foraminifera, and Bryozoa declined sharply in numbers at the Pseudofusulina* horizon, (?Orenburgian) which co.uld . imp I� onset of glaciation, and the start of a new period, supporting Option 1 of th1s d1scuss10n. Unfortunately, the Asselian values were not broken down into zones by Miloradovich ( 1 940), so that their numbers may have been inflated.

4. Biochronology for the Marine Permian
ASSELIAN STAGE
Urals
The Asselian Stage of Ruzencev ( 1937, 1952) marks the commencement of the Permian Period. It is based on rocks and faunas exposed in the Ural Mountains of Russia near the Rivers Kiya, Sintas, and Dombas, in a variety of sedimentary rock types ranging from thick shale and flysch, to polymict conglomerate with large broken block� of reefs and bioherms. Ruzencev ( 1 952) relied primarily on the incoming of four ammonoid families Perrinitidae, Metalegoceratidae, Popanoceratidae, and Paragastrioceratidae, to distinguish the Asselian faunas from the underlying Orenburgian faunas at the top of the Carboniferous Period. Three successive assemblages were recognised. The basal horizon is characterised by numerous species of Glaphyrites, a genus common also in the underlying Carboniferous, and the absence of many younger genera typical of the Permian Period. In the middle horizon Juresanites (Metalegoceratidae) and Paragastrioceras (Paragastrioceratidae) appeared for the first time. In the upper Asselian the genera Sakmarites, Tabantalites and Protopopanoceras made first appearances with a number of other species, and G/aphyrites and Neoglaphyrites disappeared.
This three-fold division of the Asselian is also reflected by faunal divisions based on Fusulinacea, abundant Schwagerinids being the most conspicuous and diagnostic fossils of the stage in the Ural Mountains. Rauser-Chernossova (e.g. 1 937, 1940) recognised three zones on the Russian Platform: (Top) 3. Schwagerina• sphaerica Raus.-Chern. & Schwerb., Pseudofusulina• firma
Sham. 2. Schwagerina• moelleri Raus.; Pseudofusulina• fecunda Sham. & Schwerb. I . Pseudofusulina• vulgaris (Schell.) and Schwagerina• fusiformis Krot.
This three-fold subdivision is best expressed by use of the names of the horizons within the Asselian beas, following Likharev ( 1966): (Top) 3. Kurmain-Kurmaian Substage.
2. Uskalik-Uskalikan Substage 1 . Suren-Surenan Substage The three-fold subdivision may be traced widely around the world, and it is
proposed that the three be elevated to substage rank, within the Asselian Stage. The brachiopod faunules of Asselian age in the Ural Mountains are very large, and
have been described as part of the Uralian assemblage in numerous studies, notably by Chernyshev ( 1 902) and Stuckenberg ( 1 898), as summarised by Miloradovich ( 1949) and Stepanov ( 1 95 1). Genera include Enteletes, Orthotichia, Derbyia, Meekella, Kochiproductus, Linoproductus, Ambocoelia, Phricodothyris and Tomiopsis. Unfortunately they have not been re-examined for many years, and we do not know details of the faunal succession within the Asselian Stage. Indeed it is scarcely possible to discriminate Asselian brachiopods from those of Sakmarian age, perhaps because of difficulties for me in coping with Russian literature. Therefore it has been necessary to rely on other brachiopod sequences, correlated directly or indirectly with the type Asselian by means of Fusulinacea, Ammonoidea, or successional detail.
Many world sequences show a three-fold subdivision, which is assumed to match


Asselian Stage 55
the three-fold subdivision found in Ammonoidea and Fusulinacea in the Urals. The basal and topmost subdivision, assigned to Asselian 1 and 3, are characterised widely by A_tten_uatella, . and fr��uently are a�companied by Kochiproductus and especially Tomzopszs and L1charewud genera, part1cularly in the later Asselian or Kurmaian Substage. Such genera, especially Tomiopsis, are suggestive of Australian-type faunas, and so indicate cool conditions (Waterhouse, 1971 ). The intervening faunas, believed to be equivalent to the Uskalikian Substage, have various genera also found in the Carboniferous, such as Orthotichia and Kutorginella, and have large coral reefs suggestive of warm-water conditions. ·
Donetz Basin, North Russia, Siberia
To the south of the Urals a sequence chiefly of carbonates up to 3000m thick in the Donetz Basin near the Don River (Fig. 1 6) contain the Schwagerines typical of the Asselian Stage, with brachiopod faunas described and listed by Yakovlev ( 1 9 12) and Gr�bau ( 1 9 3 1 , p.450). A three-fold division is recognised: the Kartamish Suite, with plants, and correlative Kalitven Suite, with marine faunas, perhaps equivalent to the Surenan Substage; the Pokrov Suite, and overlying Vrynetzev Suite, perhaps of Kurmaian age. Detailed ages are not clearly established, pending full analysis, which requires re-examination of the faunas. According to Likharev (1 959), small Foraminifera suggest a middle and upper Asselian age, but this is not sustained by Likharev ( 1 966). Some discussion has centered around the age of the underlying so-called Araucaria Suite, with plant remains, and the Fusulinacean Triticites and Pseudofusulina*, regarded as Permian by some authorities, but now generally classed in the Carboniferous by Russian workers. The presence of saline and copper deposits in this sequence suggests the possibility of a basal Permian age, correlative with the basal Permian ice-sheets of Gondwana, by analogy with the copper beds and salt deposits that formed in the Zechstein Group during episodes of mid-Permian glaciation.
From Timan, north of the Urals, the lndiga Limestone about 30m thick is correlated with the Surenan Substage and the Nenetz beds are correlated with the Uskalikian and Kurmaian Substages on the basis of Fusulinacea (Barkhatova, 1964). Brachiopods were listed and described by Barkhatova ( 1964, 1968).
At Pai Hoi, east of the Petchora Basin and west of the Ob River the Yunargin Series contains the Zaostren Horizon at the base (Table 14), with Pseudofusu/ina* krotowi (Schell.) and Schwagerina* fusiformis Krot., indicative of an Asselian age, with several brachiopods, including Spiriferella mica Barkhatova. The overlying Sezim Suite, 6 to 40m thick, has a rich brachiopod fauna with Neog/aphyrites and Glaphyrites, suggestive also of an Asselian age, supported by Yakovlevia, Brachythyris ufensis Chernyshev, and Ura/oproductus stuckenbergianus (Krotow). However the Sezim Suite was assigned to the Sakmarian Stage by Likharev ( 1966).
From Taimyr Peninsula (Fig. 1 6), the faunas of the Turuzov Suite, described by Ustritsky and Chern yak ( 1963, 1 967), contain G/aphyrites, with Linoproductus achunowensis Stepanov (also in the Sezim beds of Petchora), Licharewia, Paeckelmanella and Tomiopsis, to suggest a probable early or perhaps late Asselian age (Table 14), for at least part of the beds. Menner et a/. ( 1970) showed most of the Turuzov Suite as Late Carboniferous (Table 1 5). At the Haraulakh region near the mouth of the Lena River, the Tiksin Suite, although assigned to the Permian in Likharev (1 966, p.343), has ammonoids of Carboniferous aspect, including Eoshumardites and Owenoceras, with Spiriferella gjeliensis Stepanov, which, if correctly identified, is chiefly Carboniferous (especially Kasimovian-Gshelian), but ranges into Asselian deposits. The Tiksin Suite was shown as principally Lower Carboniferous, ranging up to Middle Carboniferous, by Menner et a/. ( 1 970), as in Table 1 6.
The Djuptagin Suite of Set Davan contains small faunas with Jakutoproductus and

56 Biochronology for the Marine Permian
Stage or Substage Serie s
Ufimian
E-1 ::::> � l:t:
Nevolin 0 ::>
Irenian
?Filippovian
Krasnoufimian
z Sarginian H
(!) l:t:
?Aktastinian 1':( z ::::>
Sakmarian >t
?Kurmaian
?Sure nan
Beds
Petchor 1100 - 33 00 m
In tin Suite 2 70 - 1100 m
Q) Rudnitz +J subsuite ·.-I ==' Ul 150 - 5 00 m
+J � � Ayachargin 0 > sub suite Q) H 2 50 - 700 m
Undifferentiated
Tala tin Suite 450 m
Belkov Subsuite 450 - 800 m
Upper 500 m Gus in
Lower Suite
40 m
( =Glini Suite)
Sezim Suite 40 m
Zaostren Suite 30 - 150 m
Table 14 . Correlation of Permian at Pai Hoi , north Russia (Likharev , 1966 , Table 9 ) Northwe st Petchora
Basin

Asselian Stage 57
Cancrinella, correlated with the Irbichan and upper Turuzov faunas by Abramov ( 1 970).
Part of the Verchoyan Suite would appear to be Asselian, as Menner et a/. ( 1 970) recorded Neog/aphyrites. The bulk of the faunas, judged from lists in Likharev ( 1 966), would appear to be Sakmarian.
In North Verchoyan, the rocks assigned to the Verc .. ..,yan �;uite includes Kochiproductus porrectus (Kutorga), with Jakutoproductus verchoyanicus, and other wide-ranging species such as Cancrinel/a janischewskiana (Step.), Neospirifer subfasciger (Likharev), Pseudosyringothyris inopinatus Sol. and ammonoids Uraloceras simense Ruz., Paragastrioceras and Neopronorites. Some of the brachiopod species persist into the overlying Megen, Echi and Haldjin Suites, of Sakmarian age.
The Imtandjin Suite of south-east Verchoyan contains indecisive brachiopods such as Jakutoproductus, Cancrinella, Pseudosyrinx (Kashirtsev, 1959), and the ammonoids Somoholites and Shumardites. It is not possible to be certain of the ag�from mid Carboniferous, to as young as upper Asselian-Menner et a/. ( 1 970) preferred a middle Carboniferous age. The Ekachan Suite of Set Davan at first sight appears to be Asselian, with Kochiproductus, Tomiopsis, and Uraloproductus, allied to a late Asselian species of Oregon and Yukon Territory, Canada, and ammonoids Owenoceras, Somoholites and Stenopronorites ekatshanensis Popov, unlikely to be as young as Kurmaian age. The beds were regarded as Late Carboniferous by Menner et a/. ( 1970), and this may be correct because 'Kochiproductus' is incorrectly identified by Abramov (1 970, pl. l 3, fig. 1 -5).
Further east in the Kolyma River-Omolon Plateau region (Table 1 5), the Paren Horizon, 25- 1 OOOm thick, is likely to be upper Asselian, judged from the brachiopods Attenuate/la omolonensis Zavodowsky and 'Pterospirifer terechovi Zavodowsky, with Paeckelmanella, Tornquistia, Anidanthus, Linoproductus, Jakutoproductus, and Martiniopsis. The fauna could be as young as Sterlitamakian, within the Sakmarian Stage as shown by Likharev ( 1966) or Zavodowsky ( 1970), but it seems unlikely. Underlying the Paren Horizon is the Burgali Horizon or Suite, containing a generalised Permian-type brachiopod fauna, with Tomiopsis, but without Attenuatella. The presence of a marginiferid, Septacamera and Choristitinids, together with the ammonoid Owenoceras suggests that the fauna could be Late Carboniferous, if Omphaloirochus can be taken as that old. A Stepanovie/la-Iike form described by Zavodowsky ( 1968, 1 970) as Linoproductus popowi Zav., Sajakel/a zyriankensis Zav. like a Spanish species, together with many Tomiopsis and Brachythyris ufensis, on the other hand allow a Permian age, both early and middle Asselian. Unfortunately, the report of Kochiproductus by Zavodowsky ( 1 970) fails to finally confirm a Permian age, because Kochiproductus levinsonlessingi Zav. is far from typical of the genus.
Other small Permian faunas that are likely to be of Early Permian age in east Siberia were described from east Zabaikal by Kotlyar & Popeko ( 1967) in the Shazagaitui Suite. Brachiopods could prove to be Permian, though correlated with the Carboniferous by these authors, perhaps correctly if they were right in identifying Levipustula, for this has been assumed to typify early Moscovian faunas in both hemispheres. The fauna is somewhat like that of the Burgali Suite of the Kolyma River region, sharing Jakutoproductus, Cancrinella cancriniformis (Chernyshev}, Orulgania, Tomiopsis, and Taimyrella. Andrianov ( 1 963, 1 966) correlated the fauna with those of the Uchagan beds of west Verchoyan which include supposed 'Ambocoelia', an ally of Attenuate/la. Further links to Orulgania (Ypenchin, Suyorgan, and Haldan Suites) and Haraulakh (Tugasir Suite) are suggested by ammonoids. The Chiron Suite of this area contains Early Permian brachiopods (Kulikov, 1 959, 1 965a, b), including Martiniopsis and other forms suggestive of an Asselian age. The Hiposhin Suite of Zabaikal has Anidanthus indicating an Asselian or Sterlitamakian age (Kotlyar & Popeko, 1 967, p.22). West Zabaikal yielded a small fauna in the Chikoiskogo Suite, possibly as young

58 Biochronology for the Marine Permian
World Standard Central SW Verchoyan
Substage Taimyr Peninsula
( Dorashamian Stage) Lower Tufolavov beds
(Dj ulfian Stage) Chernoyar ?Dulgalach Sui te
Chhidruan Horizon 600 - 1600 m
Kalabagh ian Upper Baikur Horizon
Sosnovian Lower Baikur Horizon
Ka linovian Delenj in Sui te
.. . 600- 700m � ..... k Irenian 41 en
Fi li ppovian ... " .0 ....
Krasnoufimian Sokolin Horizon '0 Tumarin Sui te 1:: lzl 500 700 m
Sarginian
Aktas tinian Birrang Horizon ., 41 ..... k
Ster litamakian 41 Echi Sui te en 200-900 m
1:: Tastubian ( ? Evenk Horizon ..... Kigi l tass 1).,
w . Taimyr) e Sui te 0 !-< 600 - l l OOm
Kurmaian
Uskalikian
Surenan Turuzov Horizon Imtandjin Suite s 1 900 - 1200 m
Carboniferous Tiksin Suite 2000 - 3000 m
E Verchoyan Kolyma , Omolon Set-Davim
Imtachan Sui te 300 700 m
Hivach Hori&on 200 - 2000m
Gij igin Horizon 70m
Chambin Sui te Omolon Horizon 500 - lOOm 100 lOOOm
Menkechen - - - - - - - - - - - - - - - - - - - - -- ·
Sui te 900 1 100 m
Dj igdalin Horizon
Dibbin Sui te Djeltin Horizon 500-700m 40m
(Tiri Bas in)
Munugudjak Horizon 35 - 60 m
Kukkan Suite Yasachnin Horizon 1 100- 1300 m 1 10m •
(Tompo Basin) Irbichan Suite 800 lOOOm
Par en Suite
Burgali Suite 200m
?Ekachan Suite
Table 15 . Correlation of Siberian sequences (see Fig . 16) .

I
�
r.:l
AI
� AI
:;:J
1-1
� r.:l
AI
� � 0
..:I �
r.:l
AI
(/)
� :;:J
� r.:l
�
1-1 �
r.:l s
ill Q
6
1-1
:E:
�
�
..:I
w
Ha
ra
ul
ak
h
Or
ul
ga
ni
a
Ve
rc
ho
ya
n
Se
t
Da
va
n
Ta
im
yr
Du
lg
al
ak
h
Du
lg
al
ak
h
Im
ta
ch
an
Ch
er
no
ya
r
Sy
ur
en
D
el
en
ji
n
Ch
ambi
n
Ha
ra
ul
ak
h
Bd
ik
ur
Tuma
ri
n
Me
nk
ec
he
n
--
--
--
-
Ha
ld
ji
n
Di
bi
n
So
ko
l i
n
Ec
hi
B
ir
ra
ng
Ve
rc
ho
yan
E
ch
i
Me
g e
n
Ki
gi
lt
as
s
Ku
kk
an
Su
rk
ec
ha
n
Tu
ru
zo
v
Ha
ld
an
So
lo
n c
ha
n
Su
yo
rg
an
E
ka
ch
an
Im
ta
nd
ji
n
Ha
ta
li
n
Ma
ka
ro
v
--
--
--
--
So
yb
ol
Y
up
en
ch
in
Ha
ti
na
kh
S
et
ac
ha
n
? T
ik
si
n
Bi
li
ka
t
Ob
la
ch
an
Lo
we
r
Ca
rb
on
if
er
ou
s
Ta
bl
e
16
. C
or
re
la
ti
on
o
f
La
te
P
al
ae
oz
oi
c
su
it
es
i
n
Si
be
ri
a
af
te
r
Me
nn
er
e
t
al
. -
-
19
70
, T
ab
le
2
, p
. 54
. "
Up
pe
r
Pe
rm
ian
"
co
mme
nc
es
a
t
Ka
za
ni
an
St
ag
e.
I
:to. � - i:i
' ::s
!'.!
�
� VI
\0

60 Biochronology for the Marine Permian
as Sakmarian (Kozubova & Kulikov, 1958; Kulikov, 1 969; Maslennikov, 1 957). Odontospirifer is present with Jakutoproductus.
In summary, most of these faunas from Siberia are difficult to correlate, partly because of the high northern palaeolatitude, and partly because the Late Palaeozoic rocks and faunas of Baikal are still incompletely monographed. It must be-recalled that most of the extensive studies commenced only in the last few years, and may require revision through more refined collection from more closely controlled stratigraphic horizons. Thus the recognition of the Paren fauna in Zavodowsky (1970) marked a considerable improvement over slightly earlier work.
From Kazakhstan the large Kokpecten fauna described by Sarytcheva et a/. ( 1 968) is judged to be early Asselian in age, with Attenuatella and Tomiopsis, although it retains many Gshelian-Orenburgian genera. In · south Fergana the very rich Karachatirian faunas fall into three divisions f, g, and h, possibly correlative with the three Asselian substages (Table 1 7), both on the basis of Fusulinacea and Brachiopoda (Likharev, 1 946), with additions from Likharev ( 1966). However the faunas were shown as Sakmarian by Ustritsky ( 197 1). Th� basal horizon f has Buxtoniids, Scacchinella
Local Suite World Standard Stage
Pamirian Chapsai Suite, lOOm ? Djulfian
Shakharsev Suite 400m
Murgabi11n Iollikhar Suite ?Kazanian 20-2000m
Gundarin Suite Baigendzinian 30-700m
Safetdaron Suite Aktastinian 30-lSOOm
Darvasian Chelamchin Sui te ? S ter li tamakian
200-400m
Zigar Suite Tas tubian l50- l200m
Upper Kurmaian
Karachatirian Karacha tirian Middle Uskalikian Suite
150- lOOOm Lower Surenan
Table 1 7 . Darvas Permian , South Fergana (Likharev 1966 . Table 14) .
and A:fa�tinia; the .middle horizon is ncb in Entetetes, Meekella, Isogramma, Teguliferma, Hustedza, and Notothyris, suggestive of warm waters; the upper horizon

Asselian Stage 61
includes more Buxtoniids, Linoproductus, Martinia and lacks such notably warm-water genera as Scacchinella and Teguliferina. Fusulinacea are highly significant, with Rugosofusulina complica (Schell.), Pseudoschwagerina ex gr. uddeni (Beede & Kniker) from the basal Karachatirian; Paraschwagerina pseudomira Mikl.-Makl., and Robustoschwagerina tumidiformis Mikl.-Makl. from the middle Karachatirian and Sphaeroschwagerina carnica (Schell.) and !Pseudofusulina* ex. gr. vulgaris (Schell.) from the upper Karachatirian fauna.
Vol gin ( 1 960) has also described a Karachatirian faunule from south F ergana. The underlying Late Carboniferous Dastar horizon C23 includes Rugosofusulina moderata (Raus.) and R. alpina (Schell.) as if to suggest a possible Early Permian age also, by comparison with the Carnian Alps of Europe, with possible Sajakella ('Muirwoodia') aurita (Volgin) and Spiriferella asiatica Volgin like an Asselian species of Canada. Otherwise the Dastar fauna does not resemble Asselian faunas very closely. Underlying beds contain an echinoconchid very like a Missourian-Moscovian species from Canada and the United States.
From Pasha Asha, Tian (or Tien) Shan (Fig. 1 7), comes another brachiopod fauna here correlated with the late Asselian on the basis of Cancrinella, Martiniopsis, Martinia, Brachythyris and Linoproductus species (Likharev, 1 946). Hayasaka ( 1 922, p. l 6) and Keidel ( 1 906) discussed faunas of the Koktan Ridge, south Tien Shan, and Loweneck ( 1 932) recorded several collections including a large fauna from Kukurtuktal in the Tien Shan, with supposed Pseudoschwagerina princeps, Rhipidomella, and numerous other brachiopods like the Asselian faunas of the Urals.
Karakorum Range, Tibet
Karakorum faunas to the south are somewhat similar. A fauna at Gilgal Camp, Gasherbrun, was recorded by Merla ( 1934) and redescribed by Sestini ( 1965d, p. l67). It is likely to be either late Asselian (probably) or Sterlitamakian, judged from Tomiopsis sokolovi (Chernyshev), Crurithyris, regarded as an ecologic and temporal as well as morphologic ally of Attenuatella, syringothyrinids; and a general similarity to the Pasha Asha fauna of the Tien Shan. Species are allied to those of the Kalkarme beds of Austria, to favour a possible Asselian age. On the other hand the fauna differs from the Shaksgam Valley faunas of the Karakorum, described by Renz (1 940a, b), here believed to be of definite Asselian age. The Shaksgam faunas are very large and accompanied by Fusulinacea identified as Triticites, Pseudofusulina, and Pseudoschwagerina. A middle or late Asselian age seems likely, in view of the presence of Linoproductus cora (d'Orb.), Cancrinella cancriniformis (Chernyshev), Ambocoelia, and Martinia.
Near by in Tibet, north of the Tsang Po or Indus suture (lat. 3 1 o 1 0' N; long. 8 7° E), Reed ( 1930) described Early Permian faunas in black limestones with Fusulinacea, including recorded but not illustrated 'Pseudoschwagerina princeps' and 'Schwagerina
fusiformis', the latter typical of the basal and middle Asselian according to Likharev ( 1 966). The collections, examined. by the writer at the Geological Survey of India, Calcutta, include Attenuatella, described as Dielasma cf. plica not Kut. by Reed ( 1 930, pl. 1 , fig. l 9). The brachiopods have much in common with species from Kokpecten, Kazhakstan, Gilgal Camp of Gasherbrun, and Shaksgam Valley.
Mongolia, China
Chao ( 1 965) recorded an Asselian fauna from Mongolia that included Triticites,
Schwagerina, and Quasifusulina, with possible Attentuatella figured as C:rurith!ris.
Specific affinities amongst the brachiopods l�e. to some extent w1th .m1ddle Karachatirian; but chiefly with the upper Karachatman faunas, and a late Assehan age is most likely.

62 Biochronology [or the Marine Permian
system series Formation Rock Character
Trias Lower Thin-bedded limestone or shales
0'1 4J Grey to deep grey -� § thick-bedded , finely Ill ..., crystalline iimestone 0'1 Ill � -� with chert nodules
ij tj ..:1 95-lOSm '" 0'1 � "" Grey, thick-bedded· .s 0'1 4J subcrystalline lime--� �
1t B stone , seldom with Ill chert nodules '" � or .<: cherty bands 0 "" ::1 ..:1 3: 370-330m
Light grey massive limestone with some chert nodules
35-l72m
z 4J Light grey , thick-..: � bedded limestone 0 intercalated with ..., siliceous lime-H Ill stone ::1: 4J
a l90-265m D: '"
.:: ..:1 111 [il '"
Grey to deep grey ::1 .:: thick-bedded , finely 0 "' .... ,;( crystalline lime-Ill 0 0'1 stone 111 2 37- 305m .::
., ::1:
><
4J Deep grey massive § cherty limestone ..., Ill fi l24-280m .... ..:1 ., .... Ill .<: .... 6
Carbon- Upper Maping Whitish grey massive iferous limestone limestone
200m
Fusulinid zones Standard Stages
(As in Shenl]) ·?-
Pal-a£o fusul.ina Zone Dorashamian Pa1-a£ofusuUna Reiche Una NankineUa
CodonofusieUa Zone Djulfian Codonofusie Ua Reiche Una Nankit�el-1-a Spha£r-u'Lina
:tabeina Zone Punjabian :tabeina NeoschuJageZ'ina Neomisel-Una KahZ.eZ'ina ChuseneHa Lantschichites
NeoschuJageZ'ina Zone Kazan ian NeoschuJageZ'ina Verbeekina SwnatZ'ina Afghanel-1-a PseudtJda Uo Una Chusenel.l.a SchuJageZ'ina PseudtJ fusu'Lina
CanceHina Subzone Kungurian Cancel. Una :tangchienia Parafusul.ina Verbeekina RussieHa 4J PseudtJfusul.ina § lSI Chusenel.l.a
<::! SchuJageZ'ina � PseudtJdaZ.iol.ina ., N ;:! �sel.l.ina Subzone IV Baigendzinian <Q ¢; �sel.Zina
� Parafusul.ina SchuJageZ'ina III ToZ'iyamaia
sakmarian NagatoeH.a II Staffel.l.a Nankinel.l.a PseudtJ fusul.ina I
Pseudaschwagerina Zone Asselian
Table 18. Sequence of Permian in South China , following Sheng (196 3 , p. 142) .

S EAi
tp�l
�TIN
SI(
,. ...
........ .
= ::· ·
·· )J
····
..
. . · · . ..
.... : .
. ··
I
N
G �
f<
.•··
•·••
. '•/
••••••••''•
·.... , •• ·
··
)
o '
'
G
E .1)
.....
. , ..
·· .......
.-I
/ S
y "
�-.. �
I N
-··1''''
" _,
-.J
. ·"
"'""
' '
·..
\ ,. . ..
.,..:..... (
·... ,_.
....
" ..
......
.
Lwntk-.
�\a
i Mts
�
,
· ·.. .
<::·;· ;· ;· :· :· �
��,, ,./,
0 ·u>· �""· -· · ;:J
/._l
.. .. ...
...... fjf' ... ······
.. . ·
-·w
en l
0
.,. '
'
0''"
J-
····
·; .
..;,�···· . .
.' /
. ..
. ..
......
. · .
·-
··
/
i."''
M f
�:
W•••
• (Loo."
� •
/ Paote
......J
•-•��o
�ouch.!Cif'9
. ... ..
......... '.
�7
; .
. ··
-·�·
"'
.
• . ••
r,-
:· ····
Toov•••
� IH
LI
E T
... 1•'
}_-0 K�
�
la pa i�· ··.·
· � • •
T B
\./\ \1
·-.:
•::
·: ···· ....
S 1> 11
�. • C
� i lic
hun
c;6\..
. : •• • • • ·
· � •
• � ••
,'So,.
··�
�alo
hal
T tr.,
� E
T •••
ICu "'oo.; �
::..
. •
p
( � ,
Dol.
..
. ....
• •• •o
.,�;
:
o ·· ..
"---.)r--·
··· · .:� p
·� ··:
'• !
• •
S H
E N
S I
'• , ., •
· ····· ·· �
···c
�i;·:�
··
.::··'·�. o
" •
'!.·i'f'
a��;;;,:�,.,
•
····.),:
······· ....
. :· ; ."'"'JI"of
�:;f ank
� 5
Z
�:''' ..
. H
0 p
E H
••:
', "'
E CH
uA
:. .si'JI'I
� �
\ _,.�
c",.sing
·.
(o,
·. !J
: .
. ....
. . ·•• ••••
t' Q·
�·· ......
.... ·····
· ..... ···
I A.
• luc
•� • .;.··
·!" ·fl • .1-
�t � 1
1·.·
'11i
urA
N\.
··
• ., A
•, (
It e
... .
. .
"l ..q
· ·
· ......
.. ··
CHIN
A an
d sur
roun
ding
region
s .: To
Jr.,.. ao
:".;: ·: YUNNAN
'So·.
�
' • :" ··•
'•• • Yan
gslflt
. . ····.
:· :
. ··
Tun glze ·
�: ':
' •," •• lA
NG
; H
U N A N
:' Lo;in ,: •• ::. KIA
NOSI
•:q,, !,,
•• ,,, �
' "
•• ·• ..�H
uong-T�-Pu
,:__�
·········
····:
:
'-4-(J
'I'-'
1-·
� =
.. � · · .:::.:
f ..
··· ·· ·······
···. ��
� V
RMOSA
..
Fig
. 17
. C
hin
a an
d su
rrou
ndin
g re
gio
ns,
red
raw
n fr
om
Gra
bau
(19
24
, pl.
4).
::t.. � � s· ;::s t"'l
�
0\
w

64 Biochronalogy for the.Marine Permian
The Maping Limestone of China is a white, massive pure limestone with chert horizons, about 200m in thickness (Table 1 8). The formations contain Schwagerinids in three zones named as follows by Chan ( 1963): (Top) 3. Paraschwagerina inflata (Chan)
2. Pseudofusulina(' Leeina') valida(Lee) (Base) 1 . Pseudoschwagerina parasphaerica (Chan)
Both Chan ( 1 963) and Sheng ( 1964) referred the formation to the Carboniferous on the basis of Pseudoschwagerina, but this is a matter of definition, not an alternative correlation. Several monographs have described the_ brachiopods (Chao 1 927b, 1928; Huang, 1 932b, 1 933 ; Grabau, 1 934, 1 936), with the latter work devoted largely to species from the Maping Formation, which had previously been regarded as young Permian, chiefly in the Kwangsi and Kiangsi provinces (Fig. 1 7). About fifty species are shared with Uralian faunas. Other large faunas come from the Payu Limestone and Lungli Limestone of K weichow with more chonetids and fewer Productacea, originally described by Huang ( 1933) and Grabau ( 1 934). The faunas are close to the upper Karachatirian faunas of Fergana (twelve restricted species), and less to the middle Karachatirian (eight restricted species) and also there are similarities to the Shaksgam faunas of the Karakorum. In addition a number of species are shared with the faunas of Tibet and the Gilgal Camp of Gasherbrun. The faunas came chiefly from the middle Maping Limestone according to Huang ( 1 933), and as they do not have late Asselian key genera, may be middle Asselian.
From north China, Chao ( 1 927) described part of the brachiopod faunules from the Taiyuan Limestone in Kansu and Shansi, probably correlative with the Maping Limestone. Further affinities are equally divided between Karachatirian horizons f and g, though considering only key species, most affinities lie with the f faunas, five species, against two for Karachatirian g.
In Manchuria, beds more or less equivalent to the Taiyuan Formation near Shihpu-tsui have yielded a fauna probably incompletely listed by Ozaki ( 1 93 1 ), and large but similar faunas are listed from near Poshan City by Ozaki ( 1 93 1 , p.10), and Hayasaka ( 1922). The preponderance of choristitinids (nikitini, baschkirica) from the Poshan beds suggest a possible Sakmarian age. Meagre faunas are found with 'Pseudoschwagerina' (s.l.) in the Koten 'Series' of Korea.
South-east Asia, Japan
Further large Asselian faunas are found in south-east Asia, including Fusulinacea and Brachiopoda from Kham Kheut, and Muong The, and other localities in Laos and Tonkin (Mansuy, 1 9 1 3 , 1 9 1 9, 1 920) as set out in Table 1 9. A small fauna at Tran-ninh lies at the base (Mansuy, 1 9 1 3 ; Deprat, 1 9 1 3 , p.74), but is judged to be of the same zone as the others by Waterhouse ( 1973a). The faunas are associated with supposed Pseudoschwagerina princeps (not Ehren burg ( =S. moelleri Raus. ?) thought to be Zellia by Kahler & Kahler ( 1 966). They were regarded as late Asselian or Kurmaian (Waterhouse, 1 97 1), on the presence of Crurithyris, Licharewiinids, Martiniopsis, and Tomiopsis. From Thurn Nam Maholan, central Thailand, a somewhat similar fauna was thought to be as young as Artinskian by Yanagida ( 1 966), but Waterhouse & Piyasin ( 1 970) preferred an Asselian age. Other faunas likely to be of identical age are mentioned by Gobbett ( 1 968, p. 1 9) from the Kinta Valley of central Malaysia.
In the Sakamotozawan Series or Stage at the base of the Permian in Japan, Fusulinacea identified as !Pseudofusulina* vulgaris (Schell.) and Robustoschwagerina schel�wieni (Yabe) in the middle of the stage are accompanied by a few brachiopods (Monkawa, 1 967; Kanmera & Nikami, 1 965) which are incompletely described, or very poorly preserved (Table 20). A brachiopod faunule has been recorded from southwest Japan in the Nakakubo Formation by Yanagida & Hirata ( 1 969), with several

Sta
nd
ar
d
Sub
sta
ge
Gr
iesb
ac
hia
n
Ob
gin
an
Ve
dia
n
Ba
isa
lia
n
Ur
ush
ten
ian
Ch
hid
rua
n
Ka
lab
ag
hia
n
Sosn
ov
ian
Ka
lin
ov
ian
Ire
nia
n
Fi
lipp
ov
ian
Kr
asn
ou
fim
ian
Sar
gin
ian
Atk
as
tin
ian
Ste
rli
tam
akia
n
Ta
stu
bia
n
Ku
rma
ian
Usk
ali
kia
n
Sure
nan
Salt
Ran
ge
Fo
rma
tio
ns
Ka
thw
ai
Memb
er
Ch
hid
ru F
m
Kal
abag
h
Memb
er
Mid
dle
W
arg
al
Fm
Low
er
Basa
l
Amb
Fm
Sar
di
, W
arch
ha
Fn
(Con
u'Lazti
a
be
ds)
(Eu
ryde
sma
b
ed
s)
Ta
lch
ir
Fm
Tim
or
Fau
na
l T
hai
lan
d
br
ac
hio
-Ca
mbo
dia
H
or
izo
ns
po
d &
fu
suli
nid
S
iso
pho
n
loc
ali
ti
es
Ish
ii e
t a
l.
(196
9 )
Amar
ass
i M
embe
r D
Ba
s le
o
Pe
tch
ab
un
M
emb
er
C
Mem
ber
B
?T
ae
Wei
K
hao
Ph
rik
Bi
tau
ni
Kh
ao
Ph
lon
g P
rab
Mem
ber
A
Somoh
ole
T
hi.Uil
Nam
M
aho
lan
N
oan
kow
tok
Tab
le
19.
Pe
rmia
n c
orr
ela
tion
s fo
r so
uth
-ea
st A
sia
, b
ase
d o
n W
ate
rh
ou
se
(197
3 a
).
Fm
-F
or
mat
io
n
Lao
s D
ep
ra
t 191
3 .
Lu
ang
Pra
dan
g
7 L
eve
l 6
Lev
el
5 L
eve
l 4
Lev
el
3 L
eve
l 2
Lev
el
1
"'
Mu
ong
Th
e K
ham
Kh
eut
Chin
a F
tm
Chan
g s
ing
r--
--
?--
--
-
Wu
chia
pi�
--
--
?--
--
-
Mao
ko
u ?
Ch
ihsi
a
Map
ing
�
� s: ::s
Cl:l
� � 0\
VI

66 Biochronology for the Marine Permian
species shared with the Maping and Taiyuan beds of China. Accompanying Fusulinacea are correlated with the Pseudoschwagerina minatoi Kanmera Subzone or Zone of the Yayamadake Formation of Kyushu, allotted a vague Sakmarian-Asselian age, but most likely mid-Asselian.
Turkey, A ustria, Spain
A fauna described by Enderle ( 1 90 1 ) from Balia Maaden, north-west Turkey, included fusulines identified as Pseudoschwagerina princeps though the brachiopods suggest only a general Asselian-Sakmarian age.
In Austria (Table 23) the base of the Permian has been assumed to lie at the base of the Rattendorfer beds (Kahler & Pray, 1 963), but underlying faunas of the Auernig complex contain some brachiopods of Permian aspect. However plants and Fusulinacea are usually judged to indicate a Carboniferous age.
Pasini ( 1963) showed that there are three Fusuline communities in the Auernig beds, a basal faunule with Pseudostaffella, Ozawainella, Fusu/inella, Wedekindellina, and Quasifusulinoides, indicating a Moscovian or more probably Kasimovian age, a much higher faunule with Quasifusulina longissima (Moeller), Triticites, and Pseudofusu/ina (Daixina) alpina alpina (=Rugosofusulina in Kahler & Kahler), classed as Gshelian, and a topmost faunule with Nankinella, Sphaerulina, Fusulinella, Quasifusulina and Pseudofusulina, classed as Orenburgian or probably Permian. The latter faunule occurs in bed n of Frech ( 1 894), in the upper kalkreiche beds. The middle faunules occur in beds g-i at the base of the upper kalkreiche beds, and the basal faunule occurs below bed a of Frech ( 1894). The stratigraphic position of the latter fusulines is not entirely clear-probably at the base of the middle kalkarme beds, or in the underlying horizon.
From the accounts in Heritsch ( 1 934) and Kahler & Pray ( 1963), the Auernig is subdivided into several units, especially well displayed along the Garnitzer ridge. The lower kalkarme beds with the Waidegger fauna is stated to be Moscovian (Mayachkovian-Samaran), although the abundance of marginiferids suggests to me a possible Kasimovian age. From the overlying lower kalkreiche beds, Heritsch ( I 933, p. 1 66) recorded Pseudofusu/ina and Triticites with various choristitinids that point to aCarboniferous rather than Permian age. Many compound rugose corals are also present, with Quasifusulina tenuissima (Schellwien). The middle kalkarme beds contain large brachiopod faunules, including Neochonetes, Chaoiella, Linoproductus, and 'horizon a' is correlated with the Krone bed 6 containing the Spirifer fauna of Schellwien, including A ttenuatellafrechi (Schell.), Plicatocyrtia zitteli (Schell.), Sergospirifer carnicus (Schell.), choristitinids and Martinia, with Stenoscisma a/pina (Schell.). Apart from Attenuatella, the brachiopods would appear to have a Late Carboniferous rather than Early Permian aspect, but this judgement is not based on intimate knowledge of the species. They apparently overlie the basal Fusuline fauna described by Pasini ( 1963). Wagner & Prins ( 1970, p.503) stated that this horizon a of Frech and Krone bed 6 contained the Moscovian-Kasimovian Fusulinacea described by Pasini ( 1963), but from the account in Heritsch ( 1934) I would gather that the brachiopod faunas were slightly higher in the Auernig sequence. This would seem to be confirmed from the overall stratigraphic columns and thicknesses involved. Overlying Fusulines Pseudofusulina (Daixina) longissima a/pina (Schell.) with Triticites and Quasifusulina longissima (Moeller) at the g horizon, or base of the upper kalkreiche group suggest, as in Pasini ( 1963), a probably Gshelian age, but an early Asselian age could not be ruled out.
The overlying upper kalkreiche beds appear, from data in Heritsch ( 1 933, 1 934),t� match be�s 1 -n up to bed r of the Monte Auernig section, and thus include thehtghest Fusulme faunule described by Pasini ( 1963). This could be Permian in age, and

Low
er
T
ri
as
si
c
Car
bo
ni
-
fe
ro
us
Up
pe
r
Mi
dd
le
Low
er
Up
pe
r
St
and
ar
d
Fu
su
li
ni
d
So
ut
h K
yu
shu
K
wan
to
M
as
si
f
Se
ct
io
n
Sub
zo
ne
K
uma
M
as
si
f
(co
mp
os
it
e)
.
Yab.
yasu
baen
sis
Kum
an
K
uma
F
m
Lep
. to
riy
amai
V.
ver
beek
i -
Kam
ik
uz
u Fm
sh
irai
wen
sis
Aka
sa
kan
K
oz
ak
i
Fm
Ns
. cr
atiau
Uf
era
As
ak
ai
do
F
m
Nab
ey
aman
N
s.
simp
lex
S
hi
mo
ku
zu
Fm
Paf
. ka
erim
izen
sis
Sh
im
o-
da
ke
Fm
Ha
na
gi
ri
Sa
kam
ot
o-
Pss
. m
orik
atVai
Y
ay
a-
za
wan
m
ad
-Fm
a
ka
Hi
ka
wa
n
Lms
t
---
---··----�'-.
.--
---
---·-
Tab
le
2
0.
Pe
rm
ia
n
of
J
ap
an,
fo
ll
ow
in
g
Ta
ka
i
et a
Z.,
19
63
, f
ig
. 3
a.
Cg
l
-c
on
glo
me
ra
te
; Fm
-
Fo
rm
at
io
n;
Gp
-
Gr
ou
p;
Lm
st
-
Lim
es
to
ne
.
Iw
ai
za
ki
Iw
ai
za
ki
Li
me
-
st
on
e
Ki
ta
kam
i
Ma
ss
if
'
In
ai
G
p
To
yo
ma
G
p
Us
ug
in
u
Cg
l
Yu
ki
-
sa
w a
K
ano
ku
ra
Fm
Gp
S
ak
amo
to
-z
aw
a
Fm
:to.
!
s·
;:s
t'.)
S' � C\
-..J

68 Biochronology for the Marine Permian
to• t s• 20° 2 s•
. �
ao•
FO R L
VESTSPI TSBERGEN
\. SCrkapp
0
B J O R N O Y A
2 0°
1o•
D
I
K m .
100 200
Fig. 1 8. Distribution of Permian rocks at Spitsbergen, simplified from Gobbett (1964, fJg. 1 , p. 18).

Asselian Stage 69
contains Pseu�ofusulina. The brachiopods from the upper kalkreiche beds are very like thos� of.the m�ddle kalkarme beds, according to Heritsch ( 1934) who lists a number of spectes, m�ludmg Attenuate /Ia frechi (Schell.). Plants and compound rugose corals are present, wtth Rugosofusulina alpina communis (Schell.) and other Fusulines, and the age may be as young as Early Permian, as suggested by Pasini ( 1963), to Orenburgian. The presence of Attenuate/la might be taken to imply that these beds belonged to the same zone as either the underlying middle kalkarme beds (more likely in view of overall faun�l similariti.es) or with the overlying Pseudoschwagerina beds, but detailed study is requtred to venfy the exact age. We know so little of the range and communal and ecologic associations of the key species that we cannot rule out an Uskalikian age for the upper Kalkarme, which might be supported simply from stratigraphic and sequential evidence.
At the top of the Auernig sequence the upper kalkarme beds contain numerous compound rugose corals, and plants assigned to Westphalian E(Reichardt, 1 932). But evidence for a Late Carboniferous age is weak, and the beds could be as young as Uskalikian. The lower 'Pseudoschwagerinakalk', 1 35m thick, of the basal Rattendorfer beds contains rare Pseudoschwagerina alpina (Kahler & Kahler) and various brachiopods and is here considered Asselian. The writer found Attenuate/la at this horizon at the Tresdorfer hohe, to increase the likelihood of a Surenan or Kurmaian age. Various faunas have been described (Schellwien, 1 892; Seelmeier, 1937; Metz, 1935 ; Heritsch, 1 935 ; Gauri, 1 965). Auernig-type brachiopod faunas are recorded from Yugoslavia (Croatia) by Simic ( 1937) with 'Ambocoelia', Choristites fritschi, Brachythyrina carnicus and Stenoscisma alpina.
From the supposedly Stephanian A Branosera Formation at Barruelo de Santulla, province of Palencia, north-west Spain, Wagner & Prins ( 1 970) have recorded a midAuernig fauna, including Attenuate/la cf. frechi (Schell.), and Karawankina (initially described from the Trogkofel of Yugoslavia), with some species identified with shells from the Miaohou Limestone at the base of the Taiyuan 'Series' in north China. The fauna is obviously close to that of the 'Spiriferenschicht' and layer 6 of Schellwien in the Carnian Alps, including such species as Attenuatellafrechi (Schell.), Plicatocyrtia zitteli (Schell.), Choristites fritschi (Schell.), Martinia karawanica Volgin, Brachythyrina carnica (Schell.), and others. Many of these forms are shared with, or are closely allied to Kasimovian, Gshelian, and Asselian species.
Spitsbergen, Canada, United States
A large brachiopod faunule from the Treskelloden beds of Hornsund, west Spitsbergen (Fig. 1 8, Table 25) was ascribed by Czarniecki ( 1969) to the Gshelian -8tage, but the fauna could be of basal Asselian age, judged from Tomiopsis petrenkoi Czarniecki and T. lata Czarniecki, species which closely resemble T. ovulum Waterhouse in the Asselian of the Yukon Territory, as well as Crurithyris, or a broad Attenuatella, and Linoproductus coralineatus Ivanov. Orthotetes and Krotovia suggest an Orenburgian age, but may have persisted into the Surenan Substage, as did a number of brachiopod species in the Kokpecten fauna of Kazakhstan. The fauna is likely to to be correlative with the Attenuatella frechi fauna in the Auernig beds of Austria.
The Triticites Zone of the Cadellfjellet Member in the lower Wordiekammen Limestone on Bunsowland (inner lsfjorden), Spitsbergen, has yielded Pseudofusulina together with a modest brachiopod faunule of general Permi�n aspect assigned to .the Orenburgian by Cuthill & Challinor ( 1 965, p. 1 9), but herem regarded as Permtan, perhaps basal Asselian. The middle member, or Brucebyen beds of the Nordenskioldbreen Formation has a marginiferid, and the upper 'Pseudoschwagerina Limestone' or Tyrellfjellet Member is possibly late Asselian, but likely to be Sakmarian, as in Cuthill & Challinor ( 1 965), judged from the brachiopods recorded by Gobbett

Ga
mi
tz
er
A
ue
mi
g
Pr
of
il
e
Se
qu
en
c
Rat
te
nd
or
fe
r
Fr
ec
h
Be
ds
1
89
4
Up
pe
r K
al
ka
rme
s
Be
ds
r
Up
per
K
al
k-
n
re
ic
he
b
ed
s
g-
i
Mid
dl
e
Ka
lk
arme
f
Be
d's
a
Lo
we
r
Ka
lk
-?
re
ich
e b
ed
s
Low
er
Ka
lk
arme
b
ed
s
-
Fa
una
l
Se
qu
en
ce
P
as
in
i
Ka
hl
er
&
Po
ss
ib
le
(i
n
pa
rt
) 1
96
3
Pr
ay
a
ge
-
ag
es
1
96
3
mi
ni
mum
Ps
eu
do
sc
hw
age
ri
na
a
l pi
na
P
er
mian
K
urma
ian
Us
ka
li
ki
an
Qu
as
if
us
ul
in
a,
Ps
eu
do
fu
sul
in
a
Pe
rmian
Qu
as
if
us
ul
in
a
lon
gi
ss
ima
, R
u �o
so
fus
ul
in
a
al �
in
a
Gsh
el
ian
S
ur
en
an
Ka
si
mov
ian
-A
tt
en
ua
te
ll
a
fr
ec
hi
e
tc
. G
sh
el
ian
W
ed
ek
in
de
ll
in
a,
Qu
as
if
us
ul
in
oi
de
s
Mo
sc
ov
ian
-P
se
ud
os
ta
ff
el
la
K
as
imo
vi
an
?G
sh
el
ian
? ?
�u
as
if
us
ul
in
a
te
nu
is
si
ma
"W
ai
de
gg
er
" f
aun
a
Mos
co
vi
an
?Ka
si
mov
ian
L_
Tab
le
21
. A
uem
ig
s
eq
uen
ce
i
n
Carn
ian
Al
ps
, A
us
tr
ian
-i
ta
li
an b
or
de
r
Po
ss
ibl
e
ag
e
maxim
um
?A
ss
el
ian
Gs
he
li
an
?Ka
si
mo
vi
an
Mos
co
vi
an
-...J
0
b::l
o·
�
:::s- � ::
:! c
0 � C)> .... � (II � ....
:;·
(II � ....
�
E· ;:j

MACK ENZIE
BAY
Asselian Stage 71
134°
LEGEND
..,.,.. . . . . . . c:::::>
\ ) I
I
66"
65°
Fig. 1 9. Location of Carboniferous and Permian outcrops, north Yukon Terri tory . Fro m B a m ber & Waterhouse ( 1 97 1 , i1g. 2).

72 Biochronology for the Marine Permian
(1964, p.20), apart from Linoproductus dorotheevi Frederiks, a form typical of late Asselian faunas in Canada.
A small ammonoid faunule has been described from the Hare Fiord Formation of Ellesmere Island by Nassichuk & Spinosa ( 1972), with Somoholites cf. artus, Kargalites, Neopronorites, and Agathiceras indicating an Asselian-Sakmarian age. Accompanying Fusulinacea were assessed as Asselian-Wolfcampian by Wilde (in Nassichuk & Spinosa, 1 972, p.538) with Pseudofusulinella and Schwagerina, apparently like forms from the lower McCloud Limestone of California. As yet undescribed Brachiopoda listed by Waterhouse in Nassichuk & Spinosa ( 1 972) might be as old as Uskalikian, to Tastubian, in age.
The three Asselian substages are well represented by brachiopods in northern Yukon Territory (Fig. 1 9) in the Jungle Creek Formation (Bamber & Waterhouse, 1 97 1) : Table 22. The basal Eka Zone has A ttenuatella and Kochiproductus (Sarytcheva & Waterhouse, 1 972), overlain by a thick sequence of clastics with Orthotichia, Kutorginella, and other species, (Eo Zone), followed by the Attenuatella-Tomiopsis (Eta) Zone of Bamber & Waterhouse ( 1 97 1 ). The latter zone is accompanied by ammonoids Neopronorites and Somoholites. The A ttenuatella-Tomiopsis Zone is judged to be of Kurmaian age, and extends widely to the south in the Rocky Mountains (Waterhouse, 1 97 1 ) into the Telford Formation of Alberta and eastern British Columbia (Logan & McGugan, 1 968).
Further south in Oregon State the Coyotte Butte Formation, 300m thick, yielded a large brachiopod fauna which, although assigned to the Word (mid-Permian) by Cooper ( 1 957), is partly upper Asselian in age (Bamber & Waterhouse, 1 97 1 ). There is strong faunal support for correlation with the Kurmaian AttenuatellaTomiopsis Zone of the Jungle Creek Formation in the Yukon Territory from Dyoros, Tubersulculus, Anidanthus minor Cooper, Kochiproductus transversus Cooper, Yakovlevia, and Attenuatella as 'Crurithyris' sp. Spiriferella draschei (not Toula) of Cooper, and S. parva Cooper, both allied to or identical with S. pseudodraschei Einor, support a Sakmarian age. Wilde ( 1 97 1) assigned Coyotte Butte Fusulinacea to the Decie Ranch-Skinner Ranch faunas of Texas, believed to be of Sakmarian age. Detailed stratigraphic analysis is required to disentangle the mixed faunas.
From south-east Wyoming the Casper Formation has yielded a silicified fauna in limestone (Pederson, 1 954), including Triticites ventricosus (Meek) found in the Neal Ranch Formation of Texas. A Surenan or Uskalikian age is likely, depending on the identity of 'Composita ovata' Mather, which looks somewhat like an Ambocoelid. Ross ( 1 963, p.49) concluded that primitive Schwagerina indicated a pre-Neal Ranch correlation, equivalent to bed 2 of the grey limestone member in the upper Gaptank of the Glass Mountains, Texas, likely to be of Surenan age.
The Earp Formation of south-east Arizona consists of shales, siltstones and limestones close to the Carboniferous-Permian boundary. Brachiopods and other fossils are listed in Gilluly et al. ( 1 954), and Ross ( 1 973) recorded Early Permian Pseudoschwagerina uddeni (Beede & Kniker), and species of Triticites and Schwagerina, overlain by a further Schwagerina-Triticites assemblage that indicated a middle or early late Wolfcampian (presumably Sakmarian) age.
In the Glass Mountains, west Texas (Fig.23, Table 26), the base of the Permian is taken as the Uddenites-bearing shale member of the Upper Gaptank Formation, hitherto often assigned to the Pennsylvanian Period. The problem is carefully discussed by Cooper & Grant ( 1 973, p.366), who point to many Permian elements amongst the brachiopod component, including the first appearance of Kochiproductus. In addition, Bostwick ( 1962) recorded Schwagerina from the shale, and Douglass (in Mudge &�ochelson 1 963, p. 1 20) :ecorded Pseudofusulina. The overlying Neal Ranch Formation of Ross ( � 963) conststs of about 1 6 cyclothems of shale passing up to calcareniteand shell debns, 1 20 to 1 50m thick, with the brachiopod fauna summarized by Ross

Asselian Stage 73
World S tandard Brachiopod North Yukon Archipelago Substage Biozones
Symbol Formation Formation
Kalinovian Canarine Z. Z.oides Gc Trold Fiord
Ufimian - ?-
Elkin .
Irenian Nevolin Lissoahonetes Fl Tahkandit Van Hauen
Filippovian Neoahonetes Fn
I Assistance =Pseudo syrinx Fps
Krasnoufimian Sowerbina Fs
Sarqinian Antiquatonia Fa
Aktastinian J akutoproduatus Ej
Sterlitamakian Tornquistia Et Jungle Belcher Attenuate Z.Z.a Ea Creek Channel
Tastubian Yakov Z.evia Ey
Kurmaian Tomiopsis Eta Attenuate Z. z.a
Uskalikian Orthotiahia Eo
S urenan Koahiproduatus Eka
Attenuate Z. Z.a
Table 2 2 . Permian correlations for Arctic Canada , based principally on
Bamber & Waterhouse ( 19 71) , and S arytcheva & Waterhouse ( 1972) • The
formations of the Arctic Archipelago are complexly interrelated , with
facies transgressing time , so that the Van Hauen Formation for instance ,
is older as well as younger than the typical Assistance Formation at
Blind Fiord (Dr w.w. Nassichuk , pers . comm. ) .

74 Biochronology for the Marine Permian
·-.
o· ........ . -.
.�·
..... ) .....
,/ ---.., .ZZ//.7.,} � BASIN
�MALVINAS IS.
SOUTH AMERICA
SCA L E �j)-"'-400>:!':::-..._":'!li6o lun
- ·· - - -· ·-·-- -- ·--- -- -
Fig. 20. Delimitation of late Palaeozic basins in South America, following R.ocha-Campos (1973,fig. 2, p. 400).

Asselian Stage 75
( 1 963) from various descriptions, chiefly in R.E. King ( 193 1). Further species and genera
. a�e described or listed by Cooper & Grant ( 1969, 1 973). The complete
redescnpttons of Glass Mountain brachiopods of Permian age by Drs G.A. Cooper andR.E. Grant are now in press, and will substantially increase understanding of these faunas. The presence of Orthotichia, Derbyia, Marginiferids, and Hustedia in the Neal Ranch faunas suggest possible correlation with the Eo (Uskalikian) faunas of the Yukon Territory in Canada. Unlike many regions, there is no clearly established three-fold faunal division for the Early Permian in the Glass Mountains but, until the brachiopods are described, it is difficult to ascertain if a substage is missing. On present evidence, the Kurmaian Substage may be absent, and Furnish ( 1 973) also noted that only part (unspecified) of the Asselian appeared to be represented by the Neal Ranch ammonoids. On the nther hand Wilde ( 197 1 ) reported two Fusulinacean zones in the Neal Ranch Formation, which might suggest that the Uskalikian and Kurmaian Substages are present.
In Kansas, as summarized by Mudge & Yochelson ( 1 963), the Permian System commences within the Admire Group, with Pseudofusu/ina entering the Five Point Limestone. Various thin carbonates and shales contain scattered brachiopods, including Crurithyris. Wilde ( 1 97 1) matched the Admire Group with the Gaptank, and the Chase-Council Grove Groups with the Neal Ranch Formation of west Texas (Table 24).
Various other Early Permian faunas have been described in the mid-continental stages of Oklahoma, Kansas and Texas, in the Luta Limestone of the Summer Group or Big Blue 'Series' of Kansas, and in Nebraska (Dunbar & Condra, 1 932). Cooper & Grant ( 1 973) provided evidence that the Virgilian Stage may prove to be Permian.
Bolivia, Peru
The Copacabana Group in the Peruvian-Bolivian basin (Fig.20) has yielded large brachiopod collections described chiefly by D'Orbigny (1 842), Kozlowski ( 1 9 14), and Samtleben ( 1 97 1). Four zones through some 300m to 3,000m of carbonate with minor shale were recognised by Newell et a/. ( 1953), the Silvaseptopora Zone, the Triticites opimus (Dunbar & Newell) Zone, the Pseudoschwagerina uddeni (Beede & Kniker) Zone, followed by barren shales and then in south central Peru by the Parqfusulina Zone of Sakmarian or Baigendzinian age. The earliest zone contains Pseudoschwagerina broggii (Roberts), indicative of a Permian age, with Peruvispira delicatula Chronic, member of a gastropod genus very widespread in the Early Permian faunas of Gondwana. A brief fie1d trip to the area by the writer yielded a Late Carboniferous fauna with Duartia, but this fauna does not seem to have been described as yet. The overlying Triticites Zone is accompanied by Pseudofusulina and Pseudoschwagerina with a few brachiopods, followed by the Pseudoschwagerina uddeni Zone, based on a species also found in the Neal Ranch Formation of the Glass Mountains. Once again a three fold zonation is present, suggesting that all three Asselian substages might be present, but little is known of the association of brachiopods within each of these zones. A few faunas of possible Permian age are recorded in Branson ( 1948) from Brazil, (e.g. Oliviera, 1 936), but some of the better known brachiopod faunas, as described by Derby (1 874) and Mendes ( 1 947) are clearly of Late Carboniferous age, with brachiopod genera Duartia and Brasilioproductus. Early Permian Pseudofusulina and Schwagerina are recorded from Madre de Dios, Chile, by Douglass & Nestell ( 1 974).
Pakistan, India, Himalayas, A.fghanistan
Widespread but small late Asselian (Kurmaian) faunas are known from v�rious segments of Gondwana, as summarized by Waterhouse ( 1 97 1). They share the biValve

-
Perth
Bas
in
Are
as o
f O
utcr
op o
f Pe
rmia
n Se
dim
ents
and
Vol
cani
cs
Perm
ian
Sedi
men
ts a
nd V
olca
nics
In
the
Sub
surf
ace
Sprin
gsur
e Co
rfield
She
lf
Lake
Eyr
e Ba
sin
� Murra
y Ba
sin
� Ma
nning
Ma
cleay
Bas
in
Sydn
ey B
asin
��asmania
Fig.
21.
Di
strib
utio
n of
Perm
ian
strat
a in
Aus
trali
a, fr
om B
rown
eta/
. (19
68, fi
g. 7,
2, p
. 18
1).
....;)
0\
ttl Ci' �
;:s- ti ;:s
c c � '0' .... ..... ;:s- "' � .... ;;· "' � .... �
s· ;:s

Asselian Stage 77
E�ry�e�ma, the gastropo� Peruvispira and brachiopod Tomiopsis, and are associatedwtth tllhte a�d othe� glac1gene rock, having lived in cold waters probably around theedge of glacters, or m areas previously occupied by ice-sheets. In the Indian subcontinent Eurydesma faunas occur below the Speckled or Warchha Sandstone above theTal�hir Conglomerate in the Salt Range, Pakistan (Waagen, 1 89 1 ; Reed, 1 936) Table27 ; m t�e �urydesma b�ds of Bren Spur, Kashmir (Reed, 1932; Waterhouse, 1 970a); innorth Stkktm of the Htmalayas (Sahni & Svrivastava, 1 956) and in central India at Manendragargh (Sahni & Dutt, 1959).
Asselian faunas are found in the lower part of shales and sandstones and a carbonate bank, in all totalling about 1 km in thickness near Wardak (Table 28) south of Kabul, Pakistan (Termier et a/. 1 974). The fossils include Tomiopsis close to T. ovulum Waterhouse, though identified with T. angu/ata (Campbell) by Termier et a/. ( 1974), together with Punctocyrtella nagmagensis (Bion), and the significant bivalves Eurydesma mytiloides Reed, Atomodesma, and Heteropecten. A late Asselian age seems likely, in view of the strong cold-water and Gondwanan affinities.
South Africa, Argentina, Australia The upper Dwyka Tillites of South Africa have yielded the characteristic
Eurydesma bivalve fauna, with other Mollusca including an ammonoid identified as Glaphyrites. This would permit an Asselian age though interpreted as Carboniferous by Martin et a/. ( 1970). In the Sierra de Ia Ventana, Argentina (Fig.20), Harrington ( 1955) described faunas from the Bonete Formation, which appear to be early Permian rather than the mid-Permian age preferred by Amos & Rocha-Campos ( 197 1). Other meagre faunas, including Attenuatel/a, come from Brazil (Fig.20) within the diamictite complex of the Parana Basin (Rocha-Campos, 1 97 1), but these occurrences could be as young as Sakmarian. The fauna described by Reed ( 1927b) from the Andean foothills at Barrealas, Argentina, is considered to be Carboniferous rather than Permian, because it contains the Moscovian genus Levipustula.
In Western Australia (Fig.2 1), late Asselian faunas are found in the upper Lyons Group and Carrandibby Shale of the Carnarvon Basin (Dickins & Thomas, 1959; Konecki et a/., 1 958; Thomas, 1959, 1967; Coleman, 1957 ; Thomas & Dickins, 1954, pp.73-4); Table 29. In eastern Australia the Allandale Formation of the Hunter Valley, north Sydney Basin, and Conjola Formation of the south Sydney Basin also contains the same Eurydesma fauna (Runnegar, 1 969a; Waterhouse 197 ld) : Table 40. Tomiopsis is present, allied to Asselian species from the northern hemisphere (Waterhouse, 1 97 1). Maxwell ( 1964) has described a somewhat richer fauna from the Burnett Formation of the Yarrol Basin, Queensland, of Kurmaian and perhaps greater age (Table 35). The Kurmaian Quam by Group of Tasmania also has a few brachiopods (Banks, 1 96 1 , 1 962; p. 1 98): Table 40.
Earlier Asselian deposits appear to be rare in Gondwana, possibly because of cannabalisation of early Asselian ,deposits by late Asselian glaciation (Burke & Waterhouse, 1 973), and partly because some early Permian deposits may have beenmisdated as Late Carboniferous. Meagre warm-water faunas from the middle LyonsGt-oup of Western Australia (Thomas, 1958) suggest a mid-Asselian Uskalikian age andthe Lochinvar Formation with scattered fossils in the Hunter Valley, New South Wales,may also be Uskalikian from sequential evidence. The barren Joe Joe Formation of theSpringsure Shelf of Queensland is also likely to be Early Permian. It is noteworthy thatEurydesma has been recorded at Stanhope, Hunter Valley, just above the Seahamvarves and tillites, which though assigned to the Carboniferous in Packham et a/. (1 969,p.252), could prove to be of early Asselian age. . . .
No Asselian brachiopods are known in New Zealand. The Crotstlles VolcamcComplex of an ophioloid suite of ultramafics, tholeiites, and volcanic-derived sediment

78 Biochronology for the Marine Permian
over IOOOm thick near Nelson city, contains the bivalve A tomodesma, and a gastropod near the top considered to be of late Asselian age by Waterhouse (1969b).
SAKMARIAN STAGE
Urals
The Sakmar Substage of Ruzencev ( 1952) is based on fine sediment passing up to coarser sediment in a sequence about 800m thick in the Aktyubin region of the Urals (Table 1 2). Fine sandstone and shale are common, with some carbonate and coarser detritus. According to Ruzencev ( 1 952), the Sakmar}an is subdivided into the Tastubian, first used by Rauser-Chernossova ( 1 940) and Sterlitamakian (RauserChernossova, 1937). However, it is here proposed to modify the stage by incorporating the overlying Aktastinian Horizon or Substage of Ruzencev ( 1934, 1 956), exposed near the Aktasty River, south Urals, in 800m of clastic sediment with thin fossiliferous carbonates, called the Burtsev and Irgin beds.
The placement of the Aktastinian Substage in the Sakmarian Stage rather than Artinskian Stage differs from some Russian usage, and is based primarily on the interrelationships between brachiopods. Aktastinian brachiopods from most world faunas are very close to those of Sterlitamakian and Tastubian age, and differ from those of Baigendzinian age (Bamber & Waterhouse, 1 97 1 ). It is considered that the sheer numerical preponderance and wide geographic distribution of brachiopods makes them of prime significance in deciding where stage boundaries should be drawn. After detailed assessment of the very rich faunas of the Glass Mountains, west Texas, Cooper & Grant ( 1973) also drew the Wolfcamp-Leonard boundary, equivalent of the Sakmarian-Baigendzinian boundary, in what I take to be the same position.
Aktastinian ammonoids indicate a different pattern. They include three distinct genera, and share fourteen new genera with the Baigendzinian Stage. The Aktastinian Substage also sees the introduction of 'Parafusulina' s.I., also characteristic of the Baigendzinian faunas, but Rauser-Chernossova ( 1949) initially classed the Aktastinian fusulines with Sterlitamak Fusulinacea. Moreover, the 'classical' Artinskian, of the Arti region, is entirely Baigendzinian (Ruzencev, 1 956). According to Dunbar ( 1940), 'Artinskian' is largely a facies term, and varies in age in different areas, so that its passing from chronologie literature need not cause regret. The following ammonoid horizons were recognised by Ruzencev ( 1 952, 1956). (top) 6. Aktastinian : characterised by entry of three new ammonoid genera,
Aktubinskia, Agathiceras, Neoshumardites. (Presumably the significance of Agathiceras is local, or climatic, for it started in the Moscovian Stage).
5. Sterlitamakian: ammonoids the same as those in upper Tastubian, no newgenera. Five species only persist into Artinskian. Ammonoids do not define this substage clearly.
4. Upper Tastubian : First appearance of many new genera, such as Synartinskia; Medlicottia; Metalegoceras; Uraloceras; Propopanoceras. Only two species persist from underlying lower Tastubian fauna.
Lower Tastubian: or Karamurum 'Series', with Juresanites karakhorum, an advanced member of the genus, and many Asselian survivors. It is hard to see why this is not Asselian on ammonoid evidence. Once again as for the Asselian Stage, the Fusulinacean-not the Ammonoidea-seem to have been decisive in Ruzencev's delineation of boundaries. '
Nautiloidea are also useful in distinguishing the faunal horizons. The Aktastinian Substage includes various Pseudofusulina•, such as P. • karogasensis Raus., P.*paraconcessa Raus., and the first appearance in the Urals of Parafusulina (s.l.).

Sakmarian Stage 79
Sterlitamakian Fusulinacea include Pseudofusulina• verneuili (Moell.), P.• devexa Raus., P. • bashkirica Korzh., and Tastubian faunas include Pseudofusulina• verneuili (Moell.), !P.• moelleri (Schell.) and Rugosofusulina serrata Raus. Schwagerina and allies are missing, but this is apparently only a local phenomenon.
Brachiopod faunas are very extensive and diverse, but have not been comprehensively revised in recent years. To judge from summaries by Miloradovich ( 1949), Stepanov ( 195 1), and Likharev (1966), the Tastubian brachiopods include Schizophoria juresanensis Chernysftev, species of Isogramma and Reticulatia, Rhynchopora nik/tini Chernyshev, Septacamera krotovi (Chernyshev), Cyrolexis superstes (Verneuil) and species of 'Spirifer and Martinia. The Sterlitamakian Substage has a very large fauna with a number of unique species including Rhipidomella ura/ica Chernyshev, Aulosteges uralicus Chernyshev, Derbyia, many Productacea, Camerisma sella (Kutorga), Mattiniopsis orienta/is Chernyshev, and several licharewinid or Spiriferid species.
Various faunas have been recorded from the general Urals region. Mironova (1960) described a small Sakmarian-Artinskian fauna from the Glini beds of PetchoraPai Hoi; and Frederiks (1926) provided a faunal list from sandy argillites of the River Kejim-Terovey, considered to be Aktastinian because of the presence of 'Chonetes' so/ida Krotow, Medlicottia and certain bivalves. Further Sakmarian (Tastubian) brachiopods were described from two horizons in the Ishimbaevo oilfield by Kulikov (1938).
North Siberia, Pamirs
A number of faunas have been described from Novaya Zemlya by Likharev & Einor (1939) and Miloradovich ( 1935). They are judged to be Sakmarian on the basis of their affmities to faunas from the Urals and Yukon Territory in Canada, and lack of Asselian key species, but little data is available on their stratigraphic interrelationships. A late Asselian age is possible for some of the faunas, i.e. from Cape Loushkin, in the Sedov and Barentz 'Series', but these are more likely to be Sakmarian, with species of such genera as Orthotichia, Derbyia, Juresania, Waagenoconcha, Kochiproductus, Linoproductus, Anidanthus, Yakovlevia, Septacamera, Pterospirifer, Phricodothyris and Choristites, the latter showing a resurgence in Sakmarian times, after <the Carboniferous. A Sakmarian age is supported by specific comparisons amongst species of the genus Spiriferella for several of the localities.
In central and west Taimyr Peninsula (Table 1 5), the Birrang Horizon was correlated with the Sakmarian and Artinskian Stages by Ustritsky & Chernyak ( 1967) and with the early Lower Permian by Menner et al. ( 1970). The upper part includes Jakutoproductus, Rhynchopora, Attenuatella, Taimyrella and Tomiopsis (Ustritsky & Chernyak, 1963), suggestive of a Sterlitamakian or even Kurmaian age. This horizon is perhaps preceded by a fauna listed as Evenk and Upper Turuzov in beds 850- 1 100m thick from east Taimyr by Ustritsky & Chernyak ( 1963), containing such species as . Echinalosia delicatula, Rugivestis species, Yakovlevia species and Rhynchopora nikitini which suggest a Tastubian age, by comparison with faunas of the Yukon Territory, Canada, discussed below. A farge Sakmarian fauna was described from west Taimyr by Einor ( I 939, 1946).
In the thick terrigenous sequences of the Verchoyan Mountains in north-east Siberia the lower part of the Tompin Series yields faunas of Sakmarian age. In the Haraulakh area near the mouth of the Lena River, the Verchoyan Suite, 650m to 2,000m thick, contains Sakmarian brachiopods such as Yakovlevia mammatiformis (Fred.), Waagenoconcha, Jakutoproductus verchoyanicus (Fred.) and Spiriferella saranae (Verneuil) with the ammonoids Paragastriocerasjossae subtrapezoidalis Max. & Chern., Uraloceras ex. gr. belgushkense Ruzh. and other species (Likharev, 1 966).

80 Biochronology for the Marine Permian
Menner et a/. ( 1970) reported Neoglaphyrites which would suggest an Asselian age in part, with Agathiceras uralicum (Karpinsky). In a closely divided sequence of southeast Verchoyan (Table 14), the Kigiltass Suite 600- l lOOm thick includes Rhynchopora nikitini Chernyshev with Derbyia, Jakutoproductus and linoproductinids, followed by thin argillites of the Echi horizon, l OOm thick, containing numerous Jakutoproductus and other distinctive brachiopods with the ammonoids Paragastrioceras and Neoshumardites triceps hyperboreus Ruzh. These faunas are clearly Sakmarian, by superposition and general faunal affinities. In the Kolyma River and Omolon Plateau, the Irbichan, Munugudjak, and Yasachnin Horizons (Table 1 5), are likely to be Sakmarian, though referred to the Artinskian by Likharev ( 1 966). The lrbichan Horizon appears to be Tastubian, or at least correlative with the Yakovlevia Zone of the Yukon (Bamber & Waterhouse, 1 97 1). It includes Yakovlevia mammatiformis (Frederiks), Cancrinella cancriniformis (Chernyshev) and species of Jakutoproductus, Anidanthus and Tomiopsis. The overlying Yasachnin Horizon has a few different species with Martinia and Spiriferella, in some l OOm of sediment. Although no direct connection with the Yukon or Urals sequence is apparent, its stratigraphic position would suggest a Sterlitamakian age. The overlying Munugudjak Horizon, 35-60m thick, contains Tornquistia, Jakutoproductus and A nidanthus to suggest correlation with the higher Yukon faunas, of approximately Aktastinian age. Evidence for correlation is not strong, but is consistent with stratigraphic position, and there is no contrary evidence. Faunas have been described chiefly by Zavodowsky ( 1968, 1970).
From the Ungadu Suite, 800- 1300m thick, in the Chiron area of west Baikal, Kulikov ( 1959) listed a small brachiopod fauna of Sakmarian age, with species allied to Verchoyan forms, such as 'Chonetes' paraconvexa Maslennikov and Cyrtospirifer khauraulakhensis Frederiks. The beds overlie the Chiron Suite.
In the northern Pamirs, especially Darvas, the upper part of the Lower Permian is referred to the Darvasian Stage with four subdivisions (Table 1 7). The Zigar Horizon at the base has fusulines Pseudofusulina kraffti (Schell.) and !P.* vulgaris (Schell.) and Parqfusulina ferganica (M.Makl.) and ammonoids Propinacoceras, Popanoceras (supposedly limited to the Artinskian), and Metalegoceras. A number of Russian experts have placed the Zigar faunas in the Aktastinian Substage and the slightly older Karachatirian faunas in the Sakmarian (i.e. lower Sakmarian) Stage. However, the Zigar Horizon is here regarded as early Sakmarian on the basis of sequence and fossils present. Parafusulina enters the Lenox Hills Formation of west Texas at a comparable early Sakmarian age, as shown by Cooper & Grant ( 1973), assuming identifications are correct. The presence of a few persistent ammonoids through thick sections with several brachiopods or fusuline zones strongly suggests that the ammonoids were, as usual, long ranging, and slightly different in age from the Urals, of a different palaeogeographic realm. It seems most unlikely that the Zigar, with overlying faunas, are all Aktastinian in age.
The overlying Chelamchin Suite of the same region has Pseudofusulina kraffti and Misellina. A large brachiopod fauna is listed by Likharev ( 1966) for the succeeding Safetdaron unit with numerous Fusulinacea, Nankinella, Pseudofusulina ('Leeina') exigua (Schell. & Dyhr.), Pseudofusulina kraffti, Cht�senella, and Parafusulina afT. japonica Gumb. Kalmikova ( 1967, p. 142) described a large fauna from Tangi-Gor, Darvas, in the Safetdaron Suite, with Schwagerina crassitectoria Dunbar & Skinner (found also in the Decie Range Member of west Texas), Schwagerina* fusiformis (Schell.) and !Pseudofusulina* vulgaris (Schell.). The brachiopods include numerous overtoniids, Camerisma sella (Kutorga), many Choristites, and Martinia, and are regarded as Aktastinian, with warm waters suggested by the presence of Parakeyserlingina. On the other hand Schwagerina crassitectoria is more likely to be Sterlitamakian in the Glass Mountains and Hueco Mountains of Texas as discussed below. Ammonoids in the overlying Gundarin Horizon include Medlicottia aff. artiensis

Substage/ or
Stage
Baisalian Subst.
Urushtenian Subst.
?
Punjabian Stage
?Kazanian
Stage
Kungurian
Stage
Baigendzinian Stage
Aktastinian
Substage
S ter li tamakian
Substage
Cl.l bO Ill .u
en
'"' •.-l
ij P-<
Cl.l bO
' Ill .u
en
.a Ill bO '"'
£
Cl.l bO Ill .u
en
� Ill
•.-l Cl) Ill
� Ill
�
Sakmarian Stage 81
s w Pamirs
Upper - - - -?- - - - - -
g Middle Codonofusie Ua� ::::>�
Reiche Una� P':)E-<
�� PaZaeofusulina� ::<:en
Lower CoZanieUa < E-<
Lower Codonofusie Z Za Tlo-i ,.]M:tina._
Neoschwagerina � E-< margaritae� H ::::> Yabeina archaica en
Neoschwagerina � (.!) schuberti
Lower
Neoschwagerina simplex
Kubergandin
CanceUina Suite
MiseUina
Upper MiseUina Darvasites
z H ? ;:3[1:1 Middle HE-t
PseudofusuZina ,_.,H � p ...:IU) Parafusulina? H Lower N H ::<:
Bazardarin
Suite
Table 23. Permian succession in Fergana, from Grunt & Dim etriev (1973).

82 Biochrono.logy for the Marine Permian
timorensis Han. and Propinacoceras bornemani Toum., here judged to be of Baigendzinian age.
Grunt & Dimetriev ( 1973) described the early Permian Bazardarin Suite in thePamirs (Table 23) with a relatively rich ammonoid faunule, including Metapronorites timorites Haniel, Synartinskia, Agathiceras, Tabantalites, Marathonites and Waagenina dieneri Smith, and a mod�st brachiopod con;tponent including Derbyia grandis Waagen, Taeniothaeru_s rustzcus. Grunt, Cancrznel�a,. and Punctocyrtella spinosa Plodowski. These brachtopod spectes are strongly remtmscent of Gondwanan faunas of early Sakmarian age, and differ from those of the nearly Safetdaron fauna,believed to be slightly younger. Grunt & Dimetriev ( 1973) assigned a Sakmarian-lower Aktastinian age (Table 23).
The overlying Kizildjilgin Horizon in Grunt & Dimetriev (1973) is of early Darvasian age, with Pseudofusulina and Parafusulina, overlain by beds with Mise/Una andDarvasites. The lower part of the formation includes brachiopods identified as Globiella rossiae Sestini, supposed Lamnimargus himalayensis (Diener), Martinia, andCamerisma murgabica Grunt and a species of Spiriferella, unnamed, but close to Spiriferella salteri Chernyshev from the Sakmarian Stage and S. waageni (Chernyshev) from the Artinskian facies of the Urals. Compound rugose corals and Artinskian ammonoids such as Agathiceras mediterraneus Toum., Gaetanoceras martini (Han.), G. timorensis (Han.), Perrinites and Marathonites also occur. Several of these ammonoids persist from the underlying faunule into the middle Kizildjilgin faunas, with Monodiexodina. The upper Kizildjilgin faunas, in 20- 1 20m of sediment, include Pseudofusulina,Parafusulina and Misellina. The faunas are here judged to be Aktastinian from brachiopods, but perhaps they are BaigendziDian.
Europe
In the Carnian Alps of Austria, Italy and Yugoslavia, the Grenzland band of the middle Rattendorfer beds may be early Sakmarian in age, with a few brachiopods including !§_ogramma paotechowensis Grabau, many compound rugose corals, indicative of warm waters, and Pseudoschwagerina extensa Kahler & Kahler, Pseudoschwagerina turbida Kahler & Kahler, and notably Rugosofusulina alpina (Schell.). The upper 'Pseudoschwagerina limestone', totalling with the Grenzland beds about 65m in thickness has Pseudoschwagerina pulchra Kahler & Kahler, and Schwagerina• nitida Kahler & Kahler, with a number of compound rugose corals. The faunas are here regarded as Sakm�ian (Table 24).
The overlying Trogkofel beds are also probably Sakmarian, possessing the ammonoid Medlicottia and a rich brachiopod faunule including Enteletes, Meekella, Scacchinella, over_toniids, Camerisma sella (Kut.), Licharewia carnicus (Schell.) and Brachythyrina, described by Schellwien ( 1 900) and Heritsch ( 1938), with close general similarities to the Safetdaron fauna of the Pamirs and Sakmarian faunas of the Urals. Fusulinacea suggest correlation with the upper Sakamotozawan Stage of Japan. The age may be as young as Aktastinian, although Tastubian affinities may also be noted. Kahler & Prey ( 1963, p.44) placed the fauna in the middle Permian. Ramovs (1963) described Productida including Karavankina and Scacchinella from the Trogkofel beds in Yugoslavia. According to the correlation proposed herein the Tarviser Breccia in the Carnian Alps lies at the same horizon as great breccias in the Hueco Mountains in T�xa� (Cooper & Grant, 1 973), suggesting the possibility of a sharp fall in sea-level at thts ttme, related to glacio-eustatic fall of sea-level, or widespread orogenic spasm.
Armenia, Iran Afghanistan, Karakorum, Chitral
The D�valin �uit� of Armenia (Table 45) contains Nankinella and may be as old as Sakmanan, wtth Schwagerina• afT. fusiformis (Schell.); !Pseudo/usu/ina• ex gr.

Sakmarian Stage 83
stage Substage Group Unit
Scythian Dienerian ?Werfer- (Servino) ? Ftm. ?
Dorashamian Griesbachian ---
-
Ogbinan
Vedian Bellerophon
Baisalian Limestone
Djulfian Urushtenian 200m ' Punjab ian
Kazan ian
Baigendzinian ?Grodener 30-40m
Aktastinian
Sakmarian S ter li tamakian Trogkofel 200-400m
Upper Pseudoschwagerina limestone
Tastubian Rattendorfer Grenzland banke 70m
Kurma1an Lower Pseudosc�erina limestone
Asselian Uskalikian Auernig Upper kalkarme beds 200m Upper kalkreiche beds
Surenan Middle kalkarme beds
ate Carboniferous -?Middle kalkarme btds Lower kalkreiche eds
Period Lower ka lkarme beds
Table 24 . Correlation of Permian sequence in Carnic Alps , Austria, assessed
from Kahler & Prey (1963) . The Auernig b�ds are retained partly as in
the German fo� the sake of brevity .

84 Biochronology for the Marine Permian
l'ulgaris (Schell.) and Triticites (Arakelyan e� a!. 1 964, p. 1 24). Nanki�ell� al�o occurs inthe Aktastinian faunas of the Safetdaron Sutte of Darvas and the Chthsta Ltmestone of China.
Douglas ( 1936, 1 950) listed and partly described a small brachiopod faunule from south-west Iran, indicative of an Asselian-Sakmarian age, including Schizophoria juresanensis Chernyshev of Tastub�an age in the yrals, and Canc;inetla cancriniformis (Chernyshev). The fauna is overlam by beds wtth Pseudofusulma, !Pseudofusulina*
vulgaris, Triticites simplex (Schell.) and Rugosofusulina alpina of Sakmarian age. From north Iran Sestini ( 1 966) described the Geirud Member D fauna in cherty
limestone, shown by Waterhouse ( 1970a) to be probably Sterlitamakian on the basis of brachiopods (Table 3 1). The fauna, not fully collected according to Dr R. Assereto, University of Milan (pers. comm., 1 97 1), includes such Sakmarian, and especially Sterlitamakian keys as buxtoniid Reedoconcha iranicus Sestini, Stepanoviella,widespread in beds of this age throughout Gondwana (Waterhouse, 1 970a), Punctocyrtella, found also in the Pamirs, and Martiniopsis orienta/is Chernyshev, also found in the Urals.
From the Haushi Formation over 200m thick at Haushi and Wadi Lusaba, Saudi Arabia, Hudson and Sudbury ( 1 959) described another apparently Sterlitamakian fauna with Licharewia and Juresania, correlative with the Geirud D fauna (Waterhouse 1970a). The ammonoid Metalegoceras is found just above. An overlying fauna in the Lusaba Limestone 33m thick is more diverse with Juresania and marginiferids, and may, from superposition and its warm water affinities, be as young as Aktastinian. Parafusulina beds lie above.
To judge from preliminary accounts by Siehl ( 1 967) and Karanatev & Leven ( 1973), there are extensive fossiliferous sequences of Permian carbonates in Afghanistan, rich in Fusulinacea and Brachiopoda. Partially described brachiopod faunules from central west Afghanistan include spiriferids such as Orulgania, Permospirifer and Punctocyrtella (Legrand-Blain, 1 968; Plodowski, 1 970), like species from the Haushi and Geirud D faunas. They may be Sterlitamakian, although the report of Haydenella suggests a younger, Aktastinian age, by comparison with the Lusaba fauna, with its marginiferids and Chonetenellids. Accorqing to Termier .et a/. (1974, pp.39,40), Sakmarian faunas near W ardak south of Kabul, Pakistan, (Table 35) include a lower fauna with Stepanoviella umariensis (Reed), also found i� the Umaria fauna of the Indian Peninsula, Tomiopsis, Martiniopsis and other forms, and an upper fauna with Neochonetes, Taeniothaerus (or Reedoconcha) permixtus Reed, Punctocyrtella spinosa Plodowski, and Tomiopsis related to Kashmir species from the Agglomeratic Slate. Both faunas may be of Sterlitamakian age, in view of the presence of Tomiopsis, or the lower faunas may prove to be as old as Tastubian.
The Aktastinian Substage may be represented in Afghanistan by black carbonates with Fusulinacea including Parafusulina ferganica Mikl.-Makl., and Pseudofusulina* procera Leven, with brachiopod species of Karavankina, Sergospirifer and Notothyris, and the gastropod Bellerophon. Species of the latter two genera were ascribed, probably mistakenly, to forms named from the Middle and Upper Productus Limestones of the Salt Range in Pakistan.
From the Shaksgam Valley, Karakorum Range, Renz ( 1 940b) and Sestini ( 1965d) recorded a few so-called upper Uralian species, too few and nondescript to date accurately, but likely to be Lower Permian, and probably Sakmarian in age. The species show little in common with the faunas from Iran, Afghanistan, or Kashmir, but orthotetids, Krotovia and marginiferids suggest some approach to the Lusaba brachiopods of Arabia, and so are perhaps of Aktastinian age.
Early Permian faunas in quartzites and limestones from Chitral Pakistan espe�ially near Baroghil Ailak, contain SchWagerinids, supposedly Pseudoschwag�rina prmceps (Ehrenberg?) and Schwagerina• fusiformis Krot. pointing to an Asselian age

Sakmarian Stage 85
(Reed, 1 925)., They probably require revision and were not figured. Accompanying Kochiproductus and Crurithyris also suggest an Asselian or mid Sakmarian age, and numerous specimens of Spiriferella look like Spiriferella saranae (Verneuil) or S. kolmaensis Zav. from Sakmarian faunas of the Yukon, Canada and Siberia. Perhaps this younger age is supported by the presence of so-called Parafusulina (=Praeparafusulina) cf lutugini (Schell.), and shells allied to Stepanoviella, as if the faunas were Sterlitamakian and Aktastinian. The faunas were collected mostly from loose blocks with poor stratigraphic control and could have come from two or more substages.
China, Japan
In China, the Chihsia Limestone is a black thick bedded limestone 1 20-300m thick, shaly near the base, lymg over shale with Propopanoceras (Table 1 8). Four Fusulinid zones were recognised by Chan ( 1 963) : Baigendzinian 7-8 Schwagerina (=Pseudofusulina) chihsianensis Lee-Rugosofusulina
multiseptata (Schell.) Sakmarian 6 Nankinella orbicularia Lee-Mu.fushanella mufushanensis Sakmarian 5 Misellina claudiae (Deprat) Sakmarian 4 Pseudofusulina* tschernyschewi (Schell.)
These zones probably embrace the Sakmarian and Baigendzinian Stages, although fusuline correlations are not entirely clear, for S. chihsianensis at the top of the Chihsia beds is reported also from the mid-Karachatirian faunas, regarded as Asselian in age. However Misellina claudiae is widespread in Artinskian faunas and Nankinella enters the Safetdaron fauna of the Pamirs, regarded as Aktastinian. The distribution of
· brachiopods within the Chihsia Limestone is poorly known, and has not been studied inrelationship to the Fusulinacea (Grabau, 1 93 1 ; Ching, 1 963 ; Chao, 1 927, p. 1 78 ;Huang, 1 933, p.96). Orthotichia suggests a general Sakmarian age, and correlation withthe Safetdaron and Trogkofel faunas is supported by Phricodothyris waageni and Martinia.
In Japan the upper Sakamotozawan Stage of Toriyama ( 1 967), with the !Pseudofusulina* vulgaris Zone (Table 20,p. I 2 1), overlain by the Pseudofusulina ambigua Deprat Zone = P. kra.ffti locally, is approximately Sakmarian in age, though !Pseudofusulina* vulgaris also characterises the middle Maping Limestone. Kanmera ( 1963) matched the two zones with a lower Misellina Zone, followed by an upper Misellina claudiae Zone. Ueda ( 1963) listed some brachiopods from the Nishikori Formation with Fusulinacea that appear to belong to the P. ambigua Zone.
Spitsbergen, Canada, A Iaska
From Spitsbergen, Bunsowland (inner lsfjorden on Fig. 1 9), the upper Wordiekammen Limestone or Tyrellfjellet Member of the Nordenskioldbreen Formation in the Pseudoschwagerina Zone is correlated with the Asselian-lower Sakmarian by Cuthill &Challinor ( 1 965) and with the Asselian-Orenburgian by Czarniecki ( 1 969, p.256). Perhaps the fauna is Uskalikian in age, or more likely Tastubian (Table 24), in view of the absence of Attenuatella and Tomiopsis. The upper 'Gypsiferous Series' has similar species (Gobbett, 1 964).
Permian brachiopods have also been identified by Gobbett ( 1964) from the Cora Limestone in Bj0rn0ya, allegedly below the Pseudoschwagerina beds. Cuthill & Challinor ( 1 965) assigned this limestone to the Sakmarian and Upper Wolfcamp Monodiexodina Zone which overlaps with Parafusulina and showed it to be younger than the Nordenskioldbreen Formation. The brachiopod genus Tityrophoria is found in the fauna, suggesting an Aktastinian age by correlation with faunas in the Yukon Territory, Canada (Bamber & Waterhouse, 1 97 1). Parafusulina has been reported by Forbes et

Bj91
rn91y
a
Spir
ife
r Lm
st
.
Cora
Lms
t
Fusu
lina
Lm
s t
,
Bil
lefj
orde
n
Hor
nsu
nd
For
bes
et
aZ
. �
1958
Cu
thil
l an
d Ch
all
inor
Brac
hio
pod
Upp
er
Hov
tind
en
Lim
es
ton
e M
idd
le
Kap
p M
emb
er
Brac
hio
pod
Sta
ros
tin
Sv
ensk
eegg
a Ch
erts
Fo
rma
tion
M
embe
r
Low
er
v;r
inge
n
Lim
est
one
Mem
ber
Upp
er G
yp
sife
rou
s G
ipsh
uken
Fo
rma
tion
Se
rie
s ?C
ancr
ine
lla
Ims
1i!!
!.e!_t£
n!. ]!
__
_ :
L.!,m!,
S!,O,!
!e_B:
l::l
Wor
die
-Ty
re
llfj
ell
et
0 .....
Mem
ber
�
k.
ammen
U
pper
e
--
--
--
--
Lim
e-Br
uce
byen
Bed
s 0
Mid
r:r.
. -
--
--
--
-
sto
ne
l::l Low
er _
__
_ �
--
--
--
--
� 1-1 Ca
dell
fje
lle
t ,0
't:l
T
resk
ell
oden
M
emb
er
.-4 0 •..-!
,.!G
bed
s B
lack
Cr
ag
Bla
ck C
rag
(I) l::l � 't:l 1-1
Min
kin
fje
llet
0 z
M
emb
er
Tab
1e
2
5.
P
ermi
an
ro
ck
s
of
S
pi
ts
ber
ge
n
and
B
jp
rnp
ya
.
Sub
stag
e
?Ka
lin
ovia
n
Iren
ian
(N
evo
lin)
Fi
lip
pov
ian
Akta
stin
ian
i
S te
r li
tam
akia
n
?Ta
stub
ian
?Usk
ali
kia
n
?Sur
enan
Carb
onif
erou
s
00
0\
t:x:l
o· g. � ::s
c c � '0> .... :;. � � ::s.
::s
� � � s·
::s

Sakmarian Stage 87
a/. ( 1958) and Ross ( 1965), though doubted by Cuthill & Challinor ( 1965) and Czarniecki (1969). The Canctinella Limestone, above the Treskelodden beds of west Spitsbergen, has a small fauna of possible Sterlitamakian, or Kurmaian age, with Paeckelmanella.
Four Sakmarian brachiopod zones have been recognised in the Yukon Territory of Canada by Bamber & Waterhouse ( 1 97 1). The Yakovlevia (By) Zone lies at the base (Table 22), correlated with the Tastubian Stage, and accompanied by rare Metalegoceras. The overlying Attenuatella (Ea) and Tornquistia (Et) zones, better considered as subzones, for they are clearly of two communities in different facies, are judged to be Sterlitamakian in age. Ammonoids include Eoasianites, Tabantalites bifurcatus Ruz. afT. trapezoidalis Max., Properrinites, Somoholites cf. belvensis (Haniel), Prothalassoceras, Medlicottia and Uraloceras. Although these were assessed as Tastubian in age by Nassichuk ( 1 97 1 ), Furnish ( 1 973, p.53 1) preferred a Sterlitamakian age as supported by the brachiopods (Bamber & Waterhouse, 197 1 ). The Jakutoproductus (Ej) Zone has various species related to Siberian species, especially Jakutoproductus as in the Echi faunas of Verchoyan, Anidanthus, Yakovlevia, and notably Martinia, so common in Aktastinian faunas. A laskanella yukonensis (Skinner & Wilde) from the Jakutoproductus Zone has been interpreted as indicating correlation with the Lenox Hills Formation by Ross ( 1967a), and Tastubian Substage by Mamet & Ross (in Bamber & Waterhouse, 1 97 1), but it is here considered that brachiopods and stratigraphic position above Sterlitamakian brachiopods and ammonoids require an Aktastinian correlation. Brachiopods of the Jakutoproductus Zone are also found in Bjome Peninsula, southwest Ellesmere Island, Canadian Arctic Archipelago, in beds perhaps erroneously referred to the Assistance Formation by Thorsteinsson (1 974). The brachiopods are found with ammonoids Neoshumardites cf sakmarae (Ruzh.), Uraloceras, and Paragastrioceras afT. jossae (Vern.) which were regarded as late Sakmarian (Sterlitamakian) or Aktastinian, by Nassichuk et a/. ( 1965). Accompanying Fusulinacea support an Artinskian age, and an Aktastinian age is likely. Lengthy faunal lists, compiled chiefly by G.H. Girty, suggest that Sakmarian faunas are widespread in Alaska, for example in the Chitina Valley of south-east Alaska (Moffitt, 1 938a) and in the upper Copper River District and east Alaska Range (e.g. �offitt, 1 938b).
Fusulinacean and brachiopod faunules from the upper Telford Formation, and from the Ross Creek Formation of South-east Alberta near Crowsnest have been discussed by Logan & McGugan ( 1968), and assigned a general Sakmarian age.
A briefly described fauna from the Buttle Lake Formation of Vancouver Island (Yole, 1963) is probably Sakmarian, judged from brachiopods Horridonia, Spiriferella and Tomiopsis like Tastubian forms of the Yakovlevia Zone in the Yukon Territory, Canada. Schwagerina and Pseudofusulinella are present, the latter known from high Wolfcamp (i.e. Lenox Hills) and Leonard according to Monger & Ross ( 1 97 1 ).
United States
In south-east Arizona, the upper Earp Formation has Sakmarian Fusulinacea, as discussed by Ross ( 1973), and higher beds contain an ammonoid Perrinites or Properrinites indicating a Wolfcamp or Leonard correlation. The overlying Colina Formation contains gastropods that suggest a late Wolfcampian age according to Knight in Gilluly et a/. ( 1954), but brachiopods were interpreted as showing a mixture of affinities. It seems likely that at least the lower fauna may be approximately equivalent to the Skinner Ranch Formation of the Glass Mountains, and roughly Aktastinian or Sterlitamakian in age or both. Perhaps the upper beds are as young as the Cathedral Mountain Formation of the Glass Mountains (i.e. Baigendzinian), but with the faunas not described, and revision of the Glass Mountains brachiopods pending, it is difficult to be sure.

Gu
ad
alu
pe
Sie
rr
a
Sta
ge
S
ubs
tag
e D
ela
wa
re
Dia
blo
G
�ass
M
oun
ta
i.ns
Dj
ulf
ian
Ur
ush
ten
ian
? L
amar
McC
�II
IDS .-!
1:
Rad
er
.-! 0
Pun
jab ian
c
apit
an
Gl >.
C
ap
itan
P
ine
ry
Ill ij
Gl u
§'
}leg
ler
.-!
Goa
t
Man
zan
ita
�
6 Goa
t
Vid
rio
Ill
Ka
zan
ian
'g
See
p
Sou
thw
el
ls
�
>.Se
ep
?S
osn
ov
ian
g G
et
away
fia
? Ap
pe
� Ra
ncn
Q
Ka
lin
ov
ian
Sa
nd
st
on
e T
on
gu
e S
s T
ong
ue
Ill
w�J.
J.�s
1<an
cn
0 3: C
hin
a
Nev
oli
n
!Brush
y
Ca
ny
on
T
ank
K
ung
urian
F
il
ipp
ovia
n
Cu
to
ff
Cu
to
ff
Ro
ad
Can
yon
'0
Vic
to
rio
V
ict
or
io
Ca
th
ed
ra
l I<
Peak
"C
ut
of
f"
Pe
ak
Mo
unt
ain
Ill
'9a
ige
nd
zin
ian
6 B
on
e Gl
Bo
ne
Bo
ne
..:I
Spr
ing
Sp
rin
g
Spr
ing
W
ed
in
Akt
ast
inian
H
ue
co
Low
er
S
ul
liv
an
Mo
unt
ain
s M
as
siv
e �
Pe
ak
Gl
.<::
1: 0
Du
go
ut
Mt
n
�
��
Sak
ma
rian
S
te
rli
ta
-A
lacr
an
B
on
e P
op
lar
T
ank
(/)
D
ecie
Ra
nch
S.
cras
site
ctor
ia
mak
ian
g.
Mo
unt
ain
Sp
rin
g
0 T
ast
ubia
n
�
� C
er
ro
Alt
o
Hu
e co
L
eno
x
0 0
Hu
e co
11-1
0
.-!
8 C
any
on
G
ro
up
Hi
ll
s
0 3: :I:
Po
wwo
w C
glm
P
owwow
?Kur
ma
ian
A
ss
el
ian
U
ska
lik
ian
S
ure
nan
N
ea
l R
anch
Udde
nite
s Sh
ale
Mag
da
len
a
( Or
do
vi
ci
an)
Gap
tan
k
�
----
� �
��-
� ,�
�
oo�
�ov
��
ha
ao
M
nn
r
nn
n�
r
£ r.
ra
nt
11
97
3.
fi
a_
2)
a w
it
h
t�
nt
at
iv
e
Tab
le
2
6.
Ea
rl
y
wo
rl
d
co
rr
el
at
io
ns
. X
n
my v
ie
w,
the
Le
on
ar
d
Gu
ad
al
upe
b
oun
da
ry
s
ho
ul
d
li
e
� t
he
R
oa
d
ca
ny
on
F
or
mat
io
n,
as
i
n
Ki
ng
19
3�.
00
00
t.:l:! (5• (') �
...
<::>
:::s
<::> c � � ...
.... �
(I) � ...
;;· (I) � ...
�
�S·
:::s

0 50
NORTHWESTERN
SHELF
N
t 100
I
Kilometres
Sakmarian Stage 89
MEXICO
Fig. 22. Palaeogeographic setting of the Permian in west Texas, according to Cooper & Grant (1973a, fig. 27, p. 97). Reproduced with permission of authors.

90 Biochronology for the Marine Permian
West Texas provides superb faunal sequences (Table 26). The Lenox Hills Formation of the Glass Mountains (Fig.22) includes Pseudoschwagerina and Monodiexodina with a number of Ammonoidea, such as Metalegoceras, Properrinites, Eoasianites and Artinskia and Brachiopoda listed in part by Ross ( 1963). Ross ( 1963, p.43) matchedthe Leno� Hills Formation with the Asselian Stage, on the basis of Pseudoschwagerina which persists to the top of the formation. Yet Parafusulina is also present in the upper Lenox Hills beds. As here interpreted, the ranges of both these genera overlapped in faunas of Sakmarian age, though not in the Urals.
The Lenox Hills, Skinner Ranch, Hess and Bone Spring brachiopods are currently being restudied by Cooper & Grant (in press) and it is not possible to provide a full analysis until the survey is published. A few initial studies in the Sierra Diablo and Glass Mountains by Stehli ( 1954), Cooper & Grant ( 1969) and others provided no wideranging correlations. Cooper & Grant ( 1973) made it clear that the Skinner Ranch brachiopods, previously treated as Leonard, were close to the Wolfcamp forms in general appearance. Cooper & Grant ( 1964, 1 973) have shown that the Hess facies of the Glass Mountains matches the Decie Ranch beds and in part the Lenox Hills Formation, as well as the lower Bone Spring Formation of the Sierra Diablo (Fig.22). Brachiopods from this latter formation were described by Stehli ( 1954), and assigned to the Leonard, in conformity with interpretations based on Fusulinacea, Stehli making no attempt to assess the age independently. But brachiopods clearly suggest a Wolfcamp age (Cooper & Grant, 1 973). To the writer, the presence of Attenuatella in the Bone Spring fauna suggests the possibility of a Sterlitamakian age, and the remainder of the brachiopods described would permit this correlation. A Sterlitamakian age for the Bone Spring basal beds would agree with the ammonoid evidence, which suggests correlation between the Lenox Hills Formation and Tornquistia Zone of Canada. It would also agree fairly well with the position of the Safetdaron fauna in Darvas, mid Asia, where a rich brachiopod fauna regarded by the writer as most likely to be Aktastinian but possibly slightly older is accompanied by Schwagerina crassitectoria of the lower Skinner Ranch Formation. Such an interpretation would appear to agree with the general concept implied by Cooper & Grant ( 1 973) that the faunal succession is continuous. By contrast, Ruzencev ( 1 952), Ross ( 1 963), and Glenister & Furnish ( 1 96 1) indicated a lengthy unconformity above the Lenox Hills Formation, although Furnish ( 1973) later modified his view by referring the Skinner Ranch ammonoids to the Aktastinian, as in Bamber & Waterhouse ( 1 97 1). Further brachiopods have been reported in the Cerro Alto Limestone and the Hueco Canyon Formation of the Delaware Basin, New Mexico, by Williams ( 1 963, p. 1 7). On the basis of Fusulinacea, Wilde ( 1 97 1 ) correlated the lower Hueco, consisting of the Powwow Conglomerate, and Hueco Canyon and Cerro Alto Formations, with the Neal Ranch Formation of the Glass Mountains, whereas Williams ( 1 963) correlated all of the Hueco Group, except the upper part of the Alacran Formation, with the Lenox Hills Formation. The latter correlation is supported by brachiopods, according to Cooper & Grant ( 1 973, p.368). The difficulties may be underlined by noting the occurrence in the Hueco Canyon Formation of the ammonoid Eoasianites, generally regarded as typical of Asselian and earlier faunas, with genera so typical of the Sakmarian Stage in the Urals, such as Metalegocer-as and Synartinskia.
Brachiopods from the light grey limestone of the Alacran Mountain Formation were listed by Williams ( 1 963), and correlated with the Schwagerina crassitectoria bed of the Decie Ranch Member at the base of the Skinner Ranch Formation in the Glass Mountains.
Wilde ( 197 1 ) correlated the Summer Group of Kansas with the Lenox Hills and lower Skinner Ranch formations of west Texas on the basis of Fusulinacea, and m�tched the overlying Stone Coral Formation and Nippewalla Group with the upper Skmner Ranch beds. The Schwagerina crassitectoria horizon was considered to be missing here, but was observed at the base of the Clyde Formation of north-central Texas.

Sakmarian Stage 91
McCloud Ls Glass Mountains North-Central
Kansas "Series n·
Fusulinid West Texas Texas
Zones
Skinner Ranch Clyde Nippewalla Gr Leonardian H Formation (restricted) Formation Stone Corral
Formation
SchuJagerina S.crassitectoria crassi tectoria
?
Decie Ranch Member Belle Plains Wolf-G Formation Stml!Der
Skitlner Ranch Forma tiro Group
camp-?
Lenox Hills Admiral ian F Formation Formation
E Basal Lenox Hills
Congl.
D Neal Ranch Moran & Putnam Chase-Formation Formation Council Grove
Groups
B-C King's Bed 3 ? Salt Creek Bend Eskridge Shale Shale
King's Bed 2 Pueblo Admire Formation Group
A of
"Gray Limestone"
Table 27. Correlation of McCloud fusulinid zones of California with other lower
Permian sequences of United States, following Wilde (1971, fig. 8).

92 Biochronology for the Marine Permian
In the same region the Belle Plains Formation and Admiral Formation were referred to the Lenox Hills and lower Skinner Ranch beds, as shown in Table 27. Ammonoids as summarized by Furnish ( 1 973) support these correlations well, with Properrinites boesi (Plummer & Scott) and P. denhami (Miller & Furnish) from the Admiral Formation, considered to be at a Tastubian phase of evolution. From the Clyde Formation, species of Medlicottia, Metalegoceras, Popanoceras and Metaperrinites suggest an Aktastinian age. None of the species are identical with Russian forms, but they are judged to be at the same stage of evolution.
Bolivia, Madagascar, India
In south central Peru the upper Copacabana Group contains primitive Parafusulina with a few brachiopods, too few to 'provide any meaningful analysis (Newell et al., 1 953).
The Sakoa Group of the Onilahy region in south-west Madagascar commences with tillite of Kurmaian age, followed by coal beds, red sandstones and clays, and then the Vohitolia marine limestones with a few productids, apparently of Sterlitamakian age judged from Brachythyrina rectangulus (Kut.). The fauna is too incomplete to analyse properly.
Elements of the preceding faunas have been traced widely through the southern hemisphere by Waterhouse ( 1 970a) as the Stepanoviella-Taeniothaerus (or Reedoconcha) fauna, already briefly discussed for Iran, Saudi Arabia and Afghanistan. The Conularia beds, 25m thick, in the Salt Range, Pakistan, may belong here (Table 18) with Discinia, Neochonetes, Sulciplica, and Tomiopsis, though the fossils are so few that they could be as young as Sterlitamakian or as old as Kurmaian. The Sakmarian Stage is well represented in the Taeniothaerus or Reedoconcha faunas of the Agglomeratic Slate of Kashmir, including Bren Spur (Bion, 1 928; Reed, 1932; Kotlyar, 1964 ; Waterhouse, 1970a), with 'Streptorhynchus' bioni Reed, Neochonetes spp., Stepanoviella, Punctocyrtella nagmagensis (Bion), and Tomiopsis spp., suggesting a Sterlitamakian age.
Elements of the same Sakmarian fauna appear at Subansiri in Sikkim (Sahni &Srivastava, 1956; Singh, 1 973). From Peninsula India Reed ( 1928) described a small fauna at Umaria, regarded as Sterlitamakian in age by Waterhouse ( 1 970a) because of the presence of Stepanoviella, Tomiopsis and rare specimens of the bivalve Eurydesma, which also appeared at this horizon in New South Wales. A similar fauna was found in western Rajasthan by Sastry & Shah ( 1 964), sharing species found at Nagmarg and Yal Nal, Kashmir, recorded by Bion ( 1928). Spores also correlate the Conularia beds of the Salt Range with the Umaria beds (Ghosh & Basa, 1 969, p.407), within the lower 'Karhabari floral zone'.
Australia, New Zealand
In Western Australia (Fig.2 1) the Beckett Member of the Holmwood Shale, Perth Basin (Table 29) has a Tastubian ammonoid fauna (Glenister & Furnish, 1961) with Juresanites jacksoni (Etheridge) and a small distinctive brachiopod fauna (Waterhouse, 1970a) allied to those of the Conularia beds in the Salt Range and Subansiri, Sikkim. The slightly younger Callytharra Formation of the Carnarvon Basin, Western Australia, possesses a large brachiopod fauna (Dickins & Thomas, 1959; Thomas, 1958, 1967; Coleman, 1957; Prendergast, 1 943 ; Campbell, 1965), with related faunas in the Nura Nura Member of the Poole Sandstone in the Fitzroy Basin. Various ammonoids from the Nura Nura Member, including advanced Propopanoceras rhuzencevi (Glenister & Furnish) and Metalegoceras, were assigned to the Sterlitamakian Substage by Glenister & Furnish ( 1 96 1 ). Correlative faunas are found in the Fossil Cliff Sandstone of the Irwin or Perth Basin. Overlying faunas are meagre, but are so close to the
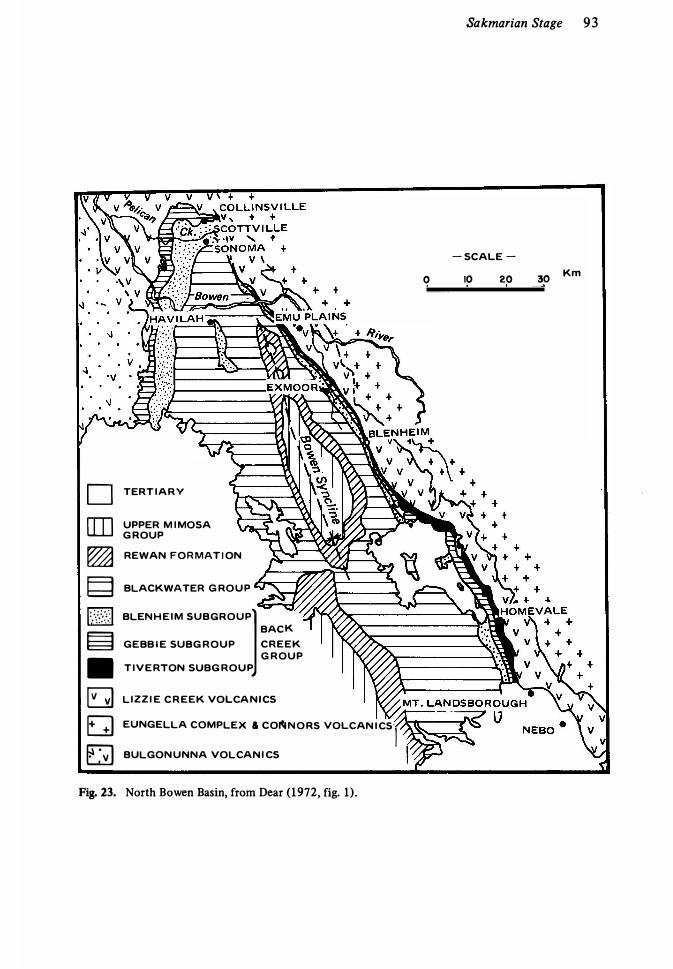
D
ITIJ
�
�
EEl
�
-
u
TERT I A R Y
UPPER M I MOSA G ROUP
REWAN F O R MAT I ON
BLACKWATER G RO U P
BAC K BLEN H E I M SUBGROUP]GEBB I E SUBG ROUP CREEK
G R OUP T I V E RTON SUBG ROUP
L I ZZ I E C R EEK VOLCA N I CS
Sakmarian Stage 93
- SCALE -
0 10 20 30 Km
+ +
Q
Q
EUNGELLA COMPLEX & CO� N O RS VOLCAN I CS·,/,k�""""'.-=""1
BULGON UNNA VOLCAN I CS
Fig. 23. North Bowen Basin, from Dear (1972, fig. 1).

94 Biochronology for the Marine Perwirm
170°
/•
North Auckland
NORTH ISLAND 3 8 °
4 2 °
SOUTH I S LAND
MTS Mataura Island
178•
TAKITIMU�· .
Southland Synclonorium
170° 174 ° 178° Fyfe's Lone t sl
42°
46°
Fig. 24. Distribution of Permian in New Zealand, on a craton at Tak�ka, along a volcanic arc and trough (arc-trench gap) in between Fyfe's Line and Tapanui Zone in the South Auckland-Nelson and Southland synclinorium, a mid-geosynclinal ridge of Clinton-Waipahi, and sporadically in a trough (with Fusulinacea more common) containing the Torlesse deposits in Otago and North Auckland. From Waterhouse (1973d, flg. 2, p. 35). The Alpine Fault transects the Permian, and is of late TertiarY - Recent age.

Baigendzinian Stage 95
Callytharra faunules that they are likely to be Aktastinian in age (Waterhouse, 1 970a). The largest of these faunas comes from the basal siltstone member of the One Gum Formation in the Wooramel Group, Carnarvon Basin (Dickins in Konecki et al., 1 958, p.33), characterised by Stepanoviella flexuosa Waterhouse.
Sakmarian fossils are widespread in eastern Australia. In Queensland (Table 36), Fauna 1 of Dickins (in Malone et al. 1 967, p.70) in the Lizzie Creek Volcanics in north Bowen Basin (Fig.23) could be Tastubian, though lumped with Fauna II by Runnegar (1969a). There is one distinctive species of Notospirifer. The Cattle Creek Formation of the Springsure shelf area, the Tiverton Formation of the Bowen Basin and the Yarrol Formation of the Yarrol Basin (Maxwell, 1 964) have yielded large correlative faunas, including ?Lissochonetes yarrolensis Maxwell, Echinalosia preovalis (Maxwell), Anidanthus springsurensis (Booker), Terrakea pollex Hill, Attenuatella australis Armstrong & Brown; syringothyrinids and Tomiopsis spp. (see Etheridge, 1 892; Campbell, 1 960, 196 1 , 1 965), here judged to be chiefly of Sterlitamakian age (Table 36). Eurydesma occurs in some of the faunas, suggesting cold waters. Species of Uraloceras, vaguely located in the stratigraphic column, were assigned by Armstrong et al. ( 1966) and Dear ( 1969) to the Aktastinian Substage, but this age requires verification from the associated faunas, which are yet to be described. The upper Cattle Creek Shale and Sirius Shale of the south-west Bowen Basin are possibly as young as Aktastinian, as the brachiopods differ slightly from those of the Tiverton faunas, and match those faunas in New Zealand (Waterhouse, 1 964, 1 969c). The Buffel Formation of south-east Bowen Basin is conceivably of the same age.
Moderately rich brachiopod faunas of much the same age as the Tiverton faunas are found in the Farley Formation in the Hunter Valley, Sydney Basin, of New South Wales (Table 40) with ' Uraloceras' characterised by an unusual suture. Somewhat allied faunas are found in the Ravensfield Sandstone, at the base of the Farley Formation. The underlying Rutherford Formation yields faunas too meagre to be assessed but, from stratigraphic position, could be Tastubian. Much the same fauna is found in the Golden Valley Group, Tasmania, with small Foraminifera studied by Crespin ( 1 958) and brachiopods summarized in Banks ( 1 96 1 , p.333 ; 1 962, p. 199), Johnston ( 1 888), and Waterhouse ( 1970d).
In the Takitimu Group of southern New Zealand (Table 38), the Telford Stage has two brachiopod zones containing such species as Echinalosia preovalis (Max.); and Terrakea pollex Hill, closely allied to those of the upper Cattle Creek Shale of Springsure, Queensland, and tentatively considered to be of Aktastinian age (Waterhouse, 1964, 1 968, 1 969b, 1 973c). The faunas are found in volcanic breccias, lOOOm thick (Fig.24).
BAIGENDZINIAN STAGE
Urals
The classical section of the Arti region in the Urals (Nalivkin, 1 949; Miloradovich, 1949; Stepanov, 1 95 1 ), commences with the Belokatai Suite ofpolymict conglomerates, 600- 1 OOOm thick, and the overlying Gabrashitov or Kashkabash Suite 100-200m thick, both containing upper Artinskian ammonoids (Ruzencev, 1 956). To the south in the lshibeisk pre-Urals the Burtsev and lrgin limestones with Aktastinian ammonoids are overlain by the Baigendzinian beds of coarsely fragmental limestone, and much argillite, also containing ammonoids. Characteristic ammonoid genera include Sakmarites, Propinac?ceras, Waagenina and Neocrimites with several other genera appearing for the first time. Metalegoceras and Paragastrioceras are especially abundant. Baigendzinian Fusulinacea include Pseudofusulina* insignita Viss., and Praeparafusulina

96 Biochronqlogy for the Marine Permian
futugini (Schell.), first appearing in the Burtsev Suite. Miloradovich ( 1949) recognised 1 3 7 species of brachiopods, of which 100 persisted from the Aktastinian Substage, and 3 7 appeared for the first time. However there have been no modern studies.
Two major divisions are recognised in the Baigendzinian Stage, the Sarginian followed by the Saranin (Nalivkin, 1 949; Likharev, 1 966, Table 4). The Sarginian, here treated as a substage, is characterised in part by Parafusulina solidissima Raus., Pseudofusulina* makarovi Raus., Cyrtella kulikiana Frederiks and several ammonoid species, including Medlicottia orbignyana Vern., and Waagenina subinterrupta Krotow. The Saranin is characterised by Parafusu/ina so/idissima Raus., Chonetina artiensis (Krot.), other chonetids, and Muirwoodia artiensis (Chernyshev). Miloradovich (1949) called the upper part Krasnoufimian after Frederiks ( 1 932), and this name is preferred for a substage over Saranin as the Krasnoufimian area yields a large brachiopod fauna (Frederiks, 1 934a), including some overtoniids, Uraloproductus stuckenbergianus (Krot.), marginiferids, Reticulatia, many Cancrinella, Anidanthus, Yakovlevia, Cleiothyridina, Purdonella, Martinia and Spiriferella. Judged from charts, there seem to be virtually no Saranin or Krasnoufimian ammonoids. Ruzencev ( 1956) dismissed the significance of the subdivisions, yet had no supporting evidence one way or other from ammonoids. Little value can be placed on such procedures.
From supposed Kazanian beds of the Aktubin Urals, Lipatova reported brachiopods that appear in fact to be of Kungurian or Baigendzinian age (Kulikov, 1 960). A huge brachiopod fauna from the Kolwa River region described by Stepanov ( 1 934, 1 93 7b) appears to be Krasnoufimian in age, judged from the presence of species of Reticulatia, Uraloproductus stuckenbergianus, Cancrinella, Martinia, Spiriferella pseudotibetana and other species. In Pai Hoi, .Petchora, the Gusin Suite at the base of the Yunargin Series has a few brachiopods, of perhaps early Artinskian age (Table 13). The overlying Belkov Suite ( 450-750m) has brachiopods, Sarginian in aspect, including Uraloproductus stuckenbergianus (Krot.), Waagenoconcha irginae Stuck., Spiriferella pseudotibetana etc. and various ammonoids, especially Waagenina supporting a Baigendzinian age (Solomina, 1 960). Ifanova & Semenova ( 1972) also recorded Chonetina artiensis (Krot.). The overlying Talatin Suite (350-700m) has similar faunas apparently correlative with the Krasnoufimian Substage. There are many links to the Bardin faunas associated with the Manchez reefs of Krasnoufimian age in the Ufimian amphitheatre. Species include many members of Streptorhynchus, Derbyia regularis Waagen, Chonetina spp., Kochiproductus, Waagenoconcha irginae (Stuck.), large Linoproductus, Reticulatia, Chaoiella, Sowerbina, Yakovlevia, and Cyrtella kulikiana (Fred.). The middle Talatin fauna is correlated with the basal Saranin (Krasnoufimian) by Gorsky & Guseva ( 1 973, p. 1 70). However Ustritsky ( 197 1} referred the Talatin Fauna to the Kungurian Stage. Perhaps the Filippovian Substage may be represented in the upper Talatin faunas because Spiriferella kei/havii (von Buch) in the fauna is typical elsewhere of Kungurian deposits, and ostracods support a Kungurian age (Gorsky & Guseva, 1 973), as do some of the brachiopods, such as Pseudosyrinx kolymensis Tolm. However a Kungurian correlation is not confirmed from recent studies by If an ova & Semen ova ( 1 972), who showed the Talatin Suite as Artinskian .
.
North Siberia, Pamirs, Afghanistan, Armenia (Table 1 5)Beds are thick but not very fossiliferous in south-east Verchoyan, Siberia. The
lower Endibal Series or Tumarin Suite, 1 ,300m thick, contains such ammonoids as Neouddenites adrianova Ruz. and Popanoceras tumarense Ruz. (Likharev, 1966, p.348). In the Tompo Basin of east Verchoyan the Kukkan Suite, 1 , 1 00- 1 ,300m thick,1s followed by the Dibin Suite (500-700m) with ' Chonetes' brama Frederiks, Stepanoviella, Jakutoproductus and Spiriferella, apparently Baigendzinian in age.

Baigendzinian Stage 97
In the Kolyma River sequence the Djeltin Horizon of plant beds or equivalent Kuktui beds may be correlative, with Horridonia and Spiriferella vaskovskii Zavodowsky, the latter also found in Baigendzinian beds of Canada. Zavodowsky (1970) assigned the beds to the Kungurian Stage.
The middle Darvasian Stage of the Pamirs and south-central Asia is approximately Baigendzinian, on the basis of Parafusu/ina and stratigraphic position. The Gundarin Horizon is Baigendzinian (Dronov & Leven, 196 1 ; Likharev, 1 966) with Nankinella, !Pseudofusulina* vulgaris, Triticites contractus (Schell.), Pseudofusulina kraffti (Schell.), Parafusulina, and Misellina and brachiopods Derbyia regularis Waagen, Marginifera and Choristites. Kalmikov ( 1964) described two zones in the Darvas region, the lower zone with Fusulinacea Pseudofusu/ina kraffti (Schell.) and Parafusu/ina, and Ammonoidea Propinacoceras bornemani Toum., Medlicottia cf artiensis timorensis, Agathiceras uralicus Karp., and Popanoceras /ahuseni Karp., the upper zone with Misellina claudiae (Deprat) and Parafusulina (Table 1 7).
Part of the Kubergandin Suite of the south-west Pamirs, some 50- lOOm thick, includes Popanoceras and Metalegoceras. Grunt & Dimetriev ( 1973) recorded a species of Terrakea, but it is too poorly preserved and decorticated to compare with Australasian species. Itrwould well be as young as Kungurian (Table 22), for Terrakea is principally, though not exclusively a cold-water genus (Waterhouse, 1 97 1), and so may beexpected to appear in paleotropical realms during the cold Kungurian Stage. Moreover Kubergandin Fusulinacea described by Leven ( 1 967) are of Kungurian age.
In Afghanistan south of Kabul (Termier et a/., 1974), the black carbonates have yielded Parqfusu/ina ferganica Mikl.-Makl., and could be as young as Baigendzinian. From Mt Bamyan, Termier & Termier ( 1970) have described Bamyaniceras bornemanni (Toum.) with Artinskia, and Popanoceras and Perrinites hilli afghana Termier &Termier.
In Armenia the Sasnian Horizon, S0- 150m thick may be of Baigendzinian age but could be Kungurian (Table 44). It contains Pseudofusulina* afT. gregaria (Lee), Nankinella caucasica Dutk., and compound rugose corals. The Armin Horizon, S0-200m thick, is lithologically the same with Nankinella, Sphaerulina and corals like those of the Sasnian faunas. Brachiopods are few and have not been described, so that correlation must depend on Fusulinacea, with support from corals.
China, Japan
In China, the fourth and uppermost Fusulinacean zone of the Chihsia Limestone (Table 18) has Pseudofusulina chihsianensis (Lee) and Rugosofusulina multiseptata (Schell.) of possible Baigendzinian age (Chan, 1963), above a zone with Nankinella orbicularia Lee, and below the Maokou Limestone of Kungurian age. The upper part of the Chihsia Limestone in the Tianwo district of northern Kwangsi yielded Artinskia, Neocrimites and Agathiceras (Chao, 1 966, p. 1 8 1 5). But as shown below, at least part of the upper Chihsia Limestone is definitely Kungurian in age.
The Nabeyaman Stage of Japan, based on black or dark grey carbonates with shale in the Y amasuge Member of the Aso Group in K wanto Province, coincides with the Parqfusulina yabei Hanzawa Zone, containing Pseudoschwagerina and Pseudofusulina• krotowi (Schell.) (Toriyama, 1 967; Fujimoto, 1 96 1 ). Various fusulinid biofacies are recognised (Igo, 1 964), with a few brachiopods (Hayasaka, 1922, 1 923, 1925).
Canada, United States, Mexico
In Canada, the Baigendzinian Stage is represented by the Antiquatonia brachiopod Zone in the basal 20m of the Tahkandit Formation (Table 22), with Derbyia cf grandis

98 Biochronology for the .Marine Permian
waagen, huge Linoproductus and Cyrt.ella, foll�we� by the Sowerbina. Zone at 70-1 OOm above the base with Muirwoodza cf artzenszs (Krotow) found m the Urals. Bathymyonia sterlitamakensis (Stepanov), Reticulatia u�?lensis (Likharev),Linoproductus, Spiriferella pseudotibetana Step., S. vaskovsku Z�v. and . Cyrtellaspecies are allied to species in the Talatin beds o� Petchora. The brach10p�?s differ considerably from those of underlying Yukon zones m the absence of Ove�tonnds and othe�early Permian species and genera. Ross ( 1967a) corr�lated S�hwagerma from the.Antz
quatonia Zone with Lenox Hills or late Wolfcampian specimens of Texas. This age seems too great. But Alaskanella yukonensis (S�inner &; �ilde), from the sligh�lyyounger Sowerbina Zone was correlated, c�rrectly m �y opmion, by Ross ( 1967a) With the upper Leonard, i.e. Cathedral Mountai� Formation of the Glass M?untams. TheTastubian age assigned to the same Fusuhnacea by Mamet & Ross (m Bamber & Waterhouse 1 97 1) cannot be reconciled with brachiopod evidence and stratigraphicposition above Sakmarian brachiopods and ammonoids. Neouddenites caurus Nassichuk et al. ( 1965, p.4 1) from northern Yukon Territory possibly came from beds ofthe same age, and was compared with N. andrianovi Ruzencev from the Endibal Series(Baigendzinian-Kazanian) of Verchoyan.
Similar brachiopods are found in the Belcher Channel Formation of Devon Islandand elsewhere in the Sverdrup Basin, Canadian Arctic Archipelago, accompanied bySchwagerina hyperborea (Salter). This species was matched with Praeparafusulina lutugini by Harker & Thorsteinsson ( 1960) and was regarded as early Artinskian (i.e. Aktastinian), but P. lutugini ranges into the Baigendzinian Stage.
Fairly large brachiopod faunas have been collected from the Halleck Formation of the Alaska Panhandle, in dark-grey calcareous siltstone and calcareous sandstone, with detrital chert and volcanics (Buddington & Chapin, 1 929; Muffier, 1967). The brachiopods were assessed as 'Leonardian' by Dutro in Muffier ( 1967), and Parafusulina has been reported, but a more precise determination must await description of the faunas.
The Grandeur Member of the Park City Formation in Wyoming, United States, may be of Baigendzinian age, with Productacea such as Squamaria ivesi (Newberry), Rugatia occidentalis (Newberry), and other species pointing to correlation with the Cathedral Mountain Formation of the Glass Mountains in west Texas (Yochelson, 1 968).
A moderate sized brachiopod fauna from the Toroweap Formation of Arizona (McKee, 1 938; McKee & Breed, 1 969) may be correlative with the Cathedral Mountain Formation, though it has many links to the younger faunas of the Word. According to Gilluly et a/. ( 1954), faunas of at least the upper part of the Colina Formation in south Arizona might be Leonardian. Fossils listed from the overlying Epitaph Dolomite are somewhat non-descript but may from stratigraphic position be correlative with the Cathedral Mountain Formation of west Texas, and the Toroweap Formation of north Arizona.
The Cath�ral Mountain Formation of the Glass Mountains, west Texas, is up to 500m thick, anti highly siliceous, with orange-coloured shale and thick sandstone near the top, bearing scattered bioherms (Fig. 29, Table 26). It is characterised by the brachiopod Institella leonardensis (King), and coincides with the fusulinacean zones proposed by Ross ( 1 962), based on Parafusulina spissisepta Ross and P. durhamiThompson. & Miller (Cooper & Grant, 1 964). A number of brachiopods are described by �.E. �I�g ( 193 1) and Cooper & Grant ( 1 969), but the bulk awaits publication in amaJor revisiOn by Cooper & Grant (in press). A few known speCies are comparable to thos� of the lower Tahkandit beds in the Yukon Territory of Canada (Antiquatonia bassz McKee, An�monaria 'pseudohorrida' (not Wiman) and Anidanthus), and the Cathedral Mountam faunas show the same disappearance of early Permian genera(Cooper & Grant, 1 973) as in the Yukon. Typical ammonoids include Medlicottia

Baigendzinian Stage 99
costellifera Miller & Furnish; Neocrimites newelli (Miller & Furnish), and P�rrinitesvidriensis Bose. Various other Baigendzinian ammonoid faunas are recogmsed by Ruzencev ( 1956) in the Blain Formation, north-central Texas; and San Andreas Formation, New Mexico, with Perrinites, Eumedlicottia, Agathiceras, and Stacheoceras, differing from the Urals faunas. Some brachiopods are known (Clifton, 1 96 1). As the brachiopods await publication by Cooper & Grant (in press), it is not possible to see if they show a two fold subdivision like those of most regions, and it is not clear if both Baigendzinian substages are present. However Ross ( 1962) did report two Fusulinacean zones.
The lower part of the sequences of lavas, conglomerates, limestones and clastics making up the Las Delicias Syncline in western Coahuila, Mexico, (King et a!. 1944) may be of Baigendzinian age (Table 35). A few brachiopods are listed by King, but await description. Ammonoids include Bitaunioceras, Perrinites and Medlicottia in 'beds' or bands 20 and 1 8 of the Difunta sections, and Parafusulina sellardsi Dunbar &Skinner occurs in 'bed' 14.
Pakistan, Australia, New Zealand
In the Salt Range of Pakistan, the Lower Productus Limestone or Amb Formation, up to 50m thick, contains a Monodiexodina fauna near the base, recently revised by Douglass ( 1970). It may be allied to the Parafusulina faunas in Saudi Arabia discussed by Hudson & Sudbury ( 1 959). Two faunas may be recognised by analysis of the Amb brachiopods described by Reed ( 1944, p.374) Table 28. The lower fauna is like those of the Antiquatonia fauna of Canada, and lower Talatin fauna of Pai Hoi, Russia, in the abundance of dictyoclostids, numerous Streptorhynchus and Lissochonetes, with Marginifera in place of the closely related genus Anemonaria. The upper fauna differs more and has no specific links. A general Baigendzinian age is suggested by Derbyia regularis Waagen, Rhynchopora variabilis Stuck., Lissochonetes, Cleiothyridina semiovalis (Waagen), Spiriferella, Pterospirifer, and Neospirifer marcoui (Waagen), with Chaoiella as in the Baigendzinian sequences of the Yukon Territory.
Correlative faunas of Western Austnilia are represented in the Lower Byro. Group of the Carnarvon Basin (Waterhouse, 1 970c, d), corresponding to Fauna D 1 of Dickins ( 1963). Two distinct faunas are recognised by analyses of the descriptions and distributions in Prendergast ( 1943), Thomas ( 1958, 1 967), Coleman ( 1957), Campbell (I 965), and other works mentioned by these authors (Table 29). The lower faunas, notably from the Madeline and Coyrie Formations, include Wyndhamia, Kiangsiella, Cancrinella, Licharewia?, Neospirifer, Taimyrella, and Martiniopsis. The upper fauna, in the Bulgadoo, Cundlego, Wandagee, and Quinnanie Formations, has · Streptorhynchus, strophalosiids including Echinalosia prideri (Coleman), many Aulostegids, including Aulosteges ingens Hosking, Taeniothaerus, various Productacea, Cleiothyridina, Fusispirifer, and Dielasmatidae . .The two faunas thus match the twofold subdivisions of faunas in the Urals, Canada and Pakistan. Thomas & Dickins (1954) have stressed brachiopod and bivalve links with the Amb Formation of Pakistan. These include, from the writer's studies, rare Overtoniids, Costiferina, Pterospirifer, and Neospirifer marcoui (Waagen). The Bulgadoo Shale, Quinnanie and Wandagee Formations include a few ammonoids with Baigendzinian affinities (Glenister & Furnish, 1961).
Probably Fauna Ilia of Dickins (in Dickins, 1 964; Runnegar, 1 969a) in Queensland is Baigendzinian. It is found in the lower Gebbie Formation (Fig. 22) of the Bowen Basin (Table 36), and contains a few brachiopods including Terrakea dickinsiDear, bivalves, and the ammonoid Neocrimites atT.fredericksi (Erne!.) of Baigendzinian age. A large fauna is found in the Berriedale Limestone of Tasmania (Table 40), correlated with the lower Byro Group of Western Australia by means of Taeniothaerus

100 Biochronology for the Marine Permian
Stage Substage Group
(Triassic)
Dorashamian? or
Djulfian Urushtenian
Punjab ian Chhidruan
ZALUCH Kalabagh ian
Kazan ian
Kungurian 50-300m
Krasnoufimian Baigendzin-
ian Sarginian
Sakmarian NILAWAN
Asselian Kurmaian 200-300m
a ·.-I .. Ill Ei 1-1 0 rz.
·.-I .... Ill 3: fa
·.-I ::e:
Formation Earlier Names
Narmia Topmost Limestone Member Dolomite
Mittiwali Ceratite Beds Member
Kathwai Upper Productus Member Limestone
Chhidru Formation
Kalabagh M .
Middle Productus War gal
Li���estone Limestone
(plant beds)
Anlb Lower Productus
Formation Limestone
Sardi Lavender Clay
Formation
Warchha Speckled Sandstone
Sands tone
Conu Za:roia beds Eurydesma beds
Tobra Talchir Boulder Beds
Formation
Table 28 . Permian and Triassic Formations , Salt Range , Pakistan .

-W
or
ld
S
ubs
ta
ge
s D
ick
ins
' F
aun
al
Pe
rt
h
Ba
sin
(I
r-
ca
rna
rv
on
B
asi
n
(No
rt
h
Can
nin
g
Ba
sin
S
ta
ge
s
win
R
ive
r A
re
al
en
d
of
Ken
ne
dy
Ra
ng
e)
(Fi
tzr
oy
R
ive
r
Ar
ea
)
Gr
ie
sb
ach
ia
n
Ko
ck
a t
ea
S
ha
le
Og
bi
nan
V
ed
ian
'R. ..
;.,,,
;.,,.,
Ur
ush
te
ni
an
.Chh
idr
uan
F
U
pp
er
m
ar
in
e
be
ds
i
nc
l.
Ha
rdm
an
Me
mbe
r
Ka
lab
ag
hi
an
B
in
th
al
ya
Fm
. L
iv
er
in
ga
Fm
!
So
sn
ov
ian
E
M
ung
ad
an
Ss
m
id
dl
e
Ka
li
no
vi
an
8J (?
Uf
im
ian
'
El
ki
n)
-�
-Gl
(Ne
vo
li
n)
Ir
en
ian
s::
Coo
lki
ly
a
Gw
Li
gh
tj
ac
k
& B
al
go
2
s:: � M
emb
er
s
Fi
lip
po
vi
an
Ba
ke
r
Fm
-
--
--
--
--
--
--
Kr
as
no
uf
im
ian
D
W
ag
in
a
Ss
N
al
bi
a Gw
W
and
ag
ee
Fm
1
8J
Cun
dl
eg
o
Fm
No
on
ka
nb
ah
Fm
0 B
ulg
ad
oo
S
ha
le
Sar
gi
ni
an
Min
ge
ne
w
Fm
�
Mal
le
ns
Gw
I:Q
Coy
ri
e
Fm
C
ary
ng
in
ia
Fm
P
oo
le
S
s Alt
ta
st
in
ian
c
Irw
in
R
iv
er
C
oa
l
Wo
or
ame
l
Gp
(u
pp
er
p
ar
t)
Me
as
ure
s
Hi
gh
C
li
ff
S
s
? S
te
rl
it
ama
ki
an
B
Fo
ss
il
Cl
if
f
Fm
Ca
ll
yt
ha
rr
a
Fm
Nur
a
Nur
a
Me
mbe
r
of
P
oo
le
S
s
·ra
st
ubi
an
Ho
lmw
oo
d
Sh
al
e
Kurma
ian
N
an
ge
tt
y G
la
ci�
C
ar
ra
nd
ib
b y Fm
A
Ly
on
s
Gp
1 U
sk
al
ik
ian
Gr
an
t
Fm
Su
re
nan
Ta
bl
e
29
. P
er
mi
an
c
or
re
la
ti
on
f
or
We
ste
rn
A
us
tr
al
ia
, em
en
de
d
fr
om
Di
ck
in
s
19
67
, T
abl
e
1.
Fm
Fo
rma
ti
on
; G
p
a
Gr
ou
p;
Gw •
G
re
yw
ac
ke
; M
em
Me
mbe
r;
Ss
S
an
ds
to
ne
.
� oq"
�
;:s fr ;;· s·
;:s
t'-1
� � .....
0

102 Biochronology for the Marine Permian
subquadratus (Morris) and ? Wyndhamia jukesi (Eth.) by Waterhouse ( 1969b, 1970d). Runnegar ( 1969a) referred the fauna to the Sakmarian Stage, equivalent to the Farley and Tiverton Fauna II, by using a very crude subdivision of time and faunas, but the fauna is definitely younger than Fauna II, and roughly equivalent to Fauna Ilia of Queensland. It is one of the most diverse of Permian faunas found in Tasmania, and oxygen isotope values suggest temperatures of up to 1 8oC (Dorman & Gill, 1961). World evidence also suggests warming during this time. However, Tasmania lay so close to the South Pole that its fauna was still of low diversity, and the value does seem high.
The Mangapirian faunas of New Zealand, found in the upper Telford Group of the Takitimu Mountains, Southland, have two divisions (Table 38). On the whole, the lower fauna, found in volcanogenic sandstones and conglomerate 7 ,OOOm thick, suggests correlation with the Berriedale Limestone of Tasmania, and the basal Byro Group of Western Australia through Wyndhamia jukesi (Etheridge), and Taeniothaerus subquadratus (Morris) and is considered to be Sarginian in age. The upper fauna in volcanogenic sandstones and siltstones 8,000m thick has affinities with higher Byro faunas with Echinalosia prideri (Coleman), Taeniothaerus afT. miniliensis Coleman and Aulosteges ingens Hoskings, and is considered to be Krasnoufimian in age (Waterhouse, 1973c).
KUNGURIAN STAGE
Russian Plaiform, Urals
Ammonoid workers in North America such as Glenister & Furnish ( 196 1) and Nassichuk ( 1 970; et a/. 1965) have regarded the Kungurian as part of the Artinskian Stage. This cannot be sustained from detailed faunal studies. The Kungurian Stage has distinctive brachiopods and bivalves and correlative faunas have distinctive ammonoids. It is true that Ruzencev ( 1 956) mentioned the possibility that the Kungurian ammonoids belonged to the Baigendzinian Stage, but he pointed out that no final judgement could be based on ammonoids, as they were so few in the type Kungurian Stage.
A second source of discussion concerns the general relationships of the Kungurian faunas. Russian, and recently, some North American workers, have classed the Kungurian, or early Word (that is Road Canyon) equivalents in North America, with the early Permian, on the basis that so many Baigendzinian forms persisted into the Kungurian Stage or in the case of west Texas, basal Kungurian· (i.e. Filippovian) Stage. However all stages contain species surviving from underlying stages. What is more significant is the entry for the first time in the Kungurian of many forms that characterised the Middle Permian Series. As Stepanov ( 1 973, p. 125) wrote, the faunas were rejuvenated. Other workers thoroughly experienced with the Kungurian faunas came to the same conclusion, including Gerassimov ( 1956). Brachiopod species are characteristic and significant Fusulinacea such as Neoschwagerinids (see Waterhouse, 1973a, p. l 88) entered palaeotropical waters, with the first members of the ammonoid family Cyclolobidae (Waterhouse, 1 972a).
Stratigraphy and faunas of the Kungurian Stage have been summarized by Zolotova et a/. ( 1966). The type sections are in the Kama River region, on the Russian Platform and eastern rim of the sub-Urals depression (Gorsky & Guseva, 1 973). Other stages have been proposed as substitutes for the Kungurian, including the Svalbardian Stage, based on Spitsbergen, by Stepanov ( 1 957), and the Paikhoyan Stage based on outcrops and faunas in Pai Hoi, by Ustritsky ( 1 97 1). The Svalbardian Stage appears to be of Kungurian to Kazanian age, though intended for pre-Kazanian rocks and faunas. It is incomplete, or not thoroughly studied, so that Elkin and Ufimian faunas have not been detected. A name to cover the Kungurian to Kazanian faunas would be useful but

Kungurian Stage 103
better based on Russian rather than Spitsbergen outcrops. The Paikhoyan Stage is richly fossiliferous and well controlled, though Filippovian and Kazanian faunas appear to be poorly represented. It is not clear why it is needed as a substitute, even though the Paikhoyan faunas are rich at some horizons. Another name that appears to have been largely ignored is the Kamian Stage, proposed by Zalessky ( 1930) for the Kungurian plus Ufimian. The Kungurian and Ufimian Stages are combined herein, as proposed by Gorsky & Guseva ( 1973), following Russian workers who appear to prefer to emend the Kungurian Stage rather than allow the name to lapse.
At the base of the Kungurian Stage, the Filippovian Horizon has a distinctive fauna that can be traced around the world, and so may be elevated to the status of a substage. It is formed in the type area of limestone, dolomite and sandstone, and has few Ammonoidea and no Fusulinacea. Small Foraminifera and Brachiopoda are diagnostic, with the first entry of species characteristic of the early mid-Permian, up to and including the overlying Kazanian Stage, such as Streptorhynchus pelargonatus (Schloth.), Neochonetes variolata (Vern.), 'Marginifera' septentrionalis Chernyshev, S]Jiriferellina cristata (Schloth.) and Dielasma elongatum (Schloth.). The fauna changed sharply after the underlying Baigendzinian, reflecting increased salinity and lowered sea-level that led to the death of Baigendzinian coral reefs (Kulikov, 1 943), suggesting onset of mid-Permian glaciation, revealed by cold water genera in Australia.
The overlying Irenian subdivision, here emended and elevated to a substage, is composed of salts, sulphates and carbonates. The lower horizon, the Nevolin, provides a distinctive and rich fauna in dolomite, with Pseudofusulina, and species of Derbyia, Chonetinella, Paeckelmanella expansa (Chernyshev), Cleiothyridina pectinifera (Sow.), Horridonia borealis (Haughton), A nemonaria species, and the ammonoid Paragastrioceras kunguriensis Mirsk. The fauna is very diverse, and involved the return of a number of Baigendzinian genera, and development of new species.
Overlying dolomites and limestones contain the meagre Elkin fauna, with a distinctive fauna of small Foraminifera, including the unique genus Kunguria. Many brachiopods disappeared. A few characteristic species include Reticulatia uralicus (Chernyshev) and Horridonia pseudotimanicus Gerassimov and distinctive nautiloids. Basically the fauna is an impoverished Nevolin fauna. It has been difficult to recognise elsewhere in the world, and may be of negligible significance, perhaps a local facies. However, the fauna was so short-lived that it will be discovered only by very careful work, and of course, might easily be represented in many sequences by unconformity, or barren beds. Waterhouse ( 1 973a, c) has suggested that correlative horizons exist in south-east Asia, east Australia and New Zealand. The horizon has not been recognised in the Arctic, where there may have been a sedimentary hiatus during this time.
Above the Elkin horizon comes the Ufimian unit, elevated to full stage status by Likharev ( 1 966, p.7 1), but difficult to recognise without detailed study. It has distinctive conchostracans and small Foraminifera, and is here placed in the Irenian Substage of the Kungurian Stage. Admittedly elements of its micro- and macro-faunas are Kazanian in aspect, but by retaining it in the Kungurian Stage, entry of the Kazanian brachiopod fauna remains a distinctive and useful time-line, a practise also supported by Stepanov ( 1973, p. 1 26), and Gorsky & Guseva ( 1 973). Alternative procedures would involve either the naming of a new unit for Irenian plus Ufimian, or, the recognition of Ufimian as a substage of the Kazanian Stage (Table 30). The Ufimian Horizon commences with the Solikamian salt deposits, about 200m thick, followed by the Shishmin Horizon with faunas towards the top, including Cancrinella koninckiana (Keys). Ostracods link the Solikamian to the rest of the Ufimian Horizon.
Kungurian brachiopods have been studied chiefly during the last .. century. More recently, a large fauna from the west slope of the Urals was listed by Kulikov ( 1 94 7); Chalishev ( 1966) recorded faunas from the northern Urals, and Likharev ( 1 9 1 3) redescribed a fauna from Kirillow. Grabau ( 1931 ) reported species from near

Ad
op
te
d
he
re
in
f
or
m
app
in
g
and
P
ri
or
it
y
of
N
ome
n-
Fa
un
al
a
ff
in
it
y
Sta
ge
� H
� � � H
8 �
sim
pl
ic
it
y"
cl
at
ure
em
pha
si
se
d
emph
as
is
ed
Sub
st
ag
e
So
sn
ov
ian
Kal
in
ov
ia
n
Ir
en
ian
Fi
lip
po
vi
an
Ho
ri
zo
n
Sub
st
ag
e
St
ag
e
So
sn
ov
ian
Ka
zan
ian
K
al
in
ov
ia
n
St
ag
e
Uf
im
ian
U
fim
ian
Ne
w
El
ki
n
Nam
e
Ne
ed
ed
I
J;e
ni
an
Ne
vo
lin
K
ung
ur
ia
n
St
ag
e
Fi
li
ppo
vi
an
--
---
--
Tab
le
30
. A
lt
ern
at
iv
e
pr
oc
ed
ur
es
f
or
n
om
en
cl
at
ure
o
f
ea
rl
y
Mi
dd
le
P
erm
ian
un
it
s
Sub
st
ag
e
So
sn
ov
ia
n
Ka
li
no
vi
an
Uf
im
ia
n
Ir
en
ia
n
Fi
lip
pov
ian
I I
- � b:l c· (")
::
:s..
.. 0 :s 0 c � � .... ;;. � � .... s· � � .... �· s· :s

Kungurian Stage 1 05
Novgorod, with the significant species ·crurithyris clannyana (King). C. acutirostris
(Krot.) has been listed by Likharev ( 1 966, p.226) in the Southern Urals, possibly but not certainly from Nevolin faunas. Ammonoids are rare in the Russian Kungurian, but include Neopronorites permicus Chernow, Uraloceras suessi (Karp.}, Artinskia artiensis (Gruen.) and Bitaunioceras krotowi (Dewing).
Kanin Peninsula, Pai Hoi, Kolyma, Sikhote Alin
Small Kungurian faunas were described from Kanin Peninsula by Frederiks (1934a) with typical Streptorhynchus, Echinalosia, Aulosteges and Tomiopsis. Kungurian beds are also known in Pai Hoi (Table 1 3) in the Vorkut Series described in part by Solomina ( 1960) and Mironova ( 1 960). The faunas were correlated with the Upper Artinskian and Kungurian by Likharev ( 1 966}, and revised by Ifanova &Semenova ( 1972}. The lower Levorkut Suite is subdivided into the Ayachyargin beds with Neopronorites permicus (Chernow) and Artinskia artiensis (Gruen.) and brachiopods such as Arctochonetes postartiensis (Ustrit.), Horridonia borealis (Haughton) and Paecke/manella expansa (Chernyshev), overlain by the Rudnitz Horizon, that shares many species with the Ayachyargin beds, especially chonetids. To the writer the two faunas look essentially similar, and are considered to be Nevolin in age. Ustritsky (1971) however placed the entire Vorkut Series in the Ufimian Stage. A Nevolin age would suggest that the Filippovian Substage was either missing, or represented by the meagre faunas of the underlying beds over the Talatin faunas with no distinctive species. Perhaps this is supported by a preponderance of linoproductids, and by the opinions of Mironov, Ustritsky and others as summarized by Likharev ( 1959) that the upper Talatin beds with Chonetina artiensis (Krot.) and Pseudosyrinx kolymensis Fred. are Kungurian (=Filippovian?}. lt would also explain why nearby supposedly correlative beds contain Licharewia ex gr. rugu/atus (Kut.) and L. stuckenbergi (Nechaev). These are normally Kazanian, but are here considered to be Nevolin, for the Licharewiinae also enter the Nevolin Substage in New Zealand and Australia (Waterhouse, 1968). The Intin beds, 250- 1 , lOOm thick, overlie the Rudnitz beds, and are dated as Ufimian by conchostracans.
A small fauna from the south island of Novaya Zemlya, described by Miloradovich ( 1936), appears to be of Kungurian age but one species alferovii Miloradovich may be Monticulifera (=Cancrinel/oides), a genus more typical elsewhere of the Kazanian Stage, unless it is Terrakea, which is widespread in faunas of Kungurian age. In central-west Taimyr Peninsula the Sokolin Horizon, 500-SOOm thick, has a small brachiopod fauna (Ustritsky & Chernyak, 1 963), assigned to the Ufimian but perhaps older Kungurian. The Sokolin fauna includes Tomiopsis mergensis Chernyak that resembles Tomiopsis from Nevolin beds in Arctic Canada and east Australia (Waterhouse, 197 1) and from Ufimian faunas in New Zealand (Clark et a/. 1 967). The Sokolin fauna also contains Chonetina, suggesting a Nevolin to Elkin age, Elkin being preferable. The underlying Efremov Suite of east Taimyr is approximately Kungurian, with a fauna so small that it is difficult to correlate: it includes Chonetina, permitting a Nevolin age.
In the Verchoyan Mountains of north-east Siberia the Dibin Suite, 500-700m thick, has Baigendzinian or Kungurian faunas to north and south, including Jakutoproductus and Tornquistia, found also in north Russia. To the south supposedly correlative beds also include Stepanoviella corcodonensis Likharev, member of a genus common in the Kazanian and Baigendzinian Stages. Indeed, one accompanying species Spiriferella saranae (Vern.), if correctly identified, suggests a Baigendzinian age, although the typically Kungurian ' Chonetes' brama Fred. is also reported.
In the Kolyma River area (Fig. 1 6) of north-east Siberia (Table 1 5) the Djigdalin Horizon a few metres thick has Kungurian brachiopods recorded in Likharev ( 1 966)

106 Biochronology for the Marine Permian
and Zavodowsky ( 1968, 1 970), with the age discussed by Waterhouse ( 1969c). Tornquistia perhaps favours a Nevolin age, supported by ammonoids Strigogoniatites zavodowskii Popov and 'Altudoceras' cf. roadense Bose. The Berein fauna of the Chiron district in Zabaikal is possibly of Nevolin age, with Pterospirifer alatus (Sow.), Pseudosyrinx, Tomiopsis and Licharewia (Kulikov, 1 965). But Neospirifer profasciger Likharev was also listed, perhaps incorrectly, for this species was recorded by Kotlyar & Popeko ( 1967) in faunas that appear to be of middle Carboniferous age.
A fauna from Cape Kalouzin, Sikhote Alin (Fig. 1 6) in the Kalouzin Suite, 800m thick, studied by Frederiks ( 1 925) was assigned to the Late Permian by Likharev ( 1966), but is more likely to be Nevolin in age (Waterhouse, 1 973a, p.200). It has 'Chonetes' brama Fred., 'Anidanthus aagardi gallatinense Girty' which is close to Chianella chianensis (Chao) of China, and several species related to Nevolin species of Arctic Can ada, including Timaniella harkeri Waterhouse, identified as S. vercherei not Waagen by Frederiks, Chonetina, Terrakea kozlowskianus (Fred.) cf T. arcticum Waterhouse, and Echinalosia cf E. maxwelli (Wat.) of Elkin age in New Zealand.
A fauna from nearby Ussuriland (Frederiks, 1 924) has been less well illustrated, but shares some species, and is said to contain Pterospirifer alatus (Sowerby), which if correctly identified, indicates a likely Nevolin age. Moreover Spiriferella /ita Frederiks is apparently allied to Spiriferella loveni (Diener), an early Kungurian species, chiefly of Filippovian age, from the Arctic. This Ussuriland fauna occurs in sandstone and siltstone, 500-700m thick, of central Sikhote Alin, shown as approximately Kazanian in Likharev ( 1966, Table 1 8). Underlying plant beds were assigned to an approximately Kungurian age, forming an extensive horizon in west Sikhote Alin, but may prove to be Baigendzinian. In turn they overlie Sakmarian Fusulinacean faunas in the Sibaigou Suite with Pseudofusulina, Parafusulina, ? Triticites, and Misellina claudiae (Deprat), (See Table 42).
Pamirs, Afghanistan, Iran
In the Pamirs of central Asia (Table 23) the Kungurian Stage is best displayed by Fusulinacean faunules described by Leven ( 1967, p.23). The upper Kubergandin Formation of limestone and shale 1 00- 150m thick contains a faunule of Pseudofusulina chihsianensis Lee, Parafusulina spp., Maklaya pamirica (Leven), M. cutlaensis (Leven) and Cancel/ina primigena (Hayden), overlain by beds with Pseudodoliolina, and then by beds with Maklaya cutlaensis and M. pamirica. These faunas were referred by Waterhouse ( I 973a, p.320, Table 3) to the Filippovian Substage. The overlying Gan Limestone, I 0- 120m thick, contains Neoschwagerina simplex Ozawa and Neofusulinella lantenoisi Deprat, according to Leven ( 1967), and was tentatively correlated with the Nevolin horizon by Waterhouse ( 1 973a). The Gan beds are overlain in turn by beds of presumably Kazanian age with Neoschwagerina craticu/ifera (Schwager). Grunt & Dimetriev ( 1973, Table 1) presented an amended sequence, as in Table 23. Certainly east Asian Fusulinacea suggest fine zonation comparable with that of the type Kungurian foraminiferal and brachiopod faunules (Waterhouse, 1973a), but it is not possible yet to securely match Asian with Russian zones, because Asian brachiopods, although listed, have not been described. Moreover the zonation is so fine that faunas were possibly mixed through sedimentary reworking or faulty collecting. Chief question must concern the age of Neoschwagerina craticulifera, or specimens so identified. Is the species early Kazanian as generally supposed, or did it enter in the Kungurian, especially the Irenian (?Nevolin) Substage?
Possibly the Terrakea species described from the Pamirs by Grunt & Dimetriev ( I 973) came from Kungurian beds. The species looks moderately like T. concavum Waterhouse in shape, but is so decorticated that the crucial spine pattern cannot be ascertained.

Kungurian Stage 107
Reed ( 1 93 1 b) described a fauna from Afghanistan near the Khojagar Dar a Dam with Overtoniids, many spiriferids such as Martinia and Martiniopsis, and Neoschwagerina craticulifera (Schwager), presumably indicative of a Kazanian age, although Waterhouse ( 1973a, p.200) preferred a Nevolin correlation. Indeed, nearby Cancel/ina primigena (Hayden) points to a Kungurian age, and also 'Fusulina' or Polydiexodina elongata (Shumard) occurs with Neoschwagerina craticulifera. Other fusulines, described by Thompson ( 1946) in the Bamian Series of Afghanistan, include Afghanella schencki Thompson and so-called Cancel/ina, referred to Maklaya afT. sethaputi by Kanmera & Toriyama ( 1 968), regarded as early Kungurian by Waterhouse ( 1973a). As a slight adjustment, perhaps Neoschwagerina commenced in the Ufimian Horizon, and M. sethaputi Kanmera & Toriyama should be regarded as Elkin, or mid-Irenian.
A fusuline fauna from Afghanistan, at Tezak south of Kabul, with Neoschwagerina simplex Ozawa, Polydiexodina, Pseudodoliolina, Russiella pulchra Mikl.-Makl., and other species occurs in fine-grained quartz with calcareous cement, and includes brachiopods identified with Enteletes conjunctus Reed, and Purdonella semiovalis (Reed) (originally described as Elivina), the former from the Warga! Formation of the Salt Range, Pakistan, the latter also occurring in the Chhidru beds of the Salt Range (Termier et al., 1974, p. 1 14). The basal beds contain Cancel/ina dutkevitchi Leven, with Stacheoceras rothi Miller and Furnish; followed by beds with C. dutkevitchi, Armenina asiatica Leven, Neofusulineila lantenoisi Deprat; and finally by a late Kubergandian fauna with Cancel/ina pamirica Leven, C. dutkevitchi, Armenina asiatica, and a number of ammonoids that show links with the Sosio fauna of Sicily. Species include Daraelites meeki Gemm., Paranorites konincki Gemm., Propinacoceras beyrichi Gemm., Neogeoceras trautscholdi (Gemm.), Eoglyphioceras meneghinii (Gemm.), Stacheoceras meditteraneum Gemm. and Popanoceras. The uppermost beds have yielded Neofusulinella lantenoisi, Praesumatrina neoschwagerinoides (Deprat) and Neoschwagerina simplex Ozawa (Termier et al., 1972).
Small Permian faunas from scattered localities in south-east Iran were described by Douglas ( 1936, 1 950), with Polydiexodina persica (' Skinnerina?) below ?Kazanian or late Kungurian beds with Neoschwagerina craticulifera and Cancel/ina primigena. To the writer some brachiopods listed by Douglas ( 1950, p.5) appear consistent with a Nevolin age, in view of the presence of Lissochonetes which is widespread in world faunas of Nevolin age; supposed Mentzelia, which might be allied to an Arctic Nevolin species Spirelytha schei (Chernyshev & Stepanov, 1 9 1 6), Spirigerella and Leptodus. Species from the Baktibari Range, south Iran, include Costalosia argentea Waterhouse & Shah from Malakabad which is allied to a B.urmese species C. bifurcata Waterhouse & Shah, that is accompanied by Polydiexodina. Other brachiopods are allied to species from the lower Wargal Limestone of Pakistan, and Cancrinella resembles an Arctic species of Filippovian age. Neochonetes deremsii (Douglas) suggests a possible Filippovian or Ufimian age. The fauna is 6verlain by beds with Neoschwagerina craticulifera and Cancel/ina.
From north Iran the Dorud fauna of the upper Djadjerud and Lar Valleys may be correlative (Table 3 1) but is so small and seems to have such an unusual combination of genera that certainty is lacking whether it is Kungurian or Kazanian. It underlies the Ruteh Formation of Punjabian age, with which it shares five of eight species, to perhaps support a Kazanian or early Kalabaghian age. Although it has Orthotichia indica (Waagen) and marginiferids like the south Iranian faunas, Neochonetes is present (typically Filippovian and lower Kazanian), with Orbicoelia, typical of Nevolin and younger faunas. On the whole the evidence suggests a lower Kazanian age. Sestini (1965c) assigned the fauna to the Darvasian Stage (Sakmarian, Artinskian).

108 Biochronology for the Marine Permian
S ub s tage
Ogbinan
Vedian
B aisalian
Urushtenian
?Chhidruan
Kalabaghian
S osnovian
Kalinovian
Krasnoufimian
Sarginian
Aktastinian
S terlitamakian
Iran , E lburz Mts
-
-
Upper Ne sen Ftm
-
? Ruteh 5
Ruteh 1 , 2
-
Dorud?
-
-
-
Ge irud D
Table 31 . Faunas in E lbur z Moun tains , north Iran .

Kungurian Stage 109
China, Japan, south-east Asia, Pakistan
In China, the Maokou Limestone above Chihsia Limestone consists of light grey thick bedded limestones 400-675m thick, (Table 1 8) devoid of ammonoids, but rich in fusulines, corals and brachiopods. There are three fusuline zones, of which the lowest is the Cancel/ina Zone, or Cancellina subzone in the Parafusulina Zone (in Sheng, 1 964) of approximately, or at least in part, Kungurian age. It is associated with Yanchienia, Verbeekina, Pseudodoliolina, and Chusenella, which entered the faunas for the first time with Parafusulina, and Schwagerina, which persisted from below. According to Sheng & Lee ( 1974) Cancellina occurs in the upper part of the Chihsia 'Stage', and is correlated with the Neoschwagerina simplex Zone of Japan. It seems possible that early Kungurian brachiopods are found at the top of the Chihsia Limestone of southern Anhui in the so-called upper 'lydite horizon'. Huang ( 1932a, p.26) and Chao (l927b, p. 179) listed a distinctive fauna including so-called 'Plicatifera' ( =? Overtoniina perhaps,or Neoplicatifera according to Sheng & Lee, 1974) huangi Ustritsky, with Tyloplecta nankinensis (Frech), T. yangzteensis (Chao) and Chianella chianensis (Chao). The latter species recalls 'Anidanthus aagardi gallatinense' (not Girty) of Frederiks ( 1925) from Cape Kalouzin. A slightly different fauna called the Hsiaochang fauna by Huang (1932b, p.96; Chao, 1927b, p. 1 79) occurs in the lower Yangzte Valley, especially Kiangsi, southern Anhui, Szechuan and southern Kwangsu (Fig. 1 7). Key forms include Chianella chianensis (Chao), Tyloplecta nankinensis (Frech), and Edriosteges medlicottianus (Waagen). Grabau ( 1934) also noted Ambocoelia or Crurithyris, suggestive of a Nevolin age. The fauna is accompanied by the coral Wentzelella subtimorica Huang which forms a zone at the base of the Maokou Limestone. Yet a similar fauna was listed from the top of the Chihsia Limestone in west Szechuan by Huang (1932a, p.27). Chan & Lee ( 1962) described a related fauna from Shansi, north China, with Orthotichia indica (Waagen), Chonetinella, 'Neoplicatifera' huangi (Ustritsky), Krotovia janus Huang, Chianella chianensis (Chao), Monticulifera sinensis (Frech), and Crurithyris. Underlying beds contain the Fusulinacea Cancel/ina schwellwieni (Deprat), (now = Praesumatrina Toumanskaya 1 95 1) and Neoschwagerina, and overlying beds contain Neoschwagerina, Verbeekina, Misellina, and Pseudodoliolina. Waterhouse (1973a) suggested that the older lydite fauna might be Filippovian, the younger Hsiaochiang fauna Nevolin. The Shansi fauna was also considered Nevolin. But, with Monticulifera sinensis (Frech) which is closely allied to and probably congeneric with Cancrinelloides of the Arctic Kazanian, the fauna could be upper Kazanian: which would allow Neoschwagerina to be early Kazanian in age.
A large ammonoid faunule is found in the Kufeng Shale of central Kiangsi, containing two significant primitive cyclolobids, l(ufengoceras and Shengoceras, marking the start of the Middle Permian Series (Waterhouse, 1 972a). All are referred to the Kufengoceras Zone, correlated with the Cancellina Zone by Chao ( 1 966), but with the Yabeina-Neoschwagerina Zones by Sheng & Lee ( 1974). Another ammonoid fauna occurs in carbonaceous shale of the lower Tingchiashan Formation of west Chekiang and north-east Kiangsi with Daubichifes shoutangensis (Chao), Waagenoceras and Mexicoceras. 'Paragastrioceras' or Altudoceras roadense Bose and other species have been recorded from much the same horizon. The faunas appear to be typically Kungurian, Daubichites being found in the Filippovian Assistance Formation of Canada, and Mexicoceras with Waagenoceras entering Nevolin faunas of west Texas.
A different suite of ammonoids was matched with the Cancellina-Neoschwagerina simplex Zone by Sheng & Lee ( 1974), with Pseudohalorites, Artinskia and Neocrimites. Amongst conodonts described by Ching ( 1 960) from the lower Maokou Limestone, Gondolella nankinensis has also been reported by Clark & Behnken ( 197 1 ) in the Streptognathodus sulcoplicatus and Gondolella idahoensis faunas of the Meade Peak Member in Idaho (of early Filippovian-Nevolin age). However Kozur ( 1974, p.49) and

1 10 Biochronology for the /f1arine Permian
Ching ( 1 960) considered that th� lo�er Mao�ou was older than �asal Gu.adalupian, though this · is difficult to reconcile w!th . fusuhne, coral, and brachiopod �vtdence.
As noted in discussion of the PunJabtan Stage, the faunas of the lower Jtsu Honguer beds in Mongolia may prove to be Kungurian.
In Japan, the Neoschwagerina simplex Zone is close to Filippovian or Nevolin in age, judged by its fauna and stratigraphic position (Table 1 7). It is classed by Toriyama ( 1 967) as part of the Nabeyaman Stage, and was correlated with the Cancel/ina Zone of China. The Kozaki Formation includes Skinnerella cf. sapperi (Schell. & Staff.), Verbeekina, Cancel/ina and Neoschwagerina simplex Ozawa.
A Nevolin brachiopod fauna has been described from Khao Phrik, south Thailand, by Waterhouse & Piyasin ( 1 970) and Yanagida ( 1970), with the age assessed on the basis of Brachiopoda by Waterhouse ( 1 973a, p. 1 94}. Some of the species of Streptorhynchus, Cleiothyridina, and Spiriferella a�e even related to those of Russia, and Orbicoelia is closely allied to Crurithyris found in contemporary faunas. Orthotichia indica (Waagen), Asperlinus asperulus (Waagen), ?Rugaria speciosus (Wat. & Piyasin) and Uncinella are other significant species. Sakagami ( 1968) recorded Ozawainella and Neofusulinella, supposedly of early Artinskian age, but this age is not acceptable.
Mansuy ( 1 9 14) recorded a number of brachiopods from Cambodia (Table 19) with some definite Kungurian - probably Nevolin - forms at Phnom Miai such as Uncinella shared with Khao Phrik and the Warga! Formation, Salt Range. A fauna from Phnom Ta Maol is either Filippovian or Nevolin, and others may be slightly younger, as at Phnom-Tu-Kreem. Some of the Cambodian brachiopods, Orthotichia indica (Waagen), ?Chaoiel/a margaritatus (Mansuy), Tyloplecta nankingensis (Frech), Urushtenia costulata (Thuan) and Chroistites are also found at Level A of Sisophon, Cambodia, at the base of the sequence discussed by Ishii et al. ( 1969), accompanied by Parafusulina padangense (Lange), and Pseudodoliolina pseudolepida (Deprat), of Kungurian age.
Several faunas from the Tonbo Limestone, Shan States, Burma, may be approximately of this age, judged in part by brachiopod affinities (Waterhouse, 1973a), though revision is needed to clarify the identities. One fauna was recorded from Namun, Shan States, with both Neochonetes and Lissochonetes, Orthotichia indica, mid-Permian Neospirifer, and ?Krotovia, of perhaps late Kungurian age. A larger fauna from Kehsi Mansam (Diener, 1 9 1 1 , p.68) has Orthotichia indica, Krotovia burmana Diener, Costa/asia bifurcata Waterhouse & Shah, Martinia, Martiniopsis /atouchei Diener and other typically Kungurian species, accompanied by 'Polydiexodina' elongata (Shumard), regarded as early mid-Permian by Douglas ( 1936, 1 950). The species may be S kinnerina according to Skinner ( 19 7 1 ), but is regarded as typical Polydiexodina by Dr D.J. Gobbett (pers. comm.). A Nevolin age appears likely. A related fauna from Mong Pawn, Burma, has some similar species with Rugaria as at Khao Phrik, Thailand, and Polydiexodina (Diener, 1 9 1 1 ).
In the Salt Range, Pakistan, the lower Wargal Limestone has a meagre fauna that includes Uncim!lla (Table 28). Most other species persist into higher stages, but Uncinella apparently indicates a Kungurian and perhaps chiefly Nevolin age. Other taxa in�lude Rhipidomella, Streptorhynchus, ?Rugaria strophomenoides (Waagen) (like the Khao Phrik species), Richthofenia, Spiriferella, Composita, and rare Hustedia. The basal coaly shales of this unit contain an even more impoverished fauna including species of Orthotichia, Streptorhynchus, Aulosteges, Cleiothyridina and Dielasma, generically quite close to the early Kungurian of Russia. It could be Filippovian, judged from faunal summaries based on Reed ( 1 944) and Waagen ( 1 89 1), but revision ·is needed to confirm the species ranges and identifications.
North-west Europe
In northern Europe (Fig. 25), the faunas of the lower Zechstein or Werra Formation in Germany and Lower to Middle Magnesian Limestone in England have been
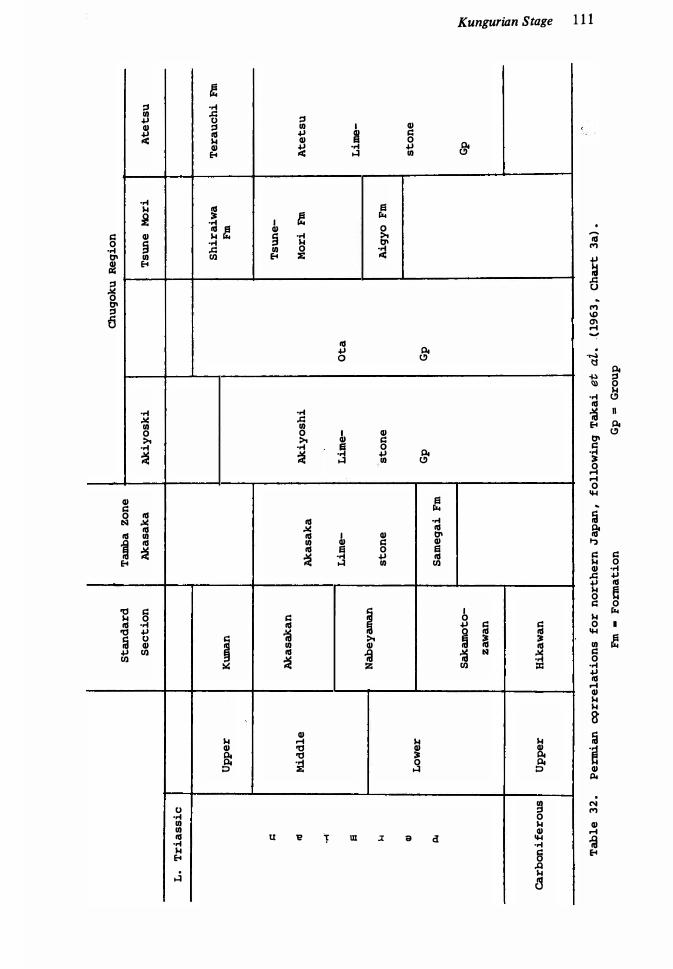
Chu
go
ku
R
eg
io
n
St
and
ar
d
Tamb
a
Zo
ne
Se
ct
io
n
Aka
sa
ka
Ak
iy
os
ki
T
sun
e
Mor
i
L.
Tr
ia
ss
ic
Sh
ir
aiw
a
Up
pe
r
Kuman
Fin
�
Ts
une
-Ak
as
aka
n
111 Mi
dd
le
Ak
iy
os
hi
M
or
i
Fin
Aka
sa
ka
. .-�·
Li
me
-L
im
e-
Ot
a
a N
abe
yaman
1-l
st
on
e
st
on
e
Aig
yo
F
m
Q) L
ow
er
G
p
Gp
llo
S
am
eg
ai
F
m
Sak
amot
o-
zaw
an
car
bo
ni
fe
ro
us
Up
pe
r
Hi
ka
wa
n
Tab
le
3
2.
Pe
rmian
CQ
rr
el
at
io
ns
f
or
n
or
th
er
n J
apan
, f
ol
low
in
g
Tak
ai
et
ai
•.
(1
96
3,
Ch
art
3
a)
.
Fm •
F
orm
at
io
n
Gp
=
Gr
ou
p
At
et
su
Te
ra
uc
hi
Fin
At
et
su
Li
me-
st
on
e
Gp
� i s
· ::
:s "-l
s
� .....
.....
.....

1 12 Biochronology for the Marine Permian
0 SOO km I
. ..:r·:; ··: · · • • •
. � � . .
Fig. 25. The late Permian Zechstein Sea in north-west Europe. Stippled areas, land. After Pattison et al. (1973, fig. 1).

Kungurian Stage 1 13
studied by many authors, including King ( 1850), with useful summaries in Mahlzahn (1937), Pattison ( 1970), Pattison et al. ( 1973), and Jordan ( 1969). The basal meagre fauna of the Zechstein is possibly Filippovian, and the well developed overlying Werra faunas of the Zechsteinkalk appear to be Irenian (Table 33) and specifically Nevolin by direct comparison with the sequences of Russia and the Canadian Arctic. Streptorhynchus pelargonatus (Schloth.), Horridonia horrida (Sow.), Cleiothyridina pectinifera (Sow.), Crurithyris clannyana (King), Spiriferellina cristata (Schloth.), and Dielasma elongatum (Schloth.) and many other species are found. The lower and middle Magnesian Limestones of England appear to have much - but not completely - the same faunas. There is one anomaly, in the presence of Neochonetes in the Magnesian Limestone, which might indicate an Ufimian age. Although it is here preferred to regard the occurrence as exceptional, due to unusually high salinity, an Ufimian correlation was strongly and perhaps correctly urged by Frebold ( 1933) and Likharev ( 195 9). The age of the overlying Zechstein beds cannot be established by brachiopods but the rhythmic cycles of evaporites suggest a climatic control which it should be possible to match with the glacial cycles of Gondwana and Siberia, to facilitate good correlation. There are no known Fusulinacea or Ammonoidea.
In recent years the faunas have been assigned a much later age, as Kazanian, or even Tatarian, but the positive identifications suggest a Kungurian age, as also con·cluded by Likharev (1 959) from a thorough comparison with the Russian faunas. Certainly this appears to be supported by the absence of well established Kazanian key genera, such as Cancrinelloides and Licharewia. A Kungurian age lies in sharp disagreement with spore studies summarized by Visscher ( 1973), who argued for a Tatarian age for the entire Zechstein Group. The upper Zechstein may well be late Permian in age, for marine faunas provide no control. But such a young age for the early faunas seems unacceptable. Although one must grant the possibility of error in interpreting marine faunas it must be pointed out that Palaeozoic spore correlations are not always well founded. For instance, an even greater margin of error has been documented for floras of the Canadian Arctic, in which numerous spore samples of indisputably Carboniferous age were confidently and quite wrongly assigned to the Permian Period, as pointed out by Bamber & Waterhouse ( 197 1). Presumably insufficient account was taken of different floral biomes and provinces. Support for the Kungurian age of the early Zechstein here adduced is provided by several Russian experts, including Gerassimov ( 1956), Stepanov ( 1957) and Likharev ( 1959).
Conodont authorities have also claimed a late Permian age for the entire Zechstein, but their correlations seem extremely ill-founded, too deferential, as Grant &Cooper (1973) devastatingly commented, to the one school of correlation that claims Cyclolobus was limited to the late Permian (e.g. Kummel & Teichert, 1 964 fT.) Conodont species are apparently shared between the Zechstein and east Greenland beds (Kozur, 1974), as would be expected, and this has been thought to require a late Permian age. On the contrary, the brachiopods and bivalves of Greenland show a Kungurian age. Moreover, a key species Neogondella divergens (Bender & Stoppel) from the basal Zechstein is also found in the upper Gerster beds of north-east Nevada, of Kungurian-Kazanian age, but wrongly assumed to be Capitan by Clark & Behnken ( 1971) - as discussed previously (Table 7). Conodonts are so widespread that they hold very high promise for correlation, but studies are just commencing, and, with so many apparently long-ranging species, cannot as yet match the degree of refined correlation achieved through study of Fusulinacea, Mollusca or Brachiopoda.
Spitsbergen, Greenland
. Faunas somewhat related to those of the Zechstein Sea are found in the SpiriferLtmestone or Voringen Member of the Kapp Starostin Formation, Spitsbergen, es-
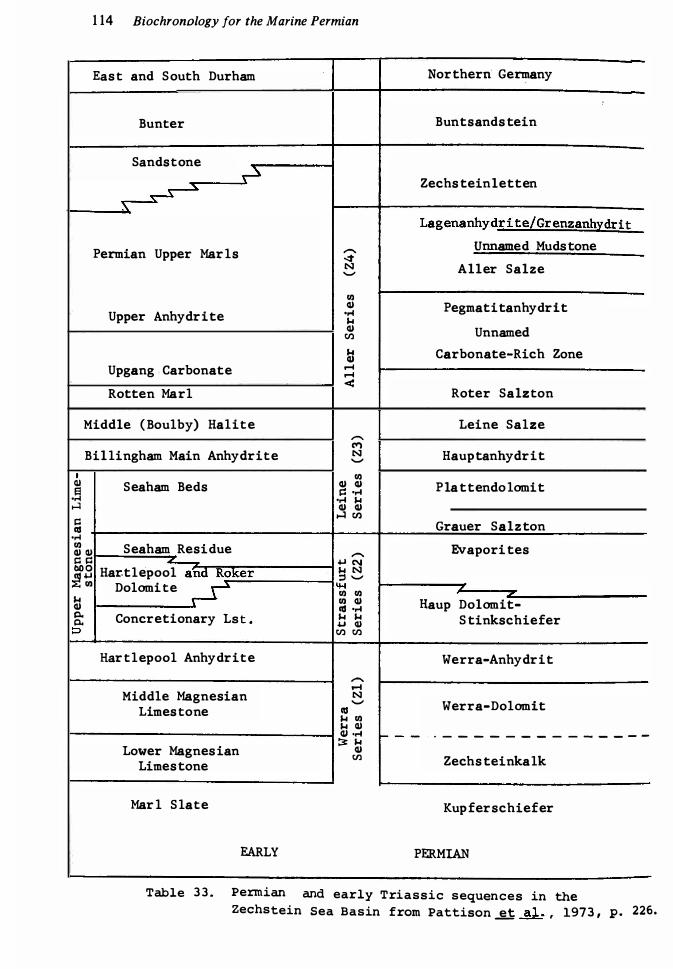
1 14 Biochronology for the Marine Permian
Eas t and South Durham
Bunter
Sandstone �
Permian Upper Mar ls -..;t N -U) QJ
Upper Anhydrite •.-I � QJ til
� Upgang Carbonate �
� <
Rotten Marl
Middle ( Boulby) Halite -M
Bil lingham Main Anhydrite N -I U) Cll Seaham Beds QJ QJ a s:: •.-1 •rl •.-1 � H QJ QJ s:: H tll Ill •rl U) Seaham Residue CII CII -s:: s:: "'--:£. .j.J NbO O Har.tlepool and Roker � N� � ::� -
Dolomi te 4-1 U) U) � U) QJ QJ Cll •.-1 Cl. Concretionary Ls t . � � Cl. .j.J Cll :;::1 til til
Har tlepool Anhydrite -�
Middle Magnesian N -Limes tone Cll � U) � Cll GJ •.-1
� � Lower Magnes ian QJ
til Limes tone
Mar l S late
EARLY
Northern Germany
: Buntsands tein
Zechs teinletten
LagenanhydritelGrenzanh�drit
Unnamed Muds tone
Aller Salze
Pegmatitanhydrit
Unnamed
Carbonate-Rich Zone
Roter Salzton
Leine Salze
Hauptanhydrit
Pla ttendo lomit
Grauer Salz ton
Evapori tes
Haup Dolomit-S tinkschiefer
Werra-Anhydrit
Werra-Dolomit
- - - · · - - - - - - - - - - - - - -
Zechs teinkalk
Kup ferschiefer
PERMIAN
Table 3 3 . Permian and early Triassic sequences in theZechstein Sea Basin from Pattison � a l . ; 1973 , p . 226.

Kungurian Stage 1 1 5
Productus Ls
Martinia Shale
, J\
25m Posidonia beds
20 1 5 1 0
5 0
Fig. 26. Permian and Triassic section at River 14, east Greenland, (Permian after Mayne, 1942; Triassic from Teichert & Kummel [ 1973, fig. 4, p. 282 ) ).

1 1 6 Biochronology for the Marine Permian
pecially at Bunsowland (Table 25), in grey or black limestone with sandstone and at Miseryfjellet, south-east Bj9)m9)ya (Fig. 1 8) see Gobbett ( 1 964, p.38) & Grabau ( 1931). A number of species are shared with the well dated Assistance Formation of the Canadian Arctic Islands to suggest a Filippovian age, including A rctitreta triangularis (Wiman), Grumantia (or A rctitreta) kempei (Andersson), Waagenoconcha,Thuleproductus arcticum (Whitfield), Spiriferella loveni (Diener), Pseudosyrinx wimani Gobbett, and Neospirifer striatoplicatus (Gobbett). There is no clear development of any younger Kungurian fauna, but it is possible that horizons have been mixed, for several key Nevolin species are present, especially the smooth chonetids and Licharewiids, either allegedly from the Spirifer Limestone, or more usually from the overlying lower and middle Brachiopod Chert or upper Kapp Starotsin Formation. Even Elkin or Ufimian faunas may be present, but these particular substages lack distinctive key species, and will only be detected by study of stratigraphically well controlled collections.
The rich brachiopod fauna from the Foldvik Creek Formation at Cape) Stosch, east Greenland (Fig. 26), occurs in 200-300m of interdigitating facies. It has been fully described by Dunbar ( 1955) to demonstrate a close similarity with Zechstein faunas of Europe. Significant genera and species include A rctitreta kempei (Andersson), widespread in Kungurian faunas of Arctic Siberia and Canada, Lissochonetes toulai Dunbar, as in Nevolin faunas elsewhere, Cancrinella, Liosotella, Craspedalosia pulchella Dunbar, Krotovia licharewi Frebold, Odontospirifer mirabilis Dunbar, Spiriferella keilhavii (von Buch), Sowerbina, and Neospirifer striatoparadoxus (Toula), which is the same as a Spitsbergen species, and differs from the Filippovian form N. striatoplicatus (Gobbett). The fauna is Nevolin in age_, by direct correlation with the type Russian sequences, and also with Canadian faunas. A Cyclolobid species is found, (e.g. in Nassichuk et a/., 1 965) and has been interpreted as of very late Permian age 'Chhidruan' (Furnish, 1 966), but the brachiopods incontrovertibly suggest an early midPermian age. Waterhouse ( 1972a) considered that the Cyclolobid was primitive, and used the generic distinction Godthaabites Frebold for species with such a simple cyclolobid suture. Conodonts of the Neogondolella rosenkrantzi (Bender & Stoppel) Zone are found in the Posidonia Shale, Productus Limestone and M.artinia Shale, (Sweet, 1973, p.64 1), with species known to range from Kungurian to Kazanian. Several brachiopod species from N athorsts Fiord, northern Greenland, recorded by Frebold ( 193 1), were considered by Dunbar ( 1 955, p. 1 8) to belong to the same fauna.
Canada
The brachiopods and molluscs from the green sandstones of the Assistance Formation at Grinnell Peninsula, Devon Island, Canadian Arctic Archipelago (Table 22) were initially described by Harker in Harker & Thorsteinsson ( 1 960). More brachiopods have been described by Waterhouse (e.g. 1 969a; 1 97 1 ; Bamber &Waterhouse, 1971 , and Sarytcheva & Waterhouse, 1 972). Several species, including Arctitreta pearyi (Whitfield), Neochonetes, Thuleproductus arcticum (Whitfield), Muirwoodia mammatus (Keys.), Pseudosyrinx, Spiriferella loveni (Diener), and Neospirifer striatoplicatus (Gobbett), suggest a Filippovian age, as confirmed bystratigraphic position, and by the sudden disappearance of Fusulinacea, as in Russia.Ammonoids described by N assichuk ( 1 970; et al. 1 965), and correlated with the RoadCanyon Formation of Texas, include Daubichites fortieri (Harker), Sverdrupites harkeri (Ruz.), Synartinskia belcheri Nassichuk, and Popanoceras cf sobolewskyanum(Vern.). Although Nassichuk ( 1970) favoured an Artinskian age, .as well as a Road Canyon c�rrelation, an anal�sis of the ammonoid affinities shows a wide range of possiblecorrelation, fr�m .sakmanan to Kazanian, and closest resemblances lie with Kunguriangenera (Daubzchztes, Popanoceras, and Sverdrupites) in China Russia and WesternAustralia.
'

A
NOU
HWES
T
I NO
RTH
WE
ST
ER
N
CA
RB
ON
AT
E
IE
LT 1BAS
INA
L C
LAS
TIC
A
ND
EVA
PO
RA
TIV
E
IEL
T
15.E.CARI
ON
ATE
'MARG
INA
L C
LAST
IC l
IEL
T AND
CAR
BONA
TE
u ;;; � iii ....
;-:/_; _i./;1£
·: C·::.+nFY
<� ;! � \'( �-; ;_ ::
::·:.-:::::-A ·
IEL
T •
IJO
RN
E
•
z c i • �
_..
.,.r
.//
............
-./_
�
........r-r
L..,-o.
:?'
�.-�
�
��
�� �
NANSEN
• 0�
"'-T-'"
.
. ..,.
.... .
... u
!!:
MA
RGIN
AL
CL
AS
TIC
I
ELT
. DA
TUM
Kl-
lrH
21
10
Fig
. 2
7.
Perm
ian
form
atio
ns o
f th
e C
anad
ian
Arc
tic
Arc
hipe
lago
, fro
m N
assi
chuk
et
al. (
197
3, fi
g. 2
, p. 2
88).
Red
raw
n w
ith
perm
issi
on o
f au
thor
s.
I SO
UTHE
AST f§ .
..
E L 1 0 t 1 . L I
� � E;• ;:s t'.l S' � .....
.....
......;)

1 1 8 Biochronology for the Marine Permian
Nevolin faunas are also widely present in the Arctic (Fig. 27) especially in the Van Hauen Formation, and part of the Degerbols Formation on Ellesmere and Axel Heiberg islands. The fauna is ch·aracterised by a new species of Thuleproductus, Pterospirifer alatus (Sow.), Chonetinella, Lissochonetes, and various other species. One well preserved brachiopod faunule was described by Whitfield ( 1908), another by Stehli & Grant ( 1 97 1) from somewhere (not known) on Svartevaeg, Axel Heiberg Island, with no stratigraphic detail provided. The age was stated to be Kazanian by Grant & Cooper ( 1973), but this is in error: the fauna is Kungurian. Possibly the fauna recorded from Great Bear Cape by Chernyshev & Stepanov ( 1 9 1 6) is correlative, bearing the typical Nevolin species of Thuleproductus. Although the presence of Overtoniids point to a greater age, such have also been reported from Pai Hoi.
The middle Tahkandit Formation exposed near the Arctic Circle in Yukon Territory has two Kungurian brachiopod zones in rubbly limestones and breccias, and allied faunas are found to the north in the Richardson Mountains (Bamber &Waterhouse, 197 1). The lower zone includes the widespread key Neochonetes, Cancrinella phosphatica (Girty) allied to an Iranian species, Thuleproductus arcticum as in Arctic faunas, Anidanthus cf eucharis (Girty), Pseudosyrinx and Neospirifer striatoplicatus (Gobbett), and other species shared with Kungurian faunas of the Arctic. The overlying fauna includes Timaniella harkeri Waterhouse as in the Kamchatka fauna of Frederiks ( 1926), and Lissochonetes and Crurithyris as in many Nevolin faunas over the world.
Various lists of faunas by Girty in Mertie ( 1930) and Brabb & Grant ( 1971) suggest that the Kungurian Stage is represented in the Tahkandit Formation of Alaska, though no detailed age studies have yet been published. From the Alaska Panhandle the Pybus Formation of chert and dolomitic limestone has yielded brachiopods listed by Girty in Buddington & Chapin ( 1929) and assessed as Roadian (=Filippovian) by Grant ( 1 97 1 , p.3 1 6), which would be in conformity with the presence of Septacamera, so far not reliably known in younger rocks. However Loney ( I 964, pp.95-96) recorded ?Licharewia and ?Crurithyris to suggest a Nevolin age.
A few brachiopods in the Ranger Canyon Chert in the Rocky Mountains of west Alberta indicate a possible Kungurian age, with Anidanthus, Cancrinella and Terrakea species like those of the Tahkandit Formation, Alaska-Yukon, and the Phosphoria complex in north-west United States (Logan & McGugan, 1 968).
The Cache Creek Group of central British Columbia is thick and contains a number of Fusulinacean zones, including a well defined Yabeina assemblage of Chhidruan-Kalabaghian age in the Marble Canyon limestone (Monger & Ross, 1971 , p.270). The only sizable brachiopod faunule from near Kamloops; as described by Crockford & Warren ( 1 935), is probably of Nevolin age, judged from examination by the writer. The fauna is like that of south Iran, with Juresania, marginiferids, and two Spiriferellinids of Kungurian age in ? Timaniella cf harkeri Wat. from the Canadian Arctic and Spiriferella cf supplanta Wat. from New Zealand. Accompanying ammonoids were assessed as Word 2 or 3 (China Tank and Willis Ranch Members) i.e. early Kazanian, late Kungurian by Dr W.W. Nassichuk, Geological Survey of Canada, Calgary, pers. comm. The fauna lacks Leptodus, unlike a tiny (?Punjabian) faunule recorded by Kindle ( I 926) from elsewhere in the Cache Creek Group.
United States, Guatemala
From the Nosoni Formation of northern California, Coogan ( I 960) recorded advanced Parafusulina and Monodiexodina with Lissochonetes, Chonetinella, Crurithyris and Cancrinella phosphatica (Girty), all showing specific links with the Nevolin faunas of the Canadian Arctic.
Mills & Davis ( 1962) recorded a few brachiopods in Washington State, including

Kungurian Stage 1 19
Anidanthus minor (Cooper) in siltstone and limestone, with Pseudofusulinella, Schwagerina and Parafusillina antimonionensis Dunbar, suggestive of an approximately Kungurian age.
The Phosphoria Rock Complex of Idaho, Wyoming, and Montana, (Table 34), has yielded small correlative brachiopod faunas described by Girty ( 1 9 1 0, 1 927), Branson ( 1930) and others, as summarized by Yochelson ( 1968) and McElvey et al. ( 1959). A number of species range from the Arctic to this region, such as Lissochonetes ostiolatus (Girty), Cancrinella phosphatica (Girty), Anidanthus eucharis (Girty), Yakovlevia geniculata (Girty), and Crurithyris arcuata (Girty). The Meade Peak member is probably Nevolin judged from the presence of Lissochonetes and Crurithyris. Furnish ( 1973, p.354) referred the Meade Peak faunas to the slightly older 'Roadian Stage', i.e. Filippovian, on the basis of Glassoceras bransoni (Miller & Cline) and Spirolegoceras fischeri Miller et al. Furnish ( 1 973) also referred Verchoyan ammonoids to the same fauna, but associated Verchoyan brachiopods are clearly Kazanian in age, suggesting that the ammonoids are long ranged from Filippovian through Irenian to Kazanian. The Rex Chert just above the Meade Peak unit could be either Nevolin or Elkin - its fauna is not large, and includes Kochiproductus, Anidanthus and Muirwoodia, all suggestive of a cool episode and impoverished fauna, as in the Elkin faunas of Russia, Australia and New Zealand. The lower Shedhorn Sandstone of a correlative but different facies has a meagre fauna with Neospirifer jJseudocameratus (Girty) that resembles early midPermian Neospirifer (fasciger type) rather than the moosakhailensis type, and so likely to be Baigendzinian or Kungurian. Accompanying species on the whole suggest an Elkin age. The fauna from the Retort Member has a small fauna with no obvious keys, and could be Ufimian or lower Kazanian. However, there is little value in these correlations, for the faunas need to be revised, and then compared with the Glass Mountain faunas, which await publication.
-
The Franson Member in the middle of the Park City Group (Yochelson, I 968; Williams in McKelvey et al. 1 959) is not easy to date. It intertongues with the Meade Peak and Rex Chert beds, and its fauna shows a mixture of affinities, with Derbyia magna Branson, Sphenosteges hispidus (Girty) and Sphenalosia smedleyi Muir-Wood and Cooper, related to Kazanian specimens in the high Ranger Canyon beds of Alberta and Trold Fiord beds of the Canadian Arctic. On the other hand Neospirifer pseudocameratus (Girty) is like N. striatoplicatus (Gobbett) from the Filippovian, and Cancrinella phosphaticus appears to be typical of the Meade Peak or Nevolin fauna. This mixture of affinities suggests an age somewhere between that of the Meade Peak and perhaps Rex Chert on the one hand, and the Tosi Chert on the other. Moreover a few species, such as Antiquatonia cf. sulcatus Cooper are shared with the Rex and Retort members. The overlapping ranges of the species suggest an Ufimian age, but further study is needed to determine limits closer than late Kungurian or early Kazanian.
The Kaibab Formation of north Arizona has a number of species said to be identical with Cathedral Mountain forms, such as Dyoros, Quadrochonetes and Peniculauris bassi (McKee) but the fauna is apparently to be correlated with the Road Canyon Formation, according to McKee & Breed ( 1969). Peniculauris bassi is also present with other key brachiopods in the Concha Formation of south Arizona (Williams in Gilluly et al. 1 954).
The lower middle part of the Las Delicias Permian in Mexico is probably Kungurian with Bitaunioceras (King et al. 1 944). Dating is not under good control as yet, but it appear� likely that 'beds' 8, 1 1 , 1 2 and I4 of the Malascachas sections are approximately Kungurian in age (Table 35). Bed 14 with Parafusulina sellardsi Dunbar &Skinner could be correlative with the Cathedral Mountain Formation. By comparison with other faunas, Bed I I could be Filippovian, as it contains Neochonetes and Bitaunioceras, and shares several species with underlying 'Cathedral Mountain' or

St
and
ar
d
Ka
zan
ian
S
tag
e
Ire
ni
an
Sub
st
ag
e
So
sno
vi
an
Sub
sta
ge
Ka
lin
ov
ian
S
ubs
ta
ge
?Ufi
mian
?Elk
in
Ne
vo
lin
?Fi
lip
po
vian
S
ubs
ta
ge
Ba
ig
en
dz
in
ian
St
ag
e
Up
pe
r
20m
Sh
edh
or
n
San
ds
to
ne
low
er
15
m
low
er
? ?
Ph
osp
ho
ri
a C
omp
lex
�ar
k
Cit
y
Fo
rm
at
ion
To
si
Ch
er
t M
emb
er
30
-S
Om
Er
va
y M
emb
er
5
-30
m
20m
ch
er
ty
sh
ale
m
emb
er
Re
to
rt
P
ho
sph
at
ic
S
ha
le
Me
mbe
r
20
-30
m
�--------------------------------!Fr
ans
on
10
-7
0mM
eml?
er
Re
x C
he
rt
Me
mbe
r
6m
-20
m
Me
ad
e
Pe
ak
e
Ph
osp
ha
ti
c
Sh
ale
M
emb
er
30
-6
5m
Lo
we
r
Ch
er
t
Me
mbe
r
Gr
an
de
ur
M
emb
er
1
5-
12
5m
.. ' ·-'1
Ta
bl
e
34
. T
en
ta
ti
ve
c
or
re
la
ti
on
s
of
t
he
P
ho
sp
ho
ri
a
Ro
ck
C
om
pl
ex
o
f
:Id
ah
o,
Wy
om
in
g,
Mo
nt
ana
.
.... � b:l c:;· g. � :::s 0 � 'C' "'f :;.
� � s· � � � Ej• :::s

I I ? Kazanian I
? lrenian - -_ , _
- -- -
.- -- -
- -
= ·,;...;_ - -
- -
- -- -
- - - - - - - - - - --
Filippovian - - -- -
- - -- -
- - -
Kungurian Stage 12 1
SP-iriferell ina
Compos ita
Cancrinella-
LeiorhY-nchoidea
Dictyoclostus
Parafusulina
Anidanthus (2)
Anidanthus ( I )
Feet 250 200 150 100 50 0
Scole
Fig. 28. Columnar section of the upper part of the Permian strata in the Monos Hills, El Antimonio, Mexico, from Cooper et al. (1953, fig. 3), with principal genera. Age tentative.

1 22 Biochronology for the Marine Permian
Baigendzinian faunas in bands 1 2 and 1 4. Description of the Las Delicias faunas and comparison with the Glass Mountain faunas may establish that a full Kungurian sequence is present.
At El Antimonio, Mexico (Fig. 28), 500m of shales and limestones of the Monos Formation are in part Kungurian (Cooper et a/. 1 953), with a sequence of brachiopod faunas outlined by Cooper. The basal Anidanthus Zone might be of Filippovian age, perhaps with the overlying Dictyoclostus Zone, which has Neochonetes and Anidanthus. Muirwoodia and Cancrinella occur in the Dictyoclostus and overlying Cancrinella Zones. Species of Spiriferella appear to be typically early Middle Permian, with Spiriferella scobinoidea Cooper from the A nidanthus Zone resembling the Arctic S. keilhavii (von Buch) and Spiriferella sp. B from the Cancrinella-Leiorhynchoidea Zone like the Arctic S. loveni (Diener). Neospirifer from the latter zone has high sharply defined plicae as in Arctic and other Neospirifer of Kungurian age. Perhaps the Cancrinella and Composita Zones are late Kungurian, i.e. Irenian in age, but detailed correlation must await description of the nearby brachiopod faunules from the Glass Mountains. Large Parqfusulina occurs just above the Anidanthus Zone.
The Road Canyon Formation of Texas, named by Cooper & Grant (1964, 1966) for Limestone no. 1 at the base of the Word Formation of P.B. King ( 193 1 ) is probably Kungurian, possibly Filippovian, to judge from Fusulinacean and ammonoid studies (Table 26). It has an early Cyclolobid ammonoid genus called Glassoceras and advanced Parafusulina. Other ammonoids include Eumedlicottia burckhardti (Bose) and Perrinites hilli (Smith). Some brachiopods have been described by Muir-Wood &Cooper ( 1960) and Cooper & Grant ( 1 969, 1 973), pending publication of Cooper &Grant (in press). Conceivably the overlying Word Limestone no. 2 or China Tank Member is Nevolin, but few brachiopods have yet been described (see Cooper & Grant, 1 973).
The Chochal Limestone of Guatemala has yielded a modest brachiopod fauna, correlated with the Cathedral Mountain or Road Canyon Formation by Stehli & Grant ( 1970). On the whole detailed affinities, especially for Edriosteges, suggest a Road Canyon age, as reinforced by discussion with Dr R.E. Grant, Smithsonian Institution. A number of species do suggest a slightly greater age (Echinauris, Hercosestria, Hercosia, Peniculauris, and Chonosteges), but the latter occurs in the Nevolin fauna of Thailand (Waterhouse & Piyasin, 1 970). Fusulinacea indicate a Leonard or possibly Word age the very indecision pointing to a Road Canyon correlation.
Sumatra, Timor
From the Djambi tuffs, andesitic lavas and limestones of Sumatra, faunas recorded by Roemer ( 1 880, p.85), Fliegel ( 1 901) and Lange ( 1 925, see. p.28 1) include Orthotichia, Tyloplecta sumatrensis (Roemer) and marginiferids suggestive of a Kazanian or Kungurian age. The presence of Crurithyris in one list implies a Nevolin age (see Roemer, 1 880, p.85). Fusulinacea include Verbeekina verbeeki (Geinitz). Brachiopods from two fusuline limestones in the Airkunung beds in the Djambi Residency of Sumatra (Marks, 1957) as described by Meyer ( 1 922) could well be Nevolin in age, and are somewhat like those recorded by Roemer ( 1 880).
The Bitauni fauna of Timor (Tables 1 9, 44) includes a number of brachiopod species (Broili, 1 9 16 ; Hamlet, 1 928) of Kungurian age, although retaining links with the Baigendzinian, such as 'Productella' patula Hamlet (not Girty) which is an overtoniid (cf Overtoniina?) species allied to 'Plicatifera' huangi from Kungurian faunas of China, Stepanoviella allied to species from the Arctic Kungurian (Waterhouse, 1 970), Trigonotre!a or . .
Licharewiinid as Spirifer simaanensis Hamlet, and ?Rugaria molengra)!l (Brmh), perhaps a senior synonym of R. speciosus (Waterhouse & Piyasin)from Tha1land. Martinia resembles Kungurian specimens described by Diener ( 1 9 1 1)

Ma
la
sc
ac
ha
s D
if
unt
a
Ki
ng
et
a�
. 1
94
4
Be
d
Be
d
Th
ick
ne
ss
in
M
et
res
1.
Vo
lc
ani
c
ro
cks
3
20
2.
Kin
gooe
ras
s
ha
le
7
0
1
3.
Vo
lc
ani
c
z:o
ck
s 2
20
2
4-
7
Tim
o:ri t
es s
ha
le
1
60
3-
17
a-
24
Waagen
ocer
as
sh
al
e
Pam
fusu
Un
a
21
30
vo
lc
ani
c
ro
ck
s
18
-2
0
25
-2
6
Perri
ni
tes
sh
ale
2
30
27
-3
3
Co
ng
lo
mer
at
e an
d
40
0
vo
lc
ani
c
ro
ck
s
To
ta
l
35
30
Tab
le
3
5.
Se
qu
en
ce
a
t
La
s D
el
ic
ia
s,
No
rth
Me
xi
co
.
Di
fun
ta
, N
ew
el
l,
19
57
Be
d
Thi
ckn
es
s
in
Me
tr
es
59
-6
4
28
0
57
-5
8
11
0
44
-5
6
30
0
43
1
20
1-
42
1
64
0+
24
50
Te
nt
at
ive
S
ta
ge
?D
jul
fi
an
7oju
lfian
?P
unj
abi
an
?K
az
ani
an
-K
ung
uri
an
Ba
ig
en
dzi
ni
an
� ·;::
�
.... e; · ::s til
�
.....
N
w

1 24 Biochronology for the Marine Permian
from Burma. A few species have Baigendzinian affinities such as Costiferina spiralis (Waagen) found also in the Amb Formation of Pakistan, and a dictyoclostid close to Reticulatia callytharrensis (Prendergast), chiefly from the Sakmarian and Baigendzinian beds of the Carnarvon Basin, Western Australia. Both Ruzencev ( 1956) and Glenister & Furnish ( 196 1) preferred a Baigendzinian age on the basis of ammonoids Agathiceras, Stenopronorites, Gaetanoceras timorensis (Haniel), etc, but Ruzencev did allow that the beds might be Kungurian. As noted by Waterhouse ( 1 970a), the only ammonoid to firmly point to a Baigendzinian age appeared to be Propinacoceras, a genus also known from Kungurian-Kazanian at Kamloops, British Columbia, and from the Kazanian of Sicily, according to Dr W.W. Nassichuk, pers. comm. Related faunas have been recorded from nearby Letti (Broili, 1 9 1 5), including Rugaria strophomenoides (Waagen) of the lower Wargal beds in Pakistan. A similar fauna at Hato Dame, Portuguese Timor (Shimizu, 1 966), includes so-c�ed 'Plicatifera minor (Schell.)' obviously close to 'Neoplicatifera' or ?Overtoniina huangi (Ustritsky) of the Chinese Kungurian. A few other species, including Stepanoviella, were found at nearby localities. The overall age was discussed in relationship to other south-east Asian fauna by Waterhouse ( 1973a, p. 194), who concluded that the faunas were most likely to be Filippovian or Nevolin.
A ustralia
The early mid-Permian sequence is more complete in the Carnarvon Basin of Western Australia (Fig. 20), where Fauna D2 of Dickins ( 1 963) in the upper Byro Group (Table 29) records the disappearance of many genera, especially those indicative of warmer waters (Waterhouse, 1970c). Brachiopods include Taeniothaerus, and Pseudosyrinx, as discussed in Thomas ( 1958, 1 967), Coleman ( 1957), Campbell ( 1965) and Waterhouse ( 1970d), but complete descriptions are not yet available, although it has been possible for the writer to examine faunas kept at the Bureau of Mineral Resources, Canberra. The faunal drop in diversity is most noticeable in the Baker (=Nalbia) Formation and Norton Greywacke. Diversity then increases again in the overlying Coolkilya Greywacke (Dickins, 1 963). On the basis of bivalve studies these formations are considered Kungurian (Dickins, 1956), and the general faunal relationships and succession suggest that the Baker and Norton are possibly Filippovian and the Coolkilya with its more diverse fauna is Nevolin. Spiriferella australasica (Eth.) is allied to Kungurian species from the Arctic, such as S. polaris (Wiman) and. other forms. A highly plicate Cancrinella from the Nalbia (=Norton) beds resembles C. magniplica Campbell ( 1953) from Nevolin beds of Queensland and New Zealand. Amongst species present in both Baker and Coolkilya faunas, Undulomya suggests correlation with Malbina A of Tasmania, and Atomodesma exaratum Beyrich is shared with Fauna 1 1 1b (Filippovian) of Queensland. In reviewing the few ammonoids found in the extensive Coolkilya Formation as proposed by Teichert, prior to a precise definition by Condon ( 1 967), Glenister & Furnish ( 1961 ) assigned the upper Coolkilya to the Wordian. Propinacoceras australe Teichert is present, member of a genus regarded as key to the Baigendzinian in the Urals, but also occurring in 'Wordian' strata elsewhere (Dr W.W. Nassichuk, Geological Survey of Canada, pers. comm.). Paragastrioceras is found also in the Coolkilya Formation. It is typical of Baigendzinian or earlier faunas in Siberia and Canada. Glenister & Furnish referred the lower 100 ft. of the Coolkilya (�Bak�r, see Dic�ins, 1 963) to the Baigendzinian Stage. Helicoprion is present, found wtdely m Kungunan beds of Russia and North America. In fact the ammonoids show a very 'Yide scatter of ages, leaving one free to choose what seems the most significant. N ass tchuk et al . ( 1 9 6 5 ) pointed to s ignific ant s imi laritie s between 'Pseudogast�ioceras' goochi (Teichert) (now assigned to Daubichites - pers. comm., Dr W.W. Nasstchuk) and Daubichites fortieri (Harker) of the Assistance Formation,

Wo
rld
Sub
sta
qe
Ya
rr
ol
Ba
sin
G
ympie
B
asi
n
No
rth
Oqb
inan
?
Ved
ian
Up
per
Bow
en
Ba
isa
lian
C
oal
Me
asur
es
ur
ush
ten
ian
?Ta
ma
ree
Fm
Chhid
ruan
So
ut
h C
urra
L
imes
to
ne
llo
?Ha
vil
ah F
aun
a
Ka
laba
gh
g "' tJ>
Sosn
ov
ian
-§ P
el
ican
Cr
eek
Ill -�
Faun
a
Ka
sim
ov
ian
Ql Sc
ot
tv
il
le
.<: 1: Ql
Fa
una
..
... Ill U
fim
E
xmo
or
Fa
una
Ire
nian
E
lkin
Ne
vo
lin
II
IC
.
Fil
ipp
ov
ian
§' G
len
do
o
IIIB
0 �
Memb
er
Ql Kr
asn
ou
fimi
an
..... A A
Sar
gin
ian
� Il
iA
Akt
ast
inian
OW
l C
ree
k
Vo
lca
nic
s
St
er
lita
mak
ian
Ya
rr
ol Flnl
?So
uth
cu
rra
T
iver
to
n
Lim
est
one
Sub
gr
oup
Ta
stub
ian
Rammu
tt
Fin
L
izz
ie C
ree
k
Vol
can
ics
Ku
rmaia
n
Bur
net
t F
tm
Yo
ulamb
ie
Con
glo
m.?
Usk
ali
kian
Sur
en
an
Ran
ds
Fin?
Tab
e
36
. T
enta
tiv
e co
rr
ela
tio
ns
for
( Q
Fin =
F
orma
ti
on,
Ss
•
Sand
ston
e.
B. o
we
n
B
a
s i
nSp
rin
qsur
e so
uth
-we
st
?Rew
an
up
per
Bla
ck
wa
ter
Bla
ck A
lle
y
Sha
le
Peaw
add
y F
in
Ing
elar
a S
ha
le
Fr
ei
tag
Fm
Ald
ebar
an
Fm
·sir
ius
Sha
le
Sta
irca
se
Ss
St
anle
igh
Fin
Or
ion S
ha
le
?
Jo
e J
oe F
in
--
sou
th-e
ast
?Rew
an
Bar
alab
a C
oal
Mea
sure
s
Gy
ran
da
Fin
--
--
--
--
--
-
Fla
t T
op Fm
Ba
rfi
eld
Fm
?Ox
tr
ack
Fm
?Bu
ff
ell
Fin
Camb
oon
An
desi
te
?
� � � IS·
::s
Cl:l
�
- tv
U'l

1 26 Biochronology for the Marine Permian
Canada, indicative of a 'Road Canyon' or Filippovian correlation. This is confirmed by the bivalves- and brachiopods, and stratigraphic position.
The Lightjack Member of the Lower Liveringa beds of the Fitzroy Basin, Western Australia, shares fifteen species with the Coolkilya and only four with the Baker, though Dickins ( 1 963) preferred 1- Baker correlation. Daubichites goochi is also- found in .the Lightjack Member. 'Propinacoceras' has been reported from nearby, but may have come from the underlying Nooncanbah Formation.
In Queensland, Fauna 1 1 1b of Dickins ( 1 964; Runnegar, 1 969a) is tentatively considered to be Filippovian, on the basis of stratigraphic position, and indications of cold water, which would .explain the wide-spread reduction in faunal diversity observed in Asia, the Arctic, and Western Australia. The fauna is best represented in the Glendoo Member of the Gebbie Formation of the Bowen Basin (Table 36). The Eurydesmatid bivalve Glendella Runnegar is present, with a few brachiopods including Wyndhamia dalwoodensis Booker and Aperispirifer wair'akiensis (Waterhouse). A . wairakiensis is closely related to shells from Kungurian faunas of New Zealand, Timor, and Burma, and is close to the Arctic species Neospirifer striatoplicatus (Gobbett). Fauna 1 1 1c of Queensland, also incompletely described, may be equivalent to Irenian faunas of Russia. It possibly includes part of the lngelara fauna of Campbell ( 1 953), in which the species from the sandstone are perhaps ?Filippovian and the species from the overlying fine calcareous argillite may be Nevolin or younger Irenian.
A few occurrences of the ammonoid Neocrimites ar� recorded by Armstrong et a[. ( 1 967), regarded by them as Baigendzinian, but age and faunal relationships remain to be clarified. The use by these authors and Runnegar ( 1 969b) of a mere 3 or 4 subdivisions for the entire Permian of eastern Australia provides only an extremely crude correlation.
Long studied faunas and rocks of the Hunter Valley, New South Wales, suggest a fairly complete Kungurian sequence (Table 40). Basal Kungurian, probably Filippovian, faunas are found in the Elderslie Sandstone, now called the Cessnock Sandstone at the base of the 'Upper Marine' or Branxton Group (see Packham et al. 1 969). These and subsequent faunas are assessed chiefly from collectipns by the author and from collections at the Bureau of Mineral Resources, Canberra, as listed in Waterhouse (1 970d). Significant species include Wyndhamia dalwoodensis Booker, Aperispirifer wairakiensis (Wat.), and Echinalosia maxwelli (Wat.), all indicative of a Kungurian age, with Eurydesma. In opposition, Dickins ( 1968) and Runnegar ( 1 969b) considered these Kungurian faunas to be of the same general age and faunal unit as the underlying Farley (Sterlitamakian) faunas, based chiefly on the ·occurrence of a few bivalves, such as Eurydesma. In my opinion the brachiopods differ considerably and consistently, and should be granted prime value in assessing correlation and differentiation. Neocrimites meridionalis Teichert & Fletcher from the Elderslie Formation did not indicate any precise age, according to Glenister & Furnish ( 1 96 1 , p.689).
The over.lying Fenestella Shale in the Hunter Valley contains a distinctive Neospirifer found also in an Irenian faunal unit in New Zealand, believed to be possibly Ufimian in age. In the south Sydney Basin the Wandrawandrian Siltstone at Ulladulla (see Campbell in Packham et al. 1 969, p.362; Dickins et al. 1 969) or 'Ulladulla mudstone' contains good indices for an Irenian age in Echinalosia maxwelli (Wat.), Anidanthus so/ita Wat. and Terrakea concavum Wat. with Sulciplica. and members of the Licharewiinid suite, that by comparison with the Pai Hoi and New Zealand Permian suggest a Nevolin age. Other Kungurian faunas are likely to be discovered when the sequence is studied palaeontologically.
In Tasmania (Table 40), the upper Grange Mudstone has Eurydesma as in the Cessnock Sandsto?e of New South Wales, and may be Filippovian in age. Overlying faunas from Malbma A and B beds, as well as the Dabool Sandstone and Mistletoe Sandstone, all units proposed by Banks (see 1962), have Irenian faunas,
'assessed by the

Kazanian Stage 1 27
writer from collections either kept at the University of Tasmania, Hobart, or made by the writer. Species include Echinalosia maxwelli (Wat.), Wyndhamia dalwoodensis Booker, Terrakea concavum Wat., etc. Licharewia clarkei (Koninck) enters Malbina A and Dabool beds, suggesting direct correlation with the Nevolin faunas of northern Russia, which also contain Licharewia (Likharev, 1 966).
New Zealand
The Kungurian Stage is well represented in the foothills of the Takitimu Mountains, New Zealand (Fig. 23), although there are hazards in assessing faunas so remote from the northern hemisphere (Table 3 7). As a consequence we have to rely heavily on stratigraphic sequence and general indications of faunal diversification or impoverishment akin to those of the northern hemisphere, discussed as 'Faunal Signature' in Chapter 1 (see Fig. 7). Even so, some Productid genera ( Wyndhamia, Echinalosia, Terrakea) and some spiriferid genera (Licharewia, Spiriferella, ?Aperispirifer, and Tomiopsis) are closely allied to species in the Canadian and Siberian Arctic. A small fauna in the uppermost Takitimu Group of southern New Zealand may be of Filippovian age (Waterhouse, 1 969b) with Terrakea dickinsi Dear and Notospirifer, unless it is infaulted (Mutch, 1 973). Overlying beds include thick and large tilloidal breccia conglomerate. A moderately rich fauna at the base of the overlying Productus Creek Group is comprised chiefly of brachiopods with Spiriferella supplanta Wat. allied to S. keilhavii (von Buch) of Arctic Kungurian faunas. A number of species are shared with east Australian faunas, especially Wyndhamia dalwoodensis (Booker), Cancrinella magniplica Campbell, Terrakea concavum Wat., Tomiopsis and Aperispirifer wairakiensis (Wat.). Licharewiriiids suggest a likely Nevolin age. The succeeding faunas, exceptionally well-preserved in favourable mudstone facies, partly as Echinalosia bioherms, were of low diversity, with a number of species shared with underlying faunas. The fauna includes species allied to northern forms (Echinalosia, Tomiopsis) and is regarded as possibly Elkin, by superposition. Succeeding collections from sandstone include Neochonetes, and also coral bioherms with a distinctive Neospirifer found also in the Fenestella Shale of the Hunter Valley. This small fauna is correlative with a more complete fauna from the Flowers Formation of the Parapara Group near Takaka in north-west Nelson (Clark et al., 1 967) (Table 40). Both are regarded as Ufimian in age. The Flowers fauna contains many Irenian species mixed with several species that flourished in Kazanian faunas of east Australia and New Zealand, thus supporting Russian evidence for placing the Ufimian Horizon with the Kazanian Stage. But, as discussed previously, it may prove easier to map the start of the Kazanian Stage at the start of the Kalinovian Substage.
KAZAN/AN STAGE
Russian Plaiform, Urals, north Russia
The Kazanian Stage is one of the long-established units of the Russian Permian, and can be recognised with relative ease by means of its brachiopods, and some extent, bivalves in high latitudes, but not so readily in the palaeotropics. Probably the conodonts, ostracods and small Foraminifera will also prove useful. The type sequences lack ammonoids and Fusulinacea, and have no marine top. The succession is composed chiefly of limestones a few hundred feet thick, with a lower fauna dominated by brachiopods, and an upper fauna dominated by bivalves. Although this division is clearly due to ecological controls, well exposed and complete Kazanian faunas elsewhere in the world show a similar two-fold subdivision, as yet without any formal names. The names Kalinov and Sosnov applied to lithological and faunal divisions for the Kazanian

1 28 Biochronology for the Marine Permian
World Substages Taki timu Mts . East Nelson (and Gore , in part)
Diener ian Wairaki Breccia 200m
Griesbachian ?
Ogbinan upper Stephens Fm 500-700m
Vedian Hawtel Fm lower Stephens Fm 250 m 200m (Tilloid)
Waiua Ftm 700m
Baisalian (AG6) 200m Greville Fm 700-SOOOm
urushtenian Little Ben Ss- (AG5) 200m
Chhidruan Glendale Lmst 350m (AG4 Lmst 170m)
Kalabaghian sandstone lOOm Wooded Peak Lmst 200-1200m
sosnovian Weetwood Tuff lOOm Patuki Volcanic
Mangarewa Fm Melange 0-300m
Kalinovian 220m Dun Mountains Ultramafics 0-1500m
Ufimian Letham Fm Patuki
Elkin Irenian ?Om Volcanic Melange
Nevolin 2000- 3000m
Filippovian (conglomerate tilloid)
Krasnoufimian Takitimu Group Rai Sandstone
Sarginian 200-600m
Aktastinian Croisilles Volcanic
16 , OOOm Sterli tamakian
Tastubian Complex
Kurmaian 2000-3000m
Uskalikian
Surenan
Faunal Zone
WairakieZZa rostrata
-
Aperispirifer neZsonensis
DurviZZeoceras woodmani
Spinomartinia spinosa
PZekone ZZa muZticostata Martiniopsis woodi
Terrakea brachythaerum
EchinaZosia ovaZis
�otospirifer spinosa �eri�irillr �· �chinaZosia maxweZZi
lppirifereZZa suppZanta
ITerrakea e:anoorensis
�chinaZosia prideri
�artinia adentata
�otostrophia homeri � zeaZandicus
Atomodesma?
MourZonia impressa
-
Atomodesma Table 37 . New Zealand correlat�ons
Fm - Formation ; Lmst - Limestone ; Ss - Sandstone .

Kazanian Stage 1 29
of the Russian Platform are here proposed as substages. The Kalinov Suite has Productida dominated by Aulosteges and Stepanoviella as significant genera and its Spiriferida are characterised by members of the Licharewiinae, such as Licharewia and Permospirifer (Slusareva, 1 960; Grigorieva, 1 962). The overlying Sosnov Suite is dominated by bivalve faunas including Procrassatella, Stutchburia (Netschajewia), and Pseudomonotis, with brachiopods Cancrinella, Aulosteges and Cleiothyridina. It may be noted that the nomenclature needs clarification and modification, because a thin intervening Godrohimiches Suite is present between the two, barren, and not faunally significant. In my view it should be treated as a member of one or other horizon after study of the microfossils. The correlative Yardren dolomite has Productacea in common with both suites, but is close to the upper or Sosnovian Substage in lacking Spiriferida. In other basins, as in that of the Mezen and Kulor Rivers, Blasispirifer blasii (Vern.) is present in the Sosnov beds. The same faunal subdivisions persist north into Timan, where the lower Kazanian faunas are found in the Chevyu Horizon and overlying upper Kazanian faunas in the Vesliyan Horizon.
Faunal descriptions of brachiopods are provided especially by N echaev ( 1 9 1 1 ). A so-called 'Zechstein' fauna recorded by Mirchink ( 1 938) from Pinega River is clearly Kazanian. Other somewhat incomplete faunas are recorded from northern Russia by Likharev ( 1 93 1 ) at Vologrod and Archangel, and from west pre-Timan by Plotnikov & Molin ( 1 969).
Northern Siberia, Caucasus, Crimea, Afghanistan
The same Kazanian suite of brachiopods extends across the Arctic in Siberia and North America. Several such faunules were recorded from the Barents and Sedov Series of Novaya Zemlya, as at Russian Harbour ar:td Borzov Bay (Likharev & Einor, 1 939). In Taimyr (Table 1 5), the Baikur Horizon of ustritsky & Chernyak ( 1963) has somewhat the same faunas in two levels, as in other Kazanian faunas. However socalled Neospirifer bambadhuriensis (Diener) and Brachythyrina suggest a younger, Kalabaghian age for the upper zone (Waterhouse, 1969c), which lacks a number of species present in the lower fauna, notably Attenuatella stringocephaloides (Likharev & Chernyshev). Moreover the lower fauna has Chonetina, not Neochonetes, which is otherwise common in lower Kazanian faunas. U stritsky ( 1971 ) referred the lower fauna to the 'upper Ufimian Stage', and the upper to the lower Kazanian, but his Late and even Middle Permian are so compressed, as discussed previously, that his correlations must be set aside. Earlier, Ustritsky & Chernyak ( 1 963) had assigned the lower beds to the Kungurian Stage.
Allied faunas occur in the Barain Series of the Verchoyan Massif east of the Lena River (Likharev, 1966, p.349), in the lower Haraulakh Suite ( l ,000- l ,900 m thick) in the north, and upper Endibal Series or Delenjin Suite ( 600-700 m thick) to the west. The latter has yielded the ammonoid Spirolegoceras, compared by Nassichuk et al. ( 1 965) with a Filippovian species from the Canadian Arctic, but it must be considered that the ammonoid was long-lived, for the upper Endibal faunas are clearly post-Kungurian. The Menkechen Suite (900- 1 , 1 00 m) of east Verchoyan has Licharewia, and the overlying Cham bin Suite has Cancrinelloides, suggestive at least in a general way of the Kalinov and Sosnov subdivisions of the Kazanian in the Russian Platform. Unfortunately the faunas are rather small, and no more precise correlation is yet possible.
A fine fauna occurs to the east in the Omolon Horizon of the Kolyma River and Omolon Plateau region (Zavodowsky, 1970, p. 1 9), including the Licharewia complex, smooth chonetid (Lissochonetes) and Attenuatella as in the lower Baikur beds of Taimyr Peninsula. The overlying Gijigin Suite is of considerable interest, because it includes a tillite according to Ustritsky ( 1 973). The accompanying fauna is very small, Zavodowsky ( 1 970) recording Tornquistia and Licharewia, which suggest a Kazanian

1 30 Biochron9logy for the Marine Permian
World
Stage
Dorashamian
Djulfian
Punj abian
Kazan ian
Kungurian
Baigendzinian
or
Aktastinian
Sakmarian
Asselian
Late
Carboni ferous
Standard Dacht-e.-Nawar Wardak
Substage and Tezak
- ? -
Pamiran
Kalabaghian A Zatoconcha vampyra Marginifera typica beds beds
Perrrrundaria sisophonensis beds
Irenian Kubergandian (NeoschJ.Jagerina)
Filippovian Parapronori tes (Canoe Z Una)
"Artinskian"
Sterlitamakian Doni Yarchi Series Taeniothaerus permi�tus horizon
?Tastubian Bini Darzak Horizon
Stepanovie Z Za umariensis
Tomiopsis sp
Bokan Series Horizon with Kurmaian
Cancrine Z Za Zyoni Eurydesma mytiZoides
Asyri�
Table 3 8 . Upper Palaeozoic sequence o f Afghanistan , after Termier � �' 1974 ( see Fig . 4) .

Kazanian Stage 1 3 1
age. But a Kalabaghian age seems possible, as discussed above for the upper Baikur Horizon.
Similar Kazanian faunas extend into east and west Baikal at Lake Chironskogo (Maslennikov, 1 960; Kotlyar & Popeko, 1 967, p. l O) and are represented in the Haranor Suite ( 1 ,600- 1 ,700 m thick) of the Borzin region, with Licharewia and other forms (Likharev, 1 966).
In the Murgab Stage of the Greater Caucasus, the Gutan (8-28 m) and Nikitin (7-30 m) Horizons include Tylop/ecta yangtzeensis (Chao), Leptodus nobilis (Waagen) and other species, with Palaeofusulina nana Likh. and Reiche/ina cribroseptata Erk. To the south correlative beds contain Neoschwagerina margaritae (Deprat), and Nankinella orienta/is Mikl.-Makl. These beds are equivalent to brachiopod Horizon A of Likharev ( 1937). Brachiopod species belong to Derbyia, Meekella, Scacchinella, (Waagen), marginiferids, with Chonetella nasuta Waagen, Linoproductus lineatus (Waagen) and Leptodus nobilis (Waagen) found also in the Wargal Formation, Salt Range, and Ty/op/ecta yangtzeensis (Chao) from the Maokou beds of China. A related fauna from the Crimea Peninsula has many of the same species, but also has Cancel/ina, perhaps indicative of a slightly greater age.
A large fauna has been described as lower Murgabian by Termier et a/., ( 1 974) from Afghanistan south of Kabul (Table 38) in bioclastic carbonates and shaly limestone, called the Permundaria sisophonensis beds. Numerous brachiopods include Rugaria simulata (Reed), (Amb or Wargal), species of Waagenoconcha, Permundaria, Kutorginella, Chonetella nasuta Waagen, Costiferina redacta (Reed), Paeckelmanella (=Pterospirifer?), Purdonella lunwalensis Reed, spiriferellids and Martinia sp., many also found in the Wargal Formation of the Salt Range. Correlation with the lower Wargal appears acceptable. None of the typical Arctic Kazanian species are present, underlining the need for clarification of which brachiopod species typified warm-water tropical faunas of the Kazanian Stage. For the Afghanistan fauna the age appears confirmed by stratigraphic position over Kungurian Fusulinacea, and below Punjabian brachiopods. Nearby in the Tazak section, Sosio ammonoids have been discovered (Termier et a/., 1 974, p. l 14), but these seem to be late Kungurian.
China, Japan, south-east Asia
The Neoschwagerina Zone in China (Table 1 8) is the middle of three Fusulinacean zones in the Maokou Limestone, above the Nankinella-Cancellina ·zone, and below the Yabeina Zone (Sheng, 1 964). Associated Fusulinacean genera include Verbeekina, Pseudodoliolina, Sumatrina, Chusenella, and Parafusu/ina. Cancel/ina and Yangchiena are no longer present. Sheng ( 1 964) matched the fauna with that · of thelower Nikitin Formation of the Rek Basin in the northern or Greater Caucasus. A few brachiopods have been recorded from the zone by Huang ( 1 932a), including species of Haydenel/a and Marginifera as in Sisophon member B of Cambodia (see below).
As noted previously, Chao ( 1 966) recorded an ammonoid fauna with possibly Daubichites, Waagenoceras and Mexicoceras from the carbonaceous shale in the lower Tingchiashan Formation of western Chekiang and north-east Kiangsi, indicative of the Waagenoceras ammonoid Zone and correlated with the Neoschwagerina Zone. This is probably of Kazanian age, although the close comparison between one ammonoid Daubichites and a Canadian Arctic species of Daubichites (Nassichuk et a/., 1965) suggests the possibility of a Kungurian age. However Dr W.W. Nassichuk pers. comm. considers · the Chinese 'Daubichites' could be A ltudoceras. The other genera Waagenoceras and Mexicoceras occur in the Word Limestone no. 2, of Nevolin age in the Glass Mountains, Texas, as well as in higher Kazanian faunas of the Word in Texas.
Reed ( 1927a) described several collections from green shales and fine sandstone in Yunnan, southeast China, of which one at Tai-li-shao (see Fig. 1 7) is. large enough to tentatively assign to the Kazanian Stage or perhaps Kalabaghian Substage, on the

1 32 Biochronology for the Marine Permian
presence of Neochonetes, Stepanoviella, an? Martiniopsis _talishaoensi� Reed. R.eed
( 1 927a, p. 164) recorded Elivina tibetana (Dtener) nearby wtth Stepanovzella, posstbly suggestive of a Kalabaghian age, if Elivina is correctly identified. A few other species from the Tai-li-shao fauna also suggest a Kalabagh age, such as Brachythyrina, Martinia semiplana Waagen and Tschernyschewia, whereas Spirifer peregrinus Reed looks like Sulciplica, a genus widespread in the Kazanian and Kungurian Stages.
The upper Jisu Honguer faunas of Mongolia could be of Kazanian age, as discussed later.
In Japan (Tables 20, 32), the Neoschwagerina margaritae Zone of the Akasaka Stage, typified by massive light and dark grey limestone, is regarded as Kazanian. The key species is locally replaced by Neoschwagerina douvillei Ozawa, and is underlain by the Neoschwagerina craticulifera Zone, also classed in the Akasaka Stage. A few brachiopods in sandstones of the Tenjinnoki Formation, 1 50 m thick, in the southern Kitakami Mountains (Ueda, 1963, p. 1 50) are accompanied by Monodiexodina matsubaisha (Fujimoto), and regarded as correlative with the Neoschwagerina fauna.
In Sisophon member B of Ishii et a!. ( 1 969) in western Cambodia, brachiopods of the Tyloplecta nankinensis Zone are somewhat like those of the Kungurian member A (Table 19). They include Chonetinella, Costiferina cf indica (Waagen), Tyloplecta nankinensis
' (Frech), Leptodus, Choristites and a distinctive dictyoclostid ?Chaoiella margaritatus (Mansuy), with Marginifera and Haydenella related to Maokou species of China. Fusulinacea seem to form two zones or communities, the lower beds with Nankinella, Schubertella, and Chusenella species, the upper with Neoschwagerina douvillei Ozawa, N. aff. margaritae Deprat and Yabeina asiatica Ishii. Yabeina asiatica is generally considered to be a primitive Yabeina, descended from Neoschwagerina douvillei according to Ishii ( 1 966, p. 1 32). It perhaps indicates a late Kazanian or early Kalabaghian age. From the generalised column in Deprat ( 1 9 13 , p.76) as repeated in Table 42 herein, level 3 with Spiriferella and Licharewinid and level 4 bearing Fusulinacea are likely to be Kungurian. Level 5 at Lang-nac has Neoschwagerina margaritae Deprat and N. megasphaerica Deprat, of Kazanian age, conceivably high Kazanian as N. craticulifera normally precedes this zone. Level 7 at Pong Oua has Kazanian-Kalabaghian species of Orthotetes, Leptodus, Linoproductus, and Neoschwagerina margaritae and Yabeina cf. globosa (Yabe), which is also found in the overlying horizon. Other brachiopods described by Mansuy ( 1 9 1 2) from Phom-TaKreem and Phnon-Ta-Maio may be roughly Kazanian in age. It is not clear whether the faunas with Y. globosa are late Kazanian or early Kalabaghian, but Japanese workers regard Y. globosa as indicative of their lower Kuman (Kalabaghian-Chhidruan) Stage. Unfortunately there are few accompanying brachiopod or ammonoid faunules to test various possible correlations, and at present we are faced with several alternatives : 1 . that Neoschwagerina craticulifera is Kungurian (as seems likely in Iran and
Pamirs), and N. margaritae and even Y. globosa are Kazanian ; 2. that N. craticulifera and N. margaritae are Kazanian and Y. globosa is early
Kalabaghian ;3 . that N. craticulifera and N. margaritae are Kazanian, and Y. globosa forms a third
zone between Kazanian and Kalabaghian ;4. that Y. globosa is insignificant, and overlaps with either Yabeina or
Neoschwagerina to indicate a Kalabaghian or Kazanian age.Similar problems concern the age of Yabeina asiatica. In the present account, high
value is given to Japanese studies on the Fusulinacea, which would suggest that Yabeina globosa is Kalabaghian, and Neoschwagerina margaritae is Kazanian. Yabeina a�iatica is tentative�y r�garded as Kazanian, but the relationship between the late Kazaman and Kalabaghtan IS under poor control, and there might well be a zone not as yet well delineated, typified in part by Yabeina asiatica. Neoschwagerina craticu/ifera or shells s�-called, tho�gh �egarde? as typically lower Kazanian, also appears in faunas of Kungunan age. Thts bemg so, tt may be that primitive Yabeina (=asiatica), and allies

Ch
i-
Th
ua
n
19
61
Be
d
5 s
il
ic
eo
us
b
la
ck
c
ar
bo
na
te
sB
ed
4
si
li
ce
ou
s
ca
rb
on
at
es
Be
d
3 r
ed
s
ha
le
, n
ot
f
os
si
li
fe
ro
us
B
ed
2
r
ed
c
ar
bo
na
te
s,
w
it
h
Yabei
na·Le
pido
lina
B
ed
1
c
ri
no
id
al
l
im
es
to
ne
, g
re
y
ma
rl
.
Is
hi
i
et a
l. 1
96
9
Me
mbe
r
D m
ud
dy
l
im
es
to
ne
an
d
li
me
st
on
e
Me
mbe
r
C
re
d
=
be
d
5-7
of
S
aur
in
. r
ed
n
od
ul
ar
m
ud
st
on
e.
re
d
sh
al
e.
Yabei
na�
Neos
ahwag
erin
a
Me
mbe
r
B
cr
in
oi
da
l
li
me
st
on
e
Me
mbe
r
A t
uf
f
Tab
le
3
9.
Co
mp
ar
is
on
o
f
Si
so
ph
on
m
id
-P
er
mi
an
s
eq
ue
nc
es
,
fr
om
W
at
er
ho
us
e
(19
73
a,
T
abl
e
27
).
� t: :::s i:i'
:::s C'.!
� � .....
w
w

1 34 Biochronology for the Marine Permian
typify the later Kazanian. Such speculation underlies the need for .detaile� analysis ofpalaeotropical brachiopods, for which there are few modern studtes avadable.
Large brachiopod faunas from Sisiphon and especially Phnom Tup are described by Chi-Thuan ( 1 96 1 ) with ?Chusene/la afT. tunetana (Douville), and Yabeina johannis Saurin (Table 19). Overlying beds contain chiefly Lepidolina multiseptata Deprat and Yabeina afT. khmeriana Saurin. As shown by Waterhouse ( 1 973a, p. 1 96), there are difficulties in reconciling Chi-Thuan's account of the stratigraphy with that outlined by Ishii et al. ( 1 969). Chi-Thuan's fauna appears to have come from the upper part of the B member, and so may be late Kazanian or early Kalabaghian age. I prefer the latter. ChiThuan herself stated that the faunas came from two horizons, but rnade no separation of localities in the text; she further stated that the faunas came from the Yabeina Zone below Lepidolina multiseptata. Many of her species are recorded in Ishii et a/. ( 1969) in Sisophon member C, which would agree with many of the known species ranges elsewhere. Yet Costalosia, Waagenites, Unoinella, and Spiriferella cambodgensis point to a Kungurian correlation, whereas Brachythyris, Tyloplecta, and Monticulifera are perhaps younger Kazanian. Still others of the species identified, perhaps incorrectly, by Chi-Thuan, suggest a Kalabaghian age by correlation with the faunas of Basleo, Himalayas, and Salt Range. The problem is insoluble at present, but it appears likely, if we can accept the identifications, which are frequently dubious and not supported by illustrations, that the collections have unwittingly crossed two or even three condensed zories, of Kungurian, Kazanian and even Kalabaghian age.
Pakistan, Iran, Mediterranean
The middle part of the Wargal Formation of the Salt Range, Pakistan (Table 28), has a fauna distinguished by a number of species of Enteletes, Orthis, Streptorhynchus, Derbyia, chonetids, Echinalosia indica (Waagen), Choristitella wynnei (Waagen), 'Spirifer' oldhamianus Waagen, Martinia semiplana Waagen and a few other forms, according to W aagen ( 1 89 1 , p. 1 1 1 , 200). Some ranges have to be amended, especially on the basis of the work by Reed ( 1944), but there appear to be Kazanian faunas at this position, below the Kalabagh Member at the top of the Wargal Formation. The fauna is richer than the lower Wargal faunas, thought to be of Kungurian age, and less diverse, but with fewer cool-water elements than in the Kalabaghian faunas. There are virtually none of the species and genera that characterise Arctic or Australian faunas, so that the stratigraphic limits of the fauna await detailed study. No Fusulinacea or significant Ammonoidea are found.
A Permian fauna from south Iran described by Douglas ( 1 936, 1950) includes Neoschwagerina craticulifera (Schwager), Cancellina primigena (Hayden), Verbeekina verbeeki (Gein.) below a fauna with Yabeina inouyei Deprat and above a fauna with Polydiexodina (=Skinnerina?), but the fauna could be as old as Nevolin, in having the brachiopods Lissochonetes and Mentzelia.
Part of what promises to be a very rich fauna has been described or listed chiefly as new Aulostegid genera, Leptodus, 0/dhamina, Derbyia, and Orthotetes, from Hydra in the Aegean Sea by Grant ( I 972). Neoschwagerina craticulifera and Verbeekina verbeeki are known widely in this area.
A huge fauna has been recorded from the Sosio beds of Sicily. by Gemmellaro ( 1 899), Greco ( I 942, 194 7), Gregorio ( 1 930) and Rudwick & Cowen ( 1968) with the ammonoid Waagenoceras, and Neoschwagerinid Fusulinacea regarded as keys to palaeotropical equivalents of the Kazanian Stage. The writer has examined little of the actual material, and so is not sure if the Kazanian age is completely correct. There are many species of Enteletes, and Martinia, with Scacchinella, Streptorhynchus, Leptotid, richthofeniids, spiriferinids, a few choristitinid and syringothyrinid forms, and many terebratulids, such as Heteralasma, Jisuina and Notothyris. The fauna is in general

Kazanian Stage 1 35
rather like those of the Sakmarian Tr.ogkofel Formation of Austria, but also resembles the Kazanian 'Horizon A' brachiopods from the north Caucasus (Likharev, 1 932), which probably occupied a similar palaeotropical station. But a high Kungurian age cannot yet be ruled out.
Spitsbergen, Greenland, North America The Brachiopod Chert of Isfjorden and Hornsund, Spitsbergen (Table 25) has
yielded large brachiopod faunules that share many species with Kazanian faunas of Russia and Siberia, such as Cancrinelloides and Licharewia. The lists are summarised from various authors by Gobbett ( 1 964). However, it must be noted that stratigraphic details are not well controlled, and it appears likely that Nevolin (Irenian) faunas, though not described from any clearly distinguished horizon, may well be present, and have been intermixed with genuine Kazanian horizons. Such forms as Chonetina superba Gobbett, and Pterospirifer cordieri (Robert) suggest the likelihood of Nevolin equivalents. This seems to be counter-balanced by the report of Licharewia (Gobbett, ,1 9.64), but we do not know if this is an early appearance of the genus, as in Nevolin faunas of Timan and New Zealand, or due to mixture of Nevolin and Kazanian faunas. The Ufimian and Elkin Horizons, requiring careful field collecting and subtle faunal analyses, have not been recognised but may well be present. This failure to distinguish the Nevolin level may have led to misleadingly high faunal diversities.
Brachiopods briefly described and listed from Horizons G and H from Amdrupsland, north Greenland by Frebold ( 1950) may be of Kazanian age, but could also be Kungurian. So few species are figured, and so many assignments are dubious, that it is difficult to tell.
Kazanian faunas with Cancrinelloides and Licharewia are wiaespread through the Canadian Arctic Islands (Fig. 27), in the Trold Fiord green sandstones, and the coarsely granular limestones of the Degerbols Formation, as well as in sandstones of the Richardson Mountains, and cherts and limestones of the upper Tahkandit Formation (Table 22) in northern Yukon Territory (Waterhouse, 1 969a; Bamber & Waterhouse, 197 1). Only one zone is present, presumably of lower Kazanian or Kalinovian age. Nassichuk et a/. ( 1965) described Neogeoceras macnairi from the Trold Fiord Formation and assigned a general Guadalupian (=basal Kungurian to Urushtenian) age. The same typically Kazanian fauna is found in the upper Tahkandit Formation of Alaska (Brabb & Grant, 197 1). To the south allied faunas occur in the Fantasque Chert of British Columbia and the upper Range Canyon Chert in Alberta as reviewed by Logan & McGugan (1968).
In the Phosphoria Rock Complex of Idaho, Montana, Wyoming, and Utah (Table 34) the Tosi Chert Member has Lissochonetes, Sphenosteges, Echinauris and Leptodusof apparent Kazanian age, though thought to be possibly younger by Y ochelson ( 1 968). Several species are shared with the Ervay Carbonate Rock Member at the top of the nearby Park City Group. Conceivably Lissochonetes implies a late Kazanian age. In northern Utah, the topmost unit of the Park City Group consists of the Gerster Formation (the Kaibab lies at the base), with a modest brachiopod faunula, including Sphenosteges hispidus (Girty), Waagenoconcha montpelierensis (Girty), Bathymyonia nevadensis (Meek), Muirwoodia multistriata (Meek) and Neospirifer pseudocameratus (Girty). The fauna was correlated by Dr G.A. Cooper with the Appel Ranch Member at the top of the Word, that is, late Kazanian or Sosnovian (Bissell, 1 973, p.324). The Gerster in places overlies the Rex Chert, which is here regarded as Elkin in age, to imply that the Ufimian and lower Kazanian are missing.
Brachiopods· from the Word and equivalents of the Glass Mountains, Texas (Table 26), have been described by R.E. King ( 193 1), Muir-Wood & Cooper ( 1 960) and Cooper & Grant ( 1 969). Horizons include the Appel Ranch and Willis Ranch Limestone Members of the upper Word in the Glass Mountains (Fig. 29), and the

136 Biochronology for the Marine Permian
�------�------�2------�3 M�
.. C] Ill .. .. I52IZill � 103"15'
�o(jVo l c a n i c � Post-Word 8 PrP. -. r--""1 Goptonk L__j .Ch1no Ton k [3 Word Shale
Appel Ranch
Word (W1Ihs R . ) EJ Rood Canyon
0
() 3rd S 4th Leonard
l imestones
Cathedra I Mtn.
Hess
Dugout Member of Skinner Ranch
Skinner Ranch
Lenox H i l l s
10f10' Fig. 29. Geological map of part of the Galss Mountains, west Texas, from Cooper & Grant (1973a, fig. 25, p. 73). Reproduced with permission of the authors.

Kazanian Stage 1 3 7
Getaway and Cherry Canyon Formations of the Guadalupe Mountains and Sierra Diablo. It is impossible to fully comprehend the faunal affinities until the brachiopod descriptions are published by Cooper & Grant (in press), but large 'Grandaurispina' (=Terrakea) from locality -706e at the top of the Willis Ranch Member for example suggests correlation with the Kazanian Stage of Australia and New Zealand. The report of Spiriferinaella in the Appel Ranch Member (Cooper & Grant, 1973) probably implies an upper Kazanian age. Ammonoids are present in the Wordian by the hundreds according to Furnish { 1 973), and include species of Waagenoceras, Agathiceras, Popanoceras, Stacheoceras, Neocrimites and Propinacoceras. Apparently no detailed zonation has been achieved for ammonoids, the faunas from Nevolin to upper Kazanian, i.e. China Tank to Appel Ranch Members all being treated as one unit.
Clastics, carbonates and lavas of probably Kazanian age in the Las Delicias syncline of north Mexico (Table 35) include faunas in the dark shale of Band 5 in the Malascachas section (King et a/., 1 944, p. lO) and in shale, greywacke, and limestone of beds 14, 15 and ? 1 5 in the Difunta section (King et a!., 1 944, p. 14). Parafusulina deliciasensis Dunbar & Skinner occurs with Meekella, Neochonetes, Prorichthofenia, Leiorhynchoidea, Paranorella, Waagenoceras dieneri Bose, W. guadulupensis Girty, Epithalassoceras ruzencevi Miller & Furnish, Stacheoceras gemmellaroi Miller and other species.
The Spiriferellina Zone at the top of the Monos Formation, north Mexico (Fig. 28), is probably Kazanian in age with a brachiopod faunule described by Cooper (1953). The faunule is too small to decipher with confidence, but contains Neochonetes which might suggest an Ufimian or lower Kazanian age. Also the low interarea and shape of a strophalosiid recall Echinalosia minima (Maxwell) of Kazanian and Ufimian age in Australia and New Zealand (Dear, 197 1 ; Waterhouse & Vella, 1 965). Waagenoceras dieneri is present.
Timor, Australia, New Zealand
No well established brachiopod fauna of Kazanian age is known from Timor (Table 16, Table 44). The Waagenoceras ammonoid fauna from Tei Wei was placed in the Baigendzinian Stage by Ruzencev ( 1956) and Glenister & Furnish ( 1 96 1 ), but is probably late Kungurian or Kazanian.
Kazanian faunas are widely represented in eastern Australia where they are called Fauna IV by Dickins { 1964 ; Runnegar, 1 969b). An outstanding summary of the Queensland successions by Dear ( 197 1 , 1 972) goes far beyond the crude recognition of Fauna IV, and names four successive faunas, the Exmoor, Scottville (=Peawaddy Formation), Pelican Creek (=Streptorhynchus pelicanensis bed) and Havilah faunas in the Blenheim Subgroup of the north Bowen Basin (Fig. 23, Table 36, Table 40). From superposition and correlation with New Zealand faunas, the Exmoor fauna is here considered to be Ufimian, the next two faunas Kazanian, and the Havilah fauna may be post-Kazanian. The Kazanian age is based on superposition, reinforced by the presence of a number of brachiopod genera and species found in the Arctic, such as Wyndhamia, Terrakea, Cancrinella, etc., and members of the Licharewiinae. Allied Kazanian faunas are found in the Muree beds and Mulbring (at least lower Mulbring) Subgroup of the Hunter Valley, New South Wales, and in Malbina D-E and lower Ferntree and correlative beds of Tasmania (Table 40).
In New Zealand two brachiopod teilzones are found in the Takitimu foothills, the Echinalosia ova/is Zone, followed by the Terrakea brachythaerum Zone, both with well developed Kazanian faunas (Table 36), and many forms shared with eastern Australia, especially Queensland (Waterhouse, 1964, 1973c ). Thus the two-fold division suggests likely correlation with the Kalinovian and Sosnovian Substages of the standard Kazanian of the Russian Platform. The lower zone includes such species as Grumantia

1 3 8 Biochronology for the Marine Permian
world Substage
Griesbachian
Ogbinan
Vedian
Baisalian
urushtenian
Chhidruan
Kalabaghian
Sosnovian
Kalinovian
Ufimian
Elkin I ren
Nevolin ian
Filippovian
Krasnoufimian
Sarginian
Aktastinian
Sterlitamakian
Tastubian
Kurmaian
Uskalikian
Surenan
Ms
New Zealand Tasmania New South Wales Queensland Western Austra-Formations , Hobart , Hunter Valley Bowen & (Gympie) lia Carnarvon Nelson (up- Poatina Basins Basin per), Southlan�
?Brooweena
stephens ?Clifton SubgrOUI Blackwater Kockatea Shale
Cygnet Coal Waiua Measures
( ?Traveston)
Greville Newcastle Coal
Black Alley Measures
Little Ben Tomago (Tamaree) (Hardman Member) Coal
Wooded Peak .Flat Top & upper (Liveringa)
Lmst Fern tree Measures s . Curra lower E lsdun Group
Barfield Mangarewa Mulbring Ss
?Risdon Ss Muree Ss Peawaddy
Member Malbina D , E Wollong Ss
Member Malbina c Fenestella Silt- Catherine Ss
Letham stone Member
Malbina B , ? Ingelara
Coolkilya A - - - - Shale - - - -
Grange Ms Cessnock Ss Freitag Baker Member
Wandagee , Quinn-Berriedale Greta Coal Aldebaran anie ?Bulgadoo
Takitimu I.mst Measures Ss Cundlego Coyrie , Mallens
Nassau Sls ? Cattle Creek Wooramel Group
Group Faulkner Farley ?Reids Callytharra
Group Dome
Bundella Ms beds Lmst
Darlington Rutherford o:Lizzie Creek Beckett Shale Lmst
Volcanics (part)
Quamby Ms Allandale Burnett Lyons Group
Wynyard Tillite
Lochinvar
Seaham
Table 40 . Intercorrelations between New Ze aland and Australian Formations . mudstone ; S s - sandstone ; Sls - siltstone ; Lmst - Limestone .

Tabl
e 41
. Co
rrel
atio
n of
Quee
nsla
nd P
ermia
n ac
cord
ing
to D
ear,
197
2, T
able
1,
p. 4
. �
/ Fa
unal
Faun
al
Nort
h So
uth-
west
So
uthe
ast
Yarr
ol
Gymp
ie
Sydn
ey
Basi
n Su
bdiv
isio
n Su
bdiv
isio
n Bow
en
Bowen
Bas
in
Bowen
Ba
sin
Basi
n Di
ckin
s 19
64
' Ba
si·n
Spri
ngsu
re An
.Ba
sin
Hunt
er V
alle
y So
uth
Coas
t
Uppe
r Bow
en
Blac
k Al
ley
Gyra
nda
.. Tom
ago
Coal
Cum
berl
and
Coal
Mea
s.
Shal
e Fo
rmat
ion
Meas
. Ha
vila
h Pe
awad
dy
Tama
ree
Subg
roup
Fa
una
Form
atio
n Fl
at T
op
Form
atio
n Mu
lbri
ngGe
rrin
gong
Pe
lica
n Bl
enhe
im
Cath
erin
eFo
rmat
ion
Form
atio
n Vo
lcan
ics
Sand
ston
e Fa
una
Cree
k In
gela
ra
Berr
y Fa
una
C>.
:::1 Fo
rmat
ion
Barf
ield
Fo
rmat
ion
0 Sc
ottv
ille
"'
Sub-
Frei
tag
Form
atio
n (.!)
IV
Fa
una
Form
atio
n So
uth
Mure
e Now
ra
..
Exmoo
r Gr
oup
Uppe
r Ox
trac
k Fo
rmat
ion
Form
atio
n Al
daba
ran
:>.
Faun
a Fo
rmat
ion
....
Sand
ston
e ..
... Cu
rra
� Lo
wer
e Up
per
Wand
rawa
nd-
Faun
a Fa
una
Gebb
ie
0 Al
deba
ran
:>.
�
Branx
ton
ian
Sand
-II
I II
I Cll
Subg
roup
..
.. r::
� Sa
ndst
one
.....
0 Su
b��;ro
uo
ston
e e
g Li
me-
Siri
us
0 �
::>
Fene
stel
la
Ulla
dull
a T·i
v;ert
on
Form
atio
n r::
Shal
es
Muds
tone
0 u
ston
e Su
bgro
up
ID Fa
una
Homev
ale
Stai
rcas
e ...
.. Low
er
�
II
Faun
a �
Sand
ston
e Bu
ffel
Br
anxto
n Su
bgro
up
� Fo
rmat
ion
Gret
a Co
al
(Inc
orpo
rat-
Meas
. Co
njol
a -
ing
Lizz
ie
Lizz
ie
Stan
leig
h Ow
l Fa
rley
Fa
una
I)
Cree
k Gu
lly
RaiiDI
Iutt
Form
atio
n Ca
mboo
n Vo
lcan
-Ru
ther
ford
Fo
rmat
ion
Volc
anic
s ic
s Fo
rmat
ion
Form
atio
n an
d Ya
rrol
Al
land
ale
Form
at-
Form
atio
n io
n Ca
rmil
a Be
ds
Orio
n An
desi
te
Burn
ett
High
bury
Lo
chin
var
:>.
....
.....
Shal
e Fo
rmat
-Vo
lcan
ics
Form
atio
n e
ion
0 � s::
Seah
am B
eds
0 u s:: ::>
Faun
al
Subd
iv-
is io
n Ru
nneg
ar
1969
Faun
a
IV
Faun
a II
I
Ulladl
lla
Faun
a
Fauna
II
Alla
ndale
Fa
una
� � ;:s e;· ;:s t:l:l i:1' � ......
w
\0

1 40 Biochronology for the Marine Permian
pelicanensis (Flet.), Neochonetes beatusi Wat., Lethamia ligurritus Wat., Terrakea elongatum (Eth. & Dun), Plekonella south/andensis (Flet.), Aperispirifer lethamensisWat. and Tomiopsis mantuanensis (Campbell). The upper zone includes Wyndhamiac/arkei gattoni (Max.), Plekonella acuta Campbell, and Tomiopsis costata Wat.
PUNJAB/AN STAGE
Pakistan
The Punjabian Stage contains the youngest widespread and richly fossiliferous rocks of the Permian Period, characterised by the Yabeina-Lepidolina fusulinacean assemblage, the Timorites-Cyclolobus ammonoid suite, and an array of brachiopod species varying in different palaeolatitudes. Stepanov ( 1 973) revived the name Punjabian Stage originally proposed by Schenk et a!. ( 1 94 1 ) for this part of the column. Stepanov ( 1 973) showed the Punjabian Stage as pre-Djulfian, and partly Guadalupian. It is adopted herein, and divided into the Chhidruan and Kalabaghian Substages, as defined by Waterhouse ( 1 972a). This Punjabian Stage is the same as the Chhidruan Stage of Furnish ( 1966), and Glenister & Furnish ( 1 970), in which the initial Chhidruan Stage of Furnish & Glenister ( 1 96 1) was expanded beyond the faunal and lithostratigraphic limits of the Chhidru Formation t9 incorporate the underlying Kalabagh Member of the Warga! Formation. Here it is preferred to retain the original definition of Chhidruan as in Furnish & Glenister ( 196 1 ), and allow the substages to agree in name with their lithological units. A Chhidruan Stage as in Furnish & Glenister ( 1 970) that differs so much from its original definition, and incorporates a Chhidru Formation and Kalabagh Member can only confuse.
The Chhidruan and Kalabaghian Substages are based on exposures in the Salt Range of West Pakistan (Table 28) and as such have been assessed as Late Permian (Miller & Furnish, 1 940; Glenister & Furnish, 1 96 1 ; Schindewolf, 1 954; Ross, 1967b; Kummel & Teichert, 1 964 ; Teichert et a/., 1 973 ; Ruzencev & Sarytcheva, 1965). Waterhouse ( 1 966) and Grant ( 1 968) showed that this was not correct, as reinforced by ammonoid studies (Tozer, 1969; Chao,- 1 966; Furnish, 1 966, 1 973 ; Spinosa et a/., 1970) and fusuline studies of important sequences in China (Sheng, 1 964). It ·may well be time to abandon the Kalabaghian and Chhidruan Substages, for they have occasioned a regrettable diversion of opinion over correlation that may take years to resolve. Other than for the Salt Range sections, there is virtually no disagreement whatsoever between brachiopod and ammonoid correlations - for instance, between such important sequences as those of we.st Texas, Mexico, China, Armenia, and Timor. But because of the Salt Range names, this close measure of agreement is not apparent. If, for example, we used Gnishik and Hachik as the basis for substages, our correlation tables would agree well except for the disputed Salt Range column, which after all, is only one of many sequences. Then also we would have a number of substages and stages in sequence, and thus reduce correlation problems, and allow one country to be custodian for standard sequences of the Permian Period.
Kozur ( 1 974) has also avoided tne diihculty, by using the Guadalupian 'Stage' with Wordian and Capitanian substages as world standards. In a considerable improvement over earlier summaries (e.g. Kozur, 1973a, b), the Kalabaghian is shown as equivalent to low Capitanian, and the Chhidruan as equivalent to upper Capitanian, largely following Waterhouse ( 1 972b), but he also correlated the Chhidruan with the 'Araksian' (=Baisalian of Waterhouse, 1972b, not Araksian of Furnish, 1973), passing up to . early Do:ashamian. In an appendix, Kozur ( 1 97 4, p.50) preferred correlation at least m part With the Abadehian Substage of Taraz ( 1 973).
The upper division of the Warga! Formation, called the Kalabagh Member by

Punjabian Stage 1 4 1
Waagen ( 189 1), (see Teichert, 1 965, p. 1 2), contains a fauna that can be distinguished from the faunas above and below. Waagen ( 189 1 ) recorded several distinct species,Enteletes l�evissimus Waagen, Enteletina /atisinuatus Waagen, Megasteges dalhousei(Dav.), Terebratuloidea depressa Waagen, Spirigerella a/ata Waagen, Notothyris lenticularis Waagen, and several nautiloids. Waagen ( 189 1 , pp. 1 98-200) listed some 18 species that appeared last in the member and some 1 8 species that entered the sequence, including Derbyia hemisphaerica Waagen, Waagenoconcha purdoni (Dav.), (assigned to a new genus Septiconcha by Termier et a[., [ 197 4 ]), 'R hynchonel/a' wynnei W aagen, Uncinunellina theobaldii (Waagen), U. jabiensis (Waagen), Spirigerella hybrida Waagen, S. ovoidalis Waagen, Notothyris subvesicularis (Dav.), and Spirifere/lina multiplicata (Sow.). Most persisted into the Chhidru beds.
The fossil lists from the Wargal Limestone in Reed ( 1944) cover 80 pages and are detailed, but provide little stratigraphic information, apart from an indication of basal faunas, and topmost faunas. Intermediate faunas are seldom described, and other faunas are described as 'upper' in contrast to 'top'. Subject to this reservation, the topmost Wargal beds are very rich compared with the middle Wargal, with numerous Enteletes, no Orthotichia indica (?) or other Orthotichia except derbyi, Meekella punjabica Reed, a number of Streptorhynchus, Schuchertella, Kiangsiella, Derbyia, Waagenites, Tschernyschewia, Lyttoniids as in lower beds, restricted Megasteges da/housei (Davidson) (as in Waagen, 1 89 1), Chonetella and Marginifera as in upper beds, restricted 'Dictyoclostid' celsus Reed; Anidanthinid 'weyprechti (Toula)', ?Stepanoviella lahuseni (Likharev), S. kulikii (Fred.) and Compressoproductus mongolicus subcircularis (Reed), Wellerel/a spp. Pugnoides, ?Purdonella scopulosus (Reed), P. limitaris Reed, P. conformis Reed, Tomiopsis punjabica (Reed), and species of Martinia, Dielasmina and Notothyris. A number of coolwater genera are present, including numerous linoproductinids, Tomiopsis, Martinia, and Dielasmina, which is related to Fletcherithyris of Australia. Perhaps this cooling was contemporaneous with the Gijigin tillite of north-east Siberia.
The overlying Chhidru Formation, previously called the Upper Productus Limestone, is 90 m thick, and composed chiefly of limestone and sandy limestone (Teichert, 1965). Compared with the underlying Kalabagh Member, Chhidruan faunas have fewer Enteletes, no Streptorhynchus, Plicatoderbyia, Chonetina, Aulosteges, fewer Tschernyschewia, no Juresania, ?Chianella, Stepanoviella, Compressoproductus, Uncinunellina, Hustedia, Tomiopsis, Heterelasma or Gefonia. New genera include Sphenalosia, Tyloplecta, Fredericksia, and Cryptacanthia - new, that is, for the Salt Range, but found in older beds elsewhere. New species include 'Chonetes' squama Waagen, Strophalosia blandfordi Reed, Sphenalosia salmunensis (Reed), and Costiferina aratus (Waagen). Thus even basing an assessment of faunal studies more than 30 year� old it is possible to distinguish the two faunas.
No Fusulinacea are present in either Kalabagh or Chhidru faunas, but the significant ammonoids Xenodiscus and Cyclolobus are present although discrimination between the substages does not appear pos'sible (Furnish & Glenister, 1 970), perhaps because specimens are so few. Just how extensively these ammonoids, particularly Cyclolobus, en�ble correlation has been debated. It appears probable that Cyclolobus of this age lived in a geographically restricted area, around the margin of the present Indian Ocean, now separated as the Himalayas, Salt Range, Timor and Madagascar (Waterhouse, 1972b). Even the Himalayan correlation has now come into question from Grant & Cooper, 1973 (see Table 9), though their view is not supported by brachiopod evidence.
Himalayas
According to most published studies, Punjabian faunas are wide-spread in the Himalayas of Kashmir, Kumwaon, Nepal and southern Tibet, as reviewed and summarized by Waterhouse ( 1 972a).

142 Biochronology for the. Marine Permian
C h h idruan 1- •
• •
. . . . .
. . . .
. .
. . . . . .
•
. .
. .
. . . .
lfi11fll - - - - - - - - - .... '"',·:"": .'"'"::·=:.·"':;"":::'"'�·""::""::··:-o..:,'-'-:,""'.•·�: .'"'·:·.,.·::ul,.
. . .
Ka labagh ian
?
. . � ·� (··:�Y; .. ·..:·:\:::..·: .·:·.·. � •, '\"'l•r•::: ···�·· . . . . . . :.·:,·.·.<:;.·:::".·.� ... ·• :.-.·::.·::•":'·" :, ..... !·�: :: .
. . .
. .
.
. . :=.::i;::i.�'�!::;!�::�·:::f;:-):·;::,.:�':':..i).���"i�
.
. .
- - - - - - - - -·- lf--r--r=.r-r •,"--,r-r-r-"1 J.J.J. l l l l l Devonian T T T T TT T T
Otoceras
t
Echinalasia kaljkotei Zone
LIMESTONE
SANDSTONE
SILTSTONE
DOLOMITE
bivalve
I Lamoim�rg� himalayensis
1
li!i2il � m .
�
� r30 m
0. m
Z o ne
Fig. 30. Simplified stratigraphic column of Permian and Triassic beds in Panjang Kola, Dolpo District, north-west Nepal, from field work by the writer. Ages for the Otoceras and Lamnimargus himalayensis beds are well established, but the ages of the intervening bivalve (Pyramus) and 'Echinalosia' kalikotei faunas require further study.

Punjabian Stage 1 43
The fullest Punjabian sequences apparently occur in the Dolpo district of northwest Nepal (Fig. 1 7, 30) judged from field work and preliminary laboratory studies as yet unpublished by the writer, within the broadly defined Chini-Thu Formation of Fuchs (see Waterhouse, 1 966). Micaceous dark silty shales and quartzites of the lower part of the group contain Lamnimargus himalayensis (Diener), Marginifera typica Waagen, Costiferina a/atus Water�ouse, Anidanthusfusiformis Wat., and Fusispirifer nitiensis (Diener) and other brachiopod species. The Lamnimargus himalayensis Zone is also found in sandstone of the Lachi Group of North Sikkhim (Muir-Wood &Oakley, 1941), in the Selung Group on the Tibetan side of Mt Jomlo Lungma (Everest) (Mu Antze et a/., 1973), and the Productus Shales in the Lissar Valley (2 miles south of peak Darma no. X 1) (Diener, 1903), and the Productus Shales of Byans (Diener, 1903). Other localities in the Productus Shales lack the key species, but on the whole share more species with the Lamnimargus himalayensis Zone, than with the Echinalosia kalikotei Zone, discussed below. However the beds at the Kiunglung camping ground (Diener, 1 897b, 1 903) could be slightly younger. The zone is well represented at Spiti in the Kuling Shale, of calcareous sandstone at the base and chiefly shale. Underlying sandstone has yielded few fossils.
The brachiopods of the Zewan beds of Kashmir have been described principally by Davidson ( 1 862) and Diener ( 1899, 1 9 1 5). Diener ( 19 15) arranged the faunas into six horizons, of which the lower two have few brachiopods, and may be little older than the overlying horizons. They contain Spinomarginifera? and Leptodus, but do not share significant species with the lower Spiti sandstone. Underlying beds contain Glossopteris plants. The main fossiliferous horizons are those of Zewan 3 and 4, essentially similar to each other. The lower horizon contains Lamnimargus himalayensis and various dictyoclostid and retariid Productacea. The overlying horizon is dominated by Spiriferella rajah, but has no unique species. Zewan 5 is a minor horizon. In short, Zewan 3-5 are a complex of several communities dominated by different species at different levels, but all apparently belong to one Lamnimargus himalayensis Zone. The topmost or Zewan 6 fauna is provisionally assigned to the uppermost Permian, as discussed later. A number of species from Zewan 3-5 are shared with those of the Kalabagh Member of the Wargal Formation in the Salt Range (Diener, 1 903, 1 9 1 5 ; Waterhouse, 1 966, 1 972a).
No Fusulinacea are known, but ammonoids are found at several localities. Furnish et a/. (1973) have recorded Cyclolobus walkeri Diener from some 20 m below Otoceras beds at Guryul Ravine in the Zewan beds of Kashmir. Diener ( 1903) also recorded species of Cyclolobus and Xenaspis (=Xenodiscus) from the Kuling Shale, some 10 m below the Otoceras beds, and other specimens have been collected by a Danish expedition. All are referred to Cyclolobus walkeri Diener by Furnish & Glenister ( 1 970, p. 165), thus suggesting correlation with the Chhidruan fauna of the Salt Range, whichcontains C. cf. walkeri. Diener referred some of the species kraffti Diener and haydeni Diener from the Kuling Shales to a new subgenus or genus Krafftoceras, on the basis of a complexly ramnified siphonal saddle, but this is discounted by Furnish & Glenister (1970), and Furnish ( 1 966) synonymised Krafftoceras with Cyclolobus.
A number of tiny ammonoids are found at Lilinthi in the Productus Shales of Byans, identified by Diener ( 1903, p. l l 5) as Hyattoceras n. sp. ex aff. H. cumminsi White, Gastrioceras sp. ind. ex aff. G. marianum Vern., Lilinihiceras n. gen. sp. ind., and Nomismoceras smithii Diener. They include a form ascribed to Hyattoceras, now generally synonymised with Waagenoceras, but suggested to be possibly of Kungurian rather than Kazanian age. Lilinthiceras, with no species named, is said to be close to Triassic forms in shape and sculpture. Presumably the Byans ammonoids were essentially contemporaneous with Cyclo/obus from the Kuling Shales, as Waagenoceras is known elsewhere to occur with Timorites, in United States, and with Cyclolobus in Timor.
Amongst Permian faunas of the Himalayas, the richest comes from a limestone

1 44 Biochronology for the.Marine Permian
crag Chitichun no. l in southern Tibet, chiefly from talus at the base, thus precluding any detailed stratigraphic study (Diener, 1 897a, 1 903). The crag itself is part of the topmost nappe of the Himalayas, and is believed to have travelled southwards with enclosing ultramafics from the vicinity of the Indus suture. The outstanding fossils are Strophalosiina tibetica (Diener) and Elivina tibetana (Diener), also reported from faunas of apparent Punjabian age in south-east Asia, especially Cambodia, and Timor, but not known in the 'Tibetan' Zone or nappe of the Himalayas. Other important forms include Chaoiella chitichunensis (Diener) and 'Spirifer' wynnei Waagen. Many species were identified by Diener ( 1 903) with shells from the Sosio beds, of supposed Kazanian age, but it must be noted that internal details were seldom available in Chitichun material, so that identifications, especially generic, lack certainty. Moreover, faunas like those of the Sosio beds also occur in slightly younger faunas, believed to be of Punjabian age on the basis of 'Attenuatella and Yabeina, in Tunisia. Diener ( 1 903, p.53) argued that the fauna was to be correlated with the Wargal, and especially Kalabaghian Member of the Salt Range, on the basis of some 2 7 species. Of these 23 were also shared with the Chhidru Formation, but four were exclusive to the Wargal. Although some of the identifications may be questioned, this still appears to be an acceptable conclusion. But some reservation must be retained because stratigraphic control within the crag has not been achieved; and the overall geological setting of the limestone points to possible origin from a carbonate bank on a guyot or mid-oceanic ridge, under conditions of slow sedimentation and high likelihood of faunal condensation or intermixing. Such can be resolved only by re-examination of the fauna, with attention to internal detail. Strong support for a Punjabian age for the Chitichun no. 1 fauna is offered by the ammonoid Cyclolobus walkeri Diener at Chitichun no. 1 , which Furnish & Glenister ( 1 970, p. 1 72) have recognised as C. cf walkeri in the Chhidru Formation of the Salt Range
In north-west Nepal, the Lamnimargus himalayensis Zone is overlain by some 1 00-200 m of calcareous sandstones, coquinites, shales and other lithologies with rich brachiopod faunas of the Echinalosia kalikotei Wat. Zone (Fig. 30). Some species are shared with the underlying zone, but many additional species appear as well in a complex of lithotopes and communities, including new species of Rugaria, Krotovia, Cancrinella, Platyconcha grandis Waterhouse, Quinquenella glabra Waterhouse, Megasteges nepalensis Wat., Transennatia gratiosus (W aagen), Pterospirifer, abundant Spiriferella rajah (Salter), and Neospirifer ravana (Diener). The zone lacks ammonoids apart from fragments of what appear to be Xenodiscus, but there are some strong faunal links with the underlying Lamnimargus Zone, that the zone is conceivabiy only slightly younger, and therefore upper Punjabian or Chhidruan in age. This appears to be reinforced by the presence of some Chhidruan species, as well as the absence of any Djulfian key species and genera. Thus the Nepalese succession provides some support for the assertion by Grant ( 1 968, 1 970) and Waterhouse ( 1 972a) that the Kalabaghian and Chhidruan faunas are closely related, but show zonal differences, as expressed herein by the use of substages for the two units, placed in one stage.
In the Thakkola region of north-east Nepal, Bordet et a/. ( 1 9 7 1 ) assigned almost 600 m of coarse and fine clastics with some carbonate to the Chini Thu Formation, overlying early Carboniferous carbonates of the Tilicho Lake Formation. The topmost fauna contained Spiriferella rajah, suggestive of a Punjabian and perhaps Chhidruan age, but the lack of accompanying fossils prevents assignment to one of the zones found in west Nepal. Fossils from the underlying beds display, from the lists in Bordet et al. ( 1 97 1 ), a bewildering array of affinities and ages, so that assessment must await formal description. It was suggested that the beds represent in part the great unconformity between Chini Thu and Ice Lake beds to the west (Bordet et a/., 1 9 7 1 , p. 1 16). The spiriferids, as. prov�sionally identified by Mme Legrand, suggest a lower Permian age, or even correlation with the Fenestella Shales of Kashmir. The accompanying Productida
'

Punjabian Stage 1 45
were provisionally identified by Mme G. Termier with early Middle and early Carboniferous genera, and the Permian genus Stepanoviella. The faunal sequences promise to be of high value when described.
The upper faunas from the Selung Formation of south-west Tibet north of Mt Jomlo Lungma include Neochonetes, Chonetella nasuta, Transennatia. Stenoscisma gigantea (Diener), and Phricodothyris (Mu An-tze et a/., 1973) which - may be of Chhidruan age.
It would thus appear that modern studies have on the whole reinforced early work by Diener ( 1 903, 1 9 1 5), with the recognition of two major faunas, rather closely allied, as in Waterhouse ( 1 972a).
Grant & Cooper ( 1973, fig. 4, see Table 9 herein) offered another view, showing the Kuling Shales (with Cyclolobus) as very late Permian (or Changshingian Stage i.e. Dorashamian), the Productus Shales (with Waagenoceras?) as Capitan-Word (i.e. Kazanian-Punjabian), and the entire Zewan fauna (with Xenodiscus, Cyclolobus) as approximately Kungurian-Baigendzinian (Leonardian). Yet all fall in one Lamnimargus himalayensis brachiopod zone.
Karakorum, Afghanistan
Several faunas from the Karakorum Range are similar. From the upper Hunza Valley Sestini ( 1965b) recorded two small faunas as probably Lower Permian in age. They include Neochonetes, Linoproductus lineatus (Waagen), Elivina tibetana (Diener) and Fusispirifer nitiensis (Diener) as in the lower Punjabian faunas of the Himalayas.
From the Camp Staghar glacier in the Karakorum Range, Merla ( 1 934) and Sestini ( 1965d) recorded a fairly large brachiopod fauna in white fine limestone, supposedly of Lower Permian age, and with a number of early Permian species, but also with many Salt Range Wargal species. Merta 0 934) called the fauna 'Lopingian'. Renz (1940b, p.277) described another large fauna from the Shaksgam Valley of the Karakorum, P.ssigning it to the Upper Artinskian. But brachiopods, even though in need of revision, and including many tiny specimens, strongly suggest a Kazanian or Kalabaghian age, probably confirmed by accompanying large Parajusulina. The fauna includes Compressoproductus, Choristitella wynnei (Waagen), Martinia, ?Chaoiella margaritatus (Mansuy) and ?Attenuatella. Attenuatella occurs in Chhidruan faunas of Mexico, New Caledonia, New Zealand and Australia and in ?Kalabaghian or late Kazanian faunas of Tunisia and perhaps Taimyr Peninsula. On the whole the fauna would appear to be late Kazanian or Kalabaghian in age. Sestini ( 1 965d, p. 1 68) added various species, including some Sosio forms, and others found in the Wargal Formati�n of the Salt Range, such as Marginifera typica Waagen and Costiferina spiralis (Waagen). There are further species in common with Himalayan forms, such as Anidanthusfusiformis Waterhouse, Megasteges dalhousei (Dav.) and Stenoscisma purdoni (Dav.). Parafusulina and Pseudofusulina occur rather than Neoschwagerina or Yabeina, perhaps as in the Leptodus beds of Japan. However there is some doubt over the correlation. Whilst it is , true that there appear to be strong faunal links with the Wargal Formation, and few with the Amb Formation, the illustrations, descriptions, and especially stratigraphy are not well enough established to allow confidence, and few of the species identifications can be sustained without re-examination of the material. At a generic' level, the faunas approach the late Sakmarian faunas of the Trogkofel Limestone of Austria, and Safetdaron fauna of Fergana. It is considered that the similarity is due to similar palaeogeographic conditions, but further study is needed.
Carbonates in the upper part of the Permian sequence described by Termier et a/. (1974) from south of Kabul, Afghanistan, are likely to be of Kalabaghian age, with Marginifera typica Waagen.

146 Biochronology for the Marine Permian
Southeast Asia, China, Japan
Levels 7 and 8 of Cammon, Cambodia, as set out by Deprat ( 1 9 1 3), contain Yabeina globosa (Yabe) and may be Kalabaghian, or perhaps late Kazanian, as discussed previously. A Kalabagh age is preferred from Japanese evidence.
Member C of Ishii et a[. ( 1969) at Sisophon, west Cambodia (Table 38) contains Yabeina asiatica Ishii and Lepidolina multiseptata (Deprat), with a fairly large brachiopod faunule of Kalabaghian age, including species typical of the Himalayan faunas, such as Strophalosiina tibetica (Diener), Lamnimargus himalayensis (Diener) and Elivina tibetana (Diener). A number of species occur in common with the Kalabagh fauna of the Salt Range (see also Waterhouse, 1972a, 1 973a). Many of the species recorded by Chi-Thuan ( 1 96 1) from supposedly slightly older beds are listed in Member C . Allied faunas were recorded by Deprat ( 1 9 1 3) for Indochina, with Lepidolina mu/tiseptata found at level 9 at Sisophon (Table 1 9, Table 42).
The high level 10 recorded by Deprat ( 1 9 1 3) is typified by distinctive brachiopods from Luang Prabang described by Mansuy ( 1 9 1 2), including ?Monticulifera (or Choanoproductus) paviei (Mansuy), ?Krotovia nystianus (De Kon.) and Stepanoviella mekongensis (Mansuy). These in turn are overlain by bivalve beds with Liebea, and similar brachiopod species at level 1 1 , possibly also Chhidruan in age on the basis of superposition and world brachiopod distributions. Similar brachiopods occur at Bannahai (Mansuy, 1 9 1 9, p.9 1). At Sisophon, the correlative Permophricodothyris grandis Zone, or Member D of Ishii et a/. ( 1 969, p.48) is typified by Echinauris khmerianus (Mansuy), and possibly identical 'Monticulifera', identified by Ishii et a/. ( 1 969) with sinensis (Frech), a species from the mid-Maokou Limestone of China. Permophricodothyris grandis (Chao) is found as in the Chhidru and Choutang beds. Fusulinacea include species of Parafusulina, with Lepidolina multiseptata gigantea (Gubler), and Yabeina minuta Thompson and Wheeler.
From black argillites in Quang Tri, central Vietnam, Chi-Thuan ( 1 962) recorded a small fauna that included several chonetid species originally described by Chao in the Wuchiaping and Changsing beds of China. A nearby richer fauna of Vietnam included Leptodus nobilis, Schuchertella semiplana, alleged Lamnimargus himalayensis, Uncinunellina jabiense and a Changsing species Spinomarginifera kweichowensis Huang. The latter fauna appears to be Kalabaghian and several of the species are identified with species found at Horizon C of Sisophon. But the argillite may be as young as Djulfian.
A small Punjabian collection described from Petchabun, Thailand, by Yanagida ( 1964), includes species of Orthotichia, Orthotetina, Tyloplecta nankinensis (Frech), T. yangtzeensis (Chao), Marginifera banphotensis Yanagida, and Haydenel/a kiangsiensis (Kayser) (Table 19).
From Bukit Tengku Lembu, Perlis, Malaysia, Ishii et al. ( 1972) recorded sandstones with the chonetid Micraphelia, known also in Guadalupian faunas of Texas, and Monticulifera, with a few other forms. Correlation with Member D of Sisophon, as suggested by Ishii et a/. ( 1 972), seems eminently acceptable, and helps correlate the Chhidruan Substage with part of the Guadalupian succession of Texas.
From Htam Sang in Burma, Reed ( 1 933) described a fauna that may be approximately Punjabian in age with Martiniopsis and Leptotids, and a number of Chhidru as well as Kalabagh species. The record of Krotovia, Martinia, Phricodothyris, Martiniopsis and several Notothyris, mostly not figured, so not available for assessment, suggests a Kalabaghian age as most likely, without being able to rule out a midKungurian age. Another Wargal equivalent is reported by Reed ( 1 93 3) from the road to T�ang-gyi, Mong Pawn, with Strophalosiina cf. tibeticus (Diener), typical of the lower Himalayan fauna, and Schuchertella semiplana (Waagen).
The. Yabeina Zone is well developed in the upper Maokou Limestone of China,overlappmg the range of Neoschwagerina, and accompanied by Verbeekina, Chusenel-

Punjabian Stage 147
Level Faun a , locality Correlation
11 Luang-Prabang greywackes , ?Krotovia nystianus Chhidruan (De Kon . ) Liebe a Substage
10 MonticuLifera paviei (Mansuy) , ?Krotovia nystianus (De Kon . )
9 LepidoLina muLtiseptata (Deprat) , S isophon Kalabagh Substage
8 Pong OUa Limestone , Yabeina gLobosa (Yabe) e arly Kalabagh-ian?
7 Pong Oua Limestone , Laos , Neosahwagerina margaritae Deprat upper? Kazanian
N. cf gLobosa (Yabe) , Leptodus� Linoproduatus Stage dielasmatids
6 Black limestone , Cammon , Pseudofusu Una subcy Undriaa (Deprat)
5 Langnac carbonate Neosahwagerina margari tae Deprat Kazanian Stage
4 Limestone , Neomise LLina Lepida S chwag. � Verbeekina verbeeki (Gein . ) , NeofusuLine LLa Lantenoisi Deprat Ufimian level
3 Limestone , Spiri fere Ua� Licharewiniid ?Elkin level
2 Limestone at Cammon , NeofusuLine LLa Lantenoisi ?Nevolin level
Deprat , ParafusuUna riahthofeni (Schwag. )
1 Carbonate at Cammon and Cambodia , Unainune L Lina tirrvrensis (Beyrich) , Chaoie LLa sumatrensis
?Filippovian aambodgensis (Mansuy) � SpirifereUina
?Septaaamera garoudi (Mansuy) Substage
Table 42 . Sequence in Indo-China , simplified from Deprat ( 1913 , p . 76)

148 Biochronology for the Marine Permian
Ia, Neomisellina, Kahlerina, Lantschichites and Wutuella. No associated ammonoids are found. Only a few brachiopods were mentioned by Huang ( 1932a,b; 1 933, p.97), reflecting the small amount of study on the faunas. The species may indicate a general correlation with the Kalabaghian of the Salt Range and Himalayas, for they include Neochonetes, Waagenites, Compressoproductus mongolicus (Diener), Hustedia grandicosta (Dav.), Terebratuloidea depressa Waagen, T. davidsoni Waagen, Phricodothyris and Martinia, with Krotovia janus Huang.
Although Sheng & Lee ( 1 974) appear to provide a disconcertingly different account of the faunas, showing Yabeina to be correlative with faunas containing Kufengoceras, Mexicoceras, Altudoceras and brachiopods Neoplicatifera huangi and Cryptospirifer, the correlations may be lumping ammonoid and brachiopod faunas. Their text does not explain the position of K1.4fenl{oceras.
The Kuman Stage of Toriyama ( 1967) is found in clastics and black limestone some 900 m thick (Table 20, Table 3 2), structurally complicated and represented by several 'biofacies' or communities. The lower part of the Kuman Stage, as herein restricted to exclude the Gujo fauna following Waterhouse ( 1969b), is equivalent to the Punjabian Stage. This involves the Lepidolina toriyamai Zone and elsewhere the Yabeina shiraewensis Zone, both said to overlap somewhat with the Yabeina globosa Zone. Not all faunas can be dated accurately, but most show a general Punjabian appearance, Shimizu ( 1 963) recording upper Wargal brachiopod species from the Mitsu area, and Yanagida ( 1958) recording a few brachiopods in the Mizukoshi Formation in shales about 100 m below the top of the upper member, just above an assemblage with Lepidolina toriyamai and Yabeina cf gubleri. A number of brachiopods have been described from the Kitakami massif, where the upper Kanokura 'Series' has been subdivided into lower Lyttonia (=Leptodus) and upper Lepidolina faunas (Kanmera, 1 953). Timorites occurs low in the Kanokura beds (Furnish, 1 966; Waterhouse, 1972a) to suggest a Captanian (=Punjabian) age. The Katchizawa Member has Monodiexodina persisting from older rocks with Leptodus richthofeni Kayser and other brachiopods (Minato et al., 1 954; Nakamura, 1 959). It is correlated with a Neoschwagerina fauna in the Kamiyase-Kesenmura area by Onuki ( 1956), who records other brachiopods also in the Maiya area. The overlying Iwahata Limestone has the Lepidolina multiseptata fusulinacean assemblage with compound rugose corals. Hayasaka ( 1925 ; 1 963, p.753) also reported on the Lyttonid beds and brachiopods in the southern Kitakami region and from the Kamiyase region in the Shigejizawa Sandstone Member, with Yabeina (fide Murata, 1 964, p.2 1), Verbeekina, and Parafusulina, and a somewhat non-descript ammonoid fauna of early mid-Permian appearance, including Propinacoceras and Stacheoceras, like the Iwaizaki Limestone which has Yabeina shiraiwensis, with Propinacoceras and Stacheoceras (Murata, 1 964, p.23). The Shigejizawa brachiopods include Leptodus richthofeni Kayser and others in some need of revision (Hayasaka, 1 960). Another Leptodus richthofeni fauna was described by Imamura ( 1 953) in the largely clastic Karita Formation, lithologically like the Maizuru Group. From the Maizuru Group, Shimizu et al. ( 1 962) recorded a Lepidolinatoriyamai Kanmera fauna or Lepidolina kumaensis - L. multiseptata according to Kanmera & Nakazawa ( 1973, p. 1 04). Sandstones and shales of about the same age contain the Kawahigashi brachiopod faunule (Shimizu, 1961 a), with some brachiopod species of the Chhidruan Substage, including Derbyia altestriata Waagen, D. cf. grandis Waagen, Lissochonetes bipartita (Waagen), L. morahensis (Waagen), Hustedia indica Waagen, a Wuchiaping species Haydenella kiangsiensis (Kayser) and Megasteges dalhousei (Dav.), which is found in Punjabian faunas of the Himalayas andPakistan.
Kalabaghian brachiopods are found in the Takauchi fauna of the Maizuru Zone(�hir?izu, �961a) with undoubted Himalayan affinities displayed by Strophalosiina ttbettca (D1ener), Schuchertella semiplana (Waagen), and Martinia elegans Diener.

Punjabian Stage 1 49
Leptodus riahthofeni Kayser is also present, providing a link to other Japanese faunas. In summary, it appears that at least two faunas are present. The lower Yabeina
shiraiwensis Ozawa fauna (Yabe, 1 964), perhaps represented elsewhere by the Leptodus richthofeni brachiopod faunule, is overlain or partly overlapped by the Lepidolina toriyamai Kanmera - L. kumaensis Kanmera subzone or communities, associated with Chhidruan brachiopods. But this is a very coarse simplification, because many Fusulinacean communities were present, with complex and overlapping interrelationships.
South Primoyre, Kolyma River
In south Primoyre of western Siberia, the Chandalaz Suite, 700-800 m thick, is likely to be of Punjabian age, with Verbeekina verbeeki (Gein.), Yabeina, Lepidolina ussurica (Dutk.), and species of Enteletes, Prorichthofenia, Tyloplecta yangtzeensis (Chao) and stenoscismatids (Likharev, 1 966, table 1 8).
In the same general region at Kur Ufiimsk the Cereduchin Suite (950- 1000 m) is of Kalabaghian age, with the ammonoid Timorites and a few brachiopods overlying the Babstov Suite of Kazanian and Kungurian age with a few more listed brachiopods in beds 190 m thick (Table 43). The Tirin Horizon of north and west Verchoyan, 1 00-700 m thick, may be Punjabian in age (Likharev, 1 966, p.35 1 ), containing numerous Atomodesma variabilis Wanner and A. costata Popov, reminiscent of the Puruhauan faunas of New Zealand and Basleo - Amarassi faunas of Timor. Atomodesma and Kolymia also occur in the Chenoyar Suite of Taimyr Peninsula (Likharev, 1 966). Conceivably the underlying upper Baikur horizon of Taimyr Peninsula is as young as Kalabaghian, in view of supposed Neospirifer bambadhuriensis (Diener) identified by Ustritsky & Chernyak ( 1963), and other species, as discussed previously.
The Gijigin fauna of the Kolyma River area may be Kalabaghian, as outlined above. Perhaps the youngest Arctic faunas of Permian age are found in north-east Siberia (Likharev, 1 966; Zavodowsky, 1 968, 1 970) in the Hivatch Suite 1 64 m thick in Omolon, 2000 m thick in the Kolyma River area (Table 1 5). The fauna is moderately large, with species of 'Strophalosia', Linoproductus, Stepanoviella, Chivatschella, and Neospirifer, but as the fauna is younger than any other large fauna of northern latitudes there is no basis for comparison, and little evidence about its upper age limits. It may be Punjabian, even as old as Kalabaghian, but could be even late Permian. Kolymia inoceramiformis Likh. is present, as in many Punjabian and/or Djulfian faunas of Siberia, Timor, Australia and New Zealand, with other members of the Atomodesminae that are too poorly figured to allow close comparison. However Etheripecten volucer (Likharev) is very close to E. striatura Waterhouse of New Zealand, suggesting a possible Chhidruan age, and perhaps a Chhidruan age is supported by the large Strophalosiid species, such as S. chivatschensi's Zavodowsky, like Echinalosia kalikotei Wat. from Chhidruan beds of north-west Nepal. On the other hand the numerous linoproductinids suggest a Kalabaghian age.
Mongolia, Pamirs, Armenia, Iran
The young Permian faunas of Mongolia from the Jisu Honguer Formation were assessed as allied to both the Wargal faunas of the Salt Range, and the Lopingian of China (Grabau, 1 9 3 1). The writer has not seen the collections, and can offer no final decision, but as a suggestion, it appears possible that the lower beds are Punjabian, and the upper faunas are Djulfian, as discussed later, in fair measure of agreement with Grabau.
The lower faunas, believed to be Punjabian, come from localities 1 190 and 1 1 92, and have numerous Enteletes, Geyerella, Aulosteges, Kochiproductus, YakovleviaMuirwoodia, Compressoproductus, Leptodus nobilis, Spiriferella including a transverse

St
ag
e
Pu
nj
abi
an
Ka
zan
ian
Kun
gu
ri
an
?B
ai
gen
dz
in
ian
Sak
ma
ri
an
-
As
se
li
an
Ku
r
Uf
im
S
ig
ni
fi
ca
nt
F
os
si
l
Ce
re
du
ch
in
S
ui
te
T
imor
ite
s 9
50
-l
OO
Om
Up
pe
r
mi
dd
le
Pe
sch
ani
k
=
Bab
st
ov
Su
it
e
Su
it
e
lo
we
r
Un
gu
n
Su
it
e
75
0-
SO
Om
Ca
na
r S
ui
te
1
70
0-
lS
OO
m
Kuk
an
S
ui
te
=
?Si
ba
ig
ou
P
seu
do
fu
suZi
na
Can
aeZZ
ina
B
OO
m
Su
it
e
kraf
fti
�
seZZ
ina
aZau
diae
Ul
kuk
S
ui
te
=
Ka
va
ler
ov
S
ahw
ag
erin
a*
11
20
-1
15
0
m
Su
it
e
fu
ssif
orm
is
,
S .*
moe
zte
ri
---
--
---
�
Tab
le
4
3.
Co
rr
el
at
io
n o
f P
er
mian
a
t
Ku
r
Uf
im
, S
ik
ho
te
A
li
n
(fr
om
Li
kh
are
v,
19
66
, T
abl
e
18
).
VI
0
�
�· g. � ::s
c 0 � '0' ...
...... ::s- 11) � ... :;· 11) � ...
�
i:j•
::s

Punjabian Stage 1 5 1
form like S. turusica Einor, and Not�thyris. The Linoproductids with Kochiproductus suggest cool water faunas; as in the Kalabaghian Substage. A number of species range higher, but younger faunas lack the distinctive productids, except in the immediately overlying locality 1 196 (Table 46) which might be high Punjabian, as it lacks significant Linoproductidae.
The basal faunas with Linoproductinids are followed by faunas in cherty limestone, some 275m higher at localities 1 205- 1 2 10 (Table 47). Here Spiriferella mongolica Grabau is abundant, a distinctive form ascribed to various species by Grabau, with projecting ventral umbo, long concave posterior walls, simple plicae, and closely costate dorsal fold. This species is difficult to match with other known forms, and may be post-Chhidruan or late Chhidruan in age. Rare shells like Spiriferella rajah are found also, suggesting a late Punjabian age, almost exactly comparable in position to late Chhidruan Spiriferella rajah communities in Nepal, but perhaps the Spriiferella mongolica community is slightly younger. Other species, including Linoproductids, Marginifera and Streptorhynchus, are rare.
However, I am not satisfied that the preceding ages have been finally established. There is the disconcerting occurrence of what appears to be Timaniella, a KungurianKazanian genus, at locality 1209 in the Spiriferella bed. The shell was misidentified by Grabau { 193 1 , pl. 23, fig. Sa-c) as Neospirifer moosakhailensis, and indicates an IrenianLower Kazanian age. If this horizon were really Nevolin (Table 47), the underlying faunas with Kochiproductus and Yakovleviinae at Joe. 1 1 92 would be Filippovian, and the overlying faunas at loc. 1 1 93 and 1 1 94 with numerous Leptodus and Martinia would compare with the Sosio fauna. That so great a range of possibilities is still open underlines the need for detailed and specific re-examination of the faunas of the Jisu Honguer beds.
Punjabian faunas appear to be scarcely represented in the Pamirs, north Caucasus or Crimea Peninsula (Table 1 8). Grunt & Dimetriev { 1973, table 1 , p.2 1 ) showed Yabeina archaica Dutk. with Neoschwagerina margaritae near the top of the Murgab Stage and Yabeina has been recorded from the uppermost Murgabian beds above the Nikitin (Kazanian) faunas of the Greater Caucasus (Likharev, 1 966, p.394).
Iran and Armenia offer thin beds with significant faunas of Punjabian age (Table 46). On the Armenian side of the border at the Araxes River (RI.izencev & Sarytcheva, 1965; Waterhouse, 1972a) the Gnishik Horizon contains a number of Fusulinacea, including Polydiexodina persica Kahler, Verbeekina, Nankinella, Chusenella, and Pseudofusu/ina and many brachiopods such as Edriosteges, Orthotetina, Leptodus richthofeni Kayser, Richthofeni lawrenciana (Koninck), Phricodothyris indica (Waagen), and Terebratuloidea species in common with the Wargal Formation, and Japanese faunas of Kalabaghian age. Various other species indicate a more general Punjabian age (Haydenella tumida, Leptodus nobilis). The only puzzling occurrence is that of Martiniopsis injlata Waagen, which typifies the Chhidruan Substage in the Salt Range. But an allied Kalabaghian form occurs in New Zealand, suggesting that reexamination of the specimens may change the identification or extend the time range. Similar faunas are found on the Iranian side of the border at Kuh-e-Aii Bashi (Stepanov et a/. , 1969).
The overlying Hachik Horizon of shales and limestone 70- 100 m thick has many of the same species, with only four distinct-Rhipidomella vediensis Sok., Orthotetina arakeljani Sok., 0. dzhu/fensis Sok. and Septospirigerella megridagica Grunt. On the Iranian side of the border Martiniopsis ex. gr. injlata is found at Kuh-e-Ali Bashi (Stepanov et a/., 1 969, pp.22, 23) at unit 23 and unit 28. The Hachik fauna is likely to be of Chhidruan age, chiefly from its place in the stratigraphic column and the distinctiveness of its fauna from the Gnishik or Kalabaghian fauna. My Armenian colleagues are convinced that the Hachik and Gnishik faunas are more distinct than suggested by the studies in Ruzencev & Sarytcheva ( 1 965).

1 52 Biochronology for the Marine Permian
Over fifty species of Brachiopo_ds, Bryozoa and Mol!usca have been described from the Ruteh Formation (Table 3 1 ) of the Elburz Mountains, north Iran, by Sestini ( 1 965a). Almost 40% are found also in the Wargal Limestone of Pakistan. Nine species from Ruteh carbonates at levels 1 and 2 suggest or permit a Kalabaghian age, including Derbyia altestriata, Schuchertella semiplana, species of Cleiothyridina, and the presence of Stepanoviella and Compressoproductus. A few species are also found in the Himalayas. Level 5 has species identified as Marginifera typica Waagen and Costiferina indica (Waagen), which would suggest a Kalabaghian age if correctly identified, but the stratigraphic position of level 5 is not clear. There is little in common with the faunas described by Douglas ( 1 936) from south Iran, presumably because the ages differ.
The Permian of Djebel Tebaga, Tu.nisia, (Glinzboeckel & Rabate, 1 964) has a brachiopod fauna of modest size high in the Permian sequence in carbonates, clays and grits, with Yabeina syrtalis (Douville) and Yabeina globosa Yabe, suggesting a horizon close in age to Punjabian faunas of south-east Asia, and Japan. This does not accord well with the very large number of Sosio brachiopod species recorded by Termier &Termier ( 1957) which would imply a slightly earlier Kazanian age, but the Djebel fauna does have an Ambocoeliid ?Attenuatella unknown at Sosio, implying a late Kazanian or Kalabaghian age by analogy with occurrences in Taimyr Peninsula and OmolonKolyma River, or even Chhidruan age by analogy with Mexico and New Zealand. Unfortunately the Tunisian brachiopods are sketched, not photographed, so that it is difficult to check identifications. Renewed study of the mid-Permian brachiopods from the Mediterranean, including Sicily, Tunisia, and the Aegean is required to clarify the sequences and correlations. Miller & Furnish ( 1 957) recorded various ammonoids, including Agathiceras, Popanoceras, Peritrochia and probable Stacheoceras from essentially the same horizon. They assigned an early Guadalupian ag�, and considered the fauna to be correlative with that of Sosio, as most of the brachiopod identifications would also suggest. But the presence of Yabeina and ?Attenuatella appear to be decisive in suggesting a slightly younger age. It is interesting to note the generic similarity of the ammonoids to those described from the Shigejizawa beds of Japan, of Kalabaghian age.
United States, Mexico
The Capitanian Stage of the Guadalupian Series is mostly of Kalabaghian and Chhidruan age (Table 26). Faunas are restricted largely to the Delaware Basin and Guadalupe Mountains (Fig 2 1). Fusulinacea as summarized by Ross ( 1 967b, p. 1352)are typified by advanced Polydiexodina with rare but advanced Yabeina (Ross & Nassichuk, 1970) as well as Leella, Codonofusiella, Reichelina, Lantschichites, etc., most of them known also in older rocks. Brachiopods collected from the middle Capitan of Guadalupe Peak were listed as the typical Guadalupian fauna by Girty ( 1909, p. 16),with a few additional forms since described by Cooper & Grant ( 1 969) and others. It isnot possible to assess faunal assemblages or precise age until the redescriptions arepublished .by Cooper & Grant (in press). Noteworthy is the presence of Orthotetina (as 10 Armema), and supposed Elivina (as in the Himalayas), together with Oldhamina, Au�ostegiids, many . linoproductinids, including Compressoproductus and Anidanthus, vanous Wellerella-hke forms, Crurithyris, Martinia and Heterelasma. The ammonoidsare . refer�ed. to Timorites, Medlicottia, Kingoceras, Xenodiscus, Xenodiscites,Strtg?gomatlles, Waa?enoceras, Epadrianites, and first appearance of Episagiceras. Furmsh. ( 1 973, p.539) mcluded the Manzanita Member at the top of the Cherry Canyon�orma�10n, and the Hegler Member at the base of the overlying Bell C anyon Formabon, w1th most �f the res� of the formation up to the Lamar Member, through some300m of strata, 10 a restncted Timorites or Capitan Zone.
At Las Delicias, west Coahuila (Table 35), Mexico, brachiopods have been

Punjabian Stage 1 53
described by Cloud (in King et al., 1 944). No specific age limits were provided by Cloud, but it is likely that the ammonite shale of band or 'Bed' 5 of King ( 1944) in the Difunta section is Punjabian, with Neochonetes, Prorichthofenia, Leiorhynchoidea, Paranorella and Attenuatella attenuata (Cloud). Band 2 has a Buxtoniid and Leptodus, with Fusulinacea Polydiexodina capitanensis Dunbar and Rauserella erratica Dunbar. Band 7 of King ( 1944, p. 14) with Polydiexodina shumardi is at the same horizon, according to Spinoza et a!. ( 1970). Ammonoids include Timorites schucherti Miller & Furnish, Strigogoniatites kingi Miller, Stacheoceras toumanskiae Miller & Furnish, and Waagenoceras dieneri Bose. The ammonoids are regarded as correlative in a general way with those of the Hegler Limestone in the Capitan of the Guadalupe area by Spinoza et a!. ( 1 970}.
Attenuatella attenuata (Cloud) from Mexico is closely allied to A. incurvata Wat. of Chhidruan and Urushtenian age in New Zealand (Waterhouse, 1 964) and especially to Attenuatella convexa Waterhouse ( 1967b) not Armstrong ( 1 968) from New Caledonia in beds close to the boundary between Atomodesma trechmanni (Marwick) and Atomodesma cf trabeculum Waterhouse. These two species are restricted in New Zealand to the Chhidruan and Urushtenian Substages respectively, and their occurrence together in New Caledonia suggests faunal intermixing, either by collecting or sedimentation, at a time close to the Punjabian-Djulfian boundary. The Mexican species is possibly of the same age, i.e. about late Chhidruan.
Madagascar, Timor, Australia, New Zealand
In north Madagascar, Astre ( 1934) described a small fauna from the 'Productus limestones' of the lower Sakamena Group. The fauna has various brachiopods suggestive in a general way of a Punjabian or younger age, including a number of supposedly unique species of Enteletes, Liosotella, Uncinunellina, and Spiriferellina, with Neospirifer moosakhailensis (Dav). and other forms. Overlying shaly sandstones contain two additional brachiopod species, including a sub-species or variety of Waagenoconcha abichi (Waagen), a species which is especially characteristic of Chhidruan faunas, and the ammonoids Cyclolobus walkeri Diener and supposed Xenodiscus carbonarium Waagen (see Waterhouse, 1 973b)
Two rich faunas come from Timor, but unfortunately exposures are poor and the structure complex (Table 1 9, Table 44). The lower or Basleo fauna is clearly Kalabaghian in age (Waterhouse, 1 972a), with such diagnostic species as Strophalosiina tibetica (Diener), Megasteges dalhousei (Davidson), Chonetella nasuta Waagen, Chaoiella chiticunensis (Diener), Spiriferella rajah (Salter), and Elivina tibetana (Diener), all identical with Himalayan species, together with , the additional species of note, Camarophoriina antisella (Broili). The faunas are described chiefly by Broili ( 1 9 1 6}, Hamlet ( 1 928) and Wanner & Sieverts ( 1 935). The ammonoid Timorites is present. Furnish ( 1973) also stated that 'Wordian' (that is Nevolin to upper Kazanian) ammonoids were present in the Basleo collections. Whether this means that the ammonoids were long ranging, or that the Basleo faunas include two horizons, as could easily be the case, is not clear.
The Amarassi faunas, described by Rothpletz ( 1 892}, Broili ( 1 9 1 6) and Hamlet ( 1928) are fairly close to those of the Basleo faunas, but show more affinities with the Chhidru and fewer with the Kalabagh faunas of the Salt Range. However the fauna is not clearly Chhidruan and has few distinctive species (Timoriina broili; Orthotetina altus Hamlet), leading Waterhouse ( 1972b) to suggest that it was early Chhidruan in age. Many of Rothpletz's species were not mentioned by Hamlet ( 1 928) and it is not certain whether they were discounted, or as seems more likely simply not rediscovered. An unusual ammonoid Cyclolobus persulcatus Rothpletz is found in the Amarassi fauna, with Timorites. Spinoza et a!. ( 1970, p.73 1) correlated the Amarassi fauna by means of Stacheoceras cf. tridens Rothpletz with the Lamar Limestone at the top of the

1 54 Biochronology for the Marine Permian
S tage Sub s tage
Dorashamian
Djul fian
Pun jab ian Chhidruan
Ka labaghian
Kazanian
Kungur ian Irenian
F i lippov ian
Baigendzinian
Sakmarian Aktas t inian
S terli tamakian
Tas tubian
As s el ian
Fauna Brachiopods Auanonoids
-
-
Amara s s i Amarassi
Ba s l eo Bas l eo
Bas leo
? Tei Wei
Bi tauni
Bi tauni
Bi tauni
-
S omoho le
Table 44. Correlation of faunas from Timor , with occurrences of ammonoids recorded by Furnish ( 19 7 3 , p . 524) . See Table 19 .
'

<. 0 t""
(')
Punjabian Stage 1 55
L E G E N D
c=J RECENT TO JURASSIC
t=::=:.E===� TRIASSIC
PERMIAN - Bryneira Group
Productus Creek Group
jjjjjjjjjjjjjjj!!jjjj Artnurton Group
F;:;:;:;�;�:;:;:;:;� Kuriwao Group
� Eglinton Volcanics. Takitimu G.roup
� Waipahi Group
IGNEOUS - Basic
- Basic and Ultrabasic
? CAR B O N I F E ROUS � Tuapeka Group
? P E R M I A N - TRIASSIC Bl Otago Schist
LOWER PALEOZOIC OF CRATON.
AND CR ETACEOUS GRANITE
0 5 10 1 5 20 M I LES
Fig. 3 1. Geological map of Southland, New Zealand, showing distribution of Permian rocks and formations, with facies that changed during the period. From Waterhouse (1973c, fig. 3 , p. 36). Reproduced with permission of Dr W. Tovell, Director, Royal Ontario Museum.

1 56 Biochronology for the Marine Permian
Capitan in Texas-New Mexico and the upper La Colorada beds of Mexico, but it is believed thar the beds in North America, at least those of Mexico, may be slightly younger, for they contain primitive Araxoceratidae, suggesting a post-Chhidruan age (Waterhouse, 1 972a).
In Western Australia the sequence is largely terrestrial in the middle mtd late Permian, but a small fauna from the Hardman Member at the top of the Liveringa Formation in the Fitzroy Basin is probably Chhidruan (Table 29), containing Derbyia grandis Waagen, Waagenoconcha imperfecta Prendergast, Cleiothyridina gerardi (Diener), and Neospirifer ravana (Diener) according to Thomas ( 1967). Dickins ( 1 963, p.67) recorded the Amarassi bivalve Atomodesma undulatum Rothpletz in the upper Liveringa beds, and Muir-Wood & Cooper ( 1 960) comoared Waagenoconcha imperfecta to the Amarass1 species W. waageni Rothpletz). Two small faunas are known from the Port Keats area in the Bonaparte G:ulf, northwest Australia (Thomas, 1957, p. 1 8 1). Fauna C of Thomas has Martiniopsis and Neospirifer, perhaps suggesting aKalabaghian age, pending full description, and the overlying Fauna D may be Chhidruan, with Waagenoconcha imperfecta and Leptodus nobilis, amongst other species yet to be described.
In eastern Australia a Chhidruan fauna appears to be present as Fauna V of Waterhouse ( 1969c), in the middle Flat Top Formation and upper South Curra Limestone of the Bowen and Gympie Basins, Queensland (Table 38, Fig. 2 1 ). Characteristic species include Capillonia brevisulcus (Waterhouse), Plekonella multicostata Wat., Stenoscisma papilio Wat. and Attenuatella incurvata Wat., (Runnegar, 1969b; Dear, 1 969 ; Runnegar & Ferguson, 1 969), all found as well in a fuller stratigraphic sequence in New Zealand. Runnegar ( 1969b) correlated the fauna with the Kazanian Fauna IV, but this is not acceptable, because many new species are present in addition to some species lingering on from the underlying stage. However the limits of the faunas are not clearly established as yet (Dear, 1 972). Conceivably the Havilah fauna, with Tomiopsis havilensis (Campbell) is of this age or slightly older (?Kalabaghian).
New Zealand faunas of Kalabaghian and Chhidruan age are widespread in thick sequences, especially well exposed around the coast-line (Waterhouse, 1 973c). Fig. 3 1 . The sequence is as follows :
Age Formation Urushtenian Little Bed
Sandstone Chhidruan Tramway
Sandstone Chhidruan Wooded Peak &
Upper Glendale limestones
Chhidruan upper AG4 Formation
Kalabaghian lower AG4 Formation
Faunal Zone Spinomartina spinosa
Atomodesma trechmanni
Spiriferella (subzone)
Plekonella multicostata
Martiniopsis woodi
Thickness 100 - 100 m
1 50 - 700 m
? + I m
50 m
30 m
The limestone containing the three lower zones varies from 50m to 1 ,000m in thickness.
Hrachiopods within the Martiniopsis woodi Wat. Zone (Table 3 7) are rather few in num�er, perhaps because diversity was inhibited by flourishing bioherms of rugose corals (m North Auckland) and stenoporid Bryozoa. They are accompanied by

Djulfian Stage 157
Fusulinacea, including Neoschwagerina margaritae Deprat, Yabeina, and Lepidolina multiseptata (Deprat) (see Waterhouse, 1 964, p. 1 3). As L. multiseptata is generally regarded as an advanced Yabeina a slight unconformity or barren interval may separate the fauna from the underlying Terrakea brachythaerum Zone of late Kazanian age. The immediate overlying faunas are more diverse and widespread, with several Kalabaghian and a few Chhidruan links, especially displayed by Cleiothyridina laqueata Wat., Rhynchonellid afT. wynnei Waagen, and Tomiopsis para/lela (Wat.) like T. punjabica (Reed) from the Salt Range. Filiconcha n. sp. is close to Monticulifera from the Chhidruan Pong Oua and Luang Prabang faunas of southeast Asia. Attenuatella incurvata Wat. entered the sequence, and is found in faunas of Punjabian age in New Caledonia and Mexico, and perhaps in Tunisia, as well as Queensland.
Above this fauna in New Zealand comes Spiriferella representing a late Chhidruan community or subzone. Spiriferella is also particularly characteristic of what appear to be late Chhidruan faunas in the Himalayas at least in north-west Nepal. Its age in New Zealand, though not finally established, could be identical.
DJULFIAN STAGE
Armenia, Iran, Caucasus
The Djulfian Stage of various authors is redefined following Waterhouse ( 1972a) to embrace two substages, characterised by Araxoceratidae and by the presence of Codonofusiella, persisting from older faunas, and absence of Neoschwagerininae. For die lower of these substages, the name Urushtenian Substage is used, based on faunas and outcrops in the Northern or Greater Caucasus. The name is only provisional, on the basis that it has long been applied to the most diverse faunas yet described. No designated type section is offered until the Russians decide if they want the name. Taraz ( 1971) has proposed the Abadehian Stage, based on outcrops and faunas in central Iran, which correspond to the same interval. Since the Abadehian unit does not appear to be so richly fossiliferous, its use is deferred pending international discussion, but the name has advantages of being associated with faunas close to those of the overlying type Baisalian and younger beds, and also enjoys priority. The substage overlying the Urushtenian = Abadehian is the Baisalian unit as defined by Waterhouse ( I 972a) with faunal content described by Ruzencev & Sarytcheva ( I 965), and type section offered by the section of Djulfa Gorge, Armenia.
In the Greater Caucasus, the Urushtenian Horizon, 1 2- 1 28 m thick, has very large brachiopod and fusuline faunas described in part by Likharev (-1 932, 1937) and treated as lower Pamiran by Likharev & Mikluko-Maklay ( 1 964) for their faunal subdivisions in central Asia. Small Foraminifera include Reichelina minuta Erk., Cribrogenerina permica Lange, and Pachyphloia .paraovata maxima Mikl.-Makl. Brachiopods of Horizon B of Likharev ( 1 932, 1 937) include Punjabian species Leptodus richthofeni, Strophalosiina tibetica, Stepanoviella, Compressoproductus, and Echinauris opuntia reinforced by Ombonia and Dorashamia to suggest the likelihood of an early Djulfian age. One of the most characteristic species is Crurithyris (=Orbicoelia) tschernyschewi Likharev. Ruzencev & Sarytcheva ( 1 965, table 2, opp. p.32) showed the Urushtenian as lower Djulfian on the basis of Fusulinacea, and the brachiopod faunas support this correlation.
In the Armenian region, the Urushtenian Substage is poorly represented by Codonofusiella (Ruzencev & Sarytcheva, 1965). Arakeljan et a/. ( 1 964) and Stepanov et a/. ( 1 969, p.27) however argued strongly to include this band in the upper Hachik horizon. The overlying faunas of the Baisalian Substage are well represented in Armenia

158 Biochronology for the Marine Permian
Stage Substage Unit Thick-ness
Dorashamian Griesbachian
Ogbinan 20m
VedHm 4.5m
Djulfian Baisalian
47m
urushtenian ?
Punjabian Chhidruan Hachik beds 70-lOOm
Kalabaghian Gnishik beds 70-lOOm
Kazan ian Armik
Kungurian Asni
Baigendzinian -
Sakmarian Davalin
Asselian
Significant Fossils
Claraia
Paratirolites
Come licania� Janiceps� Phisonites
Haydene lla, Vedioceras,
Dzhulfoceras� Oldhamina
Araxi levis, Ara.xoceras � Vescotoceras
(upper Hachik, lower Djulfian of Ruzencev and Sarytcheva, 1965) Codonofusiella Reichelina
C husenella, Nankinella
Nankinella
Pseudofusulina, ParafUiJulina
t Table 45. Sequence in Armenia, Djulfa Gorge, Dorasham Siding II
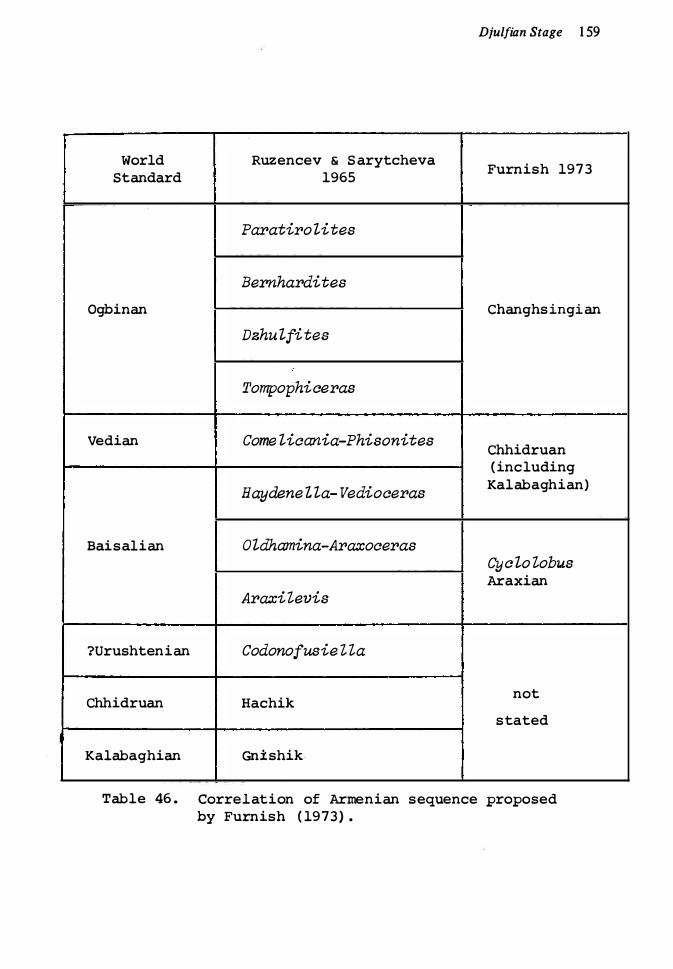
Djulfian Stage 1 59
World Ruzencev & S arytcheva Furnish 1973
Standard 1965
Paratiro Zites
Bernhardites
Ogbinan Changhs ingi an
Dzhu Zfites
Torrrpophic:eras
Vedian Come Zic:ania-Phisonites Chhidruan ( inc luding
Haydene Z Za- Vedioc:eras Kalabaghian)
Bai s alian OZdhamina-Araxoc:eras Cy c: Zo lobus Araxian
Araxi levis
?Urushtenian Codonofusie l la
Chhidruan Hachik not
s tated
Kalabaghian Gni shik
Table 46 . Corre l ation of Armenian s equence proposedby Furnish ( 1973) .

AG
E
(Gra
nt.
1970
) LI
THIC
SE
QU
ENC
E UN
ITS
AG
E (f
urni
sh a
nd G
leni
ster
. 19
70)
KATH
WA
I LI
MES
TON
E TR
IASS
IC
DZH
ULF
IAN
AN
D H
IATU
S KA
THW
AI
DO
LOM
ITE
� "C
hang
hsin
gian
" an
d W
hite
sand
ston
e . ..
....
.. .
L Do
rash
amia
n mi
ssing
CH
HID
RU
·cH
HID
RU
AN
.
GU
AD
ALU
PIA
N
Ran
ge o
f Cy
clolob
us
(=Ca
pita
n)
FOR
MA
TIO
N
_j_
WAR
GAL
I G
UA
DALU
PIA
N
LIM
ESTO
Nl
Fig
. 32
. D
iagr
amm
atic
co
mp
aris
on
of
th
e in
terp
reta
tio
ns
of
the
up
per
par
t o
f th
e S
alt
Ran
ge s
equ
ence
off
ered
by
Gra
nt
(19
70
) o
n t
he
left
sid
e, a
nd
by
Furn
ish &
Gle
nis
ter
(19
70
) o
n t
he
righ
t si
de.
The
cen
tre
par
t is
a s
ket
ch o
f th
e li
thic
seq
uen
ce,
and
th
e str
atigr
aph
ic
no
men
clat
ure
; th
e ve
rtic
al ar
row
dep
icts
th
e ra
nge
of
the
amm
on
oid
Cycl
olo
bu
s. F
rom
Gra
nt
& C
oo
per
(1
97
3,
f'Jg.
3, p
. 5
81)
. R
epro
d
uce
d w
ith
perm
issi
on
of
the
auth
ors
.
- g) ttl c· (")
::- <3 ::s <:) � 'Q> .... :;. !I> � .... s· !I> � ....
�
s· ::s

Djulfian Stage 1 6 1
(Table 45, p.272) as elaborated by Ruzencev & Sarytcheva ( 1965). Characteristic ammonoids include Vedioceras and Araxoceras, and correlative zones are recognised across the border in northern Iran by Stepanov et a/. ( 1969) with an additional Tyloplecta Zone. The same brachiopod faunas range westwards into the Bellerophon Limestone faunas of Yugoslavia and Austria, described by Heritsch (1934), Simic (e.g. 1935b), Stojanovic-Kujenko ( 1 963) and others, with the report of Crurithyris to suggest Urushtenian as well as Baisalian faunas. In central Iran, Taraz ( 1971) has recorded Djulfian faunas, and proposed an Abadehian Stage for the lower faunas between the Hachik and later Djulfian. His unit 6 (Taraz, 1971) contains Eoaraxoceras and Kingoceras (Furnish, 1 973, p.542), indicative of an Urushtenian = Abadehian age.
Furnish ( 1973) offered a different interpretation of these faunas (Table 10, Table 46). The lower and middle Baisalian faunas were assigned to an Araksian Stage and separated from the overlying Vedioceras-Haydenella fauna although there is no significant faunal distinction that I can discern, indeed as Stepanov et a/. ( 1969) also confirmed. The Vedioceras-Haydenella fauna was regarded as one with the overlying Comelicania-Phisonites fauna, although in fact the brachiopods and ammonoids are very different. Species of Xenodiscus from the Chhidru Formation, Pakistan, were matched by Furnish with species from the Phisonites Zone, and Vedioceras was correlated with a fauna of no clear stratigraphic position in China (Chao, 1 966), and with the Amilobe beds of Madagascar. The Vedioceras correlations are acceptable, but not stratigraphically significant, and the link proposed by Xenodiscus seems dubious, for Xenodiscus is a long ranging genus (Middle Permian into Triassic), and cannot be reliable except at specific levels.
Djulfian faunas are well established in the Elburz Mountains of north Iran (Table 3 1 ). A small fauna described from level 3 and ?3 in carbonates by Sestini ( 1 965a) is Urushtenian judged from brachiopods, including Orthotetina and Orbicoelia tschernyschewei (see Waterhouse, 1 972a). Level S of Sestini ( 1965a), p.24) could be Baisalian, for its stratigraphic position is not certain, but its species suggest a Punjabian age, as discussed previously. A typical Baisalian brachiopod faunule has been described from 'bed 8' of the upper Nesen Formation in the same region by Sestini & Glaus ( 1966):
Pakistan
Elements of the same Urushtenian fauna, including Orthotetina cf arakeljani (low in the Djulfian of Armenia), Chonetella, Spinomarginifera, Crurithyris, (?Orbicoelia) extima Grant, and Martinia are found in the basal Kathwai Member of the Mianwali Formation of the Salt Range, Pakistan (Fig. 32), immediately above the Chhidru Formation (Table 28). The brachiopods are clearly Permian in age, as well demonstrated by Grant (1970), and Grant & Cooper ( 1973). These authors assigned a latest Permian ('Changhsingian') age, or earlier late Djultian age. This could be correct, but not on the bases so far published. Waterhouse ( 1972a) pointed out that this late age was based on misinterpretation by Grant ( 1970) of the Ruteh faunas set out by Sestini ( 1965a). My preference for a lower Djulfian age is based primarily on the occurrence according to Grant {1970) of Orthotetina cf: arakeljani like a Urushtenian or early Baisalian species from Armenia, Crurithyris or Orbicoelia, widespread in Urushtenian faunas, and Martinia like a late Lamar Limestone species (?Urushtenian) of New Mexico. If these have been misidentified, the age could be younger, perhaps as young as late Dorashamian, i.e. Griesbachian, for Crurithyris and possible Orthotetina have been found by the writer above Otoceras in north-west Nepal. A Changhsingian age (= ?Vedian) would seem less likely, as it would imply that the Kathwai unit was bounded above and below by paraconformities, but Grant ( 1 970) noted a strong resemblance of Kathwai Crurithyris to C. speciosa Wang from the Changhsingian faunules of China.
The Kathwai fauna has occasioned violent debate about its age, particularly from

Sa
It R
an
ge
, P
ak
ista
n C
ono
do
nts
)
'Tri
ass
ic I
Am
mo
no
ids
) 1
I K
ATH
WA
I M
EMB
ER
s I
Me
tre
I -J
0 1
/
/I
/
/L
-. . '·' �·· :: ::·· .
. . .
.
. .
..
. .
...
.
. .
.. '
. .
. .
. .
. .
. .
.
.....
. -
'Pa
lae
ozo
ic I
Bra
chio
po
ds
;
.. "'
�-
,....
. ,
. _
_
,....
'_
, ..
......
I '
..
.......
, ..
.. -
/ ,.
--
-..
--
......
. _ ..
.. ,
Fig
. 33
. Pe
rmia
n -
Tri
assi
c bo
undar
y be
ds i
n th
e Sa
lt R
ange
, Wes
t Pa
kist
an,
wit
h th
ree
inte
rpre
tati
ons
of t
he b
est
plac
emen
t of
the
boun
dary
. A
. B
ound
ary
sele
cted
by
Kum
mel
& T
eich
ert
(19
70);
B. I
nter
pret
atio
n of
Gra
nt (
1970
); C
. Per
mia
n at
th
e to
p of
the
Kath
wai
m
embe
r. F
rom
New
ell
(19
73,
fig.
2, p
. 6).
Rep
rodu
ced
with
per
mis
sion
fro
m D
r N
.D. N
ewel
l.
- 0\
N
a,
c·
(";
:::r-
... 1:1 ;::s 1:1 � � '0' ...
....
:::r-
<1>
c � ..
. :;
· <1>
� ...
�
s· ;::s
8 A

Djulfian Stage 1 63
Kummel & Teichert ( 1964, 1 973 and other papers of similar content), who have strongly stated that the brachiopods are Triassic in age, and attacked the integrity of opposition to their view. Kummel & Teichert ( 1 973, p.23) stressed that underlying white sandstone had similar brachiopods, and deduced that this implied a Triassic age for the overlying dolomite, without demonstrating why this does not suggest that the white sandstone is also Triassic, or alternatively, ensure a Permian age for the brachiopod faunules. According to Grant (in Kummel & Teichert, 1 970, p. 35), the white sandstone brachiopods are Chhidruan, not Kathwai, in affinity. Conodonts however range through both units. The brachiopod faunule is so convincingly Permian that one must wonder why a Triassic age should ever have been entertained. The reason is provided by the discovery by Schindewolf ( 1954) of a well preserved specimen of Ophiceras connectens Schindewolf, with no accompanying bracntopods, and vartous poor moulds (Kummel, 1 970) suggestive of Ophiceras and alleged Glyptophiceras in the lower Kathwai beds. Ophiceras connectens, it was believed, demanded an early Triassic age for the entire Kathwai fauna; though even the ammonoid evidence is not totally convincing (Newell, 1 973), especially since the discovery of Durvilleoceras woodmani Waterhouse, with an Ophiceratid suture in rocks of Permian age in New Zealand (Fig. 29). Perhaps the readiest solution to the problem is to consider that the brachiopods were of one age, and the ammonoid of another, and that the two have been mixed (though not occurring together), by reworking of sediment and faunas near wave-base, as suggested by Waterhouse ( 1 972a, b). But the most competent analyses of the problem are provided by Grant & Cooper ( 1973) and Newell ( 1973) who have carefully discussed alternative hypotheses to explain the apparent concurrence of fossils of conflicting age implications, making no prior assumptions, unlike their colleagues. They concluded that the ammonoid, at least the only well-preserved one, was probably 'precocious'. They further noted that both the Chhidruan and dolomitic beds lay within one conodont zone (Sweet, 1 970). The whole question ofwhether brachiopods and ammonoids occur in the same unit underlines the need for modern field mapping, careful collecting and up-to-date carbonate studies in the Salt Range. (Fig. 33).
Pamirs, Mongolia, China
The Pamiran Stage of the Pamirs and Fergana has Palaeofusulina, Reiche /ina and Codonofusiella, with large brachiopod faunules recently described by Grunt &Dimetriev ( 1973) in two zones, presumably equivalent to the Urushtenian and Baisalian Substages (Table 23). · The lower Taktabulak Suite includes restricted species of Notothyris, with Martinia and Permophricodothyris, and shares several species with the middle Taktabulak Suite, Haydenella tumida ( Waagen), Parenteletes ruzhencevi Sok., and Stenoscisma armenica Sok., found in the Baisalian faunas of Armenia. Middle Taktabulak species include Richthofenia caucasica Likharev, Strophalosiina multicosta Likh. and Krotovia jisuensiformis Sarytcheva, as a mixture of Urushtenian and Baisalian species, with supposed Enteletes dzhalgrensis Sok. of Obginan age in Armenia. The claim by Taraz ( 1973) that the Pamiran 'Stage' is pre-Djulfian cannot be sustained.
In the Jisu Honguer beds of Mongolia (Grabau, 1 93 1 ), the younger faunas with Spiriferella rajah may be late Punjabian in age, as outlined previously. A still younger fauna at localities 1 2 1 1 , 1 1 93, 1 194 from carbonate lenses in shales, has numerous Martinia and Derbyella, and is quite possibly Djulfian in age. Slight support is offered from the second highest fauna by Martinia mongolica Grabau, which was also identified by Waterhouse ( 1964) from a fauna in New Zealand that is now thought to be lower Djulfian, i.e. Urushtenian in age. Codonofusiella has also been recorded in the Jisu Honguer beds, supporting a Djulfian age. But a Kazanian age cannot be ruled out, as discussed previously.

1 64 Biochronology for the Marine Permian
Locality
1186
12 11 , 11�3,.3 , 1194
1205-
1208
1209 '
12 10
1196
1190 '
1192
Fauna and Lithology Possible Age Greater Following Grabau Possible Age
?Waagenoconcha, limestone bed in shale ?
( 150 m approx. )
EnteZetes sp . ; Derbye Z Za, Leptodus
nobi Zis, Spinomarginifera jisuensis, Urushtenian
Martinia mongoZica, Martinia spp.
limestone beds in shale
( 70 m interval approx. ) -
Orthotetina, $pinomarginifera, Urushtenian or
Spirifere Z Za aff. mongoZica, S. aff late Punjabian
"rajah" in thick limestone
( 200 m interval approx. ) -
Linoproductus, Compressoproductus, Urushtenian or ?Nevolin
Spirifere Z Za mongoZica in dark late Punjabian (Timanie LLa>
cherty limestone
( 70 m interval approx. ) -
Ente Zetes, Richthofenia, ?Punj abian
Spinomarginifera Hemiptychina in ?Chhidruan
carbonate beds in sandstone
( 200 m interval approx. ) -
Orthote tina, Geyre Z Za, Richthofenia, Punj abian ?Filippovian
Leptodus nobi Zis, Kochiproductus, -?Kalabaghian
Muirwoodia, Compressoproductus ,
Spirifere L La "rajah", Phricodo thyris
indica carbonate reefs in sandstone
Table 4 7 . Sequence of faunas in the Jisu Honguer Formation ,
Mongolia , summarized from Grabau ( 19 31) , with
alternative correlations .

Djulfian Stage 1 65
In China the Wuchiaping Limestone above the Maping Limestone is richly fossiliferous and is correlated with sandstone, shale and coal measures of the lower Yangtse Valley. It contains the Codonofusiella Fusulinacean Zone of Sheng (1964). Characteristic Baisalian ammonoids such as Vescotoceras, Araxoceras, Prototoceras, and Pseudogastrioceras occur especially in the upper Laoshan Shale, in the Loping Coal Measures (Chao, 1 966). Large brachiopod faunas have been described from the Choutang Limestone, and Wuchiaping Limestone (Huang, 1 933), and Sheng & Lee (1974) listed as characteristic Edriosteges poyangensis (Kayser), Rugaria soochowensis (Chao), Tylop/ecta yangtzeensis (Chao), and 'Squamularia' grandis (Chao), approacl;ling or identical to Armenian forms. Overall the faunas look like those of the Kalabaghian and Chhidruan Substages, perhaps due to failure to correctly assess the faunas, which probably need re-evaluation from first-hand examination. Alternatively, the Choutang Limestone is really Chhidruan, or the Chhidru faunas persisted in this region. Some caution must be retained over the stratigraphy, which involves considerable facies changes. Chao ( 1 966, p. 1 8 1 8) noted that diagnostic ammonoids were missing from the Wuchiaping Limestone, and rare in the Hoshan Limestone. He further noted that the Loping Coal Measures rested disconformably on Maokou Limestone with Verbeekina and Neoschwagerina, as if to imply that the Yabeina Zone was missing, or conceivably represented by Early Lopingian beds (Chao, 1947).
From Jinxian, Anhui, Zhang & Ching ( 196 1) recorded a brachiopod faunule from low in the Upper Permian (i.e. post Maokou) that is typically Djulfian, and includes Crurithyris or Orbicoelia, suggesting an Urushtenian age.
Japan, south-east Asia
It seems likely that at least meagre Djulfian faunas are present in Japan (Table 48). Toriyama ( 1973, p.509) suggested that the Lepido/ina kumanensis Zone with Lepidolina multiseptata of the Kuman Stage was Djulfian. It includes highly evolved Neoschwagerinids and Verbeekinids with Codonofusiella, Reichelina and Dunbaru/a. Kanmera & Nakazawa ( 1973) agreed, and noted that horizons of probable correlative age included Palaeofusulina in the Maizuru belt. The Kawahigashi brachiopod faunule of Shimizu ( 1962) was regarded as Lopingian. Unfortunately, Codonofusiella, Reichelina and Palaeofusulina do not offer indisputable evidence of age, for all three genera are widespread in older deposits, as discussed by Toriyama (1973), and they are valuable only through the absence of other forms. The presence of Yabeina and Lepido/ina would suggest to the writer that some of these beds are pre-Djulfian, as seems to be confirmed by the brachiopods. But higher faunas are found, and these appear to be of genuinely Djulfian age, as in the upper part of the Kuma Formation (see Kanmera & Nakazawa, 1973, p. lOS) with Wuchiaping Fusulinacea, and perhaps the Mitai Formation, with Codonofusiella near the base and Palaeofusulina above. The Tsukumi Limestone Group is also possibly Djulfian. Unfortunately, no significant brachiopods are found with these faunas.
It seems highly likely that Djulfian faunas will be discovered, or rather recognised as such in south-east Asia. Kanmera & Nakazawa ( 1973, p. 1 1 3) have pointed to the likelihood of Djulfian or Dorashamian fossils in the Jenka Pass region of Malaysia, where Nakazawa ( 1973) identified a distinctive transverse Orthotichia with 0. dorashamensis Sok. from the mid-Baisalian Substage of Armenia. A number of bivalves are also present. The fauna lies in purplish shale over plant beds, in turn above beds with brachiopods including Spinomarginifera likely to be of Punjabian age. Nearby limestones contain Yabeina asiatica Ishii. Thailand and Vietnam ( cf Chi-Thuan, 1 961 ) may well have such Lopingian faunas. Kanmera & Nakazawa (1973) implied that Lepidolina multiseptata faunas at the top of the Sisophon sequence were of Djulfian age, but this is unlikely.

1 66 Biochronology for the Marine Permian
United States, Mexico
Beds of possible, but rather uncertain Urushtenian age are present in Mexico and upper Capitanian of Texas and New Mexico. Spinosa et a/. ( 1 970) described a primitive Araxoceratid from the upper La Colorada beds of Las Delicias as Eoaraxoceras ruzhencevi Spinosa et a/., which may signify an Urushtenian age (Waterhouse, 1972a). It is accompanied by Stacheoceras cf tridens Rothpletz, Timorites, Episagiceras cf nodosum Wanner and Kingoceras kingi Miller. To judge from descriptions by Newell ( 1957), this appears to be approximately the topmost shale of the sequence, in 'Bed 1 ' of King et a/. ( 1 944, p. 1 3) of the Difunta section (Table 34). Polydiexodina mexicana Dunbar is present, with Chonetina and an overtoniid, together with a few brachiopods as yet undescribed. the writer collecting Leiorhvnchoidea for instance. Spinosa et a/. 0 9/UJ correlated the fauna With tne Amarassian beds of Timor, which was considered to be younger than the typical Capitan, as represented by the Hegler Limestone. Only one ammonoid species with Amarassian affinities is found in Texas, in the Lamar Member, called Strigogoniatites fountaini Miller & Furnish.
The Lamar Limestone (Table 26) at the top of the Capitanian, may be as young as Urushtenian, for Grant ( 1970) stated that a peculiar species of Martinia in the Kathwai fauna of Pakistan closely resembled a peculiar species from the Lamar Limestone. The correlations should become clear when the brachiopod descriptions are published by Cooper & Grant (in press), but Furnish ( 1973) has added strong support for the La Colorada-Lamar correlation from ammonoid evidence.
The Rustler Formation in the Delaware Basin of Texas has yielded a small brachiopod faunule (Walter, 1 953), but there is little evidence for age, pending revision, for none of the genera that characterise late Permian faunas of Asia are known. The fauna is correlated by means of bivalves with the Whitehorse Sandstone of the midcontinent in Kansas, Nebraska, Oklahoma and Texas (Newell et a/., 1 940), with a few brachiopods, including the unusual Dielasmatid Pseudodielasma Brill. Newell et a/. ( 1 940) were undecided whether the Whitehorse faunas were correlative with or younger than the Capitan. But the Rustler fauna lies well above Capitanian faunas, and is probably no older than Djulfian and possibly as young as Dorashamian.
New Zealand, Queensland
In New Zealand, the AG 5 sandstone of Wood ( 1 956) near Clinton, Southland (Fig.24), contains brachiopods of the Spinomartinia spinosa Zone, and elements of the fauna are found widely throughout the South Island, in the Little Ben Sandstone near Nelson City, the upper Annear Sandstone of Eglinton Valley, and Pine Bush Formation of Mataura Island, Southland, the fauna being one of the most widespread of Permianfaunas in New Zealand (Table 3 7, Fig. 24, 3 1 ). Crurithyris suggests a direct link with the Urushtenia.n, Ruteh 3 and Kathwai faunas, as confirmed by stratigraphic position. Martinia cf mongolica Grabau supports correlation with the upper beds of Mongolia described by Grabau ( 1 93 1). Atomodesma trabeculum Waterhouse is found also in New Caledonia (Waterhouse, 1967b). The overlying Greville Formation has scattered occurrences of the ammonoid Durvilleoceras woodmani Waterhouse 1 973b probablyof Baisalian age. '
In the Gympie Basin of Queensland (Fig. 2 1), the Tamaree Formation contains a gastro�od figured as P!atyteichum sp. by Ru:�negar & Ferguson ( 1 969, pl.3 , fig. 1 5- 1 6)that m1g�t b� A nanazs campbelli Waterhouse. This species is characteristic of theUrushteman m New Zealand, and occupies a similar stratigraphic position in Queensland (Table 36).
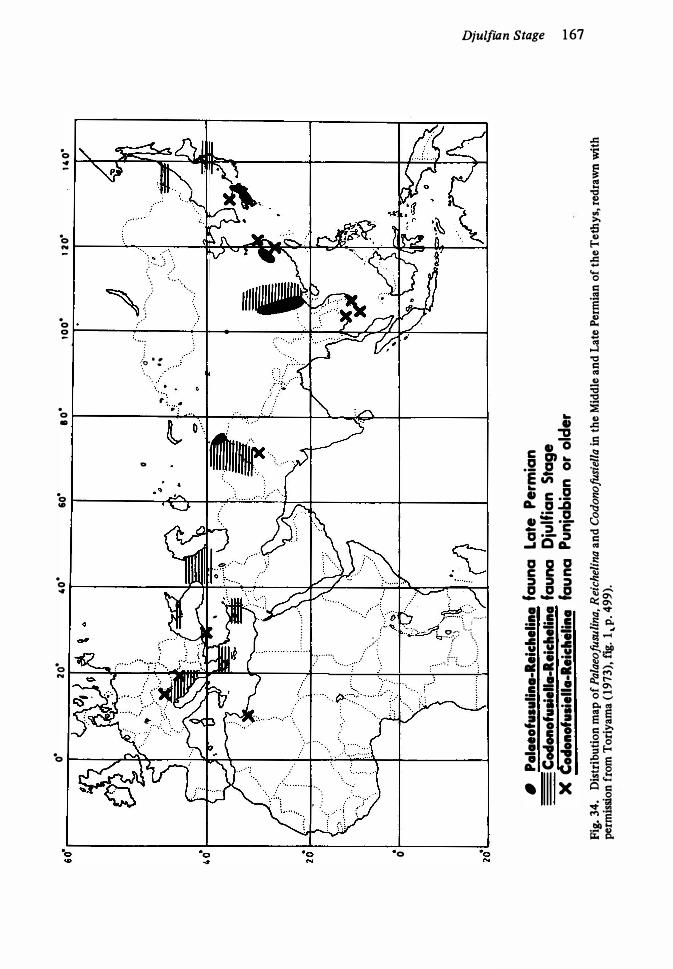
o•
zo•
40•
so•
ao•
1oo•
12
0·
140.
60
. 1 .
_
I �
r'l .rJ
. ) I
.u
ft I
I I
I· I
I I
0 . .. ··
a· ...... !
· ... f
.... , 11
. · ... ·
.....
. , · ...
.... ···. ,:" •
= --· · .
.. �-.. �� ·:�
...1 �� f�t,
{': \l-
1,(2$ I
· .. ·�
.1.:
, . . ·
... •··.
2 01 r.
·· , . .. : . . .
.. '= 1
. . -�··· · \:······· ·1
·· . .".1=· ..... �
71
\
vr-
\D
... r · �O
J)·
I
I · ·�
. . /.�
. ...-···· l
· \ . . ... · � ·.:
· 1
I f. ·'
>. '
I .n.
.. P /
I
I ')��
c:
··· ��
�
:;:J
I 0
l ..
: i',.(
' ··
lT
"\ �
Ri
a�.
;::!·····
<:�, ... �� : :· ······
2o•l
I '
[""::.. . .. ..... ··
··· .. { · ··
/1
\ I
I I
I I'
I
'-I
•
Palae
afus
ulina
-Reic
helin
a fa
una
==
= Codon
ofusi
ella-R
eiche
lina
fauna
)(
Coclo
nofu
siella
-Reic
helin
a fa
una
Late
P
erm
ian
Diu
lfia
n S
tag
e
Puni
ab
ian
or
old
er
Fig
. 34
. D
istr
ibut
ion
map
of
Pal
aeofu
sulin
a, R
eich
elin
a an
d C
od
onofu
siel
la in
th
e M
iddl
e an
d L
ate
Perm
ian
of t
he
Teth
ys,
red
rawn
with
pe
rmis
sion
fro
m T
oriy
ama
(197
3), ftg
. l,
p. 4
99).
� � s;
E; · ;::s t'.l
�
.....
0\
-...J

1 68 Biochronology for the Marine Permian
DORASHAMIAN STA GE
Vedian and Ogbinan Substages
Armenia, Iran
The Dorashamian Series of Rostovtsev and Azaryan ( 1973) includes the upper part of the former Djulfian Stage or Series, and is here treated as a stage, with two or three substages, the V edian at the base, the Ogbinan, and perhaps the Griesbachian, or part thereof, at the top. Two distinct faunas, here treated as substages, were recognised for the lower part by Waterhouse ( 1972a} on the basis of work by Ruzencev & Sarytcheva ( 1965} from sequences of south Armenia, and bv Stepanov et al. ( 1969) in northern Iran. The type section ts offered by Dorasham Siding II for the V edian Substage (see both papers) and for the Ogbinan Substage by"the Djulfa Gorge (Waterhouse, 1 972a) though Rostovtsev & Azaryan ( 1973) suggested the Dorasham II section. The stage has been referred entirely (Stepanov et al., 1 969), or partly (Ruzencev & Sarytcheva, 1 965 ; Glenister & Furnish, 1 96 1 , 1 970) to the Triassic, in the face of a Permian age clearly indicated by brachiopods, as sustained by Y akovlev ( 19 3 1 ), Chao ( 1 966), Waterhouse ( 1 967a, 1 969b) and Tozer ( 1969}.
The Vedian Substage is exposed in red shales and dark red limestone up to 5 mthick (Table 45). It is characterised by the ammonoid Phisonites, and the brachiopod genera Comelicania and Janiceps. Phisonites and Comelicania are also found across the border in Iran in beds 5 m thick (Stepanov et al., 1 969). Perhaps only the brachiopods will prove diagnostic, for Rostovtsev & Azaryan ( 1973, p.9 1) considered that Phisonites was only a compressed Xenodiscus.
The Ogbinan Substage has species of brachiopod genera Orthotichia, Enteletes and Araxathyris with characteristic ammonoids Dzhulfites and Paratirolites. Rostovtsev & Azaryan ( 1 973) synonymised Dzhulfites with Paratirolites. Pleuronodoceras, Pseudotirolites and Pseudostephanites were recognised by Teichert et a!. ( 1 973) in the late Ali Bas hi Formation of Kuh-e-Ali Bas hi, Iran. Somehow Teichert eta/. ( 1973) failed to find the critical Phisonites-Comelicania Zone, even though it was recognised in the area by Stepanov et al. ( 1969), to imply that the ranges of the species they did find lack a rigorous stratigraphic framework.
Much the same faunal sequence is found at Abadeh in central Iran (Taraz, 1971 , 1 973, fig.3, p. 1 1 28), with Dzhu/fites (Vedian) beds overlain by beds with Paratirolites (Ogbinan).
Europe
A limestone fauna in the upper Permian of the Bukk Mountains of Hungary (Shreter, 1963) has Martinia and Comelicania, with many Djulfian species as well, including Licharewinids and even Crurithyris. Whether collections have spanned some interval of time, as seems likely, or come really from only one zone cannot be ascertained but Assereto et al. ( 1 973) considered these faunas were mostly older than the faunas of the Carnian Alps. The same key brachiopods Comelicania and Janiceps are found in the upper Bellerophon Limestone (Table 24) of the south Tyrol close to the Italian, Austrian and Yugoslav borders (Stache, 1 878 ; Diener, 1 9 10; Merla, 1 93 1 ; Accordi &Loriga, 1 968 ; Assereto et al., ( 1 973). These faunas are accompanied by a very large molluscan element, including Paraceltites sextensis Diener, but chiefly gastropods and bivalves (Gortani, 1906a, b, etc.).
China, Japan
The Dorashamian Stage is represented in south China by the Changsing

Dorashamian Stage 1 69
Limestone (Table 1 8), a light grey well bedded limestone 25- 100 m thick in Chekiang, north-west Kweichow, Kwangsi and Szechuan, above rocks of Djulfian age (Sheng; 1964; Chao, 1 966}. Palaeofusulina is a predominant foraminifer and may be accompanied by Codonofusiella (Wang, 1966). Pseudotirolites and Pseudogastrioceras occur in the lower part, and Changhsingoceras, Trigonogastrites, Stacheoceras and other genera in the upper part. There are many brachiopods, few obviously distinctive, and many like those of the underlying Wuchiaping fauna, including Anidanthus sinosus (Huang) and ?Rugaria soochowensis (Chao). Sheng & Lee ( 1974) listed Crurithyris speciosa Wang and Araxathyris ·araxensis Grunt. Other ammonoids and brachiopods are found in the limestones, shales and sandstones of the Talung Formation in south China in Kwangsi and southern Kuiechow. The Talung Formation is either correlative with or slightly younger than the Changsing Limestone. Further ammonoid species were Ilsted 6y Chao (I966) in northern Szechuan. Palaeofusulina (Fig. 34) is also found in the Liangfengpo Shale in western Kueichow. No fauna clearly correlative with the Vedian Substage has yet been �cognised, but stratigraphic and faunal relationships are not under satisfactory control, so that it is not certain whether the Vedian Substage is missing, or has not been recognised. Indeed there is the possibility that it is represented by the upper Changsing Limestone. This contains the Cyclolobid ammonoid Changhsingoceras Chao, which is related to Godthaabites and Cyclolobus, genera that are on the whole typical of temperate latitudes. So the presence of Changhsingoceras in south China, which occupied the palaeotropics during the Permian (Waterhouse & Bonham-Carter, 1972, 1 975), might indicate Vedian temp<?ratures which were cooler than normal. This might be supported by the record of Eurydesma inflatum (author not stated} in Sheng & Lee ( 1974), for Eurydesma is otherwise restricted to faunules known to have been cold-water.
Faunas of Japan may be correlative but there are problems, for only Fusulinacea are abundant, and those of late Permian age are survivors from earlier faunas. However, Palaeofusulina and Codonofusiella occur in the lower to middle part of the Mitai Formation (Kanmera & Nakazawa, 1 973, p. 107) and may well be lower Changhsingian, whatever that be.
The Gujo bivalve fauna, described by Nakazawa & Newell ( 1 968) is probably Vedian in age because a number of the bivalves which dominate the fauna are obviously closely related to bivalves from the upper Bellerophonkalk of Europe, as first pointed out by Waterhouse ( 1 969b). Few brachiopods are present (Shimizu, 196Ib). Table 48.
Madagascar, New Zealand
The ammonoid Paratirolites has been found in beds of otherwise uncertain age in north Madagascar (Tozer, 1 969). Near Nelson City, New Zealand, (Fig. 24, Table 37), the local Waiitian faunas belong to the Vedian Substage, with brachiopod faunas described by Waterhouse ( 1964, 1 967a). The faunas occur in extensive carbonate lenses varying from a few metres to 700 m in thickness, above the Greville Formation with the Baisalian ammonoid Durvilleoceras. The fauna is correlated chiefly by stratigraphic position and evidence of cooling, for no well dated faunas of comparable age are known from such high latitudes in the late Permian. However a species of the gastropod, Spirovallum Waterhouse is close if not identical to a species from the Gujo fauna of Japan (Waterhouse, 1 969b). Underlying red and green shales and siltstones probably belong to the same stage, and were apparently deposited in shallow cold waters. Overlying beds include tilloidal rock, and slump breccias suggestive of sharply lowered sea level (Burke & Waterhouse, 1 973}, implying glacio-eustatic influence.
Breccias and carbonates with scattered fossils from the Takitimu foothills, Southland, New Zealand, in the Hawtel Formation, are likely to be correlative. No wellestablished Ogbinan fauna is found in New Zealand.
Waterhouse ( 1 969b) drew attention to possible tillite in the upper Ferntree Form

Ei r.. 0 Ul · nt � ·.-I :I:
-- &: � .0 �
_E-<_
f 111 1-1 :::1 ,l( 0 � :.I
So
ut
h-
we
st
Ja
pan
N
or
the
as
t
Ja
pan
I
nne
r
Zo
ne
0
u
t
e
r
z o
n
e
Ki
tak
ami
M
ai
zu
ru
C
hih
ib
u b
el
t
Kum
a
Ts
ukum
i T
ak
ac
hi
ho
Tr
ia
ss
ic
-
--
--
--
--
--
--
--
--
--
-L
eiop
hy U
i te
s N
uau.
Z.ana
nog
amii
(?)
. &:
&: 111
. t-
--
--
--
-0
1--
--
--
--
--
--
�
Mee
koce
ras
§
Pse
udo
sagec
eras
�
Mee
koae
ras
� Me
ekoc
eras
:.I
Bi
va
lv
es
--
--
--
--
><
--
--
--
--
--
--
Gl.y
pto
phi
aera
s N
eosa
hizo
dus
CZ.ar
aia
Bakc
wie
ZZ.a
�
.
G
uj
o
(fo
ss
il
s
no
t
Sta
ffe
Ua
S
taff
el.Z
a
fa
una
f
oun
d)
Ql
Nan
kine
ZZa
N
anki
neU
a
s:: L
epto
dus
P
aZae
ofus
uU
na
0
--
--
--
--
--
--
--
--
--
.j.l
Pa
l.aeo
fusu
Un
a
sin
ensi
s Ul
CoZ.an
ieU
a
m c
f
sin
ensi
s Ei
cf
si
nen
sis
r..
·.-I
Reia
hel.i
na
f-
--
--
--
-r-
--
--
--
--
-�
·.-I Le
pido
Zin
a
ahan
ghsi
ng
ensi
s 111
PaZ
aeof
us
min
ima
Codo
nofu
sie
Ua
.B
.j.l
chan
ghsi
nge
nsi
s
rrruZt
isep
tata
Re
iche
Un
a
Cod
onof
usi
e U
a
•iJ Co
dono
fus
ieZZ
a
--
--
--
--
Lep
idoZ.i
na
kum
aen
sis
� 3
kuei
ahow
ens\
s ku
eiah
owen
sis
Ul L
epido
Un
a
Codo
nof
usie
Ua
Le
pido
Un
a
E-t &
!mu
Ztis
epta
ta
e kum
aen
sis
shir
aiw
ensi
s :::1
Codo
nof
usie
l.Za
�
N
Reic
he U
na
.
.
· .-I f-
--
--
--
--
--
-�
Y
abei
na
gZ.o
bosa
Y
abei
na
ka
toi
� .
Neo
sahw
ager
ina
Lo
ph
op
hy
ll
oi
d
co
ra
l
&: Y
abei
na
gZo
bosa
a
kato
i r.
. m
arga
ritae
� _
__
__
__
__
_,
--
--
--
--
--
0 -
--
--
--
--
.j.l
Neo
sahw
afJel'i
na
Neo
schW
afJel'
ina
111
§ 3:
Neo
sahW
O!Jel'i
na
><
ma
rgar
itae
H
-T
abl
e
48
. c
or
re
la
ti
on
o
f
Up
pe
r p
ar
t
of
Pe
rm
ian
, an
d Lo
we
r
Tr
ia
ss
ic
o
f J
ap
an
and
so
me
ot
he
r
ar
ea
s
of
t
he
T
et
hy
s
re
gi
on
b
y
di
ag
no
st
ic
g
en
er
a
or
s
pec
ie
s.
(Fr
om
K
anmer
a
& N
aka
zaw
a,
1973
, p
. 11
5,
fi
g.
6.
)
f
-f
au
lt
F
m -
Fo
rma
ti
on
G
p
-G
ro
up
- -.J
0
b::l c;· (";
::::- � ::s
c
B' � � ....
...
::::
� � .... s· � � � E;· ::s

Dorashamian Stage 1 7 1
Fig. 35. Present geographic distribution of the ammonoid genus Otoceras, emended from Kummel (1973, fig. 5 , p. 568) .
Metres
.-�--------------------��------� c )
•
( • I •
"Palaeozoic"' Brachiopods )
"Triassic" Ammonoids
�����-+�-----------------------i A . :
. . . ·J . . . . .
. :
:
·
.... �:> . . . . . . . . .
· . . . · · •' . : . . . .
. r
Guryul Ravine, Kashmir
Fig. 36. Permian - Triassic boundary beds in Guryul Ravine, Kashmir. A, the boundary drawn by Nakazawa et a/. (1970) below the first occurrence of Claraia stachei. B , an alternative boundary at the base of a stratum bearing the Otoceras - Ophiceras assemblage; C, boundary favoured by NeweU (1973) and the writer (Waterhouse, 1973), at the top of the ammonoid genus Otoceras with Ophiceras transitional between Permian and Triassic. Dots represent single occurrences of ammonoid genera. Redrawn with permission from NeweU (1973, f"J.g. 3, p. 7).

1 72 Biochronology for the Marine Permian
Group and to the possible presence of Tomiopsis like a late Permian species from New Zealadd, and supposed Eurydesma. Clarke ( 1 973) strongly d�ubted the presence of tillite or glacial sediment, but other accounts record at least tee-rafted pebbles (Jago, 1972). Unfortunately we still await clarification of the Ferntree correlation. Clarke (1 973) asserted most strongly that the fauna was identical in age with that of the late Mal bin a Formation, but his lists proviqe little indication of any positive age or basis for correlation. Not even a refined age for the Malbina E is provided, other than 'Kazanian', which in the terminology of Clarke ( 1973), following Runnegar ( 1 969b), ranges from Nevolinian (Kungurian Stage) to Vedian (Dorashamian Stage).
Dulhunty & Packham ( 1 962) have described glacial sediments near Mudgee, New South Wales, regarded as correlative with the late Middle Permian Capertree Group = Wandrawandrian beds or even ?Late Permian Lithgow Coal Measures.
Griesbachian Substage
Rocks and faunas generally referred to the basal Triassic, or what used to be called the Scythian Stage, contain Permian-type brachiopods, associated with the ammonoids Otoceras and Ophiceras, and the bivalve Claraia (Waterhouse, 1 973b). The basal unit of the Scythian, called the Griesbachian Stage by Tozer ( 1967), based on outcrops and faunas of the Canadian Arctic Archipelago, should almost certainly be placed in the Permian Period where we use faunal affinities rather than historical precedence as the guide. Otoceras itself at the base of the Griesbachian Stage is distinctly Permian in affinities, and even Ophiceras is morphologically close to the Permian genus Durvilloceras Waterhouse ( 1 973b). Waterhouse ( 1973b) suggested that the boundary should possibly be drawn in the Dienerian Stage, but this is subject to further enquiry.
Kozur ( 1 973a) has argued strongly that the Otoceras faunas, or basal Griesbachian or Tozerian Stage should be treated as topmost Permian. The overlying upper Griesbachian and Dienerian Stages were referred by Kozur ( 1973a) to a new Brahmanian Stage, typified by ammonoids of Triassic aspect, including Ophiceras. In 1974 Kozur showed the top of the Permian to be represented by the Otoceras concavum and 0. boreale zones, underlain perhaps by the Hypophiceras martini and H. triviale zones of Greenland, in a Gangetian Substage, above the Dorashamian. The zone of Ophiceras commune was assigned to the Ellesmerian Substage at the base of the Triassic Period. Newell ( 1 973) has suggested drawing the boundary at the top of the Griesbachian Stage, that is, taking in the Ophiceras as well as Otoceras beds (see Fig. 36).
Clearly the decision must be by international agreement, after general discussion. What is apparent is that we may now delineate an overall geological picture after a decade of confusion caused by the allegation that 'Permian-type brachiopods' survived only locally in rocks crudely dated as Triassic or 'Scythian' by Kummel & Teichert ( 1 964, 1 970, 1 973) and their colleagues. The late Permian�arly Triassic involved a succession of faunas and geological events, commencing with world-wide basal Griesbachian, or Gangetian transgression after long emergence, with faunas characterised in part by Otoceras, closely related to Permian Otoceratidae, the bivalve Claraia, closely related to the Permian genus Pseudomonotis, and various brachiopod families, identical with preceding Permian families. The later Griesbachian apparently saw the disappearance of Otoceras, and supposed incoming of Ophiceras and otherwise little change amongst the macro-invertebrates. During the Dienerian 'Stage', the ammonoids developed from upper Griesbachian forms. Brachiopods changed drastically within the Dienerian, as far as can be ascertained, from those of Permian type, to those of �h� more restricted Triassic type. Overlying ammonoids and brachiopods of the Smtthtan Stage were typically Triassic in appearance. The Smithian ammonoids are obviously close to
_Dienerian and Griesbachian forms, whereas the Smithian brachiopods
represent an ennchment of only some of the families present in Griesbachian and possi-

Smithlan Stage and younger ? Diener ian aubatage
Doraahamian Stage
Glendale Li maatona
Dorashamian Stage 1 73
fzs
Punjabian Stage
Kazarian and earlier Permian
Fig. 3 7. Permian - Triassic boundary beds in the foothills of the Takitimu Mountains, Southland, New Zealand, redrawn from Waterhouse (1973d, fig. 6 , p. 454). Localities in the Wairaki Breccia (black spots) contain ?Dienerian brachiopods, and are overlain by rocks and faunas of Smithian age. No Griesbachian faunas are known.
Cape Stallworthy
Dienarian
Upper Griesbachian
lower Griasbachian
-unconformity -
tranian
Griesbach Creek
r.-.-.-. .-.-.-.-.-.-.-.-;·:.1 Svardrupi Zona
(:::::�::::::::::::::::::] Red and araan sandstone
Silhtona Commune ad
;• trigatvs zonas
Shale
Concavum and boreal• z.onas
Van Hauan chart
Fig. 38. Stratigraphic columns of the Blind Fiord Formation on Axel Heiberg Island, Canadian Arctic Archipelago, traditionally regarded as basal Triass!c, but here considered Late Permian in part. Redrawn from data supplied by Dr E. T. Tozer in Waterhouse ( l 972b).

1 74 Biochronology for the. Marine Permian
ble early Dienerian faunas ; other typically 'Palaeozoic families' have essentially vanished.
Faunistically one may argue for placing the boundary at the incoming of Ophiceras, or the incoming of Triassic brachiopods especially in the Smithian Stage. Or we may emphasise the disappearance of Otoceras, or disappearance of Permian brachiopod families. From the viewpoint of mapping a boundary, the base of the Griesbachian, or base of the Smithian are generally easy to recognise. The last appearance of Otoceras is particularly difficult to ml:!,p, judged from my own experience in the Himalayas. Otoceras was very widespread, but is comparatively rare in various regions, possibly because its shell was easily destroyed. The ammonoid data summarised by Waterhouse ( 1 973b) from Tozer ( 1 967, 1 97 1) suggests that the commencement of the Smithian Stage would offer a good and ·easily recognisable base for the Triassic Period, but it now appears that the ranges of a number of ammonites have to be extended downwards (Kozur, 1 973a, b; Dr E.T. Tozer, Geological Survey of Canada, pers. comm.).
In the meantime, only one modest brachiopod fauna has been completely described, by Waterhouse ( 1 967a) from the Wairaki Breccia, Takitimu Mountains, Southland (Fig. 3 1 , 36), above Vedian rocks and faunas, and below Smithian (i.e. upper Scythian) faunas (Table 37). It is thought by Waterhouse ( 1973b) to be as young as Dienerian, in view of the absence of key Permian families, but its genera are still of Permian type, so that faunistically it possibly lies right at the top of the Permian, or base of the Triassic : historically it is Triassic. Other modest brachiopod faunules are found, none described fully. In the western United States, the Dinwoody Formation with Claraia (Newell & Kummel, 1 942) contains numerous scattered brachiopods including Leptodidae, Overtoniidae, Ambocoeliidae and Dielasmatidae, with other taxa (Waterhouse, 1 973b; Grant & Cooper, 1 973), some misidentified as Triassic by Kummel (in Newell & Kummel, 1 942).
Another extensive fauna has been found in north-west Nepal by the writer in 1973, with Orthotetacea, Chonetidae, Strophalosiidae, Productacea, Rhynchonellida and Ambocoeliidae. All await determination and description. They are found with numerous 'early Triassic' ammonoids, including rare Otoceras (Fig. 30).
Other approximately correlative faunas are scattered through the Himalayas, as reviewed in Waterhouse ( 1973b), including briefly documented occurrences in the Pahlgam-Aru basin, and in Indochina (Waterhouse, 1 973b, p.3 1 1 ).
The topmost zone of the Zewan sequence in Kashmir, named the lamellibranch zone by Diener ( 19 1 5) (Fig. 3 1 ), or Zewan 6, is found two miles north of Barus and in the Guryul Ravine. Diener ( 1 9 1 5) recorded Xenodiscus cf. carbonarium and so-called Pseudomonotis and Eumicrotis that might prove to be Claraia with such typically midPermian brachiopods as Lamnimargus himalayensis and Costiferina with Lissochf!netes. Nakazawa et al. ( 1970) listed other Permian brachiopods, including 'Pustula', with Claraia and Griesbachian ammonoids such as Ophiceras. We await description of the fauna to see if it was reworked, at least in part, from underlying Zewan faunas, or represented new Griesbachian brachiopods of Permian aspect. Furnish et al. ( 1 973) have shown that so-called Xenaspis cf: carbonaria is likely to be an indeterminate Triassic ophiceratin, possibly Glyptophiceras, and have considered that socalled Popanoceras of Diener ( 1 9 1 5) is a Triassic genus.
Permian Overtoniid brachiopods occur at comparable horizons in Japan (Nakazawa, 1 97 1) and the Canadian Arctic (Waterhouse, 1 972b) (Fig. 38) and Rhynchonellids are found in Spitsbergen, Greenland. A few species are reported from the Werfener beds of Austria and especially Italy (Sherlock, 1 955). Here the barren Teser? Horizon of oolitic limestone and marly micrite up to 6 m thick is overlain by the Mazz1� Member of marly micrite 30 - 70 m thick, with Bellerophon vaceki Bittner, also found m the Otoceras beds of Shalshal Cliff; and conodonts Anchignathodus typicalis

Dorashamian Stage 1 75
Sweet and Ellisonia teicherti S�eet that are especially typical of Griesbachian faunas. Claraia enters the overlying Siusi Member, and was assumed to indicate an upper Griesbachian age by Assereto et al. ( 1973, p. 1 90). Kiparisova et al. ( 1 973, p. 14 1) recorded Crurithyris and Fletcherithyris with such bivalves as Atomodesma in the early Triassic of the Soviet Union, without specifying locality.
Various faunas are discussed by Tozer ( 1967). Dickins & MacTavish ( 1 963) recorded 'Otoceratan' fosils in the Kockatea Shale of the Perth Basin.
Conodonts are of high promise for recognition of the Otoceras and allied horizons, but the exact range of key species is not clear. Anchignathodus typicalis Sweet and Ellisonia teicherti Sweet are significant species of the lower Kathwai Member of the Salt Range, that- range down into the overlying white sandstone at the top of the Chhidru Formation (Sweet, 1 970b). These species are also found near the Permian boundary in Guyrul Ravine (Sweet, 1 970a), and in the matrix of Otoceras specimens from Spiti. But, as shown by Grant & Cooper ( 1973), the range at Guyrul is so great that the species appear to be both Permian (apparently Middle Permian) and Triassic, wherever the boundary is drawn, and species with shorter ranges would be more useful. Sweet ( 1973) has also recorded the two species in the Djulfian Ali Bashi Formation of north-west Iran, so that the species clearly range from the Punjabian to late Dorashamian.
Note added in proof.
During September, 1975, I attended the Carboniferous Congress at Moscow, to learn that the "Orenburgian Stage" was under severe criticism from stratigraphers and palaeontologists. Some would deny its validity as a unit, others maintained that it formed the base of the Permian System as a distinct unit, or as part of the Asselian Stage.
It is a pleasure to draw attention to the text Gondwana Geology, Australian National University Press, 1975, edited by Dr K.S.W. Campbell. Amongst other articles of value, M.J. Clarke and M.R. Banks in their article "The stratigraphy of the Lower (Permo-Carboniferous) parts of the Parmeener Super-Group, Tasmania" (pp.453-467), have listed faunal distributions for the Permian of Tasmania. Moreover, Runnegar and McLung ( 1975, p.43 1) in the article "A Permian time scale for Gondwanaland" (pp. 425-44 1), have abandoned attempts to subdivide the east Australian Permian through bivalves, and used brachiopods instead, following Campbell ( I 953), Maxwell ( I 954), Dear ( I 971 , 1972) and other workers. They have thus provided valuable data in support of the refined classification in this text.

5. Summary of Permian Stages and Substages: Problems and Proposals
Asselian Stage (Table 49) As summarized in Chapter 3 , difficulties still beset the base of the Permian Period,
because it may be fixed at various levels. The full range of critical Fusulinid genera, Schwagerina, Pseudofusulina, and Pseudoschwagerina has not been established and the problem is exacerbated by nomenclatural confusion. Yet we have been forced to rely on Fusulinacea, because the 'type Asselian' abounds in these fossils, and they have been closely studied. The Fusulinacea show a three-fold zonation which appears to be reflected widely by Fusulinacea and Brachiopoda over the rest of the world. But many sequences lack Fusulinacea, and the brachiopod faunas, though widespread and moderately well-known, cannot be accurately matched with those of the Urals, because there have been few modern Russian studies. The solution to most of our problems for the early Permian lies in the provision of brachiopod studies from the type and nearby Asselian Stage. Key brachiopod genera may include Kochiproductus, MuirwoodiaYakovlevia and perhaps Attenuatella. The genus A ttenuatella is especially widespread, but has been reported from deposits of Spain, Austria, and Verchoyan which may prove to be late Carboniferous. Ammonoidea play a curious role in defining the Asselian Stage. Although mid and late Asselian genera and species are characteristic, early Asselian, or Surenan Ammonoidea differ little from underlying Ammonoidea (Ruzencev, 1 952), and so suggest that the basal Asselian may differ little from the Orenburgian, unless the Ammonoidea failed to change with the rest of the faunas. This means that the base of the Permian has to be assessed from other fauna, particularly Fusulinacea and Brachiopoda. We still wait for this clarification.
Marine rocks of the Asselian Stage were widespread in all continents save the Antarctic, and record one of the most widely distributed glacial episodes known, in the late Asselian Kurmaian Substage. Faunal evidence strongly suggests that the period commenced with Gondwanan glaciation, and relicts of such glaciation are possibly found in eastern Australia, with rare fossils, though the beds have usually been interpreted as Carbc�;�erous (eg. by Runnegar, 1 969b).
In· ervening beds and faunas, and world-wide correlative faunas suggest warm conditions,. so that the Asselian may record two glacial episodes, with an intervening warm episcd;. If so, the Permian Period, although at first sight arbitrarily delimited, may have commenced at the onset of very widespread climatic cooling that must have strongly affected rocks and life more or less simultaneously (within the limits of monitoring Permian time), as the first extensive glaciation to follow the early Moscovian or Bashkirian glacial episode (see Bamber & Waterhouse, 1 97 1 ).
Sakmarian Stage (Tables 49, 50)
Fusuline, brachiopod and ammonoid species and genera delineate three substages for the Sakmarian Stage, in good and widespread agreement. The only question lies in treatment of the Aktastinian Substage, which is classed with the younger Baigendzinian Stage on ammonoid evidence, and with the Sakmarian on brachiopod evidence. Fusuline authorities initially supported the latter position in Russia and now follow th� .former classification. The difference may surely be resolved by discussion, and is not cnttcal : I have based the assessment on brachiopods because they were most

Sub
st
ag
e
Fu
su
li
na
ce
a
Br
ac
hi
op
od
a
Bi
va
lv
ia
Am
mon
oi
de
a
Ta
st
ubi
an
Rug
oso
fusi
lin
a s
pp
. f
ir
st
Sp
irif
ere
lla
s
ta
rt
o
f
Syn
arti
nsk
ia
Pse
udo
fu
suli
na
s
pp
. p
seud
odr
as
oh
ei
Me
dU
oot
tia
f
ir
st
S
. s
alt
eri
Me
tale
goo
era
s N
oto
spir
ifer
Ur
alo
oer
as
Pr
opop
anoo
er
as
Ku
rma
ian
S
ohw
ag
erin
a
wi
de
sp
re
ad
E
ury
des
ma
f
ir
st
Sa
kma
ri
tes
Pse
udofu
suli
na
sp
p.
At
ten
ua
te U
a
Delt
opeo
ten
T
aban
ta
U t
es
Tom
iop
sis
P
rot
opop
an
ooer
as
Us
ka
li
ki
an
S
ohw
ag
erin
a
Or
tho
tio
hia
f
ir
st
J
ure
sani
tes
Su
re
nan
Pse
ud
ofus
uli
na
sp
p.
Ku
tor
gin
ell
a
Pa
ra
ga
str
iooe
ra
s
Soh
wa
ger
ina
f
ir
st
K
ooh
ipr
odu
otu
s E
uryde
sma
G
la:phy
rite
s
sp
.
Pse
udof
usu
Zin
a
Ya
kov
lev
ia
Pse
ud
oso
hwa
geri
na
r
ev
iv
al
o
r
fi
rs
t
At
ten
ua
teU
a
spp
. r
ev
iv
al
To
mio
psi
s
Tab
le
4
9.
So
me
si
gn
if
ic
ant
m
ar
in
e
in
ve
rt
eb
ra
te
g
en
er
a
an
d
sp
ec
ie
s
fo
r
Ea
rl
y
Pe
rmi
an.
I
� ;!i ;!i � � � � � s · ;::s �
� � ....
-...)
-..l

Sub
stag
e
Kr
asn
ou
fim
ian
Sa
rg
inian
Akt
ast
in
ian
St
er
li t
amak.i
an
Fu
suli
nac
ea
A�as
kan
eU
a
yu
kon
ens
is
?�s
e�
�in
a
a�a
ud
iae
?Pa
raf
usu
Zin
a
so
lid
is
si
ma
Pse
udo
fu
su
Un
a
mak
aro
vi
Pa
ra
fu
su
�in
a
spp
.
Mo
no
die
xo
din
a
spp
.
en
try
o
f
Pr
aep
ar
af
us
u�i
na
�u
tug
ini
Sa
hwa
ger
ina
ar
as
si
te
ato
ria
Ps
eu
do
fu
su
�in
a s
pp
.
Pse
udo
fu
su
Zin
a
Br
ach
iop
od
a
Ea
hi
na
�os
ia
pr
ider
i
Au
�os
teg
es
ing
ens
Ura
�op
rod
ua
tus
stu
ak
enb
erg
ianu
s
Sow
erb
ina
fa
una
Cho
ne
tin
a a
rti
en
si
s
Wyn
dham
ia j
uke
si
In
st
it
eU
a
An
tiq
ua
ton
ia
faun
a
MUiPW
oo
dia
ar
tien
sis
Sp
irif
ere
Ua
sa
ran
ae
Sa
aa
ah
ine
�la
cho
net
el
lid
s K
ro
tov
ia
m
arg
in
ife
ri
ds
Ste
pan
ov
ie
��a
fle
xuo
sa
Ti
ty
rop
ho
ria
M
ar
tin
i a
Ea
hin
a�o
si
a p
re
ov
ali
s
Re
edo
aon
ah
a
An
idanth
us
spr
ing
su
ren
sis
At
ten
ua
teU
a
Pun
ato
ayr
tel
la n
ag
ma
gen
si
s
To
miop
si
s
Biv
alv
ia
Eu
ryde
sm
a
Ammo
no
ide
a
Wa
age
nin
a
spp
.
Me
d�i
ao
tti
a
spp
.
Per
rin
it
es
Pr
opin
aa
oa
era
s
Sa
kma
ri
te
s
spp
.
Pr
oper
rin
ite
s b
oe
ss
i
Ag
ath
iaer
as
spp
.
Ak
tub
in
sk
ia
Tab
le
SO
. S
ome
s
ig
ni
fi
can
t
ma
ri
ne
i
nv
er
te
br
at
e
ge
ne
ra
an
d
sp
ec
ie
s
of
t
he
E
ar
l.y
P
er
mi
an
Pe
ri
od
.
- -.J
00
t.:l ;:: �
�
1::1 q � � ....
�
l:i" ;:s t.:l
.... � �
..
.

Summary of Permian Stages 1 79
widespread. Faunal difficulties that beset Sakmarian species are much the same as those interfering with our understanding of the Asselian Stage. Fusulines have nomenclatural problems, and ranges vary locally. Brachiopoda have not been recently monographed from the type Sakmarian and there are no widespread typical new genera, and Ammonoidea show changes slightly out of phase with benthos, in that the lower Tastubian beds of the Urals have few outstanding species and no new genera, and the Sterlitamakian ammonoids are not very distinct. But these difficulties appear to have been overcome, to judge from the widespread agreement over correlation.
Sakmarian marine rocks and faunas are almost as widespread as those ot Asselian age. They are more definitely established in north-east Siberia, but somewhat less extensive in parts of Gondwana, notably South Africa and South America and central Asia.
Faunal evidence based on the appearance of Licharewia, Tomiopsis, Attenuatella, and the bivalve Eurydesma in the Sterlitamakian interval suggests a mid-Sakmarian cooling, but evidence for this in eastern Australia is restricted to occasional ice-rafted boulders, with no well developed tillite or varves as far as I am aware. Tillite has been reported from Sakmarian beds (sl.} in Verchoyan (Ustritsky, 1 973). It appears that the Sakmarian Stage witnessed extensive cooling during the Sterlitamakian Substage, but was on the whole warmer than the Asselian Stage, leading to the return and proliferation of some late Carboniferous genera, such as Choristitinids. Reduced glaciation allowed isostatic emergence of parts of Gondwana, leading to terrestrial conditions and formation of some coal measures.
Baigendzinian Stage (Table 50}
The Baigendzinian Stage delimits a distinctive phase of earth history, marked by withdrawal of the seas from extensive regions, especially over Gondwana, with the development of widespread coal measures and terrestrial sediments. Coarsening of sediments or unconformity also occurred in tropical and northerly realms, including the Glass Mountains of Texas, and northern Yukon of Canada. The climate appears to have been relatively warm, with evidence of glaciation restricted to Tasmania, which then lay very close to the South Pole (Waterhouse, 1 974a). Under warm conditions, Fusulinacea became widespread, and new lineages developed amongst Brachiopoda.
Kungurian Stage (Table 5 1}
Because of recent Russian studies the Kungurian Stage is relatively well controlled for faunas and rocks of the northern, Arctic, and southern hemisphere. That there has been controversy over the stage cannot be denied, but this has stemmed partly from the paucity of ammonoids in the type Kungurian, and partly from a historical development that failed to understand the uniqueness of Kungurian faunas. It is now well recognized by many authorities that the Kungurian Stage saw the revival of many early Permian genera, and entry of many new and widely dispersed brachiopod species, coinciding with the appearance of new Fusuline genera in the palaeotropics. These new faunas coincided with a new widely transgressive phase that commenced during the basal Kungurian and continued until the late Kazanian Stage. Understanding of critical Fusulines and Ammonoidea is still incomplete. On the whole, evidence suggests that Neoschwagerina craticulifera may be as old as Irenian, or upper Kungurian, and that Waagenoceras may be as typical of Irenian faunas as of the Kazanian. In earlier literature these forms are widely regarded as 'Wordian', a very broad and long-ranging time span, or sometimes as Kazanian, which may be imprecise.

Sub
st
ag
e
Fu
sul
in
ac
ea
B
ra
ch
iop
od
a
Bi
va
lv
ia
Am
m::moid
ea
So
sno
vi
an
?Ne
osah
wag
erin
a
Grum
anti
a p
eli
aanen
sis
Stu
tahb
ur
ia
mar
gari
tae
?smo
ot
h
ch
one
ti
ds
T
omio
psi
s a
osta
ta
Ka
li
no
vi
an
?Ne
osahw
ag
erin
a
Neo
ahon
ete
s N
eog
oaer
as
ara
tia
uZi
fera
A
ulo
ste
ges
S
taah
eoa
eras
Can
arin
e l
loid
es
Ste
pan
ovie
lla
L
iah
are
wia
Ap
eri
spir
ifer
le
tham
ensi
s
Ire
ni
an
Po
lydi
exod
in
a
Lis
soah
onet
es
an
d
smoo
th
fi
rs
t
ap
pe
ar
an
ce
I
elo
ng
ata
c
ho
ne
ti
ds
Go
dth
aab
ite
s
Neo
sahw
ag
erin
a
Eah
ina
losi
a m
axwe
lli
M
axi
aoae
ra
s Ch
iane
lla
ah
ianen
sis
Waag
enoa
eras
si
mple
x?
An
idan
thu
s eu
aha
ris
, so
lita
, m
inor
T
err
ak
ea a
onaa
vum
Un
ain
ell
a
Crur
ithy
ris
Pte
ros
pir
ifer
ala
tus
Pae
ake
lman
ell
a
Fi
li
pp
ov
ian
f
ir
st
ap
pe
ar
anc
e
Str
epto
rhy
nah
us
pe
lag
ona
rtu
s G
lend
ell
a
fi
rs
t
ap
pe
ar
anc
e
Chus
ene
lla
N
eoa
hon
etes
Da
ubi
ahit
es
Canae
lZin
a
Thu
lep
rod
ua
tus
ara
tiau
m
Sve
rdr
upit
es
Verb
eek
ina
T
err
akea
dia
kin
si
Chian
ell
a
Mak
laya
C
leio
thyr
idin
a p
eati
nif
era
P
seu
dos
yrin
x
Spiri
fer
el
la p
ola
ris
, S
. lo
ven
i,
kei
lha
vii
Tim
anie
lla
ha
rke
ri
Spir
ifer
ell
ina
ari
sta
ta
Die
Zasm
a e
ton
ga
tum
Tab
l.e S
l..
some
s
ig
ni
fi
can
t
in
ver
te
br
at
e
{ma
ri
ne
) g
en
er
a
and
s
pe
ci
es
o
f
th
e
ea
rl.
y
Mi
dd
l.e
P
erm
ian
P
er
io
d.
.......
00
0
�
;:: 3 3 s:::. q � � ..., 3 �· ;:s �
�
""

Summary of Permian Stages 1 8 1
As noted previously, nomenclature for the upper Kungurian is not fully established, and should receive attention from the Permian Subcommission, as several horizons have been only provisionally incorporated into one improperly named substage in the present study.
Climatically, the early Kungurian appears to have been cold, with evidence for glaciation in New Zealand and south-east Australia. This is supported by worldwide evidence for cool-water faunas. The later 'lrenian' Substage commenced with very much more diverse faunas rather allied to those of Baigendzinian age, and probably warm-water in relative attributes. Slightly later (Elkin and Ufimian) faunas were somewhat impoverished, with suggestions of moderate withdrawal of epicontinental seas.
Kazanian Stage (Table 5 1)
The Kazanian Stage is well defined in rocks and faunas from high and temperate Permian latitudes by a suite of brachiopod specie�, but is not so well controlled in palaeotropical regions. The subdivision into two stages rests almost entirely on Brachiopoda; and Ammonoidea provide little evidence for subdivision as far as is known. Fusuline evidence is not clear : there are suggestions -of a two-fold zonation, but correlation is not finally established. Mid-Permian transgression continued into the lower Kazanian or Kalinovian Substage, and widespread withdrawal of seas ensued in the Sosnovian or upper Kazanian Stage, starting in the Arctic. Climates are not clearly delimited, but the presence of glacial erratics in south-east Australia, and some worldwide faunal attributes suggest a moderately cool, but not strongly glaciated, episode. Both substages were somewhat similar in this regard.
Pu'fiabian Stage (Table 52)
The Punjabian Stage is characterized principally by Yabeina and Lepidolina amongst the Fusulinacea, Cyclolobus and Timorites amongst the Ammonoidea, and n�merous brachiopod species. The claim that the Punjabian Stage lay at or close to the top of the Permian Period is now abandoned, except by a few ammonoid specialists, but problems remain, particularly with regard to the age of Siberian faunas, regarded as either Kazanian or Kalabaghian (i.e. basal Punjabian), as well as certain palaeotropical faunas with primitive Y..abeina.
Climatically, the Punjabian Stage is not well understood. The Kalabaghian faunas frequently include brachiopod genera of cold-water attributes, to suggest a cold episode possibly correlative with Gijigin tillite reported from Verchoyan Mountains, north-east Siberia, but the tillite could be as old as upper Kazanian : this marks a prime question to be resolved. The Chhidruan Substage appears to be relatively warm, but there is evidence for several horizons still awaitin·g discrimination, with evidence over parts of Asia and New Zealand for a distinctive late Chhidruan fauna. Depending on correlations, it appears that withdrawal of seas continued extensively, especially after the Kazanian or Kalabaghian in north Siberia. But a Kalabaghian transgression invaded much of the Himalayas prior to · widespread Chhidruan emergence, and a Chhidruan transgression overlapped Kazanian sediments and faunas in Queensland, Australia.
Djulfuzn Stage (Table 52)
Fusulinacea and a number of brachiopod families began to diminish severely just before or within Djulfian Stage, but new ammonoid genera and families were generated. Long established brachiopod and fusuline families assumed new importance. There is fair uniformity over the classification of the stage, though the names are still to be

Sub
st
ag
e
Fu
su
li
na
ce
a
Br
ac
hi
op
od
a
Bi
va
lv
ia
Am
mono
id
ea
Gan
ge
ti
an
n
on
e
kn
ow
n
va
ri
ou
s
Pr
od
uc
ta
ce
a
Cla
ra
ia
Ot
ooe
ra
s
Og
bi
nan
P
ro
du
ct
id
s
pe
ci
es
P
ar
at
ir
oli
te
s
Ve
di
an
Pa
laeof
usu
Zin
a
Com
eli
oan
ia
� J
anic
eps
?Eu
ryd
esm
a
Ph
iso
ni
te
s
wi
de
sp
re
ad
T
omio
ps
is
?Ch
ang
hsi
ng
oo
eras
Ba
is
al
ia
n
Co
don
o f
us
ie
l l
a
Ar
axi
lev
is
Ar
axo
oer
as
Ved
io
oe
ra
s
Du.r
vi
lle
oo
er
as
Ur
us
ht
en
ian
P
oly
die
xo
din
a
Crur
ith
yr
is
At
om
od
es
mi
na
e
Str
igo
go
ni
at
it
es
m
exi
oan
a
Sp
ino
ma
rti
nia
E
oar
axo
oer
as
Ma
rt
in
ia
s
pp
.
Chh
id
ru
an
Lep
ido
Zin
a
Ter
ra
ke
a m
ul
tisp
in
osa
A
tom
ode
sma
Cy
olo
lob
us
Yab
ei
na
M
ont
io
uli
fer
a-F
ili
oo
no
ha
tr
eoh
man
ni
Tim
ori
te
s
At
ten
ua
tell
a i
no
ur
va
ta
Sp
ir
if
ere
lla
r
aja
h
Ma
rti
ni
op
sis
in
fla
ta
Ka
la
ba
gh
ian
L
epi
doZi
na
M
eg
aste
ge
s a
alh
ou
se
i
A.
wo
odi
Cyo
lolo
bu
s
Yab
ei
na
S
tro
ph
alo
si
in
a
tib
et
io
a
Tim
or
it
es
Le
pto
du
s ri
ohth
of
en
ii
Eli
vi
na
ti
be
tana
Marti
niop
sis
wood
i -
Tab
�e
52
. S
ig
ni
fi
can
t
(ma
ri
ne
i
nv
er
te
br
at
e)
ge
ne
ra
an
d
sp
ec
ie
s
of
Re
L
at
e
and
l
at
e
Mi
dd
le
P
er
mi
an.
I I
.....
00
N
Vol
l:: ::I 3 1':l q .:?., � ., 3 i:S"
;:s Vol
�
...

Summary of Permian Stages 1 83
resolved, as a relatively minor problem involving a choice between Urushtenian and Abadehian, and between Baisalian and Araksian. Climatically, there is no known evidence for glaciation, from either sedimentary or faunal analyses, and the climate may be assumed to have been warm. The Urushtenian or Abadehian Substages witnessed the cessation of deposition in Nepal-Tibet, the last major marine deposition in Texas, Mexico, Mongolia, and arguably in the Salt Range of Pakistan, but Djulfian marine sediments are widely developed in China, New Zealand, perhaps Queensland, Pamirs, Armenia, Iran, and were extensively transgressive in Yugoslavia-Italy-Austria. Coal measures possibly developed extensively in eastern Australia, but are poorly dated.
Dorashamian Stage (Table 52)
The Dorashamian Stage commenced with the highly distinctive Vedian Substage characterized by the brachiopods Comelicania and Janiceps, and various species, and by the ammonoid Phisonites. Faunas of the overlying Ogbinan Substage are less distinctive but include the ammonoid Paratirolites, and brachiopod species rather like those of Baisalian age apparently, though few have yet been described. Younger horizons are still in a state of nomenclatural flux, because studies were up till recently, uncritically based on ammonites and historical interpretation, with little consideration for overall faunal affinities. New names proposed by Kozur ( 1973b) better express the faunal subdivisions, and there can 6e little question that the Gangetian 'substage' or horizon faunally belongs to the upper Dorashamian Stage, characterized as it is by Otoceras, and various Permian-type brachiopod species as ytt undescribed. Only arguments based in historical usage can be offered against treating the Gangetian as Permian. But the position of the overlying upper Griesbachian and Dienerian units, or Brahmanian 'Stage' is still not clear, and requires further faunal studies before international discussion is worth undertaking.
Both the Vedian and Ogbinan Substages were virtually as restricted in distribution as the Baisalian Substage, being limited largely tp a 'Tethyan' belt from the south-east Mediterranean through Armenia-Iran-Pamirs, south China, and New Zealand, with other reports not yet substantiated. Apparent tillite is found at the Vedian horizon in New Zealand, and would support faunal evidence for climatic cooling at this time (with a sharp fall in temperature affinities even for Armenia shown by Waterhouse &Bonham-Carter, ( 1975), and the record of Changsingoceras and even Eurydesma (? !) in China. Later horizons appear to have been warm (Waterhouse, 1973d).
The Gangetian horizon saw the start of world-wide transgression that was most noticeable in the Arctic in covering marine beds as old as Kungurian-Kazanian, and also transgressed large tracts of North America, Himalayas, north Siberia, and elsewhere. Transgression appears to have continued into the Triassic Period.

References
Accordi, B. & Loriga, C.B. 1 968. Formazione a Bellerophon. In Le Dolomiti, Ed. P. Leonardi : 85- 10 1 . Inst. Geol. Univ. Ferrara.
Abramov, B.S. 1 970. Biostratigraphy of the Carboniferous of the Set Dahan Range (southern Verchoyan). C.R. Congr. Int. Strat. Geol. Carbon. 6th. 2 : 373-376 (In Russian).
Amos, A.J. & Rocha-Campos, A.C. 1 97 1 . A Review of South American Gondwana geology 1 967- 1 969. Second Gondwana Symposium, S. Afr. Proc. Pap.: 1 - 10.
Anderson, J.B. 1 973. The Biostratigraphy of the Permian and Triassic. Palaeont. afr. 1 6 : 59-83.
Andrianov, V.N. 1 963. On the question of fossils in Permian beds of west Verchoyan (Yakut SSR). Mat. geol. Soil, Yakut. SSR 1 1 .
--- . 1 966. Upper Palaeozoic deposits of the western Verchoyan region; Nauka, Moscow. (In Russian).
Arakelyan, R.A., Rausser-Chernossova, D.M., Reitlinger, E.A., Sherbovich, S.F., &Efrimova, N.A. 1 964. Significance of Permian Foraminifers of the TransCaucasus for the correlation of Permian deposits within the Tethys. In Stratigraphy of the Upper Paleozoic and Mesozoic of the southern biogeographical Provinces. Int. Geol. Congr. 22 Rep. Sov. Geol. Probl. 1 6a: 63-74. Nauka, Moscow, (In Russian).
Armstrong, J.D. 1 968. The unusual brachial skeleton of Attenuatella convexa sp. nov. (Brachiopoda). Palaeontology 1 1 : 783-792.
Armstrong, J.D. & Brown, C.D. 1 968. A new species of Attenuatella from the Permian of Queensland. Mem. Qd. Mus. 1 5 : 59-63 .
Armstrong, J.D., Dear, J.F. & Runnegar, B. 1 967. Permian ammonoids from eastern Australia. J. geol. Soc. Aust. 14 : 87-97.
Assereto, A., Bosellini, A., Sestini, N., Fantini & Sweet, W.C. 1973 . The PermianTriassic boundary in the southern Alps (Italy). Mem. Can Soc. Petrol. Geol. 2: 1 76- 199.
Astre, G. 1 934. La Faune permienne des gres a Productus d'Ankitokazo dans le nord de Madagascar. Annis. geol. Serv. Mines Madagascar 4 : 63-96.
Balme, B.E. 1 970. Palynology of Permian and Triassic strata in the Salt Range, west Pakistan. Stratigraphic Boundary Problems : Permian and Triassic of West Pakistan. Ed. Kummel, B. & Teichert, C . Spec. Pubis. Univ. Kansas 4 : 306-453.
Bamber, E.W. & Waterhouse, J.B. 1 97 1 . Carboniferous and Permian stratigraphy of paleontology, northern Yukon Territory, Canada. Bull. Can Petrol. Geol. 19 ( l ) : 29-250.
Banks, M.R. 196 1 . Age and relationship of Tasmanian fossil faunas and floras. Proc. 9th Pacific Sci. Congr. Geol. Geophys. 12 : 324-339.
--- . 1 962. Permian. In Geology of Tasmania. J. geol. Soc. A ust. 9 (2): 1 89-2 16. Barkhatova, V.P. 1 964. The status of the Schwagerina beds in connection with the
problem of the boundary between the Carboniferous and the Permian. C.r. Congr. Int. Strat. Geol. Carbon., 5th, Paris 1 963 1 , 273-282.
--- . 1 968. Spiriferella, Timaniella, Parachoristites. New species of polar plants and animals from USSR. 2 : 1 60- 1 67 (In Russian).
Beede, J.W. & Kniker, H.T. 1 924. Species of the genus Schwagerina and the stratigraphic significance. Bull. Univ. Texas 2433 : 1 -96.

References 1 85
Bion, H.S. 1928. The fauna of the Agglomeratic Slate Series of Kashmir. Mem. geo/. Surv. India Palaeont. indica 1 2 : 1 -42.
Bissell, H.J. 1973. Permian-Triassic boundary in the eastern Great Basin Area. Mem. Can. Soc. Petrol. Geol. 2: 3 18-334.
Bordet, P., Colchen, M., Krummenacher, D., Lefort, P., Mouterde, R. & Remy, M. 197 1 . Recherches geologiques dans /'Himalaya du Nepal, region de Ia Thakkhola. Centre Nat. Recher. Sci. Paris, 279pp.
Bostwick, D.A. 1962. Fusulinid stratigraphy of beds near the Gaptank-Wolfcamp boundary, Glass Mountains, Texas. J.Paleont. 36 : 1 1 89- 1 200.
Brabb, E.E. & Grant, R.E. 1 97 1 . Stratigraphy and paleontology of the revised type section for the Tahkandit Limestone (Permian) in east-central Alaska. Prof. Pap. U.S. geol. Surv. 703 : 1 -26, 2 pl.
Branson, C.C. 1930. Paleontology and stratigraphy of the Phosphoria Formation. Univ. Mo. Stud., 5 (2).
--- . 1 948. Bibliographic Index of Permian Invertebrates. Mem. geol. Soc. Am. 26 : . 1 - 1049.
Broili, F. 19 15 . Permische Brachiopoden de Insel Letti. Jaarb. Mijnw. Ned. 0-Ind. 43 : 187-207.
--- . 1 9 16. Die Permischen Brachiopoden von Timor. Palaont. Timor 7 ( 12): 1 - 104. Brown, D.A., Campbell, K.S.W. & €rook, K.A.W. 1968. The geological evolution of
Australia and New Zealand. Pergamon Press, Oxford. Buddington, A.F. & Chapin, T. 1 929. Geology and mineral-deposits of south-eastern
Alaska. Bull. U.S. geol. Surv. 1 24 1 C : 1 -52. Burke, K. & Waterhouse, J.B. 1 973. Saharan glaciation dated in North America.
Nature 24 1 (5387) : 267-268. Campbell, K.S.W. 1 953 . The fauna of the Permo-Carboniferous Ingelara beds of
Queensland. Pap. Dep. Geol. Univ. Qd. 4 (3): 1 -44. --- . 1 960. The brachiopod genera Ingelarella and Notospirifer in the Permian of
Queensland. J. Paleont. 34: 1 106- 1 1 23 . ·
--- . 1 96 1 . New species of the Permian Spiriferoids lngelarella and Notospirifer from Queensland and their stratigraphic implications. Palaeontographica 1 1 7 A : 159- 1 92.
--- . 1 965. Australian Permian Terebratuloids. Bull. Bur. Miner. Resour. Geol. Geophys. Aust. 68.
Chalishev, V.I. 1966. Upper Permian beds of the northern pre-Urals. Paleozoic beds of the northern pre-Urals. Trudy lnst. geol. Nauk. Mosk. 73-95. (In Russian).
Chan, Lin Sin. 1 963. Upper Carboniferous Fusulinids from Kelpin and nearby Sinkiang. Acta palaeont. sin. 1 1 ( 1 ): 36-70. (In Chinese and Russian).
Chan Li-Pei & Li Li 1962. Early Permian brachiopods from the Maokou Suite of east Uchatka Chin-Lin. Acta palaeont. sin. 10 (4) : 472-50 1 . (In Chinese and Russian).
Chao, King-Koo 1 947. Stratigraphical development in Kwangsi. Bull. geol. Soc. China 27: 32 1 -346.
--- . 1 966. The Permian ammonoid bearing formations of South China. Scientia sin. 14 ( 12): 1 8 1 3 - 1 825.
Chao, Y.T. 1 925. On the age of the Taiyuan Series of North China. Bull. geol. Soc. China 4 ( 1 ) : 22 1 -249.
--- . 1927a. Brachiopod fauna of the Chihsia Limestone. Bull. geol. Soc. China 6 : 83- 1 20.
--- . 1 927b. Productidae of China Pt. 1 . Producti. Palaeont. sin. B 5 (2). --- . 1 928. Productidae of China, Pt. 2. Palaeont. sin. B 5 (3). --- . 1 929. Carboniferous and Permian Spiriferids of China. Palaeont. sin. B 1 1
( 1 ). Chernyshev, T. 1 889. Allgemeine geologische Karte von Russland, Blatt 1 39.

1 86 References
Beschreibung des Central-Urals and des Westabhanges. Russia. Trudy geol. Kom.3 (4). (In Russian and German).
--- . 1 902. Die obercal'bonischen Brachiopoden des Ural und des Timan. Trudy geo/. Kom. 1 6 (2). (In Russian and German). ·
--- . 1 9 14 . Die Fauna der oberpalaeozoischen Ablagerungen des Darvas, Iieferung 1 . Trudy geo/. Kom. 1 04. (In Russian and German).
Chernyshev, T. & Stepanov, P. 1 9 1 6. Obercarbonfauna vom Konig Oscars und vom Heibergs Land. Rep. Second Norwegian Arctic Expedition in the Fram, 1 898-1 902, 34: 1 -67.
Chi-Thuan, T.T. 1 96 1 . Les brachiopodes permiens du Phnom-Tup, (SisophonCambodge). Annis. Fac. Sci. Saigon, 1 96 1 , 6 : 257-308.
--- . 1 962. Les Brachiopodes permiens de C am-lo (Province de Quang-Tri). Annis. Fac. Sci. Saigon, 1 962, 7 : 485-498.
Clark, D.L. & Behnken, F.H. 197 1 . Conodonts and biostratigraphy of the Permian. Mem. geol. Soc. Am. 1 27 : 4 15-439.
Clark, D.L. & Ethington, R.L. 1 962. Survey of Permian conodonts in western North America. Geol. Stud. Brigham Young Univ. 9 (2) : 102- 1 14.
Clark, R.H., Vella, P. & Waterhouse, J.B. 1 967. The Permian at Parapara Peak, Northwest Nelson. N.Z. J. Geol. Geophys. 10 (1) : 23 1 -246.
Clarke, M.J. 1973 . Faunas from the Ferntree Group of south-eastern Tasmania. Tech. Rep. Dep. Mines Tasmania 16 : 4 1 -65.
Clifton, R.L. 1 942. Invertebrate faunas from the Blaine and the Dog Creek Formations of the Permian Leonard Series. J.Paleont. 1 6 (6) : 685-699.
Coleman, P.J. 1 957. Permian Productacea of Western Australia. Bull. Bur. Miner. Resour. Geo/. Geophys. Aust. 40.
Condon, M.A. 1 967. The geology of the Carnarvon Basin, Western Australia Part 2: Permian stratigraphy. Bull. Bur. Miner. Resour. Geo/. Geophys. Aust. 71.
Coogan, A.H. 1 960. Stratigraphy and paleontology of the Permian Nosoni and Dekkas Formations, (Bollibokka Group). Univ. Calif. Pubis. geo/. Sci. 36 (5): 243-3 16.
Cooper, G.A. 1957. Permian brachiopods from central Oregon. Smithson. Misc. ColIns. 1 34 ( 12) : 1 -79.
Cooper, G.A., Dunbar, C.O., Duncan, H., Miller, A.K., & Knight, J.B. 1 953 . Permian fauna at El Antimonio, Western Sonora, Mexico. Smithson. Misc. Co/Ins. 1 19 (2): 2 1 -77.
Cooper, G.A. & Grant, R.E. 1 964. New Permian stratigraphic units in Glass Mountains, west Texas. Bull. Am. Ass. Petrol. Geol. 48 (4): 1 5 8 1 - 1 588.
--- . 1 966. Permian rock units in the Glass Mountains, West Texas. Bull. U.S. geol. Surv. 1244 E: 2-9.
Cooper, G.A. & Grant, R.E. 1969. New Permian brachiopods from west Texas. Smithson. contrib. Paleobiol. 1 : 1 -20.
--- . 1973. Dating and correlating the Permian of the Glass Mountains in Texas. Mem. Can. Soc. Petrol. Geol. 2 : 363-377.
--- . (in press). Permian brachiopods of Texas. Smithson. contrib. Paleobiol. 14-19. Crespin, I. 1958. Permian Foraminifera of Australia. Bull. Bur. Miner. Resour. Geol.
Geophys. Aust. 48: 1-207. ·
Crockford, M.B.B. & Warren, P.S. 1 935 . The Cache Creek Series of British Columbia. Proc. Trans. R .Soc. Can. 29: 149- 1 6 1 .
Cut bill, J .L. & Challinor, A . 1 965. Revision of the stratigraphical scheme for the Carboniferous and Permian rocks of Spitzbergen and Bj�rn�ya. Geol. Mag. 1 02: 4 18-439.
Czarniecki, S . 1 969. Sedimentary environment and stratigraphical position of the Treskelodden Beds (Vestspitsbergen). Pr. Muz. Zieml 1 6 : 201 -286.
Dagis, A.S. & Ustritsky, V.I. 1973 . Main relationships between the changes in marine

References 1 87
fauna at the close of the Permian and the beginning of the Triassic. Mem. Can. Soc. Petrol. Geol. 2 : 64 7-654.
David, T.W.E. 1 907. The geology of the Hunter River Coal Measures, New South Wales. Mem. geol. Surv. N.S. W. : Geol. 4.
Dear, J.F. 1966. An Ammonoid from the Permian of Queensland. Mem. Qd. Mus. 14 (5): 199-204.
--- . 1 969. A new Queensland Permian Linoproductid with Arctic affinities. Stratigraphy and Paleontology. Essays in honour of Dorothy Hill. Ed. K.S.W. Campbell : 299�306. Aust. Nat. Univ. Press, Canberra.
--- . 1 97 1 . Strophomenoid brachiopods from the higher Permian faunas of the Back Creek Group in the Bowen Basin. Pubis. Geol. Surv. Qd. 34 7 : Palaeont. Pap. 2 1 : 1 -39, 7 pl.
--- . 1972. Preliminary biostratigraphic subdivision of the Permian brachiopod faunas in the northern Bowen Basin and their distribution throughout the basin. Rep. geol. Surv. Qd. 49: 1 - 1 9.
Deprat, J. 1 9 1 3 . Les series stratigraphiques en Indochine et au Yunnan. Mem. Serv. geol. Indoch. 2 (2) 4 :
Derby, O.A. 184 7. On the carboniferous Brachiopoda of Itaituba, Rio Tapajos, Province of Para, Brazil. (Morgan Expedition, 1870-7 1 , to charge Cp. Fred. Hartt). Bull. Cornell Univ. Sci. 1 (2): 1 -63.
Dickins, J.M. 1 956. Permian pelecypods from the Carnarvon Basin, Western Australia. Bull. Bur. Miner. Resour. Geol. Geophys. Aust. 29.
--- . 1 963. Permian pelecypods and gastropods from Western Australia. Bull. Bur. Miner. Resour. Geol. Geophys. Aust. 63.
--- . 1 964. Appendix in Malone, E.J., Corbett, D.W.P. & Jensen, A.R. Geology of the Mt Coolon 1 :250,000 sheet area. Rep. Bur. Miner. Resour. Geol. Geophys. Aust. 64.
--- . 1968. Correlation of the Permian of the Hunter Valley, New South Wales and the Bowen Basin, Queensland. Bull. Bur. Miner. Resour. Geol. Geophys. Aust. 80 (3): 29-44.
Dickins, J.M. & McTavish, R.A. 1963. Lower Triassic marine fossils from the Beagle Ridge (BMR 10) Bore, Perth Basin, Western Australia. J. geol. Soc. Aust. 10 ( 1 ): 1 23-140.
Dickins, J.M., Gostin, J.A. & Runnegar, B. 1 969. Correlation and age of the Permian sequence in the southern part of the Sydney Basin, New South Wales. Stratigraphy and Paleontology: Essays in Honour of Dorothy Hill. Ed. K.S.W. Campbell: 2 1 1 -226, Aust. Nat. Univ. Press Canberra.
Dickins, J.M. & Thomas, G.A. 1 959. The marine fauna of the Carnarvon Basin, Western Australia. Bur. Miner. Resour, Geol. Geophys. Aust. 38 : 65-96.
Diener, C. 1 897a. The Permo-Carboniferous Fauna of Chitichun No. 1 . Mem. geol. Surv. India Palaeont. indica Ser. 1 5 , 1 (4).
--- . 1897b. The Permian fossil!> of the Productus shales of Kumaon and Garwhal. Mem. geol. Surv. India Palaeont. indica Ser. 1 5 , 1 (4).
--- . 1 903. Permian fossils of the central Himalayas. Mem. Geol. Surv. India Palaeont. indica Ser. 15 1 (5).
--- . 1 909. Lower Triassic Cephalopoda from Spiti, Mala Johar and Byans. Mem. Geol. Surv. India Palaeont. indica Ser. 1 5 6 ( 1 ).
--- . 1 9 10. Die Brachiopodenfauna des Bellerophonkalkes von Schaschar und Schonbrunn. Jb. geol. Bundesant., Wien. 60: 298-309.
--- . 1 9 1 1 . Anthracolithic fossils of the Shan States. Mem. Geol. Surv. India Palaeont. indica ser 3 (4).

1 88 References
--- . 1 9 1 5 . The Anthracolithic Faunas of Kashmir, Kanuar and Spiti. Mem. geol. Surv. India Palaeont. indica n.s. 5 (2).
D'Orbigny, A. 1 842. Voyages dans !'Amerique meridional 3 . Mem. Soc. geol. Fr. Paleont. : 50-56.
Douglas, J.A. 1 936. A Permo-Carboniferous fauna from S.W. Persia {Iran'). Mem. geol. Surv. India Palaeont. indica n.s. 22 (6).
--- . 1 950. The Carboniferous and Permian faunas of South Iran and Iranian Baluchistan. Mem. geol. Surv. India Palaeont. indica 22 (7).
Douglass, R.A. 1 970. Morphologic studies of Fusulinids from the lower Permian of west Pakistan. Prof. Pap. U.S. geol. Surv. 643-G: 1 - 1 1 .
Douglass, R.C. & Nestell, M.K. 1 974. Fusulinid Foraminifera from southern Chile. An. Acad. brasil. Cienc. ( 1 972) (suplemento) 44 : 1 l9- 1 23.
Dronov, V.E. & Leven, E.Y. 196 1 . On the problem of the geological problems of the south-east Pamirs. Geol. lit. SSR 1 1 . (In Russian).
Dunbar, C.O. 1 940. The type Permian : its classification, and correlation. Bull. Am. Ass. Petrol. Geol. 24 (2) : 237-28 1 .
--- . 1 955 . Permian brachiopod faunas of central east Greenland. Meddr. Gr�nland Bd. 1 10 (3): 1 - 169.
Dunbar, C.O. & Condra, G.E. 1 932. Brachiopoda of the Pennsylvanian System of Nebraska. Bull. geol. Surv. Neb. 2 {5}: 1 -317.
Dunbar, C.O. & Skinner, J.W. 193 1 . New Fusulinid Genera from the Permian of WestTexas. Am. J. Sci. 22: 252-268.
--- . 1 936. Schwagerina verus Pseudoschwagerina and Paraschwagerina. J. Paleont. 10: 83-9 1 .
Dunbar, C.O. 1 960 E T AL. 1 960. Correlation of the Permian formations of North America. Bull. geol. Soc. Am. 7 1 : 1 763-1 806.
Einor, O.L. 1 939. Some brachiopods from the Lower Permian of Taimyr {basin of the Piassina River). Trudy arkt. nauchno-issled Inst. 1 3 5 : 1 - 1 50. (In Russian).
Einor, O.L. 1 946. Brachiopods from the Lower Carboniferous and Lower Permian of West Taimyr. Problemy Trudy Arkt. nauchno-issled Inst. 26. (In Russian).
Eldredge, N. & Gould, S.J. 1 975. Punctuated equilibria : An alternative to phyletic gradualism. In Models in Paleobiology, Ed. T.J.M. Schopf. Freeman, Cooper &Co. San Francisco.
Enderle, J. 1 90 1 . Ueber eine anthracolithische Fauna von Balia Maaden in Kleinasien. Beitr. Palaont. Geol. Ost.-Ung. 1 3 : 49- 109.
Etheridge, R. 1892. IN Jack, R.L. & Etheridge, R.Jnr. Geology and Palaeontology of Queensland and New Guinea. Pubis geol. Surv. Qd. 92.
--- . 1 9 14. Palaeontological contributions to the geology of Western Australia. Ser. 5. Western Australian Carboniferous fossils, chiefly from Mount Marmion, Lennard River, West Kimberley. Bull. geol. Surv. West Aust. 5 8 : 1-59.
--- . 1 9 1 9 . Occasional descriptions of New South Wales fossils, No. 7. Rec. Aust. Mus. 12 : 1 83 - 192.
Fliegel, G. 190 1. Ueber obercarbonische F aunen a us Ost und Sud Asien. Palaeontographica 48 : 9 1 - 1 36.
Forbes, C.L., Harland, W.B. & Hughes, N.F. 1 958 . Palaeontological evidence for the age of the Carboniferous and Permian rocks of central Vestspitsbergen. Geol.Mag. 95 : 465-490.
Frebold, H. 1 93 1 . Das marine Oberkarbon Ostgronlands. Leitende fauna, alterstellund, Palaeogeographie. Meddr. Gr{lnland 84 (2).
--- . 1 933. Weitene Beitrage zur Kenntnis des oberen Palaozoikums Ostgronlands. 1 . Die Fauna und stratigraphische Stellung der oberpaUiozoischen 'Weissen Blocke' (Kap Stosch Formation) Ostgronlands. Meddr. Grflnland 84 (7}.
---. · 193� . Das Festungsprofil auf Spitzbergen. IV. Die Brachiopoden - und Lamelhbranchiaten Fauna und die Stratigraphie des Oberkarbons und Unterperms

References 1 89
nebst Beschriebung anderer Vorkommen in Svalbard. Skr. Svalbard Ishavet 69. --- . 1 950. Stratigraphie und Brachiopodenfauna des marinen Jungpalaeozoikums
von Holms und Amdrups Land (NordostgrfiJnland.) Meddr. Gr�nland 1 26 (3). Frederiks, G. 1924. Upper Palaeozoic of the Ussuriland. 1 , Brachiopoda. Mater. Geol.
po/ez. Iskop. Dal'n Vost 28 : 1 -52 (In Russian). --- . 1 925. Upper Palaeozoicum of the Ussuriland. 2. Permian Brachiopoda of
Cape Kalouzin. Mater. Geo/. polez. Iskop. Dal'n Vost. 40: 1 -28.(In Russian). --- . 1 926. Materials on the fauna of the Kejim Terovey River. Izv. vses. geol.
razved. Ob'ed 45 : 8 1 -95. (In Russian). --- . 1 93 1 . The Upper Palaeozoic Fauna of Haraulakh Mountains. Bull. A cad. Sci.
USSR: 199-228. (In Russian). --- . 1 932. The Upper Palaeozoic of the western slope of the Ural. Trudy vses geol.
razv. Ob'ed USSR 106: 1 -83 (In Russian). --- . 1 934a. The Permian fauna of the Kanin Peninsula. Trudy arkt. Nauchno
issled. Inst. 1 3 : 5-42. --- . 1 934b. In Ahnert, Permocarbonic fauna of limestones from the Ashiho River
and Ert'soengtientzu railroad station. Nat. Sci. Geog. YMCA Harbin, Ann. 1 : 103- 106. (In Russian).
--- . 1 936. Material of the Permian fauna of Novaya Zemlya. Trudy arkt. Nauchno-issled Inst 49: 94- 1 09.
Fujimoto, H. 1960. Geological map witli exp/(matory text: Togichi, I :50,000. Jap. geol. Surv.
Furnish, W.M. 1966. Ammonoids of the Upper Permian Cyclolobus Zone. Neues Jb. Miner. Geol. Palaont. Abh. 1 25 : 265-296.
--- . 1973. Permian stage names. Mem. Can Soc. Petrol. Geo/. 2: 522-548. Furnish, W.M. & Glenister, B.F. 1970. Permian ammonoid Cyclolobus from the Salt
Range, West Pakistan. Stratigraphic Boundary Problems, Permian and Triassic of West Pakistan. Ed. Kummel, B. & Teichert, C. Spec. Pub/. Univ. Kansas 4: 153- 1 77.
Gabrielyan, A.A., Azaryan, N.P., Akopyan, V.T., Arakelyan, R.A. & Mesropyan, A.l. 1 964. Geology of Armenian SSR. Part 2 Stratigraphy. Acad. Sci. Armen. SSR, Geo/. Inst. : 1 -432. ( In Russian).
Gemmellaro, G.G. 1 899. La fauna dei Calcari con Fusulina della Valle dei Fiume Sosio nella Provincia di Palermo. Brachiopoda. Giorn. di Sci. Natur. ed. Econom. Palermo 22: 95-2 1 4.
Gerassimov, N.P. 1952a. Productacea from the Kungurian and Artinskian Permian of the pre-Urals. Tr. EN/ Perm. Un-ta 10, 3, 4. (In Russian).
--- . 1952b. Kungurian Stage of the Kama area in the Pre Urals. Uch. West Perm. Gos. Un-ta 7 ( I ).
--- . 1 9 56. On the problem of intercorrelation of the Kazanian Stage of the Russian Platform and western European Zechstein. Stud. west Molot. Gos. Univ. 7 (4): 1 -2 5 . (In Russian).
Gerth, H. 1 960. Die ammonoideen des Perms von Timor und ihre Bedeutung ftir die stratigraphische Gliederung der Perm formation. Abh. Neues Jb. Miner/. Geol. Palaeont. 9 1 : 233-320.
Ghosh, P.K. & Basu, A. 1 969. Stratigraphic position of the Karharbaris in the lower Gondwanas of India. UNESCO Gond. Strat. JUGS Sympo. Buenos Aires: 407-4 1 9.
Gilluly, J., Cooper, J.R. & Williams, J.S. 1 954. Late Paleozoic Stratigraphy of central Cochise County, Arizona. Prof. Pap. U.S. geol. Surv. 266 : 1 -49.
Girty, G.H. 1 909. The Guadalupian fauna. Prof. Pap. U.S. geol. Surv. 58 : 1 -65 1 , 3 1 pl. --- . 1 9 10. The Fauna of the phosphate beds of the Park City Formation in Idaho,
Wyoming and Utah. Bull. U.S. Geol. Surv. 436.

1 90 References
--- . 1 9 1 5 . Fauna of the Wewoka Formation of Oklahoma. Bull. U.S. geo.l. Surv. 544.
--- . 1 927. Descriptions of Carboniferous and Triassic fossils. In Mansfield, G.R., Geography, Geology and Mineral Resources of part of southeastern)daho. Prof. Pap. U.S. geol. Sur,;. 1 52.
· '
Glenister, B.F. & Furnish, W.M. 1 96 1 . The Permian ammonoids of Australia. J.Paleo.nt. 35 : 673-736.
Glintz-Boeckel, C. & Rabate, J. 1964. Microfaunas et Microfacies du PermoCarbonifere du Sud Tunisien. Brill, Leiden, 263 pp.
Gobbett, D.J. 1964. Carboniferous and Permian brachiopods of Svalbard. Skr. Norsk. Polarinst. 1 27 : 1 -20 1 .
1968. The Permian System in Malaya. Buff. geol. Soc. Malaysia 1 : 17-22. Gonzalez, C.R. 1 973. Un nuevo Eurydesmidae en el Carbonico superior de Patagonia,
Argentina. Ameghiniana: Argentina 9 : 2 1 3-2 1 9. Gorsky, -V.P. & Gusseva, E.A. 1 973. The Kungurian and Ufimian Stages of the Per
mian. Mem. Can Soc. Petrol. Geol. 2 : 1 68- 1 72. Gortani, M. 1 906a. Contribuzioni allo studio del Paleozoico Carnico. 1. La fauna per
mocarbonica del Col. Mezzodi presso Formi avoltri. Palaeontogr. ita!. 12 : 1 -84. Gortani, M. 1 960b. La fauna degli strati a Belleropl;ton della Carnia. Riv. ita!. Paleont.
12 : 93- 1 3 1 . Grabau, A.W. 1 924. Stratigraphy of China. Part 1 . Palaeozoic. Geol. Surv. China,
Peking. --- . 1 93 1 . The Permian of Mongolia. Am. Mus. Nat. His(. 4 : 1 -665 . --- . 1 934. Early Permian Fossils of China, 1 : Early Permian brachiopods,
pelecypods and gastropods of Kweichow. Palaeont. sin. B 8 (3). --- . 1 936. Early Permian fossils of China, 2: Fauna of the Maping Limestone of
Kwangsi and Kweichow. Palaeont. sin. B 8 (4). Grant, R.E. 1968. Structural adaptation in two Permian brachiopod genera. Salt Range,
West Pakistan. J.Paleont. 42: 1 -32. --- . 1 970. Brachiopods from Permian Triassic boundary beds and age of Chhidru
Formation, West Pakistan. Stratigraphic Boundary Problems, Permian and Triassic of West Pakistan. Ed. Kummel, B. & Teichert, C . : Spec. Pub!. Univ. Jransas 4: 1 1 7- 1 53 .
--- . 1 97 1 . Taxonomy and autecology of two Arctic Permian Rhynchonellid brachiopods. Smithson. contrib. Paleobiol. 3 : 3 1 3-335.
--- . 1972. The lophophore and feeding mechanism of the Productidina (Brachiopoda). J. Paleont. 46 (2): 2 1 3-248.
Grant, R.E. & Cooper, G.A. 1 973. Brachiopods and Permian correlations. Mem. Can. Petrol. Geol. Soc. 2: 572-595.
Greco, B. 1942. La Fauna permiana del Sosio, etc. Parte terza, Brachiopoda, fasc. 1 , Pam. Orthidae, Strophomenidae. Palaeontogr. ita!. 40: 1 15 - 1 59.
--- . 1947. La fauna permiana del Sosio conservata nei Musei di Pisa, di Firenze e di Padova. 1 1 1 Brachiopoda Fasc. 2. Productidae, Richthofeniidae. Palaeontogr. ita!. 4 1 : 1 -22.
Gregorio, A. De. 1930. Sui Permiano di Sicilia (Fossili de calcare con Fusulina di Palazzo Adriano non descritti Prof. G. Gemmellaro conservati nel mio privatogabinetto. Annis Geol. Paleont.: Turin 52 : 1 8-32.
Grigorieva, A.D. 1962. Productida of the Kazan Stage of the Russian platform and theconditions of their existence. Trudy paleont. Inst. 92: 1 -92. (In Russian).
Gr!llnwall, K.A. 1 9 1 7. The marine Carboniferous of northeast Greenland and itsbrachiopod fauna. Meddr. Gr,nland 43 : 509-6 18 .
Grunt, T.A. & Dimetriev,V.Y. 1 973. Permian brachiopods of the Pamir. Trudy paleont.Inst. 136 : 1 -2 1 2 . ( In Russian).

References 1 9 1
Hamlet, B. 1928. Permische Brachiopoden, Lamellibranchiaten und Gastropoden von Timor. Jaarb. Mijnw. Ned-Oost-lndie. 56 (2): 1 - 1 15 .
Haniel, C.A. 19 15 . Die Cephalopoden der Dyas von Timor. Palaeont. Timor Lief 3 (6): 1 - 153 .
Hanzawa, S . & Murata, M. 1 963. The Paleontologic and Stratigraphic Considerations on the Neoschwagerinae and Verbeekininae with the descriptions of some Fusulinid Foraminifera from the Kitakami Massif, Japan. Scient. Rep. Tohoku Univ. : Sendai, Japan, Ser.2 35 ( 1) : 1 -32.
Harker, P. & Thorsteinsson, R. 1 960. Permian rocks and faunas of Grinnel Peninsula, Arctic Archipelago. Mem. geol. Surv. Can. 309: 1 -90.
Hart, G. F. 1970. The biostratigraphy of Permian palynofloras. Geoscience and Man 1 : 89- 1 3 1 .
Hayasaka, I . 1922. Palaeozoic Brachiopoda from Japan, Korea and China. Sci. Rep. Tohoku Univ: Sendai, Japan, Ser.2 6 : 1 - 1 38 .
--- . 1 923. Some Permian fossils from the Kitakami Mts. lap. J. Geol. Geog. 2. Hayasaka, I. 1925. On some brachiopods from the Lyttonia Horizon of the Kitakami
Mountains. lap. J. Geol. Geogr. 4: 89- 103. --- . 1 960. On the occurrence of Neospirifer fasciger (Keyser ling) in Japan, and a
note on some associate Permian brachiopods from around Kesen-numa city, north-east Japan. Collected Essays, Comm. Tenth Anniv. Shimane Univ. : Nat. Sci. : 34-37.
--- . 1 963. Some Permian fossils from southern Kitakami, 1 -4. Proc. Japan Acad. 39: 474-478, 479-483, 594-599, 753-757.
Harrington, H.J. 1955. The Permian Eurydesma fauna of eastern Argentina. J. Paleont. 29: 1 1 2- 128.
Hedberg, H.D. 1 972. Summary of an international guide to stratigraphic classification, terminology and usage. Lethaia 5 : 297-323.
Helby, R.J. 1969. The Carboniferous-Permian boundary in eastern Australia, an interpretation on the basis of palynological information. Spec. Pub/. geol. Soc. Aust. 2: 69-72.
Heritsch, F. 1 934. Die stratigraphie von Oberkarbon und Perm in den Karnischen Alpen. Mitt. geol. Ges. Wien 26 : 1 6 1 - 1 89.
--- . 1 935 . Neue Versteinerungen aus den Nassfeldschichten der Karnischen Alpen. Sber. Akad. Wiss. Wien Math-naturwiss. Kl. 144 : 349-375.
--- . 1938. Dje stratigraphische stellung des Trogkofelkalkes. Neues Jb. Miner. Geol. Paliiont. Beilbd. Ser. B 79: 63- 1 86.
Hill, D. 1950. The Productinae of the Artinskian Cracow fauna of Queensland. Pap. Dep. Geol. Univ. Qd. 3 (2) : 1 -36.
Huang, T.K. 1932a. The Permian Formations of southern China. Mem. geol. Surv. China ser. A 10: 1 - 1 29.
--- . 1932b. Late Permian Brachiopoda of southwestern China. Palaeont. sin. B 9 ( 1 ).
--- . 1 933 . Late Permian Brachiopoda of S.W. China. Palaeont. sin. B 9 (2). Hudson, R.G.S. & Sudbury, M. 1959. Permian brachiopods from southwest Arabia.
Notes mem. Moyen-Orient 7 : 19-55 . lfanova, V.V. & Semenova, E.G. 1 972. Middle Carboniferous and Permian
brachiopods from east and north of the European part of the USSR. Sci. Council on Problem 'Evolutionary Trends and patterns of plant and animal organisms '. Akad-Nauk. USSR: 1 - 1 6 1 . (In Russian).
lgo, H. 1 964. Permian fossils from Northern Pahang, Malaya. Geol. Palaeont. southeast Asia 1 : 1 9 1 -207.
Imamura, S. 1953. New occurrence of Lyttonia from the environs of Hiroshima, Japan, J. Sci. Hiroshima Univ. 1 (3): 1 1 - 1 5 .

192 References
Irving E. & Parry, L.G. 1 963. The magnetism of some Permian rocks from New South Wales. Geophys. J. 7 : 395-4 1 1 .
Ishii, K. 1 966. On some Fusulinids and other Foraminifera from the Permian of Pahang, Malaya. J. lnst. Polytech. Osaka City Univ. : Geoscience 9 (4): 1 3 1 - 142.
Ishii, K. , Kato, M. & Nakamura, K. 1 969. Permian limestones of West Cambodia. Lithofacies and Biofacies. Spec. Pap. palaeont. Soc. Japan Pap. 14 : 4 1 -55.
Ivanov, A. 1 935 . Brachiopod fauna of the middle and upper Carboniferous from the Moscow Basin. Part 1 Productinae. Trudy Mosk. Geol. Org. 8 : 1 - 1 34.
Ivanova, E.A. 1 958. Development of faunas through temporal and ecologic responses.·Trudy paleont. Inst. 69. (In Russian).
Ivanova, E.A. & Ivanova, I.V. 1 955 . Stratigraphy of the Middle and Upper Carboniferous of the western part of the Moscow Syncline. Trudy paleont. lnst. 53: 1-279.
Johnston, R.M. 1 888. Systematic account of the geology of Tasmania. Government Printer, Hobart, 408pp.
Jordan, H. 1 969. Zur Biostratigraphie und Fazies des Zechsteins im Germanischen Becken. Freiberger ForschHft. C 254: 27-45.
Kahler, F. & Kahler, G. 1 966. Fusulinida (Foraminifera). Fossilium Catalogue 1 : Animalia. Ed. F . Westphal. Pars 1 1 1 - 1 14, W. Junk, s'Gravenhage, 1 -974.
Kahler, V.F. & Prey, S. 1 963. Erlauterungen zur Geologischen Karte des NassfeldGartnerkofel-Gebietes in den Karnischen A /pen. Jb. geol. Bundesant., Wien.
Ka1mikova, M.A. 1 965. A revision of the groups Pseudofusulina vulgaris and Pseudofusulina kraffti (from topotype material). Izv. vses. geol. razved. Ob 'ed Micropaleont. Probl. 9 : 1 1 1 - 1 28 .
--- . 1 967. Permian Fusulinacea from Darvas. Trudy vses. nauchno-issled. geol. Inst. 1 1 6 : 1 1 6-287.
Kanmera, K. 1953. The Kuma Formation with special reference to the Upper Permian in Japan. (Geological study of the Palaeozoic in southern Kyushu, pt. 3). J. geol. Soc. Japan 59 : 449-468.
--- . 1 963. Fusulines of the Middle Permian Kozaki Formation. Mem. Fac. Sci. Kyushu Univ. D: Geol. 14 (2) : 79- 1 4 1 .
Kanmera, K . & Mikami, T. 1 965. Succession and sedimentary features of the lower Sakamotozawa Formation. Mem. Fac. Sci. Kyushu Univ. D: Geol 16 (3): 265-274.
Kanmera, K. & Nakazawa, K. 1 973. Permian�Triassic relationships and faunal changes in the eastern Tethys. Mem. Can. Soc. Petrol. Geol. 2 : 100-1 1 9.
Kanmera, K. & Toriyama, R. 1 968. Fusulinacean fossils from Thailand Part III, Maklaya, new generic description for Neoschwagerinids of the groups of Cancel/ina pamirica Leven. Geo/. Palaeont. southeast Asia 5 : 3 1 -46.
Karanetov, C .S.S. & Leven, E.R. 1 973. Upper Paleozoic deposits of central Afghanistan. Bull. Mosc. Soc. Lab. Res. Geol. 48 : 30-40.
Karpinsky, A. 1874. Geologische Untersuchungen im Gouvernment Orenburg, Verh. Min. Gesellsch. St. Petersbourg 9 : 2 1 2-3 10.
--- . 1 889. Ueber die Ammoneen der Artinsk-Stufe. Mem. A cad. Sci. (St. Petersbourg) ser. 7 3 7 (2).
Kashirtsev, A.S. 1 959. Field atlas of the fauna of the Permian deposits of the north-east of the USSR. Atlas (Fossil fauna). Leningrad Akad-nauk. USSR : 1 -85.
Kayser, E. 1 883. Obercarbonische Fauna von Lo-ping. In F. von Richthofen, China 4 (8) : 160-208.
Keidel, H. 1 906. Geologische Untersuchungen im sudlichen Tianschan nebst Beschreibung einer oberkarbonischen Brachiopodenfauna auf dem Kukurtuktal. Neues Jb. Miner. Geol. Palliont. Bei/Bd 22 : 266-384.
Kindle, E.M. 1 926. The occurrence of the genus Leptodus in the Anthracolithic fauna

References 1 93
of British Columbia. Trans. R . Soc. Can. IV: Geological Sciences and Mineralogy : 109- 1 1 1 .
King, P.B. 193 1 . The Geology of the Glass Mountains, Texas. Part I, Descriptive Geology. Bull. Texas Univ. 3038 : l - 167.
--- . 1937. Geology of the Marathon region, Texas. Prof Pap. U.S. geol. Surv. 187.
King, R.E. 193 1 . Geology of the Glass Mountains. Part 2 Faunal summary and correlarelation of the Permian formations with descriptions of the Brachiopoda. Bull. Univ. Texas 3042: 1 -245.
King, R.E., Dunbar, C.O., Cloud, P.E. & Miller, A.K. 1 944. Geology and Paleontology of the Permian area N.W. of Las Delicias, S.W. Coahuila, Mexico. Spec. Pap. Geol. Soc. Am. 52.
King, W. 1 850. A Monograph of the Permian Fossils of England. Palaeontogr. Soc. : 1 -258, London.
Kiparisova, L.D. Okuneva, T.M. & Oleynikov, A.N. 1 973. The Triassic System in the USSR. Mem. Can. Soc. Petrol. Geol. 2 : 1 37 - 149.
Konecki, M.C. Dickins, J.M. & Quinlan, T. 1 958. The geology of the coastal area between the Lower Gascoyne and Murchison Rivers, Western Australia. Rep. Bur. Miner. Resour. Geol. Geophys. Aust. 37.
Kotlyar, G.V. 1 964. New subgenera of the genus Productus from the Upper Permian of southern Primoyre. Trudy vses. nauchno issled. geol. lnst. n.s. 93 : 1 23- 1 27. (In Russian).
Kotlyar, G.V. & Popeko, L.l. 1 967. Biostratigraphy, Bryozoa and Brachiopoda of the Upper Paleozoic of Zabaikal. Zabaikal. Geogr. USSR 28 Trudy Proc. Geol. 5. (In Russian).
Kozlowski, R. 19 14. Les Brachiopodes du Carbonifere Superieure de Bolivie. A nnis. Paleont. 9 ( 1 ) : 1 - 100.
Kozubova, L.A. & Kulikov, M.V. 1959. New data on the Permian beds of west Zabaikal. DAN USSR 1 25 (5). (In Russian).
Kozur, H. l 973a. Beitrage zur Stratigraphie und Paiaontologie der Trias. Geologica Pa/aont. Mitt. Innsbruck 3 ( 1 ) : 1 -30.
--- . 1 973b. Beitrage zur Stratigraphie von Perm und Trias. Geologica, Paliiont., Mitt. Innsbruck 3 (3) : 1 -3 1 .
--- . 1974. Zur Alterstellung des Zechsteinkalkes (ca 1 ) innerhalb der 'Tethyalen' Permgliederung. Freiberger ForschHft C298 : 45-50.
Krassilov, V. 1 974. Causal biostratigraphy. Lethaia 7 (3) : 1 73 - 180. Kulikov, M.V. 1 938. Brachiopoda from the Ischimbaevo oil field. Trudy Lening. Neft.
Geol. razved. Inst. A 10 1 : 1 4 1 - 1 77, 1 88- 1 89. (In Russian and German). --- . 1942. On the nature of the Sylva reefs. C.r. Acad. Sci. USSR. 60 (9) : 375-
377. --- . 1 947. New data on the Brachiopoda of terrigenous facies of the Kungurian on.
the Western slope of the Urals. C.r. Acad. Sci. USSR n.s. 56 (6) : 63 1 -633. (In Russian).
--- . 1 950. On the systematics of Spiriferids from the Upper Permian of European SSSR. Trudy VSEGEI 1 : 3-7. (In Russian).
--- . 1 9 59. Stratigraphic outline of Permian deposits in Zabaikal. Dokl. A kad. nauk SSSR 1 25 (5) : 108 1 - 1084. (In Russian).
--- . 1 960. On the article by V.V. Lipatova 'New Data on the Kazanian Stage in the Aktyubinsk Urals'. Dokl. Akad. nauk SSSR 1 28 (4) : 804-805. (In Russian).
--- . 1 965b. On the Permian faunas from Zabaikal. Trudy vses. nauchno-issled. geol. lnst. 1 1 5 : 1 3-26. (In Russian).
--- . 1965a. On the Permian of Chironsk and its fauna. Sib. Sect. Geol. Geophys. Acad. Sci. USSR 4 : 1 25- 1 3 1 . (In Russian).

1 94 References
Kummel, B. 1 970. Ammonoids from the Kathwai Member, Mianwali Formation, Salt Range, West Pakistan. Stratigraphic boundary problems: Permian and Triassic of West Pakistan. Ed. Kummel, B . & Teichert, C . : Spec. Pub/. Univ. Kansas 4: 1 77- 1 92, pl . 1 -2 .
Kummel, B. & Teichert, C. 1 964. The Permian-Triassic boundary in the Salt Range of West Pakistan. Int. geol. Congr. Ind. 22Abstr : 1 20.
--- . 1 970. Stratigraphy and paleontology of the Permian-Triassic boundary beds, Salt Range and Trans-Indus Ranges, West Pakistan. Stratigraphic Boundary Problems. Permian and Triassic of West Pakistan. Ed. Kummel, B. & Teichert, C . : Spec. Pub/. Univ. Kansas 4 : 1 - 1 10.
--- . 1 973. The Permian-Triassic boundary in central Tethys. Mem. Can Soc. Petrol. Geol. 2 : 1 7-�4.
Lange, E. 1925. Eine mittelpermische Fauna von Guguk Bulat. (Padanger Oberland Sumatra). Verh. K. Ned. geol. mijnb. Genoot. geol. ser. 7 : 2 1 3-295.
Leffingwell, E. de K. 1 9 1 9. The Canning River region, northern Alaska. Prof Pap. U.S. geol. Surv. 1 09 : 1 -25 1 .
Legrand-Blain, M . 1 968. Spiriferacea carboniferes et Permiens de'Afghanistan central. Notes Mem. sur Moyen-Orient 9 : 1 87-253 .
Leven, E.Y. 1 967. Stratigraphy and Fusulinacea of Permian deposits of the Pamirs. Trudy geol. Inst. Acad. Nauk. USSR 167 : 1 -234.
Likharev, B.K. 1 9 13 . Die fauna der permischen Ablagerungen aus der Umgebung der Stadt Kirillow im Gouvernment Novgorod. Trudy geol. Kom. n.s. 85. (In Russian and German).
--- . 1 928. On some rare and new brachiopods from Lower Permian of north Caucasus. Izv. vses. geol. razved. Ob 'ed. 4 7 : 26 1 -296. (In Russian).
--- . 1 93 1 . Upper Permian of the northern province. Trudy glav. geol-razv. Uprav. VSNKh. 7 1 : 1 -43. (In Russian).
--- . 1 932. Fauna of the Permian deposits of Northern Caucasus. I. Brachiopods subfamily Orthotetinae Waagen. 1 1 . Family Lyttoniidae Waagen. Izv. vses. geol. razved. Ob 'ed 2 1 5 : 1 - 1 1 1 . (In Russian).
--- . 1 934. Die Fauna der permischen Ablagerungen des Kolymagebietes. Trudy Acad. Nauk. SSSR Jakut ser. 14 : 1 - 148. (In Russian and German).
--- . 1 937. Permian Brachiopods of North Caucasus. Families Chonetidae Hall and Clarke and Productidae Gray. Izv. vses. geol. paleont. raxved. Ob 'ed 3 9 ( I) : 1 - 1 5 1 . (In Russian).
1 939. Brachiopoda. In the Atlas of the leading forms of the fossil faunas of USSR Vol. V. The Middle and Upper Carboniferous. Ed. Gorsky, I : 79- 1 18.
--- . 1 946. Materials on brachiopod faunas of middle and upper Carboniferous and lower Permian of Fergana. Materialy VSEGEI 7. (In Russian).
--- . 1 959. The boundaries and principal subdivisions of the Permian System. Sov. Geol. 6 : 1 3-30. Qn Russian).
Likharev, B.K. & Einor, O.L. 1 939. Paleontology of the Soviet Arctic, Part IV : Contributions to the knowledge of the Upper Paleozoic fauna of Novaya Zemlya. Brachiopoda. Trudy arkt. naucho-issled lnst. 1 27 : 1 -245 . (In Russian, English summary).
Likharev, B.K. & Miklukho-Maklay, A.D. 1 964. Stratigraphy of the Permian System. In Stratigraphy of the Upper Paleozoic and Mesozoic of the southern biogeographical provinces. Int. Geol. Congr. 22 Rep. Sov. Geol. Probl. 1 6a: 1 2-24. (In Russian with English summary).
Logan, A. & McGugan, A. 1 968. Biostratigraphy and faunas of the Permian Ishbel Group, Canadian Rocky Mountains. J. Paleont. 42 (5): 1 1 23 - 1 1 39 .
Loney, R.A. 1 964. Stratigraphy and petrography of the Pybus-Gambier Area, Admiralty island, Alaska. Bull. U.S. geol. Surv. 1 1 78 : 1 - 103 .

References 1 95
Loweneck, B. 1 932. Beitrlige zur kenntnis des Palliozoikums im Tianshanschan. Sber. Akad. Wis. Wien (math-naturwis.) n.s. 1 1 : 5 - 140,
Mahlzahn, E. 1 937. Die Deutschen Zechsteinbrachiopoden. Preus. Geol. Landesanst. Abh. N. V. 1 8 5 : 1 -77.
Mansuy, H. I 9 I 2. Mission du Laos. 1. Geologie des environs de Luang Prabang. 2. Mission Zeil dans le Laos septentrional. Resultats paleontologiques. 1 1 Contribution a la Geologie du Tonkin. Cr. hebd. Seanc. Acad. Sci. Paris I (4) : I -9. --- . I 9 I 3 . Faunes des Calcaires a Productus de l'Indochine, Ser. l ., Mem. Serv.Geol. Indochine, C.r. hebd. Seanc. Acad Sci Paris (Fasc. 4) 2 : 23- 104; 107- 1 33 .--- . I 9 14 . Faunes des calcaires a Productus de l'Indochine. C.r. hebd. seanc.Acad. Sci. Paris 3 (3). --- . I 9 I 9. Catalogue general des fossiles recueilles en Indochine et en Yunnan. C.r.hebd. Seanc. Acad. Sci. Paris 6 (6).
--- . 1 920. Fossiles des calcaires ouralo-permiens du Tran-Ninh, Laos. C.r. hebd. Seanc. Acad Sci. Paris 7 ( 1 ) : 49-59.
Marks, P. 1 957 . Indonesia. Lex. Strat. Internal. III Asie 7a: 1 -24 1 . Martin, H., Walliser, O.H., Wilczewski, N . 1970. A goniatite from the Glaciomarine
Dwyka beds near Schlip, South West Africa. Second Gondwana Symposium S. Afr. Proc. Pap. : 62 1 -626.
Maxwell, W.G.H . . I 964. The geology of the Yarrol region. Part I Biostratigraphy. Pap. Dep. Geol. Univ. Qd. 5 (9): 1 -79.
McAlester, A.L. I 973. Phanerozoic biotic crisis. Mem. Can. Soc. Petrol. Geol. 2: I I - 16 . McGugan, A . 1 963. A Permian brachiopod and fusulinid fauna from the Elk Valley,
British Columbia, Canada. J.Paleont. 3 7 (3): 62 1 -627. McKee, E.D. 1 938. The environment and history of the Toroweap and Kaibab Forma
tions of northern Arizona and southern Utah. Pubis Carnegie Instn. 492. McKee, E.D. & Breed, W.J. 1 969. The Toroweap Formation and the Kaibab
Limestone. Spec. Pub!. New Mexico geol. Soc. 3 : 1 2-26. McKee, E.D., Oriel, S.S. & others 1 967. Paleotectonic Investigations of the Permian
System in the United States. Prof. Pap. U.S. Geol. Surv. 5 15 . McKelvey, V.E., Williams, J.S. Sheldon, R.P., Cressman, E.R., Cheney, T.M. &
Swanson, R.W. 1 959. The Phosphoria, Park City and Shedhorn Formations in the western phosphate field. Prof. Pap. U.S. geol. Surv. 3 I 3 A : I -4 7 .
Mendes, J.C. 1 959. Chonetacea e Productacea Carboniferos da Amazonica. Bolm Fac. Filos. Ci'enc. Univ. S Paulo 1 7 : 1 -83 .
Menner, V.V., Sarytcheva, T.G. & Chernyak, G.E. 1 970. Stratigraphy of the Carboniferous and Permian deposits of north Verchoyan. Trudy arkt. nauchno-issled Inst. 1 54 : 1 - I 9 1 . (In Russian).
Merla, G. 1 93 1 . La Fauna del Calcare a Bellerophon della regione dolomitica. Mem. Inst. Geo/. Min. Univ. Padova, 9 : 1 -22 1 .
--- . I 934. Fossili anthracolitici del Caracorum. Re/az. scient. Sped. ita!. Filippi Himalaia ( I 9 1 3- 1 9 1 4) ser. 2, 5 : 10 I -3 19.
Mertie, J.B. I 930. Geology of the Eagle-Circle District, Alaska. Bull. U.S. geol. Surv. 8 1 6 : I - I 68.
Metz, K. I 935. Choristiten aus den Karnischen Alpen. Sber. Akad. Wiss. Wien, mathnaturwiss Kl. I 44 : I 5 1 - 1 56.
Meyer, O.E. 1 922. Brachiopoden des Perm und Untercarbon der Residentschaft Djambi (Sumatra). Beitriige Geol. Paliiont. von Sumatra unter mitwirkung von Fachgenossen Herausgegeben von aug. Tobler, Basel No. 2.
Mikluko-Maklay, A.D. I958. On stages proposed for Moscovian to Permian beds of southern region of USSR. Doklady Acad. Sci. USSR I 20 ( I ) : 1 75- 1 78. (In Russian).

1 9 6 References
Miller, A.K. 1 93 1 . A new ammonoid fauna of late Paleozoic age from western Texas. J. Paleont. 4: 383-4 1 2.
Miller, A.K. & Furnish, W .M. 1 940. Permian ammonoids of the Guadalupe Mountain region and adjacent areas. Spec. Pap. Geol. Soc. Am. 26: 1 -244.
Miller, A.K. & Furnish, W.M. 1 957. Permian ammonoids from Tunisia. J. Paleont. 3 1 : 705 -7 1 2.
Mills, J .W. & Davis, J.R. 1 962. Permian fossils from the Kettle Falls Area, Stevens County, Washington. Contrib. Cushman Foundat. Foram. Res. 1 2 (2) : 4 1 -5 1 .
Miloradovich, B.V. 1 935 . Materials for the study of Upper Paleozoic Brachiopoda from the northern island of Novaya Zemlya. Trudy arkt. nauchno-issled Inst. 1 9 : 1 -1 76 . (In Russian).
--- . 1 936. Lower Permian fauna from the island Mejdusharsky (southern island of Novaya Zemlya). Trudy arkt. nauchno-issled Inst. 20 : 37-82. (In Russian).
--- . 1 940. Quantitative distribution in brachiopods at the C arboniferous-Permian boundary. C.r. Acad. Sci. USSR. 2 7 : 1 007- 1 0 1 0.
--- . 1 949. On the vertical extent of Upper Carboniferous and Lower Permian brachiopods from the Ufimian platform. Izvest. A cad. Nauk. SSSR Geol. ser. 1 :53-66.
Minato, M. et al. 1 954. Zur Biostratigraphe der Permischen Formation des Setamai Gelandes in Sud-Kitakami Gebirge. J. geol. Soc. Jap. 60: 378-387.
Mirchink, M. 1 938. Materials for the study of the Pinega Zechstein Brachiopoda. Trudy Geol. Inst. Mosc. Acad. Sci. USSR 7 : 1 3 1 -346.
Mironova, M.G. 1 960. Brachiopoda from the Mergelist horizon of Vorkut River, (Petchora Basin). Rep. Paleont. Biostrat. NIIGA 22. (In Russian).
--- . 1 964. On the Permian brachiopods from the Petchora Basin. Paleont. Prob. 4: 85-99.
--- . 1 967. Late Carboniferous brachiopods from Bashkiri. Pub/. Leningrad Univ. : 1 -63.
Moeller, V. von. 1 877. Uber Fusuline and ahnliche Foraminiferenformen des russischen Kohlenkalks. (Vorliiufige Notiz). N. Jb. Mineral. Geol. 1 3 9- 1 46, Stuttgart.
Moffitt, F.H. 1 938a. Geology of the C hitina Valley and adjacent area, Alaska. Bull. U.S. geol. Surv. 894: 1 - 1 37 .
--- . 1 938b. Geology of the Slana-Tok District, Alaska. Bull. U.S. geol. Surv. 904 : 1 -54.
Monger, J .W.H. & Ross, C.A. 1 97 1 . Distribution of Fusulinaceans in the western Canadian Cordillera. Can. J. Earth Sci. 8 (2) : 259-278.
Moore, R.E. (Ed.) 1 965. Treatise on Invertebrate Paleontology. Part H Brachiopoda 1 , 2 , Geol. Soc. Am. Univ. Kansas Press.
Mu An-tze, Wen Shih-Hsuan, Wang Yi-kang & Chang Ping-Kao. 1 973. Stratigraphy of the Mount Jomlo Lungma region in southern Tibet, China. Scientia Sin. 1 6 ( 1 ): 1 -1 1 1 . (In Chinese and English).
Mudge, M.R. & Yochelson, E.L. 1 963. Stratigraphy and paleontology of the uppermost Pennsylvanian and lowermost Permian rocks in Kansas. Prof. Pap. U.S. geo/. Surv. 323 : 1 -23 1 .
Muffler, L.J.P. 1 967. Stratigraphy of the Keku Islands and neighbouring parts of Kuiu and Kupreanof islands, south-eastern Alaska. U.S. geol. Surv. Bull. 1 24 1 C: 1 -52.
Muir-Wood, H.M. & Cooper, G.A. 1 960. Morphology, classification and life habits of the Productoidea (Brachiopoda). Mem. Geo/. Soc. Am. 8 1 .
Muir-Wood, H.M. & Oakley, K.P. 1 94 1 . Upper Palaeozoic faunas of North Sikkim. Mem. geol. Surv. India Palaeont. indica n.s. 3 1 ( 1) .
Murata,. M. 1 9.64. Ge?logical age of the Kanokura formation in the southern part of the
K1takamJ massif, northeast Japan. Saito Ho-on Kai Mus. Res. Bull. 33 . Murchison, Sir R.I . 1 84 1 . 'Letter to Dr. Fischer v. Waldheim', Phil. Mag. n.s. 1 9 : 4 1 8 .

References 1 97
Mutch, A.R. 1 973. GeolQgy of the Morley subdivision, Southland. Bull. N.Z. geol. Surv. 78 : 1 - 1 04.
Nakamura, K. 1959. Spinomarginifera found in Japanese Permian. Proc. Palaeont. Soc Japan n.s. 35 : 143- 146.
--- . 1960. Some Lower Permian Sakamotozawa brachiopods. J. Fac. Sci. Hokkaido Univ. ser. 4 10 ( 1) : 1 99-207.
Nakazawa, K. 1973. On the Permian fossils from Jengka Pass, Pahang, Malay Peninsula. Sci. Rep. Tohoku Univ. ser. 2, Geol. Spec. 6 : 277-296.
--- . 1 97 1 . The Lower Triassic Kurotaki Fauna in Shikoku and its allied faunas in Japan. Mem. Fac. Sci. Kyoto Univ. Geol. Miner. ser. 38 ( 1 ) : 103- 1 33 .
Nakazawa, F . & Newell, N.D. 1 968. Permian bivalves of Japan. Mem. Fac. Sci. Kyoto Univ. Geol. Miner. ser. 35 ( 1 ) : 1 - 108.
Nakazawa, K.; Kapoor, H.M., Ishii, K . ; Bando, Y. ; Maegoya, T. ; Shimizu, D. ; Nogami, Y.; Tokuoka, T. & Nohda, S. 1970. Preliminary report on the Permo-Trias of Kashmir. Mem. Fac. Sci. Kyoto Univ. Geol. Miner. ser. 37 (2) : 1 63-1 72.
Nalivkin, V.D. 1949. Stratigraphy and tectonics of Ufa Plateau and Yurezano-Sylvenian depression. Trudy VNIGRI n.s. 46 : 204. (In Russian).
--- . 1973. Geology of the USSR. Translator Rast. N. Oliver & Boyd, Edinburgh. Nassichuk, W.W. 1970. Permian ammonoids from Devon and Melville islands. Cana
dian Arctic Archipelago. J. Paleont. 44 : 77-97. --- . 197 1 . Permian ammonoids and nautiloids, southeastern Eagle Plain, Yukon
Territory. J. Paleont. 45 : 100 1 - 102 1 . Nassichuk, W.W., Furnish, W.M. & Glenister, B.F. 1 965. The Permian ammonoids, of
Arctic Canada. Bull. Geol. Surv. Can. 1 3 1 : 1 -56. Nassichuk, W.W. & Spinosa, C. 1 972. Early Permian (Asselian) ammonoids from the
Hare Fiord Formation, northern Ellesmere Island. J. Paleont. 46 : 536-544. Nassichuk,W.W. : Thorsteinsson, R. & Tozer, E.T. 1 973. Permian-Triassic boundary in
the Canadian Arctic Archipelago. Mem. Can. Soc. Petrol. Geol. 2: 286-293. Nechaev, A.V. 1 9 1 1 . Die fauna der Perm-Ablagerungen von Osten und von aussersten
Norden des europaischen Russland. Trudy geol. Kom. n.s. 6 1 . (In Russian and German.)
Newell, N.D. 1 957 . Supposed Permian tillites in northern Mexico are submarine slide deposits. Bull. Geol. Soc. Am. 68 : 1 569- 1 5 76.
--- . 1 973. The very last moment of the Paleozoic Era. Mem. Can. Soc. Petrol. Geol. 2 : 1 - 10.
Newell, N.D., Chronic;J. & Roberts, T.G. 1 953. Upper Paleozoic of Peru. Mem. Geol. Soc. Am. 58.
Newell, N.D., K night, J .B. , Moore, R.C. & Brill, K .G. 1 940. Invertebrate fauna of the late Permian Whitehorse Sandstone. Bull. Geol. Soc. Am. 5 1 : 26 1 -336.
Newell, N.D. & Kummel, B. 1 942. Lower Eo-Triassic Stratigraphy, western Wyoming, and southeast Idaho. Bull. Geol. Soc. Am. 5 3 : 937-966.
Nicol, R.S. 1 975. The effect of late Carboniferous-early Permian glaciation on the distribution of Permian conodonts in Australasia. Abst. Geol. Soc. Am. 7 (6) : 828-829.
Oliviera, E.P. DE 1 936. Urn brachiopoda Carbonifero do Rio Moa, Territorio do Acre. Brasil, Notas Ser. Geol. Miner. Prelim. Estud. 6 : 1 .
Onuki, Y. 1 956. Geology ofthe Kitakami Massif(with geological maps and explanatory text). lwate Prefecture: 1 - 1 82. (In Japanese).
Ozaki, K. 1 93 1 . Upper Carboniferous Brachiopods from North China. Shanghai Sci. lnst. Bull. 1 (6) : 1 -205 .
Packham, G.H. (Ed.) 1 969. The geology of New South Wales. J. geol. Soc. Aust. 1 6 ( 1 ). Pasini, M. 1963. Alcuni Fusulinida del Monte Auernig (Alpi Carniche) loro significato
Stratigraphico. Riv. ita/. Pa/eont. 69 (3) : 337-382.

1 98 References
Pattison, J. 1 970. A review of the marine fossils from the Upper Permian rocks ofnorthern Ireland and north-west England. Bull. Great Brit. Geo/. Surv. 32 : 1 23-1 65 .
Pattison, J., Smith, D.B. & Warrington, G . 1 973 . A review of Late Permian and EarlyTriassic Biostratigraphy in the British Isles. Mem. Can. Soc. Petrol. Geo/. 2 : 220-260.
Pavlov, A.M. 1 965. New ammonoid suites from Permian deposits of the Pamirs. Dokl. Akad. Nauk SSSR Geol. 1 64 ( 1).
Pederson, S.L. 1 954. A Permian (Wolfcampian) fauna of the Casper formation of southeastern Wyoming. J. Paleont. 28 ( 1 ) : 1 7-2 1 .
Plodowski, G. 1 970. Stratigraphie und Spiriferen (Brachiopoda) des Palaozoikums der Dascht-e-Nawar/SW (Afghanistan). Palaeontographica A 1 34 : 1 - 1 32.
Plotnikov, M.A. & Molin, V.A. 1 969. Upper Permian and Triassic deposits of west PreTiman. Com. Fi/. Inst. Geol. Leningr. : 1 - 106.
Prendergast, K.L. 1 943. Permian Productinae and Strophalosiinae of Western Australia. J. R. Soc. WAust. 28 : 1 -73.
Ramovs, A. 1 958. Die Entwicklung des Oberperms im Bergland von Skofjahoka und Polhov Gradic. W. Slowenien, Ljubljana 4 (4) : 45 1 -622.
--- . 1 963. Biostratigraphie der Trogkofelstude in Jugoslavien. N. Jahrb. Geol. Palaont. Monatsheft 1 963, 7 : 382-388.
Rauser-Chernossova, D.M. 1 937 . On Fusulinids and stratigraphic succession of the oilbearing limestone in the Sterlitakmak region. Dokl. Akad. Nauk. USSR, n.a. Geol. 1 3 (7).
--- . 1 940. Stratigraphy of the Upper Carboniferous and Artinskian Stage of the west slopes of the Urals and data on the Fusulinacean faunas. Trudy lnst. geol. Sci. Akad. Nauk SSSR 7, ser. geol. 22.
--- . 1 949. The stratigraphy of upper Carboniferous and Artinskian deposits of Bashkirian Cisuralia. Trudy lnst. Geol. Sci. Akad. Nauk SSSR 105 : 3-2 1 .
Reed, F.R.C. 1 925. Upper Carboniferous fossiis from Chitral and the Pamirs. Mem. geol. Surv. India Palaeont. indica n.s. 6 (4) : 1 - 1 54.
--- . 1 927a. Palaeozoic and Mesozoic Fossils from Yunnan. Mem. gepl. Surv. India Palaeont. indica n.s. 10 ( 1 ) : 1 -29 1 .
--- . 1 927b. Upper Carboniferous fossils from Argentina. Pubis. Carnegie. Instn. 38 1 : 1 3 1 - 1 49.
--- . 1 928. A Permo-Carboniferous marine fauna from the Umaria coal-field. Rec. geol. Surv. India 60 (4) : 367-398.
--- . 1 930. Upper Carboniferous Fossils from Tibet. Mem. geol. Surv. India Palaeont. indica n.s. 1 6.
--- . 1 93 la. New fossils from the Productus Limestone of the Salt Range. Mem. geol. Surv. India Palaeont. indica n.s. 1 7.
--- . 1 93 1 b. Upper Carboniferous fossils from Afghanistan. Mem. geo/. Surv. India Palaeont. indica 1 9.
--- . 1932. New fossils from the Agglomeratic Slate of Kashmir. Mem. geo/. Surv. India Palaeont. indica 20 ( I ).
--- . 1 933. Anthracolithic Faunas of the Southern Shan States. Rec. geol. Surv.India 67 ( I ) : 83- 1 34.
--- . 1 936. Some fossils from the Eurydesma and Conularia beds (Punjabian) of the Salt Range. Mem. geol. Surv. India Palaeont. indica 23 ( I ).
--- . 1 944. Brachiopoda and Mollusca of the Productus Limestone of the Salt Range. Mem. geo/. Surv. India Palaeont. indica n.s. 23 (2).
Reichardt, W. 1 932. Die Ostalpinen Nassfeldschichten-eine Brucke zwischen Mitteleurope und Russland. C.r. 2'me. Congr. avanc. etudes strat. Carbonifere.
Renz, C . 1 940a. Die Paliiozoischen Faunen von 1 929-30. Wissen, Ergeb. Nieder/. Ex-

References 1 99
ped. Karakorum 3 ( 1 ), Geol. Abt. 1 : 9-68, E.J. Brill, Leyden. --- . 1 940b. Die PaHtozoischen Faunen von 1 935 . Metazoa. Wissen, Ergeb.
Niederland. Exped. Karakorum 3 ( 1 ) Geol. 2 : 1 1 8-247. Rocha-Campos, A.C. 1 9 7 1 . Upper Paleozoic bivalves and gastropods of Brazil and
Argentina. A review. Proc. Pap. Second Gondwana Sympos. S. Afr. 605-6 14 . Roemer, C.F. 1 880. Ueber eine Kohlenkalk Fauna des Westkusts von Sumatra.
Palaeontographica 27 : 1 - 1 1 . 1 ·
Ross, C.A. 1 962. Fusulinids from the Leonard Formation, (Permian), western Glass Mountains, Texas. Contr. Cushman Found. Foram. Res. 1 3 : 1 -2 1 .
--- . 1 963. Standard Wolfcampian Series (Permian), Glass Mountains, Texas. Mem. Geol. Soc. Am. 88.
--- . 1 965. Fusulinids from the Cyathophyllum limestone, Central Vestspitsbergen. Contr. Cushman Found. Foram. Res. 1 6 (2): 74-86. /
--- . 1 967a. Late Paleozoic Fusulinacea from Northern Yukon Territory. J. Paleont. 4 1 (3) : 709-725.
--- . 1 967b. Development of Fusulinid (Foraminiferida) faunal realms. J. Paleont. 4 1 (6): 134 1 - 1 354.
--- . 1 973. Pennsylvanian and early Permian depo�itional history, south-eastern Arizona. Bull. Am. Assoc. Petrol. Geol. 5 7 : 887-9 12 .
Ross, C.A., Nassichuk, W.W. 1 970: Yabeina and Waagenoceras from the Atlin Horst area, northwestern British Columbia. J. Paleont. 44 : 779-780.
Rostovtsev, K.O. & Azaryan, N.R. 1 97 1 . Paleozoic-Mesozoic Boundary in Transcaucasia. Dokl. Akad. Nauk. SSSR 1 99' (2) : 4 1 8-42 1 . (In Russian).
--- . 1973. The Permian-Triassic boundary in Transcaucasia. Mem. Can. Soc. Petrol Geol. 2 : 89-99.
Rothpletz, A. 1 892. Die Perm-, Trias-, und Jura-Formation auf Timor und Rotti im indeschen Archipel. Palaeontographica 39 : 57- 106.
Rudwick, M.J.S. & Cowen, R . 1 968. The functional morphology of some aberrant Strophomenide brachiopods from the Permian of Sicily. Boll. Soc. Paleont. ita/. 6 (2).
Runnegar, B. 1 969a. Mollusca and Brachiopods from the base of the Permian sequence, South Coast of New South Wales. Stratigraphy and Palaeontology: Essays in Honour of Dorothy Hill. Ed. K.S.W. Campbell. Aust. Nat. Univ. Press, Canberra : 276-298.
--- . 1969b. The Permian faunal succession in eastern Australia. Spec. Sympos. geol. Soc. Aust. 2 : 73-98, 1 pl.
Runnegar, B. & Ferguson, J.A. 1 969. Stratigraphy of the Permian and Lower Triassic marine sediments of the Gympie District, Queensland. Pap. Dep. Geol. Univ. Qd. 6 (9) : 247-28 1 .
Ruzencev, V.E. 1 934. New data on the stratigraphy of the Artinskian Stage on the west slopes of the Urals. Oil Economy 6.
--- . 1 952 . Biostratigraphy of the Sakmar Stage in the Aktyubin Region of the Kazakhstan SSR. Trudy palaeont. lnst. 42: 1 -87. (In Russian).
--- . 1956. Lower Permian ammonities of the Southern Urals. 1 1 . Ammonites of the Artinskian Stage. Trudy palaeont. Inst. 60. (In Russian).
--- . 1962. (Ed.) Mollusca. Osnovi Palaeont. Pub. Acad. Sci. USSR. (In Russian). --- . 1 965. Principal ammonoid assemblages of Carboniferous Period. Palaeont.
Zhurn. 1 965, 2: 3 - 1 7. (In Russian). Ruzencev, V.E. & Sarytcheva, T.G. 1 965. The development and succession of marine
organisms at the Palaeozoic-Mesozoic boundary. Trudy palaeont. lnst. 108. (In Russian).
Sahni. M.R. & Dutt, O.K. 1959. Argentine and Australian affinities in a Lower Permian

200 References
fauna from Manendragarh, Central India. Rec. geol. Surv. India 87 (4) : 655-670.
Sahni, M.R. & Srivastava, J.P. 1 956. Discovery of Eurydesma and Conularia in the eastern Himalayas and description of associated faunas. J. Palaeont. Soc. Ind. 1 , ( 1 ) : 202-2 14 .
Sakagami, S. 1 968. Permian Bryozoa from Khao Phrik, near Rat Buri, Thailand. Geol.Pa/aeont. southeast Asia 6 : 45-66.
Samtleben, C. 1 97 1 . Zur kenntnis der Produktiden und Spiriferiden des Bolivianischen Unterperms. Brundes. Niedersachs. Landes. Bodenforschung Beih. 1 1 1 : 1 - 163.
Sarytcheva, T.G. (Ed.) 1 968. Brachiopods from the Upper Palaeozoic of West Kazakhstan. Trudy palaeont. lnst. 1 2 1 . (In Russian).
Sarytcheva, T.G. & Sokolskaia, A.N. 1 952. Guide to the identification of Palaeozoic brachiopods from the Moscow Basin. Trudy palaeont. lnst. 38 .
Sarytcheva, T.G. & Waterhouse, J .B. 1 972. New bral;hiopod species of the Family Retariidae from the Permian of northern Canada. Palaeont. Zhurn. 6 (4): 62-74, pl. 7, 8 (In Russian).
Sastry, M.V.A. & Shah, S.C. 1 964. Permian Marine transgression in Peninsular India. Abstr. Int. geol. Congr Ind. 22 : 1 4 1 - 1 42.
Schellwien, E. 1 892. Die fauna des Karnischen Fusulinenkalks. Palaeontographica B 39 : 9 1 - 1 36.
Schellwien, E. 1 900. Die fauna des Trogkofelschichten in den karnischen Alpen und den Karawanken. Jb. geo/. Bundesant., Wien Abh. 1 6, ( 1 ).
Schenk, H.G., et a/; Hedberg, H.D. ; Tomlinson, C .W. ; Eaton, J.E. & White, R.T. 194 1 . Stratigraphic nomenclature. Bull. Am. Assoc. Petrol. Geol. 255 : 2 1 95-22 1 1 .
Schindewolf, O.H. 1 954. Ueber die Faunenwende vom Palaozoikum zum Mesozoikum Zeitsch. Deutsch. Geol. Ges. 105 : 1 54- 1 83 .
Seelmeier, H. 1 937 . Versteinerungen aus den Rattendorfer Schichten. Mitt. natur. Ver. Steierm. 74: 98- 1 1 3 .
Sestini, N.F. 1 965a. The Geology of the upper Djadjerud and Lar Valleys (North Iran). 1 1 . Paleontology, Bryozoa, Brachiopods and Molluscs from Ruteh Limestone (Permian). Riv. ita/. Paleont. 7 1 ( 1 ) : 1 3- 1 10 .
--- . 1 965b. Permian fossils of the upper Hunza Valley. /tal. Exped. to Karakorum (K2) and Hundu Kush. Sci. Rep. IV Paleont. Zoo/. Bot. : 1 35 - 148. E.J. Brill, Lei den.
--- . 1 965c. The geology of the Upper Djadjerud and Lar Valleys (North Iran). 1 1 . Paleontology. Brachiopods from Dorud Formation. Riv. ita!. Paleont. 7 1 (3): 733-788.
--- . 1 965d. Permian fossils of the Shaksgam Valley. Ita/. Exped. to Karakorum (K2) and Hindu Kush. Sci. Rep. IV Paleont. Zool. Bot. : 1 49-2 1 5 . E.J. Brill, Lei den.
--- . 1 966. The geology of the Upper Djadjerud and Lar Valleys (North Iran) 1 1 . Palaeontology. Brachiopods from Geirud Formation, Member D (Lower Permian). Riv. ita/. Paleont. 72 ( 1 } : 9-50.
Sestini, N.F. & Glaus, M. 1 966. Brachiopods from the Upper Permian Nesen Formation (North Iran). Riv. ita!. Paleont. 72 (4) : 887-930.
Sheldon, R.P. 1 963. Physical stratigraphy and mineral resources of Permian rocks in western Wyoming. Prof Pap. U.S. geol. Surv. 3 1 3 B: 49-273.
Sheng, J .C. 1 964. Permian Fusulinids of Kwangsi, Kueichow and Szechuan, Palaeont. sin. ser. B 149 ( 1 0) : 1 29-247. (In Chinese with English summary).
Sheng, J.C. & Lee, H.H. 1 974. Recent advances in the Permian biostratigraphy of China. Mem. Nanking Inst. Geol. Palaeont. 5 : 1 1 7- 1 22.
Sherlock, R.L. 1955. The Permian-Triassic formations, a world review. Hutchinson's, London, 367 pp.

References 201
Shimizu, D. 196 1a. Brachiopod fossils from the Permian Maizuru Group. Mem. Coli. Sci. Univ. Kyoto Geol. Miner. B27 (3) : 309-342.
--- . 1 96 l b. Brachiopod fossils from the Upper Permian Gujo Formation of the Maizuru Group, Kyoto Perfecture, Japan. Mem. Coli. Sci. Univ. Kyoto ser. B 28 (4).
--- . 1 962. The Permian Maizuru Group, its stratigraphy and syntectonic succession through the latest Paleozoic orogeny. Mem. Coli. Sci. Univ. Kyoto 28 (4): 5 7 1 -609.
Shimizu, D. et a!. 1962. Stratigraphy of the Permian Maizuru Group, S.W. Japan. A study on the stratigraphy and geologic structure of the Maizuru zone. Pt. I 0. J. geol. Soc. Japan 68 (80Q): 237-247.
--- . 1 963. Permian brachiopod fossils of the Maizuru Group found on the north of Okayama City, Japan. Mem. Coli. Sci. Univ. Kyoto 30B (2) : 69-80.
--- . 1966. Permian brachiopod fossils of Timor. (Palaeontological study of Portuguese Timor, 3). Mem. Coli. Sci. Kyoto Univ. 32 B (4) : 40 1 -427.
Shreter, Z. 1 963. Die brachiopoden aus dem oberen Perm des Bukkgebirges in Nordungarn. Geol. Hungarica, S. Paleont. 28 : 79- 1 79.
Siehl, A. 1 967. Zur stratigraphie und Paleogeographie des Perm in Afghanistan. Geol. Rund. 56 (3): 795-8 1 2.
Simic, V. 1 932. Oberkarbonische Vetsteinerungen von Lika in Kroatian. Gornjokarbon. Fosili like U Hravatskoj. : 1 4 1 - 145.
--- . 1935a. Oberkarbonische Versteinerungen aus der Li-ka in Kroatien. Bull. Serv. geol. R. Yougosl. 4 : 1 4 1 - 145.
--- . 1935b. Einige Versteinerungen aus dem Oberperm von Westserbien. Bull. Serv. geol. R. Yougosl. 4 : 147- 1 56.
Singh, T. 1973. Note on the upper Palaeozoic fauna from Subansiri district, Arunachal Pradesh. Himal. Geol. 3 : 40 1 -4 10.
Skinner, J.W. 1 97 1 . New lower Permian fusulinids from Culbertson County, Texas. Pap. Univ. Kansas Paleont. Contrib. 5 3 : 1 - 1 0.
Slusareva, A.D. 1 960. Spiriferidae from the Kazanian beds of the Russian platform and the conditions .of their existence. (Genera Licharewia Einor and Permospirifer Kulikov). Trudy palaeont. Inst. 80: 1 - 1 20. (In Russian).
Solomina, R.V. 1 960. Some Permian brachiopods from Pai Hoi. Sbornik Stat. Palaeont. Biostrat. 1 9 : 24-73. (In Russian).
Spath, L.F. 1 934. Catalogue ofthe Fossil Cephalopoda in the British Museum (Natural History). Part IV The Ammonoidea of the Trias (London): 1 -52 1 . British Museum of Natural History, London.
Spinosa, C., Furnish, F.M. & Glenister, B.F. 1 970. Araxoceratidae, Upper Permian Ammonoids from the Western Hemisphere. J. Paleont. 44 (4) : 730-736.
Stache, G. 1 878. Zur Fauna der Bellerophonkalke Sudtirols. Jb. Kais. Kon. Geol. Reichsanstalt 28: 93- 1 68 .
Stehli, F.G. 1 954. Lower Leonardian Brachiopoda of the Sierra Diablo. Bull. Am. Mus. Nat. Hist. 105 (3).
Stehli, F.G. & Grant, R.E. 1 970. Permian brachiopods from Huehuetenango, Guatemala. J. Paleont. 44 ( 1 ) : 23-36.
Stehli, F.G. & Grant, R.E. 1 97 1 . Permian brachiopods from Axel Heiberg Island, Canada, and an index of sampling efficiency. J. Paleont. 45 (3): 502-52 1 .
Stepanov, D.L. 1 934. The Brachiopoda of the Bryozoa limestone of the Kolwa River region (northern Ural), Part I Strophomenacea. Trudy Neft. Geol. Razved. lnst. A 37 : 1 -63. (In Russian).
--- . 1936. Contribution to the knowledge of the brachiopod fauna of the Upper Palaeozoic of Spitsbergen. Annis. Budnoff State Univ. ser. geol. geogr. 9 (2): 1 14-1 28 . (In Russian).

202 References
--- . 1 93 7a. Permian Brachiopoda of Spitsbergen. Trudy A rkt. lnst. 76 : 105- 192. (In Russian).
--- . 1 937b. Brachiopods of the Bryozoan limestone of the Kolva area (Northern Urals), Pt. 2. Pub!. Leningrad State Univ. : 1 - 10 1 . (In Russian).
--- . 1 95 1 . Upper Palaeozoic of the western slope of the Urals. Trudy VNIGRI Geo/. lnst. n.s. 54. (In Russian).
--- . 1 95 7. A new stage of the Permian System in the Arctic. Messenger Leningrad State Univ. geo/. geog. ser. 4, 24 : 20-24. (In Russian).
--- . 1 973 . The Permian System in the USSR. Mem. Can. Soc. Petrol. Geo/. 2: 1 20- 1 36.
Stepanov, D.L., Golshani, F. & Stocklin, J . 1 969. Upper Permian and Permian-Triassic boundary in north Iran. Rep. Geo/, Surv. Iran No. 1 2 : 1 -72.
Stojanovic-Kuzenko, S. 1 963. Upper Permian brachiopods of Crikvenac (Western Serbia). Vesnik Zav. Geol. Geofiz. lstraz. A 2 1 : 1 �9- 1 80.
Sweet, W.C. 1970a. Uppermost Permian and Lower Triassic Conodonts of the Salt Range and Trans-Indus Ranges, West Pakistan. Stratigraphic Boundary Problems: Permian and Triassic of West Pakistan. Ed. Kummel, B. & Teichert, C . Spec. Pub/. Univ. Kansas 4 : 206-275.
--- . 1 970b. Permian and Triassic conodonts from a section at Guryul Ravine, Vihi District, Kashmir. Univ. Kansas Contrib. Paper Paleont. 49 : 1 - 10.
--- . 1 973. Late Permian and early Triassic conodont faunas. Mem. Can. Soc. . Petrol. Geol. 2 : 630-646.
Sweet, W.C . ; Mosher, L.C . ; Clark, D.L. ; Collinson, J .W. ; Hasenmueller, W.A. 197 1 . Conodont biostratigraphy of the Triassic. Mem. Geo/. Soc. Am. 1 2 7 : 44 1-465.
Taraz, H. 1 97 1 . Uppermost Permian and Permian-Triassic transition beds in Central Iran. Bull. A m. Assoc. Petrol. Geo/. 5 5 : 1 280- 1 294.
--- . 1 973 . Correlation of the uppermost Permian in Iran, Central Asia and South China. Bull. Am. Assoc. Petrol. Geo/. 5 7 : 1 1 1 7- 1 1 33 .
Teichert, C. 1 965. Nomenclature anp correlation of Permian 'Productus Limestone , Salt Range, West Pakistan. Rec. geol. Surv. Pakistan 1 5 ( I ) : 1 -20.
Teichert, C. , Kummel, B. & Sweet, W. 1 973 . Permian-Triassic strata, Kuh-e-Ali Bashi, northwestern Iran. Bull. Mus. Compar. Zoo/. Harvard 1 45 (8).
Termier, H. & Termier, G. 1 957 . Contribution a L'etude des Brachiopodes Permiens du Djebel Tebaga (Extreme Sud Tunisian). Bull. Soc. geo/. Fr. (6) 7 : 1 97-2 14.
--- . 1 970. Ammonoides de I' Artinskien (=Zygarien) dans Ia montagne de Bamyan (Afghanistan). Ann. Soc. Geo/. Nord. 90: 93- 100.
Termier, H . ; Termier, G . ; Desparmet, R. & Montenat, C. 1 972. Les ammonoides du Permien (Kubergandien) de Tezak (Afghanistan central). Ann. Soc. Geol. Nord. 92 : 105- 1 1 5 .
Termier, H . ; Termier, G. ; Lapparent, A.F. De; & Marin, P . 1 974. Monographie du Permo-Carbonifere de Wardak. (Afghanistan central). Doc. Labor. Geol. Fac. Sci. Lyon, Geobios ser. 2 : 1 - 1 6 7.
Thomas, G.A. 1957. Oldhaminid brachiopods in the Permian of northern Australia. J. Paleont. Soc. Ind. 2 : 1 74- 1 82.
--- . 1958 . The Permian Orthotetacea of Western Australia. Bull. Bur. Miner. Resour. Geol. Geophys. A ust. 39.
--- . 1967. The Permian brachiopod faunas of Western Australia. Simpos. lnternat.Stratig. Paleont. Gondw. Res. Assoc. Geol. A rgent. 1 96 7 : 2 1 7-234.
Thomas, G.A. & Dickins, J.M. 1 954. Correlation and age of the marine Permian formations of Western Australia. J. Sci. Aust. 16 (6) : 2 1 9-223.
Thompson, M.L. 1946. Permian fusulinids from Afghanistan. J. Paleont. 20 : 140- 1 57 . --- . 1 948. Studies of American Fusulinids. Contrib. Pa/eont. Univ. Kansas,
Protozoa Art. 1 : 1 - 1 84.

References 203
Thorsteinsson, R. 1974. Carboniferous and Permian stratigraphy of Axel Heiberg Island and western Ellesmere Island, Canadian Arctic Archipelago. Bull. Can. Geo/. Surv. 242: 1 - 1 1 5 .
Ting, Pei-Chen, 1963. Some Upper Permian brachiopods from Tibet. Acta palaeont. sin. 10 (4): 459-483 . (In Chinese).
Toriyama, R. 1963. The Permian. Geology of Japan. Ed. Takai, F., Natsumoto, T., & Toriyama, R . : 43-58. Univ. California Press, Berkeley & Los Angeles.
--- . 1967. The Fusulinacean zones of Japan. Mem. Fac. Sci. Kyushu Univ. Ser. D. : Geol. 1 8 : 3.5-260.
--- . 1973. Upper Permian Fusulinacean zones. Mem. Can. Soc. Petrol. Geol. 2: 498-5 1 2.
Tozer, E.T. 1967. A Standard for Triassic Time. Bull. Geol. Surv. Can. 156 : 1 - 103. --- . 1969. Xenodiscacean ammonoids and their bearing on the discrimination of
the Permian-Triassic boundary. Geo/. Mag. 106 (4) : 348-36 1 . 197 1 . Triassic Time and ammonoids : problems and proposals. J. Can. Earth
Sci. 8 (8) : 889- 103 1 . --- . 1973. The earliest marine Triassic rocks, their definition, ammonoid fauna, dis
tribution and relationship to underlying formations. Mem. Can. Soc. Petrol. Geol. 2: 549-556.
Ueda, F. 1963. The geological structure of the Permian and Triassic Systems in the Toyoma and Maiya districts, southern Kitakami massif, north-east Japan. J. Toyo Univ. (Nat. Sci.) 4: 1 -78. ( In Japanese and English).
Ustritsky, V.I. 197 1 . Biostratigraphy of the Upper Palaeozoic of Arctic regions. Trudy arkt. nauchno-issled Inst. 1 64: 1 -279. (In Russian).
--- . 1 973 . Permian climate. Mem. Can. Petrol. Geol. Soc. 2 : 733-744. --- . 1974. The bipolarity of late Palaeozoic faunas. Palaeont. Zhur. 1 974 (2) : 33-
3 7 . (In Russian). Ustritsky, V.I. & Chernyak, G.E. 1 963. Biostratigraphy and brachiopods of the Upper
Paleozoic of Taimyr. Trudy arkt. nauchno-issled Inst. 1 34. (In Russian). Ustritsky, V.I. & Chernyak, G.E. 1 967. Permian deposits of Taimyr Peninsula.
Palaeozoic stratigraphy of north Taimyr. A kad. Nauk. USSR Siber. Sect. : 220-223. (In Russian).
Verneuil, E. De. 1845 . Paleontologie, pp. 1 7-395. IN Murchison, R.I., Verneuil, E. de & Keyserling, A. Geologie de Ia Russie, D 'Europe et des Montagnes de l'Oura/, 2. John Murray, London and Bertrand, Paris.
Visscher, H. 1 973. The Upper Permian of Western Europe-a palynological approach to chronostratigraphy. Mem. Can. Soc. Petrol Geol. 2 : 200-2 19.
Volgin, V.I. 1960. Brachiopoda of the Upper Carboniferous and Lower Permian deposits of southern Fergana. Zhdanov Leningrad State Univ. : 1 - 1 58 . (In Russian).
Waagen, W. 1 89 1 . Salt Range Fossils. Mem. geol. Surv. India Palaeont. indica ser. 1 3 : 4 (2) : 89-242.
Wagner, R.H. & Prins, C .F.W. 1 970. The stratigraphic succession, flora and fauna of Cantabrian and Stephanian A rocks at Barruelo, (prov. Palencia), N.W. Spain. Colloquie Strat. Carbonifere Univ. Liege 5 5 : 487-5 5 1 .
Walter, J.C. 1953 . Paleontology of Rustler Formation, Culbertson County, Texas. J. Paleont. 27 : 679-702.
Wang, Y. 1956. New species of brachiopods 1 1 . Acta paleont. sin. 4 (3): 385-405 . (In Chinese).
Wanner, J. & Sieverts, H. 1935 . Zur kenntnis der Permischen Brachiopoden von Timor. Neues Jb. Miner. Geol. Paliiont. Bei/Bd 74B: 201 -28 1 .
Waterhouse, J.B. 1 963. The Permian faunal succession in New Zealand. J. geo/. Soc. A ust. 10 : 165- 1 76.

204 References
--- . 1 964. Permian brachiopods of New Zealand. Bull. Paleont. Wellington 35 : 1 -287.
--- . 1 966. Lower Carboniferous and Upper Permian brachiopods from Nepal. Jb. geol. Bundesant. Sbd. 1 2 : 5-95.
1 967a. Upper Permian (Tatarian) brachiopods from New Zealand J. N.Z. Geol. Geophys. 10 : 74- 1 1 8 .
--- . 1 967b. A new Permian fauna from New Caledonia, and its relationships to Gondwana and the Tethys. Simpos. Internal. Stratig. Paleont. Gondwana. Res. Assoc. Geol. A rgent. 1 967 : 249-272.
--- . 1 968. The classification and descriptions of Permian Spiriferida (Brachiopoda) from New Zealand. Palaeontographica 1 29A : 1 -94.
--- . 1 969a. Permian Strophalosiidae (Brachiopoda) from the Canadian Arctic Archipelago. J. Paleont. 43 : 28-40.
--- . 1 969b. World correlations of New Zealand Permian stages. N.Z. J. Geol. Geophys. 1 2 (4) : 7 1 3-737.
--- . 1 970a. Gondwanan occurrences of the Upper Paleozoic Brachiopod Stepanoviella. J. Paleont. 44 : 37-50.
--- . 1 970b. Permian Brachiopod Retimarginifera perforata n. gen. n.sp. from the Byro Group of Carnarvon Basin, Western Australia. J. Roy. Soc. W.A ust. 53: 1 20- 1 28.
--- . 1 970c. Correlation of marine Permian faunas for Gondwana. Proc. Pap. Second Gondwana Sympos. S. Ajr. : 3 8 1 -394.
--- . 1 97 1 . The brachiopod genus Tomiopsis Benediktova from the Permian of Canada. J. Paleont. 45 : 68-80.
--- . 1 972a. The evolution, correlation and paleogeographic significance of the Permian ammonoid family Cyclolobidae Zittel. Lethaia 5 : 230-250.
--- . 1 972b. A Permian overtoniid brachiopod in early Triassic sediments of Axel Heiberg Island, and its implication on the Permian-Triassic boundary. J. Can. Earth Sci. 9 : 486-499.
--- . 1 973a. Permian brachiopod correlations for south-east Asia. Bull, Geol. Soc. Malaysia 6 : 1 8 7-2 1 2.
--- . 1 973b. An Ophiceratid ammonoid from the New Zealand Permian and its implications for the Permian-Triassic boundary. Geol. Mag. 1 10 (4) : 305-329.
--- . 1 973c. Communal hierarchy and significance of environmental parameters for Brachiopods : the New Zealand Permian model. Contr. Roy. Ont. Mus. Life Sci. 92 : 1 -49.
--- . 1 973d. The Permian-Triassic boundary in New Zealand and its relationship to world climatic changes and extinction of Permian life. Mem. Can. Soc. Petrol Geol. 2 : 445-464.
--- . 1 974a. The significance of Permian faunal biomes in reconstructing Gondwana. An. A cad. brasil Crime. ( 1 972) (suplemento) 44: 365-3 8 1 .
--- . 1 974b. The Permian-Carboniferous boundary for Gondwana. An. Acad. brasil. Ctenc. ( 1 972) (suplemento) 44 : 383-39 1 .
--- . 1 975. New Permian and Triassic Brachiopod Taxa. Pap. Dep. Geol. Univ. Qd. 7 (2) : 1 -23.
Waterhouse, J .B. & Bonham-Carter, G. 1 972. Permian paleolatitudes judged from brachiopod diversities. Int. Geol. Congr. 24Can. Sect. 7 : 350-36 1 .
--- 1 975. Global distribution and character of Permian biomes based on brachiopod assemblages. J. Can. Earth Sci. 1 2 : 1085- 1 1 46.
Waterhouse, J .B. & Piyasin, S. 1 970. Mid-Permian brachiopods from Khao Phrik, Thailand. Palaeontographica 1 35A (3-6): 83- 1 97.
Waterhouse, J .B . & Vella, P.L. 1 965. Permian fossils from Parapara Peak, northwest Nelson. Trans. Roy. Soc. N.Z. geol. 3 (5) : 57-84.

References 205
Wegener, A. 1 929. Die Entstehung der Kontinente und Ozeane. Vieweg & Son. Brauschweig (4th Ed.)
Whitfield, R.P. 1 908. Notes and observations on Carboniferous fossils and semifossil shells brought home by members of the Peary expeditions of 1 905-06. Bull. Am. Mus. Nat. Hist. 24: 5 1 -58.
Wilde, G.L. 1968. Guadalupian biostratigraphic relationships and sedimentation in the Apache Mountain region, west Texas. In Silver, B.A. (Ed.) Guadalupian Facies, Apache Mountain area, west Texas Permian Basin. Sect. Soc. Econ. Paleont. Mineral. Sympos. Guidebook 68- 1 : 10-24.
--- . 197 1 . Phylogeny of Pseudofusulinella and its bearing on early Permian stratigraphy. Contrib. Smithson. Paleobiol. 3 : 363-380.
Williams, A. 1973. Distribution of brachiopod assemblages in relation to Ordovician palaeogeography. Spec. Pap. Palaeont. 1 2 : 24 1 -269.
Williams, T.E. 1963. Fusulinidae of the Hueco Group (Lower Permian), Hueco Moun-tains, Texas. Bull. Peabody Mus. nat. Hist. 1 8 .
Wood, B.L. 1956. The Geology of the Gore Subdivision. Bull. N.Z. geol. Surv. 53. Yabe, H. 1964. Lepidolina problem. Proc. Jap. Acad. 40 (3) : 2 1 4-2 1 9. Yakovlev, N.N. 1 9 1 2. Die Fauna der oberen Abteilung der Palaozoischen Ablagerungen
im Donetz-Bassin. J. Die Brachiopoden. Trudy geol. Kom. n.s. 70. --- . 193 1 . Geological investigations in Daralagese in Transcaucasia. Izv. glavn.
Geol. Razv. upr. 50: 5 1 5-52 1 . (In Russian). Yanagida, J. 1 958. Brachiopods from the Upper Permian Mizukoshi Formation, Central
Kyushu. Mem. · Fac. Sci. Kyushu. Univ. D. Geol. 14 (2) : 69-78. --- . 1964. Permian brachiopods from central Thailand. Colltributions to the
geology and palaeontology of Southeast Asia IX. Mem. Fac. Sci. Kyushu Univ. ser. D, geol. 1 5( 1 ) : 1 -22.
--- . 1 966. Early Permian brachiopods from North-central Thailand. Geol. Palaeont. southeast Asia 3 : 46-97.
--- . 1970. Permian brachiopods from Khao Phrik, near Rat Buri, Thailand. Geol. Palaeont. southeast Asia 8 : 69-96.
Yanagida, J. & Hirata, M. 1 969. Lower Permian brachiopods from Nakakubo, west central Shikoku, Japan. Trans. Proc. Palaeont. Soc. Japan, n.s 74 : 89- 1 1 1 . (Japanese summary).
Yochelson, E.L. 1 954. Some problems concerning the distribution of the late Paleozoic gastropod Omphalotrochus. Science 1 2 (3 1 10) : 233-234.
--- . 1968. Biostratigraphy of the Phosphoria, Park City, and Shedhorn formations. Prof Pap. U.S. geol. Surv. 3 1 3 D: 5 7 1 -660.
Yole, R.W. 1 963. An early Permian fauna from Vancouver Island, British Columbia. Bull. Can Petrol. Geol. (2) : 1 38- 149.
Zaleskysky, M.D. 1 930. Distribution of fossil flora, related to Gondwana on the outskirts of the western part of Eurasia. Com. AN USSR Ser. 7, Div. Phys. Math., Sci. 7 (4) : 1 -25. (In Russian).
Zavodowsky, V.M. 1 968. Superfamily Delthyriacea. New species of Permian Delthyriacea from North-east USSR. New species of polar plants and animals from the USSR 2 : 1 69- 1 73 . (In Russian).
--- . 1 970. In Atlas of Permian fauna and flora from north-east USSR. Ed. Kulikov, M.V., NW USSR Min. Geol. Magadin.
Zhang, Y. & Ching, Y.K. 1 96 1 . An Upper Permian brachiopod fauna from Jingxian, Anhui Province. Acta palaeont. sin. 9 : 40 1 -435. (In Chinese and Russian).
Zhao, T.G. 1 965. Upper Carboniferous brachiopods from the central part of the autonomous region of inner Mongolia. Acta palaeont. sin. 1 3 (3) : 420-454. (In Chinese and Russian).

206 References
Zolotova, V.P., Graifer, B.l., Federova, L.A. & Shiryeva, A.P. 1 966. Faunal characteristics of the Kungurian stage of the Permian of the Kama River region. Stratigraphy, lithology, facies and fauna of the Upper Proterozoic and Paleozoic of the Volga-Urals oil and gas beds. Geo/. Oil. Gas lnst. VNIGRI 5 1 : 247-253. (In Russian).

Geographic Index
Abadeh, 168 Aegean Sea, 134, 152 Afghanistan, 4, 20, 75, 77, 82, 92, 96, 97, 1 06,
107, 129, 131, 145: Table 38, p. 130; Fig. 4a, b, pp. 8, 9
Aktasty River, 78 Aktubin Urals, 96 Aktyubin region, 78 Alaska, 22, 23, 85, 87, 1 18, 135 Alaska Panhandle, 22, 98 Alberta, 72, 87, 1 18, 1 19, 135 Amarassi, 149, 153, 156, 166 Amdrupsland, 1 35 Andes, 77 Anhui, 109, 165 Arabia, 84 Araxes River, 151 Archanchel, 129 Arctic, 106, 109, 1 18, 1 1 9, 1 22, 124, 126, 127,
12� 131, 134, 137, 149, 181 , 183 Arctic Islands, Canada, 14, 36 Argentina, 45, 49, 77; Fig. 20, p. 74 Ar�ona, 72, 87, 98, 119 Armenia, 18, 19, 22, 23, 31 , 34, 36, 37, 82, 96,
97, 140, 149, 151 , 152, 157, 163, 165, 168, 183; Table 2a, p. xv; Table 45, p. 158
Arti region, 95 Asia, 126 Asia, central, 40, 90, 97, 1 06, 126, 157, 166,
179 Asia, south-east, 1, 16, 23, 64, 1 03, 109, 1 24,
131, 144, 146, 152, 157, 165, 181; Table 19, p. 65: Table 39, p. 133
Australia, 2, 11 , 18, 34, 39, 46, 49, 52, 55, 77, 87, 92, 97, 99, 103, 1 05, 124, 1 34, 1 37, 141, 145, 149, 153; Table 40, p. 1 38; Fig. 21 , p. 76
Australia, east, 16, 22, 23, 24, 29, 39, 45, 77, 95, 99, 1 03, 124, 127, 1 37, 156, 176, 179, 181, 183
Australia, Western, 22, 23, 77, 92, 99, 102, 116, 124, 126, 156; Table 29, p. 1 01
Austria, 2, 19, 23, 40, 44, 45, 48, 49, 52, 61 , 66, 69, 82, 135, 145, 161 , 168, 174, 183; Table 21, p. 70; Table 24, p. 83
Axel Heiberg Island, 31 , 1 1 8
Baikal (Zabaikal), 22, 23, 60, 80, 1 31; Table 13 , p. 47
Baktibari Range, 1 07 Balia Maaden, 66 Bannahai, 146 Baroghil Ailak, 84 Barrealas, 77 Barruelo de Santulla, 69 Barus, 174 Bashkiria, 42 Basin & Range Province, 22
Basleo, 29, 1 34, 149, 153 Bear Island, 22
·
Bjorne Peninsula, 87 Bj�rn�ya, 85 , 1 16: Table 25 , p. 86; Fig. 18, p.
68 Bolivia, 23, 75, 92; Fig. 20, p. 74 Bonaparte Gulf Basin, 22, 156 Borzin, 131 Borzow Bay, 1 29 Bowen Basin, 22, 95, 99, 126, 137, 156; Table
36, p. 125; Table 40, p. 133; Table 41 , p. 139; Fig. 23, p. 93
Brazil, 77; Fig. 20, p. 74 Bren Spur, 77, 92 British Columbia, 14, 72, 1 1 8, 124, 135 Bukit Tengku Lemba, 146 Bukk Mountains, 1 68 Bunsowland, 69, 85, 1 16 Burma, 23, 1 10, 1 24, 126, 146 Byans, 143
Calcutta, 61 California, 72, 118; Table 27, p. 91 Camp Staghar glacier, 145 Cambodia, 22, 1 10, 131 , 132, 144, 146; Table
19, p. 65 Cammon, 146 Canada, 14, 22, 46, 48, 49, 57, 61 , 69, 72, 75,
79, 85, 90, 97, 99, 109, 1 16, 124, 1 26, 1 79 Canada, Arctic, 20, 22, 23, 87, 98, 105, 1 1 3,
1 16, 1 1 8, 1 19, 127, 1 29, 135, 172, J74; Table 2b, p. xv; Table 22, p. 73; Fig. 2, p. 2 ; Fig. 27, p. 1 1 7; Fig. 38, p. 173
Canning Basin, 22 Cape Kalouzin, 106, 1 09 Cape Loushkin, 79 Cape Stosch, 1 1 , 1 16 Carnarvon Basin, 22, 77, 92, 95 , 99, 1 24; Table
29, p. 101", Table 40, p. 1 1 3 Carnian Alps, 19, 44, 49, 52, 6 1 , 82, 168; Table
21, p. 70; Table 24, p. 83 Caucasus, Greater (North), 22, 37, 129, 131 ,
135, 151 , 157 Caucasus, Lesser, 157 Chekiang, 109, 131 , 169 Chile, 75 China, 18, 22, 23, 3 1 , 34, 36, 37, 39, 64, 66,
84, 85, 97, 1 06, 109, 1 16, 122, 124, 1 3 1, 146, 149, 161, 163, 165, 168, 169, 183; Table 2b, p . xv; Table 18, p. 62; Table 19, p. 65 ; Fig. 17, p. 63
Chiron, 80, 106 Chitichun No. 1, 144 Chitina Valley, 87 Chitral, 82, 84 Chironskogo, Lake, 13 1 Clinton, 166 Coahuilla, 22, 23, 36, 37, 99, 152

208 Geographic Index
Cooper River Valley, 87 Crimea Peninsula, 22, 1 29, 1 3 1, 1 5 1 Croatia, 69 Croisilles, 7 7 Crowsnest, 87
Darma XI Peak, 143 Darvas, 79, 84, 90, 97; Table. 17, p. 60 Delaware Basin, 22, 26, 1 66; Table 26, p. 88 Devon i�and, 98, 1 1 6 DUUn�, 99, 1 37, 1 5 3, 1 66 Djadjerud Valley, 1 07 Djambi Residency, 1 22 Djebel Tebaga, 1 5 2 Djulfa, 1 8 Djulfa Gorge, 1 5 7 , 168; Table 4 5 , p. 1 5 8 Dolpo, 1 4 3 ; Fig. 30, p. 1 4 2 Dombas River, 53 Don River, 55 Doneu Basin, 22, 55 Dorasham Siding, 1 68
East Germany, 2 Eglinton Valley, 1 66 El Antimonio, 22, 1 22; Fig. 28, p. 1 2 1 Elburz Mountains, 22, 152, 161 Table 3 1 , p .
1 08 Ellesmere I�and, 72, 87, 1 1 8 England, 1 1 , 1 1 0, 1 1 3; Table 33, p . 1 14; Fig.
25, p. 1 1 2 Europe, 1 6, 23, 3 1 , 44, 6 1 , 82, 1 1 0, 1 1 6, 1 68,
169; Fig. 25 , p. 1 1 2 Everest, Mt., 143, 145
Fergana, 22, 49, 61, 145; Table 1 7, p. 60; Table 23, p. 81
Fitzroy Basin, 92, 1 26, 1 56; Table 29, p. 1 0 1 Foldvik Creek, 3 1 Freiburg, 26
Gasherbrun, 6 1 , 64 Germany, 1 1 0; Table 33, p. 1 14; Fig. 25, p. 1 1 2 Gilgal Camp, 6 1 , 64 Glass Mountains, 14, 22, 26, 29, 37, 46, 49, 5 2,
72, 75, 78, 80, 87, 90, 1 1 9, 1 3 1 , 1 35, 1 79; Table 26 , p. 88; Table 27, p. 9 1 ; Fig. 22, p. 89; Fig. 29, p. 1 36
Gondwana, 2, 45, 55, 75, 77, 82, 84, 98, 1 1 3, 1 22, 1 76, 1 79
Great Bear Cape, 1 1 8 Greenland, 2, 26, 3 1 , 1 1 3, 1 16, 1 35, 1 72, 1 74;
Fig. 26, p. 1 1 5 Grinnell Peninsual, 1 1 6 Guadalupe Mountains, 1 37, 1 5 2, 1 5 3; Table 26,
p. 88Guadalupe Peak, 152 Guatemala, 1 1 8, 122 Guryul Gorge (Ravine), 143, 174, 1 75; Fig. 36,
p . 171 Gympie Basin, 22, 156, 1 66; Table 36, p. 1 25:
Table 40, p. 138; Table 41, p. 139
Haraulakh, 55, 57, 79; Table 1 3, p. 47; Table 16, p. 59
Hato Dame, 1 24 Haushi, 84 Hirnalayas, 2, 16, 22, 75, 1 34, 141 , 143, 144,
145, 146, 148, 1 5 2, 1 53, 1 5 7, 1 74, 1 8 1 , 1 83 Hornsund, 69, 1 35 ; Table 25, p. 86 Htam Sang, 146 Hueco Mountains, 80, 82 Hungary, 168 Hunter Valley, 77, 95, 1 26, 137 Hunza Valley, 145 Hydra, 1 34
Idaho, 26, 1 09, 1 1 9, 1 35 ;-Table 34, p. 1 20 India, 75, 77, 84, 92, 99 Indian Ocean, 141 Indochina, 1 74; Table 42, p. 147 Indonesia, 36 Indus River, 6 1 , 144 uan, 1 8, 22, 23, 36, 37, 82, 84, 92, 1 06, 1 07,
1 1 8, 1 32, 1 34, 149, 1 5 1 , 152, 1 57, 1 6 1 , 1 68, 1 75, 1 83 ; Table 3 1 , p. 108
uwin (Perth) Basin, 92 Isfjorden, 85, 1 35 Ishibeisk Urals, 95 Ishimbaeva, 79 Italy, 19, 23, 82, 168, 1 74, 1 83; Table 21 , p. 70
Japan, 22, 23, 39, 64, 85, 97, 109, 1 1 0, 1 3 1 . . 1 3 2, 145, 146, 149, 1 5 1 , 1 5 2, 1 65 , 168, 1 74 ; Table 20, p. 67; Table 32, p. 1 1 1 ; Table 48, p . 1 70
Jenka Pass, 1 65 Jinxian, 165
Kabul, 77, 84, 97, 107, 1 3 1 , 145 Kama River region, 1 02 Kamchatka, 1 1 8 Kamiyase, 148 Kamloops, 1 1 8, 1 24 Kanin Peninsual, 1 05 Kansas, 45, 75 90 166; Table 27, p. 9 1 Karakorum Range, 2 2 , 6 1 , 8 2 , 145 Kashmir, 1 9, 22, 23, 77, 84, 92, 1 4 1 , 1 43, 144,
1 74; Fig. 36, p. 1 7 1 Kazakhstan, 49, 60, 69 Kehsi Mansam, 1 1 0 Kesenmura, 148 Kham Kheut, 64 Khao Phrik, 1 1 0 Khojagar Dara Dam, 107 Kiangsi, 64, 1 09, 1 3 1 Kinta Valley, 64 Kirillow, 103 Kitakami Mountains, 1 32, 148; Table 20, p. 67 Kiunglung, 143 Kiya River, 53 Koktan Ridge, 61 Kolwa, 96 Kolyma River, 22, 23, 57, 80, 96, 1 05 , 1 29,
149, 152; Table 13, p . 47; Table 15, p. 58 Korea, 23, 64 Kuh-e-Ali Bashi, 1 5 1 , 168 Kuiechow, 169 Kukurtuktal, 61

Kulor River, 1 29 Kumwaon, 141 Kur Ufrimsk, 149; Table 43, p. 150 Kwangri, 64, 97, 169 Kwanto Province, 97: Table 20, p. 67 Kweichow, 64, 1 69 Kyushu, 66; Table 20, p. 67
I..aos, 22, 64; Table 1 9, p. 65 Lachi, 143 I..ang-nac, 1 3 2 l.ar Valley, 1 07 Las Delicias, 36, 99, 1 1 9, 1 22, 1 37, 1 52, 1 66;
Table 35, p. 1 23 Lena River, 23, 46, 55, 79, 1 29 Letti, 1 24 Lissar Valley, 143 Linlithi, 143 Loushkin, Cape, 79 Luang Prabang, 146, 1 57
Madagascar, 92, 14 1 , 1 53, 1 6 1 , 169 Madre de Dios, 75 Maizuru belt, 1 65 Maizuru Zone, 148 Maiya area, 148 Malascachas, 1 1 9, 13 7 Malakhabad, 1 07 Malaysia, 64, 146, 165 Manchuria, 14, 64 Manendragargh, 77 Marathon Basin, 45 Mataura Island, 1 66 Mediterranean, 22 Mediterranean Sea, 1 5 2 Mexico, 22, 23, 36, 3 7 , 97, 9 8 , 1 1 9, 1 22, 1 37,
149, 145, 1 5 � 153, 1 56, 157, 1 66, 1 83: Table 3 5 , p . 1 2 3 ; Fig. 28 p . 1 21
Mezen River, 1 29 Mitsu, 148 Mongolia, 14, 22, 23, 6 1 , 1 1 0, 1 32, 149, 163,
183 ; Table 4 7, p . 1 64 Miseryfjellet, 1 1 6 Mong Pawn, 1 1 0, 146 Montana, 1 1 9, 1 35� Table 24, p. 1 20 Moscow Basin, 45 Monte Aueririg, 66 Mount Bamyan, 97 Mount Jomlo Lungma (Everest), 143, 145 Mudgee, 1 72 Muong The, 64
Nagmarg, 92 Namun, 1 1 0 Nathorsts Fiord, 1 1 6 Nebraska, 75, 1 66 Nelson, 78, 1 27, 1 66, 1 69; Table 37, p. 1 28 Nepal, 1 9, 22, 23, 141 , 143, 144, 149, 1 5 1 ,
1 57, 1 6 1 , 1 74, 1 83; Fig. 30, p . 1 4 2 Nevada, 26 New Caledonia, 145, 153, 1 57, 166 New Me�co, 5, 1 8, 3 1 , 34, 36, 49, 99, 1 24,
1 56, 1 6 1 , 1 66 New South Wales, 22, 23, 45, 95, 1 26, 1 37,
1 72 ; Table 40, p. 1 3 8
Geographic Index 209
New Zealand, 4, 14, 16, 20, 22, 23, 3 9, 77, 92, 95, 99, 1 02, 1 03, 1 05 , 1 06, 1 1 8, 1 26, 1 27, 1 35 , 1 3 7 , 145, 149, 1 5 1 , 1 5 2, 1 53, 1 56, 157, 163, 1 66: 169, 1 72, 1 74, 1 8 1 , 1 83;Table 2b, p xv; Table 6, p . 17; Table 37, p. 1 28 , Table 40, p . 1 3 8 ; Fig. 3, p . 3 ;Fig. 24, p . 94; Fig. 3 1 , p . 1 55; Fig. 37 , p. 173
North Auckland, 1 56 North America, 14, 1 8, 22, 23, 36, 40, 1 02,
1 24, 1 29, 1 35 , 156, 1 83 North Island, New Zealand, 22 Novaya Zemlya, 22, 23, 79, 1 05, 1 29 Novgorod, 1 05
Ob River, 55 Ogilvie Mts., 24, 46 Oklahoma, 75, 166 Omolon Plateau, 22, 57, 80, 1 29, 1 49, 1 52;
Table 1 5 , p. 5 8 Onilahy, 9 2 Oregon, 14, 36, 57, 7 2 Orulgania, 46; Table 1 3 , p. 47; Table 1 6 , p. 59
Pahlgam-Aru, 1 74 Pai Hoi, 22, 55, 79, 96, 99, 1 02, 1 05 , 1 1 8,
1 26; Table 14, p. 56 Pahlstan, 16, 22, L9, 36, 75, 77, t4, 92, 99,
107, 1 09, 1 10, P4, 1 34, 14'J, 148, 1 5 2, 1 6 1 , 1 66; Table 2,\ p. l Or; Fig. 3 2, p. 1 60; Fig. 33, p . 1 6:· .. '
PalenCia Province, 69 ·.
Pamirs, 22, 23, 40, 44, 49, 80, 82, 84, 85, 96, 97, 1 06 , 132, 149, 1 5 1 , 163 , 1 83 ; Table 23, p. 81
Parana Basin, 77 Pasha Asha, 61 Perlis, 146 Perm, 34 Perth Basin, 22, 92, 175; Table 29, p. 101 Peru, 23, 75, 92; Fig. 20, p . 74 Petchabun, 146 Petchora, 22, 23, 55, 80, 96, 98; Table 14, p.
56 Phnom Miai, 1 10 Phnon-Ta-Maio, 1 3 2 Phnom Ta Maol, 1 1 0 Phnom-Tu-Kreem, 1 1 0, 1 3 2 Phnom Tup, 1 34 Pinega River, 1 29 Pong Oua, 157 Port Keats, 150 Poshan City, 64 Primoyre, South, 149
Quang Tri, 146 Queensland, 22, 23, 24, 25, 77, 95, 1 02, 1 24,
1 26, 1 3 7 , 1 56, 157, 166, 1 81 , 1 83; Table 2b, p. xv Table 36, p. 1 2 5 ; Table 40, p. 1 3 8 ;Table 4 1 p . 139 ; Fig. 2 3 p . 93
Rajasthan, western, 92 Rek Basin, 1 3 1 Richardson Mountains, 1 1 8, 1 35 Rocky Mountains, 14, 22, 23, 7 2, 1 1 8

2 1 0 Geographic Index
Rusaa, 14, 1 8, 22, 34, 36, 44, S3, 5S, 92, 99, 103, 105, 1 07, 1 1 3, 1 1 6, 1 24, 1 26, 1 27, 1 35, 1 76, 1 79; Fig. 16, p. 54
Russian Harbour, 1 29 Russian Platform, 1 8, 22, 34, 39, 40, 44, 46,
53, 102, 1 27, 1 29; Table 2a, p. xv; Table 12, p. 41
Salt Range, 1, 1 9, 22, 23, 29;36, 77, 84, 92, 99, 107, 1 1 0, 1 3 1 , 140, 14 1 , 143, 144, 145, 146, 148, 149, 1 5 1 , 1 53, 1 57, 163, 175, 1 83; Table 2a, p. xv; Table 12, p. 4 1 ; Table 19, p . 65 Table 28, p. 100; Fig. 32, p. 1 60; Fig. 33, p. 162
Saxony, 2 Saudi Arabia, 84, 92, 99 Set-Davan, 46, 55, 51; Table 1 3, p. 47; Table
16, p. 59 Shaksgam Valley, 61, 145 Shalshal Cliff, 1 74 Shan States, 1 1 0 Shansi, 109 Shih-pu-sui, 64 Siberia, 1 8, 34, 39, 46, 48, 49, 57, 60, 79, 85,
87, 96, 105, 1 1 3, 1 16, 1 24, 1 27, 1 29, 1 35, 141, 149, 1 8 1 , 1 83; Table 1 3 , p. 47; Table 15, p. 58; Fig. 16, p. 54
Sicily, 26, 1 24, 1 34, 152 Sierra Diablo, 90, 1 37; Table 26, p. 88 Sierra de la Ventana, 77 Sikhote Alim , 1 05, 106; Table 43, p. 1 5 0 Sikkim, 77, 92, 14 3 Sintas River, 53 Sisophon, 131 , 1 32, 146; Table 39, p. 133 South Africa, 77, 179 South America, 1 79; Fig. 20, p. 74 Sosio, 1 3 1 , 1 34, 144, 145, 1 5 1 , 1 5 2 South Island, New Zealand, 22 Southland, 1 66, 169, 1 74; Fig. 31, p. 1 55 Soviet Union, 1 , 14, 1 8, 36, 49, 52, 1 75 South Pole, 102, 179 Spain, 45, 48, 57, 66, 69 Spiti, 143, 175 Spitsbergep, 22, 23, 3 1 , 49, 69, 85, 87, 102,
103, 1 1 3, 1 16, 1 35, 1 74; Table 25, p. 86; Fig. 1 8, p. 68
Springsure Shelf, 77, 95 Stanhope, 77 Subansiri, 92 Sumatra, 1 22 Suren River, 4 2 Svartevaeg, 31, 1 1 8; Table 9, p. 30 Sverdrup Basin, 98 Sydney Basin, 22, 77, 95, 1 26; Table 40, p. 138;
Table 4 1 , p. 139 Szechuan, 109, 160
Tai-li-shao, 1 3 1 Taimyr Peninusla, 22, 23, 46, 55, 79, l OS, 1 29,
149, 152; Table 1 3, p. 47; Table 15, p. 58; Table 16, p. 59
Takaka, 1 27 Takitimu Mountains, 1 02, 1 27, 1 37, 169, 1 74;
Table 37, p. 1 28; Fig. 37, p. 1 73
Tasmania, 22, 23, 77, 95, 99, 1 02, 1 24, 1 26, 137 , 169 . 1 72, 1 79 ; Table 40, p. 138
Texas, S, 14, 1 8, 1 9, 23, 26, 3 1 , 34, 36, 37, 39, 45, 72, 75, 78, 80, 82, 90, 98, 99, 102, 109, 1 16, 1 22, 1 3 1 , 135, 140, 146, 1 56, 166 , 1 79 Table 2b, p. xv; Table 26, p. 88; Table 27, p. 91 Fig. 22, p. 89; Fig. 29, p. 136
Tezak, 107 Tianwo District, 97 Tei Wei, 1 3 7 Thailand, 22, 64, 1 1 0, 1 22, 146, 1 65; Table 19,
p. 65 Thakkola, 144 Thurn Nam Maholan, 64 Thuringia, 2 Tianwo District, 97 Tibet, 6 1 , 64, 141 , 144, 145 Tien Shan, 6 1 Timan, 14, 22, 55, 1 29, 1 35 Timor, 1 9, 22, 23, 36, 37, 1 22, 1 24, l26, 140,
14 1 , 143, 144, 149, 1 53, 166; Table 1 9, p. 65; Table 44, p. 1 54
Tompo Basin, 96 Tonkin, 64 Trlln-ninh, 64 Tresdorfer hohe, 69 Tsang Po, 61 Taang-gyi, 146 Tunguskian Basin, 2 Turriaa, t44, 145, 1 5 2 Turkey, 66 Tyrol, 168
Uf'unian amphitheatre, 96 lilladulla, 1 26 Umaria, 84, 92 United States, 16, 1 8, 22, 23, 26, 34, 39, 52,
6 1 , 69, 87, 97, 1 1 8, 143 , 1 52, 166, 1 74; Table 2b p. xv; Table 26, p. 88; Table 27, p. 9 1 ; Table 34, p. 1 20; Fig. 29, p. 1 36
Urals, 14, 16, 18, 20, 22, 23, 26, 34, 39, 40, 44, 45, 46, 48, 49, 53, 55, 6 1 , 78, 79, 80, 84, 90, 95, 98, 99, 1 02, 1 03, 1 05 , 1 24, 1 27, 1 76, 1 79 ; Table 2a, p. XV
Ussuriland, 22, 23, 1 06 Utah, 26, 1 35
Vancouver Island, 87 Verchoyan, 22, 46, 48, 5 2, 57, 79, 80, 87, 96,
98, 1 05 , 1 1 9, 1 29, 149, 1 79, 1 81 , Table 1 3, p. 47; Table 15, p. 58; Table 16, p. 59; Fig. 22, p. 89
Vietnam, 1 1 , 146, 165 Vologrod, 1 29
Wadi Lusaba, 84 Washington State, 1 1 8Wyoming, 26, 72, 1 1 9, 1 35; Table 34, p. 1 20 Wardak, 84; Table 38, p. 1 30
Yal Nal, 92 Yangtze Valley, Lower, 109, 1 65 Yarrol Basin, 22, 77, 95; Table 36, p. 1 25;

Table 41, p. 139 Yugoda�, 23, 69, 82, 161, 168, 183 Yukon Territory, 4, 14, 22, 23, 24, 46, 57, 69,
72, 75, 79, 80, 85, 87, 98, 99, 1 1 8, 135, 17� Table 22, p. 73; Fig. 19, p. 71
Yunnan, 131
Zabaikal (see Baikal), 57, 106; Table 13, p. 47
Geographic Index 2 1 1

Stratigraphic and Biostratigraphic Index
The standard substages and stages appear on most tables and are omitted from the index. Variations in nomenclatural hierarchy, used by different authors, are generally not recorded.
Abadehian Stage, 29, 37, 140, 157, 161, 183 Admiral Formation, 92; Table 27, p . 91 Admire Group, 75� Table 27, p. 91 Afghanella Zone, Table 1 8, p . 62 Agglomeratic Slate, 84, 92 AG4 Formation, 156; Table 6, p. 1 7 AG5 Sandstone, 166 Aigyo Formation, Table 32, p. 1 1 1 Airkunung beds, 1 22 Akasaka Limestone, Table 32, p. 1 1 1 Akasaka Stage, 1 32 Ahiyoshi Limestone Group, Table 32, p. 1 1 1 Aktastinian Substage, 78, 80, 82, 84, 85, 87,
90, 92, 95, 96, 98, 1 76, A1acran Mountain Formation, 90; Table 26, p.
88 Alataconcha vampyra beds, Table 3 8, p. 130 Aldebaran Formation, Table 36, p. 1 25; Table
40, p. 1 3 8; Table 4 1 , p. 139 Ali Bashi Formation, 168, 1 7 5 Allandale fauna; Table 4 1 , p. 1 39 Allandale Formation, 77; Table 40, p. 1 3 8;
Table 4 1 , p. 1 39; Fig. 1 2, p. 50; Fig. 14, p. 51� Fig. 15, p. 51
Aller Series, Table 33, p. 1 14 Amarassi fauna; 5, 3 1 , 36, 37, 166; Table 19,
p. 65; Table 44, p. 1 54 Arilb Fo.miation, 99, 1 24, 1 3 1 , 145; Table 2a,
p. xv; Table 9, p. 30; Table 19, p. 65 ; Table 28, p. 1 00
Amilobe beds, 161 Ammonoid stages, 22 Anchignathodus fauna, Table 7, p . 27 A. parvus Zone, 29 Anidanthus zone, 1 22; Fig. 28, p. 121 Annear Sandstone, 1 66 Antiquatonia zone, 97, 99; Table 2b, p. xv;
Table 22, p. 73 Aperispirifer zone, Table 37, p . 1 28 Aperispirifer nelsonensis Zone, Table 2b, p. xv;
Table 3 7, p. 1 28 Appel Ranch Member, 26, 135, 1 3 7 ; Table 2b,
p. xv Table 26, p. 88; Fig. 29, p. 1 16
Arkasian (Araxian) Stage, 5, 37, 140, 1 6 1 , 1 83 Araucaria Suite, 55
Araxi/evis zone, Table 2a, p. xv Araxoceras Ia tum Zone, 29 Argidjin Suite, Table 13, p. 47 Armik Horizon, 97; Table 2a, p. xv Artian,JB Artinskian Stage, "Series", 14, 1 8, 64, 78, 79,
85 , 87, 95 , 96, 98, 102, 105, 1 07, 1 1 0, 1 1 6, 145� Table 10, p. 33
Asakaido Formation, Table 20, p. 67 Asni Horizon, Table 2a, p. 4
Aso Group, 97 Asselian State, 7, 1 1 , 14, 26, 36, 40, 42, 44, 45, 48, 49, 52, 53, 55, 57, 60, 6 1 , 64, 66, 69, 72, 77, 78, 79, 84, 85, 90, 1 76, 1 79
Assistance Formation, 87, 1 09, 1 1 6, 1 24, 1 29; Fig. 27, p. 1 1 7
Atetsu Limestone Group, Table 32, p. 1 11 Atomodesma zone, Table 2b, p. xv; Table 37, p.
1 28 A. trechmanni Zone, 156 A ttenuate/fa (Ea) zone, 87; Table 22, p. 73;
Figs. 1 2- 1 5 , pp. 50, 5 1 A ttenuatella Tomiopsis (Eta) zone, 72; Figs.
1 2-15, pp. 50, 5 1 Audhild Formation, Fig. 27, p . 1 1 7 Auernig Group, 44 , 66, 69; Table 24, p . 83 Autunian, Fig. 1 1 , p. 43 Ayachyargin beds, subsuite, 105; Table 14, p.
56
Babstov Suite, 149 ; Table 43, p. 150 Back Creek Group, Fig. 23, p. 93 Baigendzinian Stage, 26, 44, 75, 78, 82, 85, 87,
95, 96, 97, 98, 99, 102, 103, 1 05, 1 06, 1 1 9, 1 22, 1 24, 1 26, 1 37, 145, 1 76, 1 79, 1 81
Baikur Horizon, Suite, 1 29, 1 3 1 , 149; Table 15, p. 58; Table 16, p. 59
Baisalian Substage, 27, 29, 36, 140, 157, 161, 163, 165, 166, 169, 183
Baker (= Nalbia) Formation, 1 24, 1 26; Table 29, p. 1 0 1
Balgo Member, Table 2 9 , p. 1 0 1 Barain "Series", 1 29 Barain fauna, 96 Baralaba Coal Measures, Table 2b, p. xv; Table
36, p. 1 25 Barents Series, 79, 1 29 Barfield Formation, Table 36, p. 1 25; Table 40,
p. 1 38; Table 4 1 , p. 1 39 Bashkirian Stage, 46, 176 Basleo fauna, 29; Table 19, p. 65; Table 44, p.
154 Bazardarin Suite, 82; Table 23, p. 8 1 Beckett Member, 92; Table 4 0 , p. 1 38 Belcher Channel Formation, 98; Table 22, p.
73; Fig. 27, p. 1 1 7 Belkov Suite, 96; Table 14, p . 56 Bell Canyon Formation, 1 5 2 Belle Plains Formation, 92; Table 27, p . 9 1 Bellerophon Limestone (Kalk), 1 9, 1 6 1 , 168,
1 69; Table 24, p. 83 Belokatai Suite, 95 Berein fauna, 1 06 Berriedale Limestone, 99, 102; Table 40, p.
1 3 8 Big Blue "Series", 75

Bilikat Suite, Table 16, p. 59 Billingham Main Anhydrite, Table 33, p. 1 14 Bini Darzah Horizon, Table 38, p. 1 30 Binthalya Formation, Table 29, p. 101 Birrang Suite, 79; Table 15, p. 58, Table 16, p.
59 Bitauni beds, Table 19, p. 65 Bitauni fauna, 1 22; Table 44, p. 154 Bjorne Formation, Fig. 27, p. 1 1 7 Black Crag Member, Table 25, p. 86 Black Alley Shale, Table 36, p. 1 25 ; Table 40,
p. 138; Table 41, p. 139 Blackwater Group, Fig. 23, p. 93 Blain Formation, 99 Blenheim Subgroup, 1 37� Table 36, p. 1 25 Blind Fiord Formation, Fig. 23, p. 93; Fig. 27,
p. 1 1 7 Bokan Series, Table 3 8 , p . 130 Bone Spring Formation, 26, 90; Table 26, p.
88 Bonete Formation, 77 boreale Zone, Fig. 38, p. 1 73 Boulby Halite, Table 33, p. 1 14 Brachiopod Chert, 1 16, 135; Table 25, p. 86 Brahmanian Stage, 1 72, 1 83 Branosera Formation, 69 Brucebyen beds, 69; Table 25, p. 86 Brushy Canyon Member, Table 26, p. 88 Buffel Formation, 95; Table 36, p. 1 25; Table
41, p. 139 Bulgonanna Volcanics, Fig. 23, p . . 93 Bulgadoo Formation, 99; Table 29, p. 1 01 Bundella Mudstone, Table 40, p. 1 3 8 Burgali Suite, 57; Table 1 3, p. 4 7 ; Table 1 5 , p.
58
Stratigraphic and Biostratigraphic Index 2 1 3
Carynginia Formation , Table 29, p . 1 01 Casper Formation, 7 2 Cathedral Mountain Formation, 3 6 , 8 7 , 98,
1 19, 1 22 ; Table 2b, p . xv; Table 26, p. 88; Fig. 29, p. 1 36
Catherine Sandstone, Table 40, p. 1 38; Table 4 1 , p. 1 39
Cattle Creek Formation, 95; Table 40, p. 1 38 Cattle Creek shale, 95 Ceratite Beds, Table 28, p. 1 00 Cereduchin Suite, 149 Cerro Alto Formation, 90; Table 26, p. 88 Cessnock Sandstone, 1 26; Table 40, p. 1 3 8 Chambin Suite, 1 29; Table 1 5 , p . 5 8; Table
16, p. 59 Chandalaz Suite, 149 Changhsingian Stage, 36, 37, 145, 1 6 1 , 169;
Table 9, p. 30 Changsing (= Changhsing) Limeston�. 146, 168,
169; Table 2b, p. xv; Table 18, p. 62; Table 19, p. 65
Chapsai Suite, Table 1 7 , p. 60 Chase Group, 75; Table 27, p. 91 Chelamchin Suite, 80; Table 17, p. 60 Chenia zone, Table 1 8, p. 62 Chenoyar Suite, 149; Table 15, p. 58; Table 16,
p. 59 Cherry Canyon Formation, Member, 1 37, 1 5 2;
Table 2b, p. xv Chevyu Horizon, 1 29 Clrhidru Formation, 37, 1 07, 141 , 144, 146,
1 6 1 , 1 75 ; Table 2a, p. xv; Table 9, p. 30; Table 19, p. 65; Table 28, p. 1 00
Chhidruan Stage, 5, 1 1 , 16, 29, 3 1 , 36, 140,
Burnett Formation, 77; Table 2b, p. xv; Table 36, 145, 148, 149, 1 5 1 , 152, 153, 1 56, 157, 163, 165, 1 8 1
p . 1 25; Table 40, p . 138; Table 4 1 , p . 1 3 9 Burtsev Suite, 7 8, 9 5 , 96 Borup Fiord Formation, Fig. 27, p. 1 1 7 Buttle Lake Formation, 87 Buxtonia zone, 46 Byro Group, 99, 102, 1 24; Table 29, p. 101
Cache Creek Group, 1 1 8 Cadellfjellet Member, 6 9 ; Table 25, p. 86 Callytharra Formation, 92, 95; Table 29, p.
1 01; Table 40, p. 138 Camboon Andesite, Table 36, p. 1 25; Table 41 ,
p. 1 39 Canar Suite, Table 43, p. 150 Cancel/ina zone, subzone, 109, 1 1 0, 1 22, 1 31;
Table 1 8, p. 62; Table 23, p. 81 Cancrinella Limestone, 87; Table 25, p. 86 Cancrinella zone, Fig. 28, p. 1 21 Cancrinelloides zone, Table 2b, p. xv Canyon Fiord Formation, Fig. 27, p. 1 1 7 Capertree Group, 172 Capitan Limestone, Table 2b, p. xv Capitanian Stage, 5 , 26, 29, 3 1 , 36, 39, 1 13 ,
140, 145, 152, 153, 156, 166 Carboniferous, 1 8, 26, 40, 42, 45, 53, 55, 57,
66, 77, 79, 1 06, 1 1 3, 144, 145, 1 76, 1 7 9 , late, 20, 40, 42, 44, 45, 46, 48, 49, 52, 55 , 57, 61, 66, 75
Carmila Beds, Table 4 1 , p. 1 39 Carrandibby Shale, 77; Table 29, p. 1 01
Chihsia Limestone, 84, 85, 97, 1 09 ; Table 2b, p. xv; Table 9, p. 30; Table 1 8 , p. 62; Table 19, p. 65
Chihsia "Stage", 1 09 China Tank Member, 1 1 8, 122, 137 ; Table 2b,
p. xv; Table 26, p. 88; Fig. 29, p. 136 Chini Thu Formation, 143, 144 Chiron Suite, 79, 80 Chochal Limestone, 1 22 Choutang Limestone, 146, 165 Chusenella zone, Table 1 8, p. 62 Cisco beds, 45 Clifton Subgroup, Table 40, p. 138 Clyde Formation, 90, 92; Table 27, p. 91 Codonofusiella zone, 165; Table 1 8, p. 62; Table
23, p. 81 Codonofusiella-Reichelina fauna, Fig. 34, p. 167 Colaniella zone, Table 23, p. 81 Comelicania zone, Table 2a, p. xv Comelicania-Phisonites zone, 161 , 1 6 8 commune (Ophiceras) Zone, Fig. 3 8 , p . 173 Composita zone, 1 22, Fig. 28, p. 1 21 concavum (Otoceras) Zone, Fig. 38, p. 1 73 Concha Formation, 1 1 9 Concretionary Limestone, Table 33, p . 1 14 Conjola Formation, 77; Table 4 1 , p. 1 39 Connors Volcanics, Fig. 23, p. 93 Conularia beds, 92; Table 2a, p. xv; Table 19,
p. 65; Table 28, p. 1 00 Coolkilya Greywacke, 1 24; Table 29, p. 1 01 Copacabana Group, 92

2 1 4 Stratigraphic and Biostratigraphic Index
Cora Limestone, 85; Table 25, p. 86 Council Grove Group, 75; Table 27, p. 91 Coyotte Butte Formation, 72 Coyrie Formation, 99; Table 29, p. 101 Croisilles Volcanic Complex, 77, Table 37,
p. 1 28 Cundlego Formation, 99; Table 29, p. 1 01 Cyclolobuszone, 140; Table 1 1 , p. 35 Cygnet Coal Measures, Table 40, p. 1 38
D fauna (Canada), 48, 5 2 Dabool Sandstone, 1 26, 1 27 Dalwood Group, 45 Darlington Limestone, Table 40, p. 1 38 Darvasian Stage, 80, 1 07; Table 23, p. 81 Dastar Horizon, 44, 61 Davalin Suite, 82; Table 2a, p. xv Decie Ranch Member, 72, 80, 90; Table 26, p.
88; Table 27, p. 91 Degerbols Formation, 1 1 8, 1 35; Fig. 27, p. 1 1 7 Delenjin Suite, 1 29; Table 15 , p. 5 8; Table 16,
p. 59 Dibin Suite, 96, 1 05; Table 15, p. 58; Table 16,
p. 59 Dictyoclostus zone, 1 22; Fig. 28, p. 1 21 Dienerian Stage, substage, 172, 174, 1 83; Fig.
38, p. 173 Dinwoody Formation, 174 Djambi Tuffs, 1 22 _ _ Djeltin Horizon, 97; Table 1 5 , p. 58 Djigdalin Horizon, 97; Table 15, p. 58 Djulfian (= Dzhulfian) Stage, ("Series"), 3 1 ,
140, 144, 146, 149, 153, 157, 161, 163 ; 165, 166, 169, 1 75, 1 8 1 , 183; Table 10, p. 33
Djuptagin Suite, 55; Table 13, p. 47 Doniyarchi Series, Table 38, p. 130 Dorashamian Stage, 29, 37, 5 1 , 140, 145, 1 65,
166, 168, 172, 175, 1 83 Dorud fauna, 1 07, Table 3 1 , p. 108 Dugout Mountain Member, Fig. 29, p. 1 36 Dulgalach Suite, Table 15 , p. 58; Table 16,
p. 59; Table 26, p. 88 Dun Mountain Ultramafics, Table 37, p. 1 28 Durvilleoceras woodmani Zone, Table 2b, p. xv
Table 6, p. 1 7; Table 3 7, p. 1 28 Dwyka Tillite, 77
E fauna zones (Canada) Ea, Eta, Eka, 46, 49; Figs. 1 2-15, pp. 50, 5 1
Earp Formation, 72, 87 Echi Suite, 57, 80, 87; Table 15 , p. 5 8; Table
16, p. 59 Echinalosia zone, Table 2b, p. xv E. kalikotei Zone, 143, 144•, Fig. 30, p. 142 E. maxwelli Zone, Table 37, p. 128 E. ovalis Zone, 137, 140; Table 2b, p. xv; Table
6, p. 1 7; Table 37, p. 1 28 E. prideri Zone, Table 37, p. 1 28 Efremov Suite, 1 05 Ekachan Suite, 46, 57; Table 13, p. 47; Table
15 , p. 58; Table 16, p. 59 Elderslie Sandstone, 1 26 Elkin fauna, beds, 7, 1 02, 103, 105, 106, 1 07,
1 16, 1 1 9, 1 27, 135, 1 8 1 ; Table 1, p. xiv;
Table 30, p. 1 04 Ellesmerian Stage, 172 Elsdun Limestone, Table�O. p. 1 38 Emma Fiord Formation, Fig. 27, p. 1 1 7 Endibal "Series", 96, 98, 129; Table 1 5 , p. 58 Eotriassic, 1 9 Eoverbeekina zone, Table 1 8, p. 6 2 Epitaph Dolomite, 98 Ervay Carbonate Rock Member, 26, 1 35; Table
34, p. 1 20 Esayoo Formation, Fig. 27, p. 1 1 7 Eskridge Shale, Table 27, p. 91 Ettrain Formation, 46 Eungella Complex, Fig. 23, p. 93 Eurydesma beds, 77; Table 2a, p. xv; Table 19,
p. 65; Table 28, p. 1 00 Eurydesma mytiloides horizon, Table 38, p.
1 30 Evenk Horizon, 79; Table 15 , p. 5 8 Exmoor fauna, 137; Table 2b, p . xv; Table 36,
p. 1 25
Fantasque Chert, 1 35 Farley Formation, 95, 1 02, 1 26; Table 40, p.
1 38; Table 4 1 , p. 1 39 Faulkner Group, Table 40, p. 138 Fauna I (Australia), 95 Fauna II (Australia), 95, 1 02 Fauna Ilia (Australia), 99, 1 02 Fauna Illb (Australia), 1 26 Fauna Illc (Australia), 1 26 Fauna IV (Austrilia), 1 37 Fauna V (Australia), 156 Fauna D1 (Western Australia), 99 Fauna D2 (Western Australia), 1 24 Fenestella Siltstone Member, shale, 1 26, 1 27,
144; Table 40, p. 1 38; Table 4 1 , p. 139 Fern tree Group, 137, 169, 172; Table 40, p.
1 3 8 Filippovian Substage, 7, 26, 39, 96, 102, 1 03,
105, 106, 1 07, 1 09, 1 10, 1 1 3, 1 16, 1 1 8, 1 1 9, 1 22, 1 24 , 1 26, 1 27, 1 5 1
Five Point Limestone, 75 Flat Top Formation, 156; Table 36, p. 1 25;
Table 41, p. 139 Flowers Formation, 1 27 Foldvik Creek Formation, fauna, 3 1 , 1 16; Table
9, p. 30 Fossil Oiff Sandstone, 92; Table 29, p. 101 Franson Member, 26, 1 1 9; Table 34, p. 1 20 Freitag Formation, Table 36, p. 1 25; Table 41,
p. l 39 -
Fusulina Limestone, Table 25, p. 86
Gabrashitov Suite, 95 Gan Suite, Limestone, 1 06; Table 23, p. 81 Gangetian Stage, 1 72, 1 83 Gaptank Formation, 45, 49, 5 2, 72, 75; Table
2b, p. xv; Table 26 , p. 88; Figs. 1 2-15, pp. 50, 5 1 ; Fig. 23, p. 93
Gebbie Subgroup, 99, 1 26 ; Table 2b, p. xv; Table 36, p. 1 25
Geirud Member D, 84; Table 3 1 , p. 108 Gemmulicosta (Cg) zone, 46 Gerster Formation, 26, 1 3 5

Getaway Formation, 1 37 Gijigin Horizon, 1 29, 141, 149, 1 81; Table 15,
p. 58 Gipshuken Formation, Table 25, p. 86Glendale Limestone, 156; Table 37, p. 128Glendoo Member, 1 26; Table 36, p. 1 25 Glini Suite, 79; Table 14, p. 56 Gnishik Horizon, 36, 140, 1 5 1 ; Table 2a, p. xv Godrohiniiches Suite, 129 Golden Valley Group, 95 Gondolella bisselli Zone, Table 7, p. 27 G. carinata subcarinata Zone, 29 G. idahoensis Zone, 26; Table 7, p. 27 G. leveni Zone, 29 G. orienta/is Zone, 29 G. rosenkrantzi Zone, 26; Table 7, p. 27 Gosford Formation, Table 40, p. 1 3 8 Grandispora flora, 45 Grandeur Member, Table 34, p. 120 Grange Mudstone, 1 26; Table 40, p. 1 38 Grant Formation, Table 29, p. 101 Grenzland beds, 82; Table 24, p. 83 Greta Coal Measures, Table 40, p. 1 3 8; Table
41, p. 139 Greville Formation, 166, 169; Table 6, p. :t7;
Table 37, p. 1 28; Table 40, p. 1 38 Griesbachian Stage, 19, 22, 29, 37, 1 72, 1 74,
175, 1 83; Fig. 38, p. 1 73 Grodener Group, Table 24, p. 83 Gshelian Stage, 44, 45, 46, 49, 52, 55, 60, 66,
69; Fig. 11, p. 43; Figs. 12-15, pp. 50, 5 1 Guadalupian Series, 14, 26, 31 , 1 10, 135, 140,
146, 152; Table 10, p. 33 Gujo fauna, 148, 169 Gundarin Suite, 80, 97; Table 17, p. 60 Gusin Suite, 96; Table 14, p. 56 Gutan Horizon, 131 Gypsiferous "Series", 85; Table 25, p . 86 Gyranda Formation, Table 36, p. 1 25; Table
41, p. 1 39
Hachik Horizon, 36, 140, 151, 157, 161; Table 2a, p. xv
Haldan Suite, 48, 57; Table 13, p. 47; Table 16, p. 59
Haldjin Suite, 57; Table 16, p. 59 Halleck Formation, 98 Hanagiri Formation, Table 20, p. 67 Haranor Suite, 131 Harashibir Suite, 1 3 1 Harashibir Suite, Table 1 3, p . 47 Haraulakh (Tugasir) Suite, 1 29; Table 16, p.
59 Hardman Member, 1 56; Table 29, p. 101� Table
40, p. 138 Hare Fiord Formation, 72; Fig. 27, p. 117 Hartle Pool Dolomite, Table 33, p. 1 14 Hartle Pool Anhydrite, Table 33, p. 1 14 Hatalin Suite, Table 16, p. 59 Hatinach Suite, Table 13, p. 47; Table 16, p. 59 Haushi Formation, 84 Havililh fauna, 1 37, 156; Table 36, p. 1 25 Hawtel Formation, 169; Table 37, p. 128; Fig.
37, p. 173
Stratigraphic and Biostratigraphic Index 2 1 5
Hilton Member, Fig. 37, p. 173 Hess Formation, 90; Fig. 29, p. 1 36 Highbury Volcanics, Table 4 1 , p. 1 39 High Cliff Sandstone, Table 29, p. 101 Hiposhin Suite, 57; Table 13, p. 4 7 Hiraiso Formation, Table 1 8, p. 62 Hivach Horizon, 149; Table 15, p. 58 Holmwood Shale, 92; Table 29, p .. l O l Hoshan Limestone, 165 Hovtinden Member, Table 25 , p. 86 Hsiaochiang fauna, 1 09 Hueco Group, Table 26, p. 88 Hueco Canyon Formation, 90; Table 26, p. 88;
Table 27, p. 91
ldiognathodus ellisoni Zone, Table 7, p. 27 Imtachan Suite, Table 15, p. 58; Table 16, p. 59 lmtandjin Suite, 57; Table 15, p. 58; Table 16,
p. 59 Inai Group, Table 20, p. 67 lndiga limestone, 55 lngelara Formation, shale, 1 26; Table 36, p.
1 25; Table 40, p. 138; Table 41, p. 139 Inia Group, Table 20, p. 67 lntin beds, � 05, Table 14, p. 56 Iollikhar Suite, Table 1 7, p. 60 Irbichan Suite, 79, 80; Table 15, p. 58 Irenian Substage, 7, 1 03, 106, 107, 1 1 3, 1 19,
1 22, 126, 1 27, 1 5 1 , 1 79, 181 ; Fig. 38, p. 1 73
Irgin beds, 78, 95 Irwin River Coal Measures, Table 29, p. 101 Ishimbaevo oilfield, 79 Iwilhata Limestone, 148 Iwaizilhi Limestone, Table 20, p. 67 Iwato Formation, Table 48, p. 1 70
Jakutoproductus (Ej) zone, 87; Table 22, p. 73 Jigulevian Stage, 40, 44, 45 Jisu Honguer beds, 1 10, 1 32, 149, 151 , 163 Joe Joe Formation, 77; Table 2b, p. xv; Table
36, p. 1 25 Jungle Creek Formation, 46, 72; Table 2b, p. xv;
Table 22, p. 73
Kahlerina zone, Table 18, p. 62 Kaibab Formation, 1 19, 1 35 Kalabaghian Substage, 29, 1 29, 1 3 1 , 132, 134,
140, 143, 144, 145, 146, 148, 149, 151 , 156, 157, 165, 1 81
Kalabagh Member, 134, 140, 141 , 144, 149, 151 , 152, 153; Table 2a, p. xv; Table 9, p. 30; Table 19, p. 65; Table 28, p. 100
Kalinovian Substage, 29, 36, 107, 127, 1 29, 135, 137, 181
Kalitven Suite, 55 Kalkarrne beds, 6 1 , 66, 69; Table 24, p . 83 ; Fig.
1 3, p. 50; Fig. 14, p. 5 1; Fig. 15, p. 5 1 Kalkreiche beds, 66, 69; Table 24, p. 83; Fig. 13,
p. 50; Fig. 15, p. 51 Kalouzin Suite, 106 Kamian Stage, 1 03 Kamikuzu Formation, Table 20, p. 67; Table
48, p. 170 Hegler Limestone, 153, 166 ; Table 2b, p. xv; Table
26, p. 88 Kamura Formation, Table 48, p. 1 70 Kanokura Formation, Series, 148; Table 20, p.

2 1 6 Stratigraphic and Biostratigraphic Index
67; Table 48, p. 1 70 Kapp Starostin Formation, 1 1 3, 1 1 6; Table 25,
p. 86 Karachatirian Suite, faunas, 44, 60, 61, 64, 85;
Table 1 7, p. 60 Karamurum "Series", 78 Karhabari floral zone, 92 Karita Formation, 148 Kartamish Suite, 55 Kashirian level, 46; Fig. 1 1 , l'· 43 Kashkabash Suite, 95 Kasimovian Substage, Stage, 44, 45, 46, 49, 5 2,
55, 66, 69; Fig. 1 1 , p. 43; Fig. 1 3, p. 50; Fig. 14, p. 5 1
Kathwai Member, 161 , 163, 166, 1 75; Table 2a, p. xv; Table 19, p. 65; Table 28, p. 1 00
Kavalerov Suite, Table 43, p. 150 Kawahigashi brachiopod faunule, 1 48, 1 65 Kazanian Stage, 1 , 1 1 , 14, 16, 1 8, 26, 29, 3 1 ,
34 , 36, 96, 1 02, 103, 1 05 , 1 06, 107, 1 09, 1 1 3, 1 1 6, 1 1 8, 1 1 9, 1 22, 1 24, 127, 1 29, 1 3 1 , 1 32, 1 34, 1 35, 1 37, 143, 144, 145, 146, 149, 1 5 1 , 1 52, 153, 157, 163, 1 72, 1 79, 181 , 183
Kennedy Group, Table 29, p. 101 Kigiltass Suite, 80; Table 1 3 , p. 47; Table 15, p.
58� Table 1 6, p. 59 Kingoceras Shale, Table 35, p. 1 23 Kizildjilgin Suite, 82; Table 23, p. 81 Kochiproductus-Attenuatella (Eka) zone, 5 2,
12: Table 22, p. 73 Kochiproductus zone, Table 2b, p. xv Kockatea Shale, 1 75; Table 29, p. 1 01; Table 40,
p. 1 3 8 Kokpecten fauna, 60, 69 Koten "Series", 64 Kozaki Formation, 1 10; Table 20, p. 67 Kozlowskia zone, 46 Krasnoufimian Substage, 96, 102 Kubergandin Suite, 97, 1 06; Table 23, p. 8 1 ;
Table 3 8 , p. 1 30 Kufeng Shale, 1 09 Kufengoceras zone, 109 Kukkan Suite, 96; Table 15, p. 58; Table 16, p.
59; Table 43, p. 150 Kuktui beds, 97 Kuling Shales, 1 43, 145 Kuma Formation, 1 65; Table 20, p. 67; Table
48, p. 170 Kuman Stage, 148, 1 65 Kungurian Stage, 1 , 7, 1 1 , 14, 1 8, 29, 3 1 , 36, 3 9,
96, 97, 1 02, 103, 1 05, 106, 1 07, 1 09, 1 1 0, 1 1 3, 1 16, 1 1 8, 1 1 9, 1 22, 1 24, 1 26, 1 27, 1 29, 1 3 1 , 1 3 � 1 34, 1 35, 1 37, 143, 145 , 146, 149, 1 5 1 , 1 72, 1 79, 1 81 , 1 83
Kurmaian Substage, 1, 53, 55, 51, 64, 69, 7 2, 75, 77, 79, 87, 92, 1 76
Kurmain Formation, Figs. 1 2�15, pp. 50, 5 1 Kuseler schichten, Fig. 1 1 , p . 43
La Colorada beds, 1 55, 1 66 Lamar Limestone, 1 5 2, 153, 1 6 1 , 166; Table 26,
p. 88 Lamnimargus himalayensis Zone, 1 43, 144, 145�
Fig. 30, p. 142 Lantschichites Zone, Table 1 8, p. 62 Laoshan shale, 1 65 Leine "Series", Table 33, p. 1 14 Leiorhynchoidea zone, Fig. 28, p. 1 2 1 Lenox Hills Conglomerate, basal, Table 27, p. 91 Lenox Hills Formation, 80, 87, 90, 92, 98;
Table 2b, p. xv; Table 26, p. 88; Table ·27, p. 91; Fig. 29, p. 1 36
Leonardian Stage, 26, 34, 36, 78, 87, 90, 98, 1 22, 145; Table 27, p. 9 1
Lepidolina kumanensis Zone, 149, 1 65 L. toriyamai Zone, 149; Table 20, p. 67 Leptodus richthofeni faunule, 149 Letham Formation, Table 37, p. 1 28; Table 40,
p. 1 3 8 Levorkut Silite, 1 05 Liangfengpo Shale, 169 Lightjack Member, 1 26; Table 29, p. 101 Lissochonetes zone, Table 2b, p. xv; Table 22,
p. 7 3 Lithgow Coal Measures, 1 7 2 Little Bed Sandstone, 156, 1 66; Table 6, p . 1 7,
Table 37, p. 1 28; Table 40, p. 1 3 8 Liveringa Formation, 1 26, 156; Table 29, p.
1 0 1 Lizzie Creek Volcanics, 95 ; Table 2b, p. xv; Table
36, p. 1 25; Table 40, p. 1 38; Table 41, p. 1 39; Fig. 23, p. 93
Lochinvar Formation, 77; Table 40, p. 1 38; Table 4 1 , p. 1 39; Fig. 1 2, p. 50; Fig. 14, p. 51; Fig. 15, p. 5 1
Loping Coal Measures, 1 65 Lopingian fauna, 145, 149, 165; Table 1 8, p. 62 Lower Productus Limestone (= Amb), 99 Luang Prabang Greywacke, Table 42, p. 147 Lungli Limestone, 64 Lusaba Limestone, 84 Luta Limestone, 75 Lydite horizon, 1 09 Lyons Group, 77; Table 29, p. 101; Table 40,
p. 1 38
Madeline Formation, 99 Magar Suite, Table 1 3, p. 4 7 Magnesian Limestone, 1 1 , 1 1 0, 1 1 3; Table 33,
p. 1 14 Maitai Group, Table 2b, p. xv Maitland Group, 1 26 Maizuru Group, 148; Table 48, p. 1 70 Makarov Horizon, 46; Table 1 3 , p. 47; Table
16, p. 59 Malbina Formation, 1 24, 1 26, 1 27, 1 37, 1 72;
Table 40, p. 138 Mallens Greywacke, Table 29, p. 101 Manchez reefs, 96 Mangapirian fauna, 1 02 Mangarewa Formation, Table 6, p. 1 7; Table
37, p. 1 28; Table 40, p. 1 38 Manzanita Member, 152; Table 26, p. 88 Maokou Limestone, 97, 109, 1 1 0, 1 3 1 , 1 32,
146, 165; Table 2b, p. 5; Table 9, p. 30; Table 1 8, p. 62; Table 1 9, p. 65
Maping Limestone, 64, 66, 165 ; Table 2b, p. xv;

Table 9, p. 30; Table 1 8, p. 62; Table 1 9, p. 65
Marble Canyon limestone, 1 1 8 Marginifera typica beds, Table 38, p . 1 3 0 Martinia adentata Zone, Table 37, p. 128 Martinia shale (Greenland), 1 16 ; Fig. 26, p. 1 1 5 Martinia zone (New Zealand), Table 2b, p . xv Martiniopsis woodi Zone, 156; Table 2b, p. xv
Table 6, p. 1 7; Table 37, p. 1 28 Mayachkovian level, 46, 66; Fig. 1 1 , p. 43 Mazzin Member, 1 74 McCloud Limestone, 72 McComb's beds, Table 26, p. 88 Meade Peak Member, 26, 1 09, 1 1 9; Table 34, p.
1 20 Megen Suite, 57; Table 16, p. 59 Menkechen Suite, 1 29; Table 15, p. 58� Table
16, p. 59 Me"illina arcucristatus Zone, 26 ; Table 7, p. 27 M' divergens Zone, 26, 29; Table 7, p. 27 M galeatus fauna, 26; Table 7, p. 27 Mianwali Formation, 161 ; Table 28, p. 1 00 Miaohou Limestone, 69 Middle Kalkarme Group, 44; Figs. 1 2 - 1 5 , pp.
50, 51 •
Middle Productus {= Warga!) Limestone, 84 Mimosa Group, Fig. 23, p. 93 Mingenew Formation, Table 29, p. 101 Minkinfjellet Member, Table 25 , p. 86 Misellina subzone, Table 1 8 , p . 62; Table 23,
p. 81M claudiae Zone, 85 Missellina·Darvasites zone, Table 23, p. 81 Mistletoe Sandstone, 1 26 Mississippian Subperiod, 42 Missourian Stage, 45, 5 2, 61 Mitai Formation, 165, 169; Table 48, p. 1 70 Mittiwali Member, Tabli� 28, p. 1 00 Mizukoshi Formation, 148 Monodiexodina zone, 85, 99 Monos Formation, 1 22, 1 37 Moran Formation, Table 27, p. 91 Moscovian Stage, 40, 42, 45, 46, 48, 49, 6 1 , 66,
77, 78, 176 Mount Bayley Formation, Fig. 27, p. 1 1 7 Mourlonia impressa bed, Table 3 7, p. 1 28 Mulbring Subgroup, 1 37; Table 40, p. 1 38 Mungadan Sandstone, Table 29, p. 1 01 Munugudjak Horizon, 79; Table 15 , p. 58 Muree Sandstone Member, beds, 1 37; Table 40,
p. 1 38; Table 4 1 , p. 1 3 9Murgabian Stage, 3 6 , 1 31 , 1 5 1
Nabeyaman Stage, 9 7 , 1 00 Nagatoella subzone, Table 1 8, p. 62 Nakakubo Formation, 64 Nalbia Greywacke, 1 24; Table 29, p. 101 Namurian dolomite, Table 1 2, p. 41 Nangetty Formation, Table 29, p. 101 Nankinella zone, Table 18, p . 62 N. orbicularia Zone, 1 3 1 Nankinella - Cancellina zone, 1 3 1 Nanson Formation, Fig. 27, p . 1 1 7 Narmia Member, Table 28, p. 1 00 Nassau Siltstone, Table 40, p. 1 38
Stratigraphic and Biostratigraphic Index 2 1 7
Natalin Suite, Table 1 3, p. 47 Neal Ranch Formation, 72, 75, 90; Table 2b, p . xv;
Table 26 , p . 88; Table 27, p. 91 ; Figs. 1 2-1 5 , pp. 50, 5 1
Nenetz beds, 5 5 Neochonetes·Pseudosyrinx zone, Table 22, p.
73 Neogondolella serrata zone, 26; Table 7, p . 27 Neomisellina zone, Table 1 8 , p. 62 Neoschwagerina zone, 1 3 1 ; Table 1 8 , p. 62 N. craticulifera Zone, 1 32; Table 20, p. 67 N. margaritae Zone, 1 32; Table 23, p. 81 N. simplex Zone, 1 09, 1 1 0; Table 20, p. 67;
Table 23, p. 81 N. schuberti Zone, Table 23, p . 81 Neospathodus arcucristatus fauna, 26 Neostreptognathodus sulcoplicatus fauna, Table
7, p . 27 Nesen Formation, 1 6 1 ; Table 3 1 , p. 108 Nevolin level, fauna, 7 , 1 1 , 26, 103, 1 05, 1 06,
107, 1 09, l l O, 1 1 3 , 1 16, l l 8, 1 1 9, 1 22, 1 24, 1 26, 1 27, 1 34, 1 35, 1 37, 1 53 , 1 72; Table 1, p. 3; Table 30, p. 1 04
Newcastle Coal Measures, Table 40, p. 1 38 Nikitin Horizon, 1 3 1, 1 5 1 Nilawan Group, Table 2 8 , p. 1 00 Nippewalla Group, 90; Table 27, p. 81 Nishikori Formation, 85 Nooncanbah Formation, 1 26; Table 29, p. 101 Nordenskioldbreen Formation, 69, 85; Table 25,
p. 86 Norton Greywacke, 1 24 Nosoni Formation, 1 1 8 Notospirifer zone, Table 2b, p. xv Notostrophia zone, Table 2b, p. xv Notostrophia homeri Zone, Table 37, p. 1 28 N. zealandicus Zone, Table 37, p. 1 28 Nowra Formation, Table 41 , p. 1 39 Nuculana nogamii Zone, Table 48, p. 170 Nura Nura Member, 92; Table 29, p. 1 01
Oblachan Suite, Table 16, p. 59 Ochoan Group, Table 2b, p. xv Ochoan "Stage", 5 Ogbinan Substage, 163, 168, 169, 1 83 Omolon Horizon, 1 29; Table 1 5 , p. 58 Omphalotrochus zone, 45 One Gum Formation, 95 Ophiceras commune Zone, 29 Ordovician Period, 31 Orenburgian "Stage", Substage, 40, 42, 44, 45,
46, 49, 52, 53, 60, 66, 69, 85, 1 76; Figs. 1 2-15, pp. 50, 5 1
Orion Shale, Table 36, p . 1 25; Table 4 1 , p. 139
Ota Group, Table 32 . p. 1 1 1 Orthotichia (Eo) zone, 72; Table 2b, p . xv;
Table 22, p. 73 Orthotichia-Septospirifer (Dos) zone, 46 Otoceras concavum Zone, Fig. 38, p. 1 73 Otto Formation, Fig. 27, p. 1 1 7 Ottweiller Group, Fig. 1 1 , p . 43 Owl Creek Volcanics, Table 36, p. 1 25 Oxtrack Formation, Table 36, p. 1 25 ; Table 4 1 ,
p. 1 39

2 1 8 Stratigraphic and Biostratigraphic In dex
Paikhoyan Stage, 1 02 Palaeofusulina zone, Table 18, p. 62; Table 23,
p. 81 Palaeofusulina·Reichelina fauna, Fig. 34, p. 167 Palaeozoic, 24 Pamiran Stage, 157, 163; Table 23, p. 81 Parafusulina fauna, zone, subzone, 99, 109;
Table 11 , p. 35; Table 1 8, p. 62; Fig. 28, p. 121
Parafusulina spissisepta Zone, 98 P. durhami Zone, 98 P. kaerimizensis Subzone, Table 20, p. 67 Parapara Group, 1 27 Parapronorites beds, Table 38, p. 1 30 Paratirolites zone, Table 2a, p. xv Paren Suite, 57, 60; Table 1 5, p. 5 8 Park City Group, 26, 1 19, 1 35 Patuki Volcanic Melange, Table 37, p. 1 28 Payu Limestone, 64 Peawaddy Formation, fauna, 1 37; Table 36, p.
125; Table 40, p. 1 38; Table 4 1 , p. 139 Pelican Creek fauna, 137; Table 2b, p. xv; Table
36, p. l 25 Pennsylvanian Subperiod, 45, 72 Permian, 40, 42 Permian System, 5, 1 8, 19, 23, 25, 26, 40, 42,
75 Permian Period, 1, 7, 1 1 , 1 8, 19, 20, 22, 25, 34,
38, 1 1 3, 140, 1 72, 176 Permian early (= lower), 1 , 14, 20, 31, 34, 36,
39, 44, 45, 46, 48, 61, 66, 75, 79, 84, 106, 144, 145
Permian middle, 1 , 1 1, 16, 19, 29, 3 1 , 34, 39, 109, 1 29, 156, 1 72
Permian late (= upper), 5, 16, 1 8, 1 9, 25 , 29, 34, 36, 39, 1 1 3, 1 16, 1 29, 145, 149, 156, 168, 1 72, 1 81
Permian-Carboniferous boundary, 44, 5 2, 72, 176
Permian-Triassic boundary, 19 Permophricodothyris grandis Zone, 146 Permundaria sisophonensis beds, 1 31; Table 38,
p. 1 30 Perrinites Shale, Table 35, p. 1 23 Perrinites Zone, Table 1 1 , p. 35; Table 43, p.
159 Peschanik Suite, Table 14, p. 56 Phisonites-Comelicania zone, 161 , 168 Phisonites-Paratirolites zone, 29 Phosphoria Complex, 16, 26, 1 1 8, 1 19, 135 Pine Bush Formation, 166 Pinery Limestone, Table 26, p. 88 Plekonella multicostata Zone, 156; Table 6, p.
17; Table 17, p. 1 28 Plympton beds, 26 Podolian Zone, 46; Fig. 1 1 , p. 43 Pokrov Suite, 55 Pong Oua Limestone, Table 42, p. 147 Poole Sandstone, 92; Table 29·, p. 101 Poplar Tank Member, Table 26, p. 88 Posidonia Shale, beds, 1 16; Fig. 26, p. 1 15 Potonieisporites .flora, 45 Powwow Conglomerate, 90 Praehorridonia zone, 46 Productus Creek Group, 127; Table 2b, p. xv
Productus Limestone (Greenland), Fig. 26, p. 1 1 5
Productus Limestone (Madagascar), 153 Productus Limestone (Pakistan), Table 28, p.
1 00 Productus Shales (Himalayas), 1 16, 143, 145;
Table 9, p. 30 Properrinites zone, Table 1 1 , p. 35 Prouddenites fauna, 5 2 Pseudofusulina horizon, 52; Table 1 8, p . 62;
Fig. 1 1 , p. 43 P. ambigua Zone, 85 P. chihsianensis Zone, 85 P. * tschemyschewi Zone, 85 P. * vulgaris Zone, 85 Pseudodololina Subzone, Zone, Table 1 8, p. 62 Pseudosch»'agerina Limestone (Austria) 82;
Table 24, p. 83; Fig. 1 3, p. 50; Fig. 15, p. 5 1
Pseudoschwagerina Limestone (Spitsbergen), 69; Table 24, p. 83
Pseudoschwagerina Zone, Table 1 8, p. 62 Ps. minatoi Subzone, 66 Ps. morikawai Subzone, Table 20, p. 67 P� uddeni Zone, 15 Pseudosyrinx Zone, Table 2b, p. xv Pueblo Formation, Table 27, p. 91 Punctospirifer pulchra fauna, 26 Punjabian Stage, 1 1 , 36, 1 07, 1 10, 1 31, 140,
143, 144, 145, 148, 149, 1 5 1 , 152, 153, 157, 161 , 163, 165, 1 75, 181
Purdonella-Gibbospirifer zone, 46 Puruhauan Stage, 149 Putnam Formation, Table 27, p. 9 1 Pybus Formation, 1 1 8
Quamby Group, 7 7 Quinnanie Formation, 99
Rader Limestone, 26� Table 26, p. 88 Rai Sandstone, Table 37, p. 128 Rammutt Formation, Table 36, p. 1 25; Table
41 , p. 1 39 Rands Formation, Table 36, p. 1 25 Ranger Canyon Chert, 1 1 8, 1 19, 135 Rattendorfer Group beds, 66, 69, 82; Table 24,
p. 83 Ravensfield Sandstone, 95 Reedoconcha fauna, 92 Reichelina fauna, Table 1 8, p. 62: Table 23, p. 81·,
Fig. 34, p. 167 Reticulatia zone, 46 Retort Member, 1 1 9; Table 32, p. 1 1 1 Rewan Formation, Table-2b, p . xv; Fig. 23, p.
93 Rex Chert, 1 19, 1 35; Table 34, p. 1 20 Road Canyon Formation, 39, 102, 1 16, 1 19,
122; Table 2b, p. 5; Table 26, p. 88; Fig. 29, p. 136
Roadian Stage, 26, 3 1 , 34, 36, 39, 1 1 8, 11 9, 1 26
Roker Dolomite, Table 33, p. 114 Ross Creek Formation, 87 Rudnitz Horizon, 105; Table 14, p. 56 Russiella subzone, Table 1 8, p. 62

Stratigraphic and Biostratigraphic Index 2 1 9
Rustler Formation, 166 Ruteh Formation, 106, 152, 161, 166; Table 31,
p. 108Rutherford Formation, 95; Table 41, p. 1 39
Saarbucker Group, Fig. 11 , p. 43 Safetdaron Suite, 79, 82, 84, 85, 90, 145;
Table 17, p. 60 Sakamena Group, 153 Sakamotozawan Stage, Formation, 64, 85;
Table 20, p. 67 Sakmarian Stage, 1 1 , 14, 1 8, 26, 36, 40, 42,
44, 48, 53, 55, 57, 64, 66, 72, 75, 78, 79, 82, 84, 85, 87, 90, 92, 95, 98, 102, 1 06, 107, 116, 1 24, 135, 145, 176, 1 79; Table 10, p. 33
Sakoa Group,.92 Salt Creek Bend Shale, Table 27, p. 91 Samaran fauna, 66 Samegai Formation, Table 32, p. 1 1 1 San Andreas Formation, 99 Saranin level, 96 Sardi Formation, Table 2a, p. xv; Table 19, p.
65� Table 28, p. 100 Sarginian Substage, 96, 102 Sasnian Horizon, 97 Schwagerina "Stage", subzone, 42; Table 1 8,
p. 62Schwagerina crassitectoria bed, 40; Table 26, p.
88; Table 27, p. 91 Scottville fauna, 137; Table 2b, p. xv; Table
36, p. 125 Scythian Stage, 172 Seaham Formation, Varves, 45, 49, 52; Fig.
12, p. 50; Fig. 14, p. 5 1� Fig. 15 , p. 5 1 Seaham Residue, Table 33, p . 1 14 Sedov "Series",.79, 1 29 Selung formation, 143, 145 Setachan Suite, Table 16, p. 59 Sezim Suite, 55, Table 14, p. 56 Shakharsev Suite, Table 17, p. 60 Shazagai Suite, Table 1 3, p. 47 Shazagaitui Suite, 5 7, Table 1 3, p. 47 Shedhorn Sandstone, 1 1 9; Table 34, p. 1 20 Shigejizawa Sandstone Member, 148, 152 Shimodake Formation, Table 20, p. 67 Shimokuzu Formation, Table 20, p. 67 Shiraiwa Formation, Table 32, p. 1 1 1 · Sibaigou Suite, Table 43, p. 150 Silvaseptopora zone, 75 Sirius Shale, 95 ; Table 2b, p. xv; Table 36, p.
125 Sisophon Member A, 1 10 Sisophon Member B, 1 3 1 , 132 Sisophon Member C, 1 34, 146 Sisophon Member D, 146 Skinner Ranch Formation, 72, 87, 90, 92;
Table 26, p. 88; Table 27, p. 91� Fig. 29, p. 136
Smithian Stage, 19, 172, 1 74 ; Fig. 37, p. 173 Sokolin Horizon, Suite, 105; Table 15, p.
58; Table 16, p. 59 Solikamian salt deposits, 103 Solonchan Suite, Table 16, p. 59
Somohole fauna, Table 19, p. 65; Table 44, p. 1 54
Sosio beds, 26, 29, 1 34 Sosnovian Substage, 26, 127, 1 29, 135 , 1 37,
181 South Curra Limestone, 1 56; Table 36, p. 1 25;
Table 41, p. 1 39 South wells beds, Table 26, p. 88 Sowerbina zone, 98; Table 2b, p. xv; Table 22,
p. 73Soybol Suite, Table 1 3, p. 47; Table 16, p. 59 Speckled (= Warchha) Sandstone, 77; Table 9, p.
30; Table 28, p. 100 Sphaerulina zone, Table 18, p. 62 Spinomartinia zone, Table 2b, p. xv Spinomartinia spinosa Zone, 156, 160; Table 6,
p. 17; Table 37, p. 1 28 Spirifer Limestone, 1 1 3, 1 16; Table 25, p. 86 Spiriferella bed (Mongolia), 151 Spiriferella supplanta Zone, Table 37 , p . 1 28 Spiriferella zone, Table 6, p. 1 7 Spiriferellina zone, 1 3 7 ; Fig. 28, p. 1 21 Spiriferenschicht, 69 Spiti sandstone, 143 Staircase Sandstone, Table 36, p. 1 25; Table 41,
p. 1 39 Stanleigh Formation, Table 36, p. 1 25; Table 41 ,
p. 1 39 Stepanoviella-Taeniothaerus)(Reedoconcha)
fauna, 92 Stepanoviella umariensis, Table 38, p. 130 Stephanian Stage, 44, 45; Fig. 1 1 , p. 43 Stephanian A, 44, 49, 69 Stephanian C, 44, 49 Stephens Formation, Table 6 , p. 17; Table 37,
p. 1 28: Table 40, p. 1 3 8Sterlitamakian Substage, 5 7 , 61, 78, 79, 80, 84,
85, 87, 90, 92, 95, 1 26, 179 Stone Corral Formation, 90: Table 27, p. 91 Strassfurt Series, Table 33, p. 114 'Streptorhynchus' pelicanensis bed, 137 strigatus Zone, Fig. 38, p. 173 Suisi Member, 175 Sumatrina zone, Table 18, p. 62 Summer Group, 75, 90; Table 27, p. 91 Surenan Substage, 7, 39, 49, 52, 53, 55, 69, 72,
176� Figs. 12-15, pp. 50, 5 1 Surkechan Suite, Table 13, p . 4 7 Suyorgan Suite, 46, 48, 5 7; Table 1 3, p . 47;
Table 16, p. 59 Svalbardian Stage, 102 Svenskeegga Member, Table 25, p. 86 sverdrupi Zone, Fig. 38, p. 173 Sweetognathodus whitei Zone, 26; Table 7, p.
27; Table 16, p. 59 Syuren Suite, Table 16, p. 59
Taeniothaerus permixtus Horizon, Table 38, p. 130
Tahkandit Formation, 97, 98, 1 1 8, 1 35; Table 2b, p. xv; Table 22, p. 73
Taiyuan Limestone, 64, 66, 69 Takauchi fauna, l48 Takitimu Group, 95 , 102; Table 2b, p. xv; Table

220 Stratigraphic and Bios.tratigraphic Index
37, p. 1 28; Table 40, p. 1 38 Taktabulak Suite, 163; Table 23, p. 8 1 Talatin Suite, 96, 9 8 , 99, 1 05; Table 14, p . 5 6 Talchir Conglomerate, 77 ; Table 2a, p. xv; Table
9, p. 30; Table 1 9, p. 65; Table 28, p. 1 00 Talung Formation, 169 Tamaree Formation, 166; Table 2b, p. xv; Table
36, p. 1 25; Table 41 , p. 1 39 Tamba Zone, Table 32, p. 1 1 1 Tanquary Formation, Fig. 27, p . 1 17 Tarviser Breccia, 82 Tastubian Substage, 48, 72, 78, 79, 80, 84, 85,
87, 92, 95, 98, 179 Tatarian Stage, 5, 1 8, 34, 1 1 3 Tei Wei Fauna, Table 1 9, p . 65; Table 44, p . 1 54 Telford Formation (Canada), 72, 87, 1 02 Telford Stage (New Zealand) , 95 , 102 Tenjinnoki Formation, 132 Terauchi Formation, Table 32 , p. 1 1 1 Terrakea brachythaerum Zone, 1 37, 140, 157;
Table 2b, p. xv; Table 6, p. 17 ; Table 3 7, p . 1 28
Terrakea concavum Zone, Table 2b, p. xv T. dickinsi Zone, Table 2b, p. xv Tertiary, Fig. 23, p. 93 Tesero Horizon, 1 74 Tiksin Suite, 46, 55; Table 1 3, p. 47; Table 15 ,
p . 5 8; Table 16, p . 59 Tilicho Lake Formation, 144 Timorites Shale, Table 35, p. 1 23 Timorites zone, 140, 152; Table 1 1 , p. 35; Table
43, p. 1 50 Tingchiashan Formation, 1 09, 1 3 1 Tirin Horizon, 149 Tiverton Subgroup, 95, 102; Table 2b, p . xv;
Table 36, p. 1 25; Fig. 23, p. 93 Tobra Formation, Table 28, p. 100 Tomago Coal Measures, Table 40, p. 1 38; Table
41 , p. 139 Tomiopsis (= Eta) zone (Canada), Table 2b, p. xv Tomiopsis beds (Afghanistan), Table 38, p. 1 30 Tompin Series, Table 15 , p. 5 8 Tonbo Limestone, 1 1 0 Toriyamaia subzone, Table 18 , p. 62 Tornquistia (Et) zone, 87 , 90 ; Table 2b, p. xv;
Table 22, p. 73 Toroweap Formation, 98 Tosi Chert Member, 1 19, 1 35; Table 34, p. 1 20 Toyoma Group, Formation, Table 20, p. 67;
Table 48, p. 1 70 Tozerian Stage, 172 Tramway Sandstone, 156; Table 6, p. 17 Treskelloden beds, 69, 87; Table 25, p. 86 Triassic, 143, 161 , 1 63, 168, 172, 1 74 Triassic early, 1 9, 25, 34, 37, 163, 1 72, 175 ,
183 Triassic, pre, 1 8 Triticites zone, 6 9 Triticites opimus Zone, 7 5 Trogkofel Formation, 69, 82, 85, 1 35 , 145;
Table 24, p. 83 Trold Fiord Formation, 1 19, 1 35; Table 22, p.
73; Table 27, p. 1 1 7 Tsukumi Limestone Group, 165 Tsune-Mori Formation, Table 32, p. 1 1 1; Table
48, p. 1 70 Tufolavov beds, Table 15, p. 58 Tugasir Suite, 57 Tumarin Suite, 96� Table 15 , p. 58; Table 16,
p. 5 9 . Turuzov Suite, 55, 57, 79; Table 13, p. 47;
Table 1 5 , p. 5 8; Table 16, p. 59 Tuthaltu Suite, Table 1 3, p. 47 Tyloplecta zone, 161 Tyloplecta nankinensis Zone, 1 32 Tyrellfjellet Member, 69, 85; Table 25, p. 86
Uchagan beds, 5 7 Uchbulak Horizon, 44 Uddenites shale, 45, 52, 72; Table 26, p. 88 Uiunian "Stage" or horizon, 7, 1 1 , 26, 31, 34,
1 02, 1 03, 105, 1 07 , 1 1 3, 1 16, 1 19, 1 27, 1 29, 1 35 , 1 37, 1 8 1 ; Table 1, p. xiv; Table 30, p. 1 04
Ulkuk Suite, Table 43, p. 1 50 Ulladulla mudstone, fauna, 1 26; Table 40, p.
1 38; Table 4 1 , p. 139 Umari,a beds, 92 Ungadu Suite, 80 Ungun Suite, Table 43, p. 150 Upgang Carbonate, Table 33, p. 1 14 Upper Marine Group, 1 26 ; Table 2b, p. xv Upper Productus Limestone (= Chhidru), 84,
141 Uralian assemblage, 44, 53 , 64 Urushtenian Substage, 29, 37, 1 35 , 1 53, 156,
157, 161 , 1 63, 165, 1 66, 183 Uskalikian Substage, 7 , 53, 55, 69 , 72 , 75 , 77,
85; Fig. 1 2-15 , pp. 50, 5 1 Usuginu Conglomerate, Table 20, p . 6 7
Van Hauen Formation, 1 1 8; Table 22, p. 73; Fig. 38, p. 173
Vedian Substage, 16, 39, 1 68, 1 69, 172, 183 Vedioceras-Haydenella fauna, 1 6 1 Vedioceras ventroplanum Zone, 29 Vereyan Horizon, Fig. 1 1 , p. 43 Verchoyan Suite, 57; Table 1 3 , p. 47; Table
16, p. 59 Verbeekina zone, Table 1 1 , p. 35 ; Table 18 , p. 62 V. verbeeki-shiraiwensis Subzone, Table 20, p.
67 Vesliyan Horizon, 1 29 Victorio Peak beds, Table 26, p. 88 Virgilian Stage, 45, 5 2, 75; Fig. 1 1 , p. 43 Vohitolia marine limestone, 92 V�ringen Member, 1 13; Table 25, p. 86Vorkut Series, 105; Table 14, p. 56 Vrynetzev Suite, 55
Waagenoceras Shale, Table 35, p. 1 23 Waagenoceras zone, 1 3 1 , 1 37 ; Table 1 1 , p. 35 Wagina Sandstone, Table 29, p. 101 Waidegger fauna, 66 ; Table 2 1 , p. 70 Waiitian fauna, 169 Wairaki Breccia, 174 ; Table 37, p . 1 28 ; Fig. 37 ,
p . 1 73 Wairakiella rostrata Zone, Table 37, p. 1 28 Waiua Formation, Table 6, p. 1 7; Table 3 7, p.
1 28

Wandagee Formation, 99 Wandrawandrian Siltstone, 1 26, 1 72� Table 4 1 ,
p. 139 Warchha Sandstone, 77; Table 2a, p. xv; Table
19, p. 65; Table 28, p. 100 Wargal Formation, limestone, 107, 1 10, 1 24,
131, 134, 141, 143, 144, 145, 146, 148, 149, 15 1 , 152; Table 2a, p. xv; Table 9, p. 30; Table 19, p. 65� Table 28, p. 100
Weetwood Tuff, Table 37, p. 1 28 Werfener beds, 1 74 Werra Formation, "Series", 1 10, 1 1 3� Table 33,
p. l l4 Westphalian Stage, 69; Fig. 1 1 , p. 43 Whitehorse Sandstone, 1 66 Willis Ranch Member, 1 1 8, 1 35, 1 37; Table 2b,
p. xv; Table 26, p. 88; Fig: 29, p. 136Wolfcampian Stage, 72, 78, 85, 87, 90, 98; Table
27, p. 91� Fig. 1 1 , p . 43 Wollong Sandstone Member, Table 40, p. 1 3 8 Wooded Peak Limestone, 1 56; Table 6, p. 1 7 ;
Table 37, p . 1 28
Stratigraphic and Biostratigraphic Index 221
Wooramel Group, 95; Table 29, p. 101; Table 40, p. 138
Wordian Stage, 14, 26, 3 1 , 36, 39, 98, 1 02, 122, 124, 1 31, 1 35, 1 37, 140, 145, 153, 179
Word Formation, 18, 1 1 8, 122; Table 2b, p. xv; Fig. 29, p. 1 36
Wordiekammen Limestone, 69, 85; Table 25 , p. 86
Wuchiaping Limestone, 146, 165, 169; Table 2b, p. xv; Table 9, p. 30; Table 1 8 , p . 62; Table 19, p. 65
Yabeina zone, 1 3 1 , 1 34, 136; Table 1 1 , p. 3 5 , Table 1 8 , p. 6 2
Yabeina·Lepidolina assemblage, 140 Y. shiraewensis Zone, 149 Y. yaliUbaensis Subzone, Table 20, p. 67 Yakovlevia (Ey) zone, 80, 87; Table 2b, p. xv;
Table 22, p. 73 Yakano Formation, Table 48, p. 1 70 Yamasuge Member, 97 Yangchiena subzone, Table 18, p. 62 Yangsing Limestone, Table 9, p. 30 Yangsingian Series, Table 1 8, p. 62 Yardren dolomite, 1 29 Yarrol Formation, 95; Table 36, p. 1 25; Table
41, p. 1 39 Yasachnin Horizon, 79; Table 1 5, p. 58 Yayamadake Formation, Limestone, 66; Table
20, p. 67 Yomagu Formation, Table 48, p. 1 70 Yukisawa Group, Table 20, p. 67 Yunargin Series, 55, 96; Table 14, p. 56 Yupenchin Suite, 46, 57; Table 1 3, p. 47; Table
16, p. 59
Zaostren Horizon, 55; Table 14, p. 56 Zechstein Group, 26, 29, 55, 1 16 Zechstein (= Werra) Formation, 1 10, 1 29 Zechsteinkalk, 1 1 3 Zewan Formation, 143, 145, 1 74; Table 9 , p.
30 Zhigulian (= Jigulevian); 40, 44, 45 Zigar Suite, 80; Table 1 7, p. 60

Systematic Index for Genera and Species
Afghanella Thompson, Table 1 8, p. 62 Agathiceras Gemmellaro, 72, 78, 82, 97, 99,
1 24, 1 37, 1 5 2; Table 50, p. 178 mediterraneus Toumansky, 82 uralicus (Karpinsky), 46, 48, 80, 97
Akmilleria Ruzencev, 42 Aktubinskia Ruzencev, 78; Table 50, p. 178 Alaskanella yukonensis (Skinner & Wilde), 87,
98; Table 50, p. 178 Alatoconcha vampyra Termier et al. , Table 38,
p. 130 Aldanites rdtundus (Popov), Table 13, p. 47 "Altudoceras" cf roadense Bose, 106, 109 "Ambocoelia" Hall, 45, 48, 53, 57, 61, 69,
1 09 Ananias campbelli Waterhouse, 166 Anchignathodus Sweet, Table 7, p. 27
parvus Kozur & Pjatakova, 24 ; Table 8, p. 28 typicalis Sweet, 1 74, 1 75; Table 8, p. 28
Anemomtria Cooper & Grant, 99, 103 "pseudohorrida" (not Wiman), 98
Anidanthinid weyprechti (Toula), 141 Anidanthus Whitehouse, 57, 79, 80, 87, 96, 98,
1 1 8, 1 1 9, 1 22, 152; Fig. 28, p. 121 "aagardi gallatinense" (not Girty) of
Frederiks (1925), 106, 109 boikowi Stepanov, Table 13, p. 47 eucharis (Girty), cf 1 1 8, 1 1 9; Table 51, p. 1 80 fusiformis Waterhouse, 143, 145 minor (Cooper), 72, 1 19; Table 5 1 , p. 1 80;
Fig. 28, p. 121 sinosus (Huang), 169 so/ita Waterhouse, 1 26; Table 51, p. 180 springsurensis (Booker), 95; Table 50, p.
178 Antiquatonia Miloradovich, 46, 97, 99; Table 2b,
p. xv; Table 22, p. 73 ; Table 50, p. 178bassi McKee, 98 cf sulcatus Cooper, 1 19
Aperispirifer Waterhouse, 1 27� Table 37, p. 1 28 lethamensis Waterhouse, 140; Table 5 1 , p.
180 nelsonensis (Waterhouse), Table 2b, p. xv
Table 6, p. 17; Table 37, p. 128 wairakiensis (Waterhouse), 126, 1 27
Araxathyris Grunt, 168 araxensis Grunt, 169
Araxilevis Sarytcheva, Table 2a, p. xv; Table 45, p. 158; Table 46, p. 1 59; Table 52, p.1 82
Araxoceras Ruzencev, 161, 165 ; Table 45, p. 158; Table 46, p . 159; Table 52, p . 182 latum Ruzencev, 29
Arctutreta pearyi (Whitfield), 1 16 kempei (Anderson), 1 16 triangularis (Wiman), 1 16
Arctochonetes Ifanova, Table 5, p. 13 postartiensis (Ustritsky), 105
Aristoceras Ruzencev, 42
Armenia asiatica Leven, 1 07 Artinskia Karpinsky, 42, 90, 97, 109
artiensis (Gruenewaldt), 105 Asperlinus asperulus (Waagen), 1 1 0 Asyrinx Hudson & Sudbury, Table 3 8 , p . 130 A tomodesma Beyrich, 49, 77, 78, 149, 1 75;
Table 2b, p. xv; Table 37, p. 128; Figs. 12_:_15, pp. 50-51
costata Popov, 149 exaratum Beyrich, 1 24 trabeculum Waterhouse, 166 cf. trabeculum Waterhouse, 153 trechmanni (Marwick), 153, 156; Table 52, p.
1 82 undulatum Rothpletz, 156 variabilis Wanner, 149 wo()di Waterhouse, Table 52, p. 182
A ttenuatella Stehli, 1 1 , 14, 44, 46, 48, 49, 52, 55, 57, 60, 61 , 66, 69, 72, 77, 79, 85, 87, 90, 1 29, 144, 145, 152, 176, 179; Table 4, p. 12; Table 22, p. 73; Table 49, p. 177; Table 50, p. 1 78; Figs. 12-15, pp. 50, 5 1
attenuata (Cloud), 1 5 3 australis Armstrong & Brown, 95 convexa Waterhouse, 153 frechi (Schellwien), 44, 66, 69; Table 21, p.
70 incurvata. Waterhouse, 1 53, 156, 157; Table
52, p. 1 82 omolonensis Zavodowsky, 5 7 stringocephaloides (Likharev & Chemyshev),
1 29 Aulosteges Helmersen, 105, 1 10, 1 29, 141,
149; Table 51, p. 1 80 ingens Hosking, 99 1 02; Table 50, p. 178 uralicus Chernyshev, 79
Bakevillea King, Table 48, p. 70 Balakhonia insinuata (Girty), Table 13, p. 47 Bamyaniceras bomemani (Toumansky), 97 Bathymyonia nevadensis (Meek), 1 35
ster/itamakensis (Stepanov), 98 Bellerophon Montfort, 84
vaceki Bittner, 1 74 Bernhardites Shevyrev, Table 46, p. 1 59 Bitaunioceras Shimizu & Obata, 99, 1 18, 1 19
krotowi (Dewing), 105 Blasispirifer blasii (Verneuil), 1 29 Boloria Grunt, 19, 84 Bore/is princeps Ehren burg, 20 Brachythyrina Frederiks, 82, 129, 1 3 2
carnicus (Schellwien), 69 rectangulus (Kutorga), 92
Brachythyris M'Coy, 46, 52, 61, 134 ufensis Chernyshev, 46, 55, 5 7
Brasilioproductus Mendes, 7 5 Buxtonia Thomas, 46

Camarophoria (sic) ex. gr. pentameroides (Chernyshev), Table 1 3, p. 4 7
Camarophoriina antisella (Broili), 153 Camerisma murgabica Grunt, 82
sella (Kutorga), 79, 80, 82 Cancellina Hayden, 109, 1 1 0, 131� Table 18, p.
62; Table 23, p. 81; Table 38, p. 130; Table 43, p. 1 50; Table 5 1 , p. 180
duktevitchi Leven, 107 pam ira Leven, 107 primigena (Hayden), 106, 107, 1 34
Cancrinella Frederiks, 46, 57, 61 , 82, 96, 99, 107, 1 16, 1 1 8, 122, 124, 129, 137, 144; Fig. 28, p. 121
cancriniformis (Chernyshev), 57, 61 , 80, 84 janischewskiana Stepanov, 57 koninckiana (Keyserling), 103 lyoni (Prendergast), Table 38, p. 130 magniplica Campbell, 124, 127 phosphatica (Girty), 1 1 8, 1 19
Cancrinelloides Ustritsky, 1 1 , 105, 109, 1 13, 129, 135 ; Table 2b, p. xv; Table 22, p. 73 ; Table 51, p. 180
alferovii (Miloradovich), 105 Capillonia brevisulcus (Waterhouse), 156
toulai (Dunbar), 1 16 Changhsingoceras Chao, 31 , 169, 183; Table 52,
p. 182; Figs.8, 9, p. 32 Chaoiella Frederiks, 46, 66, 96, 99
chiticunensis (Diener), 144, 153 margaritatus (Mansuy), 1 10, 132, 145 sumatrensis cambodgensis (Mansuy), Table
42, p. 147 Chenia Sheng, Table 1 8, p. 62 Chianella Waterhouse, 141
chianensis (Chao), 106, 109; Table 51, p. 1 80 Chivatschella Zavodowsky, 149 Choanoproductus Termier et al. , 146
paviei (Mansuy), 146 Chonetella Waagen, 161
nasuta Waagen, 131, 145, 153 "Chonetes" brama Frederiks, 96, 105 , 106
paraconvexa Maslennikov, 80 solida Krotow, 79 squama Waagen, 141
Chonetina Krotow, 11, 96, 105, 106, 1 29, 141, 166 ; Table 5 , p. 13
artiensis (Krotow), 96, 105 superba Gobbett, 135
Chonetinella Ramsbottom, 103, 109, 1 1 8, 132; Table 5, p. 13
Chonosteges Muir-Wood & Cooper, 1 22 Choristitella (= ? Cartorhium Cooper & Grant ?)
wynnei (Waagen), 1 34, 145 Choristites Fischer, 46, 52, 79, 80, 97, 1 10, 132
anikeevi Einor, Table 1 3, p. 47 baschkirica Yanishevsky, 64 frischi (Schwellwien), 69 nikitini Chernyshev, 64
Chusenella Lee, 80, 109, 1 3 1 , 132, 146, 1 5 1; Table 1 8, p. 62; Table 45, p. 158; Table 51, p. 1 80
Oaraia Bittner, 19, 1 72, 1 74, 175; Table 48, p. 1 70; Table 52, p. 1 82
Systematic Index for Genera and Species 223
stachei Bittner, Fig. 36, p. 171 Qeiothyridina Buckman, 11 , 96, 99, 1 10, 1 29,
152 gerardi (Diener), 156 kuiechowensis Sheng, Table 48, p. 1 70 laqueata Waterhouse, 157 pectinifera (Sowerby), 103, 1 1 3; Table 51, p.
1 80 semiovalis (Waagen), 99
Codonofusiel/a Dunbar & Skinner, 1 5 2, 1 5 7, 163, 165, 169; Table 2a, p. xv; Table 1 8 p . 62; Table 23, p . 8 1 ; Table 4 5 , p . 158; Table 46, p. 159; Table 48, p. 1 70; Table 52, p. 1 82; Fig. 34, p. 167
? aff. tunetana (Douville), 134 Colaniella Likharev, Table 23, p. 81; Table 48,
p. 1 70 Comelicania Frech, 19, 39, 161 , 168, 183; Table
2a, p. xv; Table 45, p. 158; Table 46, p. 159; Table 52, p. 1 82
Composita Brown, 1 10, 122; Fig. 28, p. 121 ovata Mather, 72
Compressoproductus Muir-Wood & Cooper, 141, 145, 149, 152, 157; Table 47, p. 164
mongolicus (Diener), 148 mongolicus subcircularis (Reed), 141
Conularia Miller (= Paraconularia Sinclair?), 92; Table 2a, p. xv; Table 19, p. 65 ; Table 28, p. 100
Costalosia Waterhouse & Shah, 1 34 argentea Waterhouse & Shah, 107 bifurcata Waterhouse & Shah, 107, 1 10
Costiferina Muir-Wood & Cooper, 99, 1 74 alatus Waterhouse, 143 aratUs (Waagen), 141 indica (Waagen), 132, 152 redaca (Reed), 131 spiralis (Waagen), 1 24, 145
Craspedalosia pulchella Dunbar, 1 1 6 Cribrogenerina permica Lange, 1 57 Crurithyris George, 45, 46, 61 , 64, 69, 75, 85,
109, 1 10, 1 1 8, 1 1 9, 1 22, 152, 161, 165, 166, 168, 1 75; Table 51, p. 1 80; Table 52, p. 1 82
acutirostris (Krotow), 105 arcuata (Girty), 1 1 9 clannyana (King), 105, 1 1 3 speciosa Wang, 1 6 1 , 169 (? Orbicoelia) extima Grant, 161 (Orbicoelia) tschernyschewi Likharev, 157
"Crurithyris" sp. 72 Cryptacanthia White & St.John,14 1 Cryptospirifer Grabau, 148 Cyrtella Frederiks, 98
kulikiana Frederiks, 96 Cyclolobus Waagen, 5, 18, 19, 29, 31, 34, 140,
141 , 143, 145, 169, 1 81; Table 1 1 , p. 35; Table 46, p. 159; Table 52, p. 1 82; Figs. 8, 9, p. 32; Fig. 32, p. 160
haydeni Diener, 143 kraffti Diener, 143 persulcatus Rothpletz, 153 walkeri Diener, 143, 144, 153

224 Systematic Index for Genera and Species
Cyrolexis superstes (Verneuil), 79 Cyrtospirifer khauraulakhensis Frederiks, 80
Daixites Ruzencev, 42 Daraelites meeki Gemmellaro, 107 Darvasites Mikluko-Maklay, 82; Table 23, p. 81 Daubichites Popov, 1 09, 1 1 6, 1 24, 1 31 ; Table
5 1 , p. 1 80 fortieri (Harker), 1 1 6, 1 2� goochi (Teichert), 1 24, 1 26 shoutangensis (Chao), 1 09
Deltopecten Etheridge, Table 49, p. 177 Derbyella Grabau, 163 Derbyia Waagen, 75, 79, 80, 1 03, 1 3 1 , 1 34, 141;
Table 47, p. 1 64 altestriata Waagen, 148, 152 grandis Waagen, 82, 153 cf grandis Waagen, 97, 148 hemisphaerica Waagen, 141 magna Branson, 1 1 9 regularis Waagen, 96, 97, 99
Dictyloclostus Muir-Wood, 1 22; Fig. 28, p. 1 2 1 byrangi (Einar), Table 1 3 , p . 47
"Dictyoclostid" celsus Reed. 141 Dielasma King, 1 1 , 1 1 0
elongatum (Schlotheim) , 103, 1 1 3 ; Table 5 1 ; p . 1 80
cf. plica not Kutorga of Reed (=
Attenuatella), 6 1 Die/asmina Waagen, 1 4 1 Dip/anus Stehli, 45 Discinia Lamarck, 92 Dorashamia Sarytcheva, 1 57 Duartea Mendes, 46, 49, 52, 75; Figs. 1 2 - 1 5 ,
pp. 50, 5 1 Dunbarula Ciry, 165 Durvilleoceras Waterhouse, 169, 1 7 2; Table 2b,
p. XV woodmani Waterhouse, 163, 1 66; Table 6,
p. 1 7; Table 37, p. 1 28; Table 52, p.182
Dyoros Stehli, 1 1 , 1 19 ; Table 5 , p . 1 3 Dzhulfites Shevyrev, 1 68; Table 46, p . 159 Dzhulfoceras Ruzencev, Table 45 , p. 158
Echinalosia Waterhouse, 1 05, 1 27 delicatula Ustritsky & Chernyak, 79 indica (Waagen), 1 34 kalikotei Waterhouse, 143, 144, 149; Fig. 30,
p. 142maxwelli (Waterhouse), 1 06, 126, 1 27; Table
37, p. 1 28; Table 5 1 , p. 1 80 minima (Maxwell), 1 37 ova/is (Maxwell) , 1 3 7 ; Table 2b, p. xv; Table
6, p. 1 7; Table 37, p. 1 28 preovalis (Maxwell), 95; Table 50, p. 1 7 8 prideri (Coleman), 99, 1 02; Table 3 7 , p. 128;
Table 50, p. 178 Echinauris Muir-Wood & Cooper, 45, 1 22, 1 35
khmerianus (Mansuy), 146 opun tia (Waagen), 157
Edriosteges Muir-Wood & Cooper, 1 22, 1 5 1 medlicottianus (Waagen), 1 09 poyangensis (Kayser), 165
Eliva Frederiks, Fig. 6, p. 15
Elivina Frederiks, 1 07, 1 32, 1 52; Fig. 6, p. 15 (= Blasispirifer) blasii (Verneuil), Table 51,
p. 80tibetana (Diener), 1 32, 144 , 145, 146, 153;
Table 52, p. 1 82 Ellisonia teicherti Sweet, 175; Table 8, p. 28 Enteletes Fischer, 5 3 , 60, 82, 1 34, 141, 149,
1 5 3 , 168; Table 47, p. 164 conjunctus Reed, 107 dzhagrensis Sokolskaya, 163 laevissimus Waagen, 141
Enteletina latisinuatus Waagen, 141 Eoasianites Maximova, 87, 90 Eoaraxoceras Spinosa et al. , 161; Table 52, p.
1 82 ruzhencevi Spinosa et al. , 166
Eoglyphioceras meneghinii (Gemmellaro), 107 Eomarginifera migae (Chernyak), Table 1 3, p. 47 Eoparafusulina (= Alaskanella) yukonensis
(Skinner), 87, 98; Table 50, p. 178 Eoshumardites Popov, 5 5
artigensis Popov, 4 6 Eoverbeekina Lee, Table 18 , p. 62 Epadrianites Schindewolf, 1 5 2 Episagiceras Noetling, 1 5 2 Epithalassoceras ruzencevi Miller & Furnish, 137 Eridmatus Branson, Fig. 6, p . 1 5 Etheripecten striatura Waterhouse, 149 · volucer (Likharev), 149Eumedlicottia Spath, 99
burckhardti (Bose), 1 22 Eumicrotis Meek, 1 74 Eurydesma Morris, 45, 46, 49, 5 2, 77, 95, 1 26,
169, 1 72 , 179, 1 8 3 ; Table 2a, p. xv; Table 1 9, p . 65; Table 28, p . 1 00; Table 49, p. 1 77; Table 52, p. 1 82
inflatum (author not stated in Sheng & Lee), 169
mytiloides Reed, 77; Table 38, p . 1 30
Filiconcha Dear, 1 57; Table 52, p. 1 82 Fimbrinia Cooper, 45, 46, 48
cristato-tuberculata (Kozlowski), Table 13, p. 47
F1etcherithyris Campbell, 141, 1 75 Fredericksia Paeckelmann, 14 1 Fusispirifer Waterhouse, 99
nitiensis (Diener), 143, 145 Fusulina Fischer Von Waldheirn, 107 Fusulinaella Moeller, 66
Gaetanoceras martini (Haniel), 82 timorensis (Haniel) , 82, 1 24
Gangamopteris M'Coy, Table 9, p. 30 Gastrioceras sp. ind. es aff. marianum Verneuil,
143 Gefonia Likharev, 141 Gemmulicosta Waterhouse, 46 Geyerella Schellwien, 149; Table 47, p. 164 Gibbospirifer Waterhouse, 46 Glaphyrites Ruzencev, 42, 53 , 5 5 , 77; Table 49,
p. 1 77; Fig. 14, p. 5 1 Glassoceras Ruzencev, 1 22; Figs. 8 , 9 , p . 32
brasoni (Miller & Cline), 1 1 9 Glendella Runnegar, 1 26; Table 5 1 , p . 1 80

Globiella rossiae Sestini, 82 Glossopteris Brongniart, 45, 46, 52, 143; Table
9, p. 30 Glyptophiceras Spath, 163; Table 48, p. 170 Godthaabites Frebold, 31, 1 16, 169� Table 51,
p. 180; Fig. 9, p. 32 Gondolella Stauffer & Plummer, 26; Table 7, p.
27 bisselli, Table 7, p. 27 carinata subcarinata (Sweet), 29; Table 8, p.
28
Systematic Index for Genera and Species 225
22, p. 73 cheraskovi Kashirtsev, Table 13, p. 47 verchoyanicus (Frederiks), 57, 59; Table 13,
p. 47 Jimiceps Frech, 19, 39, 168, 1 83; Table 45, p.
158; Table 52, p. 1 82 Jisuina Grabau, 134 Juresania Frederiks, 49, 79, 84, 1 1 8, 141 Juresanites Maximova, 42, 53; Table 49, p. 1 77;
idahoensis Youngquist et al. , 26, 1 09; Table 7,
Figs. 1 2-15, pp. 50, 51 jacksoni (Etheridge), 92 kazakhorum Ruzencev, 78
p. 27� Table 8, p. 28 leveni Kozur et al. , 29; Table 8, p. 28 nankinensis (= Neogondolella), 109 orienta/is (Barskov & Koroleva), 29; Table 8,
p. 28 (= Neogondolella) rosenkrantzi Bender &
Stoppel, 26� Table 7, p. 27; Table 8, p. 28
(= Neogondolella?) serrata Clark & Ethington, Table 8, p. 28
Gonioloboceras Hyatt, 42 Grandaurispina Muir-Wood & Cooper, 137 Graliiiispora Hoffmeister et al. , 45 Grumantia (or Arctitreta) kempei (Andersson),
116 ("Streptorhynchus") pelicanensis (Fletcher),
137; Table 51 , p. 1 80 Gyronites Waagen, 19
Haydenella Reed, 83, 131 , 1 32, 161; Table 45, p. 158; Table 46, p. 1 59
kiangsiensis (Kayser), 146, 148 tumida (Waagen), 151, 163
Helicoprion Karpinsky, 124 Hemiptychina Waagen, Table 47, p. 164 Hercosestria Cooper & Grant, 122 Hercosia Cooper & Grant, 122 Heteralasma Girty, 134, 141, 152 Heteropecten Kegel, 77 Horridonia Chao, 87, 97
borealis (Haughton), 103, 105 horridus (Sowerby), 1 1 3 pseudotimanicus Gerassimov, 103
Hustedia Hall & Clarke, 60, 75, 1 1 0, 141 grandicosta (Davidson), 148 indica Waagen, 148
Hyattoceras GemmeUaro, 143 n. sp. ex. aff cumminsi White, 143
Hypophiceras martini Triimpy, 172 triviale (Spath), 172
ldiognathodus ellisoni Clark & Behnken, 26; Table 7, p. 27
lnstitella Cooper, Table 50, p. 178 /eonardensis (King), 98
lsarcicella (= 'Anchignathodis') isarcicus (Huckriede), 29; Table 8, p. 28
lsogramma Meek & Worthen, 60, 79 paotechowensis Grabau, 82
Jakutoproductus Kashirtsev, 46, 48, 55, 57, 60, 79, 80, 87, 96, 105 ; Table 2b, p. xv; Table
Kahlerina Kochansky-Devide & Ramovs, 148; Table 1 8, p. 62
Karavankina Ramovs, 69, 82, 84 Kargalites Ruzencev, 72
(Kargalites), Ruzencev, 42 Kiangsiella Grabau & Chao, 99, 141 Kingoceras Miller, 152, 161; Table 35, p. 1 23
kingi Miller, 166 Kochiproductus Dunbar, 1 1 , 45, 46, 48, 49, 5 2,
53, 55, 57, 72, 79, 85, 96, 1 19, 149, 1 5 1 , 176; Table 2b, p . 5; Table 22, p. 73; Table 47, p. 1 64; Table 49, p. 1 77; Figs. 1 2-15, pp. 50, 5 1
(sic) levinsonlessingi Zavadowsky, 57 porrectus (Ku.torga), 48, 57, 149 transversus Cooper, 72
Kolymia Likharev, 149 inoceramiformis Likharev, 149
Kozlowskia Frederiks, 46, 52 "Krafftoceras" (= Cyclolobus) kraffti Diener, 143 Krotovia Frederiks, 46, 5 2, 69, 1 10, 144, 146;
Table 50, p. 178; Fig. 38, p. 173 burmana Diener, 1 1 0 janus Huang, 109, 148 jisuensiformis Sarytcheva, 163 licharewi Frebold, 1 1 6 nystianus (De Koninck), 146; Table 42, p.
147 Kufengoceras Chao, 109, 148; Figs. 8, 9, p. 32 Kutorginella Ivanova, 55, 72, 131; Table 49,"p.
177
Lamnimargus himalayensis (Diener), 82, 143, 144, 145, 146, 1 74; Fig. 30, p. 142
Lantschichites Toumanskaya, 148, 152; Table 18, p. 62
Leel/a Dunbar & Skinner, 152 Leiophyllites Diener, Table 48, p. 1 70 Leiorhynchoidea Cloud, 122, 137, 153, 166 ;
Fig. 28, p. 121 Lepidolina Lee, 16, 18 , 140, 148, 165, 1 8 1 ;
Table 39, p . 1 3 3 ; Table 52, p . 1 82 kumaensis Kanmera, 148, 149, 165; Table
48, p. l 70 multiseptata (Deprat), 134 , 146, 148, 157
165; Table 42, p. 147: Table 48, p. 170
multiseptata gigantea (Gubler), 146 toriyamai Kanmera, 148, 149; Table 20, p. 67 ussurica (Dutkevich), 149

226 Systematic Index for Genera and Species
Leptodus Kayser, 107, 1 18, 132, 134, 135, 145, 151, 153; Table 42, p. 147; Table 48, p. 170
nobilis (Waagen), 1 3 1 , 146, 149, 1 5 1 , 156;Table 4 7, p. 164
richthofeni Kayser, 148, 149, 151 , 157; Table 52, p. 1 82
Lethamia ligu"itus Waterhouse, 140 Levicamera Grabau, 48 Levipustula Maxwell, 45, 57, 77 Licharewia Einor, 11, 55, 84, 99, 106, 1 13, 1 1 8,
'126, 1 29, 1 35, 179; Table 5 1 , p. 1 80 carnicus (Schellwien), 82 clarkei (Koninck), 1 27 rugulatus (Kutorga), 105 stuckenbeyti (Nechaev), 105
Liebea Waagen, 146; Table 42, p. 147 Limbella Stehli, 45 Linoproductus Chao, 46, 53, 57, 61, 66, 79, 96,
98, 1 32, 144; Table 42, p. 147; Table 47, p. 1 64
aagardiformis Seminova, Table 13, p. 47 achunowensis (Stepanov), 55; Table 13, p. 47 cora (D'Orbigny), 61 coralineatus Ivanov, 69 dorotheevi Frederiks, 72
Linoproductus popowi Zavodowsky, 5 7 lineatus (Waagen), 1 3 1 , 145
Liosotella Cooper, 1 16, 153 Lissochonetes Dunbar & Condra, 11, 99, 107,
1 10, 1 18, 1 19, 1 29, 134, 135, 1 74; Table 2b, p. xv; Table 5 , p. 1 3 ; Table 22, p. 73; Table 5 1 , p . 1 80
bipartita (Waagen), 148 morahensis (Waagen), 148 ostiolatus (Girty), 119 (= Capillonill) toulai Dunbar, 1 1 6 yarrolensis Maxwell, 95
Lilinthiceras Diener, 143 ?Lyttonia (= ?Leptodus) Waagen, 143, 148
Maklaya Kanmera & Toriyama, Table 5 1 , p. 1 80 cutlaensis (Leven), 106 pamirica (Leven), 106 aff sethaputi Kanmera & Toriyama, 107
Maorielasma Waterhouse, 14 Marathonites Bose, 42 (Almites) Toumanskaya, 42 Marginifera Waagen, 97, 99, 131, 132, 141, 1 5 1
banphotensis Yanagida, 146 septentrionalis Chemyshev, 103 typica Waagen, 143, 145, 152; Table 38, p.
130 Martinill M'Coy, 45, 46, 60, 61, 66, 79, 80, 82,
85, 87, 96, 107, 1 10, 122, 1 3 1 , 134, 141 , 145, 146, 148, 1 5 1 , 1 52, 161 , 163, 166, 168; Table 47, p. 164; Table 50, p. 1 78; Table 52, p. 1 82
adentata Waterhouse, Table 37, p. 1 28 elegans Diener, 148 karawanica Volgin, 69 mongolica Grabau, 163, 166; Table 47, p. 164 semiglobosa Gemmellaro, Table 13, p. 47 semiplana Waagen, 132, 134
Martiniopsis Waagen, 57, 61 , 64, 84, 99, 107, 146, 156
inflata Waagen, 1 5 1 ; Table 52, p. 182 latouchei Diener, 1 1 0 orienta/is Chemyshev, 79, 84 talishaoensis Reed, 1 3 2 woodi Waterhouse, 156 ; Table 2b, p . xv;
Table 6, p. 1 7 ; Table 37, p. 128; Table 52, p. 1 82
Medlicottill Waagen, 78, 79, 82, 87, 92, 152; Table 49, p. 177 ; Table 50, p. 178 aff. artiensis:timorensis Haniel, 80, 81, 97 costellifera Miller & Furnish , 98 orbignyana (Vemeuil), 96
Meekella White & St John, 5 3, 60, 82, 131, 137 punjabica Reed, 141
Meekoceras Hyatt, Table 4 8, p. 1 70 Megasteges dalhousei (Davidson), 141, 145, 148,
153 ; Table 52, p. 182 nepalensis Waterhouse, 144
Mentzelill Quenstedt, 107, 1 34 Merrillina arcucristatis (Clark & Behnken), 26;
fable 7, p. 27; Table 8, p. 28 divergens (Bender & Stoppel), 26, 29; Table
7, p. 27; Table 8, p. 28 galeatus (Bender & Stoppel) , 26; Table 7, p. 27;
Table 8 , p. 28 Metalegoceras Hyatt, 78, 80, 84 , 87, 90, 92, 95,
97; Table 49, p. 177 Metaperrinites Ruzencev, 92 Metapronorites timorites Haniel, 82 Mexicoceras Ruzencev, 1 09, 131, 148; Table 51,
p. 1 80; Figs. 8 , 9, p. 32Micraphelia Cooper & Grant, 146 Misellina Schenck & Thompson, 80, 82, 97, 109;
Table 1 8, p. 62; Table 23, p. 8 1 claudille (Deprat) , 85 , 97, 106 ; Table 43, p.
150; Table 50, p. 178 Monodiexodina Sosnina, 82, 85 , 90, 99, 1 18,
148; Table 50, p. 178 matsubaishi (Fujimoto), 132, 1 85
Monticulifera Muir-Wood & Cooper, 105, 134, 146, 157; Table 5 2, p. 1 82
impressa Waterhouse, Table 37, p. 128 (= Choanoproductus) paviei (Mansuy), 146;
Table 42, p. 14 7 sinensis (Frech), 109, 146
Mufushanella mufushanensis Lee, 85 Muirwoodia Licharev, 14, 1 19, 122, 149; Table
47, p. 164 artiensis (Chemyshev), 96; Table 50, p. 178 cf artiensis (Chernyshev), 98 mammatus (Keyserling), 1 16 multistrillta (Meek), 135 pseudoartiensis (Stuckenberg), 49
Nagatoella Thompson, Table 18 , p. 62 Nankinella Lee, 66, 80, 82, 84, 85 , 97, 1 3 1 ,
132, 15 1 ; Table 18 , p. 62; Table 45, p. 158; Table 48, p. 1 70
caucasica Dutkevich, 97 orbicularia Lee, 85, 97 orienta/is Mikluko-Maklay, 1 3 1
Neochonetes Muir-Wood, 1 1 , 6 6 , 84, 92, 107, 110, 1 13, 1 16, 1 18 , 1 19, 1 22, 127, 1 29, 132, 137 , 145, 148, 1 5 3 ; Table 5, p. 13 ; Table 22, p. 73; Table 5 1 , p. 1 80
beatusi Waterhouse, 140

Systematic Index for Genera and Species 227
deremfii (Douglas), 107 variolata (Vemeuil), 103
Neocrimites Ruzencev, 95, 97, 109, 126, 137 aff fredericbi (Emelianstev), 99 meridionalis Teichert & Fletcher, 126 newelli (Miller & Furnish), 99
Neodimorphoceras Schmidt, 42 Neo[umlinella Deprat, 1 10
lantenoisi Deprat, 106, 107; Table 42, p. 147 Neogeoceras Ruzencev, Table 51, p. 180
macnairi Nassichuk, 135 trautscholdi (Gemmellaro), 107
Noglaphyrites Ruzencev, 48, 53, 55, 57, 80 Neogondolella rosenkrantzi (Bender & Stoppel) ,
26, 116 divergem (Bender & Stoppel), 1 13 smata (Clark & Ethington), 26; Table 7, p.
27; Table 8, p. 28 Neomisellina Sheng, 148; Table 18, p. 62
lepida Schwager, Table 42, p. 147 1/eoplicatifera" Chao et al. , 124
huangi (Ustritsky), 107, 146 Neopronorites Ruzencev, 57, 72
permicus Chernow, 1 05 skvorzovi Chernow, 105
Neoshwagerina Yabe, 39, 1 09, 131 , 132, 145, 146, 165 ; Table 18, p. 62; Table 48, p. 170
craticulifera (Schwager) , 106, 107, 132, 134, 179; Table 20, p. 67 ; Table 38, p. 130; Table 39, p. 133; Table ·5 1 , p. 180
douvillei Ozawa, 132 globosa (= Yabeina) Yabe, Table 42, p. 147 margaritae Deprat, 131 , 132, 15� , 157; Table
23, p. 81 ; Table 42, p. 147; Table 48, p. 170; Table 51, p. 180
megasphaerica Deprat, 132 (= Minoella ) schuberti Kochanzky-Devide',
Table 23, p. 8 1 Simplex Ozawa, l06, 107, 109, 110; Table 20,
p. 67; Table 23, p. 81 ; Table 5 1 , p. 180
Neoschizodus Giebel, Table.48, p. 170 Neoshumardites Ruzencev, 78
cf sakmarae (Ruzencev), 87 triceps hyperboreus Ruzencev, 80
Neospathodus arcucristatus Clark & Behnken, 26
Neospirifer Frederiks, 99, 1 10, 122, 126, 127, 149, 156
bambadhuriensis (Diener), 129, 149 fasciger (Keyserling), 1 19 marcoui (Waagen), 99 mooMkhailensis (Davidson), 15 1 , 153 profasciger Likharev, 106 pseudocameratus (Girty), .1 19, 135 ravana (Diener), 144, 156 striato-paradoxus (Toula), 1 16 striJZto·plicatus (Gobbett), 1 16, 1 18, 126 subfasciger (Likharev), 57
Neostreptognathodus Clark, 26 sulcoplicatus (Youngquist et al.) , 26; Table
7 , p. 27 ; Table 8 , p. 28 Neouddenites adriJZnova Ruzencev, 96, 98
caurus Nassichuk et al. , 98 Netschajewia Likharev, 129 Noeggerathiopsis Feistmantel, 46, 48 Nomismoceras smithi Diener, 143 Norella Bittner, 19 Notospirifer Harrington, 95 , 127; Table 2b, p. xv;
Table 49, p. 177 spinosa Waterhouse & Vella, Table 37, p. 128
Notostrophia Waterhouse, Table 2b, p. xv homeri Waterhouse, Table 3 7, p. 128 zealandicus Waterhouse, Table 37, p. 128
Notothyris Waagen, 60, 84, 134, 141 , 146, 151 , 163
lenticularis Waagen, 141 subvesicularis (Davidson), 141
Nuculana nogamii, Table 48, p. 170
Odontospirifer Dunbar, 60 mirahilis Dunbar, 1 16
Oldhamina Waagen, 134, 152; Table 45, p. 158; Table 46, p. 159
Ombonia Caneva, 157 Omphalotrochus Meek, 45, 49, 57; Figs. 12, 13 ,
15 , pp. S0, 5 1 Ophiceras Griesbach, 1 9 , 163, 172, 174; Fig. 38,
p .l73 commune Spath, 29, 192; Fig. 38, p. 173 connectens Schindewolf, 163
Orbicoelia Waterhouse & Piyasin, 1 07, 1 10, 165 extima (Grant), 161 tschernyschewei (Likharev), 157, 161
Orthis not Dalman, 1 34 Orthotichia dorashamensis Sokolskaya, 165
dzhulfensis Sokolskaya, 151 Orthotetella King, 45 Orthotetes Fischer De Waldheim, 69, 132, 134 Orthotetina Schellwien, 146, 151 , 152, 161 ;
Table 47, p. 164 altus Hamlet, 153 arakeljani Sokolskaya, 151 cf arakeljani Sokolskaya, 161
Orthotichia Hall & Clarke, 46 , 53, 55, 72, 75 , 79, 85 , 1 10, 1 22, 146, 165, 168; Table 2b, p. xv; Table 22, p. 73; Table 49, p. 177
derbyi Waagen, 141 dorashamensis Sokolskaya, 165 indica (Waagen), 107, 109, 1 10, 141
Orulgania Frederiks, 48, 57, 84 tukulaensis (Kashirtsev), Table 13 , p. 4 7
Otoceras Griesbach, 19, 34, 143 , 161 , 172, 174, 175, 183; Table 52, p. 182; Fig. 26, p. 1 15 ; Fig. 30, p. 142; Fig. 35, p. 171 ; Fig. 38, p. 173
boreale Spath, 172 concavum Tozer, 172
Overtoniid nystianus De Koninck, 146; Table 42, p. 147
Overtonfina Grunt, 122 Overtoniina (or !Veoplicatifera) huangi
(Ustritsky), 109, 124, 148 Owenoceras Miller & Furnish, 46, 55, 57
orulganense Popov, 46 Ozawainella Thompson, 66, 1 10

228 Sys tema tic Index for Genera and Species
Pachyphloia paraovata maxima Mikluko-Maklay, 157
Paeckelmane/la (= better Paeckelmanne/la?) Likharev, 46, 55, 57, 87 , 1 3 1 ; Table 5 1 , p . 1 80
expansa (Chernyshev), 103, 105 Pa/aeofusulina Deprat, 163, 165, 169; Table 1 8,
p. 62; Table 23, p. 8 1 ; Table 5 2, p. 1 82 ; Fig. 3 4 , p . 1 6 7
Parafusu/ina Dunbar & Skinner, 7 8 , 80, 82, 85 , 90, 92 97, 98, 99, 106, 109, 1 1 8, 1 22, 1 3 1 , 145, 146, 148; Table 1 1 , p. 35 ; Table 1 8, p. 62; Table 23, p. 8 1 ; Table 35, p. 1 23 ; Table 45 , p. 158; Table 50, p . 178; Fig. 28, p. 1 2 1
antimoniensis Dunbar, 1 19 de/iciasensis Dunbar & Skinner, 137 durhami Thompson & Miller, 98 ferganica Miklukho-Maklay, 80, 84, 97 japonica Gumbel, 80 kaerimizensis Ozawa, Table 20, p. 67 padangense (Lange) , 1 1 0 rich thofeni (Schwager) , Table 42, p. 14 7 sellardsi Dunbar & Skinner, 99, 1 19 solidissima (Rauser) , 96 ; Table 50, p. 178 spissisepta Ross, 98 aff. yabei Hanzawa, 97
Paragastrioceras Chernow, 42, 48, 53, 57 , 80, 95, 124 ; Table 49, p. 177 ; Figs. 1 3 - 1 5 , pp. 50, 5 1
aff jossae (Verneuil) , 87 jossae subtrapezoidalis Maximov, 79 kunguriensis Mirsk, 103 (or Altudoceras) roadense Bose, 106, 109
Parakeyser/ingina Chernyshev, 80 Paranorella Cloud, 137, 153 Paranorites konincki Gemmellaro, 107 Parapronorites Ruzencev, Table 38, p . 130 Paraschistoceras Plummer & Scott, 42 Paraschwagerina inflata (Chan) , 64
pseudomira Mikluko-Maklay, 6 1 Paratirolites Stoyanov, 2 9 , 1 6 8 , 183 ; Table 2a,
p. xv; Table 45, p. 158; Table 46, p. 159; Table 52, p. 182
Parayakutoceras Popov, 46 Parenteletes King, 45
ruzhencevi Sokolskays, 163 Penicu/auris Muir-Wood & Cooper, 122
bassi (McKee), 1 1 9 Peritrochia Girty, 152 Permophricodothyris Pavlova, 163
grandis (Chao), 146 Permospiri[er Kulikov, 1 1 , 84, 129 Permundaria Nakamura et a/. , 1 3 1
sisophonensis Nakamura et a/. , 1 3 1 ; Table 38, p. 1 30
Perrinites Bose, 82, 87, 99; Table 1 1 , p. 35 ; Table 35, p. 123
hilli (Smith), 1 22 hilli afghana Termier & Termier, 97 vidriensis Bose, 99
Peruvispira Chronic, 77 de/icatula Chronic, 75
Phisonites Shevyrev, 29, 39, 1 6 1 , 168, 1 8 3 ;
Table 4 5 , p. 1 5 8 ; Table 4 6 , p. 1 5 9 ; Table 52, p. l 82
Phricodothyris George, 53, 79, 145, 1 46 , 148 indica (Waagen), 1 5 l ; Table 47, p. l64 waageni, 85
Platyconcha grandis Waterhouse, 144 Platyteichum Campbell, 166 Plekonella southlandensis (Fletcher), 140
acuta Campbell, 140 multicostata Waterhouse, 156; Table 2b, p. xv;
Table 6, p. 1 7 ; Table 37, p. 128 Pleuronodoceras, 168 "Plicatifera " (= Overtoniina, or Neoplicatifera)
huangi Ustritsky, 109, 1 22 minor Schellwien, 1 24
Plicatocyrtia zitteli (Schellwien), 66, 69 Plicatoderbyia Thomas, 141 Plicototorynifer Abramov & Solomina, 46
snjatkovi Abramov, Table 1 3 , p. 47 Polydiexodina Dunbar & Skinner, 1 10, 152
capitanensis Dunbar, 153 elongata (Shumard), 107, 1 1 0; Table 51, p.
1 80 mexicana Dunbar, 166; Table 5 2, p. 1 82 persica Kahler, 107, 1 5 1 shumardi Dunbar & Skinner, 1 5 3 (= Skinnerina Ross?), 134
Popanoceras Hyatt, 80, 90, 97, 107 , 1 16 , 137, 152, 1 74
lahuseni Karpinsky, 97 cf sobolewskyanum (V�rneuil), 1 16 tumarense Ruzencev, 96
Potonieisporites Bhardwaj , 45 Praehorridonia Ustritsky & Chernyak, 46 Praeparafusulina lutugini (Schellwien), 85 , 95,
98; Table 50, p. 1 7 8 Praesumatrina Toumanskaya, 1 09
neoschwagerinoides (Deprat), 107 schellwieni (Deprat) , 109
Procrassatella Yakovlev, 1 29 "Productella" (? Overtoniina) patula Hamlet
(not Girty) , 1 22 Properrinites.Elias, 42, 87, 90; Table 1 1 , p. 35
boesi (Plummer & Scott), 9 2 ; Table 50, p. 178
denhami (Miller & Furnish) , 92 Propinacoceras Gemmellaro, 80, 95, 126, 137,
australe Teichert, 1 24 beyrichi Gemmellaro , 107 bornemani Toumansky, 82, 97
Propopanoceras Toumansky , 85, 1 24 ; Table 49, p. 1 7 7
rhuzencevi Glenister & Furnish, 9 2 Prorich thofenia King, 1 3 7 , 149, 1 5 3 Prostacheoceras Ruzencev, 4 2 Prothalassoceras Bose, 8 7 Protopopanoceras Ruzencev, 4 2 , 5 3 , 7 8 ; Table 49,
p. l 77 Prototoceras Spath, 165 Prouddenites Miller, 42, 45 , 49 , 52; Figs. 1 2-
1 5 , pp. 50, 5 1 Pseudodielasma Brill, 166Pseudodoliolina Yabe & Hanzawa, 1 06 , 107, 109,
1 3 1

Systematic Index for Genera and Species 229
pseudolepida (Deprat), 1 10 ; Table 18, p. 62 Pseudofusulino Dunbar & Skinner.(American
Sen�), 18 , 20, 40, 42, 44, 45, 49, 52, 6 1 , 66, 69, 72, 75 , 82, 84, 103, 106, 145, 1 5 1 , 176 ; Table 18 , p . 62; Table 21 , p . 70; Table 23, p. 8 1 ; Table 45, p. 158; Table 49, p. 177; Table 50, p. 178; Figs. 12-15, pp. 50, 5 1
(Doixino) olpino alpino Pasini (= Rugosofusul-ina), 66
ombigua (Deprat), 85 chihsianensis (Lee), 85 , 97, 106 (Leeina) fusiformis (Schellwien), 80, 82 kroffti (Schellwien), 80, 85 , 97; Table 43 ,
p. 150(Doixino) longissimo olpina (Schellwien), 66 subcylindrico (Deprat), Table 42, p. 147 ( Leeino ) volida (Lee), 64
Pseudofusulina* (Soviet Sense), 20, 40, 52, 55 , 78
baschkirica Korzhenevsky, 79 devexa Rauser, 79 fecunda Shamov & Scherbovich, 53 , 79 firma Shamov, 53 aff. gregoria (Lee), 97 insignito Vissarionova, 95 korogosensis Rauser, 78 krotowi (Schellwien), 55 , 97 mokorovi Rauser, 96; Table 50, p. 178 poroconcessa Rauser, 78 procera Leven, 84 tschernyschewi (Schellwien), 85 verneuili (Moeller) , 79
!Pseudofusulina* (American & Russian Sense) vulgaris (Schellwien) , 5 3 , 64, 80, 82, 85 , 97 ex. gr. vulgaris (Schellwien), 82
Pseudofusulinella Thompson, 72, 87, 1 19 Pseudogostrioceras (= Daubichites) goochi
Teichert, 124 Pseudoholorites Yabe, 109 Pseudomonotis Beyrich, 129 , 1 72, 17 4 Pseudosogeceras Diener, Table 48, p. 170 Pseudoschwagerina Dunbar & Skinner, 20, 39,
40, 42, 44, 6 1 , 64 , 69, 75, 82, 85 , 90, 97 , 176 ; Table 1 1 , p. 35 ; Table 18, p. 62; Table 24, p. 83; Table 49, p. 177
alpino (Kahler & Kahler), 69; Table 21, p. 70 broggii (Roberts), 75 extenso Kahler & Kahler, 82 minatoi Kanmera, 66 morikawai lgo, Table 20, p. 67 parasphaerica (Chan), 64 princeps sensu Moeller, 6 1 , 66 pulchro Kahler & Kahler, 82 turbida Kahler & Kahler, 82 uddeni (Beede & Kniker), 20, 6 1 , 72, 75
Pseudostoffello Thompson, 66; Table 21 , p. 70 Pseudostephanites Chao, 168 Pseudosyringothyris inopinatus Solomina, 57 Pseudosyrinx (Kashirtsev) , 57, 106, 116 , 1 1 8,
124 ; Table 2b p. xv; Table 22, p. 73 ; Table 5 1 , p. 180
kolymensis Tolmachev, 96, 105 wimani Gobbett, 1 16
Pseudotirolites, 168, 169 Pterospirifer Dunbar, 79, 99, 1 3 1 , 144
alatus (Sowerby), 106, 1 18 ; Table 5 1 , p. 180 cordieri (Roberts) , 135 terechovi Zavadowsky, 5 7
Pugnoides Weller, 141 Punctocyrtello Plodowski, 84
nagmagensis (Bion), 77, 92; Table 50, p. 178 spinosa Plodowski, 82, 84
Punctospirifer puncta (Meek), 26 Purdonello Reed, 46, 96
conformis Reed, 141 limitaris Reed, 141 lunwalensis Reed, 131 scopulosus (Reed), 141 semiovalis Reed, 107
Pustula Thomas, 174
Quodrochonetes Ifanova, 1 1 9 Quasifusulina Chen, 6 1 , 6 6 ; Table 21 , p. 70
longissima (Moeller) , 66 ; Table 21 , p. 70 Quasifusulinoides Mikluko-Maklay, 66 ; Table 21 , p.
70 tenuissima (Schellwien), 66 ; Table 21, p. 70
Quinquenella glabra Waterhouse, 144
Rauserello errotico Dunbar, 153 Reedoconcho Kotlyar, 92; Table 50, p. 178
iranicus Sestini, 84 Reichelino Erk, 152, 163, 165; Table 18, p. 62;
Table 23, p. 8 1 ; Table 45, p. 158; Table 48, p. 170; Fig. 34, p. 167
changhsingensis Sheng & Chan, Table 48, p. 170
ctibroseptato Erk, 131 minuta Erk, 157
Reticulatia Muir-Wood & Cooper, 46, 79, 96 callytharrensis (Prendergast), 124 uralensis (Likharev), 98
Rhacopteris Schimper, 46 Rhipidomello Oehlert, 6 1 , 1 10
uralica Chernyshev, 79 vediensis Sokolskaya, 151
Rhynchopora King, 79 nikitini Chernyshev, 79, 80 variobilis Stuckenberg, 99
"Rhynchonella" wynnei Waagen, 141 Rhynchonellid aff. wynnei Waagen, 157 Richthofenia Kayser, 1 10; Table 47, p. 164
caucasica Likharev, 163 /owrenciano (De Koninck), 151
Robustoschwogerina schellwieni Yabe, 64 tumidiformis Mikluko-Maklay, 61
Rugaria Cooper & Grant, 144 molengraafi (Broili), 122 simulata (Reed), 131 soochowensis (Chao) , 165, 169 speciosus (Waterhouse & Piyasin) , 1 10, 122 strophomenoides (Waagen), 1 10, 124
Rugatia occidentalis (Newberry), 98 Rugivestis Muir-Wood & Cooper, 79 Rugosofusulina Rauser, Table 49, p. 1 77
alp ina (Schellwien), 6 1 , 66, 82, 84 ; Table 21 , p . 70

230 Systematic Index for Genera and Species
alpina communis (Schellwien), 99 complica (Schellwien), 6 1 moderato (Rauser), 61 multiseptata (Schellwien), 85, 97, 118 serrata Rauser, 79
Russiella Mikluko-Maklay, Table 1 8, p. 62 pulchra Mikluko-Maklay, 107
Sajakella aurita Volgin, 6 1 zyriankensis Zavodowsky, 5 7
Sakmarites Ruzencev, 5 3 , 9 5 ; Table 49, p. 177; Table 50, p; 178; Figs. 13 , 14, pp. 50, 5 1
Scacchinella Gemellaro, 45, 60, 6 1 , 82, 1 3 1 , 134 ; Table 50, p . 178
Schistoceras Hyatt, 42 Schizophoria juresanensis Chernyshev, 79 Schuchertella Girty, 132, 141
semiplana (Waagen), 146, 148, 152 Schwagerina Moeller (American Sense), 18, 20,
40, 49, 72, 79, 87, 98, 109, 1 18, 176; Table 18, p. 62; Table 49, p. 177
chihsianensis (Lee), 85 crassitectoria Dunbar & Skinner, 80, 90;
Table 26, p. 88; Table 27, p. 91 ; [usiformis Krotow, Table 43, p. 150 "(= Pseudofusulina-Leeina) [usiformis (Schellwien), 80, 82 princeps (Ehrenburg), 84 (= Pseudoschwagerina) uddeni Beede &
Kniker, 20 Schwagerina* Moeller (Soviet Sense), 1 10
[usiformis Krotow, 53, 55, 61 , 84 moelleri Rauser, 53 ; Table 43, p. 150 nitida Kahler & Kahler, 82 sphaerica Rauser & Scherbovich, 53
Semicostella Muir-Wood & Cooper, 46 Septacamera Stepanov, 14, 57
garoudi (Mansuy), Table 42, p. 147 krotovi (Chernyshev), 79
Septiconcha Termier et al. , 141 purdoni (Davidson), 141
Septospirifer Waterhouse, 46 Septospirigerella megridagica Grunt, 15 1 Sergospirifer Ivanova, 84
carnicus (Schellwien), 66 Settedabania Abramov, 46
stepanovi Abramov, Table 13, p. 47 Shengoceras Chao, 109; Figs. 8, 9, p. 32 Shumardites Schindewolf, 42, 57 Silvasep.topora Chronic, 75 Skinnerina Ross, 1 10, 134
cf sapperi (Schellwien & Staff), 110 Somoholites Ruzencev, 57, 72
cf artus Ruzencev, 72 cf beluensis (Haniel), 87
Sowerbina Frederiks, 96 , 98, 1 16; Table 2b, p. xv; Table 22, p. 73; Table 50, p. 178
Sphaeros�hwagerina carnica (Schellwien), 61 Sphaerulrna Lee, 66, 97; Table 18, p. 62 Sphenalosia Muir-Wood & Cooper, 141
salmunensis (Reed), 141 smedleyi Muir-Wood & Cooper, 1 19
Sphenosteges Muir-Wood & Cooper, 135 hispidus (Girty), 1 19, 135
Spinomargini[era Huang, 143, 161, 165; Table 47, p. 164 ; Table 52, p. 1 82
jisuensis (Chao), Table 4 7, p. 164 kweichowensis Huang, 146
Spinomartinia Waterhouse, Table 2b, p. xv spinosa Waterhouse, 156, 166; Table 6, p. 17
Table 37, p. 128 Spirelytha schei (Chernyshev & Stepanov), 107 "Spirifer" Sowerby, 66, 79
condor D'Orbigny, 45 oldhamianus Waagen, 134 peregrinus Reed, 132 simaanensis Hamlet, 122 wynnei Waagen, 144
Spiriferella Chernyshev, 16, 79, 80,_82, 85, 87, 96, 99, 1 10, 1 22, 127, 1 32, 149, 151 , 156, 157 ; Table 6, p . 17 ; Table 42, p. 147 ; See Fig. 6, p. 15 for all species
asiatica Volgin, 6 1 australasica (Etheridge), 1 24 cambodgensis Chi-Thuan, 134 draschei (not Toula) of Cooper, 72 (? = Eridmatus) gjeliensis Stepanov, 55;
Table 13, p. 47 keilhavii (von Buch), 96, 116 , 1 22, 127; Table
5 1 , p. 1 80 kolmaensis Zavodowsky, 85 /ita Frederiks, 106 loveni (Diener), 106, 1 16, 1 22; Table 5 1 , p.
1 80 mica Barkhatova, 55 mongolica Grabau, 1 5 1; Table 47, p. 164 parva Cooper, 72 polaris (Wiman), 1 24 ; Table 5 1 , p. 180 pseudodraschei Einor, 72 pseudosaranae Einor, Table 49, p. 177 pseudotibetana Stepanov, 96, 98 rajah (Salter), 143 , 144, 151 , 153, 163;
Table 47, p. 1 64 ; Taole 52, p. 182 salteri Chernyshev, 82; Table 49, p. 177 saranae (Verneuil), 79, 85, 105 ; Table 50, p.
178 scobinoidea Cooper, 1 22 supplanta Waterhouse, 1 18, 127; Table 37, p.
128 turusica Einor, 1 5 1 vaskovskii Zavadowsky, 9 7 , 9 8 vercherei not Waagen o f Frederiks, 106 waageni (Chernyshev) , 82
Spiriferellina Frederiks, 1 1 , 137, 153 ; Table 42, p. 147; Fig. 28, p. 1 2 1
cristata (Schlothem), 103, 1 13 ; Table 5 1 , p. 1 80
multiplicata (Sowerby), 141 Spiriferinaella Frederiks, 137 Spirigerella Waagen, 107
alata Waagen, 141 hybrida Waagen, 141 ovoidalis Waagen, 141
Spirolegoceras Miller et al. , 1 29 fischeri Miller et al. , 1 19
Spirovallum Waterhouse, 169 Squamaria ivesi (Newberry), 98 "Squamularia" grandis (Chao), 165

Systematic Index for Genera and Species 23 1
Stacheoceras Gemmellaro, 99, 137, 148, 152, 169; Table 5 1 , p. 1 80
mediterraneum Gemmellaro, 107 rothi Miller & Furnish, 107 toumanskiae Miller & Furnish, 153 cf tridens Rothpletz, 153, 166
Staffella Ozawa, Table 18, p. 62; Table 48, p. 170
gemmellaroi Miller, 13 7 Stegacanthia (?) taimyrensis (Einor), Table 13,
p. 47 Stenopronorites Schindewolf, 124
ekatshanensis Popov, 51 ex. gr. karpinskii Librovitch, Table 13, p. 47
Stenoscisma Conrad, 48 alpina (Schellwien), 66, 69 armenica Sokolskaya, 163 gigantea (Diener), 145 papilio Waterhouse, 156
• purdoni (Davidson), 145Stepanoviella Zavodowsky, 57, 84, 85, 92, 96,
122 124, 1 29, 132, 141, 145, 149, 152, 157; Table 51, p. 1 80
·
corcodonensis Likharev, 105 flexuosa Waterhouse, 95 ; Table 50, p. 1"78 kulikii Frederiks ? lahuseni (Likharev), 141 mekongensis (Mansuy), 146·umariensis (Reed) , 84 ; Table 38, p. 130
Streptognathodus sulcoplicatus , 109 Streptorhynchus King, 1 1 , 96,.99, 105, 1 10, 134,
141 , 1 5 1 bioni Reed, 92 pelargonatus (Schlotheim), 103, 1 13 ; Table
5 1 , p. 180 (? Gn.tmantia) pelicanensis Fletcher, 137;
Table 51, p. 1 80 Strigogoniatites Spath, 152; Table 52, p. 182
fountaini Miller & Furnish, 166 kingi Miller, 153 zavodowskii Popov, 106
Strophalosia King, 149 blandfordi Reed, 141 chivatschensis Zavadowsky, 149 multicosta Likharev, 163
Strophalosiina tibetica (Diener) , 144, 146, 148, 153, 157; Table 52, p. 182
Stutchburia Etheridge, 129 ; Table 51, p. 1 80 Su/ciplica Waterhouse, 92, 126, 132
peregrinus (Reed), 132 Sumatrina Volz, 131 ; Table 18, p. 62 Sverdn.tpites Nassichuk, 116; Table 51, p. 1 80
harkeri (Ruzencev), 1 16 Sweetognathodus whitei (Rhodes), 26 ; Table 7,
p. 27 Synartinskia Ruzencev, 78, 82; 90; Table 49,
p. 177 belcheri Nassichuk, 1 16
Tabantalites Ruzencev, 53, 82; Table 49, p. 177; Fig. 14, p. 5 1
bi[urcatus Ruzencev aff trapezoidalis Maximova, 87
Taeniothaen.ts Whitehouse, 92, 99, 124 aff miniliensis Coleman, 102
(= Reedoconcha) permixtus Reed, 84 ; Table 38, p. 130
n.tsticus Grunt, 82 subquadratus (Morris) , 99, 102
Taimyrella Ustritsky, 57, 79, 99 Tangshangel/a byrangi Chernyak, Table 13, p. 47 Teguliferina Schuchert & Le Vene, 45 , 48, 60,
6 1 ; Fig. 12, p. 50 Terebratuloidea Waagen, 15 1
davidsoni Waagen, 148 depressa Waagen, 141 , 148
Terrakea Booker, 97, 105, 1 18, 127, 137 arcticum Waterhouse, 106 brachythaen.tm (Morris), 137, 157 ; Table 2b,
p. xv; Table 6, p. 1 7 ; Table 37, p. 128 concavum Waterhouse, 106, 126, 127 ; Table
2b, p. xv; Table 5 1 , p. 1 80 dickinsi Dear, 99, 127 ; Table 2b, p. xv; Table
37, p. 128; Table 5 1 , p. 180 elongatum (Etheridge & Dun), 140 kozlowskianus (Frederiks), 106 multispinosa Dear, Table 52, p. 182 pollex Hill, 95
Thuleproductus Sarytcheva & Waterhouse, 1 1 8 arcticum (Whitfield), 116, 1 18 ; Table 5 1 , p.
1 80 Timaniella Barkatova, 14, 151
harkeri Waterhouse, 106, 118 ; Table 5 1 , p . 1 80
Timoriina broili Stehli, 153 Timorites Haniel, 19, 31, 34, 140, 143, 148,
149, 152, 153, 166, 1 8 1 ; Table 1 1 , p. 35 ; Table 35, p. 123; Table 52, p. 1 82; Figs. 8, 9, p. 32
schucherti Miller & Furnish, 153 Tityrophoria Waterhouse, 85 ; Table 50, p. 178 Tomiopsis Benediktova, 46, 48, 49, 52, 53, 55,
57, 60, 64, 72, 77, 79, 80, 84, 85, 87, 92, 95, 105, 106, 127, 141, 172, 179; Table 2b, p. xv; Table 22, p. 73; Table 38, p. 130; Table 49, p. 177; Table 50, p. 178 ; Table 52, p. 182
angulatum (Campbell), 77 costata Waterhouse, 140 ; Table 5 1 , p. 1 80 havilensis (Campbell), 156 lata Czamiecki, 69 mantuanensis (Campbell), 140 mergensis Chernyak, 1 OS ovulum Waterhouse, 69, 75 parallela (Waterhouse), 157 petrenkoi Czarniecki, 69 punjabica (Reed), 141 , 157 sokolovi (Chernyshev), 61
Tompophiceras, Table 46, p. 159 Tornquistia Paeckelmann, 57, 80, 87, 90, 105,
106, 1 29; Table 2b, p. xv; Table 5, p. 13 Table 22, p. 73
Toriyamaia Kanmera, Table 18, p. 62 Transennatia Waterhouse, 145
gratiosus (Waagen), 144 Trigonogastrites, 169 Trigonotreta Koenig, 1 22
simaanensis (Hamlet), 1 22 Triticites Girty, 44, 55 , 61 , 66, 72, 75, 84, 106
contractus (Schellwien), 97

232 Systematic Index for Genera and Species
opimus (Dunbar & Newell), 75, 102 simplex (Schellwien), 84 ventricosus (Meek), 72
Tropidelasma Cooper & Grant, 45 Tschernyschewia Stoyanov, 132, 141 Tubersulculus Waterhouse, 46, 48, 72 Tyloplecta Muir-Wood & Cooper, 134 , 14 1 , 161
nankinensis (Frech), 109, 1 10, 132, 146 sumatrensis (Roemer), 1 22 yangzteensis (Chao) , 109, 1 3 1 , 146, 149,
165
Uddenites Bose, 42, 45 , 49, 52, 72; Table 26, p. 88; Figs. 12-15, pp. 50-5 1
Uddenoceras Miller & Furnish, 42, 45 Uncinella Waagen, 1 1 0, 134 ; Table 5 1 , p. 1 80 Uncinunellina Grabau, 14 1 , 153
jabiensis (Waagen), 14 1 , 146 theobaldii (Waagen), 141 timorensis (Beyrich), Table 42, p. 147
Undulomya Fletcher, 124 Uraloceras Ruzencev, 48, 78, 87, 95; Table 49,
p. 177 ex. gr. belgushkense Ruzencev, 79 simense Ruzencev, 48, 57 suessi (Karpinsky), 105
Uraloproductus Ustritsky, 57 stuckenbergianus (Krotow) , 55 , 96 ; Table 50,
p . 178 Urushtenia costulata (Chi-Thuan) , 1 10
Vedioceras Ruzencev, 2, 161 ; Table 45 , p. 158; Table 46, p. 159; Table 52, p. 1 82
ventroplanum Ruzencev, 29 Verbeekina Staff, 34 , 109, 110, 121 , 146 , 148,
1 5 1 , 165 ; Table 1 1 , p. 35 ; Table 18, p. 62; Table 5 1 , p. 180
verbeeki (Geinitz) , 122, 134 , 149; Table 20, p. 67; Table 42, p. 147
Vescotoceras Ruzencev, 165 ; Table 45 , p. 158
Waagenina Krotow, 95 , 96 dieneri Smith, 82 subinterrupta Krotow, 96
Waagenites (= Rugaria) speciosus Waterhouse & Piyasin, 134, 14 1 , 148
Waagenoceras Gemmellaro, 19, 3 1 , 34 , 109, 1 3 1 , 134 , 1 3 7 143, 145, 1 5 2 , 1 7 9 ; Table 1 1 , p. 3 5 ; Table 3 5 , p . 123; Table 5 1 , p. 1 80; FigL 8, 9, p. 32
dieneri Bose, 137, 153 Waagenoconcha Chao, 14, 45 , 79, 1 16 , 1 3 1 ;
Table 47, p . 164 abichi (Waagen), 153 guadalupensis (Girty), 137 imperfecta Prendergast, 156 irginae (Stuckenberg), 96 montpelierensis (Girty), 135 (= Septiconcha) purdoni (Davidson), 141 sarytchevae (Benediktova), Table 13, p. 47 waageni (Rothpletz) , 156
Wairakiella rostrata Waterhouse, Table 37, p. 128 Wedekindellina Dunbar & Henbest, 66 ; Table 2 1 ,
p. 70
Wellerella Dunbar & Condra, 141 , 152 Wentzelella subtimorica Wang, 109 Wutuella Sheng, 148; Wyndhamia Booker, 99, 127, 137
clarkei gattoni (Maxwell), 140 dalwoodensis Booker, 1 26, 127 jukesi (Etheridge) , 102; Table 50, p. 178
Xenaspis Waagen, 143, 1 74 Xenodiscites Miller & Furnish, 152 Xenodiscus Waagen, 141 , 143, 144, 145, 152,
161 carbonarium (Waagen), 153 cf. carbonarium (Waagen), 174
Yabeina Deprat, 16, 1 8 , 34, 109, 1 18, 1 3 1 , 132, 134 ,.140, 144, 145 , 146, 148, 149, 151, 157, 165, 181 ; Table 1 1 , p. 35 ; Table 18, p . 62; Table 39, p. 1 3 3 ; Table 52, p. 182
archaica Dutkevich, 1 5 1 ; Table 23 , p. 81 asiatica Ishii, 1 32, 146, 1 65 globosa (Yabe), 1 32, 146, 148, 152; Table 42,
p. 147; Table 48, p. 1 70 cf gubleri Kanmera, 148 inouyei Deprat, 1 34 johannis Saurin, 134 katoi Ozawa, Table 48, p. 1 70 khmeriana Saurin, 132 minuta Thompson & Wheeler, 146 shiraewensis Ozawa, 148, 149; Table 20, p.
67 ; Table 48, p. 1 70 syrtalis (Douville) , 1 5 2 ussurica Dutkevich, 149 yasubaensis Toriyama, Table 20, p. 67
Yakovlevia Licharev, 49, 5 5 , 72, 80, 87, 96, 149, 176; Table 2b, p. xv; Table 22, p. 73; Table 49, p. 177 ; Figs. 1 2-15 , pp. 50, 5 1
geniculata (Girty), 1 19 mammatiformis (Frederiks), 79, 80
Yakutoceras Librovitch, 46, 79 trianguliumbicilatum (Popov), Table 13, p.
47 Yanchienia Lee, 109 ; Table 18 , p. 62
Zellia Kahler & Kahler, 64
Copyright © 2022 FDOKUMEN

