







Bahasa
Halaman
Hukum


Voiding Dysfunction
In Vitro Release of Adenosine Triphosphate from the Urothelium
of Human Bladders with Detrusor Overactivity, Both Neurogenic
and Idiopathic
Vivek Kumar a,*, Christopher R. Chapple b, Derek Rosario c, Paul R. Tophill d,Russell Chess-Williams e
a Department of Reconstructive Urology, Royal Hallamshire Hospital, Sheffield, United Kingdomb Royal Hallamshire Hospital, University of Sheffield and Sheffield Hallam University, Sheffield, United Kingdomc Royal Hallamshire Hospital, University of Sheffield, Sheffield, United Kingdomd Princess Royal Spinal Injury Unit, Northern General Hospital, Sheffield, United Kingdome Faculty of Health Sciences and Medicine, Bond University, Queensland, Australia
E U R O P E A N U R O L O G Y 5 7 ( 2 0 1 0 ) 1 0 8 7 – 1 0 9 2
ava i lable at www.sciencedirect .com
journal homepage: www.europeanurology.com
Article info
Article history:
Accepted November 25, 2009Published online ahead ofprint on December 3, 2009
Keywords:
ATP
OAB
Purinergic neurotransmission
Purinergic receptors
Neurogenic bladder
Abstract
Background: There is increased evidence to suggest a role for nonadrenergic–noncho-
linergic neurotransmission in the pathogenesis of bladder dysfunction.
Objective: In this set of experiments, we have assessed the contribution of the urothelium
to purinergic activity by quantifying the amount of adenosine triphosphate (ATP) released
from the urothelium of patients with idiopathic detrusor overactivity (IDO) and with
neurogenic detrusor overactivity (NDO) and comparing these releases to those of controls.
Design, setting, and participants: Bladder tissue with urodynamically and clinically
proven NDO (n = 8) and IDO (n = 8) were included in this study. The carefully dissected
urothelium was stimulated by mechanically stretching as well as electrically stimulating
and the ATP; thus, release was quantified.
Measurements: We used a Lucy Anthos 1 luminometre (Anthos Labtec Instruments
GmBH, Wals, Austria) to perform the assay. The results were analysed using Stingray
software (Dazdaq Ltd, Brighton, UK).
Results and limitations: Both mechanical stretch and electric field stimulation (EFS)
led to increased ATP release in both sets of tissues with overactivity compared to
the controls; this rise was even more significant for the IDO urothelium (2416.7 �479.8 pmol/g [p < 0.005]) than for the NDO urothelium (133.1 � 22.4 pmol/g [p < 0.01]);
values for the controls were 77.6 � 16.2 pmol/g. ATP release following mechanical stretch
was more sensitive to tetrodotoxin in bladders with NDO compared to those with IDO as
well as to the controls, with ATP levels falling from 233.5 � 20.7 pmol/g to 107.2 � 11.6
pmol/g, expressed as percentage of basal levels ( p < 0.002). The experiments were
performed in vitro, and the female patients were a mix of peri- and postmenopausal states.
Conclusions: These experiments suggested a significant rise in ATP release from the
urothelium of bladders with NDO as well as those with IDO in comparison to controls.
Most of the ATP released from bladders with NDO is primarily from neuronal sources.
# 2009 European Association of Urology. Published by Elsevier B.V. All rights reserved.
* Corresponding author. Department of Reconstructive Urology, Royal Hallamshire Hospital,Glossop Road, Sheffield, South Yorkshire, S10 2JF, UK.
E-mail address: [email protected] (V. Kumar).0302-2838/$ – see back matter # 2009 European Association of Urology. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.eururo.2009.11.042

E U R O P E A N U R O L O G Y 5 7 ( 2 0 1 0 ) 1 0 8 7 – 1 0 9 21088
1. Introduction
Overactive bladder (OAB) is a storage symptom complex
that refers to the symptoms of urinary urgency associated
with incontinence in up to one-third of cases, usually with
increased urinary frequency and nocturia [1]. Prevalence
rates range between 12% and 17% in North America and
Europe and are comparable among men and women [2–4].
OAB symptoms are suggestive of urodynamically de-
monstrable detrusor overactivity (DO) that may be sponta-
neous or provoked. DO may be further characterised as
neurogenic DO (NDO) or idiopathic DO (IDO). Common
neurogenic causes include stroke, Parkinson’s disease,
multiple sclerosis (MS), and spinal injury. Thus, the term
is applied to a wide range of conditions that may have a
common final pathophysiologic pathway [5].
The suburothelial layer of the bladder is rich in sensory
(afferent) nerves [6,7], and increased sensitivity of these
suburothelial sensory nerves may trigger the overactive
detrusor contraction. In 1997, Ferguson and associates [8]
demonstrated distension-evoked adenosine triphosphate
(ATP) release from the bladder urothelium. Since then,
evidence has been accumulating to support a role for
urothelial-derived ATP release in both autocrine and
paracrine signalling [9]. Furthermore, the purinergic
receptors (ionotropic purinergic receptor [P2X] and meta-
botropic purinergic receptors [P2Y]) are expressed in
various types of cells located at or near the urothelium of
the urinary bladder [10]. Local inflammation and stimula-
tion by the P2X receptor agonist can lead to increased
bladder afferent firing [11]. Thus, the stretch-evoked,
urothelial-derived ATP may stimulate the local sensory
mechanism, triggering sensations of fullness and pain that
induce changes in bladder activity [12].
When the bladder is stretched during the filling phase, it
releases several neurotransmitters that may act on recep-
tors present on the sensory nerves in the suburothelium,
thereby conveying information to the central nervous
system and initiating voiding. ATP is one such putative
neurotransmitter that contributes to the bulk of the
nonadrenergic–noncholinergic (NANC) activity, which is
responsible for the atropine-resistant detrusor contraction.
Whereas this NANC contribution in a normal human
bladder contraction is small [13], it can become significant
in functionally abnormal and aging bladders [14].
In this set of experiments, we assessed the contribution
of the urothelium to purinergic activity by quantifying the
amount of ATP released from the urothelium of patients
with IDO and NDO and comparing these releases to those of
controls.
2. Patients and methods
The experiments were performed using tissue from the dome of human
urinary bladder. Multiple full-thickness bladder tissues were extracted
from patients with urodynamically and clinically proven NDO (n = 8)
undergoing clam ileocystoplasty at the Spinal Injury Unit, Northern
General Hospital, Sheffield, United Kingdom. Those with MS (n = 6 [all
women]) and suprasacral spinal cord injury (n = 2 [one man]) were
included in this study (age range: 32–46 yr [mean: 37.5 yr]). In addition,
eight women (age range: 36–45 yr [mean: 41.5 yr]) who were clinically
and urodynamically diagnosed with IDO were also included in this
study. These patients had refractory symptoms of urinary frequency,
urgency, and urge incontinence along with DO demonstrated on video
urodynamics and had the full-thickness tissue samples taken at the time
of their clam cystoplasty.
Urothelium and suburothelial tissue were macroscopically dissected
off the samples for purposes of the experiments. The mean maximum
cystometric capacity (plus or minus standard error) of the patients with
NDO was 283 � 40 ml (range: 255–375 ml); for those patients with IDO,
capacity was 315 � 55 ml (range: 230–420 ml). Control bladders were
obtained from patients with urodynamically proven stable bladders
undergoing surgery for stress urinary incontinence (n = 4) using a cold
cup biopsy forceps (5 mm) and from patients undergoing cystectomy for
cancer (full-thickness sample) who had no history of receiving radiation
therapy and no lower urinary tract symptoms that would suggest a
functional bladder disorder (n = 5). The tissue was taken from a site distant
to the tumour from a normal-looking bladder area. The mucosa was
again macroscopically dissected off the tissues and immediately placed in
ice-cold Krebs bicarbonate solution (composition: sodium chloride,
118.4 mM/l; sodium bicarbonate, 24.9 mM/l; potassium chloride,
4.7 mM/l; calcium chloride, 21.9 mM/l; magnesium sulphate, 1.15 mM/l;
potassium dihydrogen phosphate, 1.15 mM/l; glucose 11.7 mM/l). All of the
tissues were obtained with full informed consent and with approval from
the local ethics committee.
Four tissue strips from each patient (5 � 3 mm) were placed between
two platinum-wire loop electrodes; using suture silk, the bottom end of
the tissues were secured to a tissue holder. The upper ends were
attached (with suture silk) to the UFI model 1030 transducer (UFI, Morro
Bay, CA, USA). The transducers were connected to the NeuroLog data
acquisition system (Digitimer, Welwyn Garden City, UK), allowing
changes in the tension to be recorded. Tissues were placed under 1 g of
constant resting tension in a 2.5-ml organ bath containing Krebs
bicarbonate solution at 37 8C and gassed with a mixture of 95% oxygen
and 5% carbon dioxide for 1 h, with washes at 15-min intervals.
The urothelium was mechanically stretched to 130%, and then,
following a period of rest (resting tension of 1 g), to a further 150% of the
original length. Samples (50 ml) were taken following the stretch for
quantification of ATP release. The tissues were also subjected to electric
field stimulation (EFS) at 10 Hz (40 V, 0.2 ms pulse width for 60 s) using a
Digitimer PowerStim stimulator, and ATP release was quantified using
luminometry. The ATP reagent HS luciferin–luciferase enzyme used was
obtained from BioTherma (BioTherma Luminescent Assays, Haninge,
Sweden). A Lucy Anthos 1 luminometre (Anthos Labtec Instruments
GmBH, Wals, Austria) was used to perform the assay, and the results
were analysed using Stingray software (Dazdaq Ltd, Brighton, UK).
Tetrodotoxin (5 mM) exposure to the urothelium for 20 min was used to
block the neuronal source of ATP release, thereby enabling us to measure
the non-neuronal component.
All of the tissues were weighed at the end of each experiment, and
ATP levels were calculated, as picomole per gram of tissue, as well as
relative rise, expressed as percentage of the basal level. Statistical
analysis was performed using paired t test.
3. Results
The basal release of ATP from the urothelium of control and
NDO bladders was similar (Table 1). Mechanical stretch led
to increased levels of ATP in both sets of tissues, but this
increase was significantly greater for the NDO tissues
(Table 1; Fig. 1). EFS also elicited a higher level of ATP; again,
the magnitude of this response was greater in the urothelial

Table 1 – The amount of adenosine triphosphate released, in picomoles per gram of tissue, from idiopathic and neurogenic detrusoroveractive urothelium with mechanical stretch (130% and 150%) and electric field stimulation (10 Hz)
ATP release with mechanical stretch (pmol/g tissue � SEM) ATP release with electrical stimulation(pmol/g tissue � SEM)
Baseline 130% 150% Baseline 10 Hz
Control urothelium (n = 9) 45.7 � 4.9 67.5 � 13.2 77.6 � 16.2* 45.8 � 5.1 61.7 � 10.1*
IDO urothelium (n = 8) 1064.2 � 238.9z 2103.2 � 456z* 2416.7 � 479.8z* 578.7 � 62.9z 1654.5 � 177.5z*NDO urothelium (n = 8) 57.0 � 7.9y 102.3 � 16.9* 133.1 � 22.4* 56.5 � 11.6y 124.5 � 20.7z*
ATP = adenosine triphosphate; SEM = standard error of the mean; IDO = idiopathic detrusor overactivity; NDO = neurogenic detrusor overactivity.
* p < 0.01 compared to the baseline ATP release.
y p is not significant, compared with control urothelium.
z p < 0.005 compared with control urothelium.
Table 2 – The effect of 5 mM of tetrodotoxin on the urothelium from neurogenic and idiopathic detrusor overactive bladders comparedwith normal urothelium**
ATP release with mechanical stretch(150% elongation) (% of baseline � SEM)
ATP release with electrical stimulation(10 Hz) (% of baseline � SEM)
TTX absent TTX present TTX absent TTX present
Control urothelium (n = 9) 175.4 � 21.7 147.4 � 20.1 137.9 � 4.4 30.2 � 2.2
IDO urothelium (n = 8) 227.1 � 17.9z 209.9 � 16.7z 286.6 � 33.5z 159.8 � 7.0*zNDO urothelium (n = 8) 233.5 � 20.7z 107.2 � 11.6* 221.8 � 32.4z 38.3 � 9.6*
ATP = adenosine triphosphate; SEM = standard error of the mean; TTX = tetrodotoxin; IDO = idiopathic detrusor overactivity; NDO = neurogenic detrusor
overactivity.
* p = 0.002 compared with TTX absent.
z p < 0.02 compared with control urothelium.
** The amount of ATP released is expressed as a percentage of the basal levels.
E U R O P E A N U R O L O G Y 5 7 ( 2 0 1 0 ) 1 0 8 7 – 1 0 9 2 1089
tissues taken from patients with NDO (Table 1). These
increases in ATP release were statistically significant,
whether expressed in absolute values (Table 1) or as a
percentage of the basal levels (Table 2).
In contrast, the basal release of ATP from bladders with
IDO without any stimulation was remarkably raised com-
pared to both the control and NDO bladders. In addition, this
urothelium showed significantly greater ATP release with
both mechanical stretch (130% and 150% stretch) as well as
EFS (10 Hz) when compared to the controls (Tables 1 and 2;
Fig. 2).
The ATP release from the urothelium of both NDO
bladders and controls following EFS showed a marked
sensitivity to the neurotoxin tetrodotoxin ( p = 0.002).
Fig. 1 – Amounts of adenosine triphosphate (ATP) released withmechanical stretch of the urothelium from bladders with neurogenicdetrusor overactivity (NDO) compared to controls following a basalrelease at a resting tension of 1 g.
The level of ATP release fell by 83% in NDO bladders and
by 78% in normal bladders following exposure to tetrodo-
toxin at a concentration of 5 mM (Table 2), suggesting that
EFS results in primarily neuronal ATP release. In compari-
son, the urothelial ATP release following mechanical
stretch, which may be more physiologic, was more sensitive
to tetrodotoxin in bladders with NDO compared to controls.
Tetrodotoxin reduced stretch-induced ATP release by 54% in
urothelium from bladders with NDO ( p = 0.005) and by only
16% in urothelium from controls.
Similarly, the ATP release from the urothelium of IDO
bladders following EFS showed a marked sensitivity to
tetrodotoxin ( p < 0.05), resulting in an overall reduction
of ATP levels by 45% when exposed to tetrodotoxin
(Table 2). This again confirmed that EFS predominantly
results in neuronally released ATP. However, the ATP
release following mechanical stretch of IDO bladders was
not sensitive to tetrodotoxin, with a small but similar fall
to that of the controls in the level of ATP release (8%
for idiopathic overactive bladders [p = not significant
(ns)] and 16% for controls [p = ns]) from the basal level
(Table 2).
4. Discussion
Brading and Turner [15] have emphasised that basic
myogenic changes (regardless of aetiology) underlie the
pathophysiology of DO. Furthermore, evidence of denerva-
tion is consistently found in detrusor biopsy specimens
from patients with various forms of DO [16,17], suggesting
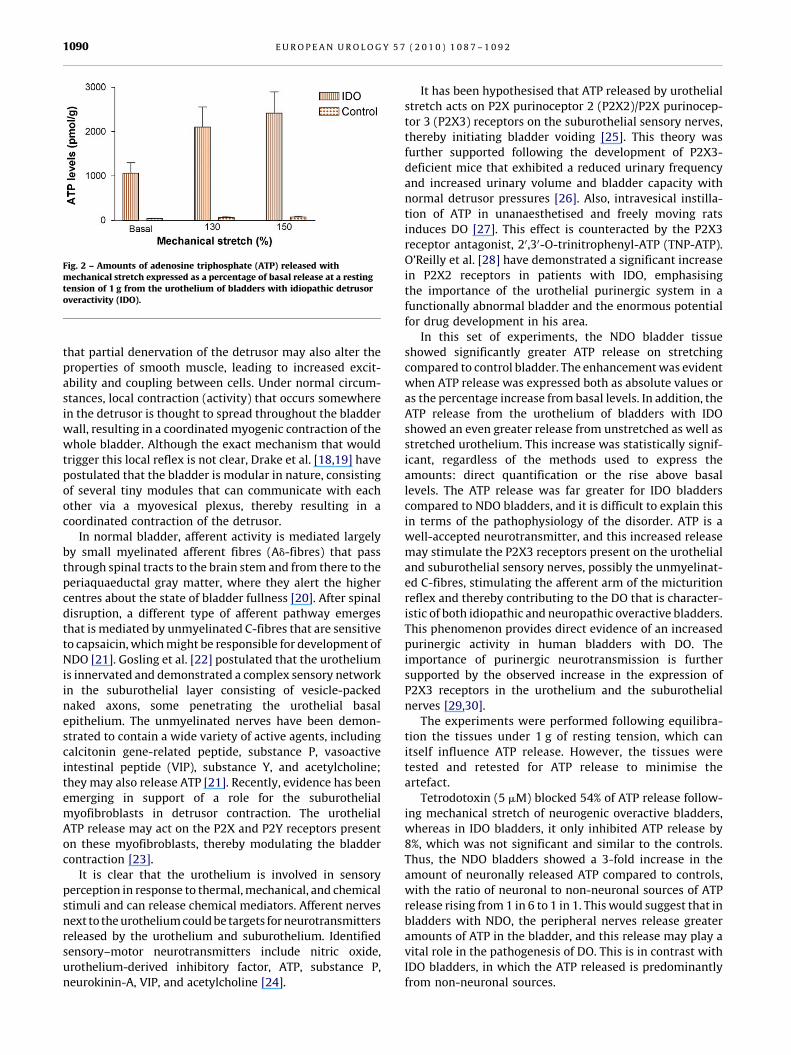
Fig. 2 – Amounts of adenosine triphosphate (ATP) released withmechanical stretch expressed as a percentage of basal release at a restingtension of 1 g from the urothelium of bladders with idiopathic detrusoroveractivity (IDO).
E U R O P E A N U R O L O G Y 5 7 ( 2 0 1 0 ) 1 0 8 7 – 1 0 9 21090
that partial denervation of the detrusor may also alter the
properties of smooth muscle, leading to increased excit-
ability and coupling between cells. Under normal circum-
stances, local contraction (activity) that occurs somewhere
in the detrusor is thought to spread throughout the bladder
wall, resulting in a coordinated myogenic contraction of the
whole bladder. Although the exact mechanism that would
trigger this local reflex is not clear, Drake et al. [18,19] have
postulated that the bladder is modular in nature, consisting
of several tiny modules that can communicate with each
other via a myovesical plexus, thereby resulting in a
coordinated contraction of the detrusor.
In normal bladder, afferent activity is mediated largely
by small myelinated afferent fibres (Ad-fibres) that pass
through spinal tracts to the brain stem and from there to the
periaquaeductal gray matter, where they alert the higher
centres about the state of bladder fullness [20]. After spinal
disruption, a different type of afferent pathway emerges
that is mediated by unmyelinated C-fibres that are sensitive
to capsaicin, which might be responsible for development of
NDO [21]. Gosling et al. [22] postulated that the urothelium
is innervated and demonstrated a complex sensory network
in the suburothelial layer consisting of vesicle-packed
naked axons, some penetrating the urothelial basal
epithelium. The unmyelinated nerves have been demon-
strated to contain a wide variety of active agents, including
calcitonin gene-related peptide, substance P, vasoactive
intestinal peptide (VIP), substance Y, and acetylcholine;
they may also release ATP [21]. Recently, evidence has been
emerging in support of a role for the suburothelial
myofibroblasts in detrusor contraction. The urothelial
ATP release may act on the P2X and P2Y receptors present
on these myofibroblasts, thereby modulating the bladder
contraction [23].
It is clear that the urothelium is involved in sensory
perception in response to thermal, mechanical, and chemical
stimuli and can release chemical mediators. Afferent nerves
next to the urothelium could be targets for neurotransmitters
released by the urothelium and suburothelium. Identified
sensory–motor neurotransmitters include nitric oxide,
urothelium-derived inhibitory factor, ATP, substance P,
neurokinin-A, VIP, and acetylcholine [24].
It has been hypothesised that ATP released by urothelial
stretch acts on P2X purinoceptor 2 (P2X2)/P2X purinocep-
tor 3 (P2X3) receptors on the suburothelial sensory nerves,
thereby initiating bladder voiding [25]. This theory was
further supported following the development of P2X3-
deficient mice that exhibited a reduced urinary frequency
and increased urinary volume and bladder capacity with
normal detrusor pressures [26]. Also, intravesical instilla-
tion of ATP in unanaesthetised and freely moving rats
induces DO [27]. This effect is counteracted by the P2X3
receptor antagonist, 20,30-O-trinitrophenyl-ATP (TNP-ATP).
O’Reilly et al. [28] have demonstrated a significant increase
in P2X2 receptors in patients with IDO, emphasising
the importance of the urothelial purinergic system in a
functionally abnormal bladder and the enormous potential
for drug development in his area.
In this set of experiments, the NDO bladder tissue
showed significantly greater ATP release on stretching
compared to control bladder. The enhancement was evident
when ATP release was expressed both as absolute values or
as the percentage increase from basal levels. In addition, the
ATP release from the urothelium of bladders with IDO
showed an even greater release from unstretched as well as
stretched urothelium. This increase was statistically signif-
icant, regardless of the methods used to express the
amounts: direct quantification or the rise above basal
levels. The ATP release was far greater for IDO bladders
compared to NDO bladders, and it is difficult to explain this
in terms of the pathophysiology of the disorder. ATP is a
well-accepted neurotransmitter, and this increased release
may stimulate the P2X3 receptors present on the urothelial
and suburothelial sensory nerves, possibly the unmyelinat-
ed C-fibres, stimulating the afferent arm of the micturition
reflex and thereby contributing to the DO that is character-
istic of both idiopathic and neuropathic overactive bladders.
This phenomenon provides direct evidence of an increased
purinergic activity in human bladders with DO. The
importance of purinergic neurotransmission is further
supported by the observed increase in the expression of
P2X3 receptors in the urothelium and the suburothelial
nerves [29,30].
The experiments were performed following equilibra-
tion the tissues under 1 g of resting tension, which can
itself influence ATP release. However, the tissues were
tested and retested for ATP release to minimise the
artefact.
Tetrodotoxin (5 mM) blocked 54% of ATP release follow-
ing mechanical stretch of neurogenic overactive bladders,
whereas in IDO bladders, it only inhibited ATP release by
8%, which was not significant and similar to the controls.
Thus, the NDO bladders showed a 3-fold increase in the
amount of neuronally released ATP compared to controls,
with the ratio of neuronal to non-neuronal sources of ATP
release rising from 1 in 6 to 1 in 1. This would suggest that in
bladders with NDO, the peripheral nerves release greater
amounts of ATP in the bladder, and this release may play a
vital role in the pathogenesis of DO. This is in contrast with
IDO bladders, in which the ATP released is predominantly
from non-neuronal sources.

E U R O P E A N U R O L O G Y 5 7 ( 2 0 1 0 ) 1 0 8 7 – 1 0 9 2 1091
5. Conclusions
These experiments suggest an increased purinergic
activity in the form of a significant rise in ATP release
from the urothelium of bladders with NDO as well as
those with IDO compared to controls. Most of the ATP
released from bladders with NDO is primarily from
neuronal sources. This observation further supports the
view that the unmyelinated C-fibres in the suburothelium
take up a more prominent role in bladders with NDO. In
bladders with IDO, ATP is being released from non-
neuronal sources, and this increased ATP release may act
as a sensory neurotransmitter and stimulate the afferent
arm of the micturition reflex, leading to a symptomatic
bladder with DO.
Author contributions: Vivek Kumar had full access to all the data in the
study and takes responsibility for the integrity of the data and the
accuracy of the data analysis
Study concept and design: Chess-Williams.
Acquisition of data: Kumar.
Analysis and interpretation of data: Kumar.
Drafting of the manuscript: Kumar, Chapple.
Critical revision of the manuscript for important intellectual content:
Chapple, Chess-Williams.
Statistical analysis: Kumar.
Obtaining funding: None.
Administrative, technical, or material support: Rosario, Tophill.
Supervision: Chapple.
Other (specify): None.
Financial disclosures: I certify that all conflicts of interest, including
specific financial interests and relationships and affiliations relevant
to the subject matter or materials discussed in the manuscript
(eg, employment/affiliation, grants or funding, consultancies, honoraria,
stock ownership or options, expert testimony, royalties, or patents filed,
received, or pending), are the following: None
Funding/Support and role of the sponsor: None.
Acknowledgement statement: The authors acknowledge Professor Ann-
Marie Surprenant of the Biomedical Department, University of Sheffield,
for her support and guidance.
References
[1] Abrams P, Cardozo L, Fall M, et al. The standardisation of terminol-
ogy of lower urinary tract function: report from the Standardisation
Sub-committee of the International Continence Society. Neurourol
Urodyn 2002;21:167–78.
[2] Milsom I, Abrams P, Cardozo L, Roberts RG, Thuroff J, Wein AJ. How
widespread are the symptoms of an overactive bladder and how are
they managed? A population-based prevalence study. BJU Int 2001;
87:760–6.
[3] Stewart WF, Van Rooyen JB, Cundiff GW, et al. Prevalence and
burden of overactive bladder in the United States. World J Urol
2003;20:327–36.
[4] Irwin DE, Milsom I, Hunskaar S, et al. Population-based survey of
urinary incontinence, overactive bladder, and other lower urinary
tract symptoms in five countries: results of the EPIC study. Eur Urol
2006;50:1306–15.
[5] Artibani W. Diagnosis and significance of idiopathic overactive
bladder. Urology 1997;50(Suppl 6A):25–32, discussion 33–5.
[6] Dixon JS, Gilpin CJ. Presumptive sensory axons of the
human urinary bladder: a fine structural study. J Anat 1987;151:
199–207.
[7] Gabella G, Davis C. Distribution of afferent axons in the bladder of
rats. J Neurocytol 1998;27:141–55.
[8] Ferguson DR, Kennedy I, Burton TJ. ATP is released from rabbit
urinary bladder epithelial cells by hydrostatic pressure changes-a
possible sensory mechanism? J Physiol 1997;505:503–11.
[9] Hanna-Mitchell AT, Birder LA. New insights into the pharmacology
of the bladder. Curr Opin Urol 2008;18:347–52.
[10] Burnstock G. Physiology and pathophysiology of purinergic neuro-
transmission. Physiol Rev 2007;87:659–797.
[11] Yu Y, de Groat WC. Sensitization of pelvic afferent nerves in the in
vitro rat urinary bladder-pelvic nerve preparation by purinergic
agonists and cyclophosphamide pretreatment. Am J Physiol Renal
Physiol 2008;294:F1146–56.
[12] Gur S, Kadowitz PJ, Hellstrom WJ. Purinergic (P2) receptor control
of lower genitourinary tract function and new avenues for drug
action: an overview. Curr Pharm Des 2007;13:3236–44.
[13] Ambache N, Aboo Zar MA. Non cholinergic transmission by post
ganglionic motor neurones in the mammalian bladder. J Physiol
1970;210:761–83.
[14] Palea S, Artibani W, Ostardo E, Trist DG, Pietra C. Evidence for
purinergic neurotransmission in human urinary bladder affected
by interstitial cystitis. J Urol 1993;150:2007–12.
[15] Brading AF, Turner WH. The unstable bladder: towards a common
mechanism. Br J Urol 1993;73:3–8.
[16] Drake MJ, Hedlund P, Mills IW, et al. Structural and functional
denervation of human detrusor after spinal cord injury. Lab Invest
2000;80:1491–9.
[17] Mills IW, Greenland JE, McMurray R, et al. Studies of the patho-
physiology of idiopathic detrusor instability: the physiological
properties of the detrusor smooth muscle and its pattern of inner-
vation. J Urol 2000;163:646–51.
[18] Drake MJ, Mills IW, Gillespie JI. Model of peripheral autonomous
modules and a myovesical plexus in normal and overactive bladder
function. Lancet 2001;358:401–3.
[19] Drake MJ, Harvey IJ, Gillespie JI, Van Duyl WA. Localised modular
contractions in the normal human bladder and in urinary urgency.
BJU Int 2005;95:1002–5.
[20] Blok BFM. Central pathways controlling micturition and urinary
continence. Urology 2002;59:13–7.
[21] Fowler CJ. Bladder afferents and their role in the overactive bladder.
Urology 2002;59(Suppl 1):37–42.
[22] Gosling JA, Dixon JS, Humpherson JA. Functional anatomy of the
urinary tract: an integrated text and colour atlas. Edinburgh, UK:
Churchill Livingstone; 1983.
[23] Fry CH, Sui GP, Kanai AJ, Wu C. The function of suburothelial
myofibroblasts in the bladder. Neurourol Urodyn 2007;26(Suppl
6):914–9.
[24] Yoshimura N. Lower urinary tract symptoms (LUTS) and bladder
afferent activity. Neurourol Urodyn 2007;26(Suppl 6): 908–13.
[25] Burnstock G. Release of vasoactive substances from endothelial
cells by shear stress and purinergic mechanosensory transduction.
J Anat 1999;194:335–42.
[26] Cockayne DA, Hamilton SG, Zhu QM, et al. Urinary bladder hypore-
flexia and reduced pain-related behaviour in P2X3-deficient mice.
Nature 2000;407:1011–5.
[27] Pandita RK, Andersson KE. Intravesical adenosine triphosphate
stimulates the micturition reflex in awake, freely moving rats.
J Urol 2002;168:1230–4.

E U R O P E A N U R O L O G Y 5 7 ( 2 0 1 0 ) 1 0 8 7 – 1 0 9 21092
[28] O’Reilly BA, Kosaka AH, Knight GF, et al. P2X receptors and their role
in female idiopathic detrusor instability. J Urol 2002;167:157–64.
[29] Brady CM, Apostolidis A, Yiangou Y, et al. P2X3-immunoreactive
nerve fibres in neurogenic detrusor overactivity and the effect of
intravesical resiniferatoxin. Eur Urol 2004;46:247–53.
[30] Popat R, Apostolidis A, Kalsi V, Gonzales G, Fowler C, Dasgupta P. A
comparison between the response of patients with idiopathic
detrusor overactivity and neurogenic detrusor overactivity to the
first intradetrusor injection of botulinum-A toxin. J Urol 2005;174:
984–9.
Top Related
Copyright © 2022 FDOKUMEN

