







Bahasa
Halaman
Hukum


Dendritic cells derived from TBP-2-deficient mice aredefective in inducing T cell responses
Aoi Son1, Hajime Nakamura2, Hiroaki Okuyama2, Shin-ichi Oka1,Eiji Yoshihara1,4, Wenrui Liu1, Yoshiyuki Matsuo1, Norihiko Kondo1,Hiroshi Masutani1, Yasuyuki Ishii3, Tomonori Iyoda5, Kayo Inaba5 andJunji Yodoi1,2
1 Department of Biological Responses, Institute for Virus Research, Kyoto University, Kyoto,Japan
2 Thioredoxin Project, Department of Experimental Therapeutics, Translational Research Center,Kyoto University Hospital, Kyoto, Japan
3 Laboratory for Vaccine Design, RIKEN Research Center for Allergy and Immunology (RCAI),Yokohama, Japan
4 Division of Systemic Life Science, Graduate School of Biostudies, Kyoto University, Kyoto, Japan5 Department of Animal Development and Physiology, Graduate School of Biostudies,Kyoto University, Kyoto, Japan
Thioredoxin-binding protein-2 (TBP-2), also known as vitamin D3-up-regulated protein 1(VDUP1), was identified as an endogenous molecule interacting with thioredoxin (TRX).Here, we show that dendritic cells (DC) derived from TBP-2-deficient mice are defective inthe function of T cell activation. To compare TBP-2–/– DC function with wild-type (WT) DC,we stimulated DC with lipopolysaccharide (LPS). Although TBP-2–/– DC and WT DCexpressed comparable levels of MHC class II and costimulatory molecules such as CD40,CD80 and CD86, the IL-12p40, IL-12p70 and IL-6 productions of TBP-2–/– DC wereattenuated. In a mixed leukocyte reaction (MLR), the concentrations of IL-2, IFN-c, IL-4 andIL-10 in the culture supernatant of MLR with TBP-2–/– DC were significantly lower thanthose in the cultures with WT DC. In MLR also, as with LPS stimulation, IL-12p40 andIL-12p70 production from TBP-2–/– DC was less than that from WT DC. Proliferation ofT cells cultured with TBP-2–/– DC was poorer than that with WT DC. In vivo delayed-typehypersensitivity responses in TBP-2–/– mice immunized with ovalbumin were significantlyreduced compared to WT mice. These results indicate that TBP-2 plays a crucial role in DCto induce T cell responses.
Key words: Dendritic cell � T cell responses � Thioredoxin-binding protein-2
Supporting Information available online
Introduction
Thioredoxin-binding protein-2 (TBP-2) was identified as anendogenous molecule interacting with thioredoxin (TRX) [1].
Since TBP-2 inhibits the reducing activity of TRX, it is consideredto be a negative regulator of the biological function of TRX. TBP-2was proved to be vitamin D3-up-regulated protein 1 (VDUP1),which was originally found as a vitamin D3-up-regulated gene inhuman HL-60 leukemia cells [2]. In our previous studies, we haveshown that TBP-2 plays an important role in the growth regulationof human T lymphocyte virus I (HTLV-I)-infected T cells [3] and
Aoi Son et al. Eur. J. Immunol. 2008. 38: 1358–13671358 DOI 10.1002/eji.200737939
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Correspondence: Aoi Sone-mail: [email protected]

that TBP-2 expression is associated with responsiveness to IL-2-dependent growth in HTLV-I-infected T cell lines [4]. We reportedthat fatty acid utilization is impaired in TBP-2–/– mice [5]. Bodnaret al. described a natural mutant mouse strain with decreasedmRNA expression for TBP-2/thioredoxin-interacting protein(Txnip), and these mice showed an abnormal lipid metabolism[6]. On the other hand, Lee et al. reported a significant decrease inNK cells and a reduction in their activity in TBP-2–/– mice [7]. It isalso known that vitamin D3 suppresses the differentiation and thematuration of dendritic cells (DC) [8–10].
DC are the most potent antigen-presenting cells capable ofstimulating naive T cells for primary immune responses, includingT cell tolerance [11]. The activation status of DC and the localcytokine milieu are important for determining the balancebetween immunity and tolerance.
IL-12 secreted fromDC polarizes naive Tcells toT helper (Th)1cells and simultaneously suppresses the differentiation of Th2cells. IL-12 also acts on resting NK cells and CD8+ T cells andinduces their cytotoxic activity. In contrast to the beneficial aspectsof IL-12 in host defense, excessive IL-12 leads to immunopathologyand host mortality, such as allograft rejection or autoimmunedisease [11–13]. DC are the most potent producers of IL-12, butnot all DC have the same capacity to produce IL-12. This is becauseDC change their effector functions depending on their maturationstage and activation signals from pathogens or cytokines [14].Thus, IL-12 produced by DC is a key cytokine for various immuneresponses, but the molecular mechanism underlying the regula-tion of IL-12 production in DC is not fully understood.
It has been described that a reducing microenvironment isrequired for lymphocyte activation [15]. Recently, it has beenreported that during antigen presentation, DC take up cystine andrelease cysteine and TRX [16]. Taken together, we hypothesizedthat TBP-2 might be involved in DC functions, such as antigenpresentation and IL-12 production. In the present study, weprepared splenic DC and bone marrow-derived DC (BMDC) fromTBP-2–/– mice, and compared their characteristics and functionswith those of wild-type (WT) mice.
Results
Reduced secretion of IL-12 and IL-6 in LPS-stimulatedTBP-2–/– DC
To examine the function of TBP-2–/– DC, first we stimulatedTBP-2–/– DC or WT DC with LPS. BMDC were differentiated fromthe bone marrow cells of WT or TBP-2–/– mice in the presence ofGM-CSF. After culture of the DC in the presence of LPS, thecytokine concentrations in the culture supernatants weredetermined. The concentrations of IL-12p40, IL-12p70 and IL-6in the supernatants of LPS-treated TBP-2–/– DC were significantlyless than those of theWT DCwhile the concentrations of IL-10 andTGF-b1 in the culture supernatants were comparable betweenTBP-2–/– and WT DC (Fig. 1A). The expression levels of CD11c,MHC class II, CD40, CD80 and CD86 on the cell surface before or
after stimulation were not significantly different between TBP-2–/–
and WT DC (Fig. 1B). These results collectively indicate that thereduction of IL-12 and IL-6 production by TBP-2–/– DC uponstimulation with LPS was not due to insufficient maturation of DCbut to defective signal transduction or secretion of IL-12 and IL-6.
Cytokine production of TBP-2–/– DC stimulated withalloreactive CD4+ T cells
To explore the cytokine production of TBP-2–/– DC with otherstimulation, we set up an allogeneic mixed leukocyte reaction(MLR) assay. BMDC were prepared from WT and TBP-2–/– miceand co-cultured with allogeneic CD4+ T cells for 48 or 72 h. Asshown in Fig. 2A, the concentrations of IL-2, IFN-c, IL-4, IL-10,IL-12p40 and IL-12p70 in the culture supernatant of MLR withTBP-2–/– DC were significantly lower than those in the cultureswith WT DC, despite the fact that the concentrations of TNF-a andTGF-b1 in the culture supernatants were comparable betweenTBP-2–/– and WT DC. From the MLR cultures, CD4+ Tcells and DCwere recovered by using magnetic beads conjugated to anti-CD4mAb and anti-CD11c mAb, respectively, and total RNA wasanalyzed by RT-PCR. Fig. 2B shows that the expression level ofIL-12p40 mRNA derived from TBP-2–/– DC was lower than thatfrom WT DC 48 h after co-culture. The expression levels of IL-4and IFN-c mRNA in CD4+ T cells cultured with TBP-2–/– DC wereremarkably lower than those with WT DC 72 h after co-culture.However, the expression levels of TRX mRNA were comparableamong the cell fractions. It is well known that mature DC canactivate naive T cells more than immature DC. To examine thepossibility that deficient maturation of DC caused the reduction inT cell activation in TBP-2–/– DC, WT DC and TBP-2–/– DC from theMLR cultures were stained with mAb specific for CD11c, MHCclass II, CD40, CD80 or CD86 and analyzed for the surfacemolecules. The fluorescence intensities of the TBP-2–/– DC werecomparable with those of the WT DC (Fig. 2C). These resultssuggest that the reduction of T cell activation capacity in TBP-2–/–
DC is not due to insufficient maturation but to other factors.
Impaired ability of TBP-2–/– DC to stimulate allo-CD4+
T cell proliferation in MLR
To investigate the proliferation of T cells in TBP-2–/– mice, weperformed MLR cultures. CD4+ Tcells were enriched by depletionof cells bound by an antibody cocktail specific for CD8a, CD11b,CD45R, DX5 and Ter-119 from spleens of BALB/c mice. DC wereprepared by anti-CD11c mAb-conjugated microbeads from thelow-density cell fraction in collagenase D-treated splenocytes ofWT mice (C57BL/6) or TBP-2–/– mice. CD4+ T cells were co-cultured with serially diluted DC for 3 days. Fig. 3A shows that[3H]thymidine uptake by the cells cultured with all dilutions ofTBP-2–/– DC was reduced by almost half compared with WT DC,but DC maturation or frequency in untreated spleens of TBP-2–/–
mice did not differ from that of WT mice (Fig. 3B). Conversely, in
Eur. J. Immunol. 2008. 38: 1358–1367 Immunomodulation 1359
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

cultures of CD4+ T cells with allogeneic CD11c+ DC, [3H]thymi-dine uptake by the T cells from TBP-2–/– mice was slightlyincreased compared with that of WT CD4+ Tcells (Fig. 3C). Theseresults indicate that, although the surface molecules on DC are notdifferent from WT DC, the ability of TBP-2–/– CD11c+ DC toactivate allogeneic CD4+ T cells might be defective.
TBP-2–/– mice were insensitive to delayed-typehypersensitivity
To test the capacity of in vivo DC function in TBP-2-deficient mice,delayed-type hypersensitivity (DTH) responses were performed.WT and TBP-2–/– mice were sensitized to ovalbumin (OVA) bysubcutaneous injection of OVA in CFA. At 7 days after immuniza-
tion, the mice were challenged by injection of OVA or the samevolume of PBS into the rear footpads. The footpad swelling wasmeasured 24 h later. The magnitude of the DTH response wasdetermined as the difference in footpad thickness before and afterchallenge. Footpad swelling in TBP-2–/– mice was significantlyreduced compared with WT mice (Fig. 4A). The expression levelsof CD11c, MHC class II, CD40, CD80 and CD86 on the cell surfacebefore or after stimulationwere not significantly different betweenpopliteal lymph nodes of TBP-2–/– and WT mice (Fig. 4B). We alsoexamined the production of antibodies. Mice were immunized i.p.with DNP-OVA in alum, and 2 wk later we took blood samples. Weanalyzed serum levels of antigen-specific IgG1 and IgG2a byELISA. In contrast to the DTH response, no changes were observedin the antibody response (Fig. 4C).
Figure 1. IL-12 and IL-6 secretions were decreased in LPS-stimulatedTBP-2–/– DC. (A) After 48-h stimulation, the levels of IL-12p40, IL-12p70,IL-6, IL-10 and TGF-b1 in culture supernatants weremeasured by ELISA.The data are representative of three independent experiments. *p<0.05.(B) Flow cytometric analysis ofWTor TBP-2–/– LPS-stimulated BMDC. DCwere stimulated with 1 lg/mL LPS for 48 h. CD11c, MHC class II, CD40,CD80 and CD86 expression on DC were analyzed by flow cytometry.
Aoi Son et al. Eur. J. Immunol. 2008. 38: 1358–13671360
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
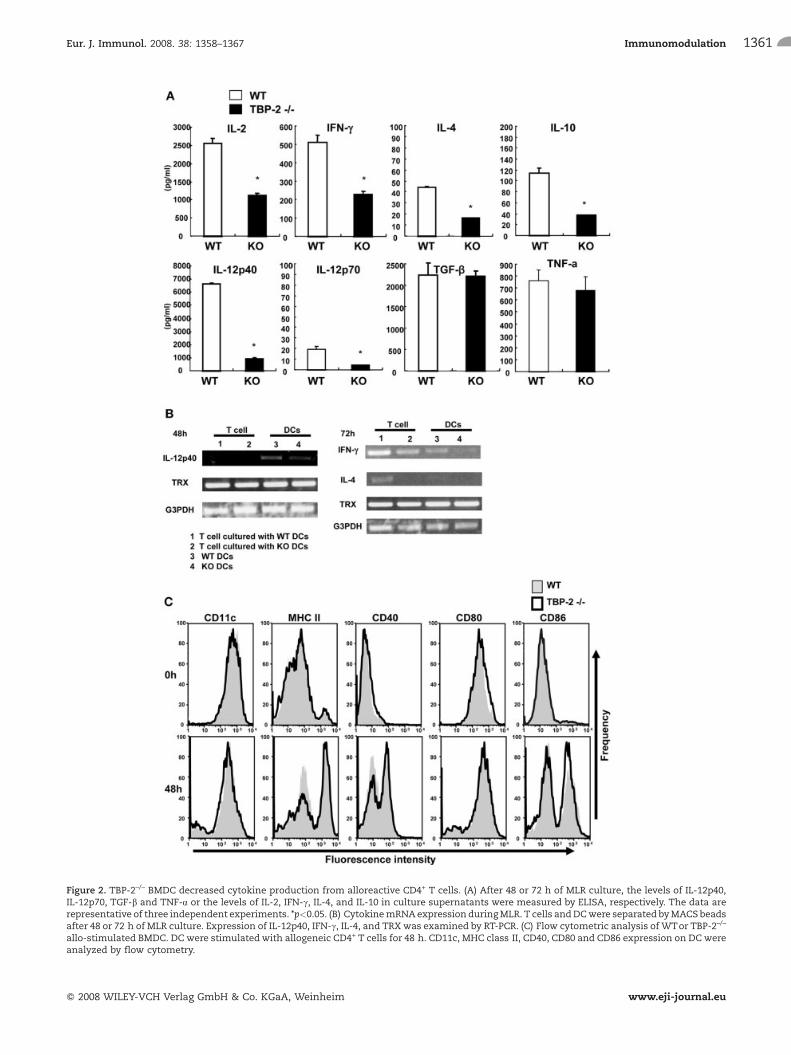
Figure 2. TBP-2–/– BMDC decreased cytokine production from alloreactive CD4+ T cells. (A) After 48 or 72 h of MLR culture, the levels of IL-12p40,IL-12p70, TGF-b and TNF-a or the levels of IL-2, IFN-c, IL-4, and IL-10 in culture supernatants were measured by ELISA, respectively. The data arerepresentative of three independent experiments. *p<0.05. (B) CytokinemRNAexpression duringMLR. T cells andDCwere separated byMACS beadsafter 48 or 72 h of MLR culture. Expression of IL-12p40, IFN-c, IL-4, and TRX was examined by RT-PCR. (C) Flow cytometric analysis of WTor TBP-2–/–
allo-stimulated BMDC. DC were stimulatedwith allogeneic CD4+ T cells for 48 h. CD11c, MHC class II, CD40, CD80 and CD86 expression on DC wereanalyzed by flow cytometry.
Eur. J. Immunol. 2008. 38: 1358–1367 Immunomodulation 1361
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Discussion
In this study, we demonstrated that TBP-2 is an importantregulator of DC-mediated T cell activation without affecting thephenotypic maturation of DC. It has been described that DCmaturation results in up-regulation of MHC class II and co-stimulatory molecules. Mature DC have a higher capacity foractivating Tcells than immature DC [11]. Although the expressionlevels of MHC class II, CD40, CD80 and CD86 of TBP-2–/– DC werenot significantly different from those of WT DC (Fig. 1B), with LPSstimulation, TBP-2–/– DC secreted less IL-12p40, IL-12p70 and IL-6thanWT DC (Fig. 1A). According to the cytokine profile elicited byMLR, cytokines secreted from T cells such as IL-2, IFN-c, IL-4 andIL-10 were attenuated in the supernatants of T cells cultured withTBP-2–/– DC (Fig. 2A). These results suggested that the ability of
TBP-2–/– DC to activate T cells is impaired. Using RT-PCR, weconfirmed the expression of IFN-c and IL-4 by T cells and IL-12 byDC in the MLR. The mRNA expression levels of IFN-c and IL-4 inT cells stimulated with TBP-2–/– DC were reduced. The mRNAexpression level of IL-12 inTBP-2–/– DCwas also reduced (Fig. 2B).Although the expression levels of MHC class II, CD40, CD80 andCD86 of TBP-2–/– DC were not significantly different from those ofWT DC, the proliferation of T cells activated by TBP-2–/– DC wasremarkably impaired (Fig. 3A and B). However, in cultures ofCD4+ Tcells with allogeneic CD11c+ DC, [3H]thymidine uptake bythe T cells from TBP-2–/– mice was conversely slightly increasedcompared with that by WT CD4+ T cells (Fig. 3C). These resultsalso suggested that TBP-2–/– DC are defective to induce T cellactivation, but it is unlikely that the inability of TBP-2–/– DC ismediated by these surface molecules.
Figure 3. TBP-2–/– DC were deficient in T cell priming. (A) TBP-2–/– DC attenuated allogeneic CD4+ T cell proliferation in MLR. CD4+ T cells (BALB/c)were cultured with WTor TBP-2–/– splenic DC for 3 days. Proliferation of T cells was determined by uptake of [3H]thymidine for the last 14 h of theculture. The figure represents the results from triplicate measurements of samples obtained from individual animals (n = 5). (B) Flow cytometricanalysis of WT and TBP-2–/– splenocytes. Splenocytes were obtained from WT and TBP-2–/– mice. CD11c, MHC class II, CD40, CD80 and CD86expression on DC were analyzed by flow cytometry. (C) TBP-2–/– CD4+ T cell proliferation in MLR was higher than for WT CD4+ T cells. CD4+ T cells(TBP-2–/– or WT) were cultured with splenic DC from BALB/c mice for 3 days. Proliferation of T cells was determined by uptake of [3H]thymidinefor the last 14 h of the culture. *p<0.05.
Aoi Son et al. Eur. J. Immunol. 2008. 38: 1358–13671362
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
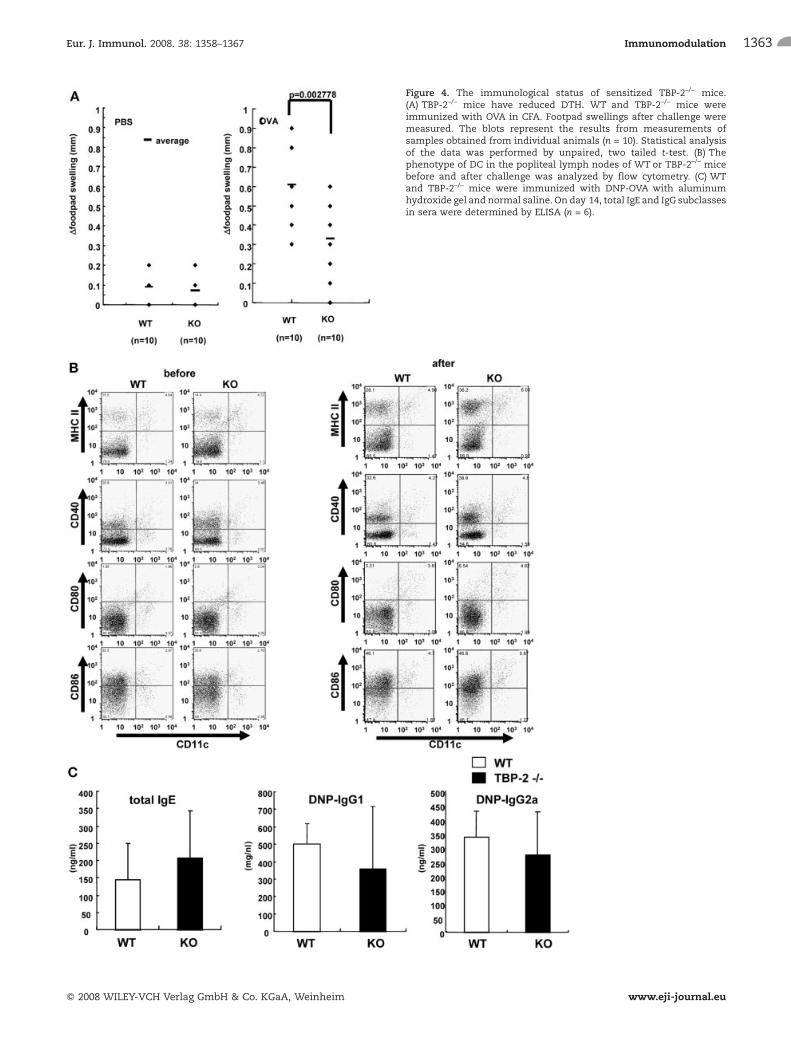
Figure 4. The immunological status of sensitized TBP-2–/– mice.(A) TBP-2–/– mice have reduced DTH. WT and TBP-2–/– mice wereimmunized with OVA in CFA. Footpad swellings after challenge weremeasured. The blots represent the results from measurements ofsamples obtained from individual animals (n = 10). Statistical analysisof the data was performed by unpaired, two tailed t-test. (B) Thephenotype of DC in the popliteal lymph nodes of WT or TBP-2–/– micebefore and after challenge was analyzed by flow cytometry. (C) WTand TBP-2–/– mice were immunized with DNP-OVA with aluminumhydroxide gel and normal saline. On day 14, total IgE and IgG subclassesin sera were determined by ELISA (n = 6).
Eur. J. Immunol. 2008. 38: 1358–1367 Immunomodulation 1363
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

The simple concept that immature DC are tolerogenic andmature DC are immunogenic has been revised by several reports.Recent studies have described tolerogenic DC and regulatory DC,which induce immunological tolerance [17]. DC can becometolerogenic when immature DC are cultured with certain cytokinesor immunosuppressive drugs. For example, vitamin D3, which up-regulates TBP-2 [2], controls DC differentiation and suppressesphenotypic and functional DC maturation [8–10, 18].
The biological effects of vitamin D3 are mediated by thevitamin D receptor (VDR), a member of the superfamily of nuclearhormone receptors. The discovery of VDR expression in a widevariety of immune cells, particularly in APC such as macrophagesand DC, as well as in both CD4+ and CD8+ T lymphocytes,prompted a number of studies investigating the capacity of VDRagonists to modulate T cell responses. Recently an increasingnumber of reports have demonstrated that VDR agonists markedlymodulate the phenotype and function of APC, in particular DC[19–21].
In vitro treatment of DCwith vitamin D3 or VDR agonists leadsto down-regulated expression of costimulatory molecules such asCD40, CD80 and CD86, markedly decreased IL-12 production andenhanced IL-10 production, resulting in inhibition of T cellactivation. The suppression of IL-12 production and the stronglyenhanced production of IL-10 highlight the important effects ofvitamin D3 and its analogs on DC [8–10, 18].
To investigate the effect of TBP-2 on the VDR signaling, wecultured bone marrow cells with vitamin D3 and developedBMDC. We observed that the decrease of MHC class II expressionon TBP-2–/– DC is comparable with that of WT DC (SupportingInformation Fig. 1). These results suggested that TBP-2 is notessential to transduce the VDR signaling.
As one group of the tolerogenic DC, there are CD11clow
CD45RBhigh DC in the spleen and lymph nodes of normal mice.CD45RB expression on TBP-2–/– DC was not higher than that onWT DC (data not shown). Vitamin D3, IL-10 and TGF-b1 can leadto the generation of tolerogenic DC [22]. These DC display animmature-like phenotype, but secrete high levels of IL-10 andinduce tolerance through the differentiation of Tr1 cells [23].
Collectively, TBP-2–/– DC are different from these regulatory ortolerogenic DC. First, the phenotype of TBP-2–/– DC was similar tothat of WT DC. The expression levels of MHC class II, CD40, CD80and CD86 on TBP-2–/– DC were not significantly different fromthose on WT DC (Fig. 1B, 2C, 3B and 4B). In addition, whenstimulated with allo-T cells or LPS, IL-10 production by TBP-2–/–
DC was not higher than that by WT DC (Fig. 1A and 2A). Takentogether, TBP-2–/– DC are not like so-called tolerogenic orregulatory DC, because the characteristics of TBP-2–/– DC werenot similar to those of tolerogenic DC.
Moreover, we examined the in vivo immunological responsesof TBP-2–/– mice. Although the expression levels of MHC class II,CD40, CD80 and CD86 of TBP-2–/– DC in the popliteal lymphnodes were not significantly different from those of WT DC(Fig. 4B), DTH responses in TBP-2–/– mice were significantlyreduced compared with those in WT mice (Fig. 4A). We alsoexamined serum levels of IgE and IgG subclasses. The levels of total
IgE, antigen-specific IgG1 and IgG2a of TBP-2–/– mice were notdifferent from those of WT mice (Fig. 4C).
TRX is a well-known inhibitor of apoptosis signal-regulatingkinase 1 (ASK-1). It has been reported that ASK-1–/– mice areresistant to LPS-induced septic shock by regulating reactive oxygenspecies (ROS)-mediated activation of TRAF6-ASK1-p38 LPSsignaling [24]. A recent report demonstrated that antigen-specific,bidirectional DC-T cell communication can be blocked byinterfering with the redox regulation pathways [25]. It has alsobeen reported that the increase in ROS was detected in the processof antigen presentation and that hydrogen peroxide activates DC,indicating that the generation of oxidants is important for DCfunctions [26, 27].
We speculate that the interaction of TRX and TBP-2 might beresponsible for the inability of TBP-2–/– DC to activate T cells.However, the expression of TRX mRNA of TBP-2–/– DC was notdifferent from that of WT DC (Fig. 2B). The TRX-reducing activityof TBP-2–/– DC was also comparable to WT DC (SupportingInformation Fig. 2). Another group has also reported that there isno difference in thioredoxin-reducing activity between livers ofTBP-2–/– mice and WT mice after an overnight fast [28].
In summary, TBP-2–/– DC attenuated T cell responses. TBP-2plays an important role in IL-12 and IL-6 production by DC.Further investigation of the molecular mechanism underlying theregulation of IL-12 and IL-6 transcription by TBP-2 in DC mayprovide a cue for understanding the decreased ability of TBP-2–/–
DC to activate naive T cells.
Mice
Female C57BL/6 and BALB/cmice 8–12 wk of age were purchasedfrom Japan SLC (Hamamatsu, Japan). TBP-2–/– mice wereestablished as described [5]. F5 or further backcrossed C57BL/6mice were used for this study. All procedures involving animalswere performed in accordance with protocols approved by theInstitute for Virus Research at Kyoto University.
Preparation of splenic DC
Spleens were treated with 1 mg/mL collagenase D (RocheMolecular Biochemicals) in PBS containing 10 mM Hepes buffer(pH 7) and incubated for 45 min at 37�C. After the recovery ofspleen cells, red blood cells were lysed by red blood cell lysingbuffer (Sigma, St. Louis, MO). The cells were subsequentlysuspended in 3 mL 30% BSA. After the addition of 5 mL RPMI1640 medium, the cell suspension was centrifuged at 1700 � g for25 min and the low-density cell fraction was recovered. CD11c+
cells in the low-density cells were isolated by using anti-CD11cmAb-conjugated MACS beads (Miltenyi Biotec, Bergisch Glad-bach, Germany).
Aoi Son et al. Eur. J. Immunol. 2008. 38: 1358–13671364
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Generation of BMDC
BMDCwere prepared as described [29]. Briefly, bonemarrow cellswere flushed from femurs and tibiae and passed through a 40 lmcell strainer. Next the cells (2 � 105/mL) were cultured in RPMI1640 medium supplemented with 10% FBS and 20 ng/mLrecombinant murine GM-CSF (eBioscience, San Diego, CA) in100-mm petri dishes. After 4 days, 10 mL of medium containing20 ng/mL GM-CSF was added to the plates. Nonadherent cellswere collected by gentle pipetting on day 8, and CD11c+ cells werepurified using anti-CD11c MACS beads (Miltenyi Biotec).
MLR assay
Splenocytes were obtained from 8–12-wk-old BALB/c, C57BL/6,or TBP-2–/– mice by passing through a cell strainer. After depletionof red blood cells in red blood cell lysing buffer, CD4+ T cells wereenriched by using a CD4+ T cell isolation MACS kit (MiltenyiBiotec), following the manufacturer's instructions. Briefly, CD4+
T cell were enriched by depletion of cells bound by an antibodycocktail specific for CD8a, CD11b, CD45R, DX5 and Ter-119 fromspleens of mice. CD4+ T cells (1 � 105/well) and allogeneicsplenic DC or BMDC were co-cultured for 3 days. [3H]Thymidinewas added for the last 14 h.
Flow cytometric analysis
For analysis of cell surface molecules, cells were stained withantibodies at 4�C for 20 min. The following antibodies fromeBioscience (San Diego, CA) were used: PE-conjugated anti-CD80(16–10A1), PE-conjugated anti-CD86 (GL1), FITC-conjugatedanti-CD40 (HM40-3), and FITC-conjugated anti-MHC class II(M5/114.15.2). Allophycocyanin-conjugated anti-CD11c mAb(HL3) was purchased from BD Pharmingen (San Diego, CA).Flow cytometric analysis was performed using a FACSCalibur (BDBiosciences, Mountain View, CA) with FlowJo software (Tree Star,Ashland, OR).
Cytokine measurement
Cytokine levels in culture supernatants were determined usingELISA kits for IL-2, IFN-c, IL-4, IL-6, IL-10, IL-12p40, IL-12p70,TNF-a and TGF-b1 (R&D Systems, Minneapolis, MN).
RT-PCR
Total RNAwas isolated fromDC or Tcells by using the RNeasyMiniKit (Qiagen, Hilden, Germany). cDNA was amplified by theSuperScript II First-strand Synthesis System for RT-PCR (Invitro-gen, Carlsbad, CA). The RT-PCR conditions were: for GAPDH,forward primer 50-ACCACAGTCCATGCCATCAC-30, reverse primer
50-TCCACCACCCTGTTGCTGTA-30, 24 cycles at 94�C for 30 s,55�C for 5 s and 74�C for 30 s; for TRX, forward primer50-ATGGTGAAGCTGATCGAGA-30, reverse primer50-CAGTAATAGAGGCTTCAAGC-30, 27 cycles at 94�C for 30 s,57�C for 45 s and 72�C for 1 min 30 s. IL-12p40, IFN-c and IL-4were assessed using appropriate pairs of primers describedpreviously [30–32].
DTH response
WT and TBP-2–/– mice were sensitized by subcutaneous injectionof 100 lL OVA (2.5 mg/mL; LSL, Japan) in CFA (ICN, Aurora,OH). At 7 days after immunization, mice were challenged byinjection of 30 lL OVA (5 mg/mL) or the same volume of PBS intothe rear footpads. Measurements of footpad swelling were taken24 h later using digital calipers (Mitsutomo, Japan). Themagnitude of the DTH response was determined by the differenceof footpad thickness before and after challenge.
Determination of serum levels of IgE and IgGsubclasses
Mice were sensitized by an i.p. injection of DNP-OVA (100 mg) inaluminum hydroxide gel and normal saline. On day 14, mice werebled and sera were prepared.
Total IgE in sera was determined by means of an ELISA kit(Morinaga, Yokohama, Japan). IgG subclasses in sera weredetermined by ELISA. Briefly, 96-well microplates (Nunc,Wiesbaden-Schierstein, Germany) were coated with DNP-BSA in50 mM carbonate buffer (pH 9.0) and incubated overnight at 4�C.After blocking, diluted samples were added and incubated for 3 hat RT. DNP-specific IgG1 or IgG2a were detected using horseradishperoxidase (HRP)-labeled anti-mouse IgG1 or IgG2a (Zymed, SanFrancisco, CA), respectively. The colorimetric reaction wasdeveloped by TNB+ substrate-chromogen (DAKO, Carpinteria,CA). The optical density of each well was determined by using amicroplate reader.
Statistical analysis
Statistical analysis was performed by Student's t-test; p values<0.05 were considered significant.
Acknowledgements: We thank Drs. Kimishige Ishizaka andChikako Nishigori for helpful discussions; Ms. Ryoko Otsuki,Ms. Suzuyo Furukawa, Mr. Masahiro Takenaka and Mr. AkiraYamada for technical assistance; Ms. Chisayo Kubo and TomoNakagawa for secretarial help. This study was supported by aGrant-in-Aid for Scientific Research from the Ministry of
Eur. J. Immunol. 2008. 38: 1358–1367 Immunomodulation 1365
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Education, Culture, Sports, Science and Technology of Japan, andby the Program for Promotion of Fundamental Studies in HealthSciences of the National Institute of Biomedical Innovation(NIBIO).
Conflict of interest: The authors declare no financial orcommercial conflict of interest.
References
1 Nishiyama, A., Matsui, M., Iwata, S., Hirota, K., Masutani, H.,Nakamura, H., Takagi, Y. et al., Identification of thioredoxin-bindingprotein-2/vitamin D(3) up-regulated protein 1 as a negative regulator ofthioredoxin function and expression. J. Biol. Chem.1999. 274: 21645–21650.
2 Chen, K. S. and DeLuca, H. F., Isolation and characterization of a novelcDNA from HL-60 cells treated with 1,25-dihydroxyvitamin D-3. Biochim.Biophys. Acta 1994. 1219: 26–32.
3 Nishinaka, Y., Nishiyama, A., Masutani, H., Oka, S., Ahsan, K. M.,Nakayama, Y., Ishii, Y. et al., Loss of thioredoxin-binding protein-2/vitamin D3 up-regulated protein 1 in human T-cell leukemia virus type I-dependent T-cell transformation: Implications for adult T-cell leukemialeukemogenesis. Cancer Res. 2004. 64: 1287–1292.
4 Ahsan, M. K., Masutani, H., Yamaguchi, Y., Kim, Y. C., Nosaka, K.,Matsuoka, M., Nishinaka, Y. et al., Loss of interleukin-2-dependency inHTLV-I-infected T cells on gene silencing of thioredoxin-binding protein-2.Oncogene 2006. 25: 2181–2191.
5 Oka, S., Liu, W., Masutani, H., Hirata, H., Shinkai, Y., Yamada, S.,Yoshida, T. et al., Impaired fatty acid utilization in thioredoxin bindingprotein-2 (TBP-2)-deficient mice: A unique animal model of Reye syndrome.FASEB J. 2006. 20: 121–123.
6 Bodnar, J. S., Chatterjee, A., Castellani, L. W., Ross, D. A., Ohmen, J.,Cavalcoli, J., Wu, C. et al., Positional cloning of the combinedhyperlipidemia gene Hyplip1. Nat. Genet. 2002. 30: 110–116.
7 Lee, K. N., Kang, H. S., Jeon, J. H., Kim, E. M., Yoon, S. R., Song, H., Lyu,C. et al., VDUP1 is required for the development of natural killer cells.Immunity 2005. 22: 195–208.
8 Hackstein, H. and Thomson, A. W., Dendritic cells: Emerging pharma-cological targets of immunosuppressive drugs. Nat. Rev. Immunol. 2004. 4:24–34.
9 Penna, G. and Adorini, L., 1Alpha,25-dihydroxyvitamin D3 inhibitsdifferentiation, maturation, activation, and survival of dendritic cellsleading to impaired alloreactive T cell activation. J. Immunol. 2000. 164:2405–2411.
10 Piemonti, L., Monti, P., Sironi, M., Fraticelli, P., Leone, B. E., Dal Cin, E.,Allavena, P. and Di Carlo, V., Vitamin D3 affects differentiation,maturation, and function of human monocyte-derived dendritic cells.J. Immunol. 2000. 164: 4443–4451.
11 Banchereau, J., Briere, F., Caux, C., Davoust, J., Lebecque, S., Liu, Y. J.,Pulendran, B. and Palucka, K., Immunobiology of dendritic cells. Annu.Rev. Immunol. 2000. 18: 767–811.
12 Kapsenberg, M. L., Dendritic-cell control of pathogen-driven T-cellpolarization. Nat. Rev. Immunol. 2003. 3: 984–993.
13 Gately, M. K., Renzetti, L. M., Magram, J., Stern, A. S., Adorini, L.,Gubler, U. and Presky, D. H., The interleukin-12/interleukin-12-receptorsystem: Role in normal and pathologic immune responses. Annu. Rev.Immunol. 1998. 16: 495–521.
14 Liu, Y. J., Kanzler, H., Soumelis, V. and Gilliet, M., Dendritic cell lineage,plasticity and cross-regulation. Nat. Immunol. 2001. 2: 585–589.
15 Ishii, T., Hishinuma, I., Bannai, S. and Sugita, Y., Mechanism of growthpromotion of mouse lymphoma L1210 cells in vitro by feeder layer or2-mercaptoethanol. J. Cell. Physiol. 1981. 107: 283–293.
16 Angelini, G., Gardella, S., Ardy, M., Ciriolo, M. R., Filomeni, G., DiTrapani, G., Clarke, F. et al., Antigen-presenting dendritic cells provide thereducing extracellular microenvironment required for T lymphocyteactivation. Proc. Natl. Acad. Sci. USA 2002. 99: 1491–1496.
17 Rutella, S., Danese, S. and Leone, G., Tolerogenic dendritic cells: Cytokinemodulation comes of age. Blood 2006. 108: 1435–1440.
18 D'Ambrosio, D., Cippitelli, M., Cocciolo, M. G., Mazzeo, D., Di Lucia, P.,Lang, R., Sinigaglia, F. and Panina-Bordignon, P., Inhibition of IL-12production by 1,25-dihydroxyvitamin D3. Involvement of NF-kappaBdownregulation in transcriptional repression of the p40 gene. J. Clin.Invest. 1998. 101: 252–262.
19 Adorini, L., Giarratana, N. and Penna, G., Pharmacological induction oftolerogenic dendritic cells and regulatory T cells. Semin. Immunol. 2004. 16:127–134.
20 Griffin, M. D., Lutz, W. H., Phan, V. A., Bachman, L. A., McKean, D. J. andKumar, R., Potent inhibition of dendritic cell differentiation and maturationby vitamin D analogs. Biochem. Biophys. Res. Commun. 2000. 270: 701–708.
21 Griffin, M. D., Lutz, W., Phan, V. A., Bachman, L. A., McKean, D. J. andKumar, R., Dendritic cell modulation by 1alpha,25 dihydroxyvitamin D3and its analogs: A vitamin D receptor-dependent pathway that promotes apersistent state of immaturity in vitro and in vivo. Proc. Natl. Acad. Sci. USA2001. 98: 6800–6805.
22 Sato, K., Yamashita, N., Baba, M. and Matsuyama, T., Modified myeloiddendritic cells act as regulatory dendritic cells to induce anergic andregulatory T cells. Blood 2003. 101: 3581–3589.
23 Wakkach, A., Fournier, N., Brun, V., Breittmayer, J. P., Cottrez, F. andGroux, H., Characterization of dendritic cells that induce tolerance andT regulatory 1 cell differentiation in vivo. Immunity 2003. 18: 605–617.
24 Matsuzawa, A., Saegusa, K., Noguchi, T., Sadamitsu, C., Nishitoh, H.,Nagai, S., Koyasu, S. et al., ROS-dependent activation of the TRAF6-ASK1-p38 pathway is selectively required for TLR4-mediated innate immunity.Nat. Immunol. 2005. 6: 587–592.
25 Matsue, H., Edelbaum, D., Shalhevet, D., Mizumoto, N., Yang, C.,Mummert, M. E., Oeda, J. et al., Generation and function of reactiveoxygen species in dendritic cells during antigen presentation. J. Immunol.2003. 171: 3010–3018.
26 Verhasselt, V., Goldman, M. and Willems, F., Oxidative stress up-regulatesIL-8 and TNF-alpha synthesis by human dendritic cells. Eur. J. Immunol.1998. 28: 3886–3890.
27 Rutault, K., Alderman, C., Chain, B. M. and Katz, D. R., Reactive oxygenspecies activate human peripheral blood dendritic cells. Free Radic. Biol.Med. 1999. 26: 232–238.
28 Chutkow, W. A., Patwari, P., Yoshioka, J. and Lee, R. T., Thioredoxin-interacting protein (Txnip) is a critical regulator of hepatic glucoseproduction. J. Biol. Chem. 2008. 283: 2397–2406.
29 Lutz, M. B., Kukutsch, N., Ogilvie, A. L., Rossner, S., Koch, F., Romani, N.and Schuler, G., An advanced culture method for generating largequantities of highly pure dendritic cells from mouse bone marrow.J. Immunol. Methods 1999. 223: 77–92.
30 Son, A., Nakamura, H., Kondo, N., Matsuo, Y., Liu, W., Oka, S., Ishii, Y.and Yodoi, J., Redox regulation of mast cell histamine release inthioredoxin-1 (TRX) transgenic mice. Cell Res. 2006. 16: 230–239.
Aoi Son et al. Eur. J. Immunol. 2008. 38: 1358–13671366
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

31 Goh, K. C., deVeer, M. J. and Williams, B. R., The protein kinase PKR isrequired for p38 MAPK activation and the innate immune response tobacterial endotoxin. EMBO J. 2000. 19: 4292–4297.
32 Ilangumaran, S., Finan, D., Raine, J. and Rottapel, R., Suppressor ofcytokine signaling 1 regulates an endogenous inhibitor of a mast cellprotease. J. Biol. Chem. 2003. 278: 41871–41880.
Abbreviations:DTH: delayed-type hypersensitivity � TBP-2: thioredoxin-binding protein-2 � TRX: thioredoxin � VDR: vitamin D receptor
Full correspondence: Junji Yodoi, Department of Biological Responses,Institute for Virus Research, Kyoto University, 53 Kawahara-MachiSyogoin Sakyo-Ku, Kyoto, 606–8507, JapanFax: +81-75-7615766e-mail: [email protected]
Supporting Information for this article is available atwww.wiley-vch.de/contents/jc_2040/2008/37939_s.pdf
Received: 23/10/07Revised: 8/2/08Accepted: 6/3/08
Eur. J. Immunol. 2008. 38: 1358–1367 Immunomodulation 1367
f 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Top Related
Copyright © 2022 FDOKUMEN

