







Bahasa
Halaman
Hukum


Journal o/ Cerebral Blood Flow and Metabolism 14:132-144 © 1994 The International Society of Cerebral Blood Flow and Metabolism Published by Raven Press, Ltd., New York
Functional Metabolic Mapping of the Rat Brain During Unilateral Electrical Stimulation of the
Subthalamic Nucleus
Minas Tzagournissakis, Catherine R. Dermon, and Helen E. Savaki
Laboratory of Physiology, Department of Basic Sciences, Division of Medicine, School of Health Sciences,
University of Crete, Crete, Greece
Summary: The alterations in local metabolic activity of several anatomically distinct brain areas were investigated by means of the quantitative autoradiographic 2-deoxY-D-[l-14C]glucose method in awake rats during unilateral electrical stimulation of the subthalamic nucleus (STH). Unilateral electrical stimulation of the STH induced local metabolic activation (by 70% as compared with the control group), as well as distal metabolic activations in the substantia nigra reticulata (by 34%), globus pallidus (by 19%), entopeduncular nucleus (by 18%), deep layers of the superior colliculi (by 15%), and parafascicular thalamic nucleus (by 18%), ipsilaterally to the stimulated side. The ventrolateral motor thalamic nucleus as well as the limbic components, posterior cingulate cortex, and anteroventral thalamic nucleus displayed bilateral metabolic activations (by 20--28%). These results indicate that, in addition to its known ipsilateral motor con-
The subthalamic nucleus (STH) is generally considered to exert control over motor functions. The association of STH hypoactivity with the clinical syndrome of hemiballismus has long been documented (Whittier, 1947; Whittier and Mettler, 1949; Carpenter and Carpenter, 1951; Hammond et ai., 1979; Mitchel et ai., 1985a; Crossman, 1987). The STH receives projections predominantly from the globus pallidus (Carpenter et ai., 1981; Van der Kooy et ai., 1981; Kita et ai., 1983a; Morizumi et ai., 1987; Groenewegen and Berendse, 1990) and
Received February 26, 1993; final revision received July 22, 1993; accepted August 9, 1993.
Address correspondence and reprint requests to Dr. H. E. Savaki at Laboratory of Physiology, Department of Basic Sciences, Division of Medicine, School of Health Sciences, University of Crete, 711 10 Iraklion, Crete, Greece.
Abbreviations used: DG, 2-deoxY-D-glucose; S TH, subthalamic nucleus.
132
nections, each STH is functionally related to the limbic system bilaterally. It is suggested that the STH is a site where the central motor information is accessible to the limbic system. Quantitative image analysis of individual serial sections in the STH, substantia nigra, and globus pallidus revealed a consistent dorsoventral pattern of topographic interrelations. Stimulation of either the dorsal or the ventral subdivision of the STH induced always stronger activation in the dorsal compartment of the substantia nigra and in the ventral compartment of the globus pallidus. These results suggest that the earlier-described inversion of the dorsoventral functional correspondence between the substantia nigra and globus pallidus may be partly mediated via the subthalamic nerve cells projecting collateral axons to both these areas. Key Words: Basal ganglia-Globus pallidus-Limbic system-Substantia nigra-Subthalamic nucleus.
the cerebral cortex (Hartmann-Von Monakow et ai., 1978; Romansky et ai., 1979; Kitai and Deniau, 1981; Afsharpour 1985; Rouzaire-Dubois and Scarnati, 1985; Canteras et ai., 1988). It also receives moderate projections from the parafascicular thalamic nucleus (Sugimoto et ai., 1983; Canteras et ai., 1990; Groenewegen and Berendse, 1990) and the pedunculopontine tegmental nucleus (Nomura et ai. , 1980; Hammond et ai., 1983a; Morizumi et ai., 1987). The STH neurons project branched axons to the substantia nigra pars reticulata, globus pallidus, and entopeduncular nucleus (Deniau et ai., 1978; Ricardo, 1980; Van der Kooy and Hattori, 1980; Hammond and Yelnik, 1983; Kita et ai., 1983b). Moreover, the STH sends sparse projections to the mesencephalic pedunculopontine tegmental nucleus, neostriatum, substantia nigra compacta, and cerebral cortex (Jackson and Crossman, 1981a,b; Saper and Loewy, 1982; Kita and

EFFECTS OF SUBTHALAMIC NUCLEUS STIMULATION 133
Kitai, 1987; Takada et aI., 1988; Granata and Kitai, 1989; Groenewegen and Berendse, 1990).
The well described neuroanatomical hardware does not necessarily represent the functionally importa,nt neuronal connections of the STH. The 2-deoxY-D-[1-14C]glucose ([14C]DG) metabolic mapping is determined by dynamic properties of neurons related to functional activity, beyond their static properties related to the anatomically traced connections (Sokoloff, 1977; Sokoloff et aI., 1977). This method has been proven useful in delineating the functional organization and topographic interrelations of the basal ganglia components in the rat (Savaki et aI., 1983a,b, 1984, 1988, 1992; Dermon et aI., 1990, 1992). Within the general framework of obtaining deeper understanding of the sophisticated neuronal network regulating extrapyramidal mechanisms of motor function, the specific aim of the present e4C]DG metabolic study was to investigate the rat brain areas functionally associated with the STH and furthermore to reveal any possible specific topographic interrelation between functional compartments of the STH and its major target areas, the substantia nigra and globus pallidus.
MATERIALS AND METHODS
Animals Experiments were carried out on 16 adult male
Sprague-Dawley albino rats (Charles River) weighing between 350 and 400 g. Animals were separated into two groups. The experimental group consisted of 11 rats, which were subjected to electrical stimulation of the STH in the right hemisphere. The control group consisted of five rats, which were simply implanted with an electrode in the STH of the right hemisphere. Experiments conducted over several days were randomized alternatively to measure the local cerebral glucose utilization in the rats of the two groups.
Surgical preparation Each rat was anesthetized with a mixture of halothane/
nitrous oxide/oxygen (1 %, 67:33 vol/vol), and polyethylene catheters were inserted in one femoral artery and one femoral vein. A bipolar stimulating electrode was vertically implanted into the right STH according to coordinates that had been previously determined experimentally (A: 5, L: 2.3, H: 2. 2). The electrode was sealed on the skull with dental cement, and the animals encased in a loose-fitting plaster cast around the lower abdomen were allowed to recover from anesthesia for 2-3 h.
Electrical stimulation of STH Electrical stimulation of the right STH started 2 min
before intravenous injections of the [14C]DG and continued during the first 15 min of the measurement of local cerebral glucose consumption. A bipolar stimulating electrode and 0.2 mm external diameter was used for the STH stimulation. The distance between the two poles of the electrode was 0.2 mm and the polarity was changing every half-minute during the 17 -min stimulating period. The
electrical stimulation consisted of trains (0.25-Hz frequency and tOO-ms duration) of 30 square pulses (300-Hz frequency, 0. 5-ms duration, and 100-f1A intensity). Stimulation with lower frequency and lower intensity did not induce any metabolic changes in pilot experiments.
Measurement of local cerebral glucose utilization The measurement of local cerebral glucose consump
tion was initiated by the intravenous injection of 100 f1Ci/ kg [14C]DG, of specific activity 50 mCi/mmol (New England Nuclear). Arterial blood samples were obtained during the succeeding 45 min and the glucose and [14C]DG plasma concentrations were measured. The rats were then decapitated and the brains were removed and frozen in isopentane cooled to - 40°C. Coronal sections 20 f1m thick were cut on a cryostat microtome at - 20°C. Autoradiograms were prepared from these sections by apposing the slides, together with precalibrated 14C_ standards, to Kodak OMI x-ray film in x-ray cassettes. Local cerebral tissue concentrations of 14C were determined from the auto radiographs by quantitative densitometry using a computer-based micro densitometer (Quanti met 970; Cambridge Instruments). The glucose consumption in discrete brain areas was calculated according to the operational equation of Sokoloff et al. (1977).
Statistical analyses Local cerebral glucose utilization values from the STH
stimulated experimental group of rats were compared with those from the control group by the Scheffe F test extension of the one-way analysis of variance. The percent increases of the significantly affected average value of local cerebral glucose utilization in the experimental group (E) over its respective value in the control group (C) were calculated as [(E - C)/C] x 100. Furthermore, ipsilateral-to-contralateral to the stimulated STH differences in glucose consumption were calculated within each one of the two groups using the paired two-tailed t test.
RESULTS
Following unilateral STH stimulation, ipsiversive head turning, accompanied by intermittent sniffing, exploring, and jaw movements, was observed in the experimental rats. The implantation of an electrode within the STH in the control rats did not induce any behavioral effect. The exact position of the stimulating electrode tract and tip was confirmed by overlaying the autoradiograms on the corresponding stained histological brain sections, and the metabolic activity within the stimulated area was determined by quantitative densitometry of the dorsal and ventral subdivisions of the STH, on computergenerated magnifications of the autoradiographic sections of interest.
Metabolic effects within STH
The auto radiograms in all STH-stimulated rats demonstrated always a well limited area of markedly increased optical density around the tip of the electrode, attesting to increased glucose con sump-
J Cereb Blood Flow Me/ab, Vol, 14, No, 1, 1994

134 M. TZAGOURNISSAKIS ET AL.
tion within the STH. Enhancement of optical density was not observed in any area adjacent to the STH (zona incerta, cerebral peduncle, or internal capsule). Quantitative analysis of the autoradiographic sections at the level of the STH revealed a 70% significant (by one-way analysis of variance) increase of local cerebral glucose utilization in the electrically stimulated STH of the experimental group as compared with the respective side of the control group. Right-to-left side comparison (by the two-tailed paired t test) demonstrated a major increase in metabolic activity in the electrically stimulated STH as compared with the intact one in the experimental group. The decrease (5%) in activity within the right STH implanted with an electrode in the control group of rats demonstrates that the electrode itself had a minor local depressive effect on metabolic activity (Table 1).
Metabolic effects in basal ganglia components
Unilateral stimulation of the STH induced significant elevation in local cerebral glucose utilization within the ipsilateral substantia nigra pars reticulata and the ipsilateral globus pallidus in the experimental group as compared with the contralateral homologous structures and the corresponding areas of the control group of animals (Table 1). Visual inspection of the magnified autoradiographic sections indicated that in all cases when (a) mostly the dorsal (Fig. 1), (b) mostly the ventral (Fig. 2), or (c) the whole dorsoventral (Fig. 3) extent of the STH was stimulated, the pattern of substantia nigra pars reticulata and globus pallidus activations was not homogeneous. Indeed, when individual local cerebral glucose utilization values separately in the dorsal and ventral subregions of the STH, substantia nigra pars reticulata, and globus pallidus (Fig. 4)
were analyzed along their rostrocaudal extent, a consistent dorsoventral pattern of topographic interrelations was revealed. Stronger stimulation of either the dorsal or the ventral subdivisions of the STH always induced a stronger activation in the dorsal compartment of the substantia nigra and in the ventral compartment of the globus pallidus. Unilateral stimulation of the STH also induced an enhancement in glucose utilization within the entopeduncular nucleus, the ventromedial part of the posterior striatum, and the substantia nigra pars compacta.
Metabolic effects in motor areas
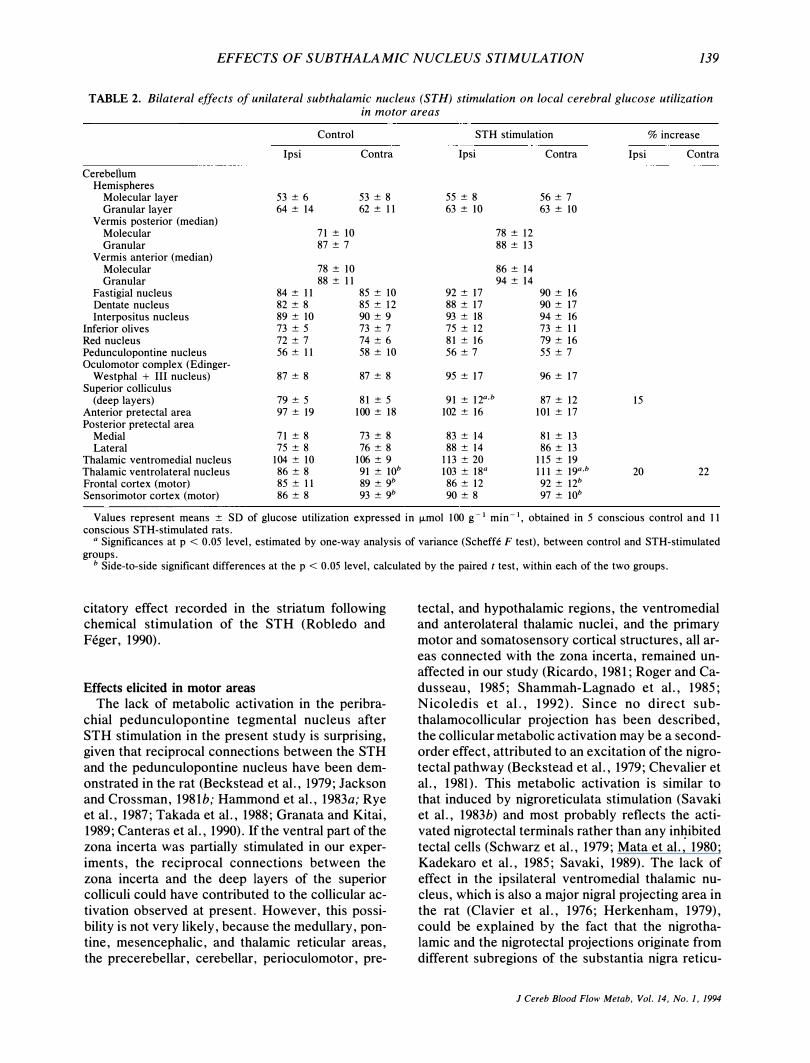
Significant elevation of glucose utilization was measured in the deep layers of the superior colliculi ipsilaterally to the stimulated STH. The ventrolateral nucleus displayed bilateral increases in glucose consumption (Table 2). The motor cortical areas demonstrated a small ipsilateral decrease in metabolic activity in the experimental as well as in the control group, which apparently reflects an electrode tract effect. There is no indication that the behavior (ipsiversive head turning) influenced the metabolic activity, because the single motor area activated (the ventrolateral thalamus) was bilaterally affected.
Metabolic effects in sensory areas
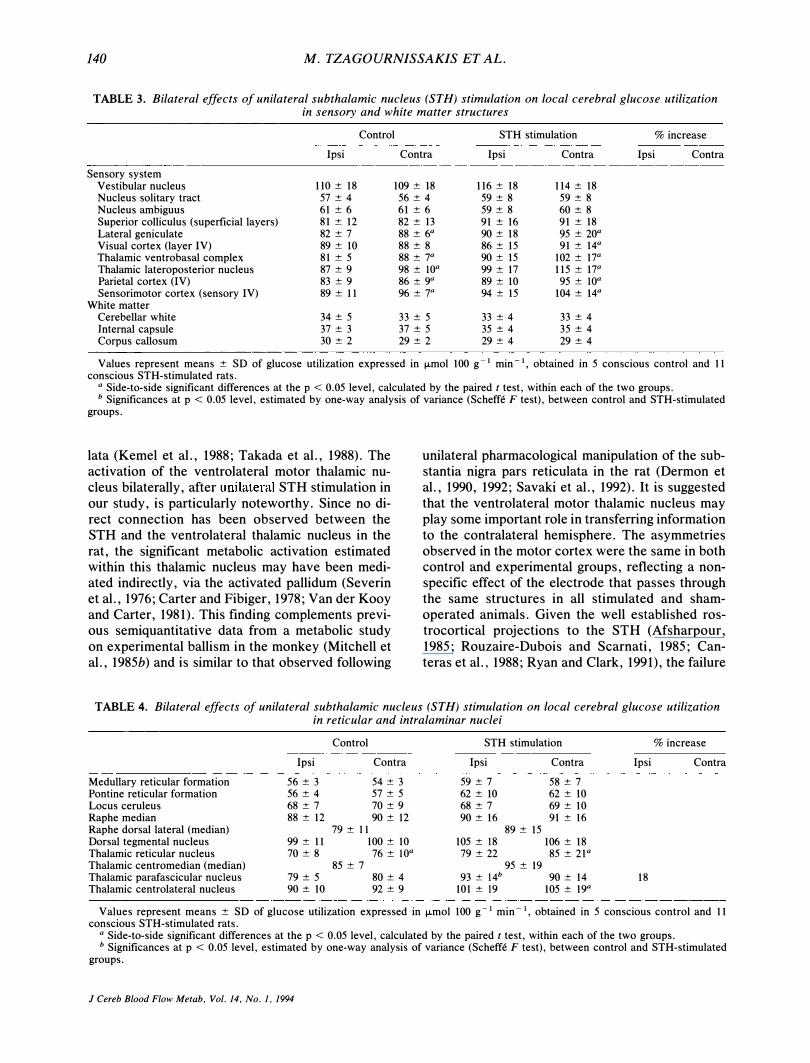
Within the sensory system, the vestibular nucleus, the nucleus of the solitary tract, the nucleus ambiguus, and the superficial layers of the superior colliculi remained bilaterally unaffected. Side-toside statistical comparison demonstrated that the ipsilateral lateral geniculate body, lateral posterior thalamic nucleus, ventrobasal thalamic complex, as well as the ipsilateral cortical parietal and somato-
TABLE 1. Bilateral effects of unilateral subthalamic nucleus (STH) stimulation on local cerebral glucose utilization in basal ganglia components
Control STH stimulation % increase -------
Ipsi Contra Ipsi Contra Ipsi Contra
Substantia nigra compacta 60 ± 10 63 ± 11 70 ± 12a 64±1O Substantia nigra reticulata 50 ± 4 51 ± 4 67 ± 17a,b 56 ± 9 34 STH 74 ± 8 78 ± 7a 126 ± 26a,b 83 ± 10 70 Posterior striatum (dorsolateral part) 80 ± 13 83 ± 14 86 ± 8 88 ± 10 Posterior striatum (ventromedial part) 82 ± 12 81 ± 11 87 ± 8a 81 ± 9 Anterior striatum (central part) 90 ± 12 92±13 92 ± 12 91 ± 9 Globus pallidus 52 ± 5 52 ± 5 62 ± lOa,b 54 ± 7 19 Entopeduncular nucleus 51 ± 5 52 ± 8 60 ± 12a 55 ± 8 18 Ventral pallidum 55 ± 5 55 ± 5 55 ± 2 55 ± 2 Accumbens nucleus 81 ± 12 80 ± 9 83 ± 10 83 ± 10
Values represent means ± SD of glucose utilization expressed in fLmol 100 g-t min-t, obtained in 5 conscious control and I I conscious STH-stimulated rats,
a Side-to-side significant differences at the p < 0, 05 level, calculated by the paired t test, within each of the two groups, b Significances at p < 0,05 level, estimated by one-way analysis of variance (Scheffe F test), between control and STH-stimulated
groups,
J Cereb Blood Flow Metab, Vol, 14, No, 1, 1994
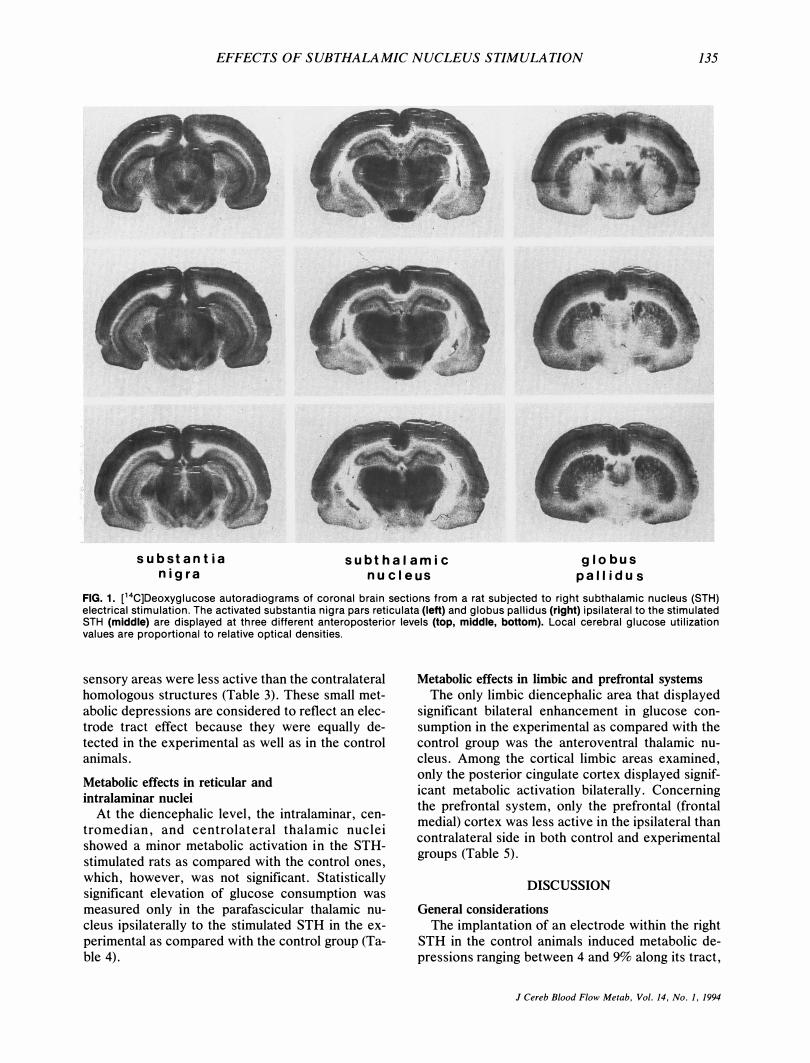
EFFECTS OF SUBTHALAMIC NUCLEUS STIMULATION 135
fl· . .-iJ.- � �\: ... '\, " , "'t;' ; . \ , ' " - �
.. . �
"
s ubstantia nig ra
s ubthalamic nucleus
globus pallidus
FIG. 1. C 4C]Deoxyglucose autoradiograms of coronal brain sections from a rat subjected to right subthalamic nucleus (STH) electrical stimulation. The activated substantia nigra pars reticulata (left) and globus paliidus (right) ipsilateral to the stimulated STH (middle) are displayed at three different anteroposterior levels (top, middle, bottom). Local cerebral glucose utilization values are proportional to relative optical densities.
sensory areas were less active than the contralateral homologous structures (Table 3). These small metabolic depressions are considered to reflect an electrode tract effect because they were equally detected in the experimental as well as in the control animals.
Metabolic effects in reticular and
intralaminar nuclei At the diencephalic level, the intralaminar, cen
tromedian, and centrolateral thalamic nuclei showed a minor metabolic activation in the STHstimulated rats as compared with the control ones, which, however, was not significant. Statistically significant elevation of glucose consumption was measured only in the parafascicular thalamic nucleus ipsilaterally to the stimulated STH in the experimental as compared with the control group (Table 4).
Metabolic effects in limbic and prefrontal systems
The only limbic diencephalic area that displayed significant bilateral enhancement in glucose consumption in the experimental as compared with the control group was the anteroventral thalamic nucleus. Among the cortical limbic areas examined, only the posterior cingulate cortex displayed significant metabolic activation bilaterally. Concerning the prefrontal system, only the prefrontal (frontal medial) cortex was less active in the ipsilateral than contralateral side in both control and experimental groups (Table 5).
DISCUSSION
General considerations
The implantation of an electrode within the right STH in the control animals induced metabolic depressions ranging between 4 and 9% along its tract,
J Cereb Blood Flow Metab, Vol, 14, No, 1, 1994
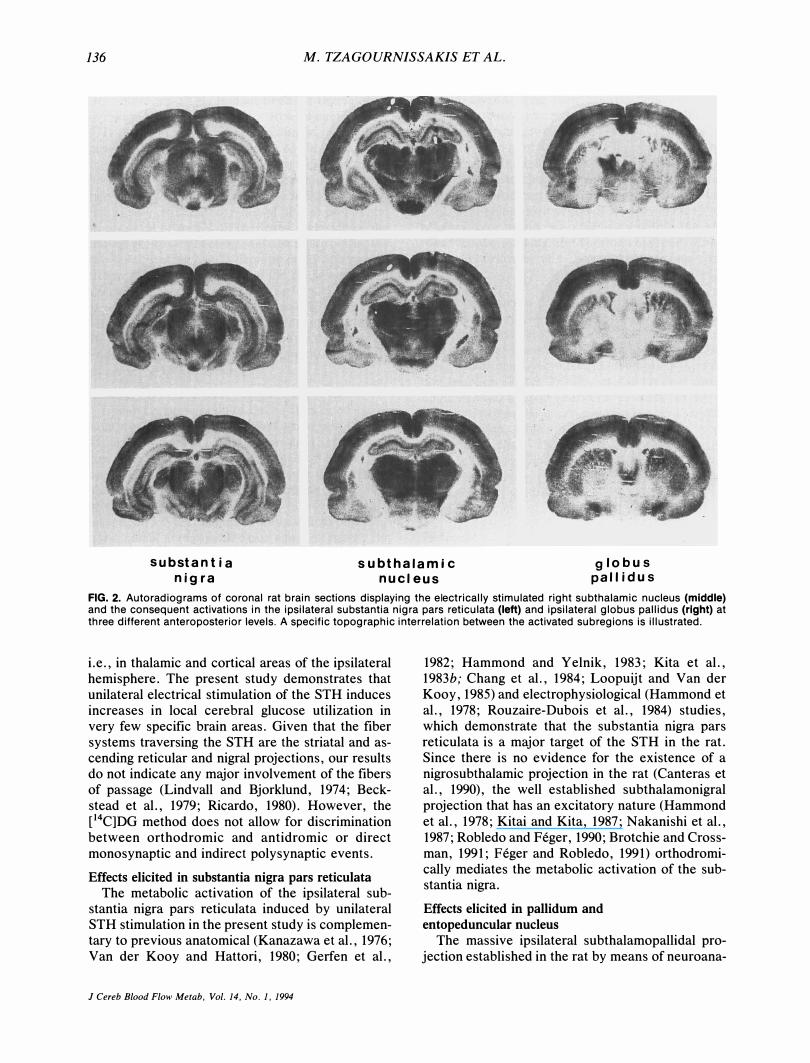
136 M. TZAGOURN1SSAKIS ET AL.
substantia nig ra
s ubthalamic nucleus
globus pallidus
FIG. 2. Autoradiograms of coronal rat brain sections displaying the electrically stimulated right subthalamic nucleus (middle) and the consequent activations in the ipsilateral substantia nigra pars reticulata (left) and ipsilateral globus pallidus (right) at three different anteroposterior levels. A specific topographic interrelation between the activated subregions is illustrated.
i.e., in thalamic and cortical areas of the ipsilateral hemisphere. The present study demonstrates that unilateral electrical stimulation of the STH induces increases in local cerebral glucose utilization in very few specific brain areas. Given that the fiber systems traversing the STH are the striatal and ascending reticular and nigral projections, our results do not indicate any major involvement of the fibers of passage (Lindvall and Bjorklund, 1974; Beckstead et al., 1979; Ricardo, 1980). However, the [14C]DG method does not allow for discrimination between orthodromic and antidromic or direct monosynaptic and indirect polysynaptic events.
Effects elicited in substantia nigra pars reticulata The metabolic activation of the ipsilateral sub
stantia nigra pars reticulata induced by unilateral STH stimulation in the present study is complementary to previous anatomical (Kanazawa et al., 1976; Van der Kooy and Hattori, 1980; Gerfen et al.,
J Cereb Blood Flow Metab, Vol. 14. No.1, 1994
1982; Hammond and Yelnik, 1983; Kita et al., 1983b; Chang et al. , 1984; Loopuijt and Van der Kooy, 1985) and electrophysiological (Hammond et al., 1978; Rouzaire-Dubois et al., 1984) studies, which demonstrate that the substantia nigra pars reticulata is a major target of the STH in the rat. Since there is no evidence for the existence of a nigrosubthalamic projection in the rat (Canteras et al., 1990), the well established subthalamonigral projection that has an excitatory nature (Hammond et aI. , 1978; Kitai and Kita, 1987; Nakanishi et al., 1987; Robledo and Feger, 1990; Brotchie and Crossman, 1991; Feger and Robledo, 1991) orthodromically mediates the metabolic activation of the substantia nigra.
Effects elicited in pallidum and
entopeduncular nucleus
The massive ipsilateral subthalamopallidal projection established in the rat by means of neuroana-
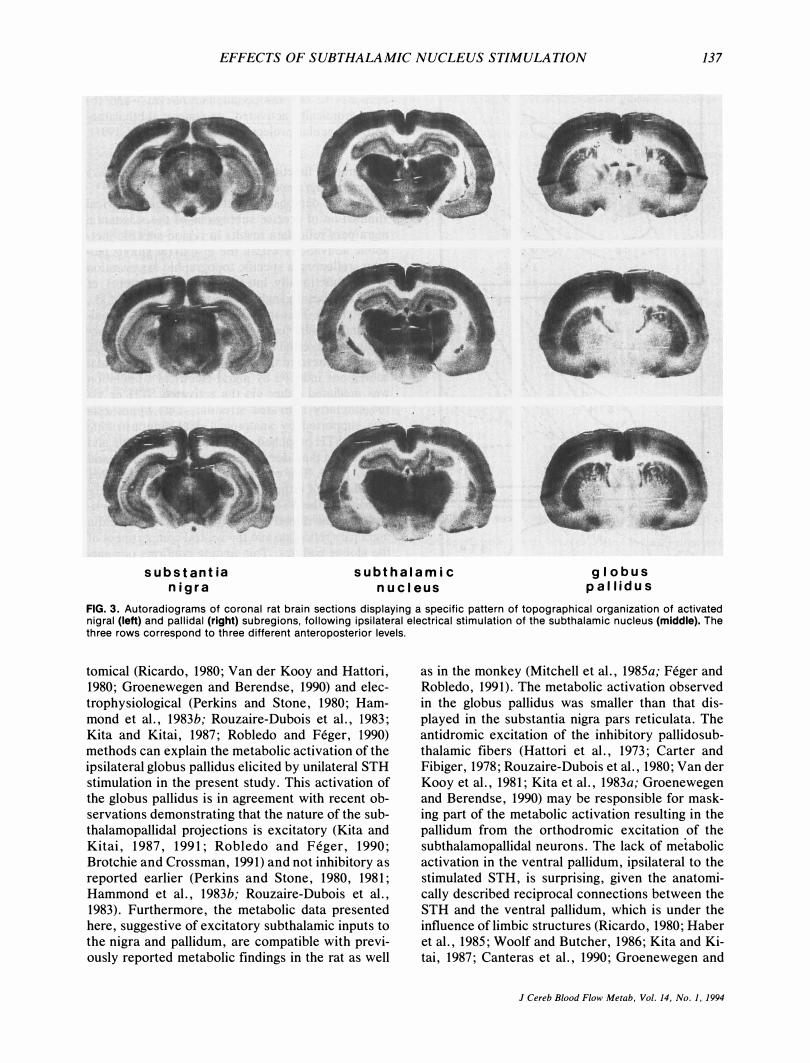
EFFECTS OF SUBTHALAMIC NUCLEUS STIMULATION 137
s ubs tantia nigra
s ubthalamic nucleus
globus pallidus
FIG. 3. Autoradiograms of coronal rat brain sections displaying a specific pattern of topographical organization of activated nigral (left) and pallidal (right) subregions, following ipsilateral electrical stimulation of the subthalamic nucleus (middle). The three rows correspond to three different anteroposterior levels.
tomical (Ricardo, 1980; Van der Kooy and Hattori, 1980; Groenewegen and Berendse, 1990) and electrophysiological (Perkins and Stone, 1980; Hammond et aI., 1983b; Rouzaire-Dubois et aI., 1983; Kita and Kitai, 1987; Robledo and Feger, 1990) methods can explain the metabolic activation of the ipsilateral globus pallidus elicited by unilateral STH stimulation in the present study. This activation of the globus pallidus is in agreement with recent observations demonstrating that the nature of the subthalamopallidal projections is excitatory (Kita and Kitai, 1987, 1991; Robledo and Feger, 1990; Brotchie and Crossman, 1991) and not inhibitory as reported earlier (Perkins and Stone, 1980, 1981; Hammond et aI., 1983b; Rouzaire-Dubois et aI., 1983). Furthermore, the metabolic data presented here, suggestive of excitatory subthalamic inputs to the nigra and pallidum, are compatible with previously reported metabolic findings in the rat as well
as in the monkey (Mitchell et aI., 1985a; Feger and Robledo, 1991). The metabolic activation observed in the globus pallidus was smaller than that displayed in the substantia nigra pars reticulata. The antidromic excitation of the inhibitory pallidosubthalamic fibers (Hattori et aI., 1973; Carter and Fibiger, 1978; Rouzaire-Dubois et aI., 1980; Van der Kooy et aI., 1981; Kita et aI., 1983a; Groenewegen and Berendse, 1990) may be responsible for masking part of the metabolic activation resulting in the pallidum from the orthodromic excitation of the subthalamopallidal neurons. The lack of metabolic activation in the ventral pallidum, ipsilateral to the stimulated STH, is surprising, given the anatomically described reciprocal connections between the STH and the ventral pallidum, which is under the influence of limbic structures (Ricardo, 1980; Haber et aI., 1985; Woolf and Butcher, 1986; Kita and Kitai, 1987; Canteras et aI., 1990; Groenewegen and
J Cereb Blood Flow Metab, Vol. 14, No. 1, 1994
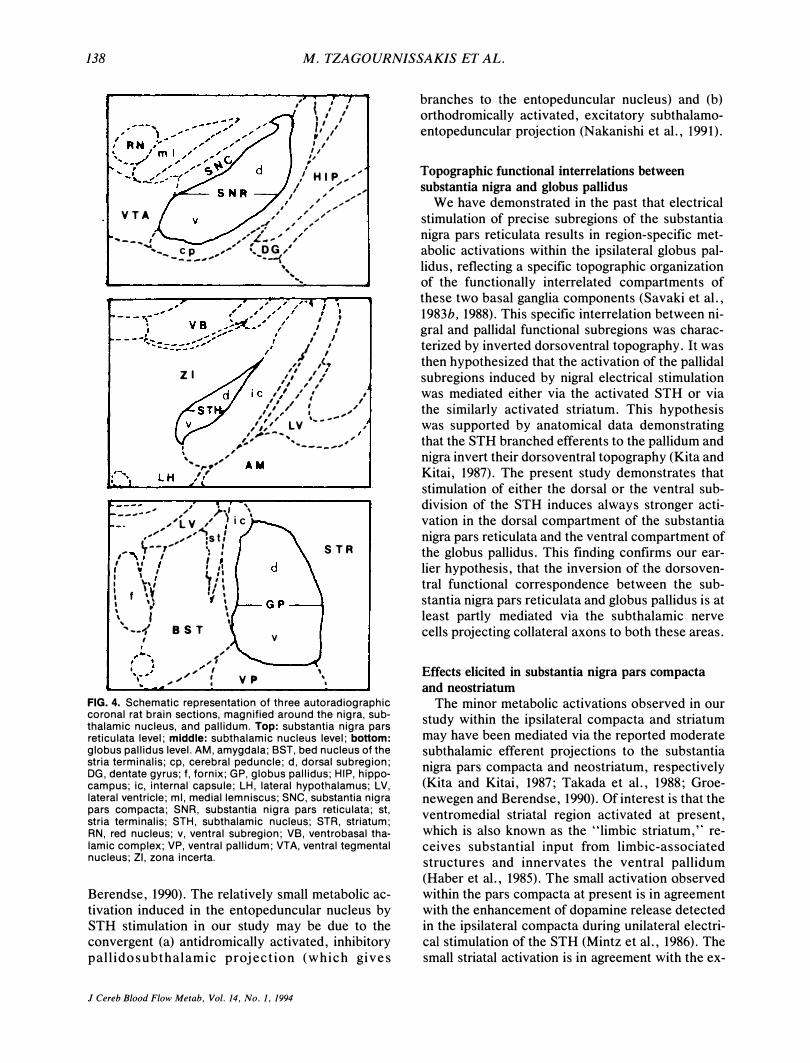
138 M. TZAGOURN1SSAKIS ET AL.
VTA
" { " I ,' ,' " I ' , " I ,
, I I, , I I HIP ,.-
,/ ,"'.... " I ' ",, ' " , ' ", "" , , " "",, ' " ,,,, '
, ,',' , , .... _ ....... � ,,,,' L .. , ' "
"__ c p , .. ' (0 G / ----� .. ' -.--{
ZI
,
�, " ".
I ; I • / : I I
J " " ,<., II " /1 ,,' ,' ,' ,' ,' , " I , I ,,' ,' I I I" • ....' " , \ ... -,,,, ,I,,' LV , , ,,,.-- ...... " ,.'.,.. ' ' ----- ,,, ' '.. ,,' A '" "',. " ... I' , ,
-- " , '" " ;-. , ', ,'L V ' � i c • , "
,,,,,, , .... '''''15 t I _ .... �--, ... ' :.! S T R , ,J I ; , , ' " "
I ", 1 :, I " I. I �., , I \ f ,� '.,' , , \ l ,
, . , \ I � "--/, B S T , v
, , , ,, " ,
.. ",/ : V p , , ,
FIG. 4. Schematic representation of three autoradiographic coronal rat brain sections, magnified around the nigra, subthalamic nucleus, and pallidum. Top: substantia nigra pars reticulata level; middle: subthalamic nucleus level; bottom: globus pallidus level. AM, amygdala; SST, bed nucleus of the stria terminalis; cp, cerebral peduncle; d, dorsal subregion; DG, dentate gyrus; f, fornix; GP, globus pallid us; HIP, hippocampus; iC, internal capsule; LH, lateral hypothalamus; LV, lateral ventricle; ml, medial lemniscus; SNC, substantia nigra pars compacta; SNR, substantia nigra pars reticulata; st, stria terminalis; STH, subthalamic nucleus; STR, striatum; RN, red nucleus; v, ventral subregion; VB, ventrobasal thalamic complex; VP, ventral pallidum; VTA, ventral tegmental nucleus; ZI, zona incerta.
Berendse, 1990). The relatively small metabolic activation induced in the entopeduncular nucleus by STH stimulation in our study may be due to the convergent (a) antidromically activated, inhibitory pallidosubthalamic projection (which gives
J Cereb Blood Flow Me/ab. Vol. 14. No. 1. 1994
branches to the entopeduncular nucleus) and (b) orthodromically activated, excitatory subthalamoentopeduncular projection (Nakanishi et aI., 1991).
Topographic functional interrelations between
substantia nigra and globus pallidus
We have demonstrated in the past that electrical stimulation of precise subregions of the substantia nigra pars reticulata results in region-specific metabolic activations within the ipsilateral globus pallidus, reflecting a specific topographic organization of the functionally interrelated compartments of these two basal ganglia components (Savaki et aI., 1983b, 1988). This specific interrelation between nigral and pallidal functional subregions was characterized by inverted dorsoventral topography. It was then hypothesized that the activation of the pallidal subregions induced by nigral electrical stimulation was mediated either via the activated STH or via the similarly activated striatum. This hypothesis was supported by anatomical data demonstrating that the STH branched efferents to the pallidum and nigra invert their dorsoventral topography (Kita and Kitai, 1987). The present study demonstrates that stimulation of either the dorsal or the ventral subdivision of the STH induces always stronger activation in the dorsal compartment of the substantia nigra pars reticulata and the ventral compartment of the globus pallidus. This finding confirms our earlier hypothesis, that the inversion of the dorsoventral functional correspondence between the substantia nigra pars reticulata and globus pallidus is at least partly mediated via the subthalamic nerve cells projecting collateral axons to both these areas.
Effects elicited in substantia nigra pars compacta
and neostriatum
The minor metabolic activations observed in our study within the ipsilateral compacta and striatum may have been mediated via the reported moderate subthalamic efferent projections to the substantia nigra pars compacta and neostriatum, respectively (Kita and Kitai, 1987; Takada et aI., 1988; Groenewegen and Berendse, 1990). Of interest is that the ventromedial striatal region activated at present, which is also known as the "limbic striatum," receives substantial input from limbic-associated structures and innervates the ventral pallidum (Haber et aI., 1985). The small activation observed within the pars compacta at present is in agreement with the enhancement of dopamine release detected in the ipsilateral compacta during unilateral electrical stimulation of the STH (Mintz et aI., 1986). The small striatal activation is in agreement with the ex-
EFFECTS OF SUBTHALAMIC NUCLEUS STIMULATION 139
TABLE 2. Bilateral effects of unilateral subthalamic nucleus (STH) stimulation on local cerebral glucose utilization in motor areas
Control
Ipsi
Cerebelium Hemispheres
Molecular layer 53 ± 6 Granular layer 64 ± 14
Vermis posterior (median) Molecular 71 ± 10 Granular 87 ± 7
Vermis anterior (median) Molecular 78 ± 10 Granular 88 ± I I
Fastigial nucleus 84 ± 11 Dentate nucleus 82 ± 8 Interpositus nucleus 89 ± 10
Inferior olives 73 ± 5 Red nucleus 72 ± 7 Pedunculopontine nucleus 56 ± I I Oculomotor complex (Edinger-
Westphal + III nucleus) 87 ± 8 Superior colliculus
(deep layers) 79 ± 5 Anterior pretectal area 97 ± 19 Posterior pretectal area
Medial 71 ± 8 Lateral 75 ± 8
Thalamic ventromedial nucleus 104 ± 10 Thalamic ventrolateral nucleus 86 ± 8 Frontal cortex (motor) 85 ± I I Sensorimotor cortex (motor) 86 ± 8
Contra
53 ± 8 62 ± I I
85 ± 10 85 ± 12 90 ± 9 73 ± 7 74 ± 6 58 ± 10
87 ± 8
81 ± 5 100 ± 18
73 ± 8 76 ± 8
106 ± 9 91 ± lOb 89 ± 9b 93 ± 9b
STH stimulation
Ipsi Contra
55 ± 8 56 ± 7 63 ± 10 63 ± 10
78 ± 12 88 ± 13
86 ± 14 94 ± 14
92 ± 17 90 ± 16 88 ± 17 90 ± 17 93 ± 18 94 ± 16 75 ± 12 73 ± I I 81 ± 16 79 ± 16 56 ± 7 55 ± 7
95 ± 17 96 ± 17
91 ± 12Q,b 87 ± 12 102 ± 16 101 ± 17
83 ± 14 81 ± 13 88 ± 14 86 ± 13
113 ± 20 115 ± 19 103 ± 18Q I I I ± 19Q,b
86 ± 12 92 ± 12b 90 ± 8 97 ± lOb
% increase
Ipsi Contra
15
20 22
Values represent means ± SD of glucose utilization expressed in �mol 100 g � 1 min � I. obtained in 5 conscious control and 11 conscious STH-stimulated rats.
Q Significances at p < 0 . 05 level, estimated by one-way analysis of variance (Scheffe F test), between control and STH-stimulated groups.
b Side-to-side significant differences at the p < 0. 05 level, calculated by the paired t test, within each of the two groups.
citatory effect recorded in the striatum following chemical stimulation of the STH (Robledo and Feger, 1990).
Effects elicited in motor areas
The lack of metabolic activation in the peribrachial pedunculopontine tegmental nucleus after STH stimulation in the present study is surprising, given that reciprocal connections between the STH and the pedunculopontine nucleus have been demonstrated in the rat (Beckstead et al., 1979; Jackson and Crossman, 1981b; Hammond et al., 1983a; Rye et al., 1987; Takada et al., 1988; Granata and Kitai, 1989; Canteras et al., 1990). If the ventral part of the zona incerta was partially stimulated in our experiments, the reciprocal connections between the zona incerta and the deep layers of the superior colliculi could have contributed to the collicular activation observed at present. However, this possibility is not very likely, because the medullary, pontine, mesencephalic, and thalamic reticular areas, the precerebellar, cerebellar, peri oculomotor , pre-
tectal, and hypothalamic regions, the ventromedial and anterolateral thalamic nuclei, and the primary motor and somatosensory cortical structures, all areas connected with the zona incerta, remained unaffected in our study (Ricardo, 1981; Roger and Cadusseau, 1985; Shammah-Lagnado et al., 1985; Nicoledis et al., 1992). Since no direct subthalamocollicular projection has been described, the collicular metabolic activation may be a secondorder effect, attributed to an excitation of the nigrotectal pathway (Beckstead et al., 1979; Chevalier et al., 1981). This metabolic activation is similar to that induced by nigroreticulata stimulation (Savaki et al., 1983b) and most probably reflects the activated nigrotectal terminals rather than any in�ibited tectal cells (Schwarz et al., 1979; Mata et al., 1980; Kadekaro et al., 1985; Savaki, 1989). The lack of effect in the ipsilateral ventromedial thalamic nucleus, which is also a major nigral projecting area in the rat (Clavier et aI., 1976; Herkenham, 1979), could be explained by the fact that the nigrothalamic and the nigrotectal projections originate from different subregions of the substantia nigra reticu-
J Cereb Blood Flow Metab, Vol. 14, No. 1, 1994
140 M. TZAGOURNISSAKIS ET AL.
TABLE 3. Bilateral effects of unilateral subthalamic nucleus (STH) stimulation on local cerebral glucose utilization in sensory and white matter structures
Control STH stimulation % increase
Ipsi Contra Ipsi Contra Ipsi Contra
Sensory system Vestibular nucleus 110 ± 18 109 ± 18 116 ± 18 114 ± 18 Nucleus solitary tract 57 ± 4 56 ± 4 59 ± 8 59 ± 8 Nucleus ambiguus 61 ± 6 61 ± 6 59 ± 8 60 ± 8 Superior colliculus (superficial layers) 81 ± 12 82 ± \3 91 ± 16 91 ± 18 Lateral geniculate 82 ± 7 88 ± 6a 90 ± 18 95 ± 20a Visual cortex (layer IV) 89 ± 10 88 ± 8 86 ± 15 91 ± 14a Thalamic ventrobasal complex 81 ± 5 88 ± 7a 90 ± 15 102 ± 17a Thalamic lateroposterior nucleus 87 ± 9 98 ± lOa 99 ± 17 115 ± 17a Parietal cortex (IV) 83 ± 9 86 ± 9" 89 ± 10 95 ± lOa Sensorimotor cortex (sensory IV) 89 ± I I 96 ± 7" 94 ± 15 104 ± 14a
White matter Cerebellar white 34 ± 5 33 ± 5 33 ± 4 33 ± 4 Internal capsule 37 ± 3 37 ± 5 35 ± 4 35 ± 4 Corpus callosum 30 ± 2 29 ± 2 29 ± 4 29 ± 4
Values represent means ± SD of glucose utilization expressed in j.Lmol 100 g - I min - I, obtained in 5 conscious control and 11 conscious STH-stimulated rats.
a Side-to-side significant differences at the p < 0.05 level, calculated by the paired t test, within each of the two groups. b Significances at p < 0.05 level, estimated by one-way analysis of variance (Scheffe F test), between control and STH-stimulated
groups.
lata (Kernel et aI., 1988; Takada et aI., 1988). The activation of the ventrolateral motor thalamic nucleus bilaterally, after unilateral STH stimulation in our study, is particularly noteworthy. Since no direct connection has been observed between the STH and the ventrolateral thalamic nucleus in the rat, the significant metabolic activation estimated within this thalamic nucleus may have been mediated indirectly, via the activated pallidum (Severin et aI., 1976; Carter and Fibiger, 1978; Van der Kooy and Carter, 1981). This finding complements previous semiquantitative data from a metabolic study on experimental ballism in the monkey (Mitchell et aI., 1985b) and is similar to that observed following
unilateral pharmacological manipulation of the substantia nigra pars reticulata in the rat (Dermon et aI. , 1990, 1992; Savaki et aI., 1992). It is suggested that the ventrolateral motor thalamic nucleus may play some important role in transferring information to the contralateral hemisphere. The asymmetries observed in the motor cortex were the same in both control and experimental groups, reflecting a nonspecific effect of the electrode that passes through the same structures in all stimulated and shamoperated animals. Given the well established rostrocortical projections to the STH (Afsharpour, 1985; Rouzaire-Dubois and Scarnati, 1985; Canteras et aI., 1988; Ryan and Clark, 1991), the failure
TABLE 4. Bilateral effects of unilateral subthalamic nucleus (STH) stimulation on local cerebral glucose utilization in reticular and intra laminar nuclei
Control
Ipsi Contra
Medullary reticular formation 56 ± 3 54 ± 3 Pontine reticular formation 56 ± 4 57 ± 5 Locus ceruleus 68 ± 7 70 ± 9 Raphe median 88 ± 12 90 ± 12 Raphe dorsal lateral (median) 79 ± I I Dorsal tegmental nucleus 99 ± 11 100 ± 10 Thalamic reticular nucleus 70 ± 8 76 ± loa Thalamic centromedian (median) 85 ± 7 Thalamic parafascicular nucleus 79 ± 5 80 ± 4 Thalamic centrolateral nucleus 90 ± 10 92 ± 9
STH stimulation
Ipsi Contra
59 ± 7 58 ± 7 62 ± 10 62 ± 10 68 ± 7 69 ± 10 90 ± 16 91 ± 16
89 ± 15 105 ± 18 106 ± 18
79 ± 22 85 ± 21a 95 ± 19
93 ± 14b 90 ± 14 101 ± 19 105 ± 19a
% increase
Ipsi Contra
18
Values represent means ± SD of glucose utilization expressed in j.Lmol 100 g-I min-I, obtained in 5 conscious control and 11 conscious STH-stimulated rats.
a Side-to-side significant differences at the p < 0.05 level, calculated by the paired t test, within each of the two groups. b Significances at p < 0.05 level, estimated by one-way analysis of variance (Scheffe F test), between control and STH-stimulated
groups.
J Cereb Blood Flow Metab, Vol. 14, No. 1, 1994

EFFECTS OF SUBTHALAMIC NUCLEUS STIMULATION 141
TABLE S. Bilateral effects of unilateral subthalamic nucleus (STH) stimulation on local cerebral glucose utilization in limbic and prefrontal nuclei
Hippocampus (moL layers) Dentate gyrus Amygdaloid complex Mamillary complex (median) Interpeduncular nucleus (median) Septum medial Lateral habenula Medial habenula Hypothalamus (posterior) Ventral tegmental area Thalamic anteromedial nucleus Thalamic anteroventral nucleus Thalamic intraanteromedian
nucleus (median) Thalamic mediodorsal nucleus Prefrontal medial cortex (IV) Anterior cingulate cortex Posterior cingulate cortex Entorhinal cortex Pyriform cortex
Control
Ipsi Contra
73 ± 7 72 ± 7 65 ± 5 64 ± 3 43 ± 4 43 ± 4
102 ± 15 96 ± 14
59 ± 8 60 ± 8 109 ± 22 114 ± 26
67 ± loa 61 ± 6 46 ± 9 46 ± 10 52 ± 8 51 ± 8
123 ± 19 124 ± 21 106 ± 22 107 ± 24
120 ± 23 106 ± 15 106 ± 15
91 ± 12 95 ± 13a 99 ± 18 101 ± 18 91 ± 14 97 ± 16a 72 ± 8 72 ± 6 62 ± 9 63 ± 7
STH stimulation
Ipsi Contra
73 ± 10 74 ± 10 65 ± 12 63 ± 9 46 ± 5 46 ± 5
114 ± 18 94 ± 13
64 ± 8 63 ± 8 119 ± 23 120 ± 22
69 ± 7a 61 ± 6 46 ± 3 45 ± 3 47 ± 8 47 ± 8
133 ± 23 133 ± 23 136 ± 22b 136 ± 24b
129 ± 24 125 ± 25 125 ± 25
91 ± 11 96 ± 11a 99 ± 12 104 ± 12a
113 ± 16b 117 ± 16a,b
72 ± 9 73 ± 9 62 ± 7 62 ± 6
% increase
Ipsi Contra
28 27
24 21
Values represent means ± SD of glucose utilization expressed in fLmol 100 g-I min -I, obtained in 5 conscious control and 11 conscious STH-stimulated rats,
a Side-to-side significant differences at the p < 0,05 level, calculated by the paired t test, within each of the two groups, b Significances at p < 0,05 level, estimated by one-way analysis of variance (Scheffe F test), between control and STH-stimulated
groups,
of the STH stimulation to induce any metabolic change in motor cortical areas indicates that the antidromic ally excited widespread projections (e. g. , corticosubthalamic) are mapped by the e4C]DG method to a much smaller degree than the orthodromic ally excited well focused projections (e.g., subthalamonigral).
Effects elicited in sensory, reticular, and
prefrontal areas
The sensory, reticular, and prefrontal areas examined in the STH -stimulated group of rats did not display any significant changes in metabolic activity when compared with the respective areas in the control group. The fact that the STH stimulation did not induce any metabolic changes in sensory and reticular regions indicates further that the electrical current did not spread in the adjacent zona incerta, which is known to be a major reticular and somatosensory relay area in the rat (Roger and Cadusseau, 1985; Shammah-Lagnado et aI., 1985; Nikoledis et aI., 1992).
Effects elicited in intralaminar and limbic areas
The metabolic activation observed in the parafascicular thalamic nucleus may have been caused by the antidromic excitation of the direct parafasciculosubthalamic projection (Sugimoto et aI., 1983; Canteras et aI., 1990; Groenewegen and Berendse, 1990). However, since the strongly activated sub-
stantia nigra is known to project to the parafascicular thalamic nucleus (Clavier et aI., 1976; Beckstead et aI., 1979; Gerfen et aI., 1982; Cornwall and Phillipson, 1988), the indirect orthodromic excitation of the subthalamonigroparafascicular polysynaptic pathway may have participated. The participation of the parafascicular thalamic nucleus in the limbic-related functions of the basal ganglia has been postulated in the past (Sadikot et aI., 1992). Furthermore, a direct relation between the STH and the limbic system is indicated by our finding that the anteroventral thalamic nucleus and the reciprocally connected with its posterior cingulate cortical area (Domesick, 1969, 1972; Jones and Leavitt, 1974; Seki and Zyo, 1984) were bilaterally activated after electrical stimulation of the STH. These effects are similar to those elicited by unilateral pharmacological manipulation of the substantia nigra pars reticulata (Dermon et aI., 1992; Savaki et aI., 1992). This limbic thalamocortical activation induced by the STH stimulation at present supports the hypothesis that the STH is associated with limbic processes (Canteras et aI., 1990; Groenewegen and Berendse, 1990).
CONCLUSIONS
The present results confirm the STH control on basal ganglia components and the specific topographic organization of the subthalamic projections
J Cereb Blood Flow Metab. Vol, 14. No, 1. 1994

142 M. TZAGOURNISSAKIS ET AL.
to the substantia nigra pars reticulata and the globus pallidus. Based on the metabolic activations induced in the nigrocollicular system as well as in the globus pallidus and the ventrolateral thalamic nucleus, we have further evidence to support the role of the STH in the realm of the motor system. Based on the metabolic activations induced in the limbicinnervated parts of the basal ganglia, entopeduncular nucleus and ventromedial striatum, as well as in the limbic-related areas, parafascicular and anteroventral thalamic nucleus and posterior cingulate cortex, we have strong evidence to suggest that the function of the STH relates to the limbic sphere of the brain. In conclusion, the present study indicates that (a) the motor nigrocollicular and pallidothalamic projecting systems and (b) the limbic thalamocortical paths share the STH as a common relay. Consequently, it can be suggested that the STH is a site where the central motor information is accessible to the limbic system.
Acknowledgment: This work was supported by the Regional Operational Program of Crete, European Community Support Framework 1989-93, and by a grant from the Greek Ministry of Health. We thank Mr. Philip Georgopoulos, Mr. Vassilis Raos, and Mrs. Patricia Pizarro for technical assistance.
REFERENCES
Afsharpour S (1985) Topographic projections of the cerebral cortex to the subthalamic nucleus. J Comp Neurol 236: 14-28
Beckstead RM, Domesic VB, Nauta WJH (1979) Efferent connections of the substantia nigra and ventral tegmental area in the rat. Brain Res 175: 191-217
Brotchie JM, Crossman AR (1991) D-[3HJAspartate and [14CJ_ GAB A uptake in the basal ganglia of rats following lesions in the subthalamic region suggest a role for excitatory amino acid but not GABA-mediated transmission in subthalamic nucleus efferents. Exp Neuroll13:171-181
Canteras NS, Shammah-Lagnado SJ, Silva BA, Ricardo JA (1988) Somatosensory inputs to the subthalamic nucleus: a combined retrograde and anterograde horseradish peroxidase study in the rat. Brain Res 458:53-64
Canteras NS, Shammah-Lagnado SJ, Silva BA, Ricardo JA (1990) Afferent connections of the subthalamic nucleus: a combined retrograde and anterograde horseradish peroxidase study in the rat. Brain Res 513:43-59
Carpenter MB, Carpenter CS (1951) Analysis of somatotopic relations of corpus Luysi in man and monkey. J Comp NeuroI95:349-370
Carpenter MB, Batton RR, Carleton SC, Keller JT (1981) Interconnections and organization of pallidal and subthalamic nucleus neurons in the monkey. J Comp NeuroI197:579-603
Carter DA, Fibiger HC (1978) The projections of the entopeduncular nucleus and globus pallidus in rat as demonstrated by autoradiography and horseradish peroxidase histochemistry. J Comp NeuroI177:113-124
Chang HT, Kita H, Kitai ST (1984) The ultrastructural morphology of the subthalamic-nigral axon terminals intracellularly labeled with horseradish peroxidase. Brain Res 299: 183-185
Chevalier G, Deniau JM, Thierry AM, Feger J (1981) The nigrotectal pathway, an electrophysiological investigation in the rat. Brain Res 213:253-263
Clavier RM, Atmadja S, Fibiger HC (1976) Nigrothalamic pro-
J Cereb Blood Flow Metab, Vol. 14, No. I, 1994
jections in the rat as demonstrated by orthograde and retrograde tracing techniques. Brain Res Bull 1:379-384
Cornwall J, Phillipson OT (1988) Afferent projections to the parafascicular thalamic nucleus of the rat, as shown by the retrograde transport of wheat germ-agglutinin. Brain Res Bull 20: 139-150
Crossman AR (1987) Primate models of dyskinesia: the experimental approach to the study of basal ganglia related involuntary movement disorders. Neuroscience 21: 1-40
Deniau JM, Hammond C, Chevalier G, Feger J (1978) Evidence for branched subthalamic nucleus projections to substantia nigra, entopeduncular nucleus and globus pallidus. Neurosci Lett 9:117-121
Dermon CR, Pizarro P, Georgopoulos P, Savaki HE (1990) Bilateral alterations in local cerebral glucose utilization following intranigral application of the GABAergic agonist muscimol. J Neurosci 10:2861-2878
Dermon CR, Tzagournissakis M, Savaki HE (1992) Bilateral cerebral metabolic effects of pharmacological manipulation of the substantia nigra in the rat: unilateral intranigral application of the putative excitatory neurotransmitter substance P. Neuroscience 50:795-809
Domesick VB (1969) Projections from the cingulate cortex in the rat. Brain Res 12:296-320
Domesick VB (1972) Thalamic relationships of the medial cortex in the rat. Brain Behav Evol 6:457-483
Feger J, Robledo P (1991) The effects of activation or inhibition of the subthalamic nucleus on the metabolic and electrophysiological activities within the pallidal complex and substantia nigra in the rat. Eur J Neurosci 3:947-952
Gerfen CR, Staines WA, Arbuthnott GW, Fibiger HC (1982) Crossed connections of the substantia nigra in the rat. J Comp Neurol 207:283-303
Granata AR, Kitai ST (1989) Intracellular analysis of excitatory subthalamic inputs to the pedunculopontine neurons. Brain Res 488:57-72
Groenewegen HJ, Berendse HW (1990) Connections of the subthalamic nucleus with ventral striatopallidal parts of the basal ganglia in the rat. J Comp Neurol 294:607-622
Haber SN, Groenewegen HJ, Grove EA, Nauta WJH (1985) Efferent connections of the ventral pallidum: evidence of a dual striatopallidofugal pathway. J Comp Neurol 235:322-335
Hammond C, Yelnik J (1983) Intracellular labeling of rat subthalamic neurons with horseradish peroxidase: computer analysis of dendrites and characterization of axon arborization. Neuroscience 8:781-790
Hammond C, Deniau JM, Rizk A, Feger J (1978) Electrophysiological demonstration of an excitatory subthalamo-nigral pathway in the rat. Brain Res 151:235-244
Hammond C, Feger J, Bionlac B, Souterand JP (1979) Experimental hemiballism in the monkey produced by unilateral kainic acid lesion in corpus Luysi. Brain Res 171:577-580
Hammond C, Rouzaire-Dubois B, Feger J, Jackson A, Crossman AR (1983a) Anatomical and electrophysiological studies on the reciprocal projections between the subthalamic nucleus and nucleus tegmenti pedunculopontinus in the rat. Neuroscience 9:41-52
Hammond C, Shibazaki T, Rouzaire-Dubois B (1983b) Branched output of the rat subthalamic nucleus: electrophysiological study of the synaptic effects of identified cells in the two main target nuclei, the entopeduncular nucleus and the substantia nigra. Neuroscience 9:511-520
Hartmann-Von Monakow K, Alert K, Kiinzle H (1978) Projections of precentral motor cortex and other cortical areas of the frontal lobe to the subthalamic nucleus in the monkey. Exp Brain Res 33:395-403
.
Hattori T, McGreer PL, Fibiger HC, McGeer EG (1973) On the source of GABA-containing terminals in the substantia nigra. Electron microscopic autoradiographic and biochemical studies. Brain Res 54: 103-114
Herkenham M (1979) The afferent and efferent connections of

EFFECTS OF SUBTHALAMIC NUCLEUS STIMULATION 143
the ventromedial thalamic nucleus in the rat. J Comp Neurol 183:487-518
Jackson A, Crossman AR (1981a) Subthalamic nucleus efferent projection to the cerebral cortex. Neuroscience 6:2367-2377
Jackson A, Crossman AR (l981b) Subthalamic projection to nucleus tegmenti pedunculoponticus in the rat. Neurosci Lett 22:17-22
Jones EG, Leavitt RY (1974) Retrograde axonal transport and the demonstration of non-specific projections to the cerebral cortex and striatum from the thalamic intralaminar nuclei in the rat, cat and monkey. J Comp Neurol 154:349-378
Kadekaro M, Crane AM, Sokoloff L (1985) Differential effects of electrical stimulation of sciatic nerve on metabolic activity in spinal cord and dorsal root ganglion in the rat. Proc Natl Acad Sci USA 82:6010-6013
Kanazawa I, Marshall GR, Kelly JS (1976) Afferents in the rat substantia nigra studied with horseradish peroxidase, with special reference to fibres from the subthalamic nucleus. Brain Res 115:485-491
Kernel ML, Desban M, Gauchy C, Glowinski J, Besson JM (1988) Topographical organization of efferent projections from the rat substantia nigra pars reticulata. Brain Res 455: 307-323
Kita H, Kitai ST (1987) Efferent projections of the subthalamic nucleus in the rat: light and electron microscopic analysis with the PHA-L method. J Comp Neurol 260:435-452
Kita H, Kitai ST (1991) Intracellular study of rat globus pallidus neurons: membrane properties and responses to neostriatal, subthalamic and nigral stimulation. Brain Res 564:296-305
Kita H, Chang HT, Kitai ST (1983a) Pallidal inputs to subthalamus: intracellular analysis. Brain Res 264:255-265
Kita H, Chang HT, Kitai ST ( I983b) The morphology of intracellularly labeled rat subthalamic neurons: a light microscopic analysis. J Comp Neurol 215:245-257
Kitai ST, Deniau JM (1981) Cortical inputs to the subthalamus: intracellular analysis. Brain Res 214:411-415
Kitai ST, Kita H (1987) Anatomy and physiology of the subthalamic nucleus: a driving force of the basal ganglia. In: The Basal Ganglia, Structure and Function, Vol 27: Advances in Behavioral Biology (Carpenter MB, Jaramayan A, eds), New York, Plenum Press, pp 357-373
Lindvall 0, Bjorklund A (1974) The organization of the ascending catecholamine neurons system in the rat brain as revealed by the glyoxylic acid fluorescence method. Acta Physiol Scand [Suppl) 412: 1-48
Loopuijt LD, Van der Kooy D (1985) Organization of the striatum: collateralization of its efferent axons. Brain Res 348: 86-99
Mata M, Fink DG, Gainer H, Smith CB, Davidson L, Savaki HE, Schwartz WJ, Sokoloff L (1980) Activity dependent energy metabolism in rat posterior pituitary primarily reflects sodium pump activity. J Neurochem 34:213-215
Mintz I, Hammond C, Guibert B, Leviel V (1986) Stimulation of the subthalamic nucleus enhances the release of dopamine in the rat substantia nigra. Brain Res 376:406-408
Mitchell IJ, Jackson A, Sambrook MA, Crossman AR (1985a) Common ventral mechanisms in experimental chorea and hemiballismus in the monkey. Evidence from 2-deoxyglucose autoradiography. Brain Res 339:346-350
Mitchell IJ, Sambrook MA, Crossman AR (1985b) Subcortical changes in the regional uptake of [3Hl-2-deoxyglucose in the brain of the monkey during experimental choreiform dyskinesia elicited by injection of a gamma-aminobutyric acid antagonist into the subthalamic nucleus. Brain 108:405-422
Morizumi T, Nakamura Y, Kitao Y, Kuro M (1987) Ultrastructural analysis of afferent terminals in the subthalamic nucleus of the cat with a combined degeneration and horseradish peroxidase tracing method. J Comp NeuroI265:159-174
Nakanishi H, Kita H, Kitai ST (1987) Intracellular study of rat substantia nigra pars reticulata neurons in an in vitro slice preparation: electrical membrane properties and response characteristics to subthalamic stimulation. Brain Res 437: 45-55
Nakanishi H, Kita H, Kitai ST (1991) Intracellular study of rat entopeduncular nuclear neurons in an in vitro slice preparation: response to subthalamic stimulation. Brain Res 549: 285-291
Nikoledis MAL, Chapin JK, Lin RCS (1992) Somatotopic maps within the zona incerta relay parallel GABAergic somatosensory pathways to the neocortex, superior colliculus, and brainstem. Brain Res 577:134-141
Nomura S, Mizuno N, Sugimoto T (1980) Direct projections from the pedunculopontine tegmental nucleus to the subthalamic nucleus in the cat. Brain Res 196:223-227
Perkins MN, Stone TW (1980) Subthalamic projections of the globus pallidus: an electrophysiological study in the rat. Exp NeuroI68:500-511
Perkins MN, Stone TW (1981) Iontophoretic studies on pallidal neurons and the projection from the subthalamic nucleus. J Exp Physiol 66:225-236
Ricardo JA (1980) Efferent connections of the subthalamic region in the rat I. The subthalamic nucleus of Luys. Brain Res 202:257-271
Ricardo JA (1981) Efferent connections of the subthalamic region in the rat II. The zona incerta. Brain Res 214:43-60
Robledo P, Feger J (1990) Excitatory influence of rat subthalamic nucleus to substantia nigra pars reticulata and the pallidal complex: electrophysiological data. Brain Res 518:47-54
Roger M, Cadusseau J (1985) Afferents to the zona incerta in the rat: a combined retrograde and anterograde study. J Comp NeuroI241:480-492
Romansky KV, U sunoff KG, Ivanov FP, Galabov GP (1979) Cortico-subthalamic projection in the cat: an electron microscopic study. Brain Res 163:319-322
Rouzaire-Dubois B, Scarnati E (1985) Bilateral corticosubthalamic nucleus projections: an electrophysiological study in rats with chronic cerebral lesions. Neuroscience 15:69-79
Rouzaire-Dubois B, Hammond C, Hammond B, Feger J (1980) Pharmacological blockade of the globus pallid us-induced inhibitory response of subthalamic cells in the rat. Brain Res 200:321-329
Rouzaire-Dubois B, Scarnati E, Hammond C, Crossman AR, Shibazaki T (1983) Microiontophoretic studies on the nature of neurotransmitter in the subthalamo-entopeduncular pathway of the rat. Brain Res 271: 11-20
Rouzaire-Dubois B, Hammond C, Velnik J, Feger J ( 1984) Anatomy and neurophysiology of the subthalamic efferent nucleus. In: The Basal Ganglia, Structure and Function, Vol 27: Advances in Behavioral Biology (McKenzie JS, Kemm RE, Wilcok LN, eds), New York, Plenum Press, pp 205-234
Rye DB, Saper CB, Lee HJ, Wainer BH (1987) Pedunculopontine tegmental nucleus of the rat: cytoarchitecture, cytochemistry and some extrapyramidal connections of the mesopontine tegmentum. J Comp Neurol 259:483-528
Sadikot AF, Parent A, Fran«ois C (1992) Efferent connections of the centromedian and parafascicular thalamic nuclei in the squirrel monkey: a PHA-L study of subcortical projections. J Comp Neurol 315:137-159
Saper CB, Loewy AD (1982) Projections of the pedunculopontine tegmental nucleus in the cat: evidence for additional extrapyramidal circuitry. Brain Res 252:367-372
Savaki HE (1989) The role of thalamic relays in the basal ganglia in interhemispheric relations, as demonstrated by the autoradiographic 14C-deoxyglucose method. In: Brain Imaging Techniques and Applications (Sharif NA, Lewis M.E, eds), Chichester, Ellis Horwood, pp 262-279
Savaki HE, Desban M, Glowinski J, Besson MJ (1983a) Local cerebral glucose consumption in the rat. I: effects of halothane anaesthesia. J Comp Neurol 213:36-45
Savaki HE, Desban M, Glowinski J, Besson MJ (1983b) Local cerebral glucose consumption in the rat. II: effects of unilateral substantia nigra stimulation in conscious and halothane anaesthetized animals. J Comp Neurol 213:46-65
Savaki HE, Girault JA, Desban M, Glowinski J, Besson MJ (1984) Adaptive reaction of nigral neurons following lesions
J Cereb Blood Flow Metab, Vol. 14, No. 1, 1994

144 M. TZAGOURNISSAKIS ET AL.
of their ventro-medial thalamic projection field. Brain Res 302: 1 90- 1 95
Savaki HE, Pizarro P, Dermon C, Arsenis S (1988) Deoxyglucose analysis of the specific topographic functional interrelations between substantia nigra and globus pallidus. Brain Res Bull 2 1 :855-863
Savaki HE, Dermon C, Raos V (1992) Bilateral cerebral metabolic effects of pharmacological manipulation of the substantia nigra in the rat: unilateral intranigral application of the inhibitory GABAA receptor agonist muscimol. Neuroscience 50:781-794
Schwartz WJ, Smith CB, Davidsen L, Savaki HE, Sokoloff L ( 1 979) Metabolic mapping of functional activity in the hypothalamo-neurohypophysial system of the rat. Science 206: 723-725
Seki M, Zyo K (1984) Anterior thalamic afferents from the mammillary body and the limbic cortex in the rat. J Comp Neurol 229: 242-256
Severin CM, Young PA, Massopust LC (1976) Pallidothalamic proj ections in the rat. J Comp Neurol 166:491-502
Shammah-Lagnado S J, Negrao N, Ricardo JA ( 1 985) Afferent connections of the zona incerta: a horseradish peroxidase study in the rat. Neuroscience 1 5 : 109-134
Sokoloff L ( 1977) Relation between physiological function and energy metabolism in the central nervous system. J Neurochern 29: 1 3-26
Sokoloff L, Reivich M, Kennedy C, Des Rosiers MH, Patlak CS, Pettigrew KD, S akurada 0 , S h inohara M ( 1 977) The [14CJdeoxyglucose method for the measurement of local cerebral glucose utilization: theory, procedure and normal val-
J Cereb Blood Flow Metab, Vol. 14, No. 1, 1994
ues in the conscious and anesthetized albino rat. J Neurochern 28:897-916
Sugimoto T, Hattori T, Mizuno N, Itoh K, Sato M ( 1983) Direct projections from the centre median-parafascicular complex to the subthalamic nucleus in the cat and rat. J Comp Neurol 2 1 4 : 209-216
Takada M, Nishihama MS, Nishihama CC, Hattori T ( 1988) Two separate populations of the rat subthalamic nucleus project to the basal ganglia and pedunculopontine tegmental regions. Brain Res 442:72-80
Van der Kooy D, Carter DA ( 1 98 1 ) The organization of the efferent projections and striatal afferents of the entopeduncular nucleus and adjacent areas in the rat. Brain Res 2 1 1 : 1 5-36
Van der Kooy D, Hattori T ( 1 980) Single subthalamic nucleus neurons proj ect to both the globus pallidus and substantia nigra in rat. J Comp Neurol 192:751-768
Van der Kooy D, Hattori T, Shannalz K, Homykiewicz 0 (1981 ) The pallido-subthalamic projection i n rat: anatomical and biochemical studies. Brain Res 204:253-268
Whittier JR (1947) Ballism and the subthalamic nucleus (nucleus hypothalamicus; corpus Luysi). Arch Neurol Psychiatry 58: 672-692
Whittier JR, Mettler FA ( 1 949) Studies on the subthalamus of the rhesus monkey. II. Hyperkinesia and other physiologic effects of subthalamic lesions, with special reference to the subthalamic nucleus of Luys. J Comp NeuroI90: 3 19-372
Woolf NJ, Butcher LL (1986) Cholinergic systems in the rat brain: III. Projection from the pontomesencephalic tegmentum to the thalamus, tectum, basal ganglia, and basal forebrain. Brain Res Bull 16:603-637
Top Related
Copyright © 2022 FDOKUMEN

