







Bahasa
Halaman
Hukum


This content has been downloaded from IOPscience. Please scroll down to see the full text.
Download details:
IP Address: 137.110.89.27
This content was downloaded on 30/07/2014 at 23:18
Please note that terms and conditions apply.
Localized increase of tissue oxygen tension by magnetic targeted drug delivery
View the table of contents for this issue, or go to the journal homepage for more
2014 Nanotechnology 25 265102
(http://iopscience.iop.org/0957-4484/25/26/265102)
Home Search Collections Journals About Contact us My IOPscience

Localized increase of tissue oxygen tensionby magnetic targeted drug delivery
Celine Liong1, Daniel Ortiz1, Eilleen Ao-ieong1, Mahantesh S Navati2,Joel M Friedman2 and Pedro Cabrales1
1Department of Bioengineering University of California, San Diego La Jolla, CA 92093 USA2Department of Physiology and Biophysics, Albert Einstein College of Medicine, Bronx, New York USA
E-mail: [email protected]
Received 28 February 2014, revised 27 April 2014Accepted for publication 30 April 2014Published 12 June 2014
AbstractHypoxia is the major hindrance to successful radiation therapy of tumors. Attempts to increasethe oxygen (O2) tension (PO2) of tissue by delivering more O2 have been clinicallydisappointing, largely due to the way O2 is transported and released by the hemoglobin (Hb)within the red blood cells (RBCs). Systemic manipulation of O2 transport increases vascularresistance due to metabolic autoregulation of blood flow to prevent over oxygenation. This studyinvestigates a new technology to increase O2 delivery to a target tissue by decreasing the Hb-O2
affinity of the blood circulating within the targeted tissue. As the Hb-O2 affinity decreases, thetissue PO2 to satisfy tissue O2 metabolic needs increases without increasing O2 delivery orextraction. Paramagnetic nanoparticles (PMNPs), synthetized using gadolinium oxide, werecoated with the cell permeable Hb allosteric effector L35 (3,5-trichlorophenylureido-phenoxy-methylpropionic acid). L35 decreases Hb affinity for O2 and favors the release of O2. The L35-coated PMNPs (L35-PMNPs) were intravenously infused (10 mg kg−1) to hamsters instrumentedwith the dorsal window chamber model. A magnetic field of 3 mT was applied to localize theeffects of the L35-PMNPs to the window chamber. Systemic O2 transport characteristics andmicrovascular tissue oxygenation were measured after administration of L35-PMNPs with andwithout magnetic field. The tissue PO2 in untreated control animals was 25.2 mmHg. L35-PMNPs without magnetic field decreased tissue PO2 to 23.4 mmHg, increased blood pressure,and reduced blood flow, largely due to systemic modification of Hb-O2 affinity. L35-PMNPswith magnetic field increased tissue PO2 to 27.9 mmHg, without systemic or microhemodynamicchanges. These results indicate that localized modification of Hb-O2 affinity can increase PO2 oftarget tissue without affecting systemic O2 delivery or triggering O2 autoregulation mechanisms.This technology can be used to treat local hypoxia and to increase O2 in tumors, enhancing theefficacy of radiation therapies.
S Online supplementary data available from stacks.iop.org/NANO/25/265102/mmedia
Keywords: paramagnets, tissue oxygen, microcirculation, hemoglobin, oxygen delivery
(Some figures may appear in colour only in the online journal)
Background
Localized increase of tissue oxygen (O2) tension (PO2) is abiomedical challenge with important clinical implications [1].Therapies to increase tissue PO2 often include vasoactivedrugs to increase blood flow and O2 supplementation byincreasing the fraction of inspired O2 (FiO2). Increased blood
flow augments O2 delivery but not tissue PO2, as tissue PO2
gradients between the blood and the tissue remain unalteredand reduce blood tissue transit time. Increased FiO2 augmentsalveolar and arterial PO2; however, the additional O2 ismostly dissolved in the plasma. Due to limited O2 solubilityin plasma, the dissolved O2 in plasma is insignificant com-pared to the O2 bound to the hemoglobin (Hb) in the red
0957-4484/14/265102+12$33.00 © 2014 IOP Publishing Ltd Printed in the UK1
Nanotechnology
Nanotechnology 25 (2014) 265102 (12pp) doi:10.1088/0957-4484/25/26/265102

blood cells (RBCs). Therefore, increasing the FiO2 does notnecessarily translate into an increase of O2 released to tissue.The clinical applicability of targeted increases in PO2 spansfrom the treatment of focal ischemia to the treatment ofcancer. The hypoxic environment of tumors is often asso-ciated with limited therapy efficacy (e.g. radiation) [2].Increasing the PO2 in tumors would allow for increasedproduction of cytotoxic reactive oxygen species (ROS) [3].
Oxygen is transported and delivered through its rever-sible binding to Hb contained in the RBCs. Normal Hb-O2
binding is controlled by allosteric effectors (e.g. 2,3-dipho-sphoglycerate, 2,3 DPG) which facilitate loading and off-loading of O2 by tuning Hb-O2 binding properties. Hb-O2
binding properties control O2 delivery, which determinestissue PO2 when O2 consumption remains unaltered. The fluxof O2 leaving a blood vessel is determined by the O2 con-sumption of the tissue surrounding the blood vessel. This fluxof O2 from arterioles and capillaries to the tissues is driven bythe PO2 gradients between the blood and adjacent cells [4].Therefore, modifying Hb-O2 affinity can manipulate the PO2
gradients required to preserve the tissue O2 consumptionwhile manipulating tissue PO2 [5]. For example, allostericeffectors that increase Hb-O2 affinity can decrease tissue PO2,as a change in Hb-O2 saturation occurs at lower PO2. Butdecreasing Hb-O2 affinity can increase tissue PO2 byachieving the required changes in Hb-O2 saturation at ahigher PO2. Our previous studies showed that a systemicincrease in Hb-O2 affinity is beneficial during anemic andhypoxic conditions by increasing O2 captured in the lungs,increasing arterial Hb-O2 saturation, and preserving O2
delivery to the microcirculation [6–7]. Systemic decreases inHb-O2 affinity facilitate the release of O2 to the micro-circulation, which triggers O2 autoregulation mechanisms,producing vasoconstriction and preventing O2 from beingtransported and offloaded to tissues [8]. To prevent metabolicO2 autoregulation mechanisms in response to tissue hyper-oxygenation, the decrease in Hb-O2 affinity has to occur afterthe arterioles, in the capillary microcirculation (50–120 μm indiameter).
A fundamental feature of the Hb-O2 affinity and coop-erativity is the action of allosteric effectors in Hb quaternaryequilibrium that favor the low-affinity T or the high-affinity Rstate. Hemoglobin is a sophisticated molecular machine ableto maintain cooperative ligand binding over a large range ofHb-O2 affinities. L35 (3,5-trichlorophenylureido-phenoxy-methylpropionic acid) is a potent Hb allosteric effector[9–10]. L35 synthesis has been described in detail before [9],and L35 binding to Hb and L35 effects on Hb structures havebeen documented before [10]. L35 favors a Hb low-affinitystate and increasing the PO2 where 50% of the Hb is saturatedwith O2 (P50). L35 binds toward the αα end of the Hb centralcavity and works synergistically with 2,3 DPG and otherorganic phosphate-based allosteric effectors to decrease Hb-O2 affinity, since it binds to the ββ end of the Hb centralcavity [11]. L35 easily penetrates the RBC membrane withoutdamaging the cell membrane; it also decreases the Hb-O2
affinity of RBC suspensions [9].
This study builds on previous studies in which oleic acidwas used to coat paramagnetic gadolinium oxide nanocrystalsto produce paramagnetic nanoparticles (PMNPs) [12–13].The oleic acid on the PMNPs can be loaded with a widevariety of hydrophobic drugs. Technically the paramagneticproperties of the gadolinium are used to localize the effects ofthe drugs to a specific site by applying a magnetic field. Thisstudy utilizes L35 loaded PMNPs (L35-PMNPs) to evaluatewhether a localized decrease of Hb-O2 affinity increases PO2
in a targeted area of tissue. We tested the hypothesis that therelease of the allosteric effectors (L35) from L35-PMNPs canbe concentrated to a specific site using a magnetic field, thusdecreasing the Hb-O2 affinity of the RBCs circulating the site,increasing tissue PO2, and preventing hemodynamic changesdue to metabolic autoregulation. The L35-PMNPs were givenas a single IV dose (10 mg kg−1) to Golden Syrian hamstersinstrumented with the dorsal window chamber model. Onegroup of animals was exposed to a strong magnetic field(3 mT) applied to the window chamber area, whereas theother group was not exposed to a magnetic field. Controlanimals did not receive L35-PMNPs. Oxygen tensions weremeasured using phosphorescence quenching microscopy(PQM) and focused on healthy tissues with well regulatedtissue PO2.
Materials and methods
L35-PMNPs preparation
The preparation of the L35 loaded PMNPs (L35-PMNPs) isbased on a protocol developed by the Einstein team to createdriamycin coated PMNPs for targeted treatment of tumors.The general process started with the preparation of the oleicacid coated paragnetic nanocrystalline core. A 10 mg sampleof commercially purchased gadolinium oxide nanocrystals(Nanostructured & Amorphous Materials, Inc., Houston,TX) with an average diameter of 30 nm (figure 1(A)) werewashed several times in 5 ml of deionized (DI) water andthen centrifuged. Next, 300 μl of oleic acid in dimethylsulfoxide (DMSO) (1:19) were mixed with water-free spundown particles, followed by vigorous sonication for 1 h in acold water bath, and left in a lab rotor overnight. Theresulting particles were spun down and washed several timeswith DI water, dried, then lyophilized for storage. They werereconstituted by mixing with an aqueous solvent and soni-cating briefly. The resulting suspension was stable with nodetectable aggregates forming over a period of several days.Dynamic light scattering measurements indicated that theresulting particles were less that 100 nm. The drug loadingprocess consisted of mixing 1 mg ml−1 of L35 solubilized in1 ml of DMSO with 10 mg ml−1 of oleic acid coated,gadolinium oxide based PMNPs at room temperature. TheDMSO mix was allowed to remain for 24 h in the dark atroom temperature. Then the suspension was centrifuged, thebrown color of the L-35 was localized, and the DMSO waspoured off. The brown colored PMNPs were washed threetimes with phosphate-buffered saline (PBS) and vortexed to
Nanotechnology 25 (2014) 265102 C Liong et al
2

re-suspend the particles. After the last wash, the excessbuffer was poured off, and the remaining brown materialwas lyophilized. The L35 coated PMNPs were then PEGy-lated by adding a small aliquot of m-PEG-DSPE (NanocsInc., New York, NY) dissolved in DMSO (derived from astock solution of 1 mg of m-PEG-DSPE dissolved in 1 ml ofDMSO). Typically, 10–50 μl of the m-PEG-DSPE DMSOstock solution was used to PEGylate 50–100 mg of L35-PMNPs. Other drugs have been prepared with loadedPMNPs in a similar manner (e.g. Adriamycin, curcumin,melanin). Exposing a cuvette containing melanin loadedPMNPs suspension to a magnet results in the accumulationof the nanoparticles adjacent to the position of the magnet(figure 1(B)).
Rhodamine labeled particles
The fluorescent dye rhodamine 110 (Sigma-Aldrich, St.Louis, MO) was bound to the surface of the L35-PMNPs bylinking the dye to carboxyl groups. Briefly, a concentrateddispersion of L35-PMNPs was prepared in a 25 mm aqueous(2-N-morpholino ethanesulfonic acid, Sigma-Aldrich) buf-fer. To the L35-PMNPs dispersion, 4 mmol of 1-3-(dime-thylaminopropyl)-3-ethylcarbodiimidehydrochloride(Sigma-Aldrich) followed by 0.25 mmol of rhodamine 110dye were added, and the reaction vial was vortexed for 4 h,
protected from the light at room temperature. The resultingrhodamine labeled L35-PMNPs were washed two timesthrough alternate cycles of vortexing and centrifugation toremove any unbound rhodamine dye.
Animal preparation
Male Golden Syrian hamsters 55–65 g (Charles RiverLaboratories, Boston, MA) were fitted with dorsal windowchambers. Animal handling and care followed the NIH Guidefor the Care and Use of Laboratory Animals, and theexperimental protocol was approved by the local animal carecommittee. The hamster window chamber model is widelyused for microvascular studies in the unanesthetized state, andthe complete surgical technique is described in detail else-where [14–15]. Briefly, the animal was prepared for chamberimplantation with a 50 mg kg−1 ip injection of pentobarbitalsodium anesthesia. After hair removal, sutures were used tolift the dorsal skin away from the animal, and one frame of thechamber was positioned on the animal’s back. The chamberconsisted of two identical titanium frames with a 15 mmcircular window. With the aid of backlighting and a stereo-microscope, one side of the skin fold was removed, followingthe outline of the window, until only a thin layer of retractormuscle and the intact subcutaneous skin of the opposing sideremained. The intact skin of the other side was exposed to the
Nanotechnology 25 (2014) 265102 C Liong et al
Figure 1. (A). Paramagnetic nanoparticles (PMNPs). Exposing the suspension of PMNPs to a powerful magnet resulted in the accumulationof the brown particles to the location where the magnetic field was applied. Other drugs have been loaded to PMNPs (e.g. Adriamycin,curcumin and melanin). (B1). Brown color solution of melanin-PMNPs in PBS buffer (without magnetic field). (B2). Melanin-PMNPssuspended in PBS buffer with a magnetic field applied. Pictures B1 and B2 were taken with melanin-PMNPs, as they are darker brown andeasier to photograph. (C). Hamster window chamber model. Implanted titanium frames on the dorsal skinfold for microvascular studies(18 mm window diameter). Intact tissue is studied in the absence of surgical trauma or anesthesia. Catheters are implanted and exteriorized atthe dorsal side and secured to the frames. A cylindrical strong magnet was used to localize the effects of the L35-PMNM to the dorsalwindow area.
3

ambient environment. Animals were allowed 2 d for recovery(figure 1(C)). Finally, catheters (PE-50) were implanted in thecarotid artery and jugular vein. Catheters were tunneled underthe skin, exteriorized at the dorsal side of the neck, andsecurely attached to the window frame.
Inclusion criteria
Hamsters were suitable for the experiments if: 1) systemicparameters were within normal range: namely, heart rate(HR) > 340 beat min−1, mean arterial blood pressure(MAP) > 80 mmHg, systemic hematocrit (Hct) > 45%, andarterial PO2 pressure (PaO2) > 50 mmHg; and 2) microscopicexamination of the tissue in the chamber observed under x650magnification did not reveal signs of edema or bleeding.Hamsters are a fossorial species with a low arterial PO2
compared to other rodents; however, the PO2s in the hamsterwindow chamber are similar to the PO2s in other rodents (e.g.mice) [16].
Systemic parameters
MAP and HR were recorded continuously (MP 150, BiopacSystem; Santa Barbara, CA). Hct was measured from cen-trifuged arterial blood samples taken in heparinized capillarytubes. Hb content was determined spectrophotometrically (B-Hemoglobin, Hemocue, Stockholm, Sweden). Arterial bloodwas collected in heparinized glass capillaries (50 μl) andimmediately analyzed for PO2, carbon dioxide partial pres-sure (PCO2), base excess (BE), and pH (Rapidlab 248, Bayer,Norwood, MA). Arterial Hb saturations were measured on theIL482 CO-Oximeter System (Instrumentation Laboratory,Lexington, MA).
Blood oxygen equilibrium curve
Oxygen equilibrium curves for hamster blood were obtainedby deoxygenation of O2-equilibrated samples in a Hemoxbuffer at 37 °C, using a Hemox Analyzer (TCS ScientificCorporation, New Hope, PA). The Hemox buffer pH wasadjusted to match the arterial blood pH of the animals usingTris and BisTris buffers. Tris and BisTris buffers were pre-pared by titrating the reagents with hydrogen chloride (HCI)before adjusting the pH of the solutions to keep the Cl—ionconcentration equal to the buffer at the pH values.
Experimental setup
The window chamber was studied using a custom intravitalmicroscope. Briefly, the restrained animals were fixed to themicroscopic stage for transillumination (BX51WI, Olympus,New Hyde Park, NY). Measurements were carried out using a40X (LUMPFL-WIR, numerical aperture 0.8, Olympus)water immersion objective. The tissue image was projectedonto a charge-coupled device camera (COHU 4815, SanDiego CA), viewed on a TV monitor, and analyzed onlineusing the following techniques.
Microhemodynamics
A video image-shearing method was used to measure vesseldiameter (D) [17]. Changes in arteriolar and venular diameterfrom baseline were used as indicators of a change in vasculartone. Arteriolar and venular centerline velocities were mea-sured online using the photodiode cross-correlation method(Photo Diode/Velocity Tracker Model 102B, Vista Electro-nics, San Diego, CA). The measured centerline velocity (V)was corrected according to vessel size to obtain the meanRBC velocity [18]. Blood flow (Q) was calculated from themeasured values as Q= π×V (D/2)2.
Microvascular oxygen tensions
High resolution non-invasive microvascular PO2 measure-ments were made using phosphorescence quenching micro-scopy (PQM) [19]. PQM is based on the relationship betweenthe decay rate of excited Palladium-mesotetra-(4-carbox-yphenyl) porphyrin (Frontier Scientific Porphyrin Products,Logan, UT) bound to albumin and the O2 concentrationaccording to the Stern-Volmer equation [20]. PO2 measure-ments by PQM were obtained by illuminating the tissue witha pulsed light at 420 nm to excite the probe into its tripletstate. The emitted phosphorescence (680 nm) was collectedand analyzed to yield the phosphorescence lifetime, whichwas then converted into PO2. The phosphorescence lifetimesare concentration independent, permitting extravascular fluidPO2 measurements. Animals received a 50 μl IV of 10mg dl−1 palladium-porphyrin complex (Frontier ScientificPorphyrin Products, Logan, UT). Porphyrin was allowed todistribute for 15 min before measurements were taken.Intravascular PO2 was measured for the same blood vesselsselected for hemodynamic analysis.
Experimental protocol
Awake hamsters, instrumented with the window chamber,were restrained in an acrylic tube with perforations to allowaccess of air. The window chamber protruded out of a slit onthe tube and was used to secure the animal to the microscopestage. Animals were given 20 min to adjust to the tubeenvironment before any measurement. Then, detailed map-pings were made of the chamber vasculature. Blood vesselswere chosen based on distinctive anatomic landmarks andoptical clarity. The same blood vessels were studied atbaseline and following time points. In each preparation, 6–8arterioles and venules were selected. Tissue PO2 was mea-sured in consecutive spots, mapping the entire center of thewindow in locations free of blood vessels. Each row consistedof 35–40 measurements and covered an area of 7.3 mm2.Rows of measurements were spaced 100 μm to preventrepeated measurements from the same spot, and 15–22 rowswere measured in each window. L35-PMNPs were suspendedin saline and vortexed for 1 min. After baseline measurements(hemodynamics and PO2s), 10 mg kg−1 of body weight ofL35-PMNPs were administered in 100 μl volume. The mag-netic field was applied around the window chamber areabefore L35-PMNPs infusion. The time points studied were
Nanotechnology 25 (2014) 265102 C Liong et al
4

baseline (before any treatment) and 1 h after treatment. Allmeasurements were completed within 30 min.
Experimental groups
A total of 24 animals were used for the study. Animals wereassigned randomly to each experimental group. Groups werelabeled based on experimental conditions: 1) in L35-PMNPswith magnetic field ON (n = 8), animals received L35-PMNPsand an applied magnetic field; 2) in L35-PMNPs with mag-netic field OFF (n = 8), animals received L35-PMNPs and noapplied magnetic field; 3) untreated control animals (n = 8)did not receive the L35-PMNPs or applied magnetic field.
Oxygen saturations
Intravascular Hb-O2 saturations were calculated using the Hb-O2 equilibrium curves and the intravascular PO2s. The O2
equilibrium curves were measured at arterial blood pH, asdescribed above. Intravascular Hb-O2 saturation for the ani-mals treated with L35-PMNPs and magnetic field ON wereassumed to follow the changes in Hb-O2 equilibrium curvesmeasured in untreated animals. This assumption may over-estimate venular Hb-O2 saturations, as the local change in Hb-O2 equilibrium curves of the blood leaving the tissue may beshifted to the right. However, micro-sampling blood fromsmall venules is extremely difficult.
Oxygen delivery and extraction
The microvascular methodology used in our studies allows adetailed analysis of O2 supply in the tissue. Calculations weremade using equations (1) and (2):
⎡⎣ ⎤⎦γ= × × ×( )O delivery RBCHb SA Q (1)2
⎡⎣ ⎤⎦γ= × × − ×( )O extraction RBCHb SA V Q (2)2
where RBCHb is the total Hb [gHb/dlblood], γ is the O2carrying capacity of saturated Hb [1.34 mlO2/gHb], SA is thearteriolar blood O2 saturation, A-V indicates the arteriolar/venular differences, and Q is the average microvascular bloodflow (arterioles and venules). Intravascular Hb O2 saturationswere calculated as described above.
Biodistribution in vivo imaging studies
The biodistribution imaging studies were performed onhamsters (n = 3) of similar size as those used for the micro-vascular study. Animals received 50 mg kg−1 IP injections ofpentobarbital sodium anesthesia. After hair removal, sutureswere used to lift the dorsal skin away from the animal, and acatheter was implanted in the jugular vein. Rhodaminelabeled L35-PMNPs (10 mg kg−1) were injected intrave-nously, and a strong magnet was placed under the skin.Dorsal skin fluorescence images were acquired on a MaestroII imaging system (Perkin Elmer, Norwalk, CT) 1 h afterinjection using a 500–720 nm emission filter with exposuretime of 1 sec.
Tissue accumulation study
Hamsters (n = 2), instrumented with the window chamber,were restrained and secured to the intravital microscope(BX51WI, Olympus, New Hyde Park, NY), as describedbefore. Rhodamine labeled L35-PMNPs (10 mg kg−1 sus-pended in 100 μl of physiological saline solution, 0.9% NaCl)were injected intravenously. Images before and after appli-cation of the magnetic field were recorded using a low lightvideo camera (ORCA 9247, Hamamatsu, Tokyo, Japan).Images were collected using a 20X (LUMPFL-WIR, numer-ical aperture 0.6, Olympus) water immersion objective andthe fluorescence filter XF100-2 (emission: 520 nm and exci-tation: 495 nm; Omega Optical, Inc. Brattleboro, VT). Themagnetic field was maintained for 1 h, then the animals wereplaced back in their cages. Additional hamsters (n = 2),instrumented with the window chamber, received the rhoda-mine labeled L35-PMNPs but were not exposed to a magneticfield. Next, 24 h after injection, major organs and the windowchamber were dissected for ex vivo imaging. Microscopicfluorescence images were taken to identify the presence ofrhodamine labeled L35-PMNPs in the tissues.
Data analysis
Results are presented as mean ± standard deviation. Datawithin each group was analyzed using analysis of variance forrepeated measurements (ANOVA, Kruskal—Wallis test).When appropriate, post hoc analyses were performed with theDunn’s multiple comparison test. Data comparison betweengroups was analyzed using two-way analysis of variance(Two-way ANOVA test). When appropriate, post-test ana-lyses were performed with the Bonferroni post-test compar-ison. Tissue PO2 frequency distribution of each group wastested for normality according to the Shapiro—Wilk test.Microhemodynamic data are presented as absolute values. Allstatistics were calculated using GraphPad Prism 4.01(GraphPad Software, Inc., San Diego, CA). Changes wereconsidered statistically significant if P < 0.05.
Results
In the study, 24 animals were divided into three groups: L35-PMNPs with magnetic field ON (n = 8), L35-PMNPs withmagnetic field OFF (n = 8), and untreated controls (n = 8). Allanimals tolerated the entire protocol without signs of dis-comfort or stress. For the biodistribution imaging (n = 3) andthe tissue accumulation study (n = 4), seven (n = 7) additionalanimals were used. All animals passed the Grubbs’ test,ensuring that all the measured values at baseline were within asimilar population (P < 0.05). Similarities between groups atbaseline for hamsters were statistically verified betweengroups (P > 0.30).
Blood chemistry and oxygen affinity
Blood basic hematology, arterial gases, and Hb-O2 affinityparameters for all groups are presented in table 1. Infusion of
Nanotechnology 25 (2014) 265102 C Liong et al
5
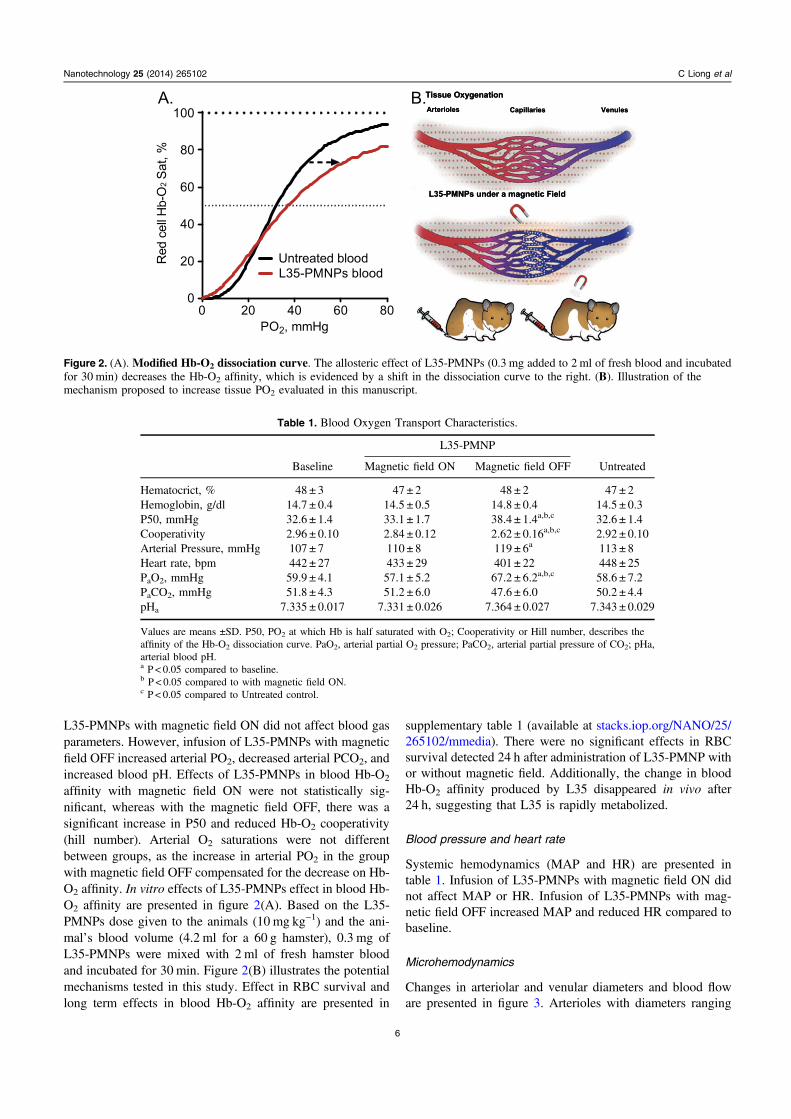
L35-PMNPs with magnetic field ON did not affect blood gasparameters. However, infusion of L35-PMNPs with magneticfield OFF increased arterial PO2, decreased arterial PCO2, andincreased blood pH. Effects of L35-PMNPs in blood Hb-O2
affinity with magnetic field ON were not statistically sig-nificant, whereas with the magnetic field OFF, there was asignificant increase in P50 and reduced Hb-O2 cooperativity(hill number). Arterial O2 saturations were not differentbetween groups, as the increase in arterial PO2 in the groupwith magnetic field OFF compensated for the decrease on Hb-O2 affinity. In vitro effects of L35-PMNPs effect in blood Hb-O2 affinity are presented in figure 2(A). Based on the L35-PMNPs dose given to the animals (10 mg kg−1) and the ani-mal’s blood volume (4.2 ml for a 60 g hamster), 0.3 mg ofL35-PMNPs were mixed with 2 ml of fresh hamster bloodand incubated for 30 min. Figure 2(B) illustrates the potentialmechanisms tested in this study. Effect in RBC survival andlong term effects in blood Hb-O2 affinity are presented in
supplementary table 1 (available at stacks.iop.org/NANO/25/265102/mmedia). There were no significant effects in RBCsurvival detected 24 h after administration of L35-PMNP withor without magnetic field. Additionally, the change in bloodHb-O2 affinity produced by L35 disappeared in vivo after24 h, suggesting that L35 is rapidly metabolized.
Blood pressure and heart rate
Systemic hemodynamics (MAP and HR) are presented intable 1. Infusion of L35-PMNPs with magnetic field ON didnot affect MAP or HR. Infusion of L35-PMNPs with mag-netic field OFF increased MAP and reduced HR compared tobaseline.
Microhemodynamics
Changes in arteriolar and venular diameters and blood floware presented in figure 3. Arterioles with diameters ranging
Nanotechnology 25 (2014) 265102 C Liong et al
Figure 2. (A). Modified Hb-O2 dissociation curve. The allosteric effect of L35-PMNPs (0.3 mg added to 2 ml of fresh blood and incubatedfor 30 min) decreases the Hb-O2 affinity, which is evidenced by a shift in the dissociation curve to the right. (B). Illustration of themechanism proposed to increase tissue PO2 evaluated in this manuscript.
Table 1. Blood Oxygen Transport Characteristics.
L35-PMNP
Baseline Magnetic field ON Magnetic field OFF Untreated
Hematocrict, % 48 ± 3 47 ± 2 48 ± 2 47 ± 2Hemoglobin, g/dl 14.7 ± 0.4 14.5 ± 0.5 14.8 ± 0.4 14.5 ± 0.3P50, mmHg 32.6 ± 1.4 33.1 ± 1.7 38.4 ± 1.4a,b,c 32.6 ± 1.4Cooperativity 2.96 ± 0.10 2.84 ± 0.12 2.62 ± 0.16a,b,c 2.92 ± 0.10Arterial Pressure, mmHg 107 ± 7 110 ± 8 119 ± 6a 113 ± 8Heart rate, bpm 442 ± 27 433 ± 29 401 ± 22 448 ± 25PaO2, mmHg 59.9 ± 4.1 57.1 ± 5.2 67.2 ± 6.2a,b,c 58.6 ± 7.2PaCO2, mmHg 51.8 ± 4.3 51.2 ± 6.0 47.6 ± 6.0 50.2 ± 4.4pHa 7.335 ± 0.017 7.331 ± 0.026 7.364 ± 0.027 7.343 ± 0.029
Values are means ±SD. P50, PO2 at which Hb is half saturated with O2; Cooperativity or Hill number, describes theaffinity of the Hb-O2 dissociation curve. PaO2, arterial partial O2 pressure; PaCO2, arterial partial pressure of CO2; pHa,arterial blood pH.a P < 0.05 compared to baseline.b P < 0.05 compared to with magnetic field ON.c P < 0.05 compared to Untreated control.
6
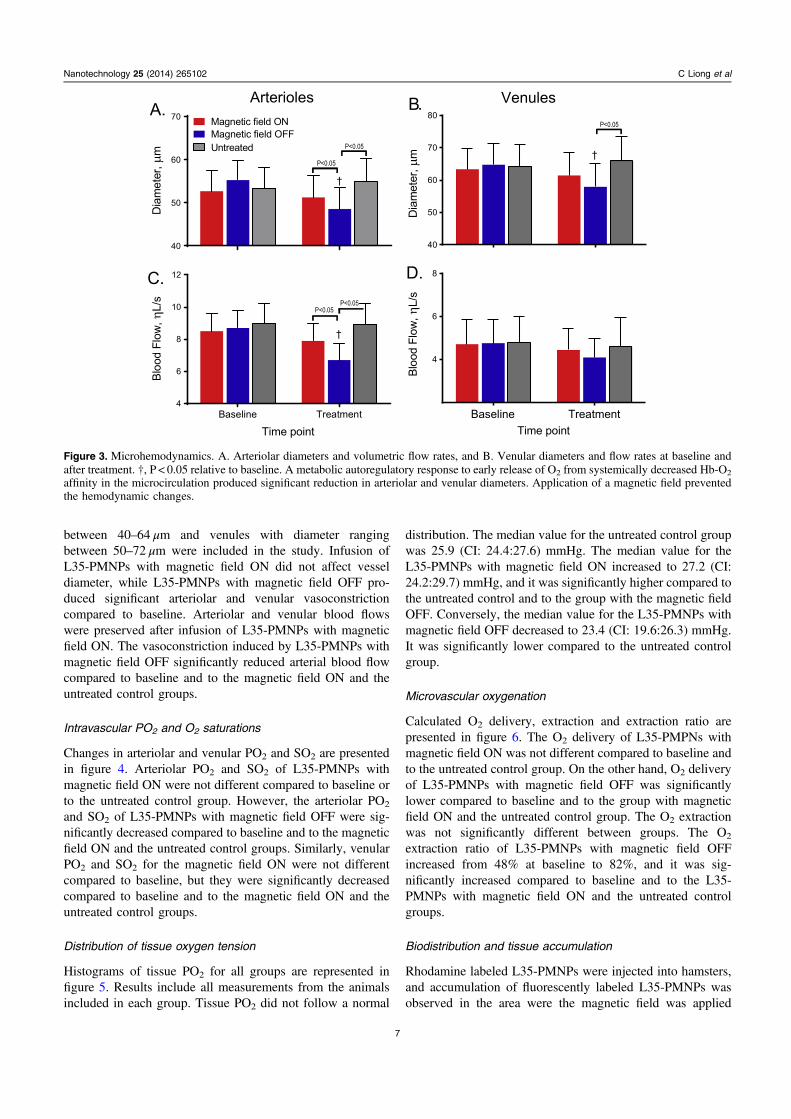
between 40–64 μm and venules with diameter rangingbetween 50–72 μm were included in the study. Infusion ofL35-PMNPs with magnetic field ON did not affect vesseldiameter, while L35-PMNPs with magnetic field OFF pro-duced significant arteriolar and venular vasoconstrictioncompared to baseline. Arteriolar and venular blood flowswere preserved after infusion of L35-PMNPs with magneticfield ON. The vasoconstriction induced by L35-PMNPs withmagnetic field OFF significantly reduced arterial blood flowcompared to baseline and to the magnetic field ON and theuntreated control groups.
Intravascular PO2 and O2 saturations
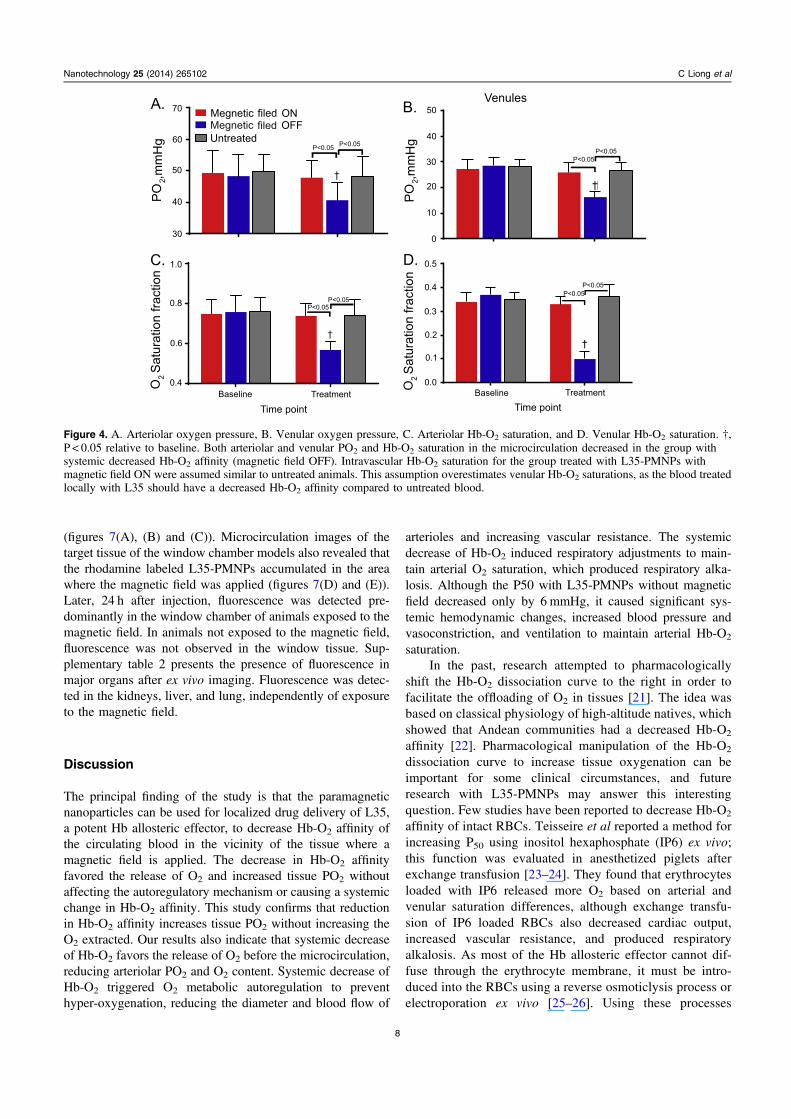
Changes in arteriolar and venular PO2 and SO2 are presentedin figure 4. Arteriolar PO2 and SO2 of L35-PMNPs withmagnetic field ON were not different compared to baseline orto the untreated control group. However, the arteriolar PO2
and SO2 of L35-PMNPs with magnetic field OFF were sig-nificantly decreased compared to baseline and to the magneticfield ON and the untreated control groups. Similarly, venularPO2 and SO2 for the magnetic field ON were not differentcompared to baseline, but they were significantly decreasedcompared to baseline and to the magnetic field ON and theuntreated control groups.
Distribution of tissue oxygen tension
Histograms of tissue PO2 for all groups are represented infigure 5. Results include all measurements from the animalsincluded in each group. Tissue PO2 did not follow a normal
distribution. The median value for the untreated control groupwas 25.9 (CI: 24.4:27.6) mmHg. The median value for theL35-PMNPs with magnetic field ON increased to 27.2 (CI:24.2:29.7) mmHg, and it was significantly higher compared tothe untreated control and to the group with the magnetic fieldOFF. Conversely, the median value for the L35-PMNPs withmagnetic field OFF decreased to 23.4 (CI: 19.6:26.3) mmHg.It was significantly lower compared to the untreated controlgroup.
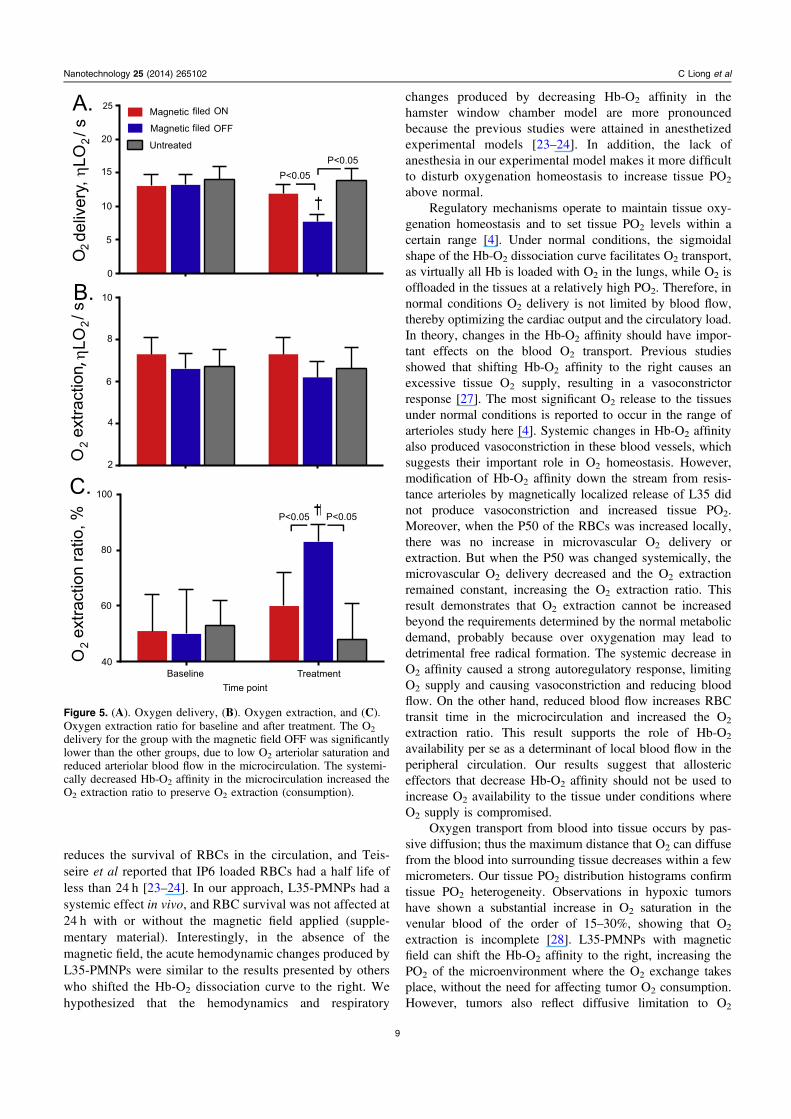
Microvascular oxygenation
Calculated O2 delivery, extraction and extraction ratio arepresented in figure 6. The O2 delivery of L35-PMPNs withmagnetic field ON was not different compared to baseline andto the untreated control group. On the other hand, O2 deliveryof L35-PMNPs with magnetic field OFF was significantlylower compared to baseline and to the group with magneticfield ON and the untreated control group. The O2 extractionwas not significantly different between groups. The O2
extraction ratio of L35-PMNPs with magnetic field OFFincreased from 48% at baseline to 82%, and it was sig-nificantly increased compared to baseline and to the L35-PMNPs with magnetic field ON and the untreated controlgroups.
Biodistribution and tissue accumulation
Rhodamine labeled L35-PMNPs were injected into hamsters,and accumulation of fluorescently labeled L35-PMNPs wasobserved in the area were the magnetic field was applied
Nanotechnology 25 (2014) 265102 C Liong et al
Figure 3. Microhemodynamics. A. Arteriolar diameters and volumetric flow rates, and B. Venular diameters and flow rates at baseline andafter treatment. †, P < 0.05 relative to baseline. A metabolic autoregulatory response to early release of O2 from systemically decreased Hb-O2
affinity in the microcirculation produced significant reduction in arteriolar and venular diameters. Application of a magnetic field preventedthe hemodynamic changes.
7
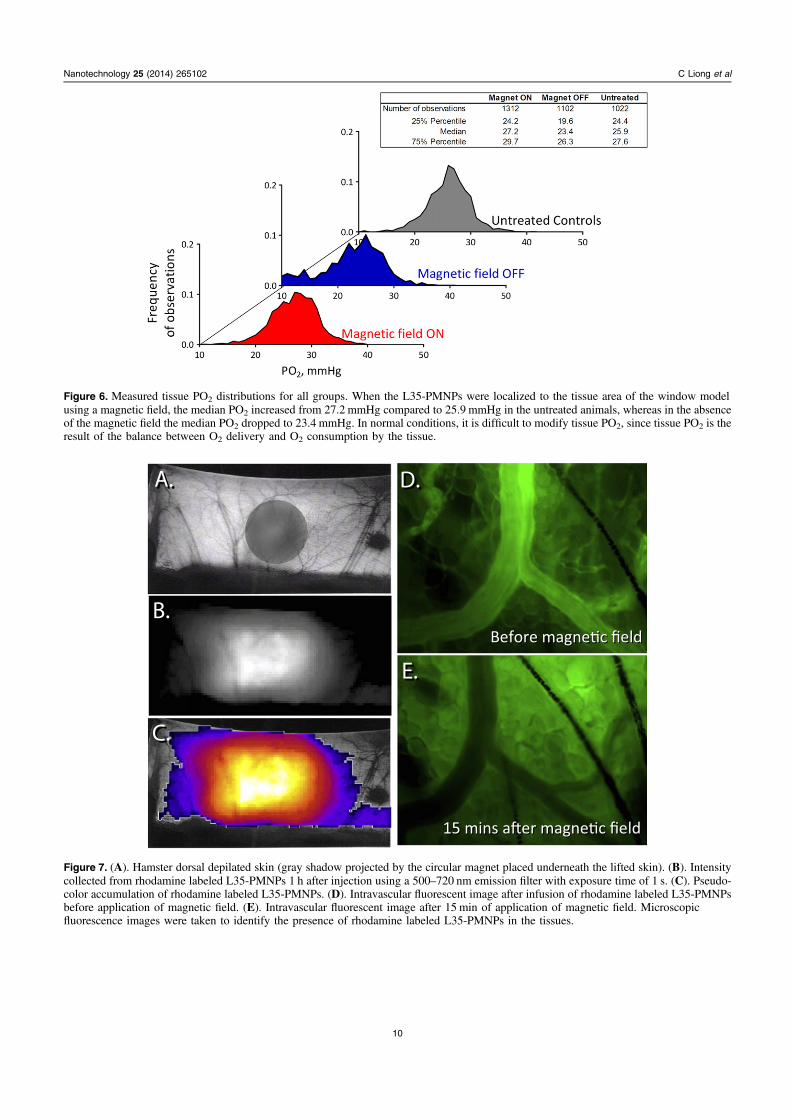
(figures 7(A), (B) and (C)). Microcirculation images of thetarget tissue of the window chamber models also revealed thatthe rhodamine labeled L35-PMNPs accumulated in the areawhere the magnetic field was applied (figures 7(D) and (E)).Later, 24 h after injection, fluorescence was detected pre-dominantly in the window chamber of animals exposed to themagnetic field. In animals not exposed to the magnetic field,fluorescence was not observed in the window tissue. Sup-plementary table 2 presents the presence of fluorescence inmajor organs after ex vivo imaging. Fluorescence was detec-ted in the kidneys, liver, and lung, independently of exposureto the magnetic field.
Discussion
The principal finding of the study is that the paramagneticnanoparticles can be used for localized drug delivery of L35,a potent Hb allosteric effector, to decrease Hb-O2 affinity ofthe circulating blood in the vicinity of the tissue where amagnetic field is applied. The decrease in Hb-O2 affinityfavored the release of O2 and increased tissue PO2 withoutaffecting the autoregulatory mechanism or causing a systemicchange in Hb-O2 affinity. This study confirms that reductionin Hb-O2 affinity increases tissue PO2 without increasing theO2 extracted. Our results also indicate that systemic decreaseof Hb-O2 favors the release of O2 before the microcirculation,reducing arteriolar PO2 and O2 content. Systemic decrease ofHb-O2 triggered O2 metabolic autoregulation to preventhyper-oxygenation, reducing the diameter and blood flow of
arterioles and increasing vascular resistance. The systemicdecrease of Hb-O2 induced respiratory adjustments to main-tain arterial O2 saturation, which produced respiratory alka-losis. Although the P50 with L35-PMNPs without magneticfield decreased only by 6 mmHg, it caused significant sys-temic hemodynamic changes, increased blood pressure andvasoconstriction, and ventilation to maintain arterial Hb-O2
saturation.In the past, research attempted to pharmacologically
shift the Hb-O2 dissociation curve to the right in order tofacilitate the offloading of O2 in tissues [21]. The idea wasbased on classical physiology of high-altitude natives, whichshowed that Andean communities had a decreased Hb-O2
affinity [22]. Pharmacological manipulation of the Hb-O2
dissociation curve to increase tissue oxygenation can beimportant for some clinical circumstances, and futureresearch with L35-PMNPs may answer this interestingquestion. Few studies have been reported to decrease Hb-O2
affinity of intact RBCs. Teisseire et al reported a method forincreasing P50 using inositol hexaphosphate (IP6) ex vivo;this function was evaluated in anesthetized piglets afterexchange transfusion [23–24]. They found that erythrocytesloaded with IP6 released more O2 based on arterial andvenular saturation differences, although exchange transfu-sion of IP6 loaded RBCs also decreased cardiac output,increased vascular resistance, and produced respiratoryalkalosis. As most of the Hb allosteric effector cannot dif-fuse through the erythrocyte membrane, it must be intro-duced into the RBCs using a reverse osmoticlysis process orelectroporation ex vivo [25–26]. Using these processes
Nanotechnology 25 (2014) 265102 C Liong et al
70
60
50
40
30
50
40
30
20
10
0
0.5
0.4
0.3
0.2
0.1
0.0
1.0
0.8
0.6
0.4Baseline Treatment Baseline Treatment
Time point Time pointO
2 Sat
urat
ion
fract
ion
O2 S
atur
atio
n fra
ctio
n
PO
2,mm
Hg
PO
2,mm
Hg
A.
D.
B.
C.
P<0.05 P<0.05
P<0.05P<0.05
P<0.05P<0.05
P<0.05P<0.05
VenulesMegnetic filed ONMegnetic filed OFFUntreated
Figure 4. A. Arteriolar oxygen pressure, B. Venular oxygen pressure, C. Arteriolar Hb-O2 saturation, and D. Venular Hb-O2 saturation. †,P < 0.05 relative to baseline. Both arteriolar and venular PO2 and Hb-O2 saturation in the microcirculation decreased in the group withsystemic decreased Hb-O2 affinity (magnetic field OFF). Intravascular Hb-O2 saturation for the group treated with L35-PMNPs withmagnetic field ON were assumed similar to untreated animals. This assumption overestimates venular Hb-O2 saturations, as the blood treatedlocally with L35 should have a decreased Hb-O2 affinity compared to untreated blood.
8
reduces the survival of RBCs in the circulation, and Teis-seire et al reported that IP6 loaded RBCs had a half life ofless than 24 h [23–24]. In our approach, L35-PMNPs had asystemic effect in vivo, and RBC survival was not affected at24 h with or without the magnetic field applied (supple-mentary material). Interestingly, in the absence of themagnetic field, the acute hemodynamic changes produced byL35-PMNPs were similar to the results presented by otherswho shifted the Hb-O2 dissociation curve to the right. Wehypothesized that the hemodynamics and respiratory
changes produced by decreasing Hb-O2 affinity in thehamster window chamber model are more pronouncedbecause the previous studies were attained in anesthetizedexperimental models [23–24]. In addition, the lack ofanesthesia in our experimental model makes it more difficultto disturb oxygenation homeostasis to increase tissue PO2
above normal.Regulatory mechanisms operate to maintain tissue oxy-
genation homeostasis and to set tissue PO2 levels within acertain range [4]. Under normal conditions, the sigmoidalshape of the Hb-O2 dissociation curve facilitates O2 transport,as virtually all Hb is loaded with O2 in the lungs, while O2 isoffloaded in the tissues at a relatively high PO2. Therefore, innormal conditions O2 delivery is not limited by blood flow,thereby optimizing the cardiac output and the circulatory load.In theory, changes in the Hb-O2 affinity should have impor-tant effects on the blood O2 transport. Previous studiesshowed that shifting Hb-O2 affinity to the right causes anexcessive tissue O2 supply, resulting in a vasoconstrictorresponse [27]. The most significant O2 release to the tissuesunder normal conditions is reported to occur in the range ofarterioles study here [4]. Systemic changes in Hb-O2 affinityalso produced vasoconstriction in these blood vessels, whichsuggests their important role in O2 homeostasis. However,modification of Hb-O2 affinity down the stream from resis-tance arterioles by magnetically localized release of L35 didnot produce vasoconstriction and increased tissue PO2.Moreover, when the P50 of the RBCs was increased locally,there was no increase in microvascular O2 delivery orextraction. But when the P50 was changed systemically, themicrovascular O2 delivery decreased and the O2 extractionremained constant, increasing the O2 extraction ratio. Thisresult demonstrates that O2 extraction cannot be increasedbeyond the requirements determined by the normal metabolicdemand, probably because over oxygenation may lead todetrimental free radical formation. The systemic decrease inO2 affinity caused a strong autoregulatory response, limitingO2 supply and causing vasoconstriction and reducing bloodflow. On the other hand, reduced blood flow increases RBCtransit time in the microcirculation and increased the O2
extraction ratio. This result supports the role of Hb-O2
availability per se as a determinant of local blood flow in theperipheral circulation. Our results suggest that allostericeffectors that decrease Hb-O2 affinity should not be used toincrease O2 availability to the tissue under conditions whereO2 supply is compromised.
Oxygen transport from blood into tissue occurs by pas-sive diffusion; thus the maximum distance that O2 can diffusefrom the blood into surrounding tissue decreases within a fewmicrometers. Our tissue PO2 distribution histograms confirmtissue PO2 heterogeneity. Observations in hypoxic tumorshave shown a substantial increase in O2 saturation in thevenular blood of the order of 15–30%, showing that O2
extraction is incomplete [28]. L35-PMNPs with magneticfield can shift the Hb-O2 affinity to the right, increasing thePO2 of the microenvironment where the O2 exchange takesplace, without the need for affecting tumor O2 consumption.However, tumors also reflect diffusive limitation to O2
Nanotechnology 25 (2014) 265102 C Liong et al
25
20
15
10
5
0
10
8
6
4
2
100
80
60
40Baseline
Time pointTreatment
P<0.05 P<0.05
P<0.05P<0.05
Magnetic filed ON
Magnetic OFF
Untreated
A.
B.
C.
filed
O2 d
eliv
ery,
ηLO
2/ s
O2
extra
ctio
nη
LO2/ s
O2
extra
ctio
n ra
tio, %
,
Figure 5. (A). Oxygen delivery, (B). Oxygen extraction, and (C).Oxygen extraction ratio for baseline and after treatment. The O2
delivery for the group with the magnetic field OFF was significantlylower than the other groups, due to low O2 arteriolar saturation andreduced arteriolar blood flow in the microcirculation. The systemi-cally decreased Hb-O2 affinity in the microcirculation increased theO2 extraction ratio to preserve O2 extraction (consumption).
9
Nanotechnology 25 (2014) 265102 C Liong et al
Figure 6. Measured tissue PO2 distributions for all groups. When the L35-PMNPs were localized to the tissue area of the window modelusing a magnetic field, the median PO2 increased from 27.2 mmHg compared to 25.9 mmHg in the untreated animals, whereas in the absenceof the magnetic field the median PO2 dropped to 23.4 mmHg. In normal conditions, it is difficult to modify tissue PO2, since tissue PO2 is theresult of the balance between O2 delivery and O2 consumption by the tissue.
Figure 7. (A). Hamster dorsal depilated skin (gray shadow projected by the circular magnet placed underneath the lifted skin). (B). Intensitycollected from rhodamine labeled L35-PMNPs 1 h after injection using a 500–720 nm emission filter with exposure time of 1 s. (C). Pseudo-color accumulation of rhodamine labeled L35-PMNPs. (D). Intravascular fluorescent image after infusion of rhodamine labeled L35-PMNPsbefore application of magnetic field. (E). Intravascular fluorescent image after 15 min of application of magnetic field. Microscopicfluorescence images were taken to identify the presence of rhodamine labeled L35-PMNPs in the tissues.
10

transport, even when the PO2 gradients are favorable for O2
release [28]. L35 is by far the most potent allosteric effectorof Hb ever described. At a concentration of 0.1 mm, itincreases the P50 of a suspension of RBCs by 50%; at 0.2 mmit raises the P50 by 2.5-fold [9]. L35-PMNPs might findclinical use to increase PO2 of hypoxic tissues or to tumorsbefore irradiation. However, the effects of L35 are acute, asL35 appears to be metabolized and loses its activity within24 h (see supplemental material).
Biodistribution and tissue accumulation study showedthe magnetic localization of the L35-PMNPs in the targettissue. Fluorescence microscopy revealed extravasation of thefluorescence from the labeled L35-PMNPs after application ofthe magnetic field, likely due to increased extra-cellularaccumulation of fluorophore conjugate to the L35-PMNPs(figures 7(C) and (D)). In vivo imaging studies found accu-mulation of fluorescence in the tissue of the window chamberwithin 15 min (figure 7(D)). In addition to the target tissue,fluorescence was also found in the liver, lung, spleen, andkidney, which can be attributed to PMNP uptake by theKupffer cells and macrophages. The kidney could be a targetbecause of its role in elimination of xenobiotics. Particle sizemay be a major factor in the accumulation of PMNP in var-ious organs. Studies on the biological effects of nanoparticleshave found similar results [29]. Application of a magneticfield after injection of PMNP increased the translocationprocess across biological barriers. The transport of PMNPsfrom the blood and their deposit in target organs can exertpotential toxic effects which needs to be studied in detail.Preliminary studies have shown that magnetic localization ofthe PMNPs is greater in tumor tissues by the combination ofmagnetic localization and leaky tumor vasculature. It can beanticipated that the magnet induced localization of the L35-PMNPs at a tumor site will produce a more profound increasein tumor PO2.
Conclusion
This study indicates that localized delivery of a pharmaceu-tical agent using nonmagnetic nanoparticles is possible; thehydrophobic nature of L35 localization with the PMNPfacilitates the target release of cell permeable drugs. The L35loaded PMNP was able to effectively modify the Hb-O2
systemically and also localized when a magnetic field wasapplied. Systemic modifications in Hb-O2 affinity triggeredthe autoregulatory mechanism, which hindered tissue oxy-genation. Localized decrease in Hb-O2 affinity did not acti-vate the regulatory mechanism and increased tissue PO2
without affecting O2 delivery and extraction. Therefore,localized decreases in Hb-O2 affinity only affect the PO2
gradient between the blood and the tissue, without affectingO2 utilization. There are significant clinical implications oflocalized increase in PO2; however, study of PMNP phar-macokinetic and long-term safety must be occur first.
Acknowledgments
This work was partially supported by BioengineeringResearch Partnership grant Program project P01-HL071064and grants R01-HL52684, R01-HL62354, and R01-HL62318. The authors thank F Barra and C Walser for thesurgical preparation of the animals. The authors also thankDrs. Iraj and Parviz Lalezari for generously gifting the L35used in the study.
References
[1] Tsai A G, Cabrales P, Hangai-Hoger N and Intaglietta M2004 Oxygen distribution and respiration by themicrocirculation Antioxidants & Redox Signaling 61011–8
[2] Jain R K 2013 Normalizing tumor microenvironment to treatcancer: bench to bedside to biomarkers J. Clin. Oncology 312205–18
[3] Yoshimura M, Itasaka S, Harada H and Hiraoka M 2012Microenvironment and radiation therapy BioMed. Res. Int.2013 685308
[4] Tsai A G, Johnson P C and Intaglietta M 2003 Oxygengradients in the microcirculation Physiol. Rev. 83 933–63
[5] Hsia C C 1998 Respiratory function of hemoglobin N. Engl. J.Med. 338 239–47
[6] Cabrales P, Tsai A G and Intaglietta M 2008 Modulation ofperfusion and oxygenation by red blood cell oxygenaffinity during acute anemia Am. J. Respir. Cell Mol.Biol. 38 354–61
[7] Villela N R, Cabrales P, Tsai A G and Intaglietta M 2009Microcirculatory effects of changing blood hemoglobinoxygen affinity during hemorrhagic shock resuscitation in anexperimental model Shock 31 645–52
[8] Weir E K, Lopez-Barneo J, Buckler K J and Archer S L 2005Acute oxygen-sensing mechanisms N. Engl. J. Med. 3532042–55
[9] Lalezari I, Lalezari P, Poyart C, Marden M, Kister J, Bohn B,Fermi G and Perutz M F 1990 New effectors of humanhemoglobin: structure and function Biochemistry 291515–23
[10] Chen Q, Lalezari I, Nagel R L and Hirsch R E 2005 Ligandedhemoglobin structural perturbations by the allosteric effectorL35 Biophys. J. 88 2057–67
[11] Lalezari I, Lalezari P, Poyart C, Marden M, Kister J, Bohn B,Fermi G and Perutz M 1990 New effectors of humanhemoglobin: structure and function Biochemistry 291515–23
[12] Soderlind F, Pedersen H, Petoral R M Jr, Kall P O and Uvdal K2005 Synthesis and characterisation of Gd2O3 nanocrystalsfunctionalised by organic acids J. Colloid Interface Sci. 288140–8
[13] Faucher L, Tremblay M, Lagueux J, Gossuin Y andFortin M A 2012 Rapid synthesis of PEGylated ultrasmallgadolinium oxide nanoparticles for cell labeling andtracking with MRI ACS Appl. Mater. Interfaces 44506–15
[14] Colantuoni A, Bertuglia S and Intaglietta M 1984 Quantitationof rhythmic diameter changes in arterial microcirculationAm. J. Physiol. 246 H508–17
[15] Endrich B, Asaishi K, Götz A and Messmer K 1980 Technicalreport: a new chamber technique for microvascular studies inunanaesthetized hamsters Res. Exp. Med. 177125–34
Nanotechnology 25 (2014) 265102 C Liong et al
11

[16] Cabrales P, Tsai A G, Frangos J A and Intaglietta M 2005 Roleof endothelial nitric oxide in microvascular oxygen deliveryand consumption Free Radic Biol. Med. 39 1229–37
[17] Intaglietta M and Tompkins W R 1973 Microvascularmeasurements by video image shearing and splittingMicrovasc. Res. 5 309–12
[18] Lipowsky H H and Zweifach B W 1978 Application of the‘two-slit’ photometric technique to the measurement ofmicrovascular volumetric flow rates Microvasc. Res. 1593–101
[19] Kerger H, Groth G, Kalenka A, Vajkoczy P, Tsai A G andIntaglietta M 2003 pO2 measurements by phosphorescencequenching: characteristics and applications of an automatedsystem Microvasc. Res. 65 32–8
[20] Vanderkooi J M, Maniara G, Green T J and Wilson D F 1987An optical method for measurement of dioxygenconcentration based upon quenching of phosphorescenceJ. Biol. Chem. 262 5476–82
[21] Kunert M P, Liard J F, Abraham D J and Lombard J H 1996Low-affinity hemoglobin increases tissue PO2 and decreasesarteriolar diameter and flow in the rat cremaster muscleMicrovasc. Res. 52 58–68
[22] Reynafarje C and Hurtado A 1971 Adaptation to high altitudeN. Engl. J. Med. 285 862
[23] Benesch R, Edalji R and Benesch R E 1976 The allostericeffect of inositol hexasulfate on oxygen binding byhemoglobin Biochemistry 15 3396–8
[24] Gibson Q H and Gray R D 1970 The reaction of inositolhexaphosphate with hemoglobin Biochem. Biophys. Res.Commun. 41 415–20
[25] Mouneimne Y, Barhoumi R, Myers T, Slogoff S and Nicolau C1990 Stable rightward shifts of the oxyhemoglobin dissociationcurve induced by encapsulation of inositol hexaphosphate in redblood cells using electroporation FEBS Lett. 275 117–20
[26] Bruggemann U, Roux E C, Hannig J and Nicolau C 1995 Low-oxygen-affinity red cells produced in a large-volume,continuous-flow electroporation system Transfusion 35478–86
[27] Ledingham J M 1989 Autoregulation in hypertension: a reviewJ. Hypertens Suppl. 7 S97–104
[28] Helmlinger G, Sckell A, Dellian M, Forbes N S and Jain R K2002 Acid production in glycolysis-impaired tumorsprovides new insights into tumor metabolism Clin. Cancer.Res. 8 1284–91
[29] Sayes C M, Reed K L and Warheit D B 2007 Assessingtoxicity of fine and nanoparticles: comparing in vitromeasurements to in vivo pulmonary toxicity profiles Toxicol.Sci. 97 163–80
Nanotechnology 25 (2014) 265102 C Liong et al
12
Top Related
Copyright © 2022 FDOKUMEN

